
Induction and Spatial Organization of Polyamine Biosynthesis During Nodule Development in Lotus...
7
Genes coding for a putative cell-wall invertase and two putative monosaccharide/H + transporters are expressed in roots of etiolated Glycine max seedlings § Maria Dimou a , Emmanouil Flemetakis a , Costas Delis a , Georgios Aivalakis a , Karoline G. Spyropoulos b , Panagiotis Katinakis a, * a Department of Agricultural Biotechnology, Agricultural University of Athens, Iera Odos 75, 11855 Botanikos, Athens, Greece b Department of Botany, Faculty of Biology, School of Sciences, National and Kapodistrian University of Athens, Panepistimiopolis, Zografou, 15771 Athens, Greece Received 8 February 2005; received in revised form 26 May 2005 Available online 22 June 2005 Abstract Sink tissues depend on the supply of sugars produced by source tissues. Cell-wall invertases (EC 3.2.1.26) are considered to have a pivotal role in supplying sink tissues with carbohydrates via an apoplasmic pathway, while associated monosaccharide/H + transporters take up the produced hexoses. In this study, we characterized genes coding for a putative cell-wall invertase (GmCWINV1) and two putative monosaccharide/H + transporters (GmMST1 and GmMST2) of Glycine max. Semi-quantitative RT-PCR analysis revealed that GmMST1 and GmMST2 are expressed in various plant organs. In situ hybridization revealed that they are expressed in different root tissues. These results propose that different monosaccharide/H + transporters may play different roles in source and sink organs. In addition, the temporal and spatial expression of GmCWINV1, as was determined by semi-quantitative RT-PCR and in situ hybridization analyses, was detected in tissues where GmMST1 and GmMST2 were also expressed, indicating that, at least part of the sucrose unloaded from soybean phloem may be hydrolyzed into hexoses before being transported from the apoplasmic space into the respective sink cells. # 2005 Elsevier Ireland Ltd. All rights reserved. Keywords: Cell-wall invertase; Glycine max; In situ hybridization; Monosaccharide/H + transporter; Root; Phloem unloading 1. Introduction The assimilated carbon is exported from the source tissues and transported to the non-photosynthetic sink tissues via phloem, mainly in the form of sucrose. Phloem unloading to the sink tissues may take place either symplasmically via plasmodesmata or apoplasmically across the plasma membrane [1]. In the latter case, sucrose may be taken up by sink cells either intact via sink specific sucrose transporters or after extracellular hydrolysis by cell- wall invertases (EC 3.2.1.26), to glucose and fructose, via monosaccharide/H + transporters, referred as sugar transport proteins (STP) [1,2]. Genes and cDNA clones encoding cell-wall invertases have been identified and characterized from a variety of plants, and are often found as multigene families that show sink tissue-enhanced expression pattern [3,4]. It has been suggested that they may play a role in photoassimilate partitioning and source/sink regulation [5,6]. However, the importance of cell-wall invertases came from the analysis of the miniature1 mutant of maize, which due to the abolishment of an endosperm specific cell-wall invertase shows a small seed phenotype [7]. Furthermore, antisense inhibition of cell-wall invertase in carrot resulted in the www.elsevier.com/locate/plantsci Plant Science 169 (2005) 798–804 Abbreviations: BCIP, 5-bromo-4-chloro-3-indolyl-phosphate; BSA, bovine serum albumin; EDTA, ethylenediaminetetraacetic acid; EST, expressed sequence tag; NBT, nitro blue tetrazolium Nucleotide sequence data are available in the EMBL database under accession numbers AJ563366 (GmMST1), AJ563365 (GmMST2), AJ563368 (GmCWINV1). * Corresponding author. Tel.: +30 210 529 4342; fax: +30 210 529 4314. E-mail address: [email protected] (P. Katinakis). 0168-9452/$ – see front matter # 2005 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.plantsci.2005.05.037
Transcript of Induction and Spatial Organization of Polyamine Biosynthesis During Nodule Development in Lotus...

www.elsevier.com/locate/plantsci
Plant Science 169 (2005) 798–804
Genes coding for a putative cell-wall invertase and two putative
monosaccharide/H+ transporters are expressed in roots
of etiolated Glycine max seedlings§
Maria Dimou a, Emmanouil Flemetakis a, Costas Delis a, Georgios Aivalakis a,Karoline G. Spyropoulos b, Panagiotis Katinakis a,*
a Department of Agricultural Biotechnology, Agricultural University of Athens, Iera Odos 75, 11855 Botanikos, Athens, Greeceb Department of Botany, Faculty of Biology, School of Sciences, National and Kapodistrian University of Athens,
Panepistimiopolis, Zografou, 15771 Athens, Greece
Received 8 February 2005; received in revised form 26 May 2005
Available online 22 June 2005
Abstract
Sink tissues depend on the supply of sugars produced by source tissues. Cell-wall invertases (EC 3.2.1.26) are considered to have a pivotal
role in supplying sink tissues with carbohydrates via an apoplasmic pathway, while associated monosaccharide/H+ transporters take up the
produced hexoses. In this study, we characterized genes coding for a putative cell-wall invertase (GmCWINV1) and two putative
monosaccharide/H+ transporters (GmMST1 and GmMST2) of Glycine max. Semi-quantitative RT-PCR analysis revealed that GmMST1
and GmMST2 are expressed in various plant organs. In situ hybridization revealed that they are expressed in different root tissues. These
results propose that different monosaccharide/H+ transporters may play different roles in source and sink organs. In addition, the temporal and
spatial expression of GmCWINV1, as was determined by semi-quantitative RT-PCR and in situ hybridization analyses, was detected in tissues
where GmMST1 and GmMST2 were also expressed, indicating that, at least part of the sucrose unloaded from soybean phloem may be
hydrolyzed into hexoses before being transported from the apoplasmic space into the respective sink cells.
# 2005 Elsevier Ireland Ltd. All rights reserved.
Keywords: Cell-wall invertase; Glycine max; In situ hybridization; Monosaccharide/H+ transporter; Root; Phloem unloading
1. Introduction
The assimilated carbon is exported from the source
tissues and transported to the non-photosynthetic sink
tissues via phloem, mainly in the form of sucrose. Phloem
unloading to the sink tissues may take place either
symplasmically via plasmodesmata or apoplasmically
across the plasma membrane [1]. In the latter case, sucrose
Abbreviations: BCIP, 5-bromo-4-chloro-3-indolyl-phosphate; BSA,
bovine serum albumin; EDTA, ethylenediaminetetraacetic acid; EST,
expressed sequence tag; NBT, nitro blue tetrazolium
Nucleotide sequence data are available in the EMBL database under
accession numbers AJ563366 (GmMST1), AJ563365 (GmMST2),
AJ563368 (GmCWINV1).
* Corresponding author. Tel.: +30 210 529 4342; fax: +30 210 529 4314.
E-mail address: [email protected] (P. Katinakis).
0168-9452/$ – see front matter # 2005 Elsevier Ireland Ltd. All rights reserved
doi:10.1016/j.plantsci.2005.05.037
may be taken up by sink cells either intact via sink specific
sucrose transporters or after extracellular hydrolysis by cell-
wall invertases (EC 3.2.1.26), to glucose and fructose, via
monosaccharide/H+ transporters, referred as sugar transport
proteins (STP) [1,2].
Genes and cDNA clones encoding cell-wall invertases
have been identified and characterized from a variety of
plants, and are often found as multigene families that show
sink tissue-enhanced expression pattern [3,4]. It has been
suggested that they may play a role in photoassimilate
partitioning and source/sink regulation [5,6]. However, the
importance of cell-wall invertases came from the analysis of
the miniature1 mutant of maize, which due to the
abolishment of an endosperm specific cell-wall invertase
shows a small seed phenotype [7]. Furthermore, antisense
inhibition of cell-wall invertase in carrot resulted in the
.

M. Dimou et al. / Plant Science 169 (2005) 798–804 799
arrest of taproot formation [8]. In both cases, provision of
apoplasmic hexoses is apparently required for normal
development.
Hexoses liberated by cell-wall invertases are taken up by
associated monosaccharide/H+ transporters and a functional
coupling is supported by a temporal and spatial association
of gene expression [2,9–12]. Genes encoding plant
monosaccharide/H+ transporters are grouped in multigene
families with different members being up-regulated in
various sinks [1,13,14]. The observations that different
Arabidopsis thaliana sugar transport proteins (AtSTPs) have
been shown to catalyze the uptake of exogenous sugars in
heterologous systems [15], and that a knock-out mutation in
the AtSTP1 gene resulted in a decrease in uptake of
exogenous monosaccharides by A. thaliana seedlings [16],
are consistent with AtSTPs functioning in the plasma
membrane to import monosaccharides from the apoplasmic
space. It remains to be determined whether all 14 putative
AtSTPs catalyze monosccaharide transport, and what
physiological functions these and other related proteins
have [15,16].
In the present work, we report the expression of a putative
cell-wall invertase and two putative monosaccharide/H+
transporter genes (designated as GmCWINV1, GmMST1 and
GmMST2, respectively) during primary and lateral root
development of etiolated soybean seedlings. Based on the
temporal and spatial accumulation of transcripts, possible
functions of putative cell-wall invertase and monosacchar-
ide/H+ transporter isoforms are discussed.
2. Materials and methods
2.1. Plant material and growth conditions
Soybean (Glycine max cv. Williams) seeds were pre-
germinated on two sheets of moist paper, in Petri dishes, in
darkness, at 26 8C for 2 days. After germination, the
seedlings were transferred in a modified 0.5 � Hoagland
solution, under a 16-h light/8-h dark cycle, at 26 8C. Young
leaves (YL), mature leaves (ML) and stems (STEMS) of 30
days old plants were used to isolate total RNA. In addition,
some seedlings were left in Petri dishes, in darkness, at
26 8C for 6 more days, and total RNA was isolated from
hypocotyls (HYP), cotyledons (COT) and different parts of
roots, representing the region of cell division (RT), the root
region of elongation (MRI) and the region of laterals
emergence (MRII). For in situ hybridization, the roots from
the 8 days old seedlings were used.
2.2. Characterization of cDNA clones coding for
GmMST1, GmMST2 and GmCWINV1
By performing BLAST searches [17] among the
databases of the American Public Soybean Expressed
Sequence Tag Project, two cDNA clones coding for putative
monosaccharide/H+ transporters, and one cDNA clone
coding for a putative cell-wall invertase were identified
and obtained from ResGen, Invitrogen Corp. Their
nucleotide sequences were determined and as parts of their
50 ends were missing, total RNAwas extracted from the roots
of 8 days old seedlings [18] and a 50 RACE-PCR was
performed with the SMART 50 RACE kit from Clontech
(Westburg, NL). The amplified fragments were cloned into
the pBlueScriptKS+ plasmid vector (STRATAGENE) and
fully sequenced. The full-length cDNA clones were
designated as GmMST1, GmMST2 and GmCWINV1. The
clustal procedure was used for both analysis of the sequences
data and determination of the relatedness of the deduced
polypeptides to related sequences from other species.
2.3. RT-PCR analysis of GmMST1, GmMST2 and
GmCWINV1
Semi-quantitative RT-PCR analysis was performed using
total RNA extracted from the tissues of interest as described
above and quantified by spectrophotometry and agarose gel
electrophoresis. All RNA samples were treated with DNase I
(PROMEGA) according to the manufacturer’s instructions to
eliminate DNA contamination. In each case, the RT-PCR
reactions were performed, according to the manufacturer’s
directions (QIAGEN) using gene-specific oligonucleotides
as primers (GmCWINV1F: 50-CTGCTTGGCTAGGCAA-
AG-30, GmCWINV1R: 50-ACCATTCACCGACGTGTC-30,GmMST1F: 50-CTGAGACACCAAACAGCC-30, GmMS-
T1R: 50-CACAGCCAAGACTGCACC-30, GmMST2F: 50-AGGCTGTAGTTGCAGCTG-30 and GmMST2R: 50-TAAT-
CATCATGCTCCACG-30). RNase inhibitor was obtained
from PROMEGA. For normalization of the different RNA
preparations, a 260 bp fragment of GmUBQ was amplified,
using two gene-specific oligonucleotides as primers
(GmUBQF: 50-GGGTTTTAAGCTCGTTGT-30 and GmU-
BQR: 50-GGACACATTGAGTTCAAC-30). The reverse
transcriptase reactions were performed in a thermal cycler
at 50 8C for 30 min, followed by PCR amplification of 25–30
cycles at 94 8C for 1 min, 54 8C for 1 min and 72 8C for 1 min.
A complete final extension for the PCR products was
performed at 72 8C for 10 min. Amplified products were
separated on 1.5% agarose gels, blotted on to nylon
membranes and hybridized with the respective digoxi-
genin-11-rUTP labeled cDNA probes.
2.4. In situ mRNA hybridization
Root segments were fixed in 4% (w/v) paraformaldehyde
supplemented with 0.25% (v/v) glutaraldehyde in 10 mM
sodium phosphate buffer (pH 7.4), for 1 h, in a vacuum
aspirator, dehydrated with ethanol, and then exchanged with
xylene before embedding in paraffin. Sections (8 mm) were
mounted on poly-L-lysine slides, digested with proteinase K
for 30 min at 37 8C, treated with acetic anhydride, dried in
ethanol, and then hybridized with appropriate gene specific
M. Dimou et al. / Plant Science 169 (2005) 798–804800
digoxigenin-labeled probes overnight, at 42 8C. After
washing with 4 � SSC containing 5 mM dithiothreitol,
the slides were treated with RNase A for 30 min, at 37 8C,
washed again at 37 8C with RNase A buffer (500 mM NaCl,
1 mM EDTA, 10 mM Tris–HCl, pH 7.5) containing 5 mM
dithiothreitol, and then processed for revealing the
digoxigenin antigen. This involved blocking with blocking
reagent and BSA, followed by incubation with an anti-
digoxigenin antibody conjugated to alkaline phosphatase,
and washing with blocking reagent. The color was revealed
by incubation in NBT and BCIP and the reactions were
stopped with water, slides dehydrated, air-dried, and then
mounted in DPX before viewing. Antisense and sense
probes labeled with digoxigenin-11-rUTP (ROCHE) were
used in parallel hybridizations. The sections were examined
using a ZEISS Axiolab (Carl Zeiss, Jena, Germany)
microscope and pictures were taken with SONY DSC-
S75 38P/45 (SONY Corporation, Japan) digital camera
system.
3. Results
3.1. Characterization of G. max cDNA clones coding
for putative monosaccharide/H+ transporters and a
putative cell-wall invertase
Systematic BLAST searches among the public databases
revealed the presence of a number of soybean ESTs derived
from root cDNA libraries showing high homology to
monosaccharide/H+ transporter genes from different plant
species. Several of these cDNA clones were obtained, and
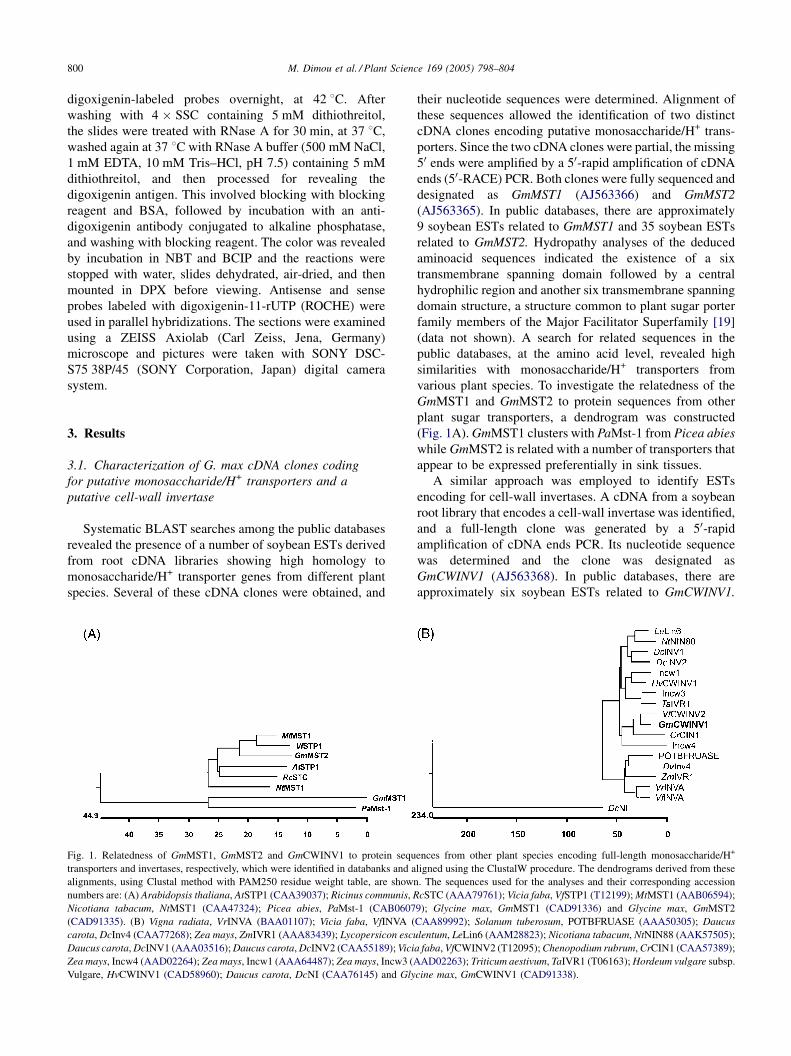
Fig. 1. Relatedness of GmMST1, GmMST2 and GmCWINV1 to protein sequ
transporters and invertases, respectively, which were identified in databanks and a
alignments, using Clustal method with PAM250 residue weight table, are shown
numbers are: (A) Arabidopsis thaliana, AtSTP1 (CAA39037); Ricinus communis, R
Nicotiana tabacum, NtMST1 (CAA47324); Picea abies, PaMst-1 (CAB0607
(CAD91335). (B) Vigna radiata, VrINVA (BAA01107); Vicia faba, VfINVA (
carota, DcInv4 (CAA77268); Zea mays, ZmIVR1 (AAA83439); Lycopersicon escu
Daucus carota, DcINV1 (AAA03516); Daucus carota, DcINV2 (CAA55189); Vici
Zea mays, Incw4 (AAD02264); Zea mays, Incw1 (AAA64487); Zea mays, Incw3 (A
Vulgare, HvCWINV1 (CAD58960); Daucus carota, DcNI (CAA76145) and Gly
their nucleotide sequences were determined. Alignment of
these sequences allowed the identification of two distinct
cDNA clones encoding putative monosaccharide/H+ trans-
porters. Since the two cDNA clones were partial, the missing
50 ends were amplified by a 50-rapid amplification of cDNA
ends (50-RACE) PCR. Both clones were fully sequenced and
designated as GmMST1 (AJ563366) and GmMST2
(AJ563365). In public databases, there are approximately
9 soybean ESTs related to GmMST1 and 35 soybean ESTs
related to GmMST2. Hydropathy analyses of the deduced
aminoacid sequences indicated the existence of a six
transmembrane spanning domain followed by a central
hydrophilic region and another six transmembrane spanning
domain structure, a structure common to plant sugar porter
family members of the Major Facilitator Superfamily [19]
(data not shown). A search for related sequences in the
public databases, at the amino acid level, revealed high
similarities with monosaccharide/H+ transporters from
various plant species. To investigate the relatedness of the
GmMST1 and GmMST2 to protein sequences from other
plant sugar transporters, a dendrogram was constructed
(Fig. 1A). GmMST1 clusters with PaMst-1 from Picea abies
while GmMST2 is related with a number of transporters that
appear to be expressed preferentially in sink tissues.
A similar approach was employed to identify ESTs
encoding for cell-wall invertases. A cDNA from a soybean
root library that encodes a cell-wall invertase was identified,
and a full-length clone was generated by a 50-rapid
amplification of cDNA ends PCR. Its nucleotide sequence
was determined and the clone was designated as
GmCWINV1 (AJ563368). In public databases, there are
approximately six soybean ESTs related to GmCWINV1.
ences from other plant species encoding full-length monosaccharide/H+
ligned using the ClustalW procedure. The dendrograms derived from these
. The sequences used for the analyses and their corresponding accession
cSTC (AAA79761); Vicia faba, VfSTP1 (T12199); MtMST1 (AAB06594);
9); Glycine max, GmMST1 (CAD91336) and Glycine max, GmMST2
CAA89992); Solanum tuberosum, POTBFRUASE (AAA50305); Daucus
lentum, LeLin6 (AAM28823); Nicotiana tabacum, NtNIN88 (AAK57505);
a faba, VfCWINV2 (T12095); Chenopodium rubrum, CrCIN1 (CAA57389);
AD02263); Triticum aestivum, TaIVR1 (T06163); Hordeum vulgare subsp.
cine max, GmCWINV1 (CAD91338).

M. Dimou et al. / Plant Science 169 (2005) 798–804 801
A search for related sequences in the public databases
revealed strong similarities with cell-wall invertases. An
alignment of the deduced GmCWINV1 cell-wall invertases
from other plant species revealed a conserved b-fructosidase
(Asn54, Asp55, Pro56, Asn57, Gly58) and a catalytic cysteine
motif (Met233, Trp234, Glu235, Cys236, Pro237). The fifth
residue in the cysteine motif is a proline that is specific to
cell-wall invertases [4]. A basic pI of 8.84 was calculated
using the ExPasy sequence analysis tool, suggesting that in a
cell-wall environment, where the pH is acidic, the
GmCWINV1 will be ionically bound with cell-wall compo-
nents. A dendrogram was constructed in order to investigate
the relatedness of GmCWINV1 to protein sequences from
other plant invertases (Fig. 1B). The sequences were divided
into three different classes representing neutral, vacuolar and
cell-wall invertases. GmCWINV1 clusters with the group of
other cell-wall invertases.
3.2. Accumulation of GmMST1, GmMST2 and
GmCWINV1 transcripts in various G. max organs
The accumulation of GmMST1, GmMST2 and
GmCWINV1 gene transcripts in various organs of 30 days
old soybean plants (young leaves, mature leaves and stems)
and 8 days old etiolated soybean seedlings (hypocotyls,
cotyledons, the root region of cell division, elongation and
laterals emergence) was investigated using a semi-quanti-
tative reverse transcription-PCR approach. A housekeeping
gene coding for ubiquitin was used as an internal control. As
shown in Fig. 2, expression of GmCWINV1 gene was
observed in all organs examined. The lower levels of
expression were found in mature leaves of 30 days old plants
and in cotyledons of 8 days old etiolated seedlings. The
highest levels of GmMST1 transcript accumulation were
observed in the root tip region (RT) and in hypocotyls of
Fig. 2. Semi-quantitative reverse transcription-polymerase chain reaction
analysis of the expression profiles of GmMST1, GmMST2 and GmCWINV1
genes. Total RNA from young leaves (YL), mature leaves (ML) and stems
(STEMS) of 30 days old plants, and hypocotyls (HYP), cotyledons (COT),
and different parts of roots, representing the region of cell division (RT), the
root region of elongation (MRI) and the region of laterals emergence (MRII)
of 8 days old etiolated seedlings was subjected to reverse transcription-
polymerase chain reaction analysis. The RT-PCR product of the GmUBQ
was used as an internal control. Amplified products were separated on 1.5%
agarose gels, blotted on nylon membranes and hybridized with the respec-
tive digoxigenin-11-rUTP labeled cDNA probes.
8 days old etiolated seedlings and in stems and young leaves
of 30 days old plants. Relatively lower levels were found in
cotyledons of 8 days old etiolated seedlings and in the root
region of elongation (MRI) and the region of laterals
emergence (MRII) of 8 days old etiolated seedlings. On the
contrary, the GmMST2 transcripts were detected in all
organs examined and particularly in the root tip region of
8 days old etiolated seedlings.
3.3. Spatial expression of GmMST1, GmMST2 and
GmCWINV1 transcripts in primary and lateral roots of
etiolated G. max seedlings
Roots of 8 days old etiolated seedlings are depleted of
their own reserves, as has been judged from starch, lipid and
protein staining of root sections (data not shown), and
therefore are typical sink tissues. So we further studied the
expression of GmMST1, GmMST2 and GmCWINV1 during
root development, using the in situ RNA–RNA hybridization
method. Longitudinal and transverse sections of primary and
lateral roots of 8 days old etiolated soybean seedlings were
hybridized with 11-digoxigenin-rUTP-labeled RNA probes
transcribed from GmMST1, GmMST2 and GmCWINV1
cDNA clones (Fig. 3). In longitudinal sections, GmCWINV1
and GmMST2 expression was detected in the root tip region
and especially in the cortex parenchyma of root tip and in the
central root cap region (Fig. 3N and I). Levels of both
transcripts, in the phloem region and in a few cell layers
neighbouring the central tissue, gradually increased from the
region of cell division to the region of elongation and then
decreased in the lateral root emergence region. As the cells
differentiated, GmCWINV1 and GmMST2 expression was
confined in the epidermis, some underlying cell layers, and
the vascular cylinder (Fig. 3N and I).
In order to identify cell types in which GmCWINV1 and
GmMST2 are expressed, we carried out in situ hybridizations
in cross-sections. On the basis of the anatomical features of
root regions we chose sections representing the zone of cell
division, elongation zone as well as the region of lateral root
emergence. As it can be observed in Fig. 3K and F, the cells
in the root cap as well as the cells in other layers in the region
of cell division were strongly stained. In the region of
elongation, the transcripts were strongly detected in cells of
the epidermis and vascular cylinder (Fig. 3L and G).
Hybridization signal was detectable in the epidermis of the
region of maturation and the cortex parenchyma (Fig. 3M
and H).
When similar sections from the same root tissues were
hybridized using an antisense probe of GmMST1, hybridiza-
tion signal was detectable in all different cell types (Fig. 3A–
D). However, root cap enlarging cells as well as enlarging
cortical parenchyma cells give higher signal.
Cross-sections of the primary root containing the lateral
root primordia also revealed that all genes were mainly
expressed in the apical region of the emerging lateral root
and in the phloem of the main root (Fig. 3E, J and O). In
M. Dimou et al. / Plant Science 169 (2005) 798–804802
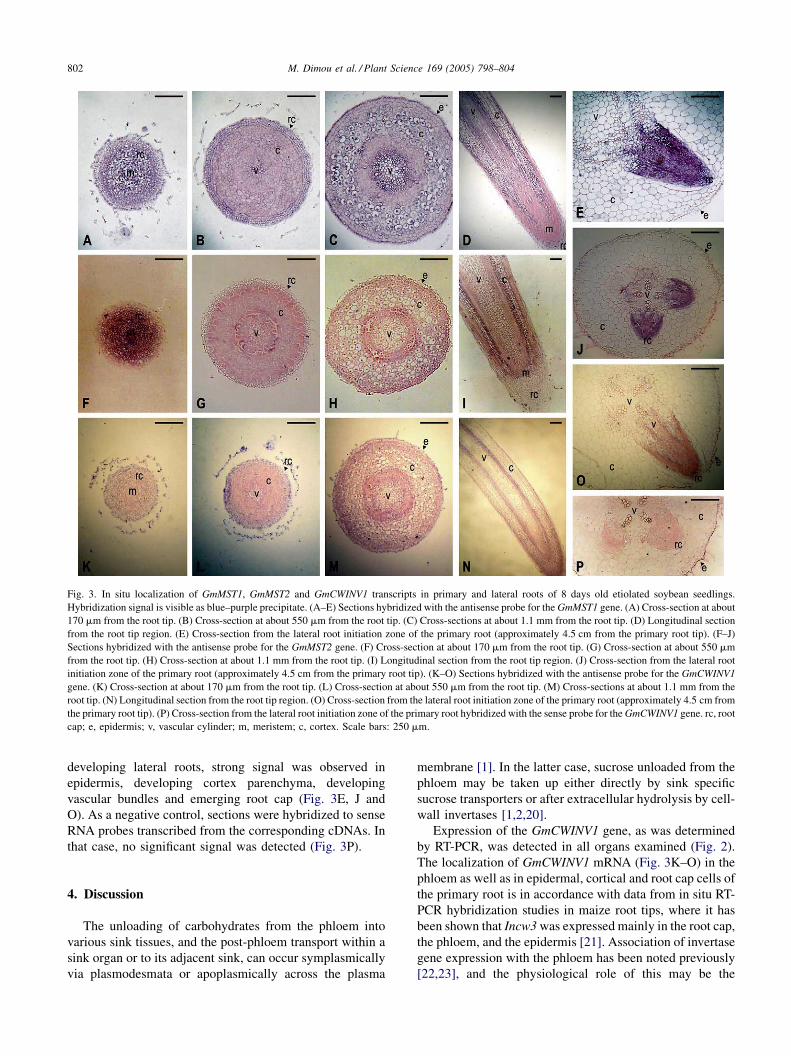
Fig. 3. In situ localization of GmMST1, GmMST2 and GmCWINV1 transcripts in primary and lateral roots of 8 days old etiolated soybean seedlings.
Hybridization signal is visible as blue–purple precipitate. (A–E) Sections hybridized with the antisense probe for the GmMST1 gene. (A) Cross-section at about
170 mm from the root tip. (B) Cross-section at about 550 mm from the root tip. (C) Cross-sections at about 1.1 mm from the root tip. (D) Longitudinal section
from the root tip region. (E) Cross-section from the lateral root initiation zone of the primary root (approximately 4.5 cm from the primary root tip). (F–J)
Sections hybridized with the antisense probe for the GmMST2 gene. (F) Cross-section at about 170 mm from the root tip. (G) Cross-section at about 550 mm
from the root tip. (H) Cross-section at about 1.1 mm from the root tip. (I) Longitudinal section from the root tip region. (J) Cross-section from the lateral root
initiation zone of the primary root (approximately 4.5 cm from the primary root tip). (K–O) Sections hybridized with the antisense probe for the GmCWINV1
gene. (K) Cross-section at about 170 mm from the root tip. (L) Cross-section at about 550 mm from the root tip. (M) Cross-sections at about 1.1 mm from the
root tip. (N) Longitudinal section from the root tip region. (O) Cross-section from the lateral root initiation zone of the primary root (approximately 4.5 cm from
the primary root tip). (P) Cross-section from the lateral root initiation zone of the primary root hybridized with the sense probe for the GmCWINV1 gene. rc, root
cap; e, epidermis; v, vascular cylinder; m, meristem; c, cortex. Scale bars: 250 mm.
developing lateral roots, strong signal was observed in
epidermis, developing cortex parenchyma, developing
vascular bundles and emerging root cap (Fig. 3E, J and
O). As a negative control, sections were hybridized to sense
RNA probes transcribed from the corresponding cDNAs. In
that case, no significant signal was detected (Fig. 3P).
4. Discussion
The unloading of carbohydrates from the phloem into
various sink tissues, and the post-phloem transport within a
sink organ or to its adjacent sink, can occur symplasmically
via plasmodesmata or apoplasmically across the plasma
membrane [1]. In the latter case, sucrose unloaded from the
phloem may be taken up either directly by sink specific
sucrose transporters or after extracellular hydrolysis by cell-
wall invertases [1,2,20].
Expression of the GmCWINV1 gene, as was determined
by RT-PCR, was detected in all organs examined (Fig. 2).
The localization of GmCWINV1 mRNA (Fig. 3K–O) in the
phloem as well as in epidermal, cortical and root cap cells of
the primary root is in accordance with data from in situ RT-
PCR hybridization studies in maize root tips, where it has
been shown that Incw3 was expressed mainly in the root cap,
the phloem, and the epidermis [21]. Association of invertase
gene expression with the phloem has been noted previously
[22,23], and the physiological role of this may be the

M. Dimou et al. / Plant Science 169 (2005) 798–804 803
maintenance of a steep sucrose concentration gradient
between source and sink regions of the plant and/or the
supply of hexoses, and thus energy, for the metabolism of
companion cells.
The expression of GmCWINV1 in the cell division area of
the root, at a stage when mitotic activity proceeds in the root
tip, might be critical for the supply of fast dividing cells with
hexoses, which in turn may act as a developmental signal.
The importance of the high ratio of hexoses to sucrose in
promoting mitotic activity has been indicated [10,24]. In the
root tip region, GmCWINV1 may, also, double the osmotic
contribution of unloaded sucrose to apoplasmic osmotic
pressures that affect sieve element turgor, and hence rates of
photoassimilate import, irrespective of the primary pathway
of unloading [5,25]. Furthermore, biochemical and physio-
logical studies provide evidence that cell-wall invertases are
most active at sites of sugar transport within rapidly
importing apoplasmic sinks [4,5].
Sink tissues can acquire the products of extracellular
hydrolysis of sucrose, via monosaccharide transport
proteins. Our in situ hybridization experiments have
indicated that the GmMST1 gene was mainly expressed in
the regions of cell division and elongation of the primary
root (Fig. 3A–E). The cells expressing the GmMST1 gene
were generally located in the vascular cylinder as well as in
the epidermis and underlying cell layers of the region of
elongation of the primary root while lower levels of
expression were detected in the cortex parenchyma.
Furthermore, expression of the GmMST1 gene was strongly
up-regulated at the lateral root primordia emerging from the
primary root.
Expression of GmMST2 was observed both in sink and
source organs, as was determined by RT-PCR analysis
(Fig. 2). In situ hybridization analysis revealed that
GmMST2 was predominantly expressed in the phloem of
the primary root as well as in most of the cells of the lateral
root primordia (Fig. 3J). In general, plant monosaccharide/
H+ transporters are thought to function in hexose import into
sink tissues [15,26], although expression associated with
source tissues has been reported [15,27], where they may
play a role in retrieval of monosaccharides lost to the
apoplasmic space by passive leakage [15].
The present study strongly indicates that the unloaded
sucrose in the apoplasmic space may be hydrolyzed into
hexoses by a cell-wall invertase, while the distribution of
the GmCWINV1 mRNA in the epidermis, cortex and root
cap cells suggests that the post-phloem long-distance
transport of sucrose may also be partly apoplasmic. There is
evidence that monosaccharide/H+ transporter gene expres-
sion is coordinately induced with that of cell-wall
invertases. This regulatory mechanism results in higher
tissue uptake of sucrose, due to the apoplasmic invertase
activity, and of hexoses, due to the monosaccharide/H+
transporter activity [9–12,28]. However, such a regulatory
mechanism in etiolated soybean roots remains to be
confirmed.
In conclusion, we have characterized three genes from G.
max that code for two putative monosaccharide/H+
transporters, GmMST1 and GmMST2, and one putative
cell-wall invertase, GmCWINV1. Our temporal and spatial
gene expression studies revealed that GmMST1 and
GmMST2 are expressed in the same organs at the same
developmental stage, albeit at different tissues, suggesting
that different monosaccharide/H+ transporters may play
different roles in both source and sink organs and in various
tissues during root development. In accordance to that, it
would be of great interest to compare the two proteins in
terms of substrate specificity and kinetic properties of
transport activity. Furthermore, these expression patterns are
similar enough with the expression pattern of the
GmCWINV1 gene, suggesting, that, at least part of the
unloaded sucrose in soybean root may be hydrolyzed into
hexoses before being transported from the apoplasmic space
into the respective sink cells.
References
[1] L.E. Williams, R. Lemoine, N. Sauer, Sugar transporters in higher
plants—a diversity of roles and complex regulation, Trends Plant Sci.
5 (2000) 283–290.
[2] W. Eschrich, Free space invertase, its possible role in phloem unload-
ing, Ber. Dtsch. Bot. Ges. 93 (1980) 363–378.
[3] D.E. Godt, T. Roitsch, Regulation and tissue specific distribution of
mRNAs for three extracellular invertase isoenzymes of tomato sug-
gests an important function in establishing and maintaining sink
metabolism, Plant Physiol. 115 (1997) 273–282.
[4] Z. Tymowska-Lalanne, M. Kreis, Expression of the Arabidopsis
thaliana invertase gene family, Planta 207 (1998) 259–265.
[5] T. Roitsch, R. Ehneb, M. Goetz, B. Hause, M. Hofmann, A. Krishna
Sinha, Regulation and function of extracellular invertases from higher
plants in relation to assimilate partitioning, stress responses and sugar
signaling, Aust. J. Plant Physiol. 27 (2000) 815–825.
[6] A. Sturm, Invertases. Primary structures, functions, and roles in
plant development and sucrose partitioning, Plant Physiol. 121
(1999) 1–7.
[7] W.H. Cheng, E.W. Talliercio, P.S. Chourey, The miniature1 seed locus
of maize encodes a cell wall invertase required for normal develop-
ment of endosperm and maternal cells in the pedicel, Plant Cell 8
(1996) 971–983.
[8] G.Q. Tang, M. Luscher, A. Sturm, Antisense repression of vacuolar
and cell wall invertase in transgenic carrot alters early plant devel-
opment and sucrose partitioning, Plant Cell 11 (1999) 177–189.
[9] V. Fotopoulos, J.K. Pittman, A.C. Marver, A.J. Buckana, N. Sauer, J.L.
Hall, L.E. Williams, The monosaccharide transporter gene, AtSTP4,
and the cell-wall invertase, Atbetafruct1, are induced in Arabidopsis
during infection with the fungal biotroph Erysiphe cichoracearum,
Plant Physiol. 132 (2003) 821–829.
[10] H. Weber, L. Borisjuk, U. Heim, P. Buchner, U. Wobus, Seed coat-
associated invertases of fava bean control both unloading and storage
functions: cloning of cDNAs and cell type-specific expression, Plant
Cell 7 (1995) 1835–1846.
[11] H. Weber, L. Borisjuk, U. Heim, N. Sauer, U. Wobus, A role for sugar
transporters during seed development: molecular characterization of a
hexose and a sucrose carrier in fava bean seeds, Plant Cell 9 (1997)
895–908.
[12] W. Weschke, R. Panitz, S. Gubatz, Q. Wang, R. Radchuk, H. Weber, U.
Wobus, The role of invertases and hexose transporters in controlling

M. Dimou et al. / Plant Science 169 (2005) 798–804804
sugar ratios in maternal and filial tissues of barley caryopses during
early development, Plant J. 33 (2003) 395–411.
[13] A. Schneidereit, J. Scholz-Starke, M. Buttner, Functional character-
ization and expression analyses of the glucose-specific AtSTP9 mono-
saccharide transporter in pollen of Arabidopsis, Plant Physiol. 133
(2003) 182–190.
[14] J. Scholz-Starke, M. Buttner, N. Sauer, AtSTP6, a new pollen-specific
H+-monosaccharide symporter from Arabidopsis, Plant Physiol. 131
(2003) 70–77.
[15] M. Buttner, N. Sauer, Monosaccharide transporters in plants: structure,
function and physiology, Biochim. Biophys. Acta 1465 (2000)
263–274.
[16] S.M. Sherson, H.L. Alford, S.M. Forbes, G. Wallace, S.M. Smith,
Roles of cell-wall invertases and monosaccharide transporters in the
growth and development of Arabidopsis, J. Exp. Bot. 54 (2003) 525–
582.
[17] S.F. Altschul, T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang, W.
Miller, D.J. Lipman, Gapped BLAST and PSI-BLAST: a new gen-
eration of protein database search programs, Nucleic Acids Res. 25
(1997) 3389–3402.
[18] J.A. Brusslan, E.M. Tobin, Light-independent developmental regula-
tion of cab gene expression in Arabidopsis thaliana seedlings, Proc.
Natl. Acad. Sci. U.S.A. 89 (1992) 7791–7795.
[19] R. Lemoine, Sucrose transporters in plants: update on function and
structure, Biochim. Biophys. Acta 1465 (2000) 246–262.
[20] N. Sauer, K. Baier, M. Gahrtz, R. Stadler, J. Stolz, E. Truernit, Sugar
transport across the plasma membranes oh higher plants, Plant Mol.
Biol. 26 (1994) 1671–1679.
[21] J.Y. Kim, A. Mahe, S. Guy, J. Brangeon, O. Roche, P.S. Chourey, J.L.
Prioul, Characterization of two members of the maize gene family,
Incw3 and Incw4, encoding cell-wall invertases, Gene 245 (2000)
89–102.
[22] P.E. Hedley, A.L. Maddison, D. Davidson, G.C. Machray, Differential
expression of invertase genes in internal and external phloem tissues of
potato (Solanum tuberosum L.), J. Exp. Bot. 51 (2000) 817–821.
[23] L. Zhang, N.S. Cohn, J.P. Mitchell, Induction of a pea cell-wall
invertase by wounding and its localized expression in phloem, Plant
Physiol. 112 (1996) 1111–1117.
[24] H. Weber, P. Buchner, L. Borisjuk, U. Wobus, Sucrose metabolism
during cotyledon development of Vicia faba L. is controlled by the
concerted action of both sucrose-phosphate synthase and sucrose
synthase: expression patterns, metabolic regulation and implications
for seed development, Plant J. 9 (1996) 841–850.
[25] S. Lalonde, M. Tegeder, M. Throne-Holst, W.B. Frommer, J.W.
Patrick, Phloem loading and unloading of sugars and amino acids,
Plant Cell Environ. 26 (2003) 37–56.
[26] S. Lalonde, E. Boles, H. Hellmann, L. Barker, J.W. Patrick, W.B.
Frommer, The dual function of sugar carriers: transport and sugar
sensing, Plant Cell 11 (1999) 707–726.
[27] A. Weig, J. Franz, N. Sauer, E. Komor, Isolation of a family of cDNA
clones from Ricinus communis L. with close homology to the hexose
carriers, J. Plant Physiol. 143 (1994) 178–183.
[28] R. Ehness, M. Ecker, E. Godt, T. Roitsch, Glucose and stress inde-
pendently regulate source and sink metabolism and defense mechan-
isms via signal transduction pathways involving protein
phosphorylation, Plant Cell 9 (1997) 1825–1841.




















