
induced immunosuppression
6
Inhibition of calcium mobilization is an early event in opiate- 2194 0892-6638/91/0005-2194/$01.50.© FASEB induced immunosuppression YOSHITATSU SEI, TODD MCINTYRE, ESTER FRIDE, KOHJI YOSHIMOTO,* PHIL SKOLNICK, AND PRINCE K. ARORA’ Laboratory of Neurosc,ence, National institute of Diabetes and Digestive and Kidney Diseases, and *Laboratory of Cerebral Metabolism, National Institute of Mental Health, National Institutes of Health, Bethesda, Maryland 20892, USA ABSTRACT Morphine administered as a subcutaneous implant inhibits the initial increase in cytoplasmic free- calcium [Ca2] induced by mitogens in mouse splenocytes. This effect was not reproduced by incubation of spleno- cytes with morphine (108_104 M). Analysis of splenocyte subpopulations demonstrates that this effect was manifest in both B and T cells. However, within T cell subpopula- tions, CD4 but not CD8 cells were affected. Adrenalec- tomy abolished this effect of morphine in CD4 T but not CD4-, CD8- spleen cells (most likely Thy 1.2- B cells). Moreover, simultaneous administration of the opiate an- tagonist naltrexone blocked the effect of morphine in CD4, CD8- spleen cells, but not in CD4 T cells. These data in- dicate that the effects of morphine on mitogen-stimulated increase in [Ca2]1 may be mediated through distinct glucocorticoid-dependent and -independent mechanisms. The morphine-induced inhibition of an increase in [Ca2J, in immune cells reported here may be an early event medi- ating opiate-induced immunosuppression. - Sei, Y.; McIntyre, T.; Fride, E.; Yoshimoto, K.; Skolnick, P.; Arora, P. K. Inhibition of calcium mobilization is an early event in opiate-induced immunosuppression. FASEB J. 5: 2194-2199; 1991. Key Words: opiates . opiate. receptors central nervous system calcium mobilization T cells B cells immunosuppression ADMINISTRATION OF OPIATES AND opioid peptides have been reported to effect a wide range of immunological changes (1, 2). For example, a subcutaneous implant of morphine com- monly used to produce tolerance and dependence in rodents (3) elicits a profound and sustained atrophy of the spleen and thymus. This atrophy was manifest within 24 h after implan- tation and was accompanied by a decreased lymphocyte con- tent in spleen nd thymus (4, 5), inhibition of mitogen- stimulated T and B cell responses (6, 7), and altered antigen- specific antibody production (8). Despite these profound im- munosuppressive effects of morphine, the sequence of cellu- lar events that mediate these phenomena has not been deduced. We examined the effects of morphine on an early event associated with activation of immune cells, the eleva- tion of intracellular calcium ions ([Ca2]1) after mitogen- stimulation (9, 10). In the present report, we demonstrate that morphine effects a robust suppression of mitogen-stimulated increases in [Ca2], in CD’I- (but not CD8) T cells and CD4; CD8 spleen cells (most likely Thy 1.2= B cells, see ref 11). This phenomenon was not reproduced by direct addition of mor- phine (108 = l0 M) to lymphocyte cultures. Studies with mice adrenalectomized before morphine implantation indi- cated that the effects of morphine on CD4 T and CD4; CD8 spleen cells are mediated via glucocorticoid-dependent arid -independent mechanisms, respectively. These findings sug- gest that the inhibition of mitogen-stimulated increases in [Ca2]1 may be an early event mediating opiate-induced im- munosuppression. MATERIALS AND METHODS Animals C57BL/6J mice (6-8 wk old) were obtained from Jackson Laboratory (Bar Harbor, Maine). Animals were housed in 32 x 25 x 15 cm transparent plastic cages with wood chip bedding material and maintained in a temperature- and humidity-controlled facility with lights on from 0600 to 1800. Rat chow and water were freely available. All animals were treated according to NIH-AALAC guidelines. Morphine pellet implantation and adrenalectomy Animals were given subcutaneous implants of either a single 75-mg morphine pellet (National Institute on Drug Abuse [NIDA],2 Rockville, Md.) or a single microcrystalline cellu- lose placebo pellet (NIDA) (3). In some experiments, the subcutaneous implantation of either a single 10-mg naltrex- one pellet (NIDA) or a single cholesterol-tristearin placebo pellet (NIDA) was also used. At 24, 48, 72, or 96 hr after im- plantation of either the placebo or morphine pellet, spleen cells were prepared for mitogen-stimulated Ca2 mobiliza- tion assay. In some experiments, animals were subjected to either adrenalectomy (Adx) or sham operation (adrenal glands were touched after laparotomy) and immediately im- planted with morphine or placebo pellets under pentobarbi- tal anesthesia. Corticosterone in mixed trunk blood was measured by a standard radioimmunoassay (Hazieton Bio- technologies, Vienna, Va.). Dexamethasone (DEX) treatment Animals were injected i.p. with 0.1 ml of DEX (10 mg/kg) (Sigma, St. Louis, Mo.) in 60% ethanol/saline or 60% ethanol/saline as vehicle control. Dual-color analysis of fluo-3 loaded cells by flow microfluorometry (FMF) An increase in [Ca2]1 was directly measured in splenocyte subpopulations by changes in the fluorescence intensity of ‘To whom correspondence should be addressed, at: Laboratory of Neuroscience, NLDDK, Bldg. 8, Room 111, National Institutes of Health, Bethesda, MD 20892, USA. 2Abbreviations: NIDA, National Institute on Drug Abuse; Adx, adrenalectomy; DEX, dexamethasone; FMF, flow microfluorome- try; FE, phycoerythrin; Con A, concanavalin A; HPA, hypothalamic- pituitary-adrenal; P1, phosphatidyiinositol; CNS, central nervous sys- tem; DTH, delayed-type hypersensitivity.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of induced immunosuppression

Inhibition of calcium mobilization is an early event in opiate-
2194 0892-6638/91/0005-2194/$01.50.© FASEB
induced immunosuppressionYOSHITATSU SEI, TODD MCINTYRE, ESTER FRIDE, KOHJI YOSHIMOTO,* PHIL SKOLNICK, AND
PRINCE K. ARORA’
Laboratory of Neurosc,ence, National institute of Diabetes and Digestive and Kidney Diseases, and *Laboratory of Cerebral Metabolism,
National Institute of Mental Health, National Institutes of Health, Bethesda, Maryland 20892, USA
ABSTRACT Morphine administered as a subcutaneous
implant inhibits the initial increase in cytoplasmic free-calcium [Ca2] induced by mitogens in mouse splenocytes.This effect was not reproduced by incubation of spleno-cytes with morphine (108_104 M). Analysis of splenocyte
subpopulations demonstrates that this effect was manifestin both B and T cells. However, within T cell subpopula-tions, CD4 but not CD8 cells were affected. Adrenalec-tomy abolished this effect of morphine in CD4 T but notCD4-, CD8- spleen cells (most likely Thy 1.2- B cells).Moreover, simultaneous administration of the opiate an-tagonist naltrexone blocked the effect of morphine in CD4,CD8- spleen cells, but not in CD4 T cells. These data in-dicate that the effects of morphine on mitogen-stimulatedincrease in [Ca2]1 may be mediated through distinctglucocorticoid-dependent and -independent mechanisms.The morphine-induced inhibition of an increase in [Ca2J,in immune cells reported here may be an early event medi-
ating opiate-induced immunosuppression. - Sei, Y.;McIntyre, T.; Fride, E.; Yoshimoto, K.; Skolnick, P.; Arora,P. K. Inhibition of calcium mobilization is an early eventin opiate-induced immunosuppression. FASEB J. 5:2194-2199; 1991.
Key Words: opiates . opiate. receptors central nervous systemcalcium mobilization T cells B cells immunosuppression
ADMINISTRATION OF OPIATES AND opioid peptides have been
reported to effect a wide range of immunological changes (1,2). For example, a subcutaneous implant of morphine com-monly used to produce tolerance and dependence in rodents(3) elicits a profound and sustained atrophy of the spleen andthymus. This atrophy was manifest within 24 h after implan-tation and was accompanied by a decreased lymphocyte con-tent in spleen nd thymus (4, 5), inhibition of mitogen-stimulated T and B cell responses (6, 7), and altered antigen-specific antibody production (8). Despite these profound im-munosuppressive effects of morphine, the sequence of cellu-lar events that mediate these phenomena has not beendeduced. We examined the effects of morphine on an earlyevent associated with activation of immune cells, the eleva-tion of intracellular calcium ions ([Ca2]1) after mitogen-stimulation (9, 10). In the present report, we demonstrate thatmorphine effects a robust suppression of mitogen-stimulatedincreases in [Ca2], in CD’I- (but not CD8) T cells and CD4;CD8 spleen cells (most likely Thy 1.2= B cells, see ref 11). Thisphenomenon was not reproduced by direct addition of mor-phine (108 = l0 M) to lymphocyte cultures. Studies withmice adrenalectomized before morphine implantation indi-cated that the effects of morphine on CD4 T and CD4; CD8spleen cells are mediated via glucocorticoid-dependent arid-independent mechanisms, respectively. These findings sug-
gest that the inhibition of mitogen-stimulated increases in[Ca2]1 may be an early event mediating opiate-induced im-munosuppression.
MATERIALS AND METHODS
Animals
C57BL/6J mice (6-8 wk old) were obtained from JacksonLaboratory (Bar Harbor, Maine). Animals were housed in32 x 25 x 15 cm transparent plastic cages with wood chipbedding material and maintained in a temperature- andhumidity-controlled facility with lights on from 0600 to 1800.Rat chow and water were freely available. All animals weretreated according to NIH-AALAC guidelines.
Morphine pellet implantation and adrenalectomy
Animals were given subcutaneous implants of either a single75-mg morphine pellet (National Institute on Drug Abuse[NIDA],2 Rockville, Md.) or a single microcrystalline cellu-lose placebo pellet (NIDA) (3). In some experiments, thesubcutaneous implantation of either a single 10-mg naltrex-one pellet (NIDA) or a single cholesterol-tristearin placebopellet (NIDA) was also used. At 24, 48, 72, or 96 hr after im-plantation of either the placebo or morphine pellet, spleencells were prepared for mitogen-stimulated Ca2 mobiliza-tion assay. In some experiments, animals were subjected toeither adrenalectomy (Adx) or sham operation (adrenalglands were touched after laparotomy) and immediately im-planted with morphine or placebo pellets under pentobarbi-tal anesthesia. Corticosterone in mixed trunk blood wasmeasured by a standard radioimmunoassay (Hazieton Bio-technologies, Vienna, Va.).
Dexamethasone (DEX) treatment
Animals were injected i.p. with 0.1 ml of DEX (10 mg/kg)
(Sigma, St. Louis, Mo.) in 60% ethanol/saline or 60%ethanol/saline as vehicle control.
Dual-color analysis of fluo-3 loaded cells by flowmicrofluorometry (FMF)
An increase in [Ca2]1 was directly measured in splenocyte
subpopulations by changes in the fluorescence intensity of
‘To whom correspondence should be addressed, at: Laboratoryof Neuroscience, NLDDK, Bldg. 8, Room 111, National Institutesof Health, Bethesda, MD 20892, USA.
2Abbreviations: NIDA, National Institute on Drug Abuse; Adx,adrenalectomy; DEX, dexamethasone; FMF, flow microfluorome-try; FE, phycoerythrin; Con A, concanavalin A; HPA, hypothalamic-pituitary-adrenal; P1, phosphatidyiinositol; CNS, central nervous sys-
tem; DTH, delayed-type hypersensitivity.
30
20
10
0
20
15
10
5
0
40
30
20
10
O()
+
0
TIME (hr)
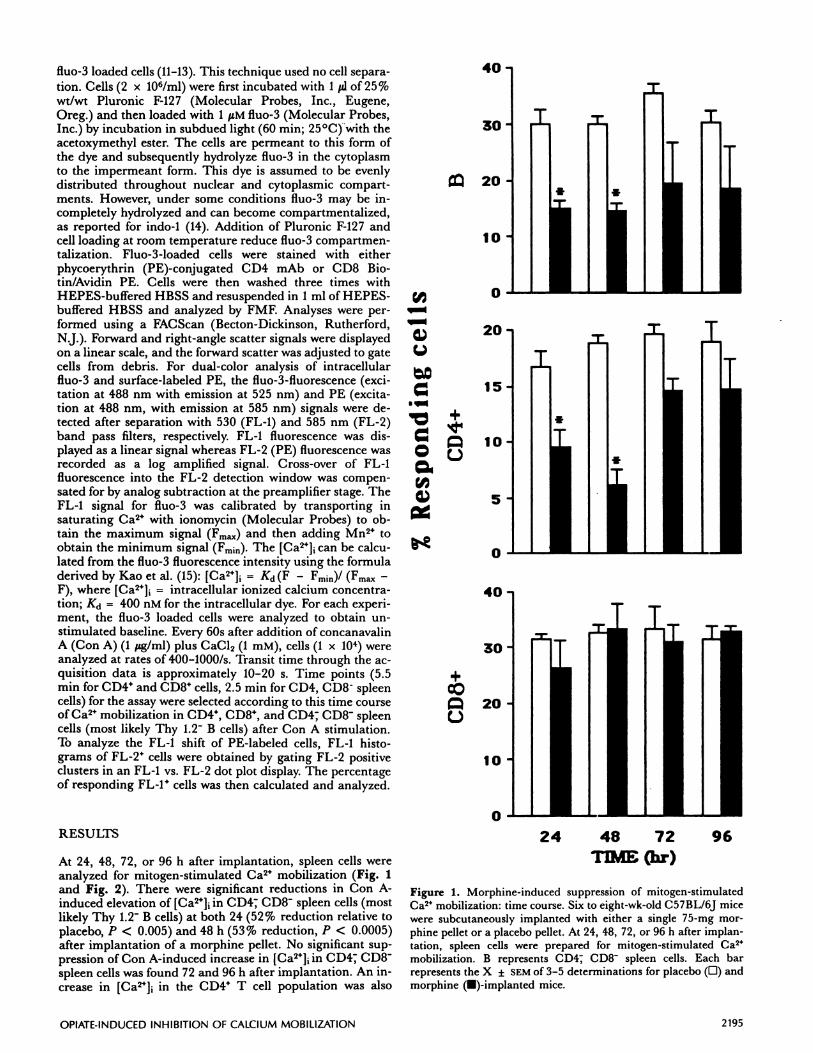
Figure 1. Morphine-induced suppression of mitogen-stimulatedCa2 mobilization: time course. Six to eight-wk-oid C57BL/6J micewere subcutaneously implanted with either a single 75-mg mor-phine pellet or a placebo pellet. At 24, 48, 72, or 96 h after implan-tation, spleen cells were prepared for mitogen-stimulated Ca2
mobilization. B represents CD4; CD8 spleen cells. Each barrepresents the X ± SEM of 3-5 determinations for placebo (D) andmorphine (U)-implanted mice.
24 48 72 96
40
OPIATE-INDUCED INHIBITION OF CALCIUM MOBILIZATION 2195
fluo-3 loaded cells (11-13). This technique used no cell separa-
tion. Cells (2 x 106/ml) were first incubated with 1 pl of 25%wt/wt Pluronic F-127 (Molecular Probes, Inc., Eugene,
Oreg.) and then loaded with 1 M fluo-3 (Molecular Probes,Inc.) by incubation in subdued light (60 mm; 25#{176}C)with theacetoxymethyl ester. The cells are permeant to this form ofthe dye and subsequently hydrolyze fluo-3 in the cytoplasmto the impermeant form. This dye is assumed to be evenlydistributed throughout nuclear and cytoplasmic compart-ments. However, under some conditions fluo-3 may be in-completely hydrolyzed and can become compartmentalized,as reported for indo-1 (14). Addition of Pluronic F-127 andcell loading at room temperature reduce fluo-3 compartmen-talization. Fiuo-3-loaded cells were stained with eitherphycoerythrin (PE)-conjugated CD4 mAb or CD8 Bio-tin/Avidin PE. Cells were then washed three times withHEPES-buffered HBSS and resuspended in I ml of HEPES-buffered HBSS and analyzed by FMF. Analyses were per-formed using a FACScan (Becton-Dickinson, Rutherford,N.J.). Forward and right-angle scatter signals were displayedon a linear scale, and the forward scatter was adjusted to gatecells from debris. For dual-color analysis of intracellularfluo-3 and surface-labeled PE, the fluo-3-fluorescence (exci-tation at 488 nm with emission at 525 nm) and PE (excita-tion at 488 nm, with emission at 585 nm) signals were de-tected after separation with 530 (FL-i) and 585 nm (FL-2)band pass filters, respectively. FL-i fluorescence was dis-played as a linear signal whereas FL-2 (PE) fluorescence wasrecorded as a log amplified signal. Cross-over of FL-ifluorescence into the FL-2 detection window was compen-sated for by analog subtraction at the preamplifier stage. TheFL-I signal for fluo-3 was calibrated by transporting insaturating Ca2 with ionomycin (Molecular Probes) to ob-tain the maximum signal (Fmax) and then adding Mn2 toobtain the minimum signal (Fmjn). The [Ca2]1 can be calcu-lated from the fluo-3 fluorescence intensity using the formuladerived by Kao et al. (15): [Ca2]1 = Kd(F - Fmin)/ (Fmax -
F), where [Ca2], intracellular ionized calcium concentra-tion; Kd = 400 nM for the intracellular dye. For each experi-ment, the fluo-3 loaded cells were analyzed to obtain Un-stimulated baseline. Every 60s after addition of concanavalinA (Con A) (1 ‘eg/mi) plus CaCl2 (1 mM), cells (1 x 104) wereanalyzed at rates of 400-1000/s. Transit time through the ac-quisition data is approximately 10-20 s. Time points (5.5mm for CD4 and CD8 cells, 2.5 mm for CD4, CD8 spleencells) for the assay were selected according to this time courseof Ca2 mobilization in CD4, CD8, and CD4; CD8- spleencells (most likely Thy 1.2 B cells) after Con A stimulation.To analyze the FL-i shift of PE-labeled cells, FL-i histo-grams of FL-2 cells were obtained by gating FL-2 positiveclusters in an FL-i vs. FL-2 dot plot display. The percentageof responding FL-P cells was then calculated and analyzed.
RESULTS
At 24, 48, 72, or 96 h after implantation, spleen cells wereanalyzed for mitogen-stimulated Ca2 mobilization (Fig. 1and Fig. 2). There were significant reductions in Con A-induced elevation of [Ca2]1 in CD47 CD8 spleen cells (mostlikely Thy 1.2 B cells) at both 24 (52% reduction relative toplacebo, P < 0.005) and 48 h (53% reduction, P < 0.0005)after implantation of a morphine pellet. No significant sup-pression of Con A-induced increase in [Ca2]1 in CD4T CD8spleen cells was found 72 and 96 h after implantation. An in-crease in [Ca2J in the CD4 T cell population was also
0
300
a
0
75U,
a,C-,
0
C-U,.0E
zU’
>
‘V
a’
75 J1
ctannel # Danne1 #
FluorescenceIntensity
T
-4
-4
C)V
0
U,C)
40
30
20
10
0
B CD4+
2196 Vol. 5 May 1991 The FASEB Journal SEI ET AL.
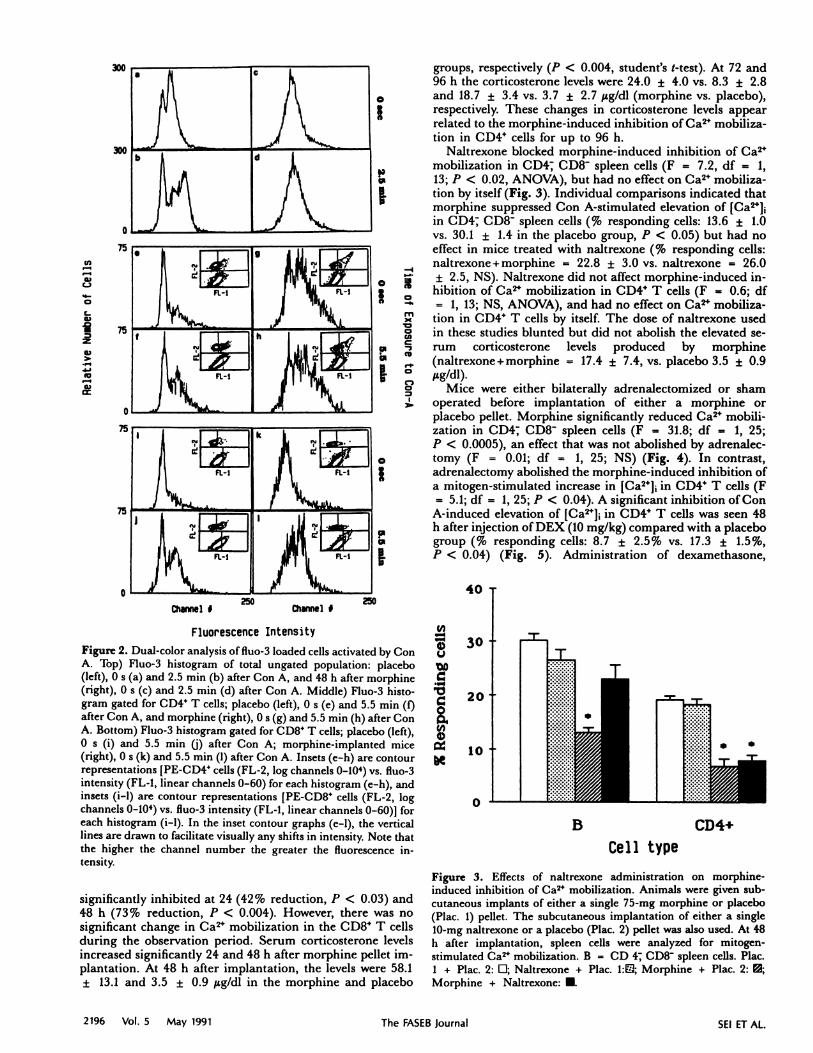
Figure 2. Dual-color analysis of fluo-3 loaded cells activated by ConA. Top) Fluo-3 histogram of total ungated population: placebo(left), 0 s (a) and 2.5 mm (b) after Con A, and 48 h after morphine
(right), 0 s (c) and 2.5 mm (d) after Con A. Middle) Fluo-3 histo-gram gated for CD4 T cells; placebo (left), 0 s (e) and 5.5 mm (f)after Con A, and morphine (right), 0 s (g) and 5.5 mm (h) after ConA. Bottom) Fluo-3 histogram gated for CD8 T cells; placebo (left),0 s (i) and 5.5 mm U) after Con A; morphine-implanted mice(right), 0 s (k) and 5.5 mm (1) after Con A. Insets (c-h) are contourrepresentations [PE-CD4 cells (FL-2, log channels 0-104) vs. fluo-3intensity (FL-l, linear channels 0-60) for each histogram (c-h), andinsets (i-i) are contour representations [PE-CD8 cells (FL-2, logchannels 0-104) vs. fluo-3 intensity (FL-I, linear channels 0-60)1 foreach histogram (i-i). In the inset contour graphs (c-i), the verticallines are drawn to facilitate visually any shifts in intensity. Note thatthe higher the channel number the greater the fluorescence in-tensity.
significantly inhibited at 24 (42% reduction, P < 0.03) and48 h (73% reduction, P < 0.004). However, there was nosignificant change in Ca2 mobilization in the CD8 T cellsduring the observation period. Serum corticosterone levelsincreased significantly 24 and 48 h after morphine pellet im-plantation. At 48 h after implantation, the levels were 58.1
± 13.1 and 3.5 ± 0.9 tg/dl in the morphine and placebo
groups, respectively (P < 0.004, student’s 1-test). At 72 and96 h the corticosterone levels were 24.0 ± 4.0 vs. 8.3 ± 2.8and 18.7 ± 3.4 vs. 3.7 ± 2.7 JLg/dl (morphine vs. placebo),respectively. These changes in corticosterone levels appearrelated to the morphine-induced inhibition of Ca2 mobiliza-
tion in CD44 cells for up to 96 h.Naltrexone blocked morphine-induced inhibition of Ca2
mobilization in CD4; CD8- spleen cells (F = 7.2, df = 1,13; P < 0.02, ANOVA), but had no effect on Ca24 mobiliza-tion by itself (Fig. 3). Individual comparisons indicated thatmorphine suppressed Con A-stimulated elevation of [Ca24]1in CD4; CD8- spleen cells (% responding cells: 13.6 ± 1.0vs. 30.1 ± 1.4 in the placebo group, P < 0.05) but had noeffect in mice treated with naltrexone (% responding cells:naltrexone+morphine = 22.8 ± 3.0 vs. naltrexone = 26.0
o ± 2.5, NS). Naltrexone did not affect morphine-induced in-hibition of Ca2 mobilization in CD4 T cells (F = 0.6; df
= 1, 13; NS, ANOVA), and had no effect on Ca2 mobiliza-‘ tion in CD4 T cells by itself. The dose of naltrexone used
in these studies blunted but did not abolish the elevated se-rum corticosterone levels produced by morphine(naltrexone+morphine = 17.4 ± 7.4, vs. placebo 3.5 ± 0.9
#{176} eg/dl).Mice were either bilaterally adrenalectomized or sham
operated before implantation of either a morphine or
placebo pellet. Morphine significantly reduced Ca2 mobili-zation in CD4; CD8 spleen cells (F = 31.8; df = 1, 25;P < 0.0005), an effect that was not abolished by adrenalec-tomy (F = 0.01; df = 1, 25; NS) (Fig. 4). In contrast,adrenalectomy abolished the morphine-induced inhibition ofa mitogen-stimulated increase in [Ca24] in CD4 T cells (F
= 5.1; df = 1, 25; P < 0.04). A significant inhibition of ConA-induced elevation of [Ca2]1 in CD4 T cells was seen 48h after injection of DEX (10 mg/kg) compared with a placebogroup (% responding cells: 8.7 ± 2.5% vs. 17.3 ± 1.5%,
P < 0.04) (Fig. 5). Administration of dexamethasone,
Celltype
Figure 3. Effects of naltrexone administration on morphine-induced inhibition of Ca2 mobilization. Animals were given sub-cutaneous implants of either a single 75-mg morphine or placebo(Piac. 1) pellet. The subcutaneous implantation of either a single10-mg naltrexone or a placebo (Plac. 2) pellet was also used. At 48h after implantation, spleen cells were analyzed for mitogen-stimulated Ca2 mobilization. B = CD 4; CD8 spleen cells. Plac.1 + Piac. 2: D; Naltrexone + Piac. 1:Z; Morphine + Plac. 2: ;
Morphine + Naitrexone: #{149}.

40-
20
however, did not affect mitogen-stimulated increase in[Ca2], in either CD8 T or CD4 CD8 spleen cells.
Subcutaneous implantation of rodents with a single mor-phine pellet has been used extensively as a model to studythe effects of morphine in vivo, as it rapidly induces toleranceand physical dependence (3). This procedure effected arobust inhibition of Con A-stimulated increases in [Ca24]1into CD4, CD8 spleen cells (most likely Thy 1.2- B cells) andCD4 (L3T44) splenic T cells within 24 h after implantation.This inhibition was fully manifest in both CD4; CD8 spleencells and CD4 T cells by 48 h, and started to return to con-trol values by 72 h after implantation. In contrast, Con A-stimulated increases in [Ca24]1 into CD84 T cells wereunaffected by morphine, indicating a differential sensitivityof CD44 and CD84 T cells which may lead to a functionalreversal of the CD44/CD84 ratio in vivo. This functionalreversal may cause suppression of mitogen-stimulated T cellresponses observed in morphine-treated mice provided cellu-lar events subsequent to protein kinase-C activation are notdifferentially affected. As high-affinity, stereospecific opiatebinding sites are present on immune cells (1, 16), experi-ments were performed to determine whether similar effectscould be obtained by incubating lymphocytes with morphinein vitro. Exposure of lymphocytes in culture to morphine(l08_104 M) for up to 24 h did not affect mitogen-stimulated increases in [Ca24]1 into either CD44 T or CD4;CD8 spleen cells (data not shown). As a 24-h in vivo ex-posure resulted in decreased Ca24 mobilization (Fig. 1), theseresults suggest that the effects of morphine on Ca24 mobiliza-tion in lymphocytes are not mediated through direct actionon opiate receptors in these cells (1).
Acute administration of morphine to rodents has beenreported to produce activation of the hypothalamic-pituitary-adrenal (HPA) axis resulting in a profound in-crease in serum corticosterone levels. (17-19). As a significantincrease in serum corticosterone levels was observed 24 and48 h after morphine pellet implantation (see Results), studieswere designed to determine whether these elevations wereresponsible for the morphine-induced inhibition of Ca24mobilization. Animals were either adrenalectomized orsham operated before morphine or placebo pellet implanta-tion. Adrenalectomy abolished the effect of morphine to in-hibit mitogen-induced elevations of [Ca24]t in CD44 T cellsbut not in CD4; CD8- spleen cells. The ability of dcx-amethasone (Fig. 5) to mimic the effect of morphine in CD44but not CD84 T cells or CD4 CD8 spleen cells supports thehypothesis that this opiate affects Ca2 mobilization in CD44T cells via an activation of the HPA axis. Because aglucocorticoid-inducible protein, lipomodulin, can inhibitphospholipase-C activity (20), an inhibition of phos-phatidylinositol (PT) hydrolysis may be responsible forglucocorticoid-mediated inhibition of Ca24 mobilization inCD4 T cells. The present findings suggest that the effects ofmorphine on inhibition of Ca24 mobilization are mediatedthrough glucocorticoid-dependent and -independent mechan-isms in CD44 T and CD4; CD8 spleen cells, respectively. Asmorphine has been reported to suppress rather than activatethe HPA axis in humans (21, 22), the glucocorticoid-dependenteffects of morphine may not be readly extrapolated to humans.Nonetheless, the presence of glucorticoid-independent effectsof morphine on CD4; CD8 spleen cell (most likely Thy 1.2B cells) functions is consistent with the increased incidenceof infections associated with illicit use of opiates (23-26).
+I-
10
0
+
0
Figure 4. Inhibition of Con A-stimulated elevation of [Ca’]1 inCD4 , CD8 spleen cells (B) and CD4 T cells: effects of adrenalec-tomy. Mice were either bilaterally adrenalectomized or sham oper-ated (adrenal glands were touched after iaparotomy) before implan-tation of either a morphine or placebo pellet. Piac. + Sham:placebo pellet implanted in sham-operated mice, Piac. + Adx:placebo pellet implanted in adrenalectomized mice, Mor. + Sham:morphine pellet implanted in sham-operated mice, and Mor. +
Adx: morphine pellet implanted in adrenalectomized mice.
Plac. Plac. Mot. Mc.+ + + +
huIk Ath
1REAThENT GROUPS
OPIATE-INDUCED INHIBITION OF CALCIUM MOBILIZATION 2197
30
10
0
20
15
5
0
40-
30-
20 -
10 -
0-
DISCUSSION

U,-4-4C)U
C‘-4
C0
U,C)
10
0#{149}
B CD4+
40 T REFERENCES
2198 Vol. 5 May 1991 The FASEB Journal SEI ET AL.
Cell typeFigure 5. Effects of DEX treatment of Ca2 mobilization. Micewere injected i.p. with 0.1 ml of DEX(lO mg/kg) in 60% ethanol/sa-line or 60% ethanol/saline as vehicle placebo control. At 48 h afterinjection, mitogen-stimulated Ca’ mobilization was examined inCD4 , CD8 spleen cells (B), CD4 and CD8 T cells. Each barrepresents the X± SEM of six determinations for placebo- (0) and
DEX-treated () mice.
Incubation of lymphocytes with morphine did not affectCa24 mobilization in either CD4; CD8 spleen cells or CD44T cells. Administration of the opiate antagonist, naltrexone,blocked the morphine-induced inhibition of Ca2 mobiliza-tion in CD4; CD8- spleen cells. These findings indicate thatin vivo effects are mediated through opiate receptors, possi-bly those located in the central nervous system (CNS) (27).Although morphine-induced activation of the HPA axis mayalso be mediated via opiate receptors in the CNS, naltrexonehad little or no effect on morphine-induced inhibition of Ca24mobilization in CD44 T cells. The dose of naltrexone usedin these studies was sufficient to blunt but not abolish theelevated serum corticosterone levels produced by morphine,and was sufficient to minimize neurological signs (hyperac-tivity, jumping, etc.) of morphine dependence evaluated bynaloxone challenge (data not shown). The reported sensitiv-ity of murine immune cells to glucocorticoids suggests thatthe modest residual elevations in circulating corticosteronemay still be sufficient to suppress Ca24 mobilization on CD44T cells. Administration of higher doses (a second 10-mgnaltrexone pellet) resulted in an unacceptably high mortalityin morphine-implanted mice, which precluded a moredetailed examination of this phenomenon. In a recent paper,Bryant and Roudebush (28) demonstrated the reversal ofmorphine effects with a single naltrexone pellet on delayed-type hypersensitivity (DTH) and in vivo lymphocyteproliferation. However, these investigators did not examinethe effects of a higher dose (i.e., two pellets) of naltrexone.
Elevation of [Ca2]1 has been proposed to be one of theearly events in a series of intracellular processes which cul-minate in lymphocyte activation and proliferation (9, 10).Our findings suggest that the morphine-induced inhibitionof Ca24 mobilization in immune cells may be one of the earlyevents mediating opiate-induced immune suppression.
1. Sibinga, N. E. S., and Goldstein, A. (1988) Opioid peptides andopioid receptors in cells of the immune system. Annu. Rev. Im-munol. 6, 219-249
2. Yahya, M. D., and Watson, R. R. (1987) Immunomodulation bymorphine and marijuana. Life Sci. 41, 2503=2510
3. Gibson, R. D., and Tingstad,J. E. (1970) Formulation of a mor-phine implantation pellet suitable for tolerance-physical depen-dence studies in mice. j Pharm. Sci. 59, 426=427
4. Arora, P. K., Fride, E., Petitto, J., Waggie, K., and Skolnick, P.(1990) Morphine-induced immune alterations in vivo. Cell. Im-munol. 126, 343-353
5. Sei, Y., Yoshimoto, K., McIntyre, T., Skolnick, P., and Arora,P. K. (1991) Morphine-induced thymic hypopiasia isglucocorticoid-dependent. j Immunol. 146, 194-198
6. Bryant, H. U., Bernton, E. W., and Holaday, J. W. (1987) Im-munosuppressive effects of chronic morphine treatment in mice.Life Sci. 41, 1731-1738
7. Bryant, H. U., Berton, E. W., and Holaday, J. W. (1988) Mor-CD8+ phine pellet-induced immunomodulation in mice: temporal
relationships. j PharmacoL Exp. Ther. 245, 913-9208. Weber, R. J., Ikejiri, B., Rice, K. C., Pert, A., and Hagan,
A. A. (1987) Opiate receptor mediated regulation of the im-mune response in vivo. Nati. Inst. Drug Abuse Res. Monogr. Set. 76,341=348
9. Linch, D. C., Wallace, D. L., and O’Flynn, K. (1987) Signaltransduction in human T lymphocytes. Immunol. Rev. 95,137=159
10. Geifand, E. W., Mills, G. B., Cheung, R. K., Lee,J. W. W., andS. Grinstein. (1987) Transmembrane ion fluxes during activa-tion of human T lymphocytes: role of Ca’, Nat/H exchangeand phospholipid turnover. Immunol. Rev. 95, 59-87
11. Sei, Y., and Arora, P. K. (1991) Quantitative analysis of calcium(Ca’) mobilization after stimulation with mitogens or anti-CD3 antibodies: simultaneous fluo-3 and immunofluorescenceflow cytometry. j ImmunoL Method. In press.
12. Tsien, R. Y. (1989) Fluorescence microscopy of living cells inculture. In Methods in Cell Biology (Taylor, D. L., and Wang, Y.,eds) Vol. 30, pp. 127-156, Academic, New York
13. Minta, A., Kao, J. P. Y., and Tsien, R. Y. (1989) Fluorescent in-dicators for cytosolic calcium based on rhodamine and fluores-cein chromophores. j BioL Chem. 264, 8171=8178
14. Luckhoff, A. (1986) Measuring cytosolic free calcium concen-tration in endothelial cells with indo-1: the pitfall of using theratio of two fluorescence intensities recorded at differentwavelengths. Cell Calcium. 7, 233-248
15. Kao, J. P. Y., Harootunian, A. T., and Tsien, R. Y. (1989) Pho-tochemically generated cytosoiic calcium pulses and their detec-tion by fluo-3. j Biol. Chem. 264, 8179=8184
16. Carr, D. J., Kim, C. H., deCosta, B., Jacobson, A. E., Rice,K. C., and Blalock, J. E. (1988) Evidence for a delta-class opioidreceptor on cells of the immune system. Cell. Immunol. 116,44=51
17. George, R., and Way, E. L. (1955) Studies on the mechanismof pituitary-adrenal activation by morphine. Br. j Pharmacol.10, 260-264
18. Gibson, A., Ginsburg, M., Hall, M., and Hart, S. L. (1979) Theeffects of opiate receptor agonists and antagonists on the stress-induced secretion of corticosterone in mice. br. j Pharmacol. 65,139=146
19. Haracz, J. L., Bloom, A. S., Wang, R. I. H., and Tseng, L. F.(1981) Effect of intraventricular beta-endorphine and morphineon hypothalamic-pituitary-adrenal activity and the release ofpituitary beta-endorphin. Neuroendocrinology 33, 170=175
20. Blackweil, G. J., Carnuccio, R., DiRosa, M., Flower, R. J.,Panente, L., and Persico, P. (1980) Macrocortin, a polypeptidecausing the anti-phospholipase effect of glucocorticoid. Nature
(London) 287, 147-14821. Deiitala, G., Mannelli, M., Giusti, M., Maggi, M., De Feo,
M. L., Giordano, G., Giusti, G., and Serio, M. (1984) Relation

OPIATE-INDUCED INHIBITION OF CALCIUM MOBILIZATION 2199
of opiates to hormonal secretion in man. In OpioidModualtion ofEndocrine Function (Delitala, G., Motta, M., and Serio, M., eds)pp. 65-79, Raven, New York
22. Kreek, M. J., Wardiaw, S. L., Hartman, N., Raghunath, J.,Friedman,J., Schneider, B., and Frantz, A. G. (1983) Circadianrhythms and levels of beta-endorphin, ACTH, and cortisol dur-
ing chronic methadone maintenance treatment in humans. LfeSci. 33 (Suppl. 1) 409-411
23. Louria, D. B., Hensle, T, and Rose, J. (1957) The major medi-cal complications of heroin addiction. Ann. Intern. Med. 67, 1-22
24. Briggs, J. H., Mckerron, C. C., Souhami, R. L., Taylor, D. J.E., and Andrews, H. (1967) Severe systemic infections com-plicating “main line” heroin addiction. Lancet 2, 1227=1231
25. Cherubin, C. E. (1971) Infectious disease problems of narcotic
addicts. Arch. Intern. Med. 128, 309-31326. Tubaro, E., Borelli, G., Crose, C., Cavallo, C., and Santiangeli,
C. (1983) Effect of morphine on resistance to infection. J. Infect.
Dis. 148, 656-66627. Weber, R. J., and Pert, A. (1989) The periaqueductal gray mat-
ter mediates opiate-induced immunosuppression. Science 245,188=190
28. Bryant, H. U., and Roudebush, R. E. (1990) Suppressive effectsof morphine pellet implants on in vitro parameters of immunefunction. J. PharmacoL Exp. Ther. 255, 410-414
Received for publication November 15, 1990.Accepted for publication March 15, 1991.




















