
Impacts of weather, shade cover and elevation on coffee leafminer Leucoptera coffeella (Lepidoptera:...
10
Impacts of weather, shade cover and elevation on coffee leafminer Leucoptera coffeella (Lepidoptera: Lyonetiidae) population dynamics and natural enemies J. Refugio Lomelí-Flores 1 , Juan F. Barrera 2 , Julio S. Bernal * Department of Entomology, MS 2475, Texas A&M University, College Station, TX 77843-2475, USA article info Article history: Received 10 December 2009 Received in revised form 10 March 2010 Accepted 14 March 2010 Keywords: Coffea arabica Ambient temperature Rainfall Predation Parasitism IPM abstract We assessed the influences of ambient temperature, rainfall, shade cover and elevation on seasonal abundance of coffee leafminer Leucoptera coffeella (Guérin-Ménèville) and its natural enemies in coffee farms in the Soconusco region of Chiapas, Mexico. Mined coffee leaves were most abundant during the rainy season (i.e. historical average rainfall >200 mm/mo, AprileNovember) compared to the dry season (<100 mm/mo, DecembereMarch), and at low (<600 m asl) relative to high (>900 m asl) elevations. The abundance of mined leaves increased with rainfall, and decreased with maximum daily temperatures. Coffee leafminer survivorship was highest during the dry season (>40%), when predation was lowest (<10%). Predation was the main source of coffee leafminer mortality, and was greatest during the rainy season (>25%) when coffee leafminer incidence was highest (>30% mined leaves per plant). None of the weather variables that were evaluated (viz. maximum and minimum temperatures, and rainfall) significantly impacted parasitism ratios. Shade cover moderated on-farm temperatures, by reducing maximum daily temperatures and any potential, direct impacts of rainfall on coffee leafminer, by providing partial shelter from rainfall, but did not significantly affect coffee leafminer incidence. In 48 h laboratory trials, coffee leafminer oviposition was highest at 28 C(w15 eggs/female), minimal at 25 C (w3 eggs) and nil at 20 C, and higher during night-time hours (>8 eggs/female/day) compared to day- time hours (<1 egg). Historical average temperatures were higher at low elevation (yearly average ca. 25 C; range ¼ 18.0e32.0 C) than at high elevation (ca. 21 C; 13.5e28.5 C), and we predicted that physical environmental conditions (i.e. night-time hours with temperatures > 20 C) were permissive of coffee leafminer oviposition during twice as many hours each year at low elevation (4060 h) compared to high elevation (2081 h). Overall, our results suggested that evident differences in the abundance of coffee leafminer between elevations may be due in considerable part to differences in ambient temperatures, particularly night-time temperatures, rather than rainfall, shade cover, or elevation per se. Ó 2010 Elsevier Ltd. All rights reserved. 1. Introduction Three sources of mortality are particularly relevant to leaf- miners: vertical sources, including natural enemies and host plant attributes, horizontal sources, such as intra and inter-specific competition, and abiotic sources, including meteorological factors, such as wind, storms, frosts, heavy rain, moisture and extreme temperatures (Auerbach et al., 1995). Though abiotic factors do not regulate populations, they may cause fluctuations in their size and limit their distribution (Hixon et al., 2002). For example, meteorological factors affect insect population dynamics via faster developmental rates with increasing temperature, and lower survival with heavy rain, extreme temperatures and low moisture (Hespenheide, 1991; Nestel et al., 1994; Tipping et al., 2005), and by negatively affecting individual reproductive behaviors (Wang et al., 1997; Saethre and Hofsvang, 2002). Similarly, meteorological factors may directly and indirectly influence the mortality ratios caused by insect natural enemies (Wang et al.,1997; Weisser et al., 1997; Stireman et al., 2005). Thus, meteorological factors may affect insect pest populations directly, through influences on reproduc- tive and mortality ratios, and indirectly, through influences on their natural enemies. Seasonal variation in weather variables, such as rainfall and temperature maxima and minima, may be the most important causes of dramatic changes in insect abundance, especially in temperate ecosystems (Cornell and Hawkins, 1995). Although the climate of tropical forest ecosystems is moderately constant (e.g., * Corresponding author. Tel.: þ1 (979) 862 8378. E-mail address: [email protected] (J.S. Bernal). 1 Present address: Instituto de Fitosanidad, Colegio de Postgraduados, Km 35.5 carretera México-Texcoco, Montecillo, Edo. de México 56230, Mexico. 2 El Colegio de la Frontera Sur, Carr. Antiguo Aeropuerto Km 2.5, Tapachula, Chiapas 30700, Mexico. Contents lists available at ScienceDirect Crop Protection journal homepage: www.elsevier.com/locate/cropro 0261-2194/$ e see front matter Ó 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.cropro.2010.03.007 Crop Protection 29 (2010) 1039e1048
Transcript of Impacts of weather, shade cover and elevation on coffee leafminer Leucoptera coffeella (Lepidoptera:...

lable at ScienceDirect
Crop Protection 29 (2010) 1039e1048
Contents lists avai
Crop Protection
journal homepage: www.elsevier .com/locate/cropro
Impacts of weather, shade cover and elevation on coffee leafminer Leucopteracoffeella (Lepidoptera: Lyonetiidae) population dynamics and natural enemies
J. Refugio Lomelí-Flores 1, Juan F. Barrera 2, Julio S. Bernal*
Department of Entomology, MS 2475, Texas A&M University, College Station, TX 77843-2475, USA
a r t i c l e i n f o
Article history:Received 10 December 2009Received in revised form10 March 2010Accepted 14 March 2010
Keywords:Coffea arabicaAmbient temperatureRainfallPredationParasitismIPM
* Corresponding author. Tel.: þ1 (979) 862 8378.E-mail address: [email protected] (J.S. Bernal)
1 Present address: Instituto de Fitosanidad, Colegiocarretera México-Texcoco, Montecillo, Edo. de México
2 El Colegio de la Frontera Sur, Carr. Antiguo AeChiapas 30700, Mexico.
0261-2194/$ e see front matter � 2010 Elsevier Ltd.doi:10.1016/j.cropro.2010.03.007
a b s t r a c t
We assessed the influences of ambient temperature, rainfall, shade cover and elevation on seasonalabundance of coffee leafminer Leucoptera coffeella (Guérin-Ménèville) and its natural enemies in coffeefarms in the Soconusco region of Chiapas, Mexico. Mined coffee leaves were most abundant during therainy season (i.e. historical average rainfall >200 mm/mo, AprileNovember) compared to the dry season(<100 mm/mo, DecembereMarch), and at low (<600 m asl) relative to high (>900 m asl) elevations. Theabundance of mined leaves increased with rainfall, and decreased with maximum daily temperatures.Coffee leafminer survivorship was highest during the dry season (>40%), when predation was lowest(<10%). Predation was the main source of coffee leafminer mortality, and was greatest during the rainyseason (>25%) when coffee leafminer incidence was highest (>30% mined leaves per plant). None of theweather variables that were evaluated (viz. maximum and minimum temperatures, and rainfall)significantly impacted parasitism ratios. Shade cover moderated on-farm temperatures, by reducingmaximum daily temperatures and any potential, direct impacts of rainfall on coffee leafminer, byproviding partial shelter from rainfall, but did not significantly affect coffee leafminer incidence. In 48 hlaboratory trials, coffee leafminer oviposition was highest at 28 �C (w15 eggs/female), minimal at 25 �C(w3 eggs) and nil at 20 �C, and higher during night-time hours (>8 eggs/female/day) compared to day-time hours (<1 egg). Historical average temperatures were higher at low elevation (yearly average ca.25 �C; range¼ 18.0e32.0 �C) than at high elevation (ca. 21 �C; 13.5e28.5 �C), and we predicted thatphysical environmental conditions (i.e. night-time hours with temperatures> 20 �C) were permissive ofcoffee leafminer oviposition during twice as many hours each year at low elevation (4060 h) compared tohigh elevation (2081 h). Overall, our results suggested that evident differences in the abundance of coffeeleafminer between elevations may be due in considerable part to differences in ambient temperatures,particularly night-time temperatures, rather than rainfall, shade cover, or elevation per se.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Three sources of mortality are particularly relevant to leaf-miners: vertical sources, including natural enemies and host plantattributes, horizontal sources, such as intra and inter-specificcompetition, and abiotic sources, including meteorological factors,such as wind, storms, frosts, heavy rain, moisture and extremetemperatures (Auerbach et al., 1995). Though abiotic factors do notregulate populations, they may cause fluctuations in their size andlimit their distribution (Hixon et al., 2002). For example,
.de Postgraduados, Km 35.556230, Mexico.
ropuerto Km 2.5, Tapachula,
All rights reserved.
meteorological factors affect insect population dynamics via fasterdevelopmental rates with increasing temperature, and lowersurvival with heavy rain, extreme temperatures and low moisture(Hespenheide, 1991; Nestel et al., 1994; Tipping et al., 2005), and bynegatively affecting individual reproductive behaviors (Wang et al.,1997; Saethre and Hofsvang, 2002). Similarly, meteorologicalfactors may directly and indirectly influence the mortality ratioscaused by insect natural enemies (Wang et al., 1997; Weisser et al.,1997; Stireman et al., 2005). Thus, meteorological factors may affectinsect pest populations directly, through influences on reproduc-tive andmortality ratios, and indirectly, through influences on theirnatural enemies.
Seasonal variation in weather variables, such as rainfall andtemperature maxima and minima, may be the most importantcauses of dramatic changes in insect abundance, especially intemperate ecosystems (Cornell and Hawkins, 1995). Although theclimate of tropical forest ecosystems is moderately constant (e.g.,

J.R. Lomelí-Flores et al. / Crop Protection 29 (2010) 1039e10481040
minimal variation in monthly average temperatures), marked localvariation in temperature and precipitation may occur due tochanges in topography, and such variation may affect insect pop-ulation dynamics, as recorded in coffee farms in Veracruz, Mexico(Nestel et al., 1994). Thus, understanding the impacts of localweather variables on insect population dynamics is important formanaging pests generally, including pests of tropical crops.
Coffee leafminer, Leucoptera coffeella (Guérin-Ménèville) (Lepi-doptera: Lyonetiidae), is a specialist herbivore known only fromCoffea species, and is an increasingly important pest of coffee in theNeotropics, including Mexico (CAB International, 2000; Ruiz et al.,2004). Previous studies have partially addressed the influences ofweather variables on coffee leafminer population dynamics, andshowed that the incidence of this pest was influenced by a suite ofenvironmental variables, such as rainfall, relative humidity, solarradiation and ambient temperature (Reis et al., 1976; Gravena, 1983;Nestel et al., 1994; Costa-Conceição et al., 2005; Teodoro et al., 2008,2009; Fernandes et al., 2009). For example, previous authors havesuggested that heavy rain was the main source of coffee leafminermortality, particularly during the rainy season (Villacorta, 1980;Campos et al., 1989; Pereira et al., 2007). Though, with the excep-tion of Pereira et al. (2007), such suggestions were based on studiesthat made comparisons among the abundances of mined leaves oncoffee plants, rather than on direct assessment of mortality sources.Using life tables, Pereira et al. (2007) found that during the rainyseason in Brazil most coffee leafminer mortality was due to physio-logical disturbances, rainfall and low egg viability, while in the dryseason it was mostly due to predaceous wasps, physiological distur-bances and parasitoids. In addition, major outbreaks of coffee leaf-miner in the Neotropics have been coincidental with reductions ofshadecoverand increasinguseofpesticidesoncoffee farms (Guharayetal., 2001;Monterreyetal., 2001;Fragosoetal., 2002;Carvalhoetal.,2005). Presumably, these factorsaffectedcoffee leafminerabundanceindirectly by reducing thepopulationsof itsnatural enemies (Fragosoet al., 2002), though this has not been confirmed.
Shade cover may influence micro-environmental conditionsprevalent within coffee farms (Velasco et al., 2001), and may indi-rectly influence the dynamics of arthropod populations withincoffee farms (Moguel and Toledo, 1999; Haggar et al., 2001). Forexample, coffee leafmineroutbreaks in Brazilwere sporadic throughca. 1970 when most coffee was grown under partial-shade condi-tions, but their intensity and frequency increased dramaticallycoincident with the introduction of highly productive coffee varie-ties that required full-sun conditions and greater use of agrochem-icals (Fragoso et al., 2002). Moreover, prior studies showed thatshade cover within coffee farmsmoderated the daily fluctuations intemperature, evapotranspiration and total radiation, and increasedsoil moisture and temperature (Barradas and Fanjul, 1986; Velascoet al., 2001). Accordingly, Haggar et al. (2001) recommended shadecover management to enhance production and facilitate pestmanagement in Nicaraguan coffee farms. Moguel and Toledo (1999)found that elimination of shade cover inMexican coffee farms led toa less stable physical environment and lower microorganismabundance and diversity. Thus, the increased abundance of coffeeleafminer in farmswhere shade coverwas reducedmaybe related tochanges in micro-environmental conditions, ensuing from theshade cover reduction, whichmay directly or indirectly affect coffeeleafminer mortality ratios (Guharay et al., 2001).
Priorobservations suggested that coffee leafminer abundancewashigher during the rainy season relative to the dry season, and in lowelevation relative to high elevation farms in the Soconusco area ofChiapas, Mexico (Lomelí-Flores et al., 2009). However, the effects oftemperature, rainfall and shade cover on the dynamics of coffeeleafminer, or its natural enemies, have not been assessed quantita-tively in southernMexico coffee farms. Thus, in the present study we
assessed the influences of ambient temperature, rainfall and shadecoveronseasonal survivorshipandabundanceof coffee leafminerandits natural enemies at two elevations in the Soconusco region ofChiapas, Mexico. Also, we assessed the oviposition rate of females inrelation to temperature and photoperiod. Based on our assessmentwe estimated the number of hours suitable for oviposition at the twoelevations to infer how elevational differences in ambient tempera-ture may affect coffee leafminer abundance in the field.
2. Materials and methods
Coffee leafminers were reared at El Colegio de la Frontera Sur(ECOSUR), Tapachula, Chiapas, Mexico on detached arabica-coffee(Coffea arabica L.) leaves, as described in Reis et al. (2000). A coffeeleafminer culture was initiated with field collected leafminers, andwas maintained in a room with constant temperature (26�1 �C),humidity close to saturation and a natural photoperiod(w13L:11D). Upon their emergence, coffee leafminer adults wereprovided with droplets of pure honey, and held as femaleemalepairs in 4 L containers for 24 h to allowmating before using them inthe experiments described below. All laboratory experiments wereconducted at ECOSUR in a roomwith the environmental conditionsdescribed above.
Field studies were conducted in three coffee farms, which rep-resented low (480e550 m above sea level) and high (w960 m asl)elevations, in Cacahoatán municipality, Chiapas, Mexico; the farmswere selected on the bases of accessibility, given terrain and roadconditions, and grower cooperation. Cacahoatán is located in theSoconusco region of Chiapas, a region comprising Pacific coastallowlands that reach the foothills of the Sierra Madre and withhigher moisture than interior regions. Low elevation coffee farmswere represented by El Encanto (14�5902100N; 92�0905500W; 480 masl) and La Gloria (15�0000700N; 92�0902400W, 550 m asl) farms, andhigh elevation farms by a farm in Ejido Alpujarras (15�0402400N;92�1001100W, 960 m asl). Comparisons between high and lowelevation farms relied on Alpujarras and El Encanto or La Gloriafarms, as available, and are noted below for each comparison. Themain coffee varieties in these farms were Catuaí, Bourbon andTypica, but all studies involved plants of the Catuaí variety. Noinsecticides were used on the farms during the study period.
Throughout this paper, “dry season” refers to the periodbetween December and March when historical average rainfall is<100 mm/mo, while “rainy season” refers to the AprileNovemberperiod when average rainfall is >200 mm/mo, unless notedotherwise. Also, all data recorded as proportions (i.e. mined leaves,coffee leafminer survival and death ratios) were arcsin Ox-trans-formed for statistical analyses and tested for normality prior toanalysis, though back-transformed (¼original) data are presentedin text, tables and figures. Nonparametric analyses were conductedas appropriate, and are noted. All statistical analyses were per-formed using SPSS 12.0 for Windows and/or Statistix 8.0 (SPSS,2005; Analytical Software, 2003), and are described in Zar (1996)and/or Sokal and Rohlf (1995).
2.1. Weather, daylength and oviposition hours at low and highelevation coffee farms
Historical weather data for the period 1961e1990 (ComisiónNacional del Agua, 2006), were used for rainfall and temperaturecomparisons between low and high elevation farms. Weather datafrom Cacahoatán (station #7018, 14�040N, 92�100W, 350 m abovesea level) were used to characterize low elevation farms, and fromEl Perú (station #7058, 15�040N, 92�130W, 800 m asl) to charac-terize high elevation farms; the distances between the Cacahoatánstation and El Encanto and La Gloria farms were w1.8 km and

J.R. Lomelí-Flores et al. / Crop Protection 29 (2010) 1039e1048 1041
w0.9 km, respectively, and between the El Perú station and Alpu-jarras farm was w5.7 km. One-sample t-tests were applied to thedifferences between Cacahoatán and El Perú monthly mean rain-falls and maximum and minimum temperatures to compareweather patterns between low and high elevation farms.
Additionally, historical weather datawere used to simulate dailyambient-temperature fluctuation at low and high elevation farms,following Tejeda-Martinez (1991). These data were then used tocalculate the monthly number of “oviposition hours,” viz. scoto-phase hours during which ambient temperature allows ovipositionby coffee leafminer females (see below, Section 2.5). The monthlynumbers of oviposition hours were compared between the low andhigh elevation farms using a Wilkinson signed rank test (Zar, 1996).
Finally, for some field studies (as noted below) daily maximumand minimum temperatures, daily rainfall and days with measur-able rainfall (>0.1 mm) were obtained from a research station(Centro de Investigación y Desarrollo de la Caña de Azúcar, TuxtlaChico, Chiapas, 14�570N, 92�100W, 366 m asl) and from rain gaugesand maximum/minimum thermometers placed within coffeefarms. The distance between the research station and El Encantofarmwas w5.4 km. Use and analyses of these data are described inthe corresponding sections below.
2.2. Effects of weather variables on coffee leafminer seasonalphenology and natural enemies
The influences of mean monthly minimum and maximumtemperatures, rainfall, days with measurable rainfall and evapora-tion on the seasonal phenology of coffee leafminer and its naturalenemies were assessed through monthly surveys at one low eleva-tion farm (El Encanto) from June 2003 to July 2004. Separately, theimpacts of mean monthly minimum and maximum temperaturesand rainfall on coffee leafminer incidence and mortality sourceswere compared between one low elevation farm (El Encanto) andthe high elevation farm through monthly surveys from March toJune 2005. For both studies,10 coffee plantswere randomly selected(exclusive of plants within two rows or 10 plants of farm margins,and separated from each other by at least 20 m or 2 rows) eachmonth and the proportions of mined leaves per plant were assessedusing nine branches per plant. In addition, 180 mined leaves werecollected monthly from both farms and examined in the laboratoryto assess coffee leafminer survival andmortality ratios. Thenumbersof mines, and coffee leafminer eggs, larvae, and pupae per each leafwere recorded, and stage-specific survival ratios and mortalityfactorswereassessed throughdissectionofminesunderadissectingmicroscope at 25� magnification. Previous studies (Lomelí-Floreset al., 2009) showed that larval and pupal coffee leafminermortality could be assignedunambiguously to one of three causes ofdeath: (i) parasitoid, (ii) predator, or (iii) factors other than parasit-oids or predators (FOPP).
Correlation analyses were conducted to find associationsbetween monthly weather variables and monthly proportions ofmined leaves per plant, and coffee leafminer mortality due toparasitoids, predators, or FOPP, or survivorship (i.e. those not killedby parasitoids, predators, or FOPP) per 180 leaf-sample. In addition,comparisons of proportions of mined leaves, and coffee leafminersurvival and mortality due to parasitism, predation, or FOPPbetween coffee plants at low and high elevation were made foreach month between March and June 2005, using t-tests.
2.3. Effects of shade cover on within-farm rainfall distribution,temperature, and coffee leafminer incidence and natural enemies
Field surveys and experiments were conducted at one lowelevation farm (La Gloria) during AprileJuly 2005. Thirty plots were
randomly selected within the farm, each plot consisting of fourplants from two contiguous rows forming a square; plots did notinclude plants within two rows or 10 plants of farm margins, andwere separated from each other by at least 20 m or 2 rows.
2.3.1. Effects of shade cover on rainfall distribution and temperatureCanopy cover, defined as the percentage of foliage cover
obscuring the open sky as viewed from the ground (Feldpauschet al., 2005), was used as an index of shade cover. Canopy coverwas assessed for each plot via analysis of digital photos with GapLight Analyzer 2.0 software (Frazer et al., 1999). Four photos weretaken at the center of each of the 30 plots at 1.5 m above the groundwith a digital camera equipped with a FC-E8 fisheye lens. Canopycover data used for analyses were the average of the four photostaken at each of the 30 plots. The 30 plots included shade coversbetween 14 and 78%, with a mean of w50%. Rainfall was measureddaily for 14 consecutive days within each of the 30 plots using four60 cm3 rain gauges at 1 m above the ground; one pair of rain gaugeswas attached to each of two wooden rods, and the rods weredistant w1.5 m from each other. Shelter from direct rainfall wasassumed to be directly related to shade cover, so the ratio of thevariance to the mean of rainfall among rain gauges per each plot(¼variance/mean ratio) was used as a rainfall dispersion index (i.e.a proxy for shelter from direct rainfall) and for statistical analyses. Amaximum/minimum thermometer was placed in the canopy of oneof the four plants in each of the 30 plots, and maximum andminimum temperatures for the preceding 24 h period were recor-ded daily between 9:00 and 10:00 h. Shade cover was assessed atthe beginning of the study (June 22, 2005), and temperatures andrainfall were recorded daily from 22 June to 5 July 2005. Simplecorrelation analyses were conducted to detect any relationshipbetween shade cover and individual weather variables (dailymaximum and minimum temperatures, and rainfall dispersionindex).
2.3.2. Effects of shade cover on coffee leafminer incidenceEight branches (branch base at ca. 1.5 m), each with four pairs of
mine-free leaves, were labeled on each of the four plants in the 30plots. Thus, a total of 64 mine-free leaves per plant, equivalent to1920 leaves on 30 plants, were labeled at the beginning of the study(April 2005). The proportion of mined leaves per plant was evalu-ated one month later following a period of low rainfall (<100 mm/month) (May 2005). Mined leaves included those having at leastone fresh mine, which was defined as any mine with a living larvaor with a dead larva and green plant tissue within. A second set ofobservations was made in the same way four months later (August2005), following a period of high rainfall (>400 mm/month).Simple correlation analyses were conducted to detect any rela-tionships between shade cover and proportion of mined leavesunder low (May 2005) or high rainfall (August 2005).
2.3.3. Effects of shade cover on coffee leafminer survival andmortality due to natural enemies
Coffee leafminer larval stage survival and mortality ratios wereassessed by establishing coffee leafminer cohorts on one plant ineach of the 30 plots. One branch without mined leaves on eachplant was selected and the apical three pairs of leaves of eachbranch were enclosed in a 30 cm nylon mesh sleeve. Two pairs ofcoffee leafminer adults (24 h old) were enclosed in each sleeve for48 h; leafminer adults were removed after this time and thenumber of eggs laid was recorded. Sleeves were left on the branchfor one week to avoid egg predation and allow larvae to initiatemines, and then removed to expose leafminer larvae to naturalenemies during their development (w1 week). Leaves were har-vested one week later and taken to the laboratory to assess larval

J.R. Lomelí-Flores et al. / Crop Protection 29 (2010) 1039e10481042
survival ratios and mortality due to predation, parasitism, or FOPP,under a dissecting microscope (see above Section 2.2). Concur-rently, coffee leafminer egg and pupa survival ratios, and mortalitydue to predation, parasitism, or FOPP were assessed by exposingcoffee leafminers from the laboratory colony to natural enemies oncoffee plants. This was done by selecting one plant from each of the30 plots and stapling five leaves with coffee leafminer eggs orpupae (<24 h old), taken from the laboratory culture, to leaves onthe selected plants. Each of the five leaves held 10e30 coffee leaf-miner eggs or 1e2 pupae, and was collected after 1 week ofexposure to assess survival and mortality ratios of coffee leafminereggs and pupae in the laboratory.
Datawere subjected to simple correlation analyses to detect anyrelationships between shade cover and coffee leafminer survivaland stage-specific mortality (¼proportions) due to parasitism,predation, or FOPP.
2.4. Rainfall and elevation effects on coffee leafminer survival andmortality factors
Stage-specific survival and mortality ratios of coffee leafminereggs, larvae and pupae were assessed during periods of low rainfall(<100 mm/mo, April 27eMay 12, 2005) and high rainfall(>400 mm/mo, June 27eJuly 12, 2005) at one low elevation farm(La Gloria) and the high elevation farm.
Average shade cover in the low elevation farm was 67%(range¼ 38e87%), and average maximum and minimum temper-atures, total rainfall, and days with measurable rainfall during theperiod of low rainfall were 32.3 �C, 18.0 �C, 80 mm and 4 d,respectively. Corresponding data during the period of high rainfallwere 29.0 �C, 20.5 �C, 413 mm and 13 d.
Average shade cover in the high elevation farm was 76%(range¼ 45e91%). Average maximum and minimum temperaturesfor the low rainfall period were 28.5 �C and 13.5 �C, and for the highrainfall period were 25.0 �C and 15.7 �C; total rainfall and days withmeasurable rainfall were not reported for the high elevation farmbecause of calibration problems with the rainfall gauge.
Tencoffeeplantswere randomlyselectedateachelevationontwodates, April 27 and June 27, and one coffee leafminer cohort wasestablished on one branch of each plant using the methodologydescribed above (see Section 2.3). Sleeves were removed frombranchesoneweek later (May4, or July 3) and thenumberof eggsperbranch was recorded. At the same time, 10 coffee leaves with coffeeleafminer eggs (4e9/leaf) or pupae (1e2/leaf) were exposed bystapling them to coffee leaves on the selected plants. At least 50 eggs(range¼ 10e30/leaf) and 10 pupae (range¼ 1e2/leaf) were thusplacedoneachplant.All coffee leafminereggs, larvaeandpupaewereexposed to mortality sources during one week, after which the cor-responding leaveswere taken to the laboratory and examined undera microscope to determine egg, larva and pupa survivorship, andmortality ratios due to parasitism, predation, or FOPP, per each plant.
The influence of elevation (low elevation¼ La Gloria; high ele-vation¼Alpujarras) and rainfall (low, <100 mm/mo, AprileMay;high, >400 mm/mo, JuneeJuly) on coffee leafminer survival ratio,and the contributions to coffee leafminer mortality of parasitoids,predators and FOPP at each of the 10 plants were compared usingthe ScheirereRayeHare extension of the KruskaleWallis test,a two-way nonparametric analysis of variance (Sokal and Rohlf,1995). Survival and mortality ratio data per each of the 10 coffeeplants were used for analyses.
2.5. Temperature effects on coffee leafminer oviposition rate
Coffee leafminer 48 h oviposition rates were assessed at threeconstant temperatures, 20, 25 and 28 �C (all� 1 �C), under laboratory
conditions. One laboratory-reared coffee leafminermaleefemale pair(24 h old) was placed inside a 0.5 L plastic cage with one coffee leaf;a droplet of pure honey was provided as a source of food for theleafminers. The coffee leafminer female was allowed to lay eggs for48 h, after which both leafminers were removed and the number ofeggs laid per leaf was recorded. Thirty-six pairs of coffee leafmineradults were monitored at each temperature. All leaves were froma single coffee plant and from the second pair on a branch, and wereheld individually on a polyurethane sponge saturated with waterinside the plastic cage. KruskaleWallis one-way nonparametricanalysis of variance, followed by Tukey’s tests, was used to compareoviposition rates among temperatures.
2.6. Circadian cycle of coffee leafminer oviposition
The circadian cycle of coffee leafminer oviposition was assessedunder laboratory conditions at constant temperature (26�1 �C)using 24 h old coffee leafminers (n¼ 24). Single coffee leafminerfemaleemale pairs were held inside 0.5 L plastic cages with a coffeeleaf following the methodology described above (see Section 2.5).Leaves were replaced and the number of eggs laid per each pairrecordedevery4 hbetweenAugust 3 andAugust 7, 2005. Leaveswerereplaced at 03:00, 07:00, 11:00, 15:00, 19:00 and 23:00 h. Solar radi-ationwas recorded at thosehours in aweather station (Vantage Pro�WeatherStation,Davis Instruments,Davis,California,USA)at ECOSUR(14�5301200N, 92�1701100W, 114 m asl). Two daily periods were estab-lished for comparisons: (i) scotophase, fromdusk throughdawn,withobservations at 19:00, 23:00 and 3:00 h, and (ii) photophase, frommorning through afternoon, with observations at 7:00, 11:00 and15:00 h. During the experimental period, morning twilight began atw6:30 h (sunrisey 6:53 h) and evening twilight ended atw17:59 h(sunsety 19:38 h). The relationshipbetween thenumberof eggs laidper female during each 4 h period and solar radiation was assessedusing the Spearman Ranks test. In addition, aWilcoxon rank sum testwas used to compare coffee leafminer oviposition (¼eggs/female)between the scotophase and the photophase.
3. Results
3.1. Weather, daylength and oviposition hours at low and highelevation coffee farms
Historical weather data (1961e1990) showed that monthlyrainfall amounts were similar at the low (Fig. 1a) and high (Fig. 1b)elevation weather stations (P¼ 0.794). In contrast, both monthlymaximum and minimum temperatures were higher [by5.7� 0.2 �C (mean� SE) and 1.6� 0.2 �C, respectively] at lowelevation relative to high elevation (P<<0.001 in each case)(Fig. 1a, b).
Oviposition hours, hereafter defined precisely as scotophasehours with ambient temperature> 20 �C (see below), variedbetween w268 h and 378 h per month at low elevation (¼4060 htotal during the year) (Fig. 1c), and between w20 h and 316 h permonth at high elevation (¼2081 h total during the year) (W¼�78,P¼ 0.004) (Fig. 1d). Overall, more than 90% of scotophase hours permonth between March and December were conducive to oviposi-tion at low elevation, while they exceeded 90% only in May at highelevation and were below 50% per month from October to March(data not shown).
3.2. Effects of weather variables on coffee leafminer seasonalphenology and natural enemies
The monthly proportion of mined leaves per plant at a lowelevation farm (El Encanto) was significantly correlated with

Jan F be Mar pA
rMay J nu Jul uA
gSep Oc
tNvo De
c0
100
200
300
400
M o n t h
yl
ht
no
mn
ae
M
om
xm
m(
ll
af
ni
ar
1-
)
0
200
400
600
800
Mean monthly temperature (oC)
16
18
20
22
24
26
28
30
32
34
36
200
400
600
800
0
a b
J na Feb MarAprMay Jun Ju
lAug eS
p OctNovDce
sr
uo
hn
oi
ti
so
pi
vO
)o
mr
ep
(
0
100
200
300
400
c d
sr
uo
hn
oi
ti
so
pi
vO
)o
mr
ep
(
yl
ht
no
mn
ae
M
om
xm
m(
ll
af
ni
ar
1-
)
Fig. 1. Mean monthly rainfall (empty columns) and maximum (filled circles) and minimum (empty circles) temperatures, and oviposition hours (¼night-time hours withtemperature >20 �C; see text) (filled columns) at (a, c) low (Cacahoatán, 14�590N, 92�100W, 350 m asl) and (b, d) high (El Perú, 15�060N, 92�160W, 800 m asl) elevations in theSoconusco region of Chiapas, Mexico. Both monthly maximum (t¼ 29.86, 11 df, P� 0.001) and minimum (t¼ 9.43, 11 df, P� 0.001) temperatures differ significantly between (a)and (b), while monthly rainfall amounts do not (t¼ 0.27, 11 df, P¼ 0.794). Monthly oviposition hours differ significantly between (c) and (d) (W¼�78, P¼ 0.004). Graphs derivedfrom historical data, 1961e1990 (Comisión Nacional del Agua, 2006).
J.R. Lomelí-Flores et al. / Crop Protection 29 (2010) 1039e1048 1043
maximum and minimum temperatures, monthly rainfall and dayswith measurable rainfall (Table 1). The proportion mined leaveswas >0.20 during the rainy season (JuneNov 2003 and MayeJul2004) and the highest proportion (w0.49) was in JuneeJuly 2004(Fig. 2a). The proportion of mined leaves during the early part of thedry season was <0.10 (Dec 2003eFeb 2004) and was w0.24 at theend of the dry season (Mar 2004) (Fig. 2a).
Coffee leafminer survival ratio was positively correlated withmaximum temperature (r¼ 0.67; P¼ 0.01), but not with minimumtemperature, monthly rainfall, nor days with measurable rainfall(Table 1). Predation, parasitism and mortality due to FOPP werenot significantly correlated with any climatic variables that wereevaluated (Table 1). Coffee leafminer survival ratio was lowest(<0.2) during the rainy season (JuleNov 2003) and early in the dryseason (Jan 2004), when monthly maximum temperatures were33e34 �C; and, was highest (>0.35) late in the dry season andearly in the rainy season (FebeJune 2004) (Fig. 3). The highestsurvival ratio (w0.65) occurred during the dry season, in March2004 (Fig. 3). Predation ratios were highest (>0.3) during the rainyseason and early dry season (Sep 2003eJan 2004 and JuneJul2004, respectively), and were lowest (<0.1) from the end of thedry season to the beginning of the rainy season (MareMay 2004)(Fig. 3).
Table 1Correlations between selected weather variables and proportions of leaves mined by cosurvival ratio and mortality factors (death due to predation, parasitism, or factors otheCacahoatán, Chiapas, Mexico, June 2003 to July 2004 (FOPP ¼ factors other than predati
Weather variables Proportion ofminedleavesa
Coffee leafminer su
Survival
r P r P
Maximum temperature �0.54 0.04 0.67 0.01Minimum temperature 0.83 <0.01 �0.01 0.97Monthly rainfall 0.76 <0.01 �0.43 0.13Days with measurable rainfall 0.85 <0.01 �0.33 0.24
a Estimated from 10 plants/month (9 branches/plant).b Estimated from 180 leaves/month.
The proportion of mined leaves during MarcheJune 2005 washigher at the low versus high elevation farm, except in April whenthe difference was not significant (Fig. 4). The proportion of minedleaves between March and June 2005 was not significantly corre-lated with maximum or minimum temperatures or monthly rain-fall at either low or high elevations (Table 2).
Coffee leafminer survival ratio did not differ, except in June,between low and high elevation coffee farms (Fig. 5a), and wasnegatively correlated with minimum temperature and rainfall, andpositively correlated with maximum temperature at the lowelevation farm, but only positively correlated with maximumtemperature at the high elevation farm (Table 2). Predation ratiosincreased between March and June, but did not differ betweenelevations (Fig. 5b). Predation ratios were positively correlatedwithminimum temperatures and rainfall, and negatively correlatedwith maximum temperatures at the low elevation farm, but onlynegatively correlated with maximum temperature at the highelevation farm (Table 2). Parasitism ratio was higher at the highelevation farm, but significantly so only in April and May (Fig. 5c),and was not correlated with maximum or minimum temperaturesat either elevation (Table 2). Coffee leafminer mortality due to FOPPwas higher at low elevation in May, but higher at high elevation inJune (Fig. 5d), and was positively correlated with minimum
ffee leafminer Leucoptera coffeella (Lepidoptera: Lyonetiidae) and coffee leafminerr than predation or parasitism), at a low elevation farm w480 m asl (El Encanto),on or parasitism; significant P-values indicated in bold type).
rvival and contributions of predation, parasitism, and FOPP to total mortalityb
Predation Parasitism FOPP
r P r P r P
�0.51 0.06 �0.25 0.38 �0.38 0.18�0.18 0.53 �0.13 0.67 0.34 0.230.26 0.36 0.06 0.83 0.34 0.230.21 0.46 0.01 0.99 0.30 0.30

Month
Jun JulAugSep Oc
tNovDec Jan Feb Ma
rAprMayJun.Jul.
)m
m(
ll
af
ni
ar
yl
ht
no
M
0
100
200
300
400
500
600
700
800
900
1000
(e
ru
ta
re
pm
et
yl
ht
no
mn
ae
Mo
)C
10
15
20
25
30
35
40
de
ni
mn
oi
tr
op
or
P
+(
se
va
el
)E
S
0.0
0.1
0.2
0.3
0.4
0.5
0.6a
b2003 / 2004
Fig. 2. (a) Proportion (�SE) leaves mined by coffee leafminer, Leucoptera coffeella(Lepidoptera: Lyonetiidae), (b) rainfall (mm) (columns) and maximum (empty circles)and minimum (filled circles) monthly temperatures (�C) at a low elevation coffee farm(El Encanto, 480 m asl), Cacahoatán, Chiapas, Mexico, 2003e2004.
Month
March April May June
+(
se
va
el
de
ni
mn
oi
tr
op
or
P)
ES
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18Low elevation
High elevation
t = 4.64*
t = 0.25
t = 4.13*
t = 5.61*
Fig. 4. Proportion leaves mined (�SE) by coffee leafminer Leucoptera coffeella (Lepi-doptera: Lyonetiidae) at low (El Encanto, 480 m asl) and high (Alpujarras, 960 m asl)elevation coffee farms, Cacahoatán, Chiapas, Mexico, in 2005. Inset statistics corre-spond to t-tests with df¼ 18; * indicates P< 0.01.
J.R. Lomelí-Flores et al. / Crop Protection 29 (2010) 1039e10481044
temperature at the low elevation farm, but not at the high elevationfarm (Table 2).
3.3. Effects of shade cover on within-farm rainfall distribution,temperature and coffee leafminer incidence and natural enemies
3.3.1. Effects of shade cover on rainfall distribution and temperatureShade cover was negatively correlated with within-farm
maximum temperature (r¼�0.38; P¼ 0.04) and rainfall dispersionindex (r¼�0.40; P¼ 0.03). Shade cover was not significantlycorrelated with within-farm total rainfall (r¼�0.28; P¼ 0.13) orminimum temperature (r¼�0.26; P¼ 0.17).
Month
uJ n uJ l AugSep O
tc NovDec aJ n Feb Ma
rAprMayJun. uJ l.
no
it
ro
po
rp
yl
ht
no
M
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7Dry season
CLM survival
Parasitism
Predation
FOPP
2003 / 2004
Fig. 3. Coffee leafminer, Leucoptera coffeella (Lepidoptera: Lyonetiidae), survival ratioand mortality ratios due to predation, parasitism, or factors other than parasitism orpredation (FOPP) at a low elevation coffee farm (El Encanto, 480 m asl), Cacahoatán,Chiapas, Mexico, in 2003e2004. Shading indicates dry season (actual monthlyrainfall< 100 mm).
3.3.2. Effects of shade cover on coffee leafminer incidenceDuring the high rainfall period, the proportion of mined leaves
was correlated with minimum temperature (P� 0.03, r��0.20),though not with maximum temperature (P¼ 0.85, r��0.02).Moreover, during periods of low or high rainfall the proportion ofmined leaves was not significantly correlated with shade cover(P� 0.12, r� 0.14), maximum temperature (P� 0.12, r��0.14), norrainfall (P� 0.10, r� 0.15).
3.3.3. Effects of shade cover on coffee leafminer survival andmortality due to natural enemies
Shade cover was significantly correlated with ratios of larvalpredation (r¼�0.48; P¼ 0.02) and death by FOPP (r¼ 0.59;P< 0.01) and pupa survival (r¼ 0.40; P¼ 0.03); maximumtemperaturewas significantly correlated with larval predation ratio(r¼ 0.59; P¼ 0.01); and, rainfall dispersion index with larvalpredation (r¼�0.42; P¼ 0.05) and pupa survival ratio (r¼ 0.45;
Table 2Correlations between selectedweather variables and the proportion of leavesminedby coffee leafminer Leucoptera coffeella (Lepidoptera: Lyonetiidae) and coffee leaf-miner survival and mortality factors (predation, parasitism, factors others thanpredation and parasitism) at low and high elevation coffee farms, Cacahoatán,Chiapas, Mexico, March to June 2005 (FOPP ¼ factors other than predation orparasitism; significant P-values indicated in bold type).
Elevation/variable Maximumtemperature
Minimumtemperature
Rainfall(mm)a
R P r P R P
Low elevation (El Encanto, 480 m asl)Mined leaves/mo (proportion)a �0.22 0.18 �0.12 0.47 0.12 0.48Survival ratiob 0.47 0.002 �0.61 <0.001 �0.67 <0.001Predationb �0.59 <0.001 0.52 0.001 0.67 <0.001Parasitismb �0.15 0.36 �0.03 0.88 0.10 0.53FOPPb 0.14 0.38 0.36 0.02 0.12 0.48
High elevation (Alpujarras, 960 m asl)c
Mined leaves/mo (proportion)a 0.28 0.09 0.18 0.28 e e
Survival ratiob 0.75 <0.001 �0.27 0.10 e e
Predationb �0.55 <0.001 0.24 0.13 e e
Parasitismb �0.10 0.56 �0.19 0.25 e e
FOPPb �0.30 0.07 0.29 0.07 e e
a Estimated from 10 plants/month (9 branches/plant).b Estimated from 180 leaves/month.c Rainfall was not recorded at the high elevation farm due to calibration problems
with the rain gauge.

+(
la
vi
vr
us
re
ni
mf
ae
le
ef
fo
C)
ES
0.0
0.1
0.2
0.3
0.4
0.5
+(
noi
ta
de
rP
)E
S
0.0
0.1
0.2
0.3
0.4
0.5Low elevation
High elevation
M o n t h
March April May June
+(
ms
it
is
ar
aP
)E
S
0.0
0.1
0.2
0.3
0.4
0.5
March April May June
+(
PP
OF
)E
S
0.0
0.1
0.2
0.3
0.4
0.5
a b
c d
t = 0.75
t = 1.29
t = 0.23
t = 4.26**
t = 0.07
t = 0.86
t = 1.61
t = 1.16
t = 1.81t = 3.08**
t = 4.00**t = 1.67
t = 0.55
t = 0.84
t = 2.64*
t = 2.19*
Fig. 5. Coffee leafminer, Leucoptera coffeella (Lepidoptera: Lyonetiidae), (a) larva survival ratios and larval mortality ratios due to (b) predation, (c) parasitism and (d) factors otherthan parasitism or predation (FOPP) at low (El Encanto, 480 m asl) and high (Alpujarras, 960 m asl) elevation coffee farms, Cacahoatán, Chiapas, Mexico, in 2005. Inset statisticscorrespond to t-tests with df¼ 18; * indicates P< 0.01 and ** indicates P< 0.001.
J.R. Lomelí-Flores et al. / Crop Protection 29 (2010) 1039e1048 1045
P¼ 0.01) (Table 3). Egg and larva survival, egg and pupa predation,larval parasitism, and egg and pupa mortality by FOPP were notsignificantly correlated with any of the weather variables that wereevaluated (Table 3).
3.4. Rainfall and elevation effects on coffee leafminer survival andmortality factors
Egg survival ratios were higher at low versus high elevation(P< 0.01), and were similar under low and high rainfall (P¼ 0.38)(Table 4). Survival ratios for coffee leafminer larvae and pupae didnot differ significantly between elevations (P� 0.16), though theywere higher under low versus high rainfall (P< 0.01) (Table 4).
Egg and larva predation ratios did not differ significantlybetween elevations (P� 0.48,) or rainfall levels (P� 0.57). Predationcontributed >90% of total egg mortality and >8% of larval totalmortality at both elevations (Table 4). Pupal predation ratiosdiffered significantly between elevations (P¼ 0.01) and rainfalllevels (P< 0.01) (Table 4). Parasitism of eggs or pupae was notdetected. Larval parasitism ratios at low elevation contributed>40% of total larval mortality, and were ca. two-fold greater than athigh elevation (P¼ 0.03), but they did not differ significantlybetween rainfall levels (P¼ 0.41) (Table 4).
The contributions of FOPP to total mortality of coffee leafminereggs did not differ significantly between elevations (P¼ 0.99) orrainfall levels (P¼ 0.82) and did not exceed 6% at both elevationsand both rainfall levels. Larval mortality ratios due to FOPP weregreater at high elevation versus low elevation (P< 0.01) and underhigh versus low rainfall levels at low elevation (P¼ 0.02), though athigh elevation they did not differ significantly between rainfall
levels (Table 4). Pupa mortality ratios due to FOPP did not differsignificantly between rainfall levels (P¼ 0.11), but were higher athigh elevation and low rainfall versus low elevation and highrainfall (P¼ 0.05) (Table 4).
3.5. Temperature effects on coffee leafminer oviposition rate
Coffee leafminer oviposition during a 48 h period differedsignificantly among temperatures. Coffee leafminer females did notlay eggs at 20 �C and laid significantly fewer eggs at 25 �C (3.0� 2.1eggs/female) than at 28 �C (15.2� 5.9 eggs/female) (c2¼ 35.82;df¼ 2; P< 0.01). Thus, temperatures above 20 �C were consideredconducive to coffee leafminer oviposition.
3.6. Circadian cycle of coffee leafminer oviposition
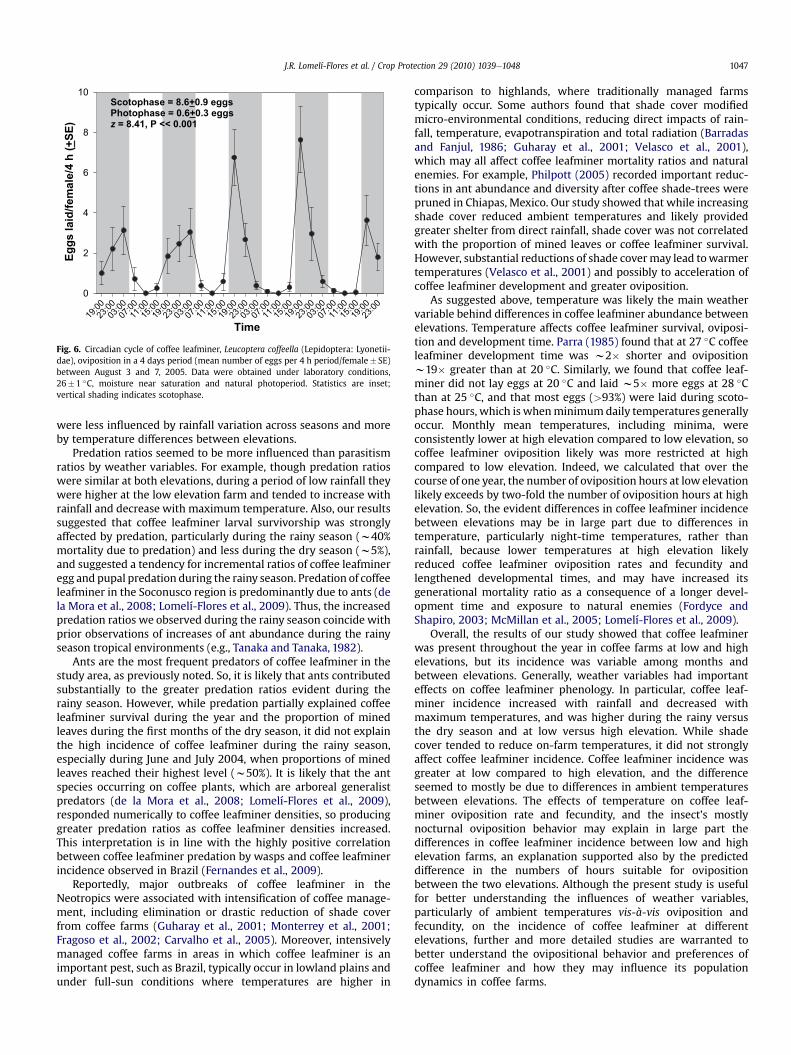
Average hourly solar radiation from sunset to sunrise was0.8� 0.2 W/m2 (range¼ 0.0e5.7 W/m2) and from sunrise to sunsetwas 451.7� 30.2 W/m2 (range¼ 55.9e882.9 W/m2). Solar radiation(4 h average) had a significantly negative influence on coffee leaf-miner 4 h fecundity (r2¼�0.60; P< 0.001; n¼ 144). Cumulativefecundity after 4 days was w14-fold higher during the scotophase(8.6� 0.9 eggs/female/day, range¼ 0e51 eggs) relative to thephotophase (0.6� 0.3 eggs/female/day range¼ 0e19 eggs)(z¼ 8.41, P� 0.001) (Fig. 6).
4. Discussion
We found that coffee leafminer was present throughout the yearin coffee farms at both low and high elevation, but its incidencewas

Table 3Correlations between selected weather variables and coffee leafminer Leucopteracoffeella (Lepidoptera: Lyonetiidae) survival and mortality ratios at a low elevationcoffee farm (La Gloria, 550 m asl), Cacahoatán, Chiapas, Mexico, June 22 to July 5,2005 (FOPP ¼ factors other than predation or parasitism; significant P-values indi-cated in bold type).
Stage/variablea Weather variables
Shade Maximumtemperature
Minimumtemperature
Totalrainfall
Rainfalldistributionb
EGGSurvival R 0.02 �0.11 0.17 0.32 0.14
P 0.90 0.56 0.37 0.08 0.45Predation R �0.04 0.08 �0.17 �0.31 �0.16
P 0.82 0.66 0.37 0.10 0.40FOPP R 0.17 0.31 0.13 0.13 0.30
P 0.36 0.10 0.51 0.48 0.11
LARVASurvival R �0.28 0.19 0.13 0.10 �0.05
P 0.20 0.38 0.56 0.66 0.82Predation R �0.48 0.56 0.11 0.15 �0.42
P 0.02 0.01 0.63 0.49 0.05Parasitism R �0.12 �0.22 0.10 0.17 �0.04
P 0.58 0.32 0.64 0.44 0.87FOPP R 0.59 �0.35 �0.17 �0.29 0.31
P 0.003 0.10 0.44 0.17 0.15
PUPASurvival R 0.40 �0.19 0.004 �0.10 0.45
P 0.03 0.31 0.99 0.59 0.01Predation R �0.23 0.18 0.09 0.11 �0.32
P 0.23 0.36 0.66 0.57 0.09FOPP R �0.04 �0.11 �0.14 �0.02 0.05
P 0.82 0.57 0.47 0.90 0.79
a Coffee leafminer survival and mortality ratios due to predation, parasitism, orFOPP were assessed in the field using coffee leafminer cohorts from a laboratorycolony.
b Coefficient of variation of daily rainfall among 4 rain gauges (¼variance/mean).
Table 4Influence of elevation (low or high) and rainfall level (low or high) on coffee leaf-miner, Leucoptera coffeella (Lepidoptera: Lyonetiidae), survival ratio and mortalitydue to predation, parasitism, or factors other than parasitism and predation(¼FOPP). Standard errors are shown in parentheses; different letters followingparentheses indicate statistically significant differences within columns; significantP-values indicated in bold type.
Elevation/rainfalla Coffee leafminer Contribution of natural enemies andother factors to total mortality
survival Predation Parasitism FOPP
(a) EGGLow/low 0.61 (0.16)a 0.94 (0.01)a 0 0.06 (0.08)aLow/high 0.42 (0.16)a 0.97 (0.06)a 0 0.03 (0.06)aHigh/low 0.13 (0.11)b 0.99 (0.02)a 0 0.003 (0.02)aHigh/high 0.13 (0.11)b 0.99 (0.03)a 0 0.009 (0.03)aElevation Hb 15.54 0.001 N/A 0.001
Pb <0.01 0.99 0.99Rainfall Hb 0.77 0.05 N/A 0.05
Pb 0.38 0.82 0.82
(b) LARVALow/low 0.39 (0.16)a 0.26 (0.15)a 0.44 (0.17)a 0.31 (0.15(c)Low/high 0.09 (0.10)b 0.08 (0.09)a 0.40 (0.16)a 0.53 (0.17)bHigh/low 0.35 (0.16)a 0.10 (0.10)a 0.19 (0.13)b 0.71 (0.15)abHigh/high 0.05 (0.07)b 0.13 (0.12)a 0.14 (0.12)b 0.73 (0.15)aElevation Hb 1.98 0.51 4.68 9.39
Pb 0.16 0.48 0.03 <0.01Rainfall Hb 15.36 0.33 0.68 5.27
Pb <0.01 0.57 0.41 0.02
(c) PUPALow/low 0.26 (0.15)a 0.91 (0.1)ab 0 0.09 (0.1)abLow/high 0.04 (0.07)b 0.97 (0.06)a 0 0.03 (0.06)bHigh/low 0.31 (0.15)a 0.65 (0.16)b 0 0.35 (0.16)aHigh/high 0.18 (0.13)ab 0.89 (0.11)ab 0 0.11 (0.11)abElevation Hb 3.28 6.76 N/A 3.89
Pb 0.35 0.01 0.05Rainfall Hb 8.58 8.58 N/A 2.52
Pb <0.01 <0.01 0.11
a The low elevation farm was represented by La Gloria (550 m asl), and the highelevation farm by Alpujarras (960 m asl), both in Cacahoatán, Chiapas, Mexico. Lowrainfall was <100 mm (April 27eMay 12, 2005) and high rainfall was >400 mm(June 27eJuly 12, 2005).
b Statistics shown are from the ScheirereRayeHare extension of the Krus-kaleWallis test.
J.R. Lomelí-Flores et al. / Crop Protection 29 (2010) 1039e10481046
variable among months and between elevations, and correlatedwith changes in temperature and rainfall. In contrast, we found thatcoffee leafminer mortality sources were minimally influenced bytemperature and rainfall, and that the influences of shade cover oncoffee leafminer and its mortality sources were relatively subtle,compared to the influences of weather variables.
The coffee leafminer population dynamics observed in our studywere similar to those reported for Colombia (Flórez and Hernández,1981), but dissimilar to those reported for Ecuador (Teodoro et al.,2009), Brazil (Villacorta, 1980; Tuelher et al., 2003; Pereira et al.,2007; Fernandes et al., 2009), Guatemala (Campos et al., 1989)and Veracruz state, Mexico (Nestel et al., 1994). In particular, weobserved higher proportions of mined leaves during the rainyseason, relative to the dry season, and these proportions decreasedwith maximum temperatures and with decreasing rainfall,whereas coffee leafminer larval survival decreased with increasingrainfall and with decreasing maximum temperatures. Also, theproportion of mined leaves was significantly greater in the lowversus high elevation farm.
Some factors that have been suggested for explaining differencesin coffee leafminer incidence between seasons and elevations includerainfall and ambient temperatures, and their effects on the coffeeplant andoncoffee leafminerand itsnatural enemies (Reis et al.,1976;Villacorta, 1980; Flórez and Hernández, 1981; Campos et al., 1989;Nestel et al., 1994; Tuelher et al., 2003; Guerreiro-Filho, 2006;Pereira et al., 2007; Teodoro et al., 2008, 2009; Lomelí-Flores et al.,2009). Coffee plants undergo significant changes in their phenology,biochemistry and physiology with changing weather conditions(Blanco et al., 2003), and those changes may affect their interactionswith both herbivores and natural enemies.Many plant-insect studiesin tropical environments showed that herbivores prefer young leaves
over mature leaves, thus higher numbers of herbivores are expectedwhenplantsproducenewfoliage,which in the tropics typicallyoccursduring the rainy season (Williams-Linera andHerrera, 2003). Thoughcoffee plants produce abundant new foliage during the rainy season(Blanco et al., 2003), coffee leafminer does not preferentially ovipositin young leaves over older leaves (Guerreiro-Filho, 2006). Thus,production of new foliage by coffee plants did not explain the higherincidence of coffee leafminer observed during the rainy season in thepresent study.
Weather variables, particularly ambient temperature and rain-fall, have been suggested to be among the primary factors affectingthe abundance of coffee leafminer natural enemies, as noted above.However, we did not find consistently significant effects of weathervariables on parasitism ratios. For example, the highest and lowestparasitism ratios we observed, 13% in March 2004 and 38% inJanuary 2004, respectively, both occurred in dry-season months,and parasitism ratios were not correlated with any of the weathervariables we evaluated. Nonetheless, whereas our field trials withlaboratory-reared coffee leafminer cohorts showed that rainfall didnot affect larval parasitism ratios, those ratios were consistentlygreater at low versus high elevation. Historical data showed thatrainfall was highly variable during the year, but did not differbetween elevations, while temperature was relatively constantduring the year, but consistently and significantly higher at lowversus high elevation. Thus, mortality ratios due to parasitism likely
Time
91: 002:3000:300 00:70
00:1100:5100:91 32: 000:300 :7000 00:11
00:5100:91 32: 000:3000:700 00:11
00:5100:9100:32 30: 000:700 :1100 00:51
00:9100:32
+(
h4
/e
la
me
f/
di
al
sg
gE
)E
S
0
2
4
6
8
10Scotophase = 8.6+0.9 eggs
Photophase = 0.6+0.3 eggs
z = 8.41, P << 0.001
Fig. 6. Circadian cycle of coffee leafminer, Leucoptera coffeella (Lepidoptera: Lyonetii-dae), oviposition in a 4 days period (mean number of eggs per 4 h period/female� SE)between August 3 and 7, 2005. Data were obtained under laboratory conditions,26� 1 �C, moisture near saturation and natural photoperiod. Statistics are inset;vertical shading indicates scotophase.
J.R. Lomelí-Flores et al. / Crop Protection 29 (2010) 1039e1048 1047
were less influenced by rainfall variation across seasons and moreby temperature differences between elevations.
Predation ratios seemed to be more influenced than parasitismratios by weather variables. For example, though predation ratioswere similar at both elevations, during a period of low rainfall theywere higher at the low elevation farm and tended to increase withrainfall and decrease with maximum temperature. Also, our resultssuggested that coffee leafminer larval survivorship was stronglyaffected by predation, particularly during the rainy season (w40%mortality due to predation) and less during the dry season (w5%),and suggested a tendency for incremental ratios of coffee leafmineregg and pupal predation during the rainy season. Predation of coffeeleafminer in the Soconusco region is predominantly due to ants (dela Mora et al., 2008; Lomelí-Flores et al., 2009). Thus, the increasedpredation ratios we observed during the rainy season coincide withprior observations of increases of ant abundance during the rainyseason tropical environments (e.g., Tanaka and Tanaka, 1982).
Ants are the most frequent predators of coffee leafminer in thestudy area, as previously noted. So, it is likely that ants contributedsubstantially to the greater predation ratios evident during therainy season. However, while predation partially explained coffeeleafminer survival during the year and the proportion of minedleaves during the first months of the dry season, it did not explainthe high incidence of coffee leafminer during the rainy season,especially during June and July 2004, when proportions of minedleaves reached their highest level (w50%). It is likely that the antspecies occurring on coffee plants, which are arboreal generalistpredators (de la Mora et al., 2008; Lomelí-Flores et al., 2009),responded numerically to coffee leafminer densities, so producinggreater predation ratios as coffee leafminer densities increased.This interpretation is in line with the highly positive correlationbetween coffee leafminer predation by wasps and coffee leafminerincidence observed in Brazil (Fernandes et al., 2009).
Reportedly, major outbreaks of coffee leafminer in theNeotropics were associated with intensification of coffee manage-ment, including elimination or drastic reduction of shade coverfrom coffee farms (Guharay et al., 2001; Monterrey et al., 2001;Fragoso et al., 2002; Carvalho et al., 2005). Moreover, intensivelymanaged coffee farms in areas in which coffee leafminer is animportant pest, such as Brazil, typically occur in lowland plains andunder full-sun conditions where temperatures are higher in
comparison to highlands, where traditionally managed farmstypically occur. Some authors found that shade cover modifiedmicro-environmental conditions, reducing direct impacts of rain-fall, temperature, evapotranspiration and total radiation (Barradasand Fanjul, 1986; Guharay et al., 2001; Velasco et al., 2001),which may all affect coffee leafminer mortality ratios and naturalenemies. For example, Philpott (2005) recorded important reduc-tions in ant abundance and diversity after coffee shade-trees werepruned in Chiapas, Mexico. Our study showed that while increasingshade cover reduced ambient temperatures and likely providedgreater shelter from direct rainfall, shade cover was not correlatedwith the proportion of mined leaves or coffee leafminer survival.However, substantial reductions of shade covermay lead towarmertemperatures (Velasco et al., 2001) and possibly to acceleration ofcoffee leafminer development and greater oviposition.
As suggested above, temperature was likely the main weathervariable behind differences in coffee leafminer abundance betweenelevations. Temperature affects coffee leafminer survival, oviposi-tion and development time. Parra (1985) found that at 27 �C coffeeleafminer development time was w2� shorter and ovipositionw19� greater than at 20 �C. Similarly, we found that coffee leaf-miner did not lay eggs at 20 �C and laid w5� more eggs at 28 �Cthan at 25 �C, and that most eggs (>93%) were laid during scoto-phase hours, which is whenminimumdaily temperatures generallyoccur. Monthly mean temperatures, including minima, wereconsistently lower at high elevation compared to low elevation, socoffee leafminer oviposition likely was more restricted at highcompared to low elevation. Indeed, we calculated that over thecourse of one year, the number of oviposition hours at low elevationlikely exceeds by two-fold the number of oviposition hours at highelevation. So, the evident differences in coffee leafminer incidencebetween elevations may be in large part due to differences intemperature, particularly night-time temperatures, rather thanrainfall, because lower temperatures at high elevation likelyreduced coffee leafminer oviposition rates and fecundity andlengthened developmental times, and may have increased itsgenerational mortality ratio as a consequence of a longer devel-opment time and exposure to natural enemies (Fordyce andShapiro, 2003; McMillan et al., 2005; Lomelí-Flores et al., 2009).
Overall, the results of our study showed that coffee leafminerwas present throughout the year in coffee farms at low and highelevations, but its incidence was variable among months andbetween elevations. Generally, weather variables had importanteffects on coffee leafminer phenology. In particular, coffee leaf-miner incidence increased with rainfall and decreased withmaximum temperatures, and was higher during the rainy versusthe dry season and at low versus high elevation. While shadecover tended to reduce on-farm temperatures, it did not stronglyaffect coffee leafminer incidence. Coffee leafminer incidence wasgreater at low compared to high elevation, and the differenceseemed to mostly be due to differences in ambient temperaturesbetween elevations. The effects of temperature on coffee leaf-miner oviposition rate and fecundity, and the insect’s mostlynocturnal oviposition behavior may explain in large part thedifferences in coffee leafminer incidence between low and highelevation farms, an explanation supported also by the predicteddifference in the numbers of hours suitable for ovipositionbetween the two elevations. Although the present study is usefulfor better understanding the influences of weather variables,particularly of ambient temperatures vis-à-vis oviposition andfecundity, on the incidence of coffee leafminer at differentelevations, further and more detailed studies are warranted tobetter understand the ovipositional behavior and preferences ofcoffee leafminer and how they may influence its populationdynamics in coffee farms.

J.R. Lomelí-Flores et al. / Crop Protection 29 (2010) 1039e10481048
Acknowledgments
We thank Messrs. Joel Herrera M., Enrique López P., RománMontes, Guillermo López G. and Heber García (all at ECOSUR) fortheir help, and Federico Ochoa (Finca Alpujarras), Alejandro Álvarez(Finca La Gloria), Humberto Prada (Finca El Encanto) and JoaquínErhlich (Finca Lindavista) for permission to work on their farms.Also, we thank the critical reviews of the earlier manuscriptversions by Drs. Carlos Bográn, Roel López, R. Wharton and J.Woolley (all at TAMU). This work was supported in part by CON-ACyT (Mexico) (Doctoral Fellowship to JRLF, #91539), El Colegio dela Frontera Sur (Fundación Produce-Chiapas project Bioecología ymanejo de plagas del café en el Soconusco y Sierra de Chiapas), TexasAgri-Life Research (Hatch Project H-8707), Texas A&M University-CONACyT Collaborative Research Grants Program, and Colegio dePostgraduados.
References
Analytical Software, 2003. Statistix 8. User’s Manual Tallahassee, Florida, USA.Auerbach, M.J., Connor, E.F., Mopper, S., 1995. Minor miners and major miners:
population dynamics of leaf-mining insects. In: Cappuccino, N., Price, P.W.(Eds.), Population Dynamics: New Approaches and Synthesis. Academic Press,San Diego, CA, pp. 83e110.
Barradas, V., Fanjul, L., 1986. Microclimatic characterization of shaded and open-growth coffee (Coffea arabica) plantations in Mexico. Agric. For. Meteorol. 38,101e112.
Blanco, M., Haggar, J., Moraga, P., Madriz, J. del C., Pavon, G., 2003. Morfología delcafé (Coffea arabica L.) en lotes comerciales, Nicaragua. AgroMeso 14, 97e103.
CAB International, 2000. Crop Protection Compendium. CD, second ed. CAB Inter-national, Wallingford, U.K.
Campos, O.G., Decazy, B., Carrillo, E., 1989. Dinámica poblacional del minador de lahoja del cafeto Leucoptera coffeella y sus enemigos naturales en la zona deNuevo San Carlos, Retalhuleu, Guatemala. Turrialba 3, 393e399.
Carvalho, G.A., Miranda, J.C., Moura, A.P., Rocha, L.C.D., Reis, P.R., Vilela, F.Z., 2005.Controle do Leucoptera coffeella (Guérin-Ménèville, Perrottet, 1842) (Lepidop-tera: Lyonetiidae) com inseticidas granulados e sus efeitos sobre vespas pre-dadoras e parasitóides. Arq. Inst. Biol. 72, 63e72.
Comisión Nacional del Agua, 2006. Normales climatológicas estándar y provisio-nales 1961e1990. Comisión Nacional del Agua, Subdirección General Técnica,Unidad del Servicio Meteorológico Nacional, Mexico. World Wide Web Elec-tronic Publication. http://smn.cna.gob.mx/productos/normales/estacion/normales.html (accessed 20.11.06.).
Cornell, H.V., Hawkins, B.A., 1995. Survival patterns and mortality sources ofherbivorous insects: some demographic trends. Am. Nat. 145, 563e593.
Costa-Conceição, C.H., Guerreiro-Filho, O., Gonçalvez, W., 2005. Fluctuação pop-ulacional do bicho-mineiro em cultivares de cafe arabica resistentes a ferrugem.Bragantia 64, 625e631.
de la Mora, A., Livingston, G., Philpott, S.M., 2008. Arboreal ant abundance and leafminer damage in coffee agroecosystems in Mexico. Biotropica 40, 742e746.
Feldpausch, T.R., Riha, S.J., Fernandes, E.C.M., Wandelli, E.V., 2005. Development offorest structure and leaf area in secondary forests regenerating on abandonedpastures in central Amazonia. Earth Interact. 9, 1e22.
Fernandes, F.L., Mantovani, E.C., Neto, H.B., Nunes, V.D.V., 2009. Efeitos de variáveisambientais, irrigação e vespas predadoras sobre Leucoptera coffeella (Guérin-Méneville) (Lepidoptera: Lyonetiidae) no cafeeiro. Neotrop. Entomol. 38,410e417.
Flórez, E., Hernández, M.R., 1981. Fluctuación de la población del minador de lashojas del cafeto, Leucoptera coffeella y de sus enemigos naturales en el Valle delCauca. Rev. Colomb. Entomol. 7, 29e38.
Fordyce, J.A., Shapiro, A.M., 2003. Another perspective on the slow-growth/high-mortality hypothesis: chilling effects on swallowtail larvae. Ecology 84,263e268.
Fragoso, D.B., Guedes, R.N., Guedes, R.N., Picanço, M.C., Zambolim, L., 2002. Insec-ticide use and organophosphate resistance in the coffee leaf miner Leucopteracoffeella (Lepidoptera: Lyonetiidae). Bull. Entomol. Res. 92, 203e212.
Frazer, G.W., Canham, C.D., Lertzman, K.P., 1999. Gap Light Analyzer (GLA): ImagingSoftware to Extract Canopy Structure and Gap Light Transmission Indices fromTrue-Colour Fisheye Photographs: Users Manual and Program Documentation(Version 2.0). Simon Fraser University/The Institute of Ecosystem Studies,Burnaby, BC, Canada/Millbrook, NY, 36 pp.
Gravena, S., 1983. Táticas de manejo integrado do bicho mineiro do cafeeiro Peril-eucoptera coffeella Guérin-Mèneville, 1842: I- Dinâmica populacional e inimigosnaturais. An. Soc. Entomol. Bras. 12, 61e71.
Guerreiro-Filho, O., 2006. Coffee leaf miner resistance. Braz. J. Plant Physiol. 18,109e117.
Guharay, F., Monterroso, D., Staver, C., 2001. El diseño y manejo de la sombra para lasupresion de plagas en cafetales de America Central. Agrofor. Am. 8, 22e29.
Haggar, J.P., Schibli, C., Staver, C., 2001. Cómo Manejar árboles de sombra en cafe-tales. Agrofor. Am. 8, 37e41.
Hespenheide, H.A., 1991. Bionomics of leaf-mining insects. Annu. Rev. Entomol. 36,535e560.
Hixon, M.A., Pacala, S.W., Sandin, S.A., 2002. Population regulation: historicalcontext and contemporary challenges of open vs. closed systems. Ecology 83,1490e1508.
Lomelí-Flores, J.R., Barrera, J.F., Bernal, J.S., 2009. Impact of natural enemies oncoffee leafminer Leucoptera coffeella (Lepidoptera: Lyonetiidae) populationdynamics in Chiapas, Mexico. Biol. Control 51, 51e60.
McMillan, D.M., Fearnley, S.L., Rank, N.E., Dahlhoff, E.P., 2005. Natural temperaturevariation affects larval survival, development and Hsp70 expression in a leafbeetle. Funct. Ecol. 19, 844e852.
Moguel, P., Toledo, V.M., 1999. Biodiversity conservation in traditional coffeesystems of Mexico. Conserv. Biol. 13, 11e21.
Monterrey, J., Suarez, D., Gonzalez, M., 2001. Comportamiento de insectos en sis-temas agroforestales con café en el Pacífico sur de Nicaragua. Agrofor. Am. 8,15e21.
Nestel, D., Dickschen, F., Altieri, M.A., 1994. Seasonal and spatial population loads ofa tropical insect: the case of the coffee leaf-miner in Mexico. Ecol. Entomol. 19,159e167.
Parra, J.R.P., 1985. Biologia comparada de Perileucoptera coffeella (Guérin-Menéville,1842) (Lepidoptera, Lyonetiidae) visando ao seu zoneamento ecológico noestado de São Paulo. Rev. Bras. Entomol. 29, 45e76.
Pereira, E.J., Picanço, M.C., Bacci, L., Crespo, A.L., Guedes, R.N.C., 2007. Seasonalmortality factors of the coffee leafminer, Leucoptera coffeella. Bull. Entomol. Res.97, 421e432.
Philpott, S.M., 2005. Changes in arboreal ant populations following pruning ofcoffee shade-trees in Chiapas, Mexico. Agrofor. Syst. 64, 219e224.
Reis, R.P., Gomez, L.J., Souza, L.J., 1976. F luctuação populacional do “bicho mineiro”das folhas do cafeeiro, Perileucoptera coffeella (Lep: Lyonetiidae), nas regioescaffeiras do Estado de Minas Gerais e identificação dos inimigos naturais. In:Congreso Brasileiro de Pesquisas Cafeeiras, 4o. Resumenes. Caxambú, Brasil, pp.105e106.
Reis Jr., R., Lima, E.R., Vilela, E.F., Barros, R.S., 2000. Method for maintenanceof coffee leaves in vitro for mass rearing of Leucoptera coffeellum(Guérin-Ménèville) (Lepidoptera: Lyonetiidae). An. Soc. Entomol. Bras. 29,849e854.
Ruiz, V.I., Barrera, J.F., Lomelí, R., Pinson, E., Herrera, J., 2004. Infestación y para-sitismo del minador de la hoja del café en el Soconusco, Chiapas, México. In:Resúmenes del I Congreso Internacional sobre Desarrollo de Zonas Cafetaleras.6e8 de Octubre de 2004, Tapachula, Chiapas, México, p. 46.
Saethre, M.G., Hofsvang, T., 2002. Effect of temperature on oviposition behavior,fecundity, and fertility in two northern European populations of the codlingmoth (Lepidoptera: Tortricidae). Environ. Entomol. 31, 804e815.
Sokal, R.R., Rohlf, F.J., 1995. Biometry: the Principles and Practice of Statistics inBiological Research, third ed. Freeman, New York, 887 pp.
SPSS, 2005. SPSS for Windows. Ver 12.0. SPSS, Inc, Chicago, Illinois, USA.Stireman III, J.O., Dyer, L.A., Janzen, D.H., Singer, M.S., Lill, J.T., Marquis, R.J.,
Ricklefs, R.E., Gentry, G.L., Hallwachs, W., Coley, P.D., Barone, J.A., Greeney, H.F.,Connahs, H., Barbosa, P., Morais, H.C., Diniz, I.R., 2005. Climatic unpredictabilityand parasitism of caterpillars: implications of global warming. Proc. Natl. Acad.Sci. U.S.A. 101, 17384e17387.
Tanaka, L.K., Tanaka, S.K., 1982. Rainfall and seasonal changes in arthropod abun-dance on a tropical oceanic island. Biotropica 14, 114e123.
Tejeda-Martinez, A., 1991. An exponential model of the curve of mean monthlyhourly air temperature. Atmosfera 4, 139e144.
Teodoro, A., Klein, A.M., Tscharntke, T., 2008. Environmentally mediated coffee pestdensities in relation to agroforestry management, using hierarchical parti-tioning analyses. Agric. Ecosyst. Environ. 125, 120e126.
Teodoro, A., Klein, A.M., Reis, P.R., Tscharntke, T., 2009. Agroforestry managementaffects coffee pests contingent on season and developmental stage. Agric. For.Entomol. 11, 295e300.
Tipping, P.W., Holko, C.A., Bean, R.A., 2005. Helicoverpa zea (Lepidoptera: Noctuidae)dynamics and parasitism in Maryland soybeans. Fla. Entomol. 88, 55e60.
Tuelher, E.S., Oliveira, E.E., Guedes, R.N.C., Magalhães, L.C., 2003. Ocorrência debicho-mineiro do cafeeiro (Leucoptera coffeella) influenciada pelo períodoestacional e pela altitude. Acta Sci. Agron. 25, 119e124.
Velasco, E., Verdecia, J., Medina, R., Rodriguez, L., 2001. Variaciones en el microclimade un cafetal en dependencia de la exposición a la radiación solar en las con-diciones del macizo de la Sierra Maestra. Cult Trop 22, 53e59.
Villacorta, A., 1980. Alguns fatores que afetam a população estacional de Peril-eucoptera coffeella (Guérin-Mèneville, 1842) (Lepidoptera: Lyonetiidae) noNorte do Paraná, Londrina, PR. An. Soc. Entomol. Bras. 9, 23e32.
Wang, B.D., Ferro, N., Hosmer, D.W., 1997. Importance of plant size, distribution ofegg masses, and weather conditions on egg parasitism of the European cornborer, Ostrinia nubilalis by Trichogramma ostriniae in sweet corn. Entomol. Exp.Appl. 83, 337e345.
Weisser, W.W., Volkl, W., Hassell, M.P., 1997. The importance of adverse weatherconditions for behavior and population ecology of an aphid parasitoid. J. Anim.Ecol. 66, 386e400.
Williams-Linera, G., Herrera, F., 2003. Folivory, herbivores, and environment in theunderstory of a tropical mountain could forest. Biotropica 35, 67e73.
Zar, J.H., 1996. Biostatistical Analysis. Prentice Hall, Upper Saddle River.622 pp.þApps.




















