
Identification of tetracycline- and erythromycin-resistant Gram-positive cocci within the fermenting...
12
ORIGINAL ARTICLE Identification of tetracycline- and erythromycin-resistant Gram-positive cocci within the fermenting microflora of an Italian dairy food product C. Devirgiliis, S. Barile, A. Caravelli, D. Coppola and G. Perozzi INRAN – National Research Institute on Food & Nutrition, Via Ardeatina, Rome, Italy Introduction Selection of antibiotic-resistant (AbR) bacteria in the intestinal microflora of livestock has occurred over the past few decades following the use of antimicrobial agents as growth promoters in animal feed (Teuber 2001; Wegener 2003). To prevent the environmental spread of AbR bacteria, this use of antimicrobials was banned in all EU member states in 2006, but an increasing number of reports describing recovery of genetically resistant strains from environmental sources raised great scientific interest (Wegener 2003), especially in the light of the intrinsic potential transmission of genetic determinants for anti- biotic resistance to human pathogens through the food chain (Mathur and Singh 2005). Commensal bacteria used as natural fermentation starters in food processing are not intrinsically pathogenic, although some genera (mostly enterococci) are capable of acquiring virulence genes. Enterococcus species represent a substantial propor- tion of the lactic acid bacteria (LAB) commonly found in foods, as they are present in raw meat and milk even when sanitary rules are respected. They can spoil pro- cessed meats, but they are also important for ripening and for the development of specific flavours in traditional cheeses and sausages, especially in those produced in the mediterranean area. Enterococci are also used as human probiotics. However, such use remains controversial in the light of the capability of most Enterococcus species to Keywords antibiotic resistance, Enterococcus, food quality, Lactococcus, Mozzarella di Bufala Campana, Streptococcus. Correspondence Chiara Devirgiliis, INRAN, Via Ardeatina 546, 00178 Rome, Italy. E-mail: [email protected] 2009 ⁄ 1514: received 28 August 2009, revised 30 November 2009 and accepted 15 December 2009 doi:10.1111/j.1365-2672.2010.04661.x Abstract Aims: Microbiological and molecular analysis of antibiotic resistance in Gram- positive cocci derived from the Italian PDO (Protected Designation of Origin) dairy food product Mozzarella di Bufala Campana. Methods and Results: One hundred and seven coccal colonies were assigned to Enterococcus faecalis, Lactococcus lactis and Streptococcus bovis genera by ARDRA analysis (amplified ribosomal DNA restriction analysis). Among them, 16 Ent. faecalis, 26 L. lactis and 39 Strep. bovis displayed high minimum inhibi- tory concentration (MIC) values for tetracycline, while 17 L. lactis showed high MIC values for both tetracycline and erythromycin. Strain typing and molecu- lar analysis of the phenotypically resistant isolates demonstrated the presence of the tet(M) gene in the tetracycline-resistant strains and of tet(S) and erm(B) in the double-resistant strains. Southern blot analysis revealed plasmid localization of L. lactis tet(M), as well as of the erm(B) and tet(S) genes. Genetic linkage of erm(B) and tet(S) was also demonstrated by PCR amplification. Conjugation experiments demonstrated horizontal transfer to Ent. faecalis strain JH2-2 only for the plasmid-borne L. lactis tet(M) gene. Conclusions: We characterized tetracycline-and erythromycin-resistance genes in coccal species, representing the fermenting microflora of a typical Italian dairy product. Significance and Impact of the Study: These results are of particular relevance from the food safety viewpoint, especially in the light of the potential risk of horizontal transfer of antibiotic-resistance genes among foodborne commensal bacteria. Journal of Applied Microbiology ISSN 1364-5072 ª 2010 INRAN Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 313–323 313
Transcript of Identification of tetracycline- and erythromycin-resistant Gram-positive cocci within the fermenting...

ORIGINAL ARTICLE
Identification of tetracycline- and erythromycin-resistantGram-positive cocci within the fermenting microflora of anItalian dairy food productC. Devirgiliis, S. Barile, A. Caravelli, D. Coppola and G. Perozzi
INRAN – National Research Institute on Food & Nutrition, Via Ardeatina, Rome, Italy
Introduction
Selection of antibiotic-resistant (AbR) bacteria in the
intestinal microflora of livestock has occurred over the
past few decades following the use of antimicrobial agents
as growth promoters in animal feed (Teuber 2001;
Wegener 2003). To prevent the environmental spread of
AbR bacteria, this use of antimicrobials was banned in all
EU member states in 2006, but an increasing number of
reports describing recovery of genetically resistant strains
from environmental sources raised great scientific interest
(Wegener 2003), especially in the light of the intrinsic
potential transmission of genetic determinants for anti-
biotic resistance to human pathogens through the food
chain (Mathur and Singh 2005). Commensal bacteria
used as natural fermentation starters in food processing
are not intrinsically pathogenic, although some genera
(mostly enterococci) are capable of acquiring virulence
genes. Enterococcus species represent a substantial propor-
tion of the lactic acid bacteria (LAB) commonly found in
foods, as they are present in raw meat and milk even
when sanitary rules are respected. They can spoil pro-
cessed meats, but they are also important for ripening
and for the development of specific flavours in traditional
cheeses and sausages, especially in those produced in the
mediterranean area. Enterococci are also used as human
probiotics. However, such use remains controversial in
the light of the capability of most Enterococcus species to
Keywords
antibiotic resistance, Enterococcus, food
quality, Lactococcus, Mozzarella di Bufala
Campana, Streptococcus.
Correspondence
Chiara Devirgiliis, INRAN, Via Ardeatina 546,
00178 Rome, Italy. E-mail: [email protected]
2009 ⁄ 1514: received 28 August 2009,
revised 30 November 2009 and accepted
15 December 2009
doi:10.1111/j.1365-2672.2010.04661.x
Abstract
Aims: Microbiological and molecular analysis of antibiotic resistance in Gram-
positive cocci derived from the Italian PDO (Protected Designation of Origin)
dairy food product Mozzarella di Bufala Campana.
Methods and Results: One hundred and seven coccal colonies were assigned
to Enterococcus faecalis, Lactococcus lactis and Streptococcus bovis genera by
ARDRA analysis (amplified ribosomal DNA restriction analysis). Among them,
16 Ent. faecalis, 26 L. lactis and 39 Strep. bovis displayed high minimum inhibi-
tory concentration (MIC) values for tetracycline, while 17 L. lactis showed high
MIC values for both tetracycline and erythromycin. Strain typing and molecu-
lar analysis of the phenotypically resistant isolates demonstrated the presence of
the tet(M) gene in the tetracycline-resistant strains and of tet(S) and erm(B) in
the double-resistant strains. Southern blot analysis revealed plasmid localization
of L. lactis tet(M), as well as of the erm(B) and tet(S) genes. Genetic linkage of
erm(B) and tet(S) was also demonstrated by PCR amplification. Conjugation
experiments demonstrated horizontal transfer to Ent. faecalis strain JH2-2 only
for the plasmid-borne L. lactis tet(M) gene.
Conclusions: We characterized tetracycline-and erythromycin-resistance genes
in coccal species, representing the fermenting microflora of a typical Italian
dairy product.
Significance and Impact of the Study: These results are of particular relevance
from the food safety viewpoint, especially in the light of the potential risk of
horizontal transfer of antibiotic-resistance genes among foodborne commensal
bacteria.
Journal of Applied Microbiology ISSN 1364-5072
ª 2010 INRAN
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 313–323 313

turn into opportunistic pathogens in immunocompro-
mised individuals, leading to nosocomial infections such
as bacteraemia or endocarditis (Bates 1997; Woodford
1998; Franz et al. 2003). It is therefore of crucial impor-
tance to identify the presence of AbR strains in fermented
foods for human consumption.
Other than enterococci, Gram-positive cocci involved
in manufacturing and preservation of fermented foods
belong to other genera, including Lactococcus and Strepto-
coccus (Liu 2003). Several examples of AbR LAB isolated
from raw meat and from cheese have been recently
reported (reviewed in Mathur and Singh 2005; Ammor
et al. 2007). Genes conferring resistance to tetracycline,
chloramphenicol, erythromycin and vancomycin have
been detected and characterized in Lactococcus lactis
(Perreten et al. 1997) and in enterococci (Giraffa and Sis-
to 1997; Teuber and Perreten 2000; Maietti et al. 2007)
isolated from fermented meat and milk products. Tradi-
tional products typical of mediterranean countries are
particularly rich in live bacteria of environmental origin,
as they do not employ selected industrial starters (Morea
et al. 1998). Such fermented foods can therefore be
considered potential vehicles for the spread of AbR LAB
to consumers through the food chain (Teuber and Perre-
ten 2000).
In our work, aimed at evaluating the presence of AbR
commensal bacteria along the manufacturing process of
the Italian traditional cheese Mozzarella di Bufala
Campana (MBC), we have established a collection of 555
LAB colonies, comprising both lactobacilli and cocci, iso-
lated from MRS (de Man, Rogosa, Sharp) plates contain-
ing tetracycline, erythromycin or kanamycin (Devirgiliis
et al. 2008). In this study, we analyse the coccal compo-
nent of our LAB collection and we report on its taxo-
nomic identity, genotypic diversity and molecular
identification of the genes responsible for the observed
phenotypic resistance to antibiotics.
Materials and methods
Bacterial strains and growth conditions
Bacterial isolation from dairy samples was previously
described (Devirgiliis et al. 2008). All cultures were
routinely grown in MRS, added or not with the antibiot-
ics under study, at 30�C (L. lactis) or 37�C (Ent. faecalis
and Streptococcus bovis) for 24–48 h, under anaerobic
conditions, and stored at )80�C in 15% (v ⁄ v) glycerol.
Enterococcus faecalis JH2-2 (LMG 19456), resistant to
rifampin and fusidic acid, grown in brain heart broth
(Merck, Darmstadt, Germany) at 37�C, was obtained
from the BCCM ⁄ LMG Bacteria Collection, Belgium
(Belgian Co-ordinated collection of Micro-organisms/
Laboratory of Microbiology, Gent University), and was
used as recipient strain in filter matings. Enterococcus
faecalis RE25 (Teuber et al. 2003), kindly provided by
G. Giraffa, Lodi, Italy, was used as positive control of
conjugal transfer.
DNA extraction and molecular analysis
Genomic DNA was extracted and amplified by PCR as
previously described (Devirgiliis et al. 2009). Plasmid
DNA was isolated with the procedure described by
Anderson and McKay (1983). Primers used are listed in
Table 1. Southern hybridization was performed by stan-
dard protocols, using probes labelled with digoxigenin-
11-dUTP (Roche Diagnostics, Milan, Italy).
Species identification by ARDRA and 16S rDNA gene
sequence analysis
The eubacterial P0–P6 primer pair (Table 1) (Invitrogen
Life Technologies, Milan, Italy) was used to amplify16S
rDNA gene fragments. Taxonomic strain identification
Table 1 Primers and conditions used in PCR amplifications
Primer pair Sequence Target gene
Annealing
T (�C)
Amplicon
size (bp) Reference for primers
P0 GAGAGTTTGATCCTGGCT 16S rDNA 55 1500 Di Cello and Fani (1996)
P6 CTACGGCTACCTTGTTAC
tetM-1 GAACTCGAACAAGAGGAAAGC tet(M) 60 740 Olsvik et al. (1995)
tetM-2 ATGGAAGCCCAGAAAGGAT
tetS7-F (FS) TTTGCCAGATGTTTATCAAG tet(S) 50 980 L. Morelli, personal communication
tetS8-R AGGCTCTCATACTGAATG
ermB1 CATTTAACGACGAAACTGGC erm(B) 55 406 Gevers et al. (2003)
ermB2 (RB) GGAACATCTGTGGTATGGCG
ermC-1 ATCTTTGAAATCGGCTCAGG erm(C) 58 294 Jensen et al. (1998)
ermC-2 CAAACCCGTATTCCACGATT
FB TTACCCGCCATACCACAGATGTTCC erm(B)–tet(S) 64 7000 This study
RS CATTTCCCACAATTACTGTCTCCCATTGTTCTGG
Antibiotic-resistant foodborne cocci C. Devirgiliis et al.
314 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 313–323
ª 2010 INRAN

was performed by the restriction of amplified fragment
with AvaII enzyme (Promega Italia, Milan, Italy). When
necessary, the DNA sequences of the amplified 16S rDNA
fragments were compared with those reported in the
Basic Blast database (Altschul et al. 1997).
Rep-PCR fingerprinting
Rep-PCR was performed with the (GTG)5 primer, as
described elsewhere (Gevers et al. 2001; Devirgiliis et al.
2009).
Minimum inhibitory concentration (MIC)
The broth microdilution method (Klare et al. 2005) was
used to evaluate MICs for each antibiotic, as previously
described (Devirgiliis et al. 2008).
Filter matings
In vitro conjugation experiments were performed as
described by Devirgiliis et al. (2009). Transconju-
gant colonies were recovered following incubation at
37�C for 24–72 h and characterized by Rep-PCR finger-
printing and PCR-based detection of the transferred
genes. Conjugation frequency is expressed as the ratio
between the number of transconjugants and donor
colonies.
Results
Species identification and strain typing
LAB colonies were isolated from raw milk (M) and nat-
ural whey starter cultures employed for fermentation
during the manufacturing process of the Italian tradi-
tional cheese MBC, as previously described (Devirgiliis
et al. 2008). Among these isolates, we selected 177 cocci
for further characterization at the species level. This was
achieved by restriction digestion of PCR-amplified 16S
rDNA (primers in Table 1) with specific endonucleases.
This approach allowed us to identify different species
within the collection, belonging to three genera: Ent.
faecalis, Strep. bovis and L. lactis. Figure 1 shows repre-
sentative ARDRA profiles resulting from digestion of the
amplified 16S rDNA fragments with AvaII, which gave
rise to distinct restriction patterns for each species. As a
control, we used the enzymes Sau3AI, to discriminate
L. lactis from Ent. faecalis and Strep. bovis and AvaI, to
further distinguish Ent. faecalis from Strep. bovis (data
not shown). Using this method, the collection of cocci
was found to contain the following species: 38 Ent.
faecalis (35 of which were isolated on MRS-Ery and 3
on MRS-Kan), 94 L. lactis (40 of which were isolated on
MRS, 21 on MRS-Ery and 33 on MRS-Tet) and 45
Strep. bovis (3 of which were isolated on MRS, 22 on
MRS-Kan and 20 on MRS-Tet). Taken together, these
results show that the majority of coccal colonies were
positively selected by erythromycin (32% of the total)
and tetracycline (30% of the total). Lactococcus lactis was
the most highly represented species (53%), followed by
Strep. bovis (25%) and Ent. faecalis (22%) (Table 2).
Using the rep-PCR technique, we obtained fingerprinting
profiles for all isolates. The dendrograms, constructed
after cluster analysis of the digitized (GTG)5-PCR finger-
prints with the unweighted pair group method with
arithmetic averages, are shown in Fig. 2a–c. The majority
of the observed fingerprinting profiles were unique,
revealing a high degree of biodiversity, which is typical
of traditional products with strong environmental influ-
ence. In particular, the 38 independent isolates of
Ent. faecalis displayed 19 different fingerprinting profiles,
resulting in 50% biodiversity (Fig. 2a and Table 2).
Streptococcus bovis and L. lactis, on the other hand,
M Sb Ef LI M
Figure 1 Species identification. Representative ARDRA profiles of the
three species, obtained by digestion of PCR-amplified 16S rDNA with
AvaII. Sb, Streptococcus bovis; Ef, Enterococcus faecalis; Ll, Lactococ-
cus lactis; M, size markers (Promega), 50-bp step ladder (left) and
1-kb ladder (right).
C. Devirgiliis et al. Antibiotic-resistant foodborne cocci
ª 2010 INRAN
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 313–323 315

exhibited lower biodiversity (about 30%), with 14 ⁄ 45
and 35 ⁄ 94 rep groups ⁄ total isolates, respectively
(Fig. 2b,c and Table 2).
Phenotypic antibiotic resistance
Each isolate was assayed for MIC for all three antibiot-
ics, irrespective of the selection medium from which
they originated. The resulting MIC values, shown in
Fig. 3 for erythromycin and tetracycline, are plotted as
overall distribution for all coccal isolates, irrespective of
species differences. As shown in the figure, we identified
98 strains (corresponding to 55Æ4% of the total) with
tetracycline MIC values above the breakpoint of 8 mg l)1.
These values, ranging between 16 and >128 mg l)1
(upper limit of test), were distributed in a bell-shaped
curve peaking at 64 mg l)1. The tetracycline MIC values
listed in Fig. 2a–c for each isolate indicate that pheno-
typically resistant strains were present in all three genera.
Among them, the majority (84Æ4%) belonged to
Strep. bovis species, 45Æ7% to L. lactis and 42% to
Ent. faecalis. On the contrary, the majority of erythro-
mycin MIC values were lower than the 4 mg l)1 break-
point, with only 17 isolates, corresponding to 9Æ6% of
the total coccal collection, all displaying the same MIC
value of 128 mg l)1 (Fig. 3b). All 17 erythromycin-resis-
tant isolates belonged to the L. lactis species, as shown
in Fig. 2c. Erythromycin-resistant L. lactis also displayed
64 mg l)1 tetracycline MIC values (framed in Fig. 2c).
Moreover, (GTG)5 fingerprinting analysis showed that
these 17 isolates grouped together, with highly similar
profiles. The erythromycin MIC values of the remaining
isolates of the collection, all below breakpoint, are not
shown in Fig. 2. MIC values for kanamycin are also not
shown, as none of the isolates displayed values above
breakpoint.
Genetic determinants of antibiotic resistance
Total DNA extracted from the phenotypically resistant
strains was PCR-amplified with primers specific for the
tetracycline and erythromycin-resistance genes most
commonly found in LAB, encoding ribosomal protection
proteins [RPP – tet(M), tet(S)] and methylases [erm(B),
erm(C)] (Table 1). In the majority of tetracycline-resistant
strains, we detected the presence of the tet(M) gene
(Fig. 2 and Table 2). In Ent. faecalis and Strep. bovis, the
presence of tet(M) determined MIC values between 16
and 64 mg l)1 (Fig. 2a,b and Table 2), with the exception
of one isolate of Strep. bovis with an MIC of 8 mg l)1
(equal to breakpoint). All isolates belonging to these two
species were susceptible to erythromycin. In L. lactis, we
detected the tet(M) gene in strains displaying tetracycline
MIC values within the range 64 to >128 mg l)1 (Fig. 2c
and Table 2). However, all isolates, within the 17 L. lactis
group phenotypically resistant to both antibiotics, shared
the same MIC value for tetracycline (64 mg l)1) and
erythromycin (128 mg l)1). These isolates were all shown
to harbour the tet(S) and erm(B) genes, while PCR ampli-
fication of the tet(M) and erm(C) genes was negative. To
investigate a possible association between erm(B) and
tet(S) genes in L. lactis, we used a PCR strategy that could
reveal, if the two genes are linked, also their orientations
relative to each other (Fig. 4a and Table 1). As shown in
Fig. 4b, we obtained the amplification of an intervening
7-kb fragment when using primer pair I (Fig. 4a and
Table 1), indicating that the two genes are indeed linked
and with the same 5¢–3¢ orientation. Previous reports
have indicated that erm genes are frequently linked to van
genes, conferring resistance to vancomycin (Jensen et al.
1998; Manson et al. 2003). However, determination of the
MIC values for vancomycin in these isolates showed that
all erm(B)-positive strains were vancomycin susceptible,
Table 2 Main features of coccal isolates
Species*
Total
rep-groups Source� Isolation medium* AbR*,�
Resistance
gene MIC range (mg l)1)
Enterococcus faecalis (38) 19 M, NWSC MRS-Ery (35) Tet (16) tet(M) 16–64
NWSC MRS-Kan (3)
Lactococcus lactis (94) 35 M, NWSC MRS (40) Tet (7) tet(M) 128
M MRS-Ery (21) Tet, Ery (17) tet(S) erm(B) 64 (tet); 128 (ery)
M, NWSC MRS-Tet (33) Tet (19) tet(M) 64 to >128
Streptococcus bovis (45) 14 M MRS (3) Tet (3) tet(M) 8–32
M, NWSC MRS-Kan (22) Tet (17) tet(M) 16–64
M, NWSC MRS-Tet (20) Tet (19) tet(M) 16–64
MIC, minimum inhibitory concentration.
*The number of isolates is reported in brackets.
�Source: M, raw water buffalo milk; NWSC, natural whey starter cultures.
�AbR, antibiotic-resistant isolates.
Antibiotic-resistant foodborne cocci C. Devirgiliis et al.
316 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 313–323
ª 2010 INRAN

with MIC values of 2 mg l)1 (data not shown). Table 2
summarizes the main features of the AbR Ent. faecalis,
L. lactis and Strep. bovis isolates. Three representative iso-
lates for each species, with distinct fingerprinting profiles,
were subjected to Southern hybridization with specific
tet(M), tet(S) or erm(B) probes. The tet(S) and erm(B)
genes in L. lactis, as well as the tet(M) genes in L. lactis,
Strep. bovis and Ent. faecalis, were all present in single
copy, as they showed a single positive hybridization band
on filters containing HindIII-digested genomic DNA
(Fig. 5, lanes 1, 3, 5, 7, 8). Furthermore, in the case of
L. lactis tet(S), erm(B) and tet(M), all three resistance
Pearson correlation (Opt: 0·35%) [0·0%–100·0%]GTG5
100
94·5
89·3
84·1
79·2
86·6
97·4
74·8
68·6 83·7
90·2
96·1
95·595
88·7
83·3
62·6
97·9
95·9
94·6
90·1
89·9
94·1
92
88·1
78·6
48·1
25·5
81·5
75·6
66·190·7
85·8 98
94·2
92·1
8983·4
90807060504030
GTG5
ID
1283 M 32 +
+++
+
++
+
+++++
+
++
–
–––––
–
–
–––––
––––
–
–
–
––
321664
222
222222
2
2
22222
2222
2
2
32
32
32323232
3232
64
64
16
16
MMMMMMMMMNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSC
NWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCMMMMM
12851282128112871289129312921296129813511353137013711373107213761380137413751378137713791367
13681369136513661364136313621361135212881291129913001284
Source MIC Tet tet (M)
Figure 2 Strain typing by rep-PCR. Composite figure of (GTG)5-PCR fingerprints of the Enterococcus faecalis (a), Streptococcus bovis (b),
Lactococcus lactis (c) isolates from raw milk or natural whey starter cultures samples. The dendrograms were constructed after cluster analysis of
the digitized (GTG)5-PCR fingerprints with the unweighted pair group method with arithmetic averages. Correlation levels are expressed as per-
centage of the Pearson correlation coefficient. Numerical designations indicate the ID number of each isolate. Minimum inhibitory concentration
values are expressed as mg l)1, and the presence ⁄ absence of the indicated antibiotic-resistant genes were determined by PCR analysis. The vertical
dotted line indicates 90% correlation (minimum level of reproducibility).
C. Devirgiliis et al. Antibiotic-resistant foodborne cocci
ª 2010 INRAN
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 313–323 317

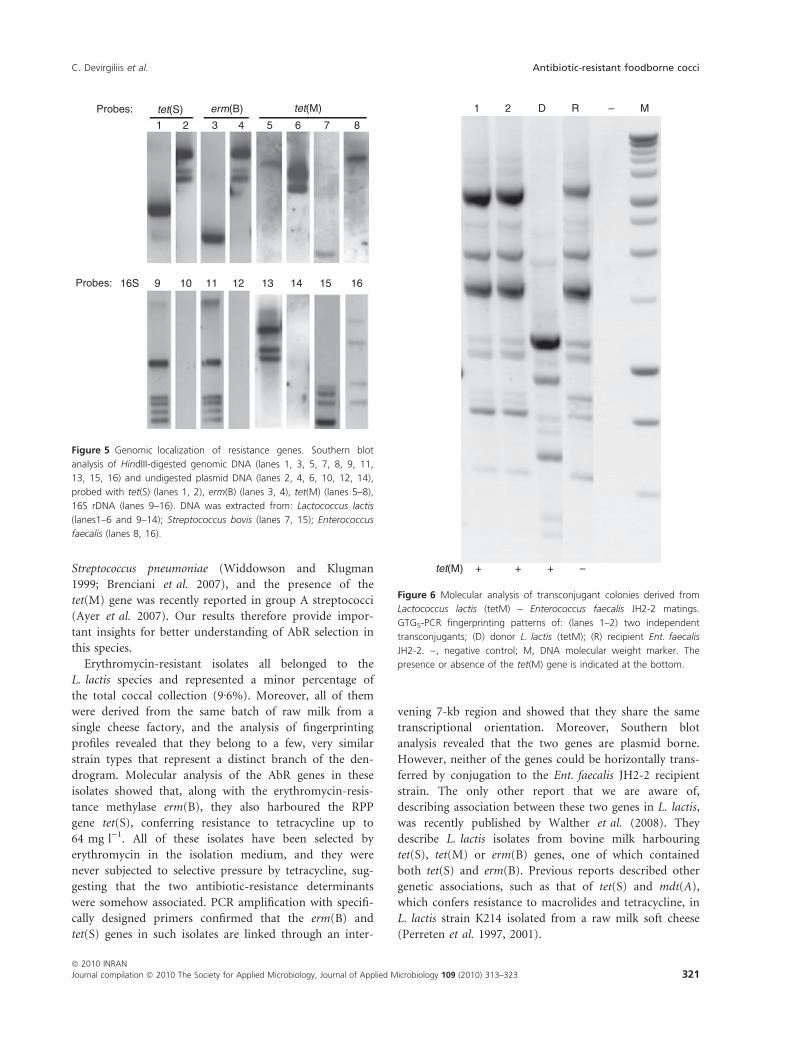
genes appeared to reside on plasmids, as positive signals
were detected in Southern hybridizations with purified
plasmid DNA preparations (Fig. 5, lanes 2, 4, 6).
To test whether any of the identified AbR genes could
be horizontally transferred, we performed conjugal trans-
fer experiments using Ent. faecalis JH2-2 as recipient
ID
1253 M32
32
32
32
323232
32
3232
3232
3232
3232
16
16
16
++++++++++++++
+
+++++++
+++++++++++
+++++
+
–
–
––
–
–
1616
16
16
1616
1616161616
8
2
2
22
2
2
6464
64646464
64
64
MMMMMMMMNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCMMMMMMMMMMMMMMMMMMM
13191317132012801316127612791271135713581398139914001395135413561350134313451347138413921349134213481059106110581064105010511052105310571278131512691273127712591268127012611252
Source MIC Tet tet (M)
Pearson correlation (Opt: 0·68%) [0·0%–100·0%]GTG5 GTG5
100
70·9
86·4
67·6
79·8 96·7
96·7
97·291·8
97·5
99·496·8
91·1
48·1
7·6
98·998·4
88·698
99.498·697·1
79·995·6
99.4
97·4
78·4
96·2
96·1
92·5
87·3 97·7
96·9
98·296·1
92·6
88·5
79·2 97·6
73·5 97·7
9591·9
90·2
76·3
80604020
Figure 2 (Continued)
Antibiotic-resistant foodborne cocci C. Devirgiliis et al.
318 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 313–323
ª 2010 INRAN

GTG5 GTG5ID
1303 M >128 +++++++++++++
+++++++
+
––––
––
–––––––––––
–––––––––––––––––––
––
–––––––––––––– +
tet (S) MIC Ery erm (B)
128128128128128128128128128128128128128128128128128
++++++++++++++++
+++++++++++++++++
––––––––––––––––
+++
+
+
>128
>128>128
128
128128128
128128
128
128128128128128128128
>128
64
64
2222
2
22
2
2
222
222222222222222
22
2222222222222
6464646464646464646464646464646464
>128>128
128
64
4
44
444
44
128
2
MMMMMMMMMMMMM
MMMMMMMMMM
M
M
MMMMMMMMMMMMM
M
MM
M
M
MMMMMMMMMMMMMMMMM
NWSCNWSCNWSC
NWSC
NWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSC
NWSCNWSCNWSCNWSC
NWSC
NWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSCNWSC
NWSC
1304130713011302130813091312131313181314131013111290132213311338125412551256125812491305130612471248128613811257138613871389139013911393139413961397124113831385138213881294129712511246996999
1000993995992994
9971004
13231260104810491046132113241325132913351333133413301336132613271328133913401295133710291031103810301039104310261044103610371040104210281034103310321056
Source MIC Tet tet (M)
20 40 60 80 100
Figure 2 (Continued)
C. Devirgiliis et al. Antibiotic-resistant foodborne cocci
ª 2010 INRAN
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 313–323 319

strain and three representative isolates for each species as
donors. The results of such filter mating experiments
showed that only plasmid-borne tet(M) from L. lactis was
transferred to Ent. faecalis at relatively high frequency
(2 · 10)5). Molecular typing of two independent trans-
conjugants obtained from such matings was performed by
(GTG)5 fingerprinting, as shown in Fig. 6. Comparison of
the resulting profiles with those of the donor and recipi-
ent strains confirms that horizontal transfer of the tetra-
cycline-resistance determinant present in L. lactis tet(M)
could confer tetracycline resistance to Ent. faecalis.
Discussion
In this study, we report the microbiological characteriza-
tion and the antibiotic resistance of a collection of 177
cocci isolated from the raw ingredients employed in man-
ufacturing of the Italian cheese MBC. Molecular analysis
of phenotypically resistant strains showed that tetracycline
and erythromycin MIC values above breakpoints were
always associated with the presence of genetic determi-
nants of antibiotic resistance. A total of 81 tetracycline-
resistant Ent. faecalis, Strep. bovis and L. lactis harboured
the tet(M) gene, confirming that it is the most common
tetracycline-resistance determinant among LAB (Ammor
et al. 2007). This gene was also reported as the most
frequent in AbR LAB isolated from poultry and swine
meat products (Aquilanti et al. 2007).
Our results show chromosomal localization of tet(M)
in the Ent. faecalis and Strep. bovis tetracycline-resistant
strains, while in the case of L. lactis the gene is likely
carried by a yet unidentified plasmid. The presence of
tet(M) in Ent. faecalis did not appear to be specifically
selected by tetracycline, as tetracycline-resistant isolates
were collected almost exclusively from erythromycin-con-
taining MRS plates. On the other hand, tet(M)-positive
L. lactis and Strep. bovis were positively selected by the
initial plating conditions, as the majority of them were
isolated on MRS-Tet plates. Tetracycline resistance in
Ent. faecalis was described in several reports (Huys et al.
2004; Rizzotti et al. 2005; Wilcks et al. 2005), while only
few studies have been published in L. lactis. Florez et al.
(2008) identified and characterized two tet(M)-positive
L. lactis strains isolated from a raw milk, starter-free
traditional cheese. On the contrary, no data are available
to our knowledge on tetracycline resistance in Strep. bovis.
Up to now, investigations were mainly focused on
pathogenic streptococci, such as Streptococcus pyogenes or
Tetracycline
Erythromycin
*
*2
807060504030
N is
olat
es
2010
0
807060504030N
isol
ates
2010
0
4 8 16 32 64
MIC (mg l–1)
MIC (mg l–1)
0·25 0·5 1 2 4 8 16 128
128 >128
(a)
(b)
Figure 3 Minimum inhibitory concentration (MIC) distribution. Bars
represent the number of isolates displaying the same MIC value,
expressed as mg l)1, for tetracycline (panel a) and erythromycin
(panel b). The data reported in each panel refer to all isolates,
rrespective of the species. Asterisks indicate antibiotic breakpoint con-
centrations added to the isolation medium.
I(a)
(b)
erm(B) tet(S)
RS
RS
FS
FB
erm(B) tet(S)
RB
erm(B)
erm(B)
MKb8-
6-
I II III IV
tet(S)
tet(S)
RB
FSFB
II
III
IV
Figure 4 Association between Lactococcus lactis erm(B) and tet(S)
genes. (a) Graphical representation of the PCR strategy employed to
investigate physical association of the erm(B) and tet(S) genes. Large
arrows represent the two genes in all possible orientations (I–IV), and
primer pairs are indicated by small arrows. F, forward primers; R,
reverse primers. (b) Agarose gel electrophoresis of PCR products
obtained with primer pairs I–IV. M, DNA molecular weight marker.
Antibiotic-resistant foodborne cocci C. Devirgiliis et al.
320 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 313–323
ª 2010 INRAN
Streptococcus pneumoniae (Widdowson and Klugman
1999; Brenciani et al. 2007), and the presence of the
tet(M) gene was recently reported in group A streptococci
(Ayer et al. 2007). Our results therefore provide impor-
tant insights for better understanding of AbR selection in
this species.
Erythromycin-resistant isolates all belonged to the
L. lactis species and represented a minor percentage of
the total coccal collection (9Æ6%). Moreover, all of them
were derived from the same batch of raw milk from a
single cheese factory, and the analysis of fingerprinting
profiles revealed that they belong to a few, very similar
strain types that represent a distinct branch of the den-
drogram. Molecular analysis of the AbR genes in these
isolates showed that, along with the erythromycin-resis-
tance methylase erm(B), they also harboured the RPP
gene tet(S), conferring resistance to tetracycline up to
64 mg l)1. All of these isolates have been selected by
erythromycin in the isolation medium, and they were
never subjected to selective pressure by tetracycline, sug-
gesting that the two antibiotic-resistance determinants
were somehow associated. PCR amplification with specifi-
cally designed primers confirmed that the erm(B) and
tet(S) genes in such isolates are linked through an inter-
vening 7-kb region and showed that they share the same
transcriptional orientation. Moreover, Southern blot
analysis revealed that the two genes are plasmid borne.
However, neither of the genes could be horizontally trans-
ferred by conjugation to the Ent. faecalis JH2-2 recipient
strain. The only other report that we are aware of,
describing association between these two genes in L. lactis,
was recently published by Walther et al. (2008). They
describe L. lactis isolates from bovine milk harbouring
tet(S), tet(M) or erm(B) genes, one of which contained
both tet(S) and erm(B). Previous reports described other
genetic associations, such as that of tet(S) and mdt(A),
which confers resistance to macrolides and tetracycline, in
L. lactis strain K214 isolated from a raw milk soft cheese
(Perreten et al. 1997, 2001).
Probes:
Probes: 16S 9 10 11 12 13 14 15 16
tet(S)
1 2 3 4 5 6 7 8
erm(B) tet(M)
Figure 5 Genomic localization of resistance genes. Southern blot
analysis of HindIII-digested genomic DNA (lanes 1, 3, 5, 7, 8, 9, 11,
13, 15, 16) and undigested plasmid DNA (lanes 2, 4, 6, 10, 12, 14),
probed with tet(S) (lanes 1, 2), erm(B) (lanes 3, 4), tet(M) (lanes 5–8),
16S rDNA (lanes 9–16). DNA was extracted from: Lactococcus lactis
(lanes1–6 and 9–14); Streptococcus bovis (lanes 7, 15); Enterococcus
faecalis (lanes 8, 16).
1 2 D R – M
tet(M) + + + –
Figure 6 Molecular analysis of transconjugant colonies derived from
Lactococcus lactis (tetM) – Enterococcus faecalis JH2-2 matings.
GTG5-PCR fingerprinting patterns of: (lanes 1–2) two independent
transconjugants; (D) donor L. lactis (tetM); (R) recipient Ent. faecalis
JH2-2. ), negative control; M, DNA molecular weight marker. The
presence or absence of the tet(M) gene is indicated at the bottom.
C. Devirgiliis et al. Antibiotic-resistant foodborne cocci
ª 2010 INRAN
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 313–323 321

Efficient gene transfer by conjugation is indicative of
the presence of either plasmids or mobile elements such
as transposons (Beaber et al. 2002; Pembroke et al. 2002;
Teuber et al. 2003; Alekshun and Levy 2007). Consis-
tently, we observed high transfer efficiency of plasmid-
borne L. lactis tet(M) to Ent. faecalis. However, our data
on the low efficiency, or lack of transfer of Ent. faecalis
and Strep. bovis tet(M), as well as of the erm(B) and tet(S)
genes of L. lactis, suggest their presence within genetic
contexts lacking mobile elements or in nonconjugative
plasmids. However, in vitro conjugal transfer is not
necessarily indicative of the in vivo situation; therefore,
we cannot directly extrapolate these results to the fre-
quency of horizontal transfer potentially occurring among
species. Moreover, we cannot exclude that species barriers
might have impaired in vitro conjugal transfer in some
cases. We have previously reported the analysis of about
300 Lactobacillus isolates from the Italian dairy product
MBC and from the raw ingredients employed for its man-
ufacturing (Devirgiliis et al. 2008). The results presented
here complete the analysis of the two predominant micro-
bial groups (lactobacilli and cocci) that constitute the fer-
menting microflora in this product and allow comparison
of their features with respect to antibiotic resistance. Con-
sidering the total microbial community, composed of 555
lactobacilli and cocci (Devirgiliis et al. 2008), AbR cocci
represented 17Æ6% of the total bacteria found in MBC
samples (14Æ6% comprising tetracycline-resistant isolates
and 3% comprising erythromycin-resistant isolates). This
value is higher than that observed for AR lactobacilli
(3Æ8% of the total bacteria), all displaying tetracycline
resistance (Devirgiliis et al. 2008). Such resistant species
most likely reflect the persistence of AR bacteria in the
environment and probably relate to the use and misuse of
antibiotics in human therapy and animal farming.
Noteworthy, tetracycline- and ⁄ or erythromycin-resistant
species were found exclusively in isolates from the raw
ingredients employed for MBC production, while absent
in the final processed cheese. This result is of particular
relevance from the food safety viewpoint, as it points to a
major role of technological procedures employed in
cheese production in influencing the composition of fer-
menting microflora and, as a consequence, also in affect-
ing the occurrence of AbR species.
Acknowledgements
The Authors thank Kariklia Pascucci for her kind support
in daily lab work. We are grateful to Consorzio MBC
(http://www.mozzarelladop.it) for cooperation in the col-
lection of samples. This work was supported by grant
ARAFOA (DM 662 ⁄ 7303 ⁄ 2003) from the Italian Ministry
of Agriculture, Food & Fisheries (MiPAAF).
References
Alekshun, M.N. and Levy, S.B. (2007) Molecular mechanisms
of antibacterial multidrug resistance. Cell 128, 1037–1050.
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang,
Z., Miller, W. and Lipman, D.J. (1997) Gapped BLAST
and PSI-BLAST: a new generation of protein database
search programs. Nucleic Acids Res 25, 3389–3402.
Ammor, M.S., Florez, A.B. and Mayo, B. (2007) Antibiotic
resistance in non-enterococcal lactic acid bacteria and bifi-
dobacteria. Food Microbiol 24, 559–570.
Anderson, D.G. and McKay, L.L. (1983) Simple and rapid
method for isolating large plasmid DNA from lactic strep-
tococci. Appl Environ Microbiol 46, 549–552.
Aquilanti, L., Garofalo, C., Osimani, A., Silvestri, G., Vignaroli,
C. and Clementi, F. (2007) Isolation and molecular charac-
terization of antibiotic-resistant lactic acid bacteria from
poultry and swine meat products. J Food Prot 70, 557–565.
Ayer, V., Tewodros, W., Manoharan, A., Skariah, S., Luo, F.
and Bessen, D.E. (2007) Tetracycline resistance in group a
streptococci: emergence on a global scale and influence on
multiple-drug resistance. Antimicrob Agents Chemother 51,
1865–1868.
Bates, J. (1997) Epidemiology of vancomycin-resistant entero-
cocci in the community and the relevance of farm animals
to human infection. J Hosp Infect 37, 89–101.
Beaber, J.W., Burrus, V., Hochhut, B. and Waldor, M.K.
(2002) Comparison of SXT and R391, two conjugative
integrating elements: definition of a genetic backbone for
the mobilization of resistance determinants. Cell Mol Life
Sci 59, 2065–2070.
Brenciani, A., Bacciaglia, A., Vecchi, M., Vitali, L.A., Varaldo,
P.E. and Giovanetti, E. (2007) Genetic elements carrying
erm(B) in Streptococcus pyogenes and association with
tet(M) tetracycline resistance gene. Antimicrob Agents
Chemother 51, 1209–1216.
Devirgiliis, C., Caravelli, A., Coppola, D., Barile, S. and Perozzi,
G. (2008) Antibiotic resistance and microbial composition
along the manufacturing process of Mozzarella di Bufala
Campana. Int J Food Microbiol 128, 378–384.
Devirgiliis, C., Coppola, D., Barile, S., Colonna, B. and Perozzi,
G. (2009) Characterization of the Tn916 conjugative trans-
poson in a food-borne strain of Lactobacillus paracasei.
Appl Environ Microbiol 75, 3866–3871.
Di Cello, F. and Fani, R. (1996) A molecular strategy for the
study of natural bacterial communities by PCR-based tech-
niques. Minerva Biotecnol 8, 126–134.
Florez, A.B., Ammor, M.S. and Mayo, B. (2008) Identification
of tet(M) in two Lactococcus lactis strains isolated from a
Spanish traditional starter-free cheese made of raw milk
and conjugative transfer of tetracycline resistance to lacto-
cocci and enterococci. Int J Food Microbiol 121, 189–194.
Franz, C.M., Stiles, M.E., Schleifer, K.H. and Holzapfel, W.H.
(2003) Enterococci in foods – a conundrum for food
safety. Int J Food Microbiol 88, 105–122.
Antibiotic-resistant foodborne cocci C. Devirgiliis et al.
322 Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 313–323
ª 2010 INRAN

Gevers, D., Huys, G. and Swings, J. (2001) Applicability of
rep-PCR fingerprinting for identification of Lactobacillus
species. FEMS Microbiol Lett 205, 31–36.
Gevers, D., Danielsen, M., Huys, G. and Swings, J. (2003)
Molecular characterization of tet(M) genes in Lactobacillus
isolates from different types of fermented dry sausage. Appl
Environ Microbiol 69, 1270–1275.
Giraffa, G. and Sisto, F. (1997) Susceptibility to vancomycin of
enterococci isolated from dairy products. Lett Appl Micro-
biol 25, 335–338.
Huys, G., D’Haene, K., Collard, J.M. and Swings, J. (2004)
Prevalence and molecular characterization of tetracycline
resistance in Enterococcus isolates from food. Appl Environ
Microbiol 70, 1555–1562.
Jensen, L.B., Ahrens, P., Dons, L., Jones, R.N., Hammerum,
A.M. and Aarestrup, F.M. (1998) Molecular analysis of
Tn1546 in Enterococcus faecium isolated from animals and
humans. J Clin Microbiol 36, 437–442.
Klare, I., Konstabel, C., Muller-Bertling, S., Reissbrodt, R.,
Huys, G., Vancanneyt, M., Swings, J., Goossens, H. et al.
(2005) Evaluation of new broth media for microdilution
antibiotic susceptibility testing of Lactobacilli, Pediococci,
Lactococci, and Bifidobacteria. Appl Environ Microbiol 71,
8982–8986.
Liu, S.Q. (2003) Practical implications of lactate and pyruvate
metabolism by lactic acid bacteria in food and beverage
fermentations. Int J Food Microbiol 83, 115–131.
Maietti, L., Bonvini, B., Huys, G. and Giraffa, G. (2007) Inci-
dence of antibiotic resistance and virulence determinants
among Enterococcus italicus isolates from dairy products.
Syst Appl Microbiol 30, 509–517.
Manson, J.M., Keis, S., Smith, J.M. and Cook, G.M. (2003) A
clonal lineage of VanA-type Enterococcus faecalis predomi-
nates in vancomycin-resistant Enterococci isolated in New
Zealand. Antimicrob Agents Chemother 47, 204–210.
Mathur, S. and Singh, R. (2005) Antibiotic resistance in food
lactic acid bacteria – a review. Int J Food Microbiol 105,
281–295.
Morea, M., Baruzzi, F., Cappa, F. and Cocconcelli, P.S. (1998)
Molecular characterization of the Lactobacillus community
in traditional processing of Mozzarella cheese. Int J Food
Microbiol 43, 53–60.
Olsvik, B., Olsen, I. and Tenover, F.C. (1995) Detection of
tet(M) and tet(O) using the polymerase chain reaction in
bacteria isolated from patients with periodontal disease.
Oral Microbiol Immunol 10, 87–92.
Pembroke, J.T., MacMahon, C. and McGrath, B. (2002) The
role of conjugative transposons in the Enterobacteriaceae.
Cell Mol Life Sci 59, 2055–2064.
Perreten, V., Schwarz, F., Cresta, L., Boeglin, M., Dasen, G.
and Teuber, M. (1997) Antibiotic resistance spread in
food. Nature 389, 801–802.
Perreten, V., Schwarz, F.V., Teuber, M. and Levy, S.B. (2001)
Mdt(A), a new efflux protein conferring multiple antibiotic
resistance in Lactococcus lactis and Escherichia coli. Antimic-
rob Agents Chemother 45, 1109–1114.
Rizzotti, L., Simeoni, D., Cocconcelli, P., Gazzola, S.,
Dellaglio, F. and Torriani, S. (2005) Contribution of
enterococci to the spread of antibiotic resistance in the
production chain of swine meat commodities. J Food
Prot 68, 955–965.
Teuber, M. (2001) Veterinary use and antibiotic resistance.
Curr Opin Microbiol 4, 493–499.
Teuber, M. and Perreten, V. (2000) Role of milk and meat
products as vehicles for antibiotic-resistant bacteria. Acta
Vet Scand Suppl 93, 75–87; discussion 111–117.
Teuber, M., Schwarz, F. and Perreten, V. (2003) Molecular
structure and evolution of the conjugative multiresistance
plasmid pRE25 of Enterococcus faecalis isolated from a
raw-fermented sausage. Int J Food Microbiol 88, 325–329.
Walther, C., Rossano, A., Thomann, A. and Perreten, V.
(2008) Antibiotic resistance in Lactococcus species from
bovine milk: presence of a mutated multidrug transporter
mdt(A) gene in susceptible Lactococcus garviae strains. Vet
Microbiol 131, 348–357.
Wegener, H.C. (2003) Antibiotics in animal feed and their role
in resistance development. Curr Opin Microbiol 6, 439–
445.
Widdowson, C.A. and Klugman, K.P. (1999) Molecular mech-
anisms of resistance to commonly used non-betalactam
drugs in Streptococcus pneumoniae. Semin Respir Infect 14,
255–268.
Wilcks, A., Andersen, S.R. and Licht, T.R. (2005) Characteriza-
tion of transferable tetracycline resistance genes in Entero-
coccus faecalis isolated from raw food. FEMS Microbiol Lett
243, 15–19.
Woodford, N. (1998) Glycopeptide-resistant enterococci: a
decade of experience. J Med Microbiol 47, 849–862.
C. Devirgiliis et al. Antibiotic-resistant foodborne cocci
ª 2010 INRAN
Journal compilation ª 2010 The Society for Applied Microbiology, Journal of Applied Microbiology 109 (2010) 313–323 323

Copyright of Journal of Applied Microbiology is the property of Wiley-Blackwell and its content may not be
copied or emailed to multiple sites or posted to a listserv without the copyright holder's express written
permission. However, users may print, download, or email articles for individual use.


















