
BPV1 E2 protein enhances packaging of full-length plasmid DNA in BPV1 pseudovirions
Upload
un-lincolnCategory
view
0download
0
ORIGINAL PAPER
In Planta Evidence for the Involvement of a UbiquitinConjugating Enzyme (UBC E2 clade) in Negative Regulationof Disease Resistance
Turgay Unver & Mine Turktas & Hikmet Budak
# Springer-Verlag 2012
Abstract One of the three main components of the ubiqui-tylation system is an ubiquitin-conjugating enzyme (UBC,E2) that attaches ubiquitin (Ub) to a substrate. A strong linkhas been discovered between ubiquitylation and regulationof plant defense responses against plant pathogens. In thisstudy, the role of UBC in response to pathogen attack isinvestigated in Arabidopsis thaliana as it has three Ubc genehomologs (Ubc1-1, Ubc2-1, and Ubc3-1). Analysis of sin-gle mutants of these three Ubc genes revealed enhanceddisease resistance. Moreover, virus-induced gene silencing(VIGS) analysis revealed an enhanced level of resistance ofNicotiana benthamiana following inoculation with avirulentPto DC3000 and virulent Pto DC3000 hopQ1-1 strains. Inaddition, RAR1, SGT1, and HSP90 mRNA levels are up-regulated following Ubc2 silencing. This study shows thatUBC2 negatively regulates disease resistance via SGT1level. Sub-cellular localization assay of UBC2 protein indi-cates its nuclear and cytoplasmic localization in planta.
Keywords Ubiquitylation . AtUbc2 . VIGS . Plant diseaseresistance . SGT1
Introduction
Commonly, proteins in plants are synthesized, folded, modi-fied and proteolized throughout their lifespans. Poly-ubiquitylation of proteins is used for selective turnover. Ubiq-uitin (Ub) is a tag used as a recognition signal; Ub is covalentlyattached to target proteins for selective proteolysis by the 26Sproteosome system, which is a highly detailed, specific, com-plex and regulated mechanism (Sullivan et al. 2003).
Ub is a 76-amino-acid protein that is expressed ubiqui-tously in eukaryotic cells. Ub labels proteins specifically formono- and poly-ubiquitylation. Ubiquitylation is involvedin many important pathways in plants, such as regulatingcell activities like signal transduction, cell cycle checks,flowering time regulation, and transcription (Cao et al.2008; Dharmasiri et al. 2005; Guo et al. 2003; Hershkoand Ciechanover 1998; Kim and Delaney 2002). There arethree main components in this system: E1 (Ub-activatingenzyme, UBA), E2 (Ub conjugating enzyme, UBC) and E3(Ub ligase). E1 activates Ub by forming an ATP-dependentthioester bond between the carboxy-terminal Gly of Ub andthe Cys residue of UBA. Activated Ub is then transferred toE2 by trans-esterification. E3 ubiquitin ligases are responsiblefor recognition of target proteins. The Ub of E2-Ub interme-diate is transferred to its target substrate by E3 via a reaction inwhich the C-terminus of Ub is linked with Lys residue oftarget protein (Vierstra 2009).
Ubiquitylation is involved in many important processes,such as plant growth, development, hormone regulation,flowering, and biotic and abiotic plant stress response(Dreher and Callis 2007). A UBC domain and active cyste-ine site were predicted in 37 Arabidopsis thaliana proteins,and E2 activity for 17 of these proteins, including AtUbc1-1,AtUbc2-1, and AtUbc3-1, was determined(Kraft et al..2005). On the other hand, it has been shown that AtUbc2-1 complements the rad6 mutant phenotype in yeast, which is
Electronic supplementary material The online version of this article(doi:10.1007/s11105-012-0502-y) contains supplementary material,which is available to authorized users.
T. Unver :M. Turktas :H. Budak (*)Biological Sciences and Bioengineering Program,Faculty of Engineering and Natural Sciences, Sabanci University,Istanbul, Turkeye-mail: [email protected]
T. UnverFaculty of Science, Department of Biology,Cankiri Karatekin University,Cankiri, Turkey
Plant Mol Biol RepDOI 10.1007/s11105-012-0502-y

defective in DNA damage-induced mutagenesis and sensitiveto DNA-damaging agents such as UV radiation/light. Xu et al.(2009a, b) showed that a Atubc1-1 and Atubc2-1 doublemutant exhibits an early-flowering phenotype with a reducednumber of rosette leaves and decreased expression levels inthe floral repressor genes, including Flowering Locus C(FLC), MADS Associated Flowering 4 (MAF4) and MAF5.
The ubiquitylation-mediated post translational modifica-tion system plays regulatory and essential roles in plantimmunity. The system components regulate plant diseaseresistances both positively and negatively. CMPG1, a U-box type E3 ubiquitin ligase, positively regulates immunity;however, RIN1, a RPM1-interacting RING finger E3 ligasewas found to negatively regulate disease response (Holt et al.2002; Yang et al. 2006) in Arabidopsis. The RING finger/Ubox type of E3 ubiquitin ligases also plays an important rolein plant disease resistance. CMPG1 (Avr9/Cf-9 rapidly eli-cited, ACRE-74) is one of the rapidly elicited U-box domaincontaining proteins in tomato and tobacco that has been prov-en to be a positive regulator for plant cell death (PCD) (Yanget al. 2006). Another example of a negative regulatory func-tion in plant immunity is SON1—an F-box containing SCFtype E3 ligase component (Kim and Delaney 2002). Spl11, aU-box type E3 ligase, is an interesting example cloned in rice.Although its homologues regulate immunity positively inArabidopsis and tobacco (U-box-containing E3 ubiquitinligases PUB17 and CMPG1, respectively), mutation inSpl11 in rice causes a hypersensitive response (HR). Spl11mutants respond efficiently to blight rice pathogen, whichmeans that SPL11 is a negative regulator for cell death andimmunity (Zeng et al. 2004). Recently, Trujillo et al. (2008)identified three homologous U-box E3s, PUB22, PUB23,PUB24, that negatively regulate PAMP triggered immunity(PTI), in Arabidopsis. The triple mutant pub22/pub23/pub24showed de-repression and decreased intensity in PTI, suggest-ing negative regulation of PTI by PAMP recognition (Trujilloet al. 2008).
The given findings suggest a regulatory role for theubiquitylation mechanism in plant immunity. Plant Sgt1has been cloned and its role in plant immunity verified(Austin et al. 2002; Azevedo et al. 2006; Tör et al. 2002).SGT1 proteins in plants are required for efficient defenseresponse against a variety of pathogens. The association ofSGT1 with SKP1 was also discovered in plants (Austin et al.2002; Azevedo et al. 2006; Peart et al. 2002). Severalputative E3 ligases have emerged as regulators for diseaseresistance (Devoto et al. 2003). F-box is a subunit of SCFtype E3 ligase; sequences of 694 potential F-box proteins inArabidopsis data have been published (Gagne et al. 2002).F-box proteins might play diverse roles in the regulation ofplant resistance. Two different F-box proteins that are im-portant for disease resistance have been identified: mutationof the Coronatine Insensitive 1 (Coi1) gene in Arabidopsis
(Xie et al. 1998) renders plants susceptible to insect, fungaland bacterial pathogen attacks (McConn et al. 1997;Thomma et al. 1998); and the COI1 protein (including anF-box motif and LRR domain) regulate disease responsesvia Jasmonic acid (Devoto et al. 2003). In contrast, mutationin SON1, another F-box protein, confers resistance to fungaland bacterial pathogen infections. A genetic screen for son1mutants showed that SON1 is a negative regulator of plantdisease resistance (Kim and Delaney 2002). Other compo-nents of E3 ligases are also functionally related to regulationof plant disease resistance. RBX1 protein has been deter-mined as a regulator of plant response to pathogens. Thesilencing of the RBX1 gene in Arabidopsis results in thesame phenotype as in coi1 mutants (Xu et al. 2002). Therole of E2 in abiotic stress responses in soybean was studiedby Zhou et al. (2010). The expression level of GmUbc2, aRad6 soybean gene homologue, was determined to be in-duced by salt and drought stresses. These molecular andphysiological results suggested that GmUbc2 is involved inthe regulation of a set of important stress-responsive genesin Arabidopsis to improve plant tolerance.
In this study, we aimed to determine the role of the Ubc2gene in plant disease resistance. A role for some of the E3ligase components of the ubiquitin pathway has been im-plied in disease resistance. On the other hand, since notmuch is known about the role of E2, which is upstream ofE3 in the ubiquitin pathway, we decided to analyze the roleof E2 in plant disease resistance. To accomplish this goal,three separate Arabidopsis Ubc gene homologue mutants(ubc1-1, ubc2-1, and ubc3-1) were determined and growthof both virulent Pto DC3000 hopQ1-1 and avirulent PtoDC3000 pathogen strains inoculated on these mutants wasmeasured. One of the Arabidopsis ubc mutants (ubc2-1) wascomplemented by the AtUbc2 gene. Overexpression of theAtUbc2 gene in the wild type Arabidopsis line was con-structed and tested with different Pseudomonas syringaestrains. The subcellular localization of the AtUBC2 proteinwas discovered. Virus induced gene silencing (VIGS) forsuppression of NbUbc2 on leaves of Nicotiana benthamianawas also analyzed. Upon silencing, some disease resistance,signaling gene expression, was noted. Pathogen growth ofboth virulent and avirulent inoculations on Ubc2 silencedleaves was measured. Transient viral-based overexpressionof the NbUbc2 gene in N. benthamiana leaves was performedfor further UBC2-SGT1 in planta interaction studies.
Materials and Methods
Plant Materials and Growth Conditions
Arabidopsis seeds from the three ubc mutants (ubc1-1,ubc2-1, ubc3-1) were obtained from NASC (European
Plant Mol Biol Rep

Arabidopsis Stock Centre, http://arabidopsis.info/), and cor-respond to T-DNA insertion lines (ubc1-1) SALK_109196,(ubc2-1) SALK_060994, (ubc3-1) SALK_045233. These T-DNA insertion lines were generated and supplied by Arabi-dopsis Biological Resource Center (ABRC) (http://abrc.osu.edu/). Since Col0 is the background for these mutants, aCol5 wild type line was also grown. Seeds were sown andplaced in pots at 4 °C for 1–2 days to synchronizegermination. The pots were then moved to a growth cabinet(Digitech, http://www.digitech-ltd.com) at 20 °C and 70 %humidity under 12 h of fluorescent light (light intensity~150 μEinsteins m−2 s−1). Identical growth chamber condi-tions were used for N. benthamiana growth.
Pathogen Tests and Growth Measurements
Pto DC3000 strains were taken out from −80 °C glycerolstocks and inoculated to a low salt Luria Bertani (LB)medium (10 g/L tryptone, 5 g/L yeast extract and 5 g/LNaCl, pH 7.0) with rifampicin (Rif) 50 (μg/ml) antibioticand grown for 2 days at 28 °C. Bacterial cultures weregrown to reach OD60001 and then centrifuged at 2,500rpm for 10 min. We suspended the bacterial culture to afinal concentration of 105cfu/ml for vacuum infiltration ofArabidopsis. Silwet L-77, a surfactant, was added to theinoculum at 0.004 % (40 μl/L) to help the bacteria invadethe apoplastic fluid. Vacuum inoculation was used for bothnon pathogenic (Pto DC3000 hrcC-) and virulent (PtoDC3000) pathogens as described in Katagiri et al. (2002).For N. benthamiana pathogen growth measurement testsavirulent Pto DC3000 and virulent Pto DC3000 hopQ1-1strains were inoculated with a blunt syringe. Strains wereinoculated at 104cfu/ml onto 6-week post germination N.benthamiana leaves. Pathogen enumeration on Arabidopsisand N. benthamiana leaves was measured according to Weiet al. (2007). Briefly, three leaves from Arabidopsis plants orthree leaf discs from N. benthamiana were ground in 300 ml10 mM MgCl2 and 100 mM sucrose, and serial dilutions(1/5 1/10, 1/20, 1/40, 1/80) were spotted onto low salt LBmedium with Rif. The cfu count of the pathogen strains wasdetermined 2 days after incubation at 28 °C. Each pathogentest was repeated independently at least three times. Thepathogen strains were kindly provided by Dr. Alan Collmerof Cornell University.
Virus-Induced Gene Silencing
To efficiently suppress NbUbc2 gene expression, primersfor cloning of Ubc2 in N. benthamiana using SpeI andHindIII overhangs, NbUbc2 F/speI 5′ GCGACTAGT ATTTTAAGCGGTTACAGCAGG and NbUbc2R/HindIII 5′GCGAAGCTT AAGACTGGATTGAAGTCAGTA weredesigned. The PCR products were cloned into pGEMT-
Easy vector. pTV:00 and pGEMT-Easy NbUbc2SpeI-Hin-dIII plasmids were then double digested with SpeI andHindIII enzymes. The 300 base long region suitable as aneffective silencing insert was ligated with digested pTV:00to obtain the TRV2:NbUbc2 (also called TRV2:UBC2) con-struct used to silence the NbUbc2 gene. The construct wasapplied to 2-week-old N. benthamiana leaves (see Unverand Budak 2009a). Briefly, pTRV-NbUbc2 transformed A.tumefaciens LBA4404 and pTRV1 transformed by electro-poration were incubated overnight at 28 °C to reach anOD600 0 0.8. These two cultures were centrifuged and re-suspended in induction medium (10 mM MgCl2, 10 mMMES, 200 mM acetosyringone) at room temperature for 2 h.TRV1 and TRV2 cultures (TRV2: Empty or TRV2: UBC2to be TRV2) were mixed in a 1:1 ratio. The culture was theninfiltrated to the underside of the leaves using a 1 ml needle-less syringe.
qRT-PCR
TRV2: Ubc2 silenced N. benthamiana plants were analyzedwith qRT-PCR to measure silencing level (using real-timePCR). First of all, TRV2: NbUbc2 and TRV2: Empty si-lenced N. benthamiana leaves were collected. Total RNAwas isolated as outlined by Budak et al. (2006). DNAsetreatment, lithium chloride precipitation were applied toRNA samples (Unver and Budak 2009b). cDNAs of si-lenced and viral control samples were synthesized from2 μg total RNA with Superscript III reverse transcriptase(Invitrogen, Carlsbad, CA) according to the manufacturer’sinstructions. qRT-PCR analysis was performed as outlinedby Unver et al. (2010). Briefly, 2 μl cDNA was amplifiedwith (1 μl) 1μM specific primers in a total volume of 20 μlusing SYBR green PCR master mix (Applied Biosystems,Foster City, CA) with Icycler Multicolor Real-time PCRDetection Systems (Bio-Rad, Richmond, CA). The quanti-fication was performed using the 18S rRNA gene as aninternal reference and three independent PCR results withacceptable efficiency (1.8–2.1) were averaged (Ergen andBudak 2009). qRT-PCR analyses were performed for triplebiological and triple technical replicates. To measure thesilencing level of the Ubc2 gene upon TRV-mediated VIGS,the following primers were used: qRT-NbUbc2 Forward 5′CTG TGA TCC AAA TCC CAA CTC 3′ qRT-NbUbc2Reverse 5′ ACA ATC TCG CGC ACC TTC 3′ (product size96 bp). Normalization experiments were performed with 18SrRNA genes 18S rRNA forward, 5′ GTGACGGGTGACGGAGAATT and 18S reverse 5′ GACACTAATGCGCCCGGTAT. RAR1, SGT1, and HSP90 gene expression levels weredetermined in Ubc2-suppressed N. benthamiana leaves. Thefollowing primers were used for RAR1 silencing level; Rar1Forward 5′ TGATGGCATGAAAGAGTGGA and Rar1 Re-verse 5′ TGGACTGGAGCTGACTTTGG, for Sgt1 level;
Plant Mol Biol Rep

Sgt1 Forward 5′AGCTTTTGCCAGGCGTATTTandReverse5′ GCGTGGTTGTTGACTTTGGT, and for Hsp90 level de-termination; Hsp90 Forward 5′ CGACCAGCACGCTCACGAT and Reverse 5′GCGATGGTCCCGAGGTTGT.
Localization Assay
We used the 35S-Ubc2-GFP-His construct to determine thesub-cellular localization of UBC2 (3′ GFP) in planta. A.tumefaciens LBA4404 strain was transformed with this con-struct, and onion inner epidermal cells were transformed asdescribed. Healthy and fresh onion scales (1–1.5×1 cm)were placed in a Petri dish and the inner surfaces wereimmersed in 10 mL resuspension Agrobacterium solution(OD60001–1.5) consisting of 5 % (g/v) sucrose, 100 mgacetosyringone/L for12h at 28 °C. The onion explants werethen transferred to a Petri dish containing MS0 [Murashigeand Skoog salts, 30 g sucrose/L and 0.7 % (g/v) agar,pH 5.7] and co-cultivated with Agrobacterium for 1–2 days.
Complementation of Mutant Plants and OverexpressorConstruction
The full open reading frame of AtUbc2 gene was amplifiedby PCR using 5′ GGGGACAAGTTTGTACAAAAAAGCAGGCTTGTCGACTCCAGCGAGGAAGA 3′ (Attb1-for-ward) and 5′GGGGACCACTTTGTACAAGAAAGCTGGGTTCTAGTCGGCAGTCCAGCTTTGT 3′ (Attb2-re-verse, stop codon containing), 5′ GGGGACCACTTTGTA-CAAGAAAGCTGGGTT GTCGGCAGTCCAGCTTTGTTCG 3′(Attb2-reverse, no stop codon containing) pri-mers containing Attb sites for gateway cloning. The resultingproduct was cloned into pEarleyGate 100 and pEarleyGate103 (C terminal GFP includes) vectors to overexpress thegene under the control of the 35S promoter (Ma et al. 2011).The pEarleyGate 100-AtUbc2 and pEarleyGate 103-AtUbc2constructs were transferred into A. tumefaciens (strainLBA4404) by electroporation. Arabidopsis plants (Col-5
ecotype and ubc2 mutant which has Col-0 background) weretransformed by the floral dip method (Clough and Bent 1998).T1 generation seeds were harvested and screened on MSmedium containing 50 mg/L kanamycin. Homozygous trans-genic lines were selected by screening for kanamycin resis-tance with BASTA spray as described by Clough and Bent(1998).
Transient Overexpression in N. benthamiana
NbUbc2 was transiently overexpressed in N. benthamianausing the viral-based vector pGR106 (the viral vector waskindly supplied by D. Baulcombe of Cambridge University).A FLAG tag was added at the 5′ of the Ubc2 gene with PCRprimers as NbUbc2ClaIFLAG 5′ AAATCGATATGGATTACAAGGATGACGACGATAAGatgtcg acaccggcgaggaagagaand NbUbc2NotI-R 5′ GGAGCGGCCGCTCAGTCTGCTGTCCAGCTCTGTTCA. The PCR products were ligat-ed with pGEMT-Easy for sequence analysis. Positive cloneswere selected and sequenced. The clone was double digestedwith ClaI and NotI for further pGR106 cloning. The ClaI-FLAG-NbUbc2-NotI insert was ligated with double digestedpGR106 and then transformed into A. tumefaciens LBA4404with electroporation. Colony PCR selection was performedfor validation of transformation. A. tumefaciens LBA440435S: NbUbc2 (pGR106) was infiltrated to 4-week-old N.benthamiana leaves.
Co-immunoprecipitation
Co-immunoprecipitation (Co-Ip) of UBC2 and SGT1 pro-teins was performed using the FLAG-tagged protein immu-noprecipitation kit (Sigma, St. Louis, MO) as describedpreviously (Tian et al. 2004). A 1 nmol quantity of purifiedFLAG tagged Ubc2 was preincubated with 500 μl N. ben-thamiana total proteins for overnight at 4 °C. A 40 μlquantity of anti-FLAGM2resin was added and incubated at4 °C overnight with gentle shaking. The precipitated protein
0
1
2
3
4
5
6
7
8
9
a
b
aa
bb
a
aa
bb
a a
0 dpi 2 dpi 4 dpi
Bac
teri
al p
opul
atio
n lo
g c
fu/m
g
Fig. 1 Growth ofPseudomonas syringae ptvtomato DC3000 is decreasedin ubc1–3 mutant Arabidopsisbut not effected in AtUbc2overexpressors (35S::UBC2and 35S::UBC2-GFPArabidopsis thaliana Col-5lines). Experiments were re-peated three times with similarresults. Means with the sameletter were not significantlydifferent at the 5 % confidencelevel based on Duncan’smultiple range test
Plant Mol Biol Rep

complexes were eluted in 40 μl with 150 ng/μl FLAGpeptide solution and were analyzed by SDS/PAGE andwestern blot.
Western Blotting
The rat anti-SGSa antibody used in this study was describedby Azevedo et al. (2006) (supplied by Ken Shirasu,RIKEN). Total protein extracts from agro-infiltrated N. ben-thamiana leaves were prepared by grinding 2 cm2 leaf tissuein 100 μl ice-cold 1 X extraction buffer [10 % glycerol,25 mM Tris-HCl pH 7.5, 1 mM EDTA, 150 mM NaCl,5 mM dithiothreitol (DTT) and 1 % of plant protease inhib-itor cocktail (Sigma)] and 4 X of SDS-PAGE sample buffer.Samples were boiled for 5 min and 10 μl run on a 12 %SDS-PAGE gel, blotted onto PVDF membranes and visual-ized using antibodies described with Pierce protein researchkit (Thermo Fischer Scientific, Waltham, MA).
Results
Detection of Three UBC Clade Mutants
The A. thalianagenome contains three RAD6 gene homo-logs encoded by At1g14400, At2g02760, and At5g62550(UBC clade). The single ubc1-1, ubc2-1, and ubc3-1 mutantlines, SALK_109196, SALK_060994 and SALK_045233,respectively, were identified from the SALK collection(Alonso et al. 2003), and homozygous mutant plants wereselected by PCR. Pathogen tests indicated enhanced resis-tance associated with ubc mutants.
To measure the level of pathogen growth on the A.thaliana ubc mutants, the disease-causing virulent inocula-tion Pto DC3000 was applied. Since Col-0 is the back-ground for ubc mutants, virulent pathogen growth wasenumerated for comparison (Figs. 1 and 2). On the otherhand, non-pathogenic Pto DC3000 hrcC- was inoculated onthe ubc mutants and Col-0 plants to observe whether resis-tance is enhanced or reduced (Fig. 3). Col-0 wild type andArabidopsis ubc mutants were inoculated with 105 cfu/mlvirulent Pto DC3000 hopQ1-1 strain using vacuum infiltra-tion. The ubc mutants showed enhanced resistance uponvirulent inoculation compared to Col-0 (Fig. 1). Upon inoc-ulation of avirulent Pto DC3000 strain on the ubc mutants,slightly elevated resistance compared to resistant wild typewas observed (Fig. 3, Suppl. Fig. 1).
AtUbc2 Overexpressor A. thaliana Lines do not AffectResistance
35S::UBC2 and 35S::UBC2-GFP plants were generated toevaluate avirulent Pto DC3000 and virulent Pto DC3000
Col-0
ubc2-2
Col-0
mutant
mock
ubc2
ubc2-3
2-1 mutant
3 mutant
Col-5
Col-5::3
control
35S:UBC2:GGFP
Col-5::3
Col-5 m
35S:UBC2
mock
Fig. 2 Representative P. syringae ptv tomato DC3000 6dpi growth.Disease symptoms are seen on wild type and overexpressors. Enhancedresistance is detected on ubc1–3 mutants. Upper panel Mutant plants,lower panel A. thaliana complemented with Ubc2 gene
Plant Mol Biol Rep

hopQ1-1 strain growth. Since the Col-5 wild type plantswere used for overexpressor line construction, pathogengrowth on transgenic lines was compared with Col-5.35S:: UBC2 and 35S:: UBC2-GFP plants were found tobe nearly as susceptible against Pto DC3000 pathogen in-oculation as Col-5 (Figs. 1 and 2). On the other hand,enhanced resistance level results from a lack of Ubc2 geneexpression, slightly altered by overexpression of Ubc2against Pto DC3000 hrcC- (Fig. 3, Suppl. Fig. 1).
Ubc2 Complementation Causes Increased Pathogen Growthon A. thaliana ubc2 Mutant
AtUbc2 gene function was complemented in Arabidopsisubc2 mutants by Agrobacterium-mediated transformationof a 35S::AtUbc2 construct. Selected complementationplants were inoculated with virulent Pto DC3000 hopQ1-1and avirulent Pto DC3000 hrcC- strains. Pathogen growth wasmeasured on Arabidopsis ubc2:35S-Ubc2 (complemented)and Col-0 (wild type) plants (Fig. 4). As expected, pathogensusceptibility was returned in complemented Arabidopsisseedlings with a gain-of-function study (Fig. 5).
Ubc2 Silencing in N. benthamiana Results in ElevatedResistance
We aimed to silence the N. benthamiana (Nb) Ubc2 gene tofunctionally analyze the role of Ubc2 in plant disease resis-tance. N. benthamiana is used as a model plant system formolecular plant microbe interaction studies. Therefore, weintended to suppress Ubc2 gene expression and inoculatewith both characterized virulent and avirulent race specificpathogens. Prior to the silencing experiment, the full-lengthN. benthamiana Ubc2 gene was cloned for the first time inthis study. It has been determined that NbUbc2 is 96 %similar to N. tabacum Ubc2 at the nucleotide level. Addi-tionally, NbUBC2 protein has 153 amino acids and a UBCcconserved domain containing protein with 96 % similarityto N. tabacum protein at the amino acid sequence level. TheTRV2: NbUBC2 silencing inoculation was constructed bycloning a partialNbUbc2 sequence into pTV:00. The silencingwas applied to 2-week-oldN. benthamiana leaves. To confirmthe negative regulatory role of the Ubc2 gene, another modelplant–pathogen system interaction—N. benthamiana- Pto
0
0,5
1
1,5
2
2,5
3
3,5
0 dpi 2 dpi 4 dpi
Bac
teri
al p
opul
atio
n lo
g c
fu/m
g
Fig. 3 P. syringae ptv tomato DC3000 hrcC inoculation does notcause suppression of resistance on Arabidopsis ubc mutants. AtUbc2overexpressors (35S::UBC2 and 35S::UBC2-GFP A. thaliana Col-5lines) exhibit a slightly more resistant phenotype compared to Col-5
against avirulent pathogen inoculation. Experiments were repeatedthree times with similar results. Means with the same letter were notsignificantly different at the 5 % confidence level based on Duncan’smultiple range test
Complemented Complemented
C D
A B
Fig. 4 a, b Arabidopsis ubc2-1 mutant complemented AtUbc2 gene(35S: ATUBC2) inoculated with P. syringae pv. tomato DC3000 (PtoDC3000) (a) or Pto DC3000 hrcC- (b). c, d Col-0 inoculated with PtoDC3000 (c), Col-0 inoculated with Pto DC3000 hrcC- (d) and picturedat 6dpi
Plant Mol Biol Rep
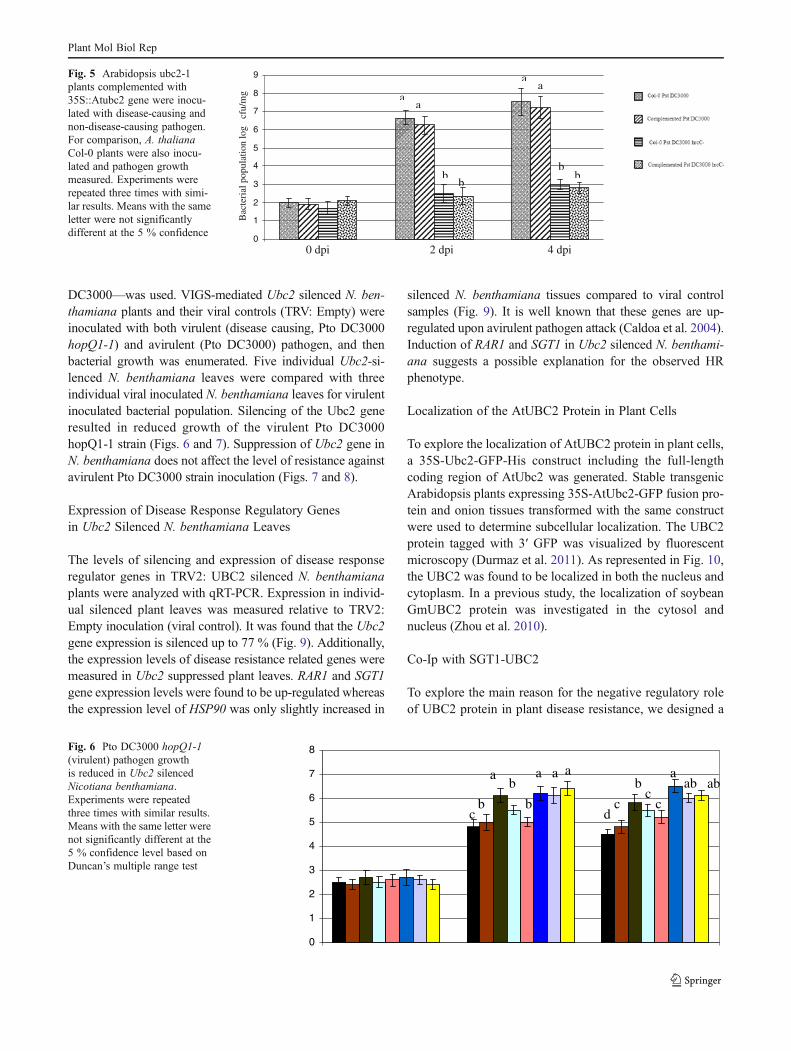
DC3000—was used. VIGS-mediated Ubc2 silenced N. ben-thamiana plants and their viral controls (TRV: Empty) wereinoculated with both virulent (disease causing, Pto DC3000hopQ1-1) and avirulent (Pto DC3000) pathogen, and thenbacterial growth was enumerated. Five individual Ubc2-si-lenced N. benthamiana leaves were compared with threeindividual viral inoculated N. benthamiana leaves for virulentinoculated bacterial population. Silencing of the Ubc2 generesulted in reduced growth of the virulent Pto DC3000hopQ1-1 strain (Figs. 6 and 7). Suppression of Ubc2 gene inN. benthamiana does not affect the level of resistance againstavirulent Pto DC3000 strain inoculation (Figs. 7 and 8).
Expression of Disease Response Regulatory Genesin Ubc2 Silenced N. benthamiana Leaves
The levels of silencing and expression of disease responseregulator genes in TRV2: UBC2 silenced N. benthamianaplants were analyzed with qRT-PCR. Expression in individ-ual silenced plant leaves was measured relative to TRV2:Empty inoculation (viral control). It was found that the Ubc2gene expression is silenced up to 77 % (Fig. 9). Additionally,the expression levels of disease resistance related genes weremeasured in Ubc2 suppressed plant leaves. RAR1 and SGT1gene expression levels were found to be up-regulated whereasthe expression level of HSP90 was only slightly increased in
silenced N. benthamiana tissues compared to viral controlsamples (Fig. 9). It is well known that these genes are up-regulated upon avirulent pathogen attack (Caldoa et al. 2004).Induction of RAR1 and SGT1 in Ubc2 silenced N. benthami-ana suggests a possible explanation for the observed HRphenotype.
Localization of the AtUBC2 Protein in Plant Cells
To explore the localization of AtUBC2 protein in plant cells,a 35S-Ubc2-GFP-His construct including the full-lengthcoding region of AtUbc2 was generated. Stable transgenicArabidopsis plants expressing 35S-AtUbc2-GFP fusion pro-tein and onion tissues transformed with the same constructwere used to determine subcellular localization. The UBC2protein tagged with 3′ GFP was visualized by fluorescentmicroscopy (Durmaz et al. 2011). As represented in Fig. 10,the UBC2 was found to be localized in both the nucleus andcytoplasm. In a previous study, the localization of soybeanGmUBC2 protein was investigated in the cytosol andnucleus (Zhou et al. 2010).
Co-Ip with SGT1-UBC2
To explore the main reason for the negative regulatory roleof UBC2 protein in plant disease resistance, we designed a
0
1
2
3
4
5
6
7
8
9
10 dpi 2 dpi 4 dpi
aa
aa
bb
bb
Bac
teri
al p
opul
atio
n lo
g c
fu/m
g
Fig. 5 Arabidopsis ubc2-1plants complemented with35S::Atubc2 gene were inocu-lated with disease-causing andnon-disease-causing pathogen.For comparison, A. thalianaCol-0 plants were also inocu-lated and pathogen growthmeasured. Experiments wererepeated three times with simi-lar results. Means with the sameletter were not significantlydifferent at the 5 % confidence
0
1
2
3
4
5
6
7
8
cb
ab
b
a a a
dc
bc
c
aab ab
Fig. 6 Pto DC3000 hopQ1-1(virulent) pathogen growthis reduced in Ubc2 silencedNicotiana benthamiana.Experiments were repeatedthree times with similar results.Means with the same letter werenot significantly different at the5 % confidence level based onDuncan’s multiple range test
Plant Mol Biol Rep
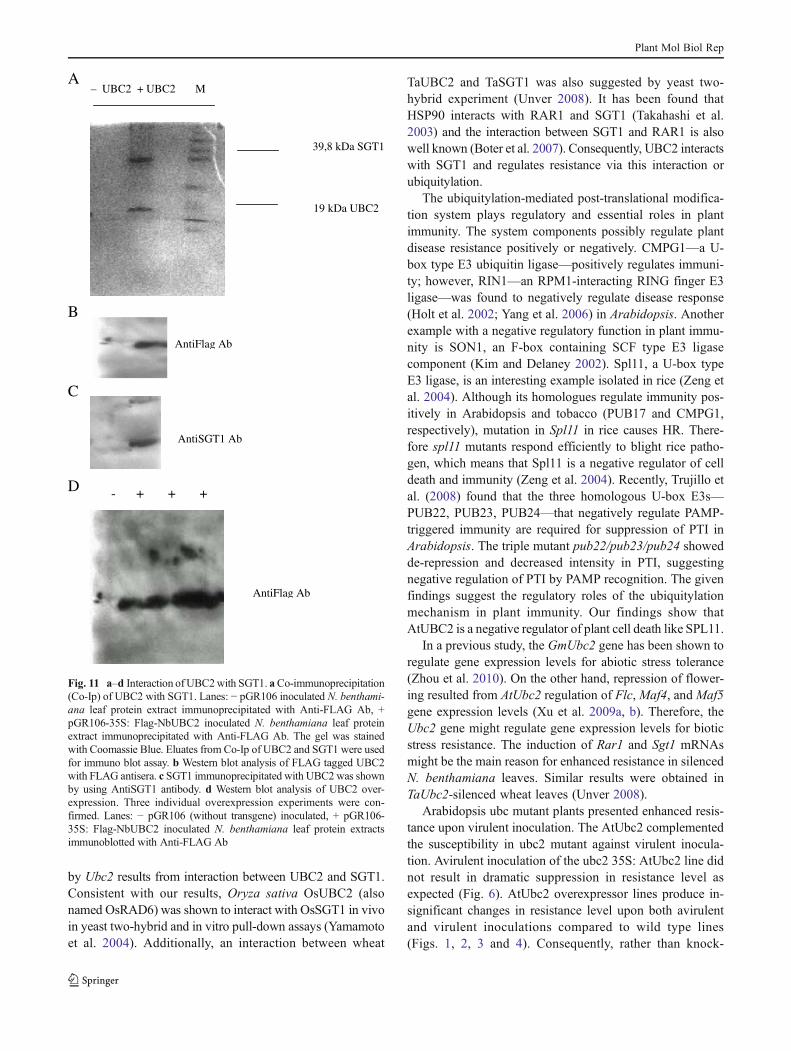
protein–protein interaction assay between SGT1 and UBC2.As a ubiquitylation pathway component, SGT1 plays im-portant roles in plant defense. Since SGT1 proteins in plantsare required for an efficient defense response against avariety of pathogens for immune activation, possibleSGT1 regulation may affect defense responses. On the otherhand we have detected up-regulation of the expression levelof Sgt1 in Ubc2-silenced N. benthamiana leaves (Fig. 9). Todetermine if there is a one-to-one interaction between UBC2and SGT1, a Co-Ip assay was performed with the N-terminalFlag tagged UBC2 and the SGS domain of SGT1; the resultsshowed immunoprecipitation of UBC2 with SGT1 (Fig. 11).Therefore, UBC2 interacts with SGT1 to regulate the plantdisease response. In Ubc2 knock-out Arabidopsis and knock-down N. benthamiana plants, SGT1 is not well regulated andplants show enhanced resistance.
Discussion
In this study, we have identified the role of the AtUbc2 genein plant disease resistance. AtUbc1, AtUbc2, and AtUbc3have a conserved catalytic E2 site (Sullivan and Vierstra1993). It has been shown that human UBC2 (HsUBC2),yeast (Saccharomyces cerevisiae) UBC2 (ScUBC2 or alsonamed RAD6), and AtUBC1, AtUBC2, and AtUBC3(AtUBC2 clade) are classified in the same group of thephylogenetic tree (Kraft et al. 2005). Among them, AtUBC2has the highest similarity with ScUBC2 at protein sequencelevel. Previous studies discovered the diverse roles ofAtUBC2, e.g., the UV-sensitive phenotype of yeast rad6mutant was rescued with AtUBC2 (Zwirn et al. 1997) Inaddition, AtUBC1, AtUBC2 and AtUBC3 proteins play arole in histone H2B monoubiquitylation in flowering timeregulation (Cao et al. 2008; Xu et al. 2009a). Additionally,soybean GmUBC2—a yeast ScUBC2 homolog—plays animportant role in salt and drought stress responses by regu-lating stress-responsive genes (Zhou et al. 2010). Here, wehave discovered the role of AtUBC2 in plant disease resis-tance by negatively regulating expression of resistance-related genes and interacting with SGT1.
It is clear that Ub-mediated degradation of proteins is alsocritical for plant defense mechanisms (Austin et al. 2002;Azevedo et al. 2006; Kim and Delaney 2002; Kitagawa etal. 1999; Trujillo et al. 2008; Xie et al. 1998; Yang et al.2006; Zeng et al. 2004). The expression level of TaUbc2 isdown-regulated upon avirulent pathogen inoculation (Unver2008), suggesting that the Ubc2 might be involved in plantdisease response to pathogen attack. Therefore, we aimed toevaluate the disease resistance level in three Arabidopsismutants. Enhanced resistance was detected in Arabidopsis
Pto DC3000 hopQ1-1
PtoDC3000
PtoDC3000
P to DC3000 hopQ1-1
A) Viral control B) Ubc2 silenced
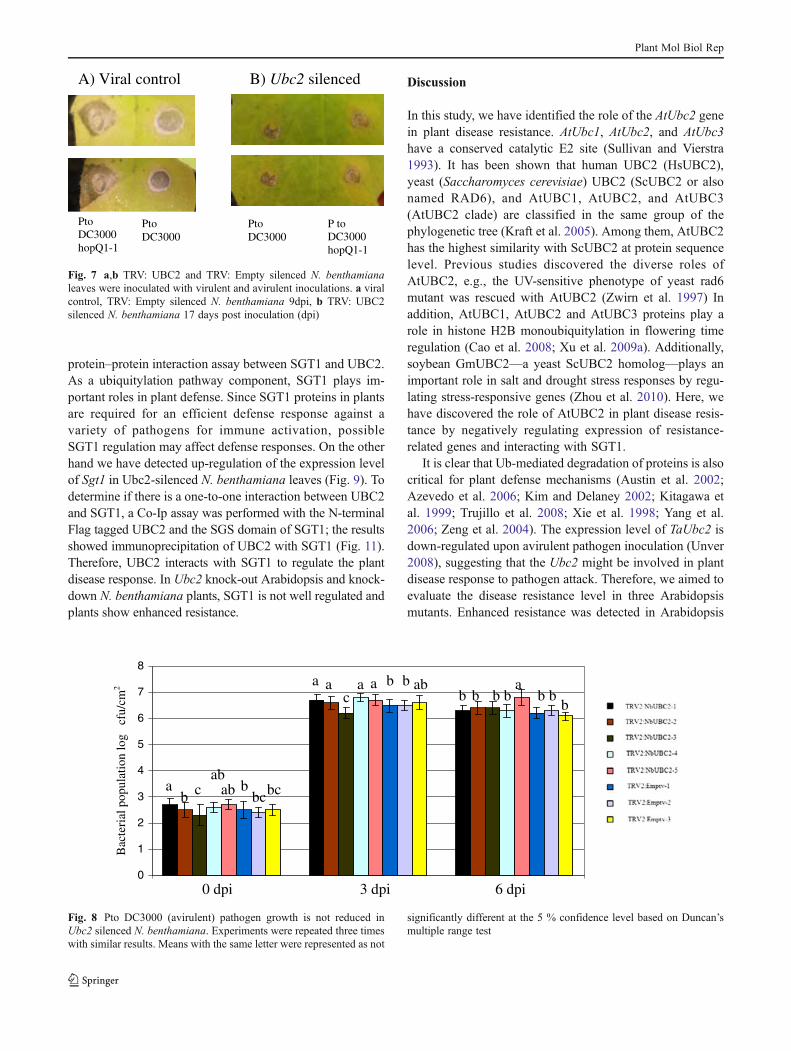
Fig. 7 a,b TRV: UBC2 and TRV: Empty silenced N. benthamianaleaves were inoculated with virulent and avirulent inoculations. a viralcontrol, TRV: Empty silenced N. benthamiana 9dpi, b TRV: UBC2silenced N. benthamiana 17 days post inoculation (dpi)
0
1
2
3
4
5
6
7
8
ab c
abab b
bcbc
a a aac
b b abb b b b
ab
bb
0 dpi 3 dpi 6 dpi
Bac
teri
al p
opul
atio
n lo
g c
fu/c
m2
Fig. 8 Pto DC3000 (avirulent) pathogen growth is not reduced inUbc2 silenced N. benthamiana. Experiments were repeated three timeswith similar results. Means with the same letter were represented as not
significantly different at the 5 % confidence level based on Duncan’smultiple range test
Plant Mol Biol Rep

ubc1, ubc2, and ubc3 mutant plants upon virulent pathogeninoculation. No repression of the resistance level was mea-sured in three Arabidopsis ubc mutants when inoculatedwith avirulent pathogen. NbUbc2-silenced N. benthamianaleaves were assessed in compatible and incompatible plant–pathogen interactions. Similar results were obtained forNbUbc2 silenced N. benthamiana leaves (Figs. 7 and 8).
RAR1, SGT1, and cytosolic HSP90, have been found tofunction in many, but not all, of the NB-LRR R-pathways(Austin et al. 2002; Azevedo et al. 2006; Tor et al. 2002). Toclarify the resistance level elevation and HR response uponknock-down of theNbUbc2 gene, the expression level ofRar1,Sgt1, and Hsp90 genes was measured. mRNA quantification
analysis showed the induction of disease resistance relatedgene expression in NbUbc2-silenced leaves (Fig. 9). IncreasedmRNA levels of Rar1 and Sgt1 results in HR since expressionlevels of these genes were up-regulated upon avirulent patho-gen inoculation in barley (Caldoa et al. 2004). Additionally,Rar1, Sgt1 and Hsp90 suppressed- Wheat and barley plantswere detected as susceptible to pathogen attack (Hein et al.2005; Scofield et al. 2005; Unver 2008; Pei et al. 2011). Hence,induction of Rar1, Sgt1 and Hsp90 expression in Ubc2-silenced tissue causes elevated resistance without pathogeninfection.
Here, we have shown the interaction of SGT1 with UBC2.The regulation of the mRNA levels of Sgt1, Rar1, and Hsp90
Rel
ativ
e ex
pres
sion
leve
l
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
Ubc2
RAR1
SGT1
HSP90
TR
V2:
NbU
BC
1-3
TR
V2:
Em
pty
TR
V2:
NbU
BC
2-1
TR
V2:
NbU
BC
1-2
TR
V2:
NbU
BC
1-4
TR
V2:
NbU
BC
1-3
TR
V2:
Em
pty
TR
V2:
NbU
BC
2-1
TR
V2:
NbU
BC
1-2
TR
V2:
NbU
BC
1-4
TR
V2:
NbU
BC
1-3
TR
V2:
Em
pty
TR
V2:
NbU
BC
2-1
TR
V2:
NbU
BC
1-2
TR
V2:
NbU
BC
1-4
TR
V2:
NbU
BC
1-3
TR
V2:
Em
pty
TR
V2:
NbU
BC
2-1
TR
V2:
NbU
BC
1-2
TR
V2:
NbU
BC
1-4
a
b bb
b
b
aa
aa
c
b
abab
a
Fig. 9 TRV-mediated silencingof Ubc2 gene in N.benthamiana and relativeexpression level changes inexpression of RAR1, SGT1,and HSP90 genes upon Ubc2silencing. Experiments wererepeated three times withsimilar results. Means withthe same letter were notsignificantly different at the5 % confidence level based onDuncan’s multiple range test
Fig. 10 a–d Nuclear andcytoplasmic localization ofAtUBC2. a, b Nuclear andcytoplasmic localization ofC-terminal GFP-tagged UBC2.c Negative control (no GFP).d GFP alone positive controlrepresentation
Plant Mol Biol Rep
by Ubc2 results from interaction between UBC2 and SGT1.Consistent with our results, Oryza sativa OsUBC2 (alsonamed OsRAD6) was shown to interact with OsSGT1 in vivoin yeast two-hybrid and in vitro pull-down assays (Yamamotoet al. 2004). Additionally, an interaction between wheat
TaUBC2 and TaSGT1 was also suggested by yeast two-hybrid experiment (Unver 2008). It has been found thatHSP90 interacts with RAR1 and SGT1 (Takahashi et al.2003) and the interaction between SGT1 and RAR1 is alsowell known (Boter et al. 2007). Consequently, UBC2 interactswith SGT1 and regulates resistance via this interaction orubiquitylation.
The ubiquitylation-mediated post-translational modifica-tion system plays regulatory and essential roles in plantimmunity. The system components possibly regulate plantdisease resistance positively or negatively. CMPG1—a U-box type E3 ubiquitin ligase—positively regulates immuni-ty; however, RIN1—an RPM1-interacting RING finger E3ligase—was found to negatively regulate disease response(Holt et al. 2002; Yang et al. 2006) in Arabidopsis. Anotherexample with a negative regulatory function in plant immu-nity is SON1, an F-box containing SCF type E3 ligasecomponent (Kim and Delaney 2002). Spl11, a U-box typeE3 ligase, is an interesting example isolated in rice (Zeng etal. 2004). Although its homologues regulate immunity pos-itively in Arabidopsis and tobacco (PUB17 and CMPG1,respectively), mutation in Spl11 in rice causes HR. There-fore spl11 mutants respond efficiently to blight rice patho-gen, which means that Spl11 is a negative regulator of celldeath and immunity (Zeng et al. 2004). Recently, Trujillo etal. (2008) found that the three homologous U-box E3s—PUB22, PUB23, PUB24—that negatively regulate PAMP-triggered immunity are required for suppression of PTI inArabidopsis. The triple mutant pub22/pub23/pub24 showedde-repression and decreased intensity in PTI, suggestingnegative regulation of PTI by PAMP recognition. The givenfindings suggest the regulatory roles of the ubiquitylationmechanism in plant immunity. Our findings show thatAtUBC2 is a negative regulator of plant cell death like SPL11.
In a previous study, the GmUbc2 gene has been shown toregulate gene expression levels for abiotic stress tolerance(Zhou et al. 2010). On the other hand, repression of flower-ing resulted from AtUbc2 regulation of Flc, Maf4, andMaf5gene expression levels (Xu et al. 2009a, b). Therefore, theUbc2 gene might regulate gene expression levels for bioticstress resistance. The induction of Rar1 and Sgt1 mRNAsmight be the main reason for enhanced resistance in silencedN. benthamiana leaves. Similar results were obtained inTaUbc2-silenced wheat leaves (Unver 2008).
Arabidopsis ubc mutant plants presented enhanced resis-tance upon virulent inoculation. The AtUbc2 complementedthe susceptibility in ubc2 mutant against virulent inocula-tion. Avirulent inoculation of the ubc2 35S: AtUbc2 line didnot result in dramatic suppression in resistance level asexpected (Fig. 6). AtUbc2 overexpressor lines produce in-significant changes in resistance level upon both avirulentand virulent inoculations compared to wild type lines(Figs. 1, 2, 3 and 4). Consequently, rather than knock-
- + + +
A
B
C
– UBC2 + UBC2 M
39,8 kDa SGT1
19 kDa UBC2
AntiSGT1 Ab
AntiFlag Ab
AntiFlag Ab
D
Fig. 11 a–d Interaction of UBC2with SGT1. aCo-immunoprecipitation(Co-Ip) of UBC2 with SGT1. Lanes: − pGR106 inoculated N. benthami-ana leaf protein extract immunoprecipitated with Anti-FLAG Ab, +pGR106-35S: Flag-NbUBC2 inoculated N. benthamiana leaf proteinextract immunoprecipitated with Anti-FLAG Ab. The gel was stainedwith Coomassie Blue. Eluates from Co-Ip of UBC2 and SGT1 were usedfor immuno blot assay. b Western blot analysis of FLAG tagged UBC2with FLAG antisera. c SGT1 immunoprecipitated with UBC2 was shownby using AntiSGT1 antibody. d Western blot analysis of UBC2 over-expression. Three individual overexpression experiments were con-firmed. Lanes: − pGR106 (without transgene) inoculated, + pGR106-35S: Flag-NbUBC2 inoculated N. benthamiana leaf protein extractsimmunoblotted with Anti-FLAG Ab
Plant Mol Biol Rep

down or knock-out, elevated AtUbc2 level does not influ-ence the pathogen response.
Plants are sessile organisms that are challenged continu-ally by biotic and abiotic stress factors. They can survivesuch unfavorable conditions using self-protecting systems.Salicylic acid (SA), jasmonic acid (JA), and ethylene (ET)are all used for response to biotic stress (Kunkel and Brooks2002). In contrast, abscisic acid (ABA) plays importantroles in both plant development and adaptation to abioticstresses (Xiong et al. 2002; de Torres Zabala et al. 2009). Onthe other hand, recent studies have shown an antagonisticinteraction between systemic acquired resistance and theABA-mediated abiotic stress response (Yasuda et al. 2008;de Torres Zabala et al. 2009). According to Zhou et al.(2010), over-expression of the GmUbc2 gene provides aresponse to salt and osmotic stresses and exogenous ABAtreatment tolerance by regulating abiotic stress-responsivegene expression. Since there is cross talk between abioticand biotic stress responses in plants, while overexpressionof Ubc2 responds to abiotic stress, knock-down or knock-out of Ubc2 might confer resistance against biotic stress.
In conclusion, the negative regulatory function of theAtUbc2 gene was observed by reverse and forward geneticapproaches in this study. The complementation study pro-vided gain-of-function and therefore loss of resistance inArabidopsis ubc2 mutants. The HR phenotype upon silenc-ing of the Ubc2 might result from upregulation ofresistance-related gene expression. The main factor explain-ing the regulatory role of UBC2 might be its interaction withSGT1.
Acknowledgments This study was partially supported by TurkishAcademy of Science (TUBA) for post doctoral research program(DSAP).
References
Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P,Stevenson DK, Zimmerman J, Barajas P, Cheuk R et al (2003)Genome-wide insertional mutagenesis of Arabidopsis thaliana.Science 301:653–657
Austin MJ, Muskett P, Kahn K, Feys BJ, Jones JD, Parker JE (2002)Regulatory role of SGT1 in early R gene-mediated plant defenses.Science 295:2077–2080
Azevedo C, Betsuyaku S, Peart J, Takahashi A, Noel L, SadanandomA, Casais C, Parker J, Shirasu K (2006) Role of SGT1 in resis-tance protein accumulation in plant immunity. EMBO J 25:2007–2016
Boter M, Amigues B, Peart J, Breuer C, Kadota Y, Casais C, Moore G,Kleanthous C, Ochsenbein F, Shirasu K, Guerois R (2007) Struc-tural and functional analysis of SGT1 reveals that its interactionwith HSP90 is required for the accumulation of Rx, an R proteininvolved in plant immunity. Plant Cell 21:1–14
Budak H, Su S, Ergen N (2006) Revealing constitutively expressedresistance genes in Agrostis species using PCR-based motif-directed RNA fingerprinting. Genet Res 88:165–175
Caldoa RA, Nettletonb D, Wise RP (2004) Interaction-dependent geneexpression in Mla-specified response to barley powdery mildew.Plant Cell 16:2514–2528
Cao Y, Dai Y, Cui S, Ma L (2008) Histone H2B Monoubiquitination inthe chromatin of Flowering Locus C regulates flowering time inArabidopsis. Plant Cell 20:2586–2602
Clough SJ, Bent AF (1998) Floral dip: a simplified method forAgrobacterium-mediated transformation of Arabidopsis thaliana.Plant J 16:735–743
DeTorres Zabala M, Bennet M, Truman WH, Grant MR (2009) An-tagonism between salicylic and abscisic acid reflects early host–pathogen conflict and moulds plant defense responses. Plant J59:375–386
Devoto A, Muskett PR, Shirasu K (2003) Role of ubiquitination in theregulation of plant defense against pathogens. Curr Opin PlantBiol 6:307–311
Dharmasiri N, Dharmasiri S, Estelle M (2005) The F-box protein TIR1is an auxin receptor. Nature 26:441–445
Dreher K, Callis J (2007) Ubiquitin, hormones and biotic stress inplants. Ann Bot 99:787–822
Durmaz E, Coruh C, Dinler G, Grusak MA, Peleg Z, Saranga Y, YaziciA, Ozturk L, Cakmak I, Budak H (2011) Expression and cellularlocalization of ZIP1 transporter under zinc deficiency in wildemmer wheat. Plant Mol Biol Rep 29:582–596
Ergen ZN, Budak H (2009) Sequencing over 13000 expressed se-quence tags from six subtractive cDNA libraries of wild andmodern wheats following slow drought stress. Plant Cell Environ32:220–236
Gagne JM, Downes, Barciszewska-Pacak K, Shiu S-H, Durski AM,Vierstra RD (2002) The F-Box subunit of the SCF E3 complex isencoded by a diverse superfamily of genes in Arabidopsis. ProcNatl Acad Sci USA 99:11519–11524
Guo FQ, Okamoto M, Crawford NM (2003) Identification of a plantnitric oxide synthase gene involved in hormonal signaling. Sci-ence 302:100–103
Hein I, Barciszewska-Pacak K, Hrubikova K, Williamson S, DinesenM, Soenderby IE, Sundar S, Shirasu K, Lacomme C (2005)VIGS-based functional characterization of genes associated topowdery mildew resistance in barley. Plant Physiol 138:2155–2164
Hershko A, Ciechanover A (1998) The ubiquitin system. Annu RevBiochem 67:425–479
Holt BF 3rd, Boyes DC, Ellerstrom M, Siefers N, Wiig A, Kauffman S,Grant MR, Dangl JL (2002) An evolutionarily conserved media-tor of plant disease resistance gene function is required for normalArabidopsis development. Dev Cell 2:807–817
Katagiri F, Thilmony R, He SY (2002) The Arabidopsis thaliana–Pseudomonas syringae interaction. In: Somerville CR, Meyero-witz EM (eds) The Arabidopsis book. American Society of PlantBiologists, Rockville
Kim HS, Delaney TP (2002) Arabidopsis SON1 is an F-box proteinthat regulates a novel induced defense response independent ofboth salicylic acid and systemic acquired resistance. Plant Cell14:1469–1482
Kitagawa K, Skowyra D, Elledge SJ, Harper JW, Hieter P (1999)SGT1 encodes an essential component of the yeast kinetochoreassembly pathway and a novel subunit of the SCF ubiquitin ligasecomplex. Mol Cell 4:21–33
Kraft E, Stone SL, Ma L, Su N, Gao Y, Lau OS, Deng XW, Callis J(2005) Genome analysis and functional characterization of the E2and RING-type E3 ligase ubiquitination enzymes of Arabidopsis.Plant Physiol 139:1597–1611
Kunkel BN, Brooks DM (2002) Cross talk between signaling pathwaysin pathogen defense. Curr Opin Plant Biol 5:325–331
Ma G, Ning G, Zhang W, Zhan J, Lv H, Bao M (2011) Overexpressionof Petunia SOC1-like gene FBP21 in tobacco promotes flowering
Plant Mol Biol Rep

without decreasing flower or fruit quantity. Plant Mol Biol Rep29:573–581
McConn M, Creelmann RA, Bell E, Mullet JE, Browse J (1997)Jasmonate is essential for insect defense. Proc Natl Acad SciUSA 94:5473–5477
Peart JR, Lu R, Sadanandom A, Malcuit I, Moffett P, Brice DC,Schauser L, Jaggard DAW, Xiao S, Coleman MJ, Dow M, JonesJDG, Shirasu K, Baulcombe DC (2002) Ubiquitin ligase-associated protein SGT1 is required for host and nonhost diseaseresistance in plants. Proc Natl Acad Sci USA 99:10865–10869
Pei D, Ma H, Wang W, Geng H, Li C (2011) Virus-induced genesilencing of a putative glutathione S-transferase gene compro-mised Ol-1-mediated resistance against powdery mildew in toma-to. Plant Mol Biol Rep 29:972–979
Scofield SR, Huang L, Brandt AS, Gill BS (2005) Development of avirus-induced gene-silencing system for hexaploid wheat and itsuse in functional analysis of the Lr21-mediated leaf rust resistancepathway. Plant Physiol 138:2165–2173
Sullivan ML, Vierstra R (1993) Formation of a stable adduct betweenubiquitin and the Arabidopsis ubiquitin-conjugating enzyme,AtUBC1. J Biol Chem 268:8777–8780
Sullivan JA, Shirasu K, Deng XW (2003) The diverse roles of ubiq-uitin and the 26S proteosome in the life of plants. Nat Rev Genet4:948–958
Takahashi A, Casais C, Ichimura K, Shirasu K (2003) HSP90 interactswith RAR1 and SGT1 and is essential for RPS2- mediated diseaseresistance in Arabidopsis. Proc Natl Acad Sci USA 100:11777–11782
Thomma BP, Eggermont K, Penninckx IA, Mauch-Mani B, VogelsangR, Cammue BP, Broekaert WF (1998) Separate jasmonate-dependent and salicylate-dependent defense-repsonse pathwaysin Arabidopsis are essential for resistance to distinct microbialpathogens. Proc Natl Acad Sci USA 95:15107–15111
Tian M, Huitema E, Da Cunha L, Torto-Alalibo T, Kamoun S (2004) AKazal-like extracellular serine protease inhibitor from Phytoph-thora infestans targets the tomato pathogenesis-related proteaseP69B. J Biol Chem 279:26370–26377
Tor M, Gordon P, Cuzick A, Eulgem T, Sinapidou E (2002) Arabidop-sis SGT1b is required for defense signaling conferred by severaldowny mildew resistance genes. Plant Cell 14:993–1003
Trujillo M, Ichimura K, Casais C, Shirasu K (2008) Negative regula-tion of PAMP-triggered immunity by an E3 ubiquitin ligase tripletin Arabidopsis. Curr Biol 18:1396–1401
Unver T (2008) Detection and characterization of plant genes involvedın various biotic and abiotic stress conditions using DDRTPCRand isolation of interacting proteins. In: Biotechnology. Ankara:Middle East Technical University
Unver T, Budak H (2009a) Virus-induced gene silencing, a post tran-scriptional gene silencing method. Int J Plant Genom 2009:1–8
Unver T, Budak H (2009b) Conserved microRNAs and their targets inmodel grass species Brachypodium distachyon. Planta 230:659–669
Unver T, Bakar M, Shearman RC, Budak H (2010) Genome-wideprofiling and analysis of Festuca arundinacea miRNAs and
transcriptomes in response to foliar glyphosate application. MolGenet Genom 283:397–413
Vierstra RD (2009) The ubiquitin-26S proteasome system at the nexusof plant biology. Nat Rev Mol Cell Biol 10:385–397
Xie DX, Feys BF, James S, Nieto-Rostro M, Turner JG (1998) COI1:an Arabidopsis gene required for jasmonate-regulated defense andfertility. Science 280:1091–1094
Xiong L, Schumaker KS, Zhu JK (2002) Cell signaling during cold,drought, and salt stress. Plant Cell 14(suppl):S165–S183
Xu L, Liu F, Lechner E, Genschik P, Crosby WL, Ma H, Peng W,Huang D, Xie D (2002) The SCFCOI1 ubiquitin-ligase complexesare required for jasmonate response in Arabidopsis. Plant Cell14:1919–1935
Xu L, Menard R, Fuchs J, Cognat V, Meyer D, Shen WH (2009a) TheE2 ubiquitin-conjugating enzymes, AtUBC1 and AtUBC2, playredundant roles and are involved in activation of FLC expressionand repression of flowering in Arabidopsis thaliana. Plant J57:279–288
Xu L, Menard R, Berr A (2009b) The E2 ubiquitin-conjugatingenzymes, AtUBC1 and AtUBC2, play redundant roles and areinvolved in activation of FLC expression and repression of flow-ering in Arabidopsis thaliana. Plant J 57:279–288
Yang CW, Gonzalez-Lamothe R, Ewan RA, Rowland O, Yoshioka H,Shenton M, Ye H, O’Donnell E, Jones JD, Sadanandom A (2006)The E3 ubiquitin ligase activity of Arabidopsis PLANT U-BOX17 and its functional tobacco homolog ACRE276 are re-quired for cell death and defense. Plant Cell 18:1084–1098
Yasuda M, Ishikawa A, Jikumaru Y, Seki M, Umezawa T, Asami T,Maruyama-Nakashita A, Kudo T, Shinozaki K, Yoshida S,Nakashita H (2008) Antagonistic interaction between systemic ac-quired resistance and the abscisic acid-mediated abiotic stress re-sponse in Arabidopsis. Plant Cell 20:1678–1692
Wei CF, Kvitko BH, Shimizu R, Crabill E, Alfano JR, Lin NC, MartinGB, Huang HC, Collmer A (2007) A Pseudomonas syringae pv.tomato DC3000 mutant lacking the type III effector HopQ1-1 isable to cause disease in the model plant Nicotiana benthamiana.Plant J 51:32–46
Yamamoto T, Kimura S, Mori Y, Oka M, Ishibashi T, Yanagawa Y,Nara T, Nakagawa H, Hashimoto J, Sakaguchi K (2004) Degra-dation of proliferating cell nuclear antigen by 26S proteasome inrice (Oryza sativa L.). Planta 218:640–646
Zeng LR, Qu S, Bordeos A, Yang C, Baraoidan M, Yan H, Xie Q,Nahm BH, Leung H, Wang GL (2004) Spotted leaf11, a negativeregulator of plant cell death and defense, encodes a U-box/arma-dillo repeat protein endowed with E3 ubiquitin ligase activity.Plant Cell 16:2795–2808
Zhou GA, Chang R, Qiu LJ (2010) Overexpression of soybean ubiquitin-conjugating enzyme gene GmUBC2 confers enhanced drought andsalt tolerance through modulating abiotic stress-responsive geneexpression in Arabidopsis. Plant Mol Biol 72:357–367
Zwirn P, Stary S, Luschnig C, Bachmair A (1997) Arabidopsis thalianaRAD6 homolog AtUBC2 complements UV sensitivity, but not N-end rule degradation deficiency, of Saccharomyces cerevisiaerad6 mutants. Curr Genet 32:309–314
Plant Mol Biol Rep
Copyright © 2022 FDOKUMEN




















