
Increased Neurogenesis and Astrogenesis from Neural Progenitor Cells Grafted in the Hippocampus of...
9
Increased Neurogenesis and Astrogenesis from Neural Progenitor Cells Grafted in the Hippocampus of GFAP / Vim / Mice ÅSA WIDESTRAND,JONAS FAIJERSON,ULRIKA WILHELMSSON,PETER L. P. SMITH,LIZHEN LI, CARINA SIHLBOM,PETER S. ERIKSSON,MILOS PEKNY Center for Brain Repair and Rehabilitation, Department of Clinical Neuroscience and Rehabilitation, Institute of Neuroscience and Physiology, Sahlgrenska Academy at Go ¨teborg University, Go ¨teborg, Sweden Key Words. Glial fibrillary acidic protein • Vimentin • Intermediate filaments • Astrocytes • Reactive gliosis ABSTRACT After neurotrauma, ischemia, or neurodegenerative dis- ease, astrocytes upregulate their expression of the inter- mediate filament proteins glial fibrillary acidic protein (GFAP), vimentin (Vim), and nestin. This response, reac- tive gliosis, is attenuated in GFAP / Vim / mice, result- ing in the promotion of synaptic regeneration after neu- rotrauma and improved integration of retinal grafts. Here we assessed whether GFAP / Vim / astrocytes affect the differentiation of neural progenitor cells. In coculture with GFAP / Vim / astrocytes, neural pro- genitor cells increased neurogenesis by 65% and astro- genesis by 124%. At 35 days after transplantation of neural progenitor cells into the hippocampus, adult GFAP / Vim / mice had more transplant-derived neu- rons and astrocytes than wild-type controls, as well as increased branching of neurite-like processes on trans- planted cells. Wnt3 immunoreactivity was readily de- tected in hippocampal astrocytes in wild-type but not in GFAP / Vim / mice. These findings suggest that GFAP / Vim / astrocytes allow more neural progenitor cell-derived neurons and astrocytes to survive weeks after transplantation. Thus, reactive gliosis may adversely af- fect the integration of transplanted neural progenitor cells in the brain. STEM CELLS 2007;25:2619 –2627 Disclosure of potential conflicts of interest is found at the end of this article. INTRODUCTION In the adult central nervous system (CNS), diverse injuries, such as neurotrauma, ischemic damage, and administration of neural grafts, result in astrocyte activation and reactive gliosis. Reac- tive gliosis was proposed to constitute an environment hostile for CNS regeneration [1– 6], but the molecular mechanism re- mains incompletely understood. Upregulation of glial fibrillary acidic protein (GFAP), vimentin (Vim), and nestin and increased abundance of in- termediate filaments are hallmarks of reactive astrocytes [1, 2]. In mice, reactive gliosis can be attenuated by genetic ablation of GFAP and vimentin [7], two intermediate fila- ment proteins that are components of the astrocyte cytoskel- eton. The attenuation of reactive gliosis in GFAP / Vim / mice slows healing after neurotrauma [7]. However, at later stages, these mice display better synaptic recovery [8] and improved axonal regeneration after spinal cord injury [9] or after severing of the optic nerve [10]. Introduction of neural transplants into the CNS also leads to a marked activation of astrocytes [11–13]. We have shown that transplanted retinal cells from postnatal mice up to 30 days of age integrate more robustly in retinas of GFAP / Vim / mice than in those of wild-type mice [14]. These results suggest that GFAP / Vim / astrocytes influence the differentiation of neural progenitor cells and that attenuation of reactive gliosis favors the differentiation and survival of neural progenitor cells in the brain. Two regions of the mammalian brain showing neurogenesis, even in the adult, are the dentate gyrus of the hippocampus and the subventricular zone of the lateral ventricles [15–17]. Hip- pocampal neural progenitor cells can be isolated, propagated, and differentiated into neurons and glial cells and are a potential source of cells for transplantation [11, 18, 19]. However, proper neuronal differentiation, integration, and survival of these cells in adult hosts are major hurdles that must be overcome before transplantation of such cells can be turned into a successful therapeutic paradigm [20, 21]. To evaluate the effect of attenuated reactive gliosis on the efficacy of graft integration, we assessed neurogenesis, astro- genesis, and oligogenesis in vitro. We also compared the num- ber of neurons and astrocytes that differentiated from grafted rat hippocampal neural progenitor cells and survived 35 days after transplantation into the hippocampus of GFAP / Vim / and wild-type mice. Signals provided by the microenvironment sur- rounding the neural progenitor cells regulate their survival, proliferation, and fate commitment, and astrocytes are often a source of such signals [22–24]. It has been shown that Wnt3 is a powerful regulator of neurogenesis from adult hippocampal progenitor cells, both in vitro and in vivo [23]. We therefore explored the expression of this factor in GFAP / Vim / and wild-type mice and in cocultures of rat hippocampal Correspondence: Milos Pekny, M.D., Ph.D., Center for Brain Repair and Rehabilitation, Department of Clinical Neuroscience and Rehabilitation, Institute of Neuroscience and Physiology, Sahlgrenska Academy, Go ¨teborg University, Medicinaregatan 9A, SE-413 90 Go ¨teborg, Sweden. Telephone: 46-31-786-32-69; Fax: 46-31-416108; e-mail: [email protected] Received February 12, 2007; accepted for publication June 27, 2007; first published online in STEM CELLS EXPRESS July 12, 2007. ©AlphaMed Press 1066-5099/2007/ $30.00/0 doi: 10.1634/stemcells.2007-0122 THE STEM CELL NICHE S TEM CELLS 2007;25:2619 –2627 www.StemCells.com
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Increased Neurogenesis and Astrogenesis from Neural Progenitor Cells Grafted in the Hippocampus of...

Increased Neurogenesis and Astrogenesis from Neural ProgenitorCells Grafted in the Hippocampus of GFAP�/�Vim�/� Mice
ÅSA WIDESTRAND, JONAS FAIJERSON, ULRIKA WILHELMSSON, PETER L. P. SMITH, LIZHEN LI,CARINA SIHLBOM, PETER S. ERIKSSON, MILOS PEKNY
Center for Brain Repair and Rehabilitation, Department of Clinical Neuroscience and Rehabilitation,Institute of Neuroscience and Physiology, Sahlgrenska Academy at Goteborg University,Goteborg, Sweden
Key Words. Glial fibrillary acidic protein • Vimentin • Intermediate filaments • Astrocytes • Reactive gliosis
ABSTRACT
After neurotrauma, ischemia, or neurodegenerative dis-ease, astrocytes upregulate their expression of the inter-mediate filament proteins glial fibrillary acidic protein(GFAP), vimentin (Vim), and nestin. This response, reac-tive gliosis, is attenuated in GFAP�/�Vim�/� mice, result-ing in the promotion of synaptic regeneration after neu-rotrauma and improved integration of retinal grafts.Here we assessed whether GFAP�/�Vim�/� astrocytesaffect the differentiation of neural progenitor cells. Incoculture with GFAP�/�Vim�/� astrocytes, neural pro-genitor cells increased neurogenesis by 65% and astro-genesis by 124%. At 35 days after transplantation of
neural progenitor cells into the hippocampus, adultGFAP�/�Vim�/� mice had more transplant-derived neu-rons and astrocytes than wild-type controls, as well asincreased branching of neurite-like processes on trans-planted cells. Wnt3 immunoreactivity was readily de-tected in hippocampal astrocytes in wild-type but not inGFAP�/�Vim�/� mice. These findings suggest thatGFAP�/�Vim�/� astrocytes allow more neural progenitorcell-derived neurons and astrocytes to survive weeks aftertransplantation. Thus, reactive gliosis may adversely af-fect the integration of transplanted neural progenitorcells in the brain. STEM CELLS 2007;25:2619 –2627
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
In the adult central nervous system (CNS), diverse injuries, suchas neurotrauma, ischemic damage, and administration of neuralgrafts, result in astrocyte activation and reactive gliosis. Reac-tive gliosis was proposed to constitute an environment hostilefor CNS regeneration [1–6], but the molecular mechanism re-mains incompletely understood.
Upregulation of glial fibrillary acidic protein (GFAP),vimentin (Vim), and nestin and increased abundance of in-termediate filaments are hallmarks of reactive astrocytes [1,2]. In mice, reactive gliosis can be attenuated by geneticablation of GFAP and vimentin [7], two intermediate fila-ment proteins that are components of the astrocyte cytoskel-eton. The attenuation of reactive gliosis in GFAP�/�Vim�/�
mice slows healing after neurotrauma [7]. However, at laterstages, these mice display better synaptic recovery [8] andimproved axonal regeneration after spinal cord injury [9] orafter severing of the optic nerve [10]. Introduction of neuraltransplants into the CNS also leads to a marked activation ofastrocytes [11–13]. We have shown that transplanted retinalcells from postnatal mice up to 30 days of age integrate morerobustly in retinas of GFAP�/�Vim�/� mice than in those ofwild-type mice [14].
These results suggest that GFAP�/�Vim�/� astrocytesinfluence the differentiation of neural progenitor cells and
that attenuation of reactive gliosis favors the differentiationand survival of neural progenitor cells in the brain. Tworegions of the mammalian brain showing neurogenesis, evenin the adult, are the dentate gyrus of the hippocampus and thesubventricular zone of the lateral ventricles [15–17]. Hip-pocampal neural progenitor cells can be isolated, propagated,and differentiated into neurons and glial cells and are apotential source of cells for transplantation [11, 18, 19].However, proper neuronal differentiation, integration, andsurvival of these cells in adult hosts are major hurdles thatmust be overcome before transplantation of such cells can beturned into a successful therapeutic paradigm [20, 21].
To evaluate the effect of attenuated reactive gliosis on theefficacy of graft integration, we assessed neurogenesis, astro-genesis, and oligogenesis in vitro. We also compared the num-ber of neurons and astrocytes that differentiated from grafted rathippocampal neural progenitor cells and survived 35 days aftertransplantation into the hippocampus of GFAP�/�Vim�/� andwild-type mice. Signals provided by the microenvironment sur-rounding the neural progenitor cells regulate their survival,proliferation, and fate commitment, and astrocytes are often asource of such signals [22–24]. It has been shown that Wnt3 isa powerful regulator of neurogenesis from adult hippocampalprogenitor cells, both in vitro and in vivo [23]. We thereforeexplored the expression of this factor in GFAP�/�Vim�/�
and wild-type mice and in cocultures of rat hippocampal
Correspondence: Milos Pekny, M.D., Ph.D., Center for Brain Repair and Rehabilitation, Department of Clinical Neuroscience andRehabilitation, Institute of Neuroscience and Physiology, Sahlgrenska Academy, Goteborg University, Medicinaregatan 9A, SE-413 90Goteborg, Sweden. Telephone: 46-31-786-32-69; Fax: 46-31-416108; e-mail: [email protected] Received February 12, 2007;accepted for publication June 27, 2007; first published online in STEM CELLS EXPRESS July 12, 2007. ©AlphaMed Press 1066-5099/2007/$30.00/0 doi: 10.1634/stemcells.2007-0122
THE STEM CELL NICHE
STEM CELLS 2007;25:2619–2627 www.StemCells.com

neural progenitor cells and GFAP�/�Vim�/� and wild-typeastrocytes.
MATERIALS AND METHODS
MiceThe GFAP�/�Vim�/� and wild-type mice used for transplantationand in vitro coculture experiments were of a mixed genetic back-ground (C57Bl/6, 129Sv, 129Ola) [25, 26]. For transplantationexperiments, we used 12 GFAP�/�Vim�/� and 13 wild-type femalemice, 4.5–5 months old. To avoid host-versus-graft rejection, thesemice were null mutants for Rag-1 [27]. The null mutations wereconfirmed as described [25–27]. The mice were housed in standardcages in a barrier animal facility and had free access to food andwater. All experimental protocols were approved by the EthicsCommittee of Goteborg University.
Neural Progenitor Cell CulturesNeural progenitor cells derived from adult rat hippocampus andexpressing green fluorescent protein (GFP) [28] were grown onlaminin/poly-L-ornithine-coated Petri dishes in Ham’s F-12 high-glucose medium (Irvine Scientific, Santa Ana, CA, http://www.irvinesci.com) with 1% N2 supplement (Gibco, Paisley, U.K.,http://www.invitrogen.com), 2 mM L-glutamine (Gibco), 20 ng/mlrecombinant human basic fibroblast growth factor (BD Biosciences,Erembodegem, Belgium, http://www.bdbiosciences.com), penicil-lin/streptomycin (100 U/0.1 mg/ml; Gibco), and Fungizone (0.25�g/ml; Bristol-Myers Squibb, Princeton, NJ, http://www.bms.com).The cells were cultured at 37°C in 5% CO2.
Assessment of Neural Progenitor Cell DifferentiationIn VitroPrimary cultures of astrocytes from 1-day-old GFAP�/�Vim�/� andwild-type mice were prepared as described [29]. At confluence,primary cultures were passaged (1:2) onto coverslips and cultured innormal serum-containing medium. Confluent cultures were washedtwice with serum-free medium (Dulbecco’s modified Eagle’s me-dium/Nut Mix F12 [1:1; Gibco], 2 mM L-glutamine, 1% N2 sup-plement, 1% penicillin/streptomycin [100 U/0.1 mg/ml; Gibco], and0.25 �g/ml Fungizone [Bristol-Myers Squibb]) and cultured inserum-free medium or medium supplemented with 1% fetal calfserum (Gibco).
The astrocyte cultures were seeded with GFP-expressing neuralprogenitor cells [24] at approximately 2.0 � 103 cells per cm2. At 6days, lineage selection was assayed. Briefly, cells were fixed (4%paraformaldehyde in phosphate-buffered saline [PBS], 4°C, 10 min-utes), nonspecific binding was blocked, and cells were immunocyto-chemically stained (1 hour, room temperature) with mouse anti-micro-tubule-associated protein 2 subunits a and b (anti-Map2ab) antibodies(1:100; Sigma-Aldrich, St. Louis, http://www.sigmaaldrich.com), rab-bit anti-GFAP antibodies (1:500; DAKO, Glostrup, Denmark, http://www.dako.com), and mouse anti-Rip antibodies (1:50; DevelopmentalStudies Hybridoma Bank, Iowa City, IA, http://www.uiowa.edu/�dshbwww) in PBS containing 3% donkey serum (Jackson Immu-noresearch Laboratories, West Grove, PA, http://www.jacksonimmuno.com) and 0.05% saponin (Sigma-Aldrich). After three washes in PBS,cells were incubated for 1 hour at room temperature with Alexa Fluor555-conjugated donkey anti-mouse or anti-rabbit secondary antibodies(1:2,000; Molecular Probes, Eugene, OR, http://www.probes.invitrogen.com) and the nuclear dye bisbenzimide from a stock at 5 �g/ml(Hoechst 33258, 1:80; Sigma-Aldrich).
Differentiation was assessed by counting GFPpos, GFPposMap2abpos,GFPposRippos, and GFPposGFAPpos cells in at least 12 randomlyselected, nonoverlapping fields using a Nikon Eclipse 80i micro-scope (Nikon, Preston, U.K., http://www.nikon.com). All differen-tiation experiments were done in triplicate (n � 4), and 200–500cells were counted in each field. Cell counts were performed onblinded samples.
For the analysis of Wnt3 expression, confluent astrocyte cul-tures and cocultures with neural progenitor cells (2.0 � 103 cells per
cm2) in 1% or 10% of fetal calf serum were fixed and immunocy-tochemically stained with goat anti-Wnt3 antibodies (1:50; SantaCruz Biotechnology Inc., Santa Cruz, CA, http://www.scbt.com)and rabbit anti-GFAP antibodies (1:500; DAKO) in PBS containing3% donkey serum and 0.1% Triton X-100 (Merck, Darmstadt,Germany, http://www.merck.com). After three washes in PBS, cellswere incubated for 1 hour at room temperature with Alexa Fluor488-conjugated donkey anti-mouse and Alexa Fluor 555-conjugateddonkey anti-goat antibodies (1:2,000; Molecular Probes). The per-centage of astrocytes displaying the filamentous pattern of Wnt3immunoreactivity was assessed by investigating 200 wild-type and200 GFAP�/�Vim�/� astrocytes. All images were processed inPhotoshop (version 8.0; Adobe Systems Inc., San Jose, CA, http://www.adobe.com) and ImageJ (version 1.30, National Institutes ofHealth, Bethesda, MD, http://www.nih.gov).
Neural Progenitor Cell TransplantationNeural progenitor cells were grown to confluence in the presence of5 �M 5-bromo-2�-deoxyuridine (BrdU) (Sigma-Aldrich) for 48hours, trypsinized, washed, resuspended (60,000 cells per microli-ter) in PBS with 0.15% glucose and 30 ng/ml of basic fibroblastgrowth factor, and kept on ice until grafting. The mice were anes-thetized with avertin and placed in a stereotactic frame, and 1.0 �lof cell suspension was slowly injected (2 minutes) unilaterally intothe hippocampus (mediolateral, �3.0 mm; anteriorposterior, �3.5mm; dorsoventral, �2.7 mm) with a 5-�l Hamilton syringe, whichwas left in place for an additional 2 minutes to prevent reflux of thecell suspension. The syringe was slowly raised 0.5 mm, 0.2 �l wasinjected, and the syringe was left in place for 1 minute. After 7hours on ice, viability of the remaining cell suspension was 90% asassessed by trypan blue exclusion.
Grafted cells were identified with rat anti-BrdU antibodies(1:100; Nordic BioSite, Taby, Sweden, http://www.biosite.se) fol-lowed by goat anti-rat Alexa Fluor 488 antibodies (1:500; Molec-ular Probes). To visualize astrocytes, we used mouse anti-GFAPantibodies (1:100; Sigma-Aldrich) and rabbit antibodies against theendothelin B receptor (ETBR) (1:100; Alomone Labs, Jerusalem,http://www.alomone.com), followed by Alexa Fluor 488 or 568goat anti-mouse or Alexa Fluor 488 donkey anti-mouse and AlexaFluor 568 goat anti-rabbit antibodies (1:500; Molecular Probes),respectively. To assess the number of immature astrocytes, we usedrabbit anti-S100 antibodies (1:200; DAKO), followed by biotinyl-ated donkey anti-rabbit antibodies (1:200; Jackson ImmunoresearchLaboratories) and streptavidin conjugated with Cy3 (1:100; Sig-ma-Aldrich) or mouse anti-glutamine synthase (GS) antibodies(1:100; Chemicon, Temecula, CA, http://www.chemicon.com)followed by biotinylated rabbit anti-mouse antibodies (1:400;DAKO) and streptavidin conjugated with Cy3 (1:100; Sigma-Aldrich). To visualize oligodendrocytes, we used mouse anti-Ripantibodies (1:20; Developmental Studies Hybridoma Bank), fol-lowed by biotinylated donkey anti-mouse antibodies (1:200;Jackson Immunoresearch Laboratories) and streptavidin conju-gated with Alexa 594 (1:1,000; Molecular Probes). To visualizeastrocyte and oligodendrocyte progenitors, we used goat anti-Olig2 antibodies (1:200; R&D Systems Inc., Minneapolis, http://www.rndsystems.com) followed by biotinylated donkey anti-goat antibodies (1:200; Jackson Immunoresearch Laboratories)and streptavidin conjugated with Cy3 (1:200; Sigma-Aldrich).To assess neuronal differentiation, we used biotinylated mouseanti-NeuN antibodies (1:100; Chemicon) followed by streptavi-din conjugated with Cy3 (1:100; Sigma-Aldrich). To study Wnt3immunoreactivity, we used goat anti-Wnt3 antibodies (1:50;Santa Cruz Biotechnology) followed by biotinylated donkeyanti-goat antibodies (1:200; Jackson Immunoresearch Laborato-ries) and streptavidin conjugated with Cy3 (1:300; Sigma-Al-drich). We used TO-PRO-3 (1:1,000; Molecular Probes) to coun-terstain cell nuclei.
Antibodies were diluted in a buffer containing 1% bovineserum albumin and 0.01% Tween 20 in PBS and mounted inMowiol (Clariant GmbH, Frankfurt, Germany, http://www.clariant.com). The sections were incubated with primary anti-bodies overnight at 4°C and with secondary antibodies for 4hours at room temperature.
2620 Neural Grafting in GFAP�/�Vim�/� Mice
To study the survival and differentiation of transplanted neuralprogenitor cells, BrdUpos transplanted neural progenitor cells werecounted in the subgranular zone, granular cell layer, and Ammon’shorn of the hippocampus. In each mouse, seven 35-�m horizontalsections through the hippocampus at 140-�m intervals were exam-ined with a Nikon Eclipse 80i epifluorescence microscope (Nikon).On the same sections, BrdUpos cells were counted in a 70,000-�m2
arc 250 �m away from the needle track and in a 120,000-�m2 arc500 �m away from the needle track. For each mouse, 25 BrdUpos
cells in the subgranular zone and 50 BrdUpos cells in the granularcell layer were examined for S100 and GS positivity (astroglialmarkers) and NeuN positivity (neuronal marker) by laser-scan-ning confocal microscopy (Leica TCS, Heidelberg, Germany,http://www.leica.com). BrdUpos cells in the granular cell layerand the subgranular zone in one and two sections per mouse wereexamined for Rip and Olig2 positivity, respectively, by laser-scanning confocal microscopy. All images were processed inPhotoshop (version 8.0; Adobe Systems) and ImageJ (version1.30; NIH).
GFPpos cells in the molecular layer with processes exceedingthree cell-body lengths were counted in seven 35-�m-thick hori-zontal hippocampal sections at 140-�m intervals; the longest pro-cess in each cell was measured, and its primary branches werecounted (Fig. 5D). We used laser-scanning confocal microscopy toproduce images at 1-�m intervals throughout the sections.
SDS-Polyacrylamide Gel Electrophoresis andWestern Blot AnalysisWestern blot was used to analyze Wnt3 protein expression in vivoand in vitro. Hippocampi from five wild-type and four GFAP�/�
Vim�/� mice, 18 months of age, were homogenized in 500 �l oflysis buffer (50 mM dithiothreitol, 25 mM Tris HCl, 35 mM Trisbase, 0.5% lithium dodecyl sulfate, 2.5% glycerol, 12.5 mM ethyl-enediaminetetraacetic acid [EDTA] and proteinase inhibitor [Com-plete Mini cocktail tablet; Roche Diagnostics, Basel, Switzerland,http://www.roche-applied-science.com]) using a motor-drivengrinder pestle. The samples were sonicated for 1 minute, agitatedfor 2 hours, and heated to approximately 80°C for 3 minutes. Thecrude protein extract was centrifuged at 13,000 rpm for 15 minutes.Primary astrocyte cultures from five wild-type and four GFAP�/�
Vim�/� mice, 1 day old, and cocultures from these cells and neuralprogenitors were lysed in lysis buffer and sonicated for 1 minute.For all samples, total protein concentration was determined withthe Bradford method using Coomassie Plus (Pierce, Rockford,IL, http://www.piercenet.com) and measuring absorbance at595 nm.
The sample volume (15–25 �l) loaded on the NuPAGE NovexBis-Tris 10% acrylamide gel (Invitrogen, Carlsbad, CA, http://
www.invitrogen.com) was adjusted to gain an equal loading of 30�g of total protein. One-dimensional gel electrophoresis was per-formed using 3-(N-morpholino)propanesulfonic acid (MOPS) SDSrunning buffer (Invitrogen) at a constant voltage of 200 V. Forquantitative Western blot analysis, protein samples were transferredfrom gels to polyvinylidene difluoride membranes (Bio-Rad, Her-cules, CA, http://www.bio-rad.com). The membranes were blocked,washed, and incubated with goat anti-Wnt3 antibodies (1:200; SantaCruz Biotechnology) or goat anti-�-actin antibodies (1:500; Abcam,Cambridge, U.K., http://www.abcam.com) followed by severalwashes and incubation with iodine-125-conjugated donkey anti-goat antibodies (1:1,000; Sigma-Aldrich). After exposure to photo-graphic film, the relative intensities of Wnt3 and �-actin proteinbands were analyzed using ImageJ (NIH).
Statistical AnalysisValues are presented as mean � SEM. Differences were evaluatedby two-tailed t test; p � .05 was considered significant.
RESULTS
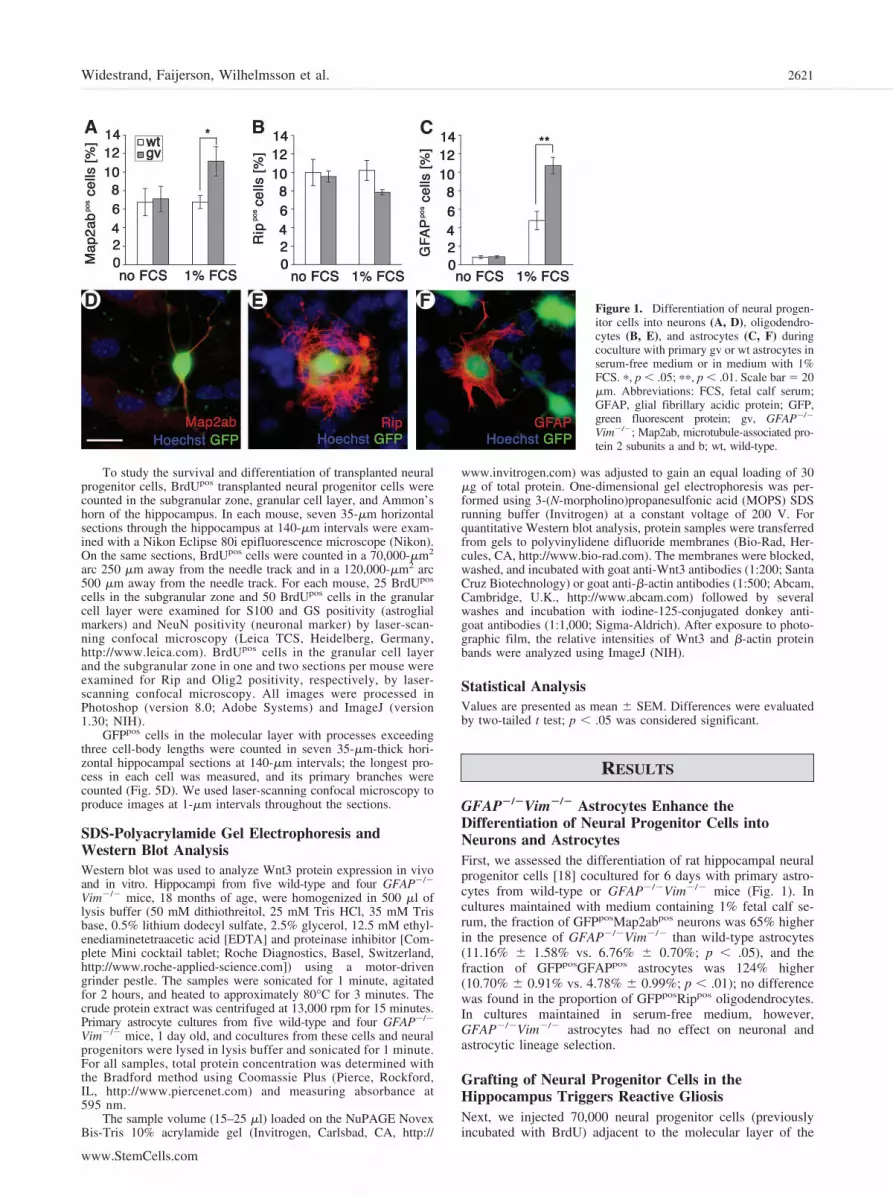
GFAP�/�Vim�/� Astrocytes Enhance theDifferentiation of Neural Progenitor Cells intoNeurons and AstrocytesFirst, we assessed the differentiation of rat hippocampal neuralprogenitor cells [18] cocultured for 6 days with primary astro-cytes from wild-type or GFAP�/�Vim�/� mice (Fig. 1). Incultures maintained with medium containing 1% fetal calf se-rum, the fraction of GFPposMap2abpos neurons was 65% higherin the presence of GFAP�/�Vim�/� than wild-type astrocytes(11.16% � 1.58% vs. 6.76% � 0.70%; p � .05), and thefraction of GFPposGFAPpos astrocytes was 124% higher(10.70% � 0.91% vs. 4.78% � 0.99%; p � .01); no differencewas found in the proportion of GFPposRippos oligodendrocytes.In cultures maintained in serum-free medium, however,GFAP�/�Vim�/� astrocytes had no effect on neuronal andastrocytic lineage selection.
Grafting of Neural Progenitor Cells in theHippocampus Triggers Reactive GliosisNext, we injected 70,000 neural progenitor cells (previouslyincubated with BrdU) adjacent to the molecular layer of the
Figure 1. Differentiation of neural progen-itor cells into neurons (A, D), oligodendro-cytes (B, E), and astrocytes (C, F) duringcoculture with primary gv or wt astrocytes inserum-free medium or in medium with 1%FCS. �, p � .05; ��, p � .01. Scale bar � 20�m. Abbreviations: FCS, fetal calf serum;GFAP, glial fibrillary acidic protein; GFP,green fluorescent protein; gv, GFAP�/�
Vim�/�; Map2ab, microtubule-associated pro-tein 2 subunits a and b; wt, wild-type.
2621Widestrand, Faijerson, Wilhelmsson et al.
www.StemCells.com

dentate gyrus in wild-type and GFAP�/�Vim�/� mice. At 35days, the transplanted cells had spread into the hippocampi ofmice in both groups (Fig. 2A, 2B, 2E, 2G). Dozens of GFPpos
cells and many more BrdUpos cells were visible on each sectionthat included the region where the neural progenitor cells wereoriginally grafted (Figs. 2A–2E, 2G, 3C).
Astrocyte reactivity was visualized by immunodetection oftwo markers of reactive astrocytes, GFAP [1, 2, 30] and ETBR[31]. In wild-type mice, astrocytes in the hippocampus aroundtransplanted cells were highly positive for GFAP (Fig. 2A, 2C)and showed prominent ETBR immunoreactivity that colocalizedwith GFAP immunoreactivity (Fig. 2E; data not shown). NoETBR immunoreactivity was observed in GFAP�/�Vim�/�
mice (Fig. 2G, 2H). Nonreactive astrocytes in the contralateraldentate gyrus of the hippocampus showed no ETBR immu-noreactivity in GFAP�/�Vim�/� mice and weak immunore-activity in wild-type mice (Fig. 2F, 2H). As expected, residentastrocytes in GFAP�/�Vim�/� mice were GFAP-negative (Fig.2B, 2D).
GFAP�/�Vim�/� Mice Have More NeuralProgenitor Cells 35 Days After GraftingTo study the survival and migration of grafted neural progeni-tors, we assessed the distribution of BrdUpos cells in the hip-pocampus. Survival of transplanted cells in the subgranularzone, granular cell layer, and Ammon’s horn did not differsignificantly between GFAP�/�Vim�/� and wild-type mice(p � .081, p � .129, and p � .074, respectively) (Fig. 3A). At250 and 500 �m from the injection site (Fig. 3C), however,transplanted neural progenitor cells were somewhat more nu-merous in GFAP�/�Vim�/� mice (799 � 31 vs. 685 � 42; p �.05) (Fig. 3B).
GFAP�/�Vim�/� Mice Have More Graft-DerivedS100pos AstrocytesTo assess the differentiation of grafted neural progenitors intoastroglial cells, we used antibodies against S100, a marker ofimmature and mature astrocytes [32, 33], and GS, a markerof mature astrocytes [34]. At 35 days after grafting, the number ofBrdUposS100pos cells was 91% higher in the granular cell layer
Figure 2. Grafting of neural progenitor cells in the hippocampal regiontriggered reactive gliosis. (A): Astrocytes were strongly GFAPpos in thevicinity of BrdU-labeled neural progenitor cells in the dentate gyrus ofwt mice 35 days after transplantation. (B): Neural progenitor cells in thedentate gyrus of gv mice; no GFAP immunoreactivity was detected.Dashed line (A, B) indicates the granular cell layer. (C, D): Higher-magnification image shows GFPpos transplanted cells in wt (C) and gvmice (D) surrounded in wt mice by reactive GFAPpos astrocytes. ETBRimmunoreactivity marks reactive astrocytes in the grafted and thecontralateral dentate gyrus of wt mice (E, F) but not gv mice (G, H).Scale bars � 100 �m (A, B, E–H) and 10 �m (C, D). Abbreviations:BrdU, 5-bromo-2�-deoxyuridine; contra, contralateral; ETBR, endo-thelin B receptor; GFAP, glial fibrillary acidic protein; GFP, greenfluorescent protein; gv, GFAP�/�Vim�/�; ipsi, ipsilateral; wt,wild-type.
Figure 3. Survival and migration of neural progenitor cells in wt andgv mice 35 days after transplantation into the hippocampus. (A): Sur-vival of graft-derived cells in the sgz, gcl, and ah of the dentate gyrus.(B): Migration of grafted cells assessed by counting BrdUpos cells 250and 500 �m away from the transplantation site (the areas indicated in[C]). �, p � .05. Scale bar � 100 �m. Abbreviations: ah, Ammon’s horn;BrdU, 5-bromo-2�-deoxyuridine; gcl, granular cell layer; gv, GFAP�/�
Vim�/�; sgz, subgranular zone; wt, wild-type.
2622 Neural Grafting in GFAP�/�Vim�/� Mice

of GFAP�/�Vim�/� than wild-type mice (91.2 � 13 vs. 47.8 �12; p � .05); no difference was found in the subgranular zone(Fig. 4A). In both regions combined, there were 83% moreBrdUposS100pos astrocytes in GFAP�/�Vim�/� mice (118 � 16vs. 64.8 � 15; p � .05).
The percentage of graft-derived S100pos astrocytes was 61%higher in the granular cell layer in GFAP�/�Vim�/� mice(20.1% � 2.3% vs. 12.5% � 2.1%; p � .05) and 50% higher inthe subgranular zone and granular cell layer combined (23.3% �2.3% vs. 15.5% � 2%; p � .05) (Fig. 4B). The percentage ofS100pos cells in the subgranular zone did not differ in wild-typeand GFAP�/�Vim�/� mice (Fig. 4B).
The number and percentage of graft-derived GSpos matureastrocytes in the granular cell layer and subgranular zone werecomparable between groups (Fig. 4D, 4E). However, GFAP�/�
Vim�/� mice tended to have more of these cells in the subgranularzone (4.1 � 0.9 vs. 1.9 � 0.7; p � .06) (Fig. 4D).
The differentiation of grafted neural progenitors intoOlig2pos cells (astrocyte progenitors and oligodendrocyte pro-genitors [35, 36]) was assessed by using antibodies againstOlig2. Thirty-five days after grafting, the number ofBrdUposOlig2pos cells in the granular cell layer was 78% higherin GFAP�/�Vim�/� mice than in wild-type mice (14.9 � 2.5vs. 8.3 � 0.8; p � .05), whereas no difference was observed inthe subgranular zone (Fig. 4G). Combined data from both re-gions displayed a 69% increase in the number ofBrdUposOlig2pos cells in GFAP�/�Vim�/� mice (21.1 � 3.2 vs.12.5 � 1.0; p � .05). The percentage of graft-derivedBrdUposOlig2pos cells was 84% higher in the granular cell layerof GFAP�/�Vim�/� mice compared with wild-type mice(11.0 � 1.7 vs. 6.0 � 0.8; p � .05); no difference was observedin the subgranular zone (Fig. 4H). When both regions werecombined, 74% more BrdUposOlig2pos cells were observed inGFAP�/�Vim�/� mice (22.5 � 3.12 vs. 12.9 � 1.5; p � .05).Consistent with the quantification of oligodendrocyte differ-entiation in vitro, no difference in the percentage of Rippos
oligodendrocytes was observed between wild-type andGFAP�/�Vim�/� mice (2.1% � 0.9% and 3.7% � 0.9%,respectively; p � .23).
GFAP�/�Vim�/� Mice Have More Graft-DerivedNeuronsNext, we assessed the differentiation of grafted neural progenitorcells into mature neurons by counting BrdUposNeuNpos cells 35days after grafting (Fig. 5). In the granular cell layer, BrdUpos
NeuNpos cells were 45% more numerous in GFAP�/�Vim�/�
mice (208 � 15 vs. 144 � 26; p � .05), but no difference was seenin the subgranular zone (21.0 � 2.8 in GFAP�/�Vim�/� vs.13.7 � 3.2 in wild-type; p � .113). In the two areas combined,mature neurons were 51% more numerous in GFAP�/�Vim�/�
mice (229 � 17 vs. 151.7 � 28; p � .05). There was no differencein the percentage of transplant-derived neurons in the granular celllayer (44.2% � 2.6% in GFAP�/�Vim�/� vs. 37.1% � 3.6% inwild-type; p � .15).
Transplanted Cells Have More Branching ofNeurite-Like Processes in GFAP�/�Vim�/� MiceTo assess the differentiation of graft-derived cells, we evaluatedthe morphology of GFPpos cells in the molecular layer near thetransplantation site that were GFPpos at 35 days (approximately3% of graft-derived cells; Fig. 5D) and whose longest cellularprocesses extended over three cell-body lengths. GFPpos cellstended to have longer neurite-like processes in GFAP�/�Vim�/�
mice (56.7 � 4.0 vs. 45.8 � 3.6 �m; p � .08). The number of cellswith neurite-like processes longer than three cell-body lengths and
Figure 4. Differentiation of grafted neural progenitor cells in the sgzand gcl of the dentate gyrus of the hippocampus 35 days aftertransplantation. Shown are the number (A) and percentage (B) ofgrafted cells that were positive for S100 (BrdUposS100pos), indicatingimmature astroglial cells and mature astrocytes, in wt and gv mice.(C): Orthogonal sections showing BrdUposS100pos cells. Shown arethe number (D) and percentage (E) of grafted cells that were positivefor GS (BrdUposGSpos), indicating mature astrocytes, in wt and gvmice. (F): Orthogonal sections showing BrdUposGSpos cells. Shownare the number (G) and percentage (H) of grafted cells that werepositive for Olig2 (BrdUposOlig2pos), indicating astrocyte progeni-tors and oligodendrocyte progenitors, in wt and gv mice. (I): Or-thogonal sections showing BrdUposOlig2pos cells. �, p � .05. Scalebars � 10 �m. Abbreviations: BrdU, 5-bromo-2�-deoxyuridine; gcl,granular cell layer; GS, glutamine synthase; gv, GFAP�/�Vim�/�;sgz, subgranular zone; wt, wild-type.
2623Widestrand, Faijerson, Wilhelmsson et al.
www.StemCells.com

at least one branch was twice as high in GFAP�/�Vim�/� mice(Fig. 5E, 5F).
Wnt3 Immunoreactivity Is Detected in HippocampalAstrocytes of Wild-Type but Not GFAP�/�Vim�/�
MiceUsing immunohistochemistry and Western blot analysis, weassessed the expression and distribution of Wnt3 protein, animportant regulator of adult hippocampal neurogenesis [23], inthe hippocampus of nongrafted wild-type and GFAP�/�Vim�/�
mice. In wild-type hippocampus, there was a distinct Wnt3immunoreactivity associated with astrocytes and blood vessels(Fig. 6A, 6C, 6E). In contrast, Wnt3 immunoreactivity wasabsent from hippocampal astrocytes of GFAP�/�Vim�/�
mice, whereas Wnt3 immunoreactivity associated with bloodvessels was retained (Fig. 6B, 6D, 6F). Western blot analysisof Wnt3 showed comparable amounts of the Wnt3 protein inthe hippocampus of wild-type and GFAP�/�Vim�/� mice(Fig. 6G, 6H).
These data suggested that the distribution of Wnt3 may bealtered in intermediate filament-free hippocampal astrocytes ofGFAP�/�Vim�/� mice. Immunocytochemical analysis was per-formed on primary astrocytes and cocultures of neural progen-itor cells and astrocytes from wild-type and GFAP�/�Vim�/�
mice. The majority of wild-type and GFAP�/�Vim�/� astro-cytes exhibited diffuse Wnt3 immunostaining throughout thecell cytoplasm. However, in approximately 3%–10% of wild-type astrocytes, Wnt3 immunoreactivity had a filamentous ap-pearance and colocalized, at least partially, with GFAP immu-noreactivity (Fig. 7A–7F). This filamentous pattern of Wnt3immunoreactivity was even more apparent when the fetal calfserum concentration was increased from 1% to 10% (Fig. 7G–7I). Quantification of astrocytes displaying such a filamentouspattern of Wnt3 immunoreactivity in primary astrocyte culturesin the presence of 10% serum showed 37% of the wild-typeastrocytes displaying such a pattern compared with 2% ofGFAP�/�Vim�/� astrocytes. Western blot analysis of Wnt3showed comparable amounts of the protein in wild-type andGFAP�/�Vim�/� astrocytes (0.42 � 0.08 AU and 0.50 � 0.10
Figure 5. Neuronal differentiation of grafted neural progenitor cells in the sgz and gcl of the dentate gyrus of the hippocampus 35 days aftertransplantation. Shown are the number (A) and percentage (B) of grafted cells that were positive for the neuronal marker NeuN (BrdUposNeuNpos)in wt and gv mice. (C): Orthogonal sections showing BrdUposNeuNpos cells. Shown are transplanted GFPpos cells with neurite-like cellular processesand their branching in wt and gv mice. (D, E): At 35 days after transplantation, the number of GFPpos cells in the molecular layer of the hippocampuswhose longest neurite-like cellular process exceeded three body lengths and had one or more primary branches was twice as high in gv mice (D) thanin wt mice (E). (F): The distribution of such cells with different number of primary branches in wt and gv mice. �, p � .05. Scale bars � 10 �m.Abbreviations: BrdU, 5-bromo-2�-deoxyuridine; gcl, granular cell layer; GFP, green fluorescent protein; gv, GFAP�/�Vim�/�; sgz, subgranular zone;wt, wild-type.
2624 Neural Grafting in GFAP�/�Vim�/� Mice

AU, respectively) and in cocultures of neural progenitor cells andastrocytes (2.55 � 0.29 AU and 2.52 � 0.05 AU, respectively).
DISCUSSION
In adults, cells grafted to the CNS survive and integrate [21] butwith low efficiency [37, 38]. Transplantation of adult neuralprogenitor cells into different brain regions has revealedneurogenic and non-neurogenic niches, underscoring the in-fluence of environment on the fate of grafted cells [11, 33,39]. Astrocytes from neurogenic regions of adult brain providepermissive or inductive environmental cues directing differen-tiation of neural progenitor cells [24]. Reactive gliosis has beenproposed to inhibit neuroregeneration after CNS injury andendogenous neurogenesis [3, 6, 40]. GFAP�/�Vim�/� mice,which have attenuated reactive gliosis [5, 7], exhibit in-creased axonal plasticity and functional recovery after spinalcord injury [9], improved synaptic regeneration after trauma[8], improved regeneration of severed optic nerve axons [10],better integration of neural grafts [14], and enhanced neuralprogenitor cell proliferation and improved neurogenesis inthe hippocampus in old age [41].
In this study, when cocultured with neural progenitor cells,GFAP�/�Vim�/� astrocytes enhanced neurogenesis by 65%and astrogenesis by 124%. In GFAP�/�Vim�/� mice, neuronsderived from neural progenitors showed improved survival andenhanced branching of neurite-like cellular processes 35 daysafter transplantation into the hippocampus. These findings sug-gest that the survival and differentiation of transplanted neuralprogenitor cells can be improved by modifying the astroglialenvironment of the host. In the presence of 1% serum to mimicconditions leading to astrocyte activation and reactive gliosis invivo, the number of astrocytes derived from neural progenitorcells increased in cocultures with wild-type or GFAP�/�Vim�/�
astrocytes, as reported before [28]. However, such astrocyteswere more than twice as abundant in cocultures with GFAP�/�
Vim�/� astrocytes, and neurogenesis was increased in cocultureswith GFAP�/�Vim�/� in the presence of serum. These findingssuggest that nonreactive astrocytes (i.e., in the absence of serum)lacking intermediate filaments have the same effect on neurogen-esis and astrogenesis as wild-type astrocytes; however, when acti-vated by serum factors, they increase the rate of neurogenesis andastrogenesis, either by increased permissiveness or instruction ofneural progenitor cell differentiation.
Previously, we reported that the retinas of GFAP�/�Vim�/�
mice are more permissive for the integration of retinal grafts[14], as shown by a sixfold increase in the number of graftedcells that migrated, differentiated into mature neurons, andintegrated into the ganglion cell layer. Consistent with thoseresults, the number of graft-derived neurons in the neurogenicsubgranular zone and granular cell layer of the hippocampuswas 50% higher in GFAP�/�Vim�/� mice than in wild-typemice. Moreover, at 35 days after grafting, the molecular layer inGFAP�/�Vim�/� mice had twice as many graft-derived cellswith long and branched neurite-like processes, implying thatthey were mature neurons. Thus, reactive gliosis may negativelyinfluence the fate of neural progenitor cells, both in culture andafter grafting, and attenuation of reactive gliosis may promoteneurogenesis from transplanted cells irrespective of whether thetransplants are immature retinal cells [14] or homogenous pop-ulations of neural progenitor cells (this study).
In the granular cell layer, the number of graft-derivedS100pos immature and mature astrocytes was 90% higher inGFAP�/�Vim�/� mice than wild-type mice. In both groups, asimilarly small fraction of these cells was GSpos, a marker ofmature astrocytes, but GFAP�/�Vim�/� mice tended to havemore GSpos cells in the subgranular zone (p � .06). Thissuggests that attenuation of reactive gliosis increases astrogen-esis, but few S100pos astrocytes complete their differentiation.
Figure 6. Wnt3 immunoreactivity was detected in astrocytes in thedentate gyrus of the hippocampus of wt (A, C) but not gv mice (B, D).(C): Higher magnification images show that astrocytes were Wnt3-positive only in wt mice, whereas Wnt3 immunoreactivity was associ-ated with blood vessels in both wt and gv mice (C, D). Astrocytespositive for GS showed immunoreactivity for Wnt3 and GFAP in wtmice (E) but not in gv mice (F). Western blot analysis showed compa-rable levels of Wnt3 in the hippocampus of wt and gv mice (G, H).Equal protein loading was confirmed by immunoblotting for �-actin (G,I). Scale bars � 100 �m (A, B) and 25 �m (C–F). Abbreviations:GFAP, glial fibrillary acidic protein; GS, glutamine synthase; gv,GFAP�/�Vim�/�; TOPRO, TO-PRO-3; wt, wild-type.
2625Widestrand, Faijerson, Wilhelmsson et al.
www.StemCells.com

Consistent with this scenario and with the paucity of transplant-derived GFAPpos cells in the dentate gyrus [11, 39], graft-derived GFAPpos cells were virtually absent in both wild-typeand GFAP�/�Vim�/� mice (not shown). In the granular celllayer, the number of graft-derived Olig2pos cells (astrocyte pro-genitors and oligodendrocyte progenitors) was 78% higher inGFAP�/�Vim�/� mice than in wild-type mice. Similar to thelow number of fully mature graft-derived astrocytes in the hosts,the number of Rippos oligodendrocytes was very low but com-parable in GFAP�/�Vim�/� and wild-type hosts.
We found that a proportion of wild-type astrocytes in pri-mary cultures or cocultures with neural progenitor cells exhib-ited a filamentous pattern of Wnt3 immunoreactivity, whichseemed, at least to some extent, to colocalize with bundles ofintermediate filaments. Astrocytes in the hippocampus of wild-type mice were clearly Wnt3-positive, but the Wnt3 immuno-staining was undetectable in astrocytes in the hippocampus ofGFAP�/�Vim�/� mice, suggesting that Wnt3 associates withintermediate filaments also in vivo. It is possible that the asso-ciation of Wnt3 with cytoskeletal components in astrocytes isimportant for the intracellular distribution of Wnt3 and mightaffect its secretion and consequently its effect on neuralprogenitor cells. Hippocampal neural progenitor cells wereshown to express receptor and signaling components for Wntproteins, and increased levels of Wnt3 were demonstrated toincrease neurogenesis from hippocampal neural progenitorcells both in vitro and in vivo [23]. Thus, the differentialdistribution of Wnt3 in wild-type and GFAP�/�Vim�/� as-trocytes could contribute to the increased neurogenesis fromneural progenitor cells observed in GFAP�/�Vim�/� cocul-tures or when grafted into GFAP�/�Vim�/� hosts comparedwith wild-type.
We conclude that inhibition of reactive astrocytes facilitates thegeneration of mature neurons and astrocytes from hippocampalneural progenitor cells in vitro and in vivo. The survival anddifferentiation of transplanted neural progenitor cells might beimproved by modifying the astroglial environment of the host.
ACKNOWLEDGMENTS
We dedicate this article to our great colleague Professor PeterEriksson, a pioneer in the field of adult human neural stem cells,who died on August 2, 2007, at only 48 years old. We thank Dr.Fred Gage for providing us with neural progenitor cells, Dr.Marcela Pekna for comments on the manuscript, Dr. SusannTeneberg for iodine conjugation, and the Swegene Center forCellular Imaging at Goteborg University. This work was sup-ported by grants from the Swedish Medical Research Council(Projects 11548 and 5174), Swedish Stroke Foundation, Torstenand Ragnar Soderberg Foundations, the region of Vastra Gota-land (RUN), Frimurare Foundation, the Swedish Society forMedicine, W. and M. Lundgren Foundation, Tornspiran Foun-dation, John and Brit Wennerstrom’s Foundation for Neurolog-ical Research, Foundation Edit Jacobson’s Donation Fund,Trygg-Hansa, Hjarnfonden, and ALF Goteborg.
DISCLOSURE OF POTENTIAL CONFLICTS
OF INTEREST
The authors indicate no potential conflicts of interest.
Figure 7. In primary cultures of wt astro-cytes, Wnt3 immunoreactivity had a fila-mentous appearance and showed partial co-localization with bundles of GFAP-positiveintermediate filaments both in 1% (A–C)and 10% (G–I) fetal calf serum. In contrast,intermediate filament-free gv astrocytes ex-hibited a diffuse nonfilamentous pattern ofWnt3 immunoreactivity (D–F). Scale bars �20 �m. Abbreviations: GFAP, glial fibrillaryacidic protein; gv, GFAP�/�Vim�/�; wt,wild-type.
2626 Neural Grafting in GFAP�/�Vim�/� Mice

REFERENCES
1 Hatten ME, Liem RK, Shelanski ML et al. Astroglia in CNS injury. Glia1991;4:233–243.
2 Eddleston M, Mucke L. Molecular profile of reactive astrocytes—Impli-cations for their role in neurologic disease. Neuroscience 1993;54:15–36.
3 Ridet JL, Malhotra SK, Privat A et al. Reactive astrocytes: Cellular andmolecular cues to biological function. Trends Neurosci 1997;20:570–577.
4 Fawcett JW, Asher RA. The glial scar and central nervous system repair.Brain Res Bull 1999;49:377–391.
5 Pekny M, Pekna M. Astrocyte intermediate filaments in CNS pathologiesand regeneration. J Pathol 2004;204:428–437.
6 Silver J, Miller JH. Regeneration beyond the glial scar. Nat Rev Neurosci2004;5:146–156.
7 Pekny M, Johansson CB, Eliasson C et al. Abnormal reaction to centralnervous system injury in mice lacking glial fibrillary acidic protein andvimentin. J Cell Biol 1999;145:503–514.
8 Wilhelmsson U, Li L, Pekna M et al. Absence of glial fibrillary acidicprotein and vimentin prevents hypertrophy of astrocytic processes andimproves post-traumatic regeneration. J Neurosci 2004;24:5016–5021.
9 Menet V, Prieto M, Privat A et al. Axonal plasticity and functionalrecovery after spinal cord injury in mice deficient in both glial fibrillaryacidic protein and vimentin genes. Proc Natl Acad Sci U S A 2003;100:8999–9004.
10 Cho KS, Yang L, Lu B et al. Re-establishing the regenerative potentialof central nervous system axons in postnatal mice. J Cell Sci 2005;118:863–872.
11 Gage FH, Coates PW, Palmer TD et al. Survival and differentiation ofadult neuronal progenitor cells transplanted to the adult brain. Proc NatlAcad Sci U S A 1995;92:11879–11883.
12 Lundberg C, Bjorklund A. Host regulation of glial markers in intrastriatalgrafts of conditionally immortalized neural stem cell lines. Neuroreport1996;7:847–852.
13 Li Y, Chen J, Zhang CL et al. Gliosis and brain remodeling aftertreatment of stroke in rats with marrow stromal cells. Glia 2005;49:407– 417.
14 Kinouchi R, Takeda M, Yang L et al. Robust neural integration fromretinal transplants in mice deficient in GFAP and vimentin. Nat Neurosci2003;6:863–868.
15 Rakic P. Adult neurogenesis in mammals: An identity crisis. J Neurosci2002;22:614–618.
16 Cameron HA, Woolley CS, McEwen BS et al. Differentiation of newlyborn neurons and glia in the dentate gyrus of the adult rat. Neuroscience1993;56:337–344.
17 Gage FH. Mammalian neural stem cells. Science 2000;287:1433–1438.18 Gage FH, Ray J, Fisher LJ. Isolation, characterization, and use of stem
cells from the CNS. Annu Rev Neurosci 1995;18:159–192.19 Palmer TD, Markakis EA, Willhoite AR et al. Fibroblast growth factor-2
activates a latent neurogenic program in neural stem cells from diverseregions of the adult CNS. J Neurosci 1999;19:8487–8497.
20 Gage FH, Kempermann G, Palmer TD et al. Multipotent progenitor cellsin the adult dentate gyrus. J Neurobiol 1998;36:249–266.
21 Lindvall O, Kokaia Z, Martinez-Serrano A. Stem cell therapy for human
neurodegenerative disorders-how to make it work. Nat Med 2004;10(suppl):S42–S50.
22 Alvarez-Buylla A, Lim DA. For the long run: Maintaining germinalniches in the adult brain. Neuron 2004;41:683–686.
23 Lie DC, Colamarino SA, Song HJ et al. Wnt signalling regulates adulthippocampal neurogenesis. Nature 2005;437:1370–1375.
24 Song H, Stevens CF, Gage FH. Astroglia induce neurogenesis from adultneural stem cells. Nature 2002;417:39–44.
25 Colucci-Guyon E, Portier MM, Dunia I et al. Mice lacking vimentindevelop and reproduce without an obvious phenotype. Cell 1994;79:679–694.
26 Pekny M, Leveen P, Pekna M et al. Mice lacking glial fibrillary acidicprotein display astrocytes devoid of intermediate filaments but developand reproduce normally. EMBO J 1995;14:1590–1598.
27 Mombaerts P, Iacomini J, Johnson RS et al. RAG-1-deficient mice haveno mature B and T lymphocytes. Cell 1992;68:869–877.
28 Palmer TD, Takahashi J, Gage FH. The adult rat hippocampus containsprimordial neural stem cells. Mol Cell Neurosci 1997;8:389–404.
29 Pekny M, Eliasson C, Chien CL et al. GFAP-deficient astrocytes arecapable of stellation in vitro when cocultured with neurons and exhibit areduced amount of intermediate filaments and an increased cell satura-tion density. Exp Cell Res 1998;239:332–343.
30 Eng LF, Ghirnikar RS. GFAP and astrogliosis. Brain Pathol 1994;4:229 –237.
31 Rogers SD, Peters CM, Pomonis JD et al. Endothelin B receptors areexpressed by astrocytes and regulate astrocyte hypertrophy in the normaland injured CNS. Glia 2003;41:180–190.
32 Hinterkeuser S, Schroder W, Hager G et al. Astrocytes in the hippocam-pus of patients with temporal lobe epilepsy display changes in potassiumconductances. Eur J Neurosci 2000;12:2087–2096.
33 Dziewczapolski G, Lie DC, Ray J et al. Survival and differentiation ofadult rat-derived neural progenitor cells transplanted to the striatum ofhemiparkinsonian rats. Exp Neurol 2003;183:653–664.
34 Norenberg MD, Martinez-Hernandez A. Fine structural localizationof glutamine synthetase in astrocytes of rat brain. Brain Res 1979;161:303–310.
35 Cai J, Chen Y, Cai WH et al. A crucial role for Olig2 in white matterastrocyte development. Development 2007;134:1887–1899.
36 Ligon KL, Fancy SP, Franklin RJ et al. Olig gene function in CNSdevelopment and disease. Glia 2006;54:1–10.
37 Takahashi M, Palmer TD, Takahashi J et al. Widespread integration andsurvival of adult-derived neural progenitor cells in the developing opticretina. Mol Cell Neurosci 1998;12:340–348.
38 Lu B, Kwan T, Kurimoto Y et al. Transplantation of EGF-responsiveneurospheres from GFP transgenic mice into the eyes of rd mice. BrainRes 2002;943:292–300.
39 Suhonen JO, Peterson DA, Ray J et al. Differentiation of adult hippocam-pus-derived progenitors into olfactory neurons in vivo. Nature 1996;383:624–627.
40 Vallieres L, Campbell IL, Gage FH et al. Reduced hippocampal neuro-genesis in adult transgenic mice with chronic astrocytic production ofinterleukin-6. J Neurosci 2002;22:486–492.
41 Larsson A, Wilhelmsson U, Pekna M et al. Increased cell proliferationand neurogenesis in the hippocampal dentate gyrus of old GFAP(-/-)Vim(-/-) mice. Neurochem Res 2004;29:2069–2073.
2627Widestrand, Faijerson, Wilhelmsson et al.
www.StemCells.com




















