
Increased gap junction assembly between cultured cells upon cholesterol supplementation
8
Increased gap junction assembly between cultured cells upon cholesterol supplementation RITA MEYER 1 -*, BARBARA MALEWICZ 2 , WOLFGANG J. BAUMANN 2 and ROSS G. JOHNSON 1 1 Department of Genetics and Cell Biology, University of Minnesota, 250 Biological Sciences Center, 1445 Gortner Avenue, St Paul, MN 55108, USA 2 The Hormel Institute, Austin, MN, 55912, USA • Author for correspondence Summary Novikoff hepatoma cells provide an excellent model system for the study of gap junction assembly, a process that could be influenced by lipids and other factors at numerous points. Since it is possible to alter the cellular levels of cholesterol in these cells, it was added to the cells in serum-supplemented me- dium and changes in gap junction assembly were evaluated. Cells were dissociated and reaggregated following exposure to a range of cholesterol concen- trations for 24 h. A five- to sixfold increase in the number of aggregated gap junction particles and a 50% increase in cellular cholesterol content were observed with 20 fiM added cholesterol. A 1-h ex- posure to added cholesterol, during cell reaggre- gation, resulted in a fourfold increase in the number of aggregated gap junction particles, demonstrating that the effect was rapid. The number of aggregated gap junction particles and formation plaque areas were used as measures of junction assembly and assayed by quantitative freeze-fracture and electron microscopy. Junctional permeabilities were evalu- ated by means of dye transfer times following the intracellular microinjection of Lucifer Yellow. Increased dye transfer was observed between chol- esterol-treated cells, which suggested that the in- crease in assembly was accompanied by an increase in junction permeability. Cells were treated with cycloheximide (lOO/jgml" 1 ) and actinomycin D (lO/jgml" 1 ) to determine whether protein and RNA syntheses were involved in the enhanced gap junc- tion assembly. Cycloheximide but not actinomycin D blocked the increased junction assembly observed with added cholesterol. These results suggested that protein synthesis, but not RNA synthesis, is necess- ary for the increased gap junction formation ob- served. Thus, cholesterol added in the presence of serum-supplemented medium resulted in a dramatic and rapid increase in the assembly of functional gap junctions between Novikoff cells. Key words: gap junction, intercellular communication, cholesterol. Introduction Gap junctions are specialized plasma membrane struc- tures that mediate the direct exchange of chemical mess- ages between cells. These membrane channels are thought to be a key factor in regulating intercellular transfer of nutrients and signal molecules involved in cell prolifer- ation, differentiation and tissue homeostasis (Warner etal. 1984; Kidder and McLachlin, 1985; Kidder et al. 1987; Fraser et al. 1987; Azarnia et al. 1988; Keane et al. 1988). The focus of our study has been on gap junction as- sembly between Novikoff hepatoma cells in which forma- tion occurs in a series of recognizable steps. Cells contact, adhere and produce matched membrane areas that con- tain clusters of 9-11 run intramembranous particles. These areas have been termed 'formation plaques' (Johnson et al. 1974; Preus et al. 1981a,i). We have suggested that formation plaques exist as plasma mem- brane domains (Johnson et al. 1989), with the 9-11 nm particles observed in freeze-fracture representing gap junction protein complexes. The flattened membrane area of the plaque may reflect an area with specialized lipids. Journal of Cell Science 96, 231-238 (1990) Printed in Great Britain © The Company of Biologiata Limited 1990 However, only a few studies have investigated the lipid components associated with gap junctions (Goodenough and Stoeckenius, 1972; Henderson et al. 1979; Hertzberg and Gilula, 1979; Hertzberg, 1984; Hertzberg et al. 1988; Malewicz et al. 1990) and the lipid composition of the gap junction formation plaque is unknown. Gap junction assembly could be influenced by cellular lipids and other factors at several stages during the formation process including: (1) the synthesis of gap junction proteins; (2) their insertion into the microsomal membrane; (3) their incorporation into the plasma mem- brane; (4) the redistribution (McCloskey and Poo, 1986; Chow and Poo, 1982, 1984) and accumulation of the precursors within the plane of the plasma membrane; and (5) junction degradation. In particular, if formation plaques are lipid domains, altered plaque or membrane lipids could affect junction formation by modifying the properties of developing junctions. To explore these ques- tions, we began growing cells in the presence of cholesterol in serum-supplemented medium to study the effects on gap junction assembly. We found that 20 /XM added choles- terol enhanced gap junction formation five- to sixfold and 231
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Increased gap junction assembly between cultured cells upon cholesterol supplementation

Increased gap junction assembly between cultured cells upon cholesterol
supplementation
RITA MEYER1-*, BARBARA MALEWICZ2, WOLFGANG J. BAUMANN2 and ROSS G. JOHNSON1
1Department of Genetics and Cell Biology, University of Minnesota, 250 Biological Sciences Center, 1445 Gortner Avenue, St Paul,MN 55108, USA2The Hormel Institute, Austin, MN, 55912, USA
• Author for correspondence
Summary
Novikoff hepatoma cells provide an excellent modelsystem for the study of gap junction assembly, aprocess that could be influenced by lipids and otherfactors at numerous points. Since it is possible toalter the cellular levels of cholesterol in these cells, itwas added to the cells in serum-supplemented me-dium and changes in gap junction assembly wereevaluated. Cells were dissociated and reaggregatedfollowing exposure to a range of cholesterol concen-trations for 24 h. A five- to sixfold increase in thenumber of aggregated gap junction particles and a50% increase in cellular cholesterol content wereobserved with 20 fiM added cholesterol. A 1-h ex-posure to added cholesterol, during cell reaggre-gation, resulted in a fourfold increase in the numberof aggregated gap junction particles, demonstratingthat the effect was rapid. The number of aggregatedgap junction particles and formation plaque areaswere used as measures of junction assembly andassayed by quantitative freeze-fracture and electronmicroscopy. Junctional permeabilities were evalu-ated by means of dye transfer times following the
intracellular microinjection of Lucifer Yellow.Increased dye transfer was observed between chol-esterol-treated cells, which suggested that the in-crease in assembly was accompanied by an increasein junction permeability. Cells were treated withcycloheximide (lOO/jgml"1) and actinomycin D(lO/jgml"1) to determine whether protein and RNAsyntheses were involved in the enhanced gap junc-tion assembly. Cycloheximide but not actinomycin Dblocked the increased junction assembly observedwith added cholesterol. These results suggested thatprotein synthesis, but not RNA synthesis, is necess-ary for the increased gap junction formation ob-served. Thus, cholesterol added in the presence ofserum-supplemented medium resulted in a dramaticand rapid increase in the assembly of functional gapjunctions between Novikoff cells.
Key words: gap junction, intercellular communication,cholesterol.
Introduction
Gap junctions are specialized plasma membrane struc-tures that mediate the direct exchange of chemical mess-ages between cells. These membrane channels are thoughtto be a key factor in regulating intercellular transfer ofnutrients and signal molecules involved in cell prolifer-ation, differentiation and tissue homeostasis (Warner etal.1984; Kidder and McLachlin, 1985; Kidder et al. 1987;Fraser et al. 1987; Azarnia et al. 1988; Keane et al. 1988).
The focus of our study has been on gap junction as-sembly between Novikoff hepatoma cells in which forma-tion occurs in a series of recognizable steps. Cells contact,adhere and produce matched membrane areas that con-tain clusters of 9-11 run intramembranous particles.These areas have been termed 'formation plaques'(Johnson et al. 1974; Preus et al. 1981a,i). We havesuggested that formation plaques exist as plasma mem-brane domains (Johnson et al. 1989), with the 9-11 nmparticles observed in freeze-fracture representing gapjunction protein complexes. The flattened membrane areaof the plaque may reflect an area with specialized lipids.
Journal of Cell Science 96, 231-238 (1990)Printed in Great Britain © The Company of Biologiata Limited 1990
However, only a few studies have investigated the lipidcomponents associated with gap junctions (Goodenoughand Stoeckenius, 1972; Henderson et al. 1979; Hertzbergand Gilula, 1979; Hertzberg, 1984; Hertzberg et al. 1988;Malewicz et al. 1990) and the lipid composition of the gapjunction formation plaque is unknown.
Gap junction assembly could be influenced by cellularlipids and other factors at several stages during theformation process including: (1) the synthesis of gapjunction proteins; (2) their insertion into the microsomalmembrane; (3) their incorporation into the plasma mem-brane; (4) the redistribution (McCloskey and Poo, 1986;Chow and Poo, 1982, 1984) and accumulation of theprecursors within the plane of the plasma membrane; and(5) junction degradation. In particular, if formationplaques are lipid domains, altered plaque or membranelipids could affect junction formation by modifying theproperties of developing junctions. To explore these ques-tions, we began growing cells in the presence of cholesterolin serum-supplemented medium to study the effects ongap junction assembly. We found that 20 /XM added choles-terol enhanced gap junction formation five- to sixfold and
231

resulted in an associated increase in intercellular dyetransfer. The effect was rapid and sensitive to cyclohexi-mide treatment, suggesting that protein synthesis isinvolved. Actinomycin D experiments indicated that RNAsynthesis may not be required.
Several studies have been done to examine the influenceof hormones (Decker, 1976; Burghardt and Anderson,1979; Burghardt and Matheson, 1982) and extracellularmatrix components (Spray et al. 1987) on a cell's comp-lement of gap junctions. While the increases observedwere presumably based on junction assembly over ratherextended periods, assembly was not directly evaluated.The levels (the number and size) of gap junctions are theresult of a dynamic relationship involving junction forma-tion and turnover. Therefore, factors that alter either ofthese parameters can have a profound effect on intercellu-lar communication. We report that communication bfe-tween Novikoff cells is enhanced due to altered gapjunction assembly over a period of 60 min, when cells aretreated with cholesterol in the presence of serum-sup-plemented medium.
Materials and methods
Dissociation, recovery and reaggregationNovikoff hepatoma cells (NlSl-67) are a cell line derived from achemically induced rat liver tumor, yet the cell type of origin isunknown. The cells are probably not a hepatocyte cell line butmay be of bile duct origin (Pitot and Potter, 1960). Cells fromsuspension culture were dissociated as previously described(Johnson et al. 1974; Preus et al. 1981a). Cells in logarithmicgrowth were centrifuged from standard growth medium(S210+10% newborn calf serum, Gibco), resuspended inS210+10mM EDTA at 6x l0 6 ce l l smr \ and placed in a 37°Cgyratory shaker incubator (200 revs min~l) for 15 min. The EDTAtreatment was repeated, resulting in greater than 95% singlecells. The cells were allowed to 'recover' in S210+5% calf serumfor 90 min, which eliminated remnants of previously existingjunctions (Preus et al. 1981a). Cells were reaggregated by centri-fugation at 30 £ for 5 min to yield loose pellets (Johnson et al.1974). A set of samples was fixed immediately following centrifu-gation (zero time control) and another set fixed after incubationfor 60 min in 50 ml round-bottomed tubes with S210+5% calfserum at 37 °C in a 5% CO2 atmosphere. These latter sampleswith cells 'reaggregated by pelleting* were used for freeze-fractureand electron microscopy. Recovered cells (2.5ml at 8xl0sml~1)were also settled out in 60 mm Petri dishes at 37 °C in 5% CO2atmosphere for 60 min and used for microinjection experiments('reaggregation by settling out').
Cholesterol supplementationA number of experiments were performed to evaluate the effect ofadded cholesterol on gap junction formation. In all cases thecholesterol was added in ethanol to heat-inactivated serum in thefollowing manner. A cholesterol stock solution (Nu Check Prep,Elysian, MN) was prepared in ethanol (Smgml"1). One ml of thestock solution was forcefully injected into 10 ml of calf serum at37 °C followed by 30 min of vortexing. The appropriate volume ofthis cholesterol-serum preparation was mixed with S210 mediumand untreated calf serum to yield media with a combined serumlevel of 5 % and the desired added cholesterol concentration. Thefinal culture medium always contained less than 0.05% ethanol.Cholesterol purity was determined by thin-layer chromatographyusing rc-hexane:diethyl etheracetic acid (70:30:1; by vol.).
Treatments of cells with cholesterol, cholesterol andcycloheximide (CHX), or cholesterol and actinomycin DFour independent sets of experiments were carried out to studythe effects of cholesterol at various concentrations on Novikoffcells. Cells were first grown for 24 h in 10% serum at added
cholesterol concentrations of 0, 10, 20, 30, 40 and 60 /JM. Thecorresponding concentration of cholesterol was also present in therecovery (90 min) and reaggregation (60 min) media.
A time course study was then performed with 0 /JM cholesterolor 20 JJM added cholesterol present in the media for 1 h (duringreaggregation), 2.5 h (recovery and reaggregation) or 26.5 h (a24h pretreatment+2.5h of recovery and reaggregation). A 60-min reaggregation period was used in all cases. Twenty-four hourcholesterol-treated samples (along with control samples) thatwere not EDTA-dissociated were also examined.
Other experiments were performed to investigate the involve-ment of protein synthesis in the increased gap junction assemblyobserved with 20/JM added cholesterol. Cells were pretreated for24 h with 20/JM added cholesterol, followed by the addition of bothcholesterol and cycloheximide (CHX, lOOmgml"1, Sigma) for a30 min dissociation, a 90 min recovery and a 60 min reaggregationperiod. Concurrent control samples included: 0 /JM cholesterol,0/JM cholesterol+CHX, and 20/iM added cholesterol withoutCHX. These samples were processed for quantitative freeze-fracture and dye injection. Similar experiments were done with-out the 24 h cholesterol pretreatment and used for dye-injectionexperiments. The transfer times were the same in the pretreatedand non-pretreated samples, so the pretreatment was dropped forsubsequent experiments.
Parallel experiments were performed to study the involvementof RNA synthesis in the enhanced junction formation observedwith 20/ai added cholesterol. Actinomycin D (lO/jgml"1) waspresent in the dissociation, recovery and reaggregation media (atotal of 3 h). Cholesterol (20 /JM) was present in the recovery andreaggregation media. The experiments included: 0 /IM cholesterol,0/JM cholesterol+actinomycin D, 20/JM added cholesterol, and20/IM added cholesterol+actinomycin D samples.
Quantitative freeze-fracture and electron microscopyPellets of cells were reaggregated for 60 min, then centrifuged at45 £ for 5 min and fixed in 2.5% glutaraldehyde in S210 mediumfor freeze fracturing (Johnson et al. 1974). Further processing wasperformed as previously reported (Sheridan et al. 1978). At leasttwo replicas per treatment were examined in each experimentand all experiments were performed at least in triplicate.
Methods of quantitation have previously been described (Preuset al. 1981a,W. Briefly, an 'interface' was defined as a fracturedmembrane area comprising at least 57/jm2 (i.e. filling the screenat x 10 000 on our microscope) and containing an indication of cellapposition. Interfaces were scored according to the presence orabsence of one or more formation plaques or gap junctions.•Formation plaques' (Johnson et al. 1974) were defined as mem-brane areas with clusters or arrays of 9-11 nra intramembranousparticles. Aggregates of six or more particles were identified asgap junctions. Data on formation plaques were tabulated on amembrane interface basis and on an individual plaque basis. Thesame conclusions were reached with the interface data and theindividual plaque results. Consequently this report describesexclusively the interface data for reasons of brevity. Formationplaque area and particle number measurements were performedas reported in detail elsewhere (Preus et al. 1981a). Measure-ments were made with the aid of an Apple II computer equippedwith a Houston Instruments Hipad digitizing tablet and morpho-metry software developed for use with the Novikoff cell system.
Microinjection of dyeCells were injected with Lucifer Yellow to determine whetherchanges in the assembly of gap junctions were accompanied bysimilar changes in junctional permeability. Dye injection studieswere performed on reaggregated cells that had been settled out inPetri dishes in contrast to reaggregation by pelleting used forfreeze-fracture. The electron microscopy required a multilayeredcell population to minimize microscope time, while cell pairs wereideal for unambiguous permeability data on a single cell-cellinterface. The two sets of data were qualitatively consistent.However, the values cannot be compared in a strict quantitativesense because the cells were not prepared in the same manner. Wefound, for example, that a 20 /JM cholesterol treatment for 1 h ledto a fourfold increase in the number of aggregated particles per
232 R. Meyer et al.

interface, while permeability was increased 2.5 times with thesame treatment. Both increases were observed consistently fromexperiment to experiment and were statistically significant (Pvalues<0.05).
Dye injection wa8 accomplished using a General Valve picos-pritzer to inject a standardized quantity of 1 % aqueous LuciferYellow CH (Sigma). Injection of the dye and subsequent transferof dye were recorded on a Zeiss IM microscope equipped with aDage SIT camera, Panasonic video recorder and monitor. Thedegree of dye transfer was estimated by measuring the timeelapsed from the beginning of dye injection into the impaled cellto detection of fluorescence in an adjacent cell. These transfertime intervals' (Atkinson et al. 1981) were determined by analysisof the videotapes.
Amino acid incorporation into proteinIncorporation of labeled amino acids into acid-insoluble materialwas measured following incubation of dissociated and treatedcells in [^SJmethionine (Amersham) for 4 h. Control cells or 20 /JMcholesterol-treated cells were incubated with CHX (100/igml"1).Concurrent samples were processed without CHX treatment.Duplicate samples were analyzed and the experiment repeated intriplicate. The cells were washed in phosphate-buffered saline(PBS) and treated with 10 % trichloroacetate (TCA; Epstein et al.1977). The precipitate was heated for 30min at 70°C, washed inTCA, dissolved in formic acid and counted in scintillation fluid.
Nucleotide incorporation into RNAIncorporation of labeled nucleotides into acid-insoluble materialwas measured following incubation of dissociated and treatedcells in [3H]uridine (Amersham) for 4h. The cells were treatedwith 0 or 20 /<M cholesterol in the presence of serum-supplementedmedium, with or without actinomycin D (lO/fgml"1). Duplicatesamples were evaluated in each of two experiments. Cells werewashed in PBS and treated with 10 % TCA. The precipitate washeated for 30 min at 70°C, washed in TCA, dissolved in formic acidand counted in scintillation fluid (Pederson et al. 1980).
Thin-section volume measurementsTransmission electron microscopy on thin sections of control and20/iM cholesterol-treated Novikoff cells was performed as pre-viously reported (Preus et al. 1981a,6). The morphometricmethods used to determine cell volume were those described byWeibel (1979).
Cell cholesterol assayCell samples for lipid analysis were washed twice with phosphate-buffered saline and the cell pellets were immediately frozen andstored at -70°C until analyzed. Novikoff cell lipids wereextracted (Bligh and Dyer, 1959) and phospholipid content wascalculated on the basis of lipid phosphorus analyses (Bartlett,1959). Cell cholesterol was separated from cholesterol esters bythin-layer chromatography using n-hexane:diethyl ethenaceticacid (80:20:1, by vol.) as the developing system and cholesterolwas assayed as reported (Rudel and Morris, 1973). A referencelipid sample consisting of cholesterolxholesterol oleate:phospha-tidylcholine (30:1:5, by mol.) was extracted and assayed with
each set of experimental samples to monitor separation efficiencyand reproducibility.
Statistical treatmentsValues were routinely expressed as means with standard errors.To determine the statistical significance of values obtained withthe various treatments, Student's t-test was applied in instanceswhere differences appeared to exist.
Results
We studied gap junction formation between Novikoffhepatoma cells in the presence of cholesterol in serum-supplemented medium. Quantitative freeze-fracture andelectron microscopy were used to measure the size andnumber of gap junctions present. Our experiments in-cluded control samples where cells were dissociated, re-covered, pelleted without reaggregation, processed, andexamined to ensure that essentially all of the junctionsobserved using our experimental procedures were newlyformed. Quantitative freeze-fracture revealed gap junc-tions on less than 2 % of the interfaces when the cells hadbeen dissociated but not reaggregated, in contrast to60-70% of the cell interfaces in preparations reaggre-gated for 60 min. In addition, the junctions between cellsthat were dissociated but not reaggregated were small(<100 particles). This indicates little carry over of 'old'junctions into the formation experiments. The fate of oldgap junction proteins upon cell dissociation is not clear. Wedo not know whether the proteins are recycled into thenewly forming junctions.
Concentration-dependent effectCells supplemented for 24 h with a range of cholesterolconcentrations were reaggregated for 60 min. Junctionassembly was monitored using freeze-fracture (Fig. 1) anddye injection methods (Fig. 2). A 4.4-fold increase in thenumber of aggregated gap junction particles (Table 1) wasobserved with this series of 20 //M cholesterol treatments.Formation plaque areas in the 20 ^M added cholesterolsamples were more than twice those of 0 /IM cholesterolsamples (Fig. 1, Table 1). Less junction formation (asmaller number of aggregated gap junction particles andsmaller formation plaque areas) was observed at choles-terol concentrations both below and above 20 J*M (Table 1).Freeze-fracture experiments with cholesterol concen-trations higher than 60 /.IM showed particle clumping.
No significant change was observed in the initiation ofgap junction assembly as reflected by a constant percent-age of positive interfaces (61-66%) with the differentconcentrations of added cholesterol. The number of forma-tion plaques per interface (control, 2.58±.25; 20 /JM choles-
Table 1. Effects of different cholesterol levels: quantitative freeze-fracture analysisTreatment*(cholesteroladded inserum)
Number ofinterfacest
Percentageof interfaces
with junctions
Formation plaquearea (/an2)
per interface0E±8.B.M.)
Number ofaggregated particles
per interface(i±8.K.M.)
0/IM10/Urf20 /IM
30 /<M
40/(M60 tin
701572133841
666466656161
0.70±0.080.52±0.081.86±0.170.38±0.060.71±0.070.31±0.04
436±44868±209
1899±214681±208445±73275 ±40
* Data are from four experiments in which cells were pretreated with cholesterol in serum for 24 ht An interface is denned as a fractured membrane area comprising at least 57 /an2 and providing an indication of cell-cell apposition.
Gap junction assembly 233

Fig. 1. Freeze-fracture micrographs of a control cell (0/JM added cholesterol) fonnation plaque (A) compared with a formation plaquebetween 20 /ZM cholesterol-treated cells (B). The 20 /uu cholesterol-treated cells showed a larger number of aggregated particles thanthe control cells, as well as larger formation plaque areas. Arrowheads point to representative examples of gap junction particleaggregates. Bar, 0.2 fan.
234 R. Meyer et al.
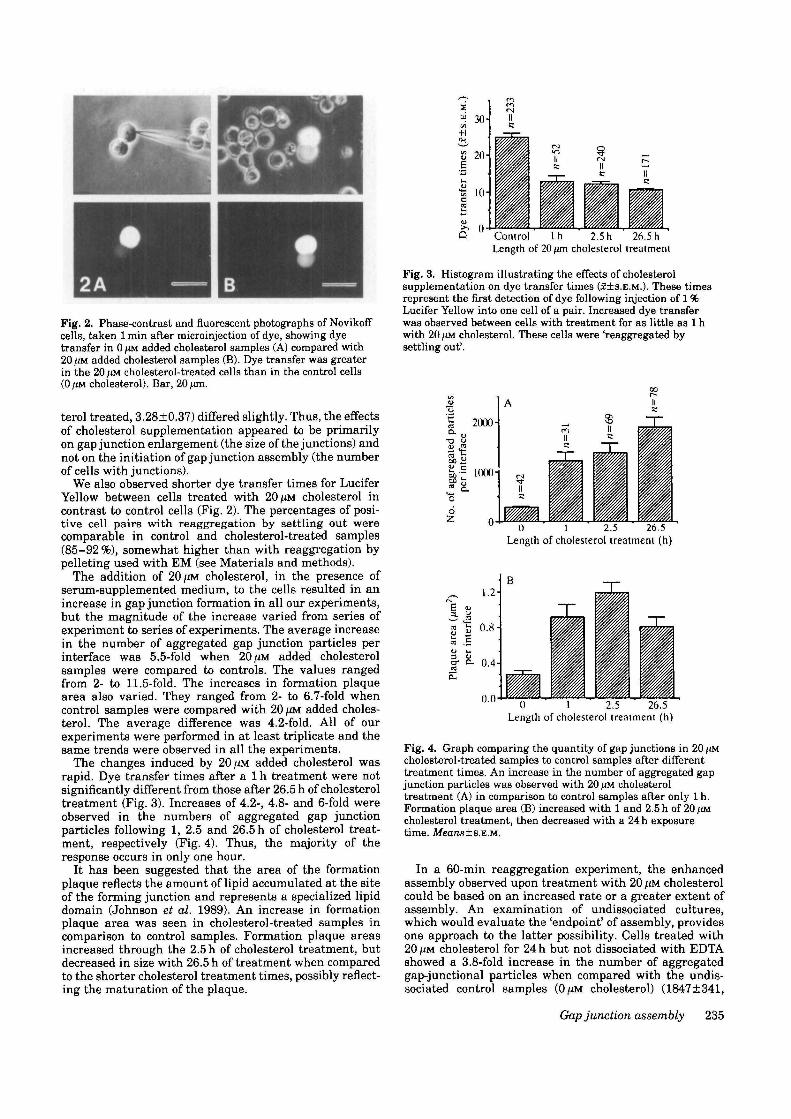
Fig. 2. Phase-contrast and fluorescent photographs of Novikoffcells, taken 1 min after microinjection of dye, showing dyetransfer in 0 /*M added cholesterol samples (A) compared with20/iM added cholesterol samples (B). Dye transfer was greaterin the 20 /(M cholesterol-treated cells than in the control cells(0 )JM cholesterol). Bar, 20 /tm.
terol treated, 3.28±0.37) differed slightly. Thus, the effectsof cholesterol supplementation appeared to be primarilyon gap junction enlargement (the size of the junctions) andnot on the initiation of gap junction assembly (the numberof cells with junctions).
We also observed shorter dye transfer times for LuciferYellow between cells treated with 20 fiM cholesterol incontrast to control cells (Fig. 2). The percentages of posi-tive cell pairs with reaggregation by settling out werecomparable in control and cholesterol-treated samples(85-92 %), somewhat higher than with reaggregation bypelleting used with EM (see Materials and methods).
The addition of 20 [XM. cholesterol, in the presence ofserum-supplemented medium, to the cells resulted in anincrease in gap junction formation in all our experiments,but the magnitude of the increase varied from series ofexperiment to series of experiments. The average increasein the number of aggregated gap junction particles perinterface was 5.5-fold when 20 HM added cholesterolsamples were compared to controls. The values rangedfrom 2- to 11.5-fold. The increases in formation plaquearea also varied. They ranged from 2- to 6.7-fold whencontrol samples were compared with 20 ^M added choles-terol. The average difference was 4.2-fold. All of ourexperiments were performed in at least triplicate and thesame trends were observed in all the experiments.
The changes induced by 20 (JM added cholesterol wasrapid. Dye transfer times after a 1 h treatment were notsignificantly different from those after 26.5 h of cholesteroltreatment (Fig. 3). Increases of 4.2-, 4.8- and 6-fold wereobserved in the numbers of aggregated gap junctionparticles following 1, 2.5 and 26.5 h of cholesterol treat-ment, respectively (Fig. 4). Thus, the majority of theresponse occurs in only one hour.
It has been suggested that the area of the formationplaque reflects the amount of lipid accumulated at the siteof the forming junction and represents a specialized lipiddomain (Johnson et al. 1989). An increase in formationplaque area was seen in cholesterol-treated samples incomparison to control samples. Formation plaque areasincreased through the 2.5 h of cholesterol treatment, butdecreased in size with 26.5 h of treatment when comparedto the shorter cholesterol treatment times, possibly reflect-ing the maturation of the plaque.
s"i 30-j[/I
-H
S3 2 0 -
10-
Q Control l h 2.5 h 26.5 hLength of 20 ^m cholesterol treatment
Fig. 3. Histogram illustrating the effects of cholesterolsupplementation on dye transfer times (£±S.E.M.). These timesrepresent the first detection of dye following injection of 1 %Lucifer Yellow into one cell of a pair. Increased dye transferwas observed between cells with treatment for as little as l hwith 20 /(M cholesterol. These cells were 'reaggregated bysettling out'.
M
' 1 2000H
as,•= 1000-1
6Z.
ii
IIB
0 1 2.5 26.5Length of cholesterol treatment (h)
1.2-
0.8-
§" ^ 0.4-K.
0.00 1 2.5 26.5
Length of cholesterol treatment (h)
Fig. 4. Graph comparing the quantity of gap junctions in 20 /uncholesterol-treated samples to control samples after differenttreatment times. An increase in the number of aggregated gapjunction particles was observed with 20 /iM cholesteroltreatment (A) in comparison to control samples after only 1 h.Formation plaque area (B) increased with 1 and 2.5 h of 20 /IMcholesterol treatment, then decreased with a 24 h exposuretime. Means±8.E.M.
In a 60-min reaggregation experiment, the enhancedassembly observed upon treatment with 20 //M cholesterolcould be based on an increased rate or a greater extent ofassembly. An examination of undissociated cultures,which would evaluate the 'endpoint' of assembly, providesone approach to the latter possibility. Cells treated with20 HM cholesterol for 24 h but not dissociated with EDTAshowed a 3.8-fold increase in the number of aggregatedgap-junctional particles when compared with the undis-sociated control samples (0/iM cholesterol) (1847±341,
Gap junction assembly 235

Table 2. Effects of inhibition of protein and RNA synthesis: quantitative freeze-fracture analysis
Treatment*^(addedcholesterol inserum)
0/ZM20 ^M
0/1M +cycloheximide
20/IM+cycloheximide
0//M +actinomycin D
20/IM+actinomycin D
Number ofinterfaces^
17917865
74
23
31
Percentageof interfaces
with junctions
656666
61
68
69
Formation plaquearea (/an3)
per interface(f±S.K.M.)
0 56±0 051.79±0.10.3±0.04
0.313±0.05
0.25±0.05
1.3±0.10
Number ofaggregated particles
per interface(J±S.B.M.)
313±312050±201263±75
169±25
227±34
1307±279
*Data are from three experiments.tData are from cells treated with cycloheximide or actinomycin D for the 3h of the experiments (dissociation, recovery and reaggregation). The
cholesterol was added in the recovery and reaggregation media.| An interface is denned as a fractured membrane area comprising at least 57 /an2 and providing an indication of cell-cell apposition.
control, n=30 versus 7113±957, 20 UM added cholesterol,n=27). An increase was also observed in plaque area percell interface when cells treated with cholesterol in thepresence of serum-supplemented medium (20 ^M) werecompared with undissociated control cells. Thus, our datasuggested that 20/JM added cholesterol influenced theextent of gap junction formation. More detailed studieswill be required to examine actual rates of assembly underthe conditions employed here.
No accumulation of lipid droplets was observed by thin-section electron microscopy when the cells were treated for2.5 h with 20 UM added cholesterol. Volume measurementswere also made of cells in control samples (766±62^m3,n=43) and 20 UM cholesterol treated samples (815±57 jon3,n=45), since cell volume could neutralize increases inassembly and leave junctional communication unaltered.However, no significant changes in cell volume wereobserved with cholesterol treatment, which suggested thatthe five- to sixfold increase in junction assembly rep-resents an increase in communication.
Protein synthesis was required for enhanced junctionformation with cholesterol supplementationCells were treated with cycloheximide to study the poss-ible involvement of protein synthesis in the enhanced gapjunction formation observed with cholesterol supplemen-tation. Cycloheximide (CHX, 100/igml"1) was added tocontrol (0/iM cholesterol) and 20/.IM cholesterol-treatedsamples for freeze-fracture, dye injection, and [36S]methio-nine studies. CHX-treated cells (both with and without20 ^M added cholesterol) showed less than 3% of thelabeled amino acid incorporation of the non-CHX-treatedNovikoff cells, demonstrating that protein synthesis wasinhibited. CHX blocked the increase in gap junctionassembly observed with 20 UM added cholesterol (Table 2).CHX treatment resulted in comparable numbers of aggre-gated gap junction particles and formation plaque areas inthe 0 UM and 20 ;<M added cholesterol samples. Initiation ofjunction formation (percentage positive interfaces) wasalso unchanged with CHX treatment (Table 2). Studies onjunction permeability were consistent with the freeze-fracture data; the transfer-time intervals were the samefor cycloheximide-treated control and 20 UM added choles-terol samples (Fig. 5).
The data indicated that the assembly of junctions wasinitiated and basal levels of junction assembly occurred inthe presence of CHX. However, the enhanced assembly
Fig. 5. Histogram illustrating the effects of the inhibition ofprotein and RNA synthesis on gap junction assembly asmonitored by dye permeability. Dye transfer times (5±S.E.M.)for 20/iM added cholesterol+cycloheximide (CHX)-treated cellswere the same as control+CHX cells, indicating that CHXblocked the increased dye transfer normally observed withadded cholesterol. Actinomycin D+cholesterol-treated cellsexhibited shorter dye transfer times than actinomycin D+0/<Mcholesterol cells, demonstrating that actinomycin D (AD) hadonly a small effect on the rapid dye transfer occurring withcholesterol supplementation.
observed with the addition of cholesterol was inhibited inthe absence of protein synthesis.
RNA synthesis may not be required for enhancedjunction formation with cholesterol supplementationCells were treated with actinomycin D (lO/Ugml"1) toexamine the involvement of RNA synthesis in theincreased gap junction formation observed with 20 IJMadded cholesterol. Actinomycin D was tested at a range ofconcentrations (0, 1, 5, 10, 20/igmT1) and the 10/igml"1
level was chosen as the most effective concentration thatwas not toxic. Actinomycin D-treated control (0 /.IM choles-terol) cells showed 12.6% of the [3H]uridine incorporationof non-actinomycin D-treated cells. The 20 JAM added chol-esterol cells treated with actinomycin D had 11.4% of the[3H]uridine labeling of 20 fm added cholesterol cells with-out actinomycin D. Thus, RNA synthesis was inhibited butwas not blocked completely.
Cells treated with 20 UM cholesterol and actinomycin Dshowed enhanced gap junction formation when compared
236 R. Meyer et al.

60
on
B
U
20
00 ' 1 ' 2.5 24
Time (h) in 20/<M cholesterol supplementation
Fig. 6. Histogram illustrating cellular cholesterol contentfollowing treatment with 20 /IM cholesterol in the presence ofserum-supplement medium for various time periods.
with controls (Table 2). A 5.7-fold increase in the numberof aggregated gap junction particles was observed insamples treated with 20 f*M added cholesterol and actino-mycin D when compared with samples treated with 0 /£Mcholesterol and actinomycin D. Formation plaque areaswere 5.2-fold larger in cells treated with actinomycin Dand 20/tM added cholesterol. The percentage of positiveinterfaces in these samples remained relatively constant(61-69 %) (Table 2) and indicated that actinomycin D hadno significant effect on the initiation of gap junctionassembly. Actinomycin D and 20 jUM cholesterol-treatedcells also transferred dye more rapidly than cells treatedwith OjtM cholesterol and actinomycin D (Fig. 5). Thus,increases in gap junction assembly and junctional per-meability occurred in the presence of limited RNA syn-thesis.
Cell cholesterol, contentIn these studies, the serum (5 %)-supplemented mediumcontained 33nmol of cholesterol ml"1. Novikoff cellsgrown in control medium were found to contain 44.6 nmolof cholesterol mg"1 protein. In cells treated with mediumsupplemented with 20fiM cholesterol, increases of 9, 15,and 21 nmol mg"1 protein were detected after 1, 2.5 and24 h of treatment (Fig. 6). Thus, most of the increaseoccurred in 2.5 h. At the same time, cell phospholipidcontent remained unchanged at about 170 nmol mg"1 pro-tein.
Discussion
We have used Novikoff hepatoma cells to examine gapjunction formation. The Novikoff cells provide a modelsystem in which the assembly process can be analyzed in aquantitative manner that permits us to obtain correlativestructural and functional data. Cells are readily dis-sociated without the use of proteases and reaggregated toform functional junctions over a time course of minutes(Johnson et al. 1974; Preus et al. 19816).
Cholesterol was added to the Novikoff cells in thepresence of serum-supplemented medium to examine poss-ible changes in gap junction assembly. We report here ourfindings, which include: (1) 20 ^M added cholesterolresulted in an increase in gap junction assembly betweenreaggregated cells, as evidenced by a five- to sixfoldincrease in the number of junctional particles per cellinterface; (2) 20 f.tM added cholesterol resulted in increasedjunctional permeability as measured by dye transfer timeintervals; (3) the increase in gap junctions was observed
with treatment for as little as lh , indicating that theprocess was rapid; (4) a 3.8-fold increase in the number ofaggregated junctional particles was observed in undis-sociated cholesterol-treated cells when compared withundissociated control cells, suggesting that cholesteroltreatment resulted in larger junctions; (5) cycloheximideblocked the enhanced formation observed with 20/.IMadded cholesterol, suggesting that protein synthesis wasinvolved in the increase in gap junction assembly;(6) actinomycin D had only a small effect on the enhancedgap junction formation observed with added cholesterol,indicating that RNA synthesis may not be required;(7) gap junction assembly varied with the concentrationsof added cholesterol; and (8) cell cholesterol content wasincreased with 20 ^M cholesterol supplementation. Thus,we demonstrated that gap junction formation was alteredby cholesterol added in the presence of serum-sup-plemented media.
Basal levels of gap junction formation were observedwhen Novikoff cells were treated with cycloheximide,which suggested that a 'pool' or a reserve of gap junctionprotein precursors exists within the cells (Epstein et al.1977). The concept of a precursor pool of junctional pro-teins was suggested by earlier studies using cultured cells(Goshima, 1971) and reinforced by work on mammalianembryos (McLachlin and Kidder, 1986) in which junctionformation occurred in the presence of protein synthesisinhibitors. In contrast, the enhanced gap junction as-sembly we observed with added cholesterol was sensitiveto cycloheximide treatment and this suggested that thesynthesis of either a gap junction protein(s) or otherprotein(s) is required for the elevated assembly. A similarresult, where new protein synthesis was necessary for gapjunction formation, was observed in rat myometrium(McKenzie and Garfield, 1986). In that study cyclohexi-mide prevented the increase in gap junctions that wasusually observed with estradiol treatment. Protein syn-thesis is also likely to be involved in other cases whereincreases have been reported in the number or size of gapjunction structures (e.g. see Decker, 1976).
The stimulated formation of gap junctions we observedwith 20/iM added cholesterol occurred in the presence ofactinomycin D, indicating that RNA synthesis was prob-ably not required. A similar result was reported in earlystage mouse embryos treated with alpha-amanitin wheregap junction formation occurred in the near absence oftranscription (McLachlin and Kidder, 1986).
To understand the mechanism of the enhanced gapjunction assembly, we need to determine whether theincrease in assembly was based on specific changes in gapjunctional proteins or whether the levels of other proteinswere altered with 20 ^M cholesterol treatment. Recentwork has identified, using Western and Northern ana-lyses, a Novikoff cell protein of approximately 43 000 Mr,which cross-reacts with antibodies to a heart gap junctionprotein (Finis et al. 1989). The rat heart gap junctionprotein has been termed connexin 43 (Beyer et al. 1987).The antibodies used on Novikoff cells were either preparedagainst a synthetic oligopeptide corresponding to anN-terminal region or against electroeluted rat heart con-nexin 43 (Yancey et al. 1989). The demonstrated cross-reactivity will allow us to study the question of whetherthe increase in junction assembly and permeability is dueto increased gap junction protein synthesis (Spray et al.1987; Traub et al. 1983,1987), reduced gap junction proteinturnover (Saez etal. 1989), the redistribution of the proteinfrom the cytoplasm into the membrane or enhanced
Gap junction assembly 237

clustering of the protein within the plane of the mem-brane.
The cause of the inhibition of gap junction assembly athigher levels of added cholesterol is unclear. There are afew studies reported that address the effects of lipidsupplementation on gap junctions (Malewicz et al. 1990).They demonstrated an inhibition of metabolic cooperationwhen cells were treated with unsaturated fatty acids(Aylsworth et al. 1986, 1987) or cholesterol epoxides(Chang et al. 1988). Since these studies do not distinguishbetween junction assembly and channel gating, theirrelevance to the present study is uncertain.
National Science Foundation grant DCB 8517726 supportedthis work. The Hormel Institute was also supported by NIHprogram project grant HL08214 and the Hormel Foundation.
References
ATKINSON, M. M., MENKO, A. S., JOHNSON, R. G., SHEPPARD, J. R. ANDSHBRIDAN, J. D. (1981). Rapid and reversible reduction of junctionalpermeability in cells infected with temperature-sensitive mutant ofavian sarcoma virus. J. Cell Biol 91, 573-678.
AYLSWORTH, C. F., TROSKO, J. E. AND WELSCH, C. W. (1986). Influence oflipida on gap junction mediated intercellular communication betweenChinese hamster cells in vitro. Cancer Res. 46, 4527-4533.
AYLSWORTH, C. F., WELSCH, C. W., KABARA, J. J. AND TROSKO, J. E.(1987). Effects of fatty acids on gap junctional communication: possiblerole in tumor promotion by dietary fat. Lipids 22, 445—464.
AZARNIA, R., REDDY, S., KMIECIK, T. T., SHALLOWAY, D. ANDLOEWKNSTEIN, W. R. (1988) The cellular arc gene product regulatesjunctional cell-to-cell communication. Science 239, 398—401.
BARTLETT, G. R. (1959). Phosphorus assay in column chromatography. J.biol. Chem. 234, 466-468.
BEYER, E. C, PAUL, D. L. AND GOODENOUGH, D. A. (1987). Connexin 43:A protein from rat heart homologous to a gap junction protein fromliver. J. Cell Biol. 105, 2621-2629.
BUOH, E. G. AND DYER, W. J. (1959). A rapid method of total lipidextraction and purification. Can. J. Biochem. Physiol. 37, 911-917.
BURGHARDT, R. C. AND ANDERSON, E (1979). Hormonal modulation ofovarian interstitial cells with particular reference to gap junctions. J.Cell Biol. 81, 104-114.
BUBOHARPT, R. C. AND MATHESON, R. L. (1982). Gap junctionamplification in rat ovarian granulosa cells. I. A direct response tofollicle stimulating hormone. Devi Biol. 94, 206-215.
CHANG, C. C, JONES, C, TROSKO, J. E., PETERSON, A. R. AND SEVANLAN,A. (1988). Effect of cholesterol epoxides on the inhibition ofintercellular communication and on mutation induction in Chinesehamster V79 cells. Mutat. Res. 206, 471-478.
CHOW, I. AND POO, M. M. (1982). Redistribution of cell surface receptorsinduced by cell-cell contact. J. Cell Biol. 95, 510-618.
CHOW, I. AND POO, M. M. (1984). Formation of electrical couplingbetween embryonic muscle cells in culture. J. Physiol., Land. 346,181-194.
DECKEH, R. S. (1976). Hormonal regulation of gap junctiondifferentiation. J. Cell Biol. 69, 669-685.
EPSTEIN, M. L., SHERIDAN, J. D. AND JOHNSON, R. G. (1977). Formation oflow-resistance junctions in vitro in the absence of protein synthesisand ATP production. Expl Celt Res. 104, 25-30.
FINIS, C. J., MEYER, R. A. AND JOHNSON, R. G. (1989). Expression ofconnexin 43 in Novikoff hepatoma cells J Cell Biol. 109, 47a.
FRASER, S. E., GREEN, C. R., BODE, H. R. AND GILULA, N. B. (1987).Selective disruption of gap junction communication interferes with apatterning process in Hydra. Science 237, 49-55.
GOODENOUGH, D. A. AND STOECKENIUS, W. (1972). The isolation of mousehepatocyte gap junction: Preliminary chemical characterization and x-ray diffraction. J. Cell Biol. 54, 646-656.
GOSHIMA, K. (1971). Synchronized beating of myocardial cells mediatedby FL cells in monolayer culture and its inhibition by trypsin-treatedFL cells. Expl. Cell Res. 65, 161-169.
HENDERSON, D., EIBL, H. AND WEBER, K (1979). Structure andbiochemistry of mouse hepatic gap junctions. J. molec. Biol. 132,193-218.
HERTZBIRG, E. L. (1984). A detergent-independent procedure for theisolation of gap junctions from rat liver. J. biol. Chem. 259, 9936-9943.
HERTZBERO, E. L AND GILULA, N. B. (1979). Isolation andcharacterization of gap junctions from rat liver. J. biol. Chem. 254,2138-2147.
HERTZBERO, E. L., SPRAY, D. C. AND COOK, R. (1988). Biochemical,immunological and topological studies of gap junctions. In GapJunctions (ed. E Hertzberg and R. G. Johnson), pp. 9-28 New York:Alan R Liss, Inc. .
JOHNSON, R., HAMMER, M., SHERIDAN, J. AND REVEL, J. P. (1974). Gapjunction formation between reaggregating Novikoff hepatoma cells.Proc. natn. Acad. Sci. USA. 71, 4536-4640.
JOHNSON, R. G., MEYER, R. A. AND LAMPB, P. (1989). Gap junctionformation: A self-assembly model involving membrane domains oflipid and protein. In Cell Interactions and Gap Junctions, vol 1., (ed. N.SperelakiB and W. C. Cole), pp. 169-179, Boca Raton, Fl.: CRC Press.
KEANE, R. W., MEHTA, P., ROSE, B., HONIG, L. S., LOEWKNSTEIN, W. R.AND RUTISHAUSER, U. (1988). Neural differentiation, NCAM-mediatedadhesion, and gap junction communication in neuroectoderm. A studyin vitro. J. Cell Biol. 106, 1307-1319.
KIDDER, G. M. AND MCLACHUN, J. R. (1985). Timing of transcription andprotein synthesis underlying morphogenesis in preimplantation mouseembryos. Devi Biol. 112, 266-276.
KIDDER, G. M., RAINS, J. AND MCKEON, J. (1987). Gap junction assemblyin the preimplantation mouse conceptus is independent ofmicrotubules, microfilaments, cell flattening, and cytokinesis. Proc.natn. Acad. Sci. U.S.A. 84, 3718-3722.
MACKENZIE, L. W. AND GARFIELD, R. E. (1986). Effects of tamoxifencitrate and cycloheximide on eatradiol induction of rat myometrial gapjunctions. Can. J. Physwl. Pharmac. 64, 703-706.
MALEWICZ, B., KUMAR, V. V., JOHNSON, R. G. AND BAUMANN, W. J.(1990). Lipids in gap junction assembly and function. Lipids 25, (inpress).
MCCLOSKBY, M. A. AND POO, M. M. (1986). Contact-induced redistributionof specific membrane components: local accumulation and developmentof adhesion. J Cell Biol. 102, 2185-2196.
MCLACHUN, J. R. AND KIDDER, G. M. (1986). Intercellular junctionalcoupling in preimplantation mouse embryos: Effect of blockingtranscription and translation. Devi Biol. 117, 146—156.
PEDERSON, D. C, SHERIDAN, J. D. AND JOHNSON, R. G. (1980). The
development of metabolite transfer between reaggregating Novikoffhepatoma cells. Expl Cell Res. 127, 159-178.
PrroT, H. C. AND POTTER, V. R. (1960). An enzymic study on the cellularorigin of the Dunning and the Novikoff hepatomas in the rat. Biochim.biophys Ada 40, 537-539.
PREUS, D , JOHNSON, R. AND SHERIDAN, J. (1981a). Gap junctions betweenNovikoff hepatoma cells following dissociation and recovery in theabsence of cell contact. J. Ultrastruct. Res. 77, 248-262.
PREUS, D., JOHNSON, R., SHERIDAN, J. AND MEYER, R (19816). Analysis ofgap junctions and formation plaques between reaggregating Novikoffhepatoma cells. J. Ultrastruct. Res. 77, 263-276.
RUDEL, L. L. AND MORRIS, M. D. (1973). Determination of cholesterolusing o-phthalaldehyde. J. Lipid Res. 14, 364-366.
SAEZ, J. C, GREGORY, W. A., WATANABE, T., DBRMIETZEL, R., HERTZBERG,E. L., REID, L. R., BENNETT, M. V. L. AND SPRAY, D. C. (1989). cAMPdelays disappearance of gap junctions between pairs of rat hepatocytesin primary culture. Am. J. Physwl. 257, Cl -Cl l .
SHERIDAN, J., HAMMER-WILSON, M., PREUS, D. AND JOHNSON, R. (1978).Quantitative analysis of low-resistance junctions between culturedcells and correlation with gap junction areas. J Cell Biol. 76, 632-544.
SPRAY, D. C, FUJITA, M , SABZ, J. C, CHOI, H., WATANABE, T.,HERTZBERG, E., ROSENBERG, L. C. AND REID, L. M. (1987).Proteoglycans and glycosaminoglycans induce gap junction synthesisand function in primary liver cultures. J. Cell Biol. 105, 641—55.
TBAUB, O., DRUOE, P. M. AND WILLECKH, K (1983). Degradation andresynthesis of gap junction protein in plasma membranes ofregenerating liver after partial hepatectomy or cholestasis. Proc. natn.Acad. Sci. U.S.A. 80, 756-759.
TRAUB, O., LOOK, J., PAUL, D. AND WILLECKE, K. (1987). Cyclic adenosinemonophosphate stimulates biosynthesis and phosphorylation of the26kDa gap junction protein in cultures of mouse hepatocytes. Eur. J.Cell Biol. 43, 48-64
WARNER, A. E., GUTHRIE, S C AND GILULA, N. B. (1984). Antibodies togap junctional protein selectively disrupt communication in the earlyamphibian embryo. Nature 311, 127-131.
WEIBEL, E. R. (1979). Stereological Methods, Vol. 1, In Practical Methodsfor Biological Morphometry. New York: Academic Press.
YANCEY, S. B., JOHN, S. A., LAL, R., AUSTIN, B. J. AND REVEL, J. P.(1989). The 43-kD polypeptide of heart gap junctions:Immunolocalization, topology, and functional domains. J. Cell Biol.108, 2241-2254.
(Received 16 January 1990 - Accepted 2 March 1990)
238 R. Meyer et al.




















