In vivo toxicity study of N-1-sulfonylcytosine derivatives and their mechanisms of action in...
10
PRECLINICAL STUDIES In vivo toxicity study of N-1-sulfonylcytosine derivatives and their mechanisms of action in cervical carcinoma cell line Jelena Kašnar-Šamprec & Ivana Ratkaj & Katarina Mišković & Marina Pavlak & Mirela Baus-Lončar & Sandra Kraljević Pavelić & Ljubica Glavaš-Obrovac & Biserka Žinić Received: 13 December 2010 / Accepted: 6 March 2011 /Published online: 22 March 2011 # Springer Science+Business Media, LLC 2011 Summary New N-1-sulfonylpyrimidines showed potent growth inhibitory activity against human and mouse tumour cells of different origin. 1-(p-toluenesulfonyl)cytosine (TsC) and 1-(p-toluenesulfonyl)cytosine hydrochloride (TsC × HCl) inhibited the growth of human cervical carcinoma cells (HeLa), and had no significant cytotoxic effects on normal human foreskin fibroblasts (BJ). TsC and TsC × HCl interfered with the HeLa cell cycle progression bringing about the accumulation of G1 phase cells and the induction of apoptosis. Antiproliferative effects of TsC and TsC × HCl were additionally confirmed by investigating de novo synthesis of RNA, DNA and proteins in HeLa cells. Monitoring gene expression using DNA Chip Analysis and quantitative PCR showed that TsC × HCl affects the expression of several cell-cycle regulating genes implying that cell cycle arrest and DNA damage-induced apoptosis might account for the observed cellular effects. In vivo experiments revealed low toxicity of TsC × HCl, as demonstrated by unaltered haematological and metabolic blood parameters. In conclusion, potent antitumour efficacy Electronic supplementary material The online version of this article (doi:10.1007/s10637-011-9657-x) contains supplementary material, which is available to authorized users. J. Kašnar-Šamprec German Heart Center, Technical University Munich, Lazarettstraße 36, 80636 Munich, Germany I. Ratkaj : S. Kraljević Pavelić Department of Biotechnology, University of Rijeka, Trg braće Mažuranića 10, 51000 Rijeka, Croatia K. Mišković : M. Baus-Lončar Laboratory of Functional Genomics, School of Medicine, J. J. Strossmayer University of Osijek, Huttlerova 4, HR-31000 Osijek, Croatia M. Pavlak Department of Veterinary Economics and Analytic Epidemiology, Faculty of Veterinary Medicine, Zagreb University, Zagreb, Croatia M. Baus-Lončar Division of Molecular Medicine, Ruđer Bošković Institute, Bijenička cesta 54, 10000 Zagreb, Croatia L. Glavaš-Obrovac Department of Clinical Chemistry and Biochemistry, School of Medicine, J. J. Strossmayer University of Osijek, Huttlerova 4, HR-31000 Osijek, Croatia L. Glavaš-Obrovac (*) Clinical Hospital Centre Osijek, Clinical Institute of Nuclear Medicine and Radiation Protection, Huttlerova 4, HR-31000 Osijek, Croatia e-mail: [email protected] B. Žinić (*) Laboratory of Supramolecular and Nucleoside Chemistry, Division of Organic Chemistry and Biochemistry, Ruđer Bošković Institute, Bijenička cesta 54, 10000 Zagreb, Croatia e-mail: [email protected] Invest New Drugs (2012) 30:981–990 DOI 10.1007/s10637-011-9657-x
Transcript of In vivo toxicity study of N-1-sulfonylcytosine derivatives and their mechanisms of action in...

PRECLINICAL STUDIES
In vivo toxicity study of N-1-sulfonylcytosine derivativesand their mechanisms of action in cervical carcinoma cell line
Jelena Kašnar-Šamprec & Ivana Ratkaj & Katarina Mišković & Marina Pavlak &
Mirela Baus-Lončar & Sandra Kraljević Pavelić & Ljubica Glavaš-Obrovac &
Biserka Žinić
Received: 13 December 2010 /Accepted: 6 March 2011 /Published online: 22 March 2011# Springer Science+Business Media, LLC 2011
Summary New N-1-sulfonylpyrimidines showed potentgrowth inhibitory activity against human and mouse tumourcells of different origin. 1-(p-toluenesulfonyl)cytosine(TsC) and 1-(p-toluenesulfonyl)cytosine hydrochloride(TsC × HCl) inhibited the growth of human cervicalcarcinoma cells (HeLa), and had no significant cytotoxiceffects on normal human foreskin fibroblasts (BJ). TsC andTsC × HCl interfered with the HeLa cell cycle progressionbringing about the accumulation of G1 phase cells and theinduction of apoptosis. Antiproliferative effects of TsC and
TsC × HCl were additionally confirmed by investigating denovo synthesis of RNA, DNA and proteins in HeLa cells.Monitoring gene expression using DNA Chip Analysis andquantitative PCR showed that TsC × HCl affects theexpression of several cell-cycle regulating genes implyingthat cell cycle arrest and DNA damage-induced apoptosismight account for the observed cellular effects. In vivoexperiments revealed low toxicity of TsC × HCl, asdemonstrated by unaltered haematological and metabolicblood parameters. In conclusion, potent antitumour efficacy
Electronic supplementary material The online version of this article(doi:10.1007/s10637-011-9657-x) contains supplementary material,which is available to authorized users.
J. Kašnar-ŠamprecGerman Heart Center, Technical University Munich,Lazarettstraße 36,80636 Munich, Germany
I. Ratkaj : S. Kraljević PavelićDepartment of Biotechnology, University of Rijeka,Trg braće Mažuranića 10,51000 Rijeka, Croatia
K. Mišković :M. Baus-LončarLaboratory of Functional Genomics, School of Medicine,J. J. Strossmayer University of Osijek,Huttlerova 4,HR-31000 Osijek, Croatia
M. PavlakDepartment of Veterinary Economics and Analytic Epidemiology,Faculty of Veterinary Medicine, Zagreb University,Zagreb, Croatia
M. Baus-LončarDivision of Molecular Medicine, Ruđer Bošković Institute,Bijenička cesta 54,10000 Zagreb, Croatia
L. Glavaš-ObrovacDepartment of Clinical Chemistry and Biochemistry,School of Medicine, J. J. Strossmayer University of Osijek,Huttlerova 4,HR-31000 Osijek, Croatia
L. Glavaš-Obrovac (*)Clinical Hospital Centre Osijek, Clinical Instituteof Nuclear Medicine and Radiation Protection,Huttlerova 4,HR-31000 Osijek, Croatiae-mail: [email protected]
B. Žinić (*)Laboratory of Supramolecular and Nucleoside Chemistry,Division of Organic Chemistry and Biochemistry,Ruđer Bošković Institute,Bijenička cesta 54,10000 Zagreb, Croatiae-mail: [email protected]
Invest New Drugs (2012) 30:981–990DOI 10.1007/s10637-011-9657-x

and low toxicity of new compounds in comparison with thecommon chemotherapy drug 5-FU make them promisinganticancer agents. Additional pre-clinical and clinicalstudies are warranted to illuminate the mode of action ofthese newly synthesized compounds in vivo, which wouldlay the groundwork for their further optimization.
Keywords N-1-sulfonylcytosine derivatives . Antitumouragents . In vitro screening . In vivo screening
AbbreviationsTsC 1-(p-toluenesulfonyl)cytosineTsC × HCl 1-(p-toluenesulfonyl)cytosine hydrochloridePRKDC protein kinase, DNA-activated, catalytic
polypeptideNFKBIA Homo sapiens nuclear factor of kappa light
polypeptide gene enhancer in B-cells inhibitorαCCNE1 Cyclin E1COL18A1 Homo sapiens collagen, type XVIII, α1APAF1 Human apoptotic protease activating factorCHEK 2 Human CHC2 checkpoint homologITGB5 Human integrin, β5ITGB 3 Human integrin, β3 (platelet glycoprotein
IIIa, antigen CD61)MMP2 Human metallopeptidase 2SDHA Homo sapiens succinate dehydrogenase
complex, subunit A, flavoprotein (Fp)PPIA Peptidylprolyl isomerase A (Cyclophilin A)B2M β-2-microglobulinQPCR Quantitative real time PCR
Introduction
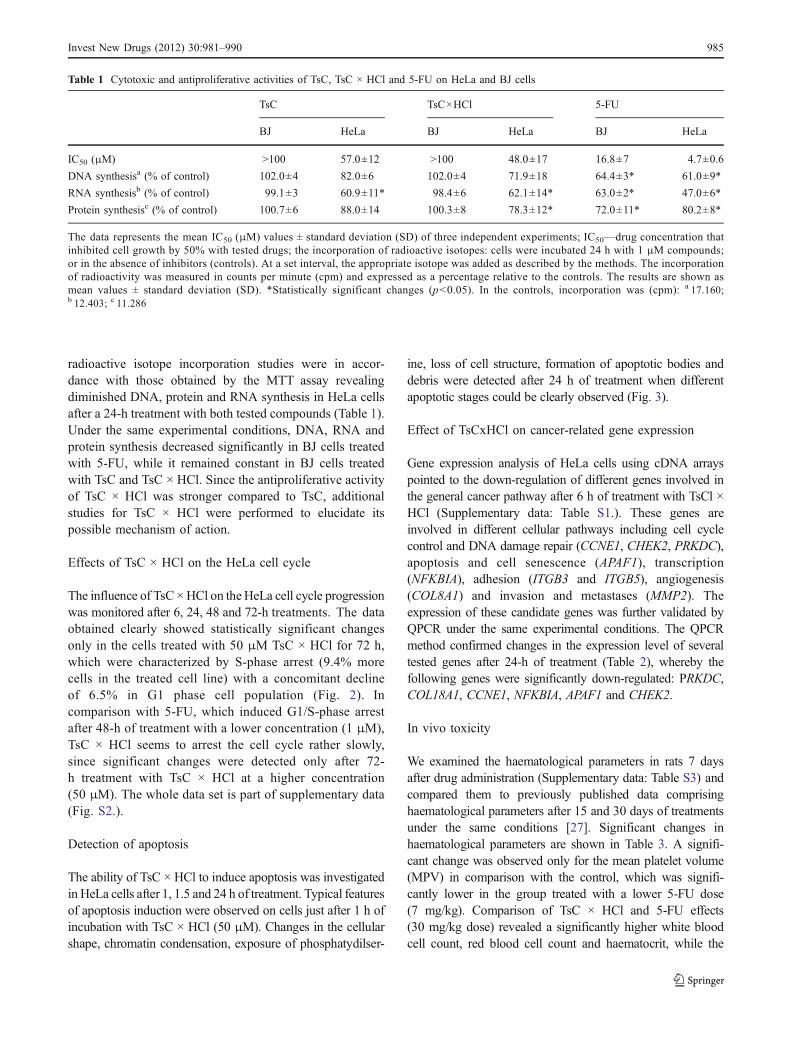
Antimetabolites are commonly used chemotherapeuticagents with the proven ability to effectively suppressuncontrolled cellular division within a heterogeneous tumorcell population. We have recently prepared a series of novelpyrimidine derivatives (Fig. 1) with a sulfonamide and anucleobase pharmacophore [1–3]. Newly synthesized N-1-
sulfonylpyrimidine compounds can be classified as anti-metabolites, a group of anticancer drugs characterized bytheir similarity to naturally occurring nucleobases. From thepoint of view of biological activity, these compounds canbe perceived as a combination of two separate biologicallyactive components: a sulfonylcyclourea fragment and anucleobase.
Sulfonylurea and its derivatives have caught the attentionof medical and agricultural researchers due to their herbicidal[4, 5], antihypoglycemic [6, 7], anticholesterolemic [8] andantitumour activity [9, 10]. In addition, it is known thatmodified nucleosides and nucleotides reduce the cell mitoticactivity resulting in antineoplastic [11–16] and antiviral [17]activity. 5-fluorouracil, a pyrimidine analogue and represen-tative antimetabolite drug, is currently used as a first-linechemotherapeutic agent for the treatment of different solidtumours [18–21]. However, the clinical efficacy of 5-fluorouracil is compromised by its propensity to causeseveral types of dose-dependent toxicity [20, 21].
New N-1-sulfonylpyrimidine derivatives showed strongcytostatic activity in human tumour cell lines in vitro [22].In comparison with 5-fluorouracil, some of these com-pounds showed 10-fold stronger antiproliferative effects,while the effects on normal human cell lines weresignificantly lower. Most of the compounds showedsignificant in vitro antitumour activity on the followinghuman tumour cell lines: pancreas (MIAPaCa-2), colon(CaCo-2 and HT-29), cervix (HeLa) and lymph nodemetastasis of human colorectal cancer (SW620) [22]. Itwas also demonstrated that N-1-sulfonylpyrimidine deriva-tives inhibit DNA, RNA and protein synthesis, and have theability to induce apoptosis in treated tumour cells [22, 23].The antimetabolic effects are partially attributable to theinhibition of de novo biosynthesis of purine and pyrimidinebases and the inhibition of some enzymes including DNApolymerase, RNA polymerase, ribonucleotide reductase,orotatphosphoribosyl transferase and thymidin kinase [22, 24].
1-(p-toluenesulfonyl)cytosine (TsC) showed good selec-tivity with regard to normal cells and detailed computationalstudies were performed as to classify them according to theirmechanism of action. Collectively, the computational resultsalong with data on cell cycle perturbation and apoptosisinduction point to an unusual mechanism of cytostatic action,possibly a combination of nucleic acid antimetabolite activityand a novel molecular mechanism [25].
TsC and its two derivatives, namely 1-(p-toluenesul-fonyl)cytosine hydrochloride (TsC×HCl) and zinc(II) com-plex [Zn(II)(TsC)2] were shown to have a strong antitumouractivity against mouse mammary carcinoma in vivo [26].Toxicological studies in vivo showed that TsC and TsC ×HCl have no significant influence on the peripheral bloodthrombocyte number, red blood cell count or haemoglobinlevels, while significant deviations from the untreated
N
NN
HN
S
O
OO
O
O
CH3
H
N
N
SO
O
O
H
R
R'
R = O or NH
N
N
S
OO
O
CH3
NH2
SULFONYLUREAN-SULFONYLPYRIMIDINES
R' = alkyl, aromatic, heteroaromaticTsC
Fig. 1 N-1-Sulfonylpyrimidine derivatives
982 Invest New Drugs (2012) 30:981–990

control group were detected in the peripheral white bloodcell count [27].
Due to the observed low cytotoxicity, we conductedadditional biological studies of these potent antimetaboliccompounds to comprehend their biological mechanisms ofaction. For that purpose, we analyzed their antiproliferative,cytotoxic and metabolic effects and time-dependent effecton the cell cycle in HeLa cells. In particular, the modulationof expression of 80 cancer-related genes was determined.Finally, we estimated the toxicological effects in an animalmodel.
Material and methods
Synthesis of test compounds
The compounds were synthesized in the Laboratory ofSupramolecular and Nucleoside Chemistry of the RuđerBošković Institute (Zagreb, Croatia). Synthesis of TsC andTsC × HCl was performed according to the procedurereported by Kašnar-Šamprec et al. [27]. Condensation ofsilylated cytosine with p-toluenesulfonyl chloride in aceto-nitrile gave 1-(p-toluenesulfonyl)cytosine (TsC) in 80%yield after recrystallization from hot methanol. Afterheating under reflux for 45 min and evaporating thesolvent, if the residue was treated with 3% HCl/CH3OH,the resulting white crystals of 1-(p-toluenesulfonyl)cytosinehydrochloride (TsC×HCl) were obtained in 75% yield.
Cell lines
Cervical carcinoma cells (HeLa) and human foreskinfibroblast (BJ; 24–27 passages) were cultured as mono-layers in Dulbecco’s modified Eagle medium—DMEM(Gibco, EU) supplemented with 10% heat-inactivated foetalbovine serum (Gibco, EU), 2 mM glutamine, and 100U/0.1 mg penicillin/streptomycin. Cells were cultured in ahumidified atmosphere (37°C/5% of CO2) in a CO2
incubator (Shell Lab, Sheldon Manufacturing, USA). Cellswere tested and authenticated prior to experiments using theshort tandem repeat profiling (STR) analysis.
Cytotoxicity evaluation
Cytotoxicity was assessed by the MTT assay [28]. TsC andTsC × HCl were prepared as stock solutions (10 mM) inwarm high-purity water. Working solutions in the concen-tration range of 10 mM–1 μM were prepared prior totesting. Cells were seeded in 96 micro-well plates (Greiner,Austria) at a concentration of 2×104 cells/mL and leftovernight in the CO2 incubator. The growth medium wasreplaced with either a medium containing tested com-
pounds or a control medium and incubated for 72 h. A freshmedium with 5 mg/mL of MTT was added to the cells andincubated for 4 h at 37°C. Upon removal of the medium,water insoluble MTT-formazan crystals that had formedinside the living cells were dissolved in DMSO, and theabsorbance at 570 nm proportional to the number of livingcells was measured on an Elisa Microplate Reader (Stat fax2100, Pharmacia Biotech, Uppsala, Sweden). All experi-ments were performed at least three times in triplicates. Thepercentage of cell growth (PG) was calculated using thefollowing equation: PG ¼ A� ABLANCK=ACONTROL�ðABLANCKÞ � 100; BLANCK = medium without cells con-taining compounds and MTT.
Incorporation of radioactive isotopes
HeLa cells were incubated for 24 h in a medium containing1 μM TsC or TsC × HCl, or in the absence of an inhibitor(control cells). The appropriate isotope was added at setintervals [29]. The amounts of incorporated radioactivity inthe cellular DNA, RNA and proteins of the control andtreated cells were determined by measuring the uptake ofthe labelled precursors [2-14C]-thymidine (0.025 μCi/well),[5-3H]-uridine (0.5 μCi/well) or L-[1-14C]-leucine(0.0125 μCi/well) into cold trichloracetic acid-insolublematerial. All of the chemicals were purchased from Sigma,USA. The incorporation of radioactivity was measured incounts per minute (cpm) and expressed as a percentagerelative to the control. The results are presented as meanvalues ± standard deviation. The statistical evaluation wasdone with Statistica™ software using the Kruskal-WallisANOVA test and a multiple comparison approach with a95% confidence interval.
Detection of apoptosis
The Annexin-V-FLUOS staining kit (Roche, Penzberg,Germany) was used for the detection of apoptosis anddifferentiation from necrosis at the single-cell level. Experi-ments were performed according to the manufacturer’sinstructions. In short, 1×105 HeLa cells were seeded onchamber slides with silicon borders, incubated in DMEMfor 24 h and then treated with TsC × HCl (10 μM and50 μM) for 1, 1.5 and 24 h. The cells were analyzed using afluorescent microscope (Axioskop 2 MOT, Carl Zeiss JenaGmbH, Jena, Germany); excitation 450–500 nm andemission 515–516 nm.
Gene expression
HeLa cells (1.4×106) were seeded in 10 mm dishes(Falcon, USA), and incubated in a humidified atmospherewith 5% CO2 at 37°C for 24 h. The growth medium was
Invest New Drugs (2012) 30:981–990 983

replaced with either TsC × HCl- supplemented (10 μM)medium or a control medium and incubated for 72 h. TotalRNA was isolated after 6 h and 24 h in three biologicalreplicates using the RNAeasy kit (QIAGEN, USA).Commercial arrays containing highly specific 96 base-pairfragments of genes related to biological pathways altered oraffected by transformation and tumourigenesis (HumanCancer Pathway Finder™ cDNK GEArray, Q Series HS-006, SuperArray, USA) were used in accordance with themanufacturer’s recommendations. A total of 3 μg of theobtained RNA was used per sample. The signal wascaptured with a digital camera (VersaDoc, Bio-Rad Labo-ratories). The obtained images were analyzed using thecommercial software GEArray Expression Analysis Suite.Reference genes (CD44, ITGA3, MICA, TGFBR1, PPIA)with a stable expression in all tested treatments were usedfor normalization, and a minimal value was used forbackground subtraction. Normalized values were statisti-cally analyzed in all groups with SPSS (SPSS®forWindows) by ANOVA (at p<0.05, Tukey’s HSD for post-hoc comparisons). The candidate genes from cDNA arrayexperiments (Supplementary data: Table S1. and Fig. S1)were further validated by the quantitative Real-time PCRmethod (QPCR) on 16 samples per group. The isolatedRNA (800 ng/reaction) was transcribed into cDNA usingthe iScript™ Select cDNA Synthesis Kit with polyA primerapproach (Bio-Rad Laboratories). We used the SyberGreendetection approach and assured optimized reactions withone specific product (confirmed by the melting curve andgel analysis) and PCR reaction efficiency of 95–100%(Details in Supplement; Table S2.). Reference genes werechosen based on pre-screening samples with the panel ofhousekeeping genes PPIA, HPRT, YWHAZ, B2M, UBC,HMBS, RPL13a, 18SRNA, SDHA and GUS. We chose 4genes: 18SRNA, SDHA, PPIA and B2M. Further analysisshowed that 18SRNA was the only stable gene, andtherefore we used it as a reference gene. The determinedct values were analyzed using the Relative ExpressionSoftware Tool (REST) [30].
Cell cycle analysis
HeLa cells (2×105 cells/well) were seeded in 6-well plates(Falcon, USA). After 24 h, TsC × HCl was added at aconcentration of 10 μM and 50 μM. Cells were harvested atdifferent intervals (6, 24, 48 and 72 h) following the drugtreatment. Floating and adherent cells were collectedseparately, then combined, washed with phosphate buffersaline (PBS), fixed with 70% ethanol, and stored at −20°C.Immediately before analysis, cell pellets were washed withPBS and resuspended in 1 μg/mL of propidium iodide and0.2 μg/μL of RNAse A. The stained cells were thenanalyzed with a Becton Dickinson FACScalibur flow
cytometer, whereby 20,000 events were recorded. Each testpoint was performed in duplicate in three individualexperiments. The results are shown as mean percentagesfrom three separate experiments for each tested group. Thepercentage of cells in each cell cycle phase was based onthe obtained DNA histograms and determined usingWinMDI and Cylchred software. A statistical analysis wasperformed in Microsoft Excel using ANOVA at p<0.05.
In vivo toxicity
Wistar rats (13–15 weeks old) were used for the experiments.This was approved by the local ethics committee andperformed in line with the guidelines of the European animalwelfare protocol. TsC × HCl (7 and 30 mg/kg) 5-fluorouracil(7 and 30 mg/kg) and isotonic solution (control) wereadministered intraperitoneally for three consecutive days intwo different concentrations to separate groups of animals (N=6). Blood samples were collected 7, 15 and 30 days after theadministration of the first compound. The following meta-bolic parameters were analyzed in serum: urea, creatinine,proteins, albumin, total bilirubin count (Tbilc), glucose,aspartate–aminotranspherase (AST), alanine–aminotranspher-ase (ALT), inorganic phosphate, magnesium, triglyceridesand cholesterol. Haematological parameters (WBC: whiteblood cells, RBC: red blood cells, Hgb: haemoglobineconcentration, Htc: haematocrit, MCV: mean cell volume,MCH: mean cell haemoglobin, MCHC: mean cell haemo-globin concentration, RDW: red cell distribution width, Plt:platelets, MPV: mean platelet volume) were analyzed onthe seventh day after the treatment and compared to thealready published results of a haematological analysisthat was done 15 and 30 days following treatment withthe same doses of applied substances [27]. A statisticalevaluation was made with Statistica™ software using theKruskal-Wallis ANOVA test and multiple comparisonapproach with a 95% confidence interval.
Results
Cytotoxic and antiproliferative activities in vitro
As evidenced from the obtained MTT results, TsC × HClexerted somewhat stronger antiproliferative effect on HeLacells than TsC (Table 1). Cytotoxic effects of TsC × HCland TsC were significantly lower in BJ cells relative toHeLa cells (IC50 >100 μM vs. IC50 ~50 μM, respectively).BJ cell survival rate ranged from 118% at the lowest(0.1 μM) to 61% at the highest applied (0.1 mM)concentration. Compared to 5-FU cytotoxicity, the testedcompounds showed significantly lower toxicity towardsboth normal and tumour cell lines. The results of the
984 Invest New Drugs (2012) 30:981–990
radioactive isotope incorporation studies were in accor-dance with those obtained by the MTT assay revealingdiminished DNA, protein and RNA synthesis in HeLa cellsafter a 24-h treatment with both tested compounds (Table 1).Under the same experimental conditions, DNA, RNA andprotein synthesis decreased significantly in BJ cells treatedwith 5-FU, while it remained constant in BJ cells treatedwith TsC and TsC × HCl. Since the antiproliferative activityof TsC × HCl was stronger compared to TsC, additionalstudies for TsC × HCl were performed to elucidate itspossible mechanism of action.
Effects of TsC × HCl on the HeLa cell cycle
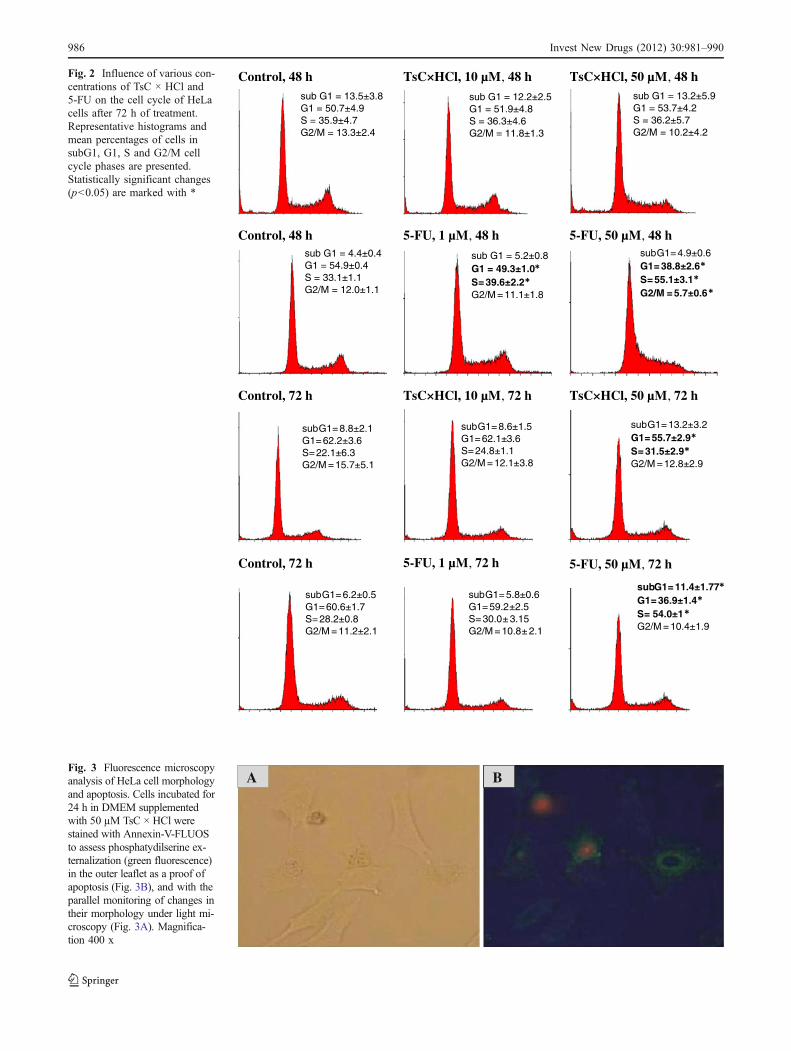
The influence of TsC ×HCl on theHeLa cell cycle progressionwas monitored after 6, 24, 48 and 72-h treatments. The dataobtained clearly showed statistically significant changesonly in the cells treated with 50 μM TsC × HCl for 72 h,which were characterized by S-phase arrest (9.4% morecells in the treated cell line) with a concomitant declineof 6.5% in G1 phase cell population (Fig. 2). Incomparison with 5-FU, which induced G1/S-phase arrestafter 48-h of treatment with a lower concentration (1 μM),TsC × HCl seems to arrest the cell cycle rather slowly,since significant changes were detected only after 72-h treatment with TsC × HCl at a higher concentration(50 μM). The whole data set is part of supplementary data(Fig. S2.).
Detection of apoptosis
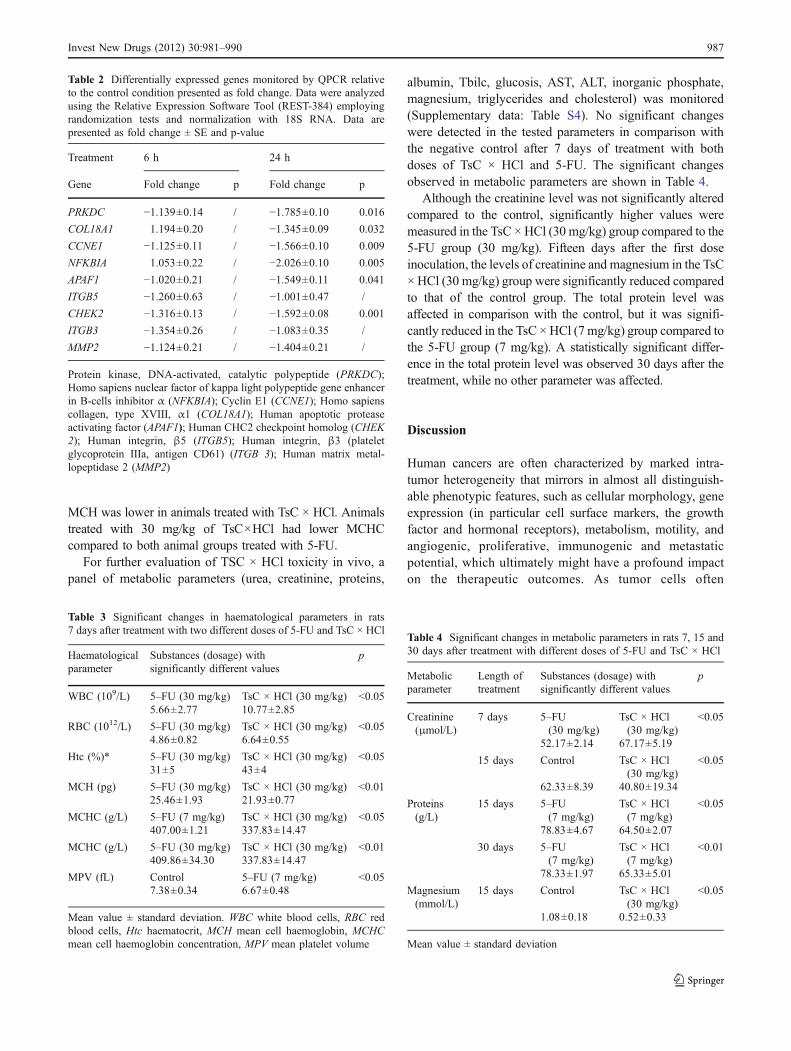
The ability of TsC × HCl to induce apoptosis was investigatedin HeLa cells after 1, 1.5 and 24 h of treatment. Typical featuresof apoptosis induction were observed on cells just after 1 h ofincubation with TsC × HCl (50 μM). Changes in the cellularshape, chromatin condensation, exposure of phosphatydilser-
ine, loss of cell structure, formation of apoptotic bodies anddebris were detected after 24 h of treatment when differentapoptotic stages could be clearly observed (Fig. 3).
Effect of TsCxHCl on cancer-related gene expression
Gene expression analysis of HeLa cells using cDNA arrayspointed to the down-regulation of different genes involved inthe general cancer pathway after 6 h of treatment with TsCl ×HCl (Supplementary data: Table S1.). These genes areinvolved in different cellular pathways including cell cyclecontrol and DNA damage repair (CCNE1, CHEK2, PRKDC),apoptosis and cell senescence (APAF1), transcription(NFKBIA), adhesion (ITGB3 and ITGB5), angiogenesis(COL8A1) and invasion and metastases (MMP2). Theexpression of these candidate genes was further validated byQPCR under the same experimental conditions. The QPCRmethod confirmed changes in the expression level of severaltested genes after 24-h of treatment (Table 2), whereby thefollowing genes were significantly down-regulated: PRKDC,COL18A1, CCNE1, NFKBIA, APAF1 and CHEK2.
In vivo toxicity
We examined the haematological parameters in rats 7 daysafter drug administration (Supplementary data: Table S3) andcompared them to previously published data comprisinghaematological parameters after 15 and 30 days of treatmentsunder the same conditions [27]. Significant changes inhaematological parameters are shown in Table 3. A signifi-cant change was observed only for the mean platelet volume(MPV) in comparison with the control, which was signifi-cantly lower in the group treated with a lower 5-FU dose(7 mg/kg). Comparison of TsC × HCl and 5-FU effects(30 mg/kg dose) revealed a significantly higher white bloodcell count, red blood cell count and haematocrit, while the
Table 1 Cytotoxic and antiproliferative activities of TsC, TsC × HCl and 5-FU on HeLa and BJ cells
TsC TsC×HCl 5-FU
BJ HeLa BJ HeLa BJ HeLa
IC50 (μM) >100 57.0±12 >100 48.0±17 16.8±7 4.7±0.6
DNA synthesisa (% of control) 102.0±4 82.0±6 102.0±4 71.9±18 64.4±3* 61.0±9*
RNA synthesisb (% of control) 99.1±3 60.9±11* 98.4±6 62.1±14* 63.0±2* 47.0±6*
Protein synthesisc (% of control) 100.7±6 88.0±14 100.3±8 78.3±12* 72.0±11* 80.2±8*
The data represents the mean IC50 (μM) values ± standard deviation (SD) of three independent experiments; IC50—drug concentration thatinhibited cell growth by 50% with tested drugs; the incorporation of radioactive isotopes: cells were incubated 24 h with 1 μM compounds;or in the absence of inhibitors (controls). At a set interval, the appropriate isotope was added as described by the methods. The incorporationof radioactivity was measured in counts per minute (cpm) and expressed as a percentage relative to the controls. The results are shown asmean values ± standard deviation (SD). *Statistically significant changes (p<0.05). In the controls, incorporation was (cpm): a 17.160;b 12.403; c 11.286
Invest New Drugs (2012) 30:981–990 985
Control, 48 h TsC×HCl, 10 µM, 48 h TsC×HCl, 50 µM, 48 h
Control, 48 h 5-FU, 1 µM, 48 h 5-FU, 50 µM, 48 h
TsC×HCl, 10 µM, 72 h TsC×HCl, 50 µM, 72 h
sub G1 = 13.5±3.8G1 = 50.7±4.9S = 35.9±4.7G2/M = 13.3±2.4
sub G1 = 12.2±2.5G1 = 51.9±4.8S = 36.3±4.6G2/M = 11.8±1.3
sub G1 = 13.2±5.9G1 = 53.7±4.2S = 36.2±5.7G2/M = 10.2±4.2
sub G1 = 4.4±0.4G1 = 54.9±0.4S = 33.1±1.1G2/M = 12.0±1.1
sub G1 = 5.2±0.8G1 = 49.3±1.0S=39.6±2.2G2/M =11.1±1.8
subG1=4.9±0.6G1=38.8±2.6S=55.1±3.1G2/M =5.7±0.6
subG1=8.6±1.5G1=62.1±3.6S=24.8±1.1G2/M =12.1±3.8
subG1=13.2±3.2G1=55.7±2.9S= 31.5±2.9G2/M =12.8±2.9
subG1=8.8±2.1G1=62.2±3.6S=22.1±6.3G2/M =15.7±5.1
Control, 72 h
Control, 72 h
5-FU, 1 µM, 72 h 5-FU, 50 µM, 72 h
subG1=6.2±0.5G1=60.6±1.7S=28.2±0.8G2/M =11.2±2.1
subG1=5.8±0.6G1=59.2±2.5S=30.0±3.15G2/M = 10.8±2.1
subG1= 11.4±1.77G1=36.9±1.4S= 54.0±1G2/M =10.4±1.9
Fig. 2 Influence of various con-centrations of TsC × HCl and5-FU on the cell cycle of HeLacells after 72 h of treatment.Representative histograms andmean percentages of cells insubG1, G1, S and G2/M cellcycle phases are presented.Statistically significant changes(p<0.05) are marked with *
A BFig. 3 Fluorescence microscopyanalysis of HeLa cell morphologyand apoptosis. Cells incubated for24 h in DMEM supplementedwith 50 µM TsC × HCl werestained with Annexin-V-FLUOSto assess phosphatydilserine ex-ternalization (green fluorescence)in the outer leaflet as a proof ofapoptosis (Fig. 3B), and with theparallel monitoring of changes intheir morphology under light mi-croscopy (Fig. 3A). Magnifica-tion 400 x
986 Invest New Drugs (2012) 30:981–990
MCH was lower in animals treated with TsC × HCl. Animalstreated with 30 mg/kg of TsC×HCl had lower MCHCcompared to both animal groups treated with 5-FU.
For further evaluation of TSC × HCl toxicity in vivo, apanel of metabolic parameters (urea, creatinine, proteins,
albumin, Tbilc, glucosis, AST, ALT, inorganic phosphate,magnesium, triglycerides and cholesterol) was monitored(Supplementary data: Table S4). No significant changeswere detected in the tested parameters in comparison withthe negative control after 7 days of treatment with bothdoses of TsC × HCl and 5-FU. The significant changesobserved in metabolic parameters are shown in Table 4.
Although the creatinine level was not significantly alteredcompared to the control, significantly higher values weremeasured in the TsC × HCl (30 mg/kg) group compared to the5-FU group (30 mg/kg). Fifteen days after the first doseinoculation, the levels of creatinine and magnesium in the TsC× HCl (30 mg/kg) group were significantly reduced comparedto that of the control group. The total protein level wasaffected in comparison with the control, but it was signifi-cantly reduced in the TsC × HCl (7 mg/kg) group compared tothe 5-FU group (7 mg/kg). A statistically significant differ-ence in the total protein level was observed 30 days after thetreatment, while no other parameter was affected.
Discussion
Human cancers are often characterized by marked intra-tumor heterogeneity that mirrors in almost all distinguish-able phenotypic features, such as cellular morphology, geneexpression (in particular cell surface markers, the growthfactor and hormonal receptors), metabolism, motility, andangiogenic, proliferative, immunogenic and metastaticpotential, which ultimately might have a profound impacton the therapeutic outcomes. As tumor cells often
Table 2 Differentially expressed genes monitored by QPCR relativeto the control condition presented as fold change. Data were analyzedusing the Relative Expression Software Tool (REST-384) employingrandomization tests and normalization with 18S RNA. Data arepresented as fold change ± SE and p-value
Treatment 6 h 24 h
Gene Fold change p Fold change p
PRKDC −1.139±0.14 / −1.785±0.10 0.016
COL18A1 1.194±0.20 / −1.345±0.09 0.032
CCNE1 −1.125±0.11 / −1.566±0.10 0.009
NFKBIA 1.053±0.22 / −2.026±0.10 0.005
APAF1 −1.020±0.21 / −1.549±0.11 0.041
ITGB5 −1.260±0.63 / −1.001±0.47 /
CHEK2 −1.316±0.13 / −1.592±0.08 0.001
ITGB3 −1.354±0.26 / −1.083±0.35 /
MMP2 −1.124±0.21 / −1.404±0.21 /
Protein kinase, DNA-activated, catalytic polypeptide (PRKDC);Homo sapiens nuclear factor of kappa light polypeptide gene enhancerin B-cells inhibitor α (NFKBIA); Cyclin E1 (CCNE1); Homo sapienscollagen, type XVIII, α1 (COL18A1); Human apoptotic proteaseactivating factor (APAF1); Human CHC2 checkpoint homolog (CHEK2); Human integrin, β5 (ITGB5); Human integrin, β3 (plateletglycoprotein IIIa, antigen CD61) (ITGB 3); Human matrix metal-lopeptidase 2 (MMP2)
Table 3 Significant changes in haematological parameters in rats7 days after treatment with two different doses of 5-FU and TsC × HCl
Haematologicalparameter
Substances (dosage) withsignificantly different values
p
WBC (109/L) 5–FU (30 mg/kg) TsC × HCl (30 mg/kg) <0.055.66±2.77 10.77±2.85
RBC (1012/L) 5–FU (30 mg/kg) TsC × HCl (30 mg/kg) <0.054.86±0.82 6.64±0.55
Htc (%)* 5–FU (30 mg/kg) TsC × HCl (30 mg/kg) <0.0531±5 43±4
MCH (pg) 5–FU (30 mg/kg) TsC × HCl (30 mg/kg) <0.0125.46±1.93 21.93±0.77
MCHC (g/L) 5–FU (7 mg/kg) TsC × HCl (30 mg/kg) <0.05407.00±1.21 337.83±14.47
MCHC (g/L) 5–FU (30 mg/kg) TsC × HCl (30 mg/kg) <0.01409.86±34.30 337.83±14.47
MPV (fL) Control 5–FU (7 mg/kg) <0.057.38±0.34 6.67±0.48
Mean value ± standard deviation. WBC white blood cells, RBC redblood cells, Htc haematocrit, MCH mean cell haemoglobin, MCHCmean cell haemoglobin concentration, MPV mean platelet volume
Table 4 Significant changes in metabolic parameters in rats 7, 15 and30 days after treatment with different doses of 5-FU and TsC × HCl
Metabolicparameter
Length oftreatment
Substances (dosage) withsignificantly different values
p
Creatinine(μmol/L)
7 days 5–FU(30 mg/kg)
TsC × HCl(30 mg/kg)
<0.05
52.17±2.14 67.17±5.19
15 days Control TsC × HCl(30 mg/kg)
<0.05
62.33±8.39 40.80±19.34
Proteins(g/L)
15 days 5–FU(7 mg/kg)
TsC × HCl(7 mg/kg)
<0.05
78.83±4.67 64.50±2.07
30 days 5–FU(7 mg/kg)
TsC × HCl(7 mg/kg)
<0.01
78.33±1.97 65.33±5.01
Magnesium(mmol/L)
15 days Control TsC × HCl(30 mg/kg)
<0.05
1.08±0.18 0.52±0.33
Mean value ± standard deviation
Invest New Drugs (2012) 30:981–990 987

acquire a resistance to anti-cancer treatment, noveltherapeutic strategies are urgently needed. Frequentlyused cytostatic drugs are antimetabolites such as 5-fluorouracil (5-FU), which incorporates into the DNAand RNA, and inhibits the enzyme thymidylate synthetase.The antitumour effect of this drug is compromised ofunwanted dose-specific side effects such as nephrotoxicityand neurotoxicity [31]. Additionally, drug resistance due tothe increased activity of thymidylate synthetase and itsaffinity for 5-FU preclude this drug from making its way intoclinical practice [32].
N-sulfonylpyrimidine derivatives appear appealingowing to two separate biologically active components: asulfonylcyclourea fragment and a nucleobase. 1-(p-toluene-sulfonyl)cytosine (TsC) and its more soluble form 1-(p-toluenesulfonyl)cytosine hydrochloride (TsC × HCl)inhibit the growth of various cancer cell lines [25, 26],and have no significant cytotoxic effect on normal humanforeskin fibroblasts. TsCl × HCl affected the HeLa cellcycle after 48 h of treatment. However, marked changeswere observed after 72 h, when the number of cells in theS-phase rose accompanied by a decrease in the number ofcells entering the G1-phase. Similarly, 5-FU induced a G1/S-phase arrest after 48 h of treatment with a lowerconcentration (1 μM). The mechanism of action of TsCand TsC × HCl might involve two distinct mechanisms.The first relies on their direct incorporation into the DNA/RNA, which reduces the rate of nucleic acid synthesis,whereas the second might involve a direct influence on theactivity of enzymes indispensable for de novo nucleic acidsynthesis. TsC was able to inhibit RNA polymerase (37%),orotate phosphoribosyltransferase (39%), uridine kinase(44%), ribonucleotide reductase (47%), and de novo purinesynthesis (61%). Antitumour activity of 1-(p-toluenesul-fonyl)cytosine (TsC) has been closely associated with itsinhibitory activity on enzymes, which plays an importantrole in the metabolism of tumour cells [23].
Cellular mechanisms that switch between survival, cellcycle arrest and death are complex and usually regulated atdifferent molecular levels. To gain a better appreciation ofthese mechanisms, we assessed the expression status ofvarious cancer-related genes in HeLa cells after 24 h oftreatment with TsCl × HCl. We showed that the expressionof several genes was significantly affected, including thegenes involved in DNA damage and repair (CCNE1,CHEK2, PRKDC), apoptosis and cell senescence (APAF1),angiogenesis (COL8A1) and transcription (NFKBIA). Themost prominent alteration (2-fold down-regulation) wasobserved for NFKBIA, which is a member of the NF-κBinhibitor family. Our results are in agreement with literaturefindings, since the NF-κB transcription factor has beenfound to regulate the expression of numerous oncogenes,tumour suppressor genes, growth factors and cell adhesion
molecules [33, 34]. Through these mechanisms, the NF-κBtranscription factor controls cell death processes (apoptosis)and has been associated with the increased survival ofapoptosis-resistant tumour cells [35, 36]. In particular, E6and E7 proteins, constitutively expressed in HeLa cellsused in our study, have also been implicated in theactivation of the NF-κB gene. This event has been shownto be associated with the development of premalignant andpapillomavirus–associated tumours [37]. Down-regulationof the NF-κB family members has already emerged as apromising targeted therapy in cancer treatment [38, 39].Furthermore, DNA-dependent protein kinase (PRKDC),which plays a key role in the process of double-strandDNA break repair and control of the DNA damagecheckpoint, was down-regulated by TsClxHCl. As shownby some authors, the mechanism of action of somechemotherapeutics involves down-regulation of PRKDC[40]. It therefore seems logical to presume that treatmentof HeLa cells with TsClxHCl triggers DNA damage-mediated apoptosis, especially when having in mind thatN-sulfonylcytosine derivatives were already shown toexert an atypical cytostatic mechanism of action [25]. Inthis study, TsCl × HCl proved to interfere with DNAsynthesis, as demonstrated by S-phase arrest, reducedDNA, protein and RNA synthesis, as well as down-regulation of CCNE1, the gene regulating the transitioninto S-phase through activation of the cyclin dependentkinases. Therefore, abrogation of the cell cycle progres-sion could be partially attributable to down-regulation ofthis gene’s expression [41]. TsCl × HCl seems toconcomitantly inhibit DNA repair processes, as witnessedby the down-regulation of the CHEK2 gene. However, theobserved changes in PRKDC expression can repress thepreferential translation of specific mRNAs, so its impacton cellular events outgrows the simple DNA repair role[42]. Apoptosis induction revealed by Annexin V assaywas in contradiction with the observed down-regulation ofAPAF1. APAF1 was previously recognized as a pro-apoptotic molecule that sets off caspase activation aftertriggering the mitochondrial pathway [43]. Nevertheless, itseems that recent data suggest other apoptotic pathways tobe activated by the concomitant down-regulation ofAPAF1, which might account for data obtained in thisstudy as well [44, 45].
N-sulfonylcytosine derivatives have already been shownto have an atypical cytostatic mechanism of action [25],which is in line with the in vitro data obtained in thepresented study. The performed computational results,together with cell cycle perturbation and apoptosis analysisfor various cell lines point to an unusual mechanism ofcytostatic action, possibly a combination of nucleic acidantimetabolite activity and a novel molecular mechanismthat needs further investigation.
988 Invest New Drugs (2012) 30:981–990

The obtained in vitro results showed pronouncedantiproliferative activity and put forward several cancermarkers as potential targets of TsC × HCl. We thereforeconducted our study on Wistar rats to assess the physio-logical effects of TsC × HCl and compare it to 5-FU. Forthat purpose, we monitored various haematological andmetabolic parameters.
Haematological parameters were examined in rats 7 daysafter the administration of drugs at two different doses andwere compared with previously published haematologicaldata [27] collected after 15 and 30 days of treatments. Ourdata, collected 7 days after treatment, showed a statisticallyhigher red blood cell count and higher haematocrite in thegroup treated with TsC × HCl (30 mg/kg) compared to thegroup treated with 5-FU (30 mg/kg). This suggests a lowertoxicity of TsC × HCl for erythropoesis. Moreover, animalstreated with 30 mg/kg of TsC × HCl displayed asignificantly higher white blood cell count on day 7 afterthe treatment compared to animals treated with 30 mg/kg 5-FU. It was previously shown that the white blood cell(WBC) count significantly increased after 15 days oftreatment with TsC × HCl and 30 days after treatment with5-FU, indicating different dynamic patterns of influence onWBC for TsC × HCl and 5-FU [27]. Interestingly, the WBCcount resumed to the control value after 30 days oftreatment with TsC × HCl, while the 30-day treatment with5-FU increased WBC values. A further toxicity evaluationin vivo was comprised of an assessment of a panel ofmetabolic parameters including urea, creatinine, proteins,albumin, Tbilc, glucosis, AST, ALT, inorganic phosphate,magnesium, triglycerides and cholesterol. A higher dose(30 mg/kg) of TsC × HCl caused a statistically significantreduction in creatinine and magnesium levels 15 days afterthe treatment in comparison with the control group. Sincethe level of creatinine is inversely proportional to glomer-ular kidney function, it can be concluded that the testeddoses of TsC × HCl do not exert toxic effects on thekidneys. This is additionally supported by the normalvalues of the blood urea. The reduction in magnesium canbe caused by various factors, including pancreatitis,thyroid dysfunction or other hormonal changes. Anotherpossible reason could be the additional activation ofenzyme pathways where magnesium plays an active role.As the reduction in the magnesium level was notobserved on day 30 following the treatment, it can beconcluded that TsC × HCl causes a temporary, clinicallyinsignificant depletion of serum magnesium. Similarly,total protein levels in the group treated with a low TsC ×HCl dose were closer to the control group values and thedifference compared to the values for the low 5-FU dosetreated animals was significant. This additionally shows alesser impact of TsC × HCl on the total protein count inblood.
In conclusion, the herein presented data from in vitroand in vivo studies provide evidence of DNA damage-mediated apoptosis induced in HeLa cells, which constitu-tively express human papilloma virus oncoproteins E6 andE7. Considering the low cytotoxic effects of tested N-sulfonylpyrimidine derivatives on normal human fibro-blasts, as well as the low toxic effects proven in the invivo rat model, further optimization of the tested deriva-tives may be pursued for the purpose of cancer treatment,either as primary cytostatic agents or as adjuvants.Additional studies might help broaden the knowledge ofthe molecular mechanism of action of N-sulfonylpyrimidinederivatives for different tumour types.
Acknowledgements This work was supported by the Ruđer Boš-ković Institute’s spin-off company BioZyne d.o.o., and the Ministry ofScience, Education and Sports of the Republic of Croatia throughGrants No. 098-0982914-2935; 219-0982914-2176; 219-0982914-2179; 335-0982464-2393. The authors are grateful to Goran Ćurić,MD, for performing the cell authentication analysis.
References
1. Kašnar B, Krizmanić I, Žinić M (1997) Synthesis of thesulfonylpyrimidine derivates as a new type of sulfonylcycloureas.Nucleosides Nucleotides 16:1067–1071
2. Žinić B, Krizmanić I, Vikić-Topić D, Žinić M (1999) 5-Bromo-and 5-iodo-N-1-sulfonylated cytosine derivatives. Exclusive for-mation of keto-imino tautomers. Croat Chem Acta 72:957–966
3. Žinić B, Žinić M, Krizmanić I (2003) Synthesis of thesulfonylpyrimidine derivatives with anticancer activity. EP 0 877022 B1 (Bulletin 1003/16; 16.04.2003)
4. Brown GM (1962) The biosynthesis of folic acid. Inhibition bysulfonamide. J Biol Chem 237:536–540
5. Hay JV (1990) Chemistry of sulfonylurea herbicides. Pestic Sci29:247–261
6. Aguilar-Bryan L, Nelson DA, Vu QA, Humphrey MB, Boyd AEIII (1990) Photoaffinity labeling and partial purification of the Bcell sulfonylurea receptor using a novel, biologically activeglyburide analog. J Biol Chem 265:8218–8224
7. Hatzidimitrou AG, Kessissoglou DP, Manoussakis GE (1993)Synthesis, characterization and molecular modeling of Cu(II) andAu(III) complexes with hypoglycemic drugs as ligands. J InorgBiochem 49:157–169
8. Slišković R, Krause BR, Picard JA, Anderson M, Bousley RF,Hamelehle KL, Homan R, Julian TN, Rashidbaigi ZA, StanfieldRL (1994) Inhibitors of Acyl-CoA: cholesterol-O-Acyl transferase(ACAT) as Hypocholesterolemic Agents. 6. The First water-soluble ACAT inhibitor with lipid-regulating activity. J MedChem 37:560–562
9. Mohamadi F, Spels MM, Grindey GB (1992) Sulfonylureas: anew class of cancer chemotherapeutic agents. J Med Chem35:3012–3016
10. Howbert JJ, Grossman CS, Crowell TA, Rieder BJ, Harper RW,Kramer KE, Tao EV, Aikins J, Poore GA, Rinzel SM, GrindeyGB, Shaw WN, Todd GC (1990) Novel agents effective againstsolid tumour. The Diarylsulfonylureas. Synthesis, activities andanalysis of quantitative structure-activity relationships. J MedChem 33:2393–2407
Invest New Drugs (2012) 30:981–990 989

11. MacCoss M, Robins MJ (1990) In: Wilman DEV (ed) Chemistryof antitumour agents. Blackie and Son, UK, p 261
12. Badawey ESAM (1996) Synthesis and in vitro evaluation of somenew pyrimidines and related condensed ring system as potentialanticancer agents. J Heterocycl Chem 33:229–233
13. Hattori H, Tanaka M, Fukushima M, Sasaki T, Matsuda A (1996)Nucleosides and nucleotides 158. 1-(3-C-ethynyl-β-D-ribopen-thofuranosyl)-cytosine, 1-(3-C-ethynyl-β-D-ribopenthofuranosyl)uracil, and their nucleobase analogues as new potential multi-functional antitumour nucleosides with a broad spectrum ofactivity. J Med Chem 39:5005–5011
14. Hatse S, De Clercq E, Balzarini J (1999) Impact of 9-(2-phosphonylmethoxiethyl)adenine on (deoxy)ribonucleotide me-tabolism and nucleic acid synthesis in tumour cells. FEBS Lett445:92–97
15. Ruiz van Haperen VWT, Peters GJ (1994) New target forpyrimidine antimetabolites for the treatment of solid tumour.Deoxycytidine kinase. Pharm World Sci 1:104–112
16. Wilt CL, Peters GJ (1994) New targets for pyrimidine antimeta-bolites in the treatment of solid tumours. Tymidilate synthetase.Pharm World Sci 16:84–103
17. Robins RK, Revankar GR (1988) In: De Clercq E, Walker RT(eds) Antiviral drug development. Plenum Press, New York, p 11
18. Tada M (1975) Antineoplastic agents. Synthesis of some 1-substituted 5-fluorouracil derivatives. Chem Lett 2:129–130
19. Tada M (1976) 5-Fluorouracil derivatives. (Mitsui Selyaku KogyoCo., Ltd.), US 3,971,784, July 27, 1976
20. Yamashita JL, Yamawaki I, Ueda S, Yasumoto M, Unemi N,Hashimoto S (1982) Studies on antitumour agents. Synthesis andantitumour activities of 5-fluorouracil. Chem Pharm Bull30:4258–4267
21. Malet-Martino M, Martino R (2002) Clinical studies of three oralprodrugs of 5-fluorouracil (capecitabine, UFT, S-1): a review.Oncologist 7:288–323
22. Glavaš-Obrovac L, Karner I, Žinić B, Pavelić K (2001)Antineoplastic activity of novel N-1-sulfonylpyrimidine deriva-tives. Anticancer Res 21:1979–1986
23. Glavaš-Obrovac L, Karner I, Štefanić M, Kašnar-Šamprec J, ŽinićB (2005) Metabolic effects of novel N-1-sulfonylpyrimidinederivatives on human colon carcinoma cells. Farmaco 60:479–483
24. Glavaš-Obrovac L, Karner I, Pavlak M, Radačić M, Kašnar-Šamprec J, Žinić B (2005) Synthesis and antitumor activity of5-bromo-1-mesyluracil. Nucleosides Nucleotides and NucleicAcid 24:557–569
25. Supek F, Kralj M, Marjanović M, Šuman L, Šmuc T, Krizmanić I,Žinić B (2008) Atypical cytostatic mechanism of N-1-Sulfonylcytosine derivatives determined by in vitro screeningand computational analysis. Invest New Drugs 26:97–110
26. Pavlak M, Stojković R, Radačić-Aumiler M, Kašnar-Šamprec J,Jerčić J, Vlahović K, Žinić B, Radačić M (2005) Antitumouractivity of novel N-sulfonylpyrimidine derivatives on the growthof anaplastic mammary carcinoma in vivo. J Cancer Res ClinOncol 131(12):829–836
27. Kašnar-Šamprec J, Glavaš-Obrovac Lj, Pavlak M, Mihaljević I,Štambuk N, Konjevoda P, Žinić B (2005) Synthesis, spectroscopiccharacterization and biological activity of N–1–sulfonylcytosinederivatives. Croat Chem Acta 78(2):261–267
28. Mickisch G, Fajta S, Bier H, Tschada R, Alken P (1999) Cross-resistance patterns related to glutathione metabolism in primaryhuman renal cell carcinoma. Urol Res 19(2):99–103
29. Freshny RI (2000) Culture of animal cells: a manual of basictechnique, 4th ed. New York, Wiley-Liss Inc, pp 89–104
30. Pfaffl MW, Horgan GW, Dempfle L (2002) Relative ExpressionSoftware Tool (REST) for group-wise comparison and statisticalanalysis of relative expression results in real-time PCR. NucleicAcids Res 30(9):e36
31. Han R, Yang YM, Dietrich J, Luebke A, Mayer-Pröschel M,Noble M (2008) Systemic 5-fluorouracil treatment causes asyndrome of delayed myelin destruction in the central nervoussystem. J Biol Chem 7(4):12
32. Mader RM, Müller M, Steger GG (1998) Resistance to 5-Fluorouracil. Gen Pharmacol Vasc Syst 31(5):661–666
33. Lee CH, Jeon YT, Kim SH, Song YS (2007) NF-κB as a potentialmolecular target for cancer therapy. Biofactors 29(1):19–35
34. Lee TL, Yang XP, Yan B et al (2007) A novel NF-kB genesignature is differentially expressed in head and neck squamouscell carcinomas in association with TP53 status. Clin Cancer Res13:5680–5691
35. Li TJ, Jia LP, Gao XL, Huang AL (2006) Gene therapy thatinhibits NF-κB results in apoptosis of human hepatocarcinomaby recombinant adenovirus. World J Gastroenterol 12(33):5287–5292
36. Wang CY, Cusack JC Jr, Liu R, Baldwin AS Jr (1999) Control ofinducible chemoresistance: enhanced anti-tumour therapy throughincreased apoptosis by inhibition of NF-kappaB. Nat Med 5:412–417
37. Ferris RL, Grandis JR (2007) NF-κB gene signatures and p53mutations in head and neck squamous cell carcinoma. Clin CancerRes 13(19):5663–5664
38. Li LW, Yu XY, Yang Y, Zhang CP, Guo LP, Lu SH (2009)Expression of esophageal cancer related gene 4 (ECRG4), a noveltumour suppressor gene, in esophageal cancer and its inhibitoryeffect on the tumour growth in vitro and in vivo. Int J Cancer 125(7):1505–1513
39. Orlowski RZ, Baldwin AS (2002) NF-κB as a therapeutic target incancer. Trends Mol Med 8(8):385–389
40. Arencibia JM, Del Rio M, Bonnin A, Lopes R, Lemoine NR,López-Barahona M (2005) Doxazosin induces apoptosis inLNCaP prostate cancer cell line through DNA binding andDNA-dependent protein kinase down-regulation. Int J Oncol 27(6):1617–1623
41. Caldon CE, Musgrove EA (2010) Distinct and redundantfunctions of cyclin E1 and cyclin E2 in development and cancer.Cell Div 5:2
42. Powley IR, Kondrashov A, Young LA, Dobbyn HC, Hill K et al(2009) Translational reprogramming following UVB irradiation ismediated by DNA-PKcs and allows selective recruitment to thepolysomes of mRNAs encoding DNA repair enzymes. Genes Dev23(10):1207–1220
43. Soengas MS, Alarcon RM, Yoshida H et al (1999) Apaf-1 andcaspase-9 in p53-dependent apoptosis and tumour inhibition.Science 284:156–159
44. Belmokhtar CA, Hillion J, Dudognon C et al (2003) Apopto-some independent pathway for apoptosis. Biochemical analysisof Apaf-1 defects and biological outcomes. J Biol Chem278:29571–29578
45. KuglerW, Buchholz F, Köhler F, Eibl H, LakomekM, Erdlenbruch B(2005) Downregulation of Apaf-1 and caspase-3 by RNA interfer-ence in human glioma cells: consequences for erucylphosphocholine-induced apoptosis. Apoptosis 10:1163–1174
990 Invest New Drugs (2012) 30:981–990