
Convergent Evolution of Escape from Hepaciviral Antagonism in Primates
Upload
independentCategory
view
4download
0
Vaccine 26 (2008) 3346–3351
Contents lists available at ScienceDirect
Vaccine
journa l homepage: www.e lsev ier .com/ locate /vacc ine
Improved HIV-1 specific T-cell responses by short-interval DNA tattooing
as compared to intramuscular immunization in non-human primatesBabs E. Verstrepena,1, Adriaan D. Binsb,1, Christine S. Rolliera, Petra Mooij a, Gerrit Koopmana,Neil C. Sheppardc, Quentin Sattentauc, Ralf Wagnerd, Hans Wolfd, Ton N.M. Schumacherb,Jonathan L. Heeneya,e, John B.A.G. Haanenb,∗
jswijkAmste
dom
deliimm
in nofold irhes
. A mthe nimat
ttooin
a Department of Virology, Biomedical Primate Research Centre, PO Box 3306, 2288 GJ Rib Division of Immunology, The Netherlands Cancer Institute, Plesmanlaan 121, 1066 CXc Sir William Dunn School of Pathology, University of Oxford, Oxford, United Kingdomd Institute of Medical Microbiology and Hygiene, Regensburg, Germanye Department of Veterinary Medicine, University of Cambridge, Cambridge, United King
a r t i c l e i n f o
Article history:Received 22 January 2008Received in revised form 18 March 2008Accepted 24 March 2008Available online 21 April 2008
Keywords:DNAHIVPrimate
a b s t r a c t
The new intradermal DNAbetween the encouragingthe poor results obtaineddemonstrate a 10- to 100-blood from DNA tattooedintramuscular (IM) routeas well as an increase infindings in non-human prare planned to validate ta
1. Introduction
With the elucidation of the genome sequences of a growingnumber of pathogens, plasmid DNA immunization is becomingan increasingly attractive strategy for the induction of protectiveimmunity [1]. Genes encoding immunologically important anti-gens can readily be inserted into DNA vaccines. These plasmidsare produced in large amounts at relatively low costs and arehighly stable. Furthermore, when DNA vaccines are used, immuneresponses are not hindered by vector specific immunity [2,3]. DNAimmunization has been demonstrated to induce protection froma variety of different infections, such as Murine Leukaemia virus,Cytomegalovirus (CMV), influenza and Lassa virus in small animalmodels [4–7]. Unfortunately, the high efficacy of DNA vaccines asobserved in murine studies was neither reproducible in non-humanprimates nor in human clinical trials [8]. This poor track record ofDNA vaccines in primates has been a major impediment for thewidespread clinical application of DNA vaccines.
∗ Corresponding author.E-mail address: [email protected] (J.B.A.G. Haanen).
1 These authors contributed equally to this work.
0264-410X/$ – see front matter © 2008 Elsevier Ltd. All rights reserved.doi:10.1016/j.vaccine.2008.03.091
, The Netherlandsrdam, The Netherlands
very technique, termed DNA tattooing might overcome the discrepancyunogenicity results obtained with DNA vaccines in murine studies andn-human primates and humans, the so called “simian barrier”. Here, we
ncrease in the magnitude of vaccine specific T-cell responses in peripheralus macaques, as compared to T-cell responses in animals immunized viaarked increase in the magnitude of the antigen specific T-cell responsesumber of animals responding to the immunogens was observed. These
es suggest that similar results may be observed in humans. Clinical trialsg as an optimal method of DNA vaccine delivery in humans.
© 2008 Elsevier Ltd. All rights reserved.
The factors responsible for the relatively weak immunogenicityof DNA vaccine candidates in humans and non-human primates
remain largely unknown. The poor immunogenicity of classicalintramuscular (IM) DNA immunization in primates has been sug-gested to be a consequence of the difference in uptake by residentantigen presenting cell (APC) populations between mice and non-human primates [9]. Differential Toll-like receptor (TLR) expressionpatterns and their function on the APCs of primates and mice maypossibly influence the immunogenicity of DNA vaccines. Further-more, in relation to body mass, the typical DNA doses used toimmunize large animals or humans have generally been 10–100times lower than those used in rodents. Collectively, these factorshave been referred to as the ‘simian barrier’ [10,11].Recently, a new intradermal DNA immunization method hasbeen developed and optimized [12]. This novel delivery strategy,termed “DNA tattooing”, uses a rapidly oscillating needle devicefor the intradermal delivery of DNA plasmids encoding vaccineantigens, involving thousands of epidermal punctures. Epidermalcells include both Langerhans cells and keratinocytes. Under nor-mal conditions Langerhans cells respond poorly to TLR9 ligandssuch as dsDNA. In mice however, keratinocytes stimulated withCpG produce cytokines that are able to increase APC function ofthe Langerhans cells [13]. This indirect enhancement of the antigen

accine
B.E. Verstrepen et al. / Vpresenting capacity of Langerhans cells might play a beneficial rolein the increased immunogenicity when administered intradermallyusing the DNA tattoo method.
DNA tattooing may have several advantages compared toparticle-mediated delivery methods, such as gene gun technolo-gies. First, DNA tattooing does not require carriers, such as thegold particles used in gene gun immunization. Second, no expen-sive equipment is required to deliver the immunogens as withelectroporation devices. Third, with this method of DNA admin-istration it would be possible to increase the amount of DNAlinear to total body surface across different species, potentiallyimproving the immunogenicity of DNA immunization in pri-mates.
Given the poor track record of DNA immunization in primateswe set out to evaluate the immunogenicity of this newly developedDNA delivery method in non-human primates.
Data revealed that intradermal DNA delivery by tattooing waswell tolerated and gave markedly superior antigen specific T-cellresponses compared to intramuscular DNA immunization. Basedon these data, clinical trials to validate these observations inhumans are warranted.
2. Materials and methods
2.1. Macaques
Captive bred, mature, outbred rhesus macaques (Macacamulatta) were housed at the Biomedical Primate Research Centre,Rijswijk, The Netherlands. This study was approved by the insti-tute’s animal ethical committee and was performed in accordancewith Dutch law and international guidelines for the use of animalsin research.
Before onset of the study, the animals were tested nega-tive for antibodies to SIV-1, Simian type D retrovirus, SimianT-cell lymphotropic virus. During the course of the study, theanimals were checked daily for appetite, general behaviour andstool consistency. Body weight and rectal temperature were mea-sured every time an animal was sedated for immunization orblood collection. During immunization periods the skin waschecked daily for adverse effects. At regular time points clini-cal chemistry and haematological parameters were monitored.Serum and peripheral blood mononuclear cells (PBMCs) were iso-lated from blood samples collected from sedated animals usingaseptic techniques (vacutainer, Becton Dickinson, Franklin Lakes,
USA).2.2. Immunogens
GMP lots of the EuroVacc HIV-1 vaccine candidates were pro-vided by the EuroVacc Foundation (Lausanne, Switzerland) andwere based on the HIV-1 clade C strain CN54 [14]. The HIV-1 gag,pol and nef expressing plasmid pORT1aCMVGPN and the HIV-1env expressing plasmid pORT1aCMVgp120 expressed HIV antigensunder the transcriptional control of a CMV promoter. GMP lotswere produced and quality controlled by Cobra Biomanufactur-ing Plc (Conner, UK) as previously described [15]. Immunizationwas performed at weeks 0 and 4. The study design is schematicallyrepresented in Fig. 1.
2.3. Immunization techniques
Intradermal delivery of the plasmid DNA vaccine was obtainedusing a tattoo device as previously described [12], with minoradaptations for use in non-human primates. In brief, the skinof each upper leg of the macaque was divided into three areas
26 (2008) 3346–3351 3347
Fig. 1. Schematic representation of the experimental setup. Arrows in the top panelindicate time points of immunization, either by DNA tattooing or by IM DNA injec-tion. The arrows in the bottom panel represent the ex vivo assays used to monitorthe induction of vaccine specific immune responses. At week 4, blood was collectedjust before DNA administration by either method.
of approximately 30 cm2 of normal skin, avoiding reproductivepigmented skin in female macaques. After atraumatic removalof hair, 650 �l of a 1 mg/ml DNA plasmid solution containingboth pORT1aCMVGPN and pORT1aCMVgp120, was applied toeach leg. The vaccine plasmids were delivered intradermally inthe non-vascularised skin layers using a standard rotary tat-too machine (Cold Skin, B&A Trading, Lijnden, The Netherlands)and a disposable 27-needle bar (Radical Clean Magnum11, Vil-leneuve Sous Dammartin, France) by a 2-min tattoo procedureper leg. As described earlier [12], antigen expression after a sin-gle intradermal tattoo immunization is short lived as comparedto expression after IM administration of a DNA vaccine. To com-pensate for this short duration of antigen expression and hencethe time in which T-cells are exposed to the encoded antigen,the DNA vaccine in the tattoo group was administered in a seriesof three applications at 3-day interval, each time using a differ-ent area of each leg. A cumulative volume of 4 ml of the vaccinewas applied in total per animal per time point or immunizationsequel.
The magnitude of DNA tattoo induced immune responses wascompared to those induced by IM injection of 4 ml of the sameGMP lot of DNA immunogens at weeks 0 and 4 in a group of 10macaques (Fig. 1). Monitoring of immune responses in the IM groupwas performed at the same time points post immunization andusing the exact same methods, peptide pools and reagents as inthe five animals that were immunized by the DNA tattoo tech-nique.
2.4. Peptides
15mer peptides with 11 amino acid overlap covering theentire sequence of the gagpolnef fusion product and gp120expression product from CN54 HIV-1 clade C were produced bySynpep, Dublin, California. To ensure a non-toxic DMSO con-centration during cellular assays, peptides, initially dissolved inhigh concentrations of DMSO and pooled together, were diluted1000-fold into RPMI shortly before use. Peptide pools were com-posed as earlier described [15,16], in brief gag-1 (aa Cg1–Cg240),gag-2 (aa Cg244–Cg486), pol-1 (aa Cgp485–Cp721), pol-2 (aaCp725–C817 and aa Cnp1017–Cp1161)), pol-3 (aa Cp1165–Cp1403),nef (aa Cn838–Cnp1030), env-1 (aa CN9–CN249), env-2 (aaCN253–CN485) and stored at −20 ◦C until use.
2.5. Cellular immune responses
To enumerate HIV-1 specific cytokine secreting cells IFN-�,IL-2 and IL-4 ELISPOT assays were performed at 4 weeks aftereach immunization according to manufacturer’s guidelines (U-Cytech, Utrecht, The Netherlands). In short, 4 × 106 freshly isolatedPBMCs ml−1 were cultured in a 24 well plate for 16 h in RPMI1640 supplemented with 5% heat inactivated pooled rhesus serum,
accine
3348 B.E. Verstrepen et al. / Vglutamine and penicillin–streptomycin in the presence of the indi-cated peptide pool at a final concentration of 2.5 �g ml−1 perpeptide. Medium was used as a negative control and ConcanavalinA (5 �g ml−1) stimulation was used as a positive control.
For detection of cytokine producing cells, non-adherent cellswere collected, washed and transferred to microtiter plates pre-coated with an antibody specific for the cytokine concerned. Cellswere subsequently incubated at 37 ◦C for 4 h (in case of IFN-� and IL-2) or 16 h (in case of IL-4). After lysing the cells and washing awaythe debris, cytokine detection was performed by adding a biotinlabelled detector antibody that was visualised using a gold stain-ing/silver enhancement technique. Spots were counted by Aelvis(Sanquin, Amsterdam, The Netherlands) and results are expressedas mean number of spot forming cells (SFCs) per million PBMCsfrom triplicate assays. Negative control values (mean medium aloneplus 2×S.D. from triplicate assays) were subtracted.
HIV specific immune responses were also characterized by thestaining of surface markers and intracellular cytokine staining (ICS)at weeks 2 and 6 following the first immunization, as describedpreviously [17]. In brief, after stimulating 1 × 106 freshly isolatedPBMCs for 16 h with either medium alone, 1.25 �g ml−1 of the indi-cated peptide pool or SEB, in the presence of aCD28 and aCD49dantibodies and Golgiplug (BD Biosciences, Franklin Lanes, USA)cells were stained with CD14TxR, CD20TxR, CD3PerCP, CD4PE-Cy7 andCD8APC-Cy7 specific antibodies. Subsequently, cells were permeabi-lized with Cytofix/Cytoperm (BD Biosciences, Franklin Lanes, USA),and levels of intracellular cytokines were determined by stain-ing with IFN-�APC, TNF-aFITC and IL-2PE specific antibodies. Flowcytometry was performed on a FACS Aria using Diva software (BD
Biosciences, Franklin Lanes, USA).2.6. Humoral responses
Serum samples were tested at weeks 0, 4 and 8 for HIV-1 enve-lope specific IgG antibodies by ELISA. Microtiterplates were coatedfor 16 h with gp140CN54 protein in 100 mM NaHCO3. Wells werethen blocked for 1 h with 2% non-fat milk before application ofserially diluted serum in 1% BSA PBS buffer. After 1 h of incubation,anti-human IgG-HRP conjugate at 1 mg ml−1 was added for anotherhour before addition of ultra-TMB ELISA development reagent. Thereaction was stopped by addition of 0.5 M H2SO4 and A450 nm wasdetermined. Between each step, plates were washed three to sixtimes with PBS.
2.7. Statistical techniques
The non-parametric Mann–Whitney test was used to comparethe median number of cytokine secreting cells between the tattooimmunized individuals and IM immunized group. A value of p < 0.05was considered significant.
Fig. 2. (a) Photographs of skin area treated by DNA tattooing of a representative animal,treatment.
26 (2008) 3346–3351
3. Results
3.1. Safety and tolerability of DNA tattoo immunization inprimates
No general adverse effects of the DNA tattoo applications wereobserved during either the first or second series of applications.In line with this laboratory tests of blood samples (haemoglobin,leukocyte counts, thrombocyte counts, creatinin, urea, and ASAT)remained within normal ranges during the experiment. To a vari-able extent, small subcutaneous haemorrhages appeared directlyafter intradermal DNA delivery (Fig. 2a and b) and in all animalsa minimal amount of yellow plasma transudate appeared on thetreated skin the day after tattooing. However, the plasma tran-sudate was short lived and disappeared within 2–5 days. Therewas no evidence that the animals had scratched or otherwisemanipulated the site of immunization and the method was welltolerated.
3.2. HIV-1 specific T-cell responses
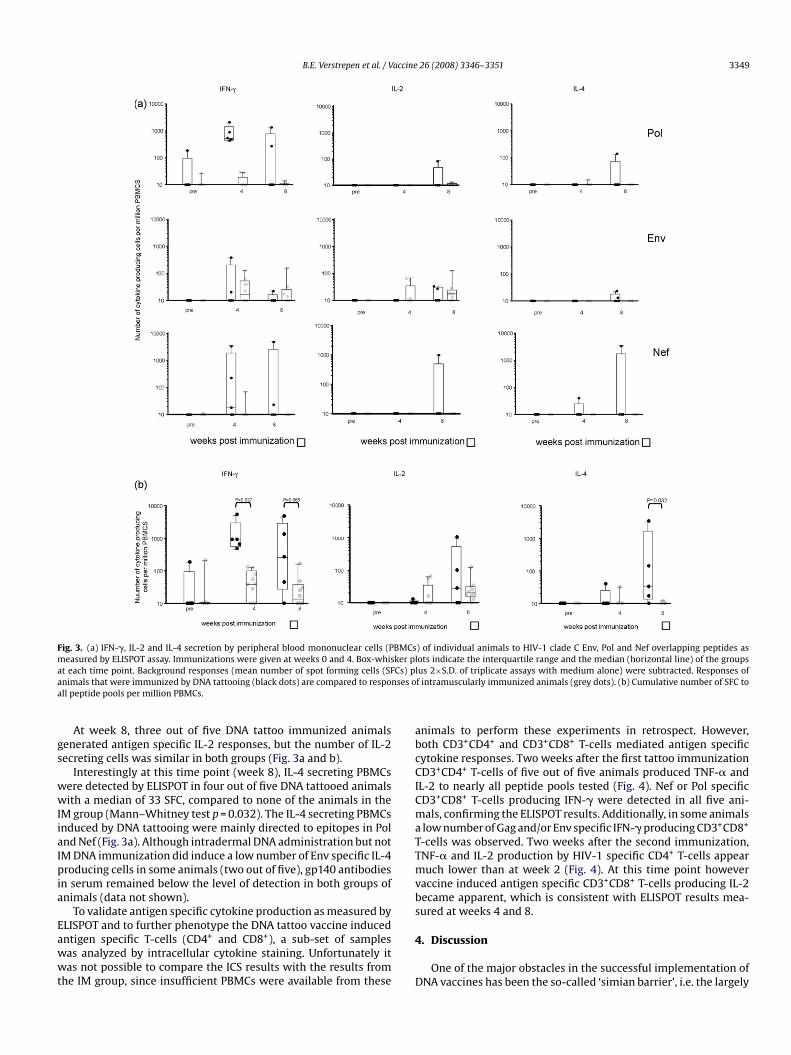
Remarkably, ELISPOT analysis at week 4 following the firstimmunization showed a strong Pol specific IFN-� response in allfive intradermally immunized animals, with a median value of 535SFC per million PBMCs, ranging from 433 to 2070 SFC per mil-lion (Fig. 3a). This compared very favourably to a median of 0(range 0–29) in the IM immunized animals (Mann–Whitney testp = 0.027). A similar observation was made when cells were stimu-lated with peptide pools derived from the Nef protein. Nef specific
IFN-� responses were observed in 3 out of 5 animals immunizedby DNA tattoo versus 1 out of 10 animals immunized by IM injec-tion. Little or no Gag responses were measured at this time point.The cumulative number of IFN-� secreting cells, i.e. the sum of thenumber of IFN-� secreting cells induced by different peptide pools,was approximately 24-fold higher in tattoo immunized animals ascompared to animals immunized via intramuscular DNA immu-nization (Mann–Whitney test p = 0.027, median 923 compared to39) (Fig. 3b). At this early time point, vaccine induced immuneresponses were largely restricted to IFN-�, almost no IL-2 or IL-4responses could be found in either of the groups.Similar to IFN-� responses measured at week 4, at week 8(4 weeks after the second immunization), the median cumula-tive number of antigen specific IFN-� secreting cells per millionPBMCs tended to be higher in the DNA tattooed group (268 SFCper million PBMCs, Fig. 3b) than in the IM immunization group(9 SFC), although this difference did not reach statistical sig-nificance (Mann–Whitney test p = 0.066). Again, IFN-� responseswere mainly directed to Nef and Pol peptide pools (Fig. 3a),little or no responses were measured directed to Gag peptidepools.
taken either directly after or (b) 3 days after intradermal vaccine delivery by tattoo
B.E. Verstrepen et al. / Vaccine 26 (2008) 3346–3351 3349
BMCsker pCs) p
nses o
Fig. 3. (a) IFN-�, IL-2 and IL-4 secretion by peripheral blood mononuclear cells (Pmeasured by ELISPOT assay. Immunizations were given at weeks 0 and 4. Box-whisat each time point. Background responses (mean number of spot forming cells (SFanimals that were immunized by DNA tattooing (black dots) are compared to respoall peptide pools per million PBMCs.
At week 8, three out of five DNA tattoo immunized animalsgenerated antigen specific IL-2 responses, but the number of IL-2secreting cells was similar in both groups (Fig. 3a and b).
Interestingly at this time point (week 8), IL-4 secreting PBMCswere detected by ELISPOT in four out of five DNA tattooed animalswith a median of 33 SFC, compared to none of the animals in theIM group (Mann–Whitney test p = 0.032). The IL-4 secreting PBMCsinduced by DNA tattooing were mainly directed to epitopes in Poland Nef (Fig. 3a). Although intradermal DNA administration but notIM DNA immunization did induce a low number of Env specific IL-4producing cells in some animals (two out of five), gp140 antibodiesin serum remained below the level of detection in both groups ofanimals (data not shown).
To validate antigen specific cytokine production as measured byELISPOT and to further phenotype the DNA tattoo vaccine inducedantigen specific T-cells (CD4+ and CD8+), a sub-set of sampleswas analyzed by intracellular cytokine staining. Unfortunately itwas not possible to compare the ICS results with the results fromthe IM group, since insufficient PBMCs were available from these
) of individual animals to HIV-1 clade C Env, Pol and Nef overlapping peptides aslots indicate the interquartile range and the median (horizontal line) of the groupslus 2×S.D. of triplicate assays with medium alone) were subtracted. Responses off intramuscularly immunized animals (grey dots). (b) Cumulative number of SFC to
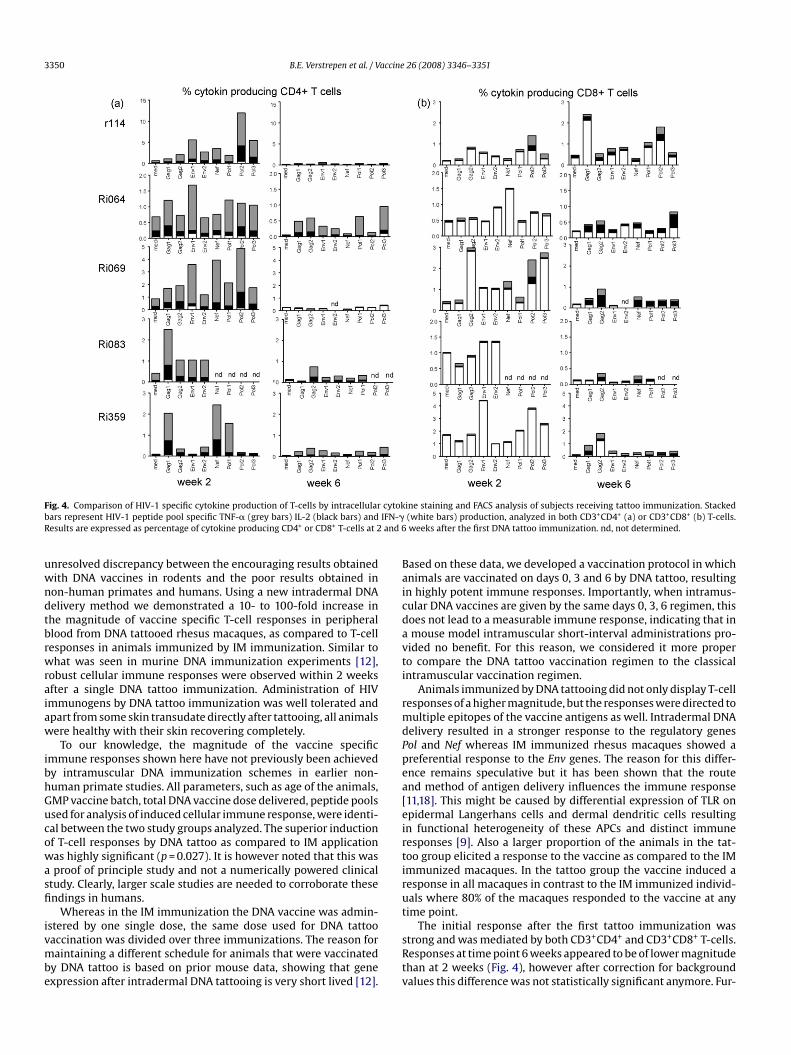
animals to perform these experiments in retrospect. However,both CD3+CD4+ and CD3+CD8+ T-cells mediated antigen specificcytokine responses. Two weeks after the first tattoo immunizationCD3+CD4+ T-cells of five out of five animals produced TNF-� andIL-2 to nearly all peptide pools tested (Fig. 4). Nef or Pol specificCD3+CD8+ T-cells producing IFN-� were detected in all five ani-mals, confirming the ELISPOT results. Additionally, in some animalsa low number of Gag and/or Env specific IFN-� producing CD3+CD8+
T-cells was observed. Two weeks after the second immunization,TNF-� and IL-2 production by HIV-1 specific CD4+ T-cells appearmuch lower than at week 2 (Fig. 4). At this time point howevervaccine induced antigen specific CD3+CD8+ T-cells producing IL-2became apparent, which is consistent with ELISPOT results mea-sured at weeks 4 and 8.
4. Discussion
One of the major obstacles in the successful implementation ofDNA vaccines has been the so-called ‘simian barrier’, i.e. the largely
3350 B.E. Verstrepen et al. / Vaccine 26 (2008) 3346–3351
cytokIFN-�and 6
Fig. 4. Comparison of HIV-1 specific cytokine production of T-cells by intracellularbars represent HIV-1 peptide pool specific TNF-� (grey bars) IL-2 (black bars) andResults are expressed as percentage of cytokine producing CD4+ or CD8+ T-cells at 2
unresolved discrepancy between the encouraging results obtainedwith DNA vaccines in rodents and the poor results obtained innon-human primates and humans. Using a new intradermal DNAdelivery method we demonstrated a 10- to 100-fold increase inthe magnitude of vaccine specific T-cell responses in peripheralblood from DNA tattooed rhesus macaques, as compared to T-cellresponses in animals immunized by IM immunization. Similar towhat was seen in murine DNA immunization experiments [12],robust cellular immune responses were observed within 2 weeksafter a single DNA tattoo immunization. Administration of HIV
immunogens by DNA tattoo immunization was well tolerated andapart from some skin transudate directly after tattooing, all animalswere healthy with their skin recovering completely.To our knowledge, the magnitude of the vaccine specificimmune responses shown here have not previously been achievedby intramuscular DNA immunization schemes in earlier non-human primate studies. All parameters, such as age of the animals,GMP vaccine batch, total DNA vaccine dose delivered, peptide poolsused for analysis of induced cellular immune response, were identi-cal between the two study groups analyzed. The superior inductionof T-cell responses by DNA tattoo as compared to IM applicationwas highly significant (p = 0.027). It is however noted that this wasa proof of principle study and not a numerically powered clinicalstudy. Clearly, larger scale studies are needed to corroborate thesefindings in humans.
Whereas in the IM immunization the DNA vaccine was admin-istered by one single dose, the same dose used for DNA tattoovaccination was divided over three immunizations. The reason formaintaining a different schedule for animals that were vaccinatedby DNA tattoo is based on prior mouse data, showing that geneexpression after intradermal DNA tattooing is very short lived [12].
ine staining and FACS analysis of subjects receiving tattoo immunization. Stacked(white bars) production, analyzed in both CD3+CD4+ (a) or CD3+CD8+ (b) T-cells.weeks after the first DNA tattoo immunization. nd, not determined.
Based on these data, we developed a vaccination protocol in whichanimals are vaccinated on days 0, 3 and 6 by DNA tattoo, resultingin highly potent immune responses. Importantly, when intramus-cular DNA vaccines are given by the same days 0, 3, 6 regimen, thisdoes not lead to a measurable immune response, indicating that ina mouse model intramuscular short-interval administrations pro-vided no benefit. For this reason, we considered it more properto compare the DNA tattoo vaccination regimen to the classicalintramuscular vaccination regimen.
Animals immunized by DNA tattooing did not only display T-cell
responses of a higher magnitude, but the responses were directed tomultiple epitopes of the vaccine antigens as well. Intradermal DNAdelivery resulted in a stronger response to the regulatory genesPol and Nef whereas IM immunized rhesus macaques showed apreferential response to the Env genes. The reason for this differ-ence remains speculative but it has been shown that the routeand method of antigen delivery influences the immune response[11,18]. This might be caused by differential expression of TLR onepidermal Langerhans cells and dermal dendritic cells resultingin functional heterogeneity of these APCs and distinct immuneresponses [9]. Also a larger proportion of the animals in the tat-too group elicited a response to the vaccine as compared to the IMimmunized macaques. In the tattoo group the vaccine induced aresponse in all macaques in contrast to the IM immunized individ-uals where 80% of the macaques responded to the vaccine at anytime point.The initial response after the first tattoo immunization wasstrong and was mediated by both CD3+CD4+ and CD3+CD8+ T-cells.Responses at time point 6 weeks appeared to be of lower magnitudethan at 2 weeks (Fig. 4), however after correction for backgroundvalues this difference was not statistically significant anymore. Fur-

accine
[
[
287–300.
B.E. Verstrepen et al. / V
thermore, in prior mouse experiments it appeared that when aprimary immune response induced by DNA vaccines is particu-larly strong, secondary responses against the same vaccine remainwithin the same range, possibly due to a more rapid clearance ofantigen-bearing DC and it seems reasonable to assume that thesame may apply here. During the course of the study both T-cellpopulations evolved to a more effector like phenotype. Initially,the HIV-1 specific CD3+CD4+ T-cells had a Th0 like profile withhigh TNF-� levels. At a later time point production of this cytokinewas much lower and a shift could be observed to IL-4 produc-tion, suggesting modulation of the CD3+CD4+ T-cell response. Thenaive, predominantly IFN-� producing, CD3+CD8+ T-cells, maturedto central memory or effector memory cells as demonstrated bythe elevated IL-2 production, indicative of increased proliferativecapacity of these cells. However, it must be stated that due to thelimited number of animals in this pilot study some apparent dif-ferences in responses between both groups were not statisticallydifferent. In addition, the current data suggest that the enhancedpotency of DNA tattoo is reflected in increased vaccine specific IL-4and IFN-� but not IL-2 production. Future studies should addressto what extent the enhanced potency of DNA vaccines delivered bythe tattoo method extends to other cytokines involved in pathogencontrol, the durability of the induced immune response, and thephenotype of the antigen specific cytokine producing cells as well asevaluation of the efficacy of the vaccine induced immune response.
In contrast to prior murine experiments, where immuniza-tion using the tattoo method induced production of antibodieswithin 2 weeks after the first immunization [12], IgG gp140 spe-cific antibodies were undetectable in the serum of the macaquesin this study. This difference may potentially be explained by thedifferent properties of the vaccines that were used in the twostudies. In earlier murine studies, a plasmid encoding a singleinfluenza derived hemaglutinin known for its antibody inducingability was used. In the current macaque study, two plasmidsencoding a total of four HIV proteins were used and, for tech-
nical reasons, only the induction of antibody responses againstthe Env protein was studied. Perhaps more importantly, the DNAvaccine used in this study was originally designed with the aimto induce HIV specific T-cell responses in a prime-boost regimenwith a pox vector encoding the same antigens. A more definitiveanswer regarding the capacity of DNA tattoo to prime vaccine spe-cific antibody responses in primates will therefore require furtherstudy.In conclusion, DNA vaccine delivery by DNA tattooing pro-vides evidence for a markedly improved immunogenicity of DNAvaccines in rhesus macaques when compared to the classicalIM route. However, this substantially enhanced potency of DNAtattoo comes at some cost, i.e. the requirement for three con-secutive applications. Both an increase of the magnitude of theantigen specific T-cell responses as well as an increase in thenumber of responders was readily established in the macaquesincluded in this pilot study. This and the fact that the tattoodevice is more practical for widespread use than the heliumrequiring instruments used for other intradermal immunizationmethods, suggests a potential clinical role for DNA tattoo-ing.
[
[
[
[
[
[
[
26 (2008) 3346–3351 3351
Acknowledgements
We thank the Animal Science Department for their excellentanimal care and support. We are indebted to the EuroVacc Foun-dation for generously providing GMP lots of their DNA vaccines aswell as peptides for T-cell analysis.
References
[1] Gurunathan S, Klinman DM, Seder RA. DNA vaccines: immunology, application,and optimization*. Annu Rev Immunol 2000;18:927–74.
[2] de Mare A, Lambeck AJ, Regts J, van Dam GM, Nijman HW, Snippe H, et al. Viralvector-based prime-boost immunization regimens: a possible involvement ofT-cell competition. Gene Ther 2007.
[3] Yang ZY, Wyatt LS, Kong WP, Moodie Z, Moss B, Nabel GJ. Overcomingimmunity to a viral vaccine by DNA priming before vector boosting. J Virol2003;77(1):799–803.
[4] Kong WP, Hood C, Yang ZY, Wei CJ, Xu L, Garcia-Sastre A, et al. Protective immu-nity to lethal challenge of the 1918 pandemic influenza virus by vaccination.Proc Natl Acad Sci USA 2006;103(43):15987–91.
[5] Rodriguez-Carreno MP, Nelson MS, Botten J, Smith-Nixon K, BuchmeierMJ, Whitton JL. Evaluating the immunogenicity and protective efficacy ofa DNA vaccine encoding Lassa virus nucleoprotein. Virology 2005;335(1):87–98.
[6] Rollman E, Mathy N, Brave A, Boberg A, Kjerrstrom A, van Wely C, et al. Evalua-tion of immunogenicity and efficacy of combined DNA and adjuvanted proteinvaccination in a human immunodeficiency virus type 1/murine leukemia viruspseudotype challenge model. Vaccine 2007;25(11):2145–54.
[7] Schleiss MR, Stroup G, Pogorzelski K, McGregor A. Protection againstcongenital cytomegalovirus (CMV) disease, conferred by a replication-disabled, bacterial artificial chromosome (BAC)-based DNA vaccine. Vaccine2006;24(37–39):6175–86.
[8] Rao SS, Gomez P, Mascola JR, Dang V, Krivulka GR, Yu F, et al. Comparative evalu-ation of three different intramuscular delivery methods for DNA immunizationin a nonhuman primate animal model. Vaccine 2006;24(3):367–73.
[9] Mitsui H, Watanabe T, Saeki H, Mori K, Fujita H, Tada Y, et al. Differential expres-sion and function of Toll-like receptors in Langerhans cells: comparison withsplenic dendritic cells. J Invest Dermatol 2004;122(1):95–102.
10] Johnston SA, Talaat AM, McGuire MJ. Genetic immunization: what’s in a name?Arch Med Res 2002;33(4):325–9.
11] McCluskie MJ, Brazolot Millan CL, Gramzinski RA, Robinson HL, Santoro JC,Fuller JT, et al. Route and method of delivery of DNA vaccine influenceimmune responses in mice and non-human primates. Mol Med 1999;5(5):
12] Bins AD, Jorritsma A, Wolkers MC, Hung CF, Wu TC, Schumacher TN, et al. Arapid and potent DNA vaccination strategy defined by in vivo monitoring ofantigen expression. Nat Med 2005;11(8):899–904.
13] Sugita K, Kabashima K, Atarashi K, Shimauchi T, Kobayashi M, Tokura Y. Innateimmunity mediated by epidermal keratinocytes promotes acquired immu-nity involving Langerhans cells and T cells in the skin. Clin Exp Immunol2007;147(1):176–83.
14] Su L, Graf M, Zhang Y, von Briesen H, Xing H, Kostler J, et al. Charac-terization of a virtually full-length human immunodeficiency virus type 1genome of a prevalent intersubtype (C/B’) recombinant strain in China. J Virol2000;74(23):11367–76.
15] Harari A, Bart PA, Stohr W, Tapia G, Garcia M, Medjitna-Rais E, et al. An HIV-1clade C DNA prime, NYVAC boost vaccine regimen induces reliable, polyfunc-tional, and long-lasting T cell responses. J Exp Med 2008;205:63–77.
16] Mooij P, Balla-Jhagjhoorsingh SS, Koopman G, Beenhakker N, van Haaften P,Baak I, et al. Differential CD4+ versus CD8+ T-cell responses elicited by differ-ent poxvirus-based human immunodeficiency virus type 1 vaccine candidatesprovide comparable efficacies in primates. J Virol 2008;82:2975–88.
17] Koopman G, Mortier D, Hofman S, Mathy N, Koutsoukos M, Ertl P,et al. Immune-response profiles induced by human immunodeficiencyvirus type 1 vaccine DNA, protein or mixed-modality immunization:increased protection from pathogenic simian-human immunodeficiencyvirus viraemia with protein/DNA combination. J Gen Virol 2008;89:540–53.
18] Bohm W, Mertens T, Schirmbeck R, Reimann J. Routes of plasmid DNA vacci-nation that prime murine humoral and cellular immune responses. Vaccine1998;16(9–10):949–54.
Copyright © 2022 FDOKUMEN




















