
Individual differences in the ability to identify, select and use appropriate frames of reference...
17
INDIVIDUAL DIFFERENCES IN THE ABILITY TO IDENTIFY, SELECT AND USE APPROPRIATE FRAMES OF REFERENCE FOR PERCEPTUO-MOTOR CONTROL B. ISABLEU, a,b * T. OHLMANN, c J. CREMIEUX, d N. VUILLERME, a,e B. AMBLARD f AND M. A. GRESTY g a University Joseph Fourier, TIMC-IMAG Laboratory, UMR CNRS 5525, Grenoble, F-38706, France b University Paris-Sud, UR CIAMS-Motor Control & Perception team, Orsay, F-91405, France c University Pierre-Mendès-France, Neurocognition and Psychology Laboratory, UMR 5105, BP 47, Grenoble, F-38040, France d University du Sud Toulon-Var, Handibio EA 4322, La Garde, F-83957, France e CIC-IT 805, INSERM/AP-HP, Hôpital Raymond Poincaré, EA 4497, Garches, France f CNRS, Institute of Physiological and Cognitive Neurosciences, Mar- seille, F-13402, France g Imperial College London, MRC Spatial Disorientation Group, Charing Cross Hospital, U.K-W68 RF, London, UK Abstract—The causes of the interindividual differences (IDs) in how we perceive and control spatial orientation are poorly understood. Here, we propose that IDs partly reflect preferred modes of spatial referencing and that these preferences or “styles” are maintained from the level of spatial perception to that of motor control. Two groups of experimental subjects, one with high visual field dependency (FD) and one with marked visual field independency (FI) were identified by the Rod and Frame Test, which identifies relative dependency on a visual frame of reference (VFoR). FD and FI subjects were tasked with standing still in conditions of increasing postural difficulty while visual cues of self-orientation (a visual frame tilted in roll) and self-motion (in stroboscopic illumination) were varied and in darkness to assess visual dependency. Postural stability, overall body orientation and modes of seg- mental stabilization relative to either external (space) or ego- centric (adjacent segments) frames of reference in the roll plane were analysed. We hypothesized that a moderate chal- lenge to balance should enhance subjects’ reliance on VFoR, particularly in FD subjects, whereas a substantial challenge should constrain subjects to use a somatic-vestibular based FoR to prevent falling in which case IDs would vanish. The results showed that with increasing difficulty, FD subjects became more unstable and more disoriented shown by larger effects of the tilted visual frame on posture. Furthermore, their preference to coalign body/VFoR coordinate systems lead to greater fixation of the head–trunk articulation and stabilization of the hip in space, whereas the head and trunk remained more stabilized in space with the hip fixed on the leg in FI subjects. These results show that FD subjects have difficulties at identifying and/or adopting a more appropriate FoR based on proprioceptive and vestibular cues to regu- late the coalignment of posturo/exocentric FoRs. The FI sub- jects’ resistance in the face of altered VFoR and balance challenge resides in their greater ability to coordinate move- ment by coaligning body axes with more appropriate FoRs (provided by proprioceptive and vestibular co-variance). © 2010 IBRO. Published by Elsevier Ltd. All rights reserved. Key words: interindividual differences, postural control, frames of reference, static and dynamic visual cues. The ability to adapt sensory–motor control to challenging situations by selection and weighting of alternative frames of reference is considered as one of the main issues for postural control (Carver et al., 2006; Maurer et al., 2006; McGuire and Sabes, 2009). Recent models of multisen- sory combination based on neurophysiological (Brandt et al., 1998), perceptual (Ernst and Bulthoff, 2004) and be- havioural evidence (Oie et al., 2002) propose that adapta- tion to environmental changes depends upon sensory re- weighting to optimize the relevance of individual sensory inputs (maximize the reliability of our estimates). The re- weighting is determined by maximum likelihood estimation (MLE) so that the brain appears as a near-optimal Bayes- ian estimator of object properties (Ernst and Banks, 2002). Static, tilted and moving visual frames of reference can induce postural tilt or sway in a standing observer who is otherwise passive. These visual cues to spatial orientation conflict with information about that orientation and motion of body segments which are perceived in an ‘egocentric space’ via a kinematic proprioceptive chain (Roll and Roll, 1988) linking the eye to the foot, paralleled by a kinetic chain of force and load-related signals (Dietz, 1993; Horst- mann and Dietz, 1990; Maurer and Peterka, 2005) stem- ming from the joints and terminating in the receptors of the plantar sole which relate the body to the external world (Ting and Macpherson, 2004). A relative assessment of visual versus somatosensory cues to orientation and bal- ance and weighting of their inputs is necessary so that muscular commands can regulate efficiently the coaligne- ment of the main axis of body FOR with the most appro- priate exocentric frames of reference (e.g., visual direction, gravito-inertial force ‘Gif’ direction, foot support). Of pertinence, numerous studies have observed large interindividual differences (IDs) in postural responses to *Correspondence to: B. Isableu, Universite Paris Sud, UR CIAMS- Motor Control & Perception Team, UFRSTAPS. Bt 335, 91405 Orsay, France. Tel: 33-1-69-15-78-82; fax: 33-1-69-15-62-22. E-mail address: [email protected] (B. Isableu). Abbreviations: AI, anchoring index; FD, field dependency; FI, field independency; FOR, frame of reference; HSTS, head stabilization on the trunk strategy; IDs, interindividual differences; MR, monopodal Romberg; NR, normal Romberg; RFT, rod and frame test; SR, sharp- ened Romberg; TC, tilted frame continuous vision; TS, tilted frame strobe vision; VC, vertical frame continuous vision; VFOR, visual frame of reference; VS, vertical frame strobe vision. Neuroscience 169 (2010) 1199 –1215 0306-4522/10 $ - see front matter © 2010 IBRO. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.neuroscience.2010.05.072 1199
Transcript of Individual differences in the ability to identify, select and use appropriate frames of reference...

IAP
BNa
5b
Oc
Ld
Fe
Gf
sg
C
Aium“tomRatdtwPmcplpsFrbet
*MFEAitResf
Neuroscience 169 (2010) 1199–1215
0d
NDIVIDUAL DIFFERENCES IN THE ABILITY TO IDENTIFY, SELECTND USE APPROPRIATE FRAMES OF REFERENCE FOR
ERCEPTUO-MOTOR CONTROLlsrldFljcm(©
Kf
TsopMsahtwiw(i
iocos1cmmp(vammpg
. ISABLEU,a,b* T. OHLMANN,c J. CREMIEUX,d
. VUILLERME,a,e B. AMBLARDf AND M. A. GRESTYg
University Joseph Fourier, TIMC-IMAG Laboratory, UMR CNRS525, Grenoble, F-38706, France
University Paris-Sud, UR CIAMS-Motor Control & Perception team,rsay, F-91405, France
University Pierre-Mendès-France, Neurocognition and Psychologyaboratory, UMR 5105, BP 47, Grenoble, F-38040, France
University du Sud Toulon-Var, Handibio EA 4322, La Garde,-83957, France
CIC-IT 805, INSERM/AP-HP, Hôpital Raymond Poincaré, EA 4497,arches, France
CNRS, Institute of Physiological and Cognitive Neurosciences, Mar-eille, F-13402, France
Imperial College London, MRC Spatial Disorientation Group, Charingross Hospital, U.K-W68 RF, London, UK
bstract—The causes of the interindividual differences (IDs)n how we perceive and control spatial orientation are poorlynderstood. Here, we propose that IDs partly reflect preferredodes of spatial referencing and that these preferences or
styles” are maintained from the level of spatial perception tohat of motor control. Two groups of experimental subjects,ne with high visual field dependency (FD) and one witharked visual field independency (FI) were identified by theod and Frame Test, which identifies relative dependency onvisual frame of reference (VFoR). FD and FI subjects were
asked with standing still in conditions of increasing posturalifficulty while visual cues of self-orientation (a visual frameilted in roll) and self-motion (in stroboscopic illumination)ere varied and in darkness to assess visual dependency.ostural stability, overall body orientation and modes of seg-ental stabilization relative to either external (space) or ego-
entric (adjacent segments) frames of reference in the rolllane were analysed. We hypothesized that a moderate chal-
enge to balance should enhance subjects’ reliance on VFoR,articularly in FD subjects, whereas a substantial challengehould constrain subjects to use a somatic-vestibular basedoR to prevent falling in which case IDs would vanish. Theesults showed that with increasing difficulty, FD subjectsecame more unstable and more disoriented shown by largerffects of the tilted visual frame on posture. Furthermore,heir preference to coalign body/VFoR coordinate systems
Correspondence to: B. Isableu, Universite Paris Sud, UR CIAMS-otor Control & Perception Team, UFRSTAPS. Bt 335, 91405 Orsay,rance. Tel: �33-1-69-15-78-82; fax: �33-1-69-15-62-22.-mail address: [email protected] (B. Isableu).bbreviations: AI, anchoring index; FD, field dependency; FI, field
ndependency; FOR, frame of reference; HSTS, head stabilization onhe trunk strategy; IDs, interindividual differences; MR, monopodalomberg; NR, normal Romberg; RFT, rod and frame test; SR, sharp-ned Romberg; TC, tilted frame continuous vision; TS, tilted frame
itrobe vision; VC, vertical frame continuous vision; VFOR, visualrame of reference; VS, vertical frame strobe vision.
306-4522/10 $ - see front matter © 2010 IBRO. Published by Elsevier Ltd. All rightoi:10.1016/j.neuroscience.2010.05.072
1199
ead to greater fixation of the head–trunk articulation andtabilization of the hip in space, whereas the head and trunkemained more stabilized in space with the hip fixed on theeg in FI subjects. These results show that FD subjects haveifficulties at identifying and/or adopting a more appropriateoR based on proprioceptive and vestibular cues to regu-
ate the coalignment of posturo/exocentric FoRs. The FI sub-ects’ resistance in the face of altered VFoR and balancehallenge resides in their greater ability to coordinate move-ent by coaligning body axes with more appropriate FoRs
provided by proprioceptive and vestibular co-variance).2010 IBRO. Published by Elsevier Ltd. All rights reserved.
ey words: interindividual differences, postural control,rames of reference, static and dynamic visual cues.
he ability to adapt sensory–motor control to challengingituations by selection and weighting of alternative framesf reference is considered as one of the main issues forostural control (Carver et al., 2006; Maurer et al., 2006;cGuire and Sabes, 2009). Recent models of multisen-
ory combination based on neurophysiological (Brandt etl., 1998), perceptual (Ernst and Bulthoff, 2004) and be-avioural evidence (Oie et al., 2002) propose that adapta-ion to environmental changes depends upon sensory re-eighting to optimize the relevance of individual sensory
nputs (maximize the reliability of our estimates). The re-eighting is determined by maximum likelihood estimation
MLE) so that the brain appears as a near-optimal Bayes-an estimator of object properties (Ernst and Banks, 2002).
Static, tilted and moving visual frames of reference cannduce postural tilt or sway in a standing observer who istherwise passive. These visual cues to spatial orientationonflict with information about that orientation and motionf body segments which are perceived in an ‘egocentricpace’ via a kinematic proprioceptive chain (Roll and Roll,988) linking the eye to the foot, paralleled by a kinetichain of force and load-related signals (Dietz, 1993; Horst-ann and Dietz, 1990; Maurer and Peterka, 2005) stem-ing from the joints and terminating in the receptors of thelantar sole which relate the body to the external worldTing and Macpherson, 2004). A relative assessment ofisual versus somatosensory cues to orientation and bal-nce and weighting of their inputs is necessary so thatuscular commands can regulate efficiently the coaligne-ent of the main axis of body FOR with the most appro-riate exocentric frames of reference (e.g., visual direction,ravito-inertial force ‘Gif’ direction, foot support).
Of pertinence, numerous studies have observed large
nterindividual differences (IDs) in postural responses tos reserved.
pvscf1aatdLt2wFt(c(tfAedsmmi1dvso
tehmM1m((ctta(
p(stpftdto
ppa
i
Md
t9vvgso(msii1da“aosptwmwtm
ma
B. Isableu et al. / Neuroscience 169 (2010) 1199–12151200
erturbation of the gravitational frame of reference in eitherisual or somatosensory modalities. Preferred modes ofpatial referencing have been identified in both the visualontribution (Chiari et al., 2000; Collins et al., 1995; Ehren-ried et al., 2003; Guerraz et al., 2001a; Isableu et al.,997; Lambrey and Berthoz, 2003; Streepey et al., 2007b)nd somatosensory contribution to postural control (Is-bleu and Vuillerme, 2006; Kluzik et al., 2005). A tradi-ional interpretation of IDs is that subjects vary in theegree to which they weight sensory inputs (Peterka andoughlin, 2004) or adopt a preference or a hierarchy be-ween spatial frames of references (Maurer and Peterka,005; Ohlmann, 1988; Ohlmann and Marendaz, 1991),hich operate as vicarious processes (Reuchlin, 1978).rom this viewpoint, the suitability of the selected FoR to
he task should constrain the efficiency of sensoryre)weighting and hence adaptation to current conditionsan be achieved. The vicarious theory of FoR selectionOhlmann, 1988; Ohlmann and Marendaz, 1991) predictshat in less demanding situations, similar behavioural per-ormance should be observed whatever the FoR selected.s a consequence, large interindividual differences maymerge (i.e., different groups of subjects each preferring aifferent FoR). Conversely, coping with more demandingituations is effective if—and only if—subjects select theore appropriate FoR. This reduction in the possibleodes of spatial referencing should lead subjects to sim-
lar behaviour and the disappearance of IDs (Marendaz,989; Ohlmann and Marendaz, 1991). In overview, bothifficulty of postural task and misleading distortions of theisual frame of reference should provoke shifts towards theelection of more appropriate FoRs in an attempt to obtainptimality in sensory–postural control.
There is evidence that demanding postural equilibriumasks elicit changes in the use of sensory modalities (Vuill-rme et al., 2006, 2001a) for example, towards an en-anced reliance to visual cues in sedentary subjects (Cre-ieux and Mesure, 1994) and in older subjects (Lord andenz, 2000; Lord and Webster, 1990; Sundermier et al.,
996). This enhanced visual dependency may be alsoodulated by the level of subject’s sensorimotor expertise
e.g., accomplished gymnasts vs. novices). Vuillerme et al.2001b) have shown that, although gymnasts cannot fullyompensate for the lack of vision, their capability to exploithe remaining sensory modalities when in unstable pos-ures is higher than non-gymnasts (Vuillerme et al., 2001a)nd that they respond more rapidly to proprioceptive cuesVuillerme et al., 2001b).
The aim of this study was to identify whether subjects’referred modes of spatial referencing (field independencyFI) vs. field dependency (FD)) as identified in a test of theubjective vertical (rod and frame test) are maintainedhrough test situations involving increasing difficulties ofostural control and with full access to all sensory inputsrom which to select cues to balance and orientation. Pos-ural tasks of increasing difficulties progressively enhanceynamics of balance as compared to the rod and frameest where dynamics balance are minimal. Following this
verview of the literature one might expect two opposite tatterns of behaviour in response to changes in difficulty ofostural challenge in the presence of misleading alter-tions of VFoR cues.
i) The first hypothesis predicts a maintenance or a rein-forcement of preferential modes of spatial referencing(from perceptual to postural levels). The FD subject’sreliance on the VFoR should lead to larger posturalreorientation towards a tilted frame, in an attempt torestore body/VFOR coordinate systems coalignment,increasing postural instability and an enhanced inter-segmental stiffening. Conversely, FI subjects shouldbetter resist alterations of the VFoR due to their greaterability to use alternative and more appropriate FoR(ground feet support, GIF direction), sensed via so-matosensory, vestibular and touch-pressure cues, fromwhich the body can be aligned.
i) The second hypothesis, based on the operation of vi-carious processes of FoR (Marendaz, 1989; Ohlmannand Marendaz, 1991), predicts that task demandsshould reduce the variability in modes of spatial refer-encing and to promote the emergence of unique andappropriate mode of spatial referencing, suppressingIDs. A shift toward the most appropriate FoR, shouldexploit reliable somatosensory signals of body orien-tation and downregulate misleading visual cues toorientation.
EXPERIMENTAL PROCEDURES
aterials and procedures for the fieldependence–independence screening test
Subjects. With a view to selecting subjects on the basis ofheir dependence–independence with respect to static visual field,7 healthy young men (mean age 23�3 years) without history ofestibular or ocular disorders and with normal or corrected normalision gave informed consent prior to participation according to theuidelines of the local CNRS Ethics Committee. Each of the testedubjects was naive about the experimental hypotheses at the timef initial testing. All subjects undertook the rod and frame testRFT apparatus, (Oltman, 1968)). In this test, the observer esti-ates the subjective vertical (SV) by adjusting the orientation of a
traight rod which could be rotated remotely in right–left tilt aboutts centre and which was enclosed within a square frame, whichtself may be either tilted to the right or to the left (Isableu et al.,997). An error in setting the rod to the SV typically occurs in theirection of the frame tilt. Subjects’ frame effect score show clearnd stable differences establishing the well-known dimension offield dependence–independence” (Asch and Witkin, 1948; Witkinnd Wapner, 1950; Witkin et al., 1991). Thus, an observer ratedn this dimension is characterized by the degree to which hisettings of the rod vary from the gravitational direction (field inde-endent) or to the geometrical “visual vertical” axes of the laterallyilted frame (field dependent). In this RFT, both rod and frameere initially tilted at 18° (at which the frame effect reaches aaximum (Zoccolotti et al., 1992)). The “frame effect” score,hich is the error in estimates of the subjective vertical due to the
ilted frame, was calculated according to Nyborg and Isaksen’sethod (Nyborg and Isaksen, 1974).
From this large tested population and for postural experi-ents, we could define two contrasting groups of subjects (Is-bleu et al., 1997): those who had the highest and lowest errors in
heir subjective vertical, respectively, yielding eight dependent
(e1sevrttad(nnFsys1rGtss1eadrv3sdcIasRvpsaw
P
F(bbvtvefhtvcpttisbbsa
am
sistotnssdla
Cfad
D
Bsralmhrnate
D
O
T(1ahTtdonabtftcj
P
Tetrpsoic
B. Isableu et al. / Neuroscience 169 (2010) 1199–1215 1201
FD) and 10 independent (FI) subjects. The corresponding meanrrors in FD and FI groups were respectively 7.4° (SD: 1.3°) and.7° (SD: 0.8°). The rational is based on the hypothesis thatubjects ranged in tails of the distribution of perceptual frameffect scores should rely in a more systematic manner on either aisual (FD subjects) or non-visual modes (FI subjects) of spatialeferencing. Hence, subject’s selection has to obey to some cri-erion. FD and FI subjects were selected assuming similar ampli-ude in verticality estimates in both conditions of frame lean (i.e.,bsolute errors measured in both frame lean conditions should notiffer) and weak intra-individual variance. The remaining subjects79), who had intermediate degrees of dependency, were elimi-ated insofar as they probably used a mixture of these visual andon-visual modes of spatial referencing. Thus, only the extremeD and FI subjects undertook the postural experiments. Theelected participants were right-handed and aged 22.5 (�2)ears. Given that static visual cues may contribute primarily to thelow reorientation of the upper part of the body (Amblard et al.,985), we have hypothesized that the RFT would predict a similareliance on the use of static visual cues to control body orientation.iven that dynamic visual cues may serve for rapid stabilization of
he whole body (Amblard et al., 1985) subjects were alsocreened for their dynamic visual dependency using the rod andpinning disc test (RDT) (Guerraz et al., 2001b; Isableu et al.,998). In this test (Guerraz et al., 1998), subjects were asked tostimate their subjective vertical by means of a little bar in front ofcircular visual display of randomly positioned photo-luminescentots subtending 28° at the eye and rotating either to the left or theight at a constant velocity of 30°/s. Setting errors of the perceivedertical increase with the angular velocity to reach a maximum at0°/s (Dichgans et al., 1972). In this test, subjects make alsoystematic spatial-orientation errors that are predictable from theirection of stimulus rotation. The Disc effect was calculated ac-ording to the same method used for the frame effect (Nyborg andsaksen, 1974). We have hypothesized that the RDT would predict
similar reliance regarding the use of dynamic visual cues totabilize the body. The Pearson’s correlation analysis between theFT and RDT perceptual scores revealed that static and dynamicisual field dependence–independence were significantly andositively correlated (r�0.64). This indicates that by and large, aubject who was dependent with respect to static visual field waslso dependent with respect to dynamic visual field, and the sameas true for an independent subject.
ostural tasks
D and FI subjects stood barefoot either in the normal RombergNR, weakly modified i.e., feet together), or in sharpened Rom-erg (SR, heel-to-toe or tandem posture) or in monopodal Rom-erg’s (MR) posture, on solid ground. They were presented with aisual scene at 0.7 m distance from the eyes, which was struc-ured by a fluorescent square frame whose sides subtended 50° ofisual field (1�1 m2, borders thick: 2 cm). The frame was orientedither vertical (V) or tilted (T) (18° to the left). The distance of 0.7 mor the frame from the subject’s eyes was decided firstly since itas been shown that visual objects must be at �2.5 m to stabilizeheir stance (Bles et al., 1980; Paulus et al., 1984, 1989) andisual “locking” to the environment is easier when the scene islose to the subject (Dijkstra et al., 1992). Secondly, the visualattern of the rod and frame test was situated at 0.7 m in front ofhe subject’s eyes, which makes both perceptual test and posturalask comparable as regards visual processing. The frame waslluminated by means of either an ultraviolet fluorescent bulb or atroboscopic visible light. In darkness, the ultraviolet black-lightulb illuminated the invisible frame to glow by fluorescence. Stro-oscopic vision (S) (about 2.8 flashes/s) was used to suppresselectively dynamic visual cues (Amblard and Cremieux, 1976),
nd was compared with continuous vision (C). Darkness (D) was pcontrol condition (see, Fig. 1 for an illustration of these experi-ental conditions).
In each trial, on a verbal signal from the experimenter, theubject released a manual support and was requested to remainn equilibrium for at least 14 s. During this period, he would looktraight ahead, with his arms relaxed along the trunk. The instruc-ions were to stand upright keeping optimal balance. Moreover,bservers were subjected to darkness during the first 4 s of eachrial by a liquid-crystal spectacles apparatus (Translucent Tech-ologiesTM, “Plato’s spectacle”) which is computer controlled andwitches from opaque to clear in 2 ms (Milgram, 1987). Thishutter was then open during the last 10 s of the trial, excepturing the control trials in darkness (14 s). Trials where the subject
ost his balance were repeated. For each experimental conditionnd subject, four successful trials were run for averaging.
Each of the five visual situations Darkness (D), Vertical frameontinuous vision (VC), Vertical frame Strobe vision (VS), Tilted
rame Continuous vision (TC) and Tilted frame Strobe vision (TS)nd postural situations (NR, SR and MR) was presented in aifferent random order for each subject.
ata collection
ody movements in the frontal plane were measured by the ELITEystem (Ferrigno and Pedotti, 1985) with the seven sphericaletroflective markers fixed over the mastoid bone of the head (1,2),cromion (3,4), posterior iliac spine (5,6) and lateral malleolus of the
eft back foot (7). With this particular arrangement of the markers, weeasured lateral angular rotations of the head (1,2), shoulder (3,4),ip (5,6) and leg (5–7) around the anterior–posterior axis (i.e., in theoll plane), at a sampling frequency of 100 Hz. Digital filtering foroise reduction was with a Finite Impulse Response filter (D’Amicond Ferrigno, 1990). System accuracy was estimated at 1/3000 ofhe maximal dimension of the useful acquisition volume yielding anrror of less than 2 mm on a markers 2D position.
ata analysis
rientation analyses
he mean orientation of the head (or whole body) in the roll axisfrontal plane) was taken as the average from second 4 to second4 of the corresponding corrected measurements (orientation rel-tive to darkness). In a given experimental condition, the subject’sead or whole-body orientation was then averaged over four trials.he head (or whole-body) orientation in darkness (from second 4
o second 14) was then subtracted from that of each visual con-ition (VC, VS, TC and TS), to obtain the head (or whole body)rientation relative to darkness. In some cases, there was aon-visual slight shift of the head or body orientation in darkness,fter the first 4 s. The difference between the head (or whole-ody) orientation relative to darkness in front of a vertical and ailted visual frame was called the head (or whole-body) posturalrame effect. The postural frame effect is thus the reorientation ofhe head (or whole-body) induced by a visual tilted frame, byomparison with a vertical one. We postulated that the FD sub-ects would lean laterally towards the frame tilt.
ostural performances
he head and body recordings (between seconds 4 and 14 ofach trial) were also subjected to a spectral analysis. For eachrial, the power spectrum of the component frequencies of theotations was obtained by a fast Fourier transform program. Theostural performance was calculated as the logarithm of the powerpectrum between 0 and 2.5 Hz (0.1-Hz bins). The overall measuref a subject’s head (or whole-body) postural performance (or stabil-
ty) was thus the average of this mean power from four trials in similaronditions (Amblard et al., 1985). A decrease in this averaged mean
ower reflects an increase in the postural stability.
As
TAhstnsab(c
sorli
lclbmdb
mtfsstA
Ts
Ntl1lccl1sclsmmg
S
Ts
Fi er is refe
B. Isableu et al. / Neuroscience 169 (2010) 1199–12151202
ssessing the strategies of intersegmentaltabilization
he anchoring index (AI; see (Amblard et al., 1997; Assaiante andmblard, 1993)) of body segments measured (head, shoulder andip) was used to assess the subject’s strategies of segmentaltabilization, in any given experimental condition, with respect towo different frames of reference (exocentric vs. egocentric). Thisormalized AI was used to compare the stabilization of a givenegment with respect both to external space and to the underlyingnatomical segment (Amblard et al., 1997; Assaiante and Am-lard, 1993). In the example of the head AI, the angular orientationaround the roll axis) of the head relative to the shoulder was firstalculated each 10 ms during a trial using the formula:
� rH �� a
H �� aS (1)
Where �Hr is the angular orientation of the head relative to the
houlder; �Ha and �S
a are the absolute head and shoulder angularrientations, respectively. For a given trial, the variance of theelative angular distribution �2(�H
r) and the variance of the abso-ute angular distribution �2(�H
a) were then calculated. The normal-zed AI of the head was then calculated by the following formula:
AI(H)���2�� rH ���2�� a
H �� ⁄ ��2�� rH ���2�� a
H �� (2)
Similarly, the shoulder AI was calculated by comparing angu-ar orientations of the shoulder and hip, whereas the hip AI wasalculated by comparing angular orientations of the hip and lefteg. The AI is thus a description of the degree of dependencyetween two consecutive segmental movements. Values of AIay vary between �1 and �1. For any given experimental con-ition, a positive value of AI for a given segment would indicate a
CONTINUOUS VISION (C)
ig. 1. Experimental conditions. Frame orientation (vertical vs. tilted),nterpretation of the references to color in this figure legend, the read
etter stabilization in space than on the inferior supporting seg- t
ent, whereas a negative value would indicate a better stabiliza-ion on the inferior segment than in space. The AIs were Z trans-ormed before conducting statistical analyses. The AI of the hip,houlder and head were tested for significance using a one-ample analysis (t-test) against the null hypothesis. This was doneo identify the inter-segmental strategies (Amblard et al., 1997;ssaiante and Amblard, 1993).
emporal and spatial relationships betweenegmental coordinated movements
ormalized cross-correlation functions were also calculated be-ween head and shoulder, shoulder and hip and between hip andeft leg angular movements about the roll axis (Amblard et al.,994, 1997). This is usually done to assess quantitatively the time
ag (abscissa of the cross-correlation function peak) between theoordinated movements of each pair of segments and the coeffi-ients of correlation at the abscissa zero, to validate the AI calcu-ations (Assaiante and Amblard, 1993; Amblard et al., 1994,997). In-phase and anti-phase movements between pairs ofegments give positive coefficients of correlation and negativeoefficients of correlation, respectively. Correlation analyses al-owed us to confirm the validity of the AIs (none of them corre-pond to significantly negative correlation between pairs of seg-ents at zero time lag, meaning that AIs were never overesti-ated) (Assaiante and Amblard, 1993). Detailed analyses areiven in the appendix of supplementary data.
tatistics
he orientation and stability as well as AIs performances wereubjected to an appropriate ANOVA to make comparisons be-
BE VISION (S) DARKNESS (D)
mination (continuous vs. strobe) and stances (NR, SR and MR). Forrred to the Web version of this article.
STRO
visual illu
ween experimental (visual and postural) situations and groups.
TfatfpatHrw
E
PAMtBFFrewNePtcoidbwwt
rt
[hpiPsismt1cb
crtActrtwswp
P
Tst
Fi figure leg
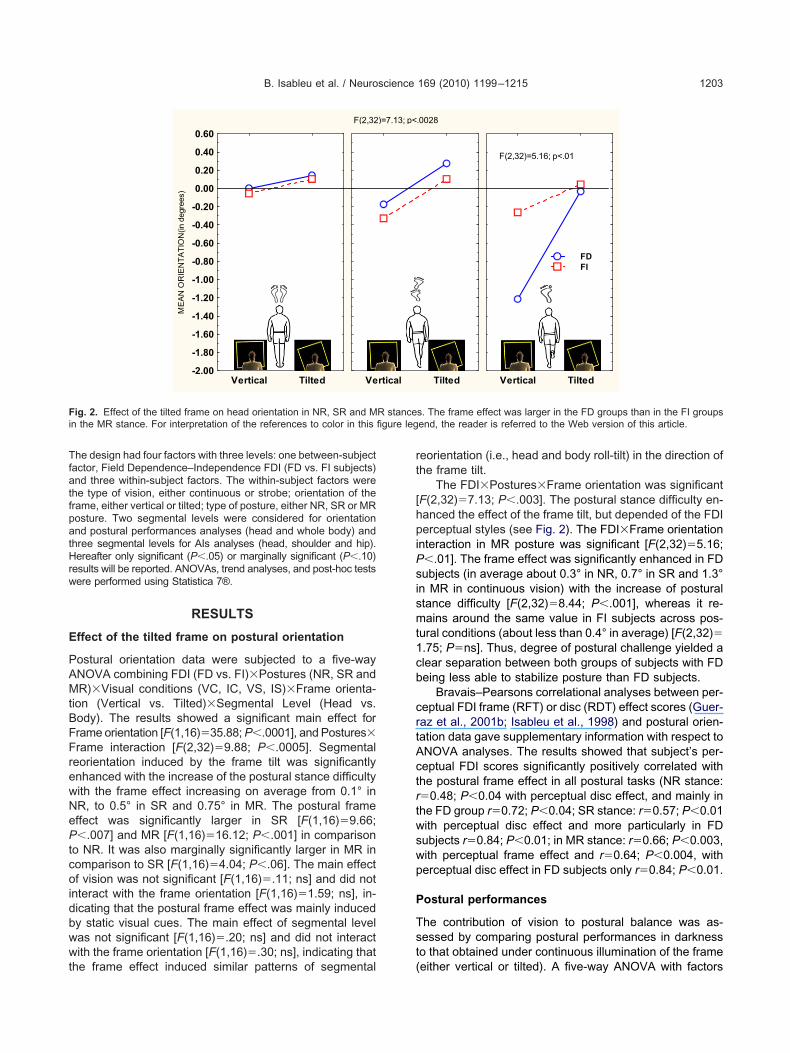
B. Isableu et al. / Neuroscience 169 (2010) 1199–1215 1203
he design had four factors with three levels: one between-subjectactor, Field Dependence–Independence FDI (FD vs. FI subjects)nd three within-subject factors. The within-subject factors were
he type of vision, either continuous or strobe; orientation of therame, either vertical or tilted; type of posture, either NR, SR or MRosture. Two segmental levels were considered for orientationnd postural performances analyses (head and whole body) and
hree segmental levels for AIs analyses (head, shoulder and hip).ereafter only significant (P�.05) or marginally significant (P�.10)
esults will be reported. ANOVAs, trend analyses, and post-hoc testsere performed using Statistica 7®.
RESULTS
ffect of the tilted frame on postural orientation
ostural orientation data were subjected to a five-wayNOVA combining FDI (FD vs. FI)�Postures (NR, SR andR)�Visual conditions (VC, IC, VS, IS)�Frame orienta-
ion (Vertical vs. Tilted)�Segmental Level (Head vs.ody). The results showed a significant main effect forrame orientation [F(1,16)�35.88; P�.0001], and Postures�rame interaction [F(2,32)�9.88; P�.0005]. Segmentaleorientation induced by the frame tilt was significantlynhanced with the increase of the postural stance difficultyith the frame effect increasing on average from 0.1° inR, to 0.5° in SR and 0.75° in MR. The postural frameffect was significantly larger in SR [F(1,16)�9.66;�.007] and MR [F(1,16)�16.12; P�.001] in comparison
o NR. It was also marginally significantly larger in MR inomparison to SR [F(1,16)�4.04; P�.06]. The main effectf vision was not significant [F(1,16)�.11; ns] and did not
nteract with the frame orientation [F(1,16)�1.59; ns], in-icating that the postural frame effect was mainly inducedy static visual cues. The main effect of segmental levelas not significant [F(1,16)�.20; ns] and did not interactith the frame orientation [F(1,16)�.30; ns], indicating that
F(2,32
ME
AN
OR
IEN
TA
TIO
N(in
deg
rees
)
Vertical Tilted-2.00
-1.80
-1.60
-1.40
-1.20
-1.00
-0.80
-0.60
-0.40
-0.20
0.00
0.20
0.40
0.60
Ve
ig. 2. Effect of the tilted frame on head orientation in NR, SR and Mn the MR stance. For interpretation of the references to color in this
he frame effect induced similar patterns of segmental (
eorientation (i.e., head and body roll-tilt) in the direction ofhe frame tilt.
The FDI�Postures�Frame orientation was significantF(2,32)�7.13; P�.003]. The postural stance difficulty en-anced the effect of the frame tilt, but depended of the FDIerceptual styles (see Fig. 2). The FDI�Frame orientation
nteraction in MR posture was significant [F(2,32)�5.16;�.01]. The frame effect was significantly enhanced in FDubjects (in average about 0.3° in NR, 0.7° in SR and 1.3°
n MR in continuous vision) with the increase of posturaltance difficulty [F(2,32)�8.44; P�.001], whereas it re-ains around the same value in FI subjects across pos-
ural conditions (about less than 0.4° in average) [F(2,32)�.75; P�ns]. Thus, degree of postural challenge yielded alear separation between both groups of subjects with FDeing less able to stabilize posture than FD subjects.
Bravais–Pearsons correlational analyses between per-eptual FDI frame (RFT) or disc (RDT) effect scores (Guer-az et al., 2001b; Isableu et al., 1998) and postural orien-ation data gave supplementary information with respect toNOVA analyses. The results showed that subject’s per-eptual FDI scores significantly positively correlated withhe postural frame effect in all postural tasks (NR stance:�0.48; P�0.04 with perceptual disc effect, and mainly inhe FD group r�0.72; P�0.04; SR stance: r�0.57; P�0.01ith perceptual disc effect and more particularly in FDubjects r�0.84; P�0.01; in MR stance: r�0.66; P�0.003,ith perceptual frame effect and r�0.64; P�0.004, witherceptual disc effect in FD subjects only r�0.84; P�0.01.
ostural performances
he contribution of vision to postural balance was as-essed by comparing postural performances in darknesso that obtained under continuous illumination of the frame
.0028
Tilted Vertical Tilted
FD FI
F(2,32)=5.16; p<.01
s. The frame effect was larger in the FD groups than in the FI groupsend, the reader is referred to the Web version of this article.
)=7.13; p<
rtical
R stance
either vertical or tilted). A five-way ANOVA with factors
[(m[77saeP
(PcmctFPiP
Fs
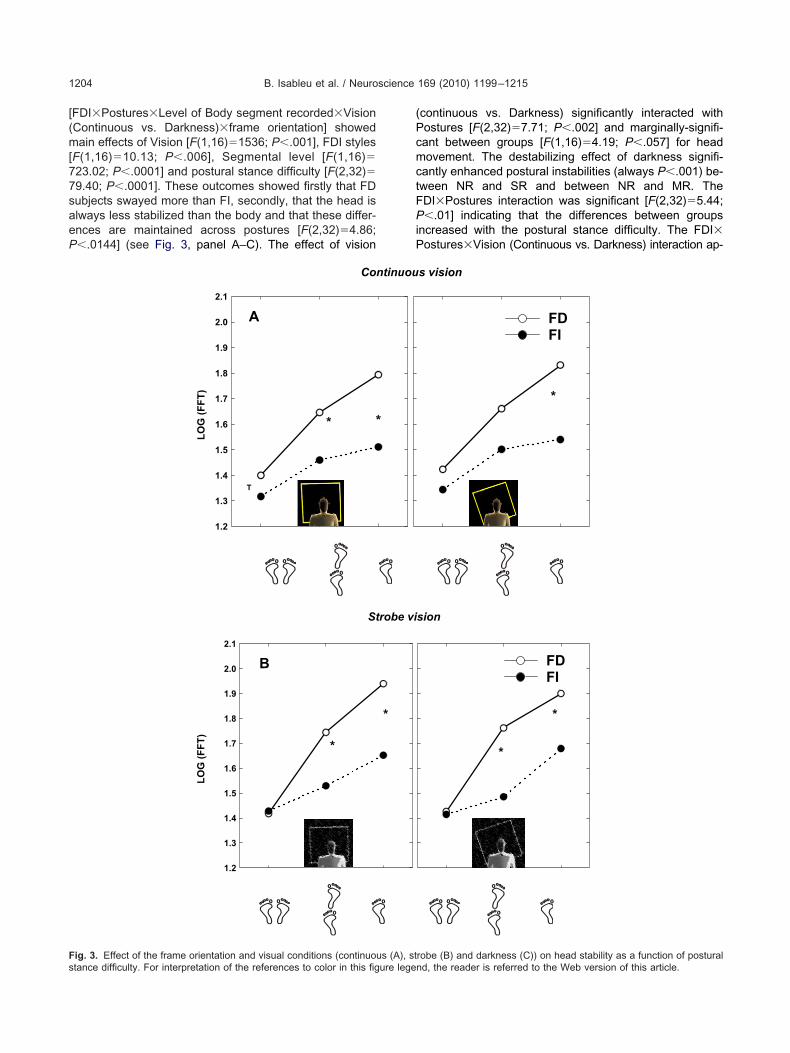
B. Isableu et al. / Neuroscience 169 (2010) 1199–12151204
FDI�Postures�Level of Body segment recorded�VisionContinuous vs. Darkness)�frame orientation] showedain effects of Vision [F(1,16)�1536; P�.001], FDI styles
F(1,16)�10.13; P�.006], Segmental level [F(1,16)�23.02; P�.0001] and postural stance difficulty [F(2,32)�9.40; P�.0001]. These outcomes showed firstly that FDubjects swayed more than FI, secondly, that the head islways less stabilized than the body and that these differ-nces are maintained across postures [F(2,32)�4.86;�.0144] (see Fig. 3, panel A–C). The effect of vision
LOG
(FFT
)
Con
1.2
1.3
1.4
1.5
1.6
1.7
1.8
1.9
2.0
2.1
*
T
*
A
LOG
(FFT
)
St
1.2
1.3
1.4
1.5
1.6
1.7
1.8
1.9
2.0
2.1
*
B
ig. 3. Effect of the frame orientation and visual conditions (continuous (A), sttance difficulty. For interpretation of the references to color in this figure legen
continuous vs. Darkness) significantly interacted withostures [F(2,32)�7.71; P�.002] and marginally-signifi-ant between groups [F(1,16)�4.19; P�.057] for headovement. The destabilizing effect of darkness signifi-
antly enhanced postural instabilities (always P�.001) be-ween NR and SR and between NR and MR. TheDI�Postures interaction was significant [F(2,32)�5.44;�.01] indicating that the differences between groups
ncreased with the postural stance difficulty. The FDI�ostures�Vision (Continuous vs. Darkness) interaction ap-
s vision
*
FDFI
sion
*
*
FDFI
tinuou
robe vi
*
robe (B) and darkness (C)) on head stability as a function of posturald, the reader is referred to the Web version of this article.
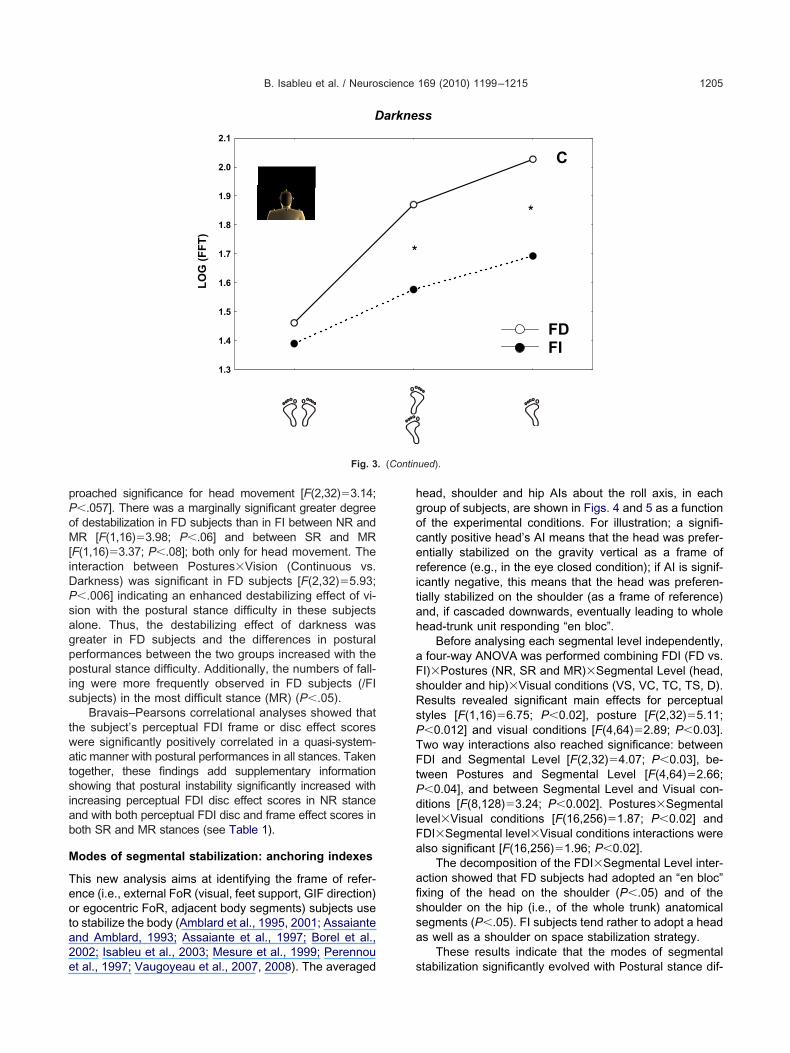
pPoM[iDPsagppis
twatsiab
M
Teota2e
hgoceritah
aFsRsPTFtPdlFa
afissa
. (Contin
B. Isableu et al. / Neuroscience 169 (2010) 1199–1215 1205
roached significance for head movement [F(2,32)�3.14;�.057]. There was a marginally significant greater degreef destabilization in FD subjects than in FI between NR andR [F(1,16)�3.98; P�.06] and between SR and MR
F(1,16)�3.37; P�.08]; both only for head movement. Thenteraction between Postures�Vision (Continuous vs.arkness) was significant in FD subjects [F(2,32)�5.93;�.006] indicating an enhanced destabilizing effect of vi-ion with the postural stance difficulty in these subjectslone. Thus, the destabilizing effect of darkness wasreater in FD subjects and the differences in posturalerformances between the two groups increased with theostural stance difficulty. Additionally, the numbers of fall-
ng were more frequently observed in FD subjects (/FIubjects) in the most difficult stance (MR) (P�.05).
Bravais–Pearsons correlational analyses showed thathe subject’s perceptual FDI frame or disc effect scoresere significantly positively correlated in a quasi-system-tic manner with postural performances in all stances. Takenogether, these findings add supplementary informationhowing that postural instability significantly increased withncreasing perceptual FDI disc effect scores in NR stancend with both perceptual FDI disc and frame effect scores inoth SR and MR stances (see Table 1).
odes of segmental stabilization: anchoring indexes
his new analysis aims at identifying the frame of refer-nce (i.e., external FoR (visual, feet support, GIF direction)r egocentric FoR, adjacent body segments) subjects useo stabilize the body (Amblard et al., 1995, 2001; Assaiantend Amblard, 1993; Assaiante et al., 1997; Borel et al.,002; Isableu et al., 2003; Mesure et al., 1999; Perennou
LOG
(FFT
)
D
1.3
1.4
1.5
1.6
1.7
1.8
1.9
2.0
2.1
Fig. 3
t al., 1997; Vaugoyeau et al., 2007, 2008). The averaged s
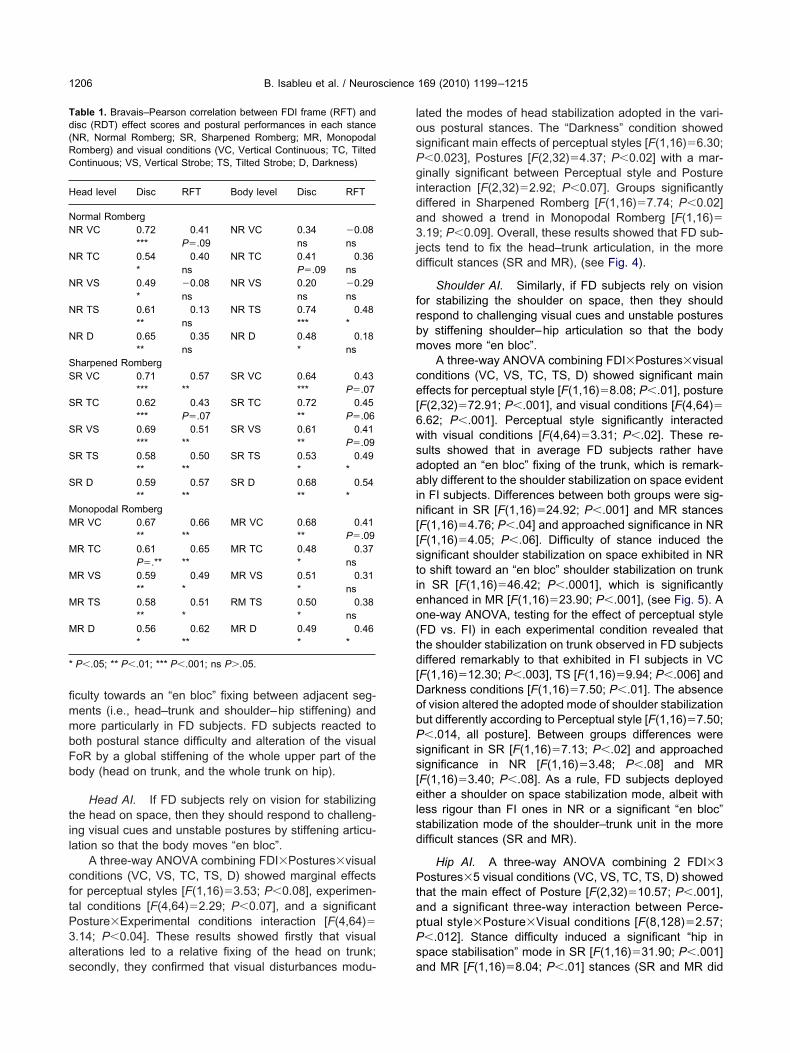
ead, shoulder and hip AIs about the roll axis, in eachroup of subjects, are shown in Figs. 4 and 5 as a functionf the experimental conditions. For illustration; a signifi-antly positive head’s AI means that the head was prefer-ntially stabilized on the gravity vertical as a frame ofeference (e.g., in the eye closed condition); if AI is signif-cantly negative, this means that the head was preferen-ially stabilized on the shoulder (as a frame of reference)nd, if cascaded downwards, eventually leading to wholeead-trunk unit responding “en bloc”.
Before analysing each segmental level independently,four-way ANOVA was performed combining FDI (FD vs.I)�Postures (NR, SR and MR)�Segmental Level (head,houlder and hip)�Visual conditions (VS, VC, TC, TS, D).esults revealed significant main effects for perceptualtyles [F(1,16)�6.75; P�0.02], posture [F(2,32)�5.11;�0.012] and visual conditions [F(4,64)�2.89; P�0.03].wo way interactions also reached significance: betweenDI and Segmental Level [F(2,32)�4.07; P�0.03], be-
ween Postures and Segmental Level [F(4,64)�2.66;�0.04], and between Segmental Level and Visual con-itions [F(8,128)�3.24; P�0.002]. Postures�Segmental
evel�Visual conditions [F(16,256)�1.87; P�0.02] andDI�Segmental level�Visual conditions interactions werelso significant [F(16,256)�1.96; P�0.02].
The decomposition of the FDI�Segmental Level inter-ction showed that FD subjects had adopted an “en bloc”xing of the head on the shoulder (P�.05) and of thehoulder on the hip (i.e., of the whole trunk) anatomicalegments (P�.05). FI subjects tend rather to adopt a heads well as a shoulder on space stabilization strategy.
These results indicate that the modes of segmental
ss
*
C
FDFI
ued).
arkne
*
tabilization significantly evolved with Postural stance dif-
fimmbFb
til
cftP3as
losPgida3jd
frbm
ce[6wsaain[[stieo(td[DobPss[elsd
PtapPs
Td(RC
H
NN
N
N
N
N
SS
S
S
S
S
MM
M
M
M
M
*
B. Isableu et al. / Neuroscience 169 (2010) 1199–12151206
culty towards an “en bloc” fixing between adjacent seg-ents (i.e., head–trunk and shoulder–hip stiffening) andore particularly in FD subjects. FD subjects reacted tooth postural stance difficulty and alteration of the visualoR by a global stiffening of the whole upper part of theody (head on trunk, and the whole trunk on hip).
Head AI. If FD subjects rely on vision for stabilizinghe head on space, then they should respond to challeng-ng visual cues and unstable postures by stiffening articu-ation so that the body moves “en bloc”.
A three-way ANOVA combining FDI�Postures�visualonditions (VC, VS, TC, TS, D) showed marginal effectsor perceptual styles [F(1,16)�3.53; P�0.08], experimen-al conditions [F(4,64)�2.29; P�0.07], and a significantosture�Experimental conditions interaction [F(4,64)�.14; P�0.04]. These results showed firstly that visuallterations led to a relative fixing of the head on trunk;
able 1. Bravais–Pearson correlation between FDI frame (RFT) andisc (RDT) effect scores and postural performances in each stanceNR, Normal Romberg; SR, Sharpened Romberg; MR, Monopodalomberg) and visual conditions (VC, Vertical Continuous; TC, Tiltedontinuous; VS, Vertical Strobe; TS, Tilted Strobe; D, Darkness)
ead level Disc RFT Body level Disc RFT
ormal RombergR VC 0.72 0.41 NR VC 0.34 �0.08
*** P�.09 ns nsR TC 0.54 0.40 NR TC 0.41 0.36
* ns P�.09 nsR VS 0.49 �0.08 NR VS 0.20 �0.29
* ns ns nsR TS 0.61 0.13 NR TS 0.74 0.48
** ns *** *R D 0.65 0.35 NR D 0.48 0.18
** ns * nsharpened RombergR VC 0.71 0.57 SR VC 0.64 0.43
*** ** *** P�.07R TC 0.62 0.43 SR TC 0.72 0.45
*** P�.07 ** P�.06R VS 0.69 0.51 SR VS 0.61 0.41
*** ** ** P�.09R TS 0.58 0.50 SR TS 0.53 0.49
** ** * *R D 0.59 0.57 SR D 0.68 0.54
** ** ** *onopodal RombergR VC 0.67 0.66 MR VC 0.68 0.41
** ** ** P�.09R TC 0.61 0.65 MR TC 0.48 0.37
P�.** ** * nsR VS 0.59 0.49 MR VS 0.51 0.31
** * * nsR TS 0.58 0.51 RM TS 0.50 0.38
** * * nsR D 0.56 0.62 MR D 0.49 0.46
* ** * *
P�.05; ** P�.01; *** P�.001; ns P�.05.
econdly, they confirmed that visual disturbances modu- a
ated the modes of head stabilization adopted in the vari-us postural stances. The “Darkness” condition showedignificant main effects of perceptual styles [F(1,16)�6.30;�0.023], Postures [F(2,32)�4.37; P�0.02] with a mar-inally significant between Perceptual style and Posture
nteraction [F(2,32)�2.92; P�0.07]. Groups significantlyiffered in Sharpened Romberg [F(1,16)�7.74; P�0.02]nd showed a trend in Monopodal Romberg [F(1,16)�.19; P�0.09]. Overall, these results showed that FD sub-
ects tend to fix the head–trunk articulation, in the moreifficult stances (SR and MR), (see Fig. 4).
Shoulder AI. Similarly, if FD subjects rely on visionor stabilizing the shoulder on space, then they shouldespond to challenging visual cues and unstable posturesy stiffening shoulder–hip articulation so that the bodyoves more “en bloc”.
A three-way ANOVA combining FDI�Postures�visualonditions (VC, VS, TC, TS, D) showed significant mainffects for perceptual style [F(1,16)�8.08; P�.01], posture
F(2,32)�72.91; P�.001], and visual conditions [F(4,64)�.62; P�.001]. Perceptual style significantly interactedith visual conditions [F(4,64)�3.31; P�.02]. These re-ults showed that in average FD subjects rather havedopted an “en bloc” fixing of the trunk, which is remark-bly different to the shoulder stabilization on space evident
n FI subjects. Differences between both groups were sig-ificant in SR [F(1,16)�24.92; P�.001] and MR stancesF(1,16)�4.76; P�.04] and approached significance in NRF(1,16)�4.05; P�.06]. Difficulty of stance induced theignificant shoulder stabilization on space exhibited in NRo shift toward an “en bloc” shoulder stabilization on trunkn SR [F(1,16)�46.42; P�.0001], which is significantlynhanced in MR [F(1,16)�23.90; P�.001], (see Fig. 5). Ane-way ANOVA, testing for the effect of perceptual styleFD vs. FI) in each experimental condition revealed thathe shoulder stabilization on trunk observed in FD subjectsiffered remarkably to that exhibited in FI subjects in VCF(1,16)�12.30; P�.003], TS [F(1,16)�9.94; P�.006] andarkness conditions [F(1,16)�7.50; P�.01]. The absencef vision altered the adopted mode of shoulder stabilizationut differently according to Perceptual style [F(1,16)�7.50;�.014, all posture]. Between groups differences wereignificant in SR [F(1,16)�7.13; P�.02] and approachedignificance in NR [F(1,16)�3.48; P�.08] and MRF(1,16)�3.40; P�.08]. As a rule, FD subjects deployedither a shoulder on space stabilization mode, albeit with
ess rigour than FI ones in NR or a significant “en bloc”tabilization mode of the shoulder–trunk unit in the moreifficult stances (SR and MR).
Hip AI. A three-way ANOVA combining 2 FDI�3ostures�5 visual conditions (VC, VS, TC, TS, D) showed
hat the main effect of Posture [F(2,32)�10.57; P�.001],nd a significant three-way interaction between Perce-tual style�Posture�Visual conditions [F(8,128)�2.57;�.012]. Stance difficulty induced a significant “hip inpace stabilisation” mode in SR [F(1,16)�31.90; P�.001]
nd MR [F(1,16)�8.04; P�.01] stances (SR and MR did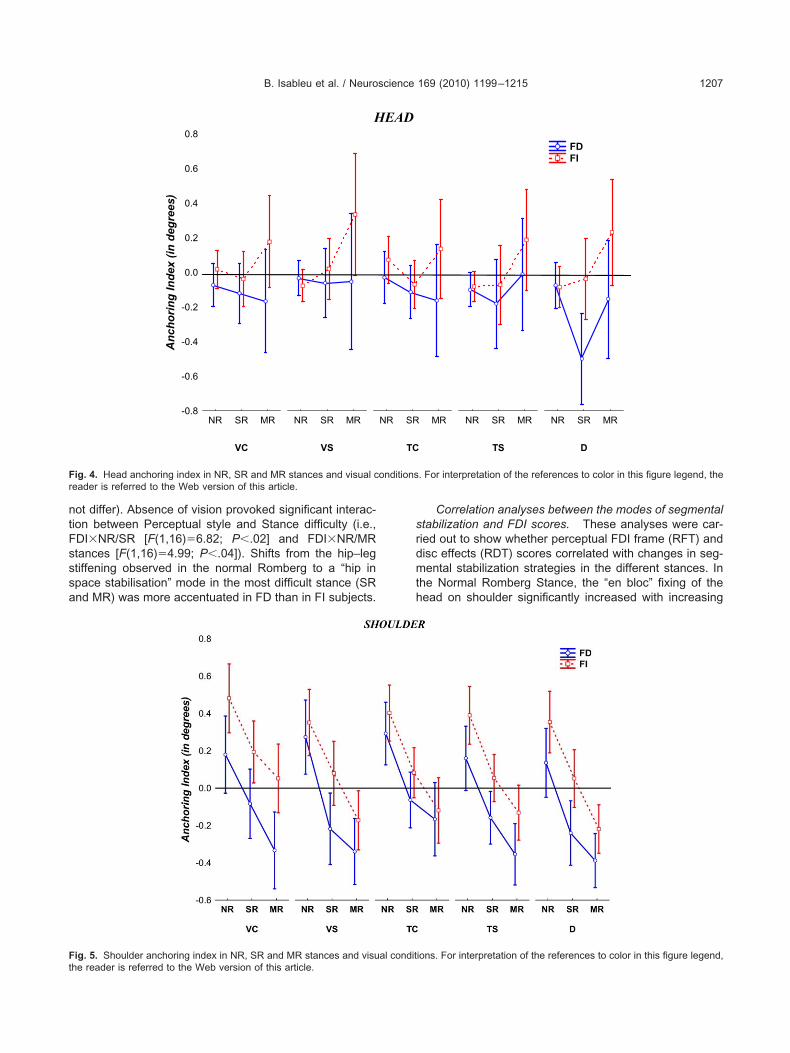
ntFsssa
srdmth
Fr
Ft
B. Isableu et al. / Neuroscience 169 (2010) 1199–1215 1207
ot differ). Absence of vision provoked significant interac-ion between Perceptual style and Stance difficulty (i.e.,DI�NR/SR [F(1,16)�6.82; P�.02] and FDI�NR/MRtances [F(1,16)�4.99; P�.04]). Shifts from the hip–legtiffening observed in the normal Romberg to a “hip inpace stabilisation” mode in the most difficult stance (SRnd MR) was more accentuated in FD than in FI subjects.
H
VC
NR SR MR-0.8
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
Anc
horin
g In
dex
(in d
egre
es)
VS
NR SR MR
ig. 4. Head anchoring index in NR, SR and MR stances and visual ceader is referred to the Web version of this article.
ig. 5. Shoulder anchoring index in NR, SR and MR stances and visual condithe reader is referred to the Web version of this article.
Correlation analyses between the modes of segmentaltabilization and FDI scores. These analyses were car-ied out to show whether perceptual FDI frame (RFT) andisc effects (RDT) scores correlated with changes in seg-ental stabilization strategies in the different stances. In
he Normal Romberg Stance, the “en bloc” fixing of theead on shoulder significantly increased with increasing
MR
TS
NR SR MR
D
NR SR MR
FD FI
. For interpretation of the references to color in this figure legend, the
EAD
TC
NR SR
onditions
ions. For interpretation of the references to color in this figure legend,

FrtpTsssvsstaPchlfih
ehfaAciPrphsdo
os(sbtutbtiiwsT(cpsvc(
OmtcvvctbfFsetdatdIhul
tvttp1dIFbtpotipbeS2tsohwsmarftov
B. Isableu et al. / Neuroscience 169 (2010) 1199–12151208
DI disc effect scores in TS (r��.72; P�.001). Theseesults also extended to the trunk. The “en bloc” fixing ofhe shoulder–hip unit went hand in hand with increasingerceptual FDI frame effect scores in VC (r��.54; P�.03),S (r��.51; P�.03) and D (r��.52; P�.03). These re-ults showed that increasing perceptual field dependencecores corresponded to higher (negative AI) values ofhoulder on hip stabilization (i.e., trunk stiffening). Con-ersely, increasing perceptual field independencecores corresponded to higher values (positive AI) ofhoulder on space stabilization (trunk articulated func-ioning). Interestingly, the pattern of results is reversedt the hip level in TC (r�.51; P�.033) and D (r�.50;�.03). These positive correlations indicate that in-reasing perceptual field dependence scores go withigher values of hip stabilization on space (hip articu-
ated functioning). Conversely, increasing perceptualeld independence scores are tied to higher values ofip on leg stabilization (hip stiffening).
The same pattern of result also emerged in the Sharp-ned Romberg Stance where the “en bloc” fixing of theead significantly increased with increasing perceptual FDIrame effect scores in TS (r��.54; P�.02). These resultslso extended to the trunk (shown by the negative shoulderI). The “en bloc” fixing of the shoulder–hip unit signifi-antly increased with increasing field dependence scoresn almost all the experimental conditions (VC: r��.53;�.03; VS: r��.64; P�.005; TS: r��.66; P�.005 and D:
��.79, P�.000) except in TC (ns.). In darkness, highererceptual field dependence scores corresponded toigher values of shoulder on hip stabilization (i.e., trunktiffening). Conversely, higher perceptual field indepen-ence scores corresponded to higher values of shouldern space stabilization (trunk articulated functioning).
In the Monopodal Romberg Stance, the “en bloc” fixingf the head on trunk significantly increased with increasingcores of static perceptual visual field dependence in VCr��.50; P�.03). Higher perceptual field dependencecores corresponded to high values of head on trunk sta-ilization (i.e., head stiffening). Conversely, higher percep-
ual field independence scores corresponded to high val-es of head on space stabilization (head articulated func-
ioning). These results also extended to the trunk (attestedy the negative shoulder AI). The “en bloc” functioning of
he shoulder–hip unit significantly increased with increas-ng frame visual field dependence field dependence scoresn VC (r��0.67; P�.002) and TS (r��.50; P�.04) andith motion visual field dependence field dependencecores in VS (r��.50; P�.002) and TC (r��.49; P�.04).he pattern of results is reversed at the hip level in TCr�.51; P�.033) and D (r�.50; P�.03). These positiveorrelations indicated that increasing perceptual field de-endence scores corresponded to higher values of hiptabilization on space (hip articulated functioning). Con-ersely, increasing perceptual field independence scoresorresponded to higher values of hip on leg stabilization
hip stiffening). nDISCUSSION
ur experiment addressed the issue of adaptation ofechanisms of postural control and multisensory integra-
ion to maintain balance and orientation in stances of in-reasing difficulty and in environmental conditions whereinisual cues of self-orientation (coordinate system of theisual FOR) and self-motion (dynamics visual cues) be-ome misleading or are suppressed. It addressed the ex-ent to which the enhanced prominence of dynamics ofalance in difficult postures (Riccio et al., 1992) is a keyactor in identifying the coordinate systems of appropriateORs and downregulates the influence of misleading vi-ual FOR. Our study was aimed also at identifying to whatxtent the idiosyncratic selection of FOR in spatial percep-ion, hypothesized to be a main source of interindividualifferences in balance control, is a rigidly fixed or adapt-ble property of the individual. The experiment tested, (i)he putative covariation between the visual field depen-ence/independence identified from the RFT test and theDs in the visual FOR influence on postural control, and (ii)ow the idiosyncratic modes of spatial referencing evolvednder increasing pressure of balance challenge and mis-
eading visual cues.Earlier reports have provided evidence that postural
ask demands can influence “perceptual styles” as re-ealed in perceptive–cognitive tasks (e.g., in Kopfermannest, see (Marendaz, 1989; Ohlmann, 1995a,b) accordingo a vicarious process. Within this framework, vicariousrocesses (Marendaz, 1989; Ohlmann and Marendaz,991; Reuchlin, 1978) predict, on the one hand, that un-emanding postural tasks should be associated with largeDs due to the equi-efficiency of the multiple availableoRs. On the other hand, the enhancement of dynamics ofalance with demanding postural stances should unravelhe different available FORs and disclose the more appro-riate ones. Hence, demanding postural stances shouldblige subjects to rely on the same, hopefully most effec-ive mode of spatial referencing, leading hence ironing outnterindividual variation. Conversely, other studies haverovided evidence that increasing difficulties in posturalalance enhance rather than suppress individual differ-nces (Cremieux and Mesure, 1994; Mesure et al., 1997;treepey and Angulo-Kinzler, 2002; Streepey et al.,007a,b; Vuillerme et al., 2001a). Given the ambiguity inhe literature regarding how perceptual and sensorimotortyles should evolve with changing levels of task demand,ur experiment was directed to probe these alternativeypotheses. Regarding the idiosyncratic FORs reliance,e have hypothesized that (i) FD subjects a rapid regres-ion of the strategies of segmental stabilization toward aore “en bloc” response of the body, when confronted tolteration of self-orientation and self-motion visual cues,esulting in increased postural instability and “posturalrame” effects. This behavioural pattern should reflect at-empts to preserve the visual coalignment of the main axesf the coordinate systems of the body with those of theisual FoR (ii) FI subjects to rely mainly on appropriate
on-visual FORs for strategies of segmental stabilization
llckc
Ps(
PopMoj1esbavscnofitcto
joeppebittfisb
ametedtrtvict
orhsosSsi(iass0wcmTit(
dsggistirac(hassspm
sosTicsdssnc2aa
B. Isableu et al. / Neuroscience 169 (2010) 1199–1215 1209
eading them hence to be less sensitive to postural chal-enges reflecting their greater ability to use both proprio-eptive, somatosensory as well as vestibular kinetic and/orinematic cues of body alignment with the support surfaceontact and/or with GIF direction.
ostural orientation, stability and strategies ofegmental stabilization relative to either externalspace) or egocentric (adjacent segments) FORs
ostural orientation measurements clearly revealed that,n average, there was a larger visually-frame inducedostural leaning in the more demanding task (i.e., in theonopodal stance), particularly in FD subjects. The meanrientation change induced by the tilted frame in FD sub-
ects is about 0.3° in NR and evolved to 0.5° en SR and to.4° en MR (up to 2.4° in the more FD subjects in MR,quivalent to a head deviation of about 4 cm). In the FDubjects correlation analyses revealed close covariationetween the strength of the perceptual frame or disc effectnd postural reorientation toward the tilted frame. Con-ersely, the postural frame effect in FI subjects remainedimilar (about 0.5°) in NR and SR and even slightly de-reased in the MR posture to 0.2–0.4° (according to theature of the visual cues available). Differential posturalrientation responses seem to be strongly related to visualeld dependency and differences become more marked inhe more difficult postural stances. Thus, reliance on aertain mode of spatial referencing in spatial perceptualasks seems to be maintained also for regulating posturalrientation.
Stabilisation findings provide evidence that, (i) FD sub-ects always swayed always more than FI. (ii) the difficultyf stance adopted enhanced, most of the time, the differ-nces between groups. (iii) Darkness or the selective sup-ression of either dynamic or static visual cues alteredostural stability to a greater extent in FD subjects andven more with increasing difficulty of the task. The num-er of falling was more frequently observed in FD subjects
n the most difficult stance (MR). Nevertheless, given thathe contribution of vision to stability is larger in FD subjects,he differences between both groups were still present withull vision. Thus, a greater contribution of non-visual cuesn FI subjects for control of postural stability seems to be atrong argument for explaining the observed differencesetween groups.
These differences in postural performances could belso attributable to the use of preferential modes of seg-ental stabilization relative to either external (space) orgocentric (segmental) FoRs, (of which the relative effec-iveness being possibly different). Thus, analyses of strat-gies of segmental stabilization could provide further un-erstanding on how sensorimotor styles (non-disorien-ed-stable vs. disoriented-unstable subjects) in posturalegulation emerge and need to be addressed in relation tohe question of the physical vs spatial FOR selected (visuals. non-visual) by subjects. The main features were thatncreasing stance difficulty elicited: (1) In FD subjects, ahange from a shoulder on space strategy in NR stance
oward a progressive enhancement of the “en bloc” fixing af the shoulder–hip unit (whole trunk) in SR stance, thateached a maximum in the MR stance associated with aead–trunk “en bloc” fixing. These head and shouldertabilization strategies were associated with a strategyf stabilizing the hip in space. (2) In FI subjects, the shouldertabilization in space, observed in NR was maintained inR stance (whereas FD subjects adopted whole trunktiffening) and changed for a shoulder–hip stiffening in MR
n association with a stabilization of head in space strategywhereas FD subjects have adopted a head–trunk stiffen-ng in the MR stance). Cross correlation analyses (seeppendix) showed that angular movements of the head,houlder and hip were significantly coordinated in theame direction (in-phase), without time lag (peak close to). In addition, these positive coefficients of correlationere significantly higher in FD than in FI subjects, thusonfirming an “en bloc” operation of the head–trunk unit, aseasured by the AIs in the FD subjects (see above).aken together, FI’s head, shoulder and hip moved rather
ndependently from each other (articulated functioning ofhe whole body), largely independently of visual conditionssee appendix).
Correlation analyses between perceptual visual fieldependence–independence scores and AIs scoreshowed that head in space and shoulder in space strate-ies were adopted in most of our FI subjects, indicating areater reliance on vestibular cues for stabilizing their head
n space and on proprioceptive cues for stabilizing theirhoulders in space. This suggests that both spatial rela-ionships stemming from the body-in-space (top anchor-ng) and the body-on-support (down anchoring) frames ofeference were effectively exploited in FI subjects, whichfforded them independent control over the two body massomponents with their respective degrees of anchoringsee appendix). Conversely, the more FD subjects showedigh level of head-shoulder and whole trunk stiffening (seeppendix). In FD subjects, the stabilization of the hip inpace, associated with an “en bloc” operation of the head–houlder and/or shoulder/hip unit, which induced a corre-ponding stability of the whole trunk and head, was therimary response to the increase of postural challenge andisleading visual cues.
What are the causes underlying these two distinctiveensorimotor patterns? (i) The “multi-articulated fixing, dis-riented-unstable” FD’s subjects and (ii) the “head inpace-articulated, non-disoriented-stable” FI’s subjects.he central nervous system has to coordinate sensory
nputs in order to gather relevant and reliable information toontrol alignment of axes of the egocentric FoR with re-pect to the external FOR (GIF, ground support, visualirection). In order to maximize information reliability, sen-ory inputs have to be integrated to improve the ratioignal/noise by (i) combining as many sensory cues asecessary, (ii) adaptively downweighting noisy sensoryues and overweighting reliable ones (Ernst and Banks,002; Ernst and Bulthoff, 2004; Hillis et al., 2002; Jeka etl., 2000; Kiemel et al., 2002; Kording et al., 2004; Kordingnd Wolpert, 2004; Oie et al., 2002; Vuillerme et al., 2007)
nd (iii) dynamically reweighting them when changes oc-
cTdtcmFttatkF
lmciiapisa
Sc
TtpBEhdiEaK2bobbtso2KS(fiIiipdeb
gtssTriarltiviwindaartaobcjaAcpHmotetbsiiag
Ssc
Hpwomm(tuas
B. Isableu et al. / Neuroscience 169 (2010) 1199–12151210
ur in the environmental, task or subject’s internal state.his challenge raises the issue of the subject’s ability toetect changes of reliability in sensory inputs. To maximize
he relevance of information, sensory inputs have to beoordinated to select the most appropriate FORs and,ore specifically, the most relevant axes of the egocentricOR (Isableu et al., 2009), which have to be coaligned with
he most relevant axes of the exocentric FORs. This raiseshe further issue of the subject’s ability to (i) detect theppropriateness of FORs and relevance of axes of rota-
ion, and to (ii) coordinate spatiotemporal relationships ofinematic or kinetic nature according to the nature of theOR selected.
Thus, interindividual differences can appear at manyevels. Given that we have manipulated both visual self
otion (decreased reliability) and visual self orientationues (Visual FOR), and mechanical stance configuration tonduce FORs change, we argue that the issue of maximiz-ng the relevance of the information that is, to detect, selectnd exploit an appropriate FOR(s), could be viewed as therimary cause of IDs and intervenes upstream of sensory
ntegration. With this view, selecting inappropriate FORshould inevitably lead to produce suboptimal perceptualnd sensorimotor behaviours (Avillac et al., 2005).
ensory reweighting to optimize perception andontrol of self-motion (stability) and self-orientation
here is considerable evidence stemming from perceptualo postural control studies to suggest that the brain usesrobabilistic models of multisensory integration like MLE orayes’ rule or Kalman like filters (Ernst and Banks, 2002;rnst and Bulthoff, 2004; Hillis et al., 2002), so that theuman perceptual system dynamically adjust weights onifferent information sources in proportion to their reliabil-
ty, often in a near-optimal way (Carver et al., 2005, 2006;rnst and Banks, 2002; Ernst and Bulthoff, 2004; Hillis etl., 2002; Jeka et al., 2000, 2006; Kiemel et al., 2002;ording et al., 2004; Kording and Wolpert, 2004; Oie et al.,002; Vuillerme et al., 2007). One hypothesis put forwardy various authors is that threshold variability, enhancingr decreasing responsiveness to visual information coulde the result of a shift in the “central set point” (i.e., feed-ack gain, see Welch and Ting, (2009)) that signals wheno alter the response to specific sensory pathways (e.g.,hifting from a reliance on visual information to a reliancen vestibular or proprioceptive information) (Kluzik et al.,005, 2007; Maurer et al., 2006; Streepey and Angulo-inzler, 2002; Streepey et al., 2007a,b). For instance,treepey and Angulo-Kinzler (2002), Streepey et al.
2007a,b) recently showed that the thresholds for shiftingrom a reliance on visual information to somatosensorynformation can differ even within a healthy population.nterestingly, Scrivens et al. (2008) showed that modulat-ng both feedback gain or stance configuration invoke sim-lar changes in the dynamic system response and morearticularly in the stiffness of the system in the context of aelayed-feedback system for lateral balance control. Forxample, increasing stance width while holding the feed-
ack gain constant or independently increasing feedback dain at a given stance width both provoke increasing sys-em stiffness (i.e., intrinsic stiffness of the musculoskeletalystem). In both cases, the increase in the stiffness of theystem results in faster and more oscillatory responses.his suggests that stiffness in the dynamics of the posturalesponse is linked to both parameters and emphasizes themportance of coordinated co-modulation of stance widthnd feedback gain for postural stability. Following thisationale, the observed intersegmental stiffness (whichikely involves a corresponding increase in the stiffness ofhe musculoskeletal system) associated with enhancednstability with postural demands in our FD subjects pro-ides evidence that they have difficulty in maintaining sim-
lar dynamic responses across different stance widths,hereas FI ones are able to do so. Scrivens et al. (2008)
ndicated that changes in stance width have to be coordi-ated with a change in feedback gains to produce similarynamic responses across different stance widths. Welchnd Ting, (2009) showed that the feedback gains (e.g.,cceleration information, Lockhart and Ting, 2007), whicheflect the “central set” of a subject, determine the ampli-ude of postural responses to perturbation, but can bedjusted, mainly in predictable environments. This is againne source of difference between FD subjects (governedy the various changes of the visual frame orientation andonditions of illumination in our experiment) and FI sub-
ects (relying on predictable relationships between massnd inertia within an egocentric FOR and the GIF vector).lternatively, if idiosyncrasies do exist at the level of theseentral set points, it appears that shift in the central setoint did not occurred in some subjects (like in FD ones).ow can the central set point be modified? The VFOR leanay optically evoke a fall (acceleration) of the body in thepposite direction of the frame tilt. Hence, “antifalling” pos-ural reactions are automatically generated, resulting innhanced stiffness of the whole body and postural orien-ation corrections in the opposite direction to the perceivedody lean (fall). In the case of visual disturbances, as in ourtudy, one way to deal with sensory conflicts would consist
n establishing new relationships with other available phys-cal FORs via motor-somesthetic and vestibular cues (e.g.,ngular coding of the body orientation with respect to theround support or GIF vector).
electing appropriate FOR to coordinate optimallyelf-motion and self orientation cues to posturalontrol
owever, an important aspect of the present findings sup-orts the hypothesis that optimal multisensory reweighting,hich consist of reducing the influence of noisy inputs andf enhancing weight of reliable ones to provide reliableultimodal percept of self-orientation and self-motion,ainly depends on the selection of appropriate FORs
Avillac et al., 2005; McGuire and Sabes, 2009). Amonghe subjects tested, FD subjects seem to fail to identify andse relevant sensory inputs to reweight alternative (moreppropriate) FORs. The selection of inappropriate FORshould lead mechanism of multisensory integration to pro-
uce suboptimal perceptual and sensorimotor behaviours,
etestclsrtbodaecct
A
IvpvsthifvtcdiislivTrrccsatstomswAwiiwt
hitdtSRiivctvbssjcm“uss“spt
Nsp
Smtcr2duFab(
En(hisnppvKvku
B. Isableu et al. / Neuroscience 169 (2010) 1199–1215 1211
ven if the respective degree of reliability of each input isaken into account. Thus “perceptive styles” or prior knowl-dge (top–down processes) are preferred over the as-essment of the appropriateness of non visual FORs andhe reliability of current sensory inputs (bottom–up pro-esses). As a consequence, the maintenance of the se-ected FORs from spatial perception to postural controltrongly supports top–down idiosyncratic modes of spatialeferencing. These FOR preferences, and their persis-ence, despite the enhanced prominence of dynamics ofalance, strongly suggest that the preferential coalignmentf the body direction of balance with some specific coor-inates systems within the external word was inappropri-tely proactive. Such excessive reliance on “prior” knowl-dge (i.e., hypothesizing that the visual FOR is stable andoaligned with G vector) impaired their ability to adapt tohallenging sensory-motor coordination tasks and dis-orted visual cues.
bility to detect appropriateness of non visual FORs
t is tempting to propose that, in the absence of adequateisual cues, FD subjects have locked most of their joints,erhaps due to difficulty in interpreting proprioceptive andestibular cues. By locking the major body segments, FDubjects have to deal with larger inertia loads whose con-rollability is more difficult. Nevertheless, a reason why theead stabilization on the trunk strategy, “HSTS”; (Assa-
ante and Amblard, 1993) may have been chosen underull vision could be that FD subjects needed to enhanceisual feedback to improve the visual contribution to pos-ural control. Another, but not exclusive interpretation,ould be that the “en bloc” fixing of the head-trunk unit mayirectly provide the subject with visual (and vestibular)
nformation concerning the trunk imbalance, without hav-ng to take into account the head-on-the-trunk signal. Aimilar HSTS is adopted in Parkinson’s disease duringocomotion under full vision as well as in stroboscopicllumination (Mesure et al., 1999), these patients being alsoisually dependent on the RFT task (Azulay et al., 1999).he equivalent HSTS of our FD subjects in darkness mayather correspond to a simplifying strategy aimed, firstly, ateducing the number of degrees of freedom (dof) to beontrolled simultaneously in the absence of any visualues. Secondly, it may have been aimed at providing theubject with vestibular cues concerning trunk imbalance,lso without having to take into account the head-on-the-runk signal. It could be also argued that inter-segmentaltiffening may be understood as a sensorimotor strategyhat could potentially lead FD subjects toward a better usef proprioceptive cues of angular variations of body seg-ents relative to the support surface, provided that inter-
egmental interactions (like head-on-the-trunk signal)ere withdrawn (i.e., reducing the processing complexity).llowing increasing motion (instability) of an assembly inhich many of the possible dof have been “frozen” so that
t moves simply “en bloc” could be seen as a way ofmproving the signal/noise ratio and emphasizing theeight of proprioceptive cues, for example, from foot con-
act over visual ones. Such adaptation of exploratory be- p
aviour by using of a stiffer body, may simplify the dynam-cs of balance and thereby enhance the likelihood of de-ecting more readily relevant spatial invariants (e.g.,iscovering the most appropriate axis of rotation) to helphem balance (Riccio, 1995; Riccio et al., 1992; Riccio andtoffregen, 1990; Riley and Turvey, 2002; Stoffregen andiccio, 1988; van Wegen et al., 2002). The hip stabilization
n space of the FD subjects deprived of visual cues may benterpreted as an indication that they shifted from a usualisual FOR to a non visual one linked to the hip level. Thisould be a gravitoinertial FOR based on motor-propriocep-ive loops, similar to that adopted by the toddlers at theery beginning of their walking experience, as suggestedy Assaiante and Amblard (1993). Nevertheless, our FDubjects fell more frequently in the more demandingtances. This observation suggested that these FD sub-
ects had disorientation problems, in spite of the simplifi-ation of sensory processing afforded by such inter-seg-ental stiffening strategy. It can thus be concluded that the
en bloc” fixing of the head-shoulder and/or shoulder/hipnit associated to the hip stabilization on space of FDubjects was a less efficient strategy than the head inpace and shoulder in space strategies associated with theen bloc” fixing of the hip on the legs observed in FIubjects. Similar trunk stabilization in space has beenroposed to be the main variable for control in posturalasks (Gurfinkel et al. (1995).
eglect of kinetic–kinematic signals ofelf-orientation and of self-motion (viaroprioceptive, somethestic, vestibular inputs)
ubjects have to select self orientation non visual cues toaximize their relevance. They have to gather propriocep-
ive, tactile, somatosensory and vestibular cues and tooordinate them to sense body mass or inertial or geomet-ical axes of rotation (Fourré et al., 2009; Isableu et al.,009) and to coalign one of them with external FOR coor-inate systems (support surface, GIF). Do the causesnderlying this difficulty to shift toward more appropriateORs originate (i) from a difficulty to detect FOR appropri-teness? or (ii) from a kind of neglect to sense and exploitody mass or inertial or geometrical axes of rotationFourré et al., 2009; Isableu et al., 2009)?
Two alternative interpretations could explain our data.ither FD subjects have difficulty shifting from what isormally their best FOR towards more adequate onesGround or GIF) in the case of visual perturbations, or theyad difficulty in coordinating non visual inputs (somesthet-
cs-proprioceptive and vestibular) and motor commands toense the most relevant body axes of rotation to coalignon visually with ground or GIF FORs. Could the betterostural control observed in FI subjects, despite visualerturbations, be explained by a better exploitation of non-isual inputs and ability to assess FOR appropriateness?luzik et al. (2005) and Isableu and Vuillerme (2006) pro-ided evidence of an idiosyncratic use of kinematic vsinetic proprioceptive cues to postural control related to these of the ground FOR. In both studies, subjects relying on
roprioceptive angular coding of the body with respect to
ttloasMradtjdriom2tfb2LarsljatscdtlatagssctirotdtspolhtG
ts
tioh(cwpcmpuaPa
Kn
TpcgsiosTmfi2lstsaa
ehtbasvobmaokoetsccCw
B. Isableu et al. / Neuroscience 169 (2010) 1199–12151212
he support surface (altered via a foam interposition or ailting of the support) were the more perturbed. Subjectsikely relying on mechanical-mass based variables (centref mass) (Bernardin et al., 2005; Fourré et al., 2009; Is-bleu et al., 2009) or vestibular cues could be immune touch visual alterations. In haptic perception of rod length,achon et al. (1998) have reported that FI subjects accu-
ately perceived rod length via kinaesthesis despite anlteration of either the first moment of their arm’s massistribution (either weightless) or of the coefficient of fric-ion during arm displacements. On the contrary, FD sub-ects exhibited systematic impaired performances, evi-encing their greater difficulty in using kinaesthetic cueselated to dynamics of movements efficiently. A betterntegration of force related cues to compensate alterationf limb’s mass distribution in 3D multijoint reaching move-ent was also reported in FI subjects (Bernardin et al.,005). Hence, we may assume that in both spatial percep-ion and motor control, FI subjects take a greater benefitrom force-related cues and particularly kinaesthetic mass-ased variables (Bernardin et al., 2005; Isableu et al.,009; Pagano, 2000; Pagano and Turvey, 1995; van deangenberg et al., 2007, 2008). The ability to control massnd inertia of our limbs with the ground reaction forceesultant via proprioception (Ting and MacPherson, 2004)hould be particularly important in the case of an articu-ated intersegmental functioning, as observed in FI sub-ects. Isableu et al. (2009) recently provided evidence thatngular variations of our joints break the colinearity be-ween axes of minimum rotation of different coordinateystems (mass, inertia tensor and articular geometry) thatan be detected and exploited via kinesthesia. Such coor-inate systems misalignments require subjects to choosehe adequate rotational axis to control rotation of theirimbs. It appears that the kinesthestic reliance on massnd/or inertial axes allows the minimization of muscleorque by using interaction torque to assist coordination. Itlso immunizes subjects from the disturbing effect of an-ular velocity and acceleration on limbs rotational axistability. Taken together, the results of the present studyhow that FI subjects were able to sacrifice a geometrical,ephalocaudal alignment with the GIF vector, suggestinghat they have a preference to match the axes of mass ornertial coordinate systems with GIF vector. Following thisational, it could be suggested that the “en block” co-rdination observed in FD subjects consist at maintaininghe congruency between the different proprioceptive coor-inate systems (mass, articular and inertia axes of rota-ion) to simplify their processing. This hypothesis is con-istent with the difficulty, which FD subjects have, in usingroprioceptive cues and/or to detect the more appropriater reliable rotation axes via proprioception when chal-
enged with misleading spatial visual cues. FD subjectsave simplified the problem by relying on and controllinghe alignment of spatial geometrical axes of the body withIF.
The proprioceptive neglect hypothesis in perception ofhe “appropriateness” of non visual FOR is supported by
everal results. For instance, protocols using supine pos- hure aim at limiting both motor components and the prom-nence of proprioceptive/somesthetic and vestibular cuesf dynamics of balance. In the RFT task, supine postureas been shown to enhance the reliance on the visual FORGoodenough et al., 1981; Templeton, 1973). These per-eptual results are in line with those of Bray et al. (2004),hich provided evidence that dynamics of balance duringostural stance reduce field dependency of about 30%, butould not totally overcome a misleading visual FOR in theore field independent subjects. These results add sup-ort to the hypothesis that some subjects cannot make fullse of kinematic–kinetic, non visual relationships of bodylignment with the support surface and/or GIF FORs.hysiologically, this information is present but unavailable;form of neglect.
inetic–kinematic proprioceptive and vestibulareglect and neural processing
herefore, our data suggest that relying on visual expro-rioception (FD subjects, i.e., a kinematical mode of spatialontrol) to control upright posture or to set a rod to theravitational vertical surrounded by large tilted frameeems to preclude the use of kinetic non visual cues. An
nability to master these motor-somesthetics cues shouldriginate in the reinforcement of other preferred visualpatial relationships perceived a priori, as more reliable.he development of less fine and structured cortical spatialaps should invariably occur as a consequence of less
nely tuned proprioceptive information (Coq and Xerri,001; Elbert et al., 1998; Pantev et al., 2001). This should
ead in turn to “neglect” these non visual FORs and sen-ory channels associated. This neglect would correspondo a difficulty in allocating attentional, sensorimotor re-ources to coordinates systems of non visual FORs. Thexes of these non visual FORs would be perceived a priori,s being noisier than visual ones.
fMRI studies carried out by Brandt et al. (1998); Diet-rich and Brandt (2000) and Deutschlander et al. (2002)ave shown that the (re)weighting mechanism subservinghe multi-sensory integration for self-motion perception isased on reciprocal inhibitory interaction. This mechanismllows a shift of the dominant sensorial weight duringelf-motion perception from one sensory modality (visual,estibular or somatosensory; Bense et al., 2001) to thether, depending on which mode of stimulation prevails:ody acceleration (vestibular input) or constant velocityotion (visual input). Taken together, a systematic reli-nce on visual FOR could lead thus to forsaking of the usef other spatial relationships (e.g., covariation pattern ofinetic, inertial, tactile, articular cues associated with axesf rotation controlling body mass distribution; (see, Isableut al., 2009)), and to neglect sensory channels by whichhey are conveyed. If the neglect of these non visual sen-ory channels produces less developed and less resolutiveortical motor-somesthetics maps, then it would reduce theapacity of FD subjects to resist to visual disturbances.onsequently, this central proprioceptive subdevelopmentould reduce the efficiency with which a subject can move
is body and control dynamics (torques) of mass distribu-
teetv
mtditlepgifpO(vps
Ombbcahbnwvapbmpastfioa
At
A
A
A
A
A
A
A
A
A
A
B
B
B
B
B
B
B
C
C
C
C
C
C
D
B. Isableu et al. / Neuroscience 169 (2010) 1199–1215 1213
ion from axes of rotation tied to inertia or mass FORsither with respect to ground surface and/or GIF refer-nces. Nevertheless, we hypothesize that appropriate
raining should modify this neglect toward an efficient nonisual control of body/exocentric FORs coalignment.
On the whole, our results allows us to assume that aotor-somesthetics “neglect” or a lack of mastering of
hese sensory inputs rather than a greater visual fieldependence in FD subjects would generate these inter-
ndividual differences in both spatial perception and pos-ural balance. This proprioceptive “neglect” is assumed toead FD subjects toward a difficulty in detecting and thus instablishing relationships with non-visual FORs, whereasroprioceptive dominance would lead FI subjects to areater ability in selecting the adequate FOR in the case of
nter-sensory disturbances. Our results are in line with theramework of the kinematic vs. kinetic organization of theerception and control of spatial orientation put forward byhlmann and Luyat (2001) and Gresty and Ohlmann
2002). According to these authors, selective use of kineticersus kinematic (movement related) modes of informationrocessing could partly explain interindividual variability inpatial orientation tasks.
CONCLUSION
ur results provide evidence that the effectiveness ofechanisms of multisensory integration to control posturalalance and orientation in circumstances of increasingalance difficulty and misleading static and dynamic visualues depends on the subject’s ability to identify and exploitppropriate non visual FORs. Indeed, the present resultsave shown that even when the dynamics of balanceecome more prominent, emphasizing the importance ofon visual FORs, they failed to be exploited by FD subjectsho, correspondingly, failed to down-weight misleadingisual FORs. Finally, this study also provides evidence fornew interpretation of the visual field dependence–inde-
endence in both spatial perception and postural controlased on neglect or difficulty in using non-visual cues andore particularly proprioceptive ones. Assessment of theerceptual dependence on -independence of visual FORsppears to be a practical and easy means of predicting theubject’s preferences to stabilize and orient his body rela-ive to visual or non-visual FORs. We believe that thesendings go part way to explain the individual differences inutcome in patients who undergoing rehabilitation for bal-nce disorders (Bronstein, 2004).
cknowledgments—This research was supported by a grant fromhe Centre National de La Recherche Scientifique.
REFERENCES
mblard B, Assaiante C, Fabre JC, Martin N, Massion J, Mouchnino L,Vernazza S (1995) Voluntary head stabilization in space duringtrunk movements in weightlessness. Acta Astronaut 36:415–422.
mblard B, Assaiante C, Fabre JC, Mouchnino L, Massion J (1997)Voluntary head stabilization in space during oscillatory trunk move-ments in the frontal plane performed in weightlessness. Exp Brain
Res 114:214–225.mblard B, Assaiante C, Vaugoyeau M, Baroni G, Ferrigno G, PedottiA (2001) Voluntary head stabilisation in space during oscillatorytrunk movements in the frontal plane performed before, during andafter a prolonged period of weightlessness. Exp Brain Res 137:170–179.
mblard B, Cremieux J (1976) Role of visual information concerningmovement in the maintenance of postural equilibrium in man.Agressologie 17:25–36.
mblard B, Cremieux J, Marchand AR, Carblanc A (1985) Lateralorientation and stabilization of human stance: static versus dy-namic visual cues. Exp Brain Res 61:21–37.
sch SE, Witkin HA (1948) Studies in space orientation. II. Perceptionof the upright with displaced visual fields and with body tilted. J ExpPsychol 38:455–475.
ssaiante C, Amblard B (1993) Ontogenesis of head stabilization inspace during locomotion in children: influence of visual cues. ExpBrain Res 93:499–515.
ssaiante C, McKinley PA, Amblard B (1997) Head-trunk coordinationduring hops using one or two feet in children and adults. J VestibRes 7:145–160.
villac M, Deneve S, Olivier E, Pouget A, Duhamel JR (2005) Refer-ence frames for representing visual and tactile locations in parietalcortex. Nat Neurosci 8:941–949.
zulay JP, Mesure S, Amblard B, Blin O, Sangla I, Pouget J (1999)Visual control of locomotion in Parkinson’s disease. Brain 122:111–120.
ense S, Stephan T, Yousry TA, Brandt T, Dieterich M (2001) Multi-sensory cortical signal increases and decreases during vestibulargalvanic stimulation (fMRI). J Neurophysiol 85:886–899.
ernardin D, Isableu B, Fourcade P, Bardy BG (2005) Differentialexploitation of the inertia tensor in multi-joint arm reaching. ExpBrain Res 167:487–495.
les W, Kapteyn TS, Brandt T, Arnold F (1980) The mechanism ofphysiological height vertigo. II. Posturography. Acta Otolaryngol(Stockh) 89:534–540.
orel L, Harlay F, Magnan J, Chays A, Lacour M (2002) Deficits andrecovery of head and trunk orientation and stabilization after uni-lateral vestibular loss. Brain 125:880–894.
randt T, Bartenstein P, Janek A, Dieterich M (1998) Reciprocalinhibitory visual-vestibular interaction. Visual motion stimulationdeactivates the parieto-insular vestibular cortex. Brain 121:1749–1758.
ray A, Subanandan A, Isableu B, Ohlmann T, Golding JF, Gresty MA(2004) We are most aware of our place in the world when about tofall. Curr Biol 14:R609–R610.
ronstein AM (2004) Vision and vertigo: some visual aspects of ves-tibular disorders. J Neurol 251:381–387.
arver S, Kiemel T, Jeka JJ (2006) Modeling the dynamics of sensoryreweighting. Biol Cybern 95(2):123–134.
arver S, Kiemel T, van der KH, Jeka JJ (2005) Comparing internalmodels of the dynamics of the visual environment. Biol Cybern92:147–163.
hiari L, Bertani A, Cappello A (2000) Classification of visual strate-gies in human postural control by stochastic parameters. Hum MovSci 19:817–842.
ollins JJ, De Luca CJ, Pavlik AE, Roy SH, Emley MS (1995) Theeffects of spaceflight on open-loop and closed-loop postural con-trol mechanisms: human neurovestibular studies on SLS-2. ExpBrain Res 107:145–150.
oq JO, Xerri C (2001) Sensorimotor experience modulates age-dependent alterations of the forepaw representation in the ratprimary somatosensory cortex. Neuroscience 104:705–715.
remieux J, Mesure S (1994) Differential sensitivity to static visualcues in the control of postural equilibrium in man. Percept MotSkills 78:67–74.
’Amico M, Ferrigno G (1990) Technique for the evaluation of deriv-
atives from noisy biomechanical displacement data using a model-
D
D
D
D
D
E
E
E
E
F
F
G
G
G
G
G
G
H
H
I
I
I
I
I
J
J
K
K
K
K
K
L
L
L
L
M
M
M
M
M
M
M
M
N
O
O
O
O
B. Isableu et al. / Neuroscience 169 (2010) 1199–12151214
based bandwidth-selection procedure. Med Biol Eng Comput28:407–415.
eutschlander A, Bense S, Stephan T, Schwaiger M, Brandt T, Diet-erich M (2002) Sensory system interactions during simultaneousvestibular and visual stimulation in PET. Hum Brain Mapp 16:92–103.
ichgans J, Held R, Young LR, Brandt T (1972) Moving visual scenesinfluence the apparent direction of gravity. Science 178:1217–1219.
ieterich M, Brandt T (2000) Brain activation studies on visual-vestib-ular and ocular motor interaction. Curr Opin Neurol 13:13–18.
ietz V (1993) Gating of reflexes in ankle muscles during humanstance and gait. Prog Brain Res 97:181–188.
ijkstra TM, Gielen CC, Melis BJ (1992) Postural responses to sta-tionary and moving scenes as a function to the scene. Hum MovSci 11:195–203.
hrenfried T, Guerraz M, Thilo KV, Yardley L, Gresty MA (2003)Posture and mental task performance when viewing a movingvisual field. Brain Res Cogn Brain Res 17:140–153.
lbert T, Candia V, Altenmuller E, Rau H, Sterr A, Rockstroh B, PantevC, Taub E (1998) Alteration of digital representations in somato-sensory cortex in focal hand dystonia. Neuroreport 9:3571–3575.
rnst MO, Banks MS (2002) Humans integrate visual and hapticinformation in a statistically optimal fashion. Nature 415:429–433.
rnst MO, Bulthoff HH (2004) Merging the senses into a robust per-cept. Trends Cogn Sci 8:162–169.
errigno G, Pedotti A (1985) ELITE: a digital dedicated hardwaresystem for movement analysis via real-time TV signal processing.IEEE Trans Biomed Eng 32:943–950.
ourré B, Isableu B, Bernardin D, Gueguen M, Giraudet G, VuillermeN, Pagano C, Amorim MA (2009) The role of body centre of masson haptic subjective vertical. Neurosci Lett 465:230–234.
oodenough DR, Oltman PK, Sigman E, Cox PW (1981) The rod-and-frame illusion in erect and supine observers. Percept Psychophys29:365–370.
resty MA, Ohlmann T (2002) Spatial orientation and disorientation,the fundamental theory. Curr Psychol Cogn 21:479–495.
uerraz M, Gianna CC, Burchill PM, Gresty MA, Bronstein AM(2001a) Effect of visual surrounding motion on body sway in athree-dimensional environment. Percept Psychophys 63:47–58.
uerraz M, Poquin D, Ohlmann T (1998) The role of head-centricspatial reference with a static and kinetic visual disturbance. Per-cept Psychophys 60:287–295.
uerraz M, Yardley L, Bertholon P, Pollak L, Rudge P, Gresty MA,Bronstein AM (2001b) Visual vertigo: symptom assessment, spa-tial orientation and postural control. Brain 124:1646–1656.
urfinkel VS, Ivanenko YuP, Levik YuS, Babakova IA (1995) Kines-thetic reference for human orthograde posture. Neuroscience68:229–243.
illis JM, Ernst MO, Banks MS, Landy MS (2002) Combining sensoryinformation: mandatory fusion within, but not between, senses.Science 298:1627–1630.
orstmann GA, Dietz V (1990) A basic posture control mechanism: thestabilization of the centre of gravity. Electroencephalogr Clin Neu-rophysiol 76:165–176.
sableu B, Ohlmann T, Cremieux J, Amblard B (1997) Selection ofspatial frame of reference and postural control variability. Exp BrainRes 114:584–589.
sableu B, Ohlmann T, Cremieux J, Amblard B (1998) How dynamicvisual field dependence-independence interacts with the visualcontribution to postural control. Hum Mov Sci 17:367–391.
sableu B, Ohlmann T, Cremieux J, Amblard B (2003) Differentialapproach to strategies of segmental stabilisation in postural con-trol. Exp Brain Res 150:208–221.
sableu B, Rezzoug N, Mallet G, Bernardin D, Gorce P, Pagano CC(2009) Velocity-dependent changes of rotational axes in the non-visual control of unconstrained 3D arm motions. Neuroscience
164:1632–1647.sableu B, Vuillerme N (2006) Differential integration of kinaestheticsignals to postural control. Exp Brain Res 174:763–768.
eka J, Allison L, Saffer M, Zhang Y, Carver S, Kiemel T (2006)Sensory reweighting with translational visual stimuli in young andelderly adults: the role of state-dependent noise. Exp Brain Res174 (3):517–527.
eka J, Oie KS, Kiemel T (2000) Multisensory information for humanpostural control: integrating touch and vision. Exp Brain Res134:107–125.
iemel T, Oie KS, Jeka JJ (2002) Multisensory fusion and the sto-chastic structure of postural sway. Biol Cybern 87:262–277.
luzik J, Horak FB, Peterka RJ (2005) Differences in preferred refer-ence frames for postural orientation shown by after-effects ofstance on an inclined surface. Exp Brain Res 162:474–489.
luzik J, Peterka RJ, Horak FB (2007) Adaptation of postural orienta-tion to changes in surface inclination. Exp Brain Res 178:1–17.
ording KP, Ku SP, Wolpert DM (2004) Bayesian integration in forceestimation. J Neurophysiol 92:3161–3165.
ording KP, Wolpert DM (2004) Bayesian integration in sensorimotorlearning. Nature 427:244–247.
ambrey S, Berthoz A (2003) Combination of conflicting visual andnon-visual information for estimating actively performed body turnsin virtual reality. Int J Psychophysiol 50:101–115.
ockhart DB, Ting LH (2007) Optimal sensorimotor transformations forbalance. Nat Neurosci 10:1329–1336.
ord SR, Menz HB (2000) Visual contributions to postural stability inolder adults. Gerontology 46:306–310.
ord SR, Webster IW (1990) Visual field dependence in elderly fallersand non-fallers. Int J Aging Hum Dev 31:267–277.
achon S, Ohlmann T, Belloni A, Masson K, Pinson E (1998) Im-provement of haptic spatial capacity by the reduction of motorconstraints. Representation and blindness. San Marino, Italie. Reftype: Generic.
arendaz C (1989) Selection of reference frames and the “vicariance”of perceptual systems. Perception 18:739–751.
aurer C, Mergner T, Peterka RJ (2006) Multisensory control ofhuman upright stance. Exp Brain Res 171:231–250.
aurer C, Peterka RJ (2005) A new interpretation of spontaneoussway measures based on a simple model of human posturalcontrol. J Neurophysiol 93:189–200.
cGuire LM, Sabes PN (2009) Sensory transformations and the useof multiple reference frames for reach planning. Nat Neurosci12:1056–1061.
esure S, Amblard B, Cremieux J (1997) Effect of physical training onhead-hip co-ordinated movements during unperturbed stance.Neuroreport 8:3507–3512.
esure S, Azulay JP, Pouget J, Amblard B (1999) Strategies ofsegmental stabilization during gait in Parkinson’s disease. ExpBrain Res 129:573–581.
ilgram PA (1987) A spectacle-mounted liquid-crystal tachistoscope.Behav Res Methods, Instruments, & Computers 19:449–545.
yborg H, Isaksen B (1974) A method for analysing performances inthe rod-and-frame test. II. Scand J Psychol 15:124–126.
hlmann T (1988) La perception de la verticale. Variabilité interindi-viduelle dans la dépendance à l’égard des référentiels spatiaux.Thèse pour le Doctorat d’état. Université de Paris VIII. Ref type:Thesis/Dissertation.
hlmann T (1995a) Selection of spatial reference. Symposium postureand gait: from representation to control. Marseille, France: CNRS.Ref type: Personal communication.
hlmann T (1995b) Théorie neutraliste de l’évolution et dynamiquedes processus vicariants: une nécessaire convergence. In: Univer-sel et différentiel en psychologie (Lautrey J, ed), pp 77–105. Paris:Presses Universitaires de France.
hlmann T, Luyat M (2001) La posture référencée et la posture sourcede références. In: Posture et équilibre (Lacour M, ed), pp 15–37.
Montpellier: Sauramps Médical.
O
O
O
P
P
P
P
P
P
P
R
R
R
R
R
R
S
S
S
S
S
S
T
T
v
v
v
V
V
V
V
V
V
W
W
W
Z
S
S
B. Isableu et al. / Neuroscience 169 (2010) 1199–1215 1215
hlmann T, Marendaz C (1991) Vicarious processes involved in se-lection/control of frames of reference and spatial aspects of fielddependence-independence. In: Field dependence-independence:cognitive style across the life span (S, Wapner J, eds), pp Demick105–129. Hillsdale, NJ: Lawrence Erlbaum Associates.
ie KS, Kiemel T, Jeka JJ (2002) Multisensory fusion: simultaneousre-weighting of vision and touch for the control of human posture.Brain Res Cogn Brain Res 14:164–176.
ltman PK (1968) A portable rod-and-frame apparatus. Percept MotSkills 26:503–506.
agano CC (2000) The role of the inertia tensor in kinesthesis. CritRev Biomed Eng 28:231–236.
agano CC, Turvey MT (1995) The inertia tensor as a basis for theperception of limb orientation. J Exp Psychol Hum Percept Perform21:1070–1087.
antev C, Engelien A, Candia V, Elbert T (2001) Representationalcortex in musicians. Plastic alterations in response to musicalpractice. Ann N Y Acad Sci 930:300–314.
aulus W, Straube A, Krafczyk S, Brandt T (1989) Differential effectsof retinal target displacement, changing size and changing dispar-ity in the control of anterior/posterior and lateral body sway. ExpBrain Res 78:243–252.
aulus WM, Straube A, Brandt T (1984) Visual stabilization of posture.Physiological stimulus characteristics and clinical aspects. Brain107:1143–1163.
erennou DA, Amblard B, Laassel el M, Pelissier J (1997) Hemi-spheric asymmetry in the visual contribution to postural control inhealthy adults. Neuroreport 8:3137–3141.
eterka RJ, Loughlin PJ (2004) Dynamic regulation of sensorimotorintegration in human postural control. J Neurophysiol 91:410–423.
euchlin M (1978) Processus vicariants et différences individuelles.Journal de Psychologie 2:133–145.
iccio GE (1995) Coordination of postural control and vehicular con-trol: implications for multimodal perception and stimulation of self-motion. In: Local applications of the ecological approach to human-machine systems (Hanckock P, Flach J, Caird J, Vicente K, eds),pp 123–181. Hillsdale, NJ: Erlbaum Associates.
iccio GE, Martin EJ, Stoffregen TA (1992) The role of balancedynamics in the active perception of orientation. J Exp PsycholHum Percept Perform 18:624–644.
iccio GE, Stoffregen TA (1990) Gravitoinertial force versus the direc-tion of balance in the perception and control of orientation [com-ment]. Psychol Rev 97:135–137.
iley MA, Turvey MT (2002) Variability of determinism in motor be-havior. J Mot Behav 34:99–125.
oll JP, Roll R (1988) From eye to foot: a proprioceptive chain involvedin postural control. In: Posture and gait: development, adaptationand modulation (Amblard B, Berthoz A, Clarac F, eds), pp 155–164. Amsterdam: Elsevier (Biomedical Division).
crivens JE, DeWeerth SP, Ting LH (2008) A robotic device forunderstanding neuromechanical interactions during standing bal-ance control. Bioinspir Biomim 3:26002.
toffregen TA, Riccio GE (1988) An ecological theory of orientationand the vestibular system. Psychol Rev 95:3–14.
treepey JW, Angulo-Kinzler RM (2002) The role of task difficulty inthe control of dynamic balance in children and adults. Hum Mov Sci21:423–438.
treepey JW, Kenyon RV, Keshner EA (2007a) Field of view and baseof support width influence postural responses to visual stimuli
during quiet stance. Gait Posture 25:49–55. ttreepey JW, Kenyon RV, Keshner EA (2007b) Visual motion com-bined with base of support width reveals variable field dependencyin healthy young adults. Exp Brain Res 176:182–187.
undermier L, Woollacott MH, Jensen JL, Moore S (1996) Posturalsensitivity to visual flow in aging adults with and without balanceproblems. J Gerontol A Biol Sci Med Sci 51:M45–M52.
empleton WB (1973) The role of gravitational cues in the judgementof visual orientation. Percept Psychophys 14:451–457.
ing LH, Macpherson JM (2004) Ratio of shear to load ground-reac-tion force may underlie the directional tuning of the automaticpostural response to rotation and translation. J Neurophysiol92:808–823.
an de Langenberg RW, Kingma I, Beek PJ (2007) Perception of limborientation in the vertical plane depends on center of mass ratherthan inertial eigenvectors. Exp Brain Res 180:595–607.
an de Langenberg RW, Kingma I, Beek PJ (2008) The perception oflimb orientation depends on the center of mass. J Exp PsycholHum Percept Perform 34:624–639.
an Wegen EE, van Emmerik RE, Riccio GE (2002) Postural orienta-tion: age-related changes in variability and time-to-boundary. HumMov Sci 21:61–84.
augoyeau M, Viel S, Amblard B, Azulay JP, Assaiante C (2008)Proprioceptive contribution of postural control as assessed fromvery slow oscillations of the support in healthy humans. Gait Pos-ture 27:294–302.
augoyeau M, Viel S, Assaiante C, Amblard B, Azulay JP (2007)Impaired vertical postural control and proprioceptive integrationdeficits in Parkinson’s disease. Neuroscience 146:852–863.
uillerme N, Burdet C, Isableu B, Demetz S (2006) The magnitude ofthe effect of calf muscles fatigue on postural control during bipedalquiet standing with vision depends on the eye-visual target dis-tance. Gait Posture 24:169–172.
uillerme N, Chenu O, Pinsault N, Boisgontier M, Demongeot J,Payan Y (2007) Inter-individual variability in sensory weighting of aplantar pressure-based, tongue-placed tactile biofeedback for con-trolling posture. Neurosci Lett 421:173–177.
uillerme N, Danion F, Marin L, Boyadjian A, Prieur JM, Weise I,Nougier V (2001a) The effect of expertise in gymnastics on pos-tural control. Neurosci Lett 303:83–86.
uillerme N, Teasdale N, Nougier V (2001b) The effect of expertise ingymnastics on proprioceptive sensory integration in human sub-jects. Neurosci Lett 311:73–76.
elch TD, Ting LH (2009) A feedback model explains the differentialscaling of human postural responses to perturbation accelerationand velocity. J Neurophysiol 101:3294–3309.
itkin HA, Demick J, Wapner S (1991) Field dependence-indepen-dence: cognitive style across the life span. Hillsdale, N.J.; London:L. Erlbaum Associates.
itkin HA, Wapner S (1950) Large oscillating visual displays increasepostural instability. Am J Psychol 63:385–392.
occolotti P, Antonucci G, Goodenough DR, Pizzamiglio L, Spinelli D(1992) The role of frame size on vertical and horizontal observersin the rod-and-frame illusion. Acta Psychol (Amst) 79:171–187.
APPENDIX
upplementary data
upplementary data associated with this article can be found, in
he online version, at doi: 10.1016/j.neuroscience.2010.05.072.(Accepted 28 May 2010)(Available online 4 June 2010)




















