
Induction of diploid gynogenesis using interspecific sperm and production of tetraploids in African...
11
Ž . Aquaculture 173 1999 401–411 Induction of diploid gynogenesis using interspecific sperm and production of tetraploids in African ž / catfish, Clarias gariepinus Burchell 1822 Laszlo Varadi ) , Istvan Benko, Janos Varga, Laszlo Horvath ´ ´ ´ ´ ´ ´ ´ ´ ´ Laboratory of Fish Culture, Institute of Animal Husbandry, Godollo UniÕersity of Agricultural Sciences, ¨¨˜ H-2100 Godollo, Pater K.u. 1., Hungary ¨¨˜ ´ Abstract Ž Homozygous diploid gynogenetic and tetraploid progeny of African catfish Clarias gariepi- . nus were produced by the suppression of the first mitotic division by applying thermal shock. The Ž . duration of one mitotic cycle during synchronous cell divisions t -value was used as unit after 0 fertilization for expression of the time of development. After determination of sensitive and non-sensitive periods, a temperature of 40.58C for 2 min was subsequently used. Diploid Ž . gynogenesis was obtained 6.34 "2.35% survivors at swimming stage when eggs were fertilized Ž . with genetically inactivated rosy barb Barbus conchonius sperm. We have not found any embryos using intact interspecific sperm. When African catfish sperm was used for fertilization a Ž . ratio of 9.2% tetraploids was obtained mixed eggs from four females . The highest frequency of both gynogens and tetraploids was observed when the heat shock was initiated at 1.4 t -value. We 0 used electrophoresis, chromosomes and nucleoli counting for identification of homozygosity and tetraploidy. q 1999 Elsevier Science B.V. All rights reserved. Keywords: Tetraploidy; Interspecific gynogenesis; Rosy barb; African catfish; Clarias gariepinus 1. Introduction Gynogens are obtained from shocks to fish ova activated with genetically inactivated sperm. Mitotic gynogenetic diploids are of great interest because completely homozy- Ž . gous lines clones can be produced in two generations, whereas with meiotic gynogene- Ž sis it requires several generations to achieve near, but complete homozygosity Nagy and . Csanyi, 1982 . Inbred lines have a wide variety of practical and theoretical applications, ´ such as the production of superior lines by crossbreeding and the study of single locus ) Corresponding author. E-mail: [email protected] 0044-8486r99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved. Ž . PII: S0044-8486 98 00467-0
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Induction of diploid gynogenesis using interspecific sperm and production of tetraploids in African...

Ž .Aquaculture 173 1999 401–411
Induction of diploid gynogenesis using interspecificsperm and production of tetraploids in African
ž /catfish, Clarias gariepinus Burchell 1822
Laszlo Varadi ), Istvan Benko, Janos Varga, Laszlo Horvath´ ´ ´ ´ ´ ´ ´ ´ ´Laboratory of Fish Culture, Institute of Animal Husbandry, Godollo UniÕersity of Agricultural Sciences,¨ ¨ ˜
H-2100 Godollo, Pater K.u. 1., Hungary¨ ¨ ˜ ´
Abstract
ŽHomozygous diploid gynogenetic and tetraploid progeny of African catfish Clarias gariepi-.nus were produced by the suppression of the first mitotic division by applying thermal shock. The
Ž .duration of one mitotic cycle during synchronous cell divisions t -value was used as unit after0
fertilization for expression of the time of development. After determination of sensitive andnon-sensitive periods, a temperature of 40.58C for 2 min was subsequently used. Diploid
Ž .gynogenesis was obtained 6.34"2.35% survivors at swimming stage when eggs were fertilizedŽ .with genetically inactivated rosy barb Barbus conchonius sperm. We have not found any
embryos using intact interspecific sperm. When African catfish sperm was used for fertilization aŽ .ratio of 9.2% tetraploids was obtained mixed eggs from four females . The highest frequency of
both gynogens and tetraploids was observed when the heat shock was initiated at 1.4 t -value. We0
used electrophoresis, chromosomes and nucleoli counting for identification of homozygosity andtetraploidy. q 1999 Elsevier Science B.V. All rights reserved.
Keywords: Tetraploidy; Interspecific gynogenesis; Rosy barb; African catfish; Clarias gariepinus
1. Introduction
Gynogens are obtained from shocks to fish ova activated with genetically inactivatedsperm. Mitotic gynogenetic diploids are of great interest because completely homozy-
Ž .gous lines clones can be produced in two generations, whereas with meiotic gynogene-Žsis it requires several generations to achieve near, but complete homozygosity Nagy and
.Csanyi, 1982 . Inbred lines have a wide variety of practical and theoretical applications,´such as the production of superior lines by crossbreeding and the study of single locus
) Corresponding author. E-mail: [email protected]
0044-8486r99r$ - see front matter q 1999 Elsevier Science B.V. All rights reserved.Ž .PII: S0044-8486 98 00467-0

( )L. Varadi et al.rAquaculture 173 1999 401–411´402
effects. Clonal lines have been produced in a variety of commercial and non-commercialŽ . Žfish species e.g., in zebrafish Danio rerio, Streisinger et al., 1981 , medaka Oryzias
. Ž .latipes, Naruse et al., 1985 and carp Cyprinus carpio, Komen et al., 1991 . The maininterest in tetraploids is the production of auto-allotriploids by mating tetraploids withnormal diploids. On the other hand progenies from tetraploid matings had levels ofheterozygosity higher than those consistent with tetrasomic inheritance.
Mitotic gynogens have already been obtained in different fish species includingaquacultured and non-commercial ones; examples include mitotic gynogens in ayuŽ . ŽPlecoglossus altiÕelis, Taniguchi et al., 1990 , koi carp Cyprinus carpio, Cherfas et
. Ž .al., 1993 various salmonid species Onozato, 1984; Chourrout, 1984 and polyploids inŽ .rainbow trout Oncorhyncus mykiss, Thorgaard et al., 1981; Chourrout, 1984 channel
Ž . Žcatfish Ictalurus punctatus, Bidwell et al., 1985; Wolters et al., 1981 and tench Tinca.tinca, Flajshans et al., 1993 .
The only chromosome set manipulations reported in African catfish have beenŽinduced triploidy and meiogynogenesis Henken et al., 1987; Volckaert et al., 1993,
.1994, 1997; Galbusera, 1997 despite their economical importance and easy propagation.Our aims included the development of the most effective and repeatable timing of heatshock for suppression of the first mitotic cleavage of the embryo. We postulated thatrosy barb sperm would be suitable for mitotic gynogenesis because it can activate catfishegg, but would not produce viable hybrids. Use of heterologous sperm to assure thatproduction of viable fish are only gynogens has precedence in many previous studies,
Ž . Ž .e.g., Na-nakorn et al. 1993 and Chourrout 1982 . Tetraploid progenies were plannedto be obtained by using shock when catfish eggs were fertilized with intact catfishsperm.
2. Materials and methods
2.1. Egg collection, artificial fertilization and larÕaculture
Ž . Ž .African catfish Cl Clarias gariepinus Burchell, 1822 were obtained from TEHAG,Ž .Szazhalombatta Hungary . Sexually mature African catfish females were injected with´
Ž y1 .dried carp hypophysis 5–5.5 mg kg body weight dissolved in 0.65% saline atapproximately 10 h prior to stripping at 268C. The handstripped eggs were fertilizedartificially with sperm obtained from surgically removed testis.
Ž . ŽRosy barb B Barbus conchonius broodstock were kept in 40 l tanks 10–12 fish per.tank , at 268C, on a 14–10 h light–dark periods. Sperm was obtained by stripping using
a pipette and diluting 1:10 in 1% NaCl solution. Hatched larvae were reared inŽ .flow-through water at 268C . Larvae were fed with Artemia and trout starter food
Ž . Ž .Tagger, Austria ; fry to the age of 4 weeks were fed with cut Tubifex and from age 4weeks to maturity, they were fed with Tubifex and trout food.
2.2. Heat shock and genetic inactiÕation of the sperm
Ž .The eggs were placed in a hot bath every 0.2 t from 0.6–1.8 . Temperature during0
heat shock was 39–428C, and the shock had a duration of 2 to 3 min in the different

( )L. Varadi et al.rAquaculture 173 1999 401–411´ 403
experiments. The heat shock was applied by submerging the basket in a temperature-Ž .controlled water bath "0.28C . The time of shock initiation was expressed in the
Ž .relative dimensionless unit t -value according to Cherfas et al. 1993 which is0
equivalent to the duration of the mitotic cycle during synchronous cell divisions at theŽinitial stages of embryogenesis in our experiments one t -value was from the onset to0
.the end of second mitotic cleavage . The first cleavage is completed at 2.2–2.3 t . The0Ž .diluted milt was exposed to ultraviolet irradiation VL-6C UV lamp 7.5 W, 254 nm at
0.5 J cmy2 in a glass tube on ice, continually stirred by hand. Such treatment makes thesperm genetically inactive but retains most of its activating capacity. The survival ofembryos and the motility of the sperm was tested after irradiation by microscopicexamination.
2.3. Production of tetraploids
The eggs from four females were mixed. Approximately 300 eggs in each experimentwere fertilized with African catfish sperm and heat shocked at 0.6, 0.8, 1.0, 1.2, 1.4, 1.6or 1.8 t after fertilization.0
2.4. Production of gynogens
Three females were used in this study. The approximately 300 eggs in eachexperiment were fertilized with non-irradiated or irradiated sperm from African catfishor rosy barb males and heat shocked at 1.4 t for 2 min after fertilization.0
2.5. Electrophoretic analyses
We scored the gynogenetic progeny for genetic variation at biochemical loci.Ž .Electrophoretic analysis of the lactate dehydrogenase LDH , malate dehydrogenase
Ž . Ž .MDH and esterase EST loci was used for testing the introgression of the malegenome of rosy barb. A fin clip from 2 to 3 week-old individuals was removed and
Ž .analyzed by vertical PAGE electrophoresis as published by Morizot and Schmidt 1990Ž .with the modification of Marian et al. 1984 . Identification and nomenclature of´ ´
Ž .enzymes followed Shaklee et al. 1990 . It was not difficult to differentiate the twospecies with the help of electrophoresis because there was a great difference in themobility of the proteins of the African catfish and the rosy barb.
2.6. Determination of ploidy status
Chromosome and nucleolus counting were used for verifying the ploidy status. TwoŽ y1 .to ten days post-hatch fry were treated by vinblastine REANAL 10 mg ml for 2 h
Ž .then incubated in a hypotonic solution distilled water for 20 min and fixed overnight ina 3:1 mixture of methanol and acetic acid. Air-dried slides were prepared according to
Ž .Yamazaki et al. 1981 and stained with 5% Giemsa. Ploidy status of each individual
( )L. Varadi et al.rAquaculture 173 1999 401–411´404
were determined based on at least 10 countable metaphase spreads. The number ofŽ .nucleoli per nucleus was examined and analyzed as described by Cherfas et al. 1993 .
3. Results
3.1. Optimization of heat-shock parameters
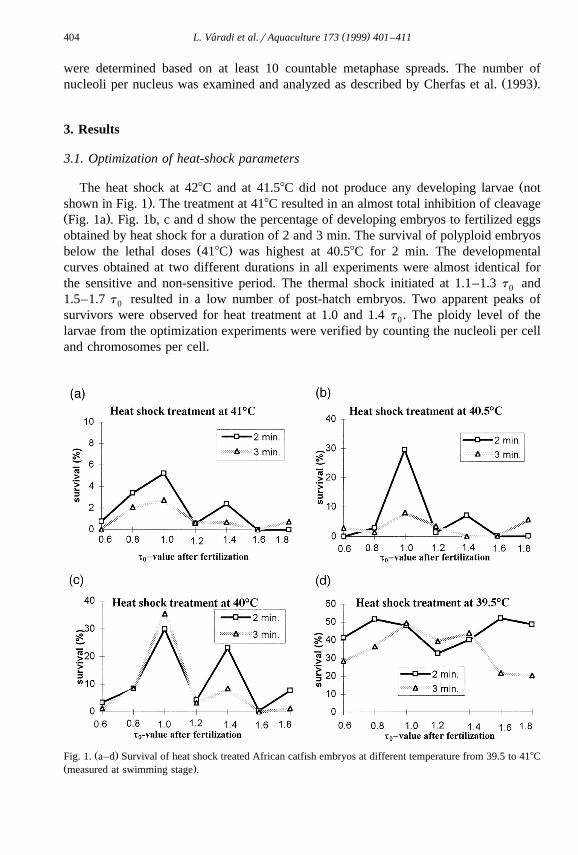
ŽThe heat shock at 428C and at 41.58C did not produce any developing larvae not.shown in Fig. 1 . The treatment at 418C resulted in an almost total inhibition of cleavage
Ž .Fig. 1a . Fig. 1b, c and d show the percentage of developing embryos to fertilized eggsobtained by heat shock for a duration of 2 and 3 min. The survival of polyploid embryos
Ž .below the lethal doses 418C was highest at 40.58C for 2 min. The developmentalcurves obtained at two different durations in all experiments were almost identical forthe sensitive and non-sensitive period. The thermal shock initiated at 1.1–1.3 t and0
1.5–1.7 t resulted in a low number of post-hatch embryos. Two apparent peaks of0
survivors were observed for heat treatment at 1.0 and 1.4 t . The ploidy level of the0
larvae from the optimization experiments were verified by counting the nucleoli per celland chromosomes per cell.
Ž .Fig. 1. a–d Survival of heat shock treated African catfish embryos at different temperature from 39.5 to 418CŽ .measured at swimming stage .
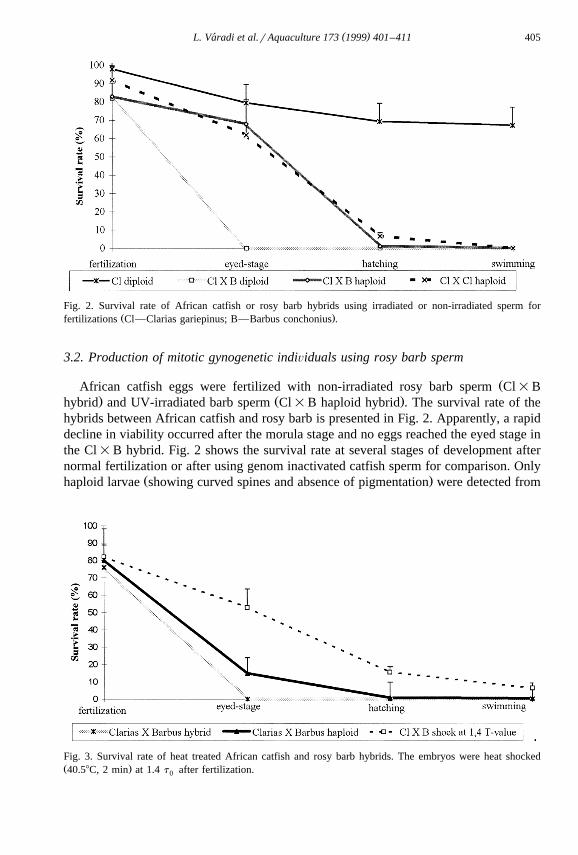
( )L. Varadi et al.rAquaculture 173 1999 401–411´ 405
Fig. 2. Survival rate of African catfish or rosy barb hybrids using irradiated or non-irradiated sperm forŽ .fertilizations Cl—Clarias gariepinus; B—Barbus conchonius .
3.2. Production of mitotic gynogenetic indiÕiduals using rosy barb sperm
ŽAfrican catfish eggs were fertilized with non-irradiated rosy barb sperm Cl=B. Ž .hybrid and UV-irradiated barb sperm Cl=B haploid hybrid . The survival rate of the
hybrids between African catfish and rosy barb is presented in Fig. 2. Apparently, a rapiddecline in viability occurred after the morula stage and no eggs reached the eyed stage inthe Cl=B hybrid. Fig. 2 shows the survival rate at several stages of development afternormal fertilization or after using genom inactivated catfish sperm for comparison. Only
Ž .haploid larvae showing curved spines and absence of pigmentation were detected from
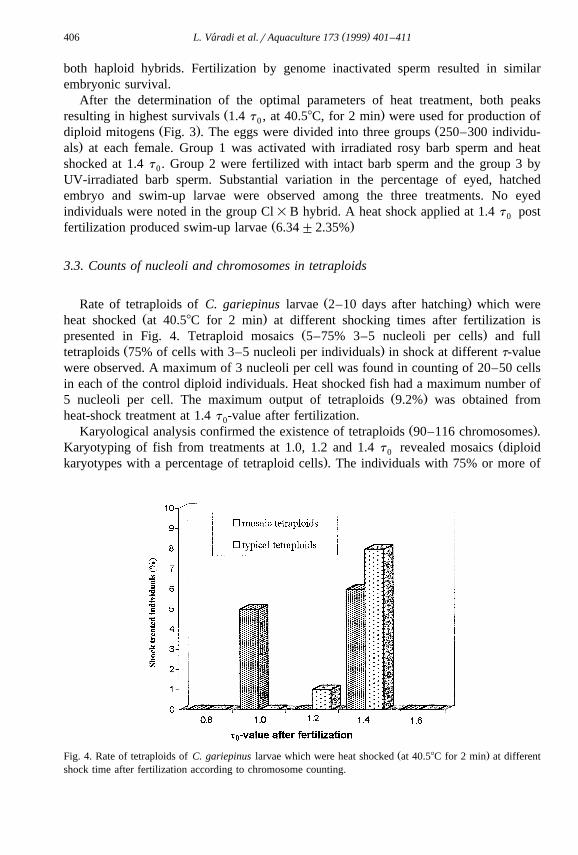
Fig. 3. Survival rate of heat treated African catfish and rosy barb hybrids. The embryos were heat shockedŽ .40.58C, 2 min at 1.4 t after fertilization.0
( )L. Varadi et al.rAquaculture 173 1999 401–411´406
both haploid hybrids. Fertilization by genome inactivated sperm resulted in similarembryonic survival.
After the determination of the optimal parameters of heat treatment, both peaksŽ .resulting in highest survivals 1.4 t , at 40.58C, for 2 min were used for production of0
Ž . Ždiploid mitogens Fig. 3 . The eggs were divided into three groups 250–300 individu-.als at each female. Group 1 was activated with irradiated rosy barb sperm and heat
shocked at 1.4 t . Group 2 were fertilized with intact barb sperm and the group 3 by0
UV-irradiated barb sperm. Substantial variation in the percentage of eyed, hatchedembryo and swim-up larvae were observed among the three treatments. No eyedindividuals were noted in the group Cl=B hybrid. A heat shock applied at 1.4 t post0
Ž .fertilization produced swim-up larvae 6.34"2.35%
3.3. Counts of nucleoli and chromosomes in tetraploids
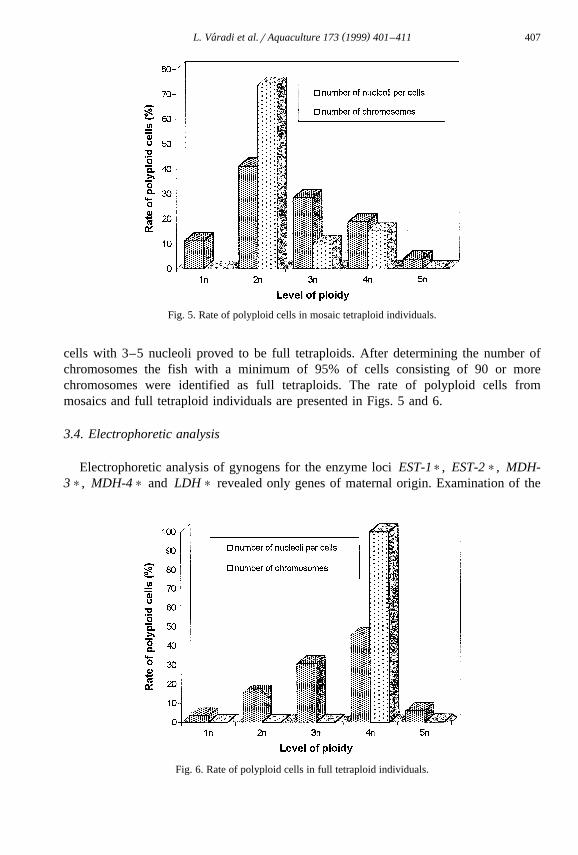
Ž .Rate of tetraploids of C. gariepinus larvae 2–10 days after hatching which wereŽ .heat shocked at 40.58C for 2 min at different shocking times after fertilization is
Ž .presented in Fig. 4. Tetraploid mosaics 5–75% 3–5 nucleoli per cells and fullŽ .tetraploids 75% of cells with 3–5 nucleoli per individuals in shock at different t-value
were observed. A maximum of 3 nucleoli per cell was found in counting of 20–50 cellsin each of the control diploid individuals. Heat shocked fish had a maximum number of
Ž .5 nucleoli per cell. The maximum output of tetraploids 9.2% was obtained fromheat-shock treatment at 1.4 t -value after fertilization.0
Ž .Karyological analysis confirmed the existence of tetraploids 90–116 chromosomes .ŽKaryotyping of fish from treatments at 1.0, 1.2 and 1.4 t revealed mosaics diploid0
.karyotypes with a percentage of tetraploid cells . The individuals with 75% or more of
Ž .Fig. 4. Rate of tetraploids of C. gariepinus larvae which were heat shocked at 40.58C for 2 min at differentshock time after fertilization according to chromosome counting.
( )L. Varadi et al.rAquaculture 173 1999 401–411´ 407
Fig. 5. Rate of polyploid cells in mosaic tetraploid individuals.
cells with 3–5 nucleoli proved to be full tetraploids. After determining the number ofchromosomes the fish with a minimum of 95% of cells consisting of 90 or morechromosomes were identified as full tetraploids. The rate of polyploid cells frommosaics and full tetraploid individuals are presented in Figs. 5 and 6.
3.4. Electrophoretic analysis
Electrophoretic analysis of gynogens for the enzyme loci EST-1), EST-2), MDH-3), MDH-4) and LDH) revealed only genes of maternal origin. Examination of the
Fig. 6. Rate of polyploid cells in full tetraploid individuals.

( )L. Varadi et al.rAquaculture 173 1999 401–411´408
five loci resulted in no traces of paternally inherited barb genes. Analysis of heat treatedoffspring for EST-1) and EST-2) revealed the expression of homozygous genotypesonly.
4. Discussion
In our experiments the effect of heat shock on embryo survival and ploidy level wasmonitored in gynogenetic offspring and polyploids. A dimensionless unit of embryologi-
Ž . Žcal age t -value was used to express the time of heat shock application Cherfas et al.,0.1993 . The heat shock was initiated within the limits of 0.6–1.8 t after fertilization.0
Using the t -value we could eliminate the maternal effects and any differences in the0
temperature of the incubation water. The heat shock temperature was 39 to 428C, andlasted for to 2 to 3 min, in different experiments. Embryo survival is characterized byalternating ‘sensitive’ and ‘non-sensitive’ periods with the lowest survival when theinitiation of heat shock corresponds to 1.1–1.3 and 1.5–1.7 t . These two peaks were0
Ž .also observed in carp Nagy et al., 1984; Cherfas et al., 1993 .After determination of ‘sensitive’ and ‘non-sensitive’ periods a temperature of 40.58C
applied for 2 min was subsequently used. A heat shock temperature at 41.58C or moreinduced mortality in early developing stages. The highest frequency of both gynogensand tetraploids was observed when the heat shock was initiated at 1.4 t . The consensus0
is that the sublethal doses are the most effective for producing genome manipulatedŽ . Žembryos Chourrout, 1987 . According to other publications Streisinger et al., 1981;
.Gomelsky and Rekubratsky, 1990; Cherfas et al., 1993 this time has proved to beŽ .approximately 60% of the first mitotic division. May et al. 1988 found the time most
effective for genome duplication at 50% of first mitotic division in fresh water fish.When African catfish sperm was used for fertilization 9.2% tetraploids were pro-
duced. The maximum numbers of nucleoli can be used to differentiate diploid individu-als in yolk sac fry where chromosome counting is difficult to do. The distribution of
Ž .nucleoli per cell in heat-treated tetraploid larvae demonstrated a large variability.These individuals could consist of polyploid embryos including mainly tetraploids,triploids and mosaics. The larvae with 5 to 75% of 3 to 5 nucleoli per cells perindividuals were identified as mosaics, although individuals with 75% or more of cellswith 3 to 5 nucleoli proved to be full tetraploids after determining the number of the
Ž .chromosomes. Cherfas et al. 1993 observed that about 80% of cells in heat-treatedŽ .embryos had 3 or 4 nucleoli per nucleus with a mean of 3.2–3.3. May et al. 1988
found maximum numbers of 6 nucleoli in pressure-blocked first mitotic rainbow trouts.Ž .Disney and Wright 1987 noted a maximum of six nucleoli per cell as well as
intraindividual variation in number in three pre-yearling brook trout females. Moreaccurate ploidy determinations and chromosome counts should be done on embryos
Ž .prior to the eyed stage Chourrout and Quillet, 1982 , or by flow cytometry after the fryŽ .have started to grow Thorgaard et al., 1982 .
Ž .Karyological analysis was performed on C. gariepinus by Ozouf-Costaz et al. 1990 .They revealed an identical diploid karyotype, 2ns56 with a pair of heteromorphic sex
Ž .chromosomes ZW and a nucleolus organizer location in a single pair of chromosomes.

( )L. Varadi et al.rAquaculture 173 1999 401–411´ 409
Ž .This is confirmed by our results in a highly inbred broodstock with approximately 43%Ž .3 of 7 fish matured females and 57% males among gynogenetic progeny. Theseindividuals obtained by heat shock of first mitotic cleavage of catfish eggs wereactivated by UV irradiated rosy barb sperm. However various authors e.g., GalbuseraŽ . Ž .1997 have strong evidence that the male carries the heterogametic sex XXrXY .Karyological analysis of both normal progenies and mitotic gynogens presented 56"7.8
Ž .chromosomes. Heat treated eggs from a normal fertilization exhibited diploid 56 ,Ž . Ž . Žtriploid 84–90 , and tetraploid 90–116 karyotypes. Typical tetraploids 95% of cells
.consisted of 90 or more chromosomes were observed only from heat shock at 1.4t -value post-fertilization. The cells containing 4 nucleoli was at a maximum of 45% in0
Ž .these fish. Shock at earlier stages 1.0–1.2 t -value resulted in mosaic individuals only.0Ž .May et al. 1988 observed an apparent additional earlier optimum at 3 h post-fertiliza-
tion which was 5 h before the halfway point to the first cell division. Similar tests onŽ . Ž .rainbow trout by Parson and Thorgaard 1985 and Chourrout 1984 were begun only 1
Žand 2 h before the half cycle optimum. The most probable explanation May et al.,.1988 is that DNA duplication has not begun at 3 h post-fertilization and that a shock at
this time leads to haploid individuals. Once DNA duplication begins, any attempt toblock the first mitotic division may result in individuals with unbalanced numbers ofeach chromosome or incompletely duplicated chromosomes in the complementŽ .aneuploid .
Eggs of African catfish activated by rosy barb sperm yielded progenies whichsuccumbed before hatching at the eyed-stage. Although the fertilization rate was high,
Ž .more than 80% Fig. 2 of the genome from rosy barb sperm associated to the genomeof African catfish eggs induced early and total mortality because of their geneticincompatibility.
On the other hand, fertilization with genetic inactivated African catfish or rosy barbsperm produced typical haploids showing the haploid syndrome. Approximately 10% ofthese embryos were hatched but none of them could swim or feed. Diploid gynogenesis
Ž .was obtained 6.34"2.35% survivors of treated eggs at feeding stage when eggs werefertilized with genetically inactivated rosy barb sperm and heat shock was initiated at 1.4t . Both protein gel electrophoresis at five loci and chromosome counts confirm diploid0
homozygosity. None of these fish showed any supernumery fragments of chromosomesor the presence of enzymes typical for rosy barb. The low survival rate may be due tothe deleterious effects of the heat shock itself, slight interanimal variations in the
Žsensitivity, and were due to the unmasking of recessive lethal genes Horvath and´.Orban, 1995 . The fish surviving in our experiments should not carry any such recessive´
lethals.Gynogenesis and androgenesis can be useful for generating inbred strains of fish
because the gynogenetic or androgenetic offspring of homozygous diploid individual areŽ .isogenic Streisinger et al., 1981; Parson and Thorgaard, 1985 . Androgenesis could
have significant advantages in fish species in which males are capable of maturingŽ .earlier than females e.g., salmonids because the time required for producing the inbred
Ž . Ž .strain can be reduced Parson and Thorgaard, 1985 . Ihssen 1976 argued that theoptimal breeding strategy in fish would be to maximize heterozygosity to achieveheterosis or hybrid vigour. The most effective means of maximizing heterozygosity is to

( )L. Varadi et al.rAquaculture 173 1999 401–411´410
cross divergent inbred lines. Alternative approaches such as gynogenesis and androgene-sis may provide more effective mechanism for the development of inbred lines.
Acknowledgements
ŽWe express our thanks to N.B. Cherfas and B. Gomelsky Fish and Aquaculture.Research Station, Dor, Israel for their invaluable advice and suggestions. I wish to
thank F. Muller for the kind help in improving the English of this manuscript and to¨Makadine Farkas Anasztazia for technical assistance.´ ´ ´
References
Bidwell, C.A., Chrisman, C.L., Libey, G.S., 1985. Polyploidy induced by heat shock in channel catfish.Aquaculture 51, 25–32.
Cherfas, N.B., Hulata, G., Kozinsky, O., 1993. Induced diploid gynogenesis and polyploidy in ornamentalŽ .koi carp, Cyprinus carpio L.: 2. Timing of heat shock during the first cleavage. Aquaculture 111,281–290.
Chourrout, D., 1982. Gynogenesis caused by ultraviolet irradiation of salmonid sperm. J. Exp. Zool. 223,175–181.
Chourrout, D., 1984. Pressure-induced retention of second polar body and suppression of first cleavage inrainbow trout: production of all-triploids, all-tetraploids and heterozygous and homozygous diploidgynogenetics. Aquaculture 36, 111–126.
Ž .Chourrout, D., 1987. Genetic manipulation in fish. Review of methods. In: Thiews, K. Ed. , SelectionHybridization and Genetic Engineering in Aquaculture II. Elsevier, pp. 112–122.
Chourrout, D., Quillet, E., 1982. Induced gynogenesis in the rainbow trout: sex and survival of progeniesproduction of all-triploid population. Theor. Appl. Genet. 63, 201–205.
Disney, J.E., Wright, J.E. Jr., 1987. Cytogenetic analyses of a SalÕelinus hybrid reveal evolutionaryrelationship between parental species. Cytogenet. Cell Genet. 45, 196–205.
Ž .Flajshans, M., Linhardt, O., Kvasnicka, P., 1993. Genetic studies of tench Tinca tinca L. : induced triploidyand tetraploidy and first performance data. Aquaculture 113, 301–312.
Galbusera, P., 1997. The genetic variability of wild and inbred populations of African catfish Clariasgariepinus. PhD thesis, Katholieke Universiteit Leuven, Leuven, Belgium.
Gomelsky, B.I., Rekubratsky, A.V., 1990. On the possibility of spontaneous andro- and gynogenesis in fishes.Ž .Voprosy Ikhtiologii 30 4 , 698–700.
Henken, A.M., Brunink, A.M., Richter, C.J.J., 1987. Differences in growth rate and feed utilization betweenŽ .diploid and triploid African catfish, Clarias gariepinus Burchell, 1822 . Aquaculture 67, 1–14.
Horvath, L., Orban, L., 1995. Genome and gene manipulation on carp. Aquaculture 129, 157–181.´ ´Ihssen, P., 1976. Selective breeding and hybridization in fisheries management. J. Fish. Res. Board Can. 33,
316–321.Komen, J., Bongers, A.B.J., Richter, C.J.J., van Miuswinkel, W.B., Huisman, E.A., 1991. Gynogenesis in
Ž .common carp Cyprinus carpio L. : II. The production of homozygous gynogenetic clones and F1 hybrids.Aquaculture 92, 127–142.
May, B., Henley, K.J., Krueger, C.C., Gloss, S.P., 1988. Androgenesis as a mechanism for chromosome setŽ .manipulation in brook trout SalÕelinus fontinalis . Aquaculture 75, 57–70.
Marian, T., Krasznai, Z., Bakos, J., 1984. Results of investigations of transferrin polymorphism in Hungarian´ ´carp populations. Halaszat 77, 112–113, in Hungarian.´
Morizot, D.C., Schmidt, M.E., 1990. Starch gel electrophoresis and histochemical visualization of proteins. In:Ž .Whitmore, D.H. Ed. , Electrophoretic and Isoelectric Focusing Techniques in Fisheries Management.
CRC, Boston, pp. 23–81.

( )L. Varadi et al.rAquaculture 173 1999 401–411´ 411
Nagy, A., Csanyi, V., 1982. Changes in genetic parameters in successive gynogenetic generations and some´calculations for carp gynogenesis. Theor. Appl. Genet. 63, 105–110.
Ž .Nagy, A., Rajki, K., Horvath, L., Csanyi, V., 1984. Investigation on carp Cyprinus carpio gynogenesis. J.Fish. Biol. 13, 215–224.
Na-nakorn, U., Rangsin, W., Wishasunkul, S., 1993. Suitable conditions for induction of gynogenesis in thecatfish, Clarias macrocephalus, using sperm of Pangasius sutchi. Aquaculture 118, 53–62.
ŽNaruse, K., Ijiri, K., Shima, A., Egami, N., 1985. The production of cloned fish in the medaka Oryzias.latipes . J. Exp. Zool. 236, 335–341.
Parson, J.E., Thorgaard, G.H., 1985. Production of androgenetic rainbow trout. J. Hered. 76, 177–181.Onozato, H., 1984. Diploidization of gynogenetically activated salmonid eggs using hydrostatic pressure.
Aquaculture 43, 91–97.Ozouf-Costaz, C., Teugels, G.G., Legendre, M., 1990. Karyological analysis of three strains of the African
Ž .catfish Clarias gariepinus Clariidae used in aquaculture. Aquaculture 87, 271–277.Shaklee, J.B., Allendorf, F.W., Morizt, K.C., Whitt, G.S., 1990. Gene nomenclature for protein coding loci in
fish. Trans. Am. Fish. Soc. 119, 2–15.Streisinger, G., Walker, C., Dower, N., Krauber, D., Singer, F., 1981. Production of clones of homozygous
diploid zebra fish. Nature 291, 293–296.Taniguchi, N., Hatanaka, H., Seki, S., 1990. Genetic variation in quantitative characters of meiotic and mitotic
gynogenetic diploid ayu, Plecoglossus altiÕelis. Aquaculture 85, 223–233.Thorgaard, G.H., Jazwin, M.E., Stier, M.W., 1981. Polyploidy induced by heat shock in rainbow trout. Trans.
Am. Fish. Soc. 110, 546–550.Thorgaard, G.H., Rabinovitch, P.S., Shen, M.W., Gall, G.A.E., Propp, J., Utter, F.M., 1982. Triploid rainbow
trout identified by flow cytometry. Aquaculture 29, 305–309.Volckaert, F.A.M., Galbusera, P., Heremans, K., Ollevier, F., 1993. Pressure-induction of diploid gynogenesis
Ž .in the eggs of African catfish Clarias gariepinus . In: Proc. 3rd Int. Meeting on High Pressure Biology.Univ. of Rochester Press, USA, pp. 73–81.
Volckaert, F.A.M., Galbusera, P., Hellemans, B., Van den Haute, C., Vanstaen, D., Ollevier, F., 1994.Ž .Gynogenesis in the African catfish Clarias gariepinus : I. Induction of meiogynogenesis with thermal and
pressure shocks. Aquaculture 128, 221–233.Volckaert, F.A.M., Van den Haute, C., Galbusera, P., Ollevier, F., 1997. Gynogenesis in the African catfish
Ž .Clarias gariepinus Burchell, 1822 . Optimizing the induction of polarbody gynogenesis with combinedpressure and temperature shocks. Aquaculture Research 28, 329–334.
Wolters, W.R., Libey, G.S., Chrisman, C.L., 1981. Induction of triploidy in channel catfish. Trans. Am. Fish.Soc. 110, 310–312.
Yamazaki et al., 1981.



















