
Inducible bacteriocin production in Lactobacillus is regulated by differential expression of the pln...
12
Molecular Microbiology (2003) 47 (2), 483–494 © 2003 Blackwell Publishing Ltd Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Publishing, 200347Original Article Inducible bacteriocidin production in LactobacillusD. B. Diep et al. Accepted 10 October, 2002. *For correspondence. E-mail dzung. [email protected]; Tel. ( + 47) 64 948 544/9469; Fax ( + 47) 64 941 465. Inducible bacteriocin production in Lactobacillus is regulated by differential expression of the pln operons and by two antagonizing response regulators, the activity of which is enhanced upon phosphorylation Dzung B. Diep,* Ronny Myhre, Ola Johnsborg, Ågot Aakra and Ingolf F. Nes Laboratory of Microbial Gene Technology, Department of Chemistry and Biotechnology, Agricultural University of Norway, PO Box 5051, N-1432 Ås, Norway. Summary Expression of the five ( pln ) operons involved in the bacteriocin production of Lactobacillus plantarum C11 is regulated by a so-called pheromone-based signal-transducing network, in which the peptide pheromone (PlnA) induces bacteriocin production through the action of a histidine protein kinase (PlnB) and two antagonizing response regulators (PlnC as an activator and PlnD as a negative regulator). All pln -regulated promoters contain a conserved pair of direct repeats that serve as binding sites for PlnC and PlnD. In the present work, we show that the five PlnA- responsive operons are differentially expressed with regard to both timing and strength, and that the pher- omone triggers a strong autoactivating loop of the regulatory unit ( plnABCD ) during an early stage of induction that gradually leads to enhanced activa- tion of the other operons. The transport operon ( plnGHSTUV ), which is involved in the secretion of the pheromone and bacteriocins, is also expressed relatively early upon induction, but is quickly turned off soon after peak expression. Further investigation of the various promoters revealed that, although sub- tle differences within the promoter regions could account for the observed differential regulation, the presence of a downstream promoter-proximal se- quence in one promoter was found to cause delayed peak activity. How phosphorylation regulates the activity of the pln response regulators was also accessed by direct mutagenesis at their phosphory- lation sites. It was found that the two response regu- lators exert activity at two different levels: a low level when they are not phosphorylated and an elevated level when they are phosphorylated. The present data demonstrate that bacteriocin production in L. plan- tarum C11 is a highly regulated process, in which different regulatory mechanisms are applied to fine tune the timing and strength of expression of the five pln operons. Introduction Recently, an increasing number of Gram-positive bac- teria have been reported to apply a so-called peptide pheromone-based signal-transducing pathway to regulate various cellular activities, including competence develop- ment ( com ) (Håvarstein and Morrison, 1999; Lazazzera et al ., 1999), secretion of staphylococcal toxins during stationary growth ( agr ) (Novick, 1999) as well as the pro- duction of antimicrobial peptides, also known as bacteri- ocins, from numerous lactic acid bacteria (Nes et al ., 1996; Dunny and Leonard, 1997; Kleerebezem et al ., 1997; Nes and Eijsink, 1999). Such a regulatory network, which can be regarded mechanistically as a more special- ized form of the classical two-component regulatory sys- tems (Stock et al ., 1989; 1995; Parkinson, 1995), normally involves a tight interplay of three genetically linked components: a secreted peptide pheromone, a histidine protein kinase (HPK) and a response regulator (RR) (Diep et al ., 1995; Eijsink et al ., 1996; Nes et al ., 1996; Kleerebezem et al ., 1997; Nes and Eijsink, 1999). The HPK component serves as the receptor for the peptide pheromone (Brurberg et al ., 1997; Håvarstein and Morrison, 1999; Diep et al ., 2001). Based on analogy with other two-component regulatory systems (Stock et al ., 1989; 2000), it is believed that the signal provided by the pheromone is then processed and transferred to the cognate RR through a series of phosphorylation reac- tions. Once phosphorylated, the RR protein triggers an output such as expression of a subset of genes. This type of regulation, often referred to as quorum sensing, allows bacteria in a population to act collectively in a concerted manner in response to environmental changes (Håvarstein and Morrison, 1999).
Transcript of Inducible bacteriocin production in Lactobacillus is regulated by differential expression of the pln...

Molecular Microbiology (2003)
47
(2), 483–494
© 2003 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382XBlackwell Publishing, 200347Original Article
Inducible bacteriocidin production in LactobacillusD. B. Diep et al.
Accepted 10 October, 2002. *For correspondence. E-mail [email protected]; Tel. (
+
47) 64 948 544/9469; Fax (
+
47) 64 941 465.
Inducible bacteriocin production in
Lactobacillus
is regulated by differential expression of the
pln
operons and by two antagonizing response regulators, the activity of which is enhanced upon phosphorylation
Dzung B. Diep,* Ronny Myhre, Ola Johnsborg, Ågot Aakra and Ingolf F. Nes
Laboratory of Microbial Gene Technology, Department of Chemistry and Biotechnology, Agricultural University of Norway, PO Box 5051, N-1432 Ås, Norway.
Summary
Expression of the five (
pln
) operons involved in thebacteriocin production of
Lactobacillus plantarum
C11 is regulated by a so-called pheromone-basedsignal-transducing network, in which the peptidepheromone (PlnA) induces bacteriocin productionthrough the action of a histidine protein kinase (PlnB)and two antagonizing response regulators (PlnC asan activator and PlnD as a negative regulator). All
pln
-regulated promoters contain a conserved pair ofdirect repeats that serve as binding sites for PlnC andPlnD. In the present work, we show that the five PlnA-responsive operons are differentially expressed withregard to both timing and strength, and that the pher-omone triggers a strong autoactivating loop of theregulatory unit (
plnABCD
) during an early stage ofinduction that gradually leads to enhanced activa-tion of the other operons. The transport operon(
plnGHSTUV
), which is involved in the secretion ofthe pheromone and bacteriocins, is also expressedrelatively early upon induction, but is quickly turnedoff soon after peak expression. Further investigationof the various promoters revealed that, although sub-tle differences within the promoter regions couldaccount for the observed differential regulation, thepresence of a downstream promoter-proximal se-quence in one promoter was found to cause delayedpeak activity. How phosphorylation regulates theactivity of the
pln
response regulators was alsoaccessed by direct mutagenesis at their phosphory-lation sites. It was found that the two response regu-lators exert activity at two different levels: a low level
when they are not phosphorylated and an elevatedlevel when they are phosphorylated. The present datademonstrate that bacteriocin production in
L. plan-tarum
C11 is a highly regulated process, in whichdifferent regulatory mechanisms are applied to finetune the timing and strength of expression of the five
pln
operons.
Introduction
Recently, an increasing number of Gram-positive bac-teria have been reported to apply a so-called peptidepheromone-based signal-transducing pathway to regulatevarious cellular activities, including competence develop-ment (
com
) (Håvarstein and Morrison, 1999; Lazazzera
et al
., 1999), secretion of staphylococcal toxins duringstationary growth (
agr
) (Novick, 1999) as well as the pro-duction of antimicrobial peptides, also known as bacteri-ocins, from numerous lactic acid bacteria (Nes
et al
.,1996; Dunny and Leonard, 1997; Kleerebezem
et al
.,1997; Nes and Eijsink, 1999). Such a regulatory network,which can be regarded mechanistically as a more special-ized form of the classical two-component regulatory sys-tems (Stock
et al
., 1989; 1995; Parkinson, 1995), normallyinvolves a tight interplay of three genetically linkedcomponents: a secreted peptide pheromone, a histidineprotein kinase (HPK) and a response regulator (RR)(Diep
et al
., 1995; Eijsink
et al
., 1996; Nes
et al
., 1996;Kleerebezem
et al
., 1997; Nes and Eijsink, 1999). TheHPK component serves as the receptor for the peptidepheromone (Brurberg
et al
., 1997; Håvarstein andMorrison, 1999; Diep
et al
., 2001). Based on analogy withother two-component regulatory systems (Stock
et al
.,1989; 2000), it is believed that the signal provided bythe pheromone is then processed and transferred to thecognate RR through a series of phosphorylation reac-tions. Once phosphorylated, the RR protein triggers anoutput such as expression of a subset of genes. This typeof regulation, often referred to as quorum sensing,allows bacteria in a population to act collectively in aconcerted manner in response to environmental changes(Håvarstein and Morrison, 1999).

484
D. B. Diep
et al.
© 2003 Blackwell Publishing Ltd,
Molecular Microbiology
,
47
, 483–494
Lactobacillus plantarum
C11 regulates its bacteriocinproduction through such a pheromone-based regulatorymechanism (Diep
et al
., 1994; 1995; 1996; Anderssen
et al
., 1998). The bacteriocin locus contains five operons:two of them (
plnEFI
and
plnJKLR
) code for bacteriocinsand immunity proteins, one operon (
plnGHSTUV
) for anABC transport system to secrete peptides containing dou-ble-glycine N-terminal leaders (Håvarstein
et al
., 1995),one operon (
plnABCD
) for the signal-transducing pathway,and the last operon (
plnMNOP
) containing genes withunknown functions in bacteriocin synthesis. In the regula-tory operon,
plnA
codes for the peptide pheromone PlnA,
plnB
for a HPK and
plnCD
for two RRs. All
pln
operonsare repressed during non-producing conditions. Uponinduction by PlnA, transcription of all five operons is reg-ulated in a co-ordinated manner: they are all activatedduring the exponential growth phase of the bacteria andsomehow downregulated before the stationary growthphase (Diep
et al
., 1996). This co-ordinated expressionsuggests a common regulatory network for all five oper-ons. This view is supported by the fact that the promotersassociated with these operons share some common fea-tures: They all contain a poorly conserved
-
35 region andare preceded by a conserved pair of direct repeats servingas regulatory elements (Diep
et al
., 1996; 2001). We havedemonstrated recently that the two
pln
RRs bind directlyon these conserved repeats as homodimers in a co-operative manner to regulate gene expression (Risøen
et al
., 1998; 2001). Interestingly, although the two RRproteins are very similar at the amino acid level (over75% similarity) and even recognize the same genetic reg-ulatory elements, they differ from each other in function:PlnC activates transcription and bacteriocin production,whereas PlnD somehow represses both processes (Diep
et al
., 1994; 2001). The exact mechanism(s) that controltheir activity so that they do not antagonize each other inan uncontrolled manner are still poorly understood.
In the present work, we sought to gain further insightinto the different mechanisms that control gene expres-sion during the induction process. We report that thefive promoters, despite their overall similarity, are differen-tially regulated in both strength and timing. We also pro-vide evidence showing that phosphorylation enhancesthe activity of the two
pln
regulators, which otherwisehave some low activity when they are in their non-phosphorylated form.
Results
The
pln
promoters are differentially regulated with regard to timing and strength
To study promoter activity of the five
pln
operons, weselected DNA fragments ranging from position
-
125/
-
126upstream of their transcription start (referred to as
+
1) to
the start codon of their first gene to represent the differentpromoters (see Table 1). The selected DNA fragments,which contain the promoter core region (
-
35 and
-
10boxes) and the upstream regulatory repeats, are hence ofabout the same length with regard to the region upstreamof the transcription start (
+
1) but differ in length in the(non-coding) region downstream of this site (see Table 1).The different promoter fragments, termed P
A
, P
E
, P
G
, P
J
and P
M
, derived from the operons
plnABCD
,
plnEFI
,
plnGHSTUV
,
plnJKLR
and
plnMNOP
, respectively, wereobtained by polymerase chain reaction (PCR) and thenligated just upstream of the promoterless
cat
in pGKV210,which was used as a reporter plasmid. After transforma-tion into C11 cells, the different clones were induced withPlnA (200 ng ml
-
1
), and CAT activity was then assayed asdescribed in
Experimental procedures
.As shown in Fig. 1A, P
A
of the regulatory operon wasstrongly activated during the early stage of induction. Itreached peak activity between time points 2 h and 3 hbefore declining gradually later in growth. The activityprofiles for P
G
and P
E
somewhat resemble that for P
A
,albeit with some differences: their peak activity appearedto be weaker and slightly delayed and, in the hours afterpeak expression, their activity also declined more quickly.In contrast, the activity profiles for P
M
and P
J
differ remark-ably from the others: the first was significantly delayed,peaking about 6 h after induction, whereas the latter dis-played little or no activity at all.
The five promoters examined above contain non-codingsequences located just downstream of their transcriptionstart sites; these associated sequences are of differentlengths (see Table 1). To test whether any of these ele-ments, especially those of P
M
and P
J
, could account forthe differential regulation of their promoter activity, wedesigned shorter promoter fragments in which most of thedownstream non-coding sequences were omitted. Thus,the new promoter fragments started at the same positionas before, but all ended at position
+
13, giving rise to DNAfragments (termed P
¢
E
, P
¢
G
, P
¢
J
, P
¢
M
) of the same lengthas P
A
, which has a minimal non-coding sequence (seeTable 1). The activity of the new promoter fragments isshown in Fig. 1B. It can be seen that the removal of thenon-coding sequence from P
M
(resulting in P
¢
M
) gave riseto a stronger and earlier peak activity. On the other hand,the removal of the non-coding sequence from P
J
(resultingin P
¢
J
) increased the activity only slightly; this resultimplies that the poor activity of P
J
was not entirely causedby the associated non-coding sequence. For the remain-ing promoters, P
¢
E
and P
¢
G
, no significant difference wasobserved with regard to timing. However, unlike P
¢
E
, thestrength of which was comparable to that of P
E
, thestrength of P
¢
G
was significantly reduced (
ª
50%) com-pared with that of P
G
.In view of the above results, we hypothesized that P
A
,

Pheromone-regulated bacteriocin production
485
© 2003 Blackwell Publishing Ltd,
Molecular Microbiology
,
47
, 483–494
Tab
le 1
.
Pln
A-r
espo
nsiv
e pr
omot
ers
and
thei
r se
quen
ce.
Pro
mot
ers
LRR
R
-
35
-
10T
S
P
A
(GA
ATT
) C
TT
TG
AC
ATT
CAT
CA
AAT
ATT
GAT
TG
ATA
GC
ATA
GT
TG
GA
ATT
TC
ATG
GT
GAT
TC
AC
GT
TTA
AAT
TTA
AA
AA
ATG
TAC
GT
TAAT
AG
AA
ATA
ATT
CC
TC
CG
TAC
TT
CA
AA
AA
CA
CAT
TAT
CC
TA
AA
AG
C
G
AG
GT
GAT
TAT
T
A
(G
GAT
CC
) (1
2)
P
M
(GA
ATT
C)
TT
GA
CT
TTA
ATT
GT
TG
GT
TAA
AC
GT
CTA
TT
TT
CG
TG
GTA
TC
GAT
GA
ATTA
TT
GTA
CG
ATA
ATA
TC
TAA
AA
ATAT
TAC
GT
TTA
TAA
AA
ATAT
CG
TAC
AAT
GAT
TC
GA
CTA
AG
CG
GTA
TAT
TA
AA
AG
CG
TAG
AG
AG
AG
TG
ATAT
CA
AG
CA
GT
TC
GAT
TG
AAT
TG
AAT
GA
AA
GC
GAT
TT
CT
TG
TG
TG
TG
AG
TC
TATA
TTA
AA
AAT
AAT
TG
TT
GT
TTA
GA
AA
GC
AG
TG
GG
AA
G
A
(G
GAT
CC
) (1
03)
P
G
(GA
ATT
C)
AA
CG
TC
AA
ATA
CC
AAT
GT
TAG
CTA
GT
TG
ATG
TT
TAA
CC
GT
TAA
GC
CT
GAT
GA
GG
AC
ATT
TAT
CAT
AA
AAT
TAT
GTA
CG
TTA
ATA
GAT
AG
TT
GG
CA
TAC
GAT
AA
C
ATT
TG
TTA
GC
CC
ATA
ATTA
AAT
A
G
ATA
CC
GAT
AC
TTA
ATA
ATT
GG
TG
AG
GG
GA
GTA
CA
AG
T
A
(G
GAT
CC
) (3
8)
P
J
(GA
ATT
C)
GC
AA
AA
ATA
ATC
CC
ATTA
ATG
TTA
TG
GG
ATC
GT
CA
GA
CAT
TC
AA
CT
TT
CA
AG
TTA
CG
TTA
AA
TC
GAT
TAA
ATA
GTA
CG
ATA
AC
aaatttaaaataattttttttaaatt
GTA
GC
GTA
TAT
TA
ATA
AG
TG
CAT
TAA
AA
ATA
CG
TAG
CG
CAT
AG
AA
CC
ATT
TT
TAA
CT
GC
TAC
AG
TAA
AG
GG
AG
AG
GT
A (
GG
ATC
C)
(58)
PE
(GA
ATT
C)
AAT
GT
CC
TC
ATC
AG
GC
TTA
AC
GG
TTA
AA
CAT
CA
AC
TAG
CTA
AC
ATT
GG
TAT
TT
GA
CG
TTA
AG
AG
AA
CG
TT
TT
TT
TAC
TT
TTA
TA
ATT
TT
TT
CA
AC
AAT
CT
GG
TAA
AA
AA
ATA
AAT
TAA
AC
TA
AAT
TT
GT
TC
CA
AAT
AA
CAT
CAT
AC
AA
GG
GG
GAT
TAT
TT
A (
GG
ATC
C)
(33)
P¢ M
(GA
ATT
C)
TT
GA
CT
TTA
ATT
GT
TG
GT
TAA
AC
GT
CTA
TT
TT
CG
TG
GTA
TC
GAT
GA
ATTA
TT
GTA
CG
ATA
ATAT
CTA
AA
AAT
ATTA
CG
TT
TAT
AA
AA
ATAT
CG
TAC
AAT
GAT
TC
GA
CTA
AG
CG
GTA
TAT
TA
AA
AG
CG
TAG
AG
AG
AG
TG
A(G
GAT
CC
)
P¢ G
(GA
ATT
C)
AA
CG
TC
AA
ATA
CC
AAT
GT
TAG
CTA
GT
TG
ATG
TT
TAA
CC
GT
TAA
GC
CT
GAT
GA
GG
AC
ATT
TAT
CAT
AA
AAT
TAT
GTA
CG
TTA
ATA
GAT
AG
TT
GG
CA
TAC
GAT
AA
CAT
TT
GT
TAG
CC
CAT
AAT
TAA
ATA
GAT
AC
CG
ATA
CTA
(G
GAT
CC
)
P¢ J
(GA
ATT
C)
GC
AA
AA
ATA
ATC
CC
ATTA
ATG
TTA
TG
GG
ATC
GT
CA
GA
CAT
TC
AA
CT
TT
CA
AG
TTA
CG
TTA
AA
TC
GAT
TAA
ATA
GTA
CG
ATA
ACaaatttaaaataattttttttaaatt
GTA
GC
GTA
TAT
TA
ATA
AG
TG
CAT
TAA
AA
ATA
A (
GG
ATC
C)
P¢ E
(GA
ATT
C)
AAT
GT
CC
TC
ATC
AG
GC
TTA
AC
GG
TTA
AA
CAT
CA
AC
TAG
CTA
AC
ATT
GG
TAT
TT
GA
CG
TTA
AG
AG
AA
CG
TT
TT
TT
TAC
TT
TTA
TA
ATT
TT
TT
CA
AC
AAT
CT
GG
TAA
AA
AA
ATA
AAT
TAA
AC
TA
AAT
TT
GT
TC
CA
AAT
AA
CA
(G
GAT
CC
)
PA
M(G
AAT
T)
CT
TT
GA
CAT
TC
ATC
AA
ATAT
TG
ATT
GAT
AG
CAT
AG
TT
GG
AAT
TT
CAT
GG
TG
ATT
CA
CG
TT
TAA
ATT
TAA
AA
AAT
GTA
CG
TTA
ATA
GA
AAT
AAT
TC
CT
CC
GTA
CT
TC
AA
AA
AC
AC
ATTA
TC
CT
AA
AA
GC
GTA
GA
GA
GA
GT
GAT
ATC
AA
GC
AG
TT
CG
ATT
GA
ATT
GA
ATG
AA
AG
CG
ATT
TC
TT
GT
GT
GT
GA
GT
CTA
TAT
TAA
AA
ATA
ATT
GT
TG
TT
TAG
AA
AG
CA
GT
GG
GA
AG
A (
GG
ATC
C)
PA
J(G
AAT
T)
CT
TT
GA
CAT
TC
ATC
AA
ATAT
TG
ATT
GAT
AG
CAT
AG
TT
GG
AAT
TT
CAT
GG
TG
ATT
CA
CG
TT
TAA
ATT
TAA
AA
AAT
GTA
CG
TTA
ATA
GA
AAT
AAT
TC
CT
CC
GTA
CT
TC
AA
AA
AC
AC
ATTA
TC
CT
AA
AA
GC
GC
ATTA
AA
AAT
AC
GTA
GC
GC
ATA
GA
AC
CAT
TT
TTA
AC
TG
CTA
CA
GTA
AA
GG
GA
GA
GG
TA
(G
GAT
CC
)
Left
(LR
) an
d rig
ht (
RR
) re
peat
s an
d pr
edic
ted
-35
and
-10
regi
ons
are
unde
rline
d, w
here
as t
rans
crip
tion
star
t (T
S)
and
the
first
bas
e (A
) of
the
sta
rt c
odon
are
indi
cate
d by
bol
d le
tters
. Seq
uenc
es in
bra
cket
s w
ere
adde
dto
inc
lude
res
tric
tion
site
s E
coR
I (G
AAT
TC
) or
Bam
HI
(GG
ATC
C).
Num
bers
at
the
end
of t
he fi
rst
five
prom
oter
s in
dica
te t
he l
engt
h of
non
-cod
ing
sequ
ence
fro
m T
S t
o th
e fir
st s
tart
cod
on (
ATG
). In
the
fus
ion
prom
oter
s(P
AM a
nd P
AJ)
, th
e no
n-co
ding
seq
uenc
es a
re d
eriv
ed f
rom
pln
M a
nd p
lnJ
prom
oter
s (in
dica
ted
by it
alic
lette
rs).
Not
e th
at o
nly
PJ
and
P¢ J
cont
ain
an e
xten
sive
AT-
rich
sequ
ence
of
26 n
ucle
otid
es (
smal
l let
ters
) lo
cate
d ju
stdo
wns
trea
m f
rom
the
rig
ht r
epea
t (R
R).
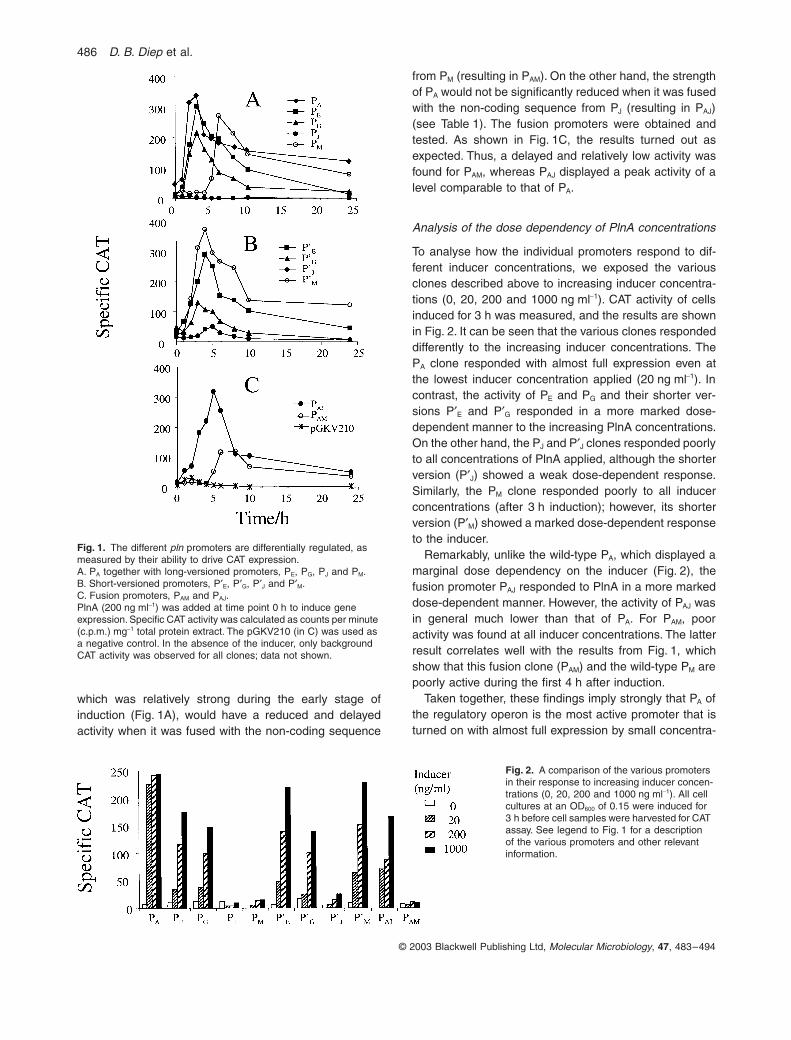
486 D. B. Diep et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 483–494
which was relatively strong during the early stage ofinduction (Fig. 1A), would have a reduced and delayedactivity when it was fused with the non-coding sequence
from PM (resulting in PAM). On the other hand, the strengthof PA would not be significantly reduced when it was fusedwith the non-coding sequence from PJ (resulting in PAJ)(see Table 1). The fusion promoters were obtained andtested. As shown in Fig. 1C, the results turned out asexpected. Thus, a delayed and relatively low activity wasfound for PAM, whereas PAJ displayed a peak activity of alevel comparable to that of PA.
Analysis of the dose dependency of PlnA concentrations
To analyse how the individual promoters respond to dif-ferent inducer concentrations, we exposed the variousclones described above to increasing inducer concentra-tions (0, 20, 200 and 1000 ng ml-1). CAT activity of cellsinduced for 3 h was measured, and the results are shownin Fig. 2. It can be seen that the various clones respondeddifferently to the increasing inducer concentrations. ThePA clone responded with almost full expression even atthe lowest inducer concentration applied (20 ng ml-1). Incontrast, the activity of PE and PG and their shorter ver-sions P¢E and P¢G responded in a more marked dose-dependent manner to the increasing PlnA concentrations.On the other hand, the PJ and P¢J clones responded poorlyto all concentrations of PlnA applied, although the shorterversion (P¢J) showed a weak dose-dependent response.Similarly, the PM clone responded poorly to all inducerconcentrations (after 3 h induction); however, its shorterversion (P¢M) showed a marked dose-dependent responseto the inducer.
Remarkably, unlike the wild-type PA, which displayed amarginal dose dependency on the inducer (Fig. 2), thefusion promoter PAJ responded to PlnA in a more markeddose-dependent manner. However, the activity of PAJ wasin general much lower than that of PA. For PAM, pooractivity was found at all inducer concentrations. The latterresult correlates well with the results from Fig. 1, whichshow that this fusion clone (PAM) and the wild-type PM arepoorly active during the first 4 h after induction.
Taken together, these findings imply strongly that PA ofthe regulatory operon is the most active promoter that isturned on with almost full expression by small concentra-
Fig. 1. The different pln promoters are differentially regulated, as measured by their ability to drive CAT expression.A. PA together with long-versioned promoters, PE, PG, PJ and PM. B. Short-versioned promoters, P¢E, P¢G, P¢J and P¢M.C. Fusion promoters, PAM and PAJ. PlnA (200 ng ml-1) was added at time point 0 h to induce gene expression. Specific CAT activity was calculated as counts per minute (c.p.m.) mg-1 total protein extract. The pGKV210 (in C) was used as a negative control. In the absence of the inducer, only background CAT activity was observed for all clones; data not shown.
Fig. 2. A comparison of the various promoters in their response to increasing inducer concen-trations (0, 20, 200 and 1000 ng ml-1). All cell cultures at an OD600 of 0.15 were induced for 3 h before cell samples were harvested for CAT assay. See legend to Fig. 1 for a description of the various promoters and other relevant information.

Pheromone-regulated bacteriocin production 487
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 483–494
tions of the inducer. On the other hand, the promoters ofthe other operons become highly active only at a muchhigher inducer concentration, suggesting that these pro-moters reach their peak activity at a later stage of induc-tion when the inducer has accumulated to higher levels.
Transcription analysis of the various pln operons in C11
Given that the five PlnA-responsive promoters are differ-entially regulated with regard to both timing and strength(Figs 1 and 2), we wanted to examine how these resultscorrelate with the transcription of their respective operonsin C11. RNAs obtained from induced and non-inducedC11 cells were analysed with DNA probes specific to theindividual pln transcripts. As shown in Fig. 3, transcriptionof all five operons could only be detected in induced cells,but not in non-induced cells, and the amounts of thetranscripts and the timing of gene expression variedbetween the different operons. For the regulatory operon(plnABCD), its transcript appeared to be highly expressedat all time points investigated, with peak expressionextending over a prolonged period of time, i.e. from timepoint 2 h to time point 5 h (Fig. 3A). For the plnMNOPoperon, its transcription seemed to follow a stepwise fash-ion: a very low but detectable level of expression wasobserved during the first 2 h after induction; in the next3 h (time points 3–5 h), the expression was increased toa significantly higher level before it diminished abruptly attime point 6 h (Fig. 3B). For the two bacteriocin operons
plnJKLR and plnEFI, their expression profiles seemed tobe quite alike: the amounts of their transcripts increasedgradually after the addition of the inducer, and they bothreached peak expression at time point 3 h before decreas-ing gradually during subsequent growth (Fig. 3C and D).Lastly, the fifth operon (plnGHSTUV) resembles the reg-ulatory operon (plnABCD), in that it also reached peakexpression relatively early, i.e. only 2 h after induction;however, from thereon, it differs from the others in that itwas downregulated earlier and also more quickly(Fig. 3E). It is noteworthy that the transport transcript(plnGHSTUV) was expressed at very low levels through-out the course of the experiment. The relative amounts ofthe different transcripts with regard to their individual peakexpression are shown in Fig. 3F.
To summarize, the transcription patterns of plnABCD(strong and early), plnMNOP (strong but delayed) andplnEFI (strong and intermediate in timing) seem to corre-late well with the activity profile of their respective promot-ers (Figs 1 and 2). On the other hand, the transcriptionpatterns of plnJKLR (relatively strong) and plnGHSTUV(poor) are not in accordance with the poor activity of PJ
and the moderate activity of PG (Fig. 1). Possible mecha-nisms responsible for this discrepancy in gene expressionare discussed below.
Analysis of non-phosphorylatable regulators
Most response regulators become activated upon phos-
Fig. 3. Northern analysis of the various pln transcripts in C11 cultures when grown with (+I) or without (–I) induction (200 ng ml-1 PlnA). RNA was isolated at the indicated time points, and three different amounts of RNA were spotted onto membranes (0.2, 1 and 5 mg). In addition, equal amounts (25 ng) of non-labelled probe DNAs (J, A, E, G and M) were spotted onto each membrane (before hybridization) and used as controls for specificity. Membranes were probed with probe A specific to plnABCD (A), probe M to plnMNOP (B), probe J to pln-JKLR (C), probe E to plnEFI (D) and probe G to plnGHSTUV.F. Relative amounts of the various transcripts in relation to their peak activity (indicated by arrows), as scanned by phosphorimager Typhoon 8600.

488 D. B. Diep et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 483–494
phorylation; this modification takes place at a conservedaspartate (D) residue located within their N-terminal half(Sanders et al., 1989; Stock et al., 1989). A sequencealignment of PlnC and PlnD with some known RRs (suchas CheY, NtrC, NodW and OmpR), the phosphorylationsites of which have been characterized, identified the Dresidue at position 59 (D59) as the potential phosphory-lation site for both PlnC and PlnD (see Fig. 4A). To gaininsight into how phosphorylation regulates the functional-ity/activity of PlnC and PlnD, we changed, by directmutagenesis, their D59 to an asparagine (N), which issimilar to D but cannot be phosphorylated. To study theserecombinant genes, we took advantage of a reportersystem that has been used in our recent study on theinterplay between the kinase PlnB and its RRs in theheterologous host Lactobacillus sakei Lb790 (Diep et al.,2001). The reporter system consists of two plasmids: pJB-GB, which contains the reporter gene gusA and the kinasegene plnB, each being preceded by its own plnA pro-moter; and, pJB37-C or pJB37-D, which contains eitherplnC or plnD driven by a constitutive promoter P32 (seeTable 3 and Fig. 4B). In such a reporter system, the het-erologous cells expressing plnC respond to the exoge-nously added inducer with a strong GUS expression,whereas cells expressing the other regulator gene (plnD)somehow respond to the inducer with a low but detectableGUS expression despite the fact that overexpression ofplnD in its homologous host L. plantarum C11 repressesboth bacteriocin production and transcription of theendogenous plnABCD operon (Diep et al., 2001). Some
of these results can also be seen in Figs 4 and 5. Whenthe wild-type regulator genes were replaced with theircorresponding D59N mutant genes (resulting in pJB37-Cmut and pJB37-Dmut), we could no longer observe anyincrease in GUS expression in response to theinducer (Fig. 4C). These findings indicate that non-phosphorylated PlnC and PlnD have little or no ability toactivate the reporter gene.
Next, we examined whether any of the mutated regula-tors can act as a negative regulator or an activator inbacteriocin production by overexpressing their genes intheir native bacteriocin producer C11. Such an experimenthas been performed previously with the wild-type genes,and that was how we established different functions ofPlnC and PlnD, the first being an activator and the secondacting as a negative regulator (Diep et al., 2001). Theeffects on bacteriocin production of the mutant genes andwild-type genes are compared in Fig. 5. It can be seenthat the overexpression of plnDmut also repressed theinduction of bacteriocin production because cells overex-pressing this gene required a relatively high inducer con-centration (50 ng ml-1 or more) to establish bacteriocinproduction. In comparison, the control clone, which con-tains only the vector (V), required much less inducer,i.e. 10 ng ml-1, to induce bacteriocin production. In fact,we have shown in other experiments that as little as1 ng ml-1 was sufficient to induce bacteriocin productionin this control clone (data not shown). However, therepression by plnDmut is clearly weaker than that exertedby the wild-type PlnD, as cells overexpressing the wild-
Fig. 4. Analysis of the mutant regulators in their ability to drive GUS expression in the het-erologous host L. sakei Lb790.A. Alignment of the response regulators PlnC and PlnD together with some known regulators whose phosphorylation sites (D) are character-ized (indicated in bold letters); OmpR (Delgado et al., 1993), NodW (Loh et al., 1997), NtrC Miranda-Rios et al., 1987) and CheY (Bourret et al., 1990). Only the residues flanking the phosphorylation sites are shown here. Number-ing indicates the position of the last residue. Stars indicate identical residues, whereas dots indicate conserved residues.B. The two-plasmid reporter system containing the reporter pJB-GB together with pJB37-C (plnC), pJB37-D (plnD), pJB37-Cmut (plnCmut), pJB37-Dmut (plnD) or pMS37c (V). gusA and plnB in pJB-GB are independently controlled by two separate inducible PlnA promoters (rectan-gles), whereas the regulator genes are con-trolled by a constitutive P32 promoter (triangle).C. GUS expression by the various clones. Cells were induced for 3 h with indicated inducer con-centrations before GUS activity was assayed. Specific GUS activity was calculated as GUS value (measured at 405 nm) per cell density (at 600 nm). Experiments were performed in dupli-cate, and mean values are shown here.

Pheromone-regulated bacteriocin production 489
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 483–494
type gene needed at least fivefold more inducer(250 ng ml-1) to establish bacteriocin production. Similarly,like that observed with the wild-type gene plnC, overex-pression of the mutant plnCmut, which has poor activity inLb790 (Fig. 4), also triggered bacteriocin production in theabsence of PlnA (Fig. 5); this finding suggests that PlnCmut
could have some trace activity that could have beenamplified by the intact autoregulatory network in C11(see Discussion).
Taken together, these studies strongly imply that bothPlnC and PlnD in their non-phosphorylated form possesssome low activities in gene regulation, but these activitiesare significantly enhanced upon phosphorylation by thekinase PlnB.
Discussion
In the present study, we have shown that the five plnoperons are differentially regulated with regard to bothstrength and timing. Most notably, the promoter of theregulatory operon (plnABCD) appeared as the most activeduring the early stage of induction, whereas the promotersof the other operons became highly active only at laterstages of induction. Based on the transcription analysis(Fig. 3), the early expression of the regulatory operonseems to be closely followed by the expression of theoperon plnGHSTUV, which codes for a transport systemdedicated to maturation and secretion of not only thebacteriocin peptides but also the inducer (Håvarsteinet al., 1995). Hence, it is reasonable to believe that theconcerted expression of these two operons is required tobring about a strong autoactivating loop of the regulatoryunit during the early stage of induction. In effect, thiswould lead to a rapid increase in the inducer concentrationin the culture, so that the expression of the other operonsthat require high inducer concentrations could beenhanced.
The two operons plnMNOP and plnJKLR differ signifi-cantly from the others in promoter activity; the promoter(PM) of the first operon regulates a much delayed activity,whereas the latter (PJ) hardly shows any activity at all(Fig. 1). In addition, these two promoters are flanked attheir 3¢ ends by relatively long non-coding sequences.Further characterization of these non-coding sequencesrevealed that only the one associated with PM could havea strong regulatory effect, whereas the other did not(Fig. 1). One possible explanation for the poor activity ofP¢J or PJ is the presence of a remarkably long AT-richstretch (26 nucleotides) located in the core region of thepromoter (see Table 1). It is possible that this AT-rich ele-ment might cause reduced transcription by destabilizingor preventing transcription initiation. However, furtherinvestigation is required for a more definitive answer.
We observed that the levels of the plnGHSTUV andplnJKLR transcripts did not correlate well with the resultsfrom the promoter study (Figs 1 and 3). Although the exactmechanism responsible for this discrepancy is unclear,these results might suggest an unknown regulatory mech-anism. It should be emphasized that there are importantdifferences in the way in which these two experimentswere set up. In the promoter study, the individual promot-ers were subcloned (in plasmids) and studied one by one.In contrast, the transcription analysis was performed to
Fig. 5. The effects of the wild-type regulator genes (plnC and plnD) and their non-phosphorylatable mutants (plnCmut and plnDmut) on bac-teriocin production in L. plantarum C11. Clones harbouring pJB36-C (C), pJB36-D (D), pJB36-Cmut (Cmut; two isolates), pJB36-Dmut (Dmut; two isolates) or pMG36e (V, used as a control) were spotted directly onto agar plates containing indicator cells (strain 965) and increasing concentrations of the inducer (0, 10, 50, 250 and 500 ng ml-1). Bacteriocin production is seen as inhibition zones surrounding the producer cells.

490 D. B. Diep et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 483–494
reveal transcription patterns of the individual operons intheir natural genetic environment, where in cis interactionsbetween proximal regulated promoters of the pln regulonmight occur and affect gene expression. Such a regulatorymechanism, which involves interactions between proximalDNA regulatory elements belonging to the same regula-tory network, has been identified in prokaryotes (exempli-fied by the ara regulon; Harmer et al., 2001) and even inviruses (such as the genetic switch in lambda; Ptashne,1992). Both the regulated promoters of plnGHSTUV andplnJKLR are in fact in proximal distance from the regu-lated promoters of plnEFI and plnMNOP respectively (seeFig. 6) and, therefore, it is tempting to speculate that thesetwo operons could be subject to such an in cis regulation.Another possible explanation is that the low levels of theplnGHSTUV transcript could result from an intrinsicmRNA instability. Evidence supporting this notion comesfrom a previous study (Diep et al., 1996), in which adetailed Northern analysis revealed that the plnGHSTUVtranscript was rapidly degraded, whereas the other PlnA-induced transcripts were relatively more stable. A lowexpression of the ABC transport system has also beenreported for the sap regulon, which regulates a similarinducible bacteriocin production in L. sakei through apheromone-based signalling pathway (Risøen et al.,
2000). As bacteriocin production is often regarded as anaccessory and energy-costly process, it is therefore fea-sible that a limited expression of the pheromone-secretingsystem could tightly regulate expression of the genesinvolved.
The second part of this work dealt with the pho-sphorylation of the regulators. We constructed non-phosphorylatable versions of the regulator genes by directmutagenesis to examine how phosphorylation affects theirfunctionality. Although the mutant genes were found to befunctionally active under certain conditions, they are ingeneral less active than their wild-type counterparts. Thestudy in the heterologous expression host clearlydemonstrated that only the phosphorylatable PlnC, but notthe non-phosphorylatable PlnCmut, could activate GUSexpression in response to the added inducer (Fig. 4).Similarly, although PlnDmut was found to be capable ofrepressing the induction of bacteriocin production, therepression caused by the corresponding wild-type pro-tein was much stronger (Fig. 5). In a recent study byRisøen et al. (2000), it was demonstrated that the non-phosphorylated form of either regulator could bind specif-ically to DNA regulatory repeats, but the DNA-bindingaffinity was enhanced for both regulators when the pro-teins were phosphorylated. These findings together sug-gest that phosphorylation enhances the activities of theregulators by increasing their DNA affinity. The fact thatPlnC or PlnCmut was capable of triggering bacteriocin pro-duction in C11 in the absence of the added inducer is alsoconsistent with the idea that the non-phosphorylated PlnCpossesses low activity. This is based on the followingreasoning. The activity of the non-phosphorylated PlnC(or by PlnCmut) might be low but sufficient to trigger theendogenous autoregulatory network (PlnABCD) in C11,the expression of which is then instrumental in the estab-lishment of bacteriocin production. Such a functional auto-regulatory network was absent in the heterologous hostLb790 and, therefore, the expression of plnC alone (with-out added inducer) or plnCmut (with added inducer) wasnot sufficient to drive GUS expression. A schematic modelsummarizing the possible functions of the different com-ponents of the pln regulatory network is shown in Fig. 6.It is pertinent to mention here that a number of regulators,e.g. PhoB (Makino et al., 1988), NodW (Loh et al., 1997)and OmpR (Igo et al., 1989), have been reported to reg-ulate genes in such a manner, i.e. low regulatory activitybeing converted to high activity upon phosphorylation.
In the present work, we have shown two different mech-anisms applied in the pln regulon: one involving sequencedifferences between the promoters that lay down the basisfor their differential regulation and the second involvingphosphorylation of two antagonizing regulators to regulatetheir activities. Apparently, a combination of these twomechanisms is necessary to fine tune expression of the
Fig. 6. Schematic model of the pln regulatory network. Interaction of the inducer PlnA (A) with the membrane-located HPK PlnB (B; unfilled) leads to autophosphorylation of PlnB (filled) (step 1). In the following step (2), the activated PlnB transfers the phosphatidyl group from its cytoplasmic domain to either PlnC or PlnD (unfilled), which is then converted to a fully active form (filled). Phosphorylated PlnC and PlnD bind the DNA regulatory elements of the pln promoters strongly (small filled rectangles in the pln locus) to regulate gene expression, PlnC as an activator (step 3) and PlnD as a negative regulator (step 4). The non-phosphorylated PlnC and PlnD possess some low but specific affinity to DNA and can also regulate gene expression to some extent, but not as strongly as their phosphorylated forms (steps 3¢ and 4¢). Organization of the five PlnA-responsive operons is shown.

Pheromone-regulated bacteriocin production 491
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 483–494
five pln operons in L. plantarum C11, not only during theactivation of bacteriocin production but also during thedownregulation process. Our study also implies that otherunknown mechanisms must be involved in order to bringabout a complete and dynamic regulation of the pln reg-ulon. For instance, it is challenging to unravel the mech-anism(s) that control the timing of PlnC and PlnD so thattheir antagonizing actions do not outdo each other in anuncontrolled manner.
Experimental procedures
Bacterial strains and growth conditions
Lactobacillus plantarum strains 965 and C11, L. sakei Lb790(LMG collection, Norway) and their derivatives were grown inMan–Rogosa–Sharpe (MRS) medium at 30∞C without shak-ing. Where appropriate, erythromycin and/or chlorampheni-col were added at 5 mg ml-1.
Plasmid construction and transformation
Most molecular techniques were performed as described bySambrook et al. (1989). PCR primers and all plasmids usedin this study are described in Tables 2 and 3. All primers wereobtained from Gibco BRL. Primers dbd62 and dbd63 wereused for PA, dbd64 and dbd65 for PM, dbd66 and dbd67 for
PG, dbd68 and dbd69 for PJ, dbd70 and dbd71 for PE, dbd64and dbd72 for P¢M, dbd66 and dbd73 for P¢G, dbd68 anddbd74 for P¢J and dbd70 and dbd75 for P¢E. For hybrid pro-moters, a two-step PCR protocol was used (Higuchi, 1990).In the first step, dbd62 and dbd77 were used for isolating thePA part, dbd76 and dbd65 for the non-coding sequence ofPM, and dbd78 and dbd69 for the non-coding sequence of PJ;in the second step, the PA part was fused with the non-codingsequences to obtain PAM and PAJ. In all cases, genomic DNAfrom C11 was used as template DNA in PCR. The resultingpromoter fragments, the sequences of which are shown inTable 1, were ligated into the reporter vector pGKV210 infront of the cat gene between restriction sites EcoRI andBamHI. Primers dbd60 and dbd61 were used to introducethe substitution D59N into plnC and plnD, giving rise toplnCmut and plnDmut. Correct sequence of all clones was con-firmed by DNA sequencing. Transformation of Lactobacillusstrains was performed as described by Aukrust et al. (1995).
Northern analysis
Growing cell cultures (OD600 of 0.15) were induced with PlnA(200 ng ml-1). Cells were then harvested at the indicated timepoints after induction, and DNA-free RNA was isolated usingan RNA isolation kit (Qiagen). RNA was disposed onto acellulose membrane using a vacuum slotblotter (Bio-Rad).Briefly, RNA dilutions (100 ml) were applied into wells followedby the addition of 200 ml of 10 mM NaOH and 1 mM EDTAto each well to denature. Membranes were then washed
Table 2. Primers.Primers Primer sequence
dbd60a GGSCTKTTYTTYTTGAATATGGAAATTGGKGAdbd61a TCMCCAATTTCCATATTCAARAARAAMAGSCCdbd62 GATCGAATTCTTTGACATTCATCAAATATTGATdbd63 GATCGGATCCTAATAATCACCTCGCTTTTAGGATdbd64 GATCGAATTCTTGACTTTAATTGTTGGTTAAACdbd65 GATCGGATCCTCTTCCCACTGCTTTCTAAACAdbd66 GATCGAATTCAACGTCAAATACCAATGTTAGCdbd67 GATCGGATCCTACTTGTACTCCCCTCACCAATdbd68 GATCGAATTCGCAAAAATAATCCCATTAATGTTATdbd69 GATCGGATCCTACCTCTCCCTTTACTGTAGCdbd70 GATCGAATTCAATGTCCTCATCAGGCTTAACdbd71 GATCGGATCCTAAATAATCCCCCTTGTATGATGTdbd72 GATCGGATCCTCACTCTCTCTACGCTTTTAATATdbd73 GATCGGATCCTAGTATCGGTATCTATTTAATTATGdbd74 GATCGGATCCTTATTTTTAATGCACTTATTAATATACdbd75 GATCGGATCCTGTTATTTGGAACAAATTTAGTTTAATdbd76 CAAAAACACATTATCCTAAAAGCGTAGAGAGAGTGATATCAAGCdbd77 CGCTTTTAGGATAATGTGTTTTTGdbd78 CAAAAACACATTATCCTAAAAGCGCATTAAAAATACGTAGCGCATdbd87 AGATGCATTTGCACCTATTTCTdbd88 CTTTGGCGAGGGTAACGGTTdbd89 TGGAAAGAGTAGTGCGTATTCdbd90 TTGGTTGGCACTTTTCCTTAATdbd91 ATGCTACAGTTTGAGAAGTTACdbd92 TTAGGTAGTAAATAGCTGTGATTdbd93 GTTGCGCAAGTTGACGAGATdbd94 AACCAGTCGGCGCTGCTTTdbd95 TACTGCCTATTTAGCATTTAAACdbd96 ACTATTGACTAGTCCATCATTAT
a. Degenerate primers: S, G or C; K, G or T; Y, C or T; M, A or C; R, A or G.

492 D. B. Diep et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 483–494
gently with 2¥ SSC for 1 min before baking for 2 h at 80∞C.DNA probes were obtained by PCR using primers dbd87 anddbd88 for probe J, dbd89 and dbd90 for probe A, dbd91 anddbd92 for probe E, dbd93 and dbd94 for probe G and dbd95and dbd96 for probe M. The various primers are listed inTable 2. Each probe was of ª 0.5 kb and specific to the 5¢end of the detected transcript. Probes were labelled with 32Pusing a random labelling kit (Amersham). Hybridization wascarried out as described by Church and Gilbert (1984) andLillehaug et al. (1991).
CAT assay
Growing cell cultures at an OD600 of 0.15 were induced withPlnA (200 ng ml-1), and samples (10–50 ml) were then har-vested at the indicated time points by centrifugation for10 min at 4∞C. Cell pellets were washed once with 8 ml ofice-cold 0.9% NaCl before cell-free extracts were obtained bydisrupting cells with glass beads as described previously byvan de Guchte et al. (1991). The CAT assay was based onliquid scintillation counting (LSC) of CAT reaction products,using a protocol provided by the supplier (Promega). Briefly,CAT reaction was carried out using [3H]-chloramphenicol andn-butyryl-CoA as substrates, and the resulting butylated [3H]-chloramphenicol was separated from the non-reacted sub-strates by xylene extraction; the extracted product was readby LSC. Samples were assayed in duplicate, which yieldedstandard deviations of <10% of the value. Protein concentra-tion was determined by the method described by Bradford(1976) using BSA (Bio-Rad) as standard.
Other techniques
Bacteriocin agar plate assay and induction on agar plateswere performed as described by Diep et al. (1995). For GUS
experiments, cells were grown and harvested, and GUS wasassayed as described previously (Diep et al., 2001).
Acknowledgements
The authors wish to thank Linda Godager for excellent tech-nical assistance. This work was supported by grants from theNorwegian Research Council (NRC).
References
Anderssen, E.L., Diep, D.B., Nes, I.F., Eijsink, V.G., andNissen-Meyer, J. (1998) Antagonistic activity of Lactobacil-lus plantarum C11: two new two-peptide bacteriocins, plan-taricins EF and JK, and the induction factor plantaricin A.Appl Environ Microbiol 64: 2269–2272.
Aukrust, T.W., Brurberg, M.B., and Nes, I.F. (1995) Transfor-mation of Lactobacillus by electroporation. Methods MolBiol 47: 201–208.
Bourret, R.B., Hess, J.F., and Simon, M.I. (1990) Conservedaspartate residues and phosphorylation in signal transduc-tion by the chemotaxis protein CheY. Proc Natl Acad SciUSA 34: 41–45.
Bradford, M.M. (1976) A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal Biochem 72: 248–254.
Brurberg, M.B., Nes, I.F., and Eijsink, V.G. (1997) Phero-mone-induced production of antimicrobial peptides in Lac-tobacillus. Mol Microbiol 26: 347–360.
Church, G.M., and Gilbert, W. (1984) Genomic sequencing.Proc Natl Acad Sci USA 81: 1991–1995.
Delgado, J., Forst, S., Harlocker, S., and Inouye, M. (1993)Identification of a phosphorylation site and functionalanalysis of conserved aspartic acid residues of OmpR, a
Table 3. Plasmids used in this study.
Plasmids Relevant characteristics References
pMG36ea Shuttle vector; containing P32 promoter; EmR van de Guchte et al. (1989)pMS37ca Shuttle vector derived from pMG36e; containing P32 promoter; CmR M. Skaugen (unpublished)pGKV210 Reporter vector containing the promoterless cat gene van der Vossen et al. (1987)p210-A pGKV210 with PA in front of cat This workp210-M/M¢ pGKV210 with PM or P¢M in front of cat This workp210-G/G¢ pGKV210 with PG or P¢G in front of cat This workp210-J/J¢ pGKV210 with PJ or P¢J in front of cat This workp210-E/E¢ pGKV210 with PE or P¢E in front of cat This workp210-AM pGKV210 with PAM in front of cat This workp210-AJ pGKV210 with PAJ in front of cat This workpELS200 Shuttle vector L. Axelsson and M. Skaugen
(unpublished)pJB-GB pELS200 containing the reporter gene gus and the histidine protein kinase gene plnB Diep et al. (2001)pJB37-C pMS37c with plnC cloned just downstream of P32 Diep et al. (2001)pJB37-D pMS37c with plnD cloned just downstream of P32 Diep et al. (2001)pJB36-C pMG36e with plnC cloned just downstream of P32 Diep et al. (2001)pJB36-D pMG36e with plnD cloned just downstream of P32 Diep et al. (2001)pJB37-Cmut pMS37c with plnCmut cloned just downstream of P32 This workpJB37-Dmut pMS37c with plnDmut cloned just downstream of P32 This workpJB36-Cmut pMG36e with plnCmut cloned just downstream of P32 This workpJB36-Dmut pMG36e with plnDmut cloned just downstream of P32 This work
a. pMG36e and its derivatives were used in strain C11, whereas pMS37c and its derivatives were used in strain Lb790.

Pheromone-regulated bacteriocin production 493
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 483–494
transcriptional activator for ompF and ompC in Escherichiacoli. Mol Microbiol 10: 1037–1047.
Diep, D.B., Håvarstein, L.S., Nissen-Meyer, J., and Nes, I.F.(1994) The gene encoding plantaricin A, a bacteriocin fromLactobacillus plantarum C11, is located on the same tran-scription unit as an agr-like regulatory system. Appl Envi-ron Microbiol 60: 160–166.
Diep, D.B., Håvarstein, L.S., and Nes, I.F. (1995) A bacteri-ocin-like peptide induces bacteriocin synthesis in Lactoba-cillus plantarum C11. Mol Microbiol 18: 631–639.
Diep, D.B., Håvarstein, L.S., and Nes, I.F. (1996) Character-ization of the locus responsible for the bacteriocin produc-tion in Lactobacillus plantarum C11. J Bacteriol 178: 4472–4483.
Diep, D.B., Johnsborg, O., Risøen, P.A., and Nes, I.F. (2001)Evidence for dual functionality of the operon plnABCD. theregulation of bacteriocin production in Lactobacillus plan-tarum. Mol Microbiol 41: 633–644.
Dunny, G.M., and Leonard, B.A. (1997) Cell-cell communi-cation in Gram-positive bacteria. Annu Rev Microbiol 51:527–564.
Eijsink, V.G., Brurberg, M.B., Middelhoven, P.H., and Nes,I.F. (1996) Induction of bacteriocin production in Lactoba-cillus sake by a secreted peptide. J Bacteriol 178: 2232–2237.
van de Guchte, M., Kok, J., and Venema, G. (1991) Distance-dependent translational coupling and interference in Lac-tococcus lactis. Mol Gen Genet 227: 65–71.
van de Guchte, M., van der Vossen, J.M., Kok, J., andVenema, G. (1989) Construction of a lactococcal expres-sion vector: expression of hen egg white lysozyme in Lac-tococcus lactis subsp. lactis. Appl Environ Microbiol 55:224–228.
Harmer, T., Wu, M., and Schleif, R. (2001) The role of rigidityin DNA looping-unlooping by AraC. Proc Natl Acad SciUSA 98: 427–431.
Håvarstein, L.S., and Morrison, D.A. (1999) Quorum-sensingand peptide pheromones in streptococcal competence forgenetic transformation. In Cell–Cell Signaling in Bacteria.Dunny, G.M., and Winans, S.C. (eds). Washington, DC.American Society for Microbiology Press, pp. 9–26.
Håvarstein, L.S., Diep, D.B., and Nes, I.F. (1995) A family ofbacteriocin transporters carry out proteolytic processing oftheir substrates concomitant with export. Mol Microbiol 16:229–240.
Higuchi, R. (1990) Recombinant PCR. In PCR Protocols. AGuide to Methods and Applications. San Diego, CA:Academic Press, pp. 177–183.
Igo, M.M., Ninfa, A.J., Stock, J.B., and Silhavy, T.J. (1989)Phosphorylation and dephosphorylation of a bacterial tran-scriptional activator by a transmembrane receptor. GenesDev 3: 1725–1734.
Kleerebezem, M., Quadri, L.E., Kuipers, O.P., and de Vos,W.M. (1997) Quorum sensing by peptide pheromones andtwo-component signal-transduction systems in Gram-positive bacteria. Mol Microbiol 24: 895–904.
Lazazzera, B.A., Kurtser, I.G., McQuade, R.S., andGrossman, A.D. (1999) An autoregulatory circuit affectingpeptide signaling in Bacillus subtilis. J Bacteriol 181: 5193–5200.
Lillehaug, D., Lindqvist, B., and Birkeland, N.K. (1991)
Characterization of phiLC3, a Lactococcus lactis subsp.cremoris temperature bacteriophage with cohesivesingle-stranded DNA ends. Appl Environ Microbiol 57:3206–3211.
Loh, J., Garcia, M., and Stacey, G. (1997) NodV and NodW,a second flavonoid recognition system regulating nod geneexpression in Bradyrhizobium japonicum. J Bacteriol 179:3013–3020.
Makino, K., Shinagawa, H., Amemura, M., Kimura, S.,Nakata, A., and Ishihama, A. (1988) Regulation of thephosphate regulon of Escherichia coli: activation of pstStranscription by PhoB protein in vitro. J Mol Biol 203: 85–95.
Miranda-Rios, J., Sanchez-Pescador, R., Urdea, M., andCovarrubias, A.A. (1987) The complete nucleotidesequence of the glnALG operon of Escherichia coli K12.Nucleic Acids Res 34: 2757–2770.
Nes, I.F., and Eijsink, V.G.H. (1999) Regulation of group IIpeptide bacteriocin synthesis by quorum-sensing mecha-nisms. In Cell–Cell Signaling in Bacteria. Dunny, G.M., andWinans, S.C. (eds). Washington, DC: American Society forMicrobiology Press, pp. 175–192.
Nes, I.F., Diep, D.B., Håvarstein, L.S., Brurberg, M.B.,Eijsink, V., and Holo, H. (1996) Biosynthesis of bacterio-cins in lactic acid bacteria. Antonie van Leeuwenhoek 70:113–128.
Novick, R.P. (1999) Regulation of pathogenicity in Staphylo-coccus aureus by a peptide-based density-sensing mech-anism. In Cell–Cell Signaling in Bacteria. Dunny, G.M., andWinans, S.C. (eds). Washington, DC: American Society forMicrobiology Press, pp. 129–146.
Parkinson, J.S. (1995) Genetic approaches for signalingpathways and proteins. In Two-Component Signal Trans-duction. Hoch, J.A., and Silhavy, T.J. (eds). Washington,DC: American Society for Microbiology Press, pp. 9–23.
Ptashne, M. (1992) The Genetic Switch. Cambridge, MA,USA: Cell Press and Blackwell Scientific Publications.
Risøen, P.A., Håvarstein, L.S., Diep, D.B., and Nes, I.F.(1998) Identification of the DNA-binding sites for tworesponse regulators involved in control of bacteriocin syn-thesis in Lactobacillus plantarum C11. Mol Gen Genet 259:224–232.
Risøen, P.A., Brurberg, M.B., Eijsink, V.G., and Nes, I.F.(2000) Functional analysis of promoters involved in quo-rum sensing-based regulation of bacteriocin production inLactobacillus. Mol Microbiol 37: 619–628.
Risøen, P.A., Johnsborg, O., Diep, D.B., Hamoen, L.,Venema, G., and Nes, I.F. (2001) Regulation of bacteriocinproduction in Lactobacillus plantarum depends on a con-served promoter arrangement with consensus bindingsequence. Mol Gen Genet 265: 198–206.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) Molecu-lar Cloning: A Laboratory Manual, 2nd edn. Cold SpringHarbor, NY: Cold Spring Harbor Laboratory Press.
Sanders, D.A., Gillece-Castro, B.L., Stock, A.M., Burlingame,A.L., and Koshland, D.E., Jr (1989) Identification of the siteof phosphorylation of the chemotaxis response regulatorprotein, CheY. J Biol Chem 264: 21770–21778.
Stock, A.M., Robinson, V.L., and Goudreau, P.N. (2000) Two-component signal transduction. Annu Rev Biochem 69:183–215.

494 D. B. Diep et al.
© 2003 Blackwell Publishing Ltd, Molecular Microbiology, 47, 483–494
Stock, J.B., Ninfa, A.J., and Stock, A.M. (1989) Protein phos-phorylation and regulation of adaptive responses in bacte-ria. Microbiol Rev 53: 450–490.
Stock, J.B., Surette, M.G., Levit, M., and Park, P. (1995) Two-component signal transduction systems: structure-functionrelationships and mechanisms of catalysis. In: Two-Component Signal Transduction. Hoch, J.A. and Silhavy,
T.J. (eds.), Washington, DC: American Society for Microbi-ology Press, pp. 25–51.
van der Vossen, J.M., van der Lelie, D., and Venema, G.(1987) Isolation and characterization of Streptococcus cre-moris Wg2-specific promoters. Appl Environ Microbiol 53:2452–2457.




















