
In vivo imaging of tumor apoptosis using histone H1-targeting peptide
9
In vivo imaging of tumor apoptosis using histone H1-targeting peptide Kai Wang a,d,f , Sangeetha Purushotham a,f , Ji-Young Lee a,f , Moon-Hee Na a,f , Hyekyung Park a,f , Sun-Jeong Oh a,f , Rang-Woon Park a,f , Jae Yong Park b,f , Eungbae Lee c , Byung Chae Cho d,f , Mi-Na Song e , Moon-Chang Baek e , Wonjung Kwak e , Jeongsoo Yoo e , Allan S. Hoffman g,h , Yu-Kyoung Oh i , In-San Kim a,f, ⁎, Byung-Heon Lee a,f, ⁎ a Department of Biochemistry and Cell Biology, School of Medicine, Kyungpook National University, Daegu, Republic of Korea b Department of Internal Medicine, School of Medicine, Kyungpook National University, Daegu, Republic of Korea c Department of Thoracic & Cardiovascular Surgery, School of Medicine, Kyungpook National University, Daegu, Republic of Korea d Department of Plastic & Reconstructive Surgery, School of Medicine, Kyungpook National University, Daegu, Republic of Korea e Department of Molecular Medicine, School of Medicine, Kyungpook National University, Daegu, Republic of Korea f Cell & Matrix Research Institute, School of Medicine, Kyungpook National University, Daegu, Republic of Korea g WCU Program, School of Medicine, Kyungpook National University, Daegu, Republic of Korea h Department of Bioengineering, University of Washington, Seattle, WA, USA i College of Pharmacy, Seoul National University, Seoul, Republic of Korea abstract article info Article history: Received 8 June 2010 Accepted 12 September 2010 Available online 30 September 2010 Keywords: Apoptosis Histone H1 In vivo imaging Phage display Targeting peptide In vivo imaging of apoptosis could allow monitoring of tumor response to cancer treatments such as chemotherapy. Using phage display, we identified the CQRPPR peptide, named ApoPep-1(Apoptosis-targeting Peptide-1), that was able to home to apoptotic and necrotic cells in tumor tissue. ApoPep-1 also bound to apoptotic and necrotic cells in culture, while only little binding to live cells was observed. Its binding to apoptotic cells was not dependent on calcium ion and not competed by annexin V. The receptor for ApoPep-1 was identified to be histone H1 that was exposed on the surface of apoptotic cells. In necrotic cells, ApoPep-1 entered the cells and bound to histone H1 in the nucleus. The imaging signals produced during monitoring of tumor apoptosis in response to chemotherapy was enhanced by the homing of a fluorescent dye- or radioisotope-labeled ApoPep-1 to tumor treated with anti-cancer drugs, whereas its uptake of the liver and lung was minimal. These results suggest that ApoPep-1 holds great promise as a probe for in vivo imaging of apoptosis, while histone H1 is a unique molecular signature for this purpose. © 2010 Elsevier B.V. All rights reserved. 1. Introduction Apoptosis plays an important role in the pathogenesis of a variety of diseases, such as cancer, myocardial and cerebral ischemia, and neurodegenerative diseases [1]. Since the induction of apoptosis is an essential event in cancer treatments, such as chemotherapy and irradiation, imaging apoptosis would provide valuable information in regards to monitoring tumor response to different therapies. Annexin V has been most extensively used to image apoptosis in diverse clinical circumstances [2]. Annexin V is a protein with a molecular weight (MW) of 36 kDa and has a high-affinity to phosphatidyserine (PS) that was exposed on the surface of apoptotic cells. It has been reported, however, that the in vivo performance of annexin V still needs improvement in terms of signal to background ratio and specificity to apoptotic cells over non-apoptotic cells [3,4]. As an alternative to bulky proteins or antibodies, small peptides may have properties as an ideal imaging probe. They have rapid clearance from blood circulation, efficient tissue penetration due to their small sizes, low immunogenicity, and low production cost [5,6]. Moreover, peptides are relatively easy to be chemically modified for coupling to imaging agents or drugs. Many peptides have already been in clinical use and under investigation in clinical trials. For example, they include the Octreoscan for the diagnosis of neuroendocrine tumors, cyclic RGD peptide for head and neck cancers, and Exendin-4 for insulinoma [6]. Cells in a disease microenvironment put their own molecular signatures on their cell surface. Angiogenic tumor endothelial cells over-express novel proteins such as the αvβ3 integrin. Macrophages in a tumor microenvironment express genes such as mannose receptor C type I, which is distinct from conventional pro-inflammatory macro- phages [7]. Such molecular signatures in diverse disease microenviron- ments have been successfully exploited by screening phage-displayed peptide libraries [8–14]. Given that cells from primary tumor tissues would more closely mimic the cells in a tumor microenvironment than do cultured cell lines, we screened a phage-displayed peptide library in order to select Journal of Controlled Release 148 (2010) 283–291 ⁎ Corresponding authors. Department of Biochemistry and Cell Biology, School of Medicine, Kyungpook National University, Daegu 700-421, Republic of Korea. Tel.: + 82 53 420 4824; fax: +82 53 422 1466. E-mail addresses: [email protected] (I.-S. Kim), [email protected] (B.-H. Lee). 0168-3659/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.jconrel.2010.09.010 Contents lists available at ScienceDirect Journal of Controlled Release journal homepage: www.elsevier.com/locate/jconrel
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of In vivo imaging of tumor apoptosis using histone H1-targeting peptide

Journal of Controlled Release 148 (2010) 283–291
Contents lists available at ScienceDirect
Journal of Controlled Release
j ourna l homepage: www.e lsev ie r.com/ locate / jconre l
In vivo imaging of tumor apoptosis using histone H1-targeting peptide
Kai Wang a,d,f, Sangeetha Purushotham a,f, Ji-Young Lee a,f, Moon-Hee Na a,f, Hyekyung Park a,f,Sun-Jeong Oh a,f, Rang-Woon Park a,f, Jae Yong Park b,f, Eungbae Lee c, Byung Chae Cho d,f, Mi-Na Song e,Moon-Chang Baek e, Wonjung Kwak e, Jeongsoo Yoo e, Allan S. Hoffman g,h, Yu-Kyoung Oh i, In-San Kim a,f,⁎,Byung-Heon Lee a,f,⁎a Department of Biochemistry and Cell Biology, School of Medicine, Kyungpook National University, Daegu, Republic of Koreab Department of Internal Medicine, School of Medicine, Kyungpook National University, Daegu, Republic of Koreac Department of Thoracic & Cardiovascular Surgery, School of Medicine, Kyungpook National University, Daegu, Republic of Koread Department of Plastic & Reconstructive Surgery, School of Medicine, Kyungpook National University, Daegu, Republic of Koreae Department of Molecular Medicine, School of Medicine, Kyungpook National University, Daegu, Republic of Koreaf Cell & Matrix Research Institute, School of Medicine, Kyungpook National University, Daegu, Republic of Koreag WCU Program, School of Medicine, Kyungpook National University, Daegu, Republic of Koreah Department of Bioengineering, University of Washington, Seattle, WA, USAi College of Pharmacy, Seoul National University, Seoul, Republic of Korea
⁎ Corresponding authors. Department of BiochemistMedicine, Kyungpook National University, Daegu 700-4253 420 4824; fax: +82 53 422 1466.
E-mail addresses: [email protected] (I.-S. Kim), leebh@
0168-3659/$ – see front matter © 2010 Elsevier B.V. Aldoi:10.1016/j.jconrel.2010.09.010
a b s t r a c t
a r t i c l e i n f oArticle history:Received 8 June 2010Accepted 12 September 2010Available online 30 September 2010
Keywords:ApoptosisHistone H1In vivo imagingPhage displayTargeting peptide
In vivo imaging of apoptosis could allow monitoring of tumor response to cancer treatments such aschemotherapy. Using phage display, we identified the CQRPPR peptide, named ApoPep-1(Apoptosis-targetingPeptide-1), that was able to home to apoptotic and necrotic cells in tumor tissue. ApoPep-1 also bound toapoptotic and necrotic cells in culture, while only little binding to live cells was observed. Its binding toapoptotic cells was not dependent on calcium ion and not competed by annexin V. The receptor for ApoPep-1was identified to be histone H1 that was exposed on the surface of apoptotic cells. In necrotic cells, ApoPep-1entered the cells and bound to histone H1 in the nucleus. The imaging signals produced during monitoring oftumor apoptosis in response to chemotherapy was enhanced by the homing of a fluorescent dye- orradioisotope-labeled ApoPep-1 to tumor treated with anti-cancer drugs, whereas its uptake of the liver andlung was minimal. These results suggest that ApoPep-1 holds great promise as a probe for in vivo imaging ofapoptosis, while histone H1 is a unique molecular signature for this purpose.
ry and Cell Biology, School of1, Republic of Korea. Tel.: +82
knu.ac.kr (B.-H. Lee).
l rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
Apoptosis plays an important role in the pathogenesis of a variety ofdiseases, such as cancer, myocardial and cerebral ischemia, andneurodegenerative diseases [1]. Since the induction of apoptosis is anessential event in cancer treatments, such as chemotherapy andirradiation, imaging apoptosis would provide valuable information inregards tomonitoring tumor response to different therapies. Annexin Vhas been most extensively used to image apoptosis in diverse clinicalcircumstances [2]. AnnexinV is a proteinwith amolecularweight (MW)of 36 kDa and has a high-affinity to phosphatidyserine (PS) that wasexposed on the surface of apoptotic cells. It has been reported, however,that the in vivo performance of annexin V still needs improvement intermsof signal to background ratio and specificity to apoptotic cells overnon-apoptotic cells [3,4].
As an alternative to bulky proteins or antibodies, small peptidesmayhave properties as an ideal imaging probe. They have rapid clearancefrom blood circulation, efficient tissue penetration due to their smallsizes, low immunogenicity, and low production cost [5,6]. Moreover,peptides are relatively easy to be chemically modified for coupling toimaging agents or drugs. Many peptides have already been in clinicaluse and under investigation in clinical trials. For example, they includethe Octreoscan for the diagnosis of neuroendocrine tumors, cyclic RGDpeptide for head and neck cancers, and Exendin-4 for insulinoma [6].
Cells in a disease microenvironment put their own molecularsignatures on their cell surface. Angiogenic tumor endothelial cellsover-express novel proteins such as the αvβ3 integrin. Macrophages ina tumor microenvironment express genes such as mannose receptor Ctype I, which is distinct from conventional pro-inflammatory macro-phages [7]. Such molecular signatures in diverse disease microenviron-ments have been successfully exploited by screening phage-displayedpeptide libraries [8–14].
Given that cells from primary tumor tissues would more closelymimic the cells in a tumor microenvironment than do cultured celllines, we screened a phage-displayed peptide library in order to select

284 K. Wang et al. / Journal of Controlled Release 148 (2010) 283–291
peptides that specifically bind to macrophages isolated from primarytumor tissues. Interestingly, we identified a peptide that was able totarget apoptotic and necrotic cells in tumor tissue by binding tohistone H1 on the surface of apoptotic cells and in the nucleus ofnecrotic cells. Here, we suggest that the histone H1-targeting peptideis a promising probe for imaging apoptosis in vivo.
2. Materials and methods
2.1. Screening of a phage peptide library
A phage library based on T7 415-1b phage vector displaying CX7C(C, cysteine; X, any amino acid residue) random peptides wasconstructed according to the manufacturer's instruction (Novagen,Madison, WI, USA). The library had a diversity of 5×108 plaqueforming unit (pfu). Primary human lung squamous carcinoma tissuesand adjacent normal tissues were obtained by surgical resection oftumor for the treatment of patients with cancer. Cell suspension wasprepared by gently homogenizing tissue using Medimachine (DAKO,Carpinteria, CA, USA). Phages (5.5×1010 pfu) were incubated with10 μl (packed cell volume) of cells from tumor tissue at 4 °C for 1 h.Macrophages were isolated from the cell suspension by using 25 μl(1×107 beads) of magnetic beads conjugated with anti-human CD14antibody (Dynal, Oslo, Norway) at 4 °C for 30 min. Macrophage-boundphages were eluted by lysing the cells with 100 μl of 1% NP-40 andadding a 900 μl culture of BL21 bacteria to the lysates. For subtraction,the elutedphage solutionwas incubatedwith cells fromnormal tissueat4 °C for 30 min and then the supernatant containing unbound phageswas recovered. The recovered phages were propagated in host bacteriaand used for the next round of selection. The output titer of phages wasdetermined by counting the number of plaques. After three rounds ofscreening, phage clones were randomly selected from the second andthird round and their DNA inserts were sequenced. Amino acidsequences were analyzed by the Clustal W program.
Peptides were synthesized by Peptron Inc. (Daegeon, Korea) andAnygen Inc. (Gwangju, Korea). Fluorescein isothiocyanate (FITC), biotin,or Cy7.5 dye (GE Healthcare, Piscataway, NJ, USA) was conjugated atN-terminal of peptides.
2.2. Immunofluorescence and flow cytometry analysis of peptide binding
A549 and H460 lung tumor cells and MDA-MB-231 breast tumorcells were maintained in RPMI-1640 medium. L132 normal humanlung epithelial cells, RAW human macrophage cells, and HeLa humancervical cancer cells were maintained in high-glucose Dulbecco'smodified Eagle's Medium. All culture media were supplemented with10% fetal bovine serum and penicillin/streptomycin.
Apoptosis was induced by incubating cells with 50 μM etoposidefor the indicated time periods. Necrosis was induced by incubatingcells with glucose-depleted medium for 10–15 h. Apoptotic stageswere determined by staining the cells with annexin V and propidiumiodide (PI). Live cells exclude cationic dyes such as PI, while dead cells(necrotic or late apoptotic cells) do not.
For immunofluorescence analysis, cells were incubated with 1%bovine serum albumin at 37 °C for 30 min for blocking and then with10 μM FITC-conjugated peptide at 4 °C for 1 h. Cells were then co-stained with Alexa-594-annexin V (Invitrogen, Carlsbad, CA, USA) for15 min at room temperature in binding buffer (10 mMHEPES, pH 7.4,140 mM NaCl, and 2.5 mM CaCl2). For competition with annexin V,cells were pre-incubated with unlabeled annexin V at excess molarconcentrations for 30 min at 37 °C and then incubated with Alexa-594-annexin V or FITC-ApoPep-1. For the staining of neuropilin-1(NRP-1), cells were incubated with a rabbit monoclonal anti-humanNRP-1 antibody (1:100 dilution; Abcam, Cambridge, MA, USA). Afterfixation, cells were incubated with 4′,6′-diamidino-2-phenylindole(DAPI, Sigma-Aldrich, St Louis, MO, USA) for nuclear staining and a
mounting solution before observation with a fluorescent microscope(Zeiss, Oberkochen, Germany).
For flow cytometry analysis, cells (1×106 cells) in suspensionwere incubated with FITC-peptide or Alexa-488-annexin V and thensubjected to flow cytometry. To see if cells were at late stage ofapoptosis or necrosis, cells were co-stainedwith 1.5 μMPI for 5 min atroom temperature.
2.3. Affinity chromatography and mass spectrometry
ApoPep-1 was conjugated with a biotinylated, photoreactive cross-linker, sulfo-N-hydroxysuccinimidyl-2-(6-[biotinamido]-2-(p-azidobenzamido)-hexanoamido) ethyl-1,3′-dithioproprionate (sulfo-SBED,Pierce, Rockford, IL), for 60 min at room temperature in the dark. Theconjugated ApoPep-1 was purified by removing unincorporated sulfo-SBED using 1 kDa Tube-O-dialyzer (Biosciences, St Louis, MO, USA) for6 h. The peptide solution was incubated with apoptotic cells(5×107 cells) at 37 °C for 60 min in serum-free medium and thenexposed to ultraviolet light at 0.04 J for 15 min using Stratalinker(Stratagene, La Jolla, CA, USA) to photoactivate the sulfo-SBED. Cellswere lysed and ApoPep-1 (bait) and binding protein (prey) complexwas immobilized by incubating the cell lysates with monomeric avidinbeads (Pierce) at 4 °C for 1 h. ApoPep-1 was separated by reducing thecomplex with 50 mMdithiothreitol (DTT). The prey protein was elutedwith 2 mM D-biotin followed by treatment with 100 mM glycine, pH3.0. Elutedproteinswere resolvedbySDS-PAGEand stainedwitha silverstaining reagent compatible with mass spectrometry. Protein bandswere excised from the silver-stained gel and subjected to massspectrometry.
For in-gel trypsin digestion, an excised gel was cut into smallpieces, destained in 30 mM potassium ferricyanide/100 mM sodiumthiosulphate, andwashedwith 100% acetonitrile/100 mMammoniumbicarbonate. The gel pieces were dried by vacuum centrifugation afterdehydrated with acetonitrile, reduced with 10 mM DTT at 56 °C for45 min and alkylated with 55 mM iodoacetamide for 30 min at roomtemperature. The gel pieces were then dried by vacuum centrifuga-tion, rehydrated with trypsin solution in 50 mM ammonium bicar-bonate, and incubated overnight at 37 °C. Peptides were extractedwith three steps: 5% formic acid/60% acetonitrile; 5% formic acid/50 mM ammonium bicarbonate; and finally 5% formic acid/100%acetonitrile. The extracts were pooled, dried by vacuum centrifugationand resolved in 10 μl of 0.1% formic acid.
The peptide extractswere separated by a nano-Acquity UPLC systemusing a Symmetry C18 5 μm, 5-mm×300-μm pre-column and anAtlantis C18 3 μm, 15-cm×75-μm analytical reversed phase column(Waters, Milford, MA, USA). Flow rate was 300 nl/min. Mobile phase Aand B are 0.1% formic acid in water and 0.1% formic acid in acetonitrile,respectively. The peptides were separated with a gradient of 3–40%mobile phase B over 60 min at a flow rate of 300 nl/min followed by a1 min rinsewith 90% ofmobile phase B. The columnwas re-equilibratedat initial conditions for 20 min. The column temperature was main-tained at 40 °C. Raw data from Q-TOF Premier mass spectrometer(Waters) were processed and identified by Protein Lynx Global Server2.3 software (Waters) and Mascot v2.0 (Matrix science, Boston, MA)using a IPI Human v3.47 (http://www.ebi.ac.uk/IPI/). For searchparameters, fixed modification was carbamidomethyl C and variablemodification was oxidation M.
2.4. Analysis of peptide binding to histone H1
Membrane fraction-enriched cell lysateswere prepared using a lysisbuffer (0.5% Triton X-100, 100 mM NaCl, 1 mM EDTA, 20 mM Tris).Biotin-eluates from affinity chromatography or cell lysates weresubjected to 15% SDS-PAGE, transferred to a nitrocellulose membrane,and immunoblotted with a mouse monoclonal anti-human histone H1(AE-4) antibody (1 μg/ml; Santa Cruz Biotechnology, Santa Cruz, CA,
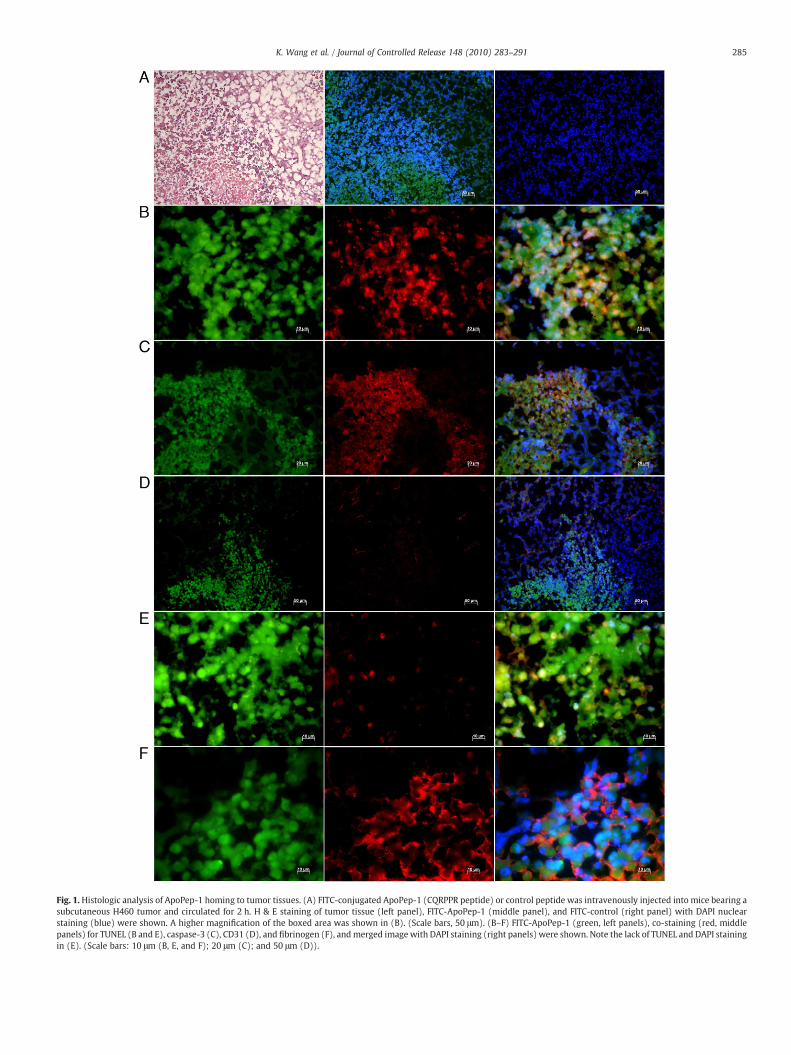
Fig. 1. Histologic analysis of ApoPep-1 homing to tumor tissues. (A) FITC-conjugated ApoPep-1 (CQRPPR peptide) or control peptide was intravenously injected into mice bearing asubcutaneous H460 tumor and circulated for 2 h. H & E staining of tumor tissue (left panel), FITC-ApoPep-1 (middle panel), and FITC-control (right panel) with DAPI nuclearstaining (blue) were shown. A higher magnification of the boxed area was shown in (B). (Scale bars, 50 μm). (B–F) FITC-ApoPep-1 (green, left panels), co-staining (red, middlepanels) for TUNEL (B and E), caspase-3 (C), CD31 (D), and fibrinogen (F), and merged image with DAPI staining (right panels) were shown. Note the lack of TUNEL and DAPI stainingin (E). (Scale bars: 10 μm (B, E, and F); 20 μm (C); and 50 μm (D)).
285K. Wang et al. / Journal of Controlled Release 148 (2010) 283–291

286 K. Wang et al. / Journal of Controlled Release 148 (2010) 283–291
USA). For peptide ligand blotting, a nitrocellulose membrane wasincubated with a biotinylated ApoPep-1 (6 μg/ml) overnight and thenwith peroxidase-conjugated streptavidin (DAKO) and developed withDAB substrate. For immunoprecipitation, recombinant human histoneH1 (rH1, 3 μg/ml, New England Biolabs, Ipswich, MA, USA) wasincubated with a biotinylated ApoPep-1 (10 μg/ml) and capturedusing monomeric avidin beads, and subjected to immunoblotting withthe anti-histone H1 antibody. For competition with the antibody, cellswere pre-incubated with the anti-histone H1 antibody (1 or 10 μg/ml)and thenwith 10 μMFITC-ApoPep-1 followed by the analysis of peptidebinding using flow cytometry.
For co-localization assays, apoptotic cellswere stainedwith the anti-histone H1 antibody (0.5 μg/ml) and then with Alexa-594-labeledsecondary antibody and 10 μM FITC-ApoPep-1. Necrotic cells wereincubated with FITC-ApoPep-1, fixed with methanol, permealized with0.1% Tween-20, and then stained with the anti-histone H1 antibody.After staining, cellswere examined under a confocalmicroscope (Zeiss).
2.5. Immunohistochemistry of peptide homing to tumor
FITC-conjugated ApoPep-1 or control peptide (100 nmol) wasintravenously (i.v.) injected into the tail vein of mice bearing asubcutaneous tumor. After 2 h of circulation, mice were perfused withphosphate-buffered saline (PBS) through the heart and then fixed with4%paraformaldehyde. Tumors andother organswere removed,fixedandprepared for frozen sections.Apoptotic areasof tissuewere stainedwith amodified terminal deoxynucleotidyl-transferase mediated UTP endlabeling (TUNEL, Chemicon, Billerica, MA) or anti-caspase 3 antibody(Cell Signaling, Danvers, MA) for 2 h. Fibrinogen was stained with theanti-mouse fibrinogen antibody (Abcam) for 1 h. Alexa-594-labeledsecondary antibodies (Invitrogen) were incubated for 30 min. Tissuesections were incubated with DAPI for nuclei staining and with amounting solution before observation under a fluorescent microscope.
2.6. In vivo fluorescence imaging of tumor apoptosis
All animal experiments were conducted according to the guidelinesof Kyungpook National University ethical committee. BALB/c nu/nufemale mice bearing a subcutaneous tumor (approximately 10 mm indiameter) were treated with doxorubicin or camptothecin (10 mg/kgbody weight; i.v. injection, three times at a two-day interval fordoxorubicin andone time for camptothecin). Three days after treatment,either FITC-ApoPep-1 (100 nmol or 126 μg) or cy7.5-ApoPep-1(100 nmol or 168 μg) was injected via the tail vein and circulated forthe indicated time periods. In vivo optical imaging was conducted byscanning themice under anesthesia usingOptix exPlore (ART,Montreal,Canada). The excitation/emission wavelengths for FITC and Cy7.5 were490 nm/520 nm and 778 nm/805 nm, respectively.
2.7. Radiolabeling of peptide, positron emission tomography (PET) imagingand biodistribution studies
In order to radiolabel ApoPep-1 with radioactive iodine (124I or 131I),one extra tyrosine residuewas introduced atN-terminal andbothN- andC-terminals were capped with acetyl and amine groups, respectively, toafford Ac-YCQRPPR-NH2. Briefly, Iodobead (Pierce) was incubated with[124I or 131I]NaI (2.3 mCi) in 100 μl of PBS. After 5 min shaking at roomtemperature, ApoPep-1 (Ac-YCQRPPR-NH2) solution (20 μg in 20 μldistilled water) was added to [124I or 131I]NaI solution. The solution wasgently shaken for 15 min. Then the bead was removed from reactiontube for terminating the reaction. [124I or 131I]ApoPep-1 was purified byFPLC with Shiseido capcell pak C-18 column at gradient condition ofwater andacetonitrile containing0.1% trifluoroacetic acid. Radiochemicalpurity of [124I or 131I]ApoPep-1wasmeasured to beN90%using radio-TLCscanner. The purified radioiodinated peptide (containing about 10% freeiodide)wasused formicroPET imaging andbiodistribution studies. There
was little degradation of the radioiodinatedpeptide in the serumstabilitytests.
For PET imaging studies, [18F]fludeoxyglucose (FDG) (18F t1/2=110 min) was i.v. injected into tumor-bearingmice thatwere untreatedor treated with doxorubicin as described above (304 and 300 μCi/0.15 mL, respectively) and imageswere taken at1 h after injectionusingamicro PET (ConcordeMicrosystems, Knoxville, TN, USA). Subsequent-ly, [124I]ApoPep-1 (124I t1/2=4.2 days) was injected into the miceuntreated or treated with doxorubicin (93 and 91 μCi/0.15 mL,respectively) and images were taken at 5 h after injection.
Biodistribution analysis was performed using 7-week old maleSprague–Dawley rats (n=4per timepoints, bodyweights: 235.9±7 g).[131I]ApoPep-1 (7.0 μCi) was injected into the tail vein of rats. Blood andorganswere isolated at 1, 4, and24 hafter injection, and the radioactivityof each organ was measured by a gamma counter (Perkin Elmer,Waltham,MA) and calculated as percent injected dose per gram(%ID/g).
3. Results
3.1. Selection of peptides that bind to macrophages from tumor tissue
A phage library containing random peptides was screened forpeptides that selectively bind to macrophages isolated from primaryhuman lung cancer tissue. The titer of phageswas significantly enrichedby approximately 200-fold after three rounds of selection (Fig. S1A).Based on the analysis of selected peptide sequences, a consensus motifcontaining R/K, P/A, P/A, and R was identified (Table S1). Evaluation ofthe individual phage clones showed that the phage clone displaying theCQRPPR peptide preferentially bound to macrophages from tumortissues of three patients with lung cancer over macrophages from thesurrounding normal tissues (Fig. S1B). Binding of insertless phage, usedas a negative control, to macrophages from either tumor or normaltissue was minimal.
3.2. Histologic analysis of ApoPep-1 homing to tumor
We examined whether the CQRPPR peptide homes to tumor andbinds to macrophages in tumor tissue. When intravenously adminis-tered into tumor-bearing mice, the CQRPPR peptide homed to tumortissue, while the NSSSVDK peptide, a control peptide from phage capsidprotein, did not (Fig. 1A). Interestingly, we found that the CQRPPRpeptide mainly localized at apoptotic areas in tumor tissue, where cellswere positively stained with TUNEL (Fig. 1B) and an anti-caspase 3antibody (Fig. 1C), while little peptide was found in the blood vessels oftumor tissue (Fig. 1D). It also localized at necrotic areas, where TUNELand DAPI staining were deficient (Fig. 1E) and fibrinogen was found inthe extravascular space (Fig. 1F). These results suggest that the CQRPPRpeptide, named ApoPep-1 (Apoptosis-targeting Peptide-1), homes totumor by binding to apoptotic and necrotic cells at tumor tissue in vivo.
We hypothesized that the selection of ApoPep-1 that binds toapoptotic cells rather than macrophages in tumor tissue was due to alarge portion of cells isolated from primary tumor tissues were alreadyundergoing apoptosis, leading to the enrichment of peptides specific toapoptotic cells. Indeed, flow cytometric analysis of cells isolated from aprimary tumor tissue showed that approximately 45% of macrophagesfrom tumor tissue were apoptotic, while less than 1% of macrophagesfrom the surrounding normal tissue were apoptotic (Fig. S2).
3.3. In vitro binding of ApoPep-1 to apoptotic and necrotic cells
The binding of ApoPep-1 to apoptotic cells was examined usingcells treated with etoposide. ApoPep-1 strongly bound to etoposide-treated, apoptotic cells, while little binding was observed inuntreated, live cells (Fig. 2A and Fig. S3). Annexin V/PI staining ofetoposide-treated cells showed that 64% and 9% of cells were in theearly (annexin V (+)/PI (−)) and late (annexin V (+)/PI (+)) stage

Fig. 2. In vitro binding of ApoPep-1 to apoptotic cells. (A) A549 cells were treated with (E(+)) or without (E(−)) 50 μM etoposide for 15 h to induce apoptosis. Cells were incubatedwith 10 μM FITC-ApoPep-1 (green) and co-stained with Alexa594-annexin V (red) and DAPI for nuclear staining (blue), and images were merged. (Scale bars, 20 μm.) (B) Cells weretreated with or without etoposide and then stained with either Alexa488-annexin V/PI or FITC-ApoPep-1/PI for flow cytometry. Numbers represent the percent of cells in thefraction. (C) Cells were incubated with FITC-ApoPep-1 and DAPI in the absence or presence of 2.5 mM CaCl2. (D) Cells were pre-incubated with the indicated concentrations ofannexin V and then incubated with Alexa594-annexin V or FITC-ApoPep-1. (Scale bars, 20 μm.)
287K. Wang et al. / Journal of Controlled Release 148 (2010) 283–291
of apoptosis, respectively (Fig. 2B). Similarly, 90% and 7% of cells werestained with ApoPep-1 (+)/PI (−) and ApoPep-1 (+)/PI (+),respectively (Fig. 2B), indicating that ApoPep-1 can recognize cellsin both early and late stage of apoptosis.
Unlike to annexin V, ApoPep-1 was able to bind to apoptotic cellsin the absence of calcium ion (Fig. 2C). Moreover, pre-incubation ofapoptotic cells with excess annexin V did not compete the binding ofApoPep-1 to apoptotic cells, while it inhibited subsequent binding ofannexin V to the cells in a dose dependant manner (Fig. 2D). Theseresults suggest that ApoPep-1 recognizes a novel epitope or receptoron apoptotic cells, which seems to be different from PS.
Mutational analysis of ApoPep-1 showed that the binding of theR3A (CQAPPR) mutant to apoptotic cells reduced to 10% of the wild-type binding (Fig. 3), suggesting that the R3 residue is the maindeterminant for ApoPep-1 binding. In addition, the binding of the C1A,Q2A, P4A, P5A, and R6A mutants as well as a scrambled peptide toapoptotic cells reduced to 25–40% of the wild-type binding. Littlechanges in the binding activity were observed with the P4G and P5Gmutants (Fig. 3).
To examinewhether ApoPep-1 binds to necrotic cells, necrosis wasinduced by glucose deprivation. Annexin V (−)/PI (+) staining,which is a known necrosis marker, was observed in approximately52% of cells (Fig. 4A). Moreover, the high-mobility group box 1(HMGB1) protein, which is released out of the nucleus of necrotic cellsbut not of apoptotic cells as a specific marker of necrosis [15], wasdetected in the culture medium of the cells (Fig. 4B). Necrotic cellswere stained with ApoPep-1 and PI but not with the control peptide(Fig. 4C). In confocal microscopic images, ApoPep-1 was observed inthe nucleus of necrotic cells and co-localized with PI (Fig. 4C).
3.4. Identification of histone H1 as the receptor for ApoPep-1
In order to identify the receptor for ApoPep-1, etoposide-treated,apoptotic cells were incubated with ApoPep-1 coupled to a biotiny-lated, photoactivatable cross-linking reagent and then exposed to theultraviolet light before cell lysis. Affinity chromatography of the celllysates using a monomeric avidin column yielded a protein band witha MW of 32–33 kDa (Fig. 5A). Using mass spectrometric analysis, the
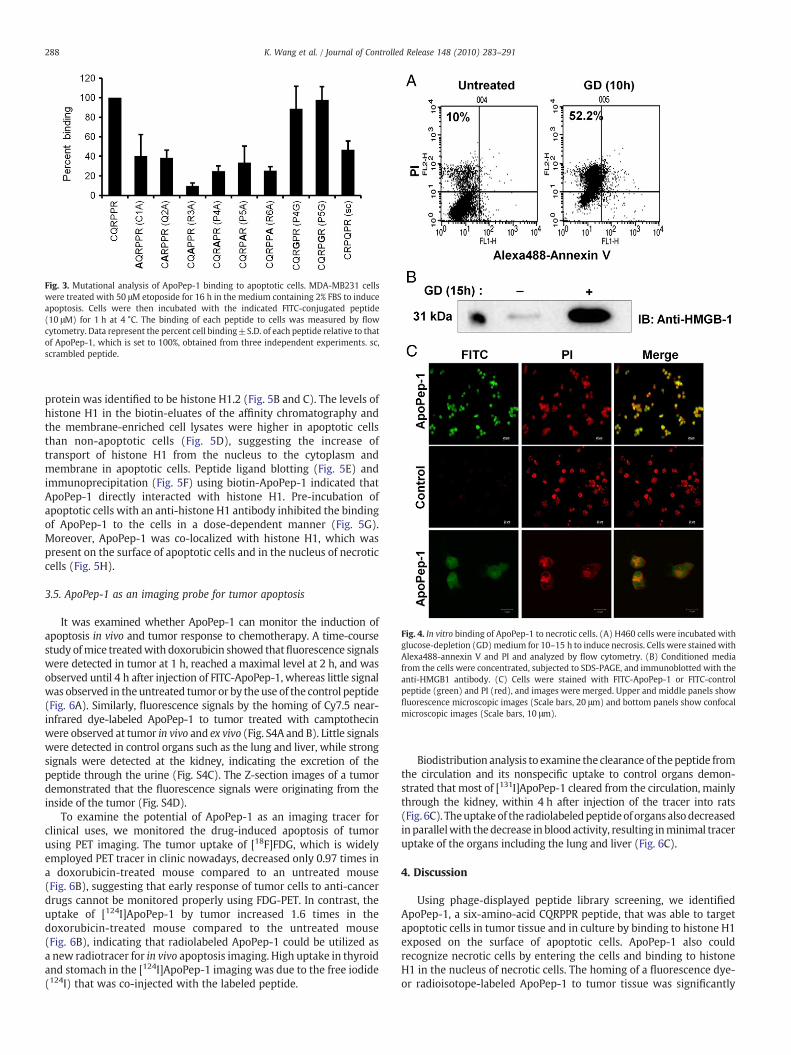
Fig. 3. Mutational analysis of ApoPep-1 binding to apoptotic cells. MDA-MB231 cellswere treated with 50 μM etoposide for 16 h in the medium containing 2% FBS to induceapoptosis. Cells were then incubated with the indicated FITC-conjugated peptide(10 μM) for 1 h at 4 °C. The binding of each peptide to cells was measured by flowcytometry. Data represent the percent cell binding±S.D. of each peptide relative to thatof ApoPep-1, which is set to 100%, obtained from three independent experiments. sc,scrambled peptide.
Fig. 4. In vitro binding of ApoPep-1 to necrotic cells. (A) H460 cells were incubated withglucose-depletion (GD)medium for 10–15 h to induce necrosis. Cells were stainedwithAlexa488-annexin V and PI and analyzed by flow cytometry. (B) Conditioned mediafrom the cells were concentrated, subjected to SDS-PAGE, and immunoblotted with theanti-HMGB1 antibody. (C) Cells were stained with FITC-ApoPep-1 or FITC-controlpeptide (green) and PI (red), and images were merged. Upper and middle panels showfluorescence microscopic images (Scale bars, 20 μm) and bottom panels show confocalmicroscopic images (Scale bars, 10 μm).
288 K. Wang et al. / Journal of Controlled Release 148 (2010) 283–291
protein was identified to be histone H1.2 (Fig. 5B and C). The levels ofhistone H1 in the biotin-eluates of the affinity chromatography andthe membrane-enriched cell lysates were higher in apoptotic cellsthan non-apoptotic cells (Fig. 5D), suggesting the increase oftransport of histone H1 from the nucleus to the cytoplasm andmembrane in apoptotic cells. Peptide ligand blotting (Fig. 5E) andimmunoprecipitation (Fig. 5F) using biotin-ApoPep-1 indicated thatApoPep-1 directly interacted with histone H1. Pre-incubation ofapoptotic cells with an anti-histone H1 antibody inhibited the bindingof ApoPep-1 to the cells in a dose-dependent manner (Fig. 5G).Moreover, ApoPep-1 was co-localized with histone H1, which waspresent on the surface of apoptotic cells and in the nucleus of necroticcells (Fig. 5H).
3.5. ApoPep-1 as an imaging probe for tumor apoptosis
It was examined whether ApoPep-1 can monitor the induction ofapoptosis in vivo and tumor response to chemotherapy. A time-coursestudy ofmice treatedwith doxorubicin showed thatfluorescence signalswere detected in tumor at 1 h, reached a maximal level at 2 h, and wasobserved until 4 h after injection of FITC-ApoPep-1, whereas little signalwas observed in theuntreated tumor or by theuse of the control peptide(Fig. 6A). Similarly, fluorescence signals by the homing of Cy7.5 near-infrared dye-labeled ApoPep-1 to tumor treated with camptothecinwere observed at tumor in vivo and ex vivo (Fig. S4A and B). Little signalswere detected in control organs such as the lung and liver, while strongsignals were detected at the kidney, indicating the excretion of thepeptide through the urine (Fig. S4C). The Z-section images of a tumordemonstrated that the fluorescence signals were originating from theinside of the tumor (Fig. S4D).
To examine the potential of ApoPep-1 as an imaging tracer forclinical uses, we monitored the drug-induced apoptosis of tumorusing PET imaging. The tumor uptake of [18F]FDG, which is widelyemployed PET tracer in clinic nowadays, decreased only 0.97 times ina doxorubicin-treated mouse compared to an untreated mouse(Fig. 6B), suggesting that early response of tumor cells to anti-cancerdrugs cannot be monitored properly using FDG-PET. In contrast, theuptake of [124I]ApoPep-1 by tumor increased 1.6 times in thedoxorubicin-treated mouse compared to the untreated mouse(Fig. 6B), indicating that radiolabeled ApoPep-1 could be utilized asa new radiotracer for in vivo apoptosis imaging. High uptake in thyroidand stomach in the [124I]ApoPep-1 imaging was due to the free iodide(124I) that was co-injected with the labeled peptide.
Biodistribution analysis to examine the clearance of thepeptide fromthe circulation and its nonspecific uptake to control organs demon-strated that most of [131I]ApoPep-1 cleared from the circulation, mainlythrough the kidney, within 4 h after injection of the tracer into rats(Fig. 6C). Theuptakeof the radiolabeledpeptideof organs alsodecreasedinparallelwith thedecrease in blood activity, resulting inminimal traceruptake of the organs including the lung and liver (Fig. 6C).
4. Discussion
Using phage-displayed peptide library screening, we identifiedApoPep-1, a six-amino-acid CQRPPR peptide, that was able to targetapoptotic cells in tumor tissue and in culture by binding to histone H1exposed on the surface of apoptotic cells. ApoPep-1 also couldrecognize necrotic cells by entering the cells and binding to histoneH1 in the nucleus of necrotic cells. The homing of a fluorescence dye-or radioisotope-labeled ApoPep-1 to tumor tissue was significantly
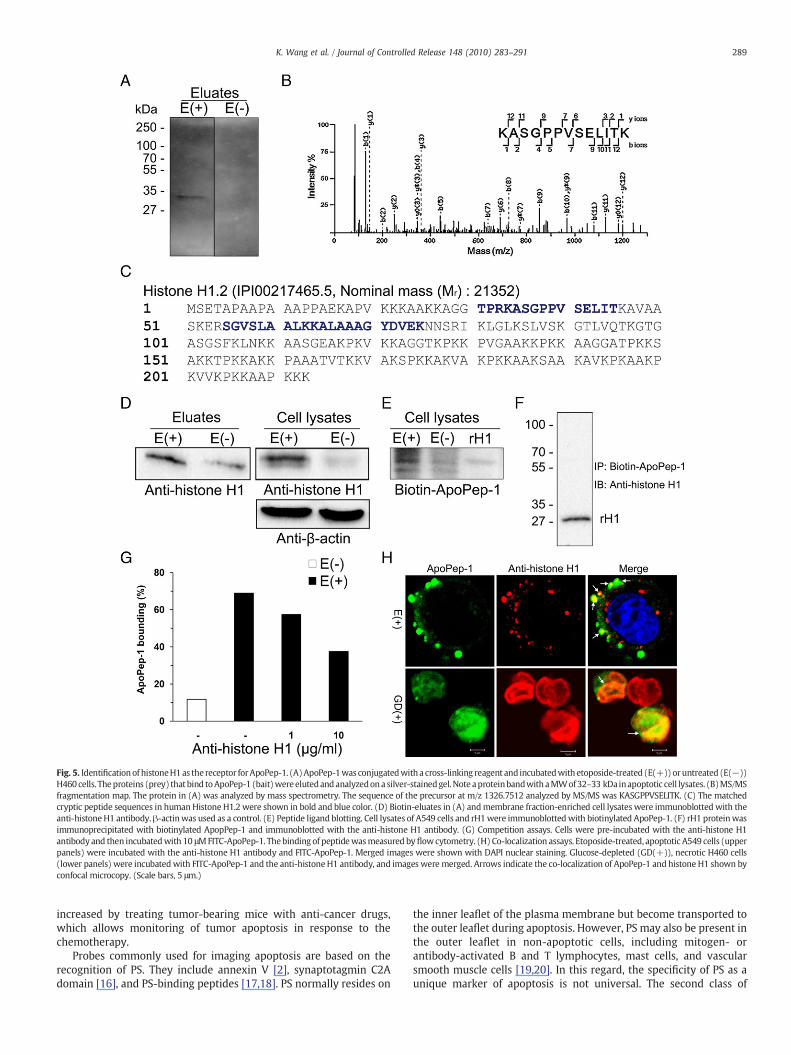
Fig. 5. Identificationof histoneH1as thereceptor forApoPep-1. (A)ApoPep-1was conjugatedwitha cross-linking reagent and incubatedwith etoposide-treated (E(+))oruntreated (E(−))H460 cells. Theproteins (prey) that bind toApoPep-1 (bait)were elutedandanalyzedona silver-stainedgel.Note aproteinbandwithaMWof32–33 kDa inapoptotic cell lysates. (B)MS/MSfragmentation map. The protein in (A) was analyzed by mass spectrometry. The sequence of the precursor at m/z 1326.7512 analyzed by MS/MS was KASGPPVSELITK. (C) The matchedcryptic peptide sequences in humanHistone H1.2 were shown in bold and blue color. (D) Biotin-eluates in (A) andmembrane fraction-enriched cell lysates were immunoblotted with theanti-histoneH1 antibody.β-actinwas used as a control. (E) Peptide ligand blotting. Cell lysates of A549 cells and rH1were immunoblottedwith biotinylated ApoPep-1. (F) rH1 proteinwasimmunoprecipitated with biotinylated ApopPep-1 and immunoblotted with the anti-histone H1 antibody. (G) Competition assays. Cells were pre-incubated with the anti-histone H1antibody and then incubatedwith 10 μMFITC-ApoPep-1. Thebinding of peptidewasmeasured byflowcytometry. (H) Co-localization assays. Etoposide-treated, apoptotic A549 cells (upperpanels) were incubated with the anti-histone H1 antibody and FITC-ApoPep-1. Merged images were shown with DAPI nuclear staining. Glucose-depleted (GD(+)), necrotic H460 cells(lower panels) were incubatedwith FITC-ApoPep-1 and the anti-histone H1 antibody, and images were merged. Arrows indicate the co-localization of ApoPep-1 and histone H1 shown byconfocal microcopy. (Scale bars, 5 μm.)
289K. Wang et al. / Journal of Controlled Release 148 (2010) 283–291
increased by treating tumor-bearing mice with anti-cancer drugs,which allows monitoring of tumor apoptosis in response to thechemotherapy.
Probes commonly used for imaging apoptosis are based on therecognition of PS. They include annexin V [2], synaptotagmin C2Adomain [16], and PS-binding peptides [17,18]. PS normally resides on
the inner leaflet of the plasma membrane but become transported tothe outer leaflet during apoptosis. However, PS may also be present inthe outer leaflet in non-apoptotic cells, including mitogen- orantibody-activated B and T lymphocytes, mast cells, and vascularsmooth muscle cells [19,20]. In this regard, the specificity of PS as aunique marker of apoptosis is not universal. The second class of
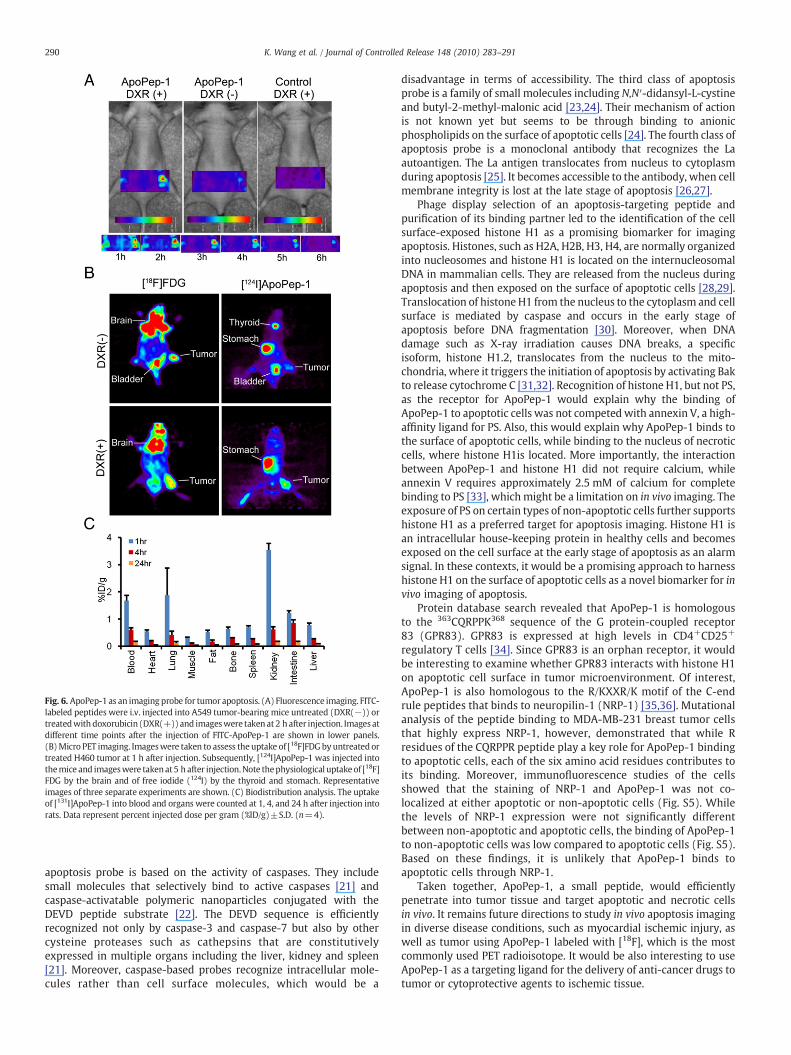
Fig. 6. ApoPep-1 as an imaging probe for tumor apoptosis. (A) Fluorescence imaging. FITC-labeled peptides were i.v. injected into A549 tumor-bearing mice untreated (DXR(−)) ortreatedwith doxorubicin (DXR(+))and imageswere taken at 2 h after injection. Images atdifferent time points after the injection of FITC-ApoPep-1 are shown in lower panels.(B)Micro PET imaging. Imageswere taken to assess theuptake of [18F]FDGbyuntreated ortreated H460 tumor at 1 h after injection. Subsequently, [124I]ApoPep-1 was injected intothemice and imageswere takenat5 h after injection.Note thephysiological uptakeof [18F]FDG by the brain and of free iodide (124I) by the thyroid and stomach. Representativeimages of three separate experiments are shown. (C) Biodistribution analysis. The uptakeof [131I]ApoPep-1 into blood and organs were counted at 1, 4, and 24 h after injection intorats. Data represent percent injected dose per gram (%ID/g)±S.D. (n=4).
290 K. Wang et al. / Journal of Controlled Release 148 (2010) 283–291
apoptosis probe is based on the activity of caspases. They includesmall molecules that selectively bind to active caspases [21] andcaspase-activatable polymeric nanoparticles conjugated with theDEVD peptide substrate [22]. The DEVD sequence is efficientlyrecognized not only by caspase-3 and caspase-7 but also by othercysteine proteases such as cathepsins that are constitutivelyexpressed in multiple organs including the liver, kidney and spleen[21]. Moreover, caspase-based probes recognize intracellular mole-cules rather than cell surface molecules, which would be a
disadvantage in terms of accessibility. The third class of apoptosisprobe is a family of small molecules including N,N′-didansyl-L-cystineand butyl-2-methyl-malonic acid [23,24]. Their mechanism of actionis not known yet but seems to be through binding to anionicphospholipids on the surface of apoptotic cells [24]. The fourth class ofapoptosis probe is a monoclonal antibody that recognizes the Laautoantigen. The La antigen translocates from nucleus to cytoplasmduring apoptosis [25]. It becomes accessible to the antibody, when cellmembrane integrity is lost at the late stage of apoptosis [26,27].
Phage display selection of an apoptosis-targeting peptide andpurification of its binding partner led to the identification of the cellsurface-exposed histone H1 as a promising biomarker for imagingapoptosis. Histones, such as H2A, H2B, H3, H4, are normally organizedinto nucleosomes and histone H1 is located on the internucleosomalDNA in mammalian cells. They are released from the nucleus duringapoptosis and then exposed on the surface of apoptotic cells [28,29].Translocation of histone H1 from the nucleus to the cytoplasm and cellsurface is mediated by caspase and occurs in the early stage ofapoptosis before DNA fragmentation [30]. Moreover, when DNAdamage such as X-ray irradiation causes DNA breaks, a specificisoform, histone H1.2, translocates from the nucleus to the mito-chondria, where it triggers the initiation of apoptosis by activating Bakto release cytochrome C [31,32]. Recognition of histone H1, but not PS,as the receptor for ApoPep-1 would explain why the binding ofApoPep-1 to apoptotic cells was not competed with annexin V, a high-affinity ligand for PS. Also, this would explain why ApoPep-1 binds tothe surface of apoptotic cells, while binding to the nucleus of necroticcells, where histone H1is located. More importantly, the interactionbetween ApoPep-1 and histone H1 did not require calcium, whileannexin V requires approximately 2.5 mM of calcium for completebinding to PS [33], whichmight be a limitation on in vivo imaging. Theexposure of PS on certain types of non-apoptotic cells further supportshistone H1 as a preferred target for apoptosis imaging. Histone H1 isan intracellular house-keeping protein in healthy cells and becomesexposed on the cell surface at the early stage of apoptosis as an alarmsignal. In these contexts, it would be a promising approach to harnesshistone H1 on the surface of apoptotic cells as a novel biomarker for invivo imaging of apoptosis.
Protein database search revealed that ApoPep-1 is homologousto the 363CQRPPK368 sequence of the G protein-coupled receptor83 (GPR83). GPR83 is expressed at high levels in CD4+CD25+
regulatory T cells [34]. Since GPR83 is an orphan receptor, it wouldbe interesting to examine whether GPR83 interacts with histone H1on apoptotic cell surface in tumor microenvironment. Of interest,ApoPep-1 is also homologous to the R/KXXR/K motif of the C-endrule peptides that binds to neuropilin-1 (NRP-1) [35,36]. Mutationalanalysis of the peptide binding to MDA-MB-231 breast tumor cellsthat highly express NRP-1, however, demonstrated that while Rresidues of the CQRPPR peptide play a key role for ApoPep-1 bindingto apoptotic cells, each of the six amino acid residues contributes toits binding. Moreover, immunofluorescence studies of the cellsshowed that the staining of NRP-1 and ApoPep-1 was not co-localized at either apoptotic or non-apoptotic cells (Fig. S5). Whilethe levels of NRP-1 expression were not significantly differentbetween non-apoptotic and apoptotic cells, the binding of ApoPep-1to non-apoptotic cells was low compared to apoptotic cells (Fig. S5).Based on these findings, it is unlikely that ApoPep-1 binds toapoptotic cells through NRP-1.
Taken together, ApoPep-1, a small peptide, would efficientlypenetrate into tumor tissue and target apoptotic and necrotic cellsin vivo. It remains future directions to study in vivo apoptosis imagingin diverse disease conditions, such as myocardial ischemic injury, aswell as tumor using ApoPep-1 labeled with [18F], which is the mostcommonly used PET radioisotope. It would be also interesting to useApoPep-1 as a targeting ligand for the delivery of anti-cancer drugs totumor or cytoprotective agents to ischemic tissue.

291K. Wang et al. / Journal of Controlled Release 148 (2010) 283–291
Acknowledgment
This work was supported by the grant from the RegionalTechnology Innovation Program of the Ministry of KnowledgeEconomy (RTI04-01-01), the grant from the National R&D Programfor Cancer Control, Ministry for Health, Welfare and Family Affairs(0720550-2), and WCU Program through the National ResearchFoundation of Korea funded by the Ministry of Education, Science andTechnology (R33-10054).
Appendix A. Supplementary data
Supplementary data to this article can be found online atdoi:10.1016/j.jconrel.2010.09.010.
References
[1] C.B. Thompson, Apoptosis in the pathogenesis and treatment of disease, Science267 (1995) 1456–1462.
[2] F.G. Blankenberg, In vivo detection of apoptosis, J. Nucl. Med. 49 (Suppl 2) (2008)81S–95S.
[3] T.Z. Belhocine, J.F. Tait, J.L. Vanderheyden, C. Li, F.G. Blankenberg, Nuclearmedicine in the era of genomics and proteomics: lessons from annexin V, J.Proteome Res. 3 (2004) 345–349.
[4] C. Mari, M. Karabiyikoglu, M.L. Goris, J.F. Tait, M.A. Yenari, F.G. Blankenberg,Detection of focal hypoxic-ischemic injury and neuronal stress in a rodent modelof unilateral MCA occlusion/reperfusion using radiolabeled annexin V, Eur. J. Nucl.Med. Mol. Imaging 31 (2004) 733–739.
[5] R.C. Ladner, A.K. Sato, J. Gorzelany, M. de Souza, Phage display-derivedpeptides as therapeutic alternatives to antibodies, Drug Discov. Today 9(2004) 525–529.
[6] S. Lee, J. Xie, X. Chen, Peptides and peptide hormones for molecular imaging anddisease diagnosis, Chem. Rev. 110 (2010) 3087–3111.
[7] F.O. Martinez, S. Gordon, M. Locati, A. Mantovani, Transcriptional profiling ofthe human monocyte-to-macrophage differentiation and polarization: newmolecules and patterns of gene expression, J. Immunol. 177 (2006)7303–7311.
[8] J.A. Joyce, P. Laakkonen, M. Bernasconi, G. Bergers, E. Ruoslahti, D. Hanahan, Stage-specific vascular markers revealed by phage display in a mouse model ofpancreatic islet tumorigenesis, Cancer Cell 4 (2003) 393–403.
[9] S.M. Lee, E.J. Lee, H.Y. Hong, M.K. Kwon, T.H. Kwon, J.Y. Choi, R.W. Park, T.G. Kwon,E.S. Yoo, G.S. Yoon, I.S. Kim, E. Ruoslahti, B.H. Lee, Targeting bladder tumor cells invivo and in the urine with a peptide identified by phage display, Mol. Cancer Res. 5(2007) 11–19.
[10] P.L. Hsiung, J. Hardy, S. Friedland, R. Soetikno, C.B. Du, A.P. Wu, P. Sahbaie, J.M.Crawford, A.W. Lowe, C.H. Contag, T.D. Wang, Detection of colonic dysplasia invivo using a targeted heptapeptide and confocal microendoscopy, Nat. Med. 14(2008) 454–458.
[11] D.K. Chang, C.T. Lin, C.H. Wu, H.C. Wu, A novel peptide enhances therapeuticefficacy of liposomal anti-cancer drugs inmice models of human lung cancer, PLoSONE 4 (2009) e4171.
[12] N. Thapa, H.Y. Hong, P. Sangeetha, I.S. Kim, J. Yoo, K. Rhee, G.T. Oh, I.C. Kwon, B.H.Lee, Identification of a peptide ligand recognizing dysfunctional endothelial cellsfor targeting atherosclerosis, J. Control. Release 131 (2008) 27–33.
[13] K. Park, H.Y. Hong, H.J. Moon, B.H. Lee, I.S. Kim, I.C. Kwon, K. Rhee, A newatherosclerotic lesion probe based on hydrophobically modified chitosannanoparticles functionalized by the atherosclerotic plaque targeted peptides,J. Control. Release 128 (2008) 217–223.
[14] H.Y. Hong, J.S. Choi, Y.J. Kim, H.Y. Lee, W. Kwak, J. Yoo, J.T. Lee, T.H. Kwon, I.S. Kim,H.S. Han, B.H. Lee, Detection of apoptosis in a rat model of focal cerebral ischemiausing a homing peptide selected from in vivo phage display, J. Control. Release131 (2008) 167–172.
[15] P. Scaffidi, T. Misteli, M.E. Bianchi, Release of chromatin protein HMGB1 bynecrotic cells triggers inflammation, Nature 418 (2002) 191–195.
[16] H.I. Jung, M.I. Kettunen, B. Davletov, K.M. Brindle, Detection of apoptosisusing the C2A domain of synaptotagmin I, Bioconjug. Chem. 15 (2004)983–987.
[17] R. Shao, C. Xiong, X. Wen, J.G. Gelovani, C. Li, Targeting phosphatidylserine onapoptotic cells with phages and peptides selected from a bacteriophage displaylibrary, Mol. Imaging 6 (2007) 417–426.
[18] N. Thapa, S. Kim, I.S. So, B.H. Lee, I.C. Kwon, K. Choi, I.S. Kim, Discovery of aphosphatidylserine-recognizing peptide and its utility in molecular imaging oftumour apoptosis, J. Cell. Mol. Med. 12 (2008) 1649–1660.
[19] K. Fischer, S. Voelkl, J. Berger, R. Andreesen, T. Pomorski, A. Mackensen, Antigenrecognition induces phosphatidylserine exposure on the cell surface of humanCD8+ T cells, Blood 108 (2006) 4094–4101.
[20] D. Smrz, L. Draberova, P. Draber, Non-apoptotic phosphatidylserine externaliza-tion induced by engagement of glycosylphosphatidylinositol-anchored proteins,J. Biol. Chem. 282 (2007) 10487–10497.
[21] L.E. Edgington, A.B. Berger, G. Blum, V.E. Albrow, M.G. Paulick, N. Lineberry, M.Bogyo, Noninvasive optical imaging of apoptosis by caspase-targeted activity-based probes, Nat. Med. 15 (2009) 967–973.
[22] K. Kim, M. Lee, H. Park, J.H. Kim, S. Kim, H. Chung, K. Choi, I.S. Kim, B.L. Seong, I.C.Kwon, Cell-permeable and biocompatible polymeric nanoparticles for apoptosisimaging, J. Am. Chem. Soc. 128 (2006) 3490–3491.
[23] R. Aloya, A. Shirvan, H. Grimberg, A. Reshef, G. Levin, D. Kidron, A. Cohen, I. Ziv,Molecular imaging of cell death in vivo by a novel small molecule probe,Apoptosis 11 (2006) 2089–2101.
[24] H. Grimberg, G. Levin, A. Shirvan, A. Cohen, M. Yogev-Falach, A. Reshef, I. Ziv,Monitoring of tumor response to chemotherapy in vivo by a novel small-moleculedetector of apoptosis, Apoptosis 14 (2009) 257–267.
[25] K. Ayukawa, S. Taniguchi, J. Masumoto, S. Hashimoto, H. Sarvotham, A. Hara, T.Aoyama, J. Sagara, La autoantigen is cleaved in the COOH terminus and loses thenuclear localization signal during apoptosis, J. Biol. Chem. 275 (2000)34465–34470.
[26] F. Al-Ejeh, J.M. Darby, M.P. Brown, The La autoantigen is a malignancy-associatedcell death target that is induced by DNA-damaging drugs, Clin. Cancer Res. 13(2007) 5509s–5518s.
[27] F. Al-Ejeh, J.M. Darby, C. Tsopelas, D. Smyth, J. Manavis, M.P. Brown,APOMAB, a La-specific monoclonal antibody, detects the apoptotic tumorresponse to life-prolonging and DNA-damaging chemotherapy, PLoS ONE 4(2009) e4558.
[28] D. Wu, A. Ingram, J.H. Lahti, B. Mazza, J. Grenet, A. Kapoor, L. Liu, V.J. Kidd, D. Tang,Apoptotic release of histones from nucleosomes, J. Biol. Chem. 277 (2002)12001–12008.
[29] M. Radic, T. Marion, M. Monestier, Nucleosomes are exposed at the cell surface inapoptosis, J. Immunol. 172 (2004) 6692–6700.
[30] S. Ohsawa, S. Hamada, H. Yoshida, M.Miura, Caspase-mediated changes in histoneH1 in early apoptosis: prolonged caspase activation in developing olfactorysensory neurons, Cell Death Differ. 15 (2008) 1429–1439.
[31] N. Yan, Y. Shi, Histone H1.2 as a trigger for apoptosis, Nat. Struct. Biol. 10 (2003)983–985.
[32] A. Konishi, S. Shimizu, J. Hirota, T. Takao, Y. Fan, Y. Matsuoka, L. Zhang, Y. Yoneda,Y. Fujii, A.I. Skoultchi, Y. Tsujimoto, Involvement of histone H1.2 in apoptosisinduced by DNA double-strand breaks, Cell 114 (2003) 673–688.
[33] I. Vermes, C. Haanen, H. Steffens-Nakken, C. Reutelingsperger, A novel assay forapoptosis. Flow cytometric detection of phosphatidylserine expression on earlyapoptotic cells using fluorescein labelled Annexin V, J. Immunol. Methods 184(1995) 39–51.
[34] W. Hansen, K. Loser, A.M. Westendorf, D. Bruder, S. Pfoertner, C. Siewert, J. Huehn,S. Beissert, J. Buer, G protein-coupled receptor 83 overexpression in naive CD4+CD25-T cells leads to the induction of Foxp3+ regulatory T cells in vivo,J. Immunol. 177 (2006) 209–215.
[35] T. Teesalu, K.N. Sugahara, V.R. Kotamraju, E. Ruoslahti, C-end rule peptidesmediate neuropilin-1-dependent cell, vascular, and tissue penetration, Proc. Natl.Acad. Sci. U.S.A. 106 (2009) 16157–16162.
[36] K.N. Sugahara, T. Teesalu, P.P. Karmali, V.R. Kotamraju, L. Agemy, O.M. Girard, D.Hanahan, R.F. Mattrey, E. Ruoslahti, Tissue-penetrating delivery of compoundsand nanoparticles into tumors, Cancer Cell 16 (2009) 510–520.




















