
Immune response to experimentally induced infection with respiratory syncytial virus: possible role...
9
Downloaded from www.microbiologyresearch.org by IP: 54.147.156.167 On: Sat, 04 Jun 2016 10:46:18 J. gen. Virol. (1989), 70, 325-333. Printed in Great Britain Key words: RSV/immune response/inflammatory cells 325 Immune Response to Experimentally Induced Infection with Respiratory Syncytial Virus: Possible Role in the Development of Pulmonary Disease By PEDRO A. PIEDRA,I'5* t GIOVANNI CAMUSSI 3'4'6 AND PEARAY L. OGRA 1'2's Departments of 1pediatrics, 2Microbiology, 3 pathology and *Medicine, State University of New York, 5Division of Infectious Diseases and Microbiology Laboratories, Children's Hospital, Buffalo, New York 14222 and 6Kidney Research Laboratory, Buffalo General Hospital, Buffalo, New York 14209, U.S.A. (Accepted 17 October 1988) SUMMARY Groups of mice were immunized intraperitoneally with HEp-2 lysate or respiratory syncytial virus (RSV)-infected HEp-2 cells with or without adjuvant(s). The animals were subsequently challenged intranasally (i.n.) with RSV and/or HEp-2 lysate and studied for RSV shedding in the lung, antigen-specific antibody response and pulmonary histopathology. A significant decrease in virus shedding was detected in each of four groups of animals immunized with a virus preparation and in two of four groups that received a HEp-2 lysate. Mice immunized with adjuvant(s) developed higher antibody-specific responses. All vaccinated animals developed pulmonary histopathology only on subsequent i.n. inoculation with RSV and/or HEp-2 tysate. Denatured extracts of purified RSV, HEp-2, BALB/c lung, cotton rat lung, Buffalo green monkey kidney and human buccal epithelium were tested for reactivity against the sera of immunized mice by an immunoblot method. Sera from all groups of immunized mice reacted with the extracts tested. The data suggest a possible role of reactivity to viral as well as non-viral components in the pathogenesis of RSV vaccine- induced pulmonary inflammation in the mouse model system. INTRODUCTION Respiratory syncytial virus (RSV) is the major pathogen responsible for bronchiolitis and other forms of acute lower respiratory tract illnesses in infants and young children (Henderson et al., 1979a; Monto et al., 1975). Considerable information about the pathogenesis and natural history of RSV disease has been collated (Hall et al., 1976, 1984; Henderson et al., 1979b; Welliver et al., 1979, 1980). In addition, purification of the virion, analysis of the viral proteins and characterization of the genome have been accomplished (Fernie et al., 1982; Huang & Wertz, 1982; Lambert & Pons, 1983; Ueba, 1978; Walsh & Hruska, 1983). In spite of the available knowledge, the development of a vaccine that is both safe and efficacious in the prevention of RSV infection has not been realized. In the early 1960s, field trials performed with a formalin-inactivated RSV vaccine were remarkable in that subjects inoculated parenterally with the vaccine developed more severe pulmonary disease on subsequent natural exposure to the wild virus than immunized controls (Fulginiti et al., 1968; Kim et al., 1968). This paradoxical phenomenon of vaccine-induced disease had not been investigated extensively, because an animal model susceptible to pulmonary infection by RSV had not been defined. Recently, Prince and colleagues duplicated a similar phenomenon in cotton rats (Prince et al., 1985). They observed that parenteral immunization with a formalin-inactivated RSV vaccine was characterized by antibody directed t Present address: Department of Microbiologyand Immunology,Baylor College of Medicine, One Baylor Plaza, Houston, Tex. 77030, U.S.A. 0000-8539 © 1989 SGM
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Immune response to experimentally induced infection with respiratory syncytial virus: possible role...

Downloaded from www.microbiologyresearch.org by
IP: 54.147.156.167
On: Sat, 04 Jun 2016 10:46:18
J. gen. Virol. (1989), 70, 325-333. Printed in Great Britain
Key words: RSV/immune response/inflammatory cells
325
Immune Response to Experimentally Induced Infection with Respiratory Syncytial Virus: Possible Role in the Development of Pulmonary Disease
By P E D R O A. P I E D R A , I ' 5 * t G I O V A N N I C A M U S S I 3'4'6 A N D
P E A R A Y L. O G R A 1'2's
Departments of 1pediatrics, 2Microbiology, 3 pathology and *Medicine, State University of New York, 5Division of Infectious Diseases and Microbiology Laboratories, Children's Hospital, Buffalo, New York 14222 and 6Kidney Research Laboratory, Buffalo General Hospital, Buffalo,
New York 14209, U.S.A.
(Accepted 17 October 1988)
SUMMARY
Groups of mice were immunized intraperitoneally with HEp-2 lysate or respiratory syncytial virus (RSV)-infected HEp-2 cells with or without adjuvant(s). The animals were subsequently challenged intranasally (i.n.) with RSV and/or HEp-2 lysate and studied for RSV shedding in the lung, antigen-specific antibody response and pulmonary histopathology. A significant decrease in virus shedding was detected in each of four groups of animals immunized with a virus preparation and in two of four groups that received a HEp-2 lysate. Mice immunized with adjuvant(s) developed higher antibody-specific responses. All vaccinated animals developed pulmonary histopathology only on subsequent i.n. inoculation with RSV and/or HEp-2 tysate. Denatured extracts of purified RSV, HEp-2, BALB/c lung, cotton rat lung, Buffalo green monkey kidney and human buccal epithelium were tested for reactivity against the sera of immunized mice by an immunoblot method. Sera from all groups of immunized mice reacted with the extracts tested. The data suggest a possible role of reactivity to viral as well as non-viral components in the pathogenesis of RSV vaccine- induced pulmonary inflammation in the mouse model system.
INTRODUCTION
Respiratory syncytial virus (RSV) is the major pathogen responsible for bronchiolitis and other forms of acute lower respiratory tract illnesses in infants and young children (Henderson et al., 1979a; Monto et al., 1975). Considerable information about the pathogenesis and natural history of RSV disease has been collated (Hall et al., 1976, 1984; Henderson et al., 1979b; Welliver et al., 1979, 1980). In addition, purification of the virion, analysis of the viral proteins and characterization of the genome have been accomplished (Fernie et al., 1982; Huang & Wertz, 1982; Lambert & Pons, 1983; Ueba, 1978; Walsh & Hruska, 1983). In spite of the available knowledge, the development of a vaccine that is both safe and efficacious in the prevention of RSV infection has not been realized.
In the early 1960s, field trials performed with a formalin-inactivated RSV vaccine were remarkable in that subjects inoculated parenterally with the vaccine developed more severe pulmonary disease on subsequent natural exposure to the wild virus than immunized controls (Fulginiti et al., 1968; Kim et al., 1968). This paradoxical phenomenon of vaccine-induced disease had not been investigated extensively, because an animal model susceptible to pulmonary infection by RSV had not been defined. Recently, Prince and colleagues duplicated a similar phenomenon in cotton rats (Prince et al., 1985). They observed that parenteral immunization with a formalin-inactivated RSV vaccine was characterized by antibody directed
t Present address: Department of Microbiology and Immunology, Baylor College of Medicine, One Baylor Plaza, Houston, Tex. 77030, U.S.A.
0000-8539 © 1989 SGM

Downloaded from www.microbiologyresearch.org by
IP: 54.147.156.167
On: Sat, 04 Jun 2016 10:46:18
326 P. A. PIEDRA AND OTHERS
to the F and G glycoproteins, low to absen t serum neut ra l iz ing act ivi ty, s ignif icant reduct ion in pu lmonary virus shedding and enhanced pu lmonary h is topathology fol lowing exposure to infect ious RSV (Price et al., 1985). E n h a n c e m e n t of lung pa thology has also been observed in mice vacc ina ted with vacc in ia virus r ecombinan t s expressing G, F or N protein, and subsequent ly chal lenged with RSV (Stott et al., 1987). The lung pa thology descr ibed consis ted pr imar i ly of pe r ib ronch io la r and per ivascular infil trates composed of po lymorphonuc lea r cells and lymphocytes .
The present study was under taken to de te rmine whe the r tissue culture componen t s , ad juvants or o ther con taminan t s present in tissue cul ture-grown RSV prepara t ions were capable o f sensi t iz ing the host to the d e v e l o p m e n t o f pu lmonary pa thology fol lowing in t ranasal (i.n.) inoculat ion with RSV. The B A L B / c mouse, a genet ical ly defined an imal permiss ive to pu lmonary repl ica t ion o f R S V (Pr ince et al., 1979; Taylor et al., 1984a, b), was used to conduct these invest igat ions.
METHODS
Animals. RSV-seronegative male BALB/c cRos mice (West Seneca Laboratory, West Seneca, N.Y., U.S.A.) age 7 to 10 weeks were used. The animals were housed in standard mouse cages at the animal facility of the Children's Hospital and were fed rat feed and water ad libitum. Animals infected with RSV were segregated and maintained in separate rooms.
HEp-2 lysate. Monolayers of HEp-2 cells were maintaned with Eagle's MEM supplemented with 2~ foetal calf serum, glutamine and antibiotics. The HEp-2 cell monolayer was disrupted with sterile glass beads and the resulting cell suspension was then sonicated, clarified by low speed centrifugation, snap-frozen and stored at - 7 0 °C. The protein concentration of the HEp-2 lysate was 1.7 mg/ml.
Virus stock. The Long strain of RSV was prepared in HEp-2 cell monolayers as previously described (Scott et al., 1978). The supernatant, containing 1 × 107 p.f.u./ml of virus, was divided into aliquots, snap-frozen and stored at - 7 0 °C. The protein concentration of the virus preparation was 2.3 mg/ml.
Purification of RSV. Semi-purified RSV (P-RSV) was prepared with a discontinuous sucrose-Tris buffer gradient (Ueba, 1978). The virus band observed at the 35 to 45 ~ sucrose interphase was collected, divided into aliquots, snap-frozen and stored at - 7 0 °C.
Preparation of vaccines. All virus vaccine preparations contained 1 x 106 p.f.u. (230 ~tg of protein) of live virion. In addition, some virus vaccine preparations contained 10 mg of AI(OH)3 (Wyeth, Philadelphia, Pa., U.S.A.) or 109 heat-killed Bordetellapertussis cells (kindly provided by J. R. Mitchell, Department of Public Health, Lansing, Mich., U.S.A.) or 10 mg of AI(OH) 3 and 109 heat-killed B. pertussis cells. A 10-fold reduction in virus viability was noted in the vaccine preparations that contained adjuvants. The virus-flee vaccine preparations each contained 170 ~tg protein of uninfected HEp-2 cell lysate and were prepared similarly to the virus vaccine preparations described above.
Experimental design. Groups of five animals were immunized with the virus or virus-free tissue culture vaccine. All animals were immunized intraperitoneally (i.p.) at 8 and 11 weeks of age. Pooled serum samples diluted fivefold in phosphate-buffered saline (PBS) were collected from each group at intervals of 1 week after the second i.p. dose and stored at - 7 0 °C. During week 16, one animal from each group was sacrificed by CO2 asphyxiation and its left lung was fixed with 10~ buffered formalin. The remaining mice were challenged with 50 gl RSV (5 x 105 p.f.u.) and sacrificed 4 or 6 days later. After the animals were sacrificed, the left lung was fixed with 10~ buffered formalin and the right lung was freshly processed for virus quantification. In addition, groups of five animals immunized with virus-free vaccine preparations according to the above protocol were challenged with 50 vtl (85 ~tg) HEp-2 lysate. The left lung from each mouse was studied for histopathology.
Antibody quantification. RSV and HEp-2 IgG antibody activities were assayed by an ELISA method employing microtitre plates coated with P-RSV (0-7 ~tg/well) or HEp-2 (0.7 ~tg/well) antigens (Welliver et at., 1985). Peroxidase-conjugated, afffinity-purified goat anti-mouse IgG (Cooper Biomedical, Malvern, Pa., U.S.A.) was the enzyme-labelled antiglobulin, o-Phenylenediamine dihydrochloride (Sigma), the substrate used for colour development, was quenched with 1 M-H2SO, after 20 min incubation in the dark at room temperature. The ELISA titre was the highest dilution that gave an absorbance value greater than three times the standard deviation of preimmune sera. The ELISA titre was expressed in ELISA units (,4 of ELISA titre x I/ELISA titre).
Virus quantification. The right lung from each mouse was homogenized in MEM containing 2~ foetal calf serum (FCS) to form a 10~ suspension. The homogenate was centrifuged at 500 g for 30 min at 4 °C, passed through a 0.45 ~tm Millipore filter, snap-frozen and stored at - 70 °C. The plaque assay as previously described (Wong et al., 1985) was used to quantify virus viability from the right lung of each animal. The results were expressed in p.f.u./g of lung tissue.
Histopathology. Sections of lung were stained with haematoxylin and eosin (H & E) and periodic acid-Schiff

Downloaded from www.microbiologyresearch.org by
IP: 54.147.156.167
On: Sat, 04 Jun 2016 10:46:18
Immune response to R S V in mice 327
(PAS). PAS-stained lung sections were useful in distinguishing collapsed from diseased parenchyma and therefore PAS-stained lung sections were used to grade the inflammatory response in the lung. All the slides were examined by the collaborating pathologist (G. Camussi), who was unaware of the study design. The pulmonary histopathology was scored by assigning a grade ranging from 0 (absent) to 3 (severe) to each of the following features: (i) inflammation around vessels, (ii) peribronchial infiltrates, (iii) septal thickening and (iv) the magnitude of interstitial cellular inflammatory response. Based on this point system, the pulmonary histology was defined as normal (0 to 2-0) or mild (2.1 to 6.0) or moderate (6-1 to 10-0) or severe (10-1 to 12-0) pathology.
Immunoblot assay. The cellular antigens used for the assay were prepared in the following manner. Established HEp-2 and Buffalo green monkey kidney cell (BGM) monolayers were maintained with MEM with 2~0 FCS and supplemented with antibiotics. The monolayers were washed extensively with PBS, harvested, sonicated and clarified at 10000g for 30 rain. The supernatant obtained was stored in aliquots at - 20 °C. Lungs from cotton rats and BALB/c mice were processed after the animals had been sacrificed by CO 2 asphyxiation. The right ventricle as well as the bronchial tree were perfused with 20 ml of PBS. Lungs were minced and filtered over a wire screen. The cells were washed with PBS, sonicated and clarified. The aliquots were stored at - 20 °C. Human buccal epithelial (HBE) cells collected by scraping the buccal epithelial lining were washed in PBS, sonicated and clarified. The supernatant was stored at - 20 °C. All antigen preparations were used at a concentration of 50 ~tg/ml except for HBE which contained 15 gg/ml.
Separation of proteins was performed on a 4 ~ stacking polyacrylamide gel in combination with a 12.5~ separating gel, using the Laemmli buffer system (Laemmli, 1970). Electrophoresis was performed with 30 mA of constant current per gel in a vertical slab gel unit (SE 600, Hoefer Scientific Instruments, San Francisco, Ca., U.S.A.) and was stopped when the dye marker reached 2 cm from the bottom of the gel. Low Mr protein standards (Bio-Rad) were used in each run.
Electrophoretic transfer of proteins onto nitrocellulose membrane was done according to the method of Towbin et al. (1979), using a TE52 Transphor cell (Hoefer Scientific Instruments; 100 V, 90 min).
Immunogenic determinants on the immunoblots were determined by an indirect technique of antige~antibody binding. In brief, the immunoblots were quenched with 3 ~ gelatin-Tris-buffered saline (GTBS) for l h at room temperature, washed three times with a 0-05~ Tween-20 Tris buffer (TTBS) and placed on a PR-200 Deca Probe unit (Hoefer Scientific Instruments). Mouse sera diluted 100-fold in 1 ~o GTBS were incubated for 2 h on the immunoblots. After three washes with TTBS, peroxidase-conjugated, affinity-purified goat anti-mouse IgG (Bio- Rad) diluted 1/1000 with 1 ~o gelatin-TTBS was incubated for 1 h. The immunoblots were rinsed with TTBS followed by Tris-buffered saline. Colour was developed in the dark during 1 h incubation with 4-chloro-l-naphthol (Bio-Rad) and the reaction was stopped by the addition of cold distilled water. No colour development occurred with preimmune sera.
Statistical analysis. The unpaired Student's t-test was used to compare differences in virus concentration and pulmonary histopathology score for the different groups of mice.
RESULTS
Serum IgG antibody response
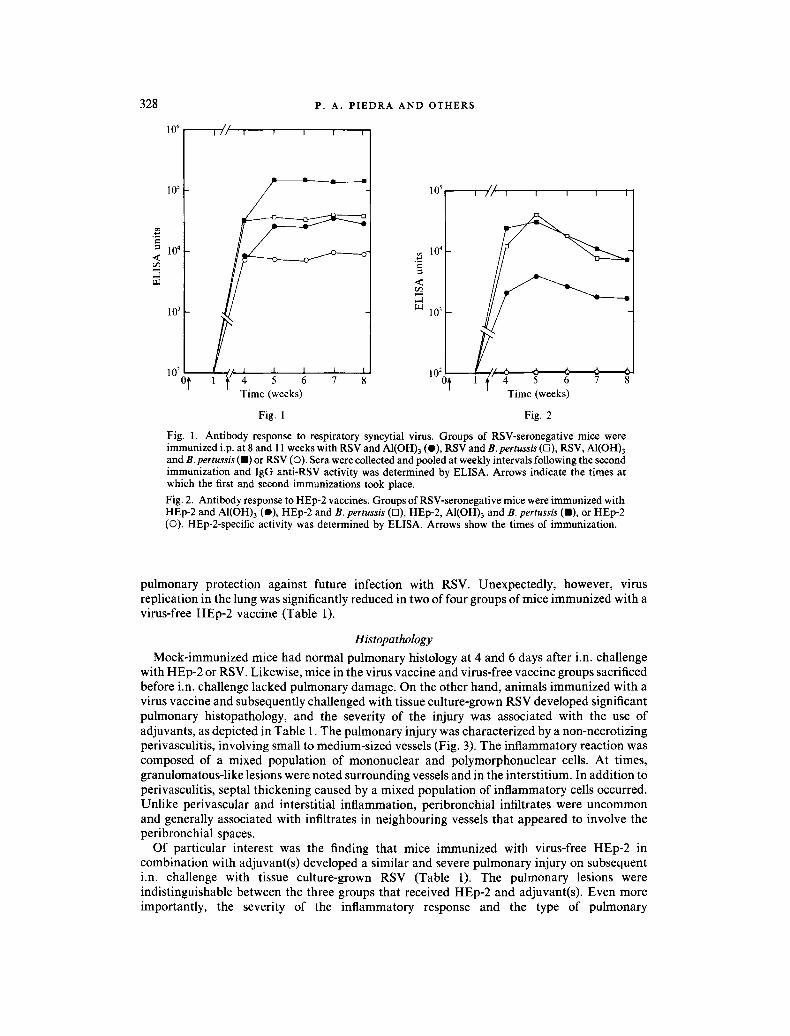
RSV-specif ic an t ibody was not de tec ted in any of the p r e i m m u n i z a t i o n samples. The RSV- specific an t ibody activity, de te rmined by E L I S A , in animals immunized i.p. wi th RSV vacc ine prepara t ions is depic ted in Fig. 1. In the virus vacc ine groups, the IgG-specif ic an t ibody response to R S V was most effectively induced in animals i m m u n i z e d with an adjuvant - con ta in ing vacc ine as compared to virus alone. The c o m b i n a t i o n of both adjuvants , B. pertussis and AI(OH)3 , incorpora ted with RSV induced the highest I g G an t i -RSV ant ibody titre. Signif icant an t ibody act ivi ty to RSV deve loped in mice i m m u n i z e d wi th RSV alone.
In contras t to the virus vacc ine groups, only an imals vacc ina ted wi th HEp-2 in the presence of" an ad juvent produced a signif icant level o f HEp-2-speci f ic I g G act ivi ty (Fig. 2). The ant ibody- specific response to tissue culture componen t s appeared greater when mice rece ived e i ther B. pertussis or B. pertussis and AI(OH)3 as adjuvants . Unexpec ted ly , HEp-2 prepara t ions alone did not induce a detectable an t ibody response to tissue cul ture components .
Pulmonary virus replication
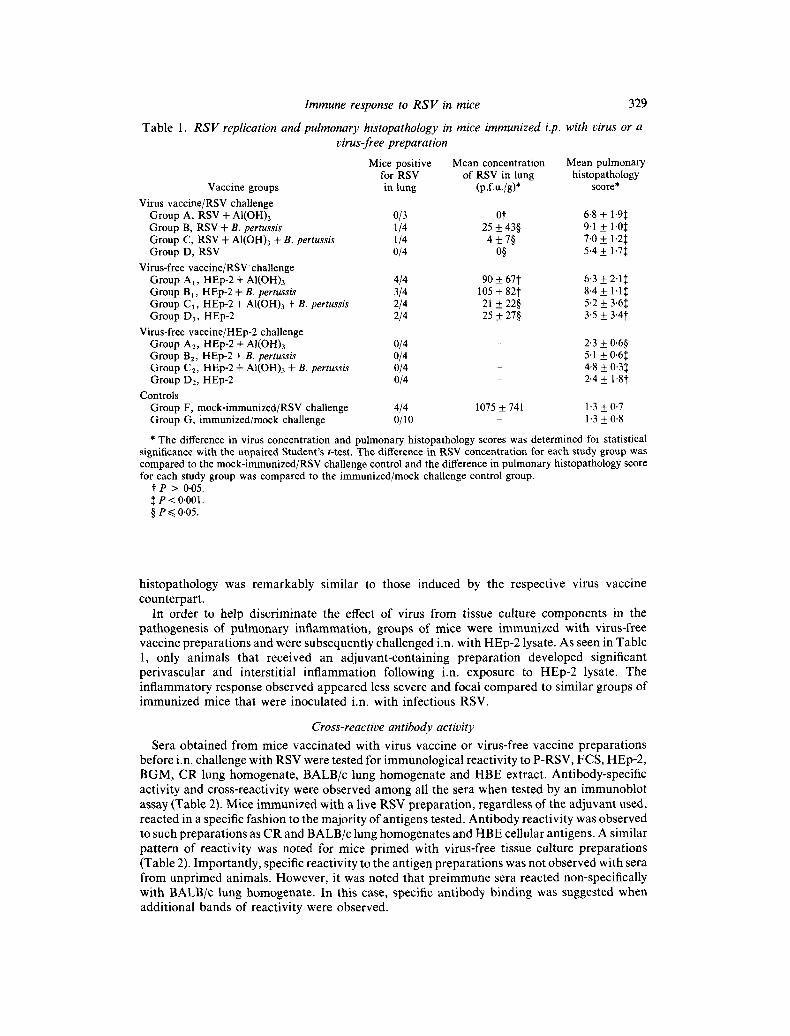
Quant i f ica t ion of virus f rom the lungs o f animals chal lenged i.n. wi th R S V is presented in Tab le 1. As noted, m o c k - i m m u n i z e d mice had a significantly h igher level o f virus shedding in the lung compared to an imals i m m u n i z e d wi th a virus prepara t ion . The v i rus -vacc ina ted animals , whe the r i m m u n i z e d in the presence or absence of an adjuvant , deve loped signif icant
Downloaded from www.microbiologyresearch.org by
IP: 54.147.156.167
On: Sat, 04 Jun 2016 10:46:18
10 5
10~ I / / I r
P. A. PIEDRA AND OTHERS
I I I
10 4 <
1 0 3
Time (weeks)
.~ 104
<
.J 10 3
328
105 I / ~ I i i
10 2 10 2 ot
I I
v t l
- ~ 1 / ~ 4 6 6 6 5 6 7 8 Time (weeks)
Fig. 1 Fig. 2
Fig. 1. Antibody response to respiratory syncytial virus. Groups of RSV-seronegative mice were immunized i.p. at 8 and 11 weeks with RSV and AI(OH)3 (O), RSV and B. pertussis (n), RSV, AI(OH)3 and B. pertussis (11) or RSV (O). Sera were collected and pooled at weekly intervals following the second immunization and IgG anti-RSV activity was determined by ELISA. Arrows indicate the times at which the first and second immunizations took place. Fig. 2. Antibody response to HEp-2 vaccines. Groups of RSV-seronegative mice were immunized with HEp-2 and AI(OH)3 (e), HEp-2 and B. pertussis (D), HEp-2, AI(OH)3 and B. pertussis (11), or HEp-2 (0). HEp-2-specific activity was determined by ELISA. Arrows show the times of immunization.
pulmonary protection against future infection with RSV. Unexpectedly, however, virus replication in the lung was significantly reduced in two of four groups of mice immunized with a virus-free HEp-2 vaccine (Table 1).
Histopathology
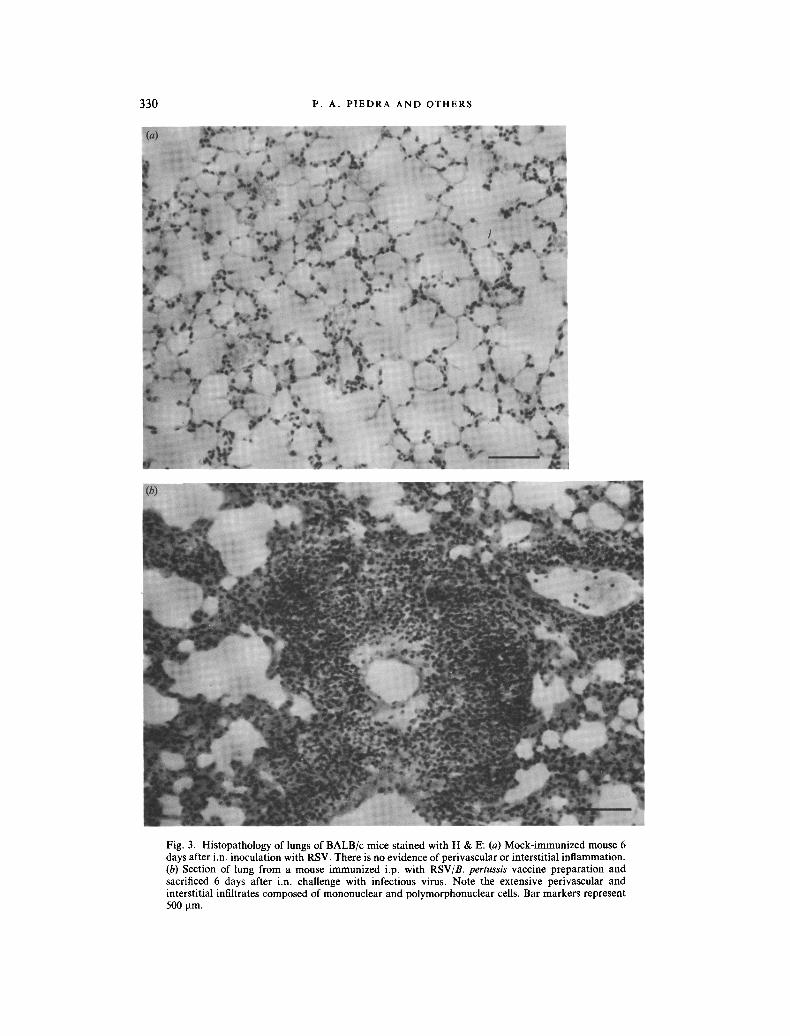
Mock-immunized mice had normal pulmonary histology at 4 and 6 days after i.n. challenge with HEp-2 or RSV. Likewise, mice in the virus vaccine and virus-free vaccine groups sacrificed before i.n. challenge lacked pulmonary damage. On the other hand, animals immunized with a virus vaccine and subsequently challenged with tissue culture-grown RSV developed significant pulmonary histopathology, and the severity of the injury was associated with the use of adjuvants, as depicted in Table l. The pulmonary injury was characterized by a non-necrotizing perivasculitis, involving small to medium-sized vessels (Fig. 3). The inflammatory reaction was composed of a mixed population of mononuclear and polymorphonuclear cells. At times, granulomatous-like lesions were noted surrounding vessels and in the interstitium. In addition to perivasculitis, septal thickening caused by a mixed population of inflammatory cells occurred. Unlike perivascular and interstitial inflammation, peribronchial infiltrates were uncommon and generally associated with infiltrates in neighbouring vessels that appeared to involve the peribronchial spaces.
Of particular interest was the finding that mice immunized with virus-free HEp-2 in combination with adjuvant(s) developed a similar and severe pulmonary injury on subsequent i.n. challenge with tissue culture-grown RSV (Table 1). The pulmonary lesions were indistinguishable between the three groups that received HEp-2 and adjuvant(s). Even more importantly, the severity of the inflammatory response and the type of pulmonary
Downloaded from www.microbiologyresearch.org by
IP: 54.147.156.167
On: Sat, 04 Jun 2016 10:46:18
Table 1.
Immune response to R S V in mice 329
R S V replication and pulmonary histopathology in mice immunized i.p. with virus or a virus-free preparation
Mice positive Mean concentration Mean pulmonary for RSV of RSV in lung histopathology
Vaccine groups in tung (p.f.u./g)* score* Virus vaccine/RSV challenge
Group A, RSV + AI(OH)3 0/3 0i" 6.8 ± 1.9:~ Group B, RSV + B. pertussis 1/4 25 + 43§ 9.1 + 1.0~ Group C, RSV + AI(OH)3 + B. pertussis 1/4 4 + 7§ 7.0 + 1.2:~ Group D, RSV 0/4 0§ 5.4 + 1.7:~
Virus-free vaccine/RSV challenge Group A l , HEp-2 + AI(OH)3 4/4 90 + 67"}" 6-3 + 2-1~ Group Bi, HEp-2 + B. pertussis 3/4 105 + 82I" 8-4 + 1.1:~ Group Cl, HEp-2 + AI(OH)3 + B. pertussis 2/4 21 + 22§ 5.2 + 3.6:~ Group D1, HEp-2 2/4 25 + 27§ 3.5 + 3"4i
Virus-free vaccine/HEp-2 challenge Group A2, HEp-2 + AI(OH)3 0/4 - 2-3 + 0.6§ Group B2, HEp-2 + B. pertussis 0/4 - 5.1 + 0.6~ Group C2, HEp-2 + AI(OH)3 + B. pertussis 0/4 - 4.8 + 0-3~: Group D~, HEp-2 0/4 - 2.4 + 1.8~
Controls Group F, mock-immunized/RSV challenge 4/4 Group G, immunized/mock challenge 0/10
1075 + 741 1.3 + 0.7 - 1 - 3 + 0-8
* The difference in virus concentration and pulmonary histopathology scores was determined for statistical significance with the unpaired Student's t-test. The difference in RSV concentration for each study group was compared to the mock-immunized/RSV challenge control and the difference in pulmonary histopathology score for each study group was compared to the immunized/mock challenge control group.
t P > 0-05. ~. P < 0.001. § e~<0-05.
histopathology was remarkably similar to those induced by the respective virus vaccine counterpart .
In order to help discr iminate the effect of virus from tissue culture components in the pathogenesis of pulmonary inflammation, groups of mice were immunized with virus-free vaccine preparat ions and were subsequently challenged i.n. with HEp-2 lysate. As seen in Table 1, only animals that received an adjuvant-containing preparat ion developed significant perivascular and interstit ial inf lammation following i.n, exposure to HEp-2 lysate. The inflammatory response observed appeared less severe and focal compared to similar groups of immunized mice that were inoculated i.n. with infectious RSV.
Cross-reactive antibody activity
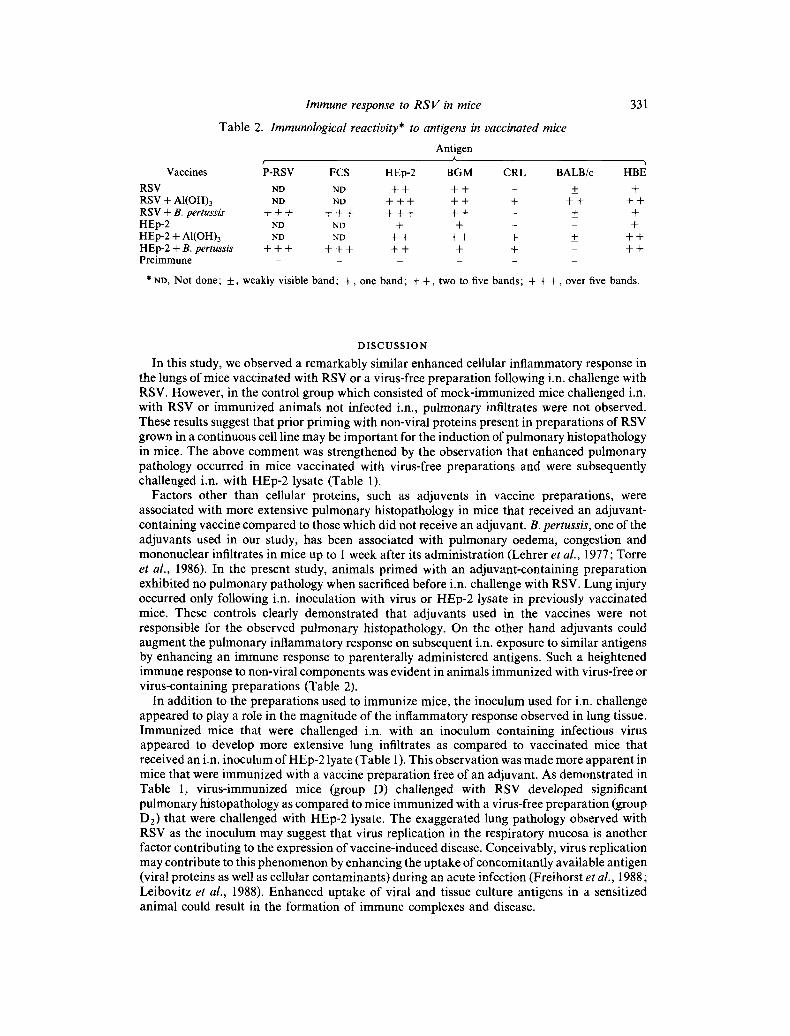
Sera obtained from mice vaccinated with virus vaccine or virus-flee vaccine preparat ions before i.n. challenge with RSV were tested for immunological reactivi ty to P-RSV, FCS, HEp-2, BGM, CR lung homogenate, BALB/c lung homogenate and HBE extract. Antibody-specific activity and cross-reactivity were observed among all the sera when tested by an immunoblot assay (Table 2). Mice immunized with a live RSV preparat ion, regardless of the adjuvant used, reacted in a specific fashion to the majori ty of antigens tested. Ant ibody reactivi ty was observed to such preparat ions as CR and BALB/c lung homogenates and HBE cellular antigens. A similar pat tern of reactivity was noted for mice pr imed with virus-free tissue culture preparat ions (Table 2). Important ly, specific reactivi ty to the antigen preparat ions was not observed with sera from unprimed animals. However, it was noted that pre immune sera reacted non-specifically with BALB/c lung homogenate. In this case, specific ant ibody binding was suggested when addit ional bands of reactivi ty were observed.
Downloaded from www.microbiologyresearch.org by
IP: 54.147.156.167
On: Sat, 04 Jun 2016 10:46:18
330 P. A. PIEDRA AND OTHERS
Fig. 3. Histopathology of lungs of BALB/c mice stained with H & E. (a) Mock-immunized mouse 6 days after i.n. inoculation with RSV. There is no evidence of perivascular or interstitial inflammation. (b) Section of lung from a mouse immunized i.p. with RSV/B. pertussis vaccine preparation and sacrificed 6 days after i.n. challenge with infectious virus. Note the extensive perivascular and interstitial infiltrates composed of mononuclear and polymorphonuclear cells. Bar markers represent 500 t.tm.
Downloaded from www.microbiologyresearch.org by
IP: 54.147.156.167
On: Sat, 04 Jun 2016 10:46:18
Immune response to R S V & mice
Table 2. Immunological reactivity* to antigens in vaccinated mice
331
Antigen A ¢
Vaccines P-RSV FCS HEp-2 BGM CRL BALB/c HBE
RSV ND ND + + + + - + + RSV + AI(OH)3 ND ND + + + + + + + + + + RSV + B. pertussis + + + + + + + + + + + - +_ + HEp-2 ND ND + + - n t- HEp-2 + AI(OH)3 ND ND + + + + + + + + HEp-2 + B. pertussis + + + + + + + + + + + + P r e i m m u n e . . . . . .
* ND, Not done; + , weakly visible band; + , one band; + + , two to five bands; + + + , over five bands.
D I S C U S S I O N
In this study, we observed a remarkably similar enhanced cellular inflammatory response in the lungs of mice vaccinated with RSV or a virus-free preparation following i.n. challenge with RSV. However, in the control group which consisted of mock-immunized mice challenged i.n. with RSV or immunized animals not infected i.n., pulmonary infiltrates were not observed. These results suggest that prior priming with non-viral proteins present in preparations of RSV grown in a continuous cell line may be important for the induction of pulmonary histopathology in mice. The above comment was strengthened by the observation that enhanced pulmonary pathology occurred in mice vaccinated with virus-free preparations and were subsequently challenged i.n. with HEp-2 lysate (Table 1).
Factors other than cellular proteins, such as adjuvents in vaccine preparations, were associated with more extensive pulmonary histopathology in mice that received an adjuvant- containing vaccine compared to those which did not receive an adjuvant. B. pertussis, one of the adjuvants used in our study, has been associated with pulmonary oedema, congestion and mononuclear infiltrates in mice up to 1 week after its administration (Lehrer et al., 1977; Torre et al., 1986). In the present study, animals primed with an adjuvant-containing preparation exhibited no pulmonary pathology when sacrificed before i.n. challenge with RSV. Lung injury occurred only following i.n. inoculation with virus or HEp-2 lysate in previously vaccinated mice. These controls clearly demonstrated that adjuvants used in the vaccines were not responsible for the observed pulmonary histopathology. On the other hand adjuvants could augment the pulmonary inflammatory response on subsequent i.n. exposure to similar antigens by enhancing an immune response to parenterally administered antigens. Such a heightened immune response to non-viral components was evident in animals immunized with virus-free or virus-containing preparations (Table 2).
In addition to the preparations used to immunize mice, the inoculum used for i.n. challenge appeared to play a role in the magnitude of the inflammatory response observed in lung tissue. Immunized mice that were challenged i.n. with an inoculum containing infectious virus appeared to develop more extensive lung infiltrates as compared to vaccinated mice that received an i.n. inoculum of HEp-2 lyate (Table 1). This observation was made more apparent in mice that were immunized with a vaccine preparation free of an adjuvant. As demonstrated in Table 1, virus-immunized mice (group D) challenged with RSV developed significant pulmonary histopathology as compared to mice immunized with a virus-free preparation (group D2) that were challenged with HEp-2 lysate. The exaggerated lung pathology observed with RSV as the inoculum may suggest that virus replication in the respiratory mucosa is another factor contributing to the expression of vaccine-induced disease. Conceivably, virus replication may contribute to this phenomenon by enhancing the uptake of concomitantly available antigen (viral proteins as well as cellular contaminants) during an acute infection (Freihorst et aL, 1988; Leibovitz et al., 1988). Enhanced uptake of viral and tissue culture antigens in a sensitized animal could result in the formation of immune complexes and disease.

Downloaded from www.microbiologyresearch.org by
IP: 54.147.156.167
On: Sat, 04 Jun 2016 10:46:18
332 P. A. P I E D R A A N D OTHERS
In each of four virus vaccine groups and two of four virus-free vaccine groups, we observed significant protection against virus replication in the lung (Table 1). This finding does not appear to be related to the method of i.n. inoculation, since unprimed animals infected i.n. using a similar technique could be infected with RSV (Table 1) and the level of virus replication observed was comparable to that observed by other investigators who used the Long strain of RSV to infect BALB/c mice (Prince et al., 1979). In addition, all animals that received either an adjuvant-containing or adjuvant-free vaccine experienced a similar level of protection against RSV infection. Thus, adjuvants did not appear to influence the course or outcome of virus infection following immunization. On the other hand, sensitization with non-viral components may have induced a significant inflammatory response that could have limited virus replication in the lungs on subsequent challenge. Alternatively, the production of HEp-2-specific antibodies could have prevented virus attachment to HEp-2 cells by blocking the putative cellular receptor(s) involved in virus-cell attachment. Such blocking antibody would thereby preclude viral replication and subsequent detection of the virus by the plaque assay employed here (Crowell et al., 1985).
We thank Dr Edward E. Walsh for constructive comments and Karen Kincade for preparing the manuscript . This work was supported by a grant from the National Institute of Allergy and Infectious Diseases (AI-15939).
R E F E R E N C E S
CROWELL, R. L., FIELD, A. K., SCHLEIF, W. A., LONG, W. L., COLONNO, R. J., MAPOLES, J. E. & EMINI, E. A. (1985). Monoclonal antibody that inhibits infection of HeLa and rhabdomyosarcoma cells by selected enteroviruses through receptor blockade. Journal of Virology 57, 438-445.
FERNIE, B. F., COTE, P. J. & GERIN, J. L. (1982). Classification of hybridomas to respiratory syncytial virus glycoproteins. Proceedings of the Society for Experimental Biology and Medicine 171, 266-271.
FREIHORST, J., PIEDRA, P. A., OKAMOTO, Y. & OGRA, P. L. (1988). Effect of respiratory syncytial virus infection on the uptake of and immune response to other inhaled antigens. Proceedings of the Society of Experimental Biology and Medicine 188, 191 197.
FULGINITI, V. A., ELLER, J. J., SIEBER, O. F., JOYNER, J. W., MINAMITANI, M. & MEIKLEJOHN, G. (1968). Respiratory virus immunization. I. Field trial of two inactivated respiratory virus vaccines: an aqueous trivalent parainfluenza virus vaccine and an alum-precipitated respiratory syncytial virus vaccine. American Journal of Epidemiology 89, 435-448.
HALL, C. B., DOUGLAS, R. C. & GEIMAN, J. M. (1976). Respiratory synctial virus infections in infants. Quanti tat ion and duration of shedding. Journal of Pediatrics 89, 11-15.
HALL, C. B., HALL, W. J., GALA, C. L., MAGILL, F. B. & LEDDY, J. P. (1984). Long-term prospective study in children after respiratory virus infection. Journal of Pediatrics 105, 358-364.
HENDERSON, F. W., CLYDE, W. A., COLLIER, A. M., DENNY, F. W., SENIOR, R. J., SHEAFFER, C. I , CONLEY, W. G. & CHRISTIAN, R. M. (1979a). The etiologic and epidemiologic spectrum of bronchiolitis in pediatric practice. Journal of Pediatrics 95, 183-190.
HENDERSON, F. W., COLLIER, A. M., CLYDE, W. A. & DENNY, F. W. (1979b). Respiratory syncytial virus infection, reinfection, and immuni ty : a prospective, longitudinal study in young children. New England Journal of Medicine 300, 530-534.
HUANG, Y. r. & WERTZ, G. W. (1982). The genome of respiratory syncytial virus is a negative stranded R N A that codes for at least seven m R N A species. Journal of Virology 43, 150-157.
KIM, H. W., CANCHOLA, J. G., BRANDT, C. D., PYLES, G., CHANOCK, R. M., JENSEN, K. & PARROTT, R. H. (1968). Respiratory syncytial virus disease in infants despite prior administrat ion of antigenic inactivated vaccine. American Journal of Epidemiology 89, 422-433.
LAEMMLI, U. K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, London 227, 680-685.
LAMBERT, D. M. & PONS, M. W. (1983). Respiratory syncytial virus glycoproteins. Virology 130, 204-214. LEHRER, S. B., NAKAMURA, R. M. & TAN, E. M. (1977). Histopathological changes in mice treated with extracts of the
histamine-sensitizing factor of Bordetella pertussis. International Archives of Allergy and Applied Immunology 54, 129-142.
LEIBOVITZ, E., FREIHORST, J., PIEDRA, P. A. & OGRA, P. L. (1988). Modulation of systemic and mucosal immune responses to inhaled ragweed antigen in experimentally induced infection with respiratory syncytial virus implication in virally induced allergy. International Archives of Allergy and Applied Immunology 86, 112-116.
MONTO, A. S., BRYAN, E. R. & RHODES, L. M. (1975). The Tecumseh study of respiratory illness. VII. Further observations on the occurrence of respiratory syncytial virus and Mycoplasma pneumoniae infections. American Journal of Epidemiology 100, 458-468.
PRINCE, G. A., HORSWOOD, R. L., BERNDT, J., SUFFIN, S. C. & CHANOCK, R. M. (1979). Respiratory syncytial virus infection in inbred mice. Infection and Immunity 26, 764-766.

Downloaded from www.microbiologyresearch.org by
IP: 54.147.156.167
On: Sat, 04 Jun 2016 10:46:18
Immune response to R S V in mice 333
PRINCE, G. A., JENSON, A. B., HEMMING, V. G., MURPHY, B. R., WALSH, E. E., HORSWOOD, R. L. & CHANOCK, R. M. (1985). Enhancement of respiratory syncytial virus pulmonary pathology in cotton rats by prior intramuscular inoculation of foimalin-inactivated virus. Journal of Virology 57, 721-728.
SCOTT, R., KAUL, A., SCOTT, M., CHIBA, Y. & OGRA, P. L. (1978). Development of in vitro correlation of cell mediated immuni ty to respiratory syncytial virus infection in man. Journal oflnfectious Diseases 137, 810-817.
STOTT, E. J., TAYLOR, G., BALL, L. A., ANDERSON, K., YOUNG, K. K. Y., KING, A. M. Q. & WERTZ, G. W. (1987). Immune and histopathological responses in animals vaccinated with recombinant vaccinia viruses that express individual genes of h u m a n respiratory syncytial virus. Journal of Virology 61, 3855-3861.
TAYLOR, G., STOTr, E. J., BEW, M., FERNIE, B. F., COTE, P. J., COLLINS, A. P., HUGHES, M. & JEBBETT, J. (1984a). Monoclonal antibodies protect against respiratory syncytial virus infection in mice. Immunology 52, 137-142.
TAYLOR, G., STOTT, E. J., HUGHES, M. & COLLINS, A. P. (1984b). Respiratory syncytial virus infection in mice. Infection and Immunity 43, 639-655.
TORRE, D. C., QUADRELLI, C., PUGLIESE, A. & MAGGIOLO, F. (1986). Effect of betamethasone in lungs of mice treated with lymphocytosis-promoting factor of Bordetella pertussis. Journal of Infectious Diseases 154, 238-244.
TOWBIN, H., STAEHELIN, T. & GORDON, J. (1979). Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proceedings of the National Academy of Sciences, U.S.A. 76, 4350-4354.
UEBA, O. (1978). Respiratory syncytial virus. I. Concentrat ion and purification of the infectious virus. Acta medicinae Okoyama 32, 265-272.
WALSH, E. E. & HRUSKA, J. (1983). Monoclonal antibodies to respiratory syncytial virus proteins. Identification of the fusion protein. Journal of Virology 47, 171 177.
WELLIVER, R. C., KAUL, A. & OGRA, P. L. (1979). Cell-mediated immune response to respiratory syncytial virus infection: relationship to the development of reactive airway disease. Journal of Pediatrics 94, 370-375.
WELLIVER, R. C., KAUL, T. N., PUTNAM, T. I., SUN, M., RIDDLESBERGER, K. & OGRA, P. L. (1980). The antibody response to primary and secondary infection with respiratory syncytial virus in kinetics of class-specific responses. Journal of Pediatrics 96, 808-813.
WELLIVER, R. C., SUN, M., RINALDO, D. & OGRA, P. L. (1985). Respiratory syncytial virus-specific IgE responses following infection: evidence for a predominantly mucosal response. Pediatric Research 19, 420-424.
WONG, D. T., ROSENBAND, M., HOVEY, g. & OGRA, P. L. (1985). Respiratory syncytial virus infection in immunosuppressed animals: implications in human infection. Journal of Medical Virology" 17, 359-370.
(Received 5 July 1988)




















