
Expression of matrix metalloproteinases during murine chorioallantoic placenta maturation

Eur. J. Immunol. 1989. 19: 2341-2348 Effect of 5-azacytidine and IFN-y on murine placental cells 2341
Irene Athanassakis-Vassiliadiso, Dimitris ThanosO and Joseph Papamatheakiso+
Institute of Molecular Biology and Biotechnology, Foundation for Research and Technologyo and Department of Biology, University of Crete+, Heraklion
Induction of class I1 major histocompatibility complex antigens in murine placenta by 5-azacytidine and interferon-y involves different cell populations*
Class I1 MHC antigen expression is required for recognition of an alloantigen and generation of immune response. In rodents as well as in humans primary trophoblasts do not express class I1 MHC antigens. In this study we focused our interest on the mechanism(s) of class 11 antigen suppression on murine tropho- blasts. First, we examined the possibility of gene inactivation by methylation and second the possibility of lymphokine regulation of the class I1 genes. The first possibility was tested by treatment of placental cells with 5-azacytidine (5-AzaC), a cytidine analog which upon incorporation into the DNA inhibits further methylation, thus leading to gene activation. In order to test the second possibility we treated placental cells with interferon-y (IFN-y) or interleukin 4 (IL4) which are known to induce class I1 antigen expression in many systems. We showed that treatment with 5-AzaC or IFN-y but not IL4 significantly increased class I1 expression on cytokeratin-positive and vimentin-negative adherent placental cells. Following placental cell fractionation we distinguished three cell subsets with different responsiveness to 5-AzaC and IFN-y. The first, characterized as placental macrophages, were induced to express class I1 MHC antigens only after IFN-y treatment. The other two subsets, characterized as trophoblasts, were isolated from the labyrinthine- and spongio-trophoblast layer of the placenta and showed class I1 inducibility to 5-AzaC and IFN-y, respec- tively. The results show that depending on the anatomical localization of trophoblasts within the placenta, various regulatory elements control gene expression, so that the placental barrier provides fetal protection at different levels.
1 Introduction
During the last few years, it has become clearer that the success of pregnancy depends on various cellular and humoral interactions taking place at the maternal-fetal interface [ 1,2]. Indeed, many experimental data support the immunotrophic hypothesis, according to which mater- nal cells at the proximity of the fetoplacental unit are stimulated by fetal alloantigens to secrete growth factors which, in turn, increase trophoblast growth improving fetal survival [3].The most important family of fetal alloantigens is represented by the class I MHC antigens expressed on the cell surface of spongiotrophoblasts [4, 51.
The development of an immunological response in most, if not all, cases requires the presence of class I1 MHC antigens. However, in the case of maternal-fetal interac- tion, class I1 MHC antigens are absent from both spongio- trophoblast and labyrinthine trophoblast cell surface [5-71. There is no evidence in the literature for the importance of this lack of class I1 antigen expression from the placenta on fetal survival. However, it has been reported that during
[I 78071
* This work was supported by grants from the Hellenic Secretariat General for Research and Technology.
Correspondence: Joseph Papamatheakis, Foundation for Research and Technology, Institute of Molecular Biology and Biotechnology, PO. Box 1527, GR-71110 Heraklion, Crete, Greece
cardiac and pancreatic allograft rejection, an increase of class I1 antigen expression in the grafted tissue is observed [8, 91. Therefore, in the case of fetal allograft, lack of class I1 antigen expression could maintain an unresponsive maternal state vis-u-vis inflammatory reaction (ADCC, opsonization, etc.).
In the present study we focused our interest on the mechanism(s) of maintenance of the class I1 negative state of cytokeratin-positive, vimentin-negative trophoblast in vitro.We tested the possibilities of class I1 gene inactivation by methylation or regulatory suppression. Methylation of the cytosine residues has been known to be one of the mechanisms that a cell may utilize to shut off gene expression, including the class I1 genes [lo, 111. This process of gene inactivation is transferred to daughter cells during DNA replication, where 5-methylcytosine residues become de novo methylated. Well-studied examples of this mechanism of gene inactivation are the globin genes, the metallothionein gene, the X chromosome and others [12-141. In all these cases, successful gene activation occurs after treatment with 5-azacytidine (5-AzaC). This pyrimi- dine is a cytidine analog with a nitrogen instead of carbon residue at position 5 which, once incorporated in the DNA instead of cytosine, inhibits further methylation [15-171.
The second possibility is positive regulation by lympho- kines known to affect class I1 gene expression. Many class 11-negative monocytic cell populations have been shown to be inducible for class I1 expression after IFN-y treatment [18]. In addition, some cell populations of the B
0 VCH Verlagsgesellschaft mbH, D-6940 Weinheim, 1989 oO14-2980/89/12 12-2341 $02.50/0

2342 I. Athanassakis-Vassiliadis, D. Thanos and J. Papamatheakis Eur. J. Immunol. 1989. 19: 2341-2348
cell lineage can be stimulated to express class I1 MHC antigens after IL4 treatment [19]. Regulatory elements at the 5' end of various class I1 genes have been found to be responsible for induction by [20]. However, such elements have not yet been described for IL4 action.
In this work, we studied the effect of either 5-AzaC, IFN-y or IL4 on placental cells in vitro. We showed that 5-AzaC and EN-y, but not IL4, can induce class I1 expression on placental cells cultured in the presence of CSF-1, that exhibit Mac-1 and cytokeratin-positive and vimentin- negative surface phenotypes. Following placental cell frac- tionation we isolated three cell populations; the first, showing monocytic characteristics, could be induced to express class I1 antigens only after IFN-y treatment. The second, characterized as spongiotrophoblast-derived tro- phoblasts, were inducible by IFN-y to express class I1 antigens. Finally, the third population, which we identified as labyrinthine trophoblast-derived trophoblasts, was stim- ulated only by 5-AzaC to express class I1 MHC antigens. This complex regulation of class I1 antigen expression on the different layers of the placenta indicates that the lack of these antigens from the materno-fetal interface should be of great importance for fetal survival.
2 Materials and methods
2.1 Animals
BALB/cJ (H-2Kd) and C3H/HeJ (H-2Kk) were maintained at the Laboratory Animal Breeding Unit at the University of Crete (Greece). Each BALB/cJ or C3H/HeJ female was checked for oestrus, caged overnight with BALB/cJ or C3H/HeJ male and examined for the presence of vaginal plug the following morning.The day on which the plug was observed was considered to be day zero of pregnancy.
2.2 Antibodies
The mouse mAb to cytokeratins (PKK1, Labsystems, Helsinki, Finland), which are members of a multigene family present only on epithelial cells, was used at the dilution of 1/10. The mouse IgG mAb to vimentin (PK-V, Labsystems), which is the main subunit protein of interme- diated filaments in fibroblastoid cells,was used at a dilution of 1/10. The anti-Mac-1 mAb, which binds to the C3bi complement receptor (CR3) on granulocytes and M@ [21], was used as concentrated culture SN conditioned by the MU70 cell line (AmericanType Culture Collection, ATCC, Rockville, MD) at a dilution of 1/100. The mAb directed against IAD, IAK, KK MHC antigens (IgGZ,, Becton Dickinson, Mechelen, Belgium) were used at a dilution of 11100. The anti-H-2KD mAb was used as concentrated culture SN conditioned by the SF1-1.1.1 hybridoma (ATCC) at a dilution of 1/50. Mouse IgG antibody from normal serum was used as negative control at the concen- tration of 0.5 mg/ml (Sigma, St. Louis, MO).
2.3 Placental cell cultures
Placentae were obtained from females on day 12-15 of pregnancy. After removing the maternally derived decidual
layer [22], the tissue was cut into small pieces and single-cell suspensions were prepared by passing through a syringe with an 18G needle into Hanks' solution (Gibco, Grand Island, NY). The cells were washed three times and cultured in RPMI 1640 (Gibco) supplemented with 10% FCS (Seralab, Sussex, GB) at 1 x loh cells/ml in 100-mm plates (Falcon, Becton Dickinson, Oxnard, CA) or 87-well charnber-slides (Lab-Tec, Miles Laboratories, Naperville, IL) with 1000 U/ml CSF-1 (stage I [23]). On the second day of culture, 2 or 5 PM 5-AzaC (Sigma), 100 U/ml rIFN-y (Holland Biotechnology, Leiden, The Netherlands) or 80 U/ml rIL4 (Genzyme, Boston, MA) were added to the cells. On the third and fifth day of culture, another stimulation with 1000 U/ml CSF-1 was given to the cells, since the addition of growth factor within 48-h intervals has been described as the best for growing fetal-adherent phagocytic placental cells [24] which in the majority are positive for cytokeratin, Mac-1, F4180 antigens and nega- tive for vimentin antigen expression [25]. On the seventh day of culture the adherent cells were either lifted off the 100-mm plates using a cell lifter (Costar, Cambridge, MA) or, in the case of chamber-slides, fixed in ice-cold methanol for 20 min and processed for marker expression analysis by immunofluorescence. In some experiments, IFN-y or IL 4 were added to cultures 48 h before harvesting, a period shown to suffice for rnRNA and surface antigen induc- tion.
2.4 Placental cell fractionation
The spongiotrophoblast (outer placental zone) was sepa- rated from the labyrinthine trophoblast (red central zone) using fine iris scissors. Cell suspensions of labyrinthine trophoblast, spongiotrophoblast or whole placentae from syngeneically or allogeneically mated females were isolated on day 12-15 of pregnancy and fractionated on a Percoll gradient. Isotonic Percoll solution (Pharmacia, Dorval, PR) was mixed with isotonic PBS to give final dilution of 209'0, 40%, 60% and 80% (e = 1.02-1.04-1.06-1.08 kgll). The gradient was prepared by layering 2 ml of each dilution in a plastic centrifuge tube.Two milliliters of cell suspension containing a total of 5 x 107-7 x lo7 cells was layered at the top of the gradient. After centrifugation at 2000 rpm for 30min at 4"C, the cells at the gradient interfaces were collected in Hanks' solution, washed three times and cultured as previously described at the concentration of 0.5 X lo6 to 1 x lo6 cells/ml in 100-mm plates. After treatment with 5-AzaC or 1FN-y the adherent cells of the 1.02-1.04 kg/l Percoll-derived interface (APC40) and 1.04-1.06 kg/l Percoll-derived interface (APC60) were processed for antigen expression analysis by immunofluo- rescence.
2.5 Indirect immunofluorescence
The adherent placental cells were washed three times in PBS (Gibco) supplemented with 1% normal goat serum (NGS), in 96-well plates (Linbro, Flow Labs, McLean,VA) at 5 x lo6 cells/ml, and incubated at 4°C for 30 min. The cells were washed once with PBS supplemented with 0.2% BSA (Sigma) and 0.01% sodium azide (Sigma; PBS- BSA-Azide) followed by the addition of 0.1 ml of test antibody, diluted as described above, and incubated for
Eur. J. Immunol. 198Y. 19: 2341-2348 Effect of 5-azacytidine and IFN-y on murine placental cells 2343
45 rnin at 4"C.The cells were then washed three times with PBS-BSA-Azide, and FITC-conjugated goat anti-mouse IgG antibody (Tag0 Inc., Burlingame, CA) was added for 45 rnin at 4"C, followed by extensive washing. The cells were then fixed with 25% glycerol, mounted on slides and fluorescence was evaluated visually using a Zeiss (Ober- kochen, FRG) fluorescence microscope. Cells with weak or no staining of all were scored as negative. Positive cells were considered those showing bright to very bright staining.
In another set of experiments the cells were first sorted, using a magnetic isolation system, and then tested for marker analysis by immunofluorescence. Thus, placental cells were treated with 5-AzaC or IFN-y as described earlier and the adherent population was first incubated for 30 min at 4°C with 1% NGS in 35-mm plates (Falcon) and then reacted with anti-IAD andlor anti-IAK mAb for 45 rnin at 4°C. After washing the cells three times with PBA- BSA-Azide, 10 pl of goat anti-mouse IgG-coated magnetic beads (Dynabeads, Dynal AS, Oslo, Norway) was added in each 35-mm plate and agitated for 15 min at 4 "C. The cell suspension was then transferred to 5-ml tubes and cell separation was achieved using a magnetic particle concen- trator (MPC1, Dynal) as described by Gaudernack et al. [26]. Briefly, the test tube containing the cells and the Dynabeads was placed in the MPCl and left for 1-2 min. During that time the cells rosetted with immunomagnetic beads were attracted to the wall of the tube. The SN containing the negative cells was pipetted out of the tube, and the remaining cells were redissolved in fresh medium. The procedure was repeated four times and then the fractions of Dynabead-rosetted or negative cells were extensively washed and processed for immunofluorescence staining.
2.6 Northern blot analysis
Adherent placental cells treated with 5-AzaC or I F N y as mentioned before were collected and washed. Total RNA extraction, agarose/formaldehyde gels and transfer to Genescreen membranes were performed as described by Maniatis et al. [27]. A nick-translated 3.5-kb Sal I fragment of the Ea gene was used for hybridizations done according to Church and Guilbert [28].
3 Results
3.1 Induction of MHC class I1 antigen expression on placental cells
Immunohistochemical studies have shown that placenta is negative for class I1 MHC antigen expression during preg- nancy [5-7,291. Some Ia+ and HLA-DR+ cells in mice and humans, respectively, appear at late-stage placentae, but these are identified as fetal fibroblasts and MQ which are not in contact with the maternal circulation [29-311.
We have previously isolated a population of fetal adherent phagocytic placental cells [25] which are trophoblasts by criteria of cytokeratin positivity and vimentin negativity. These cells also share some characteristics with MQ since they express the Mac-1 and F4180 surface antigens and are stimulated with MQ-specific growth factors [24, 251. Immunofluorescence experiments showed that these tro- phoblast cells are negative for class I1 surface antigen expression ([25],Table 1). In an attempt to induce class I1 antigen expression in these cells we have utilized either 5-AzaC, IFN-y or IL4.
Table 1. 5-AzaC and IFN-y induce class TI MHC antigen expression on adherent placental cellsa)
(BALBlcl x BALB/cl) placentaeh) Percent of fluorescent cells Antibodies Control 5-AzaC 5-AzaC IFN-yd'
(2 UM) ( 5 UM)
- If0 Anti-KD 59+5 Anti-IAD 5 + 2 Anti-IAK 3 f 1 Anti-Mac-1 75 * 3
Anti-Vim 1 5 1 Ant i-Cyt 57 f 4
Mouse IgG I f 1
I f 1 68+ 7 51 f 12 5 f 1
45+ 8 61+ 2 3 f 1
0
(BALB/cl x C3H/HeJ)Fk (placentae" Control 5-AzaC
(2 (IM)
1 f 1 7 f 1 54f 2 4 2 f 2 58+ 5 7 0 f 6 9 f 3 8f 1 68f 7 7 1 f 2 72+ 5 5 8 5 1 2 f 1 3 + 1
0 2 f 1
Mouse IgG Anti-KD Anti-IAD Anti-KK Anti-IAK Anti-Mac-I Anti-Cyt Anti-Vim
If1 81 f 5 12+5 64f0 5 f l
83 f 2 86+1 1 4 + 1
9 f 5 73-+ 14 6 5 f 6 5 6 f 14 5 5 f 1 71 f 13 6Yf 8 1 3 f 5
5 + 1 6 f I 72 f 13 7 6 f 1 1 49 f 19 Y f 6 5 7 f 7 68210 54+ 14 1 1 + 3 7 6 5 6 5 5 f 9 68+ Y 68f 9 2 9 f 6 20* 12
a) Placental cells were stimulated with CSF-1 on days 0 , 3 , 5 and the adherent cells were tested for antigen expression on day 7 of culture.
b) Antigen expression was tested on methanol- fixed adherent placental cells from BALBlcJ pregnant by BALB/cJ males.
c) Antigen expression was tested on adherent placental cells in suspension from BALBlcJ females pregnant by C3H/HeJ males.
d) IFN-y was used at the concentration of 100 Ulml.
e) IL4 was used at the concentration of 80 Ulml.
2344 I. Athanassakis-Vassiliadis, D. Thanos and J. Papamatheakis Eur. J. Immunol. 1989. 19: 2341-2348
untreated control (Table 1). The same response was seen with either syngeneically (BALBkJ x BALB/cJ) or allo- geneically (BALBkJ x C3H/HeJ)FI-derived placental cells. In the first case an increase of IAD expression was seen and in the latter a simultaneous IAD and IAK increase of expression was observed. These results show that the cells responding to these agents are of fetal origin since the K haplotype is contributed only by the paternal strain.The increase of class I1 antigen expression is not due to Fc binding, since normal mouse Ig gives background levels even after 5-AzaC or IFN-y treatment. Both 5-AzaC and IFN-y treatments did not significantly affect and in some instances rather decreased the expression of the other markers tested in these experiments.Thus, most of the cells remained positive for Mac-1, cytokeratin and class I anti- gen expression, and negative for vimentin expression.
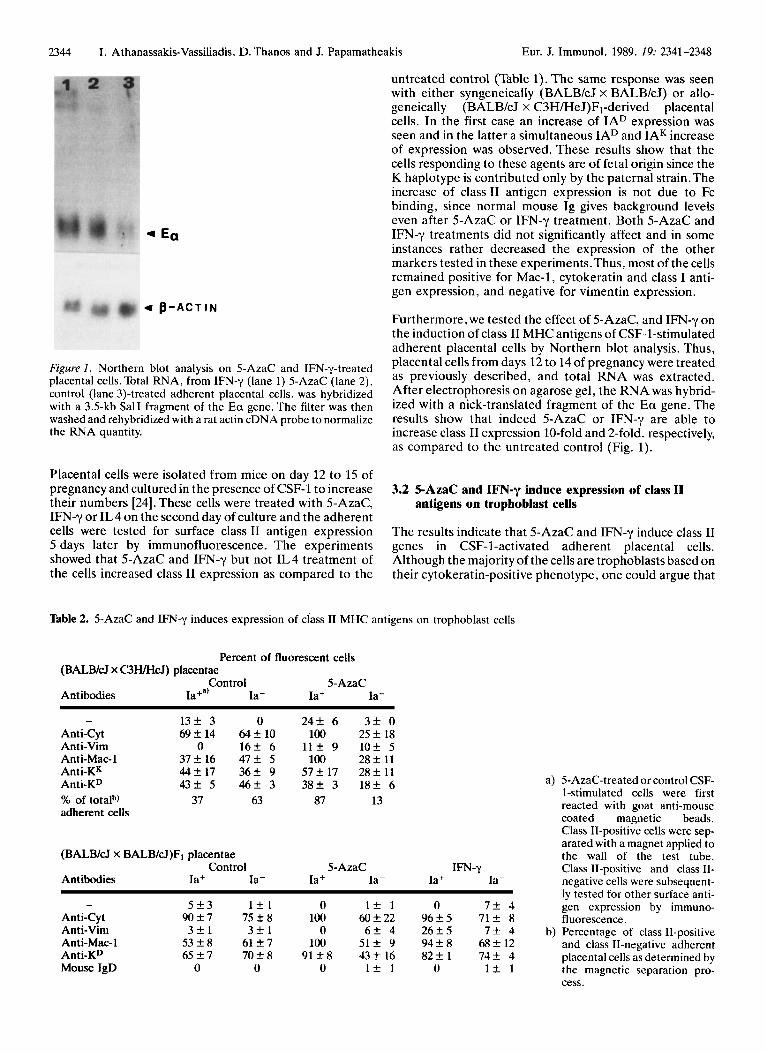
Furthermore, we tested the effect of 5-AzaC, and IFN-y on the induction of class I1 MHC antigens of CSF-1-stimulated adherent placental cells by Northern blot analysis. Thus, placental cells from days 12 to 14 of pregnancy were treated as previously described, and RNA was extracted. After On agarose gel, the RNA was hybrid- ized with a nick-translated fragment of the Ea gene. The results show that indeed 5-AzaC or IFN-y are able to increase class I1 expression 10-fold and 2-fold. respectively, as compared to the untreated control (Fig. 1).
I P - A C T I N
Figure I. Northern blot analysis on 5-AzaC and IFN-y-treated placental cells.Total RNA, from IFN-y (lane 1) 5-AzaC (lane 2), control (lane 3)-treated adherent placental cells, was hybridized with a 3.5-kb SalI fragment of the Ea gene. The filter was then washed and rehybridized with a rat actin cDNA probe to normalize the RNA quantity.
Placental cells were isolated from mice on day 12 to 15 of pregnancy and cultured in the presence of CSF-1 to increase their numbers [24]. These cells were treated with 5-AzaC, LFN-y or IL4 on the second day of culture and the adherent cells were tested for surface class I1 antigen expression The results indicate that 5-AzaC and IFN-y induce class I1 5 days later by immunofluorescence. The experiments genes in CSF-1-activated adherent placental cells. showed that 5-AzaC and IFNy but not IL4 treatment of Although the majority of the cells are trophoblasts based on the cells increased class I1 expression as compared to the their cytokeratin-positive phenotype, one could argue that
3.2 5-AzaC and IFN-y induce expression of class I1 antigens on trophoblast cells
Table 2. 5-AzaC and IFN-y induces expression of class I1 MHC antigens on trophoblast cells
Percent of fluorescent cells
Control 5-AzaC (BALB1c.I x C3WHeJ) placentae
Antibodies Ia- Ia + Ia-
- 13f 3 0 24+ 6 3+ 0 Anti-Cyt 6 9 f 1 4 64+10 100 25 + 18 Anti-Vim 0 16+ 6 I l k 9 lo+ 5 Ant i-Mac- 1 37+16 47+ 5 100 28 f 11 Anti-KK 4 4 f 1 7 3 6 + 9 5 7 f 1 7 28+11 Anti-KD 4 3 f 5 46f 3 38+ 3 18+ 6 % of totalb) 37 63 87 13 adherent cells
(BALB/cl x BALB/d)FI placentae
Antibodies Ia+ Ia- Ia+ Ia- Ia + Ia- Control 5-AzaC m - Y
- 5 + 3 1 + 1 0 1 + 1 0 7 + 4 Anti-Cyt 9 0 f 7 7 5 2 8 100 6 0 t 2 2 Y6+5 7 1 f 8 Anti-Vim 3 + 1 3 + 1 0 6 + 4 2 6 f 5 7 5 4 Anti-Mac- 1 5 3 + 8 6 1 2 7 100 51+ 9 Y4+8 688112 Anti-KD 6 5 + 7 70f8 9 1 + 8 4 3 t 1 6 8 2 + 1 7 4 t 4 Mouse IgD 0 0 0 1 + 1 0 1 + 1
a) 5-AzaC-treated or control CSF- 1-stimulated cells were first reacted with goat anti-mouse coated magnetic beads. Class 11-positive cells were sep- arated with a magnet applied to the wall of the test tube. Class 11-positive and class II- negative cells were subsequent- ly tested for other surface anti- gen expression by immuno- fluorescence.
b) Percentage of class 11-positive and class 11-negative adherent placental cells as determined by the magnetic separation pro- cess.
Eur. J. Immunol. 1989. 19: 2341-2348
the Ia+ cells are M@ which are known for their class I1 inducibility by IFN-y and growth in response to CSF-1 [18,32]. In order to investigate this possibility the class II- positive cells were first isolated using a magnetic immuno- bead selection method and then stained with different antibodies.
Effect of 5-azacytidine and IFN-y on murine placental cells 2345
Placental cells from syngeneically or allogeneically mated BALBk females were treated with 5-AzaC or IFN-y; the adherent population was first reacted with anti-IAD or anti- IAD and anti-IAK antibodies and then the class II- positive cells were selected using goat anti-mouse Ig-coated magnetic beads. The isolated class 11-positive and -negative cells were subsequently tested by immunofluorescence for the presence of trophoblast and M@ markers on their surface.
These experiments show that after 5-AzaC or IFN-y treatment the class 11-positive cells (87% of the total cells vs. 37% of the untreated cells) are 100% positive for cytokeratin and Mac-1 expression (Table 2). These results indicate that, indeed, the 5-AzaC and IFNy treatments induce class I1 expression on the cytokeratin-positive “tro- phoblasts”. Only a small fraction of the class 11-negative cell population after treatment stains positively for cytokeratin and Mac-1.
3.3 Effect of 5-AzaC and IFN-y on Percoll-fractionated placental cells
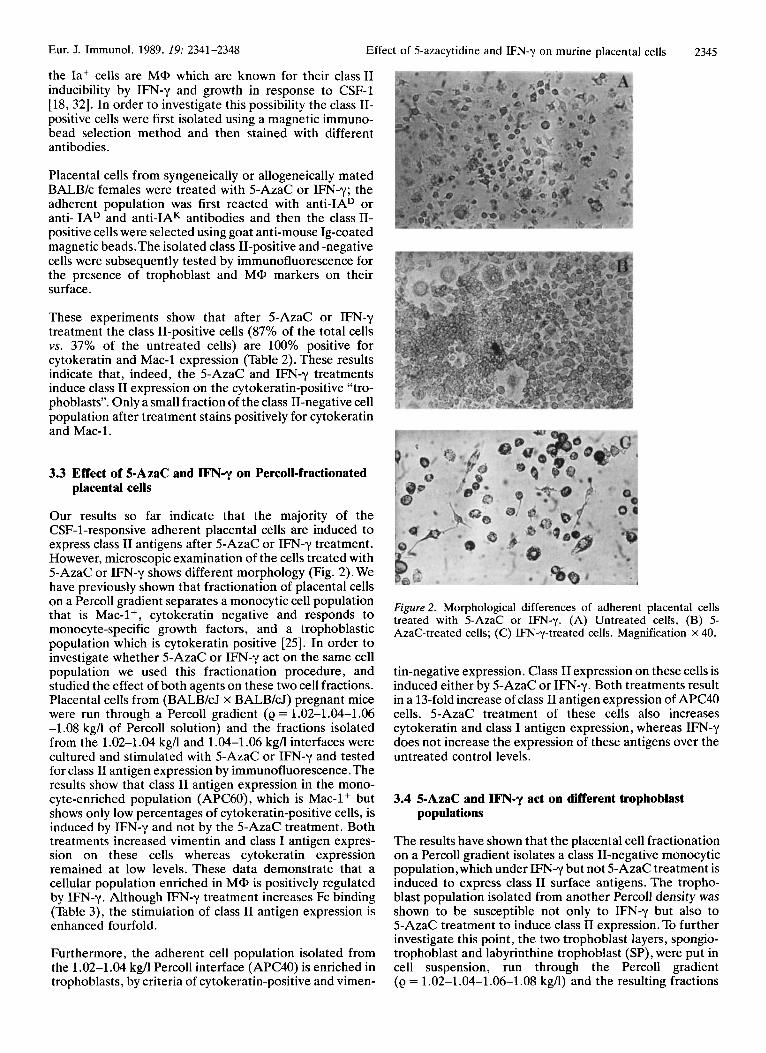
Our results so far indicate that the majority of the CSF-1-responsive adherent placental cells are induced to express class I1 antigens after 5-AzaC or IFNy treatment. However, microscopic examination of the cells treated with 5-AzaC or IFN-y shows different morphology (Fig. 2). We have previously shown that fractionation of placental cells on a Percoll gradient separates a monocytic cell population that is Mac-l+, cytokeratin negative and responds to monocyte-specific growth factors, and a trophoblastic population which is cytokeratin positive [25]. In order to investigate whether 5-AzaC or IFN-y act on the same cell population we used this fractionation procedure, and studied the effect of both agents on these two cell fractions. Placental cells from (BALB/cJ x BALBkJ) pregnant mice were run through a Percoll gradient (e = 1.02-1.04-1.06 -1.08 kg/l of Percoll solution) and the fractions isolated from the 1.02-1.04 kg/l and 1.04-1.06 kgA interfaces were cultured and stimulated with 5-AzaC or IFN-y and tested for class I1 antigen expression by immunofluorescence. The results show that class I1 antigen expression in the mono- cyte-enriched population (APC60), which is Mac-l+ but shows only low percentages of cytokeratin-positive cells, is induced by IFN-y and not by the 5-AzaC treatment. Both treatments increased vimentin and class I antigen expres- sion on these cells whereas cytokeratin expression remained at low levels. These data demonstrate that a cellular population enriched in MQ, is positively regulated by IFN-y. Although WN-y treatment increases Fc binding (Table 3), the stimulation of class I1 antigen expression is enhanced fourfold.
Furthermore, the adherent cell population isolated from the 1.02-1.04 kg/l Percoll interface (APC40) is enriched in trophoblasts, by criteria of cytokeratin-positive and vimen-
Figure 2. Morphological differences of adherent placental cells treated with 5-AzaC or IFN-y. (A) Untreated cells, (B) 5- AzaC-treated cells; (C) IFN-y-treated cells. Magnification X 40.
tin-negative expression. Class I1 expression on these cells is induced either by 5-AzaC or IFN-y. Both treatments result in a 13-fold increase of class I1 antigen expression of APC40 cells. 5-AzaC treatment of these cells also increases cytokeratin and class I antigen expression, whereas IFN-y does not increase the expression of these antigens over the untreated control levels.
3.4 5-AzaC and IFN-y act on different trophoblast populations
The results have shown that the placental cell fractionation on a Percoll gradient isolates a class 11-negative monocytic population,which under IFN-y but not 5-AzaC treatment is induced to express class I1 surface antigens. The tropho- blast population isolated from another Percoll density was shown to be susceptible not only to IFN-y but also to 5-AzaC treatment to induce class I1 expression. To further investigate this point, the two trophoblast layers, spongio- trophoblast and labyrinthine trophoblast (SP), were put in cell suspension, run through the Percoll gradient (e = 1.02-1.04-1.06-1.08 kgA) and the resulting fractions
2346
Table 3. Effect of 5-AzaC and IFN-y on Percoll fractionated placental cellsa)
I. Athanassakis-Vassiliadis, D. Thanos and J. Papamatheakis Eur. J. Immunol. 1989. 19: 2341-2348
APC40b) APCW) Control 5-AzaC IFN-y Control 5-AzaC IFN-y
- 5 f O Mac-1 71 f 7 Anti-Cyt 64f5 Anti-Vim 19 f 4 Anti-IAD 6 f 2 Anti-KD 6 5 f 5 Mouse Ig 0 Anti-IAK 9 f 3
0 82 f 9 97 * 3 10 f 3 73 f 7 82 ? 6
0 6 f 1
3 f 2 3 + 0 2 8 f 7 80213 8 2 f 4 7 1 f 4 6 3 f 8 2 4 f 1 4 1 f 3 1 3 f 5 13+ 4 74+24 8of 2 11+ 3 12+ 1 6 5 f 6 3 3 f 17 8 5 f 2
0 0 0 0 0 1 2 + 3
were treated with 5-AzaC o r IFN-y as previously described and tested for class I1 expression by immunofluores- cence .
The results show that the labyrinthine trophoblast-derived APC40 cells are induced to express class I1 MHC antigens only by 5-AzaC and not by IFN-y treatment (Table 4), whereas the spongiotrophoblast-derived APC40 cells can be stimulated only by IFN-y to express class I1 antigens (Table 4). Both APC40 populations (LAB nd SP derived) are positive for cytokeratin and negative for vimentin expression (Table 4). APC60 cells, which are positive for vimentin and negative for cytokeratin expression, are induced to express Ia antigens only by IFN-y regardless of their labyrinthine- o r spongiotrophoblast origin (Ta- ble 4).
4 Discussion
It is generally accepted that trophoblasts in the murine or human placenta d o not express class I1 MHC antigens [4-71. Some class 11-positive cells, that appear at late placentae in the subperipheral layer of the amnion and sparsely within the chorionic villous stroma, are probably fetal dendritic cells or M@ which are not exposed to the
31f 8 6 7 f 1 23+ 7 Y4+ 8 9 2 2 1 1
100 2 1 f 8 33 + 20
a) Placental cells were isolated from BALBkJ females pregnant by BALB/cJ males.
b) APC40: adherent placental cells isolated from the 1.02-1.04 kg/l Percoll interface, were stimulated with CSF-1, treated with 5-AzaC (2 p ~ ) or IFN-y (100 Ulml) and tested for antigen expression by immuno- fluorescence.
c) APC60: adherent placental cells isolated from the 1.04-1.06 kgll Percoll interface and treated as APC40.
maternal circulation [30-331. The class 11-negative state of trophoblasts may be necessary to avoid the generation of immune response towards the fetus, since an alloantigen must be recognized by immunocompetent cells in associa- tion with an l a molecule to generate such a response. In the present study we focused our interest on determining the mechanism(s) which induce the class I1 MHC antigen expression on trophoblasts. In this study, trophoblasts refer to fetal in origin adherent placental cells, which are negative for vimentin expression and positive for cytokeratin and Mac-1 antigen expression. Along this line, we have inves- tigated the action of three agents known to affect gene expression. The possibility of gene methylation was tested by treatment of placental cells with the inhibitor of DNA methylation, 5-AzaC [15]. Incorporation of 5-AzaC, which is a cytidine analog that can be integrated into DNA, leads to hypomethylation of D N A [34] as a result of the presence of nitrogen in place of carbon at the fifth position in the pyrimidine ring. In order to test lymphokine responsive- ness, placental cells were treated with IFN-y or IL4. Class I1 antigen expression can be included by IFN-y in APC, melanoma cells, some epithelial cells and other parenchymal cells [18]. Class I1 MHC antigen expression is inducible by I L 4 in resting B cells and pre-B cell lineages [19]. I n the present study we showed that the response of trophoblasts t o the 5-AzaC or I F N y treatment depends on
Table 4. Effect of 5-AzaC and IFN-y on Percoll-fractionated LAB trophoblast and Spa)
Treatment Anti- Anti-Vimentin Anti-IAD Anti-IAK N o antibody Cytokeratin
LAB@): Control 82 f 5 9 f 2 6 f l 7 + 1 6 f l y - F N 71 f 7 I1 f 1 6 + 0 l l f 0 1 5 f 4 5-AzaC 89f8 LO+ 1 7 6 f 6 7 5 f 9 4 + 0
SP4oh): Control 6 4 f 3 5 r t l 14+6 1 3 f 4 Y f 2 y-IFN 55f 1 15+4 6 0 + 2 6 0 f 2 8 f 4 5-AzaC 6 4 f 6 16+3 8 t l 1 8 + 5 4 f 0
LAB 6ob): Control 7 + 1 6 4 f 3 Y+1 5 + 0 1 0 f 1 y-IFN 1 1 f 1 92f8 6 3 f 3 7 1 f 3 0 5-AzaC 1 8 f 5 83 f 1 Y + O 7 f l 4 f 0
SPW): Control 6 f 2 62 f 8 7 + 2 6 + 2 3 f O a) The two trophoblast layers y-IFN 10 f 5 63 + 7 6 2 + 2 6 4 + 4 15 f 1 were isolated from allogeneic 5-AzaC 8 f 2 71 + 9 1 3 f 1 1 6 f 5 1 5 f 5 placentae (BALBkJ females
pregnant by C3H/HeJ males) on day 13 of pregnancy.
b) LAB 40: labyrinthine trophoblast-derived cells isolated from the 1.02-1.04 kg/l Percoll interface. LAB 60: labyrinthine trophoblast- derived cells isolated from the 1.04-1.06 kgll Percoll interface. SP40: spongiotrophoblast-derived cells isolated from the 1.02-1.04 kg/l Percoll interface. SP 60: spongiotrophoblast-derived cclls isolated form the 1.04-1.06 kg/l Percoll interface.

Eur. J. Immunol. 1989. 19: 2341-2348 Effect of 5-azacytidine and IFN-y on murine placental cells 2347
their tissue localization in the placenta.Thus, we show that spongiotrophoblast-derived trophoblasts are induced to express class I1 antigens after IFN-y treatment, whereas trophoblasts in the labyrinthine trophoblast zone express class I1 antigens in response to 5-AzaC. Furthermore, we show that adherent, Mac-1+, cytokeratin-negative CSF- 1-responsive placental MQ, express class I1 MHC antigens only after IFN-y treatment.
Placental cells from syngeneic or allogeneic pregnancies were stimulated with CSF-1 and treated with 5-AzaC, IFN-y or IL4. CSF-1 has been previously shown [24] to stimulate the growth of fetal adherent cells the majority of which are positive for cytokeratin, F4/80 and Mac-1 antigen expression, and negative for vimentin expression [25]. Treatment of the CSF-1-stimulated placental cells with 5-AzaC and IFN-y but not IL4 was able to increase approximately 10-fold class I1 antigen expression as tested by immunofluorescence (Table 1). Most of the cells that were induced to express Ia antigens were fetal in origin since they were able to bind anti-class I1 mAb of the paternal haplotype (Table 1). Further evidence for class I1 inducibility by 5-AzaC or IFN-y was obtained from Nor- thern blot analysis.
Although it is the first time to our knowledge that the effect of 5-AzaC on class I1 induction of placental cells has been studied, other investigators have reported that TFN-y is unable to induce class I1 MHC antigen expression on cultured placental trophoblast cells whereas it can increase class I MHC antigen expression in these cells [35]. How- ever, in those studies spleen cell culture SN conditioned by Con A SN were used as the source of IFN-y.Therefore, the failure of these experiments to show class I1 induction could be due to inhibition of the IFN-y effect by other factors present in the ConA SN. Furthermore, in our present study we could not detect any significant stimula- tion of the class I , cytokeratin or Mac-1 antigen expression after 5-AzaC or IFN-y treatment (Table 1).
Since placental cells were stimulated with CSF-1 to grow, one could argue that these are placental MQ, and not trophoblasts. Evidence that this is not the case was given from experiments where the class 11-positive cells were first sorted using a magnetic isolation technique and then reacted with different mAb. These experiments showed that all the 5-AzaC- or IFN-y-treated class 11-positive cells were also positive for cytokeratin and negative for vimentin expression (Table 2).
The results so far indicate that 5-AzaC or IFN-y are able to induce class I1 expression on adherent, cytokeratin-posi- tive,vimentin-negative placental cells. In addition, the cells induced with 5-AzaC are morphologically different from those induced with IFNy (Fig. 2 ) . Exposure of placental cells to these agents following fractionation has demon- strated clearly that alternate cell subsets respond to IFN-y and 5-AzaC. One subset consists of cells with the charac- teristics of trophoblasts previously described, and the second, which represents 5-10% of the total placental population, is composed of cells positive for Mac-1 and negative for cytokeratin antigens. The latter is considered to be of monocytic origin. These two fractions were treated as the unfractionated placental cells and tested for surface marker analysis by immunofluorescence. The results
showed that the monocytic subset (APC60) was induced to increase class I1 antigen expression only after IFN-y treat- ment (Table 3). This treatment considerably enhanced vimentin and class I MHC antigen expression on these cells (Table 3). In contrast to APC60 cells, the trophoblastic cell population (APC40) was induced to express Ia antigens with both 5-AzaC and IFN-y treatment (Table 3).
This effect may suggest a common regulatory pathway for 5-AzaC and IFN-y although no evidence known to us is available to support this view. Alternatively, the above effect may represent responses of different cells populating the placenta. Indeed, trophoblasts in the ectoplacental clone (7.5 days of pregnancy) are known to differentiate and give rise to two placental layers, the spongiotropho- blast and the labyrinthine trophoblast. The two placental zones were isolated with careful dissection of the mature placenta and tested for their ability to express class11 antigens after 5-AzaC or IF"-y treatment. Labyrinthine trophoblast and spongiotrophoblast zones were fraction- ated in a Percoll gradient and the derived cell populations were treated with different agents as previously described. The results showed that indeed regulation of class I1 MHC antigens in the two trophoblast populations is subject to different mechanisms. Spongiotrophoblast-derived tropho- blasts are induced to express Ia antigens only after IFN-y treatment, whereas in labyrinthine-derived trophoblasts class I1 antigens are only induced by 5-AzaC treatment (Table 4). These results and the fact that spongiotropho- blast represents only the 24% of the total placental population (data not shown) explain why 5-AzaC gives a 10-fold induction of class I1 MHC antigens, whereas IFN-y gives only 2-fold induction when tested by Northern blot analysis of unfractionated adherent placental cells. These findings agree with the pattern of class I distribution in the maternal placenta. Only the spongiotrophoblast zone is shown to express class I antigens, whereas the labyrinthine trophoblast zone is unable to bind antibodies directed against the paternal class I MHC antigens [5]. Concerning class I1 MHC antigen expression, this is forbidden in the labyrinthine trophoblast zone, since the results presented here show that the class I1 gene is only activated by 5-AzaC. On the other hand the spongiotrophoblast zone can be induced to express class I1 antigens by IFN-y treatment. Evidence in the laterature shows that IFN-y is produced at the proximity of the feto-placental unit in vivo [36]. Furthermore, human placenta has been shown to possess receptors to IFN-y [37]. However, other pregnancy pro- teins, like the a-fetoprotein, are known to inhibit class I1 induction by IFN-y [38], which provides an equilibrium as to maintaining the negative class I1 expression on spongio- trophoblast. Thus, the spongiotrophoblast layer seems to be the zone of important maternal-fetal interactions which is decisive for a successful pregnancy.
The induction of donor type class I1 antigens on grafted tissues in association to class I MHC antigens can cause rejection of the graft [8, 91. During pregnancy it is believed that class I expression on the placenta is necessary to stimulate maternal T cells for secretion of lymphokines which may improve placental and fetal growth [1]. The presence of at least two levels of regulatory restriction for class 11 expression in the placenta enables us to suggest that class I1 antigen production would render the tissue suscep- tible to maternal immune recognition which could be

2348 I. Athanassakis-Vassiliadis, D. Thanos and J. Papamatheakis Eur. J. Immunol. 1989. 29: 2341-2348
deleterious to fetal survival.The question of how important fetal survival is to the maintenance of the negative state of class I1 antigen expression on the placenta and its potential relationship with immunological abortion are currently under investigation in o u r laboratory. Preliminary experi- ments have shown that indeed treatment of pregnant females with 5-AzaC in vivo induces fetal abortion, which correlated with induction of class I1 antigen expression in the labyrinthine trophoblast zone.
We thank Catherine Liptrot for her comment.s, and Nectaria Kelaidi for typing the manuscript.
Received July 28. 1989; in revised form September 18. 1989
5 References
1 2
3
4
5
6
7
8 9
10 11 12
13 14
15
16
Wegmann,T. G., A m . J. Reprod. Immunol. 1987. 15: 67. Wegmann, T. G., Athanassakis, I.. Guilbert, L., Branch, D., Dy, M., Menu, E . andchaouat, G., Transplant. Proc. 1988.21: 566. Wegmann.T. G., Annu. Immunol. (Inst. Pasteur) 1984.1350: 309. Head, J. R., Drake, B. L. and Zuckermann, F. A., Am. J. Reprod. Immunol. 1987. 15: 12. Raghupathy, R., Singh, B., Leigh, J. B. and Wegmann,T. G.. J. Immunol. 1981. 127: 2074. Jenkinson, E. J. and Searle, R. F., J. Reprod. Immunol. 1979. I : 3. Chatterjee-Hasrouni, S. and Lala, I? K., J. Immunol. 1981.127: 2070. Milton, A. D. and Fabre, J. W., J. Exp. Med. 1985. 161: 98. Steiniger, B., Klempnauer, J. and Wonigeit, K., Transplantation 1985. 40: 234. Bird, A. A., Nature 1986. 321: 209. Cedar, H., Cell 1988. 53: 3. Busslinger, M., Hurst, J. and Flavell, R. A , , Cell 1983. 34: 197. Compkre, S. and Palmiter, R. D., Cell 1981. 23: 233. Toniolo, D., DUrso, M., Martini, G., Pcrsico, M., Tufano,V., Battistuzzi, G. and Luzzato, L., EMBO J. 1984. 3: 1987. Verolia, L., Gartler, S. M. ,Wasman, E. R. ,Yen, I?, Mohandas, T. and Shapiro, J., Proc. Natl. Acad. Sci. USA 1982. 79: 2352. Mahandas,T., Sparkes. R. S. and Shapiro, L. J., Science 1981. 211: 393.
17 McKeon, C., Ohkubo, H., Pastan, I. and De Crombrugghe, B., Cell 1982. 29: 203.
18 Skoskiewicz, M. J., Colvin, R. B., Schneeberger, E. E. and Russel, I? S., J. Exp. Med. 1985. 162: 1645.
19 Polla, B. S., Poljak, A., Ohara, J., Paul,W. E. and Glimcher, L. H., J. Immunol. 1986. 137: 3332.
20 Thanos, D., Mavrothalassitis, G. and Papamatheakis, J., Proc. Natl. Acad. Sci. USA 1988. 85: 3075.
21 Bcllcr, D. J . , Springer,T. A. and Schreiber, R. D., J. Exp. Med. 1982. 156: 1000.
22 Rossant, J. and Croy, A., J. Embryol. Exp. Morphol. 1985.86: 177.
23 Stanlcy, E. R. and Guilbert, L. J.. J. Immunol. Methods 1980. 42: 253.
24 Athanassakis, I., Bleackley, C. R., Paetkau,V., Guilbert, L., Barr, F! J. and Wegmann,T. G., J. Immunol. 1987. 148: 37.
25 Guilbert, L. J., Athanassakis, I.. Branch, D. R. , Christopher- son. R., Crainie, M. , Garcia Lloret, M., Mogil, R., Ransoon- dar, M. J. and Wegmann,T. G., in Wegman,T. G. and Gill,T. J., 111 (Eds.), The molecular and cellular immunobiology of the maternal fetal interface, Oxford University Press, New York 1989. in press.
26 Gaudernack, G., Leivestad,T., Ugelstad, J. and Thorsby, E. , J. Imrnunol. Methods 1986. 90: 179.
27 Maniatis, T., Fritsch, E. F. and Sambrook, J., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Labora- tory, Cold Spring Harbor 1982, p. 202.
28 Church, G. M. and Guilbert, W., Proc. Natl. Acad. Sci. USA 1984. 81: 1991.
29 Bulmer, J. N. and Johnson, I? M., Placenta 1985. 6: 127. 30 Bulmer, J. N. and Sunderland, C. A.. Immunology 1984. 52:
349. 31 Bulmer, J. N. and Johnson, I? M., Clin. Exp. Irnmunol. 1984.
57: 393. 32 Guilbert, L. J., in Van Furth, R. (Ed.), Mononuclear Phago-
cytes and Inflammation, Martinus Nijhoff Publishers, Dor- drecht 1985, p. 233.
33 Sutton, L., Mason, D.Y. and Redman, G. W. G., Immunology 1983. 49: 103.
34 Jones. I? A. and Taylor, S. M., Cell 1980. 20: 85. 35 Zuckermann, F. A. and Head, J. R., J. Immunol. 1986. 137:
846. 36 Chin, T. W.. Ank, B. J., Strom, S. R. and Stiehm, R., Cell.
Immunol. 1982. 113: 1. 37 Galderon, J., Sheehan, K. C. F., Chance, C.,Thomas, M. L. and
Schreiber, R. D., Proc. Natl. Acad. Sci. USA 1988. 85: 4837. 38 Lu, C.Y., Changelian, I? S. and Unanue, E. R., J. Immunol.
1984. 132: 1722.
Copyright © 2022 FDOKUMEN




















