
IgG subclass concentrations in children in health and disease
311
2-5. j,92 IgG SUBCLASS CONCENTRATIONS IN CHILDREN IN HEALTH AND DISEASE A thesis submitted by LORRAINE JOYCE BEARD, M.B.,Ch.B., F.Rá,.C.P. to the Universitv of Adelaide. South Auitralia ' For the degree of DOCTOR OF MEDICII{E Ilepartuent of Paediatrics, University of Adelaide, South Australia AugusÇ 1990
-
Upload
khangminh22 -
Category
Documents
-
view
7 -
download
0
Transcript of IgG subclass concentrations in children in health and disease

2-5. j,92
IgG SUBCLASS CONCENTRATIONS IN CHILDREN
IN HEALTH AND DISEASE
A thesis submitted by
LORRAINE JOYCE BEARD, M.B.,Ch.B., F.Rá,.C.P.
to the Universitv of Adelaide.South Auitralia '
For the degree of
DOCTOR OF MEDICII{E
Ilepartuent of Paediatrics,University of Adelaide,South Australia
AugusÇ 1990

TABLE OF CONTENTS
PAGE NO.
Ihesis Summary
Declaration
Acknowledgements
Abbreviations
Index of Tables
Index of Figurrcs
Preface
CHAPIER 1 -- Review of the literature
L.1- Introduction
L.2
L,3
8
11
72
L4
L6
19
2I
Overview of imm unsgloþtrlins
1.2.1 Immunoglobulinstructure
t.2.L.2 Immunoglobnlin classes
t.2.2 Immunoglob'rlinGsubclasses
L.2.2.L Historical bacþroundL.2.2.2 Isotlpes of IgG subclass1.2.2.3 Allotypes of IgG subclassesL.2.2.4 Relative concentrations of IgG subclasses in serumL.2.2.5 Properties of IgG subclasses
- Complement activation by IgG subclasses- Placental passage of IgG subclasses- Susceptibility of the IgG subclasses to dþestion
with proteoþtic enz5@es- Half-lives of IgG subclasses- Cellular interactions of IgG subclasses
Immunoglobulin production
1.3.1 B llmphocytes
L.3.2 Heary chain gene formation
L.3.3 The regulation of immunoglobulin production
L.3.3.L T cells- T and B cell interactions- T-independent and T-dependent antigens- T cells and histocompatibility antþens
L.3.3.2 MacrophagesL.3.3.3 Natural killer cell
22
23
23
u
n29
29
31
3l3233
33
%
37
37
TI
38
38
q
43
4444647474{ì

PAGE NO.
1.4
L.3.4 Iinmunogenetrcs
t.3.4.L Genes that influence the immune response1.3.4.2 Genes and immunoglobulin isotlpe concent¡ations1.3.4.3 Allot¡pes and disease
Evaluation of IgG subclass concent¡ations
I.4.L Quantitation of IgG subclasses
7.4.2 Normal serum concent¡ations of IgG subclasses in child¡en
IgG subclass deficiency in IgA-deficient patients
IgG subclasses and'specifi.C antibodies
I.6.I IgG subclass distribution of antibodies
1.6.1.1 Protein antþens1.6.L.2 Carbohydrate antigensI.6.1.3 Other antþensL.6.L.4 gnmm¿¡y
L.6.2 Proposed ¡ech¡nisms, of isotype restriction
L.6.2.I Models for isotype restrictionL.6.2.2 Factors that may influence the pattern of
IgG subdass production
IgG subclass concent¡ations in children with recurrent infections
IgG subdass concent¡ations in patients with recoenisedim m unodefi ciency disorders
Immunoglobulin replacement therapy
1.9.1 Intraveuous immuneglsþulin preparations
L.9.L.L Methods of preparationL.9.L.2 IgG subclass composition1.9.1.3 AntibodycontentL.9.L.4 Complement activation
L.9.2 The use qf immunogleþnlin replacement therapy i" IgGsubclass-defi cient child¡en
CHAPTER2 General Methods
2.L Immunoglobnlin quantitation
1.5
1.6
L.7
1.8
1.9
2.L.12.L.22.t.32.L.4
Selm IgA,IgG and IgMSalivary IgASerum IgEAntibody quantitation

PAGE NO.
2.2.2.L Neutrophil chemotaxis and random mobility2.2.2.2 Quantitative leucocyte io,li"ation reaction2.2.2.3 Neutrophil bactericidal and fungicidal activity2.2.2.4 Neutrophil respiratory burst
2.2.3 Lymphocytestudies
2.2.3.1 Lymphocyte phenotlping2.2.3.2 L¡mphocyte transformation to mitogens2.2.3.3 In vitro immunoglobrlin production2.2.3.4 NK cell cytotoxicity
2.2.4 Controls for cellula¡ studies
2.3 Complement
2.3.L Total haemolytic complement2.3.2 Individualcomplementcomponents
2.4 Statistical analyses
CHAPTER 3 - The development of an enz¡me-linked immunosorbent assay usingmonoclonal antibodies for the quantitation of þG subclasses
2.2 Cellular Studies
2.2.L2.2.2
3.1
3.2
3.3
3.4
3.5
3.6
3;7
Preparation of leucocytesNeutrophil studies
92
yz
94
94949696
97
97989899
101
101
101
r0z
L02
103
104
105
106
106
L09
TT4
tt4
].20
ûn
L20
L22
3.8
3.9
3.10
3.11
Rationale for developing an ELISA technique for IgG subdass quantitation
Rationale for using monoclonal antibodies
International reference standards for IgG subclasses
Overview of tÏe essential steps in ou¡ IgG subclass ELISA
Det¡ils of the IgG subclass ELISA
Accuracy of our ELISA
Comparison with other recently developed ELISA methods for IgGsubclass quantitation
Alternatives to ELISA
Comparison of RID and ELISA
Comparison of preliminary results obtained with our ELISA with thoseof other investigators
Conclusion

PAGE NO.
123
12+
125
125126
126
12ß
126VI129
150
151
152
153
L&
t67
4.L
4.2
CHAPItsR 4 - Establishing norrnal ranges for þG subclass concentrations in healthyAustralian children
Introduction and preliminary studies
Subjects and statistical analysis
4.2.L Subjects4.2.2 Statistic¿lanalysis
4.3 Results
4.4 Discussion
4.4.L Comparisons of paediatric normal rânges for IgG subclasses4.4.2 Possible rearions for differences between studies4.4.3 Conclusions
CIIAPIER 5 - ICG subclass deficiencies in lgA-delicient patients
5.1 Introduction
5.2 Patients and methods
5.3 Results
5.4 Summary
CHAPIER 6 - IgG subclass deficiency in patients with bronchiectasis
6.L Introduction
Patients
Results
Sumnary
CHAPIER 7 - IgG subclass deficiencies in infants with invasive Haemophilus influenzoetype b lnfections
7.L
6.2
6.3
6.4
168
168
L69
t70
7.2
7.3
7.4
Int¡oduction
Patients
Results
Summary
L76
t77
177
L79
184
185
186
CHAPIER I - IgG subclasses in osteomyelitis and septic arthritis
8.1 Introduction

8.2
8.3
8.4
9.1
9.2
9.3
9.4
L0.2
10.3
ro.4
1_l-.1
tL.2
tl.3
LL.4
Patients
Results
Sunmary
CHAPIER 9 - IgG subclass concentrations in children with giardiasis
Introduction
Patients
Results
Srrmmary
10.1 Introduction
Patients
Results
$nmm¿ry
Int¡oduction
Patients
Results
Summary
12.L Introduction
12.2 The composition of int¡ave¡sss imm¡asglobulin preparations
12.2.L IgG subclass content of intravenous immuûsglobulin preparations
12.2.2 Antibody content of intravenous imrnunsglsþr'lin preparations
12.3 IgG subclass replacement by immunsgls6rllin administration
12.3.L IgG subclass replacement in hlpoga--aglobulinaemic patients
12.3.1.1 Patient TN (IgA and IgG deficiency)12.3.L.2 Patient BN (IgG deficiency)
PAGE NO.
186
188
188
- IgG subclass concentrations in patients with immunological disorders
- The influence of the spleen on serum IgG subclass concenhations
- IgC subclasses and immunoglobulin replacement therapy

PAGE NO.
12-3.L.3 Patient SL (IgA and IgG deficiency)12.3.1.4 Patient MW (Ie¡A" IgG and IgM deficienry)
IgG subclass replacement i.n IgG subclass-deficient patients
?32?s
?37
231243u42A6
250
252
253
253
253254
?55
?Á2
2.63
2ß4?Á8
270n4n527621828t
2U
?35
287
311
]-2.3.2.L Patieut JB72.3.2.2 Patient RL72.3.2.3 Patient BH123.2.4 Patient AW
12.4 gumm¿¡y
CIIAPTER üt - IgG subclass concentrations in saliva and crcrebrospinat lluid
13.r Introduction
Results1i.2
L2.32
1i.2.L132.2
IgG subclasses in salivaIgG subclasses in cerebrospinal fluid
L4.T
ÍÌ3 glmm¿¡y
CHAPTER 14 - Discussiou, concrusions and Recommendations
Recotttmendations
Publications
Discussion incorporating- IgG subclass quantitation- IgA and IgG subdass deficiency- IgG4 deficiency and infection proneoess- possible -":!T^-. of infection proneness h IgG4 deficiency- treatment of IgG subclass deficiencies
ConclusionsL4.2
L43
Bibliography

B
TIIESIS STJMMARY
Athougb tle existence of fou¡ isotypes of IgG, known as IgG subclasses, has been recognised for
over 20 years, there is still a great deal of confusion about both the definition and the significance of
abnormal IgG subclass concentrations. This is partly because accurate measurement of the IgG subclasses
is <lifficult and results have often been un¡eliable. Accu¡ate normal rânges for patients of different ages
have, therefore, been difficult to establish. Studies of patients with subclass abnormalities have been largely
anccdotal. It has, however, been suggested that IgG subclass deficiencies may prove to be the most
common form of primary immr¡rsdeficiency.
In many children who suffer recurrent infections, standa¡d immuno function studies including
mcasurcmcnts of IgA, IgG and IgM, neutrophil and þmphocytc function studics and serum complement
sç¡esning fails to detect a particular immunological problem. Some investþators have suggested that an
IgG subclass deficiency may exist in a considerable proportion of tlese patients and may be associated with
impaired production of antibodies to a variety of antigens. Other studies suggest that IgG subclass
deficiency may not be an important factor. The chief aim of this project was to help to clarify ss6e sf rhis
confusion by developing a reliable enzyme-linked immunosorbent assay (ELISA) technique and using this
a) establish percentile ranges for IgG subclasses in healthyAust¡alian children.
quantitate serum IgG subclasses in groups of patients with differing types and degrees of
infection proneness.
quantitate serially serum IgG subclasses in child¡en with immuuodeficiency states who a¡e
receiving regular immr¡rsglsþrrlin infusions.
b)
These studies have increased ou¡ understandi"g of the relationship between IgG subclass
deficiencies and infection pronenoss in children, and have resulted in suggestions for directions for fu¡ther
work in this a¡ea.
to
Ð
The ELISA technique was developed using monoclonal antibodies to each of the IgG subclasses.

9
Serum samples from healthy Aust¡alian children were used to determine age-related percentfe
rånges for IgG subclasses. Previous studies have generally used less sensitive assay techniques and have not
been able to define tle lower limit of normal for IgG4 which is found in relatively low concentrations in
many normal sera. We were able to quantitate IgG4 in all ou¡ healthy subjects..
The relationship between IgA deficiency and IgG subclass deficiency was studied. In a preliminary
study we found that wbile infection-prone children with IgA deficiencies were more likely to have IgG
subclass deficiencies than were non-IgA deficient children, those with profound IgA deficiencies were less
likely to have IgG subclass deficiencies than those with less profound IgA deficiencies. In this preliminary
stud¡ IgG subclass quantitation was done by electroirnmuno¿ìfisay and we were unable to determine the
incidence of IgG4 deficiency because of limi¡¿¡i6^ in the sensitivity of the assay. In a second stud¡ we
foirnd IgG4 deficiency to be the most common IgG subclass deficiency associated with IgA deficiency.
In two other groups of patients, low IgG4 concentrations were not uncommon. Of fifteen patients
with bronchiectasß, ?JlVo were IgG4-deficient and a group of child¡en with invasive Høemophitus
influenznz type b infections had low, or relatively low, IgG4 concentrations.
Flowever, IgG subclass deficiencies were not common in groups of children with osteomyelitis or
septic arthritis or giardiasis. IgG subclass deficiencies, and particularly those of IgG4, appeared more often
amongsf patients with proneness to infections of the respiratory tract, or infections with pathogens gaining
entry via the respiratory tract, tlan amongst the patients with other types of infections.
To contribute to the understanding of the control of IgG subclass production we quantitated IgG
subclasses in children with a variety ef immunslogical disorders where control ¡nesþanisms ar€
dysfunctional.. We also studied the effect of the spleen on IgG subclass concentrations by quantitating IgG
subclasses in two groups of splenectomized patients and in a small group of child¡en with portal
hlpertension.
Finall5 we studied the IgG subclass composition of several int¡avenous immunoglobulin
preparations and then investþated the effect of immunoglobrrlin replacement therapy on IgG subclass
coucentrations in patients with either hy¡logammaglobrlinaemia or IgG subclass deficiency. While most

10
infection-prone patients with either hypogammaglobulinaemia or IgG subclass deficiency without
generalised hlpogrmmaglobuli"aemia showed marked clinical inprovement with regular immuneglsþulin
replacement therapy, the degree of improvement did not always parallel the improvement in the IgG
subclass concentrations.
Overa[ the relative frequency of IgG subclass deficiencies in the patients studied was: lgp4 44Vo,
IgG2 ZZVo,lgGL L8% and IgG3 15Vo. The frequency order was ttre reverse of the order of the IgC subclass
heavy chain gene sequence, suggesfng that the g¡eater the amount of downstream isotlpe switching
required, the more likely is a deficiency. The combination of isotype deficiencies found most frequently
was that of IgA and IgG4. While this suggests a defect in isotype switching at the genetic level the fact that
IgG2 deficiency occurred much less often indicates that more tlan only an arresfing of isotype sv,,i¡ching is
involved. Even in patients deñcient in one or more IgG subclass isoty¡res, the deficient isotype was always
detectable, indicating that the defect in production was probably regulatory ratler than st¡uctural.
While IgG subclass deficiencies a¡e associated with proneness to some types of infections i¡
children, further studies are necessary to clarify tle relationship and to develop oprim¿l ways of t¡e¿ting
infection-prone IgG subclass-deficient patients. Potentially useful areas of futu¡e study would include:-
(Ð relationships between IgG subclass deficiencies and functional antibody deficiencies.
(ü) the inportance of IgG subclasses in mucosal secretions.
(Ð -sçþanisms of controlling IgG subclass production.

11
DECI,ARATION
This thesis ç6¡fains no material which has been accepted for the awa¡d of any otler degree or
cliploma io aoy university. The work is my own except where otherwise acknowledged.
To the best of my knowledge and belief this thesis contains no material that has been published
previously or written by another person, except where due reference is made in the text of the thesis.
I consent to the thesis being made available for photocopying and loan, if applicable, if it is
accepted for the awa¡d of the degree.
LJ. BEARI)

L2
ACKNOWLEDGEMENTS
The s[udies described h this tlesis were plenned and carried out from 1984 to 1990 in the
University of Adelaide, Department of Paediatrics and the Department of Tmmunology at the Adelaide
Children's Hospital (ACH). They involved the cooperation and help of a large number of people to whom
I am deeply thanKul.
Dr Antonio Ferrante, Head of the Departms¡f sf rmmunology, ACH, provided untiring academic
stimglation, and together with Professor George Maxwelt was my supervisor in the work of this thesis.
Without his constant encouragement it would never have been completed.
The laboratory staff of the Department of rmmunologl, ACH provided technical assistance.
Brenton Rowan-Kelly and David Goh carried out tle various immune assays and Julie Hagedorn tirelessly
performed IgG subclass quantitations. The Department of Microbiolog5l, ACH, and by the rnsdtute of
Medical and Veterinary Science, Adelaide assisted by determining antibody levels to viral and microbial
antigens.
I owe very special thanks to Mrs Pru Russell-Taylor for her dedicated secretarial assistance, and to
M¡s Nance Shiell and Miss Colleen Royal for spirirual support and encouragement.
Sister Helen Forsyt\ from the Immuniza¡is¡ Çlinig ACH, provided valuable help in enrslling
child¡en for the normal range studies, and Mr Philip Leppa.4 from the Department of Statistics of the
University of Adelaide provided essential statistical help.
Many medical colleagues were involved in providing clinical information and/or referring patients
included in these studies. Dr Linda Ferris (Orthopaedic Registrar, ACÐ, Dr James Martin (Paediatric
Pulmonologist, ACÐ, Dr Rima Staugus (Paediatric Thoracic Physician, ACÐ, Dr Margaret Dean (Senior
Iæcturer, University of Adelaide), Dr Greg Smith (Visiting Paediatrician, ACH), Dr Eveþ Robertson
(Chemical Pathologist, ACH), Dr Geoffrey Davidson (Paediatric Gastroenterologist, ACH), Dr George
Kiroff (General Surgeon), Dr Ian Toogood (Paediatric Oncologist, ACH) and Dr Chris Pearson (Visiting
Medical Specialist, ACH) deserve special mention. Most of the artwork was done by Mrs Colleen Lloyd.

13
I would like to thank my colleagues in the Department of Paediatrics, University of Adelaide,
particularly Professors George Maxwell and Don Roberton who made it possible for me to do these studies
while employed in the Department. I am grateful to Dr Vivi-Anne Oxelius (University Hospita[ Lund"
Sweden) for heþ in measuring IgG subclasses when I was beginning this work and for encourâgng
discussions and to Professor Yee Hing Thong (formerly Senior Lecturer, University of Adelaide, and
currently Professor of Child Healtb, University of Queensland) for stimulating my interest in paediatric
imm¡¡6lsgy.
The work was supported" in part, by grants from the University of Adelaide and from the Channel
10 Childrens Medical Resea¡ch Foundation of South Australia.

I4
BCDFBCGFBSScCL,C2etc.CDChCMVConACPSCSFCSLCTCVCVHCVIDDEAEdlDMSODNAdPmEBVEIAELISAESRFACSFcRFCSFMLPgGmH chainHBSSH. influenzaeHibHTVHI-AHMPHRPI{ILVICSIFNIgIgAIgEIsGIsMILIMIGIU$IVIGJIL chainM.W.MCMcAbttci
ABBREVIATIONS
B cell differentiation factorB cell growth factorbalanced salt solutionconstantcomplement factors t, 2 etc.cluster differentiationchaptercytomegalovirusconcanavalinAcapsular polysaccharidecerebrospinal fluidCom m onwealth Serum Laboratoriescomputerised tomographycocfEcient of variationcommon variable hypogam-aglobrrlinaemiacomm on va¡iable ìmmunodeficiencydiversitydiethylaminoethyldecilitredi-ethyl sulfoxidedeoxyribonucleic aciddisintcgratiom pcr minutcEpstein-Barr viruselectroimmutroassayenz5meJinked immunosorbent assayerythrocyte serlim entation ratefluorescent - activated cell sorterFc receptorfoetal calf serumN- formyl L- methionyl L- leucyl L-phenylalani''eg^rrrallotype marker on human IgGheavy chainHank's balanced salt solutionH øemophilus influenzneHaemophilus influenzne type bhuman immunodeficiency virushuman leucoryte antþenhexose monophosphatehorse radish peroxidasehunan T cell leukaemia vi¡usim m r¡ros[smistry systemsinterferonimm¡agglsþnlinimmungglsþrlin Aimmunoglobulin Eimmunoglobulin Gimm¡¡sgloþulin Minterleukinintramuscular imm ¡¡sgfoþulinInternational Union of Immunological Societiesintravenous immunoglobulinjoininglit¡elight chainmolecular weightmonoclonalmonoclonal antibodiesmicrocurie

15
ABBREVIATIONS CONT.
ItgmgMHCplmlMNLNADPHngNKnm
D.S.
ODpPBSPCP. cørinü
PHAPMNLPWMRFRIARIDRNAS, øureusS, pneumoniazSDSESLESRBCTDTIURTIVVCAw/vwHo
ûucrogrammilligranmajor histocompatibility complexmicrolitremillilitremononuclear leucocytenicotine adenine diphosphate (reduced)nanogramnatural killernanometernot significantoptical densityprobabilityphosphate buffered salinepolyclonalPneumocystis cafinüphytohaemagglutininpollmorphonuclear leucocytespokeweed mitogenrheumatoid factors¡¿figìmm¡¡oãisayradial immunodiffusionribonucleic acidStøphylococcus aureusStreptococcus pneumoniaestanda¡d deviationstandard errorsystemic lupus erythematosussheep red blood cellsT cell-dependentT cell-independentupper respiratory tract infectionvariableviral capsid antþenweight/volumeWorld Health Organisation

1.1
r.2
L.3
t.4
1.5
1.6
1.7
1.8
2.1
2..2
3.1
3.2
33
4.L
4.2
4.3
4.4
45
4.6
4.7
4.8
4.9
INDEX OF TABLES
Physiochemical properties of IgG subclasses
Biological properties of IgG subclasses
Serum IgG subcless concentratioos in normal adults
Predomin¡nt isotypes of IgG antibodies to protein antþens
Predominant isotypes of IgG antibodies to carbohydrate antigens
Predominant isotypes of IgG antibodies to other antigens
IgG subclass deficiencies in patients with recurreut i¡fections
IgG subcl¡s5es in some immunodeficiency states
. Normal rângqs for ser'- Ig,\ IgG and IgM
IgE reference values for children
IgG subclass levels in reference sera
Problem results indicati-g allotype restriction for tre IgGl McAb sG16
A comparisoo of IgG subcrass measuremeuts usiog RID ¿¡d ELISA
Serum IgG subdass concent¡ations at different ages, mean+ SD
Preliminary XJVo cosfrdence limits for serum IgG subclass concent¡ationsin healthy Australi¡ n child¡.eq
Serum IgG subclass percentiles in healthy Australian adults
Serum IgG subclass X)Vo cnnfidence limits in healthy Australian malecj'ild¡en
Serym IgG subclass X)Vo coútdence limi¡s in healthyAustralian femalechild¡en
Serum IgG subdass X)Vo cnútderrçs limits in healthyAl¡5[¡.¡li¡n c],ild¡en(males and females combined)
Serm IgG subclass 95vo cnofidencc rimig in hearthyAustralian malechild¡'eu
Serym tgG subdass gsvo cnsfrdence limig in healthyAustralian femalechild¡'sa
serum IgG subclass 95vo cnsfidepçs rimits in hearthy Australian femarecbild¡eu (males and females combined)
Mean relative percentages 9f IgG subclasses in serum at di-fferent ages
Published paediatric tgG subclass normal ranges
Sr r m mary sf cl i n ical ¡ '' d immu!,ologicar abnormarities n 22 rg^-defi cient
children with recurrent or severe reipiratory i¡fections. -q - i
L6
PAGE NO.
:34
35
53
58
61
67
73
77
91
93
108
111
72L
131
732
133
1i4
ß5
lX
1i7
ß8
ü19
1¿lO
L4L
754
4.10
4.LL
5.1

5.2
5.3
5.4
5.5
6.L
6.2
6.3
6.4
6.5
7.L
7.2
7.3
7.4
7.5
8.1
8.2
9.1
9.2
10.1
L0.2
10.3
L0.4
10.5
Incidence of IgG subclass deficiencies in slmptomatic lgA-deficientpatients
þiagnoses n2i7lgÃ-defrøent patients with IgG subclass deficiencies
IgG4 deficiency n73lgA-deficient patients with and without recurrentinfections
Absolute concent¡ations of IgG4 in 73 IgA-deficient patients and inage-matched controls
Serum immunoglobulin concentrations in 15 patients with bronchiectasis
Isotlpes of IgG subclass deficiencies in patients with bronchiectasis
Serum IgG subclass concent¡ations in adults with bronchiectasis
Response to pneumocoscal immrlnisation in patient 8
Llm.phocyte and neutrophil studies in patients with bronchiectasis andin controls
Clinical details of patients with Hib disease
Conccntrations of IgA, IgG, IgM, IgE, C3 and C4 in infants withinvasive Hib infections
IgG subclass concentrations in infants with invasive Hib disease
Mean IgG subclass concent¡ations in infants with invasive Hibdisease
Lymphocyte and neutrophil studies in infants with invasive Hibi¡fections and in controls
Clinical ds¡ails of patients with osteomyelitis or septicarth¡itis
Immunoglobrlin i5ef¡çre deficiencies in patients who have had
osteomyelitis or septic a¡thritis
Serum immunoglobulin and complement concentrations in 30
children with giardiasis
Llmphocyte and neutrophil studies in children with giardiasis
and in cont¡ols
Clinical details and concentrations of immunoglobulin isot¡'pes
in symptomatic patients with varying degrees of hypogammaglob'linaemia
rmmunoglobulin isotlpe concentrations in children with recognisedimmunodeficiency disorders
Immunoglobrlin issqps concentrations in patients withi m m unoregulatory disorders
Immunoglobnlin isogpe concentrations in families ofhlpogammaglobulinaemic children
In vitro production of Ig isotypes by cultured peripheralblood lymphocytes from 3 hlpogammaglobulinaemic patients
160
r6L
t62
L63
L7L
172
173
L74
t75
t78
180
181
L82
183
L87
190
L99
200
207
248
249
2W
2L0

1L.1
Lt.2
LL.3
T2.L
L2.2
t2.3
12.4
12.5
12.6
IgA, IgG, IgM, IgE, C3 and C4 concentrations in splenectonizedadults and controls
Mean IgG subclass concentrations and percentages in 16 splenectomizedadults and controls
IgG subclass concentrations above the 95th or below the 5th percentilein splenectomized patients and in patients with portal hy¡rertension
Interstudy comparisons of IgG subclass composition of various intravenousi m m¡ssglsþrllin preparations
Mean percentages of IgG subclasses in 3 intravensgs immunsglobullnpreparations
Antibody titres in 3 int¡avenous immunoglobulin preparations
Comparison of pneumococcal antibody levels in 3 intravenous immunoglobulin
preparations
Lymphocyte, neutrophil and complement studies in hypogammaglobulinaemicpatients
Llmphocyte, neutrophil and complement studies in IgG subclass-deficientpatients
Responses to pneumococcal immunisation in 3 IgG subclass-deficient patients
IgG subclass composition of saliva and serum in healthy subjects
IgG subclass, IgG, IgA and IgM concentrations in CSF
Overall distribution of IgG subclass deficiencies in the patientgroups studied
2n
2L8
2L9
223
224
226
2n
228
239
2Q
256
257
?32
12.7
li.1
li.2
L4.L

INDEX OF FIGTJRES
Diagramatic representation of an IgGl molecule
Ig heavy chein gene sequence on ch¡omosome L4
Schematic diugru* of the organisation and assembly of the humanheavy chain gene.
Schematic represeutation of the effects of preparation procedureson immu¡sgl¡þnlins for intravenous use.
Fracturation of blood leucocytes into the main populations by therapid sinsls-step method.
Graph of the mathematical function to which NK cytotoxicity datais fitted.
schematic representation of ou¡ ELISA for IgG subclass quantitation.
The Biomek 1000 Worktation
Sl.¿rnda¡d curves showing tle relationship between OD414nm and IgGlconcentration in the ELISA.
Standard curves showing ti.e relalionship between oD414nm andlgG}concentration in the ELISA.
Standa¡d curves showing the relationship between OD414nm and IgG3concentration in the ELISA.
Standard curves showing tle relationship between OD414nm and IgG4conc€ntration in the ELISA.
schematic representation of commercially available ELISA for IgGsubclass quantitation.
IgGl and IgG2 percentiles in healthy Australian male chitd¡.ss.
IgG3 and IgG4 percentiles in healthy Australian male child¡en.
IgGl and IgG2 percentiles in healthy Australian female child¡.en.
IgG3 and IgG4 percentiles in healthyAustralian female child¡en.
IgGl and IgG2 percentiles in healthy Australian chitd¡en (malesand females combined).
IgG3 and IgG4 percentiles in healthy Australian child¡en (malesand females combined).
Lower limits for IgG subclasses in published paediatric studies.
Mean or meclian concentrations for IgG subclasses in publishedpaediatric series.
1"._* IgA IgG and IgM concentrations and associated IgG subclassdeficiencies n ?2 lg{-deficient child¡en.
Serum IgG subclass concentrations inn rgA-deficient chird¡.en withrecurrent or severe respiratory infections.
I9
PAGE NO.
25
4l
42
84
95
100
t07
1Íl
115
116
rL1
118
11,9
r4z
t43
L44
t45
146
r47
148
149
',155
156

5.3
5.4
8.L
8.2
8.3
8.4
10.1
L0.2
12:1.
12.2
12.3
12.4
12.5
L2.6
12.7
12.8
12.9
73.r
13.2
13.3
13.4
Neutrophil functions in IgA-deficient children with recurrent or severerespiratory infections.
Concentrations of the complement components C3 and C4 and haemoþticcomplement activity in the serum of lgA-deficient children with recurrentor severe respiratory infections.
Serum IgA" IgG and IgM concentrations in patients who have had osteomyelitisor septic arth¡itis.
Mitogen-induced llmphocyte prolifeation in patients who have had osteomyelitisor septic arthritis and in controls.
Mononuclear leucocyte populations in peripheral blood in patients who havehad osteomyelitis or septic arthritis and in controls.
NK cell cytotoxicity in patients who have had osteomyelitis or septic arthritisand in controls.
In vitro production of Ig isotlpes by cultured peripheral blood llmphocytesof patient JB in response to srimulation with PWM at different coucentrations.
In vitro production of Ig isotypes by cultured peripheral blood llmphocytesof patient JB in response ts sf inrul¿tiol with S. u#eus ,
NK cell cytotoxicity in patient BN, age 10 montls, and brother TN age 6 years.
Trough concentrations of IgG and IgG subclasses in patient TN.
Tro'rqh concentrations of IgG and IgG subclasses in patient BN.
Trough concent¡ations ofIgG and IgG subclasses in patient SL.
T[s sþanging pattern of Ig class deficiency in patient JB both before and¿fts¡ st¿¡fing IVIG therapy.
Trough concentrations of IgG and IgG subclasses in patient JB.
Trough concentrations of IgG and IgG subclasses in patient RL.
Trougü concent¡ations of IgG subclasses in patient BH.
Trougû concentrations of IgG and IgG subclasses in patient AW.
IgGl in saliva and serum of 19 healthy adults.
lgct2ln saliva and serum of 1-9 healthy adults.
IgG3 in saliva and serum of 19 healthy adults.
IgG4 in saliva and serum of 19 healthy adults.
158
159
L91
tyz
193
t94
2tL
212
2n
23t
233
235
v+t
212
ù+5
247
249
258
259
2û
26t

2T
PREFACE
The laboratory methods used throughout these studies a¡e described in Chapters 2 aú,3. Separate studies
form the basis of Chapters 4 to 11. The results of each of these studies are discussed in detail in Chapter 12
which integrates tle findings of all the studies. Tables and figures appear ei¡þs¡ immsdiately following
thei¡ ¡eference in the text or at the end of the appropriate chapters.
Much of tle work included in this thesis has been published or accepted for publication. A list of key
publications follows the bibliography.

CHAPTER ONE
REVIEW OF THE LITERATURE

I..1 INIR.ODUCTION
Tmm¡¡sglsþulin proteins are a major component of the body's imm¡¡o[sgical defence system.
The major classes of the immun6glsþrlins have been recogrrised and studied extensively. IgG is the
predominant immunoglobulin class in serum and is made up of four subclasses, IgG1, IgG2, IgG3 and
IgG4. Relatively little is known about the importance of each of these subclasses and tl.ere is considerable
disparity between tle conclusions that can be d¡awn fron different published studies. This chapter reviews
from the literature, the present state of knowledge, considering the structu¡e and function of
immuneglsþulins, especially IgG subclasses, and the produclion of immunsglo6ulins and IgG subclasses,
problems in IgG subclass quantitation and in the establishment of normal ranges. The suspected
association of IgG subclass deficiency with infection-proneness and with antibody deficiencies and the
inconsistency of some of the find;nss reported to date ¿¡s discussed. Finall¡ the use sf immuneglsþ'lin
replacement therapy in IgG subclass deficiency states is reviewed.
I.2 OVERVIEW OF IMMTJNOGLOBTJLINS
Immunoglob rlins a¡e glycoproteins which have key roles in immune responses. They are found in
many parts of the body, and are present in relatively large amounts in serum. Immunoglobrlins recognise
and combine with antþens either in body fluids or on cell su¡faces. They can act as specific mediators of
humoral immunity in the following ways:
1. By neutralising vi¡uses or toxins and so preventing their interaction with target cells or molecules.
They may also prevent the adhesion of pathogenic bacteria to cell su¡faces in the gastrointenstinal
and genitourinary tracts.
2. {çfiy¿fing complement to induce lysis of target cells or bacteria.
By acting with effector cells to c¿use death of target cells (i.e. antibody-dependent cellula¡
cytotoxicity). The effector cells have no immunological specificity but have receptors for the
constant (Fc) part of the IgG molecule.
3.

4.
24
By promoting phagocytosiß of micro-organisms (with or without complement) by pollmorphs or
macrophages.
The combined activity of antibody, complement and neutrophils is of major importance in
combating most bacterial infections. Opsonisation of bacteria is an essential phase in the recognition of
microorganisms before phagocytic cell killing.
In the 1930s, Tiselius (1937, L939-40), io Swedeq developed an electrophoretic technique and
separated serum proteins on tle basis of elect¡ical charge into a number of peaks (a,6 and'y globulins and
alb'min). Then, in 1939, Tiselius and Kabat (f.939) showed that the serum of a hyperimmunìsed rabbit had
an increased "y peak. Subsequently the term 'y -globnlins was used to refer to antibodies. Later, however, it
was shown that not all'¡,-fraction proteins were antibodies and that not all antibodies were in the "y-band.
Hence, tle term immunoglobulin was introduced to refer to globulins with an antigen-binding function.
Tmmunoglsþ 'lins account for a substantial proportion (li-A%) of human serum proteins (Poljak, 1983).
12.1 Immunoglobulin structure
The st¡ucture and function of immunoglobulins have been studied eÍensively. Plasmacytomas of
human and murine origin have provided homogeneous (monoclonal) immunoglobulins which have
facilitated greatly the study of tlese molecules (Poljak, 1983),
Tmm¡a6glsþulin molecules ¿tre made up of poþeptide chains. Each has a basic molecule
consisti'g of two heavy and two lþht chains. There a¡e five maitt classes sf immunoglobulins, IgG, IgM,
IgG, IÐ, IgE. IgG which is most abundant in the serum, has been studied most. IgA predominates in
mucosal secretions. Each class is defined by its unique heavy (H) chain We 0 p p,6 or e ). All classes
share the same liqht (L) chains (rc or.\).
An IgG molecule consists of two identical L chains each of molecular weight Z),000-25,000,
çs¡sisting of about 214 amì¡o acids, and two identical H chains each of M.W. 50,000-55,000 and consisting
of about 450 anino acids. Each chain c¿n be subdivided into homology regions (domains) of va¡iable
(Ð-d constant (C) sequences, each of about 100-110 amins acids (Poljaþ 1983), Fig. 1.1.

25
tH
l.Y/,t7
c\
c------>coo-
coo-I
I
I
I
<J
..1l\
-\,
r'
PAPAI N IPEPSIN
Fau F.
Fig. l.f Dlagramatlc representation of au þGl molecule indicating homologr regioos ot the beavy chalns (VH, CHl,CtI2 and CfI3), the homologr regions ot the light chaf ns (VIn CL) ioter- and intra- chal¡ dlsulphide bonds ( ¡-7_), thevariable region (V), tbe constant region (C), the hlnge regioo (H) and the cleavage polnts of papain and pepsin.
x
x

26
Light cháins consist of VL and CL regions. Heavy chains consist of one VH, and several CH
d6pains referred to as CHL, CH2, CH3 and CH4. There are five domains in the ¡¡ and e , and four in the
T and o chains. The four poþeptide chains that make up an immumoglobrrlin molecule are covalently
linked by interchain disulphide bonds. Molecules are so folded that the dsmains form globular regions.
They are held together by intrachain disulphide bonds. The zone where the V and C regions join is called
the 'switch' region. The a¡ea of the H chains in the C region between the fi¡st and second ç do¡ains (CH1
and CtI2) is called ¡¡s 'hinge' region. This a¡ea is flexible and most easily e>çosed to enz5mes and
chemicals (Goodman, L982a).
The variable regions contain the amino-terminal portion of the poþeptide çþains, and the
constant region, the carbory-termin¿[ portion. Proteolytic digestion cleaves immunoglobrlin molecules,
mostly belween the CH1 and CH2 de6ains. Papain splits the molecule on tle N-terminal side of the
interchain disulphide bonds into 3 fraqnents of similar size, i.e.2 Fab (including the entire L chain and the
VH and CH1 portions of the H shain), and one Fc portion (the C terminal parts of the H chains). Pepsin
cleaves the molecule on the C terminal side of the inter H chain disulphide bonds, resulting in a large F
(ab')2 fragment composed of roughly two Fab fragrnents The Fc fragment is extensively degraded by
pepsin (Shakib & Stanwort\ 1980a).
Antigen-bin.ling activity is associated with the VH and VL domains of the Fab fragment. There is
t¡emendous variability in the aminoacid residues in the V region, giving rise to over one million possible
antigen-combining sites. Most of tle secondary biotogical activities, such as complement fixation, are
associated with the Fc fragment. The structural heterogencity of the constant region is responsible for the
differences in molecular weight, mobility and biochemical properties, togetler with differences in effector
functions of the different immunoglobulin isotlpes (eg. Cl-q fixation, placental transfer, passive cutaneous
anaphylaxis, binding to receptors on \mphocytes, mononuclear cells, and neutrophils and binding of
rheumatoid factor).

27
12.12 Immunoglobulin classes
IcA
This is the predominant immunoglobulin in mucosal secretions where it occurs largely ¿s ¿ dime¡
containing a joining (J) chain and a glycopeptide c¡lled the secretory component, or secretory piece. Both
the J chain and the H and ¡¡" ¡ sþains of IgA are produced by lymphocytes, while the secretory component
is thought to be produced by epithelial cells.While patients with absent serum IgA almssl always have
absent secretory IgA (Amman" & Hong, 1971; Stanley & Cole, 1985), it is possible to have normal
secretory IgA but low serum IgA (Ammann & Hong, L97\), or to have low levels of secretory IgA, but
normal serum levels (Strober et al 1976). There is only one reported case of absent secretory IgA in the
presence of normal serum IgA, and this was attributed to tle absence of tle secretory piece (Stockley et al,
L981, Strober et d, L976).
Although IgA comprises only abott 15Vo of serum immunoglobrlin, over half the body's total
antibodies are IgA (Hanson et a[ 1988). In humans, it exists chiefly as a 75 monomer, but po\m.eric forms
of up to 1&S c¿n also be detected. About 9OVo of serum IgA is IgA1, whereas in secretions, IgAl and IgA2
occur in almssl equal proportions. Selective deficienry of.lgÃ2 has been reported (Van Inghem et al,
1e83).
Antigens introduced to the mucosal surface tend to evoke the secretion of IgA. Pa¡enteral
administration of the same antigen generally results in ser '- IgG and IgM with little antibody response on
tle mucosal surface (Ogfa 1-968). IgA is abundant in saliva, tears, bronchial secretions, vagin¿l secretions,
nasal mucosa" prostatic secretions and mucus secretions of the small intestine. It may function by
destroying the antigens or by preventing the access of antigens to the immune system by inhibiting
adherence, colonization or absorption (Kilian et al, 1988). The role of seru- IgA is unclear. Selective IgA
deficiency is considered to be very common, with a prevalence of between f- in 300 (Clark et a[ 1983) and 1
in 3000 (Frommell et al, L973) in different populations. IgA deficiency is often familial (Oen et al" 1982). It
seems to be in some way associated with HI-A B8 (Ambrus et al,1977, Oen et al, L982, Hamm¿¡s6s6 g
Smith, L983, Keikkila et al, 1984). Associations with autoimmune diseases (Ammann & Hong, L97t;Petty
et a7, L979), atopic allergy (Collins-Williams et al 1968) and various gastrointestinal disorders, such as
coeliac and Crohn's diseases (Hodgson and Jewell, 1977), have been established. Many IgA-deficient
subjects, however, are quite healthy.

28
Surprisingl¡ longitudinal studies on the uatural history of IgA deficiency a¡e hard to find. Gillon
et al (L986) found that 4 of. L2IgA deficient donors had normal IgA levels when re-tested 6 months later.
Laschinge et al (D8a) found fluctuating IgA levels in lgA-deficient subjects'
In families with selective IgA deficiency, abnormal concentrations of other immunoglobnlì"
isotypes are not uncommou (Burks and Steele, l-986). Ar increased incidence of IgG subclass and antibody
deficiencies has been reported i" IgA deficient subjects. This will be discussed further in section 1.5 and
1.6, and Chapters 5 and 14.
@
This accounts for only 0.2Vo of. senrm immunoglsþulin. There is evidence to indicate that its prime
role is as an antþen receptor on tle su¡face membranes of llmphocytes, along with IgM (Poljalq 1983).
IgM and IgD account for a large proportion of the surfaca immuneglobrrlin on B cells (Saxon and Stiehm,
1_e8e).
The role of serum IgD has not been defined. There is considerable biological variation between
IgD concentrations in subjects of the same age (Hiemstra et al, 1989). Raised levels of IgD occu¡ in some
imm¡¡6dsficiency states and in tle 'periodic fever hyper-IgD slmdrome' described by van der Meer (1984).
IgM
The heavy chain of this molecule fr), which has a molecular weight of 65-70000, has an additional
homology region (domain) designated CH4. IgM exists in serum as a pentåmer and so¡feins a poþeptide
J chain. IgM frequently occurs as a natural antibody (i.e. an antibody with binding activity for an antþen to
which the organism has never been exposed" e.g. naturally-occurring blood group antibodies). It is often
the first class of immunsglsþulin to be produced in the primary phase of immune response, suggesting that
it is important as a first line of humoral defence when an antþen invades the circulation. It is extremely
efEcient in activating complement. { 5ingls molecule bound to antþen can activate the complement
cascade (Goodman, L982a). IgM along with IgD serves as a receptor on the su¡face of B \mphocytes
where it occurs as a monomer.

29
4E
IgE comprises only about 0.0MVo of total serum immunsglqþrrlin. l¡ exists in senrm in monomeric
form. It binds strongly to mast cells and basophils by a binrring site in the Fc region. When it combines
with certain specific antþens (termed allergens) the mast cell releases a number of pharmacological
mediators which result in allergic reactions (Goodman, L982a). Beneficial effects of IgE may include
protection agains¡ intestinal worm parasites and against invasive parasitic infections. Studies in the rat
suggest that IgE may damage wonn parasites directly and by €using the release of pharmacological
mediators from many cells which contribute to worm e4pulsion. Schistosomes which enter the blood
stream, are more efEciently destroyed by macrophages in the presence of IgE antibodies which bind to
macrophages by their Fc portions (Saxon and Stiehm, 1989).
IgE spthesis is hþhly dependent on T cell regulation. Profound antibody and cellula¡
immunodeficiencies are associated with low IgE concent¡ations, while in some partial immunodeficiency
states eg. Wiskott-Aldrich slmdrome high concentrations of IgE are characteristic (Saxon and Stieh-,
1e8e).
Æ
This is tle predomit'ant serum immr¡nqglsbu.lin, comprising about 75Vo of. the total serum
immunoglobnlin. It is the only class of immurqglsbuli" that can cross the human placenta. It is made up
of four subclasses, desþated IgGl, IgG2, IgG3 and IgG4 (Goodman, 1982a) and will be discussed in detail
in the following section.
I22 Immunoglobin G subclasses (IgG subclasses)
122í Historicalbacþround
b L964, following the discovery of groups of anim¿l gamnaglobnlin, it became apparent that
human gammaglobulin could be subdivided into fou¡ subclasses (Ballieux 1964; Gray and Kunkel, 1964;
Terry and Fahe¡ 1964). These subclasses were füst called '¡1,'t2,'t3 and'¡4 and later as IgGl,IgG2,
1gG3 and IgG4, on the basis of their relative concent¡ations in normal serum and the relative frequencies
of their oocurrence as myeloma proteins (Kunket et at 1966).

30
The füst investþator to describe IgG subclasses was Korngold in 1f)61. In retrospect, it seems tlat
he may have recognised rc and I chains and possibly only one IgG subclass (Schur, 1988). Dra¡ in 1960,
had detected three ty¡res of IgG in human serum. b. L96r', Grey and Kunkel (1964) were the füst to
recopise four IgG subclasses in maq and soon after Terry and Fahey Gg6/.) confirmed these findings.
IgG subclasses were first defined on tle basis of antþenic differences in their heary chains.
Antisera raised in rabbits and monkeys were used to analyse these proteins, usuallyby Ouchterlony double
diffr¡sion in agat gel (Grey & Kunkel t9&), or by imm¡¡selectrophoresis (Terry & Fahey 1964).
Subsequently, radial immunodiffusion. (Yount et al" \W; Schur 1970) and haemagglutination were used
(Yount et a[ 1970). More recently, enzyme-linked immunosorbent assays have been developed @utlor et
al, 1980;^ Papadea et al, 1985; Aucouturier et al, I-985; Ferrante et al 1986). Later these subclasses were
recognised to have unique structural physiochemical, genetic and functional characteristics (Shakib and
Stanwort\ 1980 a and b).
The fi¡st reports of increased susceptibility to infections in patients with IgG subclass deficiencies
appeared in the late 1960's (Terry, 1968; Rivat et al 1969). Shortly after, abnormal IgG subclass patterns
were described in patients with h5poganmaglobulinaemia (Virella et al" L970; I*ddy et aJ,1970; Yount et
aJ\Lnq.
Schur et al, in 1970, reported tlree patients with life long susceptibility to pyogenic infections. All
tbree had various mmbinations of IgG subclass deficiencies with normal serum IgA and IgM. However,
¡fre siFificance of IgG subclass deficiency as distinct from common variable immunodeficiency was not
recognised at that time.
Ia.t974, Oxelius desc¡ibed a fanily (a mother and two children) with IgG2 and IgG4 deficiencies,
re¿urrent bronchopneumonia and otitis. The most frequently isolated causative organism was Hæmophilus
influenue. Total serum IgG was normal but there was an absence of antibodies to teichoic acid and
H. influenzac. Normal antibodyrise followed rubella immunisation.
Since 1981, when Oxelius et al reported in the New England Journal of Medicine, the finding of
IgG subclass deficiencies in patients with IgA deficienc¡ interest in the possible clinical significance of IgG

31
subclass deficiencies has increased considerably. Subtle defects in immune function are common in
patients with recurrent respiratory infections (Beard et a[ 198L) and IgG subclass deficiencies may be
particularly si gnifi cant.
1222 Isotypes of þG subclasses
The antigenic differences that charactenzn the class and subclasses of the H chains and the types
and subt¡res of the lþht chains are called isotlpes. Each isotlpe has a distinct locus in the genome. Wbile
a particular subclass isotlpe may have several alternative structures, the heavy chains of any one particular
subclass (withi" a class) are much more similar to each other than to those of the otler classes. The
alternative structures of each particular isotlpe are inherited in a Mendelian fashion. The polymorphic
forms of.the isotypes are referred to as allotlpes. Generall¡ the antþenic determinants characterisiog the
allotlpes are in the C regions. Each normal individual possesses all the immunoglobulin isotlpes of that
species but only a proportion of the possible allotypes (Van Inghem, 1-984).
There are four IgG subclass isot¡res i¡ [ 'mans, lgGL,IñZ, IgG3 and IgGa distinguished by
different H chains desig¡ated'y 1,12,13,14 respectively. These subclasses ars detected by serological and
chemical methods as their charge spectra do not differ enough for detection by electrophoresis. The
subclasses differ in the number and arrangement of thei¡ interchain disulphide bridges (Goodman, 1982å).
Amino acid sequencing has revealed strikingly high levels of homology between the different
human IgG subclasses e.g. 95-98Vo in the C'y3 domaiq depen.ling on which subclasses are compared. The
greatest differences occur in the hinge regions between the Fab and the Fc portions. The major spec'fic
antigenic determinants a¡e in the Fc region in human IgGl and lgpz and in both the Fc and Fab regions in
IgG4. In human IgG3 the specific determinant ha,s been found only in the Fd (enz¡me-susceptible) rogion
of the molecule (Shakib and Stanwort\ 1980a).
1223 Altoty¡res of þG subclasses
Allotyes to three of these subclasses a¡e defined on the basis of unique H chain aminoacid
sequences which constitute inherited genetic markers ("G" markers) Glm, G2m and G3m. Some genetic
determinants occur as allotlpe ma¡kers in one subclass but as isoallotlpic non-markers in another (i.e. all
immunoglobulins of the latter subclass bear the determinant but only some in the subclass where it serves

32
as a marker (Shakib and Stanwort\ L980a). There are two allotlpic va¡iants of IgG4 (4a and 4b). IgGl
and IgG3 sha¡e thê 4a determinant, while IgG2 shares the 4b marker (Kunkel et al 1970).
Until recently research into human IgG allotpes has been seriously hampered by a lack of t'"ing
reagents (Pandey and Fudenberg L984). Studies have progressed steadily since monoclonal antibodies for
some of the determinants have become available. Haemagglutination has been the most widely used
method of allotype determination since the fi¡st allotlpe was discovered by Grubb and Laurell in 1956,
using this method. It is not, however, particularly sensitive, and is not useful for quantitative determinations
or for use in some immunodeficient sera. Radioimmunoassays have given increased sensitivity and havo
defined more allotlpes, but currently are useful for only a few allotlpic determinants. The use of
t".o6bi .rant
DNA technolog5r has revealed hidden heterogenicity in the IgG system, and with a single DNA
probe, up to 33 haplotlpes have been revealed.
122.4 Relative concentrations of þG subclasses in serum
The relative concentrations of the four IgG subclasses in human adult seru-t have been found to
be IgGL 60-70Vo,Iñ2I+30%, IgG3 Ç8% atdlgÙ42-6%. (Yount et al 1970; Morrell et aIt97?a; Shakib
et al LTl5; Oxelius I979a; French and Harrison I984a;Aucouturier et al 1985; Ferrante et al 1986a; Mayus
et al 1-986). These vary somewh¿t ¿sco¡ding to genetic bacþound age, sex and enviro''-ental exposure to
various antigens (Shackelford et al1985, Morell et alt974 Shakib et al L982). An individual's capacity to
produce antibody of one or other IgG subclass seems to be related to the allot¡pic markers present in that
individual (see section L.3.4.2). IgG subclass concentration would thus appear to be, at least io putt,
controlled by genetic factors, although the total semm IgG concentration seems to be independent of the
IgG subclass allotypes present.
After birt\ IgG subcla.ss concenhations fa[ reaching a nadir somewhere in the first 6 montls.
Children generally have lower immunoglobulin concentrations than adults and the adult concentrations of
the various immunoglobrrlin isotypes are reached at different ages. Each IgG subclass has a different rate
of progression towards adult concentrations (Normanse[ 1987). A study by Morell et al (L972b) indic¿tes
that IgG3 concentrations reach a troueü in the first month of life and after only 3 months approach adult
concentrations. IgGl spthesis sta¡ts before 3 montls of age, and concentrations are close to adult values
by 8 months of age. Tgp2 and IgG4 concent¡ations rise more slowly IgG2 not ¡s¿ching aù¡lt

33
concentrations utrtil about L0 yeuirs of age. Not all studies agree about the ages at which adult
concent¡ations of the IgG subclasses are reached but there is agreement about the sequence in which the
va¡ious subclasses reach adult concent¡ations. Oxelius (1979a) found that IgGl and IgG3 reach aboatÍ}Vo
adult concentrations by age 3 years, whereas lgG2 and IgG4 take much longer to do so. In a given subject,
subclass concentrations are thought to remain fairly stable in the absence of a sudden infection (Shakib et
aIL97Ð. However longitudinal studies in individuats are not available and littte is knovm about the effect
of acute infection on IgG subclass concentrations. Heiner has reported considerable fluctuations in IgG4
concent¡atio¡s in two of subjects, from 0.3 to 5 and from 5 to 20 mg/ml respectively. There is great need
for serial studies of IgG subclass concent¡ations in individuals.
1225 Properties of IgG subclasses
Physiochemical properties of tle IgG subclasses Íì¡e sumnarised in Table 1.1-. Biological
properties are summarised in Table 1.2. Some of the more important properties a¡e discussed below.
Complement activation by IgG subclasses
Antibody-mediated complement activation illustrates well the subtle relationship that exists
between imm¡¡sglsþulin structuL¡e and function. Both IgM and IgG activate the classical complement
pathway.
Between the Fab and the Fc regions of IgG is a proline-rich st¡etch of heavy chain joining the C"y1
to C12 snd co¡faining the interheavy chain disulphide bridges, the hinge region, where the a"gle between
the Fab arms in the antþen IgG complexes can be va¡iable (Burtoq 1985). The length and flexibility of the
hinge vary in the different IgG subclasses and in different species. This seems an important factor in
complement activation (Feinstein et al 1986).
The initial event in complement activation is the binding of C1q to sites on the Fc portion of IgG
(or to the (Fc)5 disc of IgM). Binding of CLq to free IgG molecules is weak and ineffective in activating
C1, but binding to aggregated IgG, usually in an antþen-antibody complex, is stronger and is able to
activate Cl.

34Table 1.1
PHYSIOCIIEMICAL PROPERTES OF IgG SUBCLASSES
Molecular weightl
Electrophoresis2
Isolectric point2
Tryptic peptidedifferences¿
Interheaw chaindisulphidá bonds2
IsGl
146,000
Cathodal
8.3-9.s
++++++++
slow
slow
++
rñ2
146,(m
Anodal
7 -7.3
fast
mediun
+
Igc3
170,000
Cathodal
8.45-8.95
5-1i
++++++++++
medium
slow
+++
IgG4
1¿16,000
Anodal
2
Fc& Fab
+++++++
74 18
Fd
4
Fc
2
Fc
Susceptibility to
+++
fast
fast
Cryoglobrrlin +
1.
2.
3.
Normanselt l!97Schur,1987Shakib and Stanworth L980(a)

35
Table 12
BIOLOGICAL PROPERIES OF IgG SUBCLASSES
Complement fixation2
Bndngstaph proteinA2 +
+
IgGl
+++
+++
tL/20/?330-¿10
IgGZ
+++
Íln/?330-¿lO
IgG3
+++
22-?A
TÑ4
+
+++
Í/2n/2330-40
0
0
+
+ 0
+
+++
+
+
+
+
+
+
+ +
++
+
+
+
React withrherlmatoid factor 3
rhet matoid facto?LDH3
++
Nonspecific red cellbindingl + +
B lymphocytessu¡face im mu¡sglsþul incytoplasmic
(davÐ(duvÐ
++
++
!2ßr¿;t¡zttrc4
7-8
t3.4.
Schur,1987Shakib and Stanwort\ L980 (a)Ochs et al" L989

36
Clq bincting sites are on tle C'y2 domains of IgG. The different IgG subclasses bind C with
different efEciencies. This va¡iation seems to be due to the steric a¡rangements of the molecules, rather
than to differences in binding-site affinity. It seems that interference by the Fab arms affects the approach
of C1q to the C'y2 sites and the degree of this interference is likely to be determined by properties of the
IgG hinge region (Feinstein, et al 1,986). Hinge-dependent Fab-Fab and Fab-Fc flexibility seem to be of
particular importance in ensuring that the Cl-q binding sites on th. Ct2 a¡e available for complement
fixation. There is a correlation between the length of the upper segment o¡ 1¡s hinge and the complement-
¿çtiy¿ring activity of different IgG subclasses. Human IgGl binds complement si¡ times nore effectively
than IgG2, and C1 is not bound at all by IgG4. The upper hi"ge is virtually absent in IgG2 and consists of
only 3 residues in IgG4. One would expect flexibility between the Fab and Fc segm.ents to be greatly
reduced.in both these subclasses. Human IgG3, which has the longest hinge, is the most effective
conplement activator (Feinstein et al198ó).
Placental oassage of the IgG subclasses
All four IgG subclasses cross the placenta (Morell et aIL97l; Morell et all972b; Mellbye & Natvþ
1973). Several studies suggest that IgGl is transported more effectively than IgG2 and have fsund hishs¡
levels of IgGl in cord blood than in maternal blood (Morrell et al 1972b; Oxelius t979a; Einhorn, et al
1987). Catty et al (1977) have shown that dystrophic neonates have lower concentrations of IgG than do
normal neonates. Human placental membrane components have IgG binding properties. IgGl and IgG3
are bound more strongly than IgG2 (Md.[abb et alL976). Binding maybe to the CII2 and CH3 domains of
the Fc region (Johnson & Mat¡e 1979; Johnson & Brown 1981). Placenta cells take up immunoglobulin by
phagocytosis. Receptors are found on the surface of the microvilli and pinocytic phagosomes, and they
protect the protein from enz¡roatic degradation. Bound IgG molecules p¿ü¡s thtough the cell and a¡e
released into the foetal circulation by exocytosis (Schlamowiø L976a; Schlamowitz t976b;Wild 1979).

37
Susceptibility to proteolytic dþestion va¡ies between the subclasses, e.g. susceptibility to papain
cleavage differs markedly between IgGL/IgP3 aú IgG2/IgGa. IgGl is the most resistant to papain
cleavage (Turner et al 1970). This is of importance when the preparation of imm¡asglsþrrlin for
int¡avenous use is considered.
Half-lives of IgG subclasses
IñI,Iñ2 and IgG4 have been found to have simila¡ half-lives. Early studies (Spiegelberg et al
1968; Morell et aI L970; Schur, 1-92) suggest shorter half-lives than those found in more recent work (Ochs
et a[ 1989).
Early studies used radiolabelled myeloma proteins and found these isotypes to have a half-life of
2'1. days and IgG3 of about 7 days. Later, Ochs et al (1989) using intrave¡s¡s immunoglobnlin preparations
in patients with hy¡logammaglobllinaemia who had reached a steady state of immunoglobulin peak and
trough concentrations from regular infusions, have found longer half-lives for IgG and for IgG subclasses
than those previously reported. Half-lives for IgG subclasses, with the exception of IgG3, were between 30
and ,10 days. IgG3 appeared to consist of two populations of molecules, one showing a rapid deca¡ and the
other a half-life of. ?2-?A days. Apparent differences between the results of this and earlier studies may
suggest that radiolabelled myeloma proteins are physiolog."lly abnormal or that the patients used were
'nonequilibrated' with regard to IgG subclass concenEations.
Cellular interactions of IgG subclasses
A number of cell types are able to bind monomeúc or aggregated IgG. The cell surface molecular
species which interact with the IgG are termed 'Fc receptors' but are not necessarily identical (Burtoq
1985). These receptors are essential for such functions as phagocytosis of opsonised particles, antibody-
dependent cellular cytotoxicity and the release sf inflamm¿fory mediators (Salmon et al, 1990). GenerallS
binding of IgGl and IgG3 is shonger than binding of IgG4 which is stronger than that of. IgpZ.
Phagocytosis, like complement activation, also shows some subclass specificity. IgGl and IgG3 bind most
strongly to the Fc receptors on monocytes and polS'morphonuclear leucocytes (Shakib & Stanworth 1980a;
Rozsnyay et al 1989).

38
IgGl and IgG3, bound by their ps frngment to monocytes and macrophages, promote cytotoxic
function (Douglas f.982). Larsson et al QnÐ have shown that only IñL,I$GZ.' and IgG3 are able to bind
to human llm.phocytes via the Fc region. Huqran IgG4 is thougbt to be the only subclass with the ability to
bind to skin mast cells of subhuman primates (Stanworth & Smith LnÐ.
Igp2 aú,IgG4 exhibit a nonspecific capacity to bind to erythrocytes (Gergely et alt967). All four
subclasses bind to platelets and initiate the release sf ss¡e[enin (Shakib and Stanworth 1980a) but IgGl and
IgG3 bind most shongly (Burtoq 1985). Many cell types induding monogtes, macrophages, granulocytes,
killer cells (K cells), B cells and some T cells have membrane receptors recognising the Fc portion of
human IgG. Three distinct types of receptors have been recopnised FcRI, FcRII and FcRItr (Anderson &
Loone¡ 1986). Monocytes e{press FcRI and FcRII, neutrophils express FcRtr and FcRm, B cells elçress
FcRtr and T cells, FcRm. FcRI binds IgGl and IgG3 more strongly than IgG4 and doesn't bind IgG2
demonstrably. FcRtr binds principally to IgGl and IgG3, and FcRItr solely to IgGl and IgG3 (Rozsnyay et
al 1989). Rozsnyay et al (1989) have shown that IgG3-sensitised erythrocytes are phagocytosed by
monocytes much more efficiently than IgGl-sensitised erythrocytes, but in inducing lymphocyte (K cell) -
mediated ADCC of erythrocytes, IgGl is more effective.
1..3 IMMT]NOGLOBTJLIN PRODUCTION
1.3.1 B Lymphocytes
Imnunoglobulins are produced by antþen-specific cells of B-cell lineage. The stem cells that give
rise to B colls are füst formed in the yolk sac of the embryo, then in the foetal liver and finally in the bone
maûow. Throughout life, cells from the bone marrow move to many other orgâns to complete their
maturation. Some of these cells undergo considerable maturation in the bone ma¡row before moving to
peripheral lymphoid oÌgail¡ to complete their maturation and become responsive to antþens.
Other llmphocytes, after beginning their maturation in the bone marrow, move on to the thymus
for further matu¡ation. Most of these cells die in the th)-us and only IVo oî. them leave it. These are
known as thlm.us-derived l¡mphocytes or T cells. T cells are involved in ¿ highly complex mechanism
regulating IgG production by B cells.

39
On elçosure to an antrgeq in the presenoe of regulatory T cells, B cells proliferate and
differentiate into antibody-secre:ng B cells and plasma cells. As the B cells mature, su¡face
imm¡¡sglsþulin receptors are acquired and displayed on thei¡ membranes. IgN4 is the earliest mature
form of imnunoglobulin receptor to appear. Later, both IgM and IgD are displayed. Iglvf is the füst
immunoglobrrlin secreted. As the immune resporuie proceeds, IgG and/or IgA or IgE begins to be secreted
and the cell then displays IgG or IgE or IgA plus IgD surface receptors. Antþen must bind to a su¡face
immr¡noglsþrrlin receptor in order to activate the B ce[ þ¡t this in itself may not be a sufficient sþal for
activation. T cells are necessary to provide a second sþal in the case of many antþens.
A given B tymphocyte cårr e4press only a single set of va¡iable-region gene segments. It can,
however, express potentially two lþht chain constant- region isotopes (rc andl) and 8 different heavy chain
constant-region isotopes @,6,a, e,1t,12,13,14)- In its ultimate plasma cell form it can produce only
one t¡re of heavy chain. The steps of immunoglobulin formation dudng B cell differentiation a¡e:-
1. The appearance in the cytoplasm of a heavy chain of thep type.
The interaction of this heavy chain with a light chain to form an IgM molecule, e4pressed fi¡st in
the cytoplasm and later on the surface of the cell.
3. A parallel interaction between heavy chains ¿¡d ligh¡ chains ¡ssulting in the synthesis of
cytoplasmic and membrane IgD. The IgM and IgD have the s.me variable region sequenees, as
will all imm¡aqglsþulins 5y¡fþes,izndby that cell or all its clonal progeny.
IgM and IgD disappear from the cell surfaces when the cell encounters antigen. The cell then
begins to synthesize and secrete IgM.
The cell undergoes division and amplification with ultimate differentiation to the plasma cell form.
This clone of cells may continue to produce IgM or may switch over to one of the other heavy
chain classes. The mechanism by which a singls variable region can oscur in association with one
or other of the several heavy chain isotypes is known as heavy chain switch (Leder 1983).
2.
4.
5.

40
1.32 Heary chain gene formation
Recombinant DNA technology has shown that V and C regions are separately encoded and that
both þht chain gene formation and heavy chain formation require somatic recombination events. The
different ty¡res of heavy chain constant region genes are encoded on a single stretch of DNA (Fig. 1.2). A
singls çqnstant region gene is capable of being joined to one or many variable region genes. C and V
segments are separated in embryonic cells but joined to form a continous polpucleotide during lymphocyte
differentiation. (Hozumi and Tonegawa L976;l-ai et al 1989). Uþhly complex structu¡al rearrangement of
heavy-chain genes proceeds immunsglsþrllin production. The heavy-chain va¡iable-region is constructed by
joining 3 distinct segments of DNd a V (variabþ segment, a D (diversity) segment and a J segment. The
V and f leenelts
are like islands of genetic information adrift in a sea of nudeotides sequences. Cloning
elçeriments suggest that there are probably 6 active copies of J-region sequences and a large number of D
and V segnents. The various possible combinations and cross-over point mutations, in heavy chain gene
formation together with thousands of possible lþht chain genes allow for the possible formation of millions
of different immunoglobulin molecules (Iæder, 1983) (Fig. 1.3). V-region rearrângement takes place at the
pre-B cell and produces the complete V-region genes for the heary and lþht chains which will characterise
an individual clone permanently. CH-region ¡sarrangononts enable mature B cells to secrete their V-
regions on different IgG classes (Taussþ L988).
þsf¿þlishing tle order and structu¡e of the heavy chain constant-region genes is heþing elucidate
the mechanism by which the different isotlpes of immuLnoglobulin are produced (Frg. 1.3). Each of the
heavy chains is associated with the same va¡iable heavy chain region for a given cell (Waldman 1987) i.e.
individual B lymphocytes are genetically pre-committed to producing antibodie,s to specific antþens.
The ¡r and 6 heavy chain constant segments lie in a 5' to 3' orientation about 2 kilobases fron each
other. T$o kilobases to the 5' side of thep segment is a strip of 6 active J segments. To the 3' side of the¡r
segment are the 6 segment and the 4y segments (Fig. 1.3).
The first step in e4pression of a hearry-chain gene is a recombination event associating a V segment
with a J and a D sement close to the p segment. Ap chain is produced by transcription of the code from
its 5' side with termination at the end of the ¡r chain sequence. The original 6 chain transcript contains

4T
51 -p -6 -13-1y!-ps.6 -a'1,-ps.^r -^yZ-14-, -a2-31
Fig. 12 Ig heavy chain gene sequenoe on chromosome 14

5'-----LVHtr---a-b-c-d-x- - -I-2-34-5 -6-Ctl
l-VHl Lo"J I ,"J
42
-Q r-Qr -9 z-) +-cu-Qz----¡r(Embryonic/germli¡e heavy chain gene)
(Rearrn"ged heavyLVH_b_2_3 454_Cu_C6
chain geue)
-c¡-9s-9i-Q.
DNA rea¡rangement
tranco'in¿¡o* RNA splicing
I
\.t-9¡-9 r-Ç¡. -9 zÇ+-c. -Çz
t¡'e n sqriPtioq RNA sPlicing4LYg-b-2-3+5{-q,(IgM mRNA)
2nd DNA rea(angement (isorype switch)\
I-Y g-b-2-345-6-,-- ---e :(Rearranged heavy :hain gen$-'
or
tra nsl¿tis* processing
v(IgM heavy
VHo = multiple variable regions, J¡¡- functional joining seque¡rces, L = leader sequence, DH = fam¡lies of diversicysegmeÂts
(Adapted from Korsmeyer & Waldmann lpg| and Waldman,, 19gZ)
Fig' 13 schematic diagram of the organisatiou and assembly of the human hearl chain gene

43
both ¡r and 6 chain codes. A splic'ng mechanism 'cuts out' the ¡r chain segment and joins the V-D-J
segment to the 6 chain seCment. Subsequentl¡ by the ill-defined mechanism of heavy chain switc.b, the
same VH gene is e:çressed in combination with a different CH gene (Waldman, 1-987).
The heavy chain switch frequently occurs during þmphocyte maturation from IgM to IgG, IgA or
IgE. The heary chain gene undergoes a recombination event in which the V-D-J sequence is switched to
another site along tle cb¡omosome. This seems to occu¡ at a number of sites within the intervening
sequenc€s of thep-,'f - or o- germ line sequenoe, usually dslsring any intervening DNA. The deletion can
occur by recombination between two sequences on the same strand of DNA (deleting what lies between
them) or between sequences on different st¡ands of DNA (resulting in unequal cross-over and an apparent
rather than a real deletion).
Switch regions lie2-3 kilobases 5' from each CH gene, witl the exception of the 6 gene. They are
large seÐents composed of multiple copies of short repeated elements. These homolog¡r regions may be
important as sþals for heavy-chain class switch. Such homology regions are not found between ¡.r and 6
genes and a differeut mechanism may be involved here. Studies with leukaemic lymphocytes have
suggested that a cell can be committed to B-cell differentiation but can fail to accomplish the V-D-J
recombination needed for functional p, chatn slmthesis (Iæder, 1983; waldman 1987).
L.3.3 The regulation of immunogloþulin production
A variety of sþals a¡e involved in B cell activation, proliferation, differentiation and antibody
production. The interaction of surface immunsglsþr¡lin with antþen is thought to be one of the initial
events in B cell activation. Depending on the nature of the antþen and the phenotlpe of the B cell"
additional sigals may be required for its subsequent differentiation into an antibody secreting cell.
(Jelinek & Lipsþ, L987 ).
B cell proliferation decreases as differentiation proceeds but ongoing immunoglobulin s¡mthesis
and secretion may require some continued proliferation of the differentiated cells (Jelinek & Lipsþ, 1983).
Continued proliferation of the immunoglobulin secrering cells may be important in promoting heavy chain
isotype switching (Cebra et a[ 1.98a).

44
133.1 Tcrlls
T and B cell interactions
T cells have a key role in regulating B cell frnction. Thei¡ interaction with B cells involves both
physical contact between the cells and the production of soluble factors (lymphokines or cytokines). The
relative importance of each remains to be defined. Shinomiya et al (1939) in an ø rzÞo study with human B
cells, found that direct contact with T cells was needed to promote sq,i¡ching from IgM to IgG production.
Neither T cell supernatant nor recombinant.interleukin-z (IL2) or recombinant interferoD-gâmma (IFNf )
would suffice. Once it was thought that only a limited number of T cell products were involvd and that T
cell influences were important mainly in the differentiation stage, subsequent to activation and proliferation
(Jelinek & tq.Ot, 1987). Now, evidence is suggesting that a variety of T cetl-derived lymphokines may be
important in every step of B cell responsiveness. The efent of their role remeins to be elucidated. A T cell
llmphokine may also play a part very early in tle sequence of events ls¿ding to antibody production by
preparing s¡¿ll ¡gsting B cells to respond before the engagement of su¡face immunoglobulin receptors
(Jelinek & Lipsþ, 198Ð.
Stimulation of B cells by certain T cell clones may result in preferential production of a particular
immunsgleþulin i5st'?e. T cells may act as "switch" cells causing the differentiation of membrane IgM-
bearing cells into cells bearing IgG isotypes (Kawanishi et a[ 1983a; Benson et 4 1990) or as ,post-switch,
ceils bringing about the expansion and secretion of Ig by cells already committed to a particular isotlpe
(Kawanishi et at 1983b). Conversel¡ an individual T cell clone may provide the necessary s,ignals fer
multiple isotype expression (Teale & Abraham 1987).
Most of the work on the influence of cytokines on IgG subclass production has been done in the
mu¡ine system. The T cell cytoking interleukin-4 (II-4, BCGF or BCGFf promotes B cell maturation
and growth and also the switch from IgM to IgGl production (Snapper et a! 19SS). It also enhances IgA
and IgE production (Teale & Abrahana, L987) and can transform membrane lgM-bearing colls to
membrane IgA-bearing cells (Cotrman et a[ 198ó). [¡fs¡lerrkin-J is considered the main oblþatory
differentiation factor for IgA production- It acts on B cells that have ulreudy undergone isotype-switch;r,g
to IgA-bearing cells, promoting IgA secretion (Harriman et a! 1988). All these lymphokines are
pleiotropic in their effects and net effects may be determined by their relative concentrations, and the

4s
sequence of their production and distribution (Spickett & FarranÇ 1989). fgC subdass oçression is doqm-
rogulated via T-supþressor cells. (I¡wy and Theze, 1985).
In the human system there is preliminary evidence that cytokines are important in isotlpe
switching. The füst report to show the involvement of interleukins in human IgG subclass regulation
(Lundgren et al 1989) showed that in a mouse thymoma co-culture method IL4, caused a dramatic
increase in numbers of human B cells showing intracytoplasmic IgG4 and of hunan B cells showing
intracytoplasmic rgE. Limiting dose analysis studies for IgE suggested that fhis was due to isotype
switching for IgE but similar studies were not done for IgG4. Benson et al (1990) have demonstrated that
IL5 and Branhanellø cøtanhalis together can promote a switching from lgM-positive B cells to IgA-
positive R cells.¡¿ vitro. 'Ihß appears to be the füst human llmphokine with direct switch activity to be
described. Benson et al (190) describe the known effects of cytokines on human B cells as follows:-
r-1f-2r-3
r-4II-5
r-6fï{"f
augments B cell proliferation and differentiation but without isotype specificitypromotes proliferation and differentiation of activated B cells.s¡mergistic with IL6 and II-2 in augmenting IgG secretion. Supports IgGsecretion by transformed B cells.may increase IgG, IgG4 and IgE secretion by activated B cells.increases IgM and IgA secretion, partly by promoting switching from IgM to IgAproduction.inc¡eases immunoglobulin secretion, apparently without isotlpe specificity.acts slmergistically with Í-2to augment inmunoglobulin secretion.
Most patients with common variable hlpogammaglobulinaemia (CVH) are able to spthesize IgM
and IgG on llmphocyte surfaces. The defect may be an inability to spthesize the soluble form of IgG (and
in some patients, of IgM also). Althougþ some T cell defects have been found" the failure of B cells from
patients to secrete IgG when co-cultued with normal dend¡itic cells pulsed with pokeweed mitogen in the
presence of normal T cells, suggests an intrinsic B cell defect (Spickett & Farrant, 1989). Failure to
respond to T-independent antþens also suggests an intrinsic B cell defect. Nevertheless, there is evidence
that failu¡e to produce or to respond to lþphokines may be a contributing factor in some patients with
immunoglobulin isotlpe deficiencies as the following fi¡¡tings illust¡ate:-
(Ð B cells from CVH patients with poor responsiveness to B cell growth factor (BCGF) andimpaired production of factors by T cells (Perri & Weisdo{ 198Ð.
(ü) a heterogenicity of responses to B cell differentiation factors (BCDFÐ in patients withcornmon variable immunedeficis¡cy suggesting that T cell defecls may be a primary pathogenetic6sçþanism in this condition. Exaggerated B cell respoû;es to 3 BCDFs were found in 3 patients(Mayer et al 1984).

46
(Ð defective B-cell response to I-Z (Ariga et al, L987)
(Ð a reduction lnÍI-z receptor e4pression or peripheral llm.phocytes (Malkovsþ et a\ 1986)
(v) a patient with com-on variable immunodeficiency whose T lymphocytes released BCDFand BCGF but did not produce II-2 on stimulation with ConA (Saiki s1 al" 1984).
(Ð an infant with h¡pogem-aglob'rlinaemia whose peripheral blood mononuclear cellssecreted Í-2 andBCGF, but failed to secrete BCDF (Matheson & Green, 1987).
(vü) lymphocytes from patients with hypogammaglobulinaemia and high IgM, and from thosswith X-linked a-gammaglobnlinaemia producing some IgG when stimulated with a factor from Tcell supernatant., The factor is tot II-2, IL4, ILs or II-6 and its identity is, as yet, unknown(Benson et al, 1990).
(vüi) a doficiency of T helper cells in ataxia telangiectasia (Waldman et al 1983), a condition inwhich deficiencies of Ig,\ IgG2 and IgG4 (Oxelius et al 1982, see section 1.8) have been described.
(i*) a patient with ataxia ¡etangiectasia who lacked IL2 production and in whom sorum IgM,IgG and IgA were low. Treatment with recombinant IL2 resulted in a selective increase in serumIgM and a marked improvement inin vítro IgM production (Doi et at 1989).
G) e4perinents with recombinant interlerkins have indicated a block in the differentiation toantibody secreti'g cells in CVH which cannot be overcome with the differentiation factor lL6.(Spickett & Farrant, 1989). A defective receptor or receptor coupling for lymphokines has beenpostulated.
(ri) defective production of both BCGF and BCDF in hlpogam-aglobulinaemia (Raziuddin& TeHu" 1989).
(*ü) a patient with severe combined immunodeficiency due to a specific defect in theproduction of.II-2 (Weinberg & Parkman, 1990).
T-independent antigens and T-dependent antigens
B cell resporu;es have been categorised by their apparent need for T cell influence as T cell-
independent (Ð a"d T cell-dependent (TD) responses. Most TI antþens are linear with multiple
repeating determinants and a¡e poorly metabolised high þf.
47
required T cells or T-cell derived factors for the generation of Ig-secreting cells in man, although some
could induce B cell proliferation in a T cell-independent mânner.
TD antigens require T cell help to induce B cell differentiation into antibody producing cells. TD
responsos include both antþeuic and mitogenic responses. They may be fr¡rther subdivided u""s¡ding to
whether they require physical interaciton of B and T cells or whetler only the presence of T cell-derived
cytokines is necessary. The state of activation of the B cell subpopulation may, in part, determine the
nature of the T cell interaction required e.g. "resting" B cells may require physical interaction with T cells,
whereas "activated" B cells may be more dependent on cytokines. However, as noted eailier, Jetinek &
Lipslry's studies (198Ð suggest that cytokines affect þefþ ¡s5ting and activated B cells.
T cells and histocompatibility antigens
When T cells recognise ant'rgeq usually on the surface of antþen-presenting cells they proliferate
and begin to differentiate (Katz and Mead 1983). T cells recosnise antþen in association with the major
histocompatibility structures (HI-A-4 HI-A-8, Iil-A-D) on the surfaces of antþen-presenting cells such as
dendritic cells or macrophages (Metlay et al, 1989). T helper cells (CD4 positive lym.phocytes) which have
an important role in B cell regulation, recognise foreþ antþens in association with HI-A-D structures
(Class II antigens).
Suppressor T cells (CD8 positive þmphocytes) may hinder helper T cell interactions with B cells,
prevent B cell differentiation or dit"ctly inhibit the secretory function of fully mature plasma cells (Kaø and
Mead" 1983).
1332 Macrophages
Macrophages play a crucial role in the immune respoil;e to antþens. They process and present
antigens to T cells and induce T helper cell function. They are not thought to be specific for any given
antþen. T helper cells have specific receptors to recogni^ss carrier determinants and they also interact in a
criticål way with macrophage-associated MHC-encoded class II cell-interaction molecules. Activated T
cells then interact with B cells specific for haptenic determinants linked to the carriers. These B cells
mature into antibody-secretory pla^sma cells (Katz and Mead 1983).
The initiation of T cell activation requires both the pollpeptide qrtokine, interleukin-l (U-1),
which is produced by macroph4ges, and the triggering of T cell receptors by antþen and MHC gene

48
products. This leads to a decrease in su¡face antþen recoptors, and an increase in interleukin-z Qf-z)
receptors on the T cells. The T cells then produce and secrete IL2 which binds to thetr IL2 receptors and
promotes DNA s5mthesis and cell division (Royer and Reinherz, 1987).
1333 Natural killer (NK) cells
Natural killer cells (NK c,ells) also appear to have a role in regulating antibody spthesis. They
were first shown in vitro to suppress antibody production under certain conditions (Tilden et al 1983;
Brieva et al 198Q. In vivo studies in mice by Khater et al (L98ó) indicate that NK cells may play a major
role in down-regulating the response to pne'mococcal polysaccharides in both infants and adults. More
recently they have been shown to enhance IgG, IgA and IgM synthesis by corresponding l¡mphoblastoid B
cell lines, apparentlyby the release of soluble factors which may demonstrate isotope specificity (Kimata et
41e8Ð.
L.3.4 Immunogenetics
13.4.1 Genes that influence the immune nesponse
The production of a particular immunsgl¡þrrlin deps¡ds on both genetic and environmental
factors. Evidence indicates that genes may influence the immune response via four main mechanisms
(rù/hittingham et at 1984):-
1. Genes in the major histocompatibility complex (l!ftIc) which code for HI-A-DR and related
antþens (class II genes). These are neoessary for the cell-cell interaction that occurs when antþens are
presented to T cells by macrophages and equivalent antigen-presenting cells. Without these antþens the
imms¡s response does not occu¡. Surface st¡uctu¡es mded for by class II geues occur on B l¡mphocytes,
some monocytes/macrophages, some T cells, th¡mic epithelial cells, Langerhans cells and dendritic cells. It
appears that any cell which can e4press sufEcient MHC molecules on its surface may function as an
antþen-press¡ring cell for some antþens (Delovitch et a[ 1988).
2. Genes in the MHC which code for HLA-A, HLA-B and HLA-C antþens (class I genes). These
are necessary for cell-cell interactions where antþens (usually viral) are presented at the surfaco of cells

49
whic.h are targets for cytotoxic T cell attack. In the absence of these antþens, cytolysis will not occur, or
will be very weak. They occur on virtually every cell.
3. Genes in the MHC that code for components of the complement system.
4. Genes which speciS the st¡ucture of imm¡nsglobulins. The MHC genes are on ch¡omosome 6 in
man, while the immunoglobulin st¡uctural genes are found on th¡ee other ch¡omosomes. Genes for the H
chain are on the long arm chromosome 14 at band q32 (Croce et a15 t979; Ki¡sch et a! 1982) (see section
1.3.2). Those for therc andÀ lþht chains are on chromosomes 2 and 22 respectively. The humanrc gene
locus is located on ch¡omosome2atband2plj. The majority of humanl genes are arranged in tandem on
the longarms of ch¡omosome 22 at band ?2qLl (McBride et al, L982). Family studies have shown that loci
for heavy chain genes are closely linked. Heavy chain allotlpes Gm, Am, and Em a¡e inherited in füed
combinations referred to as haplotypes. Light chain genes and the cluster of heavy chain genes a¡e not
linked (Van Loghem, 1984). There are striking differences in the frequency and occur¡ence of various
haplotlpes in various races (Steinberg & Cooþ 198L). Certain haplotlpes a¡e cha¡acteristic for particular
races, and there is evidence for genetic control of immunoglobrllin s)mthesis associated with certain Gm
allotypes.
13.42 Genes and immunorglobulin isotype concentrations
Isolated IgA deficiency is not uncommon occurring in 1- in 300 to 1 in 3000 people (section I.2.L.2).
The defect is generally at the regulatory rather than at the structural gene level (Van Loghem, 1984).
There appears to be some association with HLA-BS/DR3 in as¡rmptomatic, not in sym.ptomatic, IgA
deficient subjects (Svejgaard" 1983).
Most IgG subdass deficiencies are relative rather than absolute and are likely to be the result of
defects in immunoregulatory mechani$ns. Cases of isolated absolute subclass deficiency are often due to
structural gene abnormalities. The absence of all allotpes of a particular subclass can indicate the deletion
of the gene for that subclass. In Ti¡nisian and I-ebanese-Irani¿¡ fanilies, segregation of Gm haplotlpes
lacking inT1, eL,12, "y4 and'¡,3 genes respectively, füst shown by alotyping and then proved by Dl.[A
analysis has been demonstrated (Lefranc, LW9, 19834 1983b).

50
Associations between IgG subclass concentrations and Gm phenotypes have been described eg.
G3m(b+) individuals þ¿vs highs¡ concent¡ations of IgG3 than do G3m(b-) individuals (Yount et aJ, L967),
and homozygotes for this marker have higher IgG3 concentratious than do heterozygotes (Shakib et al,
L982). G2m(n+) individuals tend to have higùer concentrations of IgG2 and IgG4 than do those without
this marker (Morell et al, 1972a). IgGl and IgE concentrations may be similarly affected and a gene
dosage effect has been demonstrated (Oxelius, 1Ð0). G2n(f) individuals also have slightly highs¡ lgÇl
concentrations (Lifwin &. Balaban, 1972).
The immunoglobulin response to somo antigens is associated with certain allotypes. Infectious
diseases may be among the main contributors to allotlpe selection. The facts that there is no random
selection of allotypes and that the distribution of haplotypes in ethnic g¡oups throughout the world differs
markedl¡ indicate probable selective advantage for existing haplot¡rpes. They may also reflect resistance or
susceptibility to particular diseases (Van Loghem, 1984).
13.43 Allotypes and disease
la 1977, Fa¡id et al reported that immunoglobulin allotypes may serve as markers for a disease, or
that they, like the HLA antþens, may be linked or associated with a disease. Many human diseases were
studied and fou¡ patterns of genetic involvement emerged:-
a) MHC genes only
b) Immr¡nsgl.6ulin genes only
Ð Interaction of immuneglsþrrlin and MHC genes
d) No relationship with either.
Much work has been done on the association between HI-A types and various diseases. The
relationship between immunsgleþulin ¿llqq¡pss and the level of antibody production (particularly antibody
of particular isotpes) and diseases has been less extensively studied. It is likely that the association
between antibody specificity and allotype, reported in some infectious and other diseases reflects tle
Presence of an allotlpe-linked variable region, or a linked regulator gene for variable region e:çression e.g.
The antibody response to H. influenzne type b (Hib) i¡fections is associated with G2m(n). Increased
antibody response ¡q immunis¿tion with Hib occurs in adults with G2m(n) allotype (Ambrosino et al,
Le82).

51
Since, in man, the genes for imnunoglobulin chains and HI-A antþens are on different
chromosomes, link¿ge between them is excluded. Flowever, interaction between independently segregaring
gene pairs of a phenotype is well-known, and in mice interaction of the MHC and heavy sþain genes has
been established. Genes for HI-A tlpes and Gm alleles may complement each otler so that susceptibility
to a particular disease is higher when both are present together that when either is present alone (Van
Loghem,1984).
The great polymorphism seen in MHC and imnunoglsþ'lin genes is e¡pressed in different racial
goups to varying degrees and probably represents a resporse to local envi¡onmental antigenic ehallsage.
As yet, T" T""oglobulin
region genes that code for antibody responses to particular antþenic epitopes
have not been identified in man. V region genes contributing to antibody diversity are now known to be in
linkage disequilibrium with structu¡al genes for immunoglobulin allotypes of the Gm and Km systems
(Whittingham et a[ 08a).
Very few resea¡chers have examined the association of Gm phenotlpes with infections. Nevo
(L974) demonst¡ated that subjects homozygous for Gm (3,5) responded poorly to Salmonellø typhi anttgens
compared with heterozygotes and that in subjects with t¡rphoid the relative frequency of homorygosity for
Gm (3,5) was increased, suggesring a possible ¡elationship between Gm phenotype and susceptibility to
typhoid. The complex connections between allot¡res, patterns of IgG subclass responsiveness to various
antþens and susceptibility to specific infections and other diseases ¡speins to be elucidated.
fþe imm¡¡¡globulin response to flagellin has been shown to be influenced by a number of factors
¡osluding botb Gm phenotlpe, and HI-A phenotype (Wells et al, t97t, Whittingham et al 1980). Linkage
disequilibrium between the C and V region immunoglobrliir genes is considered to be the basis of the
association with Gm phenotype. The mean effects of particular HLA and Gm antþens on the immune
resporu¡e a¡e substantially similar, but there ¡'s important interactive effects between the loci so that the
joint effects of particular HLA and Gm phenotlpes a¡e substantially greater (or less) than the sum of the
mean effects of the two loci considered separately.

52
1.4. EVALUATION OF IgG SUBCI"ASS CONCENTRATIONS
1,4.1 Quantitation of IgG subclasses
Although IgG subclasses were füst described over ?n yearc ago, progress in understanding their
sigrificance in health and disease has been hampered until recently, by difficulties in IgG subclass
quantitation due to high dsgrges of sequence homology between the heavy chain of the four IgG subclass
isotlpes. Initial studies used poþclonal antisera to each of the IgG4 subclasses for detection and
measurement. Most used radial immunodiffusiou or electroimmunoassay. There was no internationally
accepted IgG subclass standard senrm. IgG subclass concentrations on patient samples quantitated in one
laboratory were assessed accor<ling to normal rânges established in another laboratory regardless of the
fact that'they had often been established using different antisera, different techniques, different standa¡ds
and in subjects of very different genetic bacþrounds. Most assays were not sensitive enough to detect IgG4
at all in a considerable percentage ofnormal healthy subjects.
Understandably, these factors have resulted in considerable confusion in the IgG subclass a¡ea.
The problem is illustrated in Tablel3 by a comparison of normal ranges obtained in different populations
of adults, in different laboratories and by different methods. The mean IgGL,lgG} IgG3 and IgG4
concentrations in these studies vary from 5.9t-9.4,2.L7-3.æ,0.41-1.0, and 0.08-0.75 g/l respectively. The
ranges for each isotlpe also show ma¡ked differences. The fact that these differences occu¡ indicates the
need for some world-wide standa¡disation of IgG subclass quantitation. Until this fo¿s been accomplishe{
abnormalities of IgG subclass concentration would be best defined by usiog normal rânges established in
the seme laboratory with the same methods and reagents for comparison. Unfortunatelf this is not often
practicable as developing normal ranges, particularly in the paediatric population is a major undertnking
and most laboratories have attempted it.
1,42 Normal senrm concentrations of IgG subclasses in children
The measurement of serum IgG subclass concentrations is being ¡6ed i¡ç¡e¡singly to help to assess
the im¡nunological competence of infection-prone subjects. Recently, with the developnent of both
suitably standardised IgG subclass monoclonal antibodies and en4roe-linked immunosorbent assays whic.h

TABI,E 1.3
Reference
SERI]IIÍ IgG SUBCI,ASS CþNCnITRATIONS IN NORIfAL N)IILTS
Method Àntibodies Numberof sera
IgGL TgG2 rgc3 IgG4
Morell et al,I972a
RIA polyclonal L08
Shakib etr975
al, RID polyclonal 111
Oxelius,L979a EIA polyclonal 20 7 .55+2.45 3 .80+L.5( 4 .22-12 . e2) (L .7 -7 . 47 )
6 .L3+2.89 3. O0+1. 49(2.43-L3.2e) (L.2L-5.71)
5 .9I+2 .64 3 .O1+2.52(2.86-L3.46) (L.OO-7.25)
YounL970
Àucouturieret al, 1985
Ferranteet al, L986a
Shackelfordet aI, 1986
Mayuset aI, L986
â1, RID 1.0+0. 45
0.58+0. 30(0.10-2.11)
o.94(o.r_5-2.45)
o.73+O.26( 0.41-r-.2e )
o.74+O.7L(o . t6-2 .67 )
0.61+0.56(0.0e-3.r-6)
o.4L4+O.L7(o.r-2-1.14)
o.77+O.35(0.30-1.57)
+0.48
0.46(0.03-2.e0)
0. 08(0.0r-5-o.r-85)
0.55+0.63( <0. or--2 . 91)
0 .44+O .39( o. o3-r.. 38 )
0.24+0.90( <0.03-5 .39 )
0.385+0.392( <o . 0r--2 .10 )
0. 75+0 . 49(0.18-2.0r.)
Kaschka et aI RIDL982
French & RIDHarrison, L984a
Ional
polyclonal 23
McÀb ]-72
+l_.9 3.2+L.3
6.63+r.70 3.22+L.O8(3.s-11.5) .(r..3-6.8)
8.01(3 .7 -12 .3)
6 . 35+1. 50(3.0-r-0.s)
2.r7(0.46-4.3)
2.6L+t_.36(0.4-6.3)ELISA McÀb L22-L29
ELTSA MCAb 4T
RIA McAb (IgGl) 41'polYclonal (IgG2)
PCFIA MCAb ].86( rgcl. ,TgG2 ,TgG3)RIA (IgGA) 32
7 .4r+2.41, 2.55+O .94(3 .82-L4 .27 ) (L.24-4 .e6)
6.56 2.55 ND ND(3 .44-r2.07 ) ( 0.88-7 .40 )
6.03 2.49 0.52 0.34
(2.87-r2.65) (o.e2-6.72) (O.]-7-1,.62) (0.028-4.06)
innunodiffusion,' PCFIÀ Part icle concentrate fluorescence imnunoassayi EIA : electroinrnunoassay.l,leans t and rangei RIA say; ELISA = enzYme-l assayi RID = a
(¡l(,

s4
are sensitive enough to measu¡e concentrations of each of the IgG subclasses, even at the low IgG3 and
IgG4 concentratioris that are found in the sera of young children, the scope of IgG subclass as a diaenostic
tool has increased.
flowever, there are many difEculties in interpreting results, paficularly in children, as the few
paediatric normal ranges that have been published to date do not agree about the cutoff values between
normal and low concent¡ations. Using conventional statistical methods eg. mean t 2SD, even with log
transformation to normalise the data, it has not been possible to construct smooth age-related percentile
curves, partly because inadequato numbers of children in each age g¡oup have resulted in wide fluctuations
from one age group to the netr. There are considerable differences in both the mean" median or fiftieth
percentiles and in the lower limits of normal in a number of published paediatric studies. The reasons fo¡
these marked differences will be discussed in Chapter 4. It has been ass'-ed that male and female
children do not ffie¡ significantly in their IgG subclass concentrations at identical ages and comparative
normal langes for male and female children have not been published. One study (Bird et al 1985) found
no significant differences between males and females in the one to five yeal age gfoup.
Because the füst published paediatric normal rânges were determined by radial immunodiffusion,
(RID), radioimmunoasay (RIA) and electroimmunoassay (EIA) with poþclonal antibodies (Morell et al
LflZb; Van der Giesson et al t975; Oxelius t979a; Schur et al 1979; Shackelford et al 1985) and now
enz5me-linked immunosorbent assays (ELISA) and/or monoclonal antibodies are being used by an
incroasing number of investþators, different normal rânges may be necessary.
1..5 IgG SUBCLASS DEFICIENCY IN IgA-DEFICIEI\T PATIENTS
The possibility of an inportant association between IgA deficiency, IgG subclass deficiency and
infection-proneness was füst raised by Oxelius et al (1981) in a now widety quoted paper reportiog the
fioding of ILgp2 deficiency in a number of infection-prone IgA deficient patients. This suggested that IgG
subclass deficiency may be an important factor in determining why some lgA-deficient patients are
infection-prone and others are not.
After the publication of this paper, other investþators began to study IgG subclass levels in IgA-
deficient patients. In 1983, Ugazio et al reported a very high i¡side¡ce of IgG subclass deficiency in IgA-

55deficient children with recurrent respiratory infections. Subsequently, however, in a larger stud¡ the same
group (Plebani et a[ 1986) did not find ¿ signifisånf incidence of IgG subclass deficiency in IgA-deficient
patients.
Bjorkander et al (1985) found that concentrations of IgG2 or IgG3 more than 1SD below the mean
were related to impaired lug function in IgA-deficient patients, su-gesting that relatively low levels of IgG
subclasses may be contributing to their lung disease preslmably by increasing infection-proneness, although
the study did not record whether there was a difference in infection-proneness between IgA-deficient
patients with and without IgG subclass deficiencies.
IVfore recently, Klemola et al (1,988) studied a group of 60 IgA-deficient patients with a variety of
medical problems and in the lþht of previous worþ found a surprisingly low incidence of IgG subclass
deficiencies. There was, however, a significant association between IgG2 deficiency and infections. The
incidence of IgG4 deficiencies in this study did not differ significantly from that found in the healthy
Swedish controls used to establish the normal ¡angos (Oxelius, 1979a).
Aucouturier et al (L989) found a ?-ïVo ncidence of IgG subclass deficiencies in IgA-deficient
subjects which was not significantly correlated with the infection-proneness. IgG2 deficiency was the most
common vttth 63Vo of the IgG2-deficient patients also being lgG,l-deficient. However, only undetectable
IgG4levels observed together with definitely low IgG2 levels were considered as IgG4 deficiencies in this
study.
Heiner (1986) previously had reported similar incidences of. Igp2 and IgG4 deficiencies i" IgA-
deficient subjects and did find an associationbetween severe deficiencies ofthose subclasses and infection-
proneness.
It is apparent that there is considerable discrepancy between the findings of these studies.
Reported incidences of IgG subclass deficiency in IgA-deficient patients ¡a"ge from 7 to 70Vo (Klemola et
al 1988; Heiner, 1986). Some (Oxelius, 1981; Ugazio et al, 1983; Heiner, 198ó) suggest that there may be a
high incidence of IgG subclass deficiencies in IgA deficient infection-prone patients. Others (Bjorkander et
al 1985; Klemola et al 1988) found a lower incidence of IgG subclass deficiencies in such patients, but a

56
sigrificant association between IgG subclass deficiency and either impaired lung function or infections. Still
others (Plebani et af 1986; Aucoutu¡ier et al 1-989) have not found a clear relationship between IgG
subclass deficiency and recurrent infections in lgA-deficient subjects. The association between the severity
of an IgA deficiency and the occurrence of an IgG subclass deficiency has not yet been clarified.
Comparisons between studies are very difficult to make because in different studies there a¡e
di.fferences in:
(") definitionsofrecurrentinfections/infectionproneness
(b) definitions oflgA deficiency
(c) definitions oflgG subclass deficiency
(:) subjects used to establish normal rrìnges
(") ages of the patients
For those reasons, it is impossible to determine from the currently available literature the t¡ue
incidence or the clinical significarce of IgG subclass deficiency in lgA-deficient subjects. Nevertheless,
since, tle fiodiog of an IgG subclass deficiency in an IgA-deficient patient with recurrent infections is
becoming accepted more and more as an indication for intravenous immunoglsþulin replacement therapy
urgent clarification of the true situation is needed.
We need to know whether IgA deficiency really is associated with an increased incidence of IgG
subclass deficiency. To clari$ this, studies need to be done with the same laboratory techniques, the same
definitions of deficiency and preferably patients' IgG subclass concentrations compared with those of
normal subjects assessed under identical conditions. We need definite criteria for defining abnormal
proneness to infections. Pa¡ticularly in children who are normally prone to considerable numbers of
respiratory infections each year, such criteria are essential if studies relatirg infection proneness to IgG
subclass deficiencies are to þs nsaningfirl. When these conditions are met, we will be in a situation to
rearisess the incidence and significance of IgG subclass deficiency in IgA-deficient subjects Chapter 5 of this
study addresses these topics.

57
L6 IgGSUBCLASSESAI{D'SPECIFIC'AI\¡'IIBODIES
1.6.L IgG subclass distribution of antibodies
Different types of antigens often give rise to IgG antibodies of different subclasses (Yount et a!
1-968) so that specific antibodies to certain antþens may be of predominantly one or two subclasses. This is
known as isotype restriction. Although the subclass that predominates is not always the sn-e in all persons
under all ci¡cumstances, some generalis¿¡isns can be made.
1.6.1.1 Protein antþens (Table L.4)
Anti-protein antibodies are mainly of the IgGl isotlpe. These include antibodies to a wide variety
of viral bacterial and autologous antþens, (Hamm¿¡s¡om 1984b). ,Tetanus toxoid antibodies have been
found in atl IgG subclasses (Parisb 1975; Dengrove et al, 1986) but most agree that IgGl constitutes the
bulk of the post-immunization response (Yount et al 1968; Parish, 1975; Van der Giessen & Groeneboer-
Kempers, 1976; Seppala et al 1984; French & Harrison, 1985; Dengrove et al, 1986; Devey et at 1982).
Shakib and Stanworth (1980b) however, found tetanus antibody response in only the IgG3 and IgG4
subclasses after a booster injection of tetanus toxoid in 6 subjects. Schanfield et al (1979) found
responsiveness to tetanus toxoid in the population of New Guinea was related to with tle presence of
c1-(Ð alloype.
Work of flamm¿¡5f¡sm et al (1984c) suggests that antibodies to the alpha toxin of. Støphylococcus
(ntreus are restricted to IgG1, IgA1, and IgG4 and that these appear sequentially. In toxic shock slmdrome
caused by S. øtreus toxin 1, concentrations of IgG2 that are lower than in controls (Christensson et al,
1986) wbile patients show a lack of specific IgGl and IgG4 anti-toxin antibodies.
Wedege and Michaelson (1987) found that antibodies to the m.ajor outer membrane protein
antþens of meningococci were mainly of the IgGl and IgG3 isotypes. Complement dependent bactericidal
activity is considered to be the main line of immune defence agains¡ meningococcal infection (Taylor, 1983)
and these subclasses a¡e those most efficient in complement activation (section L.2.2.Ð.

TMI¿ I.'
^^t19.n
Tetanu6 toxolal
PRSÐKII{AFI rsqrÍpEs op t9ç emræoræ m pRø¿t Mrcãs 5BStqdy/Rê (er€nce
Yout er â1, t96aPàrish, l9?5Shôkib a Sfôneor!h. tgBObSepp¿ [¿ eE À1. I98aFrench ú Hârrison. l9g5Dengrove €c ¿ t. I9g6Dcvey ec à1, t98?
8¡M¿rstro¡ €r ó l, lggac
- post booster
- Éþs t boos c€ r
co@nÈ5
Àlphô toxiô ofS. aur€us
Oulcr ¡e¡braneproteln of
.cn Ingææcug
Ru-bell¡ vled
Kurp6 vfru6
Hed.g€ ¿ t{ichÄ€lsen
H.deq€ ¡ Xichôclsen
SkvÀrll, t98lSkv¿rll f Schitt. t9s.Sârnesto Gt â1, I98j
Llnde, t965LInd€, I9t5
6 veeks poscl@ql¿âtion25 ve¿¡s p¡5qt@qlzàtlon
- Ist dây otI I Lnê.s
- hcâlrty- rêcent
lnfect lon
- clinlcôliôfecÈ lon
- heÀlt¡y donors
- lndirectevldênce only
- h.À I tly- prlEry a8v
ln fect ion- reÀctlvatect EBv
infec(ion
- PostvÀccinòcion
heôIt¡y donorsO{V in(ectionre¡ctivôled cRinfec! i on
- hcÀlr¡y- prlury
Inf6ctlon- extanaled
i nfect lo n
hcôlrìyvÀrlcG I l.¿o5têr
t987
l98t
[9A2
1986¡.986
1986
Sôrnêsto êt ô1, t9€5bLind..t ô1, t9g?b
Eb¡têln krrvc^
Lind€ et.I. f9o?b
l(ô¡chà .t ¿[
Linde €t ôt
9à9ner ef ôì. 1986Hàtt er ô1. t9g6
Llnde ec ÀtLinde ct àI
LInclc er ¿l
Torfôson et .l, I9g?
Hepàtitis Isurfàc€ Àn!¡,gen
Coxsacki€ I
R.splrÀcorysyncytl! I virus
CytoEêgâ lov i ru3
XcrfÞ¡ 5l¡Þl6xvlrua
vôrlcCI l!-ao¡Èê rvlruE
Kðttil. €r.1, l9a6xorell. l9a)Skvaril a JoIler-Jeoetk¿SkvÀrt¡ r Joller-Jeoelk¿
t98a
t90a
t98l
Ssdqvi.r €r â1. t98.
Sqdqvl.q er ¡I- l9a.
tnf lu.nsrJulkqên .t öI. l9A5
+'+ ¡ þ¡t pro¡rnentt ++ ' rñÈ'rredlaca prorrnancêt + r preaênt bqt noc pro¡rncnt

59
Antibodies produced in viral infections are usually of the IgGl and IgG3 subclasses. Sarnesto et al
(1985a) studied the isotypes of antibodies produced in 20 patients with rubella" finding that during the füst
few days of the illness, IgM predomi"ated but by 10 days, IgGl- did so. Other studies also indicate that
IgGl is the major subclass produced in rubella (Skvar{ 1983; Skvaril & Schilt, 1984; Linde, 1985).
The isotype resporu¡e to mumps virus in a group of either vaccinated or infected patients has been
studied by Sarnesto et al (1985b) and found to be predominantly IgG1, both in acute and in convalescent
phase sera Linde et al (198Ð confirmed this fuding and also demonstrated a rise in IgG3 antibodies against
mumps nucleoprotein in the acute phase.
"^"0: et al (1-982) measured immunoglobulin isoty?e concent¡ations in patients with two Epstein-
Barr virus (EBV)-associated diseases, infectious mononucleosis and nasopharymgeal carcinoma. In both
groups of patients, concentrations of IgGl were significantly elevated" while l:ñz,lg}3 and IgG4 were
within the normal ratrge. Patients with infectious mononucleosis also had elevated IgM concentrations.
This study suggest that IgGl- antibodies may play an important role in the humoral immune response to
EBV determined antþens and perhaps in the control of ce¡tain infections with this virus.
Skvaril and Joller Jemelka (1984) and Mattila et al (1980), found antibodies to hepatitis B surface
antþen to be predomin¿¡¡ly IgGl following vaccination . After acute hepatitis B infection also, IgGl
antibodies predominate but with significant amounts of IgG3 (Morell 1983), or of IgG2,IgG3 and IgG4
(Skvaril & Joller Jemelka 1984). The reasorxi for the variations between these studies are unclear, although
affinity differences of the subclass reagents, the nature and amount of antþen, the genot¡re of the subjects,
and the fiming of the blood sampling may be factors.
The antibody response is predominantly of the IgGl and IgG3 isotlpes in infections with Coxsackie
B virus (Torfasson et al 1987) respiratory syncitial virus (glycoproteins F and C and fusion protein)
(Wagner et al, 1986; Watt et 4 1986) cytomegalovirus (Linde et a[ L933). Herpes simplex virus and
varicella-zoster virus may give rise to IgG4 antibodies in addition to IgGl and IgG3 (Sundqvist et al 198a).
The relative proportions of IgG subclasses in the antibody response to certain viral infections vary
¿sco¡ding to whether the infection is primary, reactivated or latent (Coleman et al" 1985; r inde et a[ 1983;
Sundqvist et al 198a).

60
Some anti-protein antibodies are predominantly IgG3. These include anti-platelet antibodies in
idiopathic thrombocytopenic purpura, some antinuclear antibodies, antfüorse immunoglobulin in serum
sickness and some antivi¡al antibodies (Shakib & Stanwort\ 1980b; Becþ 1981a).
Most studies agree that IgGL and IgG3 are tle most predominant IgG antibodies produced against
protein antigens. IgG3 may be present ¿t highe¡ concentrations than IgGl early in the infection but
generally IgGl predominates. IgG4 may become apparent, particularly after repeated antigenic sti-ulation
and always in conjunction with highe¡ concent¡ations of IgGL (Ferrante et al 1990).
1.6.L2 Carbohydrate antþens (Table L.5)
Antipolysaccharide IgG antibodies, since the pioneering work of Yount et al (1968), have generally
been considered to be predominantly of the IgG2 subclass. However, more recent studies are
demonstrating that often this is not the case in you"g child¡sq and that in adults, also, ttre pattern may vary
from individual to individual and from one polysaccharide antþen to another (Sarvis et al 1939).
Yount et al (1-968) showed that anti-dext¡an antibodies were predominantly IgGz (n 7 of 8
patients), antiteichoic acid antibodiss wers predominantly IgG2 (in 2 of. 3 patients), and antilevan
antibodies were predominantly IgG2 (in the one patient studied). liyi-anne Oxelius (L974), supported this
worþ indirectly, by findi"g that antibodies to both teichoic acid and Hib capsular polysaccharide were
absent in three IgG2-deficient subjects. A couple of years later, Reisen et al (1976) found that antibodies to
group A streptococcal polysaccharide antþen were mainly IgG2. Siber et al (1930) found a significant
correlation between IgG2 concenEations and responsiveness to both pnermococcal antigens and to
H. influenme type b (Hib) antþen.
Hammarstrom et al (1984b) found antiteiúoic acid antibodies were predominantly IgG2 and
generally absent in most lgG2-deficient donors, but substantial amounts of these antibodies in the IgGl
subclass were also found in some donors. Further work by Hamma¡strom et al (19S5a) has shown that in
the second year of life, antibodies to teichoic acid are predominantly IgG1, and that an IgG2 predominance
becomes apparent after 4 years of age. Hammarstrom et al (19S5b) have also demonstrated that specific
anti-dextran antibodies are mainly IgG2 (and IgA2) in normal donors, but somet"nes with substantial
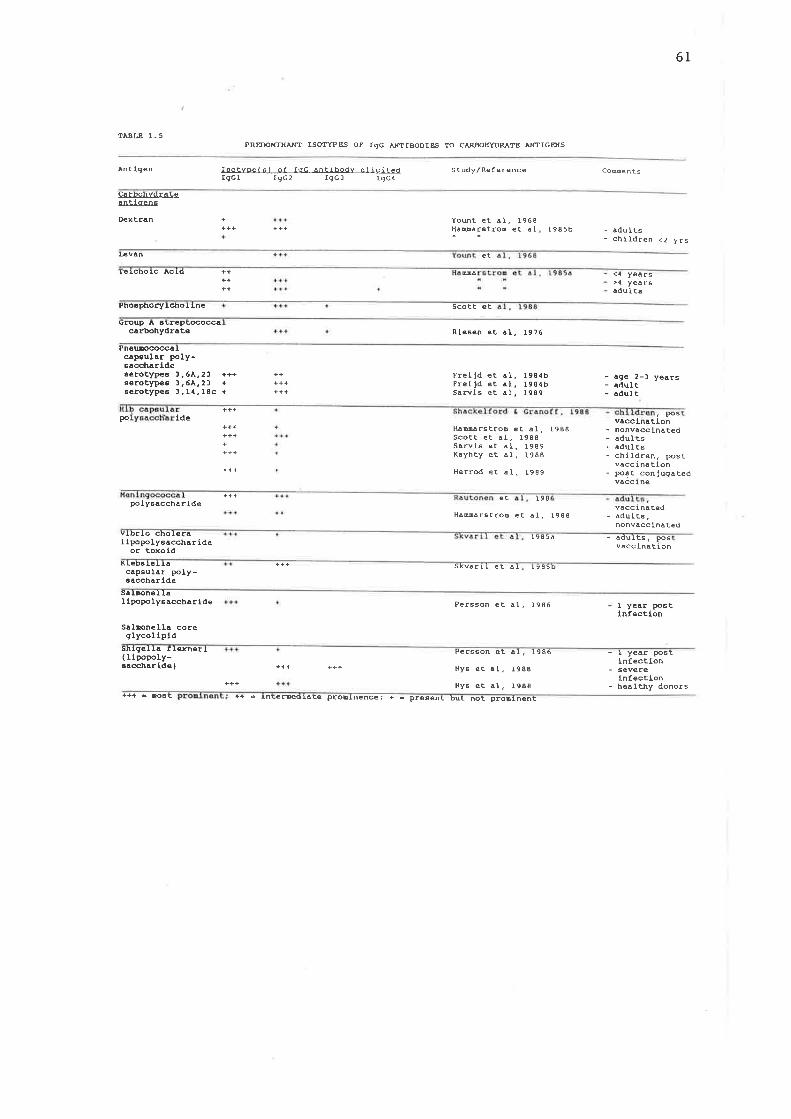
61
1À-8t 8 1.5PR-SDOICNÀ.IfT ISOTYPBS oP rgc AFTTBOOIES To C.\RBoEYDRÀTB NfTIGENS
À¡t (9en lsotvoelsl of roC Àntibodv eì icitedIgCl fgc2 IgGl IgC4
Study,/Reference Co@ents
CarbohvdrÀCeÂnÈl genE
Dextran +a++
Yoút et ÀI, 1968Hà@àr6troE et ò I t985b - .dults
- children <Z yrs[¿v n et
letcnolc Acto ++ Mâ <{ years>¡l yearsâdult6
PhoaphoryIc-t¡oI lne Scott et
9roup ^
aErepEæocqcarbohydrate Riesen eÈ ô1. t9?6
PnelJreoccalcöpaulÀr tþly-ÊaccharldeserotylE6 ¡.6À,21 +++serotl4EE 1,6À.21 +serotypes l,14. l8c +
+++FrelJd ecFrei rd etSâflls et
ô1.å1,À1,
t 984b1984br989
- age 2-l yeàrs- adulÈ- qd.ulÈ
+++po Lde
KaDürst'roD e! à1, I988Scott et àI, 1988SaRis ct ÄI. 1989Kayhty et â1, t988
Herrod ec ð1. 1989
vôcclnôt, PosIon
- nonvacclnÀced- àdults- âdulcs- chlldren, post
vacc Í na tlon- posÈ conJugàted
vÀcc lne
++++++
+++po ly6âcchÀr ide
e! t98
HàMârstroo et àI, l9govacci naÈed
- Àdults,nonvôccinâ ted
vlbrlo chol.eràI ipopolysacchÀr ide
or toxold- âdults.
vÀcc LnatPoslon
KLebaleIIà Skv¿ r!l et ôI t985bcâpsulôr poly-6accharlde
saIrcneIIaI lpopo Iy6acchar lde Persson ef al,, 1986 I yeôr po6t
InfectlonSôlrcnella coreqlycollpld
shlg6lIa flenerl( rlpopoly-saccharlde) +++
+++
Pêr66on et aI. 1986
Ny6 eÈ àI, t988
Ny6 et .1. 1988
- I yeâr postlnfectlon
- sever€ln fecÈlon
- hêalury donors+++ - rcaE ++ Ê tnEeæqlÀÈe proElnence; + r Pre€ent Þut noc pro¡lnent

62
amounts in the IgGl fraction. Most IgA-deficient donors lack anti-deÍran antibodies, but a few show anti-
dextran antibodiei in an unusual distribution, mostly IgGl and IgG3, even in ttre presence of normal IgG2
concentratio¡s. l¡ts¡sstingly, these donors all had a normal distribution of anti+eichoic acid antibodies and
generallS of anti-pneumococcal capsular polysaccharide antibodies. Anti-pneumococcal antibodies to
pneumococcal seroty¡res 3, 6A and ?3 have been found to be predominantly IgG1 in two to three year old
children and predominantlylgp2 in adults (Freijd et a[ 1984). In children with recurrent otitis media and
a mean age of 32 montls, thelgp2 antibody response was markedly deficient while the IgGl response was
nearer normal (Freijd et al ú8a).
Sarvas et al (1989) have demonstrated that pneumqcoccal vaccination in adults generally produces
a rise in IgG,JgA and/or ICM antibodies. IgG2 is the predominant subclass and IgGl is the next most
abundant, both before and after immunizatis¡. However the relative proportions of.IgG2 and IgGL
antibodies varied widely, from 1:0 to O:2. They also found a correlation between the G2m(n) allotlpe and
responsiveness in the IgG2 isot¡pe to vaccination with pneumococci tlpes lzlc and 18c. Preimmrrnization
IgG2 antibody concentrations to these serotypes were higûer in G2,m(n) positive individuals. The G2m(n)
allotype had no detectable effect on levels of IgG1, IgG4 and IgM antibodies, and possibly a weak effect on
IgG3 and IgA antibodies.
Sarvas et al (L989), in seeking to describe the isotlpe respo¡rses to diffcrent polysaccharide
antþens, standa¡dised their IgGl and IgG2 assays in relation to each other, eliminating one possible cause
of different results in diffo¡ent studies. Then, they found that pne 'mococcal IgG antibodies (to types 3, 14
and 18C) were predominantlylgp2 whereas meningococcal true A and Hib antibodies were IgGl and
lñ2 n equal mean proportions. Therefore antþenic differences between the individual polysaccharides,
rather than merely differences in methodology are likely to þe important.
Lane & Maclennan (1986) have reported four patients with IgA deficiency who failed to produce
IgG2 subclass antibodies to pneumococcal capsular polysaccharide despite normal, or near normat IgG2
levels.
Douglas et al (1983) found poor antibody responses in children younger than 5 years of age
immunised with a battery of pneumococcal capsular polysaccharidas induding tlpes 64, L4, L9F and 23F

63
which are the most likely seroty¡les to cause infection in child¡en. Cbildren between 2 and,|yoars of age
respond better thân younger children to some serot¡res (Lawrence et al 1983) but the¡ too, produce a
poor response to type 64.
While IgG antibodies to pneurnococcal polysaccharides a¡e predominantly IgG2 in adults, there is
overwhelming evidence that those against H. influenzac true b capsular polysaccharide (Hib CpS) are
predominantly IgG1. Previousl¡ because anti-Hib CPS antibody concent¡atiorxi were found to be lower in
IgG2-deficient subjects (Oxelius, L974; Siber et al 1980; Umetsu et a[ 1985) it was thought that these
antibodies were likely to be predomin¿nfly of the IgG2 isotype. Shackelford et al (1986) found reduced
baseline levels of anti-Hib CPS antibodies in only one of six IgG2 deficient children. It is of note that in the
IgG2-deficient patients who had low anti-Hib CPS concent¡ations, both Oxelius and Shackelfo¡d et al found
low IgG4 concentrations as well as low IgG2 concentrations. Umetsu et al and Siber et al did not mea¡iure
IgG4 in their patients.
When total concentrations of anti-Hib CPS antibodies are me¿uiured, a large proportion of IgA or
IgM antibodies may confi¡se the picture by obscuring an IgG isotype deficiency. Recentl¡ some
investþators have looked at isoty¡re-specific anti-Hib CPS antibodies, and, surprisingl¡ have found an IgGl
predominance in both children and adults. Shackelford and Granoff (1983), comparing the ratio of IgGl
and IgG2 anti-Hib CPS antibodies, found that IgGl antibodies predominated in bottr children and adults.
$imil¿¡l¡ Hammarstrom et al (1988) found a predominance of IgGl anti-Hib CPS in pooted sera from
both adults and children and also in sera from 14 of 16 IgG subclass deficient patients. Scott et al (1983)
found a predominance of IgGl anti-Hib CPS antibodies aftor Hib vaccination, in contrast to the usual
predominance of IgG2 for antibodies to phosphocoline and Group A streptococcal carbohydrate. One
subject, howevor, produced 46Vo of. the antiphosphocholine antibody in the IgGl subclass atd 44Vo n the
IgG3 subclass demonstrating that there can be much va¡iation between individuals. Makela et al (1937)
found equal proportions of IgGl and IgG2 anti-Hib CPS in young adults after immunization.
Subjects with G2m(n) allotlpe have been found to have higher levels of Hib antibodies and to
respond better to immunization with Hib capsular antþen than do subjects lacking this ¿llqttrps
(Ambrosino et a[ 1985).

64
The subclass restriction of specific antibodies may depend upon the adjuvant used. Attempts have
been made to modi$ polysaccharide vaccines by conjugation to protein antþens to make them T-
dependent instead of T-independent, and so to stimulate a more effective response in young children.
Shackelford and Granoff (1988) found that in children with a mean age of 10 months, initial immnnizatiea
with conjugated Hib CPS vaccine resulted in almost only IgGl, but re-immrnization with either plain or
conjugated vaccine resulted in some IgG2. Some, but not a[ Igc2-deficient child¡en had a dec¡eased
respouie to immunization with both Hib CPS and to pneumococcal capsular polysaccharide. Others
responded to the former, but not to the latter. The response was impaired in both the IgGl and IgG2
isotypes suggesting that IgG2 deficiency may be a marker of, rather than the cause of, impaired IgG
responses to Hib CPS vaccine.
IGyhty et al (1988) found the predominant IgG response to Hib CPS, or to its protein conjugate
vaccine (diphtheria-toxoid conjugated), in 24 month old childreq was predominantly IgG1. There was
relatively more IgA produced after the unconjugated vaccine. Their studies showed that the conjugated
and unconjugated vaccine induced antibodies that differed in bactericidal and/or opsonizing capacity. This
could not be e4plained by different proportions of the IgG isotlpes that were produced and may have been
related to differences in antibody classes, although these differenoes were not statistically significant.
Herrod et al (1989) found that imm 'nis¿¡isn with an Hib conjugate vaccine in 8 children over 22 months of
age who failed to respond to an unconjugated Hib vaccine resulted in a predominantly IgGl response.
None of the child¡en were IgG2-deficient. Granoff et al (1988) found no relationship between the failure of
response to Hib vaccine andlgpZ deficiency in a large group of children.
Scott et al (1988) described a gror¡p of children, who while responding poorly to carbohydrate
antþens, had normal IgG2 concentrations for their age. They also described 14 individuats with Wiskott
Aldrich Spdrome who had low or absent antibodies to 4 different carbohydrate antþens (Nahm et al
1986) but who had normal levels of Igp2 fot their age (Scott et al, 19SS). They found no correlation
between sorum IgG2levels and serum antibody titres to these carbohydrate antþens.
Antibodies of different subclasses may have different effrciencies in defence against certain micro-
organisms. Weinberg et al (1986) found, however, that þG1 and IgG2 antibodies to Hib CPS had fairly
equal functional activity when tested in an in vitro bactericidal assay.

65
Insel and Anderson (1986) described two patients with IgG2 deficiency and inability to produce
antibodies to bacterial capsular polysaccharides who were able to produce an IgGl response to a conjugate
vaccine composed of Hib CPS covalently tinked to diphtheria toxoid. Antibody titres to Hib CpS, theq
rose to what was considered q,rithin the protective range.
Briles et al (1987) suggests that the measurement of naturally occurring antibodies to
phosphocholine may be a useful indicator of carbohydrate antþen imms¡scompetence because these
antibodies to a component of the teichoic acid of pneumococci, are present at fairly constant levels in
almost all healthy individuals (except yonng children), ¿ad rnliks antibodies to Hib CpS and to group
specific streptococcal carbohydrate they are not dependent on recent illness or vaccination.
Antimeningococcal polysaccharide antibodies have been found to be both IgGl and IgG2.
(Rautonen et al 198ó; f{amm¿¡sþsm et al, 1988). The IgG response [6 galme¡ella lipopolysaccharide 0
antþns in infection is predominantly IgGl (Persson et al 1986). The response to the core glycolipid in
patients with severe infections is predominantly IgG2 and IgG3, while in healthy blood donors,
antþlycolipid antibodies are both IgGl- and IgG2 (Nys, 1988). The response to Shþella flexneri 0
Iipopolysaccharide antþens in infection is predomìnantly IgGl and IgG2 (persson et a[ 19g6). Vaccination
with either Wbrio cholerae whole cells (ie. lipopolysaccha¡ide antrgÐ or with procholeragenoid (toxoid)
produces predominantly IgGl antibody responses with some IgG2 and occasionally IgG4 responses
paficularly to the toxoid (Skvaril et 4 1985a). Klebsiella capsular polysaccharide produces a
predominantly IgG2 antibody respouie (Skvaril et al 1985b).
These few exanples serve to illustrate that the widely accepted idea that anti-polpaccharide
antibodie's are predominantly of the IgG2 isotype is an oversimplification- At some ages, and for some
carbohydrate antrgens, it is not even the rule. Some individuals have abnormal patterns of isotype
respolsiveness to certain antþens. The measu¡ement of srúclass specific antibodies to specific antþens
and studie.s of the frnctional effectiveness of such antibodies of different isotlpes, will be necesriary to heþ
clarify the situation, in both normal subjects and IgG subclass-deficient individuals.

661.6.13 Other antiçns (Table 1.6)
Some paræitic infections a¡e associated with high IgG4 concentrations. A variety of antibodies
have been identified in the IgG4 subclass. Anti-filarial antibodies of rgE and IgG4 isot¡pes have been
found in human filiariasis (Magnusson et al 1986). One study demonstrated a great preponderance of
IgG4 in 2 patients with tropical pulms¡¿ry eosinophilia and microfilaremia respectively and a relatively
high level of IgG3 in one patient with elephantiasis (Hussain et al L986). Patients with chronic
schistosomiasis have been found to have elevated IgG4 levels (Iskander et al 1981; Catty et al, 1936)
considered to represent antibodies directed against the egg which is known to cause an intense
hlpersensitivity response (C¡pess, 198Ð. IgG4 antibodies may block' the IgE-mediated h¡persensitivity
response to the parasite antþen (Hussain & Ottesen, 1986).
Antibodies to Ag 7, a major antþen of. Asperyillus fumigatus, are predominantly of the IgGl and
IgG4 isotypes (Harvey & Longbotton, 1986). Trypanosomø cruzi gives rise to IgGl and IgG3 responses
(Scott & Goss-Sampson, 1984; Romei¡o et a[ L98a) ard Plasmodium falciparum to IgGl or ILgp2.
(Wahlgren et al 1983).
Atopic and allergic patients may have high levels of IgG4 (Shakib et al, L977; Gwpn et at 19f8,
Heiner, 1984; Kemeny et al 1989b). Patients who have received immunotherapy for pollen allergies
develop dramatic increases in IgG4 (Devey et aJ, L976). Some suggest that IgG4 may be responsible for
'blocking antibody' activity decreasing allergen-induced disease in some allergic states (van der Giessen et
al" t976a; Nakagawa et at\ L987). Bee keepers often have high levels of IgG4 and bee venom specific IgG4
levels rise in patients receiving bee venom immunotherapy. The effectiveness of venom immunst¡e¡¿py
seems to correlate to some extent with rises in specific IgGa (Cheung et al, 1983; Djurup et a[ 1985 &
Urbanek et at 1986) but IgGl:IgG4 ratios may also þs important (Djurup & Osterballe, 1984). This is not
the only factor involved and patients with high lsyeþ of IgG4 may have anaphylactic reactions ¡q þss sfings
(Kemeny et a[ 1989a).
1.6.1.4 $rrmrn¿¡y
The relationship between IgG subclass deficiency and deficiency of antibodies to particular
antþens is hþþly compler Some patients have antibody deficiencies without IgG subclass deficiencies,
others have IgG subclass deficiencies with antibody deficiencies in the deficient and sometimes also in non-

67
TÀ.B LSPI@ICNN ISffiYPES OP IgG
^J{TIBoOIAS TO mEZR
^raTICEilS
A¡ti9en Predo[inànt r6otvæf s I of rqc antrbodv e I ic ¡ ledIgcl I9c2 rgc ) Igc{
Study/Reference
Filôri. X¿gnusson eL àI. 1986NussÀln et â1. t986
c¿rLy et â1, I986Àsperg I I lus
f ulgatu6Hàryey ! þngæcLoo, 1986
Trypåno6oba cruz i + 5COCt I @66-5ôoPSOn. I98{Ro¡elro e! ÀI. 1984
PIa6Ddtu(alclp.ru
+ Hôhlgren et à1, I98l
r@uoLherâ DvIC.g-P9IIÉEE:
VenoEs ch€sg N-K v.( bee keepers )Urb¿nek et àlD)urup et ô1.
et è1. t98l
I9861985
CrÀss pol Lens xvey et èl, 19760jurup ônd OsrerbàlIe, !984KeDeny €t ÀI, 1989b (àtopic)KeEeny et à1, I989b (non-òtopicl
Housèóust Dltes Nô9ôKàvà et â !, L9U /leEeny et ¿1. 1989b (ðlopicJXeDeny et ðI, 1989b (non-àÈoprc)

68deficient subclasses, and still others, it seems, a¡e subclass deficient but able to compensate by producing
protective antibodies of a different subclass. Ideally, antibody deficiency states should be detected by
measuring antibody concentrations and comparing these with normal rânges for age. However, the
quantitation of antibody tit¡es to a wide variety of antþens, both before and after irnmunization, and the
establishment of normal ¡anges at different ages for these para-eters would be a massive undertaking.
Fu¡thermore, tlere is, as yet, no consensus of opinion as to which antþens should be used given that a
patient may respond normally to one and not to anotler antþen in tle same goup. In the meantine, the
quantitation of IgG subclasses is likely to be a useful, although not absolute, goide to antibody deficiencies.
t6.2 Pmposed mechanisms of isoty¡le restriction
t.62.1 Models for isoty¡le restriction
The phenomenon of isotlpe restriction is not fully understood and its basis remains elusive. T\r,o
models have been proposed but neither explains it adequateþ According to the single-lineage model all B
cells develop in ¿ single lineage and can express all the IgG subclasses (Teale & Klinman, 1984; Gearhart et
al, 1980). Selective e:rpression of IgG subclasses is seen as the result of exogenous factors inducing the
successive expression of the IgG subclasses ¿sco¡ding to their order in the genome. I singls B cell under
experimental conditions catr express all the Ig isotlpes successively (Gearhart et al 1980; Teale 1982).
According to the mutliple-lineage model tlere are multiple B cell lineages that are pre-committed
to e4pressing particular IgG subclasses which develop at different stages in ontogeny (McKearn et al, 1982).
Mayumi et al (1983), by selectively depleting IgGl+ lgG2+ or IgG3+ cells, before the initiation of culture
and T cell stimulation, only affected the development of plasma cells bearing the honologous isotype, and
therefore suggesring that T cells were preferentially differentiation of IgG isotype precommitted
Bcells. Inhumans¡wherethe5'-3'CHregionisgeneorderisC-p,6,CI3,CI1,Cz1,Cl2,Qf 4,&,&?,the single-lineage model would predict that T-independent antþens, e.g. carbohydrate antþens, would elicit
predominantly IgG3 and IgGl, responses, since Cf 3 and C'yr are nearest the J end so that IgG3 and IgGl
production and require less isotype switching. The finding ttrat some carbohydrate antþens elicit
predominantly IgG2 antibodies, (see section L.6.1.2) is ha¡d to orplain accordi"g f6 this model Scott et al
(1988) propose that the frequent ooncurrence of IgGl-IgG3 and IgG2-IgG4 deficiencies may indicate that
separate B cell lineages express the IgGl-IgG3 or the lgG2-IgG4'sets' of subclasses and respond to

69different antþens and lymphokines. flowever, evidenco for the frequent oonclurence of the above-
mentioned deficiencies although traditionally accepted is, in fact, somewhat tenuous.
1.622 Factors that may influence the pattem of þG subclass prod.uction
(a) The tlpe of antþen. Protein antþens often elicit IgGl and IgG3 antibodies while
carbohydrate antþens more commonly elicit lgG2 and/or IgGl (section 1.6.1-).
(b) The age of the subject e.g. pneumococcal polysaccharides elicit mainly IgGl in young
children, and mainly lñ2ln most adults (Freijd et a[ 1984).
(c) Repeated immunizations may change the pattern of subclass specific antibodies (Aalberse
et al 1983).
(d) amount of the antþen given. Hþh antþen doses give rise to low affinity antibodies in
secondary immrrnizatisn and vice versa (Roitt et at 1935).
(e) The site of the immune reaction. An antþen may evoke different tlpes of imms¡s
response cells in different parts of the body:-
(Ð T cells from Peyer's patches preferentially induce an Iglvf to IgA switch in
immunoglobulin-bearing cells in mice (Kawanishi et al 1983a; Kawanishi et al
1e83b).
(ü) The distribution of sells co¡teining immunoglobulins of different isotypes is
known to vary in different orgÍìns (Partridge et al, 1984). Antigens presented
naturally may give rise to diffe¡ent IgG subclass patterns from antigens presented
by immunizatist. Anti-Rh antibodies formed in incompatible pregnancies are
generally IgGl, IgG3 and IgG4 whereas those formed after repeated
imnunization may include the IgM and IgG2 isot¡pes (Morell et ú, L9'13; Devey
&Doak,L97\.
(Ð The degree of T cell dependence. T cells and T-derived products may be a major factor in
determining which subclasses are produced by B cells (Teale & Abraham, 1987). The
same antþen may be able to interact with B cells in a T-dependent and in a T-
independent mânner, and in an MHC-restric¡ed and an MHC-unrqstricted mânner,
depending on the dose of the antþen (Asano et al 1933).
(g) Genetic differencps e.g. patients with different alloty¡les respond differently to different
antþens ( Ambrosino et a! 1985; Sarvis et al, 1989).

(ü)
(h)
70
The use of adjuvants. The effect of adjuvants on the pattern of IgG subclass isotype
fesponse fs immunization is not yet clear (Shackelford & Granoff, 1988; Kayhty et al,
1e88).
Most isotype deficiencies cannot be attributed to structural gene deletions because these
deficiencies are relative rather than absolute ie. most subjects produce small quantities of the deficient
isotpe(s). While an arrest or impairment of sequential isotype switching might explain some such
deficiencies there is evidence ffi¿t rhis is too simplistic an explanatiou to e:rplain them all because:-
(Ð Healthy subjects with partial IgA deficiencies are not generally IgG subclass-deficient. They can,
therefore, utilise CH genetic material that lies downstream to that for Igd indicating that isogrye
switching has not been merely inhibited from this point on.
(üi)
T cell-independent antþens in man do not produce predominantly IgM, IgG3 and IgGl antibodies
as one would e¡pect if absence of T cell help simply resulted in a failure to promote more
downstream switches. Rather, failu¡e to respond to T cell-independent antþens may result in a
lack of lgM,Iñ2 and IgGa aftibodies (Schur, 1987) all of which are not, in fact, t¡ansç¡iþsd frs6
adjacent parts of the CH gene.
Persson et al (1986) have showed that IgA antibodies appeü before IgG antibodies the ¡n vivo in
f,esponse to infection with Salmonella serogroup B O antþens, indicating that the temporal
appearanc€ of certain isotypes depends on some mechanism other than a sequential switching
from the production of one isotype to another ^'¡, 1,b upstream from c 1, andø2, Different clones
of B cells may be involved or one IgA - producing clone and one IgG - producing clone may have
developed from the same original B ce[, with IgG and IgA production being differently regulated.
Since specific tlpes of antþens tend to produce certain cha¡acteristic patterns of isot¡pe rosporue
it may be that isot¡pe deficiencies represent the cumulative effect of a failure to produce antibodies to a
number of antþens which would normally elicit a respou;e in the isotpe(s) concerned. Persson et al
(1986) suggest that patterns of isotlpe response may be dependent on a selective switch in precomnitted
cells, with certain V-D-J combinations preferentially combining with certain immunoglobulin hoavy chain
genes.

7T
Theoreticall¡ one can postulate a number of possible nschanisms that may result in either IgG
subclass doficiency and/or low levels of antibodies to particular tlpes of antþens.
These include:
a) Impaired antigen processing and presentation by macrophages, monocytes and B cells,
b) Impaired antþen recognition by T cells.
") rmpaired antþen recognition by B cells.
d) Defective production of regulatory (e.g. isot¡,pe switch-promsting) cytokines.
Ð A relative lack of B cells preco--itted to the production of certain isot¡pes
Ð A failu¡e of B cells to respond to certain cytokines.
g) Inhibition or arrest of isotlpe switching.
Ð Suppressor factors affecting the production of certain antibodies.
j) H¡percatabolism involving some isotypes more than others.
Evidence is lacking for tlese or other mechanisms i" IgG subclass deficiencypersø, although in
some more severe hypogemmaglobulinaemias, some of them have been demonstrated. eg.
- impaired immunoglobulin production in the bare þrm.phocyte slmdrome where the absence of
MHC antigens prevents normal antþen presentation and immune function cell interaction
(Touraine & Betue! 1983;Haas & Stiehm, L987),
- absence of B cells in Bruton's Disease.
- defective lSmphokins p¡efluçfion in some patients with hlpogammaglobulinaemia (see section
L.3.3.L).
L.7 IgG SUBCLASSES IN CHILDREN WITH RECURREI{T INFECTIONS
A great deal remains to be clarified with respect to the importance of IgG subclass levels in
relation to infection proneness. While it is knoym that some antþens stimulate IgG antibody production of
predominantly one subclass in most normal people and that subclass deficiency may be associated with
impaired specific antibody resporu¡e (see section 1.6.1) there are also reports of subclass-deficient
individuals who are not excessively infection-prone (Lefranc et a1', LV79; van Logbem et al, 1980;
Hammarstrom & Smith, 1983b; Lefranc et al 1983a). Functioning c6mpensatory immune meúanisms are
obviously important. Because most studies of IgG subclasses to date have not attempted to assess other
aspects of immune fr¡nction in the subjects studied it has not been possible for these studies to demonstrate

72whether IgG subclass deficiency alone, or IgG subclass deficiency in conjunction with some other immune
abnormality, aooorxots for the infection proneness of the patients reported.
There have been relatively few reports of IgG subclass concentrations in patients with recurrent
respiratory infections who were not selected in the basis of IgA deficiency. Table 1-.7 snmmffiss some of
these. The prevalence of IgG subclass deficiency in such patients is unknown. There have been isolated
reports of individuals with IgG subclass deficiencies and severe respiratory infections.
Studies in groups of children are relatively few. Oxelius (1984) found reduced concent¡ations of
IgG2 in a gfoup of 20 otitis-prone children whose IgGl, IgG3, and IgG4 were sinilar to cont¡ol
concentrations.. IgA concentratioû¡ were not reported. Those with IgG2 concentrations of less than 0.60
g/l at 72 montls of age had more haemophilus infections and those with hþher levels of IgG2 had more
pneumococcal infections. Although antibodies to Hib CPS are predominantly of the IgGl isotype (section
1.6.1.2), otitis media is not usually caused by encapsulated Hib.
Smith et al (198a) $udied a group of 37 non-allergic children with recurrent chest symptoms such
as bronchitis, asthmq ch¡onic cougb and recurrent respiratory infections. TWenty-three of these had
normal IgG concentrations. Of these ?3, rorre had low IgG1, 3, (1j%) had low IgG2, none had low IgG3
and 11, (45%), had low IgG4. Of the 14 who had an IgG concentration more than 2 SD below the mean,
STVowere lgGl-deficient,50Vo were IgG2-deficienq 7Vowerc IgG3-deficient and 2LVowere lgG4-deficient.
Overalt IgG2 deficiency was a little more coÍrmon than IgGl deficiency, and borderline-low IgG2
concentrations were more common than borderlinelow IgGl concentrations.
Umetsu et al (198Ð found 20 children with recurrent sino-pulmonary infections and serum IgG
concpntrationss within the normal range who were IgG subclass-deficiont. TWelve were IgG2 deficient, 5
were IgG3-deficient and 3 were deficient in both IgG2 and IgG3. Three of the Z) were IgA-deficient.
These 20 children, presented over a tlree year period to the allerg5r clinic at the Children's Hospital in
Boston because of recurrent sino-pulnonary infections. Their IgG subclass concentrations were more than
2 SD below the mean for age. Nornal values were obtained from healthy or astlmatic child¡en without
recurrent infections attending the clinic. Total numbers attending for sino-pulmonary infections are not
giveq although ens might think that either the finding of only Z) subclass deficiencies in 3 years may

73
TÀBLE 1.7rgc sul}sfl{ss DEPICIENCIES IN PÀTIEIIDS rfITE REcnRRgN.r INrEqrIoNs
study Su-bJect'a cllnlcaL flndlngs IBotop€def lc lency
Comen ts
rry,oo
ever Pa gG2 coDpacontroLs
ng e age-Da3nly girl
frequent URTIrecurrent severerespiratory, skin& urlnary lnfectlons
l9G 3I9c I
Schuet al
8y. o. boy27y -o.J.3y.o.9lrl
Recurrentpyogenlc1n fectlons
IgGl, I9G2 , IgG4IgGI, l9G2 , IgclIgGr , lgc2
conpared wlth 2o adult controlt9?0
oxelius, l9?4 I aBl Iy( not-tter É 2chl. Idren )
chron).c pY09enrc L9ç2 +I nfect ions
Heiner,198lBeck É I Pa E ents Pu r Dona ry rgGc
IgG4
r^ 4/29 päÈients
in l/l pacients
lnfectlon -including bronchiectas isBronch iectas 1 s
Bass et al, 1981 I ch l Id Recurrent Igc2neningococca J.
nenlngltls
SElth, et aI, 1984 37 cn I lClren ¡(ec[rrenE. non-allerglc chestsynptoE
IgGL t L9G2I9G3, IgG4
Helner, 1984 L2 patienes
l5 patienes
Severe orrecurrent pyogenlcInfectlonsBronchlectas is
IgG4
I9G 4 ( in 5,/ls)
oxel. lus. I984 20 children Otrtrs nedra 19G2
stanleyet al, 1984
47 patlents53 patient
chronic RTIÀcute RTl
I9G2 > I9G4*IgG2 > IgG4r
Pìebanlet aL. I985
I IÌyr olcl Recurrentinfections
lgG 2 deficit of IgG and IgHantibod ies
UEe tS uet al, 1985
20 chrldren Recurrent si.no-pulnonary lnfectlon
IgG2> I9G3r
Àm,bro s 1noet aI, 1985
Hlb lnch i ldren
Hib I9G 2
Pre.!lneret al., 1985
I âduLt Recurrentpneuococca Iin fectlons
l9G2,I9G4,IgÀ poor levels of antl-polysacchar ideantlbodles
we I tuaet al, 1986
65A() aOUr t6 Severe, rec\lrren Eor chronÍc infectlons
I9G3 > r9cr
Shacke I fordet al, 1986
lO chllctren Recuren tinf ections
IgG2 > Igc4r
Helner.1986 162 patlentsvithrecurrentlnfecÈions
Recurrent lnfectlons I9G4!I9G2 neasured IgG4 1st ancl ot¡ersc only in the lgc4-deficient
SoderstroDet aI, 1987
l7¡l sccl acluIE129 scdchlldren
Recurentinfections& otherconditlons
IgGl> IgG1> I9G2rIgG2> Igc I > Igc 1r
Read eÈ aI. 1988 z aclulLs
r lndlcates tìat deficlencles of both isotyPes occurred in t-his group of patients, one Eore frequenÈIythan tle otler -
Chronic fatiguesYndroDet r
IgGlintelrlttentfever, sore tlroåt
scd = IgG subclasa-deficient

74
indicate eitler a lower incidence of subclass deficiencies than in some other studies, or that the criteria for
defining subclass deficiency may have been stricter rhan in some other studies.
Shackelford et al (1-98ó) measu¡ed IgG subclasses in 30 children with severe recurrent infections
including multiple episodes of pneu"'onia" bacteraemiao recurrent disc.hargmg otitis media and recu¡rent
pyogenic sinusitis. Using a solid-phase competitive inhibition assa¡ they found that 7 of the 30 children had
abnormally low serum concentrations of.Igp2 (lower than 3 SD below the geometric mean for healthy
subjects). Only two had abnormally low total IgG levels. TVo had IgA deficiency. Three had very low
concentrationsof IgG4(i.e. <0.0rmg/mlatover3yearsof ageand <0.005mg/mlatunder3yearsof age).
Ability to produce specific antibodies in the 7 children varied with respect to anti-tetanus toxoid antibodies,
antibodies to_substances A and B on red blood cells and antibodies to Hib CPS. Qsmplement
concentrationss and þmphocyte subpopulations were normal in most cases. In rhis study the major
difference between theT lgp2-deficient children and the 23 who were not þG2-deficient was the severity of
tleir recurrent infections.
These studies suggest that in children with recurrent infections, IgG2 deficiencies may be the most
common IgG subclass deficiencies. In adults, some studies have found a relatively highsr incidence of IgG3
deficiencies (see Table 1.7).
Until recently, definition of IgG4 deficiency has been a problem because commonly used methods
of measu¡ement eg. RID and EIA have not been sufñciently sensitive to detect IgG4 iû considerable
numbers of nornal healthy subjects. This may have been because of limitations intrinsic to the technique
used (RID is inaccurate below levels of 0.01-0.03 g/l), or because of the eÍensively absorbed polyclonal
antisera that were used failed to detect a subpopulation of IgG4 molecules (French & Harrison, 1984a).
Oxelius $n9{ has found that levels of IgG4 vary widely in normal children. Igp2 and,IgG4 are
slower to increase with age than are IgGl and IgG3, but even in older age groups, using an 6¡r¡ ¡echnique
and polyclonal antisera, Oxelius was unable to detect IgG4 in 12-?AVo of normal controls. French and
Harrison (1984b), using RID and monoclonal antisera, found IgG4 undetectable n LÙVo of normals and
more often in females than in males, in a British population. Previousl¡ Shakib and Stanworth (1980b), in
¿ similar population, using polyclonal antisera and RID, found lower concentrations of IgGa. Shackelford

75
et al (1986), using a solid-phase competitive inhibition assay found that IgG4 was measurable, but below
O.0I g/l in 4 of 19 normal child¡en from 1 to 3 years of age.
In 1-98L, Beck and f{einer, using a sensitive radioimmunoassay, found that fou¡ of 29 patients with
recurrent pulmonary infections had an isolated IgG4 deficiency. Three of these fou¡ patients had
bronchiectasis. They found that levels under 0.0001 g/l appeared to be associated with an unusual
susceptibility to infections and that levels under 0.0L g/l usually did. Later, Heiner (1984) reported IgG4
deficiency in 5 of 35 patients with bronchiectasis, and identified 12 patients in whom an isolated dehciency
of IgG4 was associated with recurrent or chronic pyogenic infections. Two of these patients, a motler and
daughter, had an isolated absence of IgG4 and chronic sinusitis, recurrent pneumonia and bronchiectasis.
A sister had a very low but detectable IgG4level but did not suffer from excessive infections (Heiner et al,
1e83).
In a subsequent study, Heiner et al (L936) found IgG4 deficiency (defined as <0.03g/l) n?Á% of.
362 patients who were nearly all children excessively prone to bacterial respiratory t¡act infections such
sinusitis, otitis media" pneumonia and bronchiectasis. In about 25Vo, thts was the only immunoglobulin
deficiency, and in abotú 75Vo of those witl recurrent infections tlere was a deficiency of at least one other
immunoglobulin isotype. The prelimi"ary e4perience of Heiner's group suggested that isolated IgG4
dehciency may be more comrnon than isolated deficiency of any other IgG isotype. Later, however, they
reported (Heiner et al, 1988) in infection-prone patients an incidence of 38Vo of.IgG2 deficiency taking
uormal values from a composite of normal rânges from the literature, and an incidence of 26Vo IgG4
deficiency, using their own normal ranges.
Itr this later paper, several specific instances of life-threateni"g infections in patients with isolated
IgG4 deficiencies a¡e described. One such patient was a 9 year old boy with staphylocoeal septic
polyarthritis, extensive osteomyelitis and purulent pericarditis who failed to repond to intensive
antibacterial therap¡ and who improved dramatically when intrave¡s¡s imms¡oglobulin was given. The
specific antibody resporxie to the S. aureus cultured from his blood was very low. After the initiation of
gamm¿gfsþulin therap¡ his serum showed a three-fold rise in S. øtreus specific lgp2 añ a seven-fold rise
in IgG4 antibody (Schoettler et al 1-986). Another patient described is an infant aged 4 weeks who suffered
overwhelming meningococcal sepsis, ultimately requiring amputation of several extremities because of

76
e$ensive ecchynoses and tissue necrosis who had an isolated IgG4 deficiency, as did his motler (Heiner et
al,1988).
Thus, to date, studies in infection-prone children have suggested that IgG2 deficiency may be the
most common IgG subclass deficiency. Most investþators, holvever, have not been able to comnent on the
occluïense of IgG4 deficiency because of limitations in the assay systems and normal ranges used. The
overall incidence of IgG subclass deficiencies in infection-prone children is not known and neither is it
known whether IgG subclass deficiencies are more corlmon in children with susceptibility to certain types
of infections. In Chapters 5 to 12 I shall add¡ess some of these issues.
1.8 Igc SUBCI"ASS CONCENTRATIONS IN PATIENTS WITII RECOGMSEI)
IMMI]NODEFICIENCY DISORDERS
Patients with various immunodeficiency disorders provide a special resouroe to add to our
¡¡dg¡sfanding of both the development and the significance of IgG subclass deficiencies. l¡ is þesoming
realised increasingly that the quantitation of IgC subclasses may be an important part of tle assessment of
a patient's immunological status. Subclass measurements may not only enable the detection of an otherwise
undetected immunodeficiency, but also may enable a better definition of the nature of an already
recog¡ised immunodeficiency. The occurrence of certain patterns of IgG subclass concentrations in
different tlpes of immune deficiency may throw fu¡ther lþht on such factors as the possible T-cell
dependence or independence of the spthesis of IgG subclasses in humans.
Table 1.8 summarises a number of studies in which IgG subclasses have been measured in a variety
of immunodeficiency states which are discussed below. The assays and normal rânges used are different in
many instances and this may account for some of the variability in findings. Overa[ however, some
patterns appoar to emerge.
In patients with panhypogammaglobulinaemiao lgGl^,lgp2 and IgG4 levels are generally low. In
the X-linked form, IgG3 levels a¡e generally low also. In common va¡iable h¡'pogammaglobulinaemiq
however, IgG3 levels show muú variability from patient to patient, and may even be raised absolutel¡ or
relatively (Yount et aL, L970; Vi¡ella et a\ L9Ift; Aucoutu¡ier et a[ 1986). Van der Gissen et al (196b)
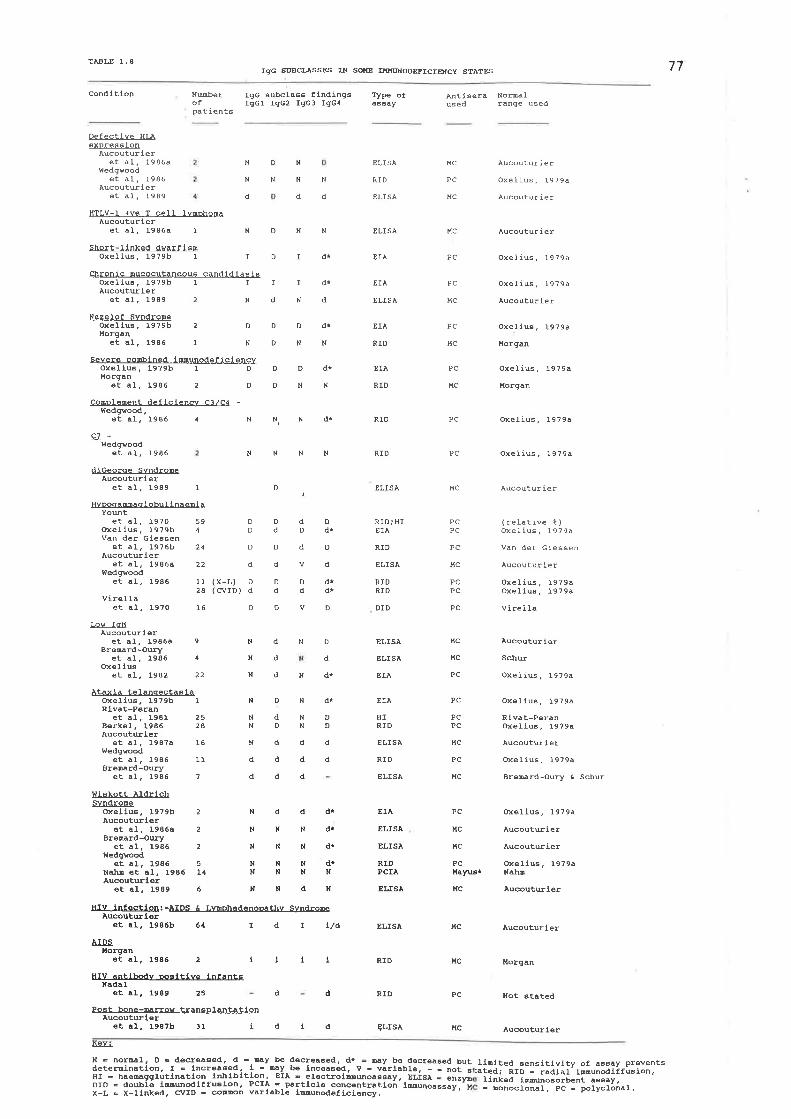
TÀ.SLB I . 877Igc SUBCIÀsses IN SoKE IxKl¡{oDEFIcIE[Cy 9IÀ1ES
cond ltlon Nu¡.b€rofPatient6
Type ofå634y
Igc Êu-bcla6Ê flndlngsIgcl lgc2 IgGS IgG4
À¡tisera NotuaLused range used
Defectlve HLAexpre g s lon
Àucouturleret al? 1986a
f,ledgvoodet âL, Ì986
Àucouturleret al, 1989
HTLV-I +ve T cell lnphonaAucouturler
et â1, I986a I
Short-Ilnked dwarflsnoxellus,1979b I
Chronic Eucocutôneou6 candldlasisOxeIIu6, 1979b I I
N
d
N
N
d
D
N
N
N
d
ELISÀ
RID
ELISÀ
ELISÀ
EIÀ
EIÀ
ELISÀ
EIÀ
RID
BIÀ
RID
HC
PC
MC
MC
PC
PC
MC
PC
MC
PC
I'fc
PC
PC
HC
PCPC
PC
Àucouturier
OxeIius, t9?9a
Àucouturier
Àucoutur i e r
OxeIius, 1979a
oxelius, I979ô
Aucoutur ier
Oxellua, I979a
HoFgan
Oxelius, 19?9â
l,forgan
Oxèlius, 19?9a
Oxel.ius, 1979a
ÀucouÈur ier
( relðtive t )Oxelius, l9?9a
Van der ciêssen
Àucoutwi,er
Oxelius, 1979aOxelius, 19794
virêi Ia
Àucoutur iêr
Schu
ûxelius ? t9?9a
Oxel-ius, I979ô
Rlvat-PeränOxelius, I979a
Àucouturier
Oxelius, I979a
BreErd-Oury & Schu
oxelius, 1979a
Àucoutuier
Àucoucurier
Oxelius, 1979aNah8
ÀucoutEler
Àucoutuier
l{or9an
Not atâted
Àucouturler
Àucouturleret al, 1989 2
NDNN
IDId*
IIdT
NdNd
Nezelof SvndropeOxeLlu. 1979b 2 O
l{organet al. 1986 1 N
Severe copbined lMuodef lclencyOxellu6, 1979b I DMorgan
et aI, 1986 2 D
Coppleuent deflclencv C3./c4 -Wedgvood ,
et ÀI. 1986 4 N
f,ledgwoodet al¿ 1986 N
diceorce SvndropeAucouturler
et al¿ 1989 I
HvMaNàql.obuI lnaeEi ÀYout
et à1, I97O 59oxeIlu6. 1979b 4van der cleasen
et aI, I9?6b 24Àucoutuler
et al, 1986a 22t'ledgYood
et al. 1986 11.2A
virellaet al, l97O 16
DDdT
DNN
DDdI
DNN
NNd*
NNN
( X-L)( cvID )
D
DddD
Dd
dv
DDdd
DV
RID; HIEIÀ
RID
ELISÀ
RIDRID
DID
RIO
RID
ELISÀ
ELISÀ
RIO
RID
ELtSÀ
DD
D
d
Dd
o
Dd*
D
d
drd*
MC
PCPC
D PC
HC
t{c
PC
PC
PCPC
HC
PC
t{c
l,fc
llc
PC
I¡w IgLlÀucoutuler
et al, 1986aBreDard-OurI¡
et al, 1986oxellu
et al. 1982 22
Àtdla telanqectðEiaoxellu6,1979b 1Rlvat-Peran
et aI, 1981 25B€rkèÌ,1986 28Aucouttl,er
et âI, 198?a 16wedgvood
et al, 1986 1lBrêerd-Oury
et al, 1986 7
wiskott ÀldrlchSvndroEe
ûxellu¿ 1979b 2Àucoutuler
êÈ aI, 1986a 2BreEard-Oury
et al, 1986 2¡{edgYood
et aI, 1986 5NaÌ¡¡ et al. 1986 14Àucoutwler
et al, ¡-989 6
ÀIDSllorgan
et aì., :.986 2
et aL, 1989 25
dND
d
dHdr
DNdT
Nddd
d
rdÍL/à
ii11
4 d
N
N
N
N
9 ELISÀ
ELISÀ
EIÀ
NdNDNDND
d
d
d
d
d
d
EtÀ
HIRID
ELISÀ
RIO
ELISÀ
EIÀ
ELISÀ
ELISÀ
RIDPCIÀ
ELISÀ
HIV lnfectlon:Àucoutwl€r
-ÀIDS Ê Lwohad€noEttrv Sndrope
et à1. 1986b 64
Halrusr
Ndddr
NNNdT
NNNd*
NNNdTNNNH
NNdN HC
t{c
t{c
PC
HC
d
HIV ÀnÈlbodv æslÈfve infantsNadaI
d
PosÈ bone-uroy tranÊplantatlonÀucoutuler
et al., I987b 31 idtd
AEY-L
N = noruI, D o decr€ãÉed, 9 -. *y b€ decreôEed, dr = Ey b€ decrea6ed but tlDiÈed aensitlvlty of as6ay preventsdeteminatlon, I = Lncreaded, 1= *y b€ IncêaEed. V - varlâble, - - not B:ated, RIO _ radial fuu;fifir.ion,Hr = hae@gglutlnatron Lr¡-l¡rbltlon , ErÀ ! el€ctroleuoaeaay, Bt is¡ - enzyn linked l@uosorbent a66ay,DtD = dou-btê lNuodlffusLon, PcrÀ - pårtlcle concêntratloñ tuuoassay,'x E Eonocronar, pc - poryclonal,X_L = X-llnked, CÍ/ID = comon varl.abl€ l@uodef iciency.

7B
found ¿ high incidence of combined IgA\ IgD, IgE and lñ4 deficiency in patients with
hlpogammaglobulenaemia and also in their ¡elatives. Relatives may also have a deficiency of only one or
two of these isotypes. Wedgwood et al (1986) point out that the IgG subclass involvement in
immunodeficiency disorders is often not proportionate. There appears to be an hierarchical order of IgG
subclass deficiency ie. IgG3 < IgGl < IgG2 < IgG4. The IgG3 - IgGl segments of the genetic code and
the IgG2 - IgG4 segments are separated by a long segment including 2 pseudo-genes and the heavy chain
constant region for the IgAl- gene. Wedryood suggests that both sequenoe and distance may affect the
hiorarchy of IgG subclass involvement. The variety of deficiences in com-on variable immunodeficiency
may reflect a progressive sequential impairment of the programrned cascade for downstrean heavy chain
constant gene rearangement, and this ma¡ at least in part be related to impaired production of or
response.to lymphokines (section t.3.3.L). Major variations in the pattern of isotlpe deficiency in individual
patients may occlrr q,rith rims. Aucoutu¡ier et al (L989) report variation from almost complete
agammaglobuli"aemia to IgA-IgG2-IgG4 deficiency, as well as tle reverse situatiou.
A few patients with low IgM and normal 6¡ high IgG and IgA have been studied (Aucouturier et
af 1986a; Bremard-Oury et al 1986). In these patients IgG4 deficiency has been a sommon fi¡ding. These
2 studies appear to include some of tle sarne patients
Many patients with IgA deficiency have been studied and these are discussed separately in section
1.5 and Chapter 5.
Ataxia telangiectasia is a rare autosomal recessive disorder in which T and B cell functions are
abnormal and there is a defect in DNA repair €using chromosome breakage and/or frequent
t¡anslocations or inversions involving several ci¡omosomal sites, corresponding to T cell recoptor and/or
immunoglobulin gene complexes (Aucouturier et al 1987a). Delayed hlpersensitivity reactioru¡, abnormal
in vítro responses to mitogens and antigens and low or undetectable concentrations of serum IgA and IgE
are usually found. The predominant findings with regard to IgG subclass levels seem to have been in good
agreement in the reported studies (Table 1.8). I-ow concent¡ations of IgpZ aú IgG4 are found in
considerable uumbers of patients. IgGl and IgG3 have been low in a few patients also (Wedgwood ot al
1986; Bremard-Oury, 1986). The subclass deficiencies a¡e not always associated with IgA deficiency
(Aucouturier et al L9üa).

79
The concurrence of IW,I$E,IgG2 and IgG4 deficiency in many eases of ataxia telangiectasia
may be related to the known deficiency of T cell function, and would be consistent with the hlpothesis that
the erçression of Ig genes which are located downstream in the CH locus (ie IgG2 and IgG4) is more
dependent on T cell help than is the e4pression of more upstreâm getres such as tlose for IgG3 and IgGl
(Aucouturier et al 1-987a).
The Wiskott-Aldrich slmdrome is another rare genetic disorder associated with abnormalities of
both B and T cell function, in addition to eczema and th¡ombocytopaenia. Tnheritance is XJinked. Serum
Iglvf is often low and IgA and IgE elevated. Patients are prone to autoantibody production and to the
development oT lymphoid malþancies. They are abnormally infection-prone and have a poor antibody
response to carbohydrate antþens (Standen, 1988). Studies of small numbers of patients have suggested
that IgG subclass deficiencies, particularly that of IgGd may not be unsommon (Oxelius, L979b;
Aucouturier et al 1986a; Wedgwood et al, 1-986; Bremard-Oury et al 1986). Nahm et al (L986) studied 14
patients and did not detect a significant incidence of IgG subclass deficiency. Fu¡thermore, this s¡dy fid
not find any strong correlation between concent¡ations of antibodies to 3 carbohydrate antþens
þhosphocholine, group specific carbohydrate of Streptococcus Wogenes and the carbohydrate capsule of I/.
influenzae qpe 6) and IgG2 subclass concentrations. The concentrations of antibodies to these antþns
were low in the presence of normal IgG2 concentrations.
Aucouturier, in a later study (1989) of 6 patients with Wiskott-Aldrich syndrome found the only
IgG subclass deficiency to be that of IgG3 in one patient. Concentrations of IgG4 of <0.01 g/l were found
in 2 of the patients but these were not noted to b.e deficient in the tef although from the data in the tables
they appear to be so.
Patients with severe combined immunodeficioncy may have low concent¡ations of all IgG
subclasses (Oxelius, 1n9b). Morgan et al (1986) however, have reported 2 with low IgGl, undetectable
IgG2 and normal IgG3 and IgG4, and another, with Nezelofs syndrome, with undetectable IgG2, but
normal IgGl, IgG3 and IgG4, suggesting that IgG2 may be disproportionately reduced. Aucoutu¡ier et al
(1939) have a patient with partial diGeorge slmdrome and IgG2 deficiency. Such a disproportionate
reduction in IgG2 is hard to explain

80
In the bare lymphocyte slmdrome defective e4pression of HI-A is associated with varying degrees
of immunodeficiency (Haas and Stiehm, 1987). Aucoutu¡ier et al (1986a) found low concentrations of IgG2
and IgG4 in 2 patients with defective HLA expression, Wedgwood et al (1986) failed to find any IgG
subclass deficiency in 2 such patients and later, Aucouterier et al (1989) found low concentrations of IgG2
in 4 patients with or without another subclass deficiency.
Oxelius (L979b) has measured IgG subclass concentrations in a patient with short-limbed
dwarfism (cartilage-hair-h1'poplasia syndrome) and found IgG2 and IgG4 undetectable but IgGl and IgG3
high. In a patient with chronic mucocutaneous candidiasis, Oxelius Qn9Ð for¡nd raised levels of IgGl,
ICCZ anf lCC": tut undetectable IgG4. Because of limitations in the assay technique used (Oxelius t979a)
these undetectable levels of IgG4 could not necessarilybe defined as deficiencies. Aucouturier et al (1989)
using a sensitive ELISA assay, found low IgG2 levels in 2 of 4 patients with chronic mucoutaneous
candidiasis, and low IgG4 in one of them. Raised levels were not distinguished.
Wedgwood et al (1986) have looked for IgG subclass deficiencies in association with serum
complement deficiencies. The possibility of IgG4 deficiency in patients utrth e/C4 deficiency should be
considered" but futher studies with more sensitive assays are needed. Oxelius et al (1986) report a family
where IgG3 and C2 deficiencies were associated. The IgG3 deficiency was found in family members
homozygous and heterorygous for C2 deficiency and also in one number without the C2 deficiency.
The possible conctulenoe of IgG subclass deficiency and neutrophil defects is yet to be
investþated. In 3 reported patients with defective leucocyte adherence protein (Aucouturier et al 1986a;
Wedgwood et a[ 1986a) no IgG subclass deficiencies have been found, although two were reported to have
defective humoral immune responses (Bowen et a[ 1982).
Some viral infections have profound effects on the imnune system. Human deficiency virus (IilÐ
infection has a devastating effect on T4 cell fi¡nction and also impairs specific antibody production despite
polyclonal B cell stimulation (Janoff et a[ 1988). Infants with HfV infection a¡e excessively prone to
infection by organisms with capsular polysaccharides as well as to the secondary infections more usually
associated with HfV infection (Kanani and Krilov, 1987). Studies of IgG subclass concentrations in HIV

81infected patients, to date, have generally been in adults. (Morgan et al, 198ó; Aucoutr¡¡ier et a[ 19S6b).
The predominent pattern seems to be raised IgGl and IgG3 Ìvith low or normal IgG2 and variable IgG4.
In seropositive infants, Igp2 and,IgG4 deficiency has also been reported (Nadal et at 1989). This may
indicate impaired isotype switching because of a lack of T helper cell fi¡nction.
Patients with Epstein-Barr virus (EBÐ infection may also deveþ hlpergammaglobulinaeniq
attributed to poþclonal B cell activation secondary to the pronouuced tropism of EBV virus for B
llm.phocytes. The CR2 (Cd3) receptor on B cells is thought to act as a receptor for EBV (Allday &
Crawford" 1988). IgGl (Kascha et al5 L982) and IgG4 (Shacks et a[ 1985) increase during infectious
mononucleosis. Deficiencies of IgG subclass that predated the onset of EBV infection have been found in
Duncan's slmdrome. This may reflect the inability to produce antibodies to EBV nuclear antþens (Purtilo
et at 1989) and may be an important factor in the exquisite susceptibility of these patients to EBV
infection.
In patients who have had bone marrow transplantation, many months after bone marrow
transplantation, Ig,\ IgG2 and IgG4 concenbations may be low with high s¡ normal concentrations of IgM,
IgGl and IgG3 many months later (Aucouturier et 4 1987b).
As can be seen from Table 1.8, many of the studies of IgG subclass concent¡ations in patients with
immunodeficiency states have used assays and normal rânges which were not capable of detecting IgG4
deficiency. With the advent of more sensitive systems, there is potential for confüming and eÉending these
studies. Chapter 10 begins fq ds this.
1.9. IMMT]NOGLOBT]LINREPLACEMENTTIIERAPY
Although imm¡neglsþrlin ¡spþcement therapy has been used to treat h¡pogammaglobrlinaemic
patients for nearly 40 years, and IgG subclass deficiencies were being reported in some patients with
recurrent infections by the early 1970s (Tet y, 1968; Schur et als 1970; Oxelius, L974), there is still much
uncertainty surrounding the use of immunoglobulin replacement therapy in IgG subclass-deficient patients.
A few studies and case reports have been published (Silk and Geh4 1987; Beard and Ferrante, 1988;
Bjorkander et at 1988; Heiner et al 1988; Beard and Ferrante, 1990) and these indicate that the use of

B2
immunoglobulin replacement in some infection-prone IgG subclass-deficient patients is probably
woflwhile. Little is known about most effective immunoglobulin preparation to use in such patients, or
the opfimum dosage or frequency of administration of the immunoglobulin preparations.
1.9.1 Intravenous immunoglobulin preparations
Immunoglobulin preparations for intramuscular use have been prepared by the Cohn ethanol
fractionation of human plasma (Cohn et a[ 1950) for nearly 40 years. The dosages of these preparations
that can be given are limited by volume constraints. The recommeuded minim¿l dose for use in
agammaglobulinaemia has been 1ü) mg/kgbody weight per mont\ which would be a volume of about 50
ml of a L6Vo pteparation per month in an adult. Larger volumes are clearly unacceptable, but may often be
necessary for optimal prophylaxis against infection. Int¡amuscular imnunoglobrlin preparations are not
suitable for int¡avenous lrrie, because although they contain mainly monomeric IgG, they also contain
variable amounts of di- and polymeric IgG (Barandun et al, f962) and such immunsglsþulin aggregates
may car¡se serious anaphylactic-type reactions by spontaneously activating complement (Skvaril & Gardi,
1e88).
A variety of procedures has been developed to modify imm¡¡egloþrlin preparations derived from
human plasma by cold ethanol fractionation in order to make them safe and suitable for intravenous
administration. The ethanol fractionation of plasma has been shoum to inactivate HfV vi¡us (Wells et al
L98ó). The ideal int¡avenous immunoglobulin ([VIG) has not yet been produced, though considerable
progress has been made. The requirements of an IVIG preparation have been formulated by the rÙVorld
Health Organization in conjunction with the General Immunology Committee of the International Union
of Tmmunological Societies (Cunningham-Rundles et al5 L982). In such a preparation:-
(a) IgG should consist of at least X)Vo morrcmeric IgG molecules, po\mers should rLot exened SVo,
and without immunoglobulin fr agments.
The 4IgG subclasses should all be present, and in proportions simil¿¡ to those in normal plasma.
IgG Fc effector function should be normal, particularly conplement activation and promotion of
phagocytosis.
(b)
(c)

(d)83
Antibody levels against at least 2 bacterial species (or toxins) and 2 viruses should be ascertained
by neutralization in tle case of viruses and additionally, at least 0.1J.U. per ml of antibody to
hepatitis B, m4 by RId a tit¡e of at least 11000 to hepatitis A virus should be present.
The biological halflife of the IgG isot5pes should be adequate.(")
1.9.1.1 Methods of Preparation
The procedure fi¡st employed to reduce complement binding was the proteolytic cleavage of all
antibodies by enz5rmes eg plasmin or pepsin. Pepsin destroys virtually all intact immunogloburin molecules,
while leaving most of the antþen-binding capacity of the F(ab')2 fragments, as rlimers and pollm.ers, intact.
(Skvaril & Ga¡di 1-988). Such preparations were well tolerated and could as¡¡'aliss toxins, but functions
that depended on the Fc part of the molecules, were largely deshoyed (Fig. 1.4) Plasmin splits the IgG
molecule into Fab and Fc fragments, but leaves a percentage of it intact (More[ 19Só) (Fig. 1.4).
Subsequently, immunoglobulin preparations were treated with various chemicals either to prevent
spontaneous aggregation or to prevent any aggregates present from activating complemeut. Procedures
employed included beta-propiolactone treatment, reduction and alþlation, and sulphitolysis. Such
procedures, also, are associated with a considerable reduction in biological activity particularly due to
gfanges in the Fc part of the imnunoglsþ,,lin molecules (Romer et al5 L982; Skvaril and Gardi, 1983) (Fig.
L.4).
Recentl¡ attempts have been made to try and preserve the physiologicat structu¡e and function of
antibodies and to prevent the formation of IgG aggregates during plasma fractionation. Adding stabilisors
such as polyethylene glycol inhibits aggregation and precipitates aggregates, but the formation of po$mers
cannot be completely prevented (Baranduq 1985; Morell, 1936).
T'activation of the complement-binding capacity of pre-formed IgG aggregates can be acåieved by
mild acid trearment, at pH4, without altering the function of monomeric molecules @arandun, 1985).
Addition of traces of pepsin (110,000) eliminates pollrmeric aggregates and prevents re-aggregation of
monomers and dimers but is thought to be insufficient to cleave peptide bonds in the native monomeric
molecule (Painter & Law, 198Ð (Fig. 1.4).

84
I
-PEPS|N+¡-
F(ab')2 + loss ot Fc
2
-PLASM|N-+
some rnlåcl
\
2 some splll rnlo
2Fab + rFc
3
fJ oroÞrolactone
Fc and Fab modrlied
1
REDUCTIOÀI T ALKYLAÍION
hydrolys6 of disulph¡de bídgesbetween polyDepl¡de cha¡ns
5
ION EXCHANGE CHROMAf OGRAPHY
o. pH{ TREATMENÍ 3 TRACES OF
PEPSIN
- inlaCt molecules
Fig. 1.4 Schemadc rfepresentadon of the efi€cts of prcparafron procedunes on lmmunoglobulin for lntravenous use.

85Ultrafiltration and ion s¡sfuange absorption of immunegloSulin isolates monomeric human IgG
and eliminates aggregates. The addition of albrmin or sugars prevents re-aggregation (Morell 1986) (Fig.
r.q.
L.9.12 IgG subclass composition
The use ef imm¡¡oglobulin replacement therapy in symptomatic IgG subclass-deficient patients
has not yet been universally accepted. An immunoglobulin preparation to be used would presumably,
need to contain adequate amounts of the subclass (es) in which the recipient was deficient. The
proportions of the IgG subclass in most preparations for intraveuouÍi use are considered to be near to
physiological.Ilowever, some prepilations have been found to be lacking in IgG3 and others low in IgG4
(Morelt .1r9f36;
Skva¡il & Gardi" l-988). Chromatographically prepared preparations have low IgG4
contents. Both IgA and IgG4 are acidic proteins and bind to anion-exchangers (eg. DEAE-exchangers)
(Skvaril & Morell, 1970). A[ rirnes, the composition of a particular fVIG preparation apears to have
shanged over a period of years (Morell, 1,98ó). It is, therefore, important that if an IgG subclass-deficient
patient is to be treated with an Ig preparation, the IgG subclass composition of that particular preparation
is known.
The comparison of IgG subcl¡ss concentrations in different IgG preparations has some limitations.
Discernible differences may be due to an alteration of subclass-specific determinants on tle molecules
(which may or may not be associated with changes in biological activiry) or to tle presenee of fraenûents
bearing subclass specific determinants (Beck & Kaiser, 1981b). In general enz5rmatically treated and
nonenz¡matically treated preparations have simila¡ subclass proportions indicating that the presence of IgG
fragmsa¡s is not a major çe¡fls¡¡rling va¡iable. IgGl and lgp2 appear to be most suspectible to
conformational cþanges leacli'g to changes in binding characteristics (Beck & Kaiser, L981b).
1.9.13 Antibodycontent
Although, logical¡ replacement of a missing IgG subclass might see¡ to be appropriate t¡eatment
for an IgG subclass deficiency, in fact, the situation may be more complex than this since some subjects
with a deficiency of one IgG subclass have been found to have an impaired ability to produce some'specifiC
antibodies of other IgG subclass isotlpes also (section 1.6.1). It is generally agreed that a deficiency of
antibody to particular pathogens is the important factor, rather than the level of any subclass per se. Itß

86
important that the IgG preparations used to treat IgG subclass-deficient patients have adequate levels of
antibodies to the pathogens to which those patients a¡e most susceptible. It is, of oourse, not possible to
measlue levels of antibodies, to all such pathogens but levels of a variety of antibodies to protein and
polysaccharide antigens can be dets¡minsd.
There are limil¿1isns inherent in assaþg IVIG preparations for specific antibody conteuts. The
presence of an antibody detected imm¡¡slsgically does not prove it is biologically intact. In a study of 5 Ig
preparations, Emm¿¡sel (1988) found no predictable relationship between the quantity of immunoreactive
CMV-specific antibody by ELISA and the in vitro functional activity as measured by neutralisation, but
noted also that there is no data i¡ þrmans to show direct correlation between an antibody's in vitro
neutralisation capacity and its possible effectiveness as a CMV prophylactic in vivo. This study also showed
considerable batch to batch reaction in some of the preparations tested.
Bender and Hetherington (198Q assessed IgG-dependent interaction between phagocytes and
Haemophilus influenzae type b (Hib) in different fVIG preparations using a lumnol-enhanced neutrophil
chemiluminescence method, and also measu¡ed antibodies to Hib and to PRP in these preparations.
Differences between opsonic capacity in the preparations studied did not correlate dite"tly with the
concentrations of either of tlese antibodies, as measu¡ed by ELISA and RIA respectively. Opsonic activity
was greatest in the preparation prepared by ion-exchange chromatograph¡ despite antibody titres being
comparable with those in treated IVIG and in heat-inactivated serum. The possibility of differences being
due to a di.fference in the subclass composition of the antibodies in the preparations was raised but as
antibody to Hib is predominantly lgG2 and,/or IgGl and the preparations tested were considered (from
previous studies) to have simil¿¡ IgGl and IgG2 proportions, this was thought ¡s þe nnlikely. Previous
studies showed 5imila¡'ly, that for S. pnuemonise alid other bacteria, ion-exchange chromatography was
apparently the most effective method for preserving opsonic activity.
Yasuda et al (L986) found that a pH4 treated preparation and a polyethylene glycol treated
preparation were superior to both sulfonated and pepsin-treated preparations in terms of opsonisation of
Eschiríchiø coli and of. Pseudomonøs aeruginosa., even when allowanoe was made for the generally lower
concentrations in the latter two preparations of antibodies to the.E colí and P. aeruginosa.

87
Van Fu¡th et al (L98a) showed pronounced differences in opsonic activity of different fVIG
preparations for different bacteria. Concentration of the immunoglobulin was not adjusted u"so¡ding to
specific antibody titre for the organism concerned but rather by equalising IgG concentratioq so results of
rhis s¡udy are not immsfi¿tsly comparable with those of Yasuda et al.
1.9.1,4 Complement activation
While IVIG for infi¡sion must be free of spontaneous anticomplementary activity which may cause
dangerous reactions, once infi¡sed, the antibodies should be able to bind antigen and to activate
complement. These criteria a¡e met in ion-exhange chromatography and pH4 treated preparations
(Morell, 1986). Measuring CH50 levels before and after IVIG infusion may give an indication of
anticomplementary activity, but needs interpreting with some caution as a fall in complement levels during
infi¡sion may also, possibly, reflect a therapautic effect.
1.92 lhe use of immunoglobulin replacement IgG subclass-defrcient children
Intramuscular immunoglobulin (tr\rflG) has been used to treat hypogammaglobnlinaemia with a
degree of sucess for nearly 40 years. More recently IVIG has been shown to be equally effective, or, in
larger doses, more effective (Ammann et aJ, L982; Cunningham Rundles et a[ 1984; Leen et al, 198Ð.
There appears to be wide variation in imms¡sglobulin concentrations from patient to patient, even when a
uniform dose of immunoglobr¡lin has been administered at a uniform rate (Leen et al 1-985). There have,
however, been relatively few reports of the treatment of isolated IgG subclass deficiencies with
imnunoglobrlin replacement. Schur et al (1970) reported two children and an adult with different
combinations of IgG subclass deficiencies and life-long susceptibility to infections who were treated with
IMIG. Oxelius (1974) reported a fanily with IgG2-IgGa deficiency, and 4 patients with IgA-IgG2
deficiency (Oxelius et at L981) who responded well to IMIG. Oxelius (1984) reported one patient with
systemic þus erythematosus, recurÌent pericarditis and infections, and a subclass deficiency who also
improved with Ig prophylaxis. Bass et al (1983) described an IgG2-deficient infant who suffered 2 episodes
sf msningoooccal meningitis prior to being given IMIG prophylaxis which proved effective. Plebani et al
(198Ð described an 11-year-old with recurrent infections and bronchiectasis who had a deficiency of.lgp2
and of specific IgG and IgM antibodies who improved on 100 mg/kg/week of subcutaneous Ig. Serum
IgG2 normalised and IgG tetanus toxoid antibodies rose significantly. Bjorkander et al (1985) described 3

88
lgÃ-lgG}-deficient patieuts with respiratory infections who benefited from imm¡qogl¡þrlin replacement
therapy.
Such studies indicate that immunoglobulin replacement is probably worthwhile in symptomatic
IgG subclass deficiency and that IMIG may, ¿[ rimss, be effective. By analogy with the situation in other
hlpogammaglobrrlinaemic states, IVIG would be elçected to be even more effective. IMIG is more
convenient, less expensive and less likely to be associated with systemic side-effects than is IVIG, and it has
not been associated with the t¡ansmission of non-A non-B hepatitis as has IVIG, albeit very rarely (Lever et
al, 1984). Some contend that IMIG is safe¡ in lgA-deficient patients who may have IgA antibodies, but
such patients may react to either form (Buckley, 198). IgA-depleted IVIG preparations have a place in
these patients (Bjorkander et a1198Ð.,
IJnansq/e¡sfl questions with regard to the use of immuLnoglobrlin ¡epl¿ssment therapy in IgG
subclass-deficient patients include:-
Which IgG subclass-deficient patients warrant immunoglobrlin fþs¡¿py!
Is the IgG subclass composition of the immunoglobrrlin preparation a critical factor?
Does the IgG subclass composition of an immuuroglobulin give any indication of its antibody
content, or should preparations be tested for specific antibody content?
Should one ai- to maintain semm IgG subclass concent¡ations in the normal range for age?
Should the amount of immunoglqþrtlin administered depend on the severity of the IgG subclass
deficiency or should it be determined by the patient's ability to produce specific antibodies?
These questions will be addressed by the studies described in Chapter 12.
1.
)
3.
4.
5.

CHAPTER TWO
GENERAL METHODS

90
2.1 IMMUNOGLOBULIN QUANTITATTON
2.1.1 Serum lgd IgG and IgM
The th¡ee major immunoglobulin isotypes Ig.\ IgG and IgM were measured by rate nephelometry
using a BeckmanAuto ICS machine (Beckman, Palo Alto, California, U.S.A..). This system depends on the
formation of insoluble complexes of antibody and antigen and the me¿ìsurement of light scatter by these
fine particles. If there is an excess of either antþen or antibody soluble complexes, no scatter light scatter
occurs. Where concentrations were too low to give reliable est"nations by this metlod, the
imm¡¡sglsþulins were measured by low concentration radial immr¡nsdiffusion (RID) plates, LC-Partþn-
IgA (Behringwerke AG, Marburg, W. Germany) so¡taining heavy chain specific antisera in agarose.
Concentrations of immunoglobrrlins were determined by comparing the diameter of the precipitation rings
formed when the immunoglobulin lgÇ diffused in the agarose with those of standards provided with the
plates.
Normal ranges for IgA, IgG and IgM were taken from Notes on Diag¡osis, number 2, "Tmm¡¡s
deficiencies in the Infant", Serological Department, Hoechst Pharmacentica based on "Les Carences
Immunitaires de I'enfant", Ed. J.P. Harpey (1977). These ranges which were based on RID studies were
converted to ranges for the Beckman ICS by studies in the Jewish Hospital in New Yorþ and provided by
Beckman. Adjustments were incorporated based on concentrations in Australian child¡en (Shelton et al,
1974). The rânges representt 2SD and are shown in Table 2.1.
Total serum IgG concentrations were double checked by using an ELISA assay (described in detail
in Chapter 3). I" a few instances, where IgA was undetectable by RID or nephelometry, an ELISA assay
like that used to check total IgG was used for IgA.
2.12 SalÍvary IgA
Saliva was microcentrifuged immediately after collection and an aliquot removed from the
supernatant. Using low concentration RID plates (Behringwerke AG, Marburg, W. Germany) and

9ITable 2.1
NORMAL RANGES FOR SERIIM' IgA IgG AND IgM
t*(e/r) rgc (g/t) telvt (e/t)
Newborn1- month2uôtrJ
+6"7-9',t0-12'
1 year2u^ltJ
4u5u
6u7n8u9u10"
00.040.Ít0.150.L70.190.22
0.540.580.630.690.75
0.780.800.820.860.86
5.¿m
4.543.242.ß2.162.ß3.m
3.894.324.æ4.975.40
5.836.056.166.266.37
6.376.,18
6.486.486.¿l{l
18.36t6.?r8.646.4{l6.708.4210.15
10.80t2.5Lt6.6114.5875.12
15.7716.2416.38L6.65L7.Ot
t7.r0t7.LOL7.28fi.?aL7.28
00.110.18o.2L0.210.n0.31
0.330.380.390.420.43
0.40.450.450.60.6
0.60.Mo.M0.60.47
0.380.470.750.91.09L.22L.32
L.321.55
1.ilr.671.69
L.72r.731.74L.761.78
0.26o.4t0.rA0.650.820.99t.t4
0.240.300.350.430.4{l
1.21t.732.t62.552.8t
3.LL3.373.633.894.t0
4.324.544.754.n5.18
11
72liL415
il
I
t.791.80L.831.851.88

92
comparing the results with those obtained using dilutions of standard sera" the concentrations of IgA in
saliva samples were able to be calculated. Values below 0.û2 gflwere considered low.
2.I.3 Serum IgE
Serum IgE was measured by a solid phase radioimmunoassay using an IgE RIACT kit (Phadebas
PRIST, Pharmacia, Sweden). Monoclonal anti-IgE, covalently coupled to the test tube *"11 and 125¡-
labelled monoclonal anti-IgE react sinultaneously with IgE i" the test sample forming a multisite compler
After incubation, unbound p51-labe[ed IgE is washed away and the radioactivity remaining is directly
proportional to the concent¡ation of IgE antibodies in the sample (Kjellmaa NM et al L976). Normal
rânges *:d fî IgE were tlose of Bhalla and Kjellman et al (1-976). The upper limi¡s s¡ these rânges are
shown nTable2.2.
2.L,4 Antibody quantitation
Antibodies to pneumococcal polysaccharides were quantitated in the Microbiolog5l Department of
the Adelaide Children's Hospital by a radioimmunoassay (Paton et al, 1-986). Antibodies titers to tetanus
toxoid were measinred by haemagglutanation and to diphtheria toxoid by en4me-linked immunosorbent
assay in the sa-e department.
2.2 CELLIJIAR STT]DIES
22.L Preparation of leucocytes
Mononuclear leucocytes and neut¡ophils were separated simultaneously from 5-10mt of
heparinized blood by the rapid one-step method (Ferrante & Thong L978). The procedure uses the
principle of density gradient centrifugation and erythrocyte aggregation. Ficoll 400, a high molecular
weight sucrose pol1m.er, is used as the erythrocyte aggregating agent. The Hlpaque is used to obtain a high
density with a relatively low viscosity. The density of the medium is 1.11-4 which is much hþher than
conventional llmphocyte separation medium (usually d=1.077). The higher density enables the retention
of neutrophils which a¡e of much hþher density than llmphocytes and monocytes. Briefly, li¡þirrm-

Table 22 93IsE REFERENCE VALIIES FOR CHILDREN
Age14
yrs
06369t234710
days wks mths mths mths yrs yrs yrs yrs yrs yrs
Geometric mean1.3L95U/ml + 2SD
6.L 3.8 L6.3 73 15.2 295 16.9 68.9 L6L 116
wks = weeks, mths = months, yrs = years.

94
heparinised blood is layered on a separation medium consisting of 20ml of 85Vo hlpaque (?Å.33Vo sodium
3,5 diacetemido -2, 4, 6- trüodobenzoate and, 56,6Vo N-methyþucamine 3,5-diacetamid6 -2, 4, 6-
träodobenzoate) and 90ml of L0% (w/v) ficoll (MW,m0,000). The blood is then centrifuged (swing out
buckets) at 4009 for 30 min at room temperature. After centrifugation the leucocytes become resolved into
two distinct bands (see Fig. 2.1). The top þ¿¡d so¡feins mononuclear cells (lymphocytes) and the second
þ¿¡d highly pure neutrophils. These are harvested carefullS washed and resuspended in tissue culture
medium or flank's balanced salt solution (HBSS).
22,2 Neutrophil studies
222.1 Neutrophil chenotaxis and random mobility
Neutrophil random mobility and chemotactic responsiveness were measured by the migration
under agarose method (Nelson ef. alo L975), with sligbt modifications (Thoog, et al 1978a; Ferrante et al
1980). N-formyl-L-methionyl-Lleucyl-Lphenylalanine (FMLP) was the chemoatt¡actant used. Agarose
plates were prepared by mixing 3ml of 2Vo agarcse solution with 3ml of 2 x normal medium 199
supplemented with lÛVoheat-nactivated foetal calf serum. Sets of three wells of 2.5mm diameter were cut
on a straight axis 2.5mm apart, measured edge to edge, with the aid of a template. To the centre well $l of
4x107 neutrophil/nl (2*1d) neutrophil suspension was added. To the outermost well ful of 1x107 FMLP
was added. To the innermost well was added Srl of control medium (DMSO diluted h 199) to the same
arnount as the FMLP. The plates were incubated at 37oC for 1.5 hou¡s in a 5Vo CO2 in higlh humidity
ahosphere. The distances travelled by the ten fastest-moving neutrophfü towards the outer well
(neutrophil chemotaxis) and the inner well (random mobility) s/as measured with the aid of an eyepiece
grid on an inverted phase contrast microscope (Nikon). The rate of mþation towards the chemotactic
agent was considered to be the chemotactic mþation. For each individual, 2 sets of 2 wells and 2 plates
were used.
2222 Quantitative leucocyte iorlination reaction
The quantitative leucocyte iodination test (Pincus & Klebanoff, 1971) modified to a semi-
automated microassay procedure (Thoog & Ferrante, 1978) was used. Neut¡ophils ¡ys¡s resuspended in
I{BSS. Dilutions of. L/4AB serum or autologous plasma were made. To eac;h, 2fu1of.7251v¡ ?fpø3i/nl
was added. To wells 1-6 of microtitre plates 5Qrl of 125I PHS was added to wells 7-72, 8ù o1 125¡

95
<- Blood Plasma
Centrif ug ation MNL
200 9 /20min <-- PMNL
-- Ficoll-Diatrizoate <- Erythrocytes
S
Fig.2.1 Fracdonaûon of blood leucocytes into the main populations by the rapid singls'sþp method.

96autologous plasma, to wells 1--3 and 7-9, 1:CQuL of HBSS and to wells 4-6 and 1-0-12, 10Qpl of z5mrosan
(zSmg/ñin HBSS). To all the wells 0.1m1 of neutrophil suspension (1xL07/mD were added. The plates
were incubated for 90 minutes at 37oC in a humidified 5% COy-atr atmosphere. The cells were harvested
onto glass-fibre filter paper with a Skat¡on cell harvester. The disç5 v,/e¡e ¡'ansferred to tubes for countirg
in an automatic gamma coutter. The amoun¡ o¡ 1ã1 incorporated was determined and the results
e4pressed as follows:
ØI uptake com in cells x 8.3 x 10
þ moËs/107 neutrophils) t"trt.p- "dd"d
where 8.3 equals the amount of iodi"e added in p moles, and l-0 represents
adjustment to 107 ûeutrophils.
2223 Neutrophil bactericidal and fungicidat activity
Methods previously described (Thoog et ú, L977; Thong et aJ, L979) were used to assay for
neutrophil bactericidal and fungicidal activity. For the bactericidal assa¡ 10Qrl of 1x107 S. aureusfml,
(1x10ó) Stuphylococcus aureus (strain NCTC 6571-) organisms were mixed with 25øl of 4x107
neutrophils/ml (1x106) neutrophils in the presence of 4qfrl of 5Vo hwanAB serum (a source of opsonins)
and made up to a total volume of 0.5m1 in HBSS in screw-capped plastic tubes. The cells were gassed with
5Vo Cþ-atr mixture and incubated at 37oC with end to end mixing on a rotating shaker. gamples were
taken at 0, L and 2 hou¡s diluted and plated on to blood agar to determine tÏe number of viable bacteria.
The results were expressed as Vobactena kille{
Vo kldled. = 0 fime counts-courts at 1 or 2b¡0 fime counts
For the fungicidal assa¡ neutrophils were tested for ability tokilToruIopsis glabrøta (a clinical isolate) by a
similar procedure. The only differences were in the sampling fimes (Q, 1.5 and 3 hours) and plating on to
Sabouraud agar.
222.4 Neutrophil respiratory burst
NADPH is produced during oxidation of glucose by the hexosemonophosphate (IilvfP) shunt in
cells. HMP shunt activity is therefore closely linked to NADPH oxidase activity. We assessed this by
quantitati"g ¡1-1aC¡-l glucose oxidation by measuring the release of radioactive Cû2 fo[owing stimulation
of cells with opsonized zymosan. Briefly a ?Ãml and a 5ml scintillation vial were used to const¡uct a

97container with an inner and an outer vessel. The outer vessel received 2rú06 PMNLs, O.OUpCiof t1-1ae-
Lglucose (3.96 mCi/mmol) and 4Vo htrnan AB serum, with or without 0.4mg of opsonised z5mosan. All
solutions were made up in glucose-free Ea¡le's BSS and the final volume in each outer vessel was 1.0m1. To
the inner vesssl Q.1ml of 5N NaOH was added to absorb the L4co2evolved. After incubation for 90 min in
a shaking water batl at 37oC, the radioactivity in the NaOH was counted by liquid scintillation (Ferrante
and Rencis, 1984).
2.2.3 LymphocyteStudies
223.L Lymphocyûe phenotyping
After purification of þmphocytes, in early studies, T cells were enumerated by spontaneous rosette
formation with sheep red cells (Scheinberg et al L976). B cells were measured by immunofluorescence
using goat anti-human immunsglsþrlin (Alexander & Sanders,L977).
Later, þmphocyte subpopulations were determined by indirect immunofluorescence and flow
cytometry r¡sing monoclonal antibodies (Ferrante et al 1-987). The monoclonal antibodies used were OKT3
(anti-CD3), OKT4 (anti-CD4), OKTS (anti-CD8) (Ortho. Diagnostic Systems, Inc. NJ.), FMC1 (which
recognises all B lymphocytes was a gift from Dr H. 7-nla, Department of Immunology, Flinders Medical
Center, South Aust¡aliq Brooks et al L983), and Iæu11, (anti-CD1,6) and LeuT (anti-CD57) (which react
with natu¡al killer cells, Becton Dickinson, Sunnyvale, CA' USA). After mononuclear leucocytes (À/NL)
were separated as outlined previously and any remaining red cells lysed by hypotonic lysis, 1x106 cells were
added to each tube and made up to 10Qrl nPBS+2Vo D-glucose (*/Ð. Then" 5r¿l of monoclonal antibody
was added to each and incubated at 4oC for 30 minutes. The cells were washed in PBS, and\tl of neat
autologous plasma followed by 5Qrl of FTTCJabelled goat anti-mouse IgG (Cappel, Malvern, Pa" U.S.A.)
diluted l/LO ta PBS was mixed with each and they were incubated a further 30 minutes at 4oC. After
w¿shingwith PBS 2@rl of L% (w/v) paraformaldehyde in PBS (pH 7.2-7.4) was added and the cells were
then kept in the da¡k until analysed, within one week b¡ indirect immunofluorescence and flow cytometry
using a FACS fV instrrrment (Becton þickinssn, Sunnyvale, Calif. U.S.A'). Fluorescenoe was excited at
ffinm and emission detected after eliminating scattered light with 520- and 5ï)-nm filters.

98
2232 Ll,mphocyte transformation to mitogens
Lymphocyte transformation to mitogens were measu¡ed by a radiometric microculture technique
(Thong & Ferrante, 1979). The mitogens used were ph¡ohaemagglutinin, PIIA (Wellcome, Sydney,
Australia), pokeweed mitogeq PWM (Gibco, Grand Island, N.Y.) and concan¿yalin { ConA
(Calbiochem, San Diego, Calif.) at concent¡ations of tWg/ú a L7o solution of lyophilised powder and
25p,g/mlrespectively. To 10Qrl of ?-rJ06 MNL per ml 10Qrl of PFIA, PWM or ConA was added made up
in RPMI Iffi + SVohvmangroupAB serum.
The cells were cultu¡ed for 72hr at 37oC n 5Vo CÛy-atr ¿¡d high humidity. Cultu¡es were pulsed
with þCi per well of 3g-tnf'miaine 13ff-fAn¡ (Amersham, Sydney, Australia) and aspirated onto glass
fibre filters using a cell harvester (Flow Laboratories, Sydne¡ Australia). fhe 3g-tdn uptake was
measured by liquid scintillation spectroscopy.
Both the dpm for tle non-s;mulated and mitogen-stimulated cells were recorded and the
stimulation index (SI) calculated:
dpm in presence of mitogenSI=
dpm in absence of mitogen
2233 In vito immunoglobulin production
In vitro production sf imm¡nsglsþrrlins by llmphocytes wÍrs carried out essentially as described
previously (Ferrante et at L98a). MNL were suspended in RPMI 1640 with FCS (10%) to a concentration
of md/mt and cultr¡red in LIIX tissue culture tubes, 16 by 125-m, (Miles Laboratories, Grand Islan{
NY) and stimulated with pokeweed mitogen at selected concentrations. The cultu¡es were incubated at
37oC in an atrnospherc of SVo CO2-95% ai¡ and high humidig, and their supernatants collected after 7
days of incubation. The supernatants were exanined for levels of IgG subclasses by the enz5rmeJinked
immunosorbent assay (see chapter 3). $imil¿¡ly, MNLs were cultured with Støphylococcus øtreus at
selected ratios of ^S.
øtteus to MNIs.

99
223.4 NK cell cytotoxicity
NK cell cytotoxicity was measured by a microcytotoxicity assay (Ferrante et al, 1-985) in which
mononuclear leucocytes were mixed with 51 CrJabelled target cells of hrman erythroid myeloid cell line
(K562) at a variety of effector: target ratios and damage to cells was measured by the relsase of 51Cr.
Briefly, the target cells were freshly cultu¡ed from cryopreserved semples in RPMI 1640 medium (RPMI)
supplemented with 5-I0Vo heat-inactivated fetal calf seru- (FCS). The cells were labelled with the isotope
51Cr by l¡çuþ¿ting 5x1-0ó ce[s/ml RMPI with 10qr Ci of Na2 51Cr04 for t hou¡ at 37oC.
The assay was carried out usi'g microtit¡e plates (Linbro, Flow Laboratories). To each well was
added 10Qrl of 51-Crlabelled target cells (1tm4 ce[s) and 10Qrl of MNL suspension to give effector:target
cell ratios of 1:1, 2:I,5;I, 10:1, 50:l- and 100:L. The cytotoxicity reaction was performed at 37oC in an
atmosphere o15% COz-aü ¿¡d high humidity. The supernatant fluids were collected after 4 hours using a
Titertek supernatant harvester (Flow Laboratories) and counted in an auto-gamma-counter. Assays were
conducted in triplicate and the degree of cytotoxicity calculated as follows:
sLCr rcLease (Vo) counts/min supernatant x 1(X)
counts/min ls1al
Lysß (%) cytotoxicity (%) = Itest release -sDontaneous release)(maximum release - spontaneous release)
It has previously been shown that if the percentage cytotoxicity is plotted against effector cell
numbers then the curye so generated fits the general equation y : CI (1- p Ð, in which x is the effeetor cell
number, y is the percentage cytotoxicity and o and p are constants which can be computed by a nonlinear
least squares technique (Ferrante et al 198ó). The limiting value of the curve is o, and p is related to the
rate at which the curve approaches a. Generall¡ curves with smaller p value approach the asymptote for
smaller values of x. Thus, given suffrcient knoum values of percentage cytotoxicity and effector cell numbers
is possible to compute cytotoxicity for the entire range as well as determine maximal lysis achieved (Fig.
2.2).
Control values are: LD25t ZSD = 75-22Vo; C-a*t 2SD : 39-88Vo.

100
y=a(t-pt)
o
v
X
ß1g,,22 Graph of the mathemadcal funcüon to qùicù NK cytotoxictty d¡ts ls ñtÛed. The unloown
¡rarametcrs to be computed are o and p . The limtting value that the curve approaches ls c , p ls rclated to
the rate that the curve apPnoachesa (Femante et al 1986).

10122.4 Controls for cellular studies
Controls for lymphocyte and neutrophil function studies have been healthy South Australian
adults. This has been necessary as it has not been possible, for practical and ethical reasons, to have age-
matched control children available for bleerling on most of the occasions when tlese studies have been run.
There appear to be no significant differences in levels and proportions of CD3+, CD4+ and CD8+ cells of
7 year old children and adults (Falcao et al 1987) and previous studies in ou¡ laboratory have shown that
there is no sig¡ificant difference between 7 and 8 year old children and adults in proportions of B cells,
Leull-+ cells, responses to PHA' PWM and ConA" and NK cell cytotoxicity (Ferrante, r'''published).
2.3 COMPLEMENT
23.1. Total haemolytic complement
Blood samples were processed usually within t hour of blood drawing and the serum stored at
-70oC. The complement assay was conducted within L week. The functional activity of the classical
complement pathway was measured by a haemolytic technique, (Mayer, 1961). Sheep red blood cells
(SRBC) sensitised with haemolysi" (IgM rabbit antibodies to SRBC) were mixed with dilutions from 1/8 to
L/12ß of. serum in isotonic veronal buffer. Non-sensitised red cells were used for the controlþlank in tubes
and incubated at 37oC for 30 minutes. The tubes were vortexed atll,?Ã and 30 minutes. After incubation,
they were centrifuged in the cold at 15009 for 10 minutes. Then, 1@rl of supernatant was mixed with 1..5m1
of distilled water and the absorbance read at 5,l0nm in the spectrophotometer. Measu¡ements are most
sensitive at5ÙVo haemolysis and results are expressed as CH5g units.
Calculations
1-. y = Vo lysis =
,,1
OD<¡n - blankOD540 1007o lysis - blank
expressed as adecimal figure
2. JL-y
Plot curve y vs recþrocal of dilution on log/log graph paper.I-y
When ¡ intercept is 1, fhis is equal to SOVo haemolysis.L-y
Total Haemolysis (CH5g units¡ = 2x5OVo haemolysis units.

to2
Results r¡re e)ipressed, as % of pooled human AB blood group serum.
The normal ¡ange for CH5g activity ß 65-ß5%, except in young i¡fants where values may be
slightly lower.
2.3.2 Individual complement components
Quantities of complement components C3 and C4 were measursd by nephelometry (ICS,
Beckman, Palo Alto, California, USA). The normal ¡angos are:- C3, 0.76-L.76g/l; C4,0.L+0.28g/l and
values do not vary signifiç¿¡¡1y with age.
Other complement components were detected by supplementation studies adtli"g patients' sera to
sera known to be lacking in certain individual complement components. Deficiencies were confirmed by
immunological methods by the Scripps Clinic and Resea¡ch Foundation, La Joll4 California, or by the
Institute of Medical Microbiology, University of Lund, Sweden (Lachman & Hobart, 1978; Truedson et at
1981).
2.4 STATISTICAL ANALYSES
Student's t test was used to oompare results of patients and controls in many of the studies where
relatively small numbers of patients were being studied. The y2 analysis was used for comparison of
discrete variables between samples where numbers permitted and where numbers were too sma[ the
Fisher exact probability test was used. ffis \¿[¡nn Whitney U test was used for comparisons of gtoups of
samples of unpaired and possibly unrelated data (e.g. salivary and serum IgG subclasses).

CHAPTTR THREE
THE DEVELOPMENT OF AN ENZYME.LINKED IMMUNOSORBENT
ASSAY USING MONOCLONALANTIBODIES FOR THE
QUANTTTATION OF IgG SUBCIASSES

104
3.L RATIONALE FOR DEVELOPING AI\ ELISA TECHNIQT]E FOR IgGsuBcrÁs s QUAI\TITATION
If measurements of IgG subclass concentration a¡e to be useful, accurate and reproducible assays
must be developed for their quantitation. Ability to separate low, but normal, f¡om subnormal
concentrations of subclasses is also essential. This is particularly important in the case of IgG4
measurements where undetectable concentrations have been found in as many as 25% of normal
individuals and in t2-2lVo of children aged over 7 yearc u5ing electroimmunoassay (EIA) or radial
immr¡nsdiffusion (RID) ¿sghniques (Oxelius, 1979a).
Today the most commonly used assays for IgG subclass quantitation are RID, radioimmunoassay
(RIA) and enzymeJinked immunosorbent assay (ELISA). The previously used EIA has generally been
replaced by RID. This is a relatively simple technique and can be done using either poþclonal antisera or
monoclonal antisera (Schur et all979; Lowe et al1982; Jefferis et al 1985; French and Harrison, 1984b).
Commercial kits for measuring IgG subclasses are available, some with polyclonal and some with
monoclonal antisera.
We have found that ¡¡p ¡echniques for IgG subclass measurements have serious limitations. Our
experience using RID plates prepared in our laboratory with Netherlands Red Cross T¡ansfusion Service
polyclonal antisera showed difEculties in the measu¡ement of IgGZ and IgG4 because the IgG2 precipitin
rings were hazy and not clearly distinguishable, and the IgG4 rings were too close to the edge of the wells
for accu¡ate ¡s¿ding. Low concentrations of IgG4 cor¡ld not be measu¡ed. A sirnila¡' observation was made
when we used com-ercial RID plates from 2 separate sources, ICN-Biomedicals (Irvine, CA, U.S"A..) and
Binding site (Birmingham, England). Because of the relatively low IgG4 concentrations in many normal
children a more sensitive method of IgG4 measurement to separate the normal from the abnormal was
required. ELISA techniques can measure concent¡ations down to l-ug/ml compared with the 0.01 g/l of
most RID assays.
The sensitivity of the ELISA technique makes it also suitable for measuring IgG subclass
concentrations in body fluids where concent¡ations are normally low such as in cerebrospinal flui{ saliva
and tears, as well as in supernatants from mitogen-stimulated lymphocytes in vitro where a sensitive assay is
essential.

105
3.2 RATIONALE FOR USING MONOCLONAL ANTIBODIES
Satisfactory polyclonal antisera for measuri.g human IgG subclass levels a¡e available from only a
few specialised laboratories. Higbly complex and meticulous procedures are required for thei¡ preparation.
Extensive adsorption procedures are necessary to guarantee specificity, high ¿¡fiþqdy tit¡e and the absence
of immune complexes. Antisera can be raised in rabbits or monkeys using myelona proteins of the
different IgG subclasses (Oxelius, 1978; Schur et aIL979). Such anim¿þ can be made tolerant to th¡ee of
the IgG subclasses prior to immrnisation with myeloma proteins of the subclass of tle antisera required.
Monoclonal antibodies (McAb) prepared by hþridoma technology for IgG subclass measu¡ements a¡e
now readily available. Prepared i" high titres they can be suitably standa¡dised and are highly specific
(Lowe et al, L982; Reimer et af 1-98a) Some have expressed reservations about the use of such antisera
(Oxelius, 1988) fearing that they may be restricted in their recognition of certain IgG subclass epitopes.
Extensive evaluation of McAb which a¡e specific for human IgG subclasses has been undertaken by an
IUIS/WHO collaborative study group (Jefferis et a[ ].985). Not all McAbs a¡e suitable for all types of
assay and some have been found to be allotype - restricted. Sixteen McAbs have been identified as suitable
for IgG subclass measu¡ements (Jefferis et al 1983). {l¡þ¡ngh both poþclonal antisera and monoclonal
antisera can be used for tle sa-e types of assays, close attention needs to be given to establishing optinal
assay condi[ions for the particular antisera used if useful comparisons a¡e to be made. Most IgG subclass
resea¡chers would agree that carefully selected monoclonal antisera are suitable for IgG subclass
measurement and there is little doubt, considering thei¡ effectiveness and availability, that they will be used
more often in the assays of the futu¡e. For this reason we used monoclonal antise¡a in ou¡ IgG subclass
ELISA. McAb recommeuded for use in IgG subclass ELISA include: IgG1, HP6012; Igp2,IIp6009,
HP6014; IgG3, IIP6010, SJ33; IgG4, I{P60L1, SK44 (Jefferis et al 1985). However, the details of rhe assay
system may influsnce profoundly the suitability of one or other antibody. McAbs which are avid and
specific fo¡ well-defined epitopes and effective in one assay system may lose their activity in another assay
system (Hussain et al 1-986).

1063.3 II\TERNATTONAL REFERENCE STANDARDS FoR rgc suBcLAssES
At present, tlere is no universal agreement as to the best method for IgG subclass quantitation.
The available reference preparations for the quantitation of IgG subclasses include World Health
Organisation (WHO) reference serum pool 67 /97 IRP (Rowe et a! 1970) (available f¡om WHO
Immunologf Research ¿¡d f¡aining Center, 1066 Epalinges, Switzerland), Fluman standa¡d serum H00-02
(Beck and Kaiser, 1981-a, Klein et al, 19SÐ (available from the Central Laboratory of the Netherlands Red
cross Blood Transfusion Service, p.o. Box 9190, Ansterdr-), u.s. Reference 151644 (Reimer et al 1_9g2)
(available from the Centre for Disease Cont¡ol Atlanta) and British Reference WHO 67/63 (Rowe et a!
1970) (available fron the National Institute for Biological Standards and Control in London). The IgG
subclass composition of some of these preparations determined into several studies is shown in Table 3.1.
Measurements of IgG subclasses have generally been done independently by several investigators using
either RID or RIA methods and polyclonal antisera. It will be noted that there is no total agreement on
the IgG subclass concentrations in these sera. Initially, we used the latter preparation, but more recently
have changed to WHO 67 /97 as it is becom;"g more universally accepted. The values we have used a¡e
those given by Klein et al, 1985 as these are tle latest proposed target values resulting from a collaborative
study in 1985. WHO reference sera6Tfß,67/95 and 67/99 should be the sâme ari 67/97, their differeut
dssig'atioris relaring to divisions of the same se¡um pool prepared in the National Institute for Medical
Research in London into portions for ease of handling (Hamilton, 1987).
In L989, a pilot project to evaluate current methods of IgG subclass quantitation as a preliminary
to establishing a new international reference serum with assþed values for IgG subclass concentrations
was undertaken by the WHO/International Union of Tmmunological Societies (WHO/IUS). Our
laboratory was one of the eleven chosen to participate in this study. The results are yet to be interpreted.
3.4. OVERVIDW OF ESSEI\TIAL STEPS IN OUR IgG SIIBCT,ASS ELISA(FIG.3.1)
(1) Microtit¡e plates are coated with goat anti-mouse (IgG) antibodies.
Subclass specific mouse anti-human monoclonal antibodies are added and captured.a)

Table 3.1 LO7
Igc SUBCLASS LEVELS IN REFERENCE SERA AS REPORTEDBY DIFT'ERENT AUTIIORS
ReferenceSerum
Report IgGL IgGzG/t)
IgG3 IgG4
WHO 67/97 Morell,et alsL97?a
wHo 67 /97 Van derGiessonet aJ,lfiS
wIIo 67 /97 Klein et alL985
0.35
0.55
0.5
0.55
0.55
0.4
2.5
2.2
2.6
5.1
4.8
5.0
0.5
0.6
0.6
0.5
0.5
o.&
3.2
3.3
2.4
5.9
6.2
7.9
HOO-02
HOO-02
HOO-02
Klei¡,et at 1985
Kleinet al L985
Dutch RedCross
USNRP(rs1644)
Reimer,1982
6.95 3.15 0.70 0.61

108
HRP-sheep anti-human Ig
Samples/standards etc
lgG subclass "specific" MoAb
Goat anti-mouse lgG(lgG3:Goat anti-mouse lgM)
rrg.3.1 schematic rrpresentation of our El,rsa for þG subclass quanfitation

(4)
109Serum samples or standards are added so that IgG subclasses bind to the appropriate
antisera.
¡¡otss¡¿rlish peroxidase (HRP)-anti-human immunoglobulin conjugate is added to detect
the bound immr¡nqglqbrrlin subclass.
The substrate,2.2-azi¡o-di-(3-ethyl-benzthiazoli¡e sulphonate-6), ABTS, is added.
Absorbance at 4t4 ''m is measu¡ed and the immunoglobulin concentration in each sample
determined by comparisons with a standard curve.
(s)
(6)
3.5 DETAILS OF TïIE IgG SIJBCI"ASS ELISA
Preliminary studies demonstrated that subclass specific McAb were not suitable for coating
microwells. Hence, an attempt was made to attach them to solid phase was made by usiog goat anti-mouse
IgG (Fc specific) antibody. This proved successful for attaching all 4 IgG subclass-specific McAb to the
polystyrene surface of the microtitre plates. Tmprovements in the assay since it was published (Ferrante et
al, 1986a) are included in the step-wise description that follows.
(1) To measure lgG1.,IgG2 and IgG4 goat anti-mouse IgG (Fc-specific) (Cappe[ Malverq Pa.
U.SA.) is bound to the surface of wells of (% well) mic.rotitre plates (Dynatech Laboratories,
Chatilly Virginia) by adding 2OQpl dilutions made up in 0.1n carbonateþica¡bonate buffer pH 9.6
to the wells and incubating the plates at 4oC for 18 to 20 h¡. To measure IgG3, goat anti-mouse
IgM is used. The dilutions used need to be oprimised for each new lot of plate-coating antibody.
RecentlS the dilutions have been 1/(n0 for IgGL, IgG2 and IgG4 and 1/500 for IgG3. The plates
are then washed with 0.1m phosphate-buffered saline (PBS) pl,rs 0.05Vo TWeen ?Ã, pH 7.4.
Origina[¡ ¿fts¡ washing, 10Qrl of IOVo sheep serum in PBS-TWeen was added to each well and the
plates are incubated for 3 hours at37oC. This step, however, proved to be u¡necessa¡y.
Fifty¡rl of McAb diluted in PBS-Tween with tOVo sheep semm ¡nthliVo merthiolate is then added.
The monoclonal antibodies used are:-
IgGl, NL 16 (Unipath, Oxoid g¿singsf6ke,, Hampshire)
IgG2, HP6014 (Biomakor, formerly Bioyeda, Rehovot, Israel)
IgG3, SJ33 (Sigma, St. Louis, USA)
(3)
(2)

(3)
110IgG4, SKzl4 (Biomakor, formerþ Bioyed4 Rehovot, Israel)
Specificity of each antibody had been confirmed by independent sources and various assay
techniques by the WHO/IUS collaborative study group (Jefferis et al 1935). The above McAbs
have been reco-mended as suitable for use in ELISA.
When we fi¡st developed the assa¡ we used SG-16 (Miles-Yeda" Israel) as the monoclonal
antibody to trap IgG1, but we found that while each of the otler antibodies performed
satisfactorily in our ELISA system SG-16 gave falsely low values for IgGl as shown by major
differences in the total IgG concent¡ation calculated from the su- of the subclasses and that
ï"^î"0 independently by ELISA or by nephelometry, in a few patients, nearly all of whom were
aboriginal (Table 3.2). This led us to suspect that SG-L6 may be allotlpe-restricted. Recent work
indicates that it is probably specific for the IgGl allotype, G1m(f) (Jefferis et al L985). We also
tried using a cocktail of SG-16 and NL16 for IgGl detection, and also using a polyclonal anti-
IgG1. Neither performed as consistently as NL16 alone.
The optimal dilutions for monoclonal antibodies must be determined for each new lot of
monoclonal antibody supplied at the time of writing, dilutions in use were:-
- for IgGL to 1 in 10,000 using NL L6
- for IgG2 to 1 in 20,000 using HP 601a
- for IgG3 to 1 in 1000 using SJ 33
- for IgG4 to 1 itr 2000 using SK zl4
The plates are incubated for 1-8 to 20 hrs at 4oC, and then washed with PBS-TWeen. Dilutions of
5Qrl of IgG subclass standard in triplicate, pooled human senlm (quality control) in duplicate and
dilutions of trnknoum sera in duplicate n L0% sheep serum in PBS-TWeen are then added to the
wells. The plates are incubated for 3 hours at 37oC and washed again.
NeÍ, 6Q.cl of 1/3000 HRPJinked sheep anti-hum¿¡ immunqgloþ'lin (Silenus, Victoria, Australia)
n LÙVo sheep serm in PBS-Tween is added to each well. The opfimnm concentration of
conjugate must be determined for each lot.
(4)

Table 32
PROBLEM RESIILTS INDICATING ALLOTYPE RESTRICTION FOR THE Igcl MONOCLONALAI\TIIBODYSG 16
111
Patient McAb IgGl lgG2 Igc3 IgG4 Sumof IgGsubclasses
s/r
TotalIgG byELISA
Total IgGby nephelometry
1.
,)
3.
4.
5.
6.
SG16NL16
0.904.61.
SG16NL16
0.653.70
L0.5210.52
8.178.L7
7.79
4.53
L4.67
7.74
L7.AZ
7.73
7.33
6.99
8.6L
t0.76
7.n7.n
L1,.20
17.70
6.87
7.87
9.10
9.57
11.90
r.r4 0.2.4 0.77 3.0r.6.72
0.65 0.09 0.02 r.4L4.52
0.574.74
0.?a 0.01.6 L.725.60
0.62 0.1-8 0.12 1.926.97
0.39 0.14 0.15 1.383.L7
0.67 0.4L 0.014 2.159.45
1.18 0.2L 0.n 2.097.54
L.Lt 0.35 3.24
8.838.83
sG16 0.86NLL6 4.74
scl_6 0.99NL16. 6.05
8.556.05
SGl-6NL1.6
0.702.49
sG16 r..06
NL16 8.3ó
sG16 0.68NL16 6.L3
7.
8.
9.
10.
LL.
12.
73.
scL6 t.45NL16 4.35
sG16 r.63NL16 8.53
sG16 1.81-
NLL6 5.98
SG16NL16
SG16NL1.6
1.42t0.7L
6.1815.47
0.30 0.04 0.04 t.824.72
0.26 0.Íì 0.03 2.04
t.13 0.34 0.1.0 3.387.55
0.% 0.48 0.009 2.8L
9.27
L.877.n
0.59 0.45 0.006 2.929.02
8.94
t.%7.82
SG16NL16
- = not measured

LI2
(Ð After a fu¡ther 3br incubation at 37oC in a moist incubator with plates i¡ 5ingte layers to allow
equal hearing tluoughsu¡ the plates -d v¿shing agair:" 10Q¡rl of substrate, ABTS is added to each
well. The colou¡ reaction is allowed to proceed at room temperature for 30 to 45 minutes.
originally the plates were incubated at 37oC but the colour changes progressed too fast for the
plate-reading to be done at the optimal time.
(6) The absorbance at oD414 nm is measured using a Titertek Multiscan MC spectrophotometer
(Boehringer, Mannheip, Dulwich, South Australia), and the immunoglsþulin concent¡ation
computed by comparison with standard curves using an Apple II computer and curve fitter
programme.
To measure total IgG by ELISd -y-chain specific sheep anti-hnman IgG (Silenus Laboratory
Victoria" Australia) is used to coat the wells of microtit¡e plates (Dynatech Labo¡atories, Australia). \0%
sheep serum is added to prevent non-specific binding of the en4rme-antibody conjugate. The amount of
IgG bound is quantitated by the addition of conjugate, HRPJinked sheep anti-human immunoglobulin,
(Amersham, Sydney, Australia) followed by the substrate, ABTS (Boehringer, Mannhei¡, Dulwich, South
Australia). The absorbance at 4L4nm was measure{ and the IgG concent¡ation in each test sample w¡rs
determined by comparison with a standard cu¡ve.
Initially we used H'man standard serum, H00-02 (Red Cross Blood Transfusion Service,
Netherlands) for our IgG subclass standard. At the end of 1-987 we çþenged to WHO reference serum
67 /97 as ttis is more commonly used internationally for IgG subclass measurements with the completion of
the wHo/rurs study (section 3.3) improved standa¡disation should be possible.
At first, all the steps of this ELISA were done manually. Later, we introduced a Biomek 1000
automated laboratory workstation (Fig. 3.2), (Beckman, Palo Alto, CA5 U.SA.) for some of the pipettin&
dituting and plate washing steps. This increased to the accuracy of the assay and made it less tedious to
perform.

113
l'
r}
..r.'ã
û,.-
Fig.32 The Biomek 1fiX) workstation.

LI43.6 ACCT]RACY OF OTJR ELISA
Standard curves were produced using reference sera (Fig. 3.3-6). They were similar in shape and
sensitivity for all 4 subclasses. Numerous e4perimental runs showed the lower limits of detection for IgGL
and IgG3 to be approxirnately 7ng/ml, and for lgGZ and IgG4 to be approximately L0ng/ml. Further
studies showed that the standard curves for each IgG subclass did not differ significantly between plates.
IgG subclass concent¡ations wers srnmated and the value compa¡ed with the total IgG
concentrations measu¡ed by ELISA and by nephelometry. The best agreement was between the sr¡m of the
subclasses and the IgG concentration measured by nephelometry. Nephelometry IgG = f2Vo + 25Vo (svm
of subclasses).
3.7 COMPARISON \ilITII OTT{ER RECEITTLY DEVELOPED ELISA METIIODS
FOR IgG SIIBCI"ASS QUAITTITATION
A number of ELISA or competitive ELISA assays for measuring IgG subclasses have been
developed recently. Problems are encountered when attempts are made to coat microtitre plates with IgG
subclass specific monoclonal antibodies and va¡ious ways of stabilising these antibodies must be found for
di¡ect ELISA. Papadea et al (1985) have described a method similar to ou¡s but using gluteraldehyde-
treated bovine serum ¿lþrrmin to coat the plates and capture the IgG subclass specific McAb, instead of the
goat anti-mouse antiserum that used in our method.
An ELISA kit for measuring IgG subclasses is co--ercially available. This uses plates coated
with antihuman immuuroglobulin tq which all the IgG subclasses bind (Fig. 3.7). Bound IgG is identified
and quantitated by using HRP subclass-specific McAb conjugates. Working with this kit, we found that
IgG2 curves were poor, and that there was great variability betrveen kits. Some of the problems were
overcome bymodi$ing the serum blocking step (Ferrante & Beard 1988).
Competitive ELISA assays for IgG subclasses have also been described (Aucouturier et at L985;
Skvaril, 1986). Aucouturier et al coated plates with purified monoclonal IgG subclass and then added a
mixture of samples, or standards, together with the IgG subclass specific McAb. IgG subclass
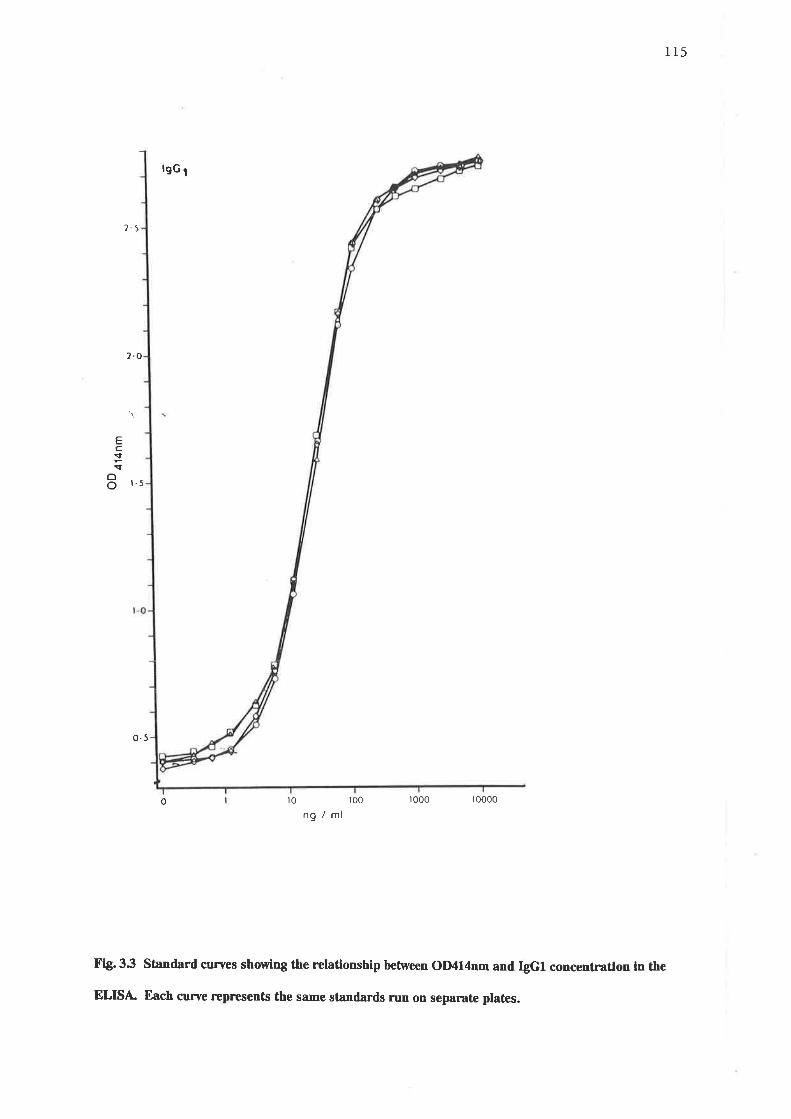
115
lgc t
2.5
2.O
Ec:çoo t-5
0.5
r0 r00 ro00 10000
ng/ml
Fig.33 Stnndard curaes showÍng the relationship between OIXl4nm and þG1 concentratlon in the
ELISÀ Each cu¡ve rcprcsents the same standards run on separate plates.
0

116
lgc 2
Ec:Io
o
25
r5
05
0 r0 t00 too0 10000
ng/ml
Fig' 3'4 stand¡rd cuFves showing the relationshlp between olXl4nm and þG2 concentratlon Ín the
ELISÀ Each curve repr€sents ttre same standards run on sepamte plates.

TI7
lgG¡
?'5
EcI1
oo t5
r.0
0.5
0 r0 00 1000 10000ng/ml
fig.35 Standard cuFves showing the relationship between OIXl4nm and þG3 conccntration ln the
ELISÀ Each cune reprcsents the same standards run on separate plates.
0

r18
lgG¡
2.5
EcIçoo 1.5
1.0
0.5
0 0 r0 r000 r0000ng /ml
Fl& 3.6 Standard curves showÍng the relationship between OD414nm and þGrl concentratlon ln the
ELISÀ Each curve reprcsents the same standards run on separate ptates.
00

119
<- HRP-subclass "specific" MoAb
-
Samples/standards
<- anti-human lg
HRP
Flg. 3.7 Schemadc rcpnesentation of commerctally avallable ELISA for þG subclass quandtadon.

L20concentration can be determined accordi'g to the ability of the serum samples to inhibit binding of the
McAb to the IgG on the plastic surface. Skvaril (1936) describes an alternative ¡eshnique in which
subclass-specific McAb a¡e bound to microtitre plates before the serum samples a¡e added. The
concentration of subclasses is determined ¿scs¡ding to the degree of inhibition of binding of alkaline
phosphatase-human myeloma IgG subclass conjugates.
3.8 ALTERNATTVES TO ELISA
ELISAs are not the only sensitive assays for IgG subclass measurement. Other assays such as
electro-immunodiffusion (sensitive down to 0.002-0.0059/l) (Skvar{ 1986), RIA,, with a higher sensitivity
*- *, (Heiner, 1984; Shackelford et al, 1986) and particle concent¡ation fluorescence immunoassays
have been developed (Mayus et at L986)
3.9 COMPARISON OF RID AND ELISA
We compared the results of RID, using polyclonal antisera" and ELISd using McAb, to measuÍe
IgG subclass concentrations in the sera of 39 adults. While relative percentages rye¡e simil¿¡, absolute
values were different (Table 3.3). This indicates that patients results need to be evaluated using a normal
range established with the same technique. Even theq a consideration should be given to the finding that
IgG subclass concentrations may, to some eÍent, be racially determined (Shackelford et al, 1-986), and also
that differences in laborator! handling and reagents may be important (Dju.up et al 1983).
3.10 COMPARISON OF PRELIMINARY RESTJLTS OBTAII\ED WITH OIJR
ELISAWITII THOSE OF OTTIER II{VESTIGATORS
Using ou¡ ELISA method we found the concentrations of IgG subclasses in the sera of a group of
healthy adults to be in general agreement with those reported by other investþators who used a variety of
techniques: RID, RIAs ELISA and particle concentration fluorescenss immuno¿ssay and either poþclonal
or moroclonal antisera. 'We were, however, unable to demonstrate tÏe very low IgG4 levels that others
such as Oxelius (1979a), French and Harrison (1984a) and Aucouturier et al (198Ð have found in a
proportion of normal healthy subjects. It is possible that our small group of subjects (ody a1) was not

L2LTable 33
A COMPARISON OF IsG SUBCIÀSS MEASUREMENTS USING RID AhID ELISA
Subclass RIDpolyclonal antisera
ELISAmonoclonal antibodies
IgGl
rñ2
IgG3
IgG4
6.12+ L.576I.29Vo
2.78+ L.7526.y%
0.62+ 0.296.3OVo
0.61-+ 0.¿m
6.07%
7.4t+ 2.4r64.(AVo
2.55+ 0.9422.32Vo
0.77+ 0.356.7ÙVo
0.75+ 0.496.35%
Results are mean+ SD in g/l of 39 healthy adults.RID was conducted with co-mercially available plates (Seroteg Oxon, Engtand). Monoclonal antibodiesused: HP6012 (IgGL), I{P6014 (rgG;2), SJ33 QgG3) and SK¿14 (Igca).

722
represontative of the general population. It would appear that the highe¡ mean values of IgG4 in our study
compared with other studies can only be partly erçlained by the different techniques used since, using the
RID method on our samples we obtained values only slightly lower tlan those that we had obtained by
ELISA. Other technical factors, such as different antisera" and different calibrators, population factors
such as race and IgG allotype patterns and geographical and envi¡onmental (Shackelford et al 1985)
factors may all have contributed to the differences.
French and Ha¡rison (1984a) found IgG4 concentrations to be slþhtly highs¡ in males than in
females. Since our group consisted of a predominance of fenales, Q7 /a\ our higher values cannot be
e4plained on the basis of having ¿ higher proportion of males. French and Harrison (1-984a) have suggested
that extensive absorption of the polyclonal antisera used in some studies may result in an IgG4 antiserrrm
which fails to detect a subpopulation of IgG4, eg. IgG4a or IgG4b subtypes, and hence gives lower IgG4
results tlan monoclonal antisera.
Subsequently, extending our studies to include a larger number of normal adults we found that
although our median value for IgG4 was closer to that in other studies, we still did not fud IgG4 as low in
any subjects ¿ìs some other investigators have done (See Ch. 4).
3.lL CONCLUSION
If IgG subclass quantitation is to be useful, it must be able to be done accurately and reproducibly.
For IgG subclasses present in low concentratio^ ¡¡ 5amples to be analysed" the sensitivity of the assay used
may be a crucial factor. This is particularly important for paediatris samples where concentrations of IgG4,
especiall¡ may be too low to detect by some commonly used assays. gimil¿¡l¡ for the measulemont of IgG
subclasses in body fluids other tlan serum, and for the measu¡ement of IgG subclass production in vitro, a
hþhly sensitive assay is required. The ELISA for IgG subclasses described in this chapter, meets all these
requirements.

CHAPTER F'OUR
ESTABLISHING NORMAL RANGES FOR IgG SUBCLASS
CONCENTRA1IONS IN HEALTITY AUSTRALIAN CHILDREN

r244.I INTIR.ODUCTION AND PRELIMINARY STTJDIES
Initiall¡ in attempting to establish paediatric normal ranges for serum IgG subclasses
concentrations in healthy Australian children and adults, we c¿lculated the mean serum concentrations and
standard deviations (SD) at each age (Table 4.1). This was not ideal as the IgG subclass concentrations did
not follow a normal distribution pattern, and the standard deviations were often extremely large.
Subsequently, we defined the 5th and 95th percentiles at each age by non-parametric methods without
reference to the data obtained for subjects of higher and lower ages (Table 4.2). This was more useful"
although there were some irregularities related to inadequate numbers of subjects at each age. Adult IgG
subclass percentiles were calculated similarly in 81 adult control subjects (Table 4.3). The adults while
basicallytealth¡ were not as carefully screened for infection proneness as were the child¡en (see section
4.2.L).
More recentl¡ with the assistance of Mr P. Leppard (Statistical Consultant, University of
Adelaide) we have been able to fit our data to a matlematical formula which has allowed the computatiou
of percentile rânges for IgG subclasses in children. The computation has taken into account data from
subjects of lower and higüer ages tlan the age in question, and so has enabled us to make optimum use of
all the data available.
This ehapter deals wtth the development of these percentile rânges and compares them with
previously published paediatric normal rânges (see Beard et al 1990a).
In the studies described in subsequent chapters, normal ranges for IgG subclass have been defined
by the 5th and 95th percentiles for age unless otherwise stated.
The measu¡ement of senm IgG subclass concentrations is being used increasingly to h9þ to assess
the immunological competence of infection-prone subjects. With the recent development of suitably
standardised IgG subclass monoclonal antibodies and of enz5tme-linked immunosorbent assays which are
sensitive s¡16troh to measure concentrations of each of the IgG subclasses, even at the low IgG3 and IgG4
concentrations that are found in yort"g childreq the scope of IgG subclass measurement as a diagnostic
tool has increased.

r25
flowever, there are many difficulties in interpreting results, particularly in children, as the few
paediatric normal rânges that have been published to date are not in agreement about the cutoff values
between normal and low concentrations. Using conventional statistical methods eg. mean* 2SD (with or
without log tr¡nsformation to normalize the data) to estimate normal values for IgG subclasses in children
of different ages, it has not been possible to const¡uct smooth age-related percentile curves, partly because
inadequate n'mbers of children in each age group have resulted in wide fluctuations from one age gtoup to
the next. It has been assumed that male and female children do not differ sig'nificantly in their IgG subclass
concentrations at identical ages and comparative normal ranges for male and female children have not been
published. Because the first published paediatric normal rânges were determined by radial
immunodiffusion, radioimmunoassay and elect¡oimmunoassay using polyclonal antibodies (Morell et al
1972b; Van der Giessen et al, t975; Schu¡ et ú, t979; Oxelius, L979a; Shackelford et al, 1985) and today
ELISA techniques and/or monoclonal antibodies are bei.g used by an increasing number of investþators,
different normal ranges may be-necessary. In this 5g¡dy we measured serum IgG subclasses by an ELISA
technique ¡5ing monoclonal antibodies. Using an age-dependent va¡iant of the Box-Cox transformation,
we then developed paediatric age-normal reference percentle ranges for healthyAustralian children.
42 STJBJECTS AND STATISTICAL ANALYSIS
4.2.L Subjects
Sera f¡om 2g}healthy child¡"n 171 males alaidl2l females, ranging from age 4 months to 15 years
were analysed. Children who were excessively infection-prone (i.e. above the 95th percentile for infection
proneness) on an assessment scale produced for use in Australian children (Roberton & Hosking, L987)
and children with immunological disorders were excluded. Child¡en with atopic rlisease, defined either
clinicallyorbyveryhighserumTgEconcentrations, (IgE > 250i.u./mlforlto2year olds, >500for2to15
year olds) were excluded also. The children were drawn fr66 immunisation clinic attenders and their
siþlings, from co-mrrnigy contacts and from hospital attenders with non-infective problems eg. short
stature.

r26
422 StatisticalanalYsis
An age-dependent variation of the Box-Cox trnnsformation (Box and Co4 19t6+) was determined
for each of the variables. The ¡ssulting transformed va¡iable was exarnined for normality and was found to
be satisfactorily so in every case. Regression of the transformed variable on age then allowed central
percentile ¡anges to be calculated.
4.3 RESULTS
The resultant serum IgG subclass concent¡ation percentiles for males and females from age 4
months to 15 years are shown in figures 4.I-4.4 and for males and females combined in figures 4.5 and 4.6.
Tables 4.+4.6 show 90Vo confidence limits 4.7-4.9,95Vo confidence limits in tabula¡ form. The mean
relative percsntage of the IgG subclasses at different ages is shown in Table 4.10. IgG of all subclasses was
detectable in all subjects although IgG4 concentrations in some child¡en less ttran two years of age were
<0.0fg/1. The sum of the IgG subclasses obtained by ELISA compared well with the total IgG
concentration es"mated by ELISA and by nephelometry in each of the samples. Fiftieth percentiles for
males and females were simila¡ for IgGl and IgG3, while that for lgp2 in males over 10 years fell
increasingly below that of females, and that for IgG4 ¡v¿s highsr for males of all ages than for females. The
lower limits (2.5th percentile) of the IgGl1 lgG2 and IgG4 ranges tended to be slightly lower in females but
for IgG3 the lower limit in males fell below that of females over 9 years of age (Fig. 4.7). A comparison of
our IgG4 ranges in males and females showed very little difference between tlem.
4.4 DISCUSSION
4.4.1 Comparisons of paediatric normal ranges for IgG subclasses
5sps dsteils of the paediatric normal ranges that have been published are shoqm in Table 4.11.
Figure 4.8 compare mean or 50th percentile concentrations of IgG subclasses in some of these studies, and
Figure 4.7 compares the lower limits of normal ranges. Only those studies in which results for children
more than 2years of age are given expressed in absolute values have been compared.

r27
IgGl: Mean or 50th perceutile values for IgGl are very similar in most of the studies Fþ. 4.8. Aucoutu¡ier
et al (1987a) found higher values than the other investþators. Our studies gave 50th percentile values
about the middle of the range shown in the other studies. The lower limits of normal given in these studies
a¡e similar (Fig. a.f, although they have been determined or calculated in different ways. The study of
Bird et al (L985) gives exceptionally low lower limits. Our lower limits for IgGl, shown as the 2.5th
percentile in both males and females are relatively highs¡ than those of most other studies.
IgG2: Mean or 50th percentile for lgp2 show much interstudy variation (Fig. 4.8) and there is a bþ
discrepancy in the lower limits of normal e:rpressed in these various studies. Values in Oxelius' study
(1979a) are considerably hþher than those in the others. Our 50th percentile values for both males and
females were within the range of those in other studies, and closest to those of Singh's study (1988). Our
lower limits for lgp2, the 2.5th percentile, were considerably lower than those of Oxelius (1979a) and
considerably higher than those of Schur et al (1,979) at most ages (Fþ. a.7).
þG3: Mean or 50th percentile values show much interstudy variation. Our values for males and females
are lower than those in most of the other studies, but comparable to those of Aucouturier (1987a) (Fþ.
4.8). Our lower limits for IgG3, 2.5th percentile, fall within the range of the lower limits defined in other
studies (F 9. 4.7).
SQ!: Mean or 50th percentile values in most studies show wide fluctuations with age (Fig. 4.8). Our study
gave values within those obtained in other studies. Oxelius (L979a), Schur et al (1979) and Bird et al (1985)
found undetectable IgG4 levels in considerable numbers of healthy children making the lower limits for
IgG4 difEcult to define. We found detectable IgG4 in all the normal children we studied but values were
below 0.0Lg/l in a number of males below 2yearc of age, and of females below 1 year of age. There was
little difference in the 2.5th percentile values for males and females.
4,4,2 Possible reasons for differences between studies
The inter-study variations in normal ranges illust¡ated here in studies using different methods,
different antisera, different reference standards and different cutoff criteria indicate the need for
standardization of both IgG subclass measurement techniques and statistical analyses if ¡eaningful

T28
comparisons of results obtained in different laboratories are to be possible. Even then, some reservations
are warranted. A recent study by Djurup et al (1988) compared IgG subclass concentrations in sera from
ZX) normal adults measu¡ed in two different laboratories using the same technique (RID), tle same
subclass specific antisera" and the same calibrator. Even with this degree of standardization, signifiç¿¡¡
differences were found. In one laboratory the dist¡ibutions of IgGl and IgG2 were normal, whereas those
of IgG3 and IgG4 deviated significantly f¡om normal, while in the other laboratory all 4 subclasses were
significantly different from a normal distribution. Since the same samples were analysed, differences may
have to be attributed [s mins¡ variations in technical performance of the assays in the two laboratories.
This shows the need for caution in interpreting patients' IgG subclass results measured in one laboratory
with reference to a normal range established in another laboratory.
Differences in tle relative proportions of males and females in different studies could be a factor
accounting for differences between studies, but its relevance cannot be assessed as published paediatric
normal range studies have not generally compared or recorded ¡6s fi¡¡tings in males and females. We
found differences in the 50th percentile values for males and females to be most ma¡ked for IgG2 and for
IgG4, and differencss in the 2.5th percentile values to be most marked for IgGl andlgp2. Shackelford et
al (1985) found by comparing mean concentrations, that female cbildren between one and 38 months of age
had higher concentrations of IgG1, than did males. Although, we did not find major differences in 50th
percentile values for IgGl in ou¡ study, in children less than 8 years of age, values *e¡e slightly highe¡ ia
females, and in older children, sþhtly higher in males.
Differences in the criteria used to select children who were defined as healthy and included in the
studies may be anotler factor accounting for some of the inter-study variations. Schur et al (L979) do not
give details about the health of the children in their study. Oxelius (1979a) included children who were
largely hospital inpatients with surgical and other non-immunological disorders. They were infection-free
and had erythrocyte serlimentation rates below tQmm/tr. Shackelford et al (L98Ð included children who
were participants in various vaccitre studies or who presented to doctors for routine preventative ca¡e. Bi¡d
et al (1985) c.hose children who attended local child welfare clinics and does not give detafü about their
health. Aucouturier et at (1987a) used child¡en who were admitted to casualty for trauma and does not
give other details of their health. Singh (1988) gives no information about the health of children included in
his study. In ou¡ study we excluded children with a history of excessive infection-proneness, or with

t29immunological disorders, and with atopic disorders defined by history or by very higlh serum IgE
concent¡ations. It can be seen that the populations ofchildren included in these studies a¡e not necessarily
strictly comparable.
Genetic factors probably play a part in the differing IgG subclass normal ranges defined by
different ¡esearchers in different parts of the world. The association between IgG subclass levels and Gm
phenotlpes indicates tþ¿t this should be so (see chapter I.3.4.2). Different populations having different
predominant Gm phenoty¡res may have different concentrations of IgG subclasses.
Environmental factors, also, are likely to influence IgG subclass levels. Hence, repeated expostue
to antþens likely to stimulate the production of antibodies of a particular subclass may result in higher
concentrations of that subclass than would otherwise be e>cpected. Eg5rptian patients *¡¡þ schisfosomiasis
have high concentrations of IgG4 (Iskander et al 1981). Over long periods of tims one would erçect to see
the effect of selective advantage on IgG subclass values in given populations. The survival of individuals
with the Gm allotypes associated with good responses to certain prevalent envi¡onmental pathogens would
be favoured. These responsos would be reflected in subclass concentrations which would therefore, vary in
populations exposed to differing antigen loads. A connection between immuneglsþulin ¿llstlpss and
immr¡n6gl6þ 'lin responses to some antþenic stimuli has been clearly demonstrated (Pandy et al, 1979;
Ambrosino et al, 1985; Granoff et al 1986a).
4.4.3 Conclusions
The wide fluctuations in concent¡ations of some IgG subclasses with age seen in many of the
studies are likely to be, to some extent, artefactual and to reflect the lack of numbers of subjects of different
ages and the use of statistical methods of analysis that have failed to take into consideration the
concent¡ations of the subclass in children of lower and higher ages than the one in question. There a¡e
ethical problems and practical difficulties in obtaining blood sanples from healthy children. By using a
statistical method that takes all the data into account, rather tþ¿n t¡s¿ting subclass concentrations at each
age independently of those at other ages, we have been able, from 2T2 serum samples, to calculate IgG
subclass age-related percentiles that vary smootlly with age for male and female children. Because such
major difference in IgG subclass concentrations have been found in different studies in children, ideally,

130
each laboratory measuring IgG subclass concentrations in paediatric patients should develop its own age-
normal reference ranges. When this is not possible, ¡anges established by similar techniques, with similar
reagents, with the same standa¡ds, and in simila¡ populations should be used.

Table 4.1 131
SERUM lgc SUBCLASS CONCENTRATIONS AT DITTERENTAGES G/l), MEAN+ SD
Age No IgGl rñ2 IgG3 lgc4
4-6mo
7-9mo
1-0-l-2mo
2.77+ L.Lz
3.t9+ 0.y2
3.37! L.rO
3.82! t.12
4.85+ r.n
4.67! L.62
5.55+ 1-.68
5.70+ t.37
6.14+ 2.&
5.34+ !.04
5.6t! t.?Á
5.90+ 1.35
6.18+ 1.36
5.75+ 1.,18
5.90+ 0.88
5.09+ 1.50
6.05+ L.24
6.02+ 0.99
s.LL! L.72
0.57+ 0.23
0.53+ 0.20
0.611 0.07
0.94+ 0.55
1.15+ 0.57
1.38+ 0.73
L.25+ 0.&
1.63+ 0.78
1.50+ 0.57
L.%! 0.42
L.99+ 0.75
L.70+ 0.70
t_.81+ 0.55
1.84+ 0.98
1.41+ 0.68
1.83+ 0.¿m
1.65+ 0.55
2.1i+ L.4t
2.77+ t.M
0.3t! 0.23
0.zi+ 0.10
0.33+ 0.04
0.30+ 0.15
0.37+ 0.18
0.381 0.26
0.37+ 0.L9
0.51+ 0.25
0.54+ 0.39
0.4t! 0.20
0.55+ 0.23
0.491 0.L8
0.41+ 0.14
0.571 0.30
0.6+ 0.12
0.¿l4t 0.18
0.47+ 02A
0.43+ 0.19
0.55+ 0.2lí
0.056 + 0.038
0.05+ 0.05
0.2r.t 0.16
0.16+ 0.17
0.33+ 0.32
0.a3! 0.37
0.35t o.z2
0.35+ 0.30
0.37+ 0.¿10
0.39+ 0.31
0.47+ 0.33
0.59+ 0.61
0.72t 0.50
0.5L+ 0.42
0.,18+ 0.,+8
0.55+ 0.47
0.691 1.45
0.4L+ 0.07
0.57+ 0.56
L-Zw
2yr
3yr
4yr
5yr
6yr
7yr
8yr
9r
1or
11yr
12yr
ljyr
L4w
15yr
Adult
25
13
4
36
?Ã
L9
?n
n14
L6
4t4
10
37
1_0
9
9
L2
49
mo = months, yr = yeafs

Table 42
PRELIMINARY9OTo CONFIDENCE LIMIS FOR SERIIM lgc SUBCLASS CONCENTRATIONSHEALTIIY IN AUSIRALIAN CHILDREN
Age(years)
No. IgGl TÑ2 Igc3 TÑ4
r32
0.3-0.5
0.5-1.0
1-2
2-3
3-5
5-8
8-11
LL-T4
L+L7
t+15
15-16
25
18
K
?n
39
6
63
51
25
9
10
1.15-5.53
L.93-5.43
2.L2-6.67
2.7r-6.&
2.6r-7.æ
3.M-7.55
3.69-7.82
2.95-7.L6
3.17-8.21
3.81-8.21
4.75-8.07
0.?Á-r.29
0.26-0.93
0.?a-1.75
0.38-2.93
0.66,3.n
0.65-3.52
0.93-3.51
o.5r-4.43
0.6r-2.59
0.73-2.59
0.97-2.M
0.13-0.67
0.13-0.43
0.10-0.61
0.08-0.75
0.1+0.90
o.l+0.96
0.26-0.Í36
0.25-0.95
0.15-0.98
0.15-0.98
0.27-0.Í36
0.005-0.Íl
0.007-0.46
0.0r-0.47
0.01-1,.25
0.07-L.n
0.09-1.91
0.0+1.63
0.I0-L.u
0.Í!-1.14
o.LL-0.92

Table 43 133
SERIJM IsG suBcr,ass PERcEllrrLEs IN TIEALTHYaUSTRALIAN AI)t LTs
Igcl TÑ2 IgG3 Igc4
slr
Pzs
P5
Pro
P25
Pso
Pls
P9g
Pgs
Pn.s
3.10
3.37
3.80
4.89
6JJ,3
7.63
8.63
8.90
9.04
L.12
L.32
L.Æ
2.8L
3.q
4.68
5.y3
6.29
8.4ó
0.16
0.22
0.26
o.y
0.4
0.58
0.79
0.90
1.00
0.06
0.09
0.12
o.n
0.43
0.74
1.06
1.65
1.69

Table 4.4 134
SERI]M IgG SUBCI"ASS9OIO CONFIDENCE LIMMS IN IIEALTITYAUSTRALIAN MALE CHILDREN
IgGl IgG2 IgG3 IÑ4Agelvea¡s)
0.33
0.50
0.75
L
2
3
4
5
6
7
8
9
10
11
a13
t4
15
t.75-6.42
LA-6.53
t.92-6.69
2.@¡6.U
2.ß-7.4r
2.88-7.9L
3.U-8.y
3.55-8.71
3.80-9.02
4.N-9.25
4.t6-9.42
4.26-9.5t
4.3L-9.53
4.32-9.47
4.?Å-9.y
4.?n-9.r4
4.09-8.87
3.95-8.55
0.33-1.60
0.*L.67
0.:x-r.79
0.38-1.90
0.ß-2.%
0.61-2.80
0.7+3.m
0.87-3.56
0.98-3.86
1.07-4.08
t.L+4.22
L.L7-4.n
L.L8-4.23
t.L7-4.tL
1.ß-3.90
t.07-3.63
1.00-3.31
0.9r-2.96
0.08-0.,l8
0.08-0.49
0.08-0.51
.0.09-0.52
0.10-0.58
0.12-0.63
0.Íì-0.68
0.t+0.72
0.15-0.75
0.16-0.77
0.r.7-0.78
0.L7-0.78
0.L7-0.77
0.L7-0.75
0.t6-0.72
0.15-0.69
0.15-0.65
0.L40.60
0.00-0.30
0.00-0.32
0.01-0.35
0.01--0.38
0.02-0.5r.
0.03-0.67
0.0+0.83
0.06-1.00
0.07-L.L6
0.09-r..30
0.t0-r.42
0.LL-r.52
0.12-L.57
0.t2-t.59
0.L1-1.56
0.10-1.50
0.08-1.39
0.M-L.26

Table 45 135
SERUM IgG SUBCI"ASSSÛTo CONFIDENCE LIMITS IN HEALTHYAUSTRALIAII FEMALECIIILDREN
IgGl Íñ2 IgG3
0.08-0.¿18
0.08-0.49
0.09-0.50
0.09-0.51
0.10-0.54
0.11-0.57
0.12-0.ffi
0.1+0.63
0.15-0.66
0.16-0.68
0.L7-0.70
o.r8-0.72
0.18-0.73
o.L9-0.74
0.2a-0.75
o.2a-0.76
0.2L-0.76
o.2r4.76
ICG4
0.01-0.26
0.0L-0.n
0.01-0.30
0.01-0.33
0.û2-0.45
0.03-0.59
0.0+0.75
0.05-0.93
0.06-1.11
0.07-L.29
0.07-1.4
0.08-1.55
0.08-1.62
0.08-1.63
0.07-1,.59
0.06-1.50
0.05-1.37
0.034.24
0.50
0.75
1
2
3
4
5
6
7
I
9
10
TL
L2
li
L4
15
t.73-6.L0
L.80-6.22
1.90-6.56
2.0-6.56
2.4t-7.?r
2.ffi-7.78
3.L6-8.29
3.47-8.72
3.73-9.08
3.9+9.%
4.09-9.55
4.t8-9.6
4.20-9.æ
4.L7-9.62
4.07-9.6
3.9L-9.?3
3.70-8.91
3.45-8.52
0.%-t.96
0.37-2.0L
0.38-2.08
0.39-2.L6
0.45-2.45
0.5L-2.74
0.58-3.03
0.65-3.32
0.73-3.59
0.80-3.84
0.87-4.07
0.9+42A
l.00-4.6
1.05-4.61
L.W-4.73
L.12-4.81
r.ru.85
1.15-4.8ó

136Table 4.6SERUM IgG SUBCI,ASS9OTo CONFIDENCE LIMITS IN HEALTTIYAUSTRALIAN CHILDREN
(MALES AND FEMALES COMBINED)
Age(years)
IgGL lgG2 IgG3 IgG4
0.33
0.50
0.75
1
2
3
4
5
6
7
8
9
10
11
L2
13
L4
L5
1.79-6.24
L.86-3.56
L.97-6.53
2.07-6.69
2.ß-7.3L
2.f36-7.87
3.2L-8.36
3.51--8.78
3.77-9.13
3.n-9.39
4.11-9.58
4.20-9.68
4.V+-6.94
4.22-9.62
4.15-9.47
4.0+9.ffi
3.88-8.92
3.69-8-53
0.33-1.80
0.35-1.86
0.%-L.96
0.38-2.05
0.47-2.4
0.57-2.8L
0.67-3.L7
0.77-3.49
0.f16-3.79
0.95-4.03
L.02-4.23
L.07-4.37
t.rL-4.44
t.1i-4.45
r.1i-4.q
1.12-4.29
r.09-4.12
1.05-3.91
0.08-0.49
0.08-0.50
0.08-0.51
0.09-0.52
0.10-0.57
0.11-0.61
0.13-0.65
0.1¿l-0.68
0.15-0.71
o.L6-0.73
0.r7-0.75
0.r7-0.76
0.18-0.76
0.18-0.76
0.18-0.75
0.18-0.73
0.18-0.71
0.17-0.68
0.006-0.29
0.007-0.30
0.009-0.33
0.010-0.36
0.018-0.49
0.0n-0.&
0.039-0.80
0.051-0.97
0.064-L.L4
0.075-L.29
0.085-r-.42
0.091-1.51
0.093-1.56
0.091-1.57
0.08¿l-1.53
0.07+L.45
0.060-1.33
0.045-1.18

Table 4.7t37
SERTJM IgG ST]BCLASS9S%|o CONFIDENCE LIMITS IN IIEALTIIYAUSTRALIATMALE CHILDREN
IgGL lgGz IgG3 IgG4Age(yea¡s)
0.33
0.50
0.75
1.
2
3
4
5
6
7
8
9
10
11_
12
13
L4
15
L.67-6.98
t.67-7.W
L.69-7.25
L.77-7.q
2.16-7.97
2.55-8.¿18
2.89-8.93
3.L9-9.3L
3.+9.62
3.*9.86
3.79-t0.03
3.90-10.ß
3.96-L0.15
3.97-L0.09
3.9+9.96
3.88-9.75
3.78-9.47
3.6-9.13
0.32-L.87
0.32-L.96
0.3+2.08
0.35-2.2t
0.43-2.7r
0.53-3.19
0.65-3.63
0.76-4.02
0.86-4.35
0.9+4.59
L.N-4.74
1.0+4.80
L.05-4.75
1..0+4.6t
1.01-4.38
0.96-4.08
0.90-3.72
0.83-3.32
0.07-0.55
0.07-0.57
0.07-0.58
0.08-0.60
0.09-0.66
0.t0-0.72
0.L1,-0.77
0.12-0.81
0.ß-0.84
0.1+0.86
0.1+0.87
0.15-0.88
0.15-0.87
0.1¿t-0.85
0.1+0.82
0.ß-0.78
0.13.0.73
0.L2-0.68
0.00-0.38
0.00+0.41
0.00-0.44
0.00-0.,+8
0.01-0.64
0.v2-0.82
0.03-1.01
0.0+r.2t
0.05-1.¿10
0.0ó-1.57
0.07-L.71.
0.08-L.8L
0.08-1.88
0.08-1.90
0.07-1,.87
o.M-L.79
0.05-1.68
0.0+1.53

Table 4.8 138
SERIIM IgG SUBCLASS9SVo CONFIDENCE LIMITS IN IüALTIIYAUSTRALIAN FEMALECHILDREN
Agelvea¡s)
0.33
0.50
0.75
1
2
3
4
5
6
7
8
9
10
LL
12
13
L4
15
IgGl IgG2 IgG3
r.49-6.61
t.55-6.74
L.63-6.9L
t.72-7.09
2.08-7.74
2.45-8.33
2.78-8.85
3.08-9.30
3.33-9.6
3.35-9.95
3.68-10.15
3.76-t0.26
3.79-L0.28
3.75-r0.ZL
3.66-10.06
3.52-9.82
3.32-9.49
3.08-9.09
0.32-2.28
0.33-2.y
0.3+2.42
0.35-2.50
0.39-2.82
o.+3.15
0.50-3.47
0.56-3.78
0.62-4.08
0.68-4.35
0.7+4.61.
0.80-4.84
0.85-5.03
0.9{J-5.2A
0.9+5.32
0.96-5.41
0.98-5.46
0.99-5.47
0.07-0.55
0.07-0.56
0.08-0.57
0.08-0.58
0.æ-0.62
0.r.0-0.65
0.L1-0.68
0.12-0.7L
0.1i-0.74
0.L+0.76
0.15-0.79
0.15-0.80
0.L6-0.82
0.17-0.83
0.17-0.84
0.L8-0.85
0.L8-0.85
0.19-0.85
IgG4
0.0L-0.34
0.01-0.36
0.01-0.39
0.01-0.43
0.02-0.59
0.02-0.78
0.03-0.99
0.wt.23
0.0+L.6
0.05-1.69
0.05-1.89
0.M-2.04
0.6-2.13
0.M-2.15
0.05-2.10
0.05-1.98
0.04-1.81
0.02-L.59

139Table 4.9
SERUM IgG SUBCLASS9STo CONFIDENCE LIMITS IN IIEALTITYAUSTRALIAN CHILDREN(MAIÆS AND FEMALES COMBINED)
Age(years)
IgGl lgGZ IgG3
0.07-0.49
0.07-0.58
0.07-0.59
0.08-0.60
0.09-0.65
0.10-0.70
0.tt-0.74
0.12-0.77
0.L3-0.80
0.121-0.83
0.1¿l-0.84
0.15-0.85
0.15-0.05
0.16-0.85
0.1-6-0.84
0.16-0.82
0.15-0.80
0.15-0.77
rgG4
0.003-0.37
0.003-0.39
0.004-0.42
0.006-0.,16
0.01L-0.62
0.018-0.80
0.0z'-1.00
0.035-L.2L
0.w1..4L
0.053-1.59
0.059-L.74
0.0úl-1-.8ó
0.M5-r.92
0.063-1.93
0.06-1.88
0.05-1.79
0.0+L.65
0.03-1,.47
0.33
0.50
0.75
1
)
3
4
5
6
7
8
9
10
11
12
1i
14
15
1.56-6.73
L.6L-6.90
1.70-7.08
L.80-7.24
2.L7-7.88
2.52-8.6
2.f36-8.96
3.15-9.39
3.39-9.75
3.59-r0.02
3.73-L0.2t
3.82-L0.32
3.86-10.y
3.85-t0.?Á
3.79-L0.LL
3.69-4.M
3.5+9.53
3.37-9.13
0.31-1.80
0.32-2.L9
0.33-2.29
0.*2.q
0.4r-2.82
0.50-3.24
0.58-3.63
0.67-3.99
0.75-4.32
0.83-4.59
0.89-4.81
0.9+4.96
0.98-5.04
1.00-5.05
t.0L-4.9
1.00-4.87
0.n-4.67
0.9+4.43

Table 4.10 140
MEAN REI,ATIVE PERCENTAGES OF IgG STJBCLASSES IN SERI]M AT DtrT.ERE}IT AGEs
Age IgGl rñ2 IgG3 IgG4
Cord
46 mo.
7-9 mo.
10-12 mo.
1yr
2yr
3yr ì
4yt
5yr
6yr
7yr
8yr
9yr
10 yt
11n
12yr
13 yr
t4w
15 yr
Adult
63.88
73.ß
78.83
76.07
74.?Á
72.99
70.04
74.L9
70.ß
7L.n
7r.63
65.%
68.01
67.29
63.97
68.50
63.90
68.82
68.L1_
57.28
?3.41
L6.57
13.32
12.75
18.07
16.74
20.26
1:6.45
L9.32
18.36
L8.A
22.70
L9.89
20.M
?r.ß
L7.02
24.02
L8.29
23.N
30.n
6.76
8.38
6.52
7.60
5.7t
5.4{t
5.41
4.78
6.10
5.75
5.43
5.80
5.7L
4.97
5.93
5.62
5.91
5.35
5.39
6.39
5.57
1.59
t.32
3.57
3.05
4.89
4.n
4.57
4.09
4.L9
4.73
5.33
6.38
7.46
5.93
5.85
6.L4
7.55
4.82
6.26

L4LTable 4.11
PUBLISHED PAEDIATRIC IeG STIBCLASS NORMAL RANGES
Study Ages Number ofcovered child¡en(years)
Method Antisera ReferenceStandard
E>çressionof normalrange
Morellet al,l972b
0-2 95 RIS PC WIIO 67 /97 a)Mean+ SDb)range
Van derGeissene¡ aJ, L975
+12 t4t RID PC Myelomaproteins
a)Meant SDb)rnnge
Schuret al,l979
0-16 281, RID PC Myeloma 95%confidencelimits
Oxelius 1979a 0.1-15 L62 EIA PC WIIO67/97 a)mean+ SDb)range
Shackelfordet al, 1985
(IgGl andIgG2 only)
0.6-L6 rr4 MC(IgG1)Pc(IgG2)
Myelomaproteins
9s%confidencelimits
RIA
Birdet al, 1-985
0.5-5 215 RID MC wfJo 67 /e7 9s%confidencelimits
Aucou¡lurier 1-16
et aJ,L987a128 ELISA MC Mean+ SD
$ingh,1988 0-15 56 RID PC wfIo 67 /97
RID = radial immunodiffusion; RIS = ¡¿disimmunosorbent assay; EIA - elestroimmunoassay;
R[4 = radioimmunoassay; ELISA = enzyme-linked immunosorbent assay.

r42
9/lto
9/t6.0
5.O
4.0
3.0
2.0
r.o
lsGl PERCENTILES - Mate
123456789
, AGE (years)
97.5
95
6
75
50
25
54
2
2-5
o
10 l.t 12 13 14 15
lgG2 PERCENTILES - Male
975
95
75
50
25
5-52
o
1 2 3 4 5 6 7 I I lO 11 12 13 14 t5
AGE (years)
Fig;4.1. þG1 and þG2 subclass percentiles in healthyAustralian male chitd¡en.

t43
9/lr.0
lgG3 PERCENTILES - Male
0.8
0.6
0.4
o-2
1234567A910il1213t415
AGE (years)
lgG4 PERCENTILES - Mole
3.O
2.
1.0
75
s/l25
5
2-5
o-o1
1 2 3 4 5 6 7 I I 10 l1 12131415AGE (yeors)
Flg.42. þG3 and þGlzl pertentiles tn heatthy australian male chlldren.
97-5
95
75
5025Ê
2.5
9795
5
0
o

r44
s/l lgGl PERCENTILES - Female
10
I
6
4
2
97. 5
95
75
50
25
2.5
1 2 3 4 5 6 7 I 9 10 11 12 13 t4 15
g/l
6.O
5.0
4.O
2-O
r.o
AGE (years)
lgG2 PERCENTILES - Female
97.5
5 6 7 I I 10 l1 12 1314 t5
AGE (years)
95
3-0
75
50
25
5
2-5
o
1234
Fig. 43. þGl and þG2 percentiles in healthy Australian female chitdren.

L4s
s/l1.2
r.0
0.8
0.6
0.4
o.2
lgG3 PERCENTILES - Femate
12345678910
AGE (years)
lsG4 PERCENTILES - Femole
97.5
95
75
50
25
5
25
0
'tl 12 13 14 15
3.0
2.O
r.097-595
s/l
5
2.50.01
o.ool1 2 3 4 5 6 7 I I 10fl 12131415
AGE (yeors)
Fig.4.4. þG3 and þGlf percenüle ranges tn healthy Australlan female children.
75
50
25o.1

s/l
10
146
B
6
4
2
lgGì PERCENTILES
AGE (yeors)
lgG2 PERCENT¡LES
97.595
75
50
25
52.5
o
1234567891011 12 13 14 15
s/l
5.0
4.O
3.O
2.O
1.O
97.5
95
75
50
25
52. 5
o
1 2 3 4 5 6 7 I I 10 11 12 13 14 15
AGE (yeors)
Fig.45 þGl and þG2 percentiles in healthyAusfratþn chll{¡ç¡ (males and females combined)
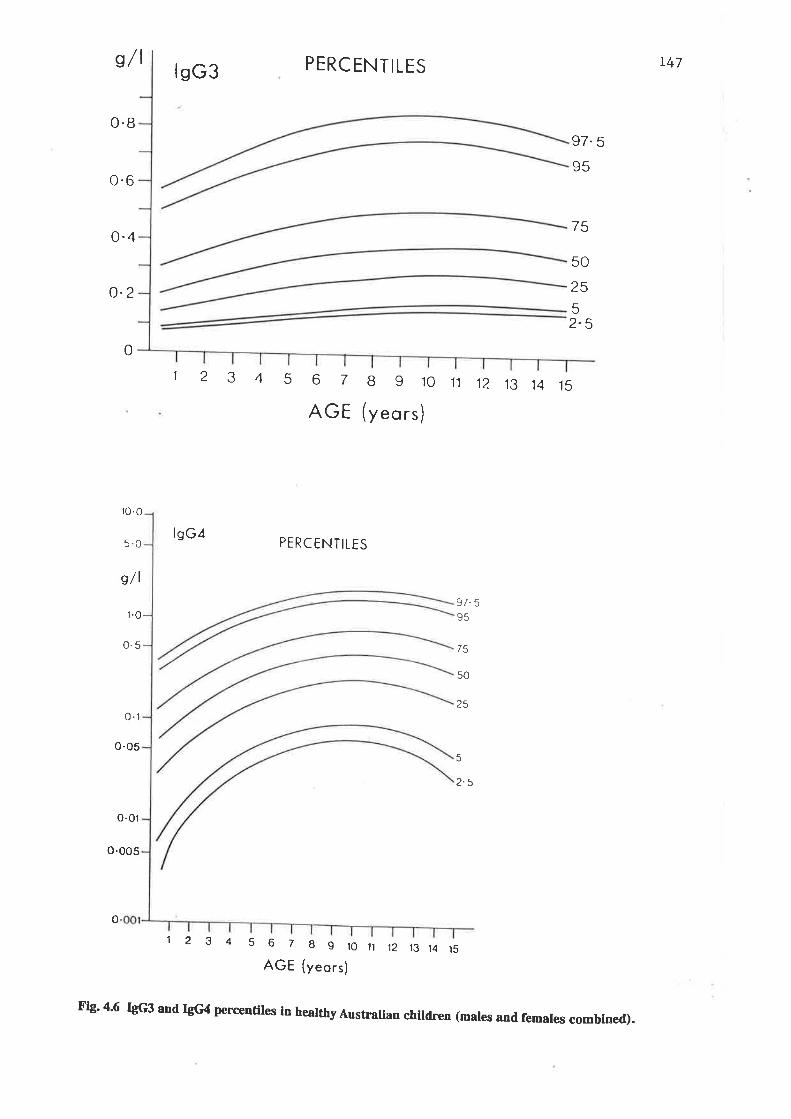
s/l
0.8
0.6
0.4
o.2
10.0
5.0
s/l
r.0
0.5
o-l
o-os
o-01
o.005
PERCENTILES
AGE (yeors)
PERCENTILES
r47lsG3
lsG¿
97.s
95
75
50
25
52-5
0
12345678910t112 13 14 15
75
50
25
97-595
2.5
5
o
12 3 4 5 6789loll12t314t5
AGE (yeors)
F¡g' 4'6 þG3 and þG4 percentiles in heatthy Australian children (mates and females combfned).

r48
B
lsGl 6
(g /l) 4
lgG2
(g/t\'
o-8
o.2
2¿-t-t-'
2
46810121416
-- -+-
AGE (yeors)
.:':- -
hr.t ñP2 5 Ma¡e
P2.5 Femaþ
P2'5 Male
P2 5 Female
4
4
6 810AGE (yeors)
12 14 16
o.6lsG3
(g/l) o'¿
P2 5 Femal€
P2. 5 Male
68 10 12 14 16
AGE (yeors)
--
Schu¡ et aJiLng?. ... ., .. " Oxelius 1979a
Shackelford et a[ 1985
-.-.- Bird et 4 1985.Ê - <- This study males
-
This stud¡ femalqs
Fig.4.7. r.ower limits for þG subclasses in published paediatric stud.ies:

lsGì
(g/t\
lsG2
(s/lJ
leG4
b/tl
B
4
r49
Aucouturier et aJ, L987 a
Schu¡ et ^1,L979. . ... Oxelius, L!79a
Shackelford et al 1985
Bird et al, 1985
810121416
6
2
0.8
o.2
l,o
o.8
o.6
2
4 6
3
4 6
AGE (yeors)
AGE (yeors)81012 14 16
l--;"-t-"=--
0.6lsG3
G/t\ 0 4
6 810124 14 16
AGE (yeors)
0.4
Singb,1988
46BlOtZ1416AGE (yeors)
Fig.4A. Mean or median concentrations for þG subclasses in published paediafric studies.
o.

CHAPTER FTYE
IgG SUBCLASS DEFICIENCIES IN IgA.DEFICIENT PATIENTS

151
5.1 INTRODUCTION
As outlined in Chapter 1.5, IgG subclass deficiency may perhaps be an important factor in the
infection proneness of some IgA-deficient patients.
Although it has become widely accepted that IgG2 deficiency is the IgG subclass deficiency most
commonly associated with IgA deficiency, in the lþht of recent studies this idea must be questioned. One
reason why IgG2 "deficiencies" may have been reported more commonly in the past may be the use of the
normal ranges defined by Oxelius (L979a) to define deficiency. As has been shown in Chapter 4, Oxelius'
normal values fq¡ rhis isotype are generally highs¡ than those determined by other investigators. Hence,
some values that would be considered normal according to otler normal ¡anges would be considered
subnormal according to Oxelius' ranges.
Since the findi"g of an IgG subclass deficiency in an IgA-deficient patient who is suffering from
recurrent infections is now considered by many as an indication to t¡eat that patient with immunoglobrrlin
replacement tlerapy, it is very inportant to be able to determine accurately whether there is an IgG
subclass deficiency in IgA-deficient patients. Under-estimation of subclass deficiency in such patients may
result in under-treatment, with some patients developing severe incapacitating infective disorders and the
coruiequences thereof. Over-esfimation of the frequency of subclass deficiency may lead to unnecessary
treatment with inLravenous immunoglobulin infusions with their attendant high time and financial costs,
and risks to the patient of anti-IgA mediated anaphylactic reaction or, more remotely, of nonA nonB
hepatitis (Bjorkander et al 1988).
The incidence and clinical siguificance of IgG subclass deficiency in IgA-deficient subjects ¡smains
to be determined. Although IgA deficiency is relatively common in healthy subjects (L500) it is much more
common amsngst patients with recu¡rent respiratory infections and atopic diseases. In order to gather
more information about the frequency and significance IgG subclass deficiency in IgA-deficient patients we
did two studies in IgA-deficient subjects, quantitating IgG subclasses and examining their relationship to
infection proneness.

r525,2 PATIENTS AI\D METIIODS
In a preliminary study (Beard et al 1-986) IgG subclass concent¡ations were measured by
electroimmunoassay (EIA) with polyclonal antisera by Dr Vivi-Anne Oxelius in Lund (Oxelius, 1978). Itr
addition to studying imm¡¡sglsþulin concentrations, we studied the patients for deficiencies of neutrophil
function and complement activity, which may have been contributing to increased infection proneness. In a
fu¡ther study (Beard and Ferrante, 1989) IgG subclass concent¡ations were measured by an ELISA and
using McAb. In rhis study, neutrophil functions and complement quantitations were not studied as the
initial study had not revealed deficiencies in these parameters.
I *". initial study, twenty-two children found to have apparently selective IgA deficiency, and
presenting witl recurrent alidf or severe respiratory infections were studied. Their ages ranged from 0.4 to
13.8 yr, with a mean of 3.5yr. TWelve were males and ten females. Seven child¡en had unusually frequent
upper respiratory tract infections, eg. sinusitis, tonsillitis, pha4mgitis, otitis media and ill-defined upper
respiratory tract infections (UTRIs), eight had ¡ecurrent lower respiratory infections, and seven in addition
to multiple respiratory tract infections, had serious or unusual infections in other sites. None of the
children had cystic fibrosis, gastroesophageal reflux or other adequate explanations for their repeated and
oft en severe infections.
In the second study seventy-three patients who were referred for immunological assessment and
were found to have concentrations of serum IgA more than 2SD below the mean for age were included. In
only four patients was IgA undetectable, ie <0.0L9/1. The ages of the patients ranged from 4 months to 60
years with a mean of 8.7 years. All but five were less than 18 years of age. Patients were grouped accorrling
to their medical histories obtained from referring medical practitioners, case notes and directly from
patients or parents by the investþators. One group was comprised of 50 patients, who had had recurrent
infections such as sinusitis, tonsillitis, otitis media, pneumonia and who were considered by thei¡ medical
practitioners to be excessively infection-protre. Many of these patients were t¡eated outside the hospital
and details of infecting pathogens were not always available. The other group, was comprised of 9 patients
who had recurrent cough and wheeze, not considered primarily infectious in origi¡r, and 14 patients with a
variety of otler conditions including celiac disease, lichen sclerosis, alphal antitr¡'psin deficiency and
epilepsy.

153
5.3 RESTJLTS
Results from the first study are summa¡ised in Table 5.1. At least nine of the 22 children were
found to have IgG subclass deficiency (deficiency here being defined as below ttre lower limit of the age-
related normal rango: Oxelius, L979a). Of these nine, snly one had a very low serum IgA concentration.
The others had less markedly reduced IgA concentrations (Fig. 5.1). Six children had undetectable
concentrations of serum IgA (<0.01-9 /l) and none of these had a detectable IgA subclass deficiency. One
qths¡ child had a very low serum IgA concentration (0.069/l) and no IgG subdass deficiency (Fig. 5.1). If a
serum IgG4 concentration of <0.01-g/l were considered as a deficiency, it was found that in the group with
markedly low IgA (<0.069/l), tlree showed IgG subclass deficiency while five were normal, and in the
group with slþh{y low IgA (slightly more tlan 2SD below the mean) L2 showed IgG subclass deficiency
while two were normal. These results suggest that IgG subclass deficiency is more likely to be associated
with slþhtly rather than with markedly low IgA concentrations (p=0.05). Most children with IgG subclass
deficiencies had total serum IgG concentrations close to 2SD below the mean (Fþ. 5.1-). None of the
children studied had serum IgG coucentrations more than the 2SD above the mean. Serum IgM
concentrations appeared to bear no relationship to tle presence of IgG subclass deficienies (Fig. 5.1).
Serum IgE concentrations also appeared u¡¡elated to any of the IgG subclass concentrations (data not
presented).
IgGL deficiency was tle most cornmon IgG subclass deficiency, occurring in six patients. In tl¡ee
it occurred as an isolated deficiency (p.9, p.D, and p.15; Table 5.1; Fig. 5.2), in anotler it was associated
with an IgG2 deficiency (p.L8; Table 5.L; Fig. 5.2) and in two with IgG4 concent¡ations of <0.01g/l (p.4 and
p.11-; Table 5.1; Fig.5.2).
Serum IgG2 subclass deficiency was found in four children. In th¡ee it was associated with a
deficiency of either IgGl. or IgG3, or with undetectable IgG4 þ.18, p.20, and p.3; Table 5.1; Fig. 5.2).
Thrce of these children also showed total IgG concentrations nearly 2SD below the mean for age.
Nine of the ?2 patients had serum concentrations of IgG4 unmeasurable by the EIA and these
included all seven child¡sa who were 3 yr old or less. Fou¡ of them had low normal IgE concentrations
(Table 5.1).

Table 5.1 Ls4
STIMMARY OF CLINICAL AND IMMTINOI,OGICAL ABNORMALITTESIN 22 IgA-DEFICENT CHILDREN
Patient(p.) Clinical problems Immunological abnormalities
1
)3
45
6
78910LI
1213
18
L9
20
2L22
L415
L6L7
URTIS, salmonellainfection, (JTILRTISLRTIS (shuntedhydrocephalus)LRTISLRTI, salmonellaand moniliainfectionsLRTIS, URTIS,asthmaURTIS, PUOLRTIS, URTISURTTSURTISURTIS, LRTIS,pneumococcal6gningitisLRTIS, URTISURTIS, herpessimplexLRTIS, URTISURTIS, as'hmaURTISLRTIS, URTIS,1¡¡6 psningrtisLRTÏS, URTIS,hay feverLRTIS, bronchiectasis,URTISURTIS, infectiousmononucleosis, IJTILRTIS, URTISURTIS, ¿sthm¿,herpes zoster
I IgG4, I bactericidal activity
+ IgA" + IgG4+IgA +Iñ2,+tñ4
+ IgGL, + IgG4t IgG4, I chemotaxis
lIgG4,lC4
+Igd +IgG4I rgE+ IgGl{C4,lgG2low normalI IgG1, + IgG4, l IgE
I IgGL, I chemotaxis+IgA
{.IgG4+IgG4r IgGl+lgGz,l chemotaxis
t IgG1, + IgG2, l IgE,{ bactericidal activity, + C+r IgA, l IgE
tIgG2,+IgG3
+IgA+IgA lIsE
* URTIS, upper respiratory tract infections, e.g. otitis media, tonsillitis, sinusitis; UTI, urinary tract
infection; LRTIS = lower respiratory tract infections; I decreased (see data in Figs); 1 increased; + IgGaindicates an IgG4 concentration of <0.01¡ng/l; +lg^ indicates <0.06mg/1.
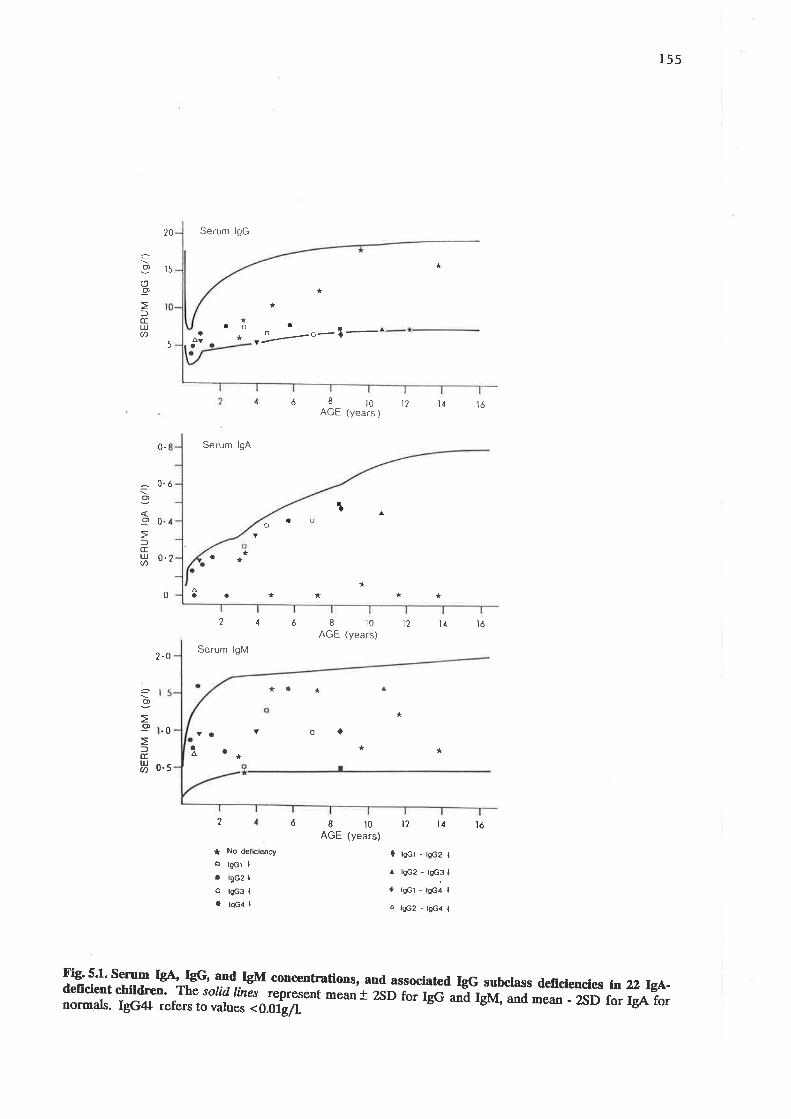
155
20
t5
Serum lgG
*
a
^Vao o-l-
Y
=o(,-9
)(I[!U)
-o)
-9
l(ILrJU)
0
0.5
âo
I
f,îLUJU)
*
*
¡0 l2 ì4
aa*q
*5
Serum lgA
AGE (years )
I r0AGE (years)
ó
6
tó
0.
0.ó
n1
oa
*
o
0.2
2.0
*a*
0 *
216
Serum lgM
*a *
ôa a
Y.a^
v
*
* No def¡cieocy
s lgct I
. l9c2 I
ô lgc3 I
a lsc4 I
*
8ì0AGE (years)
*
ì2 ì1 ìó
t2 t4 tó
o<)
*
*
0 rscl - tsc2 I
^ lgc2 - lgc3 I
I rgcl - tgc4 I
Á lgc2 - tgc4 I
*a
2
F¡g' 5'1' sen¡m IgA IgG' and IgM conoentrations, and. associated IgG subctass deficiencies ln zz W-deficlent chlld¡en' The solid /rnãs represent mear+ 2sD for rgc- -¿ Iglvf, and mean - zSD for IgA fornormals. IgG4+ refers to values <0.0ig/1.

156
qJ'dl
lgGl
m9/d
'¡{orÉ2
É:eaJfg
ot
6ao
É
ãa
1
2b
É
3c3
16z
bt
I€t2 o ót o12(
MONTHS Y€^RS MOô¡fHS YT ARSîEtdl
sllCGa
ñ9/dl
a O¡6raalourl¡or¡THs YEAÂS r¡oa{Txs YEAßS
F¡9.5¿. Serum þG subclass concenhations ln 22 child¡¡n with ¡rcur¡nt and or severe rcspiratoryinfections. The solid lines represent mean and normal range for age for IgGl, IgG} and IgG3, and meanplus upper limit of range for IgG4. CircIes represent sorum concentrations in patients atd semicirclesrepresent concent¡ations that were <0.019/1.
r¡
$)CT
Þ
oI
a
6

r57
A plot of IgG subclass concentrations against normal concentrations showed that t6,L7,15 and 1,7
respectively of the 22 patients had IgG1, IgG2,Igp3, or IgG4 concentrations below tÏe mean values for age
(Fig. s.2).
No patients demonstrated defects of either neutrophil iodination activity or fungicidal capacity
(Fig. 5.3). Three child¡en showed reduced neutrophil chemotaxis (P.5, p.12, and p.17) and at least two of
these were also IgG subclass-deficient (p.12 and p.17; Table 5.1; Fig. 5.3). Serum C3 and CH5g were within
normal limits in all patients. Reduced concent¡ations of C4 occurred in three children (p.6, p.1-0, and p.L8)
and one of these had an IgG subclass deficiency (Table 5.1; Fþ. 5.4). None of the children with extremely
low IgA concentrations had defects in neutrophil function or deficiencies of complement.
Results in the second study showed that IgG subclass deficiencies occurred n 37Vo of the 73
patients, when concent¡ations below the 5th percentile for age were considered deficiencies. lgG4
deficiency occu¡red n 26% of patients (p<0.001), and was by far the most common IgG subclass
defrciency. lgGt,lgp2 and IgG3, deficiencies occu¡red in 10 and 12 andSVo of. patients, respectively (n.s.)
(Table 5.2). None of the patients had a generalised hypogamm¿globulinaemia.
Subclass-deficient patients were classified according to clinical histories (Table 5.3). Twenty-three
of the IgG subclass-deficient patients were f¡om the group of 50 patients who had recurrent infections.
Four we¡e from the group of 23 who had other problems' There was a significantly highs¡ incidence of
IgG4 subclass deficiency among those with than among those without recurrent infections (p<0.0Ð (Table
s.4).
Absolute concentrations of IgG4 were compared in patients and in normal control subjects (Table
5.Ð. h each age gouping there was ¿ 5ignifiç¿¡tly highs¡ incidence of patients with Igg4 concentrations
below an arbitrarily chosen value of 0.\3g/lthan of conhols. This occurrednTs% of patients and?ß% of.
cont¡ols aged from 4 to 72 montls (p < 0.02), n 47% of patients and L4% of controls aged from 1 to 2 years
(p<0.02) and in 20vo olpatients andtvo of controls aged3 years and over þ<0.001).

158
U>
o(J
Í\.
IØ_9oEo.
oLUl-E.oo-ú.oC)1
r/ls
IOOINATION
o
CHEMOTAXIS
4
8
ôô
aloccîtrE
zat--
Í.I
o
2-5
2-O
t-5
l.o
o.5
loo
40
12
t0
I
6
4
2
É&ô
ôô0 aao- ----
aa
ô
ôa
1é b.o
FUNGIC¡DAL
a
BACTERICIDAL
aaao
ot
IOO
80
A
1" o3
â
óôô
tta
a
80oLl-'J¿YæèS
a
otlJJJ60Ys
ô
¿O
20 20
Fig' 53' Neutrophll functÍons in þa-deficient childr,en with recurrent or sevene respiratory infec{ions.Patients are grouped into thosesho*i"g rgc lyFl* d"il";.y ó), very low IgA levels (o), both of these
fi)¿ï.*tther of the above (o). ní" íot¡¿ line representr áJ- *d th" uiokcnn* ì æp of healthy

159
2-O
g/t
I.O
0.6
O,4
g/l
O,2
200
t50
units/ml
fu &r*
C¡oö
o
C¿Aô
8o
ôA 8o
-r---$ô
oa
a
-t-a
CH 50
¿ôÁ
o
O-r
3-¡o
ôô
-fÁa.¿r
^^atoo
50
nd C4, and haemotytÍc complement acdvltysevene rcsplratory lnfecdons. patients arelow IgA levels (O), both of thase ( I ), or

TaÞle 5.2
study
oxelius et aI198 L
cunningham-Rundleset al, L983
Bjorkander et aI,1985
INCIDENCE OF IgG SITBCLA.SS DEFICIB¡CIES IN SYIIPTOUATIC I9A-DEFICInÛI PÀTIEN1IS
No. ofPatients
IgGLNo. t
LgG2No.
IgG3No. Z
IqG4No. I
<L7* <47
notstated
<8 <22
zMethod ofMeasurement
PC
PC
EIA
EIA
PC
MC
RTD
RID
PC EIÀ
MC RTD
PC RID
MC RrA (T9GA)
MC ELISA
Nu:nber) ofControls
182(1)
182(1)
2T
449
182(1)
not stated
1s2(1)
35
361
7L9 <10* <27 Pc ErA 182(1)37
39
29
36
66
5
8
6
3 82
Ugazio et aI,
Plebani et aI,1986
t_98 3
(2) ,
tI3
310
925
58
Beard et al,(lst atudY)ChandrÀ, l-986
1986 22
2L
L988 60
6
7
27 <g* <4L
5
<8
56
26
l-8
24
7
70
L2
4
5
4
55
9
5
7
1
4
3**
_*<5
1
Klemola et aI,
Heiner, 1986
Beard & Ferrante(second studY),
4479
731e8e (3)
10 6819
***(r.(2(3
below detection range Àbbreviations:<0 . oo6llitre') oxelius. L979a) i""i"a"å tot" patients from Ugazio et a1,-1983) il; not incluãe patients fron Beard et al | 1-986
using polyclonal antiserausing monoclonal antiseraradiãI i¡nmunodiffusionelectroimmunoassaYenzlnne-linked imrnunosorbent assay
PC=MC=RID =EIÀ =ELISA = F
OìO

Ta-ble 5.3
Combin f i onc af rìef i r':ì enr:i es
IgA-IgG4
IgA-IgG2-IgG4
IgÀ-IgGL-TgG2-IgGA
IgÀ-IgGl--I94
rgÀ-IgGL
IgA-IgG3-IgG4
rqA-LgG2-rgÊ3
IgA-IgG3
TgA-19G2
IgA-rgc1-IgG3
IgÀ-IgGL-IgG2-IgG3
Total number of Patients withan IgG subclass deficiencY
DTAGNOSES DI 27 IgA-DEFICIENT PÀTIENTS I{IIE IgG SIIBCLASS DEFICIENCIES
of nati ents Di aonosesNumbe
1,2
27
L0t-t_
recurrent infectionsrecurrent feversrecurrent cough, wheeze
1 recurrent URTI1 bronchiectaslsl- recurrent sinusitis cbronchitis
1 sinusitis, bronchiectasis1 sLE, recurrent infections
recurrent infections
I asthmal- recurrent infections,arthritis
recurrent El{T infections
1 recurrent infectionsL recurrent tonsillitis,asthma
chronic cough
SLE & recurrent infections
recurrent bronchitis
recurrent URTI
3
2
1
2
1
2
1
I
t
1
URTI = Upper respiratory tract infection; SLE = systemic lupus erythematosus; EiIT : ear¡ nose and throat 3

Table 5.4 162
Igc4 DEFICIENCY IN 73 IgA-DEFICIENT PATIENTS WIIH Al{D IITIHOUT RECURRENTINTECTIONS
IsG4 deficient Not IsG4 deficient
With infections
Without infections
33
2t
17
2

Table 5.5
ABSOIJITE CþNCEN,:I1¡ÀTIONS OF I9G4 IN 73 IgÀ-DEFICIENT PATIENTS Al{D IN AGB-IIATCEED cOlflIR.oL SUÈrEefS
Age of subjects
4 to L2 monttrs 1 to 2 years Over 3 years
TgG4Concentrations(q rlitre I
<0.005
0. 006 - 0. ol-0
0.011 - 0.030
0.031 - 0.06
0.061- - 0.60
0.61- - 2.00
>2.OO
PatientsNo. I
Controls INozNozNoPatients Controls Patients Controls
No. I No. Z
l_
2
4
2
6
0
0
3
4
2
3
0
0
0
25
33
L7
25
0
o
0
t-
2
9
T6
L4
o
0
2
5
2L
38
33
0
0
0
3
t_ l_
15
6I
10
0
0
0
2
4
199
53
3
7
13
27
t3
40
0
0
0
2
7
9
38
6
0
1
3
5
2
2A
7
0
2
7
t1
4
61
l_5
0
0
0
t_
2
76
20
t
L2 L00 42 100 15 t-00 62 100 46 L00 26L 100
tso\(,

164
5.4 SIJMMARY
In the preliminary stud¡ a group of 22 children presenting with recu¡rent or severe respiratory
tract infections who had low IgA concentrations (more than 2 SD below the mean for age) were examined
for IgG subclass deficiency. Patients rvere screened for possible defects in neutrophil chemotaxis,
bactericida! fungicidal and quantitative iodination activity, as well as for complement function. The
majority of the patients showed IgG subclass concentrations below the mean for age. Nine of the child¡en
showed d.efinite IgG subclass deficiency and at least two showed definite deficiency of more than one IgG
subclass. The predominant subclass deficiency was found to be IgG1. While nine children showed IgG4
concentrations below the level detectable by the technique used" it was not possible to assess whether these
patients were deficient in this isotype since some healthy subjects also gave values below the level of
detection. Most of the patients who had very low (0.0L{.069/1) or undetectable (<0.0L9/l) concentrations
of serum IgA did not show IgG subclass deficiencies, while IgG subclass deficiencies were cornmon among
those with borderline low IgA concent¡ations (slightly more than 2 SD below the mean for age). Nine
children showed total IgG concent¡ations close to 2 SD below mean for age, and at least six of these
showed IgG subclass deficiency. The results suggest that patients with recurrent andf ot severe respiratory
infections who have borderline IgA and IgG concentrations may be more likely to have IgG subclass
deficiencies than those with very low IgA concentrations.
Like the above stud¡ most studies on IgG subclass deficiency in IgA-deficient subjects have been
unable to determine the incidence of IgG4 deficiency because the limitations of the assay metlods used
have often made a distinction between low-normal and sub-normal concentrations impossible (Table 5.2).
A more sensitive assay based on ELISA was used in a second study to attempt to resolve some of the
problems. Here, we measu¡ed IgG subclass concentrations in 73 IgA-deficient patients, the majority of
whom were children with recurrent respiratory infections. We found IgG4 deficiency occurred n26Vo of
the patients, and this was tle most common IgG subclass deficiency. There was a significantly increased
incidence of IgG4 deficiency in those patients with recurrent infections. IgG1, IgG2 and IgG3 deficiencies
occurred" respectivelS ta, ITVI, 12Vo and 8% of. the patients. Ig^-lÑ4 deficiency occurred n t6Vo of the
patients,IgA-IgG2-IgG4tn4Vo, andIgGl-IgG2-IgG4 IgA-IgGl and lgé^-lÑ2-lÑ3 each occurredn3%o.
Other subclass deficiencies or combinations of deficiencies were even less frequent. These results suggest

165
that IgG4 deficiency, even in the absence of.lgp2 deficiency, may be an important, but hitherto largely
unrecognised fac¡or in infection-proneness in some IgA-deficient patients.
(1)
(2)
(3)
The results in these two studies ¿tre not strictly comparable because:-
lhe ?2lgA-deficient patients in the first study were all children investþated because of recurrent
respiratory infections while the 73 IgA-deficient patients in the second study included not only 50
patients with recurrent respiratory infections, but also 9 patients with probably non-infective
recurrent cough and wheeze and 1-4 patients with a variety of otler disorders.
The methods of measu¡ing IgG subclasses differed. In the füst study, these were measured by EIA
using polyclonal antisera raised in rabbits, while in the second study IgG subclasses were
measured by ELISA using monoclonal antise¡a.
The normal ¡anges used in the füst study were those established by Oxelius (1979a) where the
cutoff point for the lower limit of normal at any given age was the lowest value recorded in a
healthy Swedish subject at that age. The incidence IgG4 deficiency could not be ascertahed
because due to 1¡s limited sensitivity of the assay and the fact that IgG4 was undetectable in
considerable numbers of healthy subjects. The normal ranges used in the second study were those
determined in ou¡ laboratory r¡sing our ELISA assay and sera from healthy Australian child¡s¡.
The cutoff point used for the lower limit of normal was tle 5th percentile for age.
The most l¡1e¡s5ring fiodiogs in the füst study was that IgGl deficiency was found nTlVo of The
patients and was the most commonly found IgG subclass deficiency, and that IgG subclass deficiency
occurred more commonly in patients with moderately reduced IgA concent¡ations than in those with
undetectable concentrations of IgA. In the second study, we found IgG4 deficiency to be the most oommon
IgG subclass deficiency, occurring n?ß% of the patients, while IgGl deficiencywris found in only tÙVo. ln
fact, as many as 4wo of patients in the first study had undetectable levels of IgGa, but it is not possible to
say how many of these were actually IgGzl-deficient. In both studies, the incidence of IgG2 deficiency was
comparable, tSVo and 12Vo rcspectlely. Very few patients in the second study had undetectable
concent¡ations of IgA and we were unable to draw msaningfrrl conclusions about the severity of the IgA
deficiency and the likelihood of a concurrent IgG subclass deficiency from this study.

r66Overalt ou¡ studies of IgG subclass concentrations in IgA-deficient patients confirm the work of
previous studies (Table 5.2) which indicate an increased incidence of IgG subclass deficiency amongst IgA-
deficient patients. Ou¡ work suggests that IgG4 deficiency may be the most common IgG subclass
deficiency in IgA-deficient patients, rather than IgG2 deficiency as has been previously suggested. [n our
second stud¡ we found that those IgA-deficient patients who were excessively infection-prone were more
likely to have an IgG4 deficiency than tlose who were not. We do not yet know whether or not IgA-
deficient healthy subjects have an increased incidence of IgG subclass deficiency and specificall¡ of IgG4
deficiency. A study of such subjects would be helpfuI in determining the possible role of IgG4 deficiency in
the infection proneness of some IgA-deficient subjects.

CHAPTER SfX
IgG SUBCI"ASS DEFICIENCY IN PATIENTS WITH BRONCHIECTASIS

168
6.I INTRODUCTION
Bronchiectasis is a condition cha¡acterised by dilatation of bronchi associated with inJflâmmatory
destruction of bronchial and peribronchial tissue, accumulation of exudative material in dependent bronchi
and sometimes distension of dependent bronchi. Most bronchiectasis is postnatal in origin, and results
from chronic pulms¡ary infection. The mechanisms involved in its causation are poorly understood.
Several predisposi"g caüses such as bronchial obst¡uction of various etiologies (e.g. foreign body,
luberculous nodes) followed by infection, cystic fibrosis, recurrent and chronic lung infections, measles,
pertussis, pneumonia (rarely now), sarcoidosis, neoplasms, lung abscess, localised cysts, recurrent
aspiration pneumonitis, imms¡jls cilia slmdrome and va¡ious t¡res of immunodeficiency, e.g. X-linked
agammaglobulinaem.ia have been recognised (Stern 1983; Phelan et at,]1982).
Despite this multitude of known predisposing factors, many patients who develop bronchiectasis do
not have any such recognised predisposing factors. Most of them have ¡aised levels of inmunoglobuli"s
(Horan et at, 1984). While tlere have been a number of reports of patients with IgG subclass deficiencies
and chronic or recurrent lung infections (Schur et ú" L970; Oxelius et al 1-981; Beck & Heiner, 1-98L;
Heiner et al, 1983; Ugazio et al 1983; Stanley et al, 1,984; Ileiner, l-984, Smith et aJ, 1984; Umetsu et al
L985; Bjorkander et af 1985; Plebani et af 1986; Shackelford et al 198ó), relatively few reports have
specifically identified patients with bronchiectasis. Because there is a paucity of information on immune
host defences in patients with bronchiectasis, we measured serum immunoglobulin class and IgG subclass
concentrations, and llm.phocyte and neutrophil functions in a group of such patients.
62 PATIENTS
We studied 15 patients with chronic suppurative lung disease in whom bronchiectasis had been
diagnosed by radiological studies and/or bronchoscopy. Thorough previous investþations had excluded
known causes of bronchiectasis. AII were referred to the Immunolog5l Department at the Adelaide
Children's Hospital for immunological assessment. Nine of the patients were over and six under 16 years of
age. Adults were included in this study because it was thought that if they showed a consistent IgG subclass

L69
deficiency pattern this may be an indication for seeking this pattern in children vift significant respiratory
infections and possibly i¡¡s¡vening prophylactically.
6.3 RESULTS
Nine of the 15 patients (60Vo) had a deficiency of at least one immunoglob'rlin isotype (Table 6.1).
Fou¡ had an IgA deficiency, but in fwo of these this deficiency was only marginal. Of these, one child þ.3)
and one adult (p.Íl) had reduced concentrations of IgA only. Another child @.a) had a reduced
concentration of Igd a reduced concentration of IgG Ø.72g/L; N.R. 6.¿A-17.Lg/l) aú a deficiency of IgG2
and IgG4. One adult (p.8) had a borderline low concentration of IgA" a reduced concentration of IgG
(+.i6e/\, N.R. 6.¿A-17.28e/D, a borderline low concent¡ation of IgM and a deficiency of IgGZ and IgG4.
Two patients (p.1-0 and p.15) had reduced IgGl concentrations in tle absence reduced concentrations of
total IgG. One of them had a high relative proportion of IgG2 and the other of IgG4. One child (p.2) had
a reduced IgG2 concentration with raised total IgGl and IgG concentrations. Another child (p.1) and
another adult (p.7) had reduced concentrations of IgG4 only'
Seven patients @7%) had a deficiency of at least one IgG subclass @<0.05). The types of these
subclass deficiencies are showu in Table 6.2. Fou¡ Q1%) of the patients had deficiency of IgG4 (p<0.00L)'
and three (20%) had a deficiency of IgG2 (p<0.0Ð. Th" incidence of IgGl deficiency was not statistically
significant and no IgG3 deficiency wÍrs detected. For adult patients analysed as a group, the mean IgG
subclass values of patients did not differ significantly from those of healthy controls (Table 6.3)' Similar
comparisons could not be made for children because of inadequate numbers at any particular age.
In one patient, p.8., we had the opportunity to study the pneumococcal antibody respome.
Immunization with L4-valent vaccine showed a lack of responsiveness to each of the 14 pneumococcal
serotypes included (Table 6.a). Previousl¡ this patient had been found to respond normally to
immunizationwith tetanus toxoid but not to Salmonella typhiH or O (see J.B. Ch. n.2.2.I).
None of the patients had low concentrations of C3 or C4. Three had a raised C3 concentration
and th¡ee a raised C4 concent¡ation. No deficiencies of CHSO were found. There was no difference

170
between patients and cont¡ols in percentages of T and B cells and l5rmphocyte responsiveness to mitogens,
or in neutrophil chemotaxis, ioclinati'on reaction and HMP-shunt activity (Table 6.5).
6,4 STJMMARY
Bronchiectasis is a chronic, disabling infective condition which develops in the absence of known
predisposing factors in a considerable number of patients. Data presented in this study demonstrate
normal lynphocyte mitogen rosponses, neutrophil chemotaxis, iodination, bactericidal and fungicidal
activity and respiratory burst in a group of patients with bronchiectasis. Ilowever, 60Vo of the patients had
¿¡ immunsgloþulin class and/or an IgG subclass deficiency. The most com'non deficieucy was that of Ige
and of tep+ e1h occurring n 2i7Vo of the patients. One patient was studied for the capacity to produce
antibodies, and showed inability to respond to pneumococcal antþens. Defects in humoral immunity may
be more commou than previously recognised in bronchiectasis, and in selected patients, imm¡¡sglsþulin
replacement therapy may warrant consideration.
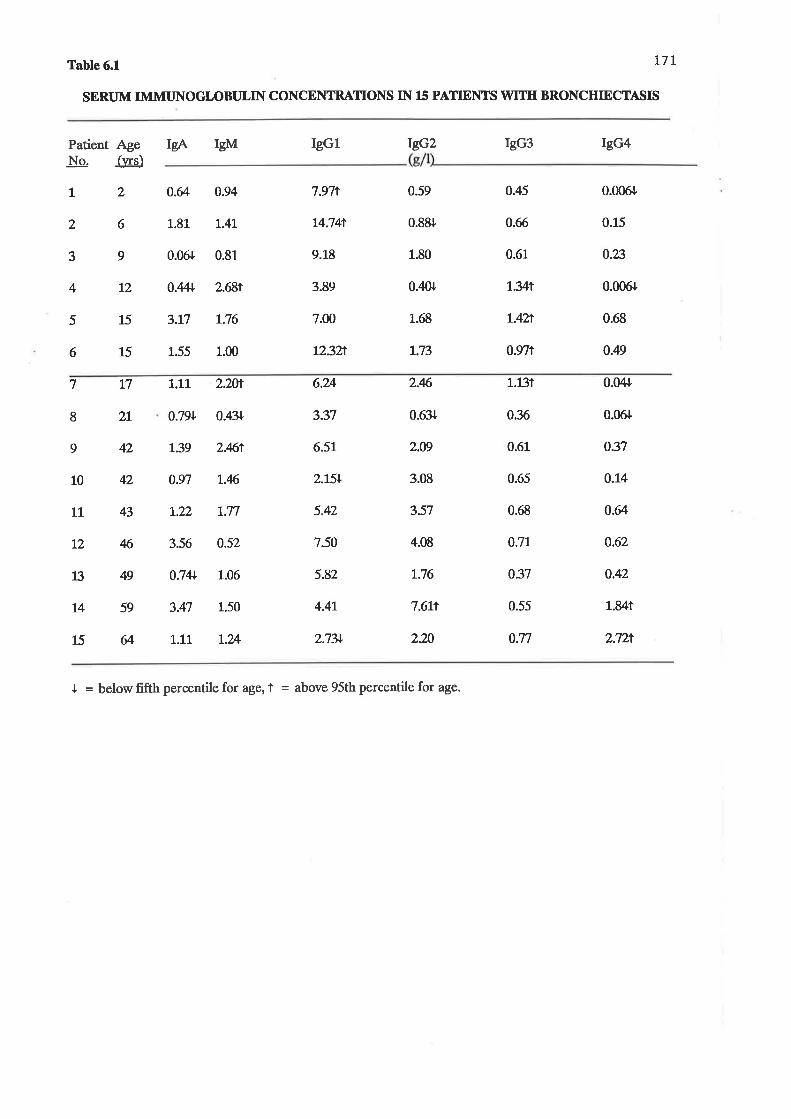
t7rTable 6.1
SERTJM IMMI]NOGI.,OBTJLIN CONCENTRATIONS IN 15 PATTENTS WT['H BRONCHIECTASIS
Patient AgeNo. fws)
IsA IsM ICGl rñ2 IgG3 IgG4
l2 o.&
L.8L
0.06r
0.4+
3.t7
1.55
0.94
l.4L
0.81_
2.681
L.76
1.00
7.97t
L4.74t
9.1_8
3.89
7.00
12.3h
)
3
4
5
6
6
9
12
15
15
0.59 0.45 0.006r
0.88r 0.66 0.15
1.80 0.61 0.23
0.¿lor L.yt 0.006r
1.68 L4Zr 0.68
r.73 0.y71 0.49
7
8
9
L7
2L
42
42
43
46
49
59
il
1.1L
0.7N
t.39
0.97
t.22
3.56
0.741
3.47
r..11
2.2t¡
0.411
2.Mt
L.46
t.77
0.52
1.06
L.50
T.?A
6.U
3.37
6.51
2.15+
5.42
7.50
5.82
4.4L
2.7N
2.6
0.63r
2.t9
3.08
3.57
4.08
L.76
7.6Lt
2.20
L.13t
0.%
0.61
0.65
0.68
0.7L
0.37
0.55
0.77
0.(X+
0.06r
0.37
0.L4
o.Ø
0.62
0.42
1.841
2.72'¡
10
LL
L2
1i
T4
15
I = below fifth percentile for age, I = above 95th percentile for age.

L72Table 62
TYPES Or IgG SUBCLASS DEFICIENCIES IN PATIENTS WIIII BRONCHIECTASIS
IpG subclass
IgG4IgGllgGzrñL/rñ2lrgp3/IgGa
lec4TÑ2rgGLIgG2Iñ3/tñ4
Vo deftctent
Children(n=6)
22n11
4
33L6
33

Table 63
SERUM lgc SUBCLASSES CONCEIì{TRATIONS IN N)ULTS WTIII BRONCIIIECTASIS
Mean* so (e/l)
IgG subclassisotvne
L73
IgGl
rgÊz
IgG3
IgG4
Patients
4.68+ 2.22
2.83+ 2.20
0.65+ 0.23
0.76+ 0.82
Controls
5.n+ 2.N
2.75+ t.43
0.58+ 0.32
0.56+ 0.58
p values
n.s.
n.s.
n.s.
n.s.

Table 6.4 174
RESPONSE TO PNET]MOCOCCAL IMMTJNISATION IN PATIENT 8
Pneumococc¿lSerotype
Antibody ("g/-l) Fold rise post-im m unization
Patient Normalmean(Paton et al, L986)
fm¡ 'nizrtiooPre- Post-
L.73.92.97.23.98.25.86.81.5
5.54.62.74.73.0
0.80.70.92.Lr..8
0.50.70.80.81.0
L.20.40.40.4
1)34678972t418
L9
2325
225188
98333341yL72226322160
120%5LMz2M
267125875696073125L76269154147
267459870

Table 65L7s
LYMPHOCYTE AND NEUTROPHIL STUDIES IN PATIENTS WTTH BRONCHIECTASISAND IN CONTROLS
Function tested
(a) L)¡mphocl¡te Studies
T cell percentage(E rosettes)
B cell percentage(Surfaceimmr¡nsglsþrlin)
Mitogen rerpo^"rlPHAPWMConA
lb) Neut¡oohil Studies
ñ.6+ L.45 57.65+ 2.07 n.s
12.29+ L.70 13.85+ 1.74 n.s
LM769+ 4L675 78L7L+ 42ñ3 n.sß26+ 76Y26 74503+ 26883 n.s9q13+ %0ß 81795+ Y948 n.s.
L.49+ 0.50 1.60+ 0.53 n.s
Patients Controls o values(meantSE) _
5.99+ 0.876.04+ 0.65
9r.o! 2.7988.0+ 1.655.44+ r.O3
NumberTested
13
13
121212
flexosemonophosphateshunt activit¡P
5.63+ 0.746.1-8+ 0.52
91.3+ 2.34
86.9+ 3.L44.28+ L.42
n.sD,.S.
n.s.
n.s,
n.s.
9
99
875
L.
2.3.4.
5.
Expressed as c.p.m.3g-tdn io-rporated into stim$ated lymphocytes.E4pressed as picomoles of øI incorporated per 10 / neutrophils per hourE4pressed as Vo S. aureus killed in 2 hou¡s.E4pressed as Vo T. glabrata killed in 2 hou¡s.Expressed as sfimulation indices.

CHAPTER SEVEN
IgG SUBCLASS DEFICIENCIES IN INFANTS \ryITH IIWASMHAEMOPHILUS INFLUENZAB TYPE B INFECTIONS

L77
7.L IIITRODUCTION
Høemophílus influenzne is a co--on pathogen in yo,t"g children. It is the most common cause of
bacterial meningitis in the first few years of life, causing deaths and neurological sequelae. Invasive H.
influenzal disease is ra¡e after age 4 or 5 years, presumably due to the acquisition of protective levels of
opsonic antibodies to H. influeruae t¡,pe b capsular polysaccharide (Hib CPS) (Peltola et al 1977). It is not
clear why some apparently normal ]orng child¡s¡ develop invasive H. influettzal disease and others do not.
Immunoglobulin deficiency has not been a eornmon finding in infants with H. influenzal infection, although
in an earlier study (Beard and Thong 1981-) in which IgG subclasses were not measured we for¡nd low or
borderline IgA and/or IgG concentrations in over SOVo of a small group of child¡e¡1 with pyogenic
neningitil, predominantly caused by H. influenzte. Recently, with the development of assays for IgG
subclass measurements, tle spectrum of recognisable types of hypoga--aglobuliuaemia has been
broadened. Since the general consensus is that polysaccharide antibodies are preferentially e:rpressed in
specific IgG subclasses (Ch. t.6.I.2), we looked for possible IgG subclass deficiency in addition to
examining lynphocyte and neutrophil functions in a group of children with invasive haemophilus disease'
7.2 PATIENTS
We studied 10 infants admitted to the Adelaide Childrens Hospital with serious invasive culture-
proven Hib infections. Ages ranged from 4 to 16 months. Nine of the children had had at least one
episode of Hib neningitis and one had had two episodes. Another child had had ¡ro episodes of
meningitis, but in only one of these was Hib isolated. One child had Hib septicaemia without nsningitis. A
summary of clinical details is given in Table 7.1-.

Table 7.1 178
CLINICAL DETAII^S OF PATIENTS WIIH HIB DISEASE
Patient (o.) Age(months)¡Ss,
Clinical Details
ffiþ psningifis x 2
Hib meningitis
Hib meningitis
Hib meningitis andpneumonia
Hib meningitis
Hib meningrtis
¡¡iþ ¡eningitis
Hib septicaemia
Hib meningifü x 1(meningitis x 2)
¡¡iþ ¡eningitis,shþella gastroenteritis,boils
t
2
3
4
4
6
8
9
10
11
LL
12
L4
16
5
6
7
8
9
L0

L79
7,3 RESULTS
None of the patients had reduced concentrations of Igd IgG, IgM or of C3 or C4. TWo þ. 1 and
p.5) had raised IgA concent¡ations. Th¡ee (p.L, p.2 and p.10) had raised IgG and raised IgM
concentrations þ<0.001). One had a raised C3 concentration (p.4) and 5 had raised C4 concentrations
(p.4, p.6, p.7, p.8 and p.9) (p<0.0001). TWo patients had raised IgE concentrations (Table7.2).
IgG subclass coucent¡ations a¡e shown in Table 7.3. Th¡ee patients had IgGl- concentrations
above the 95th percentile for age (p.l,p.2and p.10). None had reduced IgGl concentrations. TWo patients
had IgG2 concentrations below the 5th percentile þ.7 and p.8). All but two þ.7 and p.8) had IgG3
concentrations at or above tle mean for age. None had low IgG3 concentrations. Three patients (p.7 p.8
and p.9) had IgG4 concent¡ations at or below the 5th percentile for age. G<0.001). None had raised IgG4
concentrations. All but two patients (p.5 and p.6) had IgG4 concentrations well below the mean for age' In
each of the children who had a low concentrations of IgGZ there was also a low concentration of IgG4.
The mean concent¡ations of IgG subclasses in patients and in age matched control children are
compared in Table 7.4. The only findings of significanoe are a hþber mean IgG3 concentration þ<0'01)
and a lower mean IgG4 concent¡ation þ<0.05) in the ,l-9 montl old patients.
The results of cellular immune function studies are shown in Table 7.5. Patients differed from
controls only in that they had lower percentages of T cells in thei¡ peripheral blood þ<0.01).

Table 72 180
CONCENTRATIONS OF IgAe IgG,IgIVI, IgF'., C3 AND C4IN INFAITITS WTIII NVASM HIB INFECTIONS
Patient AgeNo. (months
IsA IsG IgM IgE* C3 C4
4
6
8
9
1
2
3
4
5
6
7
8
9
10
10
11
L!
12
L4
f6
1.081
0.77
0.¿10
0.26
3.57t
0.77
0.32
0.,18
0.51
L.2T
9.57t
22.tÌ
6.q
7.t9
7.18
5.31
5.20
3.96
3.96
L7.3tî
L.34t
2.18'¡
1.30
0.81
L.23
0.7L
L.09
1.01
r.32
t.7Lt
61
ND
15
272t
7
4
11
8
3
5
L.L9
ND
ND
2.lh
0.99
1.38
L.t6
r.70
r..03
0.99
0.24
ND
ND
0.571
0.19
0.471
0.751
0.6t
0.311
o.n
I = >2SD above mean for age; ND = not done; * = explessed as 1U./nl

181Table 73
IsG SUBCLASS CONCENTRATIONS IN INFANTS WTIII INVASIVE HIB DISEASES
Patient
(months)No IgGl
IgG subclass concentration (g/l)
Age TÑ2 IcG3 rñ4
L
2
4
6
7.8h
11.901 0.84
2.35
4.74
3.82
3.49
3.47
2.53
3.7L
8.361
0.68
0.57
0.84
L.28
0.18.r
0.49
0.?ß
0.4
0.v
0.30
0.043
0.010
0.037
0.0L6
0.?i0
o.2m
0.008r
0.006f
0.,1{l
0.51
0.96
8
9
10
11.
11
12
L4
L6
3
4
5
6
7
8
9
10
o.n+ 0.21
0.38 0.3L
0.67 0.41
0.007+
0.014
I = above 95th percentile for age; { = below 5th percentile for age

182Table 7.4
MEAN IgG SUBCLASS CONCENTRATIONS IN INFAI{TSWIH INVASM HIB DISEASE
To(ì Srhclass Concentration I /l\(months)
IgGl rñ2 IgG3 ICG4
Age
+9
Patients
Controls
(p)
LO-?A
Patients
Controls
(p)
2.80
2.9t
(o.r.)
4.23
3.78
(o.r)
0.83
0.56
(n.s)
0.60
0.91
(o.r)
0.49
0.29
C,>0.01)
0.v
0.30
(n.s)
0.0L9
0.055
(p<o.oÐ
0.093
0.169
(^)

Table 7.5
Function tested
(a) Llmphocyte StudiesT cell percentage(E rosettes)
LYITÍPEOCYTE A}TD NETTTROPEIL STT]DIES IN INFAtflTS WTTE I¡TVASTVE EIB ffFECIIONS AI{D IN COI{IIROIÁ;
(b) Neutrophil StudiesQuantilative iodination2
Pooled SerumAutologous Serum
t"t
B cell ercentage( Surf ace irn:rrunoglobulin )
lfitogen r""porr="=lPHAPWMCo
Bactericidal activiI\rngicidal activity
Patients
52.5+2.8
8. 2+1. 1
76955+833L43234+r5240657 42+223LO
4.5+O.44 .6+0 .5
96.L+1,.382.2+4.5
Controls
(mean + SE)
6L.O+2 .7
9. 6+1. 3
61,4L6+L77753L285+9L69507 41,+L5785
4 . 9+0.55.2+0.5
96 . 5+t_. 391.4+L. I
p Values
<0.01
n.s
n. s.n. s.n. s.
n.sn.s
n.s.n. s.
l_
234
Þ<pressed asÞ<pressed asb<pressed as
pr-z
p.m. 3u-rdn- ipcorporated into stirnu]comoles of. L¿)T incorporated per 1-0/S. aureus killed in 2 hours.
ated llnnphocytes.neutrophils per hour.
Þ<pressed as å T. glabrata killed in 2 hours.
Hoo(,

i84
7,4 STJMMARY
Hacmophilus influenzac true b causes considerable morbidity and mortality in infants and young
children. Immunity to this organism has been attributed in part to the formation and increase with age of
antibodies to the capsular polysaccharide of the bacteria. Tmmunodeficiency coul{ in some cases, explain
why some infants and young children develop invasive haemophilus disease. In this study we investigated
ten infants with invasive haemophilus disease. We found normal lymphocyte mitogen responses' neutrophil
iodination, bactericidal and fungicidal capacities in this g¡oup. While we found no deficiencies of any of the
immunoglobulin classes, three patients had an IgG4 deficiency and two of them had anlgG2 deficiency as
well. These fiodiogr suggest that in some infants who develop haemophilus infections, the measu¡ement of
IgG subclasses reveals deficiencies that may indicate immune defects that would not otherwise be apparent.

CHAPTER EIGHT
IgG SUBCI,ASSES IN OSTEOMYELITIS AND SEPTIC ARTIIRITIS

1868.1. INTRODUCTION
flaematogenous osteomyelitis and septic arthritis are serious and potentially crippling infections
most commonly caused by Staphylococcus auleus, but sometim esby Streptococcus pyogenes or Haemophilus
influenzne. Immr¡nity to the common causative organisms is considered to involve opsonisation by
complement and/or antibodies and ki[ing by phagocytic cells (Adlam and Easmon, 1-983). Surprisingly
little is known about how staphylococci colonise host tissues or the reasorui why some people become
colonised and otlers do not.
While IgA" IgG and IgM have been measured in patients with osteomyelitis (Eid et a! L980) there
are very few reports of the measurement of IgG subclasses in such patients. There is also a paucity of
information about lprphocyte function and complement activity in child¡en with osteomyelitis and septic
a¡th¡itis. Heiner et al (1-988) have reported osteomyelitis in one IgG,l-deficient patient. Since IgG subclass
deficiencies can occur in the presencô of normal total serrm IgG concentrations and IgG subclass
deficiencies can be indicative of impaired antibody production to certain tlpes of antigens (Ch. 1.6.1-) to
look for evidence of immune deficiency, we measured IgG subclass concentrations in additioo ¡o ¿sssssing a
number of lymphocyte functions and making serum complement measurements in a gtoup of children who
had had osteomyelitis and septic arth¡itis.
82 PATIENITS
We studied all children admitted to the Adelaide Child¡en's Hospital over a 6 month period in
whom a diagnosis of haematogenous osteomyelitis and/or septic arthritis was confi¡med and who were able
to return to have immunological studies done at least 6 week after the com-encement of t¡eatment
(Beard et al 1990b). The group comprised fifteen children, aged from Z) montbs to L5 years with a mean
of 8 years. Diagrnosis was based on clinical findings together with the use of radiographs, isotope bone
scans, blood cultures and drainage procedures. Thirteen of the children had had osteomyelitis and 7 of
these also had had septic arthritis. TWo had had septic arth¡itis without evidence of osteomyelitis.
Staphylococci were isolated in 9 cases, streptococci rn 2, and no organisms in fou¡. Only three of the
children had a history suggesring excessive infection proneness (Table 8.1).

r87Table 8.1
CLINICAL DETAILS OF PATMNTS WTTH OSTEOMYELIIIS ORSEPTIC ARTIIRITIS
PatientNo.
Age(Y*)
Diagnosis Organism Past History
L
2
1
2
OM femu¡
SA hip
OM os calcis
OM humerus
OM tibiaSA sacroiliac
SAhip
OM tibia
OM cuneiform &navicula¡SA cuneonavicular
OM tibia
OM tibia
S. aureus
S. øtreus
GpA streptocoocl¡s
n.i.
GpC streptococclts
S. sureus
n.i.
S. øtreus
n.i.
veryfrequenttonsillitisand earinfections
ch¡onicsinusitis
recurrent
infections
mastoiditis
n.l.
3
3
3
3
4
5
6
7
8
6
7
8
9 8
9
IL
12
7i
13
10
L1.
12
73
L4
SA knee
oM tibia
OM femurSA hip
OM femu¡
SA knee
S. aureus
S. aureus
S. aureus
S. aureus
OM T10 vertebra S. anreus
eal
and
15 15
OM = osetomyelitis; SA = septic arthritis; n.i. = not isolated; = not excqssively infection-prone

r8B
8.3 RESIJLTS
Four patients had a low concent¡ation of at least one immunoglobulin isotype (Table 8.2). Serum
IgG concentrations were within the normal ¡ange in lÍl patients, raised in one, and sþhtly low in another
(Fig. 8.1). Seru'n IgM concentrations were within the normal ¡attge in all but one patient whose level was
marginally raised. Serum IgA concentrations were low in 2 patients and in one of them IgG and IgGl
concentrations wero also low. In most patients, both serum IgA and serum IgG concentrations tended
towards the lower limit of the normal rânges. One patient had low IgGz,lgG3 and IgG4 concent¡ations.
Six had at least one IgG subclass concentration above the 95th percentile for age. Neither the incidence of
IgG subclass concentrations below the 5th nor above the 95th percentiles for age was significant. Al
patients had normal concent¡ations of C3, one had ¿ 5lightly reduced C4 concent¡ation (0.12g/l normal
¡a¡ge0.L4-0.28 g/0, and none had reduced complement activity.
There were no significant difference in lymphocyte responses to mitogens between the patient and
control groups (Fig. 8.2). The percentage of Leu-l-L+ cells was lower in patients than in cont¡ols (p<0.02)
but other differences in lymphocyte subpopulations between patients and cont¡ols did not reach statistical
significance (Fig. 8.3).
There was no difference in cytotoxicity NK cell cytotoxicity between the patient and control gfoups
(Fig. 8.4), neither was there a difference in the subgroup of patients with IgG subclass deficiencies.
8.4 SI]MMA,RY
We investþated serum Igd IgG, IgM and IgG subclass concentrations, complement activity,
tymphocyte subpopulations, tym.phocyte responses to mitogens and natural killer cell cytotoxicity in 15
children who had had osteomyelitis and septic arthritis and in a group of control subjects. At least one IgG
subclass concentration was below the 5th percentile for age in 3 patients. We found an isolated deficiencies
of IgGl and combined deficiencies of.IgG2/lgG3/ryGa andlgG2/IgQ4. Concent¡ations of IgA and of IgG
tended to be below tle mean for age. The percentage of Leu-1-1+ cells was reduced in patients. Other
immunological parameters studied were uormal. Hence our stud¡ did not provide evidence for defects in
humoral immunity or lymphocyte function in most of the patients with osteomyelitis or septic arthritis that

189
we studied. Furtler studies, particularly those desig'ed to detect ¡estricted defects in antibody production,
in patients witl osteomyelitis and septic arthritis may be more fruifi¡l in helping to determine whether or
not such defects are a predisposing factor in these infections, and whether, in selected patients at least,
standard therapeutic regimes could be improved by the addition of immunoglobrrlin replacement therapy.
Although the lymphocyte functions that we studied were not abnormal in our patients, more extensive
studies could reveal lprphocyte abnormalities particularly in the patients with IgG subclass deficiencies.

190Table 82
IMMTJNOGI.OBTJLIN ISOTYPE DEFICIENCIES IN PATIENTS WHO IIAVEHAD OSTEOMYELITIS OR SEPIIC ARTTIRITIS
Age ofpatient(years)
Immunoglobulin isotl?e
2
3
IgA IgG IsM IgGl tñ2 IgG3 rgG4
G/t)
0.ã+ 3.99t o.tt L.821 0.72 0.33 0.05
0.43 6.87 0.81 4.35 0.5,1+ 0.06+ 0.05+
1.18 7.87 L.37 8.53 0.63+ 0.45 0.W
0.16[ 12.6 t.L6 6.50 L.47 0.54 0.68
I = >2SD below tle mean for age for immunoglobulin classes, and below the 5th percentile for IgGsubclasses
9
1_L

oo o
Ser
um l
gM (
g/l)
oo
a
a
a
Ser
um l
gG (
g/l)
oôo
Ser
um l
9A (
g/l)
\ \
NoÀ
9øo
oo
N oN o
t_!
CE æ U2 (! I E O
Q P : c(¡ o Þ E È oÊ aà c rì ê Þ E E
È o E ø Þ E 19 (! Èt
1Â f Ë 0 F ¡t (lt F t9 Ê c ø (D o E 6 aÀ c I ø (! E ê
F I E I É a ÈE ã o E l! F E r1 cà ã E Fl 6 aÀ (D Èt
Èt
.D tD ll t+ h) çt2 tt
a\ a\
\
aa
a\ 1
a
a
a
a
a a a
a
\ I I I I I I I I I I I I
N o
a r\ \
¡l
N
I \
\
a a a a
C) mo
2 m >ó
!. (r)
l' I
\ \ \ I I I
a
a
\ I l. I I l. I I
a a
Ì I I I I r \
a
\ \ \
a
a
\o @
\o H

L92
400
ÕoXEo-o- 300
uJYFo-l
$ zool-
I
ICY)
500
to0
o
No PHA Ftr/M ConAm¡to€en
Flg.8¿ Mitogen'induced lynphocyte prollferatlon in patients who have had osteom¡rlitis or sepücarthriüs (sl u"a in controls (f[) (meant SE¡.

193
80
60LIJot-ztu
Poo[rJo-
20
0
FMCl cD3 cD4 cD8 Leu 1'l
Fi& 83' Mononuclear leucocyte subpopulations in perlpheral blood paüents who have had osteomyeliüsor sepdc arthrtds (Ell a"¿ in conrok (fll t-"a"l SÈ¡
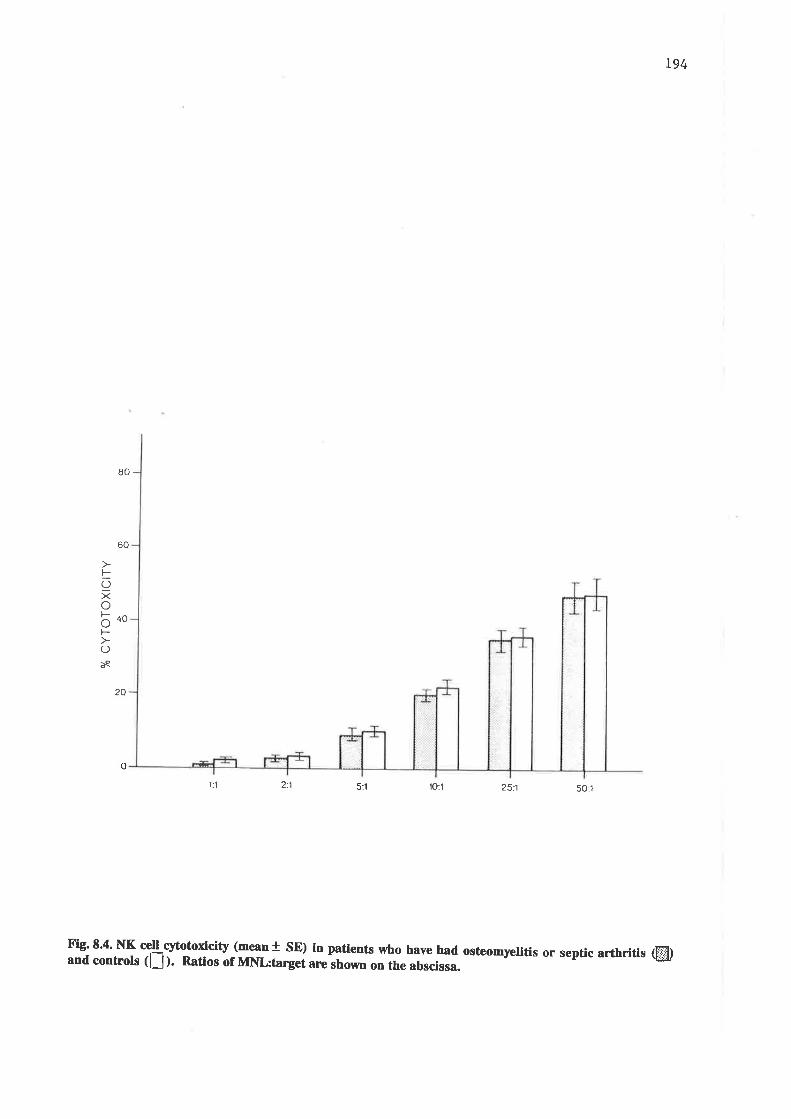
194
80
60
FOXobooF-
Oèe
20
o
't:1 2:1 5:1 'lO:l 251 5O:1
Fi8' 8'4' NK crctl cytotodcity (mean I sE) in patients who have had ostcomyelltis or septic arthr.lti, (E[¡and controls (lJ ). nauos of MNl:target arc shown on the abscissa.

CHAPTER NINE
IgG SUBCIASS CONCENTRATIONS IN CHILDREN \ryTTT{
GIARDIASIS

L96
9.L IITTRODUCTION
Giardiø lambliø is an enteric protozoa-n pathogen. It is still not known how the organism
overcomes host defences to cause disease or why the effects of i¡fection are so variable rangrng from
asymptomatic cyst passing to frank malabsorption. There is evidence that both humoral and cell-mediated
immnnity are important in defence against Giardia larnblia. Antibodies may either promote cytotoxicity by
macrophages and neutrophils s¡ inhibit attachment of parasites to the iutestinal wall.
While giardiasis is frequently present in hypogammaglobllinaemic patients with malabsorption
(Ament & Rubin, 1972; Amma¡¡ & Fudenberg,L9S2), giardiasis is a common infection and the majority of
patients with symptomatic giardia infections are not likely to be immunodeficient. Until recently neither a
deficiency of ci¡culating nor of secretory immuneglsþrrlin had been found in otherwise healthy subjects with
giardiasis (Babb et d, L97L; Jones & Brown, 1974). Circulating antigiardial antibodies have been
demonstrated in patients with giardiasis (Vinayak et aJ, L978; Smith et al, 1981) but their precise role has
not been defined. Anti-giardial antibodies in maternal milk protect ¡¡¡eanling mice agains¡ colonisation by
giardia (Andreas & Hewlett, 1981). Recently, reduced IgG4 concent¡ations in duodenal secretions of
patients with giardiasis have beeu reported (Grill et al 1985). Kumkum et al (1988) suggested that failure
to develop adequate amounts of antibody to su¡face antþen may impair ability to eradicate the parasite.
This sludy showed that patients with the ch¡onic giardiasis had lower levels of circulating antibodies to 2 of
the 3 su¡face antigens of. Giaùiø lamblia (plasma membrane and M¡82000/GISA 82) than did
asymptomatic or acutely infected patients and also found low levels of serum IgG and IgA in patients with
persistent giardiasis.
Cellular immune mechanisms also appear to be important in the clearance of giardia. Athymic
mice have an unusual susceptibility to giardiasis (Stevens et al, 1978; Roberts-Thompson & Mitche[ 1978).
Steroid therapy can decrease host resistance to the effects of giardial infection (Nair et al" 1,981). Increased
numbers of llmphocytes in jejeunal biopsies a¡e found in human giardiasis (Ferguson et al,1976; Wright &
f6mkins, L977). Theh role and function in immunity to, or imm¡pepathogenesis of, giardia infection is
unclear.

, lg7In order to try and identiS further defects in irnmune function that may predispose chìld¡en to
giardiasis we extended previous studies by measuring serum IgG subclasses, sen¡m complement,
lymphocyte responses to mitogens and neutrophil function in a group of children presenting with giardiasis.
9.2 PATIENTS
Thirty children, aged between one and 11- years, with a mean age of 2.9 years who presented with
gastrointestinal sig"s and slm.ptoms, and who had microbiologically prover infection wtth Giørdia lamblia
were studied for serum IgG subclass concent¡atiorxi, serum imm¡¡sglsþ'lins Ig\ IgG, IgNd and IgE and a
variety of lymphocyte and neutrophil functions.
9.3 REST]LTS
The serum IBA, IgG, IgM, IgE, IgG subclasses and serum C3 and C4 results are shown in Table
9.1. Two patients had reduced concentrations of both IgA and IgG, one had a reduced concent¡ation of
IgA and one had a reduced IgM concentration. Sixteen patients had raised IgE, concentrations (p<0.001).
Three had decreased C3 concentrations (n.s.), fou¡ had raised C3 concentrations (n.s.) and nins þ¿d
increased C4 concent¡ations (p<0.01). IgG1, IgG3 and IgG4 deficiencies each occurred in one patient.
Raised concentrations of IgGl occurred 'tn IL,37Vo of patients (p<0.01) and raised concentrations of IgG3
n 5, ?lVo (n.s.). Results of lymphocyte and neutrophil studies a¡e shown in Table 9.2. Patients had higher
B cell percentages than did controls and lower neutrophil chemotaxis. These differences, while statistically
significâlt, were not ma¡ked and patient values were well wifhin the normal ¡anges. Patients and controls
did not differ in T cell percentages, lymphocyte responsiveness to nitogens, neutrophil iodinatioq
bactericidal activity or fungicidal activity.
9.4 STJMMARY
A group of 30 patients who presented with proven giardiasis were studied for evidence of
underlþg immunodeficiency. Six of the 30 patients had a deficiency of one or 6s¡s immg¡oglobulin class
or subclass, IgA being low in 3 of these. The most striking fi¡dings, however, were raised concent¡ations of
IgE in 16 of the patients and of IgGl in 11 of the patients. The only IgG subclass deficiencies wero a low

198concentration of IgG3 in one patient and a low concentration of IgG4 in another. fmm¡¡s deficiency, and
in particular, IgG subclass deficiency does not appear to be a major factor in predisposing Australian
children to giardiasis. The raised IgE levels may play a protective role in thin intestinal parasitic infestation
(ch.1.2.1.2).

Tab]-e 9.1-
PatientNo.
Ageyrs
sERt[rf II,luttNoGIpBgLI}¡ A¡¡D CoMPLE¡@{T-CôNCENTIRATIO!{S IN 30 crrrLDREN WIIE GIARDTASIS
rgÀ IgG IgM I9E* IgGL TgG2 IgG3 IgG4 c3 c4
1234567I9t_0Ll_I213I4l516I7t_8)-9202L222324252627282930
11l_
t1l_
l-1Lt-1l_
22222233334456679LL
o.57o.370.56o.230.460.85o.240.36o.422.42+0.55o.320.58o.490.l_6-o.72o. 8l_o.L7-o.821.060.95t_. L00.88o.721.59o.7Lo.73o.590.48-0.80
7 .704.685.804.406.9L8.3L5.624.629.538.594.624 .437.686. t_53 .46-7 .636.138.22Lo. 007 .O5LO.208.376.086 .4012.037 .775.528.584.95-8.93
L.07o.25-L.420. 69o .62o .970. 690. 500.592.1,8+o.75o .430.860. 600.390.89o .470.95l_. l_6o .490.950.89o.751. 081.58L.20o. 59o.790.882.L2
37+1L3+242+208+27t_
6I35+1031546+9956+32+41,27 9+173+172+2L0+925+6779+88+182+47l_Llt_ 3
7 .66+5.867 .O2+4.48L2.04+7.26+6.264 .656.747.07+6.234.066.295.184 .477 .7L+7.73+5 .876.776 .248.09+L2.29+3 .614.268.36+8.t55.946.863.888.08+
0.660.84l_. 350.950.821_.29
. 1.04o. 9t_0.66o .670.98o .62L. 090.880.961. r.9o.920.83t_. 51L.7 2L. 0Lr..54L.37L.78r.972.L41.890.981. O4L.7 0
o .460.28o.97+o.250. 950.66+0.410.300.45o.79+o.25o.27L.L7+0. 36o.29o.29o. 450.51_0.360.310.30o.6l_o.26o.261. 38+o.37o.52o.L70.1_4-0.55
o.070.040.090. t_50.050.43o.020. 0to.L70. L4o.020.03o.23o.020.350. 09o. 04o.340.930.08o.L2o .440.060. t_30. 810. 63o .67o.4L0.080.03-
o .49o.94o.71,-0.861, .28l-. 83+o.97o.87r.221.t8L.O7o.72-2.2L+0.902.29+0.860.83t-. 68r.241 .010. 87L.L2L.32ndL. 02o.69-0. 800.80l_.041.86+
o.2I0.30.24o.2L0.48+o.27o.270.32+0.220.43+o.2L0.260.42+0. 36+o.270. 30+0.35+o.24o.200.14o.230. 32+0.24nd0.45+0.21o.25o.t70. L6o.26
f = >2SD aþove mean for age ror t_nmunogtoþulln classes ana aþove cne 95t.rr percentj-]-e for age f or fgc suÞclasses,-<2SD below the mean for age for inmunoglobulin classes and below the 5th percenti 1e for age for IgGsubclasses,. tt 1.u.,/nl ; nd not measured
\o\^o

TaÞle 9.2.
h¡nction tested
(a) LlnnphocYte StudiesT cell Percentage
(E rosettes)
(b)
LYITIPEOCTTß ÀIÍD NETTI'ROPEIL SjIUDIES II¡ CEILDRET YÍTTE GIÀRDIÀSIS ÀND 1]C C:OTflTROT'S
p ValuesPatients Controls(mean + SE)
NunberTested
B cell percentage( Surface ínnunoglobulin )
Mitogen r"tponteslPHAPTÍMConÀ
Neutrophil studiesChemotaxis (nn/3hr)
Quantltatíve iodlnation2Pooled serumAutologous serum
6r.0 + 2.8
]-2.5 + 0.6
19516 + 2083I),279 + 1641L7382 + 2275
2.1- + O.2
83.4 +85.0 +
59.6 + 8.0 22
r-0.1 .t 0.5 22
L8769 +L0348 +L4286 +
r.558l-1682068
202020
n.s
< 0 .001_
< 0.05
n.sn.sn.s
2.6 + 0.1
89.0 +88.1 +
19
4.96.3
n. s.n. s.
++
5 .1_
6.r++
SS
0.40.6
2.91.8
0.40.4
1.5t.5
2020
L9l-9
Bactericidal activiÇY3Fungicidal activitY"
nn
l-. Þ<pressed as c.p.m. 3H-TdR., jrpcorporated into stimulrated, lynphocytes2. n<þressed as piãomoLes of 125I iñcorporated per Lo/ neutrophils per hour.3. O<þressed as * S. aureus killed in 2 hours'4. D<iressed as å T. glabrata killed in 2 hours'
N)oo

CHAPTER TEN
IgG SUBCLASS CONCENTRATIONS IN PATIENTS WTIII
IMMUNOLOGICAL DISORDERS

2021.0.1 INTTR.ODUCTION
Because the description of IgG subclass patterns in known immunslsgical disorders will contribute
to a greater understanding of the natu¡e of these states and may also provide information about the in vivo
cont¡ol of IgG subclass production (Ch. L.8), lve measured IgG subclass concentÍations in patients with a
wide rânge of immunodeficiency and autoimmune disorders, and production of IgA, IgG, IgM, IgG1, IgG2,
IgG3 and IgGa by \mphocytes in vitro from four patients with immunoglobulin isotype deficiencies.
IO2 PATIENTS
Patients with isolated IgA deficiency formed the largest subgroup and have been discussed
separately in Chapter 5. The nerú largest subgroup of immunodeficient patients were children with reduced
concentrations of at least one effis¡ immunoglobulin class and with histories indiç¿ting excessive infection-
proneness. Fou¡ children had profound hlpoga-maglobrrlinaemia. Th¡ee of these 4 had very low IgA
concentrations and two had low IgM concentrations. Family studies in two of the 3 families involved were
also done. Another 5 children had mildly reduced IgG concentrations. Three of them also had low IgA
concentrations, and one a low IgM concentration (Table 10.1). Seven of these children have received
immun6glsþulin replacement therapy and are discussed in detail in Chapter 12. Concent¡ations of serum
immunoglobulins IgA, IgG, IgM, IgE and IgG subclasses shown in Table 10.1- were determined before
immunoglobulin therapy was started in all but one patient, MW, who had been receiving regular
int¡amuscul¿r imm¡¡sglsþrrlin for many years before IgG subclass measluements were possible in our
laboratory. Values given for this patient afe ftough concentrations obtained immediately before her 2-
weekly dose of intramuscular imm uneglsþrrlin.
Lymphocytes from three of the h1ryoga-maglobqlinaemic patients (TbI, BN and SL) and from one
patient with mildly reduced IgA and IgG concentrations (JB) were srimulated with pokeweed mitogen
(PïVM), a T cell-dependent mitogen and with S. atreus, a T cell-independent mitogen. Cells were cultured
over a 7 day perio4 with a series of concentrations of the mitogens, and immunoglobulin Qg) isotype
concentrations were measured in the supernatants.

203Other immunodeficient patients included in this study were child¡.en who presented to the
Adelaide Child¡en's Hospital with severe combined immunsde¡ciency (Roberton et a! 1986), adenosine
deaminase (ADA) deficiency, Wiskott-Aldrich syndrome, A, deficiency (Thong et al, 19g0),
myeloperoxidase defrciency, cyclic neutropaenia, diGeorge slmdrome (Thong et al, 1-978; Beard et al, 19g0),
human imm¡¡sdsficiency virus (Hf$ infection, and Ebstein Barr virus (EBÐ infection with histiocytosis
(Table 10.2).
The patient witl severe combined immunodeficiency received an HLA identical mixed leucocyte
reaction non-reactive bone marrow graft from a first cousin after T cell depletion at age 6 months. At first,
there was good evidence of engraftment (Roberton et al, 1986). Over the following years, however, there
was a gradual deterioration in his immune function, seen first in a reversed CD4:CDS lynphocyte ratio with
low numbers of CD4 cells, and then in a sequential drop in his serum IgG, IgA and IgM concentrations.
NK cell cytotoxicity and lymphocyte responses to mitogens also deteriorated. A second HI-A identical
mixed leucocyte reaction non-reactive T lymphocyte-depleted bone marrow ¡'ansplant from a cousin was
done 4.9 years after the fi¡st. CD4 numbers, CD4:CD8 ratio, and seru'r' Ig,\ IgG and IgM were low 0.g
months late¡. Both before and 0.8 6e¡rhs after the second ¡ansplant tÏe patient,s lymphocytes failed to
produce Tl'IFf on stimulation with mitogens. IgG subclass concentrations were monitored serially
tl¡ou ghout rhis period.
The patient with Wiskott-Aldrich syndrome received an Hl-A-identic¿l mixed leucocyte reaction
non-reactive bone marroy ¡ansplant from a siþling at age 10 montls. Subsequently, IgG subclasses were
serially quantitated.
Sera from patients with a variety of autoimmune diso¡de¡s - systemic þus er¡lematosus (SLE),
Henoch Schonlein purpura, juvenile rheumatoid arthritis, sclerodermq dermatomyositis and Hashimoto's
disease were also studied (Table 10.3).
10.3 RESI]LTS
IgG subclass concent¡ations in the patients with varying degrees of h¡,pogammaglobr¡linaemia are
shown in Table 10.1. Deficiencies of all four IgG subclasses occurred in the four profoundly

204
hypogamm¿gl¡þrrlinaemic children. TVo of these child¡en, TN and BN, are 5iþlings. Studies in their family
are shown in Table 10.4 and show IgG3 deficiency in their fatler, IgG4 deficiency in their mother, and no
deficiency in their sister. Patient SL is an only child. Studies in her parents revealed an IgG4 deficiency in
her mother and no deficiency in her father (Table 10.4). Llmphocytes from the 3
hypogammaglobnlinaemic patients tested did not respond by producing imm¡¡sglsbrrlins when stim¡l¿tsd
with PWM. When the cells were stinulated with ^S. øuteus, however, lym.phocytes from each of the 3
patients showed marked increases in Iglvf production, and cells from patient BN also showed a marked rise
i" IgA and IgG3 production. None showed ¿ signific¡nt rise in total IgG production or in that of IgG1,
IgG2 ot IgG4 (Table 1-0.5). These results are reasonably consistent with ths serm immunoglobulin
concentrations in these patients ie. TN and SL have normal serum IgM but very low IgA and IgG
concentrations,_and BN has normal serum IgA and IgM concentrations (see Ch. 12). Hi.IgG3, however, is
subnormal.
In the patients with mildly reduced IgG concentrations, 3 had reduced lgpl,lgG2 and IgG4
concentratiorxi, one had reduced IgGl and IgG3 and one had reduced IgG1, IgG3 and IgG4 concent¡ations.
Both of those with a low IgG3 concentration had a raised IgiE concentration.
Llmphocytes from patient JB, (see Ch. n) one of those who had mildly reduced IgA and IgG
concentrations together with reduced concentrations of IgGl, IgG2 and IgG4 responded very poorly to
stimulation with PIWM (Fig. 10.1) and showed variable, but generally better, respoû¡es to stimulation with
S. anreus. IgM and IgG3 production were better than those of control cells at some ooncentrations with S.
üneus (Fig. 10.2).
The patient with severe combined immunedeficiency was deficient in all the IgG subclasses, except
Igp4, at the time of diagnosis at age 4 months. Following a bone marrow transplant there was an ill-
sustained improvement in his IgG subclass concentrations and then a fall to subnormal levels, coinciding
with the fall is his total serum IgG concentration. Nearly five years later, a second bone marrow transplant
was done. Because of a failure to produce satisfactory immunoglobnlin concentrations he has been
receiving regular immunoglobrrlin ¡spl¿cenent therapy for the 14 months since then.

205
The patient with ADA deficiency was deficient in all IgG subclasses at tle time of diagnosis at age
14 months. Further results are not available as she died shortly after bone marrow ¡ransplantation.
The patient with the Wiskott-Ald¡ich slmdrome did not have any IgG subclass deficiency before or
after his bone ma¡roy ¡¡ansplant at age 10 months. The patient with diGeorge syndrome who had had a
thynic epithelial ¡ansplant 9 years previousl¡ had an IgG4 deficiency.
One of the 3 patients with a C2 deficiency had an IgGl deficiency. No other IgG subclass
deficiencies were detected in this goup. None of the few patients with neutrophil disorders had IgG
subclass deficiencies.
Of the 1-6 patients with SLE, scleroderm4 dermatomyositis and juvenile rheumatoid arthritis and
Hashimoto's disease, 6gVohad raised IgGl concent¡ations (p<0.001), 3L% also had raised IgG2 (n.s.), and
25Vo rused IgG3 (n.s.). The only deficiencies werc TiVo with a low IgG2 (n.s.) and 3L% wrth a low IgG4
concent¡ation (n.s.). No trends were apparent in the group with Henoch Schonlein purpura.
IO.4 SI]MMARY
IgG subclass deficiencies were found" as one would expect, in patients with known immunoglobulin
deficiencies, severe combined immunodeficiency and adenosine deaminase deficiency. They also occurred
in some patients with C2 deficiency, the diGeorge Syndrome, SLE, scleroderma" juvenile rheumatoid
arth¡itis and Hashimoto's disease. In the patients with the rJ/iskott-Aldrich syndrome, myeloperoxidase
deficiency, cyclic neutropaeni4 HIV infection, EBV infection with histiocytosis, dermatomyositis and
Henoch Scholein purpura, IgG subclass deficiencies were not found. Raised concent¡ations of IgG
subclasses, predominantly of IgGl-, were common in the patients v,/ith ¿¡tsimmune disorders.
The capacity of cells from fou¡ patients with immunoglobulin deficiencies to respond, to some
degree, to S. aureus, a T cell-independent mitogen and not at all to PWM, a T cell-dependent mitogeq
suggests that there is an impairment of T cell immr¡nsregulatory -ssþanisms in these patients.

206Similarl¡ the failure of the patient witl severe combined immunodeficiency to produce satisfactory
concentrations of IgG subclasses and other imm¡¡eglsþrrlins despite satisfactory B cell numbers and in the
presence of reduced CD4 numbers and reduced cytokine production suggests that a T cell regulatory defect
may be an important factor in his humoral immunodeficiency.
fu this very heterogeneous group of patients with disorders known to affect immunological
function, the order of frequency 6f immr¡nsglobulin isotype deficiencies was:
rsGa (aÙvo) > Igc (30%) = IgGl Qlvo) >rgG2 (n.5%) >rg{. (22.5vo) >Igc3 (n%) >rsNr
(n.5Vo)
The order of frequency of raised concentratio¡5 6f ìmmunogloblin isotypes was:-
,f, f*ø> >rgc (?SVo) >rs{ (L7.5%) = IsM (I7.5Vo) = IsG3 (I7.5Vo) >rsG2 (rsVo) >Igc4
(o.7svo)
Since patients with apparently isolated IgA deficiencies were excluded from this stud¡ the
occlurence of IgA deficiency and concurrent IgG subclass deficiency are under-represented in these figures.
As in the studies in IgA-deficient patients described in Chapter 5, fhis sg¡¿y in anotler group of patients
with immune disorders shows IgG4 deficiency to be the most common IgG subclass deficiency.
Furthermore it shows that IgG3 and IgM deficiencies are uncommon, as are raised concentrations of IgG2
and IgG4 in this group of paediatric patients. Some possible implications of these fiodi"g" will be discussed
further in Chapter 14.

207
Tô.bI€ IO. T
CLD{ICì,I. DSIAII.S MD CONCBTI1T,ÀÍrOÑS OP IIIIII]NOGLOEÛI.DI I,SOr!PAS D¡SYXPIþ'o'rIC PÀffE¡ Ts rllE VÀRYII¡G DEGR.BAI; OP EIPOGÀXXÀGLOE LIXÀ¡I(IÀ
PaCfat Àge cILnlqL IgÀdet¡1le
IgG Igt{Àt IgGr fgc2 tsca IgG¡Û rgE
lul!lt4tlng
1. t¡
B.I
6.L
T.P
À.t
?e
6yr
6@
0 . 011-
r.{l
UD-
L.47
0.5r-
78
0. l8-
0. r1-
UD-
5. r9-
5.t2
0.76
L.22
0.07-
o. 056-
0.1.6-
0.0{t-
o.078-
0.015-
o.015-
O.0¡¡7-
0. oo9a-
0.007-
0. o0l-
0. oo0l-
0 . ooo9-
0.6
o.5
<1
0.68- 0,28 0.O18- 564+
l.oa 0.ll- 0.82 ll7+
t6-
0.62- 0.51 o.o22- 9
conwl6lon6Pnewgystl6PneEonl.!Braln 6teE dlseÀ6e( ?câse )
12yr Rêc. ear and O.ol-c¡e6t lnfectlon6
Pn€Eoq!¡6tIEPnèt&nla
Rec. otltlEæd1ÀPn€Eontà x ¡
2 .2r-
5.8r-
0. 14-
o .05-
o.93-
l.l7-
0. 59-
o-77-
0. 05-
o.{o
o. 05-
0.07-
<l
<1.l2yr Rec. 6lncitls
ctrronLcbronchltl6Bronchl.ectà6 isÀs thu
lsfr Rec,6lnultis andtonslllftis
8yr Rec- Oot1t16 EedlàRec- bronchltls
t.ol
o. 55
40-
2-01-
loyl Rec.Ilyr bronchlitis
ÀstteRec . Glnarylnfectlons.
o.57-o.80
o.65o-72
2 -5t-t .29-
52II 26-
2f-oo
251+
10!E Rec. o. 15-pnelenlÀÞrdobronchlàI vebs
1.46- 0. 58 ).23-
lEEy€ar;rcG ¡ontht R€c - feclJEent >2 SD above rean for rge tor cla6€s6 ùd aævet¡e 95t-h p€rcêntlle forb€Id th.5tà p€rc.ntllafor
Àfl6rgcfor
6u¡cl.ÂÉae6, - - <2SD b€loyIgc €u¡classoa
thê ¡qn lor lge for lruoglobuLln clÀ66s. andÀ9êlor

208
Þ¡le 1O-2
IIlln]ITOGIl)q'IJN TAOI"IPE COIICEIIIßÀTIO!¡S D¡ CAILI'R.E¡ ¡I¡Tg RT€Ð@{IAÐ DIXI'XODETqIAT¡CY DIAORDEìS
DlagDoels Pat- Àge atlênt t66tlng
(years )
rgÀ IgG Iqt{ IgGl fgcz tgc3 I9c4 rsBlu,/El
Sevg€coDbl¡edl@uo-d€flcidcy
ÀÐÀdef Lclqcy
KÀ 14Eo o.l3-
0.20o.2r-0.32-0.13-T'D-UD-
0. {7-8. O03.23-1. 20-o. 87-5 .O7
o.471. 6{+0. s9o .470.02-o. 20-
0.005-0. 007-0.016-
00240
Hu
5I6577
0
0000
ITP
BLDW
T¡{
r,fr
5980+3605+l350
8Ao760
32à7¿5a3a4b
3
5562a9â
o.23-4,74
0. 12-t. 84
00
oo
o397
0405o5
0, 150. 2l0. 180.13o.70o.{o
05-08-39-
7.50+5.78
I.74
00
7757
0.r1-0 ,2I-1. 48
L9799¡l5o+0858+
000
t{IÊtott-ÀJ.drlcbSyndræ
0.¿18r.040,86o.422.43+l.{4+
3. 909.14+s.155.035.387.O0+
0.03
o .37 o.65
o. 07
1. 56- O. 28- 1. lt- 0.28_ o. o7-
1221t2
1Io
0Io
6794+74B9oo7L
006o25o150110526
<l
41+
2 35+76+{l8 3+
def iclqcy
HyeIo-IErdidå9edef lcIqqy
10110.8
7 .026.40-8.46
5.242. l0-6 .68
0. 370.24o .27
o. l4o. 211. 06
r77605
975?O+I
cYcuc t{cneutrcIE€nld À.tl
dlcoorge BDslmdFæ(post thybl.c¿râÉpLütâtlon )
9 .68 0.5{ 7. 19
0.63- 7.59 1.50
o .12 3,51
{5
0. 57 3.14 O. ¡14 o.58
0.71o.72
11.46+10 . o8+
oo
10 1,00 1.19
1.7
I1
oo
'l+8+
75+40
00
09l1
86o1
{3
I
0 025-o. 82+
HIV frfectlon LR o.7
EBV Lnfêctlon BCt hL6tLæyto6lÊ
1.1¡lo. 98
14.4+l¡-3+
2.991. s5
22
1166.7
a = Dntàs post flrst bone Érrow transplantâtloni b - EontàE Iþst socond bone @rov t¡usplantåtloni + = >2sä¡ore the æån for aqe foE i@uoglobulfn claÉ6ês, Ànd above th6 95t.L ¡Ercenttle for agê foi IgG Bu.bc1as6ês,- - <SD belos t¡¡e Eean for age for l@uoglobulon claaaes, and belor t.l¡e 5th p€rcenttle for age for lgc6u-bcla6se6i tD = udetectàble,

209
T¡-bl,e 1O.3
IXXI]IIOGI¡8I{'LII{ ISqfYPE CONCEITTRÀTIOT¡S Ill PÀTIEI'IS rIT! IXXOìIORECÛI¡TONY DISORDERS
u¡å9hoBag I9A r9c ¡9tr IgGI tgÉ ¿ rgGf IgG4NaEe Àge
05
43473t
05-o5-
40. 3+1.612.9r1{.710.311 .5+
J.I7,1+1.11.52.4t.31.12.r2.L
20oo00000
12
0I000I0
4
13
4
0142
6+53
252+4+9
4I2
2I220
0,180.200.03-0.89
o.2Lo.24o.07-0.85
0.ls-
tf56+{8l082+85-
5{05
55+577756
2850+5284
r{. fCN ].IJT 1IRf{ 14t¡ 15r¡ 16PS L7Kt 18KX ¡8
cD 11
I7 .9+8.9
37.69+9.38+6.A27.966 .16ro. 9t+9. 16+7 .47
Sclerode@
D€rutoyosicl6 cB lI1.6
1. 15
0.4a-
f.66+
0.39
0.37
l. f7+
0 .50
1.23
32l956
0.9L.6I.8+¡,.0
3.r4L5 .7 6+5.928.42
13.64+
L2-O2+
17. OO+7.7510.93+t8,68+
15.5
19,5+
4+
5+
00o0
2,I.l.1.
1.
1.3-
64
3+4+
IL22
12. 01.723 .5+17.8+
6.115.6+9.18.8
Juvenilerheuutoidart-b¡:ltls
>3.5+l.o6. O+t.9
o.a6 ,4+L.74-7
r.9
KD2ÀÀ3l{L 15vD5
colRll 5su6
13
KI, 15
l3533367
I¡enocbSchonLeinPurpra
0.{74.28+2.7+ 8. 12+9.9HaBhlrcto'6dibease '
+ Eeusu-bcla6ses; - - <2SD b€lovIgG auÞclaases.
the æan f or aq'e f or Ln cLa66ê6, and belou ths sth
DIXI'TOGIOF]LIX TSOTYPE COÙCBI¡IRÀTTONS IN FÀIET.IES OF A!'ÞOGIII0ìCI¡BI'üII{À3ITIC CEILDRSX
for age for
Îable 10. a
Patlênt Àge cllnlcaIdetå1I6
rgÀ IgG rgü IgGl tqcz lgc3 IgG4 IgE1ulEI
Fap1lv 6tudles
1. r{. . B. N.Pãtber Àd f{elll{otl¡er Àd Rec.S16ter 3]E well
T'RTIS2 .802 .32o.67
7 .436.366.50
1, 590. 05-0. 33
,90.9000
13108.
!.10
18-7024
ooo
007A95
s. ú.Fatlr€rXother
W6L1tfsII
2.331.{r 0.6s
0. 68L.270.05-
Àd - adult, Rec' uRTrê ' r€ctlmt up¡nr reeplratory t¡act lnfectlonet - - b€ror tbe stl¡ ¡þrcqttle for age.
1I .701¡.10
Àdrd 1
1 32029
3955
058l

Table 105 2LO
INWIRO PRODUCTION OF Ig ISOTYPES BY CULTURED PERIPIIERAL BLOOD LYMPIIOCIESF'ROM 3 HYPOGAMMAGLOBTJLINAEMIC PATIENTS, EXPRESSED AS FOLD RISE
Isoty¡re Stimulant PatientSLBNTN
IsG
IsA
IgM
IgGl
leGz
IgG3
IcG4
PWMS. aureus
P\ryMS. aureus
PWMS. aureus
PWMS. aureus
PWMS. aureus
PWMS. aureus
P\ryMS. aureus
r.tÉ.7)0.6Q..2)
o.e(3.0)L.8(2.8)
1.0(1.ÐL0.2Q.Ð
r.z(L.3)0.7(0.Ð
o.e(1.Ð0.6(2.7)
0.e(2.0)r.6Q.7)
t.2(L.7)0.sQ.2)
1.0(3.0)7.6(2.8)
r.3(1.7)s2.6(7.s)
1.1(1.3)0.6(0.Ð
-(1.s)-Q.1)
-(2.0)3.6(2.7)
0.6(2.0)0.e(1.0)
0.e(3.6)1.6(1.3)
1.0(5.e)11,.7(1.5)
0.6(2.e)0.8(1.3)
-(4.s)-(3.0)
0.8(2.4)L.3(0.e)
-(1.3)-(1.3)-Q.o)
-(1.3)-(2.0)
(2.4)
Control values in parentheses; PWM dilution = ILAD; S. anreus:lymphocyte ratio = 10:1; - = aso"detectable.

2LL
=E-àc
o
-E
oc
6o
Eocc)o-s
fxx,
t50=Eoc
o
Eoc
N(,o
âÊoco-9
200
()0
4
30
500
2so
0 0'2 r 5
oo.2r5
oo.2t5
o o.2 | 5
o o.2 I 5
o0-2t5
00-2ts
t5 EocIoo
Fl& 10.1 In vífro produclfon of þ tso$pes by cultured peripheral blood lymphocyteÁ of ¡nüent JB
ln response to stimul¡fion with FIYM at difierent conccnhadons.

2L2
=Eoc(,-g
=Eoc
.9
r50oc
o
Èoc
N(,o
a2ahoc
f€oo
200
I rso
loo
'l)
o rr 511251
o 1r 51¡251
O 1:r 5t I 25:1
o t:r 5:rl 25:r
o rr srl zs,
Eoc
(5-9
Eoco(9õ
f5
50
25
^
o Patient
. Control
o r:1 5tl25t
XÏ9. 102 In vifro producdon of þ isot¡pes by cultured pertpheral blood l¡mphocftes of paüent JB
ln response to stlmulaüon with S. øtreus (rafias rcpresent.S. øçeuÍs.lymphocXtes).

CHAPTER ELEVEN
THE INFLUENCE OF THE SPLEEN ON IgG SUBCIASS
CONCENTRATIONS

1.1..1, I}IIR.ODUCTION 2r4
The spleen is important in host defence in that it clears particulate matter from the blood and
produces antibodies (Sullivan et aJ, L978; Schrmacher 1-970; Hosea et al 1981; Pedersen et al, 1982; di
Padova et al, 1983; Kiroff et al 1985). The liver clears IgM-opsonised particles well and IgG-opsonised
particles not very effeclively. The spleen clears IgG opsonised particles more effi.ciently. Splenectomized
subjects have an impaired ability to clear particulate matter from the blood and to produce antibodies.
Decreased concentrations of serum IgM (Spirer, 1980; Cohen & Ferrante, 1982; Drew et al L983;
Schumacher 1970) and decreased ability to produce both IgM and IgG antibodies to pne lmococcal
capsular polysaccharides occur after with splenectomy (Hosea et aI, 1981-; Pedersen et al, L982; di Padova et
al 1983; Schwart4 1985).
Splenectomized subjects may also have a reduction in circulating T lymphocyte numbers and
fi¡nction (Ferrante et al 1985). They do not, however, show an increased tendency to malignancy or to
infections in general, but are particularly prone to infection with certain encapsulated bacteria, especially
Streptococcus pneumoniøc, for reasons which are, as yet, ill-defined.
In previous studies we found that there vve¡e simil¿¡ abnormalities in some immune function
parameters in both splenectomized adults (Ferrante et al, L985; Ferrante et at 1986) and in child¡en with
portal hlpertension (Ferrante et al L989). Both groups showed an increase in the relative percentages and
absolute numbers of peripheral blood B lymphocytes, although the increase in absolute numbers was more
marked in the splenectomized group. Both groups also showed increases in NK cells and monocvtes and
tlese were more dra-atic in tle splenectomized. Both groups showed decreased responsiveness of
peripheral blood lymphocytes to sfimul¿fis¡ with PHAs ConA and PWM. Splenectomized subjects had a
marked increase in NK cell cytotoxicity (p<0.002). Splenectomized child¡en and adults a¡e both at risk for
severe S. pneumoní¿¿ infections, and tïe risk is greater in children (Drutz & Graþill, 1982). Patients with
portal hypertension do not appear to be at risk from such infection. As discussed in Chapter L.6.L.2,lgG
antibodies to S. pneumoniae are generally of the IgGl and IgG2 isot¡'pes.

215
In order to determine whether IgG subclass deficiency migh¡ çsntri6ute to the proclivity of
splenectomized patients to serious infections with encapsulated organisms, particularly S. pneumoniøe, we
measured IgG subclass concentrations in 3 groups of patients.
a)
b)
Ð
a group of 18 splenectomized child¡en
a group of L6 splenectomized adults
a group of 6 child¡en with portal hypertension.
LIz PATIEIYTS
Eighteen children between 9 and 16 years of age splenectomized for t¡auma or for haematological
indications, L6 adults, splenectomized after traumatic rupture of the spleen and 6 children with portal
hypertension and splenomegaly were sludied.
1.1.3 RESULTS
In the adult group of splenectomized patients me¿ìn serum IgA was higher than in controls, while
mean serum IgM was lower. Mean serum C3 and C4, were lower in the patients than in the controls
(Table 1.L.1). Selm IgE was elevated n 2 of the adults. Mean serum IgGL and IgG3 were higher in
patients than in controls (Table L1-.2). No splenectomized adults had an IgG subclass concsntration below
the 5th percentle for age, whereas 5 had an IgGl and 4 had an IgG2 above the 95th percentile (Table 11-.3).
In the group of splenectomized children, 3 had low IgA concentrations (p<0.003), one had a low
IgG concentration and none had low IgM concentrations. TVo þ¿d ¡inimally raised IgM concentrations.
IgE was elevated in 6 of the 18 children in who it was measured (p<0.001). There were 4 with an IgGl and
7 with aa lgp2 concentration above the 95th percentile. There were not ¿ significant number $'ith IgG
subclass concentrations below the 5th percentile for age (Table 11.3).
In the group of chitdren with portal hypertension, there was no significant incidence of IgG class or
subclass deficiency. Fifty percent of the patients had raised concent¡ations of IgG4 G,<0.001) (Table 11.3).

2L6Qqs child had slightly low concent¡ations of IgA and IgG. AlI had normal IgM concentrations. c3 was
sþhtlyreduced in two.
LI.4 STJMMARY
Splenectomized subjects are at risk from infections with encapsulated bacteria" particglarly S.
pneumoniac, and for unknown reasons this risk is greater in child¡'en than in adults. In this study we
eÍended previous studies of immunity in subjects with absent or impaired splenic function by quantitating
IgA, IgG, IgM' (tIgE) and IgG subclasses in these subjects. There was no evidence that IgG subclass
deficiency was contributing to the proclivity to infections with encapsulated bacteria. Rather than IgG
subclass deficiencies, we found elevated levels of IgGL andrgp2in significa¡t numbers of the
splenectomized subjects. In the adult splenectomized patients, IgM tended to be lower than in controls,
and in the paediatric splenectomized patients IgA tended to be lower than in cont¡ols while IgE was often
highs¡. Previousl¡ we have found raised NK cell activity in splenectomized patients. It may be that the
abnormalities of immunoglobulin isot}pe concent¡ation in tlese patients reflects changes in regulatory
¡sçþanisps mediated in part, by NK cells. The impaired ability to produce antibodies to pne ,mococcal
polysaccharide antigens in splenectomized subjects, is not reflected in a concomitant deficiency of total
serum lgGI and,/or lgG2.

Tabte 11.1 2r7
IeA Igc, þM,IgÍ'., C3 and C4 CONCENTRATIONS INSPLENECTOIù¡IIZF,D ADI]LTS AND CONTROI,S
Parameter Patients*
2.4+ 0.L9
10.59+ 0.63
0.y2+ 0.t9
1.06+ 0.06
0.20+ 0.01
Controls**
T.7L
LO.ß
t.37
L.27
0.25
p.
<0.01
Il.S¡
<0.001
<0.01
<0.001
rgA
IgG
rsM
uu
* mean* SE
** mean

Table 112
MEAN IgG STIBCLASS CONCENTRATIONS AND PERCENTAGES IN 16 SPI,ENECTOMIZEDAI)IJLTS
mean* SF, G/t)
patients controls
IgGL 6.661 0.58 5.11+ 0.25 <0.02
rñ2 3.2L+ 0.47 2.75+ O.2O n.s.
Igc3 0.85+ 0.10 0.58+ 0.05 <0.02
lCG4 1.07+ 0.30 0.56+ 0.08 n.s.
218
p'
mean 7,t SE
p.patients
60.00+ 2.53
25.53+ 2.04
6.81+ 0.78
7.68+ 1.50
controls
57.67+ L.87 n.s.
29.y2+ t.72 <0.05
6.28+ 0.Y n.s.
6.06+ 0.87 n.s.
IcGl
IgG2
IgG3
IgG4

Table 113
þG SUnCr"nSS CONCENTRATIONS ABOVE TIÍE 95TH OR BELOWTTTE STH PERCENTILE INSPIÆNECTOMIZED PATIENTS AND IN PATIENTS WITH PORTAL ITYPERTENSION
>Po<-)-
219
1-6 splenectomized adults
IgGl
IgG2
IgG3
IgG4
s þ<0.0031 0
4 þ<0.051 0
3 [n.s.] 0
3 [n.s.] 1
ln.sJ
[o..1
lo.sJ
t"sl
18 splenecto mizÊd children
IgGl
IÑ2
IgG3
IgG4
a [p<0.01] 0
7 [p<.00q 0
0 [n.s.] L
0 [n.s.] L
lo.sJ
[n.sJ
ln.sJ
[o.sJ
6 child¡en with portal hypertension
IsGl
TÑ2
IgG3
lgG4
1- [n.s.]
0 [n.s.]
0 [n.s.]
3 <0.001
0
1,
1
0
lo.sJ
["s]
lnsJ
lo.sJ

CHAPTER TWELYE
IgG SUBCLASSES AND IMMUNOGLOBULIN REPLACEMENT
THERAPY

22Itz.L Ii\¡"IRODUCTION
Severe and recurrent infections in antibody-deficient subjects cause considerable morbidity and
mortality. IgG subclass deficiencies may be a sign of antibody deficiencies (Ch. 1.6.1; 6.3).
Early prophylactic intervention in IgG subclass - deficient subjects may help to prevent disabling
ch¡onic recu¡rent infections. Ilowever, it is not knoum what is optimal in terms of early prophylactic
intervention. As discussed in chapter r..9, there Írre many unrnswered questions regarcling the use of
immunoglob¡rlin replacement therapy i" IgG subclass-deficient infection-prone subjects. Some patients do
appear to benefit from such therap¡ but there are no clear guidelines for decirting which patients require it,
which imm¡¡oglobulin preparations a¡e most effective for different IgG subclass deficiencies, what are the
optimal dosages and frequencies of administration or whetler age-normal conceut¡ations of each IgG
subclass should be maintained.
In order to provide further i¡formation about the effect of regular immunoglob,lin replacement
therapy on infection proneness and on IgG subclass concentrations, we evaluated three intravenous
immunoglobulin (MG) preparations for IgG subclass and content of selected antibod.ies, and administs¡sfl
one of these preparations regularly to t¡eat seven patients with immunoglobulin deficiencies. Another
patient was treated with intramuscular immunoglobrrlin (IMIG).
In fhis chapter, clinical and immr¡trologicål details of each patient are described together with thd
immu¡sgl6þrrlin replacement reeimes used and the effect of tlese on infection proneness and on
imm¡¡sglsþr¡lin subclass concentrations (see Beard & Ferrante, 1990).

I:22 THE COMPOSITION
PREPARATIONS
OF INTRAVENOUS222
IMMTJNOGI.OBT]LIN
122,L IgG subclass content of intravenous immunoglobulin preparations
The IgG subclass compositions of a n'mber of different IVIG preparations prepared by a variety
of methods and analysed by a number of investþators are shown in Table 12.1. While most analyses were
made by radial immunodiffr¡sion with polyclonal antisera, we used an ELISA with monoclonal antisera and
obtained simil¿¡ results to those of other inVestþators.
We have measured and compared the IgG subclass composition of the th¡ee IVIG preparations:
Intragam (CSl Australia), Sandoglobulin (Sandoz, Basle, Switzerland) and Ge-maga¡d (Hyland-
Travenol USA). Intragam and Sandogl6þr,lin are prepared by cold ethanol fractionation of pooled sera
and t¡eated at pH4. In addition the Sandoglobulin preparation is subjected to a trace of pepsin to
eliminate polymeric aggregates and to prevent reaggregation of poþmers and ¡limers. Qemmagard is
prepared by ion exchange chromatography. A comparison of the mean IgG subclass compositions of
mnltiple lots of these tl¡ee prepartions is shown nTable 12.2. Sandoglobrlin contains relatively more
IgGl and IgGa, and relatively less IgG2 and IgG3 than does Intragam. Çamm¿g¿¡d contains relatively
more IgGl and IgG3 and relatively less ÍgG2 and,IgG4 than does Intragam. While some of these
differences are statistically signifisanÇ their clinic¿l signifiç¿¡çe is not known.
l:¿22 Antibody content of intravenous immunoglobutin preparations
Although, logically, replacement sf ¿ missitrg IgG subclass migh¡ 5ssp to be appropriate t¡eatment
for an IgG subclass deficiency, in facÇ the situation is probably much more complex ¡[¿¡ rhis. Some
subjects with a deficiency of one IgG subclass have been found to have an impaired ability to produce
antibodies of other IgG subclass isotlpes also (Shackelford et 4 1983). It is generally agreed that a
deficiency of functional antibodies against particular pathogens is the important factor, rather than the
concentration of an IgG subclass pet Ee, so it is important that IgG preparations used to treat IgG subclass -
deficient patients contain adequate concentrations of protective antibodies against the pathogens likely to
infect those patients. It is, of coluse, not possible to measure concentrations of all such antibodies, but
those to a number of protein and polysaccharide antigens can be determined.

Table fÌ.1 223
INTERSTIJDY COMPARISONS OF IgG SUBCLASS COMPOSITION OF VARIOUS INTRAVENOUSIMMTJNOGT,OBTJLIN PREPARATIONS
Preparation Study IgG subclass percentage
IgGl rgc2 IgG3 tgG4
Method ofmeasurement
Anti- Standardsera
SulfonatedTeijin, JapanTeijin, Japan
B propiolactone-treatedIntraglobin F,Biotest, Germany 3Intraglobin F 2Intraglobin F 4
Polyethyleneglpoltreated
fmmuno, Austria 2Immuno, Austria 4Greencross, Japan 2Greencross, Japan 4
PPr¡B/¿lþtrminKabiVit¡um, Sweden 5
DEAEHyland, Travenol 2Hyland, Travenol 4Gamm¿g¿¡d, Hyland" 6
Travenol
pH4, trace pepsinSandoglobulin,Sandoz, Basel
T')
2L
3
1
2%?Á
83526
302633'2Á
33
33?329
332030?323
6372
uØ69
&726267
62
627076
5769606872
<1 RIDRID
67 /e767 /e7
H0-00267 /e7
67 /e7
67 /e7
67 /e7
67 /e7
67 /e7
67 /e7H0-00267 /e7H0-00267 /e7
PCPC
PCPC
0
NephRID
RTD
RID
5 0 ErAlRrD
002
4034
565
il
2)23
24332
53276
87764
<1<1<1
PC
PC
PC
PC
MC
PCPCPCMCMC
RID
ELISA
E1A/RrDNephRIDELISAELISA
RID = radial immunodiffusion, Neph = nephelometry, EIA = elect¡oimmunoÍ¡ssay, ELISA = enzjEe-linked imm unosorbent assay
Studies: l- = Skvariletal,1980;2 = Romer etal5t982;3 = BeckandKaiser,1.981;4 = Morell,1986;5 = Oxelius, L984;6 = Our studies L989;7 = Out studies 1-987.

Table7i22 224
MEAN PERCEI\ITAGES OF IgG SUBCI,ASSES IN 3 INTRAYENOUS IMMIINOGLOBIILINPREPARATIONS
Intragan
Sandoglobulin
Çammagard
IgGl
6t.2
72.3(<o.oo1)
75.7(<o.oo1)
IgG2
33.5
23.0(<o.oo1)
19.3(<0.001)
IgG3
3.5
2.5(<o.oo1)
4.6(0.006)
IsG4
L.9
2.2(0.01)
0.4(<0.001)
Probability of difference from Intragan by independent t test shown in parentheses.

225
A number of studies l¿ys highlighted the fact that the titre of an antibody measured
immunologically io ¿¡ imms¡sglobulin preparation does not necessarily correlate with its biological
activity. (chapter 1.9.1.3) Alteration of immunogloþulin molecules du¡itrg processing, or even the different
IgG subclass composition of the antibodies present warrant consideration. with these reservations, we
have compared antibody titres to a variety of bacterial and vi¡al antþns in three intravenous
immunoglobulin preparations. Tables 12.3 and 12.4 show that there were no major differences.
t2.3 IgG suBCLAss REPLACEMENT By rMMUNocr,oBuLIN
ADMINISTRATION
IgG subclass concentrations, in IgG or IgG subclass-deficient patients, can generally be maintained
within the normal ¡ange for age by regular infusions of IVIG but we do not know whether rhis should be
fts ¡im of replacement therapy or whether it is necessary to maintain normal coucentrations of all IgG
subclasses to protect patients from frequent and severe infections The main¡g¡ance of protective antibody
titres, ratler than the maintenance of normal IgG subclass concent¡ations, should perhaps be the aim of
t¡eatment. If a patient's health clearly improves s¡ immunqglsþrrlin therapy is it necessary to monitor IgG
subclass or antibody concentrations? The following case reports illustrate dilem-as encountered in trying
to address some of these issues.
l:¿.3.1 Immunoglobulin subclass replacement in hypogammaglobulinaemic patients
1:¿3.l,l Patient TN (IgA and IgG deficiency) (see StaÌgas et at, 19gE)
Clinical hesentation. A 7 month old male infant presented with acute respiratory distress resulting from a
Pneumocystis carinü pulmonary infection was found to have profound hypogammaglob'tinaemia. He had
had a rattly couøh since age 5 months. Before this he had been well. Growth and psychomotor
development were normal.
rmmunological Investþations. He had very low concentrations of serum IgG and IgA but a normal IgM
concentration. His IgG subclass concent¡ations were all very low (Table 10.1). He had normal B and T
cell numbe¡s, proportions and subpopulations and in vitro responses to mitogens. which while low to low-
normal were comparable with those of the control of this day. (Table 12.5).

Table fl3 226
ANTIBODY TITRES IN 3 INTRAVENOUS IMMTJNOGLOBT]LIN PREPARATIONS
Antþn Assayused
Vtual
C.M.V.
Measles
Rubella
Heæe¡ simplex
Hepatitis Bsurface antþen
R.S.V.
HTLV-III
ELISA
c.F.T.
Há..I.
C.F.T.
E.I.A..
C.F.T.
E.I-A,.
R.I.P.
w.B.
-ve
160
-ve
ve
>80
80
512
>80
>3?n
+ve(false)
>80
>80
256
>80
>80
80
512
>80
-ve
>3?l
+ve(false)
>20ß
LL.6
2.7
-ve
-ve
ffi
2W
7i.4
3.0
2.2
-ve
-ve
Bacteria/Toxins
Diphtheriatoxoid
ELISA
Tet. toxoid H"4,.
Bordetella pertussis
IgG ELISA
IgM ELISA
TgA ELISA
>ffi
L.4
320
512
1i.L
4.8
3.5
CMV = cytomegalovirus; R.S.V. = respiratory syncitial virus; HA.I. = haemagglutination inhibition;c.F.T. = complement fixation test; E.I.A,. = electroi
'noÍrssalï ELISA = enã,me-linked
immunosorbent assay; HA. = haemagglutination;R.I.P. = radiåi--uno precipitationi W.B. = Westernblot

Tablet2,4 227
COMPARISON OF PNET]MOCOCCAL ANTIBODY LEVEI,S IN 3 INTRAVENOUSIMMI]NOGITOBT]LIN PREPARATIONS
PnernococcalSerotype
nø Anfihn¡lv N/mlIntragam Sandoglobnlin Çamm¿g¿¡d
t
2
3
4
6
7
8
9
12
L4
18
t9
23
25
5667
n7t
6745
89M
3550
631i
2%L
3082
3225
3383
5807
2588
59L9
M
5700
L693
313()
7ffi
1893
M7
t91i
2%2
269L
2538
3329
1830
5W2
n38
5338
28t0
5531
9130
3180
4231-
2226
?ß93
3060
2975
5078
2625
6r,90
51i2

Tab1e 12.5
Lymphocyte studiesT cells (å)-erythrocyte rosette forming-cD3-cD4-cD8B cells (8)-surface Ig
Lymphocyte transforrnatíona to-phytohaenagglutinin-pokeweed mitogen-concanavalin A
Neutrophil studiesQuantitative iocinationb-pooJ-ed serum-autologous serl:m
Bactericidal activityc(8) kiltedChe¡ootaxis (nrn/3hr)
69665515
63665548
LYMPEOCYTE, NETIIROPEIL ÀlID COMPLHTÍEI.IT STUDIES rN IrrpOGÀtfMAcr¡BULINÀ.E[IC PÀTIBTTS
TN MWBN
(s)e
0.85o.2Lt_ 18
79
15,00027,6873L,82I
91
L.7
1.300.40123
16
23,74937,54329,r3L
87
t.23
0.59(0.86,1.01)d0.11(0.16,0.13)79
89
57
roo ,4r229,5656L ,620
97
2.0r
SL
13
228,960L43,369248,563
99
t.7 4
1.330. 18L27
Normal range
45-6548-8824-699-46
5 -15
40,958-375 trr222,379-3L4,90I2L,656-229 ,479
>80
>r.2
o.76-L.760.14-0.2865-13 5
7.O6.2
6.65.7
>3.0>3.0
6.36.5
1a
6.9
Serum complernentc3 (g/L)e4 ß/r)Tota1 heroolytic
a cpn of "H-t44pipine incorporated into_stinulated lyrnphocytes¡ piconoles of'125r incorpoiated per 107 neutrophils'pãr hourc percent of Staphvloceccus aureus killed in 2 hoursd nunbers in parenthesis show resul-ts after recovery from acute illnesse percent of normal serun pool
N)N)co

229
Neutrophil numbers and functions were normal (Table 12.5). Natural killer (NK) ceu cytotoxicity
was reduced (Ftg. n.D. CMI skin testing (Multitest, CMI, Institut Merieux Lyon, France) to g antþens
was non-reactive. Salivary IgA was undetectable. Senm C3 and C4 were minim¿lly reduced during the
¿ç¡[s illns55, but subsequently normalised (Table 12.5). Serology for HrV was negative. Open lung biopsy
confirmed P. coinü pneumonia. Small llm.phoid follicles were noted, with a paucity of plasma cells.
Trcatment. Int¡avenous immunoglobulin v¿5 begun ari soon as the hypoganmaglqþrrlinaemia was
detected. He received S00mg/þ of Intragam, (CSL, Austratia) by a slow int¡avenous infusion. A
precþitous drop in his neutrophil count was detected the following day. At first, this was attributed to the
intravenous 5"þhamethoxazole-himethoprin that he was receiving for his infection and he was therefore
çþenged to pentarnidine instead. His neutrophil count returned to normal very quickly and he was able to
tolerate further 5r'lphamethoxazole-trimethoprim without adverse effects. It, therefore, seemed more likely
that the Intragam had precipitated the neutropaenia. IVIG infusions were continued but with
Sandoglobrrlin, (sandoz, Basle) at a dose of 300mg/þ/month. The effects on his seru,n IgG, IgG1, IgG2,
IgG3 and IgG4 are shown nFþtre12.2.
Results. Trotrgh concentrations of his IgG subclasses are shown in Figure 12.2. Iñz concentrations have
been well maintained" IgGl- and IgG4 have generally been in the lower part of the normal rsngê , and IgG3
troughs have always been below the normal range for age.
This child made a slow but steady rscovery from his P. ca¡inü infection. Since then, he has enjoyed
very good health lor ?n monfhs. He received prophylactic snlphamethoxazole-trimethoprim twice daily for
18 montls, and has remained well since stopping fhis ¡e¿¡.ly 12 months ago. Sym.ptom score records and
clinical assessments have not indicated any abnormal infection proneness.
U¿3.12 Patient BN (IgG deficiency)
Clinical Presentation: BN is the older brother of patient TN. His imm¡¡qglqþulin concentrations were
measured when TN's hlpoganmaglobrrlinaemia was discovered. Before the results were available BN had
developed lobar pneu-onia. He had a past history of recu¡rent otitis media until age 4 years and had had
one previous episode ofpnernonia.

230
so
40
+
{
10
025 50
MNL ( x104 )
F¡g. 12.1 NK cell cytotoxicity in patlenÇ TN, age 10 months (o) and brother, BN, age 6 years (O).
(O) mean t SD (bars) of NK cell cytotoxicity for normals (22 iudividuals). Cytotoxicity nas measurcd at
monouuclear leukocyte (MM,) targeEratios of 5:1, 10:1, 25:l and 50:1.
r
30
20
õXoo
Oòe
r
5

23r
T.N ¡mmurþgbbuf¡n ¡nftJsiqì
lil I II llI Ill Iil l II IltTotol
lsG
(s/l) '
6
5-2SO
lscl
b/tl
lsG2
h/tl
-P5
o.2
lsG3
b/ll o'i
lsG4 .b/tl
24AGE (months)
2
I
{3
2
2
t2
Fw n2 Tboueù concenEations of þG and þG subcrasses rn padent rN.

232Investig¡ations. At age 6 years, he was found to have an ortremely low serum IgG concentration with very
low concentrations of all the IgG subclasses, but with normal concent¡ations of IgA and IgM (Table 10.1).
He had normal B and T cell proportions and in wfro responses to mitogens (Table 12.$. NK cell
cytotoxicity was reduced (Fig. 12.1). Neutrophil numbers and functions were normal (Tabte 12.Ð. CMI
skin testing (Multitest, CMf, Insfi¡¡t Merieux, Lyon France) was positive to only one of the 8 antþens
tested. Salivary IgA,,8, C4 and CH50 were normal (Table 12.5).
Treatment. BN was given an initial dose of Sandoglobulin, 500mg/þ by slow intravenous infusion. His
pnerrmonia was t¡eated with cotrimoxazole. Subsequently he has received 300 mg/kg/month of
intravenous Sandoglobulin.
Results. Trongh concentrations of his IgG subclasses are shown in Figure 12.3. IgG2 and IgG4 trough
concentrations have been within the normal tange, IgGl borderline and IgG3 below tle normal range for
age. Clinicall¡ BN has been given antibiotics for bronchitic symptoms on 6 occasions in the 20 montls
5i¡çs sfafting immunsglsþulin infusions. He has had pneumonia once and, Streptococcus pneumonia¿ a¡d
Høemophilus influenme have been isolated from his sputum, each on one occasion, since he has been on
immunsglsþtrlin ¡spl¿çsment therapy. As this boy did not appear excessively infection-prone in the 20
montls immediately before sf¿¡ting immunoglobrlin replacement, and has not appeared excessively
infection-prone in the 2 yearc afterward assessment of the effectiveness sf ¡¿i¡¡aining improved IgG
subclass concentrations is diffisult to assess. The value of treatment in terms of reduced risk of serious
overwhelming infection is diÊEcult to quantitate. Nevertheless his general health is good and his lung
function excellent.
l:23.L3 Patient SL (IgAand þG deficiency)
Clinical kesentation. This infant, born to consanguinous Lebanese patents, first presented. at age 2
montls with une¡plained generalised convulsions. She was developmentally normal at that rime but a
computerised tomographic (CT) head scan showed widespread linear and nodula¡ intracerebral
calcification in all lobes of the brain except the left frontal and occipital. Serological sç¡ssning for
int¡auterine infections (toxoplasm4 cytonegalovirus, herpes simple¡C rubella and slphilis) and for HIV
infection was negative. Her convuslive tendency was controlled with oral phenoba¡bitone. She was
considered to be doing wellwith normal g¡owth and deveþment until she presente{ agaiq this time with

233
knn¡noglobul¡n infu sion
B.N. lil lllllllllil lllil6
lsGl
b/tl
Totol
lsG
b/tl
o.2lsG3
(s/l) o't
2
I
3
lgGd 6.¡
(s/l)
65 7
AGE (yeors)6
-P5
FtC. 123 f¡'sllgh concentratfons of IgG and IgG subclasses tn padent BN.

234pneumonia and hepatosplenomegaly at the age of 7 months. The aetiology u/âri uncertain and she appeared
to improve with a short course of sulphamethoxazole-trimethoprim.
Immunologtcal Investigations. At age 7 months, her serum IgA was undetectable by nephelometry, by
RID on a low concent¡ation RID IgA plate, and by an IgA ELISA. FIer serum IgG concentration was very
low as were all her IgG subclass concentrations. Her serum IgM was at tle lower part of the normal range
(Table 10.1). Anti-IgA antibodies were sought using Ouchterlany's double imm¡¡sdiffusion technique but
the result was equivocal. T and B cell proportions were normal. T cell responses to mitogens were good
(Table 12.5).
Because this patient had a total IgA deficiency and we could not exclude the possibility of anti-IgA
antibodies we elected to begin immunoglobulin replacement therapy with an immunoglobnlin preparation
so¡faining only trace amounts of IgA in order to try and preclude a possible anaphylactic reaction. The
preparation chosen 1¡r¿s Çamm¿gard (Traveno[ California).
Initial Treatment. She received a loadi.g dose of 600mg/kg over a 5 hou¡ period. Apart from a transient
tachycardia early in tle cou¡se of the infusion, tlere were no clinically apparent side effects. Her CH50
levet however, fell from l73Vo before the infusion to <l7Vo immediately afterwards. The effects on her
serum IgG and IgG subclass concentratiorui are shown in Figure 12.4.
Further Investigations and Results. Two weeks later, she developed a febrile illness with convulsions. The
initial diagnosis was otitis media. However, she remai"ed febrile after several days of antibiotic trearment,
and then bec¿me tachypnoeic and had recurrent convulsions, which were very difficult to control. Her
overall condition deteriorated profoundly. Her serum IgG and IgG subclass concentrations were still
within normal limits for age at the onset of this illness.
Chest X-ray showed eÉensive bilateral alveola¡ consolidation with air bronchograms. P. cañnü
was isolated from her respiratory secretions. Cerebrospinal fluid contained 7 lmphocytes/mm3 with a
raised protein of 1.35 g/l (normal range 0-0.4 g/l) añ a normal glucose conceut¡ation.
A CT head scan showed a gross abnormality in the nidbrain region in addition to the calcification
seen previously. There was an unusual large hlpodense lesion producing slmmetrical enlargement of the
upper brain stem extending into the thalamus and the internal capsules and corona radiata. A second
infusion sf Çamñagardlnng/kg, was given. Drug treatment included sulphamethoxazole-trimethoprim

235
immunoglobul¡n infusbns.L. llllillillltI
Totol
lsG
b/tl
lsGì
b/tl
7
6
4
3
2
6
5
4
2
1
-
_2SO
-Ps
lsG2(s/l)
o
o'3
lsG3 o'z
ls/tl o.l
t2A-GE (months)
7 r8
ßWf2,.4 Tlongh concentrations of þG and þG subclasses in patient SL.

236and folinic acid for the pneumocystis pneumonia. Pyrimethamine was added as active toxoplasmosis was
considered a possible cause of the brain stem lesion because her mother's toxoplasma titres were sligbtly
raised.
Fu¡ther immunological investþations included determination of the presence of HLA-.\B and DR
antigens on leucocytes to exclude bare llmphocyte slmdrome. Antþens of each tlpe were present, tle
pattern being HI-AAIL; A281' B35; 850; DR6; DR1-1.
Fufher progress and tr.eatment Six weeks after the onset of this illness SL was discharged. She had
made a very poor reoovery and was hypertonic and unresponsive and required nasogastric fseding. Her
convulsive tendency was controlled with phenobarbitone and pheyntoin. Her respiratory symptoms had
improved. Sulphamethoxazole-trimethoprim was continued for 6 montls. She received regular inft¡sions of
Gammagard, úffing/kg initially, and then at 2,3 and 4 weekly intervals. After 3 6e¡ths the dose was
decreased to 300mglþ, and after 9 months the frequency was decreased to 6 weekly. Trougü
concentrations are shown in Figure 72.4. Nl serum IgG subclass trougû concentrations including those of
IgG4 were maintained within the normal ¡attge for age on 4 weekly Intragam infusions, and have begun to
fall since the frequency of infusions has been decreased. {s Çemm¿gard has a very low IgG4 "oa¡s¡¡
this
is surprising and suggests a prolonged IgG4 half-life or tle spontaneous production of some IgG4 in this
patient. ClinicallS SL has not done well. The child is now microcephalic, spastic and quadriplegic. In the
L8 months since this severe ¿¡d dev¿st¿fing illns5s, SL has not shown any further sigrts of excessive infection
proneness.
123,1.4. Patient iUW (IgA IgG and þM deficiencV) (see Beard & Ferrante, 1988)
Ctinicat hesentation. This patient is now 15 years old. She presented at age 10 months with e:úensive
pneumonia and bilateral otitis media.
Immunological Investigation. Her senrm IgA concentration was low, serum IgG borderline low and serum
IgM borderline elevated (0.L g/lr2.Mg/lu andL.56g/Lrespectiveþ. She continued to suffer recurrent chest
and ear infections. At ?5 months of age, antibodies to tetanus toxoid and pertussis were undetectable
atthough she had previously received fou¡ standard doses of triple antþen. Re-immunization produced a
tetanus toxoid titre of L:32 and a pertussis titre of 1:12f1. Immunization with t¡re 3 pnenmococcal capsular
polysaccharide gave a very poor response. In her fourth year she had five episodes ofpneumonia. Serum

237IgA and IgM were by then undetectable and IgG was very low. At age 9 years, neutrophil functioq
lymphocyte ¡'ansformation to mitogens and serum complement studies were normal (Table 12.5). IgG
subclass levels were first measured in this patient at age 72 yearc immediately before one of her regular
doses of intramuscul¿¡ immunsglsþrllin. (Table 10,1) f¡6rgh concentrations have been repeated on
subsequent occasions. Concentrations of IgGl, IgG2 and IgG3 have been low on all occasions and IgG4
was low or low normal.
Treahent. MW received intramuscr¡l¿¡ imm¡nsglsþulin (IMIG) replacement therapy, 1-00mg/þ every 2
weeks from age 4 years. She elected to continue on intramuscular therapy even after IVIG became
available.
Results. After starting IMrG, MW improved considerably. For several years she continued to have one or
two brief episodes of chest infection per year, but generally, she was quite well, whereas previously she had
been chronically ill. In recent years her main problem has been as'hma which has been well controlled with
bronchodilators. She attends school and leads a normal active life. Lung function tests have shonm
reversible bronchoconstriction. The persistent changes seen in her chest radiographs in her early years
have disappeared almost completely.
This patient with common va¡iable immunodeficiency and low concentrations of IgG1, IgGZ and
IgG3 has benefited from IMIG replacement therapy. She has tolerated very large volumes of IMIG
extremely well and elected to continue with IMIG rather than to change to fVIG, largely for reasons of
convenience. On rhis treatment, she has not appeared to be excessively infection-prone.
l:¿,32. IgG subclass replacement in IgG subclass deficient patients
t232.1 Patient JB (sec rhong and Maxwell ,lllS; Beard and Ferrante, 19BE)
Clinical Presentation. This girl e:rperienced severe recurrent infections from age 4 months. These
included otitis media, sinusitis, bronchitis and pneumonia. Pyogenic organisms, generally S. pneumoniae,
H, ínfluenzae or Staphylococcus Øtrelrs were frequently isolated. When she was agedT years a lobectomy
was done for bronchiectasis. By age !9 years, she had chronic sinusitis and suppurative bronchitis which
was worsening despite the constant use of antibiotics and regular physiotherapy. Lung function tests were
poor, and x-rays indicated widespread pulnonary disease and ch¡onic sinusitis.

238
rmmunological Investigations. JB was thought, initially, to have a selective IgM deficiency with serum IgM
concentratio¡g ranging from 0-0.3g/l between ages 7 and 11 years. She responded normally to
irnmunization with tetanus toxoid but not to Salmonella typhi H or O (Thong and Maxwell, 197g).
Lymphocyte and neutrophil function studies were normal (Table 12.6). l-ater. following imm¡¡i2¿¡i¡n with
a 14-valent pneumococcal vaccine she failed to produce antibodies to all 14 serotypes (Table 12.7).
over the years the pattern of her i--unoglobulin abnormality sþnnged (Fig. 12.s). By age 19, her
senrm IgA,, IgG and IgM were all in the low to low-normal ra¡ge. df fhis tims, s/s were able to measu¡e
her senm IgG subclass concent¡ations, and found that these of IgGl, IgC2 and IgG4 were low (Table
10.1).
Treatment. IVIG therapy was begun with a loadirg dose of 300mg/þ of Normal Human Tmmungglsþrrlin
(CSL, Austrelia), follswsd by a dose of SÙmg/kg every 2 weeks for 6 montls, 5Omg/kg 3 weekly for 8
months, and50mg/kg5 weekly for 14 montls.
Results and Further Management. Wifhin fou¡ months of beginning IVIG her ch¡onic productive cor¡gh
had resolved and her exercise tolerance had improved. Her sinusitis had improved dramatically for the füst
"me she was well enotlgh to begin part-rime work. However, for JB, IVIG infusions were not problem
free. Her fea¡ of needles which was very rliffiçul¡ to overcome. During infi¡sions of Normal Human
Tmmunsgleþulin she e4perienced, invariabl¡ malaise, chest tþhtness and dyspnoea and some';mes muscle
pains. For this reasoq the dosages were restricted.
After 2 years therap¡ Sandoglob¡li¡ 3Vo (Sandoz, Basle) was substituted for Normal Htrman
rmmunoglobulino and untou,ard symptoms during infusion lessened considerably. Frequently, she
complained of aching in the arm receiving the infr¡sion but malaise, chest tþhtness, dyspnoea and muscle
pain ceased. Previously, we had had clifficulty in increasing the dose of imm¡¡sg.6ulin given because of
side effects, and even with Sandoglob'lin, doses were limited to 100mg/þ 3 weekly because of local aching
a¡ound the infusion site. The effects on her IgG subclass concent¡ationri are shown i" Fig. 12.6 which shows
a series of recent values.
IgG subclass trough concentrations were not maintained within the normal range in this patient.
She has continued to have problems with recurrent bronchitis. She also has asrhma. Her infective and
asthmatic slm.ptoms are not always easy to separate. flowever, since beginning immunoglobrllin fftç¡¿py

Talc1e 12 - 6
Lynphocyte studiesT cel1s (8)-erythrocyte rosette
forning-cD3-cD4-cD8B cells (Z)-surface Ig
Lynphocyte transformationa to-phytoha ernagg lutin i n-pokeweed mitogen-concanavalin A
LYI'IPHOCYTE, NE(]1IROPHIL AND COUPLH{EI{I STUDIES IN IgG SUBCIÀSS-DEFfCIn|1I PÀTIEIflIS
RL Norna1 rangeAWBHJB
532t
14
59,31133,39689 ,L69
48
99
L.6
50 62
L7
23), , 47 6L24,94739O ,43L
51
13
362,OgL229,728365,322
1.5
L.790.1_3L73
45-65
48-8824-699-46
5-15
40 ,958-375,L]-222,379-314,9OL2L t656-229 ,479
>80
>L.2
0.76-L.76o.I4-O.2865-l_ 3 5
232,379],44,832206,768
L.040.1_595
Neutrophil studiesQuantitative iodination-pooled serum-autologous serum
Bactericidal activityc(?) kiIledChe¡rotaxis (rnm7'3hr)
b>3.0>3.0
7.O5.6
6.1_7.9
8.37.9
2.64.6
o.77- L.7 4
Serun conplenentc3 (s/L)c4 (s/L)Tota1 henolytic (8) d
0.900.432L7
0.850.1050-
a cpm of 3H-tr1'¡pifline incorporated into-stimulated lymphocytesb picorootes of- 125r incorpoiated per 1,07 neutrophils pàr nburc percent of Staphylococcus aureus killed in 2 hoursd percent of normal senrn pool- reduced
N)(¡)\o

Table7i2.7
RESPONSES TO PNEIIMOCOCCAL IMMUNIZATION IN3 IgG SUBCLASS-DEFICIEI\T pATIENIS
240
Pne'mococcalseroty¡le
Fold rise in antibody concentration following immunization
Normal meân*
r.73.92.97.23.98.25.86.8155.54.62.74.73.0
L.62.63.614.4L.63.4L.LL.7L.8L.4L.7L.4L.4
RL
L.42.6L.03.1L.28.94.63.71.5
1.8.5
L.4t.42.0L.3
JB
0.80.70.92.L1.8
0.50.70.80.81.01.20.40.40.4
1
234678912T4
18
L9
2325
AW
*Paton et al, 1986

gA 24L
ol
60
so
30
oo
20
r.0
o
o
o 5 ro !5 20
AGE (yeors)
lsG
r5
lO
ol
(,I
IIII
vg
5 tII
5 rO
AGE (yeors)l5 20
gM
2.O
:ol
ão
o
¡5
AGE (yeorsl
Ftg.US The changtng pattern of þ class deficieucy ln patient JB both before and lmmedlaúely after
Etardry IVIG therapy. Solid llnes reprrcsent paüent values a¡d d¡shed llnes reprcsent the nonnal rangs
(meant 2SD)
5020

242
rmmunoglobultn tnluston
J8
2SO
Totol
lsG(s/t\
lgGl
b/tl
lqG3
(s/l)
P5
lsG4
G/t\o.1
o.05
23AGE (yeors)
++++{+{it++ilil1I7
6
5
4
2
I
I7
6
5
4
3
2
,|
3
\^
2
2
Fig. Í¿.6 f¡.sngh concentrations of þG and þG subclasses in patient JB.

243she has improved considerably as iirdicated by the fact that n lhe 2years before starring immunoglobuli'
therapy she spent 53 days in hospital and in the 2 years afterwards only 4 days.
Summary. This patient, origl"ally considered to have a primary IgM deficiency, was subsequently shown to
have IgG subclass deficiency. The use of IVIG has improved the quality of her life considerabl¡ but was
started too late to prevent ch¡onic lung damage. It appears that some of the side effects of IVIG which
previously limited her therapy could have been avoided by using Sandoglobulin. Brearley and Rowbotham
(1984) reported that the lgp agregate content of Sandoglobulin was considerably less than that of normal
human immursglsþuli" (CSL, Australia). The newer ¿¡d improved CSL preparation, Intraga-, was not
available when this patient began therapy.
l:2322 Patient RL
Clinical Presentation. At 16 years of age, RL was referred for assessment of a lifelong tendency to
respiratory infections. He had been receiving antibiotics almost continuously for over 6 years to try and
control his recurrent sinusitis, pharyngitis and suppurative bronchitis. Nevertheless, he was frequently ill
with infections and f6¡ fhis reason was missing a great deal of school. Also, he had a tendency to mild
¿srhm¿, and allergy to several grass pollens.
rmmunslsgical Investigations. He had a variety of immunological abnormalities. He had partial
deficiencies of serum Igd IgG, lgpL,lgG? and IgG4. Serum IgE was elevated (Table 10.1). Salivary IgA
concent¡ation was normal. Neutrophil chemotaxis was reduced on two occasions and serum C3, C4 and
CH50 were also reduced. T and B cell percentages and lymphocyte respoffies to mitogens were normal
(Table 12.6). Specific antibody titres to tetanus and diphtheria were low despite previous immunization.
There was a good response to reimmunization. (Baseline tit¡es: tetanus 16, diphtheria L0; post
¡eimmrrnization titres: tetanus 1û21, diphtheria ,10). He was immunized with Pneumovax (MSD, CSL,
Australia) and showed a moderate response to most serotypes with the exception of types 4, L6 and, L9
(Table 12.7).
Tbeatment. At age 16.2yearc he received his first inft¡sion of [VIG, Normal Serum rmmunsgleþulitr (CSL,
Australia), lffing/kg. The slow infr¡sion was unevenú¡l but within 30 minutes of its completion he began
to feel ill. He sþt for about 10 hou¡s and woke with ma¡ked lethargy, malaise, and aches and myalgia. He

244þ¿d ¿ single episode of urinary incontinence. On examination tlere were no objective signs except that his
bloodpressurewaisalittlebelowhisnormal (n/50 instead of f0a/70). Aspitio600mg6hourly,wasbegun
and he improved slowly over the next few days.
After a 2 month interval monthly infusions of Sandoglobtlì¡,?û}mgfkg were begun and continued
for 6 montls. He then elected to have a t¡ial without treatment. After the füst Sandoglobulin infusioq RL
experienced lethargy and ti¡edness but was not nearly as incapacitated as he had been after his infusion of
Normal Human Immunoglobulin. Subsequent infi¡sions were unevenû¡I.
Results. The effect of these inftrsions sa his IgG isotype concentrations is shown in Figure 12.7. IgGL and,
IgG2 concentrations normalised on t¡eatment, but IgG4 trough concent¡ations remai"ed borde¡line low. In
the 12 m,onths,after stopping immunoglobrlin treatment, his IgGl and IgG2 remained within the normal
range. Neutrophil chemotaxis and CII50 were normal 5 months after treatment was stopped. Throughout
the 8 month period of imm¡¡eglsþrrlin infusions, RL received oral sulphame¡þsxazole-trimethoprim. He
had three sinopharyngeal infections during this period requiring additional antibiotic treatment. This,
however, was an improvement 6¡ his pretreatment state. After IVIG was discontinued he showed no signs
of his earlier infection proneness.
It is impossible to assess the clinical effectiveness of IVIG in this patient. In the 12 montls prior to
treatment he had almost constant pharyrngitis and sinusitis. During treatment, he inproved and after
stopping treatment he improved even more. His IgGa levels were low or low-normal before during and
after t¡eatment.
lj2323. Patient BH
Clinical kesentation. In his first year of life, BH e4perienced bronchiolitis followed by four episodes of
right upper lobe consolidation at ages 5, 6, 7 arld 11 months. He had herpes simplex stomatitis once
S. pneumoniae and .F/. influenzae were isoliated from nasophar¡mgeal secretions. Although he was
considered ¡e þe ¿srhmatic these episodes were thougbt to be largely infective and settled with antibiotic
therapy. He was also prone to superficial skin i¡flsçfisns and had mild eczema. He did not have
gastroesophageal reflux
Immunological Studies. Tmmune function studies were normal (Table 72.6) apafi from a low serum IgA
concent¡ation, (0.09g/l; normal range 0.?-LL.L4E/D, a raised serum TgE concentration (150 i.u./ml; normal
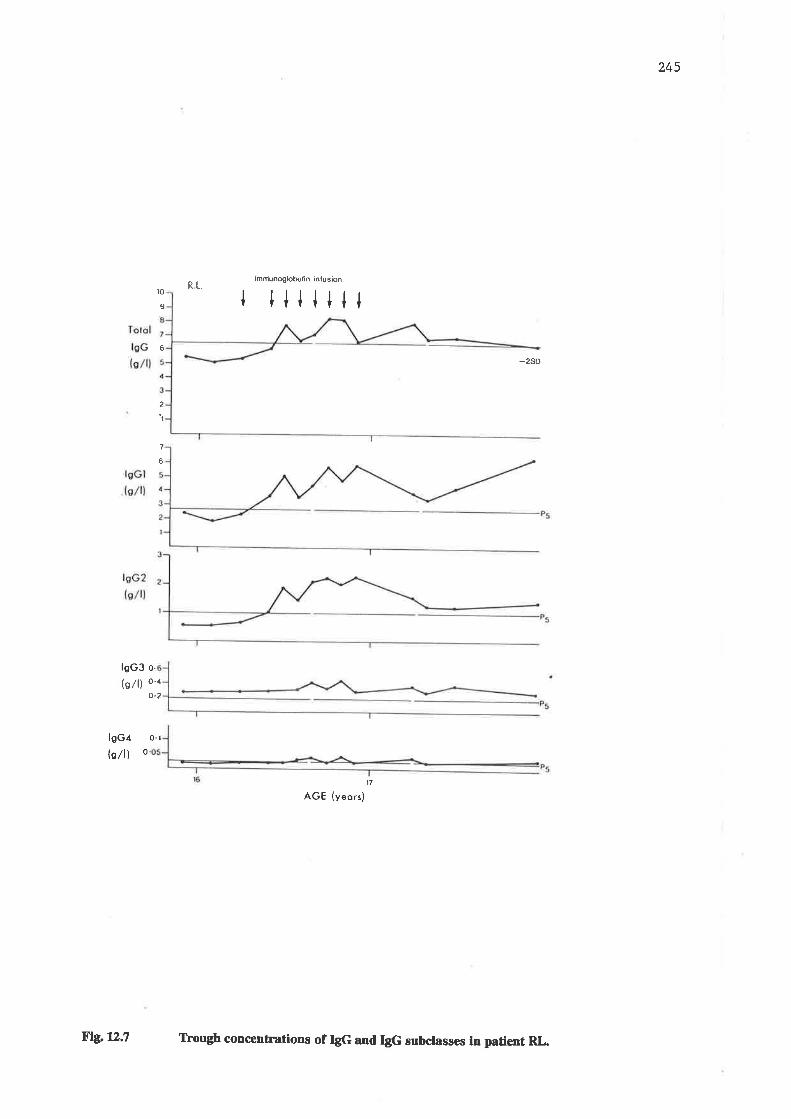
24s
¡mftiunoglobutin inlus¡onRL.
to
9 l illilll6
-2SD
2
I
7
6
lsG3 o.
(o/l) o'ro.2
lsG/
ls/ll o
0.1
t7
AGE (yeors)
ßlgx¿.7 frougb concenûations of þG and IgG subclasses in padent RL

246< 16 i.u./ml) and a borderline low IgG4 concent¡ation (0.01g/l; 5th percentile for age 0.009g/l). Initially he
was considered to be IgG,l-deficient but improvements in our normal ranges resulting from the inclusion of
more normal samples have shown that he had low-normal ratler tlan subnormal IgG4 concentrations.
Treatment. In view of his worrying clinical history and what was, at the time, considered to be an IgG4
deficienc¡ BH began monthly intravenous infusions of Sandoglobulin 300 mg/kg, at age 11 months. These
were continued for Íi moutls. The actual dose he received relative to his body weight gradually decreased
as he continued to receive 39 lots of Sandoglobrlin.
Results. The effect of these infusions on his IgG subclass conc¡nt¡ations is shown in Figure 12.S. IgGl
concentrations_were above the normal range for age before IVIG was begun and trough concentrations
have remai"ed so. IgG2 was in the low normal range prior to the commencement of therap¡ and on IVIG
trough concentrations were around the mean for age. IgG3 concentrations were pithin the normal range
for age before IVIG was begun, ¿¡d ¡¡¡ngh concentrations have remained well within this range. IgG4
concentratiorui were low uormal before fWG was begun. Trough concentrations remainsd low normal
during the early period of the IVIG infusions, but began to rise a little after 8 months.
Serum IgA was below the normal range at almost ¿ll tims5. I{owever, tlere were rises in IgA
concent¡ation detectable immsfi¿¡ely after IVIG infusions indicating tle preseuce of substantial amounts
of IgA in Sandoglobulin.
During the 13 montl period of his infusions, BH had no episodes of bacterial pne 'monia. He had
one episode of respiratory infection during which respiratory spcitial virus was isolated 8 months after the
onset of the IVIG therapy. He had several episodes of asrhma requiring hospital aclmission but without
evidence of an infective component.
In the 7 months after his Sandoglobulin infi¡sions were discontinued he had 2 episodes of right
upper lobe pneumonia, one of streptococcal pharyngitis, one of giardiasis and several viral URTIS. It was
decided ¡fr¿f ¡sgumption of IVIG infusions was warranted even although his IgG4 and also his IgA
concsntrations were now in tle normal range.
l:232.4. Patient AIV
Clinical Presentation. This 11 year old boy has a long history of coughing and whee"irg going back over
many years. At age 7 yearc, he had a persistent right niddle lobe collapse and bronchoscopy showed some

247
B.H. immunoglobulin infu sircn
il llil ltil lllltgGl
G/tl
lsG2
ls/tl
lsG3
b/tl
10
0.
o.
o. 4
P5
_Þo
o-06
o.05lsG4 ,(s/l) o
0
0.01
18
AGE (months)2112 30
Ftg. 1lÌ.8 TÞough concentrations of þG subclasses in patient BH.

248narrowing of the right middle and lower lobe bronchi. At age L0 years, he developed left upper and lower
lobe collapse with consolidation and sputum grew H. ínftuewne. Blood cultu¡es were negative and viral and
Mycoplasma pneumoniae tit¡es were not elevated. He was t¡eated aggressively with bronchodilator therapy
and antibiotics but his chest signs persisted for some weeks. Aw had a history of both ¿5rhm¿ ¿¡d
recurrent infection. He hardly had a day without cough or wheeze. When he was youngor he had been
prone to ear infections. There was no history of eczema or hay fever.
His motler has chronic chest disease and has been found to have multiple endobronchial webs.
Her respiratory function is grossly compromised and she is tachlpnoeic at rest. She has been prone to
asthma and recurrent infections all her life. His father has a history of ear problems and had mastoiditis as
a child. Like his mother, AW has been found to have endobronchial webs affecting the right middle lobe.
AW is a well groum lad on the 50th percentile for weight and height. Apart from variable chest
sig¡s at different phases of his history there have been no abnormal physical finrtin$ of note.
Chest x-rays have shown varying degrees of lobar collapse ardf or consolidation. After he had
recovered from pne'monia at age 1"0 \f2 years his ftings appeared clear apart from slþht residual
atelectasis in the lingular segment of the left upper lobe which may have represented sca¡ tissue. At age 1l-
years he developed further liogutat opacity consistent with collapse and consolidation in the left upper lobe.
Immunological Investigations. At age 10 7f2years, after the prolonged exacerbation of his chest disease,
AW was referred for immìmelogical testing. His immunoglobrrlin concentrations were abnormal in that he
had persistently low serum IgA, IgG, lgpL,lgG2 and IgG4 (Table L0.1). He was immunized with 14 valent
pneumococcal vaccine and failed to show any response to the serotypes included (Iable 12.7). His
lymphocyte functions and neutrophil functions were normal (Table 12.6).
Family studies were also done. AW's mother was found to have low levels of IgG1, IgG2 and
IgG4. She also had reduced lymphocyte responses to srimulation with the mitogens PHA and pWM.
Tleatuent. In view of AWs clinicål history and the evidence of IgG subclass deficiency and impaired
antibody production he was begun on a trial of IVIG replacement therapy at age 11 years Infr¡sions of
3ffimg/kgof Intragam (CSl Australia) were given at monthly intervals.
Results. Since starting IVIG 19 months ago, AW has had 5 further episodes of lobar collapse and,/ot
consolidation. Ifowever in between these episodes his cough and wheeze have been less prominent tlan

249
. lmmunoglobulin infusionIrilIilIIIIIIIIIo)
oI
:o)
oI
%
O)
C.loI
o,
cooI
10
B
6
4
2
0
7
6
5
4
3
2
11 12
12
12
Pz.s
5
10
10
10
10
113.0
2-O
1.0
o1.1 12
0
o.5
o'4
o-3
o.2
0.1
0
0
o.2
o'5
'11
0.1
'o5'04.03
o.02
o1
o0o
o
I.çt()I
Ã
ofo 1t 12
AGE IN YEARS
F'rc. 12.9 Trough concenEations of IgG and IgG subclasses in patieut AW.

250tley were before he started ¡y¡6 f¡sngh serum IgG concentrations have been maintained sligh¡ly below
tle normal lower limit for age. IgG2 and IgG3 trough concenhations have been within the normal range
but IgGl and IgG4 tro'gh concentrations have remained subnormal (Ftg. n.Ð.
It is difficult to assess the effectiveness of IWG replacement therapy in this patient witl common
va¡iable immunodeficiency as, despite improvements in serum immì¡nsglsþulin concentrations, he
continues to have st¡uctu¡al pathology predisposing him to respiratory infections. A longer period of
observation broken into comparable periods with the patient acting as his own cont¡ol with and without
immunoglobuli" replacement may help to clari$ the situation. Clinically, such a trial is hard to justrfy as
his risk of severe progressive lung disease is great, he has been shown to be antibody-deficient and IVIG
does appear to be helping him to some er(ent.
I:2.4 SUMMARY
(a) It was possible to maintain IgGl and IgG2 concent¡ations within the normal range for age in
hypogammaglobrrlinaemic patients with monthly infusions of intravenous immunogl6þrrlin of 200-
300mg/þ/month.
It was not possible to maintain normal IgG3 concentrations with his regime, pres'mably because
of the shorter half-life of IgG3 (Morell et al 1970). Trouoh concentrations of IgG4 were also
difficult to maintain. The low concentrations that a¡e normal in young child¡'en were achieved as
shown in TN, BN, SL and BH, but the highs¡ concentrations that are normal in older child¡en
were not achieved as shovm in JB, RL and AW.
(b) There was no detectable difference in the trough IgG subclass concentrations achieved with the
th¡ee different preparations of intravenous immunoglobulin that we used. All in 5imil¿¡. dsss5,
appeared suitable to replace IgGl- and IgG2. Neither Sandoglob,lin nor Çamm¿g¿r:d maintained
IgG3 trough concent¡ations in the normal range. Intragam was not used in IgG3-deficient
patients. Surprisingly and ine4plicabl¡ the only patient receiving Çamm¿g¿rd a preparation
sonfaining very little IgGa, naintained normal t¡sngh IgG4 concentrations. prolonged half-lives of
IgG subclasses have been reported in hypogammaglobulinaemic patients (Ochs et al, 19g6).

G)
251It is extraordinarily difficuLlt to correlate the patient's health with his/her IgG subclass
concentrations, as the following findings illustrate:-
(Ð Patients TN, BN and SL have not been excessively infection-prone while on
immunoglobt lin replacement tlerapy, despite low IgG3 concentrations.
(ü) Patient MW has been well despite subnormal concentrations of IgG subclasses.
(-) Patient JB has been less infection-prone while on treatment, despite persisting lgp1 and
IgG4 deficiencies.
(Ð Patient RL who was abnormally infection prone and lñ2-Igc+deficient prior to therapy,
improved somewhat while receiving therapy with normal IgG2 concentrations, and has not
been subject to infective problems in the 22 months since treatment was discontinued,
although IgG2 concentrations ate lower than during treatment and his IgG4
concentratio¡5 ¡s¡ein borderline low. Improvement in his neutrophil chemotaxis may be
a contributing factor in his improvement.
(v) Patient BH who had recurrent pnermonia before treatment and a low-normal IgG4
concentration had no bacterial pne'monia during the year of treatment and his IgG4
concent¡ations normalized. Subsequentl¡ despite normal IgG4 concentrations he began
to have recurrent pneu'nonia and after 7 monfhs, fVIG treatment was resumed this time
with Intragram and with good effect so far (4 months). Despite the presence of normal
concent¡ations ofIgG subclasses a restricted antibody deûciency needs to be considered in
this patient.
(oi) Patient AW has continued to have recurrent lung disease in the 19 months that he has
been t¡eated with fVIG, despite normal trough serum IgG2 concentrations and in the
preseûce of low IgG4 trough concentrations. However, he has structural bronchial
pathologywhich helps e4plain his recurrent pnenmonia and atelectasis. The improvenent
in his interval slmptoms perhaps indicates a worthwhile improvement with the use of
immun6gl6þ'lin ¡spl¿ce6e¡1.

CHAPTER THIRTEEN
IgG SUBCLASS CONCENTRATIONS IN SALTVA ANDCEREBROSPINAL FLUID

25313.1. NTR.ODUCTION
Because, in ou¡ studies, IgG4 deficiency was tle most common IgG subclass deficiency in patients
with increased susceptibiliry to respiratory infections (Ch. 5,6) and in patients with i¡vasive infections
car¡sed by H. influenzne (Ch. 7), and because IgG4 is a relatively minor isotype in serum (Ch. 1.2.2.4), we
sough¡ to address the question of whether or not IgG4, like IgA" might be predominantly a mucosal
immun6gleþrrlin. This possibility has been suggested by Keller et al (1983) who found an unexpectedly high
percentage of IgG4 in colostrum and by Merrill et al (1985) who found relatively higfr concent¡ations of
IgG4 in lower respiratory tract secretions.
In many of the patients with invasive infections caused by H. influenzae, serum IgG4 concent¡ations
were low, or relatively low (Ch. 7). Since most of these patients had msningitis, we thought it appropriate
also to determine the pattern of IgG subclass distribution in CSF to see whether there was a relative
prominence of IgG4 in this fluid which might be a sþn of a special role for IgG4 in defence against
bacterial 6sningitis.
L3.2 RESULTS
L32.I IgG subclasses in saliva
In this study we quantitated IgG subclasses and total IgG by ELISA in salivary samples f¡om 19
healthy adults. Saliva was collected and prepared as described in Chapter Z.!.2 and was frozen and re-
thawed prior to tssting. Resr¡lts were expressed as percentages of total IgG concentration because dilution
factors made meaningful comparisons of absolute concentrations impossible. Seru'n samples f¡om all the
subjects except two were also analysed.
The results a¡e summarised in Table 1Í1.1 and in Figures 13,14. Total IgG concentrations varied
considerably raning ftom 2.2 to 54.5 mg/I. It can be seen that the mean percentages of IgGL and IgG3
tended to be lower in saliva than in serum while IgG2 and IgG4 percentages were higher. There were
marked variations in concent¡ation between samples from different patients and only for IgG3 was there a
sigrificant difference between salivary and serum concent¡ations (p=O.(X, Mann ffii¡¡ey U test). In

254individual patients, the percentage of salivary IgG3 was alrns5f always the same, or lower, than that of
serum IgG3 (Fþ. 1Iì.3) and for IgG4, the reverse was the case (Fig. 1iI.4).
132.2 IgG subclasses in cerebrospinal fluid
Ssmples of cerebrospinal fluid (CSF) from 12 child¡en in whom neningrtis was suspected but
disproven by CSF microscopy and culture, were sludied for total IgA\IgG, tgM and IgG subclass content as
measu¡ed by ELISA. For IgA and IgM, polyclonal antisera (Silenus laboratory, Victoria, Australia) were
used as described for IgG in Chapter 3.5.
Tmm¡¡oglsþulin 6f each of the classes IgA IgG and IgM and of each of the IgG subclasses was
detected in the CSF of each of 12 patients (Tabte 13.2). Concentrations were about 1000 times less than
those found in seru'n.
IgG and IgA concentrations were almost equal in CSF from 2 patients, a finding which would be
most unusual in serum where IgG is relatively more abundant. IgG subclass concentrations varied
considerably, ¿¡d ¿l¡fisugh there was an overall tendency for these to i¡crease with the age of the patient,
tfts highe5t levels of IgGL andI:gG2 were found in the you"gest patient, who was 14 days otd.
The relative percentage of IgGl v¿s higher than the mean relative percentage of IgGl in serum in
child¡en of ttre same age in LL of 12 5amples. Conversely, the relative percentage of $p2 was lower than
the mean percentage of. lgp2 in serum of children of the sâme age in 11 of 12 samples. The relative
percentage of IgG3 was lower than the mean percentage of IgG3 in senun of 10 of lJ samples. The
relative percentage of IgG4 was lower than the mean percentage of IgG4 in serum of 8 of 12 samples.

25513.3 STJMMARY
Quantitation of the low concent¡ations of all the IgG subclasses in saliva and in CSF was possible
by ELISA. In neither fluid was the proportion of IgG4 hieher than in serum. In saliva, the pattern of
distribution of the IgG subclasses did not differ greatly from that in serum. This suggests either
t¡ansudation from serum or local salivary production of the IgG isotypes in simila¡ proportions to those
found in sentm. In CSF, the relative proportion of the IgG subclasses differed f¡om that i¡ ss¡nm, IgGl
þsing relatively more abundant, and IgGz and IgG3 relatively less so. No conclusions could be drawn
concerning IgG4. Concent¡ations of the IgG subclasses in the CSF were about 1000 fold less than in serum.

zs6
Table l3.lIsG suBct Ass coMPoslrloN oF SALwA AND SERUM rN HEALTHY suBJEcrs
Subject Total IgGmc/lG/t)
IgGl lgGz ICG3 ICc4Vo
Fu
Ro
Ka
Zß,
Ha
Tu
Bac
Bat
Bae
Gr
CI
AI
Fr
Pi
Ja
No
Ste
Be
Ca
44.9
(11.8)
3.2(8.8)
5.3(5.5)
2.5
(11.6)
8.7(8.1)
2.3
(5.7)
45.7
(11.8)
35.2(12.1)
5.8(7.4)
25.2(14.2)
65(7.0)
t4.7(13.3)
54.5(6.Ð
L73(e.Ð
8(7)
(ó)
4
(6)
2(3)
3
(4)
3
(4)
3(7)
')
(2)
1
(2)
4
33
(24)45
(s6)
8.8(e.0)
I4(13)
60
(61)
53
(4e)
49(42)
x(36)
31
(28)
46
(33)
34(27)
50(65)
50
(s1)
42(40)
35(u)
43
(s2)
35
(4Ð
35(3s)
34(34)
23
(30)
2A
(25)
38
(32)
61
(60)
61(6Ð
63(66)
45(52)
1A
(31)
43(s2)
5
50
(54)
55(61)
50
(4e)39(42)
49(s3)
44(56)
:
58
45(3e)
4't(34)
i28
2(3)
(5)
22(31)
i3(8)
5
(2)
ð
(7)
a(ú)
L5
(10)
15
( 15)
L5
(8)
42.6
9.9
23.8(11.3)
2.2(10.8)
1
(3)
4
(2)
8
(6)
3(2)
8
(o
4
(7)
3
(6)
J(3)
3
(6)
2(3)
5
4
I(2)
7
I'8(1)
2
$9¡um values a¡e in pareutheses; - = not quantitated
(4)10
(4)

Table üt2 257
gC SUBCT,ISS,IgG,IgAAND IgM CONCENTRATIONS IN CSF'
Age IgGl IgGz IgG3 rñ4 IgG IgA IgM
(v.)
L4 days 23192 ß52 L042
7 mo. 3978 429(84)1 (e)+
25723 2f36 874
4762 4008 L65
7823 n4 498
(3)+(6)+(78)17N(2)+
L7L(4)+
LU(4)1
L0 mo.
72mo.
?3mo.
ù1mo.
29 mo.
6 yrs.
8 yrs.
11 yrs.
15 yrs.
16 yrs.
5854(83)1
5n4Qe)t
9252(81X
95L8(78)1
L7697(8L)1
50(0.Ð+
970 2LL(14)1 (3)+
9681 3109 3592(53)+ (17)+ (n)t
4(0.Ðr
7394 300 6L6
6322 6026 285
7783 542 358
148(3)+
5612 309 2L6
12690 290 329
12?32 308 y3
1960 L8100 ¿1000 7037(LL)1
46Lt 969(73)1 (15)+
4393(7e)î
1080(16X
301(4)+
L77(3)+
939 83(16)+ (1)+
1212 359(11)+ (3)+
L690 413(14)+ (3)+
2653(n)+
565(e)1
224(4)+
5¿lO
(5)1
654(Ð+
4993 722 293(80)1 (12){ (5)+
7422 ?ÁL0(64)1 (n){
859 513 23L76 156/. twz(3)+ Q)+
1089 5n [075 262 3,18(ex (Ð+
1 = ) mean serum pefcentage for ageI = ( mean serum percentage for age

258
% lsGl
70
60
50
40
30
20
10
t-l
Mean ! SD
SALIVA
lndividualvelues
lndividual
valuesMean !SD
SERUM
Hg. 13.1 þGl Ín salÍva and serum of 19 healthy adults (expressed as percentage of total IgG).

.259
60
50
o//o lsG2
Mean tSD
SALIVA
t-
__l
SERUM
Fig. 13¿ þG2 in saliva and serum of 19 healthy adults (exprcssed as percentage of total þG).

260
o//o
10
lgG3
Mean jSD
5
ïï
Mean tSD
SALIVA SERUM
lndividualvalues
Flg. Í13 IgG3 h salÍva and serum of 19 healthy adults (expressed as percentage of totat þG).

26I
lsG4
o//o
30
20
10
Mean tSD Mean lSD
SALIVA SERUM
lndividualvalues
lndividual
values
Fig. llt.4 IgG4 in saliva and semm of 19 heatthy adults (e:rprcssed as percentages of total IgG).

CHAPTER F'OURTEEN
DISCUSSION, CONCLUSIONS AND RECOMMENDATIONS

263I4.I DISCUSSION
IgG subclass quantitation
There is much unse¡taìnqr as to the clinical usefulness of IgG subclass quantitation. This has
arisen because of discrepancies between the results of different IgG subclass assays, poor assay sensitivity
and reliability, lack of agreement as to which antisera and reference sera should be used in assays, variable
definitions of normal ranges and differences in the criteria for patient selection in different studies. A
number of these problems have been previously discussed (Ch. 3,4; Ferrante and Beard, 1988). The need
for standardization of IgG subclass measu¡ement has been recognised and is currently being addressed by
the WHO/IUIS collaborative study.
For these reasons, studies and case reports of IgG subclass deficiency in infection-prone patients
are ¡lifficult to evaluate and very rtifficult to compare with one another. It was, therefore, a primary eim of
this sf¡dy to establish a reliable and sensitive assay for IgG subclass quantitation and to develop age-normal
percentile rânges for ou¡ local South Aust¡alian populatio4 in order to be able to define IgG subclass
abnormalities and to study their occlurence and possible sipificance in a wide ratrge of patients and patient
gloups.
Initiall¡ we studied differences between polyclonal antisera and McAb, and between RID and
ELISA for IgG subclass quantitation. We found that quantitation by RID with poþclonal antisera gave
marked va¡iacions between ÍNsay nrns. SimilarlS commercial RID plates with McAb gave higlhly variable
results. Quantitation of.lgp2 was often rlifficult bec¿use of.hazy precþitation rings, and quantitation of
IgG4 was often impossible, particularly in paediatric serum samples, because tle concentration was too low
for detection. Consequentl¡ an ELISA method was established in our laboratory. The accuracy of the
method was improved by the introduction of automation (Biomek 1ü)0, Beckman, Palo Alto, Calif.) to
increase accuracy in some of the tedious diluting steps where mistakes were occasionally made.
A range of McAb were tested in our ELISA and eventually a group which were found to perform
reliably and reproducibly were chosen.

264
Although poþclonal antisera a¡e still widely used it seems inevitable that these will be replaced by
McAb in the futu¡e. There has been concern about the restriction of some murine monoclonal IgG
subclass antisera and thei¡ failu¡e to detect all the antigenic variants of all IgG subclasses (Oxelius, 1987).
For this reason, good standardisation and rigorous testing of IgG subclass-specific McAb is essential.
Antibodies recommended for use in the type of assay being done should be used eg. for ELISA: IgGl
ÍIP60L2; IgG2 tIP6009, FIP6014; IgG3 HP6010, SJ33; IgG4 HP6011, SK¿14 (Jefferis et al, 1985).
Nevertheless, the details of the assay system used may profoundly influence the stability of one or other
antibody. McAbs which a¡e avid potent and specific for well-defined epitopes may partially or completely
lose their activity in some assay systems (Hussain et al, 1986).
IgG subclasses in healthy subjects of all ages were quantitated by ELISA and age-normal
percentfe rânges with lower linits that were measurable for each IgG subclass were derived. Recently,
Plebani et al, (1989) published simil¿¡ paediatric percentile rânges for IgG1,IgG2 and IgG3 derived using
McAb and RID, but were unable to determine lower cutofflimits for IgG4.
IgA and IgG subclass defrciency
IgG subclass deficiency in patients with IgA deficiency has been proposed as the e4pla"ation for
infection proneness in some such patients. This could be the difference between IgA-deficient healthy and
IgA-deficient infection-prone individuals. Ou¡ results confirm the work of some previous investigators in
showing an increased incidence of IgG subclass deficiency in IgA-deficient patients, but differ in the pattern
of IgG subclass deficiency observed. In ou¡ preliminary study, IgGl deficiency was the the most common
IgG subclass deficiency, but, as in many previous studies, the incidence of IgG4 deficiency could not be
determined because of the lack of sensitivity of the EIA used for quantitation. In our later study, we were
able to quantitate IgG4 by ELISA" and to compare results with normal ranges established locally using the
same technique in tle sa-e laboratory. rWe then found that IgG4 deficiency wÍrs the most common IgG
subclass deficiency in lgA-deficient subjects. Fu¡thermore, tlere was a correlation between infection
proneness and IgG4 deficiency in IgA-deficient patients.
In our initial sudy of children with increased proneness to respiratory infections, IgG subclass
deficiency was more common in children with lesser rather than with greater degrees of IgA deficiency, a

26sfinding which had not previously been noted. flowever an examination of data in a study by Ugazio et al
(1983) indicates tle same pattern of deficiency. In a group of 13 lgA-deficient children with recurrent
respiratory infections, IgG subclass deficiencies did not occu¡ in 4 of the 5 children with very low IgA levels
(<0.05g/l) but were very common in child¡en with lesser degrees of IgA deficiency (Ugazio et al, 1983).
Our fi¡st study excluded concomittant defects in neutrophil or complement activity as important
son6iþ¡ring causes of infection proneness in IgA-deficient children, and suggests that very low IgA levels,
per se, may be significant in this group of patients.
Children with a profound IgA deficiency are more likely to be infection prone than those with a
partial deficiency (Burgio et al 1-980). The situation may be different in adults where as many healthy IgA-
deficient.subjects have undetectable as have sþhtly low IgA concentrations (Ropars et aJ, 1982). A low
serum IgA i" healthy adults may be associated with increased serum IgG (Ropars et al, 1-982). This may
compensate for the IgA deficiency and help to e>rplain why these subjects a¡e not infection-prone. None of
our infection prone children with very low IgA concentrations had elevated IgG concentrations. This leads
us to postulate that the ability to compensate for low IgA concentrations by increased IgG production may
be age-related.
In ou¡ studies, the incidence of lgG2 deficiency in lgA-deficient patients was consistent with the
incidence of IgG2 deficiencies reported in other published studies (Table 5.2). However our findings differ
from other studies in that (a) we found a 27Vo nøLdence of IgGl- deficiency in our preliminary study which
is considerably higher tlan others have found" and also considerably highs¡ than we found in our
subsequent larger study. It is notewortly that five of the six IgGl-deficient children in the fi¡st study had
IgGl concentrations only marginally below the age-related normal tl¡ge, and that the lower li6il 6f this
normal r¡nge (Oxelius et al L979a) yy¿s highs¡ than that of ou¡ own and other published normal ranges at
most ages (Fig. a.Ð. (b) *" found a ?ÁVo ncrdence of IgG4 deficiency in our second and major study. This
was the most common IgG subclass deficiency found in this study. Like most other investigators, in our
preliminary stud¡ we could not determine the incidence of IgG4 deficiency because of assay limi¡¿¡ie¡5.
However, in that sfidy, 4wo of patients had undetectable IgG4 concentrations and many of them may have
been IgGzl-deficient.

266The une>rpectedly high incidence of IgG4 deficiency was tle most striking fiodi"g in our major
study. This was clearly evident whether either the 5th percentile for age or an arbitrary concentration of
less than O.O3g/litre was taken as the cutoff point. If we define IgG4 deñciency accorcting to plebani et al
(1986), as <0.006g/litre, only lÙVo of our patients would be considered IgGrl-deficient. However, IgG4
concentrations below O.ÙLg/húe were not found in any of our normal subjects who were 3 years of age or
older, and only two of 26L had concentrations below O.O3g/litre. In all age goups the 5th percentile was
well above 0.006g/litre. This leads us to conclude that Plebani et al's cutoff point is not appropriate for ou¡
assay for most age groups.
Since subnormal concent¡ations of IgG4 occu¡red in 17 of 50 IgA-deficient patients with recurrent
infectionl and in only two of 23 other IgA-deficient patients, we consider a low IgG4 concentration to be
an important sþ of impaired immunologic¿l defence. In some patients with IgA deficiency production of
antibodies in other isotlpes is impaired (French & Harrison, 1984b; Lane & Macl-ennan" 1-986; Hanson et
al, 1988). The question of whether an IgG4 deficiency in an lgA-deficient patient is an indication of a
deficiency affecting antibody production in the IgG4 isotype onl¡ in other isotypes only or in both isotlpes
need to be addressed. Our fiodiogs provide ¿ st¿¡ting point for further iavestþating the links between IgA
deficiency, IgG subclass deficiency and infection proneness.
The results from ou¡ investigations support some of the ideas generated by Heiner et at (1936)
who found that low concentrations of IgG4 were more oommon in patients with recurrent infections than in
normal or atopic subjects. They found that ?ßVo percent of.362 patients witl recurrent respiratory tract
infections had IgG4 values below 0.03 g/litre compared vttrh 5.6Vo of healthy subjects. We found that
percentages of symptomatic lgA-deficient subjects with IgG4 concentrations below O.03g/litre were
signifigantly higher than those of cont¡ol subjects at all ages. In the total patient Foup, 34.2Vo of patients
had IgGa concentrations of <O.O3g/litre compared ¡ørth6.Z7Vo of controls (p<.001). Ifence, like Heiner et
al we found that a disproportionate nunber of q'mptomatic patients had low concentrations of IgG4.
Their patients differed from ou¡s in that tley were investþated primarily for IgG4 deficency and only 2IVo
of them were found to be IgA-deficient. The similarity in the frequency of low IgG4 concentrations,
however, may be of relevance. In another stud¡ Heiner (1936) found IgG4 deficiency (usi"g RIA) in
greater tJ.an 50Vo of IgA-deficient patients together with IgG2 deficiency n70Vo. The incidence of IgG4
deficiency in this study is highe¡ than that in ours, and that of [gp2 deficiency is considerably higher. The

267latte¡ is difficult to interpret, because normal rânges for IgG2 were not derived in Heiner,s own laboratory.
The ages of the patients were not stated. There were several combinations of IgG subclass deficiency in
patients in our second study. IgG2-IgG4 occurred, n 4% of patients and was tle most common deficiency
after isolated IgG4 deficiency. lgGL-IgG2-IgG4 and IgGz-lgS3 deficienies each occurred, n 3Vo of.
patients.
Oxelius et al (1981) found a relationship between lgG2 and,IgA deficiency in respiratory infection-
prone children and adults but were unable to show whether or not an IgG4 deficiency occurred
concurrently in these patients because of the limitations in assay sensitivity. Similarly, Qrnningh¿p-
Rundles et al (1983), Bjorkander et al (198Ð and Klemola et al (1983) were unable to determine the
incidence of IgG4 deficiency occurring in conjunction with other IgG subclass deficieucies.
Ugano and Plebani et al (1-983), in an initial stud¡ using polyclonal antibodies, RID and normal
rânges f¡om another laboratory found a significa¡rt incidence of Igp} and IgG4 deficiency in IgA-deficient
child¡en with increased susceptibility to infection. Flowever, in a subsequent larger study of lgA-deficient
child¡en using monoclonal antisera and their own age-nonnal percentiles, Plebeni et al (1986) found that,
although IgG2 deficiency w¿ts the most frequent concurrent deficiency, it did not occu¡ in a significant
number of patients and was not associated with IgG4 deficiency. The different fi¡rtings in these two
studies, like those in our prelimin¿¡y and subsequent stud¡ highlight the importance of methodolog5r and
normal ranges in IgG subclass studies. Unless normal ranges obtained in the seme laboratory with the
same metlods and reagents, and assays sensitive enough to detect IgG4 have been used, reports on tle
relative incidence of IgG subclass deficiencies in IgA-deficient patients a¡e of questionable validity. Our
studies, and the second study by Plebani's group, found similar incidences of.Igp2 and IgG3 deficiencies in
IgA-deficient children, but differed with regard to the reported incidence of IgG4 deficiency, largely
because of our differing definitions of IgG4 deficiency, as discussed previously.
Klemola et al (L988) found a 5imil¿¡ incidence of.IgG} deficiency in lgA-deficient children and
adults to that which Plebani et al found QVo,\Vo, respectiveþ). IgG4 deficiency could not be defined in
Klemola's study as the normal ranges used were those of Oxelius (L979a) in which the lower limit of the
normal range was below the sensitivity of the assay. In fact, in lgA-deficient subjects, Klemola et al (19SS)
found a lower incidence of 'nmeasurable IgG4 concentrations tlan did Oxelius (1979a) in healthy controls.

268Chandra (1986) found IgG2 deficiency to be the most common IgG subclass deficiency in a group IgA-
deficient patients, but used RID and did not speciS how normal ranges were determined so that the
reported incidence of IgG4 deficiency in this study is questionable. The normal IgG subclass range
established by Oxelius (L979a) has been used in at least five of these studies in IgA-defrcient subjects.
IgG4 deficiency and infection proneness
Although IgG subclass deficiency is thought to be a factor in infection-proneness in some patients,
little is known of the role of IgG subclass deficiency in increasing susceptibility to different t1ryes of
infections. Our studies have indicated that g¡oups of patients with certain types of infections are more
Iikely to thow
IgG subclass deficiencies than are those with other types of infections. We found that IgA-
deficient patients with proneness to respiratory infections, patients with invasive Hib infections and patients
with bronchiectasis had a significant incidence of IgG subclass deficiencies, while patients with osteomyelitis
and septic arthritis and giardiasis and splenectomized patients did not. In each of the groups of patients
where a sìgnificant incidence of IgG subclass deficiency was found, IgG4 deficiency was the most oommon
IgG subclass isotlpe deficiency. It is of note tlat each of the groups of patients where IgG subclass
deficiencies occurred most often were groups of patients with infections either of the respiratory tract itself
or with organisms gaining entry to the body via the respiratory tract. In those g¡oups of patients with
infections where IgG subclass defrciences were not prominent, the respiratory tract was not generally
involved. The splenectomized patients -" ¿ unique group in that although they are at risk of respiratory
and systemic infection from ^S.
pneumoniae they are not abnormally prone to otler respiratory tract
infections (Heier, 1980).
Our finding sf ¿ significant incidence of IgG4 deficiencies in patients with bronchiectasis
supplements that of several other studies. Stanley et al (1984), using RID, found undetectable, though ¡s¡
unequivocally abnormal IgG4 concentrations in 5 of 8 patients with bronchiectasis and in only one of these
5 was there a deficiency of another immunoglobulin isst'¡ps alss. Murphy et al (19S4) found 4 of 23
patients with bronchiectasis, had IgG4 levels below those of controls. A few studies using more sensitive
assay methods have associated IgG4 deficiency with severe respiratory t¡act infections, including
bronchiectasis (Heiner et al 1983). In 1981, Beck and Heiner using a RIA technique, polyclonal
antibodies, and a normal range established in the sa-e laboratory found isolated IgG4 deficiency in 4 of

269the 29 patients with recurrent pulmonary infections, but in none of 154 healthy subjects, and in none of 239
patients with suspected allergic disease. Three of 7 patients with idiopathic bronchiectasis were IgG4-
deficient. Extending these studies, 5 of 35 simila¡'patients were found to be IgGzl-deficient (Heiner, 19g4).
Qu¡ findingsr using an ELISA technique, monoclonal antibodies and age-normal range established in ou¡
laboratory provide further evidence for the possible importance of IgG4 deficiency in respiratory tract
infections, including bronchiectasis.
The reported incidence of IgG subclass deficiencies in patients with recurrent infections varies
considerably. ff IgG subclass deficiency is an important factor predisposing to respiratory infections, a
correlation between the severity of the respiratory infections and the degree and incidence of IgG subclass
deficiency mighl þe e4pected, such that, in a heterologor$ group of patients with varying degrees of
respiratory disease, less IgG subclass deficiency would be found than in a group of patients with well-
defined severe pulmonary infective disease. Reported incidences of IgG subclass deficiencies i" IgA-
deficient patients witl recurrent infections vary greatly, ranging from about L7 to 77Vo (Ugazio et a! 19g3,
Oxelius et al, 1981, Plebani et al 198ó).
Reported incidences in other groups of patients with respiratory infections also show great
differences ranging from about 5 to 60Vo (Oxelius et al, 1986; Smith et al, 1984). Some of tlese di_fferences
must be related to va¡iations in the criteria used to define both the patients and the IgG subclass
deficiencies. often, limitations of assay sensitivity have hampered IgG4 measurements and comments
about the incidence of IgG4 deficiency have been impossible. In two recent studies, using RIA and a solid
phase conpetitive inhibition ¿rssay, respectively, ¿ significan¡ number of IgG4 deficiencies have been found.
Heiner et al (1988) fotnd' ?ÁVo of 400 patients with recurrent respiratory infections to be lgGzl-deficient
when results were compared with normal rânges determined in the same laboratory. IgG2 deñciencies
were even more oommon' occurring in38% of the patients but the normal rânges used for IgG2 were not
determined in the same laboratory. Shackelford et al (1986) using their own normal ranges, found Z of 30
(23Vo) children with recurrent infections had IgG2 subclass deficiencies and three of them had probable
IgG4 deficiencies although the lower limits of normal at young ages were difficult to define. The reasons
for the apparently lower incidence of IgG2 deficiencies in ou¡ studies of respiratory infection-prone patients
are not entirely clear, particularly as the lower limi¡s qf normal for IgG2 in the study of Shackelford et al

270(1986) tend to be lower ttran ours. The differences may be apparent rather than real because of the
relatively small numbers of patients studied.
While IgG4 deficiency is obviously not tle major predisposing factor in otherwise une4plained
bronchiectasis and other respiratory tract infections, ou¡ findirgs, and those of a number of other
investþators (Beck and Heiner, 1981-; Heiner et a! 1983; Stanley et a! 1-9g4; Heiner, 19g4) suggest that this
deficiency may occur in a considerable number of such cases.
Possible mechanisms of infection-proneness in lgGlt defrciency
Although, quantitatively, IgG4 is ¿ mins¡ IgG subclass in the serum comprising onty 2-6Vo of. the
total circulating IgG, and although it does not activate the classical component of the complement casc¿de
(Bird and l¿chmann, 1988) or bind strongly to the Fc receptors of monocytes or macrophages (Stanwort\
1983)' it may play ¿ 'niquo, but as yet ill-defined role in host defence. It is possible that a serum IgG4
deficiency may be a sb of a broader defect. This could involve the production of ,specific, antibody of
other isotypes also or deficiencies in other a¡ms of ffie imm¡¡s system. [n our studies we did not find
evidence of abnormalities of lymphocyte or neutrophil function or complement deficiencies in association
with IgG4 deficiency' It therefore seems more likely that IgG4 deficiency is a sþ of antibody deficiency. Itis possible a serum IgG4 deficiency may reflect a mucosal IgG4 antibody deficiency and that IgGa, üke Ig¡\may be primarily a secretory immunoglobulin having an important role in mucosal defence.
A deficiency of one IgG subclass can be associated with deficiencies of 'specific, antibody
production both of this is6$ps and of otlers, even though the total concentrations of the other isotypes
may be within the normal range. Schoettler et al (L986) and Heiner et al (19gS) describe a patient in whom
a deficiency of IgG4 (detected by RIA) was associated with a deficiency of lgp2 antibodies to S. øtreus
even in the presence of normal totat IgG2 levels. The patient developed life-threatening systomic
staphylococcal disease, improving only after immunoglobulin replacement was added to the antibiotic
regime' An infant, aged 4 weeks, who suffered overwhelming meningococcal sepsis requiring amputation
of several limbs because of tissue nectosis, and who like his mother had an isolated IgG4 deficiency has also
been described (Heiner et at 19s8). The possible significance of the IgG4 deficiency is however, difficult to
assess in the absence of information about other aspects of imrnune function in this patient.

27L
Recently, investigators have become interested in the IgG subclass distribution of protective
antibodies to 'specifiC antþens, e4pecting to find that a deñciency of one isotype would result in proneness
to infection by organisms which normally evoke antibodies of that isotype. Flowever, preliminary studies
have shown that the situation is much more compler This is well-illustrated by studies on antibodies to fL
influenme. Althoueh antibodies to Hib CPS, which are thought to be essential for protection, are
predominantly found in the IgGl- arrdlgG2 isotlryes (Weinberg et al, 1986; Hamma¡strom et a! 1988; Scott
et al L988; Shackelford and Granoff, 1988) susceptibility to invasive Hib disease does not appear to
correlate consistently with total IgGl- or IgG2 concentrations (Ch. 1.6). Our studies suggested" rather, tlat
there may be a correlation with IgG4 concentrations as alnost all the patients in this study had low or low
normal IgG4 concentrations.
Some investigators have shown that patients with low concentrations of IgG2 have low
concentrations of antibodies to Hib CPS, either in 1þs ¡snimmunized state or ¿fte¡ immrtnization (Oxelius,
1974; Siber et al 1980; Umetsu et al, 1985). This suggests that IgG2 subclass deficiency may be a sþ of an
impaired antibody responsiveness to Hib CPS. Other investþators, however, have failed to show a
correlation between IgG2 concent¡ations and anti Hib CPS concent¡ations (Scott et al, 1988). Still others
have found no deficiency of Hib CPS antibodies in the majority of IgG2-deficient patients (Shackelford et
al, 1986). In tlose studies showing a correlation between IgG2 and Hib CPS antibodies where IgG4 was
measured elss, fhis, as well as [gp2, was low. Heiner et al (L988) found a correlation between IgGl
deficiency and impaired production of antibodies to Hib CPS. Granoff et al (1983) studied a group of
children who had been immunized with Hib CPS and yet subsequently developed invasive H. influenzac
disease. There was no evidence of.Igp2 deficiency in most of these children. This was in agreement with a
previous study (Granoff et a[ 1986;b) where Hib vaccine failu¡e child¡en were found to have normal senur
concent¡ations of IgG2 and other immunoglobr lins.
Recently, several investþators have sought to clari$ the divergent data by measuring the
concentration of IgGl andlgp2 antibodies to Hib CPS. IgGl- rather than IgG2 has been tle predominant
isotype fgund in both children (shackelford and Granoff, 1988) and adults (Ha--a¡strom et a! 19gg; Scott
et al 1988). In these studies, IgG3 or IgG4 antibodies to Hib CPS have not been measured. Shackelford
and Granoff (L988) also showed that some IgG2-deficient patients had impaired production of both IgGl

272and IgG2 antibodies to Hib CPS after imrnlrnis¿fign, but that amonøst IgG2-deficient patients some were
normally responsive. Since, therefore, a deficiency of one subclass (IgG2) can be associated with a
restricted deficiency of antibody production which may also affect another subclass (IgGl), it is conceivable
that an IgG4 deficiency, similarly could reflect an antibody deficiency affecting isotypes other than IgG4.
Concent¡ations of IgG4 may be a more sensitive indicator of impaired antibody production to a restricted
group of antigens than a¡e concentrations of IgG2. This may help to explain why various studies have given
çpnÊliçfing results concerning IgG2 concent¡ations and Hib infections.
On the other hand a reoent study by Hetherington (1989) indicates, opsonic antibodies to the
outer membrane protein of Hib are more important than anti-Hib CPS antibodies in defence ¿gainst
invasive Hib disease. This also could help to erçlain the lack of correlation of IgGl and IgG2
concent¡ations with this condition. The isotypes of these antibodies a¡e not known, although anti-protein
antibodies are often predominantly of the IgGl and IgG3 and somerimes also IgG4 isotypes (Schur, 1987).
Antibodies of the IgG4 isotype, per se, may be important, or alternatively, low IgG4 concentrations may
reflect impaired ability to produce anti-protein antibodies of other isotypes.
The significance of lower T cell proportions in ou¡ patients with Hib infection compared with
normal subjects is difficult to evaluate, especially since ou¡ results showed that the lower proportions were
seen evenly amsngst the patients irrespective of the presence of a subclass deficiency. Nevertheless, it is
interesting to note that most patients in the group had IgG4 levels well below mean for age, perhaps
indicating a correlation between T cell function and IgG4 production.
Patient JB, described in Ch. 72.3.2, illustrates how profound but restricted antibody deficiencies
may occur in the presence of IgG subdass deficiencies. This patient with slþhtly low Igd IgG and IgM and
with deficiencies of IgG1-, lñ2 and IgG4, showed a total lack of respousiveness to S. typhi and to
pne 'mococcal polysaccharide antþens, despite normal responsiveness to the protein antlgeq tetanus
toxoid. She also illust¡ates how the pattern of imm¡¡sg1sþrlin isotlpe deficiencies may change markedly
over a period of years. Roberton et al (1990) have sholvn that IgA deficiencies may not be persistenÇ
while concurrent IgG subclass deficiencies may persist. There is a great need for serial studies in
immunoglobrrlin isot¡re-deficient patients to determine the permanence of the deficiencies.

273The possibility that IgG4 may be primarily a secretory imm¡¡sglgþulin warrants serious
consideration, end ou¡ findi"g that groups of patients with infections of or gaining entry through the
respiratory tract had a sipnificant incidence of IgG4 deficiency, may be relevant in this regard. Although we
did not detect sipificant differences in the percentage of IgG4 in saliva and in serrn in a group of patients
the fin.li.g that in 76Vo the percentage yy¿5 highsr in saliva is of interest. The relative distribution of IgG
subclasses in mucosal secretions f¡om different sites may vary.
Only a few investigators have measu¡ed IgG subclass concent¡ations in mucosal secretious.
Sotomayer et al (l-989) have shown that while IgA is tle predomi"¿nf imm¡¡¡qglobulin in the secretions of
the upper respiratory tract, IgG predominates in bronchoalveola¡ lavage fluid. Merrill et al (1985) found
high concent¡ations of IgG4 in the lower respiratory tract secretions obtained from bronchial lavage fluid in
29 healthy adults measudng IgG subclasses by ELISA and found that IgGl and IgG2 were present in
relatively proportionate âmounts in senm and lavage fluid. Relative levels of IgG4 were highs¡ i¡ ¡þe
lavage fluid than in tle seru'n. The data in this sludy suggests that IgGl and IgG2 in bronchial secretions
depend largely on transudation from serum, whereas IgG4 and IgG3, in some subjects, may possibly be
either locally produced or may have a tendency to increased local accumulation eg. by attaching to
bronchial or intramural cells. Soderst¡om et al (1983) found no correlation between serum, salivary and
nasal IgG subclass-specific responsss [s immrnisation with Hib and tetanus toxoid, suggesring tlat mucosal
IgG subclass concentrations are strongly affected by local factors. They found a correlation betwsen senm
IgG2 concentrations and numbers of mucosal IgG2-producing lymphocytes, but not between those of IgGl
and IgG3.
Keller et al (1983) found the relative percentage of IgG4 in the colostlm of postpaftum women
was about fou¡ fimes that in tle serum, and subsequently (Keller et al 1938) produced data to support the
hypothesis that local mâmrnary gland production of IgG4 occurs in at least some subjects. Grill et al (1985)
suggested that the mucosal production of IgG4 may be important in defence agains¡ giardiasis by
demonstrating that patients with giardiasis had lower concentrations of IgG4 in thei¡ duodenal secretions
than did patients with other diarrhoeal dise¿ses. Serum IgG4 deficiency occurred in only one of the four
patients. The study was small and was not fully reported.

274If IgG4 is a secretory immr¡neglsþnlin, its interaction with bacteria and viruses, with alveolar cells,
macrophages and mast cells (Beck & Heiner, 1-981) or with microbial cytopathogenic products may play a
functional role in host protection at the mucosal level. While serum IgG4 concentrations may not
necessarily reflect mucosal IgG4 concentrations, it is not unlikely that, if patients who have low senrm IgG4
concentrations have impaired IgG4 production systemicall¡ they may also have impaired IgG4 production
locally. The situation may be simila¡ to that for IgA where low concentrations of both serum and secretory
forms are related (Ch.I.2.I.2).
IgG4 and lgpZ arc the predominant surface immr¡nsglqþulins on adult B lymphocytes (Yount et
al, 1980). Percentages of su¡face IgG subclasses in normal IgG-positive lymphocytes reported by Simmons
et al (1981) were IgG2, 36Vo,IgG4,29Vo,IgG1,24Vo andIgp3,LÙVo. Surface immr¡nsgleþillin is þsum 16
function as an antigen receptor, and it may be that IgGzl-deficient patients are likely to be infection prone
because of deficient antigen-recognition and present¿¡ie1 psçþanisms (Rock et ù, 1984; Abbas, L988;
Metlay et al 1-989). There are many unanswered questions in fhis ¿¡s¿.
Tleatment of IgG subclass deficiencies
The optimal management of IgG subclass-deficient patients who are unusually susceptible to
infections is of concern to clinicians. Some recommend the use of prophylactic antibiotics and others, the
administration of immunoglobulin on a regular basis.
Immunoglobnlin replacement therapy appears to benefit some patients who have IgG subclass
deficiencies. However as some patients with subnormal trough concentrations of IgG subclasses remain
well and other patients who have borderline or low normal concent¡ations of an IgG subclass aro
abnormally infection prone and heþed by immunoglobulin therapy and as infection proneness in individual
patients does not always appear to parallel their IgG subclass concent¡ations, it is evident that IgG subclass
concentrations, while a helpful guide, are not an absolute determinant of the need for immunoglobulin
replacement therapy in infection-prone patients. I¡w IgG subclass concentrations may indicate impaired
ability to produce certain'specifiC antibodies and antibody replacement is considered to be the crucial
factor in treatment, rather than merely maintaining concentrations of IgG subclasses at particular levels.

Infection-prone patients without IgG subclass deficiencies275
In the groups of patients with osteomyelitis and septic arthritis and giardiasis and insplenectomized subjects we did not find a significant incidence of an IgG subclass deficiency, but there were
some individual patients with deficiencies. Four of the 15 patients who had had osteomyelitis or septic
artbritis had a deficiency of at least one IgG subclass, possibly indicaring impaired antibody production in
these individuals' TWo of these patients also had a history of excessive infection proneness, making such a
possibility more likely' Eid et al (19s0) gnggest that there may be an impaired antibody production in
patients with osteomyelitis because they found that most fail to develop the expected imm¡¡6g16þrlin ¡iss
in response to infection. The fact that more of our patients had IgG and rgA concentrations below than
above the mean for age may reflect, simil¿¡l¡ subnormal antibody respo¡uies to some infections. Ilowever,
the 'aiseà
concentrations of at least one IgG subclass in 6 of our patients may reflect appropriate antibody
responses to recent infection in these particular patients. It is possible that we have underestimated the
incidence of immunsglobulin isotlpe deficiencies by blood sampling at a 'Ðe when an antibody response to
infection may be gvi"S falsely hieh values although it is unlikel¡ that patients who are deficient would
produce a good antibody respome in the deficient isotype.
f¡[srrgh patients with hlpogtmmaglobtlin¿epia are particularly prone to giardiasis, most
patients with giardiasis are not hypogamnaglobrrlinaemic and the reasons for their susceptibility to this
infection are unknown. ou¡ study showed that IgG subclass deficiency is not likely to be a reason. wefound une4pectedly high lr¡¡þe¡s of patients had raised concentrations of senm IgE, IgGl and IgG3. The
hrgh Ign bvels may indicate either a degree of immune dysregulation with a component of hlpersensitivity
as a possible mechanism contributing to the development of symptoms or a role for IgE in immunity to
Giordia lamblia' The high concentrations of IgGL may reflect a vþorous response to antþenic srim'l¿tisq
possibly to a protein antþn of. Gio¡diø lamblia, since it is knoqm that antibodies to protein antþens are
predominantly of this isotype (ch. 1.6.1.1). ou¡ tests of cellular immune function did not reveal any
ftmctional defects in the patient group. The percentage of B cells was sþhtly highe¡ than in controls. This,
like the raised concentrations of certain immunsglsþrlin issçss, may have reflected a respon*e to ch¡onic
antþenic stimulation.

276
The raised concentrations of serum C4 in 9 of our patients with giardiasis may have represented an
imbalance between production and cons ¡mption and been secondary to the infection. The facts that ? of 9
patients with raised C4 concentrations had raised IgE concentrations and 5 of these also had raised IgGl
concentrations are of interest, althot'gh of uncertain significance. Elevated serum complement has,
however, been found in some other disorders with gastroenterological involvement such as ulcerative colitis
and typhoid fever. The signific¿nce qf this findi"g is unclear (Stites, 1982).
Our results indicate that serum immunsglsþulin isot¡rps deficiency is not likely to be a common
factor in predisposing otherwise healthy children to symptomatic giardiasis. Kumkum et al (1988) found
low concentrations of serum IgA and IgG i" a gfoup of patients with ch¡onic giardiasis. Patient selection
criteria diff, ered from those in ou¡ study and so results a¡e not strictly comparable. The possibility of a
localised mucosal deficiency of an IgG subclass remains to be investigated.
The inlluence of the spleen on IgG subclass concentrations
Althot'gh splenectomized subjects are at increased risk of S. pneumoniac tnfectrons, in significant
numbers of these patients, we found elevated rather than reduced concent¡ations of IgG subclasses,
specificall¡ IgGl and IgG2. Defence against S. pneumon¡'a¿ involves opsonisation and phagocytosis in
which the spleen has a role in protection by producing antibodies and clearing the bacteria. Removal of the
spleen, therefore decreases the body's capacity to deal witl some organisms. For opsonisation of S.
pneumoníae, both IgM and,lgp2 antibodies are important in adults, and IgM, IgGl and lgG}n children
(Freijd et al 1984b; Sarvis et al, 1989). Low IgM concentrations have been reported post-splenectom¡ but
IgG subclass concent¡ations have not previously been measured.
While significant nunbers of splenectomizßd patients had elevated IgGL and IgG2 concentrations
this pattern was not apparent in the patients with portal hypertension where IgG4 concent¡ations were
urore commonly raised. It is of note that a tendency to increased concentrations of isotypes produced by
downstream switching of the imm¡¡eglsþulin gene code was apparent, the gene sequence for
immunoglob,,lins being IgM-IgG3-IgGL-leA.1-lñ2-IgÊ+ldE-IgA2 (Ch. 1.3.2).

277Relatively low concentrations of IgM, normal concentrations of IgG and IgG3, and raised
concentrations of IgGl, IgG2, IgG4 IgE and IgA occurred in more of the patients than e4pected.
Splenocytes produce predominantly IgGl and IgG3 in response to srimulation with mitogens (Scott &
fr[ahm, 1984). IgG1, is the predominant IgG subclass e4pressed by foetal splenic tissue (Partridge et al,
1984). Since IgGl concentrations in splenectomized patients were high rather tlan low, and IgG3
concentrations were high i" splenectomized adults ou¡ findi"gs, indicate that removal of the spleen does
not cause an imm¡¡sglsþulin subclass imbalance directly by removing B cells that produce certain IgG
subclasses.
T lymphocyte function is decreased in splenectomized patients and in patients with portal
hlpertension (Ferrante et al, 1989). In man, I$vI,IgG2 and IgG4 are considered to be largely T cell-
independent isotlpes. The IgGl respolxie is more sensitive to T cell regulation than the IgG2 response
(Morell et al, 1-981; Mayumi et a[ 1983). T cell deficiency, therefore would be ulikely to account for low
IgM concentrations, or to e¡plain the raised concentrations of IgGL, IgA and IgE which are considered to
be T cell-dependent isotypes. NK cells may have an important role in regulating immune respoilies
mediated by the secretion of cytokines such as interleukin 1, interleukin-2 atd B cell growth factor (Kimata
et al,,1987; Rodriguez et aJ, L987). Supernatant from NK cell cultu¡es enhances the production of IgE, IgA
and IgG (Kimata et al 1988). NK cell activity in mice, has been shonm to down-regulate tle production of
antibodies to pneumococcal polysaccharides in a T cell-dependent fashion (Khater et al, 1986). Studies in
ou¡ laboratory have shown that NK cell activity is increased markedly in splenectomized patients and
slightly in patients with portal hlpertension (Ferrante et al, 1985; Ferrante et al L989). This increased NK
cell activity may be a factor in the impaired antibody responses to pneumococcal polysaccha¡ide in
splenectomized subjects. It may also contribute to the abnormalities in the patterns of Ig isogpes found in
our splenectomized subjects.
The low IgM concentrations found in some splenectomized patients may reflect low concent¡ations
of IgM anti-pnerrmococcal antibodies and contribute to the proclivity of splenectomized subjects to
pneumococcal infections. We postulated that IgG subclass deficiencies, particularly of IgGL and IgG2, may
prove to be another contributitg factor. Surprisingly, we found that concentrations of these isotlpes were
not low in any of the patients, but were elevated in significant numbers. Without the measurement of the
IgG subclass of the anti-pneumococcal antibodies present in these patients we cannot exclude a deficiency

278
of isotype-specific IgG subclass anti-pneunococcal antibodies, since low concent¡ations of IgG2 anti-
pneurnococcal antibodies have been found in some patients with normal IgG2 concent¡ations (Lane &
Maclennan 1986). rWhile such a restricted dehciency is probably less likely in the face of raised
concentrations of these subclasses, the finding that there is a reduced IgG response to pneumococcal
immunization in splenectomized subjects (Hosea et al 1981) suggests that such a deficiency may indeed
exist.
IgG subclass concentrations in immunological disorders
In our four patients with hy¡roga--aglobulinaemia we found tlat concentrations of all the IgG
subclasses were very low. In three patients who had had significant chest and sinus infections we found
reduced concentrations of IgG1, IgG2 and IgG4. In two patients with a history of asthma and bronchitis we
found reduced concentrations of IgGl and IgG3.
Family studies in Cwo families, one with fwo hlpogammaglobulinaemic childreq and the other with
one, showed reduced concentrations of IgG3 in the father and of IgG4 in the mother in the former, and of
IgG4 in the mother in the latter. Isotype deficiencies in relatives of hypoga--aglobulinaemic patients have
been described previously (Van der Geissen et al, 1976;b) and may indicate heterozygosity for the condition.
In the patient witl severe combined immunodeficiency and in the one with ADA deficiency all the
IgG subclass concentrations except that of IgG4 were low at fiegnosis. This is in agreement with the
fi¡dings of Oxelius (1979b). We did not find undetectable concentrations of IgG2, IgG3 and IgG4 as
Morgan et al (L936) (Ch. 1.8) had probably because our ELISA was more sensitive than their RID assay.
Ou¡ one patient with the Wiskott Aldrich slmdrome was not deficient io aoy of the IgG subclasses
before or after a successful bone marrow transplantation. Because he was under one year of age, we did
not attempt to measure his responsiveness to pneumococcal immunisation as normal responsiveness is poor
¿t this ¿gs. Nahm et al (1936) did not find a significant incidence of IgG subclass deficiency in 14 patients
with Wiskott Aldrich syndrome, and failed to show a correlation between concentrations of IgG2 and
antibodies to three carhobydrato antigens. As levels of these antibodies were low in the presence of normal
IgG2 concentrations, the inability to respond to carbohydrate antþens was not contingent on IgG2

279deficiency. Aucoutu¡ier (1989) studied 6 patients with the Wiskott-Aldrich slmdrome and found that the
only IgG subclass deficiency was that of IgG3 in one patient (Ch. 1.8). Other studies of small numbers of
patients have suggested that IgG subclass deficiency may not be uncommon in Wiskott-Aldrich slm.drome
(Oxelius I979b; Aucouturier et al, 1986a; rWedgwood et al 1986; Bremard-Ouryet al 1986). In most of
these, however, the inability to delineate an IgG4 deficiency has been a problem as rhis has been the main
deficiency suspected. It seems likely that the impaired ability to produce antibodies to carbohydrate
antigens which is cha¡acteristic of the Wiskott-Aldrich syndrome generally occurs in the absence of an IgG
subclass deficiency, highlighfing, once again the complexity and restricted natu¡e of some such deficiencies.
Of ou¡ three C2-deficient patients, one had an IgG subclass deficiency involvinl IgG1. Several
patients have been described previously with combinations of complement and IgG subclass deficiencies
(Ch.1.8, Wedgwood et al, 1-986; Oxelius et a[ 1986). A link between complement concentrations and IgG
subclass concentrations is suggested by the conclurence of raised C4 and raised IgGl in a number of ou¡
patients with giardiasis. Bird and Lachmann (1983) describe a correlation between deficiencies of the early
components of the classical pathway (C1 to C4) and C3 and IgG4 deficiency, and propose that IgG4 may be
the product of a complement-dependent B cell which could have a negative feedback effect inhibiting
antigen localization on follicula¡ dendritic cells.
The effect of bone marrow transplantation on IgG subclass concent¡ations appears to vary from
patient to patient. In ou¡ one child who underwent a successful bone marrow transplantation from an older
siþling, IgG subclass concentrations showed no deficiencies throughout the following year, and at 10
montls post-transplantation, IgGl and IgG2 concentrations were elevated. In the child with severe
combined immunodeficiency, all IgG subclass concentrations were low after a first ,nsuccessful
transplantation anq after a secoud gs¡tinning immunsglsþulin i¡fusions have been necessary because of
low concentrations of all the IgG isot¡pes. Neither of these patients fit the patterns of IgG subclasses
previously described after bone marrow transplantation. Morell et al (Lg7Ð have described a patient in
whom IgGl- and IgG3 returned to normal $,ithin the first year after bone marrow transplantation and IgG2
and IgG4 took longer to do so. Aucouturier et al (1987b) in a group of 31 patients who had received bone
marrow transplantation also found thatlgG? and IgG4 concentratioru¡ were often low many montls later,
while IgGl- and IgG3 concentrations were sometimes elevated.

280In the one patient with diGeorge syndrome who had received a thymic epithelial6ansplant about 9
years prior to her IgG subclass quantitation, serum IgA and IgG4 concentrations were reduced. Although
she was not apparently infection-prone, this may indicate a degree of continuing immune dysfunction,
specificall¡ perhaps, a persisting abnormality of T cell function a,ffecting downst¡eam immunoglobllin
isotype switching.
Patients with a variety qf ¿utsimmune connective tissue disorders frequently showed high
concentrations of IgGl and sometimes of IgG3, and several with SLE showed IgG4 deficiency as well. This
is in accord with some previous studies in patients with SLE (Oxelius, 1984; Schur 1987). We did not find
low IgG2 concent¡ations to the e>úent that Oxelius did (1984). This may be because we used a different
assay system and defined ou¡ normal ranges differently. IgGl- and IgG3 are the most strongly complement-
binding IgG subclass isotypes (Cb.1'.2.2.5) and have been previously found to be the only IgG subclasses
detectable in immune complex-mediated cutaneous vasculitis (Shakib and Stanwort\ 1-980b). By renal
immunofluro5seaf tssring, with polyclonal antisera" Bannister et al (1983) found IgG3 the most
predominant IgG subclass found ia glomerular deposits io lupus nephritis, and IgGl tle second most
predominant.
Antinuclear antibodies appear to be predomi"antly IgGl and IgG3 although not all studies agree
about the contribution of IgG2 and IgG4 (Shakib and Stanworth, 1980b; Schur, 198Ð. Afruoth et al (1989)
found a high ¡1san serum IgGl in patients with þus glomerulonephritis according to most normal ranges,
but commented only on tle high 6san IgG4 value. The apparent increases in IgGl and IgG3 in many
patients \l/ith SLE and other autoimmune disorde¡s may indicate that IgG antibodies to autoanttgenic
protein antþens are generally of the IgGL and IgG3 isotypes as are antibodies to other proteins (Ch.
1.6.1.1). Yount et al (1988) studied the IgG subclass dist¡ibution of a variety of human autoantibodies and
found that for anti-Sm and anti-RNP, IgGl predominated. Anti-ds-DNA and anti-SS-B included some
IgG3 although IgGl predominated. Anti-RF included simil¿¡' âmounts of IgGl and IgGa. Kay et al (1988)
report a very high incidence of connective tissue disease in subjects with disproportionately high tgÇt
concent¡ations.
Our patients with Henoch Schonlein prupurq like two with Henoch Schonlein nephritis reported
byAlmroth et al (1939) did not show an IgG subclass imbalance.

28rStudies of relationships between the HLA system and immunoglobulin allotypes and isoty¡les may
help to elucidate the reasons why certain isotype patterns a¡e evident in some ¿¡fgimmuns disorders (Ch.
t.3.4).
Previous studies in common va¡iable immunodeficiency have suggested that IgG1, IgG2 and IgG4
concent¡atiorìri are more likely to be low than are those of IgG3 (Ch. 1.8). In most of tlese studies the
separation of low and low-normal IgG4 values have not been possible because of limi¡¿¡¡s* ;11 1¡"
sensitivity of the assay systems used. Numbers in ou¡ sfudy are not large enough for statistical comment,
but our findings a¡e consistent with those of previous worþ and by a more sensitive quantitation of IgG4,
we have found that this isst)?e was frequently deficient in immunodeficiency of this type. These fudiogr,
taken in conjunction with the facts that IgA deficiency is more cotnrnon than IgG deficiency which is more
common that IgM deficiency, suggest the deficiencies in immunoglob'lin isotypes may reflect a progressive
sequential impairment of the heavy chain constant region gene arrângements such that deficiencies
resulting from more downstrea- switching occur more frequently (Ch. 1.3.2).
Isotype switching and IgG subclass deficiencies
A computation of tle relative frequency of deficiencies of tle IgG subclasses was made f¡om all
the patients studied in the cou¡se of this project (excluding those IgA-deficient subjects in whom IgG
subclasses were meÍìsured by EIA where IgG4 deficiency could not be defined). The results a¡e
st'mmarised in Table l4.L IgG4 deficiency accounted fot 44Vo of the IgG subclass defi.ciencies, IgG2 for
2ZVo,IgGl for LSVo and IgG3 for 15Vo. Thus, in the disease-groups of patients studied the frequency order
of IgG subclass deficiencies was tle reverse of the order of the IgG subclass heavy chain gene arrangement
on cb¡omosome 14 (Ch. 1.3.2). Thi.s suggests that isot¡pe production which requires more downstream
switching is most likely to be impaired and highlights the need for a better understanding of the
mechanisms controlling IgG heavy chain switching so that IgG subclass deficiencies may be better
understood" better treated, and possibly even prevented.
Taken together, the relative frequencies of the different IgG subclass deficiencies, and the
association of IgA and IgG4 deficiencies (section 1-.5, Ch. 5) suggest that a defect in isotype switching would
seem a likely e4planation for many IgG subclass deficiencies. Studies to determine whether the partial IgA

Table 14.1
OVERALL DISTRIBUTION OF IgG ST]BCLASS DEFICIENCIES IN TIIE PATIENT GRotTPsSTUDIED
Patient Grouo Number of patients with deficiences
IgGl IgG2 IgG3
282
IgA-deficient(ch.5)
Osteomyelitis andseptic arthritis
Hib infection
Bronchiectasis
Giardiasis
Known immunelsgic¿ldisorders (Ch. 10)
Splenic dysfunction
Others (Ch. 12)not so fa¡ included
(6)7
(4)9
(1)6
IgG4
(<e)?Á
2I1 )
2
22
43
3
4
L
6
1
2
)
5
1
68
)
8
Total7o of deficiencies
Numbers in parentheses indicate those from the prelimi¡ary study in IgA-deficient patients in whom theincidence of IgG4 deficiency could not be determined because of limitatiõns in assay sänsitivity.
'26
22
2T
18524
18
15

283deficiencies that a¡e often associated with IgG4 deficiencies involve both IgAl aú,IW may be helpful. If
one postulates that the deficiency relates to impaired isotype switching, deficiencies of IgA1, rather than
IgA2, would be more likely in the absence of lgGZ deficiencies.
The finding that lpaphocytes from three hlpogammaglobrrlinaemic and from one IgG subclass-
deficient patient produced IgM (t IgG3) but failed to produce IgG subclasses of other isot¡res on
stimulation with a T cell-independent mitogen suggests a non-T cell-dependent defect in isotype sryitching.
Conversely, the failure to produce imm¡¡sglsþrlin of any isotype when cells were stimulated with a T cell-
dependent mitogeq suggests a defect in T cell regulation of immuneglsþr,lin production. These results,
which appear contradictory could be reconciled by postulating that although S. au¡eus is considered to be a
T cell-independent mitogen, the degree of T cell-independence that it exhibits may be inversely related to
the degree of isotype switching Decessary for production of the isotlpe in question. This being so, the
primary defect in the four patients studied and possibly in many other IgG or IgG-deficient patients is
likely to be a defect in T cell regulation of isotype switching. The finding of hypog'-maglobnlinaemia with
a CD4 deficiency and impaired cytokine production in one of ou¡ patients many monfhs after a bone
mÍurow transplantation may þ¿ve simil¿¡ implications. Studies of fi¡st the production of immunoglob,lins
by the patient's lymphocytes cultu¡ed with T cells from normal subjects, and secondly cytokine production
by the patient's T lymphocytes relative to that of T þmphocytes from normal subjects may help to clari&
this hypothesis. Specific defects in cytokine production have already been described in some
hypogammagloþr,lin ¿e¡1is patients (Ch. 1.3.3. 1).
Further elucidation of the mechanisms underlying hypogammaglob'linaemias and IgG subclass
deficiencies may pave the way for the developmen¡ o¡ lschniques to modiS immunoresponsiveness which
could be used therapeutically and ci¡cumvent the need for regular immunqglqþrrlin infusions in some
patients with defects in humoral immunity. In time, the therapeutic use of cytokines to promote antibody
production may become possible (Farrant and Webster, 1989).

L4.2 CONCLUSIONS 284
Having developed a sensitive and reliable ELISA for IgG subclass quantitation and established
age-normal percentiles for IgG subclasses for the paediatric population in South Australia, we a¡e now able
to detect abnormalities of IgG subclass concentrations in children. We have shown that subnormal
concentrations of IgG4 are the most common IgG subclass deficiencies in a number of groups of infection-
prone patients, particularly those prone to respiratory tract infections. Therefore, if the problem of IgG
subclass deficiency is to be appreciated fully it is essential tlat assays sensitive enouqh for IgG4 quantitation
be used.
Fu¡tlermore, because of the considerable differences between paediatric age-normal ¡anges
obtained in different geographical locations, by different assay tecbniques and with different reagents, the
ideal is that, until more interlaboratory standardisation is achieve{ IgG subclass concentrations should be
assessed by comparison with age-normal rânges obtained in the same laboratory, with the same technique
and the same reagents.
We have confirmed and eÍended tle work of some previous investigations showing that there is an
association between IgA and IgG subclass deficiencies and infection proneness. IgG4 deficiency was the
most commonly found deficiency in ou¡ studies both in association with IgA deficiency and apart from IgA
deficiency. Many investigators have been unable to comment on the occrrrence of IgG4 deficiency because
of limitations of the assay system and normal rânges used. To date, the mechanism by which IgG4, an
apparently minor imm¡¡eglsþnlin isstj¡ps, is important in defence against infection is unclear.
Although we recognise that antibody deficiency rattrer than IgG subclass deficiency perse is likely
to be the determining factor in infection proneness, screening for antibody deficiencies is hþtrly complex,
and age-normal ranges for concentrations and resporuies to immunisation are not generally available. The
isotype pattern of responsiveness to antigens may also þs important, but not enough data is available for
this to be clinically usefiil at this stage. Since some subjects can respond well to one antigen and not to
another, even when antþens are of the same class (e.g. polysaccharide) antibody screening tests may need
to be very extensive to detect deficiencies. At present, we consider IgG subclass quantitation as a useful
investigation in infection-prone subjects where 1þe f¡¡ding of an IgG subclass deficiency may be ¿ sign of an

285antibody deficiency state, and so give di¡ection for further antibody tesring. Some antibody-deficient
patients will be missed by this approad as not all antibody-deficient patients are IgG subclass-deficient.
Nevertleless, IgG subclass quantitation is a significant advance in the assessment of i¡fection-prone
individuals, particularly of those in whom respiratory infections are a problem.
There is potential for the prevention of infective episodes and chronic infections by
immunoglobulin replacement therapy in infection-prone IgG subclass-deficient patients. Multicentre
controlled trials a¡e needed to develop a consensus about oprimal dosages, frequency of administ¡ation and
effectiveness of this treatment. There are still many unresolved questions in this area.
The relalive frequency of IgG isotype deficiencies found in ou¡ sfudies indicates that the more
downstream isotype switching required for the production of an IgG subclass, the more likely is a deficiency
of that subclass. Studies of the mechanisms involved in the cont¡ol of isotype switching will play an
inportant role in furthering understanding of and, hopefully, i¡ e¡¡s¡ding treafment options for IgG
subclass-deficienct patients in the futue. Flowever, the evidence discussed previously indicates that defects
in isotlpe switching are likely to be only one of several ¡xesfoanisms of immunogleþulin isotype deficiencies.
I4.3 RECOMMEI\DATIONS
Until further interlaboratory standardisation of IgG subclass measurements is achieved"
laboratories measuring IgG subclasses should develop thei¡ own age-normal rânges using the same
assay and reagents as they use for patient samples.
IgG subclass assays should be sensitive and accurate enough to quantitate all the IgG subclasses in
sera from control subjects of all ages. Assay sensitivity is particularly important where IgG
subclasses in paediatric 5amples are being quantitated.
IgG subclass quantitation should be included in the immuns[qgical work-up of infection-proue
individuals, particularlS but not onl¡ where there is IgA deficiency or where respiratory infections
are prominent.
1.
2.
3.

4.
6.
5.
286Studies of 'specific' IgG subclass antibody production should be pursued at the research level with
the aim of providing a framework for the assessment of normal and abnormal antibody
responsiveness and an urderstanding of the importance of the isotlpe patterns of antibody
production in normal and abnormal individuals.
Serial studies of IgG subclass concentrations, particularly in patients in whom a deficiency has been
found at one point in time a¡e needed as the natural history of IgG subclass deficiency is largely
unknown.
Studies of the IgG subclass composition of mucosal secretions f¡om different sites should be done
to help determine whether IgG4 is primarily a mucosal inmunoglobulin.
A multicentre trial to study the optim¿l treatment of IgG subclass deficiency, particularly with
reference to the use of intraveno¡s imm¡¡sglobulin, is needed.
Studies of the mechanisms underlþg IgG subclass deficiency, particularly in the a¡ea of the effects
of cytokines in isotype switching should be pursued with the aim, Dot only of promot'ng an
u¡derstandi.g of these deficiencies, but, eventually of developing a treatment more effective tlan
the regular ¿dministration of intravenous immunoglobrrlin for patients with antibody deficiencies.
7
8.

287
BIBLIOGRAPI{Y
Aalberse RC, van der Gaag R, van der Leeuwen J. 1983. Serologic aspects of IgG4 antibodies. 1.Prolonged immunis¿fien results in IgG4-restricted response. J. Immunol. 1j0:722-7?5.
Abbas AIC 1988. A reassessme¡¡ qf msçþ¡nisms of ancþen-dependent B-cell activation. Tmmunsl. Today.9:89-94.
. Imm¡¡¡¡1' and hyp n. In: StaphylococciVol. 1. Clinical an CSF and Adlam C,n5323.
Alexander EL , Sanders SIC 197?. F(ab1)2 reagents are not required if goat rather than rabbit antibodiesafe used to detect htt-an surface immunsglsþulin. J. Immunol. 119:10g4-10gg.
Allday MI , Crawford DH. 1988. Role of epithelium in EBV persistence and pathogenesis of B-celltumors. I¿ncet. 1:855-857.
Alm¡oth G, Sjostrom P, Svalander C, Dauielsson D. 1989. Serum immunogloburins and IgG subclasses inpatients with glomerulonephritis. J. of Internal Medicine. 225:3-7.
Ambrosino D, Schiffrnan G, Gotscblich EC, Schur P, Rosenburg G, del-ange GG, van I-oghem E, SiberER. 1985. Correlation between G2m (n) immunoglobrlin ¿llstlpe and human antibody response andsusceptibility to polysaccharide encapsulated bacteria. J. Clin. Invest. 75:L935-L942.
Ambrosino DM, Siber GM, Van Loghem E , Schiffman G. LQBZ. Increased antibody respouse toimmrtnizagisn with Haemophilus influenzne type b (Iß) and pne,mococcal capsular potysãccnaride (pS)in adults with the G2m (n) allotlpe. Pediat¡. Res. 16:2344 (Abstract).
Ambrus M, Hernadi E , Batjai G. 1977. Prevalence of HI-A-41 and HI-A-88 antigens in selective IgA-deficiency. Clin. Im m unol. Im - unstathol. 7:311-314.
Ament ME, Rubin CE. 1912. Relation of giardiasis to abnormal intestinal st¡ucfu¡e and function ingastrointestinal immunodeficiency syndromes. Gastroenter ol. 62:2L6-226.
{s¡ann AJ, Ashman RF, Buckley RH, Hardie wR, Krantmann flJ, Nelson J, ochs H, Stiehm E\ TillerT, Wara DW, Wedgwood R. 1982. Use of intravenous gamm¿g[sþr¡lin in antibody immuuodeficienqnresults of a multi-centre cont¡olled trial. clh. Tmmunol. tmm¡¡sp¿thol. D:604.¡.
Ammann AJ, Fudenberg HH. 1982. Clinical lrnmunolog5¿ ¡ fmm¡¡sdeficiency diseases. In. Basic andClinical Immunolory, Stites DP, Stobo JD, Fudenberg HH. and Wells JV, eds. Lange, California. pp. 395-429.
Ammann AJ, Hong R. Lnl.literature. Medicine. 50:223-DÁ.
Selective IgA deficiency : presentation of 30 cases and review of the
Anderson Cl Looney RI. 198ó. Human leucocyte Fc receptors. Tmmunol Today. 7:26Ç266.
And¡eas JS, Hewlett EL. 1981. Protection ¿gainst infection with Giardia muris by milk çg¡tainingantibody to Giardia. J. Infect. Dß.2:?12-?-46.
ArþaT, OkanoM, TakahashiY, Sakiyama Y, Matslmoto S. L987. Analysis of B cell dysfunctioninpatients with com-on variable immunodeficiency by u"i"g recombinant i¡¡e¡lenkin 2. foh-oku J. Exp.Med.152:53{1.
Asano Y' Singer A' Hodes RJ. 1983. Role of the major histocompatibility complex in T cell activation of Bcell subpopulations. Ir gene regulation of the T cell i ependent aCtivation of disltinct B cell subpopulations.J.Immunol. 730:67-71.

288
Aucouturier P, Barra d Intrator L, Cordonnier C, Schulz D, Duarte F, Vernant J-p, preud,homme J-L.1?87b' Long lasting IgG subclass deñciency 11d__antibacteáal polysaccharide antibådy deficiency afterallogenic bone marrow ¡ansplantation. Blooã. 7 0;779 _jgi.
Aucoutu¡ier P, Bremard-Oury C, Clavel JP, Debre M, Griscelli C, Seligman M and preud,homme J-L.1986a. Serum IgG subclass levels in primary imm¡¡sdeficiency. Monogr. Alletg.?n:62_74.
Aucouturier P, Bremard-Oury C, Griscelli M, Berfhis¡ M and preud'homme J-L. !9g7a. Serun IgGsubclass defi cienry in ataxia-telan giectasia. Clin. e4p. tm m unol. g:3V2_396.
Aucoutu¡ier P, Couderc LI, Gouet D, Danon P, Gombert J, Matlerson S, Saimot AG, Caluvel Jp andPreud'homme J-L. 1986b. Serum imm¡¡sgleþutin G subclass dysbalances in the tlrmphadenopathyslmdrome and acquired immune deficiency syndrome. Clin. Exp. Irnmunol. 63:2-3L?A0.
Aucoutu¡ier P, Lacombe C, Bremard C, Lebranchu y, Seligrnann þf, Griscelli C, preud'homme J_L. 19g9.Serun IgG subclass levels in patients with primary immunoãeficiency s¡m.dromes or abnormal susceptibilityto infections. Clin. Tmmunol. Tmmrnsp¿ft ol. 5t:22_37 .
Aucoutu¡ier P, Mounir S, Preud'homme J-L. L985. Distribution of IgG subclass levels in normal adult seraas determined by a competitive ertlr]rnie imm¡¡sass¿y using monoclõnal antibodies. Diagnostic Immunol.3:l9L-I96.
Babb RR, Peck oc, vescia FG. 1971. Giardiasis. A cause of t¡aveller's diarrhoea. JAMA. 2L7:1359-L:%t.
Ballieux RE, Bernier GM, Tominaga\ Putnam FW. 19t6i{. Çammaglob'lin antigenic types defined inheavy chain determinants. Science. 145: 168-170.
Bannisfe¡ KM, Howarth GS, Clarkson AR, Woodroffe AJh u'n an glomeruloneph¡itis. Clin. Nephrol. 19: 161-165.
1983. Glomerular IgG subclass distribution in
Barandun S, Kistler P, Jeunet F, Isliker H. 1962. Intravenous a¡lminist¡.ation of human gamm¿glsþ¡lin.Vor Sang.7:157-174.
Barandun S. 1985. Imnunoglobnlins¡ history present trends and safety aspects. In intravenousimmun6glsþt'lins in immunedeficiency syndromes and idiopathic thrombocytopu""i" purpura. Waters AHand Webster ADB eds. O¡dord Uni, Oford. pp. 1-10
Bass JL, Nuss R, Menta Kd Morganelli P, Bennett L. 1983. Recurrent psningococcaemia associated withIgG2-subclass deficiency. New Engl. J. Med. 309:430.
Bea¡d LI, Ferrante A,- Hagedorn JF, Leppard P, Kiroff G. 1990a. Percentile ranges for IgG subclassconcentrations in healthyAustralian child¡en. pedi rtr. Infect. Dis. J. 9:s9-15.
Bea¡d lJ, Ferrante,t oxelius V-4, Maxwell GM. 1986. IgG subclass deficiency in chilfl¡s¡ with IgAdeficiency presenting with recurrent or severe respir lory infections. Pediatr. nesearch. 2O:937-942.
Bea¡d LI, Ferrante A. 1988. IgG replacement therapy i" IgG subclass deficient child¡sa. Monogr.Allergy. ?3:L9Ç?Ã3.
Beard LI, Ferrante A. 1989' IpS4 deficiency in IgA-deficient patients. Pediatr. Infect. Dis. J. g:705-70g.
Bea¡d LI, Ferrante A. 1990. Aspes¡s sf immunoglobrlin replacement therapy. pediatr. Infect. Dis. J.9:55¿l-61
Bea¡d LI, Ferris L, Ferrante A. 1990b. Immunoglobulin G subclasses and llmphocyte subpopulations andfunction in osteomyelitis and septic arthritis. Acta paediatr. scand. zg: in press.'
Bea¡d IJ, Maxwell GM, Thong YH. 1981. Tmrnunocompetence of children with frequent respiratoryinfections. Arch. Dis. Child. 56:101-105.

289
Bea¡d LI, Roberton EF, Thong YH. 1980. Pa¡a-influenza pnerL onia in diGeorge syndrome two yearsafter thymic epithetial t'ansplantation, Acta paediat¡. scan¿- oõ:qol_co6.
Beard LI, Thong YH' 1981. Immunological competence of children with pyogenic meningitis. Eur. J.Pediat¡. 136:Bl-2i5.
Beck CS, Heiner DC. l-981. Selective immunoglobulin G4 deFrciency and recurrent infections of therespiratory tract. A¡¡. Rev. Respir.Dts. 124:9Ç96.
Beck O, Kaiser P. 1981a. Nephelometry of hnm¿¡ IgG subclass concent¡ation in serum. Clin. Chem.17:310-313.
Beck oE, Kaiser PE' 1-981b. Distribution of human IgG subclasses in commercial intravenousimm¡¡¡sglsþ rlin preparations. A rate nephelometric method. îor sang. 4L:79-M..
Beck OE. 1981. Distribution of virus antibody activity among h,man IgG subclasses. Clin. Exp. rmm¡¡sl.43:6?Á-632.
Bender S, Hetherington S. 1987. Haemophilus influenzae type b opsonins of int¡avenous imm¡¡egleþrlins.J. Clin. Immunol. 7 :47 5-480.
Benson EM, Beìtol;'t; JN, Brodtma¡n ME. 1990. T cell regulation of immunoglobrlin is6f¡ps5 in healthand disease. Pediatr. Infect. Dis. J. 9:525-35.
Berkel AI. 198ó. Srudies of IgG subclasses in ataxia-telangiectasia patients. Monogr. Allergy. 20:100_105.
Bird D' Duffy s, Isaacs A, webster ADB. 1985. Reference ranges for IgG subclasses in preschoolchild¡en. A¡ch. Dis. Child. 6O.?ÃL?Ã7.
Bird P, Lachma"n PJ.- 1988. The regulatiol ^o-f
Ig9 subclass production in man: low serum IgG4 ininherited deficiencies of the classical pathway of c3 aãtivation. gur. J. Immunor. Lg:rzr7-r222.
Bjorkander J, Bake B, Oxelius V-An Hanso"_L+. 1985. Impaired lung function in patients with IgAdeficieucy and low levels of IgG2 or tgG3. N. Eogl. J. Med. lts:izn-lzq.
Bjorkalder J, Hamm¿¡strom L, smith ECI, Buckley RH, cumingham-Rundles c, Fl¡ns.n r-A. rgg7.Immunoglobrtlìn prophylaxis in patients with antibody deficiency .yodro-", -¿ ^ti-Iga antibodies. J.Clin. Immunol. 7:8-15.
Bjorkander J, Oxelius V-4, Soderstrom R, Hanson I-A. 19gg. Immunoglobulin t¡.e¿t¡¡snt of patients withIgG and IgA deficiency states. Monogr. Allergy. ?3:160_167.
Bowen TJ, Ochs HD, Altman LC, Price TTI,-van Epps DE, Brautþan DL, Rosin RE, perkins WD, BabiorBM, Klebanoff SJ, Wedgwood RI. 1982. severe iècurreut bactJrial infections ^.ó.iut"d
with defectiveadherence and chemotaxis in two patients with neutrophils deficient in a cell associated glycoprotein. J.Pediat¡. I0L:932-94O.
Box GE, Cox DR. 1964. Analysis of t¡ansformation. J. R. Stat. Soc. B. ?Á:211_252.
Brearley RL, Rowbofham BN. 1984. High-dose gamm¿glsþulin for idiopathic thrombocytopenic purpura.Aust. NZ. J. Med. 74:67-(ß.
Bremard-Oury C, Aucouturier P, Debre M, Preud'homme J-L, Griscelli C. 19g6. Immunoglobrlin Çsubclasses in patients with imm¡¡odeficiencies. Monogr. a[ergy. 20:75-79.
Brieva J'\ Targan S, Stevens RH. 1984' M_Td T cell subsets regulate antibody production by human lnvívo anttgen-induced lymphoblastoid B cells. J. tmm rnol. Íi2:611-615.
Briles DE, Scott G, Gray B, Crain MI, Blaese M, Nahm M, Scott V, Haber p. 19g7. Naturally occurringanti-phosphocholine antibodies: a potential index of anti-polysacch-ia" ,"spoos¡|,;;",| J. Infect. Dis.155:1307-13L4.

290Buckley RH. 1985. Gamm¿gtqþuri. replacement. Clinics in Immunol & Allerry.5:141-15g.
Burgro GR, Druse M, Monago V, Ascione A, Nespoli L. 1-980. Selective IgA deficiencp Clinical andimm¡¡6l6gical evaluation of 50 pediatric patients. Eur J. pediatr. Íi3:101-10ó.
Bu¡ks AW, Steele RW. l-986. Selective IgA deficiency (review article). Allerry. 57:3-10.
Bu¡ton DR. 1985. Imm¿¡sglsþrrlin G: fr¡nctional sites (review). Mo[ fmmunol. 2Z:I6L_?û6.
Butler JE, Cantarero I-4, McGivern PL. 1980. Measu¡ement of bovine subclass antibodies (IgGl andIgG2) using the ampliñed snztmeJinked immunosorbent assay. Mol.Immunol. L7:645_653.
Catty D, Jassin A, Hassan Ç Raykundalia C. 1986. IgG subclasses in parasitic i¡fections. Monogr.Allergy. L9:L44-155.
Catty D, Seger R, Drew R, St¡oder J, Metze H. lW, IgG subclass concentrations in cord sera frompremature, full-term and small-for-dates babies. Europ. ¡. pèdiat. l2s:g9-96.
Cebra JJ, Komisa¡ JL, Schweitzer PA. 1:gu. C¡¡ isotype 'switching' during normal BJlmphocytedevelopment. Annu. Rev. Immunol. 2:493-548
Qfoerrng N:K V,'Bles5ing-Moore J, Reid MI, Yang G. 1983. Honey bee venom specific immunoglobrli'G4 in honey bee sting allergic patients and bee keepers. Ann. Allergy. 50:155-160.
Christensson B, Johansson PJH, Oxelius VA. 1986d. Imbalanced serum IgG subclass pattern in toxicshock syndrome patients: deficiency of specific IgGl and IgG4 subclass antiboáies to toxic fuock syndrometoxin I. Clin. Exp. Tmmunol. 6.44349.
Clark JA., C¡llicoat PA, Brenner Nd Bradley CA,, Smith DM. 1-983. Selective IgA deficiencies in blooddonors. Am. J. Clin. Pathol. 80:2L0-2ß.
coffman RL, ohara J, Bondlvtw, carty J,Z.otnik d paur wE. 19g6. B cell stimulatory factor - 1euhances the IgG response of lipopolyssacharide-activated B cells. J. Immunol. 13ó:453g-4541.
Cohen RC, Ferrante A. 1982. rmm¡¡e dysfunction in the presence of residual splenic tissue. A¡ch. Dis.Child. 57:5?3-52i7.
Cohn JE', Gurd FRN, Surgenor DM, Barnes Bd Brown RI! Derouaux G, Gillespie JM, Kahnt FW,læverWF, Liu CH, Mittelmann D, Mounton RF, Schmid I! Uroma E. 1950. A system^for the separation for tlecomponents of hrlman blood: quantitative procedures for the separation of the protein components ofhuman plasma. J. Am. Chem. Soc.72:464 474
Coleman RM, Nahmias AJ, Willi¡ms SC, Phillips DJ, Black CM, Reimer CB. 19g5. IgG subclassantibodies to herpes simplex virus. J. Infect. Ois. fSf:q2q-g36.
Collins-Williams C., lamenzaC, Kokuba H. 1%8. Deficiency of IgA in serum and respiratory secretions.Can. Med. Assoc. J.99:1069-L07I.
Croce CM, Shander M, \a[¿¡'tinis J, Cicurel L, D,Ancona GG, Dolby T,W, Koprowski H. lng.Chromosomal locations of the genes for human immunoglobulin þs¿vy chains. pioc. ñat. Acad. Sci. USA.76:34t6-AL9.
Cunningham-Rundles C, Hanso¡ tA' Hitzig WH_ ef at. L982. Appropriate uses of h ,man immunoglob.linin clinicål practice. Memorandum from an IUIS/WHO 6ssting.-
-nou. wrro. 60:43-47.
Cunningham-Rundles C, Oxelius V-4" Good RA. IgG2 and IgG3 subclass deficiencies in selective IgAdeficiency in the United States. 1983. Bhth Defects Ló-1|ß-L75.
Cunningham-Rundles C, Siegal F, Smithwick EM, Lion-Boule E, Cunningham-Rundles S, O,MaIley J,Barandun S, Good R. t98/,. Efficacy of intravenous imnunogtãUuün in primary humóralimmunodeficiency clisease. Ann. Intern Med. 101:435-439.

29LCypess RH. 1985. Mechanis¡¡s of imm¡¡¡¡y to parasitic diseases. l¡; Imm¡¡slory III, Bellarti JA" ed.Philadelphia, WB Saunders Co. pp. rc3n.Delovitch Tl,-semple JW, Phillips ML. 1988. Tnfl¡s¡se of antþen processing on immune resporsiveness.Tmmunol. Today. 9:ZL6-2I7.
Dengrove J,l'f.,e EI, Heiner DC, St Jeme JW, læake R, Ba¡off LI, Ward JI. L9g6. IgG and IgG subclassspecific. altibody responses to diphtheria and tetanus toxoid in newborns and
"infants ä"* DT"immunization. Pediat¡. Res. 20:735-?39.
Devey ME, Bleasdal EK Isenberg DA. 1987. Antibody affinity and IgG subclass of responses to tetanustoxoid in patients ¡vi¡þ ¡þsnm¿toid a¡th¡itis and systemic-þus erJ,the-ãtor*. Clin. E>cp. T--unsl. 6g:562-569.
Devey ME, Doak D' 1974. A criticål study of the IgG subclasses of Rh anti-D antibodies formed inpregnaDcy and in immunized volunteers. Immunology. 2jT:I073-L079.
Devey ME, Wilson DV, Wheeler Aw. 1976. The IgG subclasses of antibodies to grass pollen allergensproduced in hay fever patients during hyposensitization. Clin. Allergy 6:227-2K.
Di Padova F, Durig M, Wadstrom J, Harder F. 1983. Role of spleen in imm¡¡s response to poþalentpne rmococcal vaccine. Br. Med. J.?Å7:1829-1g32.
Djurup R, Malling H-J, Sondergaard I, Weeke B. -1985-
The IgG and IgG subclass antibody response inpatients allergic to yellow jacket venom undergoing different regimeni of venom immunotherapy. J.Allergy Clin. Immun ol. 7 6:46-55.
Djurup R, Mans¿ B, Sondergaard I, Weeke B. 1988. IgG subclass concentrations in sera from 200 normaladults and IgG subclass determination of 106 myeloma proteins: an interlaboratory.t,ray. S.uod. J. Clin.Lab.Invest. 48:77-83.
Djurup R, osterballe o. 1984. IgG subclass antibody response in grass pollen-allergic patients undergoingspecific immunotherapy. Allergy . 39:433-441.
Doi S, Safü o, Hara T, Sugita T, Ha-Kawa K Tanaka T, Hara H, Negoro S, yabuuchi H, Kishims¡6 5.1989. Admi"ist¡ation of recombinaún-2 argrnents the level of serum IgM i" anll_Z deficient patient.Eu¡. J. Pediat¡. 148:630-633.
Douglas RM, Paton JC, Duncan SJ, Hansman DJ. 1983. Antibody response to pneumococcal vaccinationin child¡en yorìnger than 5 years of age. J. Infect. Dis. 1¿E:13j. -1j7.
Douglas SD. 1982. Development and structure of cells i¡ ús immune system. In: Basic and ClinicalImmunolog¡r, 4th edition. Stites DP, Stobo JD, Fundenberg HH and weils w, eds. r-ange, ¡.os Altos,California, USA. pp. 65-88.
Dray S. 1960. Tb¡ee gamm¿-glsþrtlins in normal human serum revealed by monkey precþitins. Science732:1313-7314.
lrew PA" Kiroff GK Ferrante A" Cohen RC. 1983. Alterations in imm¡¡sgl6þntin spthesis by peripheralblood mononuclea¡ cells from splenectomized patients with and without Srcoi. r"þo*tt. J. rmmunol.732:l9L-I96.
!rutz DJ, Graybill JR. 1982. lnfectious diss¿sss. In: Basic and clinical rmmun6l6gy, 4th edition. stitesDP, Stobo JD, Fudendberg HH and Wells JV, eds. L^¡ge,I-os Altos, California" USÃ.'pf. Sø-Sea.
Eid AM, Issa Ha¡d Dief AI. 1980. Som-e immunological aspects of staphylococcal haematogenousosteomyelitis. Æch. Orthop. Tra'ma. Surg. 96:221-2?A.
Einhs¡¡ MS, Granoff DM, Nahm MH, Qtrinn d Shackelford PG. 1987. Concentrations of antibodies inpaired maternal and infant sera. Relationship to IgG subclasses. J. pediatr. ttt:l1Z-i17.

292Emmanuel D. Ú88. Issues concerning the use of intravenous immunsglsþillins for theiS.nunop-rophylaxþ of cytomegalovirus infectioà i" allogeniclo* -ut o* 1¡ansplant rãcipients. Monogr.Allergy. ?3:2I6-2?t1.
Endres RO, Kushnir E, Kappler JW, Marrack P, Kinsky SC. 1983. A requirement for nonspecific T cellfactors in antibody responses to'T cell independent, antigens. J. Irnmunol. 130:7g1.-7g4.
Falc¿o RP, Ismael SJ, Donadi EA. 1987. Age-associated changes of T lprphocyte subsets. Clin. fmmunol.5:2A5-?n8.
Farid NR, Newton MR, Noel EP, Marshall w]d. rw. Gm phenotlpes i¡ ¿utsimmune thyroid diseases.J. Immunoge net. 4:429 -432.
Farrant J, Webster ADB. 1989. BImmunol. 78:32L-322.
cell-cytokine interactions: lessons fr66 clinisal imm¡¡slsgy. Cli". E.p.
Fei¡stein A" Richardson N, Taussþ MS. 19g6.rmmunology Today. 7 :L69-17 4.
Imm¿asglsþrlin flsxiþility in complement fixation.
Ferguson A, Mcclure JP, Townley RRw. L976. Intraepithelial lymphoryte counts in small intestinalbiopsies from child¡en with diarrhoea. Acta pediat¡. Scand. 65:54L-56.
Ferrante A" Beard LI, Feldm¿¡ ft6. 1990. IgG subclass distribution of antibodies to bacterial and vi¡alantþens. Pediatr.Infect. Dis. J. 9:St6-?A.
Ferrante A" Beard LI, Thong YH. 1980- Early decay of human neutropbil chemotactic responsivenessfollowing isolation from peripheral blood. Clin. E4p. Immunol. 39:532_537.
Ferrante A, Bea¡d lJ. 198s. IgG subclass Írssays with polyclonal and monoclonal antibodies. Monogr.Allergy. 23:61.-72.
GP, Beard LI, Goh DHB. 1989. Alterations in ftrnction and subpopulations ofonuclea¡ leucocytes in child¡en with portal hypertension. Int. A¡ch.^Ailergy Appl.
Ferrante A, Drew PA" Kiroff G,7-nlalJ. 1987. Peripheral blood leucocyte subpopulations in patientssplenectomized for t¡a'ma. Clin. Exp. Immunol. ZO:t5g-t03.
Ferrante d Kiroff GII Drew PA. 1986b. Elevated natu¡al killer (NK) cytotoxicity of mononuclea¡leucocytes from splenec¡emissd patients: increase in læu-7+ and r-eu-11i r"u*.yt".- ðí¡". n¡¡p. Immunol.64:173-t80.
Ferrante A" Ki¡off GIÇ Goh DHB, Drew PA. 1985. Elevated natu¡al killer (NK) cell activitp a possiblerole in ¡esistance to. infection and malignancy in immunodeficient splenàctoLize¿ patiénts. Med.H¡'pothesis . l6:133-L46
Ferraute A" Rencis Vo. Lgu. Enhancement of base hexose-monophosphate shunt activity of hr¡manpolymorphonuclear leucocytes by human beta-interfe¡sa. Tmms¡ol. Lett. AilS_Ztl.
Ferrante d Rowan-Kelly B, Beard LI, Maxwell GM. 1986a. An enzSme-linked immunosorbent assay forthe quantitation of human IgG subclasses using monoclonal antibodies. J. rmrnunol. Methods. g3:?n7-212.
Ferrante A, Rowan-Kelly B, Paton JC. Lg8/,. Inhibition of in vit¡o hrrman lynphocyte response bypne rmococcal toxin pneu'nolysis. Infect. Immun. 46: 5g5-5g9.
Ferrante d Rowan-Kelly B' 1983. Activation of the alternative pathway of compleme nt by Acønthanoebøcuheñsoni. Clin. Exp. Immunol. 54:477-485.
Ferrante A, Thong YH. 1ln8. A rapid one-step procedure for purification of mononuclea¡ andpollmorphonuclear leukocytes from þrrm¿¡ blood using à modificatioo ôf th" hypaque ficoll technique. J.Immunol. Methods. ?A:ß9-393.

293Freijd d rlamm¿¡stlom L, Persson MA,ql Smith cIE. 1984. Plasma ânti-pnerrmoooccal antibody activityof the IgG class and subclasses in otitis-prone child¡.en. Clin. EÀ". Tmmunol. 56:233_?5.
French MAH, Ha¡rison G. L984a. Serum IgG subclass concentra[ions in healthy adults. A study usingmonoclonal antisera. Tmmunol. 56:{73-475.
French MAH, Harrison G. 1984b. Systemi_c antibody deficiency in patients with serum immunoglob'lindeficiency or with selective IgA deficiency. Clin.Exp. rmmunol. iø:tg-n.
French MAH, Harrison G. 1985. The IgG subclasses of tetanus toxoid antibody following immnnizatis¡;studies in healthy adr¡lts and patieuts with ch¡onic respiratory tract infectio^. ¡. Clin. Lab. Tmmunol.I8:2L-?5.
Frommell D, Moullec J, Lambin P, Fine JM. 1973. Selective serum IgA deficiency. Frequency among15200 French blood donors. Vox geng. 25:51IÌ-1g.
Gearhart PJ, Hurwitz JL, Cebra JJ. 1980. Successive switching of antibody isotlpes expressed within 1¡¡".of a B cell clone. Proc. Natl. Acad. Sci. USA. 77t542+S4Zg.
Geha RS. l-988. IgG antibody responses to polysaccha¡ide in IgG subclass-deficient child¡en. Monogr.Allergy. 't?:97-102.
Gergely J, Atky L, Medgyesi GA. L967. Non specific interaction of the papain-insensitive and papain-sensitive populations of human IgG with erythrocytes. vox sang. LZ:252-2&.
Gillon J. Barclay GR, Yap PL, Ferguson A. 1-986. Food antibodies, intestinal permeability and HI-A statusin IgA-deficient blood donors identified by a new rapid screening test. Clin. ai"rgy. 16:5g3-5gg.
Goodman JW. 1982a. Immu¡oglobrlins J: Structu¡e and function In: Basic and Clinical Immunology, DpStites, JD Stobo, HH Fundenberg, JV wells, eds. Lange. california. pp.33-35.
Goorlman Jw. 1982b. I'n-unogeniciry and antigenic specificiry. In: Basic and Clinical Inmunolog¡r. Dpstites, JD stobo, HH Fundenberg w wells, eds. Lange,california. pp.21-29
Granoff DM, Shackelford PG, Pandey JP, Bites EG. L986a. Antibody responses to Haemophilusinfluenzae type b polysaccharide vaccine in relation to kn (1) and G2m (z:)'inmunoglobutin allotypes. J.Infect. Dß. 154:257 -2ß.
Granoff DM, Shackelford PG, Suarez BK Nahm MH, Cates KL, Murphy TV, Karasia R, Osterholm MT,Pandey JP, Dau'n RS and the collaborative group. 1986b. Haemophilus inÍluerase type b disease iochildren vaccinated with type b polysaccharide vaccine. N.E"gl.J. Med. ¡rs:rsa+u90.
Granoff DM, Sheetz KE, Nahm MH, Madassery JV, Shackelford pG. 19gg. Fu¡ther immunelggisinvestþation of children who develop haemophiius disease despite previous vaccination with t,,pe"bpolysaccharide vaccine. Monogr. Allergy. 23:256.2ß.
Grey HM, Kunkel HG. 1964. H chain subgtoups of myeloma proteins and normal 7S gamm¿-globrlin. J.E4p. Med. 7?Ã:253-?ß.
Grill BB, Bui HD, Jauhar R, Heiner DC. 1985. I-ow duodenal IgG4 levels in patients with giardiasis(abstr). Clini Res. 33:1%A.
Grubb R, LaurellAB. 1956. Hereditary serological hu-an semm groups. Acta pathol. Microbiol. Scand.39:390-398.
Gwpn CM, Smith JM, Iæon GI, Stanworth DR. 1978. Role of IgG4 subclas5e5 i¡ chilflhood allergy.Lancet L:910-911.
Haas A, Stiehm AR. 1987. Failure to tlrive, thrush and hlpog¡mmoglobrtinspa in a 6_month_otd infant.Ann. Allerry. 59:l4L-L44.

294Hamilton RG. 1987- fl'man IgG subclass measurements in the clinical laboratory. Clin. Chem. 33:L7Al-1725.
Hamma¡strom L, Granstrom M, Mollby R, oxelius v, Persson MA,q\ Smith CIE. 19g5a. ontogeny ofIgG2-antibodies against s. øtreus teichoic acid in n rrmal ¿¡d immunsdeficient child¡en. Acta paediatr.Sc¿nd. 74:12Á-730.
f{emm¿¡5[¡'sm L, Granstrom M, Oxelius V, Persson MAA. 1984b. IgG subclass distribution of antibodies¿gainsf s' ouretts teichoic acid and alpha-toxin in normal and iÃmunodeficient donors. Clin. Exp.Imms¡sl. 55:593-601.
Pelmblad J, Persson MA., snith cI. 19g4a. Lack of IgG in a healthy adr¡lt: abrlinemia with undetectable serum rñ, rgL2 ana-Igc. clin. Tmmunol.
Hamm¿¡'5¡'em I Inset RA¡ Persson MAAv smith cIE. 1.988. The IgGl subclass preference of selectednaturally occurring antipolysaccharide antibodies. Monogr. Allogy. 23:Lg-26.
Hamm¿¡56em L, Persson MA,\ smith cIE. 1984c. Subclass distribution of human aiúr-staphylococcusauteus alpha toxin antibodies : suggestion of IgG1, rg1rr,rñ4 switch pattern. Scand. J. immunol. ?Ã:?/,7-250.
Hamm¿¡s66m L, Persson MA,\ Smith CIE. 1985b. Imm¡¡6g16þnlin 5gþdass distribution of hnman anti_carbohydrate antibodies: aberrant pattern in IgA-deficient donõrs. Immunol. 54:K2l-g?Á.
Hamm¿¡5¡6m L, smith CIE. 1983a. HI-A-,\ B, c and DR antþens in IgA-deficiency. Tissue antigens.2l:75-79.
Flamm¡'566m L, Smith CIE. 1983b. rgc,2 deficiency in a healthy blood donor. concommitant lack ofrgo2, Ig^ and IgG immunoglobr¡lins and specific anti-carbohydrate antibodies. Clin. E>rp. tmmunol.5:600-604.
IJ¡nson I-A, Bjorkander J, Carlsson B, Roberton D, Soderstrom T. 1988. The heterogeneity of IgAdeficiency. J. Clin. Tmmunol. 8:159-L62.
Harriman GR, Kunimoto DY, Elliott JF, Paetkau V, Strober W. 1988. The role of II-5 in IgA B celldifferentiatio¡. J. Imm¡nol. t,t0:3033-3039.
"nry"t C, Longbotto- +. 1?36. Development of a sandwich ELISA to detect IgG and IgG subclass
antibodies for a major antþen (Ag7) of Aspàgtttus lumeryøns. clin. Allergy. r6:3?j.-í30.
Heier HE. 1980. splenectomy and serious infections. scand. J. Haematol .?A:5-j2.
Heiner DC, Lee SI, Kim IÇ Schoettler J. 1988. Replacement therapy in lgc+deficient patients. Monogr.Allerg5r. 73:L87-193.
Heiner DC, I-ee SI, Kim IÇ Schoettler J. 1988. Replacement therapy in IgG4-deficient patients. Monogr.Allergy. ?3:L87-LV3.
Heiner DC, Myers AS, Beck CS. 1983. Deficiency of IgG4 : a disorder associated with frequent i¡fectionsand bronchiectasis which may be familial. clin. Rev. a[ãrgy. \:259-?ß6.
Heiner DC, Sang tr-, Short JA. 1986. IgG4 deficiency slndromes. Monogr. Allergy. 20:149-156.
Heiner DC. 1984. Significance sf immunsglobr¡lin G subclasses. Am. J. Med. 76:L_6.
Heiner DC. 1986- IgG subclass deficiencies : identi$ing and treating patients at risk. Vox Sang.5l(Suppl. 2):57-62.
Herrod HG, Gross S, Inssl p. 1989. Selective antibody deficiency to Haemophitus ínfluenue type bcapsular polysaccharide vaccination in child¡'en with recurrent respiratory tract infe-ction. J. Clin. Tmmumol.9:429-434.

295
Hetherington SV. 1989. Antibody to outer membrane p.o¡"¡" is the dominant opsonic antibody in normalh'man serum agains¡fL influenzae type b. Immunology. eirïT_ù.
Hiemstra I, Vossen JM, van der Meer JwM, weemaes cMR, out TA" zngersBJM. 19g9. clinical andimmnnological studies in patients with an increased serum IgD level. J. Clin. Immunol. 9:393-4N.
Hodgson FUF, Jewell DP. 1977. Selective IgA deficiency and Crohn,s dissase : report of 2 cases. Gut.18-64+646.
Horan M,\ I-eahy BC, Fox RA, Stretton TB, Haeney M. 19g4. I--unologic¿l abnormalities in patientswith ch¡onic bronchial suppuration: a possible relationship with endotoxaemià. sr. t. Dt". Chest. 7g:66-74.
Hosea SW, Buruch CG, Brown EI, Berg RA" Frank MM. 19g1. Impaired immune response ofsplenectomized patients to polyvalent pneumococcal vaccine. Lancet. I:g04-g07.
Hozumi N, Tonegawa S. 196. Evidence for lorltic re¿urângeme1gf inmunoglobulin genes corrìng forvariable and constant regions. proc. Natl. Acad. Sci. uSA. 73:ã62g-%32.
Hussain R, ottesen EA. 1986. IgG responses in human fila¡iasis. J. Immu¡ol. 136:1g59-1g63.
Hussain È, Poiode*ter RW, wista¡ R, Reimer cB. 1986. use of monoclonal aniibodies to quantifysubclasses of huma¡ IgG. 1. Development of two-sile immunoen4momet¡ic assays for total IgG subclassdeterminatioas. Imm¡¡¡sl. Methods. g¡,Sg-ø.
Tnsel p'{, Anderson PW. L986. Response to oligosaccharide-protein conjugate vaccine ¿gsinstHøemophilus influenzae b in rwo patients with IgG2 deñciency, unresponsive to capsular poþaccharidevaccine. N. Engl. J. Med. 315:49-503.
Iskander R, Das PII Aalberse RC. 1981. IgG4 antibodies in EgSrptian patients with schis¡ssomiasis. Int.A¡chs. Allergy. Appl. Immun ol. 66.?ß-?nj.
Janoff E'N, Douglas JM Jr, Gabriel M, Blaser MI, Davidson AJ, cohn DL, Judson FN. 1ggg. classspecific antibody response to pne rmococcal capsular polysaccha¡ides in men infected with humanimmunodeficiency virus type I. J. Infect. Ois. f5g:g-gf_990.
Jefferis R, Reimer c, skvaril F, de Lange G. 19gg. wHo/ruIS progrâmme for standa¡dization ofimmunological reagents: availability of mãuse monoclonal uotibodi"* (nAb) to h,man IgG sublclasses.Immunology. 63:17I.
Jefferis R, Rei-mer cB, skvaril F et al. 1985. Evaluation of monoclonal antibodies having speci.ñcity forh 'man IgG subclasses : results of IUIS/T9HO collaborative study. tmmunol l_ett.l0:Z?S_252.
Jelinek DF, Lipsþ PE. 1983' The role of B cell proliferation in the generation of imm¡¡sglobrllin-secreti.g cells in man. J. Immunol. 130:2597-?Á04.
Jelinek DF, Lipsþ PE. 1987. Regulation of hlm¿¡ B \mphocyte activation, proliferation anddifferentiation. Adv. Immunol. 4O:L-59.
Johnss¡ PM, Brown PJ. 1981. Review article: Fc-gam¡¿ receptors in the human placenta. placenta.2:355-70.
Johnson PM, Matre R. 1979. Membrane receptors for IgG in the human placenta. In: Flennings WA. ed.Protein transmission through living membranes. Amsterdam: Elsevier/ñ"r,h-I{"il-ã. Bio. MedicalPress. pp.45-54.
Jones EG, Brown WR. 1974. Serum and intestinal fluid immunoglobllin in patients with giardiasis. Am.J. Dig. Dß. L9:79L-796.
Julkunen I, Hovi T, Seppala I, Makela o. 19q5. Immunoglobulin G subclass antibody responses ininflssnz¿ A and parainfl¡e¡2 type I infections. Clin. Ìxp. ImmLol. 60:130_133.-- --'

296
Kamani \ Krilov L- L987. AIDS and the spectru,n sf þrman immunodeficienry virus infection in chitd¡.en.Pediatric Rev. Commun. 1:101-121.
{^ùu WP, Hilgers R, Skvaril F. 1982. II ,moral imms¡e responses in Epstein-Barr vi¡us infections. I.elevated serum concent¡ations of the IgGl subclass in infectous mononucleosis and nasophaqmgealcarcinoma. Clin. EÃ". Immunol. 49:149-756.
Katz DH, Mead M. 1983- Cont¡ol of antibody slnthesis. In: Allerry, Principles and practice. MiddletonE Jr, Reed cE and Ellis ER eds. cV Mosby, st. Luis, Toronto. pp. 31-41.
-
Kawanishi H, Saltzrnan LE, Strober w. 19S?l -Mecha'isms regulating IgA class-spscifiç immutroglobntin
production in murine_gut-.associated lymphoid tissues. I. T celf, deriv"ed?om r"yã.'rl"t"nes that switchsIgM B cells to s IgA B cells in vitro. J.E4p. Med. 157:433_450.
Kawanishi H, Saltzman LE, Strober W. 1983b. Mecha'isas relatirg IgA class-specific immunoglobutinproduction in murine_gut-associated lymphoid tissues. tr - TerminJ differeutiatiàn of postswit"î .ig6-bearing Peyer's patch B cells. J. Exp. Med. 15g:649ó69.
Kay RA" wood KI, Bernstein RM, Holt PJL, Pumphrey RSH. 1988. Ao IgG subclass imbalance inconnective tissue disease. Ann. f,fisum. Dis. 47:53ó- S+t.
Kayhry H, Makela o, Eskola J, saarinen L, seppala I. 1988. Isotype distribution and bactericidal activity ofantibodies after immunization with Haemopiitus influenzae ryeä¡ vaccines at rg-?tt-ooths of age. J.Infect. Dis. 158:973-982.
Keikkila M, Korstinen J, Lohman M, Koskimies S. 1984. Increased frequency of HLA A1 and Bg inassociation with total lack, but not deficiency of serum IgA. Tissue antþens. 23,2ÄO-zSz.
Keller MA¡ Gendreau-Reid L, Heiner DC, Rodrþes A, Short JA. 19gg. IgG4 in h,ma¡ colostrum andh 'man milk continued local production or selecûvã bansport from serum. Acta paediatr. scand. 77:?A-29.
Keller MA, Heiner Dc, Kidd RM, Myers AS. 19g3. Locar production of IgG4 in h,man corost¡um. J.Tmmunol. 13¡0:165Ç1657.
Kemeny DM, Lessof MH, Patel S, Youlten LIF, Williams d Lambourn E. 19g9a. IgG and IgG antibodies¿jfts¡ immutrotherapy with bee and wasp venom. Int. Arch. ÁU".g5r. Appl. rmmun ol.&:?A7_Utt9
Kemeny DM, Urbanek R, Ewan P, McHugh S, Richards D, patel S, læssof MH. 19g9b. The subclass ofI-cG anjibody in allergic rtisease: tr the IgG subclass of antibodies produced following natural exposure todust mite and grass pollen in atopic and non-atopis infividuals. Clin. E4p. Immunol. 1õ:545_549.
Khater M, Macai J, G91lea C, Kaplin J. 1986. Natural kille¡ sell regulation of age-related and tlpe-specific variations in antibody responses to pneumococcal polysaccha¡ides] J. Erp. tr¡ef. ro¿:uo5-1515.
Kilian M,. I,vfestecþ J, Russell MW. 1988. Defense mechenisms involving Fc-dependent functions ofimmunsglsþrrlin A and thei¡ subversion bybacterial immunoglobulin A proteises. MiLobiol. Rev. 52:296-303.
Kimata H, Shanahan F, Brogan M, Targan S, Saxon A. Lg87. Modulation of ongoing humanimmunsgleþulin s)mthesis by natural killer cells. Cell. Immunol. 107:7¿f-gg.
Kimata H, Sherr EH, Særon A. 1988. Hl-- natural killer (NK) cells produce a late-acting B-celldifferentiation activity. J. Clin. rmmunsl. g:3g1-3g9.
Kir.off GIÇ Hodgen AN, Drew Pd Jemieson GG. 1985. Lack of effect of splenic regrowth on reducedantibody responses to pne'mococcal polysaccharides in splenectomized patiäots- Clt. n,p. Tmms¡sl.62:48-56.
Ki¡sch IR, Morton CC, Nakahara IÇ Leder P. Lg82. Hu-an immunoglobrlin-þe¿yy-chain genes map to aregion of t¡anslocation in malig"ant B lymphocytes. Science. 216:301_303.

297
Kjellman NM, Johansson SGo, Roth A. 1976. _Serum IgG levels in healthy children quantified by asandwich ¡echnique (PRIST). Clin. Allerg5r. 6:51-59.
Klein F, Skvaril F, Vermeeren R, Vlug A Duimel 1ry¡. 19g5. The quantification of IgG subclasses inreference preparations. Clin. Qhim. [s¡¿ l5}:l1r9-127
Klemola T, Seppala I, Savilahti G. 1988. gs¡rm IgG subclass levels in paediatric clinis p¿tisats withva¡iable degrees of IgA deficiency. J. Clin. Lab.Immuno. ?5:29_M.
Korngold L. t96L. Abnormal plasma components and thei¡ significance in disease. Ann. NIy Acad. Sci.94:110-130.
Korsmeyer SJ, Waldmann TA. 1982- Tmmunoglobrlins II. Gene organisation and assembly. In Basic andClinical Immunslsgy. eds. stites DP, Stobo JD, Fundenberg HH and wells JV, eds. Lange, california.pp.43-51.
Kumkum, I(hann¿þ R, Nain cIÇ Mehta S, vinayak vIC 19gg. Depressed humoral immune responses tosurface antþens of Gia¡dia lamblia in persistent giardiasis. Pediatr. Infect. Dis. J. 7:4V249g.
5l*:t Y9,_f"þ:yrt-, Franklin EC, osserman EF, Terry wD. 1966. Notation for human IgG subclasses.Bulletin WHO. 35 (6):81L-982.
Kunkel HG, Joslin FC, Penn GM and Natvþ JB. 1970. Genetic variants of gamm¿-64 glob,¡lin. 4 rrniquerelationship to other classes of gamm¿-Ç globulin. J. E)rp. Med. Ii2:50g_520.
Lachman PJ, Hobart MJ. 1978. Compleme_nt technology. In: Handbook of Experimental Tmmunology.3rd edition. Weir DM, ed. Blackwe[ Scientific, Offord. jA:2_S.
Lai E, Wilson RK Hood LE. 1990. Physical maps of the mouse and human immunoglobrlin-lifts lsci.Adv. Immunol. ¿16:1-59.
Lancet (ed.). 1982. Battles against giardia in gut mucosa. Lancet. 2:5?jt-5?Å.
Lane PJL, Maclennan-IcM. 1986. Impaired IgG2 anti-pne,mococcal antibody response in patients withrecurrent i¡fection and normal IgG2levels but no IgA. Clin. Exp. Tmm¡¡ ol. 65:427_433.
La¡sson A Perlmann r, N3{ig {n . rn3. C}totoxicity of human lym.phocytes induced by rabbit antibodies
to chicken erythrocytes. Inhibition by normal IgG and by human -yólo-u proteins of different IgGsubclasses. Tmmunology. 25:675-86.
Laschinge C, Gauthier D, Valet JP, Naylot DH. tgtø. þlustu¿ring levels of IgA in individuals withselective IgA deficiency. Vox Sang. 47:60-67.
Law¡ence EM, Edwards KM, Schiffman G, Thompson JM, Vaughn WK Wright pF. 19g3. pneumococcalvaccine in normal child¡en: primary and secondaryvaccinaúon. a-. ¡. Dis. Child. 1i!7:g46-g50.
Leddy JT, Deitch-an J, Bakemeir RF. L970. IgG subclasses: measurement by radioim-unoassay innormal and hypogamm aglobulinemic sera. Arth¡itis Rheum. 13:33L-332.
Leder P. 1983. Genetics of inmunoglob.rin production. In: The Biology of rmmunologic Disease. FJDixon and DW Fisher, eds. Sinaner. Sunderland, Mals. pp.3-12.
Leen CLS, Yap Pl McClelland DBL. 1985. Increase of serum immunoglob,¡lin level into the normalIu"g" i" primary hlpogammaglobrrlinaemia by dosage individualisation oiintrave¡sus immunsglobulin.Vox Sang. 5I:T78-?36.
Lefranc G, Chaabani Fr, von Loghem E, Lefranc MP, delange G, Helal AN: 1983a. Simr¡ltaneous absenceof human IeGl; IgG¿ IgG4 and IgAl subclasses: i--unãlogical and inmunã;"";d.ul considerations.Eu¡. J. rmmunol. 1i:Z4O-244.

Lefranc G, D lmitresco S-M, Salier J-P, Rivat L, delange G, van Inghem E, Loiselet J. tng. "J;ålack of the IgG3 subclass: gene elimin¿tion or tu¡ni¡ oF expression and neutral evolution io n" i-*
system. J. Immunog enet. 6:221-?5I.
LeFranc M-P, l,eFranc G, delange G. et aI. 1Lg!?b. rnstability of human heavy chain constant region locusmediated by different ch¡omosomal deletions. Mol. 3iol. ueã,.t:zm-zn.
l'ever AML, webster ADB, Brown D, Thomas HC. 1984. _ Non-A non-B hepatitis occurriag inagammaglob 'linaemic patients after int¡avenous runoglobr¡lin. Lancet Z:1062-L064',
Linde A, Sundqvist V-4" Mathiesen T, Wahren B. 1988. IgG subclasses to subviral components. Mouogr.Allergy. 23:27-32.
Linde AG. 1985. Subclass distribution of rubella virus specific ìmmunoglobrrlin G. J. Clin. Microbiol.2L:ll7-121.
Linde C. IgB7b. Immunoglobrlin class and immunoglob,lin G subclasse\TW ays comp¿ued with microneut¡:alization assays fór serodiaposis ofm'mp of imm¡¡¡¡y. J. Clin. Microbiol.25:1653_165g.
Linde GA' Hamma¡strom L, Persson MAA".Smith Cftr, Sundqvist VA5 wahren B. 19g3. Virus-specificantibody,activity of different subclasses of immunoglobulins ò and A and cytomeg;ovirus infections.Infect. ImmunôI. 42.?37 -?A4.
Litwin SD, Balaban S. 1972. A quantitative method for determining hnman gamm¿_Ç allotype antþens(Gm) tI - differences in Gm gene expression for gamm¿-Çl and gam¡1¿-G3 H chains in sera. J. Immunol.108:991-999.
Lowe J, Bird P, Hardie D, Jefferis R, Liog NR. 1982. Monoclonal antibodies (McAbs) to determinants onh'man gamma chains; properties of antibodies showing subclass rest¡ictìon or 'subclass specifici$.rm m unology . 47 :329 -336.
LowyI, ThezeJ. 1985.- Regulation of IgGl and IgG2 subclass expression by adjuvant-activated splenicsuppressor T cells. Cell Immunol. 9I:467-476.
Lundgren M, Persson [J, Larsson P, Mapusson C, Smith CIG, Hammarstrom L, Severiruon E. 19g9.Interleukin 4 induces synthesis of IgE and IgG4 in hnman B cells. Eu¡. J. Imm¡¡61. 19:1311_1315.
Magnusson CGM, Cesb¡o_n JY, Djurup R, Capron A, Johansssn SGO. j-9gó. Raised IgG4 levels inpatients with atopy and filariasis: Application of an automated particle-countiog i--uooassay usingmonoclonal antibody. Int. Arch. AllergyAppl. Tmmunol. gt:23g-?-M.
Magnusson CGM, Delacroix DL, Vaerman JP, Masson PL. 1984. Ty?-g of subclasses and light chains ofhuman monoclonal immumoglob 'lins by particle co¡nring ;--uooassày (neCn¡. J. Immunol. Methods.69:?29-?At.
pala Isotype concent¡ations of humantlpe yorhg adults immrnised with thea pr Tmmunol. 139:I999_?fr4.
Malkovsþ M, Jira M, Gao L, Loveland B, Malkovska V, Dalgtsish AG, Webster ADB. 19g6. Reducedexpression 6f i¡¡s¡lsrrkin-2 receptors in hypogammaglobulinaeÃia. A possible calrse of a higher incidenceof c¿ncer. Lancet. I:L442-I443.
Matheson D, Green BJ. L987. Defect in production of B cell differentiation factor-like activity bymononuclea¡ cells from a boywith hlpogammaglob linemia. J. rmmunol. 1ß:2Æ9_ùl:72
Matleson DS, Green BJ. 1987- Defect in production of B cell differentiation factor-like activity bymononuclea¡ cells from a boy with hypogammaglob inaemia. J. Immunol. ljg:2469_?/l:72.
Mattila PS, Pelkonen J, Makela O. 1986. Pro_portions 9f IgG subclass antibodies formed duringvaccination with hepatitis B surface antigen. Scend. J. Tmrnunol. zi.øsg-lC/.-

299Mayer L, Fu SM, Cunningham-Rundles C, Kunkel HG. Lg8y'.. Poþclonal immunoglobulin secretion inpatients with common variable immunodeficiency using monodonal B cell differentiation factors. J. Clin.Invest. 74:2L75-212A.
Mayer MM. 1961". Complement and complement fixation. In: E¡perimental immunochemistry. Kabat EAand Mayer MM eds. Charles C. Thomas, Springfield Ill.pp. 133-?Æ.
Mayumi M, Kuritani T, Kubagawa H, Cooper MD. 1983. IgG subclass e4pression by human Blymphocytes and plasma cells: B lymphocytes precommitted to IgG subclass c¿n be preferentially inducedby polyclonal mitogens with T cell help. J. Immunol. 1jO:67L-677.
Mayus J, Macke IÇ Shackelford P, Kin J, Nahm M. 1986. Human IgG subclass assays using a novel assaymethod. J. Tmmunol. Methods. 88:65-73.
McBride OW, Hieter PhA, Hollis GF, Swan D, Oety MC, I-eder P. ]1982. Ch¡omosomal location ofhuman kappa and lambda imm¡¡sglsbuli" light chain constant genes. J. Exp. Med. 155:i.¿180-1490.
McKearn JP, Paslay JP, Slack JH, Ba'm C, Davie JM. 1982. B cell subsets and differential responses tomitogens. Tmmunol. Rev. Ø:5-23.
McNabb T, Koh TY, Dorrington KI, Painter RH. 1976. Structu¡e and function ¡f imm¡¡sglsþ'lindomains. 'Binding sf immunsglsþrlin G and fragment to placental membrane preparations. J. rmmunol.117:882-888.
Mellbye OJ, Natvþ IB. L973. Presence and oriþ of human IgG subclass proteins in nevòorns. Vox Sang.?A:?16-215.
Merrill MW, Naegel GP, Olchowski JJ, Reynolds tfY. 1985. Tmmutr6gl6þulin Ç subclass proteins inserum and lavage fluid of normal subjects, quantitation and comparison with immunoglob 'lins A and E.Am. Rev. Respir. Dis. 131:584-587.
Metlay JP, Pure E, Steinman RM. 1989. Cont¡ol of the immune respoû¡e at the level of antþen-presenting cells: a comparison of the function of dendritic cells and B lymphocytes . Adv. rmmunol. 47:45-716.
Mond JJ, Farra¡ J, Paul WE, Fuller-Farrar J, Schaeffer M, Howard M. 1983. T cell dependence and factorreconstitution of ln vlrro antibody responses to TNP-B. Abortus and TNP-FicolL restoration of depletedresporuies with ch¡omatographed fractions of a T cell-derived factor. J. Tmmunol. ljt:633-637.
Morell A, Roth-Wicky B, Skvaril F. 1983. Tmmunoglobulin G subclass restriction of antibodies againsthepatitis B su¡face antþen. Infect. Tmmun.39:565-568.
Morell A, Skvaril F, Hit"ig WH and Ba¡andun S. Lnzþ). IgG subclasses: development of serumconcentration in "normal" infants and children. J. Pediat¡. 80:960-964.
Morell A' Skvaril F, Rade J, Dooran IJ, Barandun S. Lns. IgG subclass abnormalities in primaryimnunodeficiency diseases. In: Immunodeficiency in Man & Animals. Birth Defects: Original-ArticleSeries. Vol. )fl. No.1. Bergsma D, ed. Sunderland" Mass. Sinauer Associates Inc. pp. 108-120.
Morell A5 Skvadl F, Rufener J. L973. Cha¡acterization of Rh antibodies formed after incompatiblepregnancies and after repeated booster injections. Vox Sang. ?A:3?3-3j0.
Morell A' Skvaril F, Steinberg AG, van Loghem E, Terry WD. tn2@). Correlation betweon theconcentrations of the fou¡ subclasses of IgG and Gm allotlpes in normal human sera. J. rmm¡¡et. 108:195-2M.
Morell A' Skvaril 4 van Loghem E, Kleemola M. LnL. H'man IgG subclasses in maternal and fetalserum. Vox Snng. 2L:481-4T2.
Morell4 Terry WD, Waldmann TA. L970. Metabolic properties of IgG subclasses in man. J. Clin.Invest. 49.673-û0.

300Morell A. L986. Various immr¡nsglqþulin preparations for intravenous use. Vox Sang. 5L(suppl. 2):44.49.
ky RI. 1986. Heterogeneity of clinical slmdromesImmunodeficiency Research and Therapy tr. J.
ers. B.V. Biomedical Division. pp.?29-?-33.
Mu.phy MB, Reen DR, FitZgerald ND( 1984. Atop¡ immutrsl¡gical changes and respiratory function inbronchiectasis. Thorax 39:L79 -184.
Nadal D, Ilrlnziker UAo Schupbach J, Wetzel J-C, Tomasik Z, Jendis JB, Fanconi d Seger RA. 19g9.Imm¿¡s1.gical evaluation in the early diag"osis of prenatal or perinatal HIV infection. A¡ch. Dis. Child.64:662-669.
Nahm MH, Blaese RM, Crain Mf, Briles DE. 1986. Patients with Wiskott-Aldrich syndrome have normalIgG2 levels. J. Tmmunol. 1i7:348L3487.
Nair KPV, Sillen J, Fergusou A. l-981. Corticosteroid treatment increases parasite numbers in murinegiardiasis. Gttt. 22:47 5-480.
hita N, Miyanoto T, Skvaril F. L997. Changes ofantibodies during immunotherapy in patients with
82:95-99.
Nelson RD, Quie PG, Simmsns RL. 1975. Chemotaxis under agarose: a new and simple method fórmeasuring chemotaxis and spontaneous -ipration of human polymorphonuclear leukocytes
--d -ooo"yt"..J. Tmmunol. 115:1-650-1-656.
Nevo S. L974. Association of $'phoid fever and responso to vaccination with polymorphic systems in man.In: Protides Biological Fluids, Proc.)2tdColloquium, Brugge p649. Ordord. pèrgamoi press.
Normansell DE. 1987. Hu'nan immunsgl¡þnlin subclasses. Diagn. Clin. Immunol. 5:115_1ã1.
Nys M, Joassin L, Somzee d Demonty J. 1988. f,as¡rns-linked immunosorbent assay for immr¡nsglsþnlinG subclass antibobodies specific for enterobacterial re core glycolipid in healthy individuals and in latieutsinfected by gram-negative bacteria. J. Clin. Microbiol. ?Á:857-862.
Qchs HD, Fischer SH, Pyun KH, Wedgwood RJ, Morell A. 1989. Antþen-specific IgG subclassproduction and metabolism. In IgG subclass deficiencies. RI Levinsky, ed. International Cãngress andSlmposirrm Series. No. 143. Royal Society of Medicine Services Ltd. London . pp./l-35.
RI. 198ó. Survival of IgG subclasses followingts with primary immunodeficiency diseases. In:
ulins, Morell A. and Nydegger UE, eds. Interlaken,Acadenic Press, London, pp. 77 -85.
Oen IÇ Petty RE and Scb¡oeder Nn-. L982. IgA deñciengn genetic studies. Tissue Antþens. L9:L7stg2.
Ogra Pl Ka¡zon DT, Righthand F, MacGillway M. 1968. rmm¡¡sglqþrlin response in serum andsecretion after immunization with live and inactivated polio vaccine and nãtrual infectiãn. N. Engl. J. Med.219:8r3-9ffi.
Oxclius V-4, Berkel AI, Hanson I-4. L982. IgG2 deficiency in ataxia-telangiectasia. N. Engl. J. Med.300:515-517.
Oxelius V-Ao Hanson LAn Bjorkandor J, Hammarstrom ! SjohoLn A. 1986. IgG3 deficieng¡ common inobstructive lung disease. Monogr. Allergy. ?n:L06-L15.
Oxelius V-d Laurell AB, Lindquist B, Golebiowska H, Axelsson U, Bjorkander J, Hanson Lq.. 1931. IgGsubclasses in selective IgA deficiency. Importance of.Igp2-lgA deficieãcy. N.Eogl.J. Med. lM:L476-t477.

301Oxelius V'é^. 1974. Chronic infections in a family with hereditary deficiency of IgG2 and IgG4. Clio. E*p.Immun¡1. 1!7:19-2i1.
Oxelius V-4. 1978. Crossed imrnunsslectrophoresis and electroimmunoassay of hrrm¿¡ IgG subclasses.Acta Pathol. Microbiol. Scand., C.Irnmunol. 36:10g-1[].
Oxelius Y-A. 1979a. IgG subclass levels in iof-.y and childhood. Acta paediat¡. Scand. 6g:?3-2l.
oxelius V-A- Lngb' Quantitative and qualitative investigations of serum IgG subclasses inimmunodeficiency diseasq . Clin. Ex¡r. Imm¡¡ ol. %:L12-116.
Oxelius V-4. 1984. Immunoglobrlin G QgG) subclasses and hu'nan ¡tisease. Amer. J. Med. 76:7-lg.
Oxelius V-4. 1987. Antþenic va¡iants of human IgG subclasses. Restriction of murine monoclonal [gGsubclass antisera. Scand. J. Tmmunol. ?5:169-t74.
Oxelius V-4. 1988. Restrictive murine monoclonal IgG subclass antisera rsye,eling antigenic va¡iants ofhuman IgG subdasses. Monogr. Allergy. ?3:57-60.
Oxelius V-A. 1990- Gm allotlpes and gene dosage ¿ffsçting both IgG subclass and IgE levels in atopicpatients. Int A¡ch AllergyAppl. Immunol. 91:58-61
Painter RH, Law DTS' 1985. Structural and biotoglcal properties of tl¡ee int¡avenous immr¡nqglgþrlinpreparations. In: Int¡aveuous immunsglqþnlins in im'nunodeficiency syndrome and idíopathicthrombocytopenic purpura. Waters AH and Webster ADB, eds. Royal Sociery ãr Iraeaicine, InternitionalCongress and Symposium Series. pp.11-18.
PandeyJP, Fudenberg HH, Virella G, Kyong CU, l,oadholt CB, Galbraith RM, Gotschlich EC, pa¡ke JC.Jr. lng- Association between immunsgl6þ.lin ¿llsb¡pss and immune resporuies to Høemophilusinfluenzne -¿ neningococ,cus polysaccharides. Lancet L. IgO-Iy2.
Pandey JP, Fudenberg HH. 1984. Perspectives on the genetics of the Gm system and its role in immuneresponse and diseases. Clinics in Immr¡nol. and Allergy. aQ):6al-6a5.
Papadea C, Check U, Reimer CB. 1985. Monoclonal-antibody based solid-phase immunoenz)mometricassays for quantifying h 'man immunoglobulin G and its subclasses in serum. Ctio. Cn"-. 3I:I940-1M5.
Parish WE' 1-975. Some biological activities of IgG subclass antibodies after inocula[ion and in disease. In:Allergy, Ganderton MA and Frankl¿¡fl AW, eds. Pitman Med. Publ. Co. Ti¡nbridge Wells, p. 153.
Partridge I Jefferis R, Hardie D, Liog NR, Richardson P. 1984. Subclasses of IgG on tÏe surface ofhu'nan lymphocytes: a study with monoclonal antibodies. Clin. Exp. Immr¡nol.56:I67-L74.
Paton JC, Toogood IR, Cockington RAr flansmm D. 1986. Antibody response to pneu.nococcal vaccinein children aged 5 to 15 years. Am. J. Dis. Child.t40:f|5-13g.
Pedersen FK Hen¡ichsen J, Schiffman G. L982. Antibody response to vaccination with pneu'nococcalcapsular polysaccharides in spleneç¡smissd child¡en. Acta Paediatr. Scand. 7l:45I-455.
Peltola H, Kayhty H, Sivonend Makela H. Lgn. Hacmophihts influerunc tlpe b capsular poþsaccharidevaccine in child¡en: a double blind ñeld study of 100,000 vãccinees-3 -ootb" io 5 years of a-ge-in Finland.Pediatrics. 60:730 -737 .
Perri RT, Weisdorf DJ. 1985. Impaired responsiveness to B cell growth factor in a patient witl commonvariable hypogam m aglobr¡linaemia. Blood. 6:345-34¡9.
Persson MAA! Ekwall E, Hammarstrom L, LindbergAd Smith CIE. 19g6. Tmmunoglobutin G (IgG) andfgA sybclass pattero of human antibodies to Shigettø ftæteri and Salmonella sãrogroup È-and Dlipopolysaccharide O antþ . Infect. rmmun. 52:83Ç839.
Petty RE, Palmer NR, Cassidy JT et al. Lng. The association of autoimmune dise¿5ss and anti-IgAantibodies in patients with selective IgA deficiency. Clin. Exp.Immunol.3T:g3gg.

302Phelan DP, Landau LI, Olinsky A. 11982. Suppurative lung disease. In: Respiratory lllnsss in Children.Blackwell, Odord. pp. 22L-?-38.
Pincus SH, KlebanoffSJ. 1971. Quantitative leukocyte iodination. N. Engl. J. Med. ?&:74L750.
PlebaniA' Duse M, Monafo V. 1985. Recurrent infections with IgG2 deficiency. A¡ch. Dis. Child.60:670-672.
Plebani d Monafo V Avazini M,\ Ugazio AG, Burgio GR. 1986. Relationship between IgA and IgGsubclass deficiencies: a reappraisal. Monogr. Allergy. ?Ã:L7t-178.
Plebani,4" Ugazio AG,Avaz.ini MA" MassimiP,T-nnta L, Monafo V, Burgio GR. 1989. Serum IgGsubclass concentrations in healthy subjects at different age:, age normal percentile cha¡ts. Eur. J. pediat¡.I49:L64r67.
Poljak RI. 1983. rmmunsglsb'lin structure and function. In: Allergy, Principles and practice. MiddletonE, Reed CE and Elliott EE, eds. CV Moseby Toronto. pp. 19-30.
Prellner Ç Braconier JH, Sjoholm AG. 1985. Combined lgG2,lgp4 and IgA deficieucy low C1econcentrations and the presence of excess Cl-R and C1S in an adult patient with recurrent pne,mococcalinfections. Acta Pathol. Microbiol. rmmunol. Scand. (C). 93:257 -?Á3.
Purtilo DT, Grierson HL, Ochs H, Skare J. 1989. Detection of X-linked llmphoproliferative disease usingmolecular and immunovirologic markers. Am. J. Med. 87:4ZL-4V1.
Rautonen N, Pelkonen J, Sipinen S, Kayhty H, Makela O. 1986. Isot¡pe concent¡ations of h,manantibodies to group 4 neningocococcal polysaccharide. J. Immunol.3T:?Á70-2675.
Raziuddin S, Teklu B. 1989. Severe T lymphocyte immunodeficiency associated withhypogammaglobtrlinaemia: defective lymphokine secretion but enhanced autologous mixed lymphocytereaction. J. Clin. Imnunol. 9:448-453.
Read R, Spickett G, Harvey J, Edwards AJ, Larson HE. 1988. IgGl subclass deficiency in patients withchronic fatþe slmdrome. Lancet: l:VlL-?A2 (letter).
Reimer CB, Philips DJ, Aloisi CH, Moore DD, Galland GG, Wells TW, Black CM, McDougal JS. 19g4.Evaluation of thirty-one mouse monoclonal antibodies to h lman IgG epitopes. Hybridom a3:?:63-2j15.
Reimer cB, smith SJ, wells T'w, Nakamura RM, Keitges pw, Ritchie RF, williams GW, Flanso¡ þJ,Dorsey DB. 1982. Collaborative calibration of the U.S. National and the College of American Pathologistsreference preparations for specific senrm proteins. Amer. J. clin. path.77:1LLi.
Riesen WF, Skvaril F, Braun DB. Lm6. Natural infection of man with group A streptococci levels:restriction in class, subclass and tlpe and clonal appearance of polysaccharide-group specific antibodies.Scand. J. Immunol. 5:383-390.
Rivat L, Ropartz C, Burtin P, Kartizlry D. 1969. Abnormalities in the spthesis of some subclasses ofgamma-G in a family with two cases of ataxia ¡elangiectasia. Vox gang. 17:5-10.
Rivat-Peran I Buriot D, Salier JP, Rivat C, Dumitresco SM, Griscelli C. 1.981. Imm¡¡sglsþrrlins in ata¡rialelangiectasia: evidence for IgG4 aadrgnz subclass deficiencies. clin. Tmmr¡nel. T"'mr¡nopathol. 20:99-110.
Roberton DM, Coþan T, Ferrante d Jones C, Mermelsten \ Sennhauser F. 1990. IgG subclassconcentrations in absolute, partial and trnnsient IgA deficiency in childhood. pediatr. Infect. Dis. J. 9:54t-49.
Roberton DM, Georgiou GM, Tiedemann K Hosking CS, Ferrante A" Toogood I, Mcl(enzie IFC. 1986.rmmr¡nslsgical reconstitution in a patient with severe combined immunodefi"i*cy usinmg ¡6¡-5iþling bonemarrorv depleted of T cells with Hul-y-ml. Aust. NZ J. Med. 16:385-3gg.

303
Roberton DM, Hssking cs. rg87. childhood infections. In: child and Adolescent Health forPractitioners. Buchanan N, Willi¡ms & Wilkins, Sydney. pp.nS_nl.
Roberts-Thompson IC, Mitchell GF. 1978. Giardiasis in mice. I. prolonged infections l¡ ss¡tain mouses¡¡ans ând hypothymic (nude) mice. Gast¡oenterology. 75:42_46.
Rock KL, Benacerraf B, Abbas ArC 1984. Antigen presentation by hapten-specific B lymphocytes. I.Role of su¡face imm¡¡sglsþulin receptors. J. Exp. Med. too:rtoz-[rg.
Rodriguez MA., Blanco I, Baroja ML, Aram¿ S, l,eon-ponte M, Abadi I, Bianco NE. 19&7. Helper activityby human large granular lymphocytes in ¡¿ ra'tro immunoglob,,lin spthesis. J. Clin. Tmmunol. 7:35G364,
Roitt I, BrostoffJ, Male D. 1985. Immunology. Gower Medical puþlishing, London.
Romeiro SA" Takeha¡a HA" Mota I. 1984. Isotype of lytic antibodies in serum of Chagas, diss¿ss paLients.Clin. Exp. Tmmu¡ol. 55:4Íi-418.
Romer J, Morgenthaler J-J, Scherz R, Skvaril F. rg8z, Cha¡acterization of various immun6glsþrlinpreparations for intravenous application I. Protein composition and antibody content. và* suog. 42:62-73.
Ropars Ç Mull,er,Ab Paint N, Beige D, Avenard G. 1982. Large scale detection of IgA deficient donors. J.Immunol. Methods. 54: 181189.
Rowe DS, Anderson SG, Grab B. 1970. A research standa¡d for hrm¿¡ serum immunoglob.rins, IgG, Ig{and IgM. Bull. WHO. 42:535-552.
Royer HD, Reinherz\L. 1987. T' lymphocytes: ontogeny, function and relevance to cliniç¿1 disorders. N.Engl. J. I|rled,. 3L7 :II%-LL4Z.
f'ozsnyay 7-Satmay G, Walks¡ M, Maslanka I! valasek Z, Jefferis R, Gergely J. 19g9. Distinctive role ofIgGl and IgG3 isotypes in Fc gâmma-R-mediated functions. Immunol. æ:iSt_qgl.
Saiki O, Shimizu M, Saeki Y, Kishimoto S, Kishìmsto T. j.9g4. Dissociation in the production of B cell_stimnlating_ factors (BCGF and BCDF) and interleukin -2 by ^r celrs from a cornmon variabreimmunodeficient patient. J. Tmmunol. 1984. 1j3:1g?n-7g24
1fu9" JE, Edberg JC, Kimberly RP. 1990. Fc gamm¿ receptor III on h rman neutrophils. Allelic va¡iantshave functionally distinct capacities. J. clin. Invest. g5:12g7 -7295.
Sa¡nesto A" Julkunen I, Makela o. 1985b. Proportions of IgG classes and subclasses in mu-ps antibodies.Scand. J. Immunol. 22:345-350.
Sarnesto A, Ranta S, Vaananen P, Makela O. 1985a. Proportions of IgG classes and subclasses in rubellaantibodies. Scand. J. Immunol. 2I:Zl5-?Å2.
Sarvis H, Rautonen N, Sipinen S, Makela o. 1989. IgG subclasses of pne ,-ococcal antibodies - effect ofallotype of G2m(n). Scand. J.Immunol. 29:229-237.
Saxon A" Stiehm ER. 1989. The B-lymphocyte system. In: Immunologic disorders in infants and child¡sqStiehn ER, ed. Saunders, philadelphia .- pp. 4O-67.
Schanfield MS, Wells JV, Fudenberg HH. 1979. Immunoglobnlin allotypes and response to tetanus toxoidin Papua \fsv/ Çrins¿. J. Immunogènet. 6:311-315.
Scheinberg M, Blacklow NR, Goldstein AL, Parring JA" Rose FB, cathcart ES. 1926. Tnfluenza: responseof T cell lymphopenia to thlmosin. N.E"gl.J. Med. Z9a:12ng-121ri.
Schlamowiø M. L976a. Membrane receptors in the specific transfer of imm¡¡6g1sþulins from mother toyou'g. Immunol. Commun. 5:¿81-500.

304Schlamowiø M' 196b. Maternofoetal transmission of protein in the rabbiu f¡ensfs¡ in vivoand binding inviîo to the yolk sac membrane. In: Maternofoetal transmission of rmunoglob'lirrs. rt"--;'æ rW.\ ed.Cambridge. Cambridge lJniv. press, l7g_2Cfi'.
Schoettler JJ, Heiner DC' 1986. Hypercatabolism of IgG4 associated with staphylococcal infection. clin.Res. 128A:3 (absrr).
itËtriï #;"rylri serum immrneslsþulin ¿¡d transfe¡¡i¡ levets after cbldhood splenectomy. Arch.
Schur PH, Borel H, Gelford EW, Alper CA, Rosen FS. 1970. Selective gamma-G glob,rlin deficiencies inpatients witl recurrent pyogenic infeãtions. N.Engl.J. Med. zsz:slt-øy.
Schur PH, Rosen F, Norman ME. Lng. Imrn¡¡qglsþrlin subclasses in normal child¡en. pediat¡. Res.ljì:L81-183.
Schu¡ PH. lif). lltt¡n¿¡gnmm¿-Ç subclasses. prog. Clin. Imm¡¡sl. l;Jl.
Schu¡ PH. 1987. IgG subclasses - a review. Ann. Allergy. 5g:g9_100.
schur PH. 198s. IgG subclasses: a historical perspective. Monogr. Arergy. 23:r_Lr.
Schwa¡tz AD. 1989. Spleen physiology and hyposplsnisn. In: Disorders of the spleen. pochedly C, SillsRH and Schwa¡tzAD, eds. Marcel Oãi<ker, N.i. p. fSZ.
scott M'A¡ shackelford PG, Briles ME, Nehm \dp. 1988. H,man IgG subclasses and thei¡ relation tocarbohydrate antigen i m m unocormpe t ence. Dia gr¡. Cli". f --u"-J. S :?til_?Æ.
Scott MG, Nahm 1r,41¡. 1984. Mitogen-induced h,ma¡ IgG subclass e4pression. J. Immun ol. 735:?A5Ç24&.
Scott MT, Goss-s'mpson M. 1984. Restricted IgG isotype profi.les n T. cruziinfected mice and chagas,clisease patients. Cli". E*p. Immunol. 5g:372_379.
Seppala IJT, Rautonen Makela O. 19g4. The percentages of siximmunsglsþrrlin isstJps5 toxoid: standa¡'¡lization of isotype-specrfic secondantibodies in solid-phase _g75.
Shackelford PG, Granoff DM, Nahm MH, scott MG, surez B, pandey Jp, Nelson sJ. 19g5. Relation ofage' race and allotype tq immutrsglsþrlin subclass conôntratio^. p"¿iut . Re,s. 19:g46_g49.
Shackelfordand normal tibodl response of healthy adulb
Hib pS-prot type b polysaccharide vaccine or
Shackelford PG, Palmer- SH, Mayus JL, Johnson lilL, Corry JM, Nahm \d[. 19g6. Spectrum of IgG2subclass deficiency in child¡en with recurrent infections. prorí""tiu" study. J. pediatr. 10g:647-653.
shacks SJ, Heiner DC, Bahn¿ sL, Horwitz cA. 1985. Increased senrm IgG4levels in acute Epstein-Barrviral nononucleosis. Am. Allerry . 54:2g4_?ß.
?lft F, Janes K Barr D. Lggz. Gm allotlpes and IgG subclasses in Iraqi A¡abs. J. rmm¡¡sgenet. 9:149-
shakib F, Mclaughlin P, stanwortl DP, smith E, Fairburn E. 1g77. F]levated serum IgE and IgG4 inpatients with atopic dermatitis. Br. J. Dermatol.gT:59_63.
shakib F' stanworth !R, Drgw R, catty D. 1n5. A quantitative study of the dist¡ibution of IgGsubclasses in a group of normal human r"rá. J. Tmmr¡nol. Methods. g:17-2g.

305
shakib F, Stanworth DR. 1980(a). Human IgG subclasses in health and disease. part I. La Ricerca Clin.Lab.10:463-479.
Shakib F, Stanwortl DR. 19S0(b). Human IgG subclasses in health and disease. pa¡t tr. La Ricerca Clin.Lab.10:561-580.
Shelton N[I, Meek F, Geller I, Hoskins CS. 194. Serum immunoglobrrlin lsyeþ i¡ child¡.s¡. Aust. J. Med.Technol. 5:113-1-17
Shinomiya N, Kuratsuji T, Yata J. 1989. The role of T cells in immunoglqþrrlin class switchi'g of specificantibody production system in vit¡o in hnm¡ns. esll Tmmum oL TL}:239-?A9.
Siber GR, Schu¡ PH, Aisenberg AC, Weitzman SA, Schiffman G. 19g0. Correlation between serum IgG2concentrations and the antibody response to bacterial capsular polysaccharide antigens. N. Eogl,J. Àñed.303:778-182.
silk HJ, Geha RS. 1987. Inrravenous qammaglobulin (rvGG) prophylaxis in chird¡.en with IgG2deficiency. J. Allergy Çli¡. Tmm¡mol. 79:188 (abstract).
Simmons JG, Fuller CR, Buchanan PD, Yount WJ. 1981. Distribution of surface, cytoplasmic andsecreted IgG subclasses in h'man lymphoblastoid cell lines and normal periperhal tiooO fympno"yilSc¡nd. J. Immunol. L4:L-13.
Singb Vijendra IC 1988- Quantitation of age-related levels of IgG subclasses using serotec RID plates.Diagn. Clin. Immun ol. 5;414-4L6.
Skvaril F, Ctyz SJ, Furer F,. 1985b. IgA and IgG subclass antibodies i¡ i¡fiyidu¡ls vaccinated withKlebsiella capsular polysaccharide vaccine (abstract). Symp. on vaccines and vaccinations. InstitutPasteur, Paris.
skvaril F, Gardi A. 1988. Differences among available immunsgleþrlin preparations for intravenous use.Pediat¡. Infect. Dis. J. 7 :543-548.
Skvaril F, Joller-Jemelka H. 1984. IgG subclasses of anti-HBs antibod.ies in vaccinated and non-vaccinatedhdividuals and in anti-HBs immunoglobulin preparations. Int. Arch. AllergyAppL r--[ o1.73:330-337.
Skvaril F, Morell A. 190. The G4 subclass in IgG fractions prepared by ion-exchange chromatography. J.[m m unol. IM:I3L} - 1312.
lku-il F, Roth-Wicþ B, Barandun S. 1980. IgG subclasses in hrman gamm¿-gleþulin preparations forint¡avenous use and thei¡ reactivity with staphylococcus protein A. Vox Sang. 3S:ia7-6S.
Skvaril F, Schilt U. Lg8r''. Cha¡acterization of the subclasses and light chain types of IgG antibodies torubella. Cli". E.p. Immunsl. 55:67l-676.
Sl*Tl F, Stanley JC, Furer E. 1985a. Human immunoglob¡rlin G subclass distribution in response tovaccination tvt|hl4rbio cholerae vaccine and procholer"geooid. Current Microbiology .12:747-150.
Skvaril F' 1983. Human IgG subclasses in antiviral antibodies determined with monoclonal antibodies inELISA. In: Awomeas S, Druet P., Masseyeff R. and Feldman G, eds. Immunoenz)matic techniques, Vol.18, Elsevier Science puþlishing Inc., Amsterdam. pp. ?ß7-290.
Skvaril F. 1986. Immunoglobrrlin Ç subclasses. Methods of enzymatic analysis. Vol. 10. 60-71 (Vat,Verlagsgesellschaft, Weinheinn, 1986).
Sloyer JL Jr, Ploussa¡d JlI, Howie VM. 198L. Efficacy of pne,mesoccal polysaccharide vaccine inpreventing otitis media in infants in Huntsville, Alabama. Róv. Infìct. Dis. 3(suppì.¡:'Súl-rza.
lryth PD, Gillan FD, Brown WR, Nash TE. 1981. IgG antibody to Gia¡dia lamblia detected by enz5melinked immunosorbent assay. Gastroenterology. g0:1476-14{10.

Smith TF, Morris MD, Bain [P,slmptoms. J.Pediatr. 105:896-900.
306
Lgu. IgG subclasses in nonallergic chirdren with ch¡onic chest
Snapper CM, Finkle'man FD, Stefany D, Conrad DH, Paul WE. 1988. II-4 induces co-expression ofintrinsic membrane IgGl and rgE murine B cells stimulated with lipopolysaccharide. J. rmmunol. 74L:4g9-496.
Soderst¡om T' Soderstrom R, Anderson R, Lindberg J, Hanson LA. 1988. Factors influenc,ng IgGsubclass levels in serum and mucosal secretions. Monogr. Allergy. g:86-?-43.
soderstrom T, soderstrom R, Avanzini Ar Brandtzaeg p, Karlsson G, flanssa I¿.. 19g7. ImmunoglobrrinG subclass deficiencies. Int. Arch. AllergyAppl. Imnunol. g2:4764g0.
Sotomayer Jl Douglas SD, Wilmott RW. 1989. Pulmonary manifestations of immune deficiency rlisease.Pediat¡. Pnlmonol. 6:275-212.
Spickett GP, Farrant J. 1989. The role of lymphokines in common variable hlpogammaglobrrlinaemia.Immunolory Today. L0:LV2-L94.
Spiegelberg HL, Fishken BG, Grey HM. 1968. Catabolism of human gamma-G immu¡.gtsþulins ofdifferent heavy shain subclasses. I. Catabolism of gamm¿-6 myeloma p.õt"i* in man. J. Clin. Invest.47:?3?3-?330.
Spirer Z- 1980. The role of the spleen in immunity and infection. Adv. pediatr. Z7:55-gg.
Standen GR' 1988- Wiskott-Aldrich Syndrome: nelv perspectives in pathogenesis and management. J.R.Coll. Physicians Lond. 22:80-83.
Stanley PJ, Cole PH. 1985. The concentrations of IgA and free secretory piece in the nasal secretions ofpatients witl recurrent respiratory infections. Clin. Exp. Immunol. 59:lg7-W2.
Stanley PJ, Corbo G, Cole PH. 1984. Serum IgG subclasses in ch¡onic and recurrent respiratoryinfections. Clin. Exp. Immunol. 58:703-708.
Stanworth DR, Smith AIC 1qB. Inhibition of reagin mediated PEA reactions in baboons by human IgG4subclass. Clin. Allergy. 3:17-41.
Stanwortl DR. 1983. lmmunological aspects of human IgG4. Clin. Rev. Allergy. 1:183-195.
Staugas REM, Beard lJ, Simmer K, Ferrante A. 1988. Hypogammaglobulinaemia and depressed natu¡alkills¡ cell activity in a patient withPneumoqstis cañnü infection. Pediãt¡. Infect. Dis. J. 7:7?-L7?ß.
Steinberg AG, Cook CE. 1981-. The distribution of human immunegleþnlin ¿llst)¡pes. OdordMonographs on Medical Genetics. O:dord, Oford Uni. press.
Stern RC. Pulmonary suppuratioq bronchiectasis. L983. In: Nelson TeÍbook of pediatrics. Behrman RE¿¡d \'/¿ngh¿¡ VC, eds. Japan. WB Saunders. p. 1071.
Stevens DP, Frank DM, Mahmud AAF. 1-978. Thymus dependency of host resistance to Gia¡dia murßinfection studies in nude mice. J.Immunol. 72û:ß0{fj2.
Stites DP. lÇ$f,,. rmm¡¡ologic laboratory tests. Clinical laboratory methods for detection of antþens andantibodies' In: Basic and Clinical Tmmunology. Stites DP, Stobo JD, Fudenberg HH and Wells rV, eds.Lange, California. pp. 325-365.
Stockley RA, Burnett D, Afford SC. 1981-. The immunological measu¡ement of 'free' sesretory piece andits relationship to local IgA production. Clin. Exp. Tmmunol.45:72A-1j0.
Strober W, Krakauer R, Klaeverman HL, Repolds lfy, Nelson DL. tn6.deficiency. A disorder of the IgA immune system. N. Eogl.J. Med. 294:35L-356.
Secretory component

307Sullivan JL, Ochs HD, Schiff-an G, ÉIammerscblag MR, Misser J, Vichinsþ E, Wedgwood RJ. lng.rmrng¡s respoDse after splenectomy. Lancet. 1:17g_1-g1.
Sundqvist Vd Linde d trl/ahren B. 1984. Virus-specific imm¡¡sglgbrrlin G subclasses in herpes simplexand varicella-zoster virus infections. J. clin. Microbiol. }nrg+gg
Svejgaard {PlazlP, Ryder LP. 1983. HI.A and disease L9g2 _ asurvey. tmmunol. Rev. 70:193_21g.
Taussþ MJ. 1988. Molecular genetics of immuneglsþnlins. Immunol. (suppl) L:7;16.
Taylor PW. 1983. Bactericidal and bacteriolytic activity of serum ¿gainst gran_negative bacteria.Microbiol. Rev. 47:¿t6-83.
å3" -, Abraham KM. 19g7. The regulation of antibody class expression. rmmunolory Today. g:122_
Teale JM, Klinman NR' ]rgu. control of the production of different classes of antibody. In:Fundamental Imm¡¡slsgy. paul W, ed. New york Raven p;;.,;p 5D_536.
Teale JM' 1982' The potential of B lymphocytes for isotype erçression. Fed. proc. 4t:vt9g-?s01.
iffi"ff.¿ ffi.k"ff;.å,ïHläî er r'm¿¡1 gamm¿-glsþurin þ¿5sd on differences in the heavy
Terry WD. 1-968. Immunologic deficiency rlise¿5es in man. Birth Defects . 4:357_363.
i"ïr"Järl3ffi. u*"tions in the subclasses of IgG. Birth Defects. Immunological Deficiency Diseases
Lindsten T, LgE/.. Size_dependent Bof cell vo cel cycle, pioliferative
zuz. production J. rmmun õt. tszrnlz-
Thong YH, Douelas BS, Ferrante A. 198a. .Abnormal neut¡ophil chemotaxis in a slmdrome of unusualfacies, proportionate small statu¡e and sensori-neu¡al deafüess -'-utis-. Acta paediat¡. Sc¿¡d. 67: ß3-387.
Thong YH, Ferrante A, Ness D' rg77- Neutrophil phagoc¡ic and bactericidal dysfunction induced bybilirubin. Aust. PaediaÏ. J. 73:?Å7-?Å9.
Thong YH' Ferrante A' Ln8. A semi-automated nicroassay ¡schnique for quantitalive neutrophiliorlination. J. Immunol. Methods. 0:2gl-299.
Thong YH, Ferrante A. L979. Inhibition of mitogen-induced human þmphocyte proliferative responses bytetracycline analogues. Cli". E*p. Immunol. 35:443446.
Thong YH, Maxwell GM. 19?8. pti-aty selective deficiency of immunogrsþrrin M. Aust. N.Z. J. Med.8:436-438.
;TffinJ?s.3'T;î*e A. re7e. Effecr of bitirubin on rhe funsicidal capacity of h ,man neurrophfü.
Thong YH, Roberton EF, Rischbieth HG, Smith GJ, Binns GF, Cheney C, pollard AC. 197g. Successfulrestoration of immunit ' in the diGeorge slmdrome witl ret¡ "pitl"urt
tr'ansplant. À"i. oir. child. 53:5g0-584.
Thong YH' Simpson D'\ Multer-Eberha¡d ry: ury. Homozygous deficiency of the second component ofcomplement presenring with recurrent bacterial 6s,ingrtis. eri,i. ó¡r. Child. 55:47 L473.
Tilden AB, Abo T, Balch CM. 1983. Suppressor cell function sf ftum¿¡ granular lymphocytes identified byHNK-L (Leu 7) monoclonal antibody. J. Tmmunol. t3f¡:ttlt_t1¡iS.

308Tiselius A, Kabat EA. L939. An electrophoretic study of immune sera ¿ùld purified antibody preparations.J. Exp. Med. 69:1L9-131-.
Tiselius A. 1937. Elect¡ophoresis of serum globulin II: electrophoretic analysis of normal and immuneserum. Biochem. J. 3L:L46ÇL477.
Tiselius A. 1939-4[.. Electrophoretic analysis and constitution of native fluids. Harvey I*ct.35:37-X).
Torfasen EG, Reimer CB, Keyserling }JC. L987. Subclass restriction of human enterovirus antibodies. JClin. Microbíol. 25:1i7 6-1i79.
Tou¡aine JL, Betuel H. 1983. The Bare Lymphocyte Syndrome : immunodeficiency resulting f¡om the lackof e4pression of HI-A antþens. Bi¡th Defects: Original a¡ticle series, 19:83-85.
Truedsson L, Sjoholn AG, Laurell A-B. 1-981. gs¡ssning for deficiencies in the classical and alternativepathways of complement by hemolysis in gel. Acta Path. Microbiol. Sc¿nd. Sect. C. 89:161-L66.
Turner MW, Bennich HH, Natvþ JB. 1970. Pepsin digestion of human G-myeloma proteins of differentsubclasses. I. The characteristic featu¡es of pepsin cleavage as a function of time. Clin. Exp. Immunol.7:603-625.
Ugano AG, Out Td PlebaniA, Duse M, Monafo V, Nespoli L, Burgio GR. 1983. Recurrent infections inchildren with 'selective' IgA deficiency association with IgG2 and IgG4 deficiency. Birth Defects. L9:169-t7t.
Umetsu DT, Arnbrosino DM, Quinti I, Siber GR, Geha RG. 1985. Recurrent sinopuünonary infectionand impaired antibody response to bacterial capsular polysaccharide antigen in child¡en with selective IgG-subclass deficiency. N. Engl. J. Med.313:1247-1251.
Urbanek R, Kemeny DM, Richards D. 1986. Subclass of IgG anti-bee venom antibody produced duringbee venom immunotherapy and its relationship to long-term protection from bee stings and followingtermination of venom immunotherapy. Clin. Allergy. l6:3L7-322.
Van der Giessen M, Groeneboer-Kempers O. 1976. The subclasses of human IgG antibodies ag¡ins¡tetanus toxoid. Clin. E,lç. Immunol. ?5zLL7-121.
Van der Giessen M, floman'WL, van Kernebeek G. et al. L976a. Subclass typrng of IgG antibodies formedby grass pollen allergic patients d*iog immunotherapy. Int. Arch. Allergy.Appl. Immunol. 50:6?5-639.
Van der Giessen M, Reefrink-Brongers, Aþra-van Veen T. In6b. Quantitation of IgG classes andsubclasses in sera of patients with a variety of immunogloþnlin deficiencies and thei¡ relatives. Clin.rmmunol. Tmmurep¿thol. 5:388-398.
Van der Giessen M, Rossouw E, Aþra-van Veen T, Van I-oghem E, Zngerc BJM, Sander PC. L975.Quantitation of IgG subclasses in sera of normal adults and healthy child¡en between 4 and,12 years of age.Clin. Ex". Immunol. 2L:501-5ú.
Van der Meer JWM, Vossen JM, Radl J, van Nieuwkoop Jd Meyer ChJLM, Lobatto S, van Furth R.1984. Hyperimmunoglobrrlinaemia D and periodic fever: a new syndrome. Lancet 1:1087-i-090.
van Fu¡th R, Iæijh PCI, Klein F. 1984. Correlation between opsonic activity for various microorganismsand composition of gammaglobulin preparations for int¡avenous use. J. Infect. Dis. 149:511-517.
Van Loghem E, Subernik RI, Osþova LP,7-ngers BJ, Matsumoto Ff, delange G, Lefranc G. 1980. Genedeletion and gene duplication within the cluster of human heavy-chain genes. Selective absence of IgGsubclasses. J. Tmmunsgsrct. 7 :2f35-299.
Van Loghem E,7agerc BJM, Bast EIEG, Kater L. 1983. Selective deficiency of imnunoglobulin A2. J.Clin. Invest. 72:L9L8-L923.

309van l'oghem E. 191. IgG allotlpes. The_ Tmmunoglobrrlin genes: genetics, biological and clinicalsienificance. Clinics in Tmmunol. uod A¡1ergy. ae):607-6ñ,.
Vinalak VK Jain P, Naik SR. Lw}. Demonst¡ation of antibodies in giardiasis using irnm¡¡odiftision¡schnique with Giardia cyst antigen. Ann. Trop. Med. pa¡asitol.T2:5gt-5g2.
Vi¡ella G, Howard A' Cherrington J. 1970. Distribution ofåeavy chain subclasses sf immgnoglobrrlin G innormal IgG and residual IgG from hypogamm¿glsþnlin¿emis."å. CUo. Exp. Immunol. 7:671_6g0.
virella G' lnl Degradation of IgG3 monoclonal proteins during storage and in the presence of a thioi(DLpenicillamine). Experentia 2l:94196.
, Walsh , Nelson D! Chanock RM, Murphy
:.Ëff respirarory slncyrial virus F _å c
Wahlgren W, Berzìns IÇ Perlmann P, Persson M. 1983. Characterization of humoral imms¡s response inPlasmodium falciparum malaria. Clin. Exp. rmmunol. 54:l3S_L42.
Waldman TA. L987.. The arrangements of immr¡nsglobulin and T cell receptor genes in hrmanlymphoproliferative disorders. 4O:2A7 -32t.
waldman" TA! Broder s, Golrlman cIÇ Frost IÇ Korsmeyer SJ, Medici MA. 19g3. Disorders of B cellsand helper T cells in the pathogenesis of immunoglobutin deficiáocy of patients with ataxia telangrectasia.J. Clin. Invest. 7 I:282-295.
Watt PJ, 7ãdts M, Lambden PR. 1986. Age related IgG subclass response to respiratory sync¡ial virusfusion protein in infected infants. Ctin. Exp. Tm-unol. Oã,SO¡_Sæ.
wedege E, Michaelson TE. 1987. Flurn¿n immunoglobrlin Ç subclass imms¡s response to outermembrane antigens ¡¡ meningococcal group B vaccine. ¡. ctin. Microbiol. 2s:149_1jJ53.
Wedgwood RL, ochs HD, Oxelius v-4. 198ó. IgG subclass levels in the serum of patients with primaryimmunodeficiency. Monogr. Allergy. 20:g0-g9.
weinberg GA" Granoff DM, Nahm MH, Shackelford PG. 198ó. Functional activiry of different IgGsubclass antibodies against rype b capsular polysaccharide or Haemophilus influeruae. J. Immunol.1i6:4?32-4?5.
rJ/einberg Il Parkman R. 1990. Severe combined immunodeficiency due to a specific defect in theproduction sf i¡¡s¡lenkin -2. lV. EnCl. J. Med,. 322:L7 Lg-L723.
wells MA \ilittek AE, Epstein JS et al. 1986. .Inactivation and partition of hr¡man T-cell lymphocytotropic
virus q'pe m, dudng ethanol fractionation of plasma. Transfusion . ?Á:2L0-2r3.
Whitting¡¿¡ S, Mackay IR, Mathews JD. 1984. HI-A-Gn interactisns; clinical implications. Clinics inïmmunol. and Allergy. 4:6?364O.
Whittingham S, Mathews JD, Schanfield MS, Matthews JV, Tait BD, Morris pJ, Mackay IR. 19g0.Interactive effect of Gm allotlpes and HI-A-B locus antþens'on human antibody ,.rpo^" to a bacterialantþen. Cli". E*p. Immunol. ¿lO:8-15.
Wild AE. t979. Fc receptors and selective ìmmunoglob.lin [rensport across rabbit yolk-sac endoderm. In:Protein transmissisa through living memb¡¿¡s5. flsrnminos W¿, "¿. Amsterdam: Elsevier/North_Holland Biomedical Press. pp. n-K.
wrigbt sG, Tomkins AM. ln7. Quantification of the lprphocyte infilu¿¡s in jejunal epithelium ingiardiasis. Clin. Enp. Tmm¡¡sl. 29:4O8412.
Yql9 H,-Yajima T, Tnnii T, Ashiba T, Iwata M. 1986. opsonic and complement-dependent bactericidalactivities of va¡ious immunoglobrlin preparations for int¡avenous use. Vox S¡ng. 5t:270-277.

310Yount w, Dorner M, Kunkel H, Kabat E. L968. Studies on human anJibodies. VI. Selective va¡iations insubgroup compositions and genetic markers. J. Exp. Med.12ir:63k646.
Yount WJ, Cohen P, Eisenberg RA. 19gg. fStributigl of IgG subclasses among furrm¿¡ autoantibodies toSm, RNP, dsDNAb ss-B and IgG rhenmatoid factor. rvro"ofr._-e¡ã gy23:4L-56.
#åljJti.r"Hî .Distribution of rgG subclasses in h,,m¿¡ B_lymphocytes:
subpopulations. J. in su¡face and cytoplasmic IgG in -i'or BJymphocyte
Yount wJ, Hong R, Seligmann M, Good R, Kunkel HG. 1970. hlaþ*_. of gamm¿glsþrrin suþg.supgand gene defects in patients with primary hpoganmagl6þntinsmi¿. J. clin. Invest. 49:1957-LW.
Yount wJ, Kunkel GH, Litwin sD. 1962. Studies of the vi (gamm¿-f,s) subgroup of gamma-glob.rin. ArelationshiP between the concentration and genetic qp" *oì! *.-¿ ioaoião*. t. E4p. Med,.125:t7-
Tnlaschl D, Nevòy c, Parsons M, van west R, cavalli-Sforza LL,Herznnberg I-d Herzenberg I¿' 19g3.H'man immursglsþrrlin ¿llgtlPes: previousiy *"*gni."a iete minants and alleles defined withmonoclonar antibodies. proc. Nat. acaã. sa. uSA. g0:376î2 37ff..

311
PTJBLICATIONS
The following papers describing work presented in this thesis have been published or are currently in press.
Bea¡d LI, Ferrante A' Hagedorn JF, Leppard P, Kiroff G. 1990. Percentile rânges for IgG subclasscoucent¡ations in healthy Aust¡alian child¡en. pediat¡.Infect.DisJ. 9:s9-15.
Bea¡d Lf, Ferrante A" Oxelius V-A, Maxwell GM. 1986. IgG subclass deñciency in children with IgAdeficiency presenting witl recurrent or severe respiratory infections. Pediat¡.Rese a¡ch. ?n:g37-942.
Beard IJ, Ferrante A. 1988. IgG replacement tlerapy i" IgG subclass-deficient child¡en. Monogr¿llergl.23:L9L?Ã3.
Beard IJ, Ferrante A. 1989. IgG4 deficiency in lgA-deficient patients. Pediatr. Infect.DisJ. g:705-709.
Beard LI, Ferrante A. 1990. Aspects of immunoglqþulin replacement therapy. Pediatr.Infect-DisJ. 9:S5¿t-6r.
Beard LI, Ferris L, Ferrante A. 1990. Immunegloþrrlin Ç subclasses and lymphocyte subpopulations andfunction in osteomyelitis and septic arthritis. Acta paediatr.scand. 79: in pt"ir.
-
Ferrante A, Beard LI, Feldman RG. 1990. IgG subclass dist¡ibution of antibodies to bacterial and vi¡alantigens. Pediatr.Infect.DisJ. 9:St6-?A.
Ferrante d Beard LI. 1988. IgG subclass assays with polyclonal and monoclonal antibodies. Monogr.Allergy. ?3:6I-72.
Ferrante A, Davidson GP, Beard LI, Goh DHB.peripheral blood mononuclea¡ leu-kocytesIntÁrch,AllergyAppl.Im m unol. 88:38-352.
1989. Alterations in function and subpopulations ofin child¡en with portal hlpertension.
Ferrante Ao Rowan-Kelly B, Beard LI, Maxwell GM. 1986. An enzyme-linked immunosorbent assay forthe quantitation of human IgG subclasses using monoclonal antibodies. J.Tmmunol.Methods. g3:?Ã7-zb.
Staugas REM, Beard LI, gimme¡ Il Ferrante A. 1988. Hypogemm¿globulinaemia and depressed naturalkiller cell activity in a patient with Pneumocystß ca¡inü i¡fection. Pediãtr.Infect.DisJ. 7:7?A-7?3.




















