
Ilja Svetnik, BSc - unipub
74
MASTER THESIS Ilja Svetnik, BSc RED LIST AND DNA BARCODING OF CARINTHIAN AND STYRIAN CENTIPEDES (CHILOPODA) For the master’s program Ecology and Evolutionary Biology UNIVERSITY OF GRAZ - 2019
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Ilja Svetnik, BSc - unipub

MASTER THESIS
Ilja Svetnik, BSc
RED LIST AND DNA BARCODING OF CARINTHIAN AND
STYRIAN CENTIPEDES (CHILOPODA)
For the master’s program
Ecology and Evolutionary Biology
UNIVERSITY OF GRAZ - 2019

ACKNOWLEDGMENTS
This work could not have been possible without the immense support of the University of Graz and the
outstanding Professors and Researchers of the Institute of Biology, as well as its head at the time Dr.
Christian Sturmbauer.
I want to thank my supervisor Dr. Stephan Koblmüller and my mentor Lukas Zangl, MSc, who supported
me in every way possible and shaped this thesis into what it has become. I also want to thank Dr. Werner
Holzinger for his immense help and advice in creating a cohesive Red List, and his support years ago,
when I just thought about maybe investing time in researching soil arthropods.
My thanks also extend to my previous supervisor Dr. Gernot Kunz, who supported me during my
bachelor thesis and thus laid the ground stone for this thesis’ research.
I also want to thank Dr. Nikolaus Szucsish from the Vienna Natural History Museum (NHM) for providing
me with additional material from different locations, thereby adding immensely to my barcoding data
bank.
Lastly, I want to thank all the people who I had contact to and who offered their support during the
making of this thesis, including but not limited to Dr. Karin Voigtländer, Dr. Christian Wieser, and Dr.
Henrik Enghoff as well as my colleagues Michaela Bodner, David Fröhlich and Maximilian Wagner.

3
ABSTRACT
Centipedes are predatory, multi-legged arthropods inhabiting soil, litter, and rocky surfaces in all kinds of
habitats. As such they serve an important role in the global detritus chain and soil ecology. Central
European centipedes have been studied ever since the 18th century with a focus on alpine species and
species from the British Isles. Austria has been a hotspot of these studies ever since, and has been
known to be home to around 70 native species of centipedes. Unfortunately, these species have been
described multiple times with different names and have been plotted together and split apart
taxonomically various times in the last decades, leaving taxonomists confused, especially due to the lack
of consistent literature. The scarcity of taxonomic information and difficulty of evaluating population
sizes leads to a lack of red lists on centipedes, especially those with reasonable criteria. DNA barcoding is
a recent method that can assist in studying the phylogeny of any living being that is gaining popularity,
yet should be used with great care. Today many ecologists turn to analysis of DNA or RNA in order to
study the diversity of beings. We take advantage of both classic and new methods in order to resolve the
taxonomy and status of native centipede species.

TABLE OF CONTENTS
ACKNOWLEDGMENTS ................................................................................................................................... 2
ABSTRACT ................................................................................................................................................... 3
TABLE OF CONTENTS ..................................................................................................................................... 4
INTRODUCTION ............................................................................................................................................ 6
Concerning Centipedes 6
Evolutionary and taxonomic history 6
Taxonomy and morphology 7
Anatomy and physiology 9
Mating and reproduction 10
Systematics and species numbers 11
Distribution 12
Prey and predators 13
Venomousness and danger 14
Concerning Barcoding 16
The “Barcode of Life” Projects 17
Part I.
Red List of Centipedes 19
State of the art 19
Methodology 22
Disclaimer 23
Results 25
Checklist of Carinthia 25
Checklist of Styria 26
Statistical analysis 27
Legend of the Red List 28
Red List of Centipedes of Carinthia 29
Red List of Centipedes of Styria 41
Critical reflection 52
Summary and outlook 54
Part II.
DNA Barcoding of Centipedes 55

5
State of the art 55
Methodology 58
Results and discussion 60
Summary and outlook 64
APPENDIX ................................................................................................................................................. 68
BIBLIOGRAPHY ........................................................................................................................................... 69

INTRODUCTION
Concerning Centipedes
Centipedes can be generally described as predatory arthropods of the taxonomic class Chilopoda, part of
the subphylum Myriapoda, which also includes the Millipedes (Diplopoda) and two other multi-legged
arthropod groups, the Pauropods and Symphylans (ROSENBERG 2009). According to MINELLI (2011) there
are 3461 species of Centipedes described, while ADIS & HARVEY (2000) claim, there are at least 5000
species out there that have yet to be described. These numbers make the Centipedes a relatively small
group within the enormous phylum of arthropods, and yet they are quite diverse, show a wide variety of
adaptations, and are present in almost every habitat on every continent except Antarctica. They can all
be described as worm-like, multi-legged, chitinized inhabitants of the ground and litter-layers that vary in
size from a few millimeters up to more than 30 centimeters in length (ROSENBERG 2009). Despite their
name, no Centipede species has exactly one hundred legs, since all species show an odd number of leg
pairs. The number of individual legs varies from 30 to 354 depending on the genus. Inside the class of
myriapods, the Centipedes are an exclusively carnivorous clade of fast, rather strong, venomous
predators (LEWIS 2011).
Evolutionary and taxonomic history
The myriapods in general are a very old taxonomic group, with the earliest records of specimens dating
back to the late Silurian, 430 million years ago (SHEAR 1992). It is generally accepted today that the
insects and myriapods (together referred to as “Antennata”) made their way from sea to land two
separate times in evolutionary history. Convergent selection processes led to the myriapod ancestors
losing the first pair of antennae and slowly evolving a homonomic segmentation of their bodies. Thanks
to a strong drive of evolutionary forces at the end of the Silurian, a wide variety of prehistoric myriapod
species could already be found in the early Devonian. The records of these prehistoric myriapods also
present some of the largest species of land-dwelling Arthropods we know of. The genus Arthropleura
brought forward enormous, heavily armored specimens with a length of up to 2.3 meters (BRADDY et al.
2008). The heavily armored Arthropleura armata (MEYER 1854) were probably more closely related to the
Millipedes than Centipedes, but it has long been suggested by many scientists that they could have been
predators, a thesis that lacks evidence until this day (SCOTT; CHALONDER & PATERSON 1985). Records of
prehistoric Centipede species are present, but none of them represent enormous species. Predatory
Centipedes ever since filled the economic niche of ground- and litter-living macro predators and have
ever since been of greater importance in the detritus food-chain. Today, Centipedes are omnipresent in

7
almost all habitats in the world. Depictions of Centipede species can be found everywhere in recorded
history, especially in sacred texts of ancient Egypt and medical records of ancient Greek society, with the
earliest depictions dating back to our Cro-Magnon ancestors and their cave-paintings (LEWINSON &
LEWINSON 2004). It was Greek philosopher ARISTOTLE who first described Centipedes around 300 B.C.,
merely characterizing them as “wingless insects with several legs”. Hundreds of years later, in 1758,
French naturalist Carl LINNÉ used a similar description to distinguish between the genera Iulus and
Scolopendra, which LINNÉ both placed inside the order of the Aptera, or wingless insects (VERHOEFF 1928).
The genera Iulus and Scolopendra later turned into the taxonomic groups Diplopoda and Chilopoda we
use today. Modern zoologists like VERHOEFF, KOCH and ATTEMS dedicated themselves so intensively study
the myriapods, and thereby pioneered the descriptions and pre-genetic systematic order of these
animals, leading the way for today’s researchers to approach the order with modern scientific tools and
challenge the discoveries made decades ago.
Taxonomy and morphology
DNA-based research by EDGECOMBE & GIRIBERT (2002) has shown us, that the evolutionary gap between
the myriapod orders Diplopoda (Millipedes), Pauropoda and Symphyla is quite small, while there is a
huge gap between those orders and the Centipedes (Chilopoda). Together, the Diplopoda, Pauropoda
and Symphyla make up the taxon Progoneata, named after the shared position of the genital system
inside the animals’ bodies. The second characteristic these groups share, is their plant-, fungi- or
microbial-based diet (HOPKIN & READ 1992). The carnivorous Centipedes therefore have a different body
structure, one that is slicker, less stiff and makes them much faster and maneuverable.
EDGECOMB & GIRIBERT (2009) primarily distinguish the centipedes by their head-structure and branching of
the stigmata. We differentiate the Notostigmomorpha, who have short, single stigmata on the posterior
area of each dorsal plate, from the Pleurostigmomorpha, who feature deep-reaching, heavily branched
stigmata that are always present in pairs. The head structure of the Notostigmomorpha is rounded, and
(unlike all other myriapods) they feature compound eyes, and rather long, multi-segmented antennae.
This taxon only consists of one order: the Scutigeromorpha, also known as house centipedes, which
consist of 3 families, 2 of them only native to tropical areas. Only the family Scutigeridae is present in
Europe, with only one species: Scutigera coleoptrata LINNÉ 1758, thanks to its high abundance in human
populated areas also referred to as Common House Centipede. Within the Pleurostigmomorpha we find
all other taxa of Centipedes, with the main orders Lithobiomorpha (stone centipedes),
Craterostigmomorpha, Geophilomorpha (soil centipedes) and Scolopendromorpha (tropical centipedes).
There is very little known about the order Craterostigmomorpha, which is only represented by two
species found in Tasmania and New Zealand, and therefor will not be further discussed.

The Scutigeromorpha, along with the above mentioned Craterostigmomorpha, and the yet to be
discussed Lithobiomorpha make up the subphylum (or division) called Anamorpha. The group comprises
the centipedes whose number of body-segments increases as the animals mature, with the young
hatching with only 7 pairs of legs and the adults always having 15 pairs. In contrast, the young specimens
of the Epimorpha group (consisting of the Geophilomorpha and Scolopendromorpha) all hatch with their
full number of leg-pairs, which for the Scolopendromorpha is 21 or 23 (39/43 in the case of
Scolopendropsis duplicate EDGECOMB & GIRIBERT 2008) and anywhere between 25 and 177 for the
Geophilomorpha (SIRIWUT et al. 2016). Both the Lithobiomorpha, Craterostigmomorpha and
Geophilomorpha have a heteronomic body-segmentation, but while all body segments bear legs within
the stone centipedes, only every second segment bears legs within the soil centipedes. Only the
Scolopendromorpha show a homonomic body-segmentation, with every segment bearing a pair of legs.
The Scutigeromorpha have 15 segments, covered by seven tergites.
The house centipedes have compound eyes and long antennae composed of only two segments. All
other species of centipede feature a single Ocellus, a group of them, or don’t feature eyes at all. They
also feature multi segmented, (mostly) short antennae, which can be moved in all directions and serve as
sensory appendages. The heads of all centipedes are flat (except for the Scutigeromorpha), and feature
three pairs of mandibles, the latter two in the form of maxillae. The appendages of the first body-
segment have evolved into maxillipeds, in the form of strong pincers with venom glands, which serve
both to paralyze and catch prey as well as to defend against enemies. Except for the last pair of
appendages, all other extremities serve as legs. Like all other arthropods, centipede legs are made up of
several parts, and consist of a coxa, trochanter, femur, tibia and (depending on the genus) a monomial or
binomial tarsus, always equipped with a claw-like pretarsus. The last pair of legs usually serves a
different purpose and is always shaped differently from the other appendages. They usually serve as
either grabbing- or sensory extremities. The last body-segment only features a very small pair of
extremities, the gonopods, positioned close to the genital opening and used to deliver the
spermatophores or the placement of eggs. Only the Scolopendromorpha have no visible gonopods,
which are shaped like telescopes, and can therefore be retreated inside of the pregenital body-segment.
The gonopods are very diverse in shape, and therefore often used to distinguish even closely related
species. Especially the gonopods of the females are much longer and larger. They consist of three
segments, are mostly claw-shaped and often feature spurs. The gonopods of the males are much shorter
and stubbier and cover the extendable penis (ROSENBERG 2009).
One of the most interesting (and most discussed) structures of the centipede body are the coxal pores,
which are present (at least) on the last pair of extremities in almost every species in some shape or form.
Inside the Lithobiomorpha the pores are always present on the coxae of the last four leg-pairs, mostly in

9
low numbers, as a single row of elongated wells. Only few genera like Eupolybothrus feature smaller,
oval pores in much higher number, distributes scree-like on the coxae. We find similar patterns of coxal
pores within the orders Geophilomorpha and Scolopendromorpha, where the pores are not always as
easy to recognize as such. The functions and evolution of said pores has been a hot topic of discussion
ever since, and even though a multitude of studies tried to pin down its reason of existence, evidence is
sparse and often holey. Factually, the exact functions and biological meaning of said pores are yet to be
fully discovered, but today it is generally accepted, that the pores have some function in the water-
regulation of the centipedes, and serve as openings to control the intake and release of water vapor.
They might additionally release fragrances or secretes, but (most definitely) do not function as excretory
organs to release venoms, sticky secretes or slime to cover their bodies or eggs, as assumed by science in
previous decades (ROSENBERG 2009). To fully discover the functions and biological meaning of the coxal
pores, additional research and the application of new methods will be necessary. But as new are
constantly being developed and the field of science turning towards the analysis of arachnid- and insect-
secretes, we may answer this question sooner than we think.
Anatomy and physiology
The inner anatomy of centipedes is the same we find in all arthropods. They have a hemolymph which is
distributed inside the worm-like body by multiple ostia and the heart tube. A ventral nerve cord (NVC)
links the many extremities to a tritocerebrum (supraesophageal ganglion), thereby ensuring a well-
coordinated motor function. Excretion happens through a malpighian tube system, branching tubules
lined by chitin extending from the alimentary canal, found in all higher arthropods. Uric acid is produced
and released through the anal opening, water and ions are being reabsorbed at the rectum, a key
adaptation of land-dwelling arthropods, especially the thin-skinned species and those living in arid areas
such as deserts. As a fact, specimens of the Geophilomorpha lose water at a much slower rate than the
Lithobiomorpha, which may be contributed to the pleural membrane of the specimens, which shows
much higher levels of sclerotization. The size and shape of the stigmata, as well as the ability to curl, also
have an impact on the water regulation of the animals. Questionable, again, is the role of the coxal pores
in this instance. Primal myriapods also feature one or two pairs of plesiomorphic glands at the antennae
and maxillary nephridia, of the same kind found in the closely related Pancrustaceans. The exchange of
air functions just as it does within the class of insects, through trachea, which deliver oxygen directly to
the muscles (RUPPERT, FOX & BARNES 2004). As discussed earlier, the shape and position of the stigmata,
the openings of the trachea, vary from order to order and are therefore a taxonomic trait.

Mating and reproduction
Centipedes are ovipary creatures. The reproductive process happens without a real mating or
copulation. Instead the males place their spermatophores on a weave and leave it there for the females
to pick it up. A real pairing only happens within the Scutigeromrpha, where the males attract the females
with a dance, place their spermatophore on the ground once a female shows up, and in the process pull
the dancing female over the spermatophore, sticking it onto the body of the female. Very few species of
centipedes are also known to be parthenogenetic and produce offspring out of unfertilized eggs. All
other female specimens of centipedes pick up the spermatophores of the males and fertilize the eggs
inside of their bodies themselves. They then lay a small to moderate amount of eggs, ranging from ten to
50 within the Scolopendromorpha and Geophilomorpha, and even less within the Lithobiomorpha. The
Scutigeromorpha usually also lay between ten and 50 eggs, but within laboratories they have shown to
produce anywhere from 60 to 150 eggs, depending on the conditions. The egg deposition occurs in
temperate areas in spring and summer, with some species occurring earlier than others, and some
species even having two breeding events within one season. Parental care is not unusual within the
centipedes. The Geophilomorpha and Scolopendromorpha (as well as the Craterostigmomorpha)
females guard their eggs and young until several days after they hatch. The females curl around their
offspring in a protective state, with the Geophilomorpha curling around them with the dorsal side, with
the defensive secretory glands positioned to the outside, while the Scolopendromorpha (and
Craterostigmomorpha) curl around them with the ventral side, defending their offspring with the
(mostly) heavily chitinized dorsal plates. The timespan between the deposition of the eggs and the
hatching of the offspring is very variable from species to species and can take anywhere between one to
several months. Where parental care is happening, the mother will constantly lick her eggs and keep
them clean from any spores of fungi. Once the offspring hatches the young are quite inactive for a
couple of days and will only leave the protection of their mother after a couple of moultings at a more
active state. In the meantime, the young depend on their mother, and are being fed by her indirectly.
There are few species of Scolopendromorpha which are known to always be matriphagous, where the
body of the mother serves as food for her offspring. When the mother is disturbed by a predator, she
will either abandon her offspring or consume them. Abandoned eggs will quickly be destroyed by fungi
without the constant care of their mother. There is no evidence of any parental care within the
Scutigeromorpha and Lithobiomorpha. The females will place their eggs separate and either mask them
with litter material or place them inside of individual little holes in the ground (LEWIS 2007). Hatched
“larvae” of centipedes do look very similar to the adult individuals but are mostly transparent. Those of
the Lithobiomorpha, Craterostigmomorpha and Scutigeromorpha have fewer legs when they hatch, and
grow two or more pairs of legs with each moulting until they reach the full number of lags. The
Scutigeromorpha are born with only four pairs of legs, most Lithobiomorpha with seven and the

11
Craterostigmomorpha with 12. They usually reach their full number of leg pairs before they are fully
adult, since the last two to three moultings of the young serve to develop the gonopods, additional
antenna segments, different glands and some ocelli. The Gophilomorpha and Scolopendromorpha hatch
with the same number of legs they carry as an adult, and only develop the gonopods and additional
glands during each moulting until they are adults (LEWIS 2007). Usually centipedes reach sexual maturity
at an age of one year and are fully adult after one to three years. The time between the hatching of the
eggs and full adulthood can vary a lot from species to species and ecozone to ecozone, especially with
the larger species of Scolopendromorpha, who can grow to reach an age of multiple years. The average
lifespan of a centipede is somewhere between four to seven years, with Scolopendra subspinipes (LEACH
1816) reaching an age of over 10 years, according to YATES III. & JULIAN (1992). Since centipedes lay a
moderate amount of eggs, have quite long incubation times and need several years to become fully
adult, it is commonly accepted that they target K-selection, and do not target massive distribution of
their species, but rather seek to preserve a rather high number of individuals over multiple generations
(ALBERT 1979).
Systematics and species numbers
The Lithobiomorpha, or stone centipedes consist of two similar families, the Henicopidae and
Lithobiidae. The Henicopidae consists of 19 genera and approximately 120 species, while the Lithobiidae
consists of over 80 genera and around 1200 described species worldwide. The Scolopendromorpha, or
tropical centipedes, comprises five families. Even though the name suggests otherwise, the
Scolopendromorpha are not exclusively found in the tropics, but all over the world. The family
Mimopidae is the smallest, consisting of only two valid species of the order Mimops. The family
Plutoniumidae has only recently been established. There are now six species described, either from the
new genus Plutonium or the older taxon Theatops, both formerly contributed to the family Cryptopidae.
The family Cryptopidae is still quite large, and consists of almost 200 species, of which over 150 can be
contributed to the genus Cryptops. The family Scolocryptopidae consists of around 100 species, with the
two large genera Newportia (over 50 species) and Scolocryptops (20 species) contributing majorly. Lastly,
the family Scolopendridae consists of several hundred species. This family is where we find the largest
centipede species in the world, especially from the genus Scolopendra (around 100 species), which is also
where we find the only two amphibious centipedes we know of, Scolopendra cataracta SIRIWUT,
EDGECOMBE & PANHA 2016 and Scolopendra paradoxa DOMÉNECH 2018 (HOLMES 2016, BATES 2016). The
Geophilomorpha, also called soil centipedes, are another very diverse group, consisting of at least seven
families and almost 1300 species, making them the most diverse group of centipedes (MINELLI 2011). The
family Geophilidae (including the former Linotaneidae, Dignathodontidae and Macronicophilidae) alone

features over 120 genera. The remaining families Mecistocephalidae, Oryidae, Himantariidae,
Schendylidae, Zelanophilidae and Gonibergmantidae are all significantly smaller, but do add to the
extreme diversity found in soil centipedes (MYERS et al. 2013). Soil centipedes lack eyes, just like all
Cryptops and most Scolocryptops and Theatops species. Because they spend most of their life in the
ground or dark caves, they also sometimes lack pigmentation, or show reduced pigmentation and
chitinization in most areas of their bodies, save for the heads.
Distribution
Thanks to their far-reaching evolutionary history and prehistoric dominance, the centipedes can be
found all over the world, in almost every climate zone and altitude, with many species adapting to
extreme conditions such as extreme heat, high atmospheric pressure and regular flood events. Most
centipedes prefer wet, lush layers of litter as their habitat, while the tropical species live on and below
the massive vegetation of the rainforests, in caves as well as deserts, coastlines and even completely
underground. The larger species of the rainforests and desert areas usually have individual dens and lairs
inside of wood and vegetation or under rocks and larger systems of boulders (LEWIS 2007). Those species
native to central Europe also occur under rocks and dead wood but are scarce outside of these habitats
and rarely ever found on vegetation. Geophilomorpha usually live underground or inside the uppermost
layers of the ground, but there are several species that live in caves or inhabit rock systems, and a
handful of examples of species that dwell on coastlines made up from sand, gravel or boulders (LEWIS
1961). Those species occurring in higher altitudes usually show higher levels of pigmentation, a
phenomenon that can often be observed within the arthropods. There are no records of centipede
species in the snow zones of mountains or the Arctic regions, but there are species who live beyond the
arctic circle (RIEGER ET AL. 2010). Centipedes are generally sensitive to light and extreme heat but can
comprehend extreme heat much better than extreme cold. Centipedes occur in deserts, steppe areas
and savannas much more frequently than in the tundra, and there are no species present in any ice-
deserts (LEWIS 2007, ROSENBERG 2009). Those species who live in countries with cold winters like Austria,
survive the cold by retreating underground, inside protected microhabitats or spend the winter inside
dead wood or large boulders. Those centipedes that are more robust and thermophilic can also be found
in cities or man-made landscapes. These are mostly very common species with few preferences and
demands when it comes to their habitat. In central Europe these are species such as Lithobius forficatus
(LINNÉ 1758) and Scutigera coleoptrata (LINNÉ 1758), in tropical areas also the larger Scolopendra
specimens (LEWIS 2006). The thermophilic Scutigera coleoptrata (LINNÉ 1758) is often found in or near
houses in rural regions or submontane settlements and doesn’t seem to be bound to any specific
habitat. Scutigera are very active and fast hunters and can often be found under matrices, carpets and

13
pillows (RICKS 2015, LEWIS 2007). In contrast to Scutigera and some millipedes (i.e. Ommatoiulus
sabulosus (LINNÉ 1758)), Lithobiomorpha and Geophilomorpha are usually not found in houses, maybe
because they are bound to litter- and ground habitats. Native species of the Scolopendromorpha do also
appear in cities, gardens and parks, but often do not stay there for very long, unlike Scutigera does.
Especially the genus Cryptops finds at least two species in flowerbeds of large cities (even with neozoic
species like Cryptops rucneri MATIC 1967) and even Scolopendra cingulata LATREILLE 1829 can be found
breeding in gardens in the northeastern parts of our country.
Prey and predators
A key element for the presence or absence of centipedes is the abundance of possible prey and natural
enemies in a certain habitat (LEWIS 2007). Centipedes have no problem with the presence of conspecifics
or other species of centipedes in the same are, if there is enough prey for them to survive. They are less
frequent in areas where hunting-spiders and larger species of ants are present, but do live side by side
with millipedes, since they do not share a common prey but prefer similar habitats. While millipedes,
such as the Julidae, feed on fungi and dead woods, they produce perfect microhabitats for smaller
arthropods such as springtails (Collembola), which serve as a primary prey for smaller species of
centipedes, who therefore prefer to live in the very same microhabitat as its prey. They are predatory
generalists and hunt a multitude of different animals. They never consume plants but have shown to
feed on some dead plant material or fungi when brought close to starvation. They usually hunt and
consume all animals that are rather thin-skinned and not much larger than the centipedes themselves
(LEWIS 2007). The hypo- and endogenous Geophilomorpha mostly feed on smaller and larger species of
worms like Nematodes or smaller Oligochaeta, which are omnipresent in soil and easy to hunt and kill
(WEIL 1985). Thanks to their sizes, the giant centipedes within the order Scolopendromorpha are even
capable of hunting smaller vertebrates such as mice, small lizards and amphibians. They can catch bats in
Venezuelan caves out of the air by crawling onto the ceiling of the cave and letting the posterior parts of
their serpent-like bodies hang down. If they manage to grab a flying bat, they let themselves fall to the
ground and immediately paralyze the prey with a powerful bite and the injected venom (MOLINARI et al.
2005). Giant tropical centipedes such as Scolopendra gigantea (LINNÉ 1758) have even been reported to
feed on serpents, birds and bats, and there are even reports of snakes consuming entire Scolopendra
specimens alive which then proceeded to consume the snake themselves from the inside, eventually
hatching out of their predators’ body. Springtails and other small ground-arthropods serve as the main
prey for most small and mid-sized species of centipedes, but they are also intraguild predators, and are
known to hunt other centipedes, spiders and other competitors. While little is known about the

predatory behavior of the Scutigeromorpha and Craterostigmomorpha, we can assume that they are
similar to those of the Lithobiomorpha and Scolopendromorpha (LEWIS 2007).
Even though the most centipedes live a very cryptic life, they do have to face a lot of predators. They
themselves are hunted by mice, lizards and snakes, as well as birds and larger beetles. Interestingly
enough, these are the same groups of animals which serve as prey to the largest species of centipedes.
Larger centipedes are also a common prey of Mongoose. We know of two species of animals who
exclusively feed on centipedes: the African ant Stigmatomma1 pluto (GOTWALD & LEVIEUX 1972), feeding
exclusively on a species of Geophilidae, and the south-African snake Aparallactus capensis SMITH 1849
(LEWIS 2007). It has been observed time and time again that centipedes avoid the neighborhood of ants,
especially larger, colonial ants, probably because of the high aggressivity and precaution of those
species, who will attack and kill any potential danger to the colony. This claim is further supported by the
fact that millipedes can often be seen fairly close to colonial ants. The slow, xylobiontic millipedes aren’t
of any danger to the colony and might therefore be accepted as neighbors, unlike their fast, predatory
centipede relatives.
Venomousness and danger
Centipedes use their massive maxillipeds to kill their prey and defend against enemies. The maxillipeds
are formed as massive claws, which always feature glands filled with venomous substances, which
means that all centipede species are venomous, except for a few cryptic species which do not produce
venomous substances or even lack the glands to do so. In addition to these glands, the ventral plates of
the Geophilomorpha also feature microscopic glands which produce sticky secretes with venomous
components such as Benzoic acid and hydrogen cyanide. We assume that the Lithobiomorpha are also
able to produce such sticky secretes, but many descriptions of such glands date back to the time when
the above mentioned coxal pores were described as secretory glands, which they are certainly not
(EISNER T., EISNER M. & SIEGLER M. 2005). The biggest advantage of the centipedes is their immense
flexibility and speed. When being exposed to light, hearing an approaching predator or being disturbed,
they can retreat into the smallest spaces between rocks or underground in no time.
In the field of zoology, centipedes are often an overlooked group of arthropods, positioned somewhere
between the insects and arachnids, and mentioned sparsely next to venomous spiders and scorpions.
Despite that, centipedes have been known to be venomous predators just as long as spiders and
scorpions, with some species of the Scolopendra order even being of great danger to human beings
(KNOFLACH 2010). In ancient Egypt and Mesopotamia, we can find countless sculptures and depictions of
1 formerly genus Amblyopone.

15
centipedes in temples and medical reports, next to the classic images of serpents and snakes. The
ancient Egyptians even had a god by the name of Sepa resemble a centipede, who was known as a
protector against evil animals, and described as a god of death, disease and healing. NEWPORT (1844) was
the first scientist to describe the maxilliped claws of the centipedes, even though they have already been
depicted a few decades earlier in the zoological drawings of LEEUWENHOEK. The venom glands evolved
from an aggregate of epidermal glands along the cuticular canal. The glands are holocrine, and venom is
released through contraction of the muscles surrounding the glands (KNOFLACH 2010). How potent the
venom of the centipedes really is, has been a topic of discussion for decades. While most centipedes use
their venom to kill or paralyze worms, springtails and snails, larger specimens of the Scolopendromorpha
are even capable of paralyzing small vertebrates like mice, frogs, lizards, fish and bats (LEWIS 1981;
CARPENTER & GILLINGHAM 1984). The venom of Scolopendra species has undergone a lot of research in the
last decade. COHEN & QUISTAD (1998) were some of the first scientists to discover the key component of
the centipede venom. Those components are histamine, serotonin and different enzymes, which form a
complex protein mix that has both neuro- and cytotoxic attributes (GOMES et al. 1982; MOHAMED et al.
1983). In 2006 WENHUA et al. were able to isolate an unknown peptide from the venom of Scolopendra
subspinipes (LEACH 1816), given the name Scolopendrin I., and shown to have immobilizing or even
destructive effects on a wide variety of different microorganism. In 2009 GONZALES-MORALES et al.
detected the first phospholipase in the venom of Scolopendra viridis SAY 1821, and studies by RATES et al.
from 2007 revealed that at least 12 components of the Scolopendra venom have shown to be highly
toxic to most insects. A controversial topic has ever since been the effect of the Scolopendra venom on
humans. Reports of cases of death caused by centipede venom date back to the first century A.D. but are
met by modern scientists with much disbelief (BÜCHERL 1946, 1971; MINELLI 1978; LEWIS 1981). Only one
incident reported by PINEDA (1923) is to be believed. The reported death of a small child in the
Philippines caused by a bite of Scolopendra subspinipes (LEACH 1816) is accepted as fact by multiple
authors (REMINGTON 1950; BUSH et al. 2001) who examined the case. SERINKEN et al. (2005) report the
death of a man from Turkey who died after being bitten by a centipede, but explain that his death was
actually caused by an infection after the bite, not the venom or the bite itself. All other modern reports
of centipede bites do not suggest any lethal effects on mankind at all, not even the bites of the
enormous Scolopendra gigantea LATREILLE 1829 (REMINGTON 1950; SOUTHCOTT 1978; MUMCOUGLU &
LEIBOVICI 1989; MOHRI et al. 1991; RODRIGUEZ-ACOSTA et al. 2000; BARROSO et al. 2001: BUSH et al. 2001;
BOUCHARD et al. 2004; GUERRERO 2007; VARIANZADEH et al. 2007; IVOCHEVA et al. 2008). Bites of larger species
of centipedes are reported to be extremely painful and have a long-lasting burning sensation. They also
provide a large risk of dangerous inflammation and infections (KNOFLACH 2010). Bites of smaller species
are often compared to bites of ants and usually only hurt for a minute without leaving any scars for
inflammations behind. Reports of bites from Geophilomorpha are sparse, but there are several species

whose bites are known to be quite painful, often compared to the sting of a bee or wasp.
Scutigeromorpha bites are also not reported a lot, which seems odd, since they are the most abundant
species of centipedes in houses, and are known to be the most “aggressive” centipedes, or rather the
most likely to bite in defense, and release a rather large amount of venom, which results in a very
unpleasant bite, though the pain only last for a short amount of time. In general, unlike spiders and
snakes, centipedes use their venom very sparingly, and only in extreme emergencies. They are regarded
as fairly aggressive animals and use their venom mainly to paralyze and kill their prey. It is assumed that
centipedes are capable of biting without releasing any venom, which further supports the claim that
centipede venom is extremely precious (ASPÖCK 2010). Most native species of centipedes are too small to
even penetrate human skin and are therefore regarded as completely harmless. Only bites of Scutigera
coleoptrata (LINNÉ 1758) and Geophilus electricus (LINNÉ 1758) are unpleasant, with the latter species
barely coming in contact with humans.
Concerning Barcoding
DNA barcoding is a novel system designed to provide rapid, accurate, and automatable species
identifications by using short, standardized gene regions as internal species tags (HEBERT & GREGORY
2005). VALENTINI, POMPANON & TABERLET (2008) described the rise of barcoding as a tool for ecologists as a
new method that has received much attention, and will increasingly be used to not only identify a single
species from a specimen or its remains, but also determine the species composition of environmental
samples. Ecologists can even take advantage of DNA tools when only hair, feces or urine left behind by
animals are available for species identification. As a consequence, it will make the Linnaean taxonomic
system more accessible, with benefits to ecologists, conservationists, and the diversity of agencies
charged with the control of pests, invasive species, and food safety. VALENTINI et al. (2008) further claim
that “short DNA fragments persist in the environment and might allow an assessment of local
biodiversity from soil or water”. The ideal DNA barcoding marker has not yet been found or may not
even exist, because the marker would have to meet a wide variety of criteria:
(i) The gene region sequenced should be nearly identical among individuals of the same
species, but different between species.
(ii) It should be standardized, with the same DNA region used for different taxonomic groups.
(iii) The target DNA region should contain enough phylogenetic information to easily assign
unknown or not yet ‘barcoded’ species to their taxonomic group (genus, family, etc.).
(iv) It should be extremely robust, with highly conserved priming sites and highly reliable DNA
amplifications and sequencing (particularly important when using environmental samples,
containing a mixture of many species to be identified at the same time).

17
(v) The target DNA region should be short enough to allow amplification of degraded DNA.
Usually, DNA regions longer than 150 bp are difficult to amplify from degraded DNA.
The authors also note that, different groups of users will give different priorities to those categories: “for
taxonomists, criteria (i), (ii) and (iii) will be more important […], whereas ecologists working with
environmental samples will favor criteria (iv) and (v) […]”. The approach most taxonomists take is
barcoding sensu stricto, whereas ecologists favor a sensu lato approach. The now well established
initiative CBOL (consortium for the barcode of life) has been designed to fit with DNA sequencers based
on capillary electrophoresis, which yield a typical read length of 500–1000 bp. CBOL proposes the 658 bp
gene region encoding the mitochondrial cytochrome c oxidase 1 (COI) for standardized barcoding of
animals. Yet the very characteristics that make the COI gene a candidate for high throughput DNA-
barcoding, also limits its information content on a deeper phylogenetic level, as HEBERT et al. (2003) point
out. According to MORITZ & CICERO (2004) DNA barcoding should therefore not be misinterpreted as a
tool to resolve the “tree of life”. More and more sequence data for the accepted barcoding markers are
becoming available in public databases as sequencing facilities improve. VALENTINI et al. (2008) note: “[…]
the quality of the sequence data in GenBank, EMBL or DDBJ is not always perfect, either as a result of
sequencing errors, contaminations, sample misidentification or taxonomic problems”. CBOLs own
database specially dedicated to DNA barcoding called BOLD (Barcode of Life Data Systems) provides an
efficient and accurate way for species identification. Not only does the database record DNA sequences
of several individuals per species, but also contains complete taxonomic information, place and date of
collection, and specimen images.
The “Barcode of Life” Projects2
ABOL – the Austrian Barcode of Life, is an initiative that aims to generate a public database of DNA-
barcodes of all native lifeforms, by building a national network of researchers in the field of biodiversity.
Thereby the platform additionally promotes all national biodiversity research and coordinates related
projects all over the country. It has been designed as a long-term initiative of at least 10 years and began
its run in 2017, after a 3-year pilot phase from 2014 to 2017. Initiated and coordinated by the Museum
of Natural History in Vienna, its pilot phase and coordination is being financed by the Austrian Federal
Ministry of Education, Science and Research, and involves all relevant institutions involved in biodiversity
research, such as universities, museums, biosphere parks and several different research institutions in
addition to federal conservation departments and regional scientific societies. The initiative consists of
group-specific clusters, characterized by a group of experts of each specific group of lifeforms or specific
2 Source: www.abol.ac.at

orders of lifeforms. Inside the ABOL project is a specific cluster of myriapods and basal hexapods,
coordinated by Dr. Nesrine Akkari and Mag. Daniela Bartel of the Museum of Natural History in Vienna.
The coordination team of ABOL also aims to stay in contact with other national barcode-of-life-initiatives
such as iBOL, GBOL and CROBOL. Like all other BOL-initiatives, ABOL stands for quality standards. The
specimens from which the DNA is extracted have been legally collected and are deposited in a publicly
accessible scientific collection. DNA-barcodes and corresponding required meta-data are fed into the
ABLO-database. All work from the collection to the documentation, sampling and evaluation process are
carried out obeying highest carefulness and meeting laboratory standards. Remaining DNA or tissue
samples are being stored in a collection, and all barcodes are incorporated into the international BOLD©
database.

19
PART I.
RED LIST OF CENTIPEDES
State of the art
Native centipedes have been studied ever since the days of Austrian entomologist Robert LATZEL, who in
1880 published his monography on the myriapod-fauna of the then Austro-Hungarian empire. LATZEL was
the first Austrian scientist to build upon the works of pioneers such as Pierre André LATRILLE, who earlier
published works on the fauna of Austria, Hungary and parts of Czechoslovakia. LATZELS work, with
contributions by naturalists Carl Ludwig KOCH and Frederik Vilhelm August MEINERT who described
several new species native to Austria, thereby filled a huge gap in European faunistic research. LATZEL
took KOCHS and MEINERTS descriptions and documentations and combined them into one collective work,
which to this day is regarded as the most important collection of descriptions for native myriapods. The
collection consists of 130 taxa of Diplopoda (1 genus, 2 subgenera, 69 species and 56 forms) and over 40
groups of Chilopoda (2 genera, 29 species and 12 forms), as well as 4 taxa of both the Pauropoda and
Symphyla. In 1884 LATZEL published his second work on the Pauropoda, Symphyla and Diplopoda,
another work of huge importance in the field of myriapod systematics and distribution. LATZEL also
turned out to be a pioneer in the field of gonopod-research and was the first to realize that size and
shape of the gonopods are the best and often only way to determine certain species of myriapods. LATZEL
thereby majorly contributed not only to the field of Austrian faunistics, but also to the field of
entomology globally. His work was so far reaching, there have only been a dozen new native species of
centipedes described since LATZELS days. LATZELS work has been described by other entomologists as
extremely detailed and highly accurate, and his collection of myriapods is regarded as one of the most
important collections of the museum of natural history in Vienna, containing several type-species.
LATZELS impact on the field of myriapod-research will remain unforgotten, thanks to a group of
researchers who gave a group of house-centipedes the taxonomic name Latzelia. LATZEL himself always
knew there were more species to be discovered, and wanted other naturalists to become “friends” of
the myriapod-group. Only then, as LATZEL himself wrote: “[…] can we expect to achieve completion”.
LATZEL himself only published two more works in the following years, both on the myriapod-fauna of
Carinthia, and worked on the index of myriapods of Tyrol with HELLER and DELLA TORRE.

The next naturalist to contribute to the Austrian myriapod-research was German zoologist Karl Wilhelm
VERHOEFF, who worked as a lecturer at the University of Bonn in Germany at the time, where he
previously graduated. He traveled through most of the southern lands like Istria, Dalmatia and Carniola,
and visited parts of Tyrol and Styria. With his works VERHOEFF mainly aimed to describe more and more
species of myriapods, and in the process learned a lot about the affinity of certain species. At the same
time, Austrian monarch and zoologist Carl August of Attems-Petzenstein (a.i. Carl ATTEMS) began his
private studies on Styrian myriapods. ATTEMS wanted to explore the fauna of Styria, and previously
studied the gonopods of Polydesmida. In 1895 he published a revised work on the fauna of Styria, which
also featured numerous publications and findings from Lower Austria. With VERHOEFFS help, ATTEMS
described over 1800 new species of myriapods, which he collected on numerous private excursions all
around the world and published 138 scientific papers on the topic. In the following years more and more
publications described new and known species of Polydesmida native to Austria and Germany, all
building upon the foundations laid by ATTEMS, who later included said descriptions in his work on the
systematics of Styrian Polydesmida (HÖLDER 1901).
Edward Holt EASON was the next naturalist to describe several new species of myriapods in the 1960’s,
mainly species of the British Isles. He published an extraordinarily accurate determination key for
centipedes, including some of the best zoological drawings of centipede bodies and gonopods to date.
Several excellent works on the biodiversity of Austrian centipedes existed by the 1960’s, what was
missing is a uniform collection, to combine all the scattered descriptions and information that would also
clarify synonymic species, unknown species and establish a uniform taxonomy. We got that with the
infamous “Catalogus Faunae Austriae”, published in 1972 by naturalist Marcus WÜRMLI, who included a
uniform determination key like the one of EASON.
Another notable work was published in the early 1990’s by biology-professor and zoologist Armin KOREN,
who collected several species of centipedes in Carinthia and published two determination keys for the
native species, which included drawings by EASON and a large amount of new drawings by KOREN himself.
On the basis of his own publications, KOREN later with the nature conservation union and local
government, worked on Carinthias red list of centipedes, the very first red list for centipedes of Austria.
Unfortunately, the list was not very detailed, and left out questionable species and other key elements
like the responsibility for the species and abundance of the species in neighboring countries and states.
With the rise of phylogenetic methods in the last decade, the field of systematic zoology changed its
face. The systematic system that has been valid for centuries was changed from its foundations, thanks
to discoveries from genetic studies, which by now have mostly replaced the classic morphological
approach to systematics. This also impacted the field of entomology and thereby the systematics of
myriapods. Today, more and more new species are being found and described every year, while

21
phylogenetic studies indicate that different previously described species are either one and the same or
represent either sub-species or variates. This “enlightenment” led to a fundamental problem, which is
(again) the establishment of a uniform and valid systematic system. For many years this has gotten more
difficult, especially since many scientists today rely on genetic data only, even if its results heavily
contradict morphological discoveries. The growing number and lack of definition of sub-species further
complicates the establishment of a uniform system. A system that combines morphological and
phylogenetic evidence is a difficult task many entomologists face.
Since it is almost impossible to keep a record of any larger centipede population (or its area of
distribution), and the fact that this has not been done in the last 25 or more years, there is no red list of
Centipedes released under the umbrella of the IUCN. The IUCN has only evaluated ten species of
Centipedes as of January 2020, all of them of tropical origin, mostly from the Seychelle Islands. There are
checklists and regional red lists of centipedes from several German states (SPELDA 1999, 2004, 2005;
VOIGTLÄNDER 2004, 2005; REIP & VOIGTLÄNDER 2009; DECKER & HANNING 2011), as well as one for the country
of Germany based on said lists (GRUTTKE et al. 2016). Most of these lists work with data from findings and
only give abundances and responsibilities. Same goes for the only red list of centipedes in Austria from
1990, where KOREN lists the native centipedes by abundance and habitat preference to evaluate the
endangerment of at least those species with enough records.
The Austrian soil conservation law has been part of the nature conservation decree ever since its
initiation (BGBl. Nr. 491/1984), but is directed by the nine states, not the nation. The importance of its
quality and faunal diversity is often underestimated. Today we have a much clearer vision of the
biodiversity of cryptic taxa such as Collembola, Acari and Diplura, but where the taxonomy becomes
clearer every day, very little is known about the distribution and abundance of these groups of animals.
The idea behind creating a full checklist and red list of Carinthian and Styrian centipedes is to raise
awareness of the biodiversity of the taxa, importance of some of its species (especially endemic species)
and create a better understanding of the biology, ecology and role of centipedes in the environment.
Some species have the potential to be used as indicators in environmental studies or nature
conservation surveys.
Future friends of these taxa can improve upon this work and aim to one day evaluate population status
and distribution of these species throughout Austria. Only then can we work on a cohesive red list of
centipedes following IUCN criteria.

Methodology
The south of Austria is the most studied part of the country when it comes to centipedes, and therefore
shows the highest density of records and descriptions of said animals. According to RABITSCH & ESSL
(2009) the level of records of Carinthia and Styria is almost on par with those of the “British Myriapod
Survey”, yet the rest of the country is far behind. There are around 70 species of centipedes present on
Austrian soil, though statements regarding that number may vary. Pinning down the exact number of
native species is much more challenging than it would seem to be. With the rise of internet databases
such as Fauna Europeae (ENGHOFF 2007) and ChiloBase (MINELLI 2006) many species underwent
taxonomic revision. Species such as Geophilus noricus VERHOEFF 1928 and Geophilus pauropus ATTEMS
1927, which only a decade earlier had just been distinguished and morphologically described as separate
by KOREN, completely disappeared and are ever since synonymous with a third species, Geophilus
oligopus (ATTEMS 1895). Many of the native subspecies described by WÜRMLI as endemic met the same
fate. There are some species described as native that lack proper description or records, while others are
heavily implied to be native but have not been found yet. The exact species count an author will give
heavily depends on whether or not he will count said species as native or not, and if the author agrees
with all the synonyms made by ENGHOFF, MINELLI and others in the early 2000’s.
For this thesis, the number of valid described species for Austria can be set somewhere between 60 and
70. Of these species between 50 and 60 can be found in the counties of Carinthia and Styria. The species
count of the two neighboring counties is almost the same, and either one could practically have the
higher amount of species. For the county of Carinthia, we set the number at 52 certain species and an
additional 2 species with unclear status. For Styria, the count is 51 certain species with an additional 2
uncertain species. The “certainty” of said species of course is never 100%, leaving at least some leeway
as is always the case in taxonomy. With 59 described species and possibly even 60, the two counties
could be home to over 90% of all centipede species native to Austria.
A quantitative comparison and statistical analysis based on the number of individuals in a certain area is
practically impossible when it comes to centipedes. A solid basis for any analysis is the number of
discovery sites. In addition to the number of sites the bond of a certain species to a specific habitat
should be considered for evaluations. For stenoecious species this proves to be particularly difficult, as
very little is known about the preferences of most of these animals (GRUTTKE et al. 2016). Especially with
these “rare” species, another problem arises. An unconfirmed or missing entry of a discovery site may
have a large impact on any statistical evaluation of species with few localities, which (in reality) does not
impact the overall trend of the population.

23
Due to the low degree of discovery, it is impossible to formulate any trend criteria for a population. As
centipedes are k-strategists, population fluctuations are common, often extreme, and almost
unpredictable (GRUTTKE et al.. 2016).
Disclaimer
The following chapter contains the two red lists for the counties of Carinthia and Styria respectively. For
the county of Carinthia, part of the neighboring Eastern Tyrol was included for all statistical analysis.
Other authors including KOREN (1986, 1992) did the same thing, because Carinthia and Eastern Tyrol
share the same ecogeological and meteorological area.
In order to create a cohesive and logical checklist of native centipedes, research was done using the
currently valid taxonomy used on the website Chilobase© (MINELLI 2016), distribution data of native
centipedes found on the website Zobodat (REICHL 1972) and the works of KOREN (1986, 1992), and
additional information from the website FaunaEuropeae©. The author did not concern himself with the
subject of subspecies, but comments regarding important subspecies have been added to the red list
separately. One of the major problems creating any checklist of species are synonyms. Centipedes have a
long history of countless revisions and name changes. The most encountered synonyms of the native
species have been added to the list, while a complete list of synonyms can be found in the appendices.
The states of Carinthia and Styria have been divided into units of squares by drawing lines between the
latitudes and longitudes. A square was “occupied” whenever data of a species’ presence between its
borders was available. To formulate statements on the abundance of a species, the total number of
occupied spaces (squares containing data) per state was taken as reference (Figure 1, p. 27). A 5-step
logarithmic scale was formulated to determine a species’ distribution from tiny scale (1) to omnipresent
(5). For category (5) the number of occupied squares necessary was set at 80, for category (1) at least 2.
Species with data from only one square were put in category (0), a category for species with deficient
data.
The two counties have been divided into 3 geographical subareas, all covering approximately the same
number of squares for further analysis of distribution. These areas were used as additional indicators of
the species’ distribution. If a species is present in all three areas, its category will be upgraded by one (for
example going from category (2) to (3) with less than 15 occupied squares), if a species is only present in
one area its category will be downgraded by one (going from category (3) to (2) despite occupying over
15 squares). There is no downgrading a species from categories (1) to category (0), as only species with
less than 2 records will be put in the latter. There is also no upgrading for species with more than 80

occupied squares that are present in three areas. Said species will feature a (!) icon next to their
category, indicating the impossibility of an up- or downgrade.

25
Results
Checklist of Carinthia
Eupolybothrus grossipes (C. L. KOCH, 1874)
Eupolybothrus tridentinus (FANZAGO, 1874)
Harpolithobius anodus (LATZEL, 1880)
Lithobius aeruginosus* C. L. KOCH, 1862
Lithobius agilis C. L. KOCH, 1874
Lithobius austriacus* (VERHOEFF, 1937)
Lithobius borealis MEINERT, 1868
Lithobius burzenlandicus VERHOEFF, 1931
Lithobius castaneus NEWPORT, 1844^
Lithobius crassipes* C. L. KOCH, 1862
Lithobius denatus C. L. KOCH, 1844
Lithobius erythrocephalus C. L. KOCH, 1847
Lithobius forficatus (LINNÉ, 1758)
Lithobius lapidicola MEINERT, 1872
Lithobius latro MEINERT, 1872
Lithobius lucifugus C. L. KOCH, 1862
Lithobius macilentus C. L. KOCH, 1862
Lithobius melanops NEWPORT, 1845
Lithobius microps MEINERT, 1868
Lithobius moellensis VERHOEFF, 1940 ?!
Lithobius mutabilis C. L. KOCH, 1862
Lithobius muticus C. L. KOCH, 1847
Lithobius nodulipes LATZEL, 1880
Lithobius pelidnus HAASE, 1880
Lithobius piceus C. L. KOCH, 1862
Lithobius punctulatus C. L. KOCH, 1847 ?
Lithobius pygameus LATZEL, 1880
Lithobius subtilis LATZEL, 1880~
Lithobius tenebrosus MEINERT, 1872
Lithobius tricuspis MEINERT, 1872
Lithobius validus MEINERT, 1872
Lamyctes emarginatus (NEWPORT, 1844) ~
Eurygeophilus pinguis (BRÖLEMANN, 1896)
Clinopodes flavidus C. L. KOCH, 1846
Stenotaenia linearis (C. L. KOCH, 1835)
Geophilus alpinus MEINERT, 1869
Geophilus electricus (LINNÉ, 1758)
Geophilus flavus (DE GEER, 1778)
Geophilus oligopus (ATTEMS, 1895)
Geophilus pygmaeus LATZEL, 1880
Henia illyrica (MEINERT, 1870)
Henia vesuviana (NEWPORT, 1845)
Strigamia acuminata (LEACH, 1815)
Strigamia crassipes (C. L. KOCH, 1835)
Strigamia transsilvanica (VERHOEFF, 1928)
Pachymerium ferrugineum (C. L. KOCH, 1835)
Schendyla carniolensis VERHOEFF, 1902
Schendyla nemorensis (C. L. KOCH, 1837)
Schendyla tyrolensis (MEINERT, 1870)
Dicellophilus carniolensis (C. L. KOCH, 1847)
Cryptops hortensis (DONOVAN, 1810)
Cryptops parisi BRÖLEMANN, 1921
Cryptops rucneri MATIC, 1967
Scutigera coleoptrata (LINNÉ, 1758)
52 confirmed species (54 total)
*Subgenus Monotarsobius (previously Genus) ~ not yet recorded, most definitely native ? Taxonomy uncertain ?! Taxonomy uncertain, possibly endemic ! endemic to Austria !! endemic to the state

Checklist of Styria
Eupolybothrus fasciatus (NEWPORT, 1845)
Eupolybothrus grossipes (C. L. KOCH, 1874)
Eupolybothrus tridentinus (FANZAGO, 1874)
Harpolithobius anodus (LATZEL, 1880)
Lithobius aeruginosus* C. L. KOCH, 1862
Lithobius agilis (C. L. KOCH 1874)
Lithobius anisanus VERHOEFF, 1937
Lithobius austriacus* (VERHOEFF, 1937)
Lithobius borealis MEINERT, 1868
Lithobius burzenlandicus VERHOEFF, 1931
Lithobius castaneus NEWPORT, 1844 ~
Lithobius crassipes* C. L. KOCH, 1862
Lithobius curtipes C. L. KOCH 1847
Lithobius denatus C. L. KOCH, 1844
Lithobius erythrocephalus C. L. KOCH, 1847
Lithobius forficatus (LINNÉ, 1758)
Lithobius franzi ATTEMS, 1949 !!
Lithobius lapidicola MEINERT, 1872
Lithobius latro MEINERT, 1872
Lithobius lucifugus C. L. KOCH, 1862
Lithobius macilentus C. L. KOCH, 1862
Lithobius macrocentrus ATTEMS, 1949 !
Lithobius melanops NEWPORT, 1845
Lithobius microps MEINERT, 1868 ~
Lithobius mutabilis C. L. KOCH, 1862
Lithobius muticus C. L. KOCH, 1847
Lithobius nodulipes LATZEL, 1880
Lithobius peggauensis VERHOEFF, 1937 ?!
Lithobius pelidnus HAASE, 1880^
Lithobius piceus C. L. KOCH, 1862
Lithobius pygameus LATZEL, 1880
Lithobius tenebrosus MEINERT, 1872
Lithobius tricuspis MEINERT, 1872
Lithobius validus MEINERT, 1872
Lamyctes emarginatus (NEWPORT, 1844)
Clinopodes flavidus C. L. KOCH, 1846
Stenotaenia linearis (C. L. KOCH, 1835)
Geophilus alpinus MEINERT, 1869
Geophilus electricus (LINNÉ, 1758)
Geophilus flavus (DE GEER, 1778)
Geophilus oligopus (ATTEMS, 1895)
Geophilus pygmaeus LATZEL, 1880
Strigamia acuminata (LEACH, 1815)
Strigamia crassipes (C. L. KOCH, 1835)
Strigamia transsilvanica (VERHOEFF, 1928)
Pachymerium ferrugineum (C. L. KOCH, 1835)
Schendyla carniolensis VERHOEFF, 1902
Schendyla nemorensis (C. L. KOCH, 1837)
Schendyla tyrolensis (MEINERT, 1870)
Dicellophilus carniolensis (C. L. KOCH, 1847)
Cryptops hortensis (DONOVAN, 1810)
Cryptops parisi BRÖLEMANN, 1921
Scutigera coleoptrata (LINNÉ, 1758)
51 confirmed species (53 total)
*Subgenus Monotarsobius (previously Genus) ~ not yet recorded, most definitely native ? Taxonomy uncertain ?! Taxonomy uncertain, possibly endemic ! endemic to Austria !! endemic to the state

Statistical analysis
DATA:
Carinthia 9.536 km² Area 1 Area 2 Area 3 Eastern Tyrol 2.020 km² 3 360 squares 123 109 128 Styria 16.401 km² 544 squares 180 203 161
27957 km² 904 squares OCCUPIED SQUARES
4 Carinthia 189 47 47 63 Styria 163 63 72 26
352 STUDIED AREA IN % Carinthia 52,5 Styria 30,3
39
SPECIES NO. OF EACH RL CATEGORY
Cat. CAR STY CAR % STY %
0 6 10 11,5 19,6
1 5 7 9,6 13,7
2 6 3 11,5 5,9
3 17 13 32,7 25,5
4 13 10 25,0 19,6
5 5 8 9,6 15,7
Sum 52 51 100 100
3 not the entire area of Eastern Tyrol was considered for analysis, only the easternmost parts, approximately 1/3 of
the actual area. 4 Geographical subunits containing data of findings within its borders (Graphic 1).
Graphic 1: Areas with collection data in both Carinthia (left) and Styria (right).

Legend of the Red List
RED LIST CATEGORIES
Table 1: Categories and criteria of the red list
0 1 2 3 4 5
Occupied squares = 1 ≥ 2 ≥ 4 ≥ 15 ≥ 40 ≥ 80
Distribution category - tiny scale small scale normal spacious omnipresent
Abundancy category
Data deficient very rare rare moderate common very common
ADDITIONAL SYMBOLS
↑ = Category upgraded (species present in 3 areas),
↓ = category downgraded (species present in only 1 area)
! = category should be down-/upgraded, but not possible (species is already in Cat. 1 or 5)
ECOLOGY CATEGORIES
Temp = preferred temperature, Hum = preferred humidity, Alt = preferred altitude, Soil = preferred soil
acidity, Hab = preferred habitat
BOND CATEGORIES
? = questionable, ! = strong, 0 = low/none, !! = very strong5
1 2 3
Temperature cold neutral warm
Humidity low normal high
Altitude colline/sub mountain mountain/subalpine alpine/nival
Soil alkine none/either or acidic
1 2 3
Habitat open land, gardens, anthropologic areas
litter, rocks, wood, mixed habitats
deciduous forests, pine forests, mixed forests
5 Only used in Habitat category.
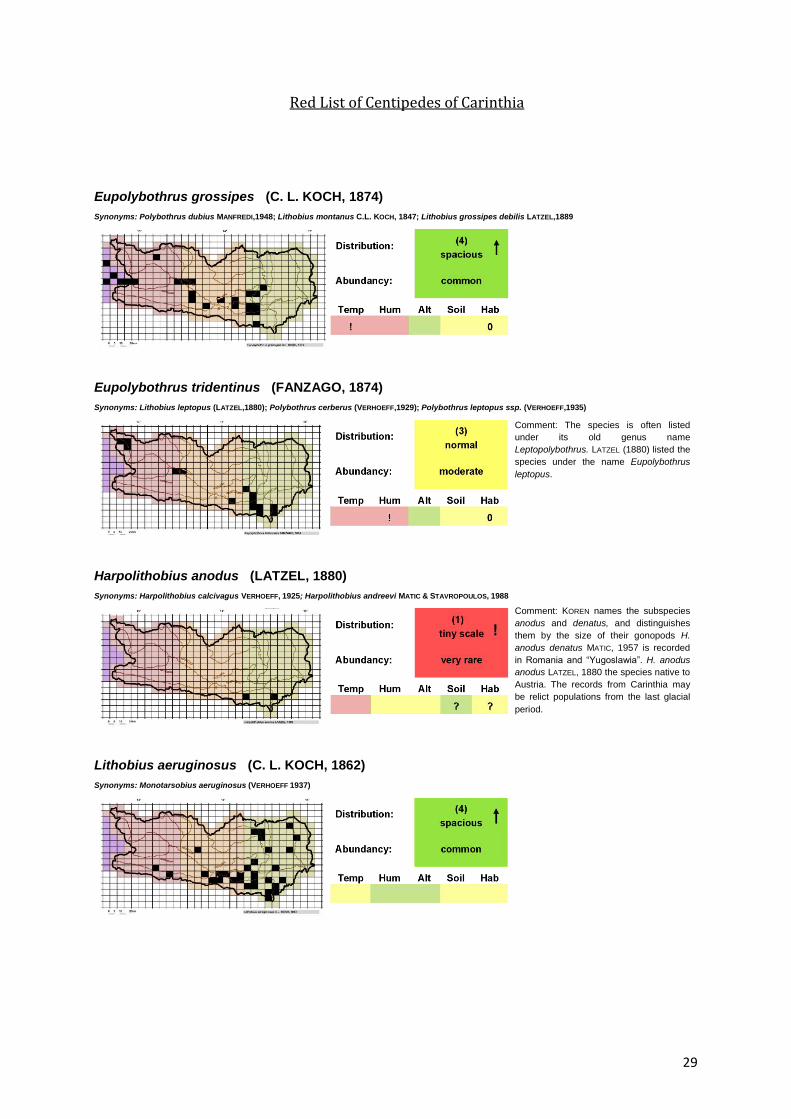
29
Red List of Centipedes of Carinthia
Eupolybothrus grossipes (C. L. KOCH, 1874)
Synonyms: Polybothrus dubius MANFREDI,1948; Lithobius montanus C.L. KOCH, 1847; Lithobius grossipes debilis LATZEL,1889
Eupolybothrus tridentinus (FANZAGO, 1874)
Synonyms: Lithobius leptopus (LATZEL,1880); Polybothrus cerberus (VERHOEFF,1929); Polybothrus leptopus ssp. (VERHOEFF,1935)
Harpolithobius anodus (LATZEL, 1880)
Synonyms: Harpolithobius calcivagus VERHOEFF, 1925; Harpolithobius andreevi MATIC & STAVROPOULOS, 1988
Lithobius aeruginosus (C. L. KOCH, 1862)
Synonyms: Monotarsobius aeruginosus (VERHOEFF 1937)
Comment: The species is often listed
under its old genus name
Leptopolybothrus. LATZEL (1880) listed the
species under the name Eupolybothrus
leptopus.
Comment: KOREN names the subspecies
anodus and denatus, and distinguishes
them by the size of their gonopods H.
anodus denatus MATIC, 1957 is recorded
in Romania and “Yugoslawia”. H. anodus
anodus LATZEL, 1880 the species native to
Austria. The records from Carinthia may
be relict populations from the last glacial
period.

Lithobius agilis C. L. KOCH, 1874
Synonyms: Lithobius paradisiacus MATIC & DARABANTU, 1971
Lithobius austriacus VERHOEFF, 1937
Synonyms: Monotarsobius austriacus (VERHOEFF, 1937)
Lithobius borealis MEINERT, 1868
Synonyms: Lithobius lusitanus würmanus (VERHOEFF 1937); Lithobius lapidicola (LATZEL 1880)
Lithobius burzenlandicus VERHOEFF, 1931
Synonyms: Lithobius gridellii MANFREDI, 1955; Monotarsobius veronensis MATIC & DARABANTU,1971
Lithobius castaneus NEWPORT, 1844
Synonyms: Lithobius lucasi NEWPORT, 1849; Lithobius meridionalis FEDRIZZI, 1877; Monotarsobius remyi VERHOEFF,1943
Comment: Very closely related to
Lithobius aeruginosus.
Comment: KOREN claims that the two
subspecies burzenlandicus and euxinicus
are recorded in Romania, while the
subspecies carinthicus is native to
Carinthia. Findings from Slovenia only
name the species with no subspecies
(KOS 1988). The findings of KOREN
generally match those made by MATIC in
1966. The valid name for the native
species is L. burzenlandicus carinthicus
nov. ssp. and can only be found in
Carinthia.
Comment: Records of this species only
exist from several areas in Carinthia, but it
is most definitely also native to Styria and
possibly even Burgenland. The species is
very circummediterrian and is distributed
throughout northern Italy, Slovenia and
western Hungary. According to MINELLI
(1987) the species inhabits temperate
oak-beech forests.

31
Lithobius crassipes C. L. KOCH, 1862
Synonyms: Lithobius (Monotasobius) crassipes C. L. KOCH, 1862; Lithobius atrifrons SILVESTRI, 1896, Lithobius podokes ATTEMS, 1903
Lithobius dentatus C. L. KOCH, 1844
Synonyms: Lithobius alpestris LATZEL, 1880
Lithobius erythrocephalus C. L. KOCH, 1847
Synonyms: Lithobius armatus SSELIWANOFF, 1880; Lithobius dubius TÖMÖSVÁRY, 1880; Lithobius dubius TÖMÖSVÁRY, 1880
Lithobius forficatus (LINNÈ, 1758)
Synonyms: Lithobius leachii NEWPORT 1844; Lithobius vulgaris LEACH, 1817
Lithobius lapidicola MEINERT, 1872
Synonyms: Lithobius pusillus (LATZEL 1880); Lithobius sulcatus (C. L. KOCH 1862)
Comment: Originally described as L.
dubius by TÖMÖSVARY, in 1880, L.
erythrocephalus has been described by
many authors all throughout Europe. In
many places this species is distinguished
into two sympatric subspecies. The
subspecies L. erythrocephalus schuleri
(MATIC, 1966 schulleri) described by
VERHOEFF (1925, 1937) is much more
common in Austria, but the exact
taxonomy of the species and its
subspecies is in need of revision.
Comment: The “common stone centipede”
is the most common species of centipedes
all throughout Europe and is omnipresent
in almost any habitat.

Lithobius latro MEINERT, 1872
Synonyms: Lithobius mutabilis transalpinus LATZEL, 1880
Lithobius lucifugus C. L. KOCH, 1862
Synonyms: Lithobius alpinus C. L. KOCH, 1862; Lithobius rupivagus VERHOEFF, 1937; Lithobius walachicus VERHOEFF, 1901
Lithobius macilentus C. L. KOCH, 1862
Synonyms: Lithobius aulacopus LATZEL, 1880
Lithobius melanops NEWPORT, 1845
Synonyms: Lithobius glabratus C.L. KOCH, 1847; Lithobius lusitanus crissolensis VERHOEFF, 1935
Lithobius microps MEINERT, 1868
Synonyms: Lithobius duboscqui BRÖLEMANN, 1896; Lithobius olivarum VERHOEFF, 1925; Sigibius puritanus CHAMBERLIN,1913
Comment: Though many sources cite
microps as missing in Austria, the species
has been recorded in Vienna, Lower
Austria and Carinthia, and is possibly
present in more states, since it is native to
most surrounding countries such as
Germany, the Czech Republic, Slovakia,
Hungary, Slovenia, Croatia, Albania, Italy
and Switzerland.

33
Lithobius moellensis VERHOEFF, 1940
Synonyms: none
Lithobius mutabilis C. L. KOCH, 1862
Synonyms: Lithobius daday TÖMÖSVÁRY, 1880; Lithobius communis C.L. KOCH, 1844; Lithobius maculatus MATIC & DARABANTU, 1971
Lithobius muticus C. L. KOCH, 1847
Synonyms: Lithobius bicolor TÖMÖSVÁRY, 1879; Lithobius cinnamomueus L. KOCH, 1862
Lithobius nodulipes LATZEL, 1880
Synonyms: Lithobius athesinus VERHOEFF, 1937
Lithobius pelidnus HAASE, 1880
Synonyms: none
Comment: L. moellensis is the possible
third endemic species of centipedes in
Austria, and the only species endemic to
Carinthia. It has only been found and
recorded once, but the specimen is
damaged and has only been recorded to
have been found “in the Glockner area,
some 1500 meters above sea level”. The
taxonomy of the species is uncertain, and
even though it is listed as its own species,
has not officially gained endemic status
yet.
Comment: VERHOEFF noticed the variety of
certain traits of this speices and described
several subspecies: L. p. allemanicus from
Schwarzwald, L. p. insubricus from Wallis,
L. p. ponalensis from Gardasee and L. p.
annulipes from Hallstadt. A new
subspecies L. p. triangulatus nov. ssp. has
been described by KOREN and found at
the Saualpe.
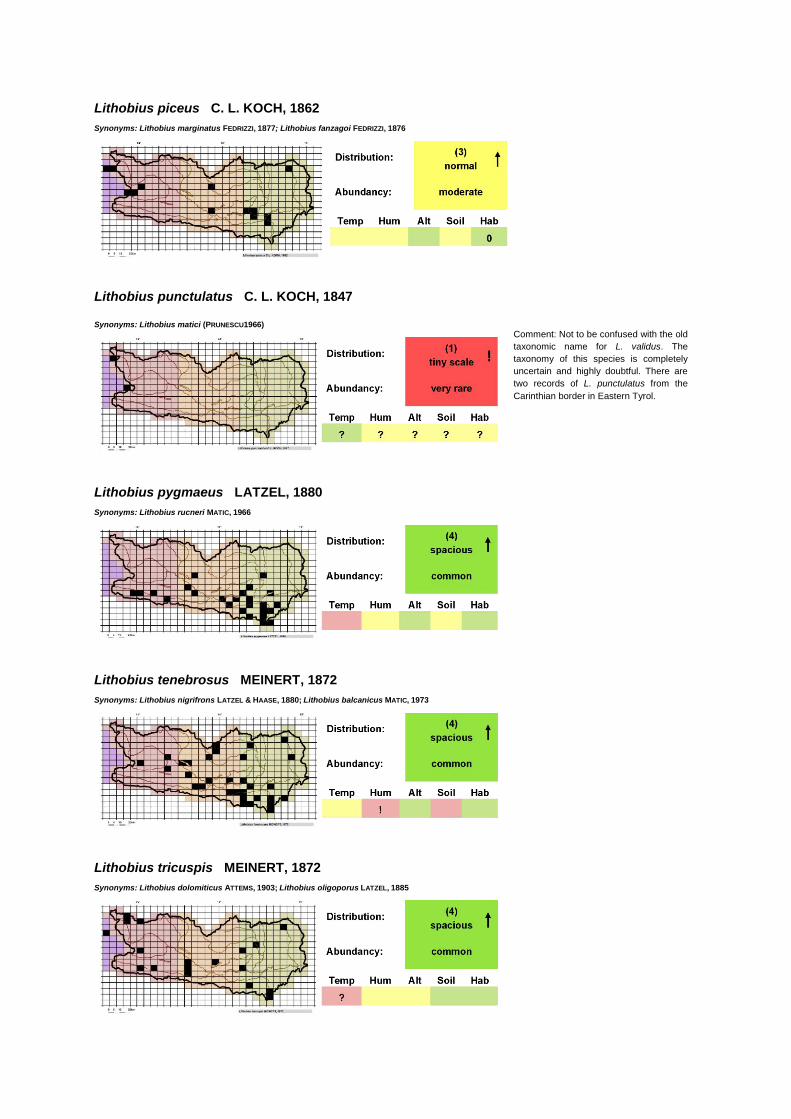
Lithobius piceus C. L. KOCH, 1862
Synonyms: Lithobius marginatus FEDRIZZI, 1877; Lithobius fanzagoi FEDRIZZI, 1876
Lithobius punctulatus C. L. KOCH, 1847
Synonyms: Lithobius matici (PRUNESCU1966)
Lithobius pygmaeus LATZEL, 1880
Synonyms: Lithobius rucneri MATIC, 1966
Lithobius tenebrosus MEINERT, 1872
Synonyms: Lithobius nigrifrons LATZEL & HAASE, 1880; Lithobius balcanicus MATIC, 1973
Lithobius tricuspis MEINERT, 1872
Synonyms: Lithobius dolomiticus ATTEMS, 1903; Lithobius oligoporus LATZEL, 1885
Comment: Not to be confused with the old
taxonomic name for L. validus. The
taxonomy of this species is completely
uncertain and highly doubtful. There are
two records of L. punctulatus from the
Carinthian border in Eastern Tyrol.
35
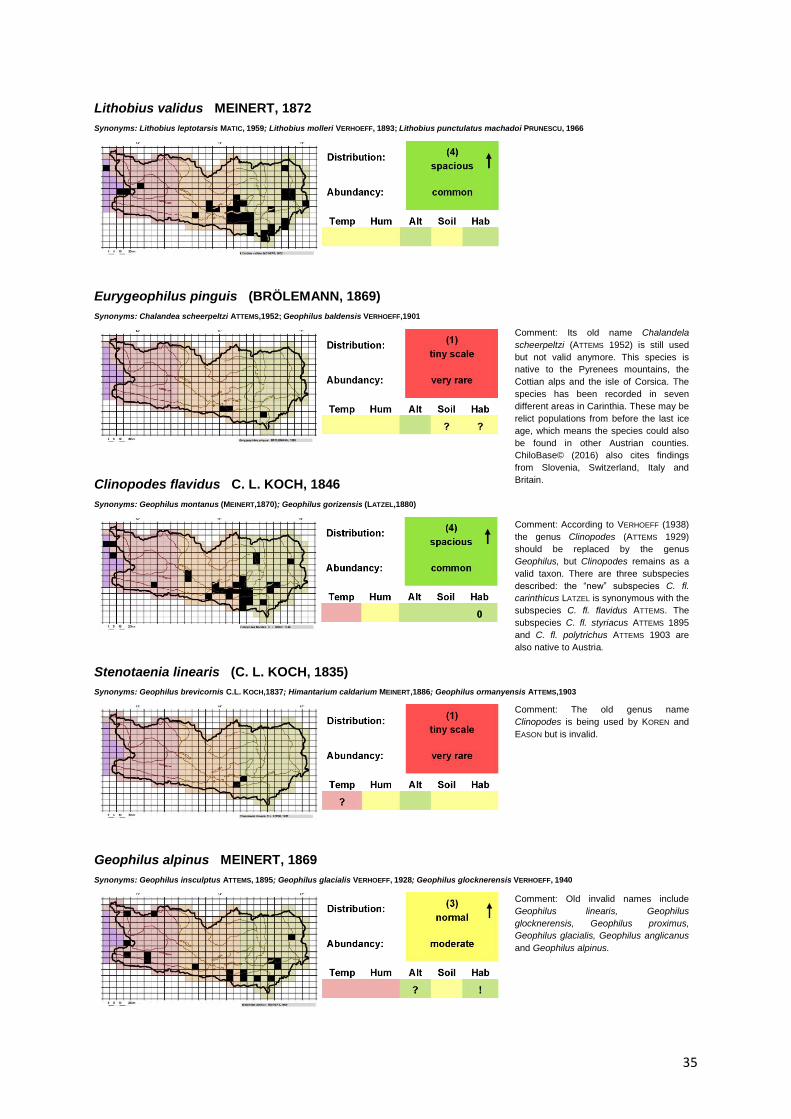
Lithobius validus MEINERT, 1872
Synonyms: Lithobius leptotarsis MATIC, 1959; Lithobius molleri VERHOEFF, 1893; Lithobius punctulatus machadoi PRUNESCU, 1966
Eurygeophilus pinguis (BRÖLEMANN, 1869)
Synonyms: Chalandea scheerpeltzi ATTEMS,1952; Geophilus baldensis VERHOEFF,1901
Clinopodes flavidus C. L. KOCH, 1846
Synonyms: Geophilus montanus (MEINERT,1870); Geophilus gorizensis (LATZEL,1880)
Stenotaenia linearis (C. L. KOCH, 1835)
Synonyms: Geophilus brevicornis C.L. KOCH,1837; Himantarium caldarium MEINERT,1886; Geophilus ormanyensis ATTEMS,1903
Geophilus alpinus MEINERT, 1869
Synonyms: Geophilus insculptus ATTEMS, 1895; Geophilus glacialis VERHOEFF, 1928; Geophilus glocknerensis VERHOEFF, 1940
Comment: Its old name Chalandela
scheerpeltzi (ATTEMS 1952) is still used
but not valid anymore. This species is
native to the Pyrenees mountains, the
Cottian alps and the isle of Corsica. The
species has been recorded in seven
different areas in Carinthia. These may be
relict populations from before the last ice
age, which means the species could also
be found in other Austrian counties.
ChiloBase© (2016) also cites findings
from Slovenia, Switzerland, Italy and
Britain.
Comment: According to VERHOEFF (1938)
the genus Clinopodes (ATTEMS 1929)
should be replaced by the genus
Geophilus, but Clinopodes remains as a
valid taxon. There are three subspecies
described: the “new” subspecies C. fl.
carinthicus LATZEL is synonymous with the
subspecies C. fl. flavidus ATTEMS. The
subspecies C. fl. styriacus ATTEMS 1895
and C. fl. polytrichus ATTEMS 1903 are
also native to Austria.
Comment: The old genus name
Clinopodes is being used by KOREN and
EASON but is invalid.
Comment: Old invalid names include
Geophilus linearis, Geophilus
glocknerensis, Geophilus proximus,
Geophilus glacialis, Geophilus anglicanus
and Geophilus alpinus.

Geophilus electricus (LINNÈ, 1758)
Synonyms: Geophilus sudeticus HAASE, 1880; Geophilus helveticus VERHOEFF, 1928
Geophilus flavus (DE GEER, 1778)
Synonyms: Necroploeophagus longicornis EASON 1960; Geophilus longicornis LEACH, 1815; G. pygmaeus styricus VERHOEFF, 1895
Geophilus oligopus (ATTEMS, 1895)
Synonyms: Geophilus noricus VERHOEFF, 1928; Geophilus pauropus ATTEMS, 1927; Geophilus insculptus ATTEMS, 1895
Geophilus pygmaeus LATZEL, 1880
Synonyms: Geophilus carnicus VERHOEFF, 1928; Geophilus cispadanus SILVESTRI, 1896; Geophilus larii VERHOEFF, 1934
Henia illyrica (MEINERT, 1870)
Synonyms: Chaetechelyne herzegowinensis VERHOEFF,1938; Henia termena CHAMBERLIN, 1952
Comment: Necroploephagus longicornis,
Geophilus longicornis, Geophilus
hortensis and Pachymerium tristanicum
are all old taxonomic names that are still
used but invalid.
Comment: Synonymous with the species
G. noricus (VERHOEFF 1928 and G.
pauropus (ATTEMS 1927), which have
been described as separate species by
KOREN and others.
Comment: According to ATTEMS 1929, G.
pygmaeus is an Illyric species native to
Slovenia, Croatia and Hungary. One
record from Carinthia and one from Styria
confirm the species is native to Austria. It
is also heavily implied that the species is
native to Upper Austria as well.

37
Henia vesuviana (NEWPORT, 1845)
Synonyms: Chaetechelyne vesuviana NEWPORT, 1845; Chaetechelyne corsica VERHOEFF,1943; Scolopendra fusca FOURCROY,1785
Strigamia acuminata (LEACH, 1815)
Synonyms: Scolioplanes italicus VERHOEFF,1928; Linotaenia rosulans C.L. KOCH, 1847; Geophilus subtilis C.L. KOCH, 1838
Strigamia crassipes (C. L. KOCH, 1835)
Synonyms: Geophilus breviceps NEWPORT,1845; Scolioplanes mediterraneus VERHOEFF,1928
Strigamia transsilvanica (VERHOEFF, 1928)
Synonyms: Scolioplanes transsilvanica (VERHOEFF, 1935)
Pachymerium ferrugineum (C. L. KOCH, 1835)
Synonyms: Geophilus caucasicus ATTEMS,1903; Geophilus paradoxus TÖMÖSVÁRY,1880; Mecistocephalus punctilabium NEWPORT,184
Comment: The old genus name
Chaetechelyne is valid no more. It’s a
Mediterranean species and native to
northern Africa, south France, Croatia and
all throughout Italy to Tyrol, Romania and
Hungary. The few records of this species
from Carinthia are probably relict
populations of warmer ages.

Schendyla carniolensis VERHOEFF, 1902
Synonyms: Schendyla (Echinoschendyla) zonalis BRÖLEMANN & RIBAUT, 1911
Schendyla nemorensis (C. L. KOCH, 1837)
Synonyms: Dicellophilus nemorensis C. L. KOCH, 1837; Geophilus gracilis HARGER,1872
Schendyla tyrolensis (MEINERT, 1870)
Synonyms: Schendyla montana ATTEMS, 1895; Brachyschendyla montana RIBAUT & BROLEMANN,1927
Dicellophilus carniolensis (C. L. KOCH, 1847)
Synonyms: Geophilus apfelbecki VERHOEFF, 1898; Geophilus austriacus MEINERT, 1886; Mecistocephalus hungaricus TÖMÖSVÁRY,1880
Cryptops hortensis (DONOVAN, 1810)
Synonyms: Cryptops aenarienis VERHOEFF, 1943
Comment: The subgenus is
Echinoschendyla (BRÖLEMANN & RIBAUT),
but the old name Echinoschendyla zonalis
is invalid.
Comment: The old genus name
Dicellophilus is invalid.
Comment: The names Brachyschendyla/
Schendyla montana are not valid
anymore.

39
Cryptops parisi BRÖLEMANN, 1921
Synonyms: none
Cryptops rucneri MATIC, 1967
Synonyms: none
Scutigera coleoptrata (LINNÈ, 1758)
Synonyms: Scutigera pretzmanni WÜRMLI, 1973; Scutigera rubrovittata VERHOEFF, 1905
Comment: Closely related to Cryptops
hortensis, and maybe even synonymous.
The identity of Cryptops rucneri is
uncertain and only very few data exists on
the species.
Comment: This species is the only repre-
senttative of the Scutigeromorpha in
Europe and very thermophile. It has
invaded almost every anthropomorphic
area thanks to carryover but has no high
abundances. It is extremely anthropo-
genic and common in houses and
manmade structures. There are only few
populations in the wild, which is why the
species is often listed as endangered.
KOREN (1992) and Zobodat© (2016) show
no records of Scutigera in Carinthia and
Styria, yet it is omnipresent. It is
discussed if the species should be
classified as a Neozoon, but experts claim
the species populated Austria from the
south by itself (ESSL &RABITSCH 2002).

1 2
3
4
5 6

41
Red List of Centipedes of Styria
Eupolybothrus fasciatus (NEWPORT, 1845)
Synonyms: Lithobius montellicus (FANZAGO, 1874); Polybothrus praecursor alarichi (ATTEMS, 1934)
Eupolybothrus grossipes (C. L. KOCH, 1874)
Synonyms: Polybothrus dubius MANFREDI,1948; Lithobius montanus C.L. KOCH, 1847; Lithobius grossipes debilis LATZEL,1889
Eupolybothrus tridentinus (FANZAGO, 1874)
Synonyms: Lithobius leptopus (LATZEL,1880); Polybothrus cerberus (VERHOEFF,1929); Polybothrus leptopus ssp. (VERHOEFF,1935)
Harpolithobius anodus (LATZEL, 1880)
Synonyms: Harpolithobius calcivagus VERHOEFF, 1925; Harpolithobius andreevi MATIC & STAVROPOULOS, 1988
Comment: The species is often listed
under its old genus name
Leptopolybothrus. LATZEL (1880) listed the
species under the name Eupolybothrus
leptopus.
Comment: KOREN names the subspecies
anodus and denatus, and distinguishes
them by the size of their gonopods H.
anodus denatus MATIC, 1957 is recorded
in Romania and “Yugoslawia”. H. anodus
anodus LATZEL, 1880 the species native to
Austria. The records from Carinthia may
be relict populations from the last glacial
period.

Lithobius aeruginosus (C. L. KOCH, 1862)
Synonyms: Monotarsobius aeruginosus (VERHOEFF 1937)
Lithobius agilis C. L. KOCH, 1874
Synonyms: Lithobius paradisiacus MATIC & DARABANTU, 1971
Lithobius anisanus VERHOEFF, 1937
Synonyms: none
Lithobius austriacus VERHOEFF, 1937
Synonyms: Monotarsobius austriacus (VERHOEFF, 1937)
Lithobius borealis MEINERT, 1868
Synonyms: Lithobius lusitanus würmanus (VERHOEFF 1937); Lithobius lapidicola (LATZEL 1880)
Comment: The taxonomy of this species is
uncertain. It is only known from one record
from Selzthal in Styria and could possibly
be another species endemic to Austria.
Comment: Very closely related to
Lithobius aeruginosus.

43
Lithobius burzenlandicus VERHOEFF, 1931
Synonyms: Lithobius gridellii MANFREDI, 1955; Monotarsobius veronensis MATIC & DARABANTU,1971
Lithobius crassipes C. L. KOCH, 1862
Synonyms: Lithobius (Monotasobius) crassipes C. L. KOCH, 1862; Lithobius atrifrons SILVESTRI, 1896, Lithobius podokes ATTEMS, 1903
Lithobius curtipes C. L. KOCH, 1847
Synonyms: Lithobius carpathicus (MATIC 1958); Monotarsobius conformatus (CHAMBERLIN 1952)
Lithobius dentatus C. L. KOCH, 1844
Synonyms: Lithobius alpestris LATZEL, 1880
Lithobius erythrocephalus C. L. KOCH, 1847
Synonyms: Lithobius armatus SSELIWANOFF, 1880; Lithobius dubius TÖMÖSVÁRY, 1880; Lithobius dubius TÖMÖSVÁRY, 1880
Comment: KOREN claims that the two
subspecies burzenlandicus and euxinicus
are recorded in Romania, while the
subspecies carinthicus is native to
Carinthia. Findings from Slovenia only
name the species with no subspecies
(KOS 1988). The findings of KOREN
generally match those made by MATIC in
1966. For the specimen found in Styria by
THALHAMER (1998) only the species is
given.
Comment: Originally described as L.
dubius by TÖMÖSVARY, in 1880, L.
erythrocephalus has been described by
many authors all throughout Europe. In
many places this species is distinguished
into two sympatric subspecies. The
subspecies L. erythrocephalus schuleri
(MATIC, 1966 schulleri) described by
VERHOEFF (1925, 1937) is much more
common in Austria, but the exact
taxonomy of the species and its
subspecies is in need of revision.

Lithobius forficatus (LINNÈ, 1758)
Synonyms: Lithobius leachii NEWPORT 1844; Lithobius vulgaris LEACH, 1817
Lithobius franzi ATTEMS, 1949
Synonyms: none
Lithobius lapidicola MEINERT, 1872
Synonyms: Lithobius pusillus (LATZEL 1880); Lithobius sulcatus (C. L. KOCH 1862)
Lithobius latro MEINERT, 1872
Synonyms: Lithobius mutabilis transalpinus LATZEL, 1880
Lithobius lucifugus C. L. KOCH, 1862
Synonyms: Lithobius alpinus C. L. KOCH, 1862; Lithobius rupivagus VERHOEFF, 1937; Lithobius walachicus VERHOEFF, 1901
Comment: The “common stone centipede”
is the most common species of centipedes
all throughout Europe and is omnipresent
in almost any habitat.
Comment: According to ATTEMS, L. franzi
is most closely related to L. walachius
(VERHOEFF, 1901) {= L. lucifugus C. L.
KOCH, 1862}. There is only one record of
this species and one type specimen, yet
its taxonomic status is unequivocally,
making it the only species endemic to
Styria and one of only two fully accepted
endemic centipedes of Austria.

45
Lithobius macilentus C. L. KOCH, 1862
Synonyms: Lithobius aulacopus LATZEL, 1880
Lithobius macrocentrus ATTEMS, 1949
Synonyms: none
Lithobius melanops NEWPORT, 1845
Synonyms: Lithobius glabratus C.L. KOCH, 1847; Lithobius lusitanus crissolensis VERHOEFF, 1935
Lithobius mutabilis C. L. KOCH, 1862
Synonyms: Lithobius daday TÖMÖSVÁRY, 1880; Lithobius communis C.L. KOCH, 1844; Lithobius maculatus MATIC & DARABANTU, 1971
Lithobius muticus C. L. KOCH, 1847
Synonyms: Lithobius bicolor TÖMÖSVÁRY, 1879; Lithobius cinnamomueus L. KOCH, 1862
Comment: This endemic species has
been found and recorded multiple times in
Styria, Upper and Lower Austria as well as
Tyrol. The species is mostly alpine and
has been found over 3000 m above sea
level, yet has also been found in sub
montane areas just 500 m above sea
level.

Lithobius nodulipes LATZEL, 1880
Synonyms: Lithobius athesinus VERHOEFF, 1937
Lithobius peggauensis VERHOEFF, 1937
Synonyms: none
Lithobius pelidnus HAASE, 1880
Synonyms: none
Lithobius piceus C. L. KOCH, 1862
Synonyms: Lithobius marginatus FEDRIZZI, 1877; Lithobius fanzagoi FEDRIZZI, 1876
Lithobius pygmaeus LATZEL, 1880
Synonyms: Lithobius rucneri MATIC, 1966
Comment: Originally found and described
in Styrian Peggau, the species has also
been recorded in Slovakia and Bulgaria,
and its taxonomic status is highly
uncertain.
Comment: VERHOEFF noticed the variety of
certain traits of this speices and described
several subspecies: L. p. allemanicus from
Schwarzwald, L. p. insubricus from Wallis,
L. p. ponalensis from Gardasee and L. p.
annulipes from Hallstadt. A new
subspecies L. p. triangulatus nov. ssp. has
been described by KOREN and found at
the Saualpe.

47
Lithobius tenebrosus MEINERT, 1872
Synonyms: Lithobius nigrifrons LATZEL & HAASE, 1880; Lithobius balcanicus MATIC, 1973
Lithobius tricuspis MEINERT, 1872
Synonyms: Lithobius dolomiticus ATTEMS, 1903; Lithobius oligoporus LATZEL, 1885
Lithobius validus MEINERT, 1872
Synonyms: Lithobius leptotarsis MATIC, 1959; Lithobius molleri VERHOEFF, 1893; Lithobius punctulatus machadoi PRUNESCU, 1966
Lamyctes emarginatus (NEWPORT, 1844)
Synonyms: Lamyctes fulvicornis (MEINERT 1868); Lithobius gracilis (PORAT 1869)
Clinopodes flavidus C. L. KOCH, 1846
Synonyms: Geophilus montanus (MEINERT,1870); Geophilus gorizensis (LATZEL,1880)
Comment: Is the only neobiotic species of
centipedes found in Austrias south (ESSL
& RABITSCH 2002). Its distribution is
primarily south-hemispheric but is often
recorded as Western-Palearctic (WÜRMLI
1972) (ZALESSKAJA & GOLOVATCH 1996)
and holarctic, and is indeed distributed
wide throughout the latter, and common in
tropical areas as well (ZALESSKAJA 1994).
It is a typical pioneering species in most
of Europe, preferring instable and
anthropogenic habitats of every kind. It
has been recorded in Styria by ATTEMS in
1895. There are no records in Carinthia
as of yet, but as the species kept
spreading in the last 20 years it should be
present in Carinthia as well.
Comment: According to VERHOEFF (1938)
the genus Clinopodes (ATTEMS 1929)
should be replaced by the genus
Geophilus, but Clinopodes remains as a
valid taxon. There are three subspecies
described: the “new” subspecies C. fl.
carinthicus LATZEL is synonymous with the
subspecies C. fl. flavidus ATTEMS. The
subspecies C. fl. styriacus ATTEMS 1895
and C. fl. polytrichus ATTEMS 1903 are
also native to Austria.

Stenotaenia linearis (C. L. KOCH, 1835)
Synonyms: Geophilus brevicornis C.L. KOCH,1837; Himantarium caldarium MEINERT,1886; Geophilus ormanyensis ATTEMS,1903
Geophilus alpinus MEINERT, 1869
Synonyms: Geophilus insculptus ATTEMS, 1895; Geophilus glacialis VERHOEFF, 1928; Geophilus glocknerensis VERHOEFF, 1940
Geophilus electricus (LINNÈ, 1758)
Synonyms: Geophilus sudeticus HAASE, 1880; Geophilus helveticus VERHOEFF, 1928
Geophilus flavus (DE GEER, 1778)
Synonyms: Necroploeophagus longicornis EASON 1960; Geophilus longicornis LEACH, 1815; G. pygmaeus styricus VERHOEFF, 1895
Geophilus oligopus (ATTEMS, 1895)
Synonyms: Geophilus noricus VERHOEFF, 1928; Geophilus pauropus ATTEMS, 1927; Geophilus insculptus ATTEMS, 1895
Comment: The old genus name
Clinopodes is being used by KOREN and
EASON but is invalid.
Comment: Old invalid names include
Geophilus linearis, Geophilus
glocknerensis, Geophilus proximus,
Geophilus glacialis, Geophilus anglicanus
and Geophilus alpinus.
Comment: Necroploephagus longicornis,
Geophilus longicornis, Geophilus
hortensis and Pachymerium tristanicum
are all old taxonomic names that are still
used but invalid.
Comment: Synonymous with the species
G. noricus (VERHOEFF 1928 and G.
pauropus (ATTEMS 1927), which have
been described as separate species by
KOREN and others.
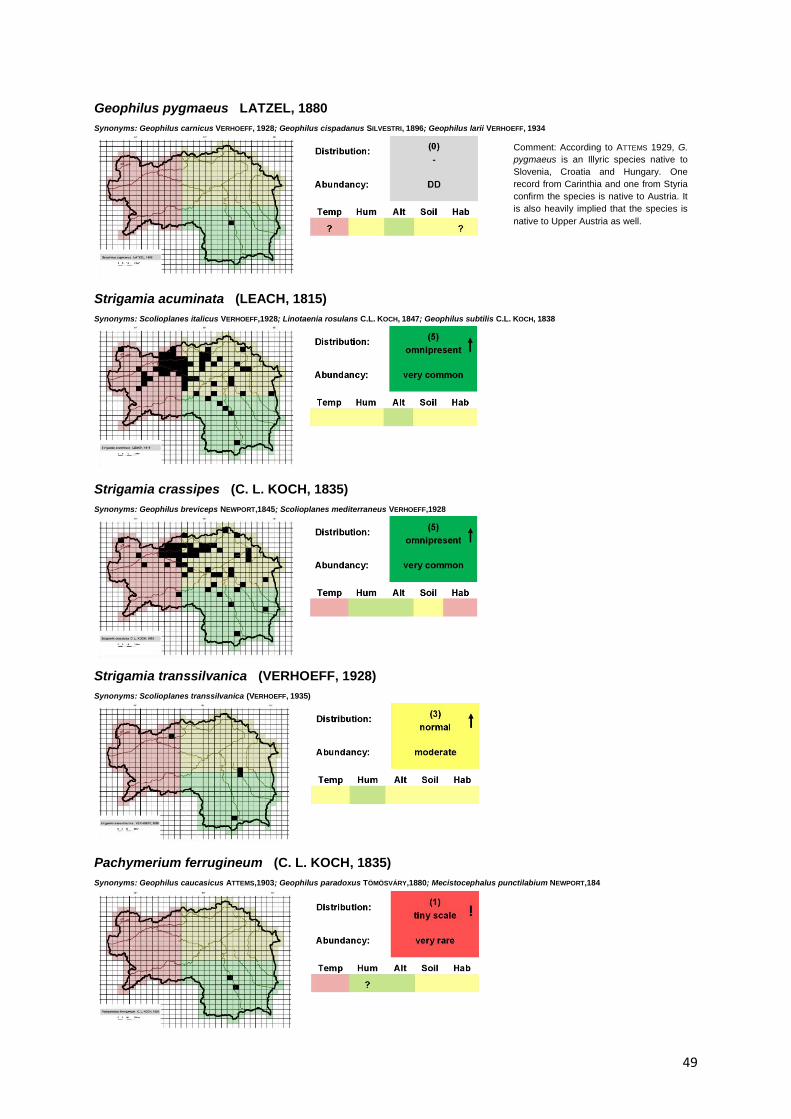
49
Geophilus pygmaeus LATZEL, 1880
Synonyms: Geophilus carnicus VERHOEFF, 1928; Geophilus cispadanus SILVESTRI, 1896; Geophilus larii VERHOEFF, 1934
Strigamia acuminata (LEACH, 1815)
Synonyms: Scolioplanes italicus VERHOEFF,1928; Linotaenia rosulans C.L. KOCH, 1847; Geophilus subtilis C.L. KOCH, 1838
Strigamia crassipes (C. L. KOCH, 1835)
Synonyms: Geophilus breviceps NEWPORT,1845; Scolioplanes mediterraneus VERHOEFF,1928
Strigamia transsilvanica (VERHOEFF, 1928)
Synonyms: Scolioplanes transsilvanica (VERHOEFF, 1935)
Pachymerium ferrugineum (C. L. KOCH, 1835)
Synonyms: Geophilus caucasicus ATTEMS,1903; Geophilus paradoxus TÖMÖSVÁRY,1880; Mecistocephalus punctilabium NEWPORT,184
Comment: According to ATTEMS 1929, G.
pygmaeus is an Illyric species native to
Slovenia, Croatia and Hungary. One
record from Carinthia and one from Styria
confirm the species is native to Austria. It
is also heavily implied that the species is
native to Upper Austria as well.

Schendyla carniolensis VERHOEFF, 1902
Synonyms: Schendyla (Echinoschendyla) zonalis BRÖLEMANN & RIBAUT, 1911
Schendyla nemorensis (C. L. KOCH, 1837)
Synonyms: Dicellophilus nemorensis C. L. KOCH, 1837; Geophilus gracilis HARGER,1872
Schendyla tyrolensis (MEINERT, 1870)
Synonyms: Schendyla montana ATTEMS, 1895; Brachyschendyla montana RIBAUT & BROLEMANN,1927
Dicellophilus carniolensis (C. L. KOCH, 1847)
Synonyms: Geophilus apfelbecki VERHOEFF, 1898; Geophilus austriacus MEINERT, 1886; Mecistocephalus hungaricus TÖMÖSVÁRY,1880
Cryptops hortensis (DONOVAN, 1810)
Synonyms: Cryptops aenarienis VERHOEFF, 1943
Comment: The subgenus is
Echinoschendyla (BRÖLEMANN & RIBAUT),
but the old name Echinoschendyla zonalis
is invalid.
Comment: The old genus name
Dicellophilus is invalid.
Comment: The names Brachyschendyla/
Schendyla montana are not valid
anymore.

51
Cryptops parisi BRÖLEMANN, 1921
Synonyms: none
Scutigera coleoptrata (LINNÈ, 1758)
Synonyms: Scutigera pretzmanni WÜRMLI, 1973; Scutigera rubrovittata VERHOEFF, 1905
Comment: This species is the only repre-
senttative of the Scutigeromorpha in
Europe and very thermophile. It has
invaded almost every anthropomorphic
area thanks to carryover but has no high
abundances. It is extremely anthropo-
genic and common in houses and
manmade structures. There are only few
populations in the wild, which is why the
species is often listed as endangered.
KOREN (1992) and Zobodat© (2016) show
no records of Scutigera in Carinthia and
Styria, yet it is omnipresent. It is
discussed if the species should be
classified as a Neozoon, but experts claim
the species populated Austria from the
south by itself (ESSL &RABITSCH 2002).

Critical reflection
As many authors previously stated, most of the native centipede species are common. Within the 3rd, 4th
and 5th category, we will find 67,3 % of all species in Carinthia and 60,8 % in Styria. The fact that only 11-
19 % of all species native to Austrias south lack an efficient amount of data for statistical analysis, is once
again prove that the chilopod-fauna of the two counties Carinthia and Styria underwent far greater study
than the rest Austria as well as most of its neighboring countries.
The numbers of Carinthia and Styria match surprisingly well. While 9,6 % of Carinthian and 13,7 % of
Styrian species were declared “very rare” (Category 1), 11,5 % of Carinthian and 5,9 % of Styrian species
were declared as “rare” (Category 2), adding up to approximately 20 % of the species of both counties in
the bottom two categories. Same goes for categories (4) and (5), where Styria has more species in
category (5) and Carinthia more in category (4).
The terms “rare”, “omnipresent” and “common” are of course chosen and categorized within the
context of this study and the data available. As mentioned in the disclaimer, the fact that a species is
declared as “omnipresent”, does not directly indicate that the species can be found anywhere. A species
may still prefer a certain habitat or completely avoid another. A species declared “very rare” is not said
to be “endangered” in any way. It could be, but as previously mentioned, due to the lack of metadata on
populations and their growth no statements will be made about the species’ “vulnerability”. The species
Geophilus electricus (LINNÉ 1758) is a great example, being declared “rare” in Styria (Data = 8) and being
uncategorized in Carinthia (Data = 1). This species is well studied and nowhere near any endangerment
within Austria. But it can only be found in certain habitats underground, making it a very rare encounter.
Another curious case is Lithobius forficatus (LINNÉ 1758), the most common species of centipedes in most
central European countries. The species is listed in both counties as “omnipresent” yet has significantly
less findings than species like Cryptops parisi BRÖLEMANN 1921 and Strigamia acuminata (LEACH 1815),
two species that are definitely less “common” than forficatus, but might be encountered more often in
certain habitats. Another important factor, especially when comparing the amount of data between
Styria and Carinthia, is the fact that the south of Styria is not well studied compared to the rest of the
county and compared to Carinthia in its entirety. Another curious case, and possibly for the above
mentioned reason, is Clinopodes flavidus C. L. KOCH 1846. The species is quite common in Carinthia
(Category 4) and has been recorded many times but has only been found once in Styria in recent years in
the very south. The species is probably quite common in Styria (at least south of Graz), but could not
have been recorded more often, since the south has only undergone studies by centipede experts in
recent years. Findings from Carinthia imply that Clinopodes, although not completely absent, avoids
mountainous habitats as found in Styrias heavily studied north. Therefore, it is quite a task evaluating the
abundance of a species. Species like Harpolithobius anodus (LATZEL 1880) are vulnerable to some degree,

53
but the few relict populations we have are quite stable, so categorizing them as “very rare” might be
misleading if the list is interpreted the wrong way. On the other hand, a species like Lithobius pygmaeus
LATZEL 1880 (Category 0 in both Carinthia and Styria) is profiting from a lack of data in this case, since
only one finding more could put the species in Category 1 (very rare) which it is certainly not, but only a
very cryptic species with distribution mainly in mountainous areas.
Furthermore, evaluating a species with uncertain taxonomy is always a difficult task, especially if sources
disagree on the status of a species. Endemic and possibly endemic species are very important here. As of
now, there are two confirmed endemic species native to Styria. One of them, Lithobius franzi ATTEMS
1949 is endemic to Styria, the other, Lithobius macrocentrus ATTEMS 1949 has also been recorded in
Tyrol, Upper and Lower Austria and is endemic to Austria. The species Lithobius anisanus VERHOEFF 1937,
whose identity is completely uncertain, has only been found in the Styrian Selzthal and might also be
endemic to Styria. Same goes for the Carinthian Lithobius moellensis VERHOEFF 1940 so far, the only
species that might be endemic to Carinthia, whose identity is uncertain as well. When it comes to
neobiotic species, the situation in Austria is quite simple. The only species of centipedes declared
neobiotic is Lamyctes emarginatus (NEWPORT 1844), which has been recorded in Styria in recent years,
after spreading all over Austria from the east. The species has not been recorded in Carinthia yet but
might already be present and could be added to the list any day. The house centipede Scutigera
coleoptrata (LINNÉ 1758) has been declared neobiotic in many northern European countries by some
experts. In Austria (especially the south), the situation is more complicated. Though the species may be
highly anthropogenic and has not been found in the wild in neither Carinthia nor Styria, there are
populations of Scutigera in the northern parts of Austria as well as in the neighboring countries Slovenia,
Croatia and Italy. Experts claim Scutigera coleoptrata has colonized Carinthia and Styria from the south
on its own and has not established populations in the wild because of its anthropogenic preferences.

Summary and outlook
Overall, the lists give a good overview of the abundancy and ecology of the native species. Especially the
ecology of most of the species is well known and could assist in further evaluating a species’
endangerment, especially if its habitat is or might be subject to environmental changes. Each map also
gives a simple overview of the species’ geographical distribution and amount of data available. The
statistics show that most of the species are common and nowhere near any endangerment. For those
species declared “rare”, comments have been added to explain the “rarity”. Future friends of this taxon
should aim to improve upon this list and expand its categorization to species of other Austrian regions.
The difficulty of this task will of course heavily depend on how the taxonomic status of the native species
will change and whether or not the status of uncertain species will be cleared.
The easiest way to improve upon or add to the status quo is by collecting and determining individuals
and thereby gaining data, especially from those regions and habitats which have not undergone intense
(or any) studies so far (i.e. Styrias south). A specific search for cryptic Geophilomorpha or rare
Lihtobiomorpha would lead to an increase in data, thereby helping us better understand the biology,
distribution and possibly taxonomic history of the species, leading to an even more cohesive red list that
could tackle the difficult task of evaluating the vulnerability or even endangerment of at least a few
native species.
A work such as this is never fully finished. One can only aim to achieve greater goals. As the taxon of
centipedes is gaining more and more friends, and more people turning towards biodiversity and
ecological studies as a hobby in recent years, maybe as a collective we will be able to fully understand
the fascinating and often cryptic life of these versatile critters.

55
PART II.
DNA BARCODING OF CENTIPEDES
State of the art
For the German Barcode of Life initiative (GBOL), WESENER et al. (2015) had already experimented with
several primer pairs when trying to barcode Stenotaenia linearis (KOCH, 1835), a native species of
Geophilomorpha. As for over 200 Myriapoda species, the aim is to sequence parts of the mitochondrial
cytochrome c oxidase subunit I gene. For PCR and sequencing HCO/LCO primer pairs (FOLMER et al. 1994)
were utilized, which showed a low success rate (<50%). The degenerate primer pair HCOJJ/LCOJJ (ASTRIN
& STÜBEN 2008) was used for further sequencing attempts, resulting in a much higher success rate
(>75%). In 2011 SPELDA et al. pioneered research in this field in Germany and sequenced many Bavarian
species of myriapods for the first time.
According to the official GBOL website 37 species of Chilopoda have been barcoded by the project since
its initiation (Dec. 2019), with 658 individuals registered in the database, some not determined further
than at family level (and some missing with actual barcode data). 658 individual centipedes have been
barcoded within the GBOL project including the following species: Lithobius forficatus (LINNÉ 1758),
Lithobius erythrocephalus C. L. KOCH 1847, Lithobius piceus C. L. KOCH 1862, Lithobius mutabilis C. L. KOCH
1862, Lithobius tenebrosus MEINERT 1872, Lithobius agilis C. L. KOCH 1874, Lithobius melanops NEWPORT
1845, Lithobius austriacus (VERHOEFF 1937), Lithobius malicentus C. L. KOCH 1862, Lithobius tricuspis
MEINERT 1872, Lithobius dentatus C. L. KOCH 1844, Lithobius muticus C. L. KOCH 1847, Lithobius borealis
MEINERT 1868, Lithobius microps MEINERT 1868, Lithobius crassipes C. L. KOCH 1862, Lithobius pelidnus
HAASE 1880, Lithobius curtipes C. L. KOCH 1847, Lithobius calcaratus C. L. KOCH 1844, Lithobius nodulipes
LATZEL 1880, Lithobius aeruginosus C. L. KOCH 1862, Eupolybothrus tridentinus (FANZAGO 1874), Cryptops
hortensis (DONOVAN 1810), Cryptops anomalans NEWPORT 1844, Cryptops parisi BRÖLEMANN 1921,
Geophilus truncorum BERGSØE & MEINERT 1866, Geophilus alpinus MEINERT 1869, Geophilus carpophagus
LEACH 1816, Geophilus flavus (DE GEER 1778), Geophilus electricus (LINNÉ 1758), Stenotaenia linearis (C. L.
KOCH 1835), Pachymerium ferrugineum (C. L. KOCH 1835), Schendyla nemorensis (C. L. KOCH 1837),
Stigmatogaster subterranean (SHAW 1974), Lamyctes emarginatus (NEWPORT 1844), Strigamia acuminata
(LEACH 1815), Strigamia crassipes (C. L. KOCH 1835), Henia vesuviana (NEWPORT 1845).

For the Austrian Barcode of Life (ABOL) project Dr. Nesrine Akkaris work group already sequenced
several species of centipedes, mainly of the genus Lithobius including the species’ Lithobius nodulipes
LATZEL 1880, Lithobius pelidnus HAASE 1880, and Lithobius pygmaeus LATZEL 1880 from Carinthia, with the
four markers COI, 16S, 18S and 28S6. An extensive study by OEYEN et al. (2014) unveiled the evolutionary
history of the Austrian Scolopendra cingulata LATREILLE 1829 population. The large gap between the intra-
and interspecific distances not only confirmed the Austrian and Hungarian populations to be relict
populations of a much wider distribution during the last post-glacial climatic optimum (which later
became isolated by the spreading of forests), but also shows that the Austrian population has a
completely unique haplotype within the species, despite its low genetic variation. A similar study by
WESENER et al. (2016), focusing on barcoding Centipedes from Austria and Germany, revealed large
intraspecific distances in several Cryptops species, with German specimens formally assigned to Cryptops
parisi BRÖLEMANN 1921 divided into three clades differing by 8.4–11.3% from one another. This study also
revealed the existence of several ghost lineages and revealed new species for Germany such as Cryptops
umbricus VERHOEFF 1931.
For this thesis 32 different species have been barcoded with the COI maker from different Austrian
areas, for 26 of which at least one barcode was successfully generated. Scolopendra cingulata LATRILLE
1829 and Geophilus electricus (LINNÉ 1758) as well as two unidentified Lithobius specimens were
collected in the state of Burgenland. From the same area in Burgenland as well as from the FH Graz
Campus the species Scutigera coleoptrata (LINNÉ 1758) has been barcoded, Pachymerium ferrugineum (C.
L. KOCH 1835), Lithobius erythrocephalus C. L. KOCH 1847, Lithobius aeruginosus C. L. KOCH 1862, Lithobius
latro MEINERT 1872,Lithobius pelidnus HAASE 1880, and Lithobius agilis C. L. KOCH 1874 from Lower
Austria, Lithobius lucifugus C. L. KOCH 1862 from Lower Austria and Carinthia, Eupolybothrus grossipes (C.
L. KOCH 1847), Lithobius macilentus C. L. KOCH 1862, Lithobius mutabilis C. L. KOCH 1862 and Schendyla
tyrolensis ATTEMS 1895 from Carinthia, Eupolybothrus tridentinus (FANZAGO 1874), Lithobius muticus C. L.
KOCH 1847, Schendyla carniolensis VERHOEFF 1902 and Cryptops hortensis (DONOVAN 1810) from Styria,
Henia illyrica (MEINERT 1870) and Henia vesuviana (NEWPORT 1845) from Vienna, Lithobius tricuspis
MEINERT 1872 from Carinthia, Clinopodes flavidus C. L. KOCH 1847 from Carinthia and Vienna, Strigamia
acuminata (LEACH 1815) from Styria, Vorarlberg and Upper Austria, Strigamia transsilvanica (VERHOEFF
1928) from Styria, Vorarlberg and Upper Austria, and from several counties the more widespread species
Lithobius dentatus C. L. KOCH 1844, Lithobius forficatus (LINNÉ 1758), Lithobius piceus C. L. KOCH 1862,
Lithobius tenebrosus MEINERT 1872, Lithobius validus MEINERT 1872, Dicellophilus carniolensis (C. L. KOCH
1847) and Cryptops parisi BRÖLEMANN 1921.
6 Data yet unpublished

57
Austrian centipede species that already underwent barcoding include Lithobius forficatus (LINNÉ 1758),
Lithobius dentatus C. L. KOCH 1844, Lithobius schuleri VERHOEFF 1925, Lithobius agilis C. L. KOCH 1874,
from Upper Austria, Lithobius glacialis (ATTEMS 1909), Strigamia acuminata (LEACH 1815) and Cryptops
parisi BRÖLEMANN 1921 from Upper Austria and Salzburg, Lithobius piceus C. L. KOCH 1862, Lithobius
tenebrosus MEINERT 1872, Lithobius aeruginosus C. L. KOCH 1862, Lithobius austriacus (VERHOEFF 1937),
Geophilus alpinus MEINERT 1870, Geophilus flavus (DE GEER 1778) and Schendyla tyrolensis (MEINERT 1870)
from Salzburg and Scolopendra cingulata LATRILLE 1829 from Burgenland, as well as 25 other species only
identified as part of the Lithobiomorpha and an additional six identified as Geophilomorpha order from
Salzburg, Upper Austria, Carinthia and Tyrol.7 Barcodes of species from Styria, Lower Austria and Vienna
(which are included in this thesis) are completely absent. Of the barcode data available on BOLD© 48
specimens are from the institution Zoologische Staatssammlung in Munich, the data of the remaining 10
specimens is retrieved from GenBank©.
7 All with COI markers; source: boldsystems.org (March 2020)

Methodology
The centipedes barcoded for this thesis were both collected by me and other researchers. Many
specimens were provided by Nikolaus Szucsich of the NHM Vienna. A complete list of all individuals,
collectors and locations can be found in the appendix.
Literature for species determination of centipedes is sparse, especially those with depictions of the
morphological structures (most only show representative drawings of the last pair of legs). Fortunately
KOREN wrote some of the best determination keys for the Geophilomorpha & Scolopendromorpha (1986)
and Lithobiomorpha (1992) for Carinthia and Eastern Tyrol, based upon and including drawings from the
works of EASON (1964). These determination keys also apply to most of the species found in Styria, since
there are only a handful species present in Styria that are absent in Carinthia (see Chapter I). For a
couple of other species, determination was done using internet research, individual papers or
descriptions and depictions of the species found in online centipede forums.
The critters were stored in 100% Ethanol in small test tubes. Some individuals have previously been
stored in 70% Ethanol but were then relocated to pure Ethanol. Individual names were attached to the
samples consisting of a code which contained the species name, sample number and location of
collection of the individual. For example, the individual number 51 is the species Lithobius forficatus
(LINNÉ 1758) and was the 4th individual collected at the location “Johnsbach”. The resulting code for the
species is 51_FOR_JB4.
Mitochondrial DNA has been extracted in the laboratory facilities of the Department of Biology.
Extraction of the material (mostly stored in 100% EtOH) was achieved by cutting off small pieces (i.e. one
leg) of the centipedes, drying the piece quickly thereby draining it from the alcohol and then placing it in
10% Chelex® solution, as was done by RICHLEN & BARBER (2005). The vortexed and centrifuged material
then underwent the extraction process for 20 minutes at 95°C.
For PCR the primer pair LCO1490‐JJ2/HCO2198‐JJ2 (ASTRIN & STÜBEN 2008) has been used, which has
shown great success in barcoding centipedes for the German GBOL project in recent years. Three
different types of Taq-polymerase had been used in the process: PhusionTM, BioThermTM and
SupraThermTM, all to great success with PhusionTM generally being the most effective and BioThermTM
the least. After trying out different annealing temperatures between 48°C and 51°C during the PCR
process results varied as well. DNA amplification followed the protocol of DUFTNER et. al 2005. The PCR
cocktail (9 µl) consisted of a Taq-polymerase (0,2 µl for PhusionTM/ 0,1 µl for Bio/SupraThermTM), the
primer pair (1 µl/ 0,5µl), dNTP’s (0,5 µl/ 0,35 µl), water (5,3 µl/ 7,5 µl) and a corresponding buffer (2 µl/ 1
µl), mixed with 2 µl of the sample. PCR was performed at a maximum temperature of 95°C and either
60°C or 72°C at the end of each cycle. The number of cycles varied as well, as 35 is the maximum number

59
for PhusionTM and 45 cycles the regular amount for primers such as BioThermTM and SupraThermTM
(KOBLMÜLLER et. al 2011).
After electrophoresis, samples with clearly visible bands underwent cleanup via ExoSAPTM, a reagent
which hydrolyzes excess primer fragments and nucleotides, followed by the sequencing reaction. For the
three-hour sequencing reaction at 94°C initial temperature and annealing temperature of 50°C, a 6 µl
cocktail of H2O (3,95 µl), sequencing buffer (1,6 µl), BigDye® terminator (0,2 µl) and the primer (0,25 µ)
was mixed with 2 µl of the sample (KOBLMÜLLER et. al 2011). After adding distilled water to the samples
and pipetting them onto SephadexTM pillars inside Eppendorf test-tubes, the samples were centrifuged at
9000rpm and thereby purified. What followed was the sequencing process using a 16-capillary-
sequencer 3130x and a 3500xl 24-capillary-sequencer by Applied Biosystems®. The resulting sequences
were (if necessary) corrected, inserted into a FASTA file and aligned using the MEGA7© program. After
shortening all the sequences to “upload length” the sequences were “blasted” using the BOLD©
(www.boldsystems.org) database, and matching sequences were downloaded from GenBank©
(https://www.ncbi.nlm.nih.gov/genbank) to be inserted into the FASTA file for comparison and further
analysis.
From there a Neighbor-Joining tree could be constructed using MEGA©, a agglomerative bottom-up
clustering method that requires knowledge of the distances between each pair of taxa and is commonly
used when working with protein-coding DNA data. A p-distance model has been chosen for substitution
with uniform rates and a 1000 bootstrap method for statistical support. In order to further estimate the
evolutionary divergences between the sequences/species, a matrix output was created displaying
between group mean distances. Maximum intraspecific and minimum interspecific differences were
evaluated manually using Microsoft Excel©. The same file was then used to find the nearest neighbor of
those species which were to further evaluate, i.e. those where data of three or more specimens was
available. For those species a distance sheet was created.

Results and discussion
Graphic 2: Complete neighbor-joining tree computed using MEGA7.

61
Graphic 3: Circle NJ tree of the Geophilomorpha computed using MEGA7.
Graphic 4: Circle NJ tree of the Lithobiomorpha computed using MEGA7.

Graphic 5: NJ tree of the Scolopendromorpha and Scutigeromorpha computed using MEGA7.
Table 2: Interspecific distances and distances to nearest neighbor (DNN) evaluated for every species with n>=3.
Species N Imax Nearest Neighbour DNN
Scutigeromorpha
Scutigeridae
Scutigera coleptrata 3 4,3 Cryptops hortensis 22,2
Scolopendromorpha
Cryptopidae
Cryptops parisi 4 9,6 Cryptops anomalans 18,4
Lithobiomorpha
Lithobiidae
Lithobius erythrocephalus 3 15,4 Lithobius piceus 12,9
Lithobius forficatus 5 14,5 Lithobius validus 16,6
Lithobius tenebrosus 3 9,3 Lithobius piceus 14,9
Lithobius validus 5 13,7 Lithobius piceus 15,6
Geophilomorpha
Geophilidae
Strigamia acuminata 9 15,7 Strigamia transsilvanica 15,4
Henia illyrica 4 11,6 Henia vesuviana 8
Henia vesuviana 4 3,9 Henia illyrica 8

63
The complete NJ-tree (Graphic 2, p. 60) shows a clear distinction between the orders Lithobiomorpha
and Geophilomorpha. The Scutigeromorpha are falsely potted with the Scolopendromorpha, rather than
being an outside group. COI barcoding data is however unreliable for evaluating and supporting larger
phylogenetic splits due to the high mutation rate of the gene, and should only be used to distinguish
closely related species. With the exception of Pachymerium ferrugineum (C. L. KOCH 1835) all members of
the Geophilomorpha order can be found potted at the bottom of the tree. The odd positioning of
ferrugineum as an outside group of the Geophilomorpha is even more evident in the Geophilomorpha
NJ-tree (Graphic 3, p. 61), but can (again) be explained away with the limitations of the barcoding data
when it comes to supporting deeper phylogenetic splits. While Pachymerium may not be closely related
to other Geophilomorpha genera such as Strigamia it is usually found closer to genera such as Geophilus.
The unknown Lithobius specimen #1 seems to be related to Lithobius forficatus (LINNÉ 1758) but doesn’t
appear to be Lithobius piceus C. L. KOCH 1862, which it shared many morphological traits with. Judging
from the tree alone, the 2nd unidentified Lithobius specimen is most probably Lithobius macilentus C. L.
KOCH 1862, a morphological analysis however strongly suggests otherwise.
The notably high distances displayed in Table 2 (p. 62) suggest high levels of cryptic diversity. Studies on
cryptic diversity in centipedes are not sparse, but usually restricted to a confined area such as islands or
archipelagos. Cryptic diversity of central European centipedes has mostly been studies by WESENER,
VOIGTLÄNDER and their associates. They all have come to conclusion that cryptic diversity in European
centipedes is not uncommon, especially in older and omnipresent species like Cryptops parisi BRÖLEMANN
1921 (WESENER et. Al 2016). The highest intraspecific distance in Lithobius erythrocephalus C. L. KOCH
1847 strangely enough appears to be lower than the distance to its nearest neighbor Lithobius piceus C.
L. KOCH 1862. The two specimens closest to each other were retrieved from GenBank, however the
distance to the other piceus specimens are all lower than the imax for erythrocephalus as well. Lithobius
erythrocephalus C. L. KOCH 1847 is a species with several subspecies in Europe, two of which are native to
Austria. Even KOREN (1992) noted that the status of erythrocepahlus and all its subspecies should be
evaluated in order to track the species’ history. Lithobius piceus C. L. KOCH 1862 is an older species and
omnipresent all throughout Europe. It is therefore probably very diverse as well. We face the same
situation with Henia illyrica (MEINERT 1870) and Strigamia acuminata (LEACH 1815). The imax of the latter
however almost matches the distance to its nearest neighbor Strigamia transsilvanica (VERHOEFF 1928).
Both species show high levels of diversity with distances ranging from 0.2 to 15.7. The species’ of the
genus Henia are known to be very closely related to each other. The high intraspecific distance of Henia
illyrica (MEINERT 1870) can be explained by the inclusion of one specimen retrieved from GenBank©
which was collected in Germany. The large geographic distance is reflected in the genetic distance,
further underlining the diversity of these critters. Greater interpretation of the data and results is not
easy due to the low number of specimens per species.

Summary and outlook
The majority of the laboratory analysis has been carried out between February and June of 2020 and
therefore has been interrupted for weeks due to Corona-Virus safety measures, when university facilities
remained closed to all non-essential staffers. DNA barcoding has been largely unsuccessful ever after the
end of the regulations in late May. The specimens have been extracted several times, primers and
polymerases were changed and experimented with in order to achieve satisfying results, yet none of the
trials proved successful. The ultimate decision was to work with what we had. Unfortunately, the lack of
a substantial amount of usable barcodes per species and location left us unable to upload any barcodes
to the ABOL data bank or GenBank©, as was previously one of the goals of this thesis. However outside
of this thesis laboratory work may continue in order to create publishable barcodes.
Today several authors note that species identification using morphological traits has several limitations
such as misidentification due to plasticity of a trait or the existence of cryptic taxa. A high level of
expertise is often required to correctly identify species with the required certainty, and even the best
morphological keys are often only of use in one particular life stage or gender if based on genitalia. The
DNA barcoding approach might currently represent the best solution for species identification, even if
barcoding itself represents some limitations (VALENTINI et al. 2008, SALVI et. al 2020). Identifying species
through DNA barcoding is also helpful for understanding interspecies interactions as researchers such as
PFENNINGER et al. (2007) and TEDERSOO et al. (2007) point out. The number of studies involving DNA
barcoding has risen sharply ever since 2017, with the rise of newer and more effective methods which
(by then) have become broader available (DE SALLE & GOLDSTEIN 2019). DNA barcoding has already led to
many important phylogenetic discoveries, as barcoding of Austrian Scolopendra cingulata LATREILLE 1892
populations by OEYEN et al. (2014) revealed the population to be an important biogeographical relic in a
possible microrefugium.
DNA barcoding is mainly limited by the system it is using, which is single locus based, even if several
regions from the organelles DNA are sequenced. It is known and has been well discussed that identical
mitochondrial DNA sequences can be present in different related species, either due to introgression or
incomplete lineage sorting since the time of speciation (BALLARD & WHITLOCK 2004). The inclusion of
mtDNA evidence in biogeographic analysis often reveals unexpected diversity or discordance with
morphology, prompting a re-evaluation of morphological and ecological characteristics and even
taxonomic revision (MORITZ & CICERO 2004). MORITZ & CICERO (2004) point out that mtDNA divergence
should not be a primary criterion for recognizing species boundaries and further claim: “[…] focusing on
mtDNA divergence as a primary criterion for recognizing species […] may lead scientist to overlook new
or rapidly diverged species, such as might arise through divergent selection or polyploidy […]”. There are
several other problems such as the accidental amplification of nuclear copies of mitochondrial DNA

65
fragments and the use of a divergence threshold for intra- & interspecies variation distinction (ZHANG &
HEWITT 1996, HEBERT et al. 2003). According to NIELSEN & MATZ (2006) several cases of species
misidentification are caused by incomplete databases, where the number of analyzed individuals per
species is often far too low and does not allow for a precise estimation of intra- and interspecific
variation. HEBERT et al. (2003) mentioned another limitation of DNA barcoding in in the length of the
sequences used, which is usually > 500 bp and thereby prevents the amplification of degraded DNA,
which many DNA barcoding applications can only be based on, especially environmental samples where
the target DNA is often from dead animals or plants (DEAGLE et al. 2006, VALENTINI et al. 2008). The rise of
NGS brought fourth even newer, more revolutionary barcoding methods such as “mini-barcoding”,
methods that are used to this very day. VALENTINI et al. in 2008 proposed NGS methods such as
pyrosequencing could counteract many of the upper mentioned problems; however pyrosequencing did
not see the rise the authors expected and was quickly replaced by cheaper NGS methods.
With many methods failing to provide cohesive results a lot of doubt has been cast on the DNA
barcoding method in the late 2000’s, yet ecologists increasingly turned to barcoding methods as a means
of species determination, because in many circumstances it represents an easy and rather quick way to
identify a species. HULLEY et. al (2018) found another use of DNA barcoding in their research on larval fish
in the great lakes. They deemed DNA barcoding: “an effective alternative to morphological identification
[…] of larval and embryonic fish, but comparisons of the two approaches with species from the Great
Lakes are limited. It may be particularly important to examine this issue in the Great Lakes because a
relatively young group of post-glacial fish species are present which may be difficult to resolve using
morphology or genetics.” Consequently both critics and champions of the barcoding method published a
wide variety of papers on the subject in the last decade. General interest in barcoding, as well as careful
and critical analysis, led to the rise of more effective methods and even approaches specific to one group
of organisms or even a single genus. Still, the accuracy of DNA barcoding tools depends on the existence
of a comprehensive archived library of sequences reliably determined at species level by expert
taxonomists, as SALVI et. al (2020) point out. Pre-barcoding morphological analysis is especially important
when working with a new or small group of animals, where new subspecies or variates are expected to
still be discovered. It is most important to always have a DNA barcode of the sample as basic data, since
the sequences are not prone to subjectivity and can be reanalyzed in the future in accordance with
improvements in taxonomic knowledge, as VALENTINI et al. (2008) point out. SALVI et. al (2020) further
discussed the limitations of barcoding and the issue of intrinsic and extrinsic errors when they explain:
“While intrinsic errors to be solved require an integrative taxonomic and evolutionary study approach,
which goes beyond the idea of barcoding as identification tool, extrinsic errors are due to human
mistakes and can be corrected much more easily. Thus, in a reference dataset free of extrinsic errors it
would be easy to spot species identification inconsistencies that require further taxonomic research, [...]

the importance of which [...] cannot be overstated”. While DE SALLE & GOLDSTEIN (2019) seem to agree
with most researchers defending the DNA barcoding method as a valuable taxonomic tool, they very
critically discuss distance measures and NJ trees derived from barcoding data as a means of species
determination. They claim: “it is important to separate the statement that NJ analyses “work” to identify
species from the supposition that they allow us to infer anything about species in the abstract” and
summarize a long discussion with: “[...] the fact that a very high proportion of diagnosable species are
captured by NJ analyses is encouraging, but not sufficient.” Thereby they reject the premise that DNA
barcoding serves to repair some inherent flaw in the practice of systematic, yet actively embrace DNA
barcoding as a near-universal advance for taxonomic research.
Generally speaking, DNA barcoding works well in most species, especially when the amount of data is
large and covers the total variability of the species’ area of distribution. Problems may still occur with
either very young species that did not accumulate or sort out enough mutations to distinguish them by
their barcodes alone or old species with large population sizes where divergent haplotypes may survive.
Hybridization and introgression events may still lead to “wrong barcodes”, but refined methods aim to
avoid those problems (ZIMMERMANN et. al 2013). As is evident, the method is an effective and powerful
one, and should therefore be used with great care and thought put into it. A critical analysis of one’s own
methods is fundamental in evolutionary biology.
As far as centipedes are concerned, the DNA barcoding approach alone will not resolve their taxonomy.
The high levels of cryptic diversity, numerous subspecies and high variability of important traits (such as
gonopods shape, number of segments etc.) make a morphological (pre)analysis essential. DNA barcoding
should however be applied whenever possible, since it is an important tool that may aid in taxonomic
determination. Another acceptable approach for example would be to simply generate barcodes with
very little pre-analysis in order to search for signs of cryptic diversity. This approach might also reveal
other details of greater importance, such as the revelations of the research by OEYEN et al. (2014) on
native Scolopendra populations, which would have remained completely unobserved without the usage
of the barcoding method.
The results of this very thesis have shown what issues may occur. The barcode of one individual may
match the barcode of a second individual, yet morphologically they can be completely different. The
problems we (possibly) face when applying or relying on the barcoding method are various and have
been discussed extensively above. Problems should always be critically examined individually. Many
taxonomists face the problem of numerous subspecies within their orders of expertise. Discussions on
the actual existence of subspecies as biological units will continue and not be resolved any time soon.
Whatever these subspecies may represent, they are proof of high levels of variety within certain species’.
As we have shown, DNA barcoding may assist in evaluating the gaps between these varieties. Knowledge

67
about the species’ biology, ecology, morphology etc. is fundamental, because it aids us in interpreting
the barcoding results and helps resolving possible issues and mismatches.
Lastly, DNA barcoding will continue to evolve and improve. We cannot predict what new methods will
arise in the future, but we shall aim to improve upon what is established. Solving the taxonomy of all
beings sharing this world with us is a massive task, but (as is evident) many scientists take on this task
with more and more joining every day. With combined efforts and many friends of different organisms
we can aim to achieve greater knowledge about the history of all kinds of organisms with the help of
present and future DNA barcoding methods.

APPENDIX
List of individual centipede specimens included in the tree
Specimen# Species Collection Site Coordinates Date o. Coll. Collector Determinatior
6_COL_TR3 Scutigera coleoptrata Thenauriegel (BL) 47.941453, 16.718001 08.06.2018 Svetnik Svetnik
48_COL_FH1 Scutigera coleoptrata FH Joanneum Graz (Stmk) 47.069168, 15.410829 04.07.2019 Lienhard Svetnik
43_VAL_E12 Lithobius validus Leechwald Graz (Stmk) 47.084431, 15.462240 01.04.2019 Svetnik Svetnik
66_VAL_X1 Lithobius validus Tainach (Ktn) 46.391780, 14330349 14.04.2017 Svetnik Svetnik
67_VAL_X2 Lithobius validus Gut Landskron (Ktn) 46.645287, 13.895456 27.07.2019 Svetnik Svetnik
68_VAL_BL6 Lithobius validus Burg Landskron (Ktn) 46.641788, 13.898047 07.08.2018 Svetnik Svetnik
36_FOR_E1 Lithobius forficatus Leechwald Graz (Stmk) 47.084431, 15.462240 01.04.2019 Svetnik Svetnik
45_FOR_NO9 Lithobius forficatus Schönau (NÖ) 48.135400, 16.613800 29.04.2018 Wagner, Spiß Svetnik
51_FOR_JB4 Lithobius forficatus Johnsbach (Stmk) 47.321630, 14.372630 19.06.2018 Svetnik Svetnik
56_FOR_NO6 Lithobius forficatus Schönau (NÖ) 48.135400, 16.613800 29.04.2018 Wagner, Spiß Svetnik
53_PIC_JB6 Lithobius piceus Johnsbach (Stmk) 47.321630, 14.372630 19.06.2018 Svetnik Svetnik
87_PIC_1960 Lithobius piceus* St. Pölten (NÖ) 47.995619, 15.456727 unknown Bartel, Böhm Svetnik
107_DEN_2330 Lithobius dentatus Johnsbach (Stmk) 47.569639, 14.583405 unknown Macek, Bartel Svetnik
27_TEN_E6 Lithobius tenebrosus Leechwald Graz (Stmk) 47.086294, 15.462648 01.04.2019 Svetnik Svetnik
58_TEN_WH17 Lithobius tenebrosus Waldhof Villach (Ktn) 46.636078, 13.896829 14.08.2019 Svetnik Svetnik
15_BOR_WF10 Lithobius borealis Waldhof Villach (Ktn) 46.635511, 13.896883 04.08.2018 Svetnik Svetnik
70_MAL_WF11 Lithobius macilentus* Waldhof Villach (Ktn) 46.635511, 13.896883 04.08.2018 Svetnik Svetnik
112_IND_2366 Lithobius indet. Gsuchmauer (Stmk) 47.547383, 14.656950 unknown Group NHM Svetnik
23_ERY_NO8 Lithobius erythrocephalus Schönau (NÖ) 48.134100, 16.606900 29.04.2018 Wagner, Spiß Svetnik
91_ERY_1955 Lithobius erythrocephalus St. Pölten (NÖ) 47.995619, 15.456727 unknown Bartel, Böhm Svetnik
83_IND_2193 Lithobius indet. Ruine Neuburg (Vbg) 47.423219, 9.622580 unknown Macek Svetnik
86_GLA_1980 Lithobius glacialis St. Pölten (NÖ) 47.914555, 15.372111 unknown Macek, Bartel Svetnik
26_HOR_E9 Cryptops hortensis Leechwald Graz (Stmk) 47.084431, 15.462240 01.04.2019 Svetnik Svetnik
46_ANO_NO10 Cryptops anomalans Schönau (NÖ) 48.135400, 16.613800 29.04.2018 Spiß Svetnik
42_PAR_E11 Cryptops parisi Leechwald Graz (Stmk) 47.084431, 15.462240 01.04.2019 Svetnik Svetnik
65_PAR_AB1 Cryptops parisi Gut Landskron (Ktn) 46.644767, 13.899715 27.07.2019 Svetnik Svetnik
88_PAR_1956 Cryptops parisi St. Pölten (NÖ) 47.995619, 15.456727 unknown Bartel, Böhm Svetnik
96_ELE_2235 Geophilus electricus Ebner Klamm (Stmk) 47.530836, 14.530836 unknown Macek, Bartel Svetnik
25_ACU_E14 Strigamia acuminata Leechwald Graz (Stmk) 47.086294, 15.462648 04.05.2018 Svetnik Svetnik
76_ACU_2044 Strigamia acuminata Tschütsch (Vbg) 47.315355, 9.646802 unknown Macek Svetnik
77_ACU_2050 Strigamia acuminata Krummberg (Vbg) 47.334980, 9.606916 unknown Macek Svetnik
84_ACU_2177 Strigamia acuminata Kloster Maria Stern (Vbg) 47.570386, 9.779252 unknown Macek Svetnik
85_ACU_2078 Strigamia acuminata Kloster Maria Stern (Vbg) 47.570386, 9.779252 unknown Macek Svetnik
97_ACU_2251 Strigamia acuminata Ebner Klamm (Stmk) 47.530836, 14.530836 unknown Macek, Bartel Svetnik
98_ACU_2267 Strigamia acuminata Ödelsteinhöhle (Stmk) 47.519166, 14.615277 unknown Macek, Bartel Svetnik
102_ACU_2274 Strigamia acuminata Ödelsteinhöhle (Stmk) 47.519166, 14.615278 unknown Macek, Bartel Svetnik
111_TRA_2358 Strigamia transsilvanica Landweg Liezen (Stmk) 47.609808, 14.740686 unknown Macek, Bartel Svetnik
71_ILL_1227 Henia illyrica Baumgartner Höhe (W) 48.210816, 16.277955 unknown Bartel, Szucsich Szucsich
74_ILL_1528 Henia illyrica Döbling (W) 48.275700, 16.351986 unknown Szucsich Szucsich
95_ILL_1599 Henia illyrica Prater (W) 48.191666, 16.445000 unknown Szucsich Szucsich
72_VES_1321 Henia vesuviana Wienflussufer (W) 48.209833, 16.212641 unknown Bartel, Szucsich Szucsich
73_VES_1442 Henia vesuviana Schlosspark Schönbrunn (W) 48.183330, 16.300000 unknown Szucsich Szucsich
81_VES_1443 Henia vesuviana Schlosspark Schönbrunn (W) 48.183330, 16.300001 unknown Szucsich Szucsich
94_NEM_2196 Schendyla nemorensis Ruine Neuburg (Vbg) 47.423219, 9.622580 unknown Macek Svetnik
78_TYR_2214 Schendyla tyrolensis Wolfsberg Graben (Ktn) 46.898333, 14.848888 unknown Walzl Svetnik
82_FLA_1530 Clinopodes flavidus Döbling (W) 48.275700, 16.351986 unknown Szucsich Szucsich
37_FER_NO1 Pachymerium ferrugineum Hafen Lobau (NÖ) 48.154200, 16.487600 30.04.2018 Wagner, Spiß Svetnik
41_CAR_E10 Dicellophilus carniolensis Leechwald Graz (Stmk) 47.084431, 15.462240 04.05.2018 Svetnik Svetnik
*reverese Taxonomy

69
BIBLIOGRAPHY
ADIS J. & M.S. HARVEY (2000): How many Arachnida and Myriapoda are there worldwide and in Amazonia?
– Studies on neotropical Fauna and Environment 35 (2).
ARTHUR W. (2002): The interaction between developmental bias and natural selection from centipede
segmentation to a general hypothesis. – Heredity 89.
ASPÖCK H. (2010): Krank durch Arthoropoden. – Denisia 30.
ASTRIN J.J. & P.E. STÜBEN (2008): Phylogeny in cryptic weevils: molecules, morphology and new genera of
western Palaearctic Cryptorhynchinae (Coleoptera : Curculionidae). – Invertebrate Systematics 22 (5).
BALLARD J.W.O. & M.C. WHITLOCK (2004): The incomplete natural history of mitochondria – Molecular
Ecology 13 (4).
BARROSO E., HIDAKA A.S., DOS SANTOS A.X., MATOS FRANҪA J.D., DE SOUSA A.M., RODRIGUES VALENTE J., AMORAS
MAGALHÃES A.F. & P.P. OLIVEIRA PARDAL (2001): Acidentes por centopéia notificados pelo „Centro de
Informaҫões Toxicológicas de Belém“, num periodo des dois anos. – Revista de Sociedade Brasileira de
Medicina Tropical 34.
BATES M. (2016): „Horrific“ First Amphibious Centipede Discovered. – National Geographic.
BOUCHARD N.C., CHAN G.M. & R.S. HOFFMAN (2004): Vietnamese centipede envenomation. – Veterinary and
human toxicology 46.
BÜCHERL W. (1946): Venomous chilopods or centipedes. – In: BÜCHERL W. & E.E. BUCKLEY (Ed.), Venomous
animals and their venoms. Academic Press, New York.
BUSH S.P., KING B.O., NORRIS R.L. & S.A. STOCKWELL (2001): Centipede envenomation. – Wilderness and
environmental Medicine 12.
CARPENTER C.C. & J.C. GILLINGHAM (1984): Giant centipede (Scolopendra alternans) attacks marine toads
(Bufo marianus). – Caribbean Journal of Science 20.
COHEN E. & G.B. QUISTAD (1998): Cytotoxic effects of arthropod venoms on various cultured cells. –
Toxicon 36.

DEAGLE B.E., EVESON J.P. & S.N. JARMAN (2006): Quantification of damage in DNA recovered from highly
degraded samples – a case study on DNA in faeces. – Frontiers in Zoology 3 (11).
DE SALLE R. & GOLDSTEIN P. (2019): Review and Interpretation of Trends in DNA Barcoding. – Frontiers in
ecology and evolution 7.
DUFTNER N., KOBLMÜLLER S. & STURMBAUER C. (2005): Evolutionary Relationships of the Limnochromini, a
Tribe of Benthic Deepwater Cichlid Fish Endemic to Lake Tanganyika, East Africa. – Journal of Molecular
Ecology 60.
EASON E.H. (1964): Centipedes of the British Isles – Warne.
EDGECOMBE G.D. & G. GIRIBERT (2002): Myriapoda phylogeny and the relationships of chilopoda. – In:
Biodiversidad, Taxonomía y Biogeografia de Arthropods de México: Hacia una Síntesis de su
conocimiento, Volumen III.. – Universidad Autónoma de México.
EDGECOMBE G.D. & G. GIRIBERT (2008): A New Zealand species of the trans-Tasmanian centipede order
Craterostigmomorpha (Arthropoda: Chilopoda) corroborated by molecular evidence. – Invertebrate
Systematics 22.
EDGECOMBE G.D. & G. GIRIBERT (2009): Evolutionary developmental biology does not offer a significant
challenge to the neo-Darvinian paradigm – In: contemporary Debates in Phylosophy of Biology (JOHN
WILEY and Sons).
EISNER T., EISNER M. & M. SIEGLER (2005): Secret Weapons: defenses of insects, spiders, scorpions and
other many-legged creatures. – Cambridge Mass.: The Belknap Press.
GOMES A., DATTA A., SARNGI D., KAR P.K. & S.C. LAHIRI (1982): Occurrance of histamine and histamine release
by centipede venom. – Indian Journal of Medical Research 76.
GRUTTKE H., BINOT-HAFKE M., BALZER S., HAUPT H., HOFBAUER N., LUDWIG G., MATZKE-HAJEK G. & M. RIES (2016):
Rote Liste gefährdeter Tiere, Pflanzen und Pilze Deutschlands Band 4: Wirbellose Tiere (Teil 2).
GUERRERO A.P. (2007): Centipede bites in Hawaii: a brief case report and review of the literature. – Hawaii
Medical Journal 66.
HEBERT P.D.N., RATNASINGHAM S. & J.R. DE WAARD (2003): Barcoding animal life: cytochrome c oxidase
subunit 1 divergences among closely related species. – The Royal Society Biological Sciences 270.

71
HEBERT P.D.N. & T.R. GREGORY (2005): The Promise of DNA Barcoding for Taxonomy. – Systematic Biology
54.
HÖLDER A. (1901): Geschichte der Zoologie in Österreich von 1850 bis 1900, Myriapoden von Dr. Karl Graf
Attems; taken from „Botanik und Zoologie in Österreich während der letzten fünfzig Jahre“; Wien/
Vienna.
HÖLLDOBLER B. & E.O. WILSON (1990): The specialized predators. – The Ants; Howard University Press.
HOLMES O. (2016): Giant Swimming venomous centipede discovered by accident in world first. – The
Guardian July 1st 2016.
HOPKIN & READ (1992): The Biology of Millipedes. – Oxford University Press.
HULLEY E.N., TAYLOR N.D.J., ZARNKE A.M., SOMERS C.M., MAZON R.G., WILSON J.Y., & R.B. DOUGLAS (2018): DNA
barcoding vs. morphological identification of larval fish and embryos in Lake Huron: Advantages to a
molecular approach – Journal of Great Lakes Research 44 (5).
IVOCHEVA M., ZLATEVA S., MARINOV P. & Y.U. SABEVA (2008): Toxoallergic reactions after a bite from
myriapoda, genus Scolopendra in Varna region during the period 2003-2007. – Journal of IMAB – Annual
Proceeding (Scientific Papers) 2008(1).
KNOFLACH B. (2010): Hundert- und Tausenfüßer und ihre humanmedizinische Bedeutung.
Biologiezentrum Linz.
KOBLMÜLLER S., SALZBURGER W., OBERMÜLLER B., EIGNER E., STRUMBAUER C. & K.M. SEFC (2011): Separated by
sand, fused by dropping water: habitat barriers and fluctuating water levels steer the evolution of rock‐
dwelling cichlid populations in Lake Tanganyika – Molecular Ecology 20 (11).
KOREN A. (1986): Die Chilopoden-Fauna von Kärnten und Osttirol – Teil 1: Geophilomorpha,
Scolopendromorpha.
KOREN A. (1992): Die Chilopoden-Fauna von Kärnten und Osttirol – Teil 2: Lithobiomorpha.
LEVINSON H. & A. LEVINSON (2004): Wehrhafte Gliederfüßer als heilige Tiere in Altägypten. –
Naturwissenschaftliche Rundschau 57 (3).
LEWIS J.G.E. (1961): The life history and ecology of littoral centipede Strigamia maritima (Leach). –
Proceedings of the zoological society of London 137.
LEWIS J.G.E. (2007): The Biology of Centipedes. – Cambridge University Press.

MINELLI A. (2011): Treatise of Zoology. – The Myriapoda Volume 1: Brill, Leiden.
MIYAZAWA HIDEYUKI, UEDA CHIAKI, YAHATA KENSUKE & ZHI-HUI SU (2014): Molecular phylogeny of Myriapoda
provides insights into evolutionary patterns of the mode in post-embryonic development. – Scientific
Reports 4 (4127).
MOHAMED A.H., ABU-SINNA G., EL SHABAKA H.A. & A.A. EL-AAL (1983): Proteins, lipids, lipoproteins and some
enzyme characterization of the venom extract from the centipede Scolopendra morsitans. – Toxicon 21.
MOHRI S., SUGIYAMA A., SAITO K. & H. NAKAJIMA (1991): Centipede bites in Japan. – Cutis 47
MOLINARI J., GUTIÉRREZ E.E., ASCENҪÃO DE A.A., NASSAR R.J., ARENDS A. & R.J. MARQUEZ (2005): Predation by
giant centipedes, Scolopendra gigantea, on the species of bats in a Venezuelan cave. – Caribbean Journal
of Science 41.
MORITZ C. & C. CICERO (2010): DNA Barcoding: Promise and Pitfalls. – POLS Biology 2 (10).
MUMUCUOGLU K.Y. & V. LEIBOVICI (1989): Centipede (Scolopendra) bite: a case report. – Israel Journal of
Medical Sciences 25 (1).
MYERS P., ESPINOSA R., PARR C.S., JONES T., HAMMOND G.S. & T.A. DEWEY (2013): Geophilidae: Classification. –
Animal Diversity Web. University of Michigan; Museum of Zoology.
NEWPORT G. (1844): Monography of the class Myriapoda, order Chilopoda; with observations on the
general arrangement of the Articulata. – Transactions of the Linnean Society of London (Zoology) 19.
NIELSEN R. & M. MATZ (2006): Statistical approaches for DNA Barcoding. – Systematic Biology 55 (1).
OEYEN J. P., FUNKE S., BÖHME W. & WESENER T. (2014): The Evolutionary History of the Rediscovered
Austrian Population of the Giant Centipede Scolopendra cingulata LATREILLE 1829 (Chilopoda,
Scolopendromorpha). – PloS One 9 (9).
PFENNINGER M., NOWAK C., KLEY C., STEINKE D. & B. STREIT (2007): Utility of DNA taxonomy and barcoding for
the inference of larval community structure in morphologically cryptic Chironomus (Diptera) species. –
Molecular Ecology 16 (9).
RABITSCH W. & F. ESSL (2009): Endemiten. Kostbarkeiten in Österreichs Pflanzenund Tierwelt. –
Arachnologische Mitteilungen 37.
REMINGTON C.L. (1950): The bite and habits of a giant centipede (Scolopendra subspinensis) in the
philippine Islands. – American Journal of Tropical Medicine and Hygene 30.

73
RIEGER J.C., SHULTZ J.W., ZWICK A., HUSSEY A., BALL B., WETZER R., MARTIN J.W. & C.W. CUNNINGHAM (2010):
Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences. –
Nature 463.
RODRIGUEZ-ACOSTA A., GASSETTE J., GONZALEZ A. & M. GHISOLI (2000): Centipede (Scolopendra gigantea LINNÉ
1758) envenomation in a newborn. – Revista do Instituto de Medicina Tropical de Sao Paulo 42 (6).
ROSENBERG J. (2009): Die Hundertfüßer: Chilopoda. – Die neue Brehm-Bücherei 285 Westarp
Wissenschaften, Hohenwarsleben.
SALVI D., BERRILLI E., D’ALESSANDRO P. & M. BIONDI (2020): Sharpening the DNA barcoding tool through a
posteriori taxonomic validation: The case of Longitarsus flea beetles (Coleoptera: Chrysomelidae). –
PLoS ONE 15 (5).
SCOTT A.C., CHANDLER W.G. & S. PATERSON (1985): Evidence of pteridophyte-arthropod interactions in the
fossil record. – Proceedings in the royal society of Edinburgh 86 B.
SHEAR W.A. (1992): Early Life on Land. – American Scientist 80.
SMITH K.F. & S.M. CARPENTER (2006): Potential spread of introduced black rat (Rattus rattus) parasites to
endemic deer mice (Peromyscus maniculatus) on the California Channel Islands. – Diversity and
Distributions 12 (6).
SOUTHCOTT R.V. (1978): Australian harmful Arachnids and their allies. A guide to the identification,
symptoms and treatment of the effects caused by scorpions, ticks, spiders, millipedes and centipedes
injurious to an in the Australian region. – Southcott, Mitcham, South Australia: 1-16.
SPELDA J., REIP H.S., OLIVEIRA-BIENER U. & R.R. MELZER (2011): Barcoding Fauna Bavarica: Myriapoda – a
contribution to DNA sequence-based identifications of centipedes and millipedes (Chilopoda,
Diplopoda). – ZooKeys 156 (123-139).
TEDERSOO L., SUVI T., BEAVER K. & KOLJALG U. (2007): Ectomycorrhizal fungi of the Seychelles: diversity
patterns and host shifts from the native Vateriopsis seychellarum (Dipterocarpaceae) and Intsia bijuga
(Caesalpiniaceae) to the introduced Eucalyptus robusta (Myrtaceae), but not Pinus caribea (Pinaceae). –
New Phytologists 175 (2).
VAZIRIANZADEH B., RHAMANEI A.H. & S.A. MORAVVEJ (2007): Two cases of chilopoda (centipede) biting in
human from Ahwaz, Iran. – Pakistan Journal of Medical Sciences 23.
WEIL E. (1958): Zur Biologie der Einheimischen Geophiliden. – Leitschrift.

WESENER T., VOIGTLÄNDER K., DECKER P., OEYEN J. P., SPELDA J. & N. LINDNER (2015): First results of the German
Barcode of Life (GBOL) – Myriapoda project: Cryptic lineages in German Stenotaenia linearis (KOCH, 1835)
(Chilopoda, Geophilomorpha). – ZooKeys 510 (15-29).
WESENER T., VOIGTLÄNDER K., DECKER P., OEYEN J. P. & J. SPELDA (2016): Barcoding of Central European
Cryptops centipedes reveals large interspecific distances with ghost lineages and new species records
from Germany and Austria (Chilopoda, Scolopendromorpha). – ZooKeys 564 (21-46).
VALENTINI A., POMPANON F. & P. TABERLET (2008): DNA barcoding for ecologists. – Trends in Ecology and
Evolution 24 (2).
ZHANG D.X. & HEWITT G.M. (1996): Nuclear integrations: challenges for mitochondrial DNA markers. –
Trends in Ecology and Evolution 11 (6).
ZIMMERMANN D., SATTMAN H. & E. HARING (2013): DNA-Barcoding - von iBOL zu ABOL. – Entomologica
Austriaca 20.




















