
IGF-1對發育時期運動神經生長轉向分子機制之探討 - 國立中山 ...
96
國立中山大學生物科學系 碩士論文 Department of Biological Sciences National Sun Yat-sen University Master Thesis IGF-1對發育時期運動神經生長轉向分子機制之探討 Study on the Molecular Mechanisms of IGF-1-induced Turning Response at Developing Motoneuron 研究生:楊昆霖 Kun-Lin Yang 指導教授﹕劉昭成 博士 Dr. Jau-Cheng Liou 中華民國105年7月 July 2016
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of IGF-1對發育時期運動神經生長轉向分子機制之探討 - 國立中山 ...

國立中山大學生物科學系
碩士論文
Department of Biological Sciences
National Sun Yat-sen University
Master Thesis
IGF-1對發育時期運動神經生長轉向分子機制之探討
Study on the Molecular Mechanisms of IGF-1-induced Turning
Response at Developing Motoneuron
研究生:楊昆霖
Kun-Lin Yang
指導教授﹕劉昭成 博士
Dr. Jau-Cheng Liou
中華民國105年7月
July 2016

i
論文審定書

ii
致謝
兩年來的研究所生涯,雖然過程中歷經許多挫折與失敗,不過在紮
實的磨練下,終於完成學位了!
在此要特別感謝我的指導教授 劉昭成老師讓我加入這個實驗室,
提供學習的機會與空間,謝謝老師在實驗研究過程中給予許多指引與
教導。另外,感謝本系所 吳長益老師和高雄醫學大學醫學系藥理學科
吳炳男老師,在我的論文上費心的批改與修正,也給予寶貴的意見。也
非常感謝實驗室的同學-柯品伃,以及學弟妹們-劉嘉恩、李宓錤、陳信
甫和洪翊筑,在這段研究過程中相互輔助,讓我的研究能夠順利完成。
最後我要感謝我的家人,謝謝你們默默的在背後支持我,作為我最
強的後盾。

iii
中文摘要
神經生長發育初期需要指引因子(guidance cues)或基質結合分子(substrate-
bound molecules)與神經生長錐(nerve growth cone)的接受器結合,接著經由一連
串的訊息傳遞路徑造成細胞骨架的重新排列,最終改變軸突的生長方向。雖然在胚
胎發育時期運動神經與肌細胞形成突觸的過程中胰島素樣生長因子(Insulin-like
growth factor-1,IGF-1)扮演重要的角色;根據研究也發現,胚胎發育時期骨骼肌
細胞中 IGF-1 表現量相當高,直到神經-肌突觸建立完成後 IGF-1 的表現量才會下
降。然而,IGF-1 對於胚胎發育早期運動神經生長方向是否具有導引作用,目前還
是不清楚的。
在本實驗中,主要探討 IGF-1 在胚胎發育時期造成神經生長錐偏轉現象
(growth cone turning),以及訊息傳遞路徑。我們利用非洲爪蟾(Xenopus Laevis)
的神經培養,以 turning assay 的實驗方式來探討單一特定分子 IGF-1 對 axon
guidance 生長呈現趨向(attraction)或逆向(repulsion)的分布。在 turning assay 是
使用細胞培養後 4 至 10 小時(胚胎發育時期)和 24 小時(成熟時期)的 culture。注
射用玻璃毛細管(直徑約 1 μm)以每秒 1 次,每次 50 msec duration 的給藥方式來
進行實驗。
研究結果顯示 IGF-1 注射下,胚胎發育時期神經生長速率較成熟時期神經快,
且轉彎角度(turning angle)也較明顯朝向注射方向偏轉。預處理 IGF-1 接受器的抑
制劑 Genistein (tyrosine kinase inhibitor)可以阻斷 IGF-1 所造成神經受到吸引而
產生偏轉,顯示 IGF-1 造成的 growth cone turning 是透過 IGF-1 receptor 的活化,
引起下游訊息傳遞路徑,最後造成神經偏轉現象。另外針對鈣離子(Calcium)的來
源,在 Ca2+-free 溶液置換和鎘離子(CdCl2)預處理下,結果發現皆影響 IGF-1 對神
經產生的偏轉作用,顯示鈣離子的流入需要透過 voltage-activated calcium
channels 提共。在細胞內儲存池 IP3R 利用 Xec 抑制劑預處理,會阻止原本 IGF-

iv
1 對神經產生的偏轉作用。分別預處理 PI 3-kinase 抑制劑 wortmannin 和
phospholipase Cγ (PLCγ)抑制劑U73122皆會影響 IGF-1對神經吸引轉向的作用。
此外,IGF-1 對神經偏轉作用也會受到 TRP channels 的抑制劑 SKF96365 或
Gadolinium (Gd3+)的處理而減少,顯示細胞內鈣離子參與其中的訊息調控。綜合
以上所述,我們的研究顯示 IGF-1 吸引神經的偏轉作用是透過 PLCγ,IP 3-kinase
以及 TRP channels 的訊息傳遞路徑所調控。
關鍵字:神經生長錐、神經導引因子、鈣離子儲存池、TRP channel、IGF-1

v
Abstract
The path-finding of developing neurons subject to the guidance of both attractive
and repulsive cues emerge in the way to their target. The discoveries of IGF-1
(insulin-like growth factor-1) expression in the developing skeletal muscle
increase with the formation of differentiated skeletal muscle fibers and decrease
to very low adult levels during the process of synapse elimination. Although
evidence suggests that insulin-like growth factor plays an important role in the
development and growth of the neuromuscular synapse, the effect of IGF-1 in the
guidance on the growth cone of a motoneuron remains unknown. Here we focus
on the possibility and underlying molecular mechanisms of IGF-1 in the guidance
of a developing neuron by using 4 to 10 hours (embryonic development) and 1-
day-old (mature stage) primary cultured motoneurons of Xenopus laevis.
The growth rate of developing axons was increased and the growth cones were
significantly attracted toward to the direction of IGF-1 application. Pretreatment
of tyrosine kinase inhibitor genistein effectively occluded growth turning response
induced by IGF-1. The IGF-1-induced guidance effect was abolished when
calcium was eliminated from the culture medium or bath application with the
pharmacological calcium channel inhibitor cadmium, indicating that calcium
influxes through voltage-activated calcium channels is required. IP3 receptor Xec
hampered the IGF-1-induced turning. Treating cells with PI-3 kinase inhibitor
wortmannin and with phospholipase Cγ (PLCγ) inhibitor U73122 abolished IGF-
1-induced axonal attraction. Moreover, IGF-1-induced turning response is
significantly occluded by store depletion-activated calcium channel (TRPC)
inhibitors either SKF96369 or Gadolinium, suggesting the involvement of internal
calcium store. Overall, results from our studies demonstrate IGF-1 has an

vi
attractive effect on axonal guidance and this is done via PLCγ, PI3 kinase and
TRPC signaling cascades.
Keywords:axon guidance, guidance cue, sarcoplasmic reticulum, TRP
channel, insulin-like growth factor 1

vii
目錄
論文審定書 ................................................................................................ i
致謝 ........................................................................................................... ii
中文摘要 .................................................................................................. iii
Abstract .................................................................................................... v
目錄 ......................................................................................................... vii
圖目錄 .................................................................................................... viii
附圖目錄 ................................................................................................... x
縮寫表 ...................................................................................................... xi
緒論 ........................................................................................................... 1
實驗目的 ................................................................................................... 8
實驗材料 ................................................................................................... 9
實驗方法 ................................................................................................. 12
結果 ......................................................................................................... 19
討論 ......................................................................................................... 29
參考文獻 ................................................................................................. 35

viii
圖目錄
Figure 1. Experimental design of pressure injection-induced microscopic gradient. ...................................................................... 45
Figure 2. Cyclic AMP microscopic gradient successfully induce attractive response on growth cone of Xenopus developing motoneuron. .................................................................................... 47
Figure 3. Attractive effect of IGF-1 on Xenopus developing spinal motoneuron. .................................................................................... 49
Figure 4. Effect of IGF-1-induced attractive turning response on Xenopus motoneuron at different developmental stages. .............. 51
Figure 5. Role of calcium on IGF-1-induced growth cone turning. .. 53
Figure 6. Involvement of internal calcium reservoir in IGF-1-induced growth cone turning response. ....................................................... 55
Figure 7. The exocytic membrane effect of IGF-1 microscopic gradient on the growth cone. .......................................................... 56
Figure 8. Role of ryanodine receptor in IGF-1-induced attractive turning response. ............................................................................ 58
Figure 9. IP3 receptor is responsible for IGF-1-induced turning response at developing Xenopus motoneuron. .............................. 60
Figure 10 . Activation of tyrosine kinase IGF-1 receptor is prerequisite for IGF-1-induced growth cone turning response........ 62
Figure 11. PLCγ activity is involvement in IGF-1-induced growth cone turning. ................................................................................... 64
Figure 12. Involvement of PI-3 kinase pathway in IGF-1-induced growth cone turning. ....................................................................... 66
Figure 13. Role of MAP kinase in growth cone turning response induced by IGF-1. ........................................................................... 68
Figure 14. Summary of the effect of pharmacological inhibitors on IGF-1-induced growth cone turning response. ............................... 70

ix
Figure 15. TRPC channel is involved in IGF-1-induced growth cone turning response. ............................................................................ 72
Figure 16. Effect of TPR channel inhibitor Gd3+ on the growth cone turning response induced by IGF-1. ............................................... 74
Figure 17. Schematic diagram illustrating in the signaling cascade underlying IGF-1-induced attractive growth cone turning response. ........................................................................................................ 77

x
附圖目錄
Table 1. ........................................................................................... 75
附圖 1. The structure of the growth cone. ...................................... 79
附圖 2. Chemorepulsion and chemoattraction. .............................. 80
附圖 3. Structure of the insulin receptor and IGF-I receptor. .......... 81
附圖 4. Representation of the expression profiles of IGFs. ............ 82
附圖 5. The system of pharmacy appliction with air pump. ............ 83
附圖 6. Procedures for the preparing Xenopus nerve culture. ....... 84

xi
縮寫表
ACh acetylcholine
cAMP cyclic adenosine monophosphate
DAG diacyl glycerol
IGF-1 insulin-like growth factor 1
IP3 inositol 1,4,5-trisphosphate
IP3R inositol 1,4,5-trisphosphate receptor
MAPK mitogen-activated protein kinase
PIP2 phospholipid phosphatidylinositol 4,5-bisphosphate
PLCγ phospholipase Cγ
RyR ryanodine receptor
TG thapsigargin
TRP transient receptor potential

1
緒論
壹、 神經細胞的探索與發育
一、 神經細胞的結構與組成
過去幾十年來,已經有許多研究在瞭解有關於神經細胞的各個生長發育階
段與機制,而神經的發育牽涉到神經的產生、型態的改變以及神經系統的建立;
雖然成千上萬的神經細胞形狀和大小截然不同,但組成結構可以定義為主要三
個區域:(1) peripheral domain (P),由 actin 組成包含 filopodia 和 lamellipodia
結構、(2) transitional domain (T),位於 P domain 與 C domain 之間,並垂直
於 F-actin 形成一個 hemicircumferential ring 和(3) central domain (C)由
microtubule 組成,負責延伸或收縮神經的長度(Fuchs and Cleveland, 1998,
Gallo and Letourneau, 2004, Mattila and Lappalainen, 2008, Lowery and Van
Vactor, 2009) (附圖 1.)。神經細胞需要利用 cytoskeletal proteins、molecular
motors 和 cell adhesion molecules (Zheng et al., 1994, Tojima and
Kamiguchi, 2015),以及 actin 和 microtubule 動態平衡(dynamics)的特性
(Schaefer et al., 2008, Conde and Caceres, 2009),來共同調控神經軸突的生
長方向及長度的延伸。
若將神經軸突以縱剖分為左右兩側,可以發現當細胞膜上其中一側的受體
與配體如滋養因子或 adhesion molecules 的吸引而產生結合,經由下游訊息
傳遞分子調控 filopodia 或 lamellipodia 的結構,能夠使細胞骨架 actin 產生重
組排列的現象,以 actin polymerization 的形式讓細胞膜向外延伸;而細胞膜
另一側則會以 depolymerization 的方式使得 actin 表現量遞減,整體而言,神
經細胞就會朝向吸引因子的一端不斷增長(Zheng et al., 1996, Dent and
Gertler, 2003, Marsick et al., 2010)。然而,在其他活體細胞影像實驗中發現,

2
actin filaments 與鈣離子之間會相互影響,其中 spine 的形態會影響
postsynaptic 鈣離子的流入(Majewska et al., 2000),而且透過神經傳遞物質
來調控鈣離子流入的時間和流量(Koch and Zador, 1993, Svoboda et al.,
1996);相對的,若調整細胞內鈣離子濃度,則會造成 spine actin 骨架的重新
排列和動態平衡的改變,導致 spine 型態和穩定性有所變化(Fischer et al.,
1998, Richards et al., 2004)。
二、 神經細胞生長需要特定指引因子
神經細胞發育的過程中,需要仰賴一些特殊分子引領它們的生長方向,而
這些引領分子被稱為:『指引因子(guidance cues)』,因子可以是吸引(attractive)
或排斥(repulsive)神經生長錐往其方向生長(附圖 2.),在環境中各種指引因子
因為距離的遠近形成濃度梯度差,並且透過兩種不同的機制影響神經錐產生偏
轉 現 象 : (1)chemoattraction 和 (2)chemorepulsion(Goodman, 1996,
TessierLavigne and Goodman, 1996)。
根據 Roger Sperry 過去針對視覺神經系統的研究(Parekh and Putney,
2005),得到了以下的結論:當視神經纖維造成損傷或斷裂時,再生的纖維具
有重新沿著原始生長軌跡連接回中腦 tectum 的能力,以及這些重建連結是建
立在有順序性的基礎之下(chemospecificity theory);換言之,每條視神經纖維
和 tectal 神經元具有 cytochemical label,這些特殊標定能夠專一的表達不同
神經細胞的類型和位置,使得相異的神經細胞具有選擇性,與目標細胞產生連
結(Parekh and Putney, 2005)。
另外,Scolnick 在嗅覺神經細胞的實驗研究也有相似的發現,在正常情況
下嗅覺神經細胞會與 lateral bulb 產生連結,不過當抑制 Insulin-like growth
factors (IGFs)訊息傳遞路徑後,因缺乏神經指引因子,最終神經細胞與 ventral-
medial 產生突觸,導致錯誤的連結(Scolnick et al., 2008)。細胞外有許多不同

3
種類的指引因子,利用膜上特異性的受體交互作用,開起胞內下游如 cascade
般的訊息傳遞路徑,改變神經細胞的生長軌跡,因此本實驗將進一步了解 IGF-
1 對於神經生長過程中導引神經生長的機制是如何產生的。
三、 神經生長偏轉
在許多研究中顯示,細胞外環境中各種吸引或排斥因子建立在濃度梯度
情況下,例如:brain-derived neurotrophic factor (BDNF)、cyclic adenosine
monophosphate(cAMP)、netrin-1 以及離子等,透過膜上特異性的受體交互
作用 (Lohof et al., 1992, Eichmann et al., 2005, Berzat and Hall, 2010,
Nieto-Estevez et al., 2016),便會啟動下游多樣化的訊息傳遞路徑,造成局
部 actin 細胞骨架的重新排列,使得神經生長的方向產生漸進式的反應
(graded response) (Dickson, 2002, Guan and Rao, 2003, Chilton, 2006),
最終改變神經生長錐的偏轉方向。
然而,神經錐生長過程中也會改變細胞膜的動態平衡,因此在近年的研究
認為造成神經細胞偏轉,可能與細胞膜的 exocytosis 和 endocytosis 囊泡運輸
機制有關(Tojima et al., 2014, Ros et al., 2015),因為許多證據顯示細胞骨架
和黏附物質的動態變化,是必須透過 exocytosis 和 endocytosis 囊泡運送
(Itofusa and Kamiguchi, 2011, Tojima et al., 2011, Vitriol and Zheng, 2012)。
雖然,目前已經發現許多的化學分子都能夠影響神經生長的方向,但確切的訊
息傳遞路徑目前為止尚未了解清楚,尤其是常見的 second messenger–鈣離
子,在複雜的機制中所扮演的角色沒有清楚的答案。
貳、 運動神經的成熟與再生
一、 運動神經與肌細胞的建立
在胚胎發育時期,當運動神經細胞與肌細胞接近時,彼此間在結構上會產

4
生非常複雜的改變:舉例來說,哺乳動物的中樞神經系統中 corticospinal tract
(CST)會形成最長的神經軸突,它起始於腦部皮質直到脊椎末梢,最後與肌細
胞形成緊密的結合,用於控制肢體肌肉組織精確的移動(Canty and Murphy,
2008)。而在周圍神經系統的發育過程中,也可以發現神經細胞會從神經脊
(neural crest)和外胚層基板(ectodermal placodes)開始進行程序性的生長、發
育和遷移(Jessen and Mirsky, 2005)。
運動神經末梢由本來負責生長與搜尋路徑的生長點轉變成具有特化結構,
形成有效率釋放神經傳遞物質的神經末梢,而肌細胞則是在神經細胞接觸之後,
會進行一些結構上的變化,如乙醯膽鹼接受體(Acetylcholine receptor,AChR)
往突觸的方向集中以及突觸凹(Junctional fold)的形成等,而這個過程的改變
與神經細胞和肌細胞彼此之間的電流訊號與滋養因子的釋放關係緊密(Sanes
and Lichtman, 1999)。當神經細胞固定於特定位置後,便開始分化出樹突
(dendrites),負責探測環境與接收訊號;當軸突(axon)與目標細胞形成連結的
特殊構造-突觸(synapse)之後,則負責傳出細胞體的訊號。然而神經生長錐尚
未與目標細胞間產生連結時,都是藉由外界環境因子和神經細胞間的交互作用,
來指引他們往特定的方向生長(Song and Poo, 2001, Gallo and Letourneau,
2004)。
二、 Insulin-like growth factors system (IGFs)
Insulin-like growth factors (IGFs) 最 早 在 1976 年 被 科 學 家 發 現
(Rinderknecht and Humbel, 1976)。IGF-1 會作用於 IGF-1 受體(IGF-1 receptor,
IGF-1R),IGF-1與 insulin的受體非常相似,均由具有 ligand結合位的α subunit
及具有 tyrosine kinase 活性的 β subunit 組成,再藉由雙硫鍵將二個 α β基本
結構的 monomer 連結成一個 α2β2的 heterotetramer receptor。IGF-1 與其受
體 α subunit 結合之後,會造成受體結構上的改變(conformational change),
而造成兩個 β subunit 上的 tyrosine 的自主磷酸化(autophosphorylation),使

5
tyrosine kinase activity 活性增加,β subunit 上的許多 tyrosine 會形成
phosphorylation site,提供下游訊息傳遞蛋白的結合位,當訊息傳遞蛋白結合
這些特定的結合位之後,便會引發細胞內下一步的訊息傳遞路徑反應,而產生
不同的生理作用(附圖 3.) (D'Mello et al., 1997, Butler et al., 1998, Adams et
al., 2000, Van Obberghen et al., 2001)。
就 IGF-1 受體而言,tyrosine 磷酸化會形成許多的 phosphorylation site,
這些 phosphorylation site 可以形成專一的 docking site,進而得到許多 insulin
receptor substrates (IRS)-1 及-2,透過與 β subunit 上 docking site 的結合使
tyrosine residue 被磷酸化,使特異性的結合位能夠與 Src-hormology domain
(SH2 和 SH3 domain) 、 Pleckstrin homology domain (PH domain) 或
phosphotyrosine binding domain (PTB domain)的蛋白接合上去而活化,因此
一但 IGF-1R 被活化之後,將會啟動 phosphoinositide 3-kinase (PI 3-kinase)
(Franke et al., 1997, Blair et al., 1999, Parekh and Putney, 2005) 、
phospholipase Cγ (Sasaoka et al., 2001),以及 mitogen-activated protein
kinase (MAPK) (Kim et al., 1997),等訊息傳遞路徑。根據文獻顯示,這些活
化的訊息傳遞路徑會產生多樣化的生理結果,如促進細胞的增生與分化、避免
神經細胞走向凋亡或形成斑塊、促進神經 neurite 向外延伸以及調節神經細胞
N 或 L type 鈣離子通道的活化等等(Blair and Marshall, 1997, Ozdinler and
Macklis, 2006, Zhang et al., 2010)。
經歷多年來的研究,科學家已經證實 IGF-1 對於細胞週期的進行、細胞的
分化、細胞的增生以及細胞的癌化扮演重要的角色(Adams et al., 2000,
Brooker et al., 2000, Sasaoka et al., 2001, You et al., 2002, Huat et al., 2014);
另外,IGF-1 在胚胎及 postnatal 時期神經的發育與生長有關,舉例來說:IGF-
1 可以增加 corticospinal motor neurons (CSMN)軸突向外擴展及生長速率
(Meir et al., 1999, Ozdinler and Macklis, 2006)、預防神經元和腦神經細胞凋
亡(Liou et al., 2003),以及 IGF-1 對於建立大腦海馬迴神經網絡的功能是不可

6
或缺的(Sosa et al., 2006)。
除此之外,研究也發現 IGF-1 對於神經損傷後,在神經功能的恢復亦扮演
關鍵的角色,舉例來說:在活體情況下使用 somatomedin C (Sm-C/insulin-like
growth factor I/IGF-I)可以讓成年大鼠坐骨神經進行再生和增加神經受損傷後
所存活的運動神經元數目(Hansson et al., 1986, Pu et al., 1999),在大鼠腦神
經損傷的治療方面,也發現 IGF-1 對於神經修復的潛力(Gluckman et al., 1992),
以及在基因轉殖鼠實驗證實 IGF-1 可以加速修復神經訊息傳遞的速率並強化
神經細胞與肌細胞間的聯繫功能(Rabinovsky et al., 2002)。而 IGF-1 引起的
所有生理作用,都是透過 IGF-1 與其受體結合,進而激活下游一連串訊息傳遞
路徑(D'Mello et al., 1997, Butler et al., 1998, Adams et al., 2000)。
三、 IGFs 促進神經傳遞物質的產生與釋放
根據研究,內源性的 IGF-1 不僅是幹細胞分化成神經元主要的調控因子,
高濃度內生性的 IGF-1 或胰島素也可以促進神經的成熟和神經傳遞物質的形
成(Brooker et al., 2000)。本實驗室也證實,在神經細胞中 IGF-1 會透過 PI 3-
kinase 和 PLCγ路徑的活化,開啟細胞內鈣離子儲存池 IP3和 ryanodine 受體,
提高神經末梢內的鈣離子濃度,進而活化 CaMK II 路徑,最終增加神經傳遞物
質的釋放(Liou et al., 2003)。一但神經細胞與肌細胞接觸後,肌細胞會釋放
IGF-1 至胞外,並作用於神經細胞使得更多神經傳遞物質釋放,來建立他們之
間的交互作用。
根據研究顯示,肌細胞在生長發育初期,尚未受到神經細胞的支配時,會
表現高濃度的 IGFs,尤其在脊椎、中腦、大腦皮層、海馬迴以及嗅球等部位
(Werther et al., 1990, Naeve et al., 2000),在這些特定部位的神經細胞中,為
了讓神經幹細胞能快速分化和增生,幼年時期 IGF-R 的表現量都較成熟時期
高,然而 postnatal 時期隨著神經完成支配肌細胞,IGF-R 的數量會略微減少

7
(Nieto-Estevez et al., 2016)。這個過程中神經細胞內包覆 acetylcholine (ACh)
的囊泡固定不完全,囊泡偶爾會融合並釋放 ACh 至胞外,偶然的作用於肌細
胞上的 ACh-R,同時肌細胞開始產生活性,並透過肌細胞的收縮釋放胞內高濃
度的 IGF-1。
一旦突觸結構發生損壞或失去支配時,肌細胞為了在與肌細胞建立連結,
所以又會表現高濃度的 IGFs 在肌細胞中(Ishii, 1989, Caroni, 1993) (附圖 4.)。
所以可以了解肌細胞內 IGF-1 的表現量與神經細胞的連結有緊密關聯,不僅在
神經細胞發育時期的存活、分化與增生有重要的功能,以及神經受損時的再生
與修復也扮演關鍵的角色。因此由以上證據得到一個暗示:IGF-1 應該在胚胎
發育過程中可能會回饋於神經細胞;不過是否吸引神經生長的作用,讓神經-
肌細胞間的關係更緊密呢?因此進行以下實驗。
本實驗可以確立 IGF-1是否對於運動神經胚胎發育時期促進突觸的建立,
同時對於神經損傷後 IGF-1 對神經修復再生及突觸的重建扮演重要的角色,所
以在 in vitro 的情況下,非洲爪蟾(Xenopus)胚胎發育時期 spinal cord 的運動
神經可以做為一項非常好的實驗動物材料。過去 Zheng 的實驗已經建立了一
套神經生長錐的實驗模組(Zheng et al., 1994),因此我們也試著在本實驗室重
建此實驗模式,用來評估 IGF-1 對神經細胞作用的訊息傳遞路徑機制。

8
實驗目的
本實驗以 turning assay 的實驗方法,來探討 IGF-1 對神經生長產生偏轉作用
中所扮演的角色。由於先前實驗證實神經滋養因子 IGF-1,能夠促進神經傳遞物質
釋放,使肌肉細胞活性的增加,所以肌細胞由負回饋機制所釋放的 IGF-1,對於肌
肉細胞與神經細胞的成熟過程是重要的因子;然而在神經-肌突觸結構建立之前,
IGF-1 對神經是否產生指引的作用,目前仍不清楚。因此,本實驗可以提供 IGF-1
對於運動神經胚胎發育時期是否促進神經-肌突觸的建立,以及對於神經損傷後
IGF-1 對神經修復再生及突觸重建的角色。

9
實驗材料
本實驗是以非洲爪蟾 (Xenopus laevis) 的運動神經元來進行,其製備方式主
要依據先前所發表的文獻報告(Spitzer and Lamborghini, 1976, Anderson et al.,
1977, Tabti and Poo, 1991) (附圖六)。 實驗過程簡略如下:取一對成熟雌、雄爪
蟾,分別在蛙體洩殖腔處施打性腺激素(Human Chorionic Gonadoyropin,HCG)
1.0 ml (500 U/ml),將其靜置於黑暗環境的水族箱中,使其自然行體外受精,第二
天 (約 24 小時左右) 收集受精卵,將其置於室溫下等待發育至 20-24 stage﹝約
一天左右;(Nieuwkoop and Faber, 1967)﹞,再進行以下步驟:
1. 消毒:
將 stage20-24 的胚胎以 75% 酒精消毒清洗數秒後,置於 10% Ringer’s
solution 中。
2. 去膜 :
在顯微鏡下以乾式滅菌器(Germinator 500)滅菌過後之鑷子將包覆胚胎外的透
明莢膜 (capsule)剝除。接著以玻璃管吸取胚胎轉置另一個 10% Ringer’s
solution 中清洗第二次, 過程中應避免胚胎接觸到空氣。
3. 剪下背脊神經與肌肉組織:
接著再以玻璃滴管將胚胎置於無鈣、鎂的 Ringer’s solution (calcium
magnesium-free Ringer, CMF)中,剪下背部神經管及其周邊的中胚層肌肉組織
後,靜置約 5 至 10 分鐘。
4. 脫皮:
此時胚胎表層皮膚已逐漸與組織分離,以鑷子將皮膚剝離,可加快剪下的組織
解離成較鬆散的細胞團塊,並降低皮膚細胞及黑色素細胞出現在 culture 的機
率,可減少干擾因素。
5. 靜置解離:

10
於 CMF solution 中靜置約 15 至 20 分鐘,等待組織解離成較鬆散的細胞團。
若是較成熟的胚胎(例如:可看得到背鰭),則需要更長的解離時間,約 30 至 40
分鐘。
6. 舖盤:
將解離成鬆散狀組織,以拉細的巴斯德玻璃管吸取,當組織通過拉細的玻璃管
尖端時有助於細胞團分散成單細胞,將這些細胞鋪於含有培養液的培養皿中,
於室溫(20-25℃) 下培養 4 小時候,即可得到爪蟾的神經細胞培養。
以上過程所需的溶液組成如下:
1. Ringer’s solution:
NaCl(115 mM), CaCl2(2 mM), KCl(2.5 mM), HEPES(10 mM), pH=7.6。
2. 無鈣、無鎂的 Ringer’s solution (Ca2+, Mg2+-free Ringer solution; CMF solution):
NaCl(115 mM), KCl(2.5 mM), EDTA(0.5 mM), HEPS(10 mM) pH=7.6。
3. 細胞培養液:
49% L-15 medium Leibovitz (Sigma), 49% Ringer’s solution, 1% fetal bovine
serum (Gibco),1% penicillin/streptomycin mixtures。
※ 以上的三種溶液皆須經過無菌過濾處理才能使用。
本實驗選擇 Xenopus laevis 之細胞培養,具有的優點如下:
1. 神經元的培養取得容易。
2. Xenopus laevis 細胞的發育階段特徵明顯且易做判斷。
3. 胚胎發育速度快,便於研究胚胎發育時期的神經細胞的特性。
4. 細胞培養可於顯微鏡下之開放空間及室溫中存活,而且不影響其生理特質,以
此進行 turning assay 極為理想。
5. Xenopus laevis 細胞本身即具有卵黃粒(yolk granules) 可供應養份,因此細胞

11
培養時只需要 1% 胎牛血清(fetal bovine serum) 即可培養細胞,較不易有汙
染的情形。

12
實驗方法
藥物前處理
本實驗的藥物會在實驗前 25 分鐘直接加入培養皿中,並且持續存在直到
實驗結束。唯有 Thapsigargin 需要在實驗前預處理 1 個小時,才能達到排空
細胞內儲存池中鈣離子的效果。
Growth cone turning assay
在室溫中培養 4 小時後神經細胞已經貼附於細胞培養皿底部,也是生長能
力最活躍的時刻,在倒立顯微鏡(TE2000-5, Nikon)下尋找適當神經細胞後,即
可進行實驗。本實驗主要是透過 turning assay 的方法,觀察細胞外神經滋養
因子造成神經生長錐偏轉的機制。依下列步驟,來建立神經細胞外環境的濃度
梯度差:
(1) micropipette 的製備:
實驗用的 micropipette,是以外徑 1.5 mm 之薄壁玻璃毛細管,以兩段式垂
直拉針器 (PC-10,Narishige) 拉成尖端內徑開口約 1 ~ 1.2 μm 的
micropipette,然後填入 IGF-1 10 μM 溶液、Ca2+-free IGF-1 10 μM,或
是經由過濾膜處理的 Ringer’s solution 作為控制組
(2) 神經細胞的選擇:
在顯微鏡視野下,選定一顆獨立生長的神經細胞,不能與其他神經細胞或
肌肉細胞形成突觸,之後將適當神經生長錐的頂點,置於電腦螢幕上虛設
的十字架中間(x=0,y=0),如此可以確保每次實驗神經細胞的初始位置是
相同的(附圖 5)。
(3) micropipette 的架設:
為了建立藥物有效的濃度梯度差,micropipette 的針尖設置在距離十字架
中心 100 μm,並且與十字架 X 軸夾 45 度角的位置;micropipette 的另一

13
端利用微調輔助系統(MS 314, WPI),將針尖盡可能調整置培養皿最底部,
使藥物形成平面的擴散作用。
(4) 空氣幫浦的設定:
我們經過多次實驗測試與參數的更改,micropipette 連接的電控式氣壓供
應系統的極佳注射條件為:每秒注射 1 次、每次注射 50 毫秒、空氣壓力 2
至 3 個 psi (pound per square inch),並且持續注射 70 分鐘至實驗結束(附
圖 5)。
影像處理
實驗前,在顯微鏡 side port 架設影像拍攝系統 CCD,攝影機另一端則傳
送影像至電腦的影像紀錄軟體(I spy);實驗中,我們每 30 秒擷取一張神經生
長錐的影像,總共紀錄時間為 70 分鐘;實驗後,拍攝的圖片集結於同一檔案
之中,待日後分析。
偏轉角度與生長速率分析
從集結的檔案資料夾中,選擇要分析的對照組或實驗組圖片,並選取單一
實驗所拍攝的第 1 分鐘及第 60 分鐘的影像,經由繪圖軟體 Adobe photoshop
(Ver 7.0, Adobe System Inc.)疊圖的功能,繪製神經生長錐初始位置和最終的
生長軌跡。
(1) 偏轉角度:
使用軟體(Image J),將 neurite 最終生長位置的頂點和實驗初始位置之頂
點連線,與 Y 軸之間所夾之角度,測量並紀錄之。
(2) 生長長度和速率:
使用 Adobe photoshop 軟體中的量測工具,沿著 neurite 的生長軌跡,從
實驗初始位置至最終的生長軌跡,測量並紀錄之;另外也透過時間的換算,

14
可以得到 neurite 的生長速率。
質體製備
1. pHluorin 質體來源為陽明大學,高閬仙老師實驗室。
2. 轉形作用
(1) 自-80℃冰箱拿出勝任細胞 DH5α後,立刻插冰上,等待 5 分鐘解凍
後取 1 μl 質體至菌管中,再用 tip 溫和地攪拌 3 圈,放冰上 30 分鐘。
(2) 使用Heat shock實驗原理,在攝氏42度加熱1分鐘,此時 competent
cell 脂雙層膜熱膨脹讓質體 DNA 進入,隨後立即插冰上 3 分鐘冷卻,
使脂雙層膜收縮以避免質體 DNA 流出;實驗前先預熱乾浴槽至 42
℃。
(3) 加入 250 μl Super optimal broth with catabolite repression (soc),
放入 37℃, 225 rpm 培養箱搖盪培養 4 個小時。
(4) 分別取 20、50、120 μl 菌液各別塗 ampicillin LA plate,並放入 37
℃培養箱培養 over night (16~18 小時),培養出單一菌落(single
colony)。
3. 菌株繼代培養
(1) pHluorin 所帶有的抗抗生素基因為抗 ampicillin,因此使用 ampicillin
LA plate 進行篩選。
(2) 將 plate 微微傾斜反面風乾,並在 plate 底部以奇異筆將其畫對半及
編號。
(3) 養菌環滅菌:養菌環沾酒精(97%)利用酒精燈燒紅等退溫(重複
3 次)。
(4) 挑菌環沾單一菌落,然後塗盤。

15
4. 小量與中量菌液培養
(1) 小量培養:3ml 的 LB 以及 3μl 抗生素加入菌管,放入 37℃、225 rpm
培養箱搖盪培養 4 個小時。
(2) 中量培養:將小量培養之菌液加入含有 250μl 抗生素的 250ml LB 錐
形瓶中放入培養箱,37℃、225 rpm 培養箱搖盪培養 16-18 個小時。
中量質體 DNA 萃取純化- Qiagen Plasmid Midi Kit
1. 將菌液平均分裝至50mL離心管中,以8000rpm、4℃離心15分鐘,之後倒
掉上清液。
2. 在離心管中加入buffer P1後pipetting總共4ml,再加入buffer P2後invert總
共4ml,直到顏色轉變成藍色,接著靜置室溫5分鐘。
3. 在離心管中加入buffer P3後invert總共4ml,直到白絲狀的染色體DNA出
現,接著置於冰上15分鐘。
4. 以8000rpm, 4℃離心30分鐘,之後抽取上清液至新管子中,再以8000rpm、
4℃離心15分鐘。
5. 取2管QIAGEN-tip架設於試管架上(下方擺放廢液杯),以4ml QBT潤洗
QIAGEN-tip。
6. 將上清液注入QIAGEN-tip中,之後以10ml QC buffer潤洗QIAGEN-tip兩
次。
7. 每管QIAGEN-tip加入5ml QF buffer沖洗,且下方必須接上新的15ml離心
管(將2管QIAGEN-tip收集於1管15ml離心管中)。
8. 加入3.5ml isopropanol並invert,接著離心30分鐘,之後倒掉上清液。
9. 加入4ml 70%酒精,接著離心10分鐘,之後倒掉上清液。
10. 倒置於試管架上,等待風乾。

16
11. 加入約200μl TE buffer並測定濃度。
12. 盡速分裝後,儲存於-20℃冰箱中。
神經細胞培養與轉染
(1) 消毒 :
取 15 隻 stage20 -24 的胚胎以 75% 酒精消毒清洗數秒後,置於 10%
Ringer’s solution 中。
(2) 去膜 :
在顯微鏡下以乾式滅菌器滅菌過後之鑷子將包覆胚胎外的透明莢膜剝除。
接著以玻璃管吸取胚胎轉置另一個 10% Ringer’s solution 中清洗第二次,
過程中應避免胚胎接觸到空氣。
(3) 剪下背脊神經與肌肉組織:
接著再以玻璃滴管將胚胎置於無鈣、鎂的 Ringer’s solution 中,剪下背部
神經管及其周邊的中胚層肌肉組織後,靜置約 5 至 10 分鐘。
(4) 脫皮:
此時胚胎表層皮膚已逐漸與組織分離,以鑷子將皮膚剝離,可加快剪下的
組織解離成較鬆散的細胞團塊,並降低皮膚細胞及黑色素細胞出現在
culture 的機率,可減少干擾因素。
(5) 細胞樣品混和:
將解離成團的細胞全部移至 eppendorf 中,使用低速離心機離心 5 秒鐘,
將所有細胞沉降至底部,之後將上清液(CMF)去除,並且加入 sterile 1X
PBS (25 μl)和 pHluorin plasmid (3 μl)。
(6) NeonTM儀器組裝
i. 將 NeonTM Tube 插入 NeonTM Pipette station 中,並加入 sterile 1X PBS
(3 ml)。

17
ii. 以NeonTM Tip吸取eppendorf中混合好的樣品,垂直插入NeonTM Tube
中。
(7) NeonTM系統設定
Electroporation操作條件為Voltage:650 V, Width:40 ms以及Pulses:
2 pulses。按 start 進行 electroporation,結束後 NeonTM 儀器會顯示
“Complete”,接著隨即將細胞樣品鋪於含有培養液的培養皿中(每盤約 5 條
spinal cord,共可鋪 3 盤),置於室溫(20-25℃)下培養一天,即可得到神經
細胞 overexpression pHluorin plasmid 的培養。
Image-Pro Plus 7.0螢光影像軟體之操作
於倒立顯微鏡(OLYMPUS IX70)下觀察帶有螢光的神經細胞,藉由顯微影
像軟體ImagePro® Plus中的Scope-Pro功能來控制Shutter開關,並可以隨著
細胞螢光亮度調整拍攝的曝光時間。我們透過縮時攝影(time lapse)的拍攝模
式,記錄單一神經細胞螢光變化,再利用該軟體的巨集程式(Macro)來定量分
析螢光亮度之變化(Density/Intensity)。
實驗數據統計分析
本實驗數據統計中,每單一實驗所計得神經生長軸若小於 5 μm,則捨棄此實
驗結果於統計之外 (Jin et al., 2005)。實驗數據以平均值±平均標準差
(mean±S.E.M)表示。而評估實驗組與對照組之間的差異,是根據 Mann-
whitney U-test 進行統計分析,以*p<0.05 代表有統計上的意義。統整匯集後
的數據利用軟體(SigmaPlot 10.0),來繪製生長速率和偏轉角度的圖表。
實驗用試劑及供應商
購自 Sigma:

18
﹝3,4,5-Trimethoxybenzoic acid-8-(Diethylamino)Octyl Ester﹞(TMB-8) 、
Thapsigargin、N6,2’-O-Dibutyryladenosine 3’,5’-cyclic monophosphate
sodium salt (cAMP)、U0126、Gadolinium (III) chlordie (Gd3+)、Cadmium
chloride-1-hydrate (Cd2+)
購自 Torcis :
Wortmannin、PD98059、Genistein、U73122
購自 abcam:
Xestospongin (XeC)、SKF 96365 hydrochloride
購自 Bio-Genesis Technologies:
Insulin-like growth factor I (IGF-1)

19
結果
一、建立壓力給藥形成濃度梯度的模式
根據過去的研究,關於神經偏轉的實驗有許多外給藥物的條件,不論是注
射的頻率、壓力、注射的時間,或是針尖口徑的大小都不盡相同(Naeve et al.,
2000, Weinl et al., 2003, Meriane et al., 2004, Joanne Wang et al., 2008,
Merianda et al., 2009, Han et al., 2011),由於這些條件會影響注射藥物濃度
梯度的建立,甚至壓力過大也會造成對神經生長點的 stress。我們實驗的第一
個步,就是要找到合適的注射條件。首先以一種化學螢光物質 FITC 裝填於
micropipette 中,由於此螢光物質透過藍光的激發會呈綠色螢光,因而極容易
在螢光顯微鏡之下紀錄 FITC 的螢光變化(Figure 1A),針尖距離神經生長點
100 μm 處並與水平 X 軸夾 45˚的位置。藉由測量螢光的強度,得知針尖開口
呈現最高的螢光數值,隨著遠離針尖開口的位置濃度逐漸下降,在多次實驗的
觀察下,發現注射 10 分鐘後距離針尖 100 μm 位置,螢光表現可以形成有效
的濃度梯度﹝約 15.6±1.7 倍稀釋 (n=4)﹞,直到約 30 分鐘後達到飽和濃度,
(Figure 1B)。
經過一連串不斷的測試和更改參數,我們最終找到極佳的注射條件為針尖
內徑開口約 1~1.2 μm、壓力 3 psi、每次注射 50 毫秒,每秒注射 1 次持續 70
分鐘,在這種情況之下每次藥物注射的體積大約增加 9.99±2.44 nl ,n=9,只占
細胞培養液(3 ml)中 0.1%的體積。接下來,我們首先驗證整套實驗注射系統運
作的可行性,利用 Ringer’s solution 作為 negative control,從實驗結果顯示
(Figure 2A),經過 1 小時的注射,Ringer’s solution 的平均偏轉角度與 control
相比沒有顯著性的差異(分別是-4.73˚±8.63˚,n=17 及-4.18˚±9.78˚,n=11),由此
可知,在壓力給藥形成濃度梯度的模式情況下,壓力並不會影響神經細胞的正
常生長。
另外在 positive control 中,我們選擇過去研究已經被證實是一種

20
chemoattraction 的化學物質 cAMP (Lohof et al., 1992, Nieto-Estevez et al.,
2016),結果顯示經過 1 小時的實驗,由 Figure 2B 可以看出整體實驗結果的
趨勢,cAMP 與 control 神經生長軌跡相比,cAMP 平均偏轉角度比 control 大
(分別是 19.37˚±8.99˚,n =12 及-4.18˚±9.78˚,n=11;*p<0.05, Mann-whitney U
test),確實 cAMP 明顯促進神經生長的偏轉,因此表示以上的注射條件是能夠
成功產生有效的濃度梯度和引導作用。
二、IGF-1 對神經生長與偏轉的影響
當突觸結構發生損壞或失去支配時,肌細胞為了在與肌細胞建立連結,所
以又會表現高濃度的 IGFs 在肌細胞中(Ishii, 1989, Caroni, 1993)。由此可知
肌細胞內 IGF-1 的表現量與神經細胞的連結關係緊密。因此由以上證據得到一
個暗示:IGF-1 應該在胚胎發育過程中可能會回饋於神經細胞;不過是否吸引
神經生長的作用,讓神經-肌細胞間的關係更緊密呢?為了瞭解神經碰觸到肌
細胞前,肌細胞釋放的滋養因子如何造成神經轉向的分子機制,我們經由 axon
guidance 的方法,紀錄神經生長過程中,指引因子會造成神經吸引或排斥的
作用,整體的神經生長軌跡可以觀察神經生長的趨勢,作為判斷此因子吸引與
否的重要指標。實際生長軌跡紀錄結果如 Figure 3B 所示,我們發現神經注射
IGF-1 其 偏 轉 角 度 明 顯 比 注 射 Ringer’s solution 的 大 ( 分 別 是
16.69˚±6.25˚,n=22 及-4.73˚±8.63˚,n=17;*p<0.05;Figure 3D),畫成累積角
度百分比的分佈曲線時亦可看出 IGF-1 的曲線明顯較 Ringer’s solution 右移
(Figure 3E),若觀察生長長度,也清楚看出注射 IGF-1 所測得的平均生長速率
顯著比 Ringer’s solution 來的快 (分別是 0.45±0.05 μm min-1,n=22 及
0.26±0.04 μm min-1,n=17;Figure 3F;#p<0.05, Student’s t test),這顯示 IGF-
1 可能是一種神經生長的指引因子。
由於先前的觀察發現神經細胞培養 4 小時後,IGF-1 能夠發揮神經指引的

21
效果,於是進一步瞭解神經在不同發育階段對 IGF-1 之間的導引情形。我們同
樣以神經的偏轉角度來判定 neurite 產生吸引與否。比較 IGF-1 對於舖盤後
4~10 小時及 1 天的神經細胞其偏轉角度,發現 1 天發育階段的神經,IGF-1
對神經生長速率已經沒有顯著的促進作用(由 0.45±0.05 μm min-1,n=22 減少
至 0.36±0.05 μm min-1,n=10;Figure 4F),而對神經偏轉作用也明顯比 4~10
小時的神經來的小(由 16.69˚±6.25˚,n=22 下降至 5.47˚±8.20˚,n=10;Figure
4D),所以往後的研究主要鎖定 4~10 小時的神經來進行實驗。
三、IGF-1 對神經的偏轉作用與鈣離子有關
由於神經細胞骨架 actin 和 microtubule 具有動態平衡(dynamics)的特性
(Vitriol and Zheng, 2012),透過鈣離子與 actin filaments 之間交互作用(Tojima
and Kamiguchi, 2015),來調控神經軸突長度的增加或減少響(Schaefer et al.,
2008, Conde and Caceres, 2009),因此在瞭解神經細胞內 IGF-1 的訊息傳遞
路徑之前,我們先闡明鈣離子的來源對神經偏轉的影響。首先在培養 4~10 小
時爪蟾之神經,藉由 turning assay 方法來觀察阻斷神經胞外鈣離子來源對
IGF-1 指引作用的情形。
當我們分別將 culture medium 置換成無鈣離子的 Ringer’s solution (Ca2+
free Ringer)或加入 0.5 mM Cd2+(一種細胞膜上鈣離子通道阻斷劑)預先處理
25 分鐘之後, IGF-1 所造成神經生長軸偏轉角度的現象被抑制了 (從
16.69˚±6.25˚,n=22 分別降至 Ca2+ free 組的-5.22˚±9.57˚,n=13 及 Cd2+組的
1.55˚±11.20˚,n=9;Figure 5E);若從單一實驗結果得到的角度分佈位置圖
(Figure 5D)中亦可發現,不論是外界環境無鈣離子的供應與廣效性鈣離子通道
阻斷劑 Cd2+的存在下,都改變原本 IGF-1 對神經偏轉吸引的作用;同樣的結
果在(Figure 5F)也可以清楚的看出整體神經轉向的趨勢,因受到阻斷胞外鈣離
子的來源而影響。另外,針對神經的回縮率和死亡率進行分析,發現不論是置

22
換成 Ca2+ free Ringer 或 Cd2+預處理情況下,與 control 相比都有極高的死亡
率(從 control 組的 31%, n=32 分別提升至 Ca2+ free 組的 70%, n=44 和 Cd2+
組的 67%, n=27),以上結果都顯示,細胞外鈣離子對神經的偏轉和生長確實
具有不可或缺的必要性。
為了瞭解其它鈣來子來源對神經指引的現象,我們進一步觀察細胞內鈣離
子儲存池所扮演的角色。我們利用 thapsigargin (TG)做預處理 1 個小時,TG
為一種細胞內鈣離子儲存池排空劑,會導致鈣離子無法回收至儲存池中,使細
胞質內的鈣離子濃度提升,而後因為濃度梯度差驅使鈣離子擴散至胞外
(Berzat and Hall, 2010, Tojima et al., 2014);結果顯示,投予 2 μM TG 的神
經,在 1 小時的平均生長速率沒有太大的影響(由 0.45±0.05 μm min-1,n=22 減
少至 0.33±0.08 μm min-1,n=6;Figure 6F),不過在平均偏轉角度上,神經生
長錐已經顯著的不再對 IGF-1 產生吸引作用(從 16.69˚±6.25˚,n=22 下降至-
11.41˚±6.60˚,n=6;Figure 6D;*p<0.05);有趣的事,同樣進行死亡率的分析
(63%, n=16)時,得到與抑制胞外鈣離子來源情況相似的結果。據此推測由 IGF-
1 產生的神經偏轉過程,細胞外或細胞內儲存池中的鈣離子都參與在其中,因
此阻斷其中一側鈣離子的來源,就無法維持原本神經的生長與偏轉。
四、IGF-1 對神經偏轉作用與囊泡釋放有關
神經錐生長過程中細胞骨架屬於動態平衡的狀態,而造成細胞膜外形時而
凸出時而縮減,因此一些研究認為造成神經細胞偏轉,可能與細胞膜的
exocytosis 和 endocytosis 囊泡運輸機制有關(Tojima et al., 2014, Ros et al.,
2015),因為許多證據顯示細胞骨架和黏附物質的動態變化,必須透過
exocytosis 和 endocytosis 進行囊泡的運送(Itofusa and Kamiguchi, 2011,
Tojima et al., 2011, Vitriol and Zheng, 2012),導致細胞膜的延伸或收縮。先
前我們的研究也顯示,IGF-1 可以促進囊泡包覆的神經傳遞物質釋放(Liou et

23
al., 2003)。而且近年已經有研究利用 pHluorin probes 的方式,來觀察神經生
長過程中單一囊泡釋放的過程(Tojima et al., 2007, Burgo et al., 2012)。
為了研究 IGF-1對神經的偏轉作用與囊泡的 exocytosis 或 endocytosis 的
相關性,因此我們藉由細胞轉染的技術,將這種酸鹼值敏感型的質體─pHluroin,
轉染進入神經細胞中並 overexpression overnight,方便我們在螢光顯微鏡輔
助下,觀察 IGF-1 對神經產生偏轉作用時,細胞內囊泡運輸機制的變化。由
(Figure 7A-B)分別可以透過白光觀察神經在尚未注射 IGF-1 前的型態,以及利
用螢光顯微鏡所發出的藍色波段激發光,來觀察 pHluorin 在神經細胞中表現
的 pattern;接著同樣利用 axon guidance 實驗方法給予 IGF-1 的注射,大約
在注射後 20 分鐘發現螢光 pattern 分佈有了改變(Figure 7C),由注射前和注
射後的結果圖比較,會發現經由壓力注射 IGF-1 的情況下,藥物漸漸在神經生
長錐前端形成濃度梯度,此時 IGF-1 與生長錐上的 IGF-1 受器結合產生作用,
藉由下由訊息傳遞路徑的活化,造成細胞內囊泡開始移動,因此可以發現當神
經以 Y 軸區分為左有兩側時,在注射面的這一側螢光表現 pattern 較明亮。從
以上證據可以推論,IGF-1 對神經產生偏轉的作用,確實是藉由細胞內囊泡的
釋放過程,導致細胞膜向外延伸。
五、阻斷胞內鈣離子儲存池 IP3受體影響 IGF-1 對神經吸引偏轉作用
由以上實驗我們可以得知,在爪蟾神經、肌肉培養中,不同來源的鈣離子
對 IGF-1 產生的神經生長錐轉向過程極為重要。由於各種來源的鈣離子受到不
同訊息傳遞路的調節,因此我們接下來釐清胞內鈣離子儲存池膜上不同受體所
調控鈣離子的釋放對 IGF-1 所造成神經偏轉的相關性。在細胞內鈣離子儲存池
的釋放主要經由兩種途徑:一種是 ryanodine receptor (RyR) (Tsien et al.,
1988),另一種為 inositol 1,4,5-trisphosphate receptor (IP3R) (Ros et al., 2015);
所以我們分別預處理 RyR 的抑制劑 TMB-8,以及 IP3R 的抑制劑 XeC,並等

24
待 25 分鐘的藥物作用,之後進行 turning assay 實驗分析。
觀察 IGF-1 所造成神經偏轉作用的情形,發現在事前給予 3 μM TMB-8 的
情況下,雖然 1 小時的平均生長速率相較 control 緩慢許多(0.26±0.03 μm min-
1,n=8;Figure 8F),但並不影響 IGF-1 對於神經轉向,甚至比 control 有更強
的吸引效果(43.22˚±7.97˚,n=8;Fig 8.D;*p<0.05);除此之外,從單一實驗角
度分佈結果圖(Figure 8C),亦可發現各個神經最終生長位置都趨向 IGF-1 注
射的這一側,顯示阻斷細胞內鈣離子儲存池膜上的 RyR,與 IGF-1 造成的神
經偏轉作用係沒有相關性。
由於 TMB-8 對 RyR 的抑制,並不會造成神經偏轉作用的改變,因此我們
的焦點轉向另外一個調控細胞內鈣離子儲存池的孔道,進一步探討 IP3R 所釋
放的鈣離子與 IGF-1誘導神經偏轉的關聯。從圖(Figure 9B右)顯示了所有 1μM
XeC 預處理的神經生長軌跡,發現經過 XeC 處理之後,其偏轉趨勢與 control
相比明顯轉向排斥的一側(-4.54˚±6.89˚,n=9;Figure 9D;*p<0.05),而 1 小時
的平均生長速率,相較於 control 也有顯著性的降低(由 0.45±0.05 μm min-
1,n=22 減少至 0.29±0.05 μm min-1,n=9;Figure 9F;#p<0.05)。綜合以上結
果,可以推論 IGF-1 造成神經轉向的作用,是經由 IP3 receptor 釋放的鈣所導
致,而與鈣離子儲存池 RyR receptor 無關。
六、IGF-1 對神經的偏轉作用與活化 Tyrosine kinase receptor 有關
根據過去的研究,已知 IGF-1 受體是一種 tyrosine kinase receptor,透過
許多的 phosphorylation site 可以形成專一的 docking site,藉此得到許多
insulin receptor substrates (IRS)-1 及-2,透過與 β subunit 上 docking site 的
結合使 tyrosine residue 被磷酸化,使特異性的結合位能夠與蛋白接合而活化
(Butler et al., 1998, Adams et al., 2000, Van Obberghen et al., 2001),開啟
像是 Phospholipase Cγ (PLCγ)、phosphoinositide 3’-kinase(PI 3-kinase)以及

25
mitogen-activated protein kinase (MAPK)等一連串訊息傳遞路徑。
在闡明神經細胞內 IGF-1 下游的分子機制,對神經生長錐產生的偏轉作用
前,我們先瞭解 IGF-1 與細胞膜上 IGF-1 受體的結合,其受體活化與否的影
響。因此我們在爪蟾神經細胞培養中,投予 tyrosine kinase 的抑制劑-Genistein,
利用 turning assay 方法進行實驗分析。發現事前預處理 25 分鐘的 10 μM
Genistein 時,的確減少 IGF-1 對神經的偏轉作用,圖(Fig 10.B)中每個單一的
Genistein 生長軌跡中,清楚看出神經不再受到 IGF-1 吸引。畫成生長最終的
角度偏轉分布圖(Fig 10.C),發現整體的分佈趨勢不在吸引的一側,另外從累
積角度百分率的分佈曲線分析,看出曲線明顯的向左移(由 16.69˚±6.25˚,n=22
減少至-2.72˚±5.84˚,n=12;Figure 10E)。由上述結果確認,IGF-1 會透過與 IGF-
1 受體的交互作用,進一步誘發 IGF-1 受體活化,造成 IGF-1 對神經產生的偏
轉作用。
七、IGF-1 對神經偏轉作用的分子機制
由於先前的實驗發現在爪蟾神經細胞培養中,當我們投予細胞內鈣離子儲
存池 IP3R 的 XeC 抑制劑時,阻斷鈣離子的釋放會造成 IGF-1 對神經產生的吸
引效果減低,而文獻中得知 IP3的產生與 PLCγ的活化有關,是細胞內鈣離子
濃度上升的重要因子(Majewska et al., 2000, Richards et al., 2004)。接下來的
實驗我們使用 turning assay 的方法,來探討 IGF-1 對神經偏轉作用與 PLCγ
路徑的關係,因此預處理 PLCγ的抑制劑 5 μM U73122 於 bath 中,並等待 25
分鐘的反應時間。由圖(Figure 11B 右)可以瞭解所有 PLCγ預處理的神經生長
軌跡,發現 IGF-1 對神經偏轉作用明顯被抑制;同樣的趨勢,亦可觀察神經生
長 的 柱 狀 統 計 圖 ( 偏 轉 角 度 由 control : 16.69˚±6.25˚,n=22 減 少 至 -
13.13˚±10.58˚,n=10;Figure 11D;*p<0.05),從生長速率分析,同樣也造成顯
著的抑制效果(由 0.45±0.05 μm min-1,n=22減少至 0.28±0.04 μm min-1,n=10;

26
*p<0.05)。顯示 PLCγ路徑的活化與否,參與 IGF-1 對神經偏轉的作用,而且
推測被抑制的 PLCγ 路徑會影響鈣離子的釋放,導致神經的生長速率因 actin
的 polymerlziation 減少而減緩。
由於 PLCγ的訊息傳遞路徑受到 PI-3 kinase 所調控,PLCγ的 pleckstrin
homology (PH) domain 與 phosphatidylinositol 3,4,5-trisphosphate(PIP3),會
藉由 PI-3 kinase 的結合 (Oertner and Matus, 2005),引發 PLCγ 水解
phospholipide 的產物,使 IP3 及 diaclglycerol (DAG)的生成。為了瞭解 PI-3
kinase 所調控的 PLCγ 路徑是否與 IGF-1 對神經偏轉作用有關,因此我們在
bath 中預處理 PI-3 kinase 的抑制 100 nM wortmannin,觀察 IGF-1 對 neurite
生長轉向情形。從(Figure 12B 右)的生長軌跡累計圖可以知道神經生長偏轉在
加入 wortmannin 之後,偏轉趨勢已經不在朝向 IGF-1 注射的一側;由統計分
析柱狀圖中,亦可以發現顯著性的差異(0.47˚±6.83˚,n=11;Figure 12D;
*p<0.05)。另外在 1 小時的平均生長速率分析,發現與 control 相比也顯著的
減緩(control 組的 0.45±0.05 μm min-1,n=22;wortmannin 處理的 0.25±0.02
μm min-1,n=11;Figure 12F;*p<0.05)。綜合以上結果,可以推測 IGF-1 對神
經的偏轉作用,乃是透過 PI 3-kinase 刺激 PLCγ的水解,使細胞內鈣離子儲
存池 IP3通道開啟,進而釋放鈣離子至細胞質中。
八、阻斷 MAP Kinase 路徑造成 IGF-1 對神經偏轉作用下降
除了 PI-3 kinase 和 PLCγ路徑之外,細胞內另一條 IGF-1 的下游訊息傳
遞路徑為 MAP Kinase 路徑,在過去一些文獻中提及,活化 MAP kinase 的路
徑有助於啟動神經生長錐生長機制(Markus et al., 2002)與促進 neurite 向外延
伸(Caroni, 1993, Kim et al., 1997)。為了進一步瞭解 MAP Kinase 是否參與
IGF-1 的神經偏轉作用,我們在爪蟾神經培養中,分別預處理 25 分鐘的 10 μM
PD98059 和 10 μM U0126 來阻斷 MAP Kinase 的活性。由結果(PD98059:-

27
14.01˚±8.39˚,n=17;U0126:5.59˚±13.58˚,n=10;Figure 13D)中可以發現神
經收吸引的偏轉角度有所改變,甚至在預處理 PD98059 後偏轉顯著減少,亦
可在統計柱狀圖 Figure 13E 中看出大部分神經軸突轉向不吸引的一方;在生
長速率方面,神經也確實因為受到訊息傳遞路徑的抑制而減緩(由 control 組的
0.45±0.05 μm min-1,n=22;減緩至 PD98059 組的 0.26±0.03 μm min-1,n=17;
U0126 組的 0.38±0.06 μm min-1,n=10;Figure 13F)。所以得知,當抑制 MAP
Kinase 的路徑時,會影響原本神經細胞的增生速度,同時也減少 IGF-1 對神
經產生的吸引效果。
利用 Figure 14 的統計柱狀圖,將 IGF-1 對神經產生偏轉作用的分子機制
進行分析;可以推論,當細胞外 IGF-1 指引因子與神經細胞膜上 IGF-1 受體結
合,產生交互作用時,會使 tyrosine kinase 活化,藉此引發 IGF-1 下游訊息傳
遞路徑 PI-3 kinase 的活性來刺激 PLCγ 的水解,透過水解的產物 IP3 使細胞
內鈣離子儲存池 IP3 channel 活化而開啟,進而釋放鈣離子至細胞質中,以利
提供更多鈣離子與 actin 進行 polymerization。
九、阻斷 TRP channels 造成 IGF-1 對神經偏轉作用下降
TRP channels 具有許多結構異構物,主要表現於神經系統中,對於神經
生長錐的吸引或排斥以及鈣離子濃度的調控具有相當廣泛的影響(Werther et
al., 1990, Li et al., 2005),當神經生長錐上的 TRP channels 活化後,使胞外
鈣離進入細胞中,進而產生神經偏轉作用、神經軸突的延伸或神經型態上的改
變(Naeve et al., 2000, Nieto-Estevez et al., 2016)。為了瞭解 IGF-1 對神經造
成偏轉與 TRP channels 活化之間的關係,所以我們使用兩種廣效性的 TRP
channels 抑制劑,分別為 25 μM SKF96365 及 10 μM Gd3+,在爪蟾神經培養
中預處理 25 分鐘。

28
由結果發現,抑制神經細胞膜上的 TRP channels 導致 IGF-1 對神經吸引作用
減少﹝(SKF96365:-22.98˚±10.69˚,n=5;Figure 15D);(Gd3+:-6.55˚±12.24˚,n=10;
Figure 16D)﹞,從累積角度百分率分佈曲線(Figure 15C, 16C),亦可看出處理組
的曲線向左偏移的趨勢。由上述結果得知,TRP channels 受到阻斷後,流入足夠
的鈣離子提供神經的生長,而 IGF-1 所引起的神經偏轉作用也將跟著消失;因此
在本實驗中,細胞膜上 TRP channles 活化與否,所供給的細胞外鈣離子來源,對
於神經的延伸或偏轉相當關鍵。

29
討論
生物的胚胎發育過程中,神經細胞如何與標的細胞建立完整的結構呢?早在
1970 年代,由於 in vitro 的實驗方法蓬勃發展,因此有利於科學家方便觀察各種細
胞的實驗,其中也包含神經細胞與肌細胞間 -肌突觸的建立 (Spitzer and
Lamborghini, 1976, Anderson et al., 1977)。直到 1980 年代,科學家在神經細胞
培養的觀察,他們認為神經生長錐需要透過細胞外環境中,其他鄰近的細胞提供化
學性或物理性的因子像是 chemotactic substances, mechanical barriers,
substratum, adhesiveness,以及 selective affinity 來誘導神經的生長(Lockerbie,
1987, Bray and Hollenbeck, 1988)。神經細胞生長發育過程中猶如雷達衛星一般,
不斷的四處搜尋周遭環境各種因子,因為他們仰賴多樣化的指引因子,來影響神經
生長錐的偏轉,有些能夠產生神經吸引作用(attractive)但有些可能造成神經排斥作
用 (repulsive),最終都是為了與標的細胞達成聯結建立 (TessierLavigne and
Goodman, 1996, Song et al., 1998);乃至於神經受損後,對於神經的再生過程中,
也需要藉由相似的機制進行修復。
為了瞭解神經指引因子對神經生長過程的重要性,所以科學家開始藉由空氣
幫浦輔助下,注射特定因子形成濃度梯度的模式(Gundersen and Barrett, 1980),
來取代過去使用擴散速率的給藥方式。在我們的預實驗中,也嘗試使用擴散速率的
實驗方法給藥,但經過多次實驗後,因為考慮藥物可能針尖口徑太小以及無法確保
每次實驗藥物都能順利擴散至神經生長點,最後選擇以空氣壓力幫浦輔助注射藥
物;Lohof 等人開始在 in vitro 情況下建立一套藥物注射系統(Lohof et al., 1992),
整套系統中不論是針尖口徑大小、針尖架設的位置、注射的壓力大小、注射的時間
長短,以及注射的頻率等,許多變數都會造成藥物濃度梯的改變。先前的實驗結果,
我們也經過多次參數的更改,才找到極佳的注射條件;我們選擇針尖開口約 1~1.2
μm,針尖距離神經生長點 100 μm 處並與水平 X 軸夾 45˚的位置,給予 1 Hz 每次

30
50 ms 的 100 psi 注射條件情況下,大約在注射時間 10 分鐘可以形成濃度梯度差,
且在注射後 30 分鐘到達穩定的飽和濃度。以上條件確立的過程中,我們同時也使
用 Ringer’s solution 和 cAMP 分別做為 negative 和 positive control,可以確保我
們使用的參數,不影響正常神經的生長。
在神經支配肌細胞所形成突觸的特殊結構中,肌肉細胞的活性可能受到本身
細胞內某些基因的轉譯所調節,另外也受神經細胞活性所調控,所以肌細胞可能因
為活性的改變,而釋放某些滋養因子扮演回饋(feedback)角色來影響神經的功能
(Eftimie et al., 1991),然而這些滋養因子像是 BDNF 和 NT-3 同時也認為具有吸引
神經生長和促進神經的再生作用(Xu et al., 1995, Genc et al., 2004);目前研究中
已經發現一些神經指引因子像是netrins family, semaphorins family, ephrins family
以及 slits family (Winberg et al., 1998, Brose and Tessier-Lavigne, 2000, Rohm
et al., 2000, Guan and Rao, 2003, Egea and Klein, 2007, Hale and Coward, 2013),
在神經生長過程中具有吸引神經偏轉的作用,亦針對其下游訊息傳遞路徑的分子
機制廣泛研究。由於相似的作用機轉也同樣發生在本研主探討的 IGF-1,本實驗室
的研究中發現在爪蟾肌肉細胞所釋放的神經滋養因子 IGF-1,能夠逆向的做用於神
經細胞上 IGF-1 受器,導致神經傳遞物質釋放量增加,進而促進肌細胞的活性(Liou
et al., 2003)。此外,IGF-1 在肌肉細胞發育過程會大量產生,而隨著細胞逐漸的
成熟 IGF-1 的濃度則會呈現遞減狀態,一旦突觸結構發生損壞或失去支配時,肌
細胞為了在與神經細胞建立連結,所以又會表現高濃度的 IGFs 在肌細胞中(Ishii,
1989, Caroni, 1993)。所以接下來本實驗透過空氣幫浦輔助下注射 IGF-1,使 IGF-
1 在 culture medium 中建立濃度梯度,結果發現神經生長發育過程中,的確造成
神經吸引偏轉的現象,而且分析生長速率與control相比也有顯著的增加,顯示 IGF-
1 透過與神經細胞膜上的 IGF-1 受體結合並產生交互作用後,誘導下游訊息傳遞
路的活化,因此得知 IGF-1 不但可以促進神經-肌細胞的活性,也有助於吸引神經
的偏轉作用和神經的生長速率。
由上述結果瞭解 IGF-1 能夠吸引神經偏轉的作用,接著觀神經細胞在發育不

31
同階段 IGF-1 對神經細胞會產生什麼樣的影響?過去文獻提及本實驗使用的爪蟾
神經細胞,在 primary culture 後 2~3 小時,大部份神經細胞與 dish 間已經貼附完
好(Wen et al., 2004),且 neurite 開始向外生長,表示此刻的神經軸突具有極佳的
能力向外延展。因此相較於鋪盤後 1 天的神經細胞培養,不但神經細胞對 IGF-1 吸
引效果不明顯而且生長速率也相對緩慢;所以本篇研究主要鎖定 culture 後 4~10
小時的神經細胞培養進行實驗,這個區間的神經細胞具有較高的生長活性,亦在
IGF-1 注射情況下也具有較強的吸引偏轉效果。
確立了 IGF-1 對神經造成偏轉的吸引現象後,我們更好奇此過程的分子機制
又是如何產生呢?或許可以透過觀察鈣離子來源的實驗瞭解其中的變化。在一些
抑制鈣離子來源的神經偏轉實驗中發現,神經軸突的延伸與收縮與細胞質的鈣離
子濃度息息相關(Naeve et al., 2000, Hutchins et al., 2012),由於細胞骨架動態平
衡的改變過程中,不論 actin 或 microtubule 都需要透過與鈣離子的交互作用,才
能進行 polymerization 或 depolymerization 使細胞骨架的長度產生改變,進而造
成細胞的移動(Yu and Bargmann, 2001, Gallo and Letourneau, 2004)。在我們置
換成無鈣離子的 Ringer’s solution (Ca2+-free Ringer)或是投予 Cd2+來抑制細胞外
鈣離子來源的實驗中發現,不但減少 IGF-1 對神經細胞的吸引能力,在神經收縮
或死亡率也相當高,顯示抑制細胞外鈣離子來源,導致鈣離子無法供給細胞質內充
足的鈣離子濃度,因此對神經細胞生長有所影響。除此之外,瞭解了細胞外鈣離子
來源的作用之後,在本實驗中我們亦可發現即使利用 TG 排空細胞內鈣離子儲存
池的鈣離子,IGF-1 對神經細胞的指引作用也受限制,不再產生吸引偏轉的效果。
我們的發現就如同一些文獻報導,神經生長錐最終產生的吸引或排斥反應,會因為
下游訊息同時受到 intracellular 或 extracellular 鈣離子濃度的變化而影響(Zheng
et al., 1994, Song et al., 1998, Nieto-Estevez et al., 2016)。
然而,近年來有些科學家認為,神經生長錐向外延伸的機制,不單只是透過鈣
離子與 actin 或 microtubule 的交互作用所產生,因為研究顯示造成細胞膜外形突
出或縮減,可能與細胞膜的 exocytosis 和 endocytosis 囊泡運輸機制相關(Tojima

32
et al., 2014, Ros et al., 2015),而且發現細胞骨架和黏附物質的移動過程中,必須
透過 exocytosis 和 endocytosis 進行囊泡運送(Itofusa and Kamiguchi, 2011,
Tojima et al., 2011, Vitriol and Zheng, 2012),許多研究也發現,當神經走向神經
指引因子的一側時需要 exocytosis,反之當神經生長錐崩解並走向排斥一方時,主
要依靠 endocytosis (Cotrufo et al., 2011, Cotrufo et al., 2012, Zylbersztejn et al.,
2012)。此外,近年也有一些神經生長錐的研究,利用囊泡分泌過程中酸鹼值產生
變化的特性,因此在神經細胞轉染培養中表現一種 pH 敏感型的探針-pHluorin,使
囊泡 VAMP2 的 C terminus 與 pHluorin 結合,藉此觀察 pHluorin 在細胞螢光表現
的特性(Miesenbock et al., 1998),來探討局部細胞膜動態平衡的改變與囊泡釋放
的過程中,pHluorin 螢光表現量的變化(Tojima et al., 2007, Burgo et al., 2012,
Nakazawa et al., 2012)。當我們同樣利用 pHluorin probe 轉染至神經細胞中進行
IGF-1 turning assay 實驗,結果中發現注射 IGF-1 所造成神經偏轉作用的過程,
細胞局部 pHluorin 螢光表現量增加,而這也表示 IGF-1 作用於神經生長錐膜上
IGF-1 受器後,引起下游訊息傳遞路徑的活化,促使細胞骨架和黏附因子開始經由
囊泡的運輸,並朝向注射的一方堆積。
瞭解 IGF-1 對神經所造成的偏轉作用中,細胞骨架動態平衡的鈣離子來源後,
我們進一步探討 IGF-1 對神經所造成偏轉現象,誘發的訊息傳遞作用機轉。實驗
設計是透過實驗前,預處理不同訊息傳遞路徑的抑制於 culture medium 中,再以
turning assay 方法觀察。首先,從文獻中已知神經細胞膜上 IGF-1R為一種 tyrosine
kinase receptor (Lemmon and Schlessinger, 2010),所以我們觀察藥物抑制
tyrosine kinase 的活性之下產生的影響,結果發現 IGF-1 所造成的神經偏轉作用
受到抑制,因此我們推斷在 IGF-1 吸引神經生長的作用中,IGF-1 確實與神經細胞
膜上的 IGF-1R 結合產生交互反應。再來,為了進一步瞭解 IGF-1R 活化後啟動的
下游路徑機制,因此我們參考文獻得知 IGF-1R 活化後,目前已知主要的三條路徑
分別為 PLCγ, PI-3 kinase 和 MAP Kinase (Foncea et al., 1997, Kim et al., 1997,
Leski et al., 2000, Hong et al., 2001, Mehrhof et al., 2001),而上述文獻中也顯示,

33
在不同類型細胞的 IGF-1 可能藉由不同訊息傳遞路徑,活化其下游轉錄因子,造
成不同生理反應。因此在本研究中分別處理這三條路徑的抑制劑,來探討 IGF-1 在
神經偏轉作中的訊息傳遞過程。由實驗結果顯示,這三條路徑都與 IGF-1 所產生
偏轉作用有關,但其中 PI-3 kinase 的下游產物會藉由 PLC 促進 phospholipid
phosphatidylinositol 4,5-bisphosphate (PIP2)水解成 diacyl glycerol (DAG)和
inositol 1,4,5-trisphosphate (IP3) (Hilgemann, 2007),所以當我們抑制 PLC 路徑
後,也影響原本 IGF-1 所產生的神經偏轉現象。
從以上結果推論,當神經細胞外 IGF-1與細胞膜上 IGF-1受器結合,使 tyrosine
kinase 的活化,便會啟動下游一連串訊息傳遞路徑的反應,經由 PI-3 kinase 的活
性來刺激 PIP2 的水解導致 IP3 產生,此時 IP3 就能夠與細胞內鈣離子儲存池膜上
IP3 receptor 結合,最後造成 IP3R 活化;也有實驗證實在大鼠大腦的 granule 細胞
中,藉由 BNDF 吸引神經的偏轉作用並活化其細胞膜 BNDF 受器,而啟動的下游
訊息傳遞路徑最終活化 IP3R,但與 RyR 活化與否無關(Li et al., 2005)。在本實驗
中也發現相同的結果,甚至當我們預處理 RyR 的抑制劑後,IGF-1 對神經所產生
的偏轉作用明顯增加,推測可能與 IP3R 活化細胞膜上的 TRP channels 有關,使
細胞外鈣離子流入,進而增加細胞質鈣離子濃度。過去在 signal channel 研究中發
現,在 IP3R 與 TRP channels 之間可相互影響,而且兩者間複合連結在一起
(Kiselyov et al., 1998);另外近年在 Yuan 等人 2003 年的研究發現,細胞膜上 TRP
channels 和 IP3R 之間會與 adapter protein-termed Homer 形成複合體,透過物理
性的交互作用促進 TRP channel 的反應產生(Yuan et al., 2003)。所以細胞質鈣離
子濃度的提升,可能是藉由 PLCγ的產物 IP3活化下游訊息傳遞路徑,並透過 IP3R
與 TRP channels 作用,使鈣離子流入(Meriane et al., 2004, Li et al., 2005)。
在神經細胞發育過程仰賴許多細胞外的因子,提供神經生長的指引與向外延
伸的訊號,且對於 axon initiation 具有影響(Barnes and Polleux, 2009),在過去的
研究證實一些訊息傳遞路徑如 Ras, Raf 和 ERK/MAPK 的活化,有助於促進神經
生長錐的生長(Markus et al., 2002)。事實上 ERK/MAPK 具有許多調節點,調控轉

34
錄因子對神經軸突生長的影響,而且神經軸突的細胞骨架中,也可以藉由
ERK/MAPK 的磷酸化作用,調節局部 microtubule associated protein (MAP)的活
性(Polleux and Snider, 2010),進一步利用 APC (Adenomatous polyposis coli)控
制神經軸圖的延長(Zhou et al., 2004),或是其他MAP像是microtubule-associated
protein 1B (MAP1B)和 Tau protein 在磷酸化情況下會減少 microtubule 的結合,
使神經骨架縮短(Sperber et al., 1995, Gonzalez-Billault et al., 2004)。本實驗預處
理 PD98059 和 U0126 情況下,IGF-1 所產生的神經偏轉作用因而趨緩,同時也降
低生長速率。
在過去文獻中指出,一些神經滋養因子如 netrin, BNDF, DDC 或 myelin-
associated glycoprotein (MAG),都會與神經細胞膜上特異性受器結合並產生交互
作用,經過下游訊息傳遞路徑的活化,導致細胞內鈣離子儲存池鈣離子的釋放,使
得細胞質中鈣離子濃度提升,透過鈣離子濃度的改變進一步誘導細胞膜上 TRP
channels開啟,最終使更多的細胞外鈣離子流入細胞質中(Naeve et al., 2000, Hale
and Coward, 2013)。在我們 TRP channels 實驗結果亦可發現相同的現象,我們
經由預處理廣效性的 TRP channels 阻斷劑情況下,IGF-1 對神經產生的偏轉作用
明顯減少。因此推測 IGF-1 所產生的神經偏轉現象中,可能也涉及上述文獻中所
發現的訊息傳遞機制,而導致神經細胞膜上 TRP channel 參與神經偏轉的過程。
由於目前發現的 TRP channel superfamily 種類繁多,依照胺基酸序列可分為
TRPC, TRPV, TRPM, TRPP, TRPML, TRPA 和 TRPN (Venkatachalam and
Montell, 2007),其中 TRPC 主要存在於神經系統中,而且表現於神經生長錐
(Werther et al., 1990);不過確切 IGF-1 所造成的神經偏轉作用引發哪一類 TRP
channels 的活化,還需更進一步的探討。
本實驗以 turning assay 的實驗方法,提供了 IGF-1 對神經產生的偏轉作用關
於鈣離子來源所扮演的角色之外,同時也證實 IGF-1 所引起 tyrosine kinase 訊息
傳遞路徑的機制,可能是經由 PLC 促進 IP3的產生,讓細胞內鈣離子儲存池 IP3R
活化,最終導致細胞膜上 TRP channels 的開啟,影響鈣離子流入細胞內。

35
參考文獻
Adams TE, Epa VC, Garrett TPJ, Ward CW (2000) Structure and function of the type 1 insulin-like growth factor receptor. Cell Mol Life Sci 57:1050-1093.
Anderson MJ, Cohen MW, Zorychta E (1977) Effects of innervation on the distribution of acetylcholine receptors on cultured muscle cells. The Journal of physiology 268:731-756.
Barnes AP, Polleux F (2009) Establishment of axon-dendrite polarity in developing neurons. Annu Rev Neurosci 32:347-381.
Berzat A, Hall A (2010) Cellular responses to extracellular guidance cues. Embo J 29:2734-2745.
Blair LAC, Bence-Hanulec KK, Mehta S, Franke T, Kaplan D, Marshall J (1999) Akt-dependent potentiation of L channels by insulin-like growth factor-1 is required for neuronal survival. J Neurosci 19:1940-1951.
Blair LAC, Marshall J (1997) IGF-1 modulates N and L calcium channels in a PI 3-kinase-dependent manner. Neuron 19:421-429.
Bray D, Hollenbeck PJ (1988) Growth cone motility and guidance. Annual review of cell biology 4:43-61.
Brooker GJF, Kalloniatis M, Russo VC, Murphy M, Werther GA, Bartlett FF (2000) Endogenous IGF-1 regulates the neuronal differentiation of adult stem cells. J Neurosci Res 59:332-341.
Brose K, Tessier-Lavigne M (2000) Slit proteins: key regulators of axon guidance, axonal branching, and cell migration. Curr Opin Neurobiol 10:95-102.
Buck KB, Zheng JQ (2002) Growth cone turning induced by direct local modification of microtubule dynamics. J Neurosci 22:9358-9367.
Burgo A, Proux-Gillardeaux V, Sotirakis E, Bun P, Casano A, Verraes A, Liem RK, Formstecher E, Coppey-Moisan M, Galli T (2012) A molecular network for the transport of the TI-VAMP/VAMP7 vesicles from cell center to periphery. Dev Cell 23:166-180.
Butler AA, Yakar S, Gewolb IH, Karas M, Okubo Y, LeRoith D (1998) Insulin-like growth factor-I receptor signal transduction: at the interface between physiology and cell biology. Comp Biochem Phys B 121:19-26.
Canty AJ, Murphy M (2008) Molecular mechanisms of axon guidance in the developing corticospinal tract. Prog Neurobiol 85:214-235.
Caroni P (1993) Activity-Sensitive Signaling by Muscle-Derived Insulin-Like Growth-Factors in the Developing and Regenerating Neuromuscular System. Ann Ny

36
Acad Sci 692:209-222. Chilton JK (2006) Molecular mechanisms of axon guidance. Dev Biol 292:13-24. Conde C, Caceres A (2009) Microtubule assembly, organization and dynamics in axons
and dendrites. Nat Rev Neurosci 10:319-332. Cotrufo T, Andres RM, Ros O, Perez-Branguli F, Muhaisen A, Fuschini G, Martinez R,
Pascual M, Comella JX, Soriano E (2012) Syntaxin 1 is required for DCC/Netrin-1-dependent chemoattraction of migrating neurons from the lower rhombic lip. The European journal of neuroscience 36:3152-3164.
Cotrufo T, Perez-Branguli F, Muhaisen A, Ros O, Andres R, Baeriswyl T, Fuschini G, Tarrago T, Pascual M, Urena J, Blasi J, Giralt E, Stoeckli ET, Soriano E (2011) A signaling mechanism coupling netrin-1/deleted in colorectal cancer chemoattraction to SNARE-mediated exocytosis in axonal growth cones. J Neurosci 31:14463-14480.
D'Mello SR, Borodezt K, Soltoff SP (1997) Insulin-like growth factor and potassium depolarization maintain neuronal survival by distinct pathways: Possible involvement of PI 3-kinase in IGF-1 signaling. J Neurosci 17:1548-1560.
Dent EW, Gertler FB (2003) Cytoskeletal dynamics and transport in growth cone motility and axon guidance. Neuron 40:209-227.
Dickson BJ (2002) Molecular mechanisms of axon guidance. Science 298:1959-1964. Eftimie R, Brenner HR, Buonanno A (1991) Myogenin and MyoD join a family of skeletal
muscle genes regulated by electrical activity. Proc Natl Acad Sci U S A 88:1349-1353.
Egea J, Klein R (2007) Bidirectional Eph-ephrin signaling during axon guidance. Trends Cell Biol 17:230-238.
Eichmann A, Le Noble F, Autiero M, Carmeliet P (2005) Guidance of vascular and neural network formation. Curr Opin Neurobiol 15:108-115.
Fischer M, Kaech S, Knutti D, Matus A (1998) Rapid actin-based plasticity in dendritic spines. Neuron 20:847-854.
Foncea R, Andersson M, Ketterman A, Blakesley V, SapagHagar M, Sugden PH, LeRoith D, Lavandero S (1997) Insulin-like growth factor-I rapidly activates multiple signal transduction pathways in cultured rat cardiac myocytes. J Biol Chem 272:19115-19124.
Franke TF, Kaplan DR, Cantley LC, Toker A (1997) Direct regulation of the Akt proto-oncogene product by phosphatidylinositol-3,4-bisphosphate. Science 275:665-668.
Fuchs E, Cleveland DW (1998) A structural scaffolding of intermediate filaments in health and disease. Science 279:514-519.
Gallo G, Letourneau PC (2004) Regulation of growth cone actin filaments by guidance

37
cues. J Neurobiol 58:92-102. Genc B, Ozdinler PH, Mendoza AE, Erzurumlu RS (2004) A chemoattractant role for NT-
3 in proprioceptive axon guidance. PLoS biology 2:e403. Gluckman P, Klempt N, Guan J, Mallard C, Sirimanne E, Dragunow M, Klempt M, Singh
K, Williams C, Nikolics K (1992) A Role for Igf-1 in the Rescue of Cns Neurons Following Hypoxic-Ischemic Injury. Biochem Bioph Res Co 182:593-599.
Gonzalez-Billault C, Jimenez-Mateos EM, Caceres A, Diaz-Nido J, Wandosell F, Avila J (2004) Microtubule-associated protein 1B function during normal development, regeneration, and pathological conditions in the nervous system. J Neurobiol 58:48-59.
Goodman CS (1996) Mechanisms and molecules that control growth cone guidance. Annu Rev Neurosci 19:341-377.
Guan KL, Rao Y (2003) Signalling mechanisms mediating neuronal responses to guidance cues. Nat Rev Neurosci 4:941-956.
Gundersen RW, Barrett JN (1980) Characterization of the turning response of dorsal root neurites toward nerve growth factor. The Journal of cell biology 87:546-554.
Hale LJ, Coward RJ (2013) Insulin signalling to the kidney in health and disease. Clinical science 124:351-370.
Han L, Wen Z, Lynn RC, Baudet ML, Holt CE, Sasaki Y, Bassell GJ, Zheng JQ (2011) Regulation of chemotropic guidance of nerve growth cones by microRNA. Molecular brain 4:40.
Hansson HA, Dahlin LB, Danielsen N, Fryklund L, Nachemson AK, Polleryd P, Rozell B, Skottner A, Stemme S, Lundborg G (1986) Evidence Indicating Trophic Importance of Igf-I in Regenerating Peripheral-Nerves. Acta Physiol Scand 126:609-614.
Hilgemann DW (2007) Local PIP(2) signals: when, where, and how? Pflugers Archiv : European journal of physiology 455:55-67.
Hong F, Moon KA, Kim SS, Kim YS, Choi YK, Bae YS, Suh PG, Ryu SH, Choi EJ, Ha J, Kim SS (2001) Role of phospholipase C-gamma 1 in insulin-like growth factor I-induced muscle differentiation of H9c2 cardiac myoblasts. Biochem Bioph Res Co 282:816-822.
Huat TJ, Khan AA, Pati S, Mustafa Z, Abdullah JM, Jaafar H (2014) IGF-1 enhances cell proliferation and survival during early differentiation of mesenchymal stem cells to neural progenitor-like cells. Bmc Neurosci 15.
Hutchins BI, Li L, Kalil K (2012) Wnt-induced calcium signaling mediates axon growth and guidance in the developing corpus callosum. Science signaling 5:pt1.
Ishii DN (1989) Relationship of Insulin-Like Growth Factor-Ii Gene-Expression in Muscle

38
to Synaptogenesis. P Natl Acad Sci USA 86:2898-2902. Itofusa R, Kamiguchi H (2011) Polarizing membrane dynamics and adhesion for growth
cone navigation. Molecular and cellular neurosciences 48:332-338. Jessen KR, Mirsky R (2005) The origin and development of glial cells in peripheral
nerves. Nat Rev Neurosci 6:671-682. Jin M, Guan CB, Jiang YA, Chen G, Zhao CT, Cui K, Song YQ, Wu CP, Poo MM, Yuan XB
(2005) Ca2+-dependent regulation of rho GTPases triggers turning of nerve growth cones. J Neurosci 25:2338-2347.
Joanne Wang C, Li X, Lin B, Shim S, Ming GL, Levchenko A (2008) A microfluidics-based turning assay reveals complex growth cone responses to integrated gradients of substrate-bound ECM molecules and diffusible guidance cues. Lab on a chip 8:227-237.
Kim B, Leventhal PS, Saltiel AR, Feldman EL (1997) Insulin-like growth factor-I-mediated neurite outgrowth in vitro requires mitogen-activated protein kinase activation. J Biol Chem 272:21268-21273.
Kiselyov K, Xu X, Mozhayeva G, Kuo T, Pessah I, Mignery G, Zhu X, Birnbaumer L, Muallem S (1998) Functional interaction between InsP(3) receptors and store-operated Htrp3 channels. Nature 396:478-482.
Koch C, Zador A (1993) The function of dendritic spines: devices subserving biochemical rather than electrical compartmentalization. J Neurosci 13:413-422.
Lemmon MA, Schlessinger J (2010) Cell signaling by receptor tyrosine kinases. Cell 141:1117-1134.
Leski ML, Valentine SL, Baer JD, Coyle JT (2000) Insulin-like growth factor I prevents the development of sensitivity to kainate neurotoxicity in cerebellar granule cells. Journal of neurochemistry 75:1548-1556.
Li Y, Jia YC, Cui K, Li N, Zheng ZY, Wang YZ, Yuan XB (2005) Essential role of TRPC channels in the guidance of nerve growth cones by brain-derived neurotrophic factor. Nature 434:894-898.
Liou JC, Tsai FZ, Ho SY (2003) Potentiation of quantal secretion by insulin-like growth factor-1 at developing motoneurons in Xenopus cell culture. J Physiol-London 553:719-728.
Lockerbie RO (1987) The neuronal growth cone: a review of its locomotory, navigational and target recognition capabilities. Neuroscience 20:719-729.
Lohof AM, Quillan M, Dan Y, Poo MM (1992) Asymmetric modulation of cytosolic cAMP activity induces growth cone turning. J Neurosci 12:1253-1261.
Lowery LA, Van Vactor D (2009) The trip of the tip: understanding the growth cone machinery. Nature reviews Molecular cell biology 10:332-343.

39
Majewska A, Brown E, Ross J, Yuste R (2000) Mechanisms of calcium decay kinetics in hippocampal spines: Role of spine calcium pumps and calcium diffusion through the spine neck in biochemical compartmentalization. J Neurosci 20:1722-1734.
Markus A, Zhong J, Snider WD (2002) Raf and akt mediate distinct aspects of sensory axon growth. Neuron 35:65-76.
Marsick BM, Flynn KC, Santiago-Medina M, Bamburg JR, Letourneau PC (2010) Activation of ADF/cofilin mediates attractive growth cone turning toward nerve growth factor and netrin-1. Developmental neurobiology 70:565-588.
Mattila PK, Lappalainen P (2008) Filopodia: molecular architecture and cellular functions. Nat Rev Mol Cell Bio 9:446-454.
Mehrhof FB, Muller FU, Bergmann MW, Li P, Wang Y, Schmitz W, Dietz R, von Harsdorf R (2001) In cardiomyocyte hypoxia, insulin-like growth factor-I-induced antiapoptotic signaling requires phosphatidylinositol-3-OH-kinase-dependent and mitogen-activated protein kinase-dependent activation of the transcription factor cAMP response element-binding protein. Circulation 104:2088-2094.
Meir A, Ginsburg S, Butkevich A, Kachalsky SG, Kaiserman I, Ahdut R, Demirgoren S, Rahamimoff R (1999) Ion channels in presynaptic nerve terminals and control of transmitter release. Physiological reviews 79:1019-1088.
Merianda TT, Lin AC, Lam JS, Vuppalanchi D, Willis DE, Karin N, Holt CE, Twiss JL (2009) A functional equivalent of endoplasmic reticulum and Golgi in axons for secretion of locally synthesized proteins. Molecular and cellular neurosciences 40:128-142.
Meriane M, Tcherkezian J, Webber CA, Danek EI, Triki I, McFarlane S, Bloch-Gallego E, Lamarche-Vane N (2004) Phosphorylation of DCC by Fyn mediates Netrin-1 signaling in growth cone guidance. The Journal of cell biology 167:687-698.
Miesenbock G, De Angelis DA, Rothman JE (1998) Visualizing secretion and synaptic transmission with pH-sensitive green fluorescent proteins. Nature 394:192-195.
Naeve GS, Vana AM, Eggold JR, Verge G, Ling N, Foster AC (2000) Expression of rat insulin-like growth factor binding protein-6 in the brain, spinal cord, and sensory ganglia. Brain research Molecular brain research 75:185-197.
Nakazawa H, Sada T, Toriyama M, Tago K, Sugiura T, Fukuda M, Inagaki N (2012) Rab33a Mediates Anterograde Vesicular Transport for Membrane Exocytosis and Axon Outgrowth. J Neurosci 32:12712-12725.
Nieto-Estevez V, Defterali C, Vicario-Abejon C (2016) IGF-I: A Key Growth Factor that Regulates Neurogenesis and Synaptogenesis from Embryonic to Adult Stages of the Brain. Frontiers in neuroscience 10:52.
Nieuwkoop PD, Faber J (1967) Normal table of Xenopus laevis (Daudin); a systematical and chronological survey of the development from the fertilized egg till the end

40
of metamorphosis. Amsterdam,: North-Holland Pub. Co. Oertner TG, Matus A (2005) Calcium regulation of actin dynamics in dendritic spines.
Cell Calcium 37:477-482. Ozdinler PH, Macklis JD (2006) IGF-I specifically enhances axon outgrowth of
corticospinal motor neurons. Nat Neurosci 9:1371-1381. Parekh AB, Putney JW, Jr. (2005) Store-operated calcium channels. Physiological
reviews 85:757-810. Polleux F, Snider W (2010) Initiating and Growing an Axon. Cold Spring Harbor
perspectives in biology 2. Pu SF, Zhuang HX, Marsh DJ, Ishii DN (1999) Time-dependent alteration of insulin-like
growth factor gene expression during nerve regeneration in regions of muscle enriched with neuromuscular junctions. Mol Brain Res 63:207-216.
Rabinovsky ED, Gelir E, Gelir S, Lui H, Kattash M, DeMayo FJ, Shenaq SM, Schwartz RJ (2002) Targeted expression of IGF-1 transgene to skeletal muscle accelerates muscle and motor neuron regeneration. Faseb J 16:53-+.
Richards DA, De Paola V, Caroni P, Gahwiler BH, McKinney RA (2004) AMPA-receptor activation regulates the diffusion of a membrane marker in parallel with dendritic spine motility in the mouse hippocampus. The Journal of physiology 558:503-512.
Rinderknecht E, Humbel RE (1976) Polypeptides with Nonsuppressible Insulin-Like and Cell-Growth Promoting Activities in Human-Serum - Isolation, Chemical Characterization, and Some Biological Properties of Forms 1 and 2. P Natl Acad Sci USA 73:2365-2369.
Rohm B, Ottemeyer A, Lohrum M, Puschel AW (2000) Plexin/neuropilin complexes mediate repulsion by the axonal guidance signal semaphorin 3A. Mech Develop 93:95-104.
Ros O, Cotrufo T, Martinez-Marmol R, Soriano E (2015) Regulation of Patterned Dynamics of Local Exocytosis in Growth Cones by Netrin-1. J Neurosci 35:5156-5170.
Sanes JR, Lichtman JW (1999) Development of the vertebrate neuromuscular junction. Annu Rev Neurosci 22:389-442.
Sasaoka T, Ishiki M, Wada T, Hori H, Hirai H, Haruta T, Ishihara H, Kobayashi M (2001) Tyrosine phosphorylation-dependent and -independent role of Shc in the regulation of IGF-1-induced mitogenesis and glycogen synthesis. Endocrinology 142:5226-5235.
Schaefer AW, Schoonderwoert VTG, Ji L, Mederios N, Danuser G, Forscher P (2008) Coordination of actin filament and microtubule dynamics during neurite outgrowth. Dev Cell 15:146-162.

41
Scolnick JA, Cui K, Duggan CD, Xuan S, Yuan XB, Efstratiadis A, Ngai J (2008) Role of IGF signaling in olfactory sensory map formation and axon guidance. Neuron 57:847-857.
Song H, Ming G, He Z, Lehmann M, McKerracher L, Tessier-Lavigne M, Poo M (1998) Conversion of neuronal growth cone responses from repulsion to attraction by cyclic nucleotides. Science 281:1515-1518.
Song HJ, Poo MM (2001) The cell biology of neuronal navigation. Nat Cell Biol 3:E81-E88.
Sosa L, Dupraz S, Laurino L, Bollati F, Bisbal M, Caceres A, Pfenninger KH, Quiroga S (2006) IGF-1 receptor is essential for the establishment of hippocampal neuronal polarity. Nat Neurosci 9:993-995.
Sperber BR, Leight S, Goedert M, Lee VM (1995) Glycogen synthase kinase-3 beta phosphorylates tau protein at multiple sites in intact cells. Neuroscience letters 197:149-153.
Spitzer NC, Lamborghini JE (1976) The development of the action potential mechanism of amphibian neurons isolated in culture. Proc Natl Acad Sci U S A 73:1641-1645.
Svoboda K, Tank DW, Denk W (1996) Direct measurement of coupling between dendritic spines and shafts. Science 272:716-719.
Tabti N, Poo M-M (1991) Culturing spinal neurons and muscle cells from Xenopus embryos. In: Cellular and molecular neuroscience series (G, B. and K, G., eds) Cambridge, Mass.: MIT Press.
TessierLavigne M, Goodman CS (1996) The molecular biology of axon guidance. Science 274:1123-1133.
Tojima T, Akiyama H, Itofusa R, Li Y, Katayama H, Miyawaki A, Kamiguchi H (2007) Attractive axon guidance involves asymmetric membrane transport and exocytosis in the growth cone. Nat Neurosci 10:58-66.
Tojima T, Hines JH, Henley JR, Kamiguchi H (2011) Second messengers and membrane trafficking direct and organize growth cone steering. Nat Rev Neurosci 12:191-203.
Tojima T, Itofusa R, Kamiguchi H (2014) Steering neuronal growth cones by shifting the imbalance between exocytosis and endocytosis. J Neurosci 34:7165-7178.
Tojima T, Kamiguchi H (2015) Exocytic and endocytic membrane trafficking in axon development. Development, growth & differentiation 57:291-304.
Tsien RW, Lipscombe D, Madison DV, Bley KR, Fox AP (1988) Multiple types of neuronal calcium channels and their selective modulation. Trends in Neurosciences 11:431-438.
Van Obberghen E, Baron V, Delahaye L, Emanuelli B, Filippa N, Giorgetti-Peraldi S,

42
Lebrun P, Mothe-Satney I, Peraldi P, Rocchi S, Sawka-Verhelle D, Tartare-Deckert S, Giudicelli J (2001) Surfing the insulin signaling web. Eur J Clin Invest 31:966-977.
Venkatachalam K, Montell C (2007) TRP channels. Annu Rev Biochem 76:387-417. Vitriol EA, Zheng JQ (2012) Growth cone travel in space and time: the cellular ensemble
of cytoskeleton, adhesion, and membrane. Neuron 73:1068-1081. Weinl C, Drescher U, Lang S, Bonhoeffer F, Loschinger J (2003) On the turning of
Xenopus retinal axons induced by ephrin-A5. Development 130:1635-1643. Wen Z, Guirland C, Ming GL, Zheng JQ (2004) A CaMKII/calcineurin switch controls the
direction of Ca(2+)-dependent growth cone guidance. Neuron 43:835-846. Werther GA, Abate M, Hogg A, Cheesman H, Oldfield B, Hards D, Hudson P, Power B,
Freed K, Herington AC (1990) Localization of insulin-like growth factor-I mRNA in rat brain by in situ hybridization--relationship to IGF-I receptors. Molecular endocrinology 4:773-778.
Winberg ML, Noordermeer JN, Tamagnone L, Comoglio PM, Spriggs MK, Tessier-Lavigne M, Goodman CS (1998) Plexin A is a neuronal semaphorin receptor that controls axon guidance. Cell 95:903-916.
Xu XM, Guenard V, Kleitman N, Aebischer P, Bunge MB (1995) A combination of BDNF and NT-3 promotes supraspinal axonal regeneration into Schwann cell grafts in adult rat thoracic spinal cord. Experimental neurology 134:261-272.
You H, Zheng HW, Murray SA, Yu Q, Uchida T, Fan DM, Xiao ZXJ (2002) IGF-1 induces Pin1 expression in promoting cell cycle S-phase entry. J Cell Biochem 84:211-216.
Yu TW, Bargmann CI (2001) Dynamic regulation of axon guidance. Nat Neurosci 4 Suppl:1169-1176.
Yuan JP, Kiselyov K, Shin DM, Chen J, Shcheynikov N, Kang SH, Dehoff MH, Schwarz MK, Seeburg PH, Muallem S, Worley PF (2003) Homer binds TRPC family channels and is required for gating of TRPC1 by IP3 receptors. Cell 114:777-789.
Zhang FJ, Wang LH, Liu Z, Xing ZY, Wang HJ, Li ZZ (2010) Insulin-like growth factor-1 modulates Ca2+ homeostasis and apoptosis of cultured dorsal root ganglion neurons with excitotoxicity induced by glutamate. Pharmazie 65:5-8.
Zheng JQ, Felder M, Connor JA, Poo MM (1994) Turning of Nerve Growth Cones Induced by Neurotransmitters. Nature 368:140-144.
Zheng JQ, Wan JJ, Poo MM (1996) Essential role of filopodia in chemotropic turning of nerve growth cone induced by a glutamate gradient. J Neurosci 16:1140-1149.
Zhou FQ, Zhou J, Dedhar S, Wu YH, Snider WD (2004) NGF-induced axon growth is mediated by localized inactivation of GSK-3beta and functions of the microtubule plus end binding protein APC. Neuron 42:897-912.

43
Zylbersztejn K, Petkovic M, Burgo A, Deck M, Garel S, Marcos S, Bloch-Gallego E, Nothias F, Serini G, Bagnard D, Binz T, Galli T (2012) The vesicular SNARE Synaptobrevin is required for Semaphorin 3A axonal repulsion. The Journal of cell biology 196:37-46.

44

45
Figure 1. Experimental design of pressure injection-induced microscopic gradient.
(A) FITC was locally-applied from a glass micropipette by pressure injector (1 Hz,
3 psi and 50 msec in duration) and images were recorded every 5 minutes from
onset (0 min) to the end (60 min). The concentration of FITC at 100 μm distal
from tip of micropipette was saturated in 10 minutes. Fluorescence images are
shown in pseudocolor, with blue and red representing lowest and highest
intensities, respectively. The left bright-field image shows the arrangement of
micropipette. Scale bar, 25 μm. (B) Composite 3D graph of time, distance and
FITC gradient from the tip of micropipette to the growth cones (100 μm).

46

47
Figure 2. Cyclic AMP microscopic gradient successfully induce
attractive response on growth cone of Xenopus developing motoneuron.
(A-B) The turning response of growth cone before (left) and after 60 min (middle)
Ringer’s solution and cAMP (25 mM), respectively, application by micropipette
with a distance of 100 μm and 45 degree angle. Superimposed traces depict the
trajectory image (right). Each line represents a single growth cone trajectory. The
arrow indicates the direction of the micropipette. Scale bar, 25 μm. (C) The open
circles depict the angle of growth cones turning at the end of experiment. (D)
Average turning angles for the data illustrated in (A-B). Data are Mean±SEM. and
*P < 0.05, Mann-whitney U test. (E) Cumulative distributions of turning angles
induced by cAMP and Ringer’s solution.

48

49
Figure 3. Attractive effect of IGF-1 on Xenopus developing spinal
motoneuron.
(A-B) The turning response of growth cone before (left) and after 60 min (middle)
Ringer’s solution and IGF-1 (10 μM), respectively. Superimposed traces (right)
depict the trajectory image. Each line represents a single growth cone trajectory.
The arrow indicates the direction of the micropipette. Scale bar, 25 μm. (C) The
open circles depict the angle of growth cones turning at the end of each
experiment. (D) Average turning angles for the data illustrated in (A-B right). Data
are Mean±SEM. Data significantly different from Ringer’s solution is marked (*P
< 0.05, Mann-whitney U test). (E) Cumulative distributions of turning angles
induced by Ringer’s solution and IGF-1. (F) Growth rate of Xenopus motoneuron
in the presence of Ringer’s solution and IGF-1 treatments.
50

51
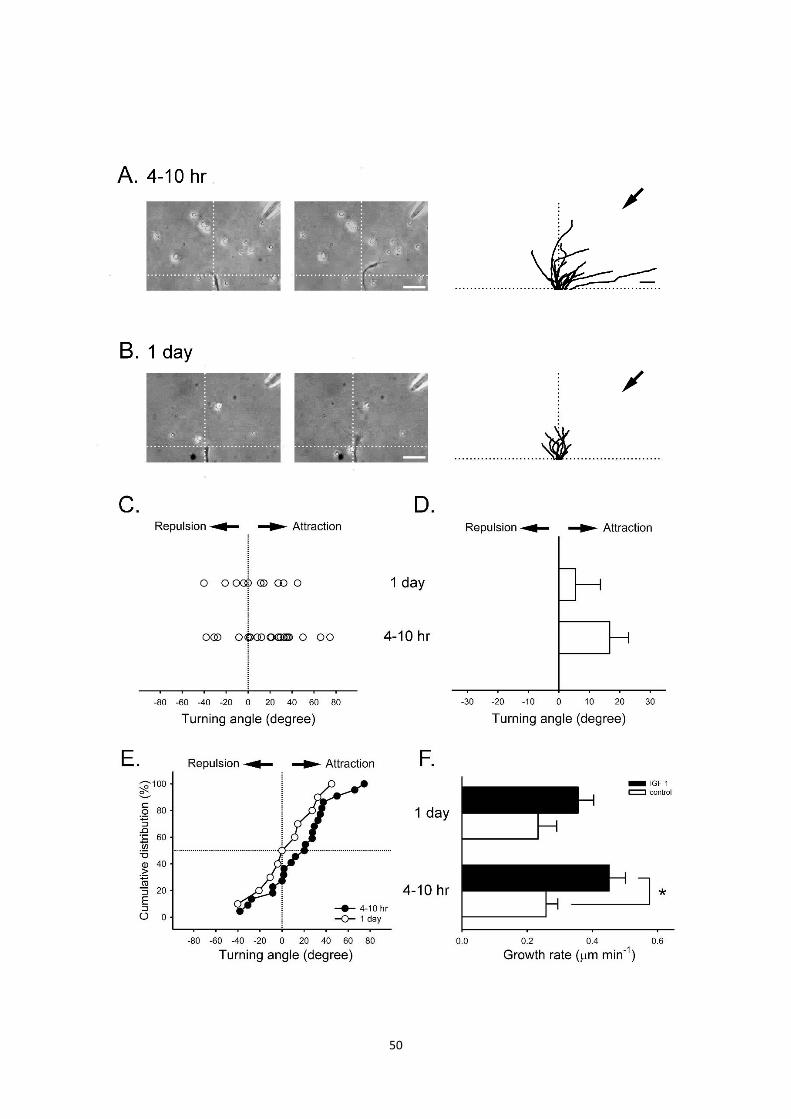
Figure 4. Effect of IGF-1-induced attractive turning response on
Xenopus motoneuron at different developmental stages.
Cultures of either 4-10 hours incubation time (A) or 1 day (B) are used in growth
con turning assay. Representative images of growth cone turning response
before (left) and after 60 min (middle) application by IGF-1 (10 μM).
Superimposed traces (right) depict the trajectory image. Each line represents a
single growth cone trajectory. The arrow indicates the direction of the micropipette.
Scale bar, 25 μm. (C) The open circles depict the angle of growth cones turning
at the end of each experiment. (D) Summary of turning angles for the data
illustrated in (A-B right). (E) Cumulative distributions of turning angles induced by
IGF-1. (F) Summary of the effect of IGF-1 on the growth rate of motoneuron at
different developmental stages. Data are Mean±SEM. Data significantly different
from that of the control are marked (*P < 0.05, Mann-whitney U test).

52

53
Figure 5. Role of calcium on IGF-1-induced growth cone turning.
(A-C) The turning response of IGF-1(10 μM) on growth cone before (left) and
after 60 min (middle) pretreatment with (B) Ca2+-free Ringer and (C) Cd2+ (0.5
mM), respectively. Superimposed traces depict the trajectory image (A-C right).
Each line represents a single growth cone trajectory. The arrow indicates the
direction of the micropipette. Scale bar, 25 μm. (D)The open circles depict the
angle of growth cones turning in the last. (E) Average turning angles for the data
illustrated in (A-C right). (F) Cumulative distributions of turning angles induced by
IGF-1 in Ca2+-free Ringer or Cd2+ pretreatment. (G) Effect of IGF-1 on the growth
rate of developing motoneuron under various condition. Data are Mean±SEM.
Data significantly different from that of the control are marked (*P < 0.05, Mann-
whitney U test).

54

55
Figure 6. Involvement of internal calcium reservoir in IGF-1-induced
growth cone turning response.
(A-B) The IGF-1-induced turning response of growth cone before (left) and after
60 min (middle) pretreatment with Thapsigargin (2 μM, TG). Superimposed traces
(right) depict the trajectory image. Each line represents a single growth cone
trajectory. The arrow indicates the direction of the micropipette. Scale bar, 25 μm.
(C) The open circles depict the angle of growth cones turning at the end of each
experiment. (D) Average turning angles for the data illustrated in (A-B right). (E)
Cumulative distributions of turning angles preloaded with TG. (F) The average
net lengths of growth cone extension for all the growth cones examined under
IGF-1 treatments. Data significantly different from that of the control are marked
(*P < 0.05, Mann-whitney U test).

56
Figure 7. The exocytic membrane effect of IGF-1 microscopic
gradient on the growth cone.
Representative bright-field (A) and fluorescence images (B-C) of motoneuron
transfected by pHluorin. Images were recorded every 30 second from onset (0
min, right) to the end (60 min, left). Note the IGF-1 induced differential increase
in pHluorin fluorescence intensity on both site of the growth cone after at 20
minute IGF-1 application. Scale bar, 25 μm.

57

58
Figure 8. Role of ryanodine receptor in IGF-1-induced attractive
turning response.
(A-B) The turning response of IGF-1(10 μM) on growth cone before (left) and after
60 min (middle) TMB-8 (3 μM) pretreatment. Superimposed traces (right) depict
the trajectory image. Each line represents a single growth cone trajectory. The
arrow indicates the direction of the micropipette. Scale bar, 25 μm. (C) The open
circles depict the angle of growth cones turning in the last. (D) Average turning
angles for the data illustrated in (A-B right). (E) Cumulative distributions of turning
angles preloaded with TMB-8. (F) The average net lengths of growth cone
extension for all the growth cones examined under IGF-1 treatments. Data
significantly different from that of the control are marked (*P < 0.05, Mann-whitney
U test).

59

60
Figure 9. IP3 receptor is responsible for IGF-1-induced turning
response at developing Xenopus motoneuron.
(A-B) The IGF-1-induced turning response of growth cone before (left) and after
60 min (middle) XeC (1 μM) pretreatment (B). Superimposed traces (right) depict
the trajectory image. Each line represents a single growth cone trajectory. The
arrow indicates the direction of the micropipette. Scale bar, 25 μm. (C)The open
circles depict the angle of growth cones turning in the last. (D)Average turning
angles for the data illustrated in (A-B right). (E) Cumulative distributions of turning
angles preloaded with XeC. (F) The average net lengths of growth cone extension
for all the growth cones examined under IGF-1 treatments. Data significantly
different from that of the control are marked (*P < 0.05, Mann-whitney U test).

61

62
Figure 10 . Activation of tyrosine kinase IGF-1 receptor is prerequisite
for IGF-1-induced growth cone turning response.
(A-B) The IGF-1(10 μM)-induced turning response of growth cone before (left)
and after 60 min (middle) pretreatment with Genistein (10 μM). Superimposed
traces (right) depict the trajectory image. Each line represents a single growth
cone trajectory. The arrow indicates the direction of the micropipette. Scale bar,
25 μm. (C) The open circles depict the angle of growth cones turning in the last.
(D) Average turning angles for the data illustrated in (A-B right). (E) Cumulative
distributions of turning angles pretreatment with Genistein. (F) The average net
lengths of growth cone extension for all the growth cones examined under IGF-1
treatments. Data significantly different from that of the control are marked (*P <
0.05, Mann-whitney U test).

63

64
Figure 11. PLCγ activity is involvement in IGF-1-induced growth cone
turning.
(A-B) The IGF-1(10 μM)-induced turning response of growth cone before (left)
and after 60 min (middle) pretreatment with U73122 (5 μM). Superimposed
traces (right) depict the trajectory image. Each line represents a single growth
cone trajectory. The arrow indicates the direction of the micropipette. Scale bar,
25 μm. (C) The open circles depict the angle of growth cones turning in the last.
(D) Average turning angles for the data illustrated in (A-B right). (E) Cumulative
distributions of turning angles pretreatment with U73122. (F)The average net
lengths of growth cone extension for all the growth cones examined under IGF-
1 treatments. Data significantly different from that of the control are marked (*P
< 0.05, Mann-whitney U test).

65

66
Figure 12. Involvement of PI-3 kinase pathway in IGF-1-induced
growth cone turning.
(A-B) The IGF-1(10 μM)-induced turning response of growth cone before (left)
and after 60 min (middle) pretreatment with wortmannin (100 nM). Superimposed
traces (right) depict the trajectory image. Each line represents a single growth
cone trajectory. The arrow indicates the direction of the micropipette. Scale bar,
25 μm. (C) The open circles depict the angle of growth cones turning in the last.
(D) Average turning angles for the data illustrated in (A-B right). (E) Cumulative
distributions of turning angles pretreatment with wortmannin. (F) The average net
lengths of growth cone extension for all the growth cones examined under IGF-1
treatments. Data significantly different from that of the control are marked (*P <
0.05, Mann-whitney U test).

67

68
Figure 13. Role of MAP kinase in growth cone turning response
induced by IGF-1.
(A-B) The IGF-1(10 μM)-induced turning response of growth cone before (left)
and after 60 min (middle) pretreatment with PD98059 (10 μM) or U0126 (10 μM).
Superimposed traces (right) depict the trajectory image. Each line represents a
single growth cone trajectory. The arrow indicates the direction of the micropipette.
Scale bar, 25 μm. (C)The open circles depict the angle of growth cones turning
in the last. (D)Average turning angles for the data illustrated in (A-B right). (E)
Cumulative distributions of turning angles for the condition shown in (A-C). (F)The
average net lengths of growth cone extension for all the growth cones examined
under IGF-1 treatments. Data significantly different from that of the control are
marked (*P < 0.05, Mann-whitney U test).

69

70
Figure 14. Summary of the effect of pharmacological inhibitors on
IGF-1-induced growth cone turning response.
Summary of growth cone turning in response to a gradient of IGF-1 in different
conditions. Data significantly different from that of the IGF-1(10 μM) are marked
(*P < 0.05, Mann-whitney U test).

71

72
Figure 15. TRPC channel is involved in IGF-1-induced growth cone
turning response.
(A-B) The IGF-1(10 μM)-induced turning response of growth cone before (left)
and after 60 min (middle) pretreatment with SKF96365 (25 μM). Superimposed
traces (right) depict the trajectory image. Each line represents a single growth
cone trajectory. The arrow indicates the direction of the micropipette. Scale bar,
25 μm. (C) The open circles depict the angle of growth cones turning in the last.
(D) Average turning angles for the data illustrated in (A-B right). (E) Cumulative
distributions of turning angles pretreatment with SKF96365. (F)The average net
lengths of growth cone extension for all the growth cones examined under IGF-1
treatments. Data significantly different from that of the control are marked (*P <
0.05, Mann-whitney U test).

73

74
Figure 16. Effect of TPR channel inhibitor Gd3+ on the growth cone
turning response induced by IGF-1.
(A-B) The IGF-1(10 μM)-induced turning response of growth cone before (left)
and after 60 min (middle) pretreatment with Gd3+ (10 μM). Superimposed traces
(right) depict the trajectory image. Each line represents a single growth cone
trajectory. The arrow indicates the direction of the micropipette. Scale bar, 25 μm.
(C)The open circles depict the angle of growth cones turning in the last.
(D)Average turning angles for the data illustrated in (A-B right). (E)Cumulative
distributions of turning angles pretreatment with Gd3+. (F)The average net lengths
of growth cone extension for all the growth cones examined under IGF-1
treatments. Data significantly different from that of the control are marked (*P <
0.05, Mann-whitney U test).

75
Tabl
e 1.
Res
pons
e of
Xen
opus
neu
ron
grow
th c
one
turn
ing
appl
ied
by d
iffer
ent p
retr
eatm
ent i
n gr
adie
nts.

76

77
Figure 17. Schematic diagram illustrating in the signaling cascade
underlying IGF-1-induced attractive growth cone turning response.
Results from our studies demonstrate IGF-1 has an attractive effect on axonal
guidance and this is done via IGF-1 receptor, PLCγ, PI 3-kinase and TRPC
signaling cascades.

78

79
附圖 1. The structure of the growth cone.
The structure of the growth cone is fundamental to its function. The growth cone
can be separated into three domains based on cytoskeletal distribution. Including
peripheral domain (P), central domain (C) and transition zone (T) (Lowery and
Van Vactor, 2009).

80
附圖 2. Chemorepulsion and chemoattraction.
The growth cone (light blue) induced by a gradient of molecules, it extends away
from chemorepulsion (red) and towards chemoattraction (green).The axon effect
is to cause a turning of the growth cone (dark blue) (Chilton, 2006).

81
附圖
3. S
truc
ture
of t
he in
sulin
rec
epto
r an
d IG
F-I
rec
epto
r.
(Buc
k an
d Z
heng
, 20
02,
Hal
e an
d C
owar
d, 2
013)
.

82
Sch
emat
ic r
epre
sent
atio
n of
the
exp
ress
ion
prof
iles
of I
GF
1 an
d IG
F2
in t
he d
evel
opin
g an
d
rege
nera
tion
neur
omus
cula
r sy
stem
(C
aron
i, 19
93).
附圖
4. R
epre
sent
atio
n of
the
expr
essi
on p
rofil
es o
f IG
Fs.
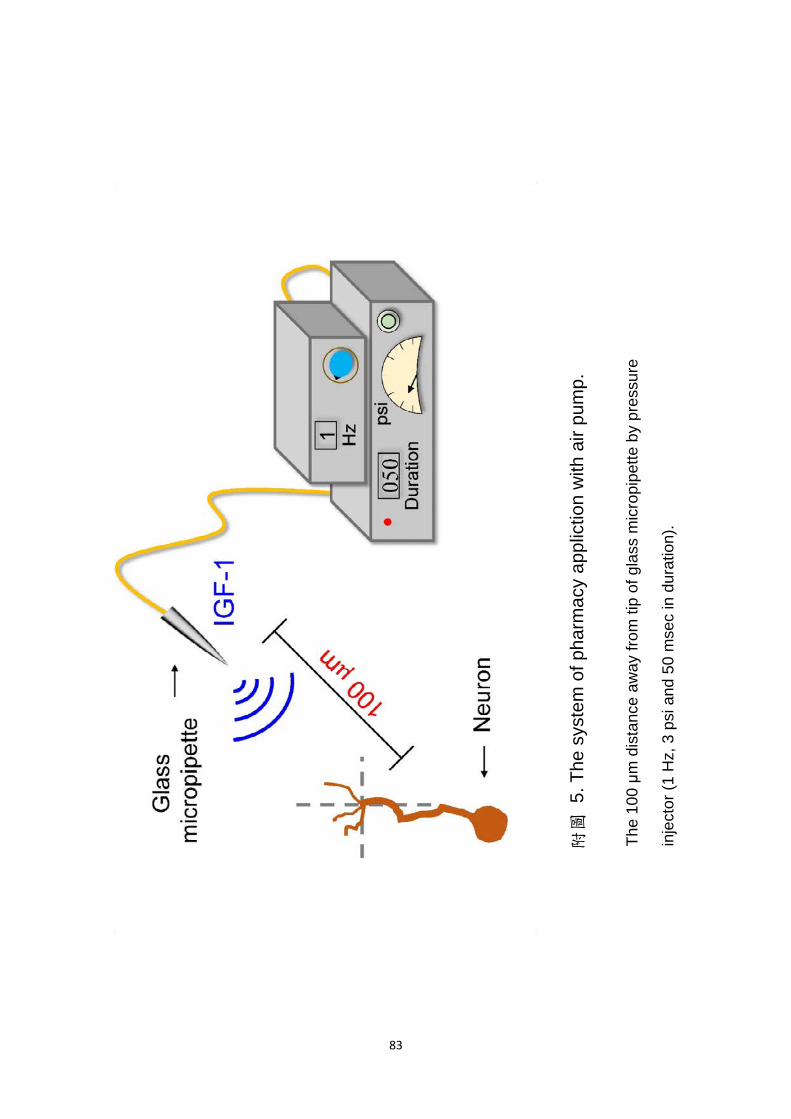
83
The
100
μm
dis
tanc
e aw
ay fr
om ti
p of
gla
ss m
icro
pipe
tte b
y pr
essu
re
inje
ctor
(1
Hz,
3 p
si a
nd 5
0 m
sec
in d
urat
ion)
.
附圖
5. T
he s
yste
m o
f pha
rmac
y ap
plic
tion
with
air
pum
p.

84
附圖
6. P
roce
dure
s fo
r th
e pr
epar
ing
Xen
opus
ner
ve c
ultu
re.










![南宋文言公案故事[通殺夫]類之敘事轉向 - 國立中興大學中國 ...](https://static.fdokumen.com/doc/165x107/63209c8b117b4414ec0b1844/--1677074896.jpg)









