
Identification of a Mammalian Homologue of the Fungal Tom70 Mitochondrial Precursor Protein Import...
10
Identification of a Mammalian Homologue of the Fungal Tom70 Mitochondrial Precursor Protein Import Receptor as a Thyroid Hormone-Regulated Gene in Specific Brain Regions Manuel Alvarez-Dolado, *²Marı´a Gonza ´lez-Moreno, *Alfonso Valencia, ‡Martin Zenke, Juan Bernal, and Alberto Mun ˜oz Instituto de Investigaciones Biome ´dicas “Alberto Sols,” Consejo Superior de Investigaciones Cientı ´ficas (CSIC)–Universidad Auto ´noma de Madrid (UAM), *Centro Nacional de Biotecnologı ´a, Consejo Superior de Investigaciones Cientı ´ficas, Cantoblanco, and ² Centro de Astrobiologı ´a, INTA, Madrid, Spain; and ‡Max-Delbru ¨ck Center for Molecular Medicine, Berlin, Germany Abstract: Thyroid hormone is an important regulator of mammalian brain maturation. By differential display PCR, we isolated a cDNA clone (S2) that is specifically up- regulated in the striatum of neonatal hypothyroid rats. S2 was identified as KIAA0719, the first human gene dis- tantly homologous to the fungal Tom70, which encodes a member of the translocase mitochondrial outer mem- brane complex involved in the import of preproteins into the mitochondria. By northern and in situ hybridization studies, KIAA0719 was found to be up-regulated in the striatum, nucleus accumbens, and discrete cortical layers of 15-day-old hypothyroid rats. In contrast, lower expres- sion was found in the olfactory tubercle, whereas no differences were detected in other brain regions. Signifi- cantly, treatment of hypothyroid animals with single in- jections of thyroxine restored the normal levels of KIAA0719 expression. Moreover, treatment of control an- imals with thyroxine led to a reduced expression, dem- onstrating a negative hormonal regulation in vivo. Thus, KIAA0719 gene expression is regulated by thyroid hor- mone in the neonatal rat brain in a region-specific fash- ion. Given the role of the homologous Tom70 gene, the alteration of KIAA0719 expression may contribute to the changes in mitochondrial morphology and physiology caused by hypothyroidism in the developing rat brain. Key Words: Thyroid hormone —KIAA0719 gene —Mito- chondrial protein import receptor—Tom70 —Striatum— Differential display. J. Neurochem. 73, 2240 –2249 (1999). Thyroid hormone [3,39,5-triiodothyronine (T3); thy- roxine (T4)] is a main regulator of growth, development, and homeostasis in higher organisms. Lack of adequate levels of thyroid hormones during the fetal and neonatal periods leads to mental deficiency in humans and multi- ple brain abnormalities in experimental animals, such as the rat (Legrand, 1984, 1986; Porterfield and Hendrich, 1993; Bernal and Nunez, 1995). These abnormalities include biochemical, metabolic, cellular, and behavioral alterations throughout the brain, including a reduction in the mean size of neuronal cell bodies, a decreased den- sity and abnormal distribution of dendritic spines, changes in the number of particular cellular populations and in synaptogenesis, reduced arborization and delocal- ization of cerebellar Purkinje cells, retarded and dimin- ished myelination, and migration defects (Schwartz and Oppenheimer, 1997; Bernal and Guadan ˜o-Ferraz, 1998). Thyroid hormone acts by regulating target gene ex- pression through binding to its high-affinity receptor [thyroid receptor (TR)], which belongs to the superfam- ily of nuclear hormone receptors that function as ligand- regulated transcription factors (Sap et al., 1986; Wein- berger et al., 1986). The different TR isoforms are dis- tinctly expressed in a time- and region-specific fashion in the developing brain, presumably regulating the tran- scription of a particular set of genes (Mellstro ¨m et al., 1991; Bradley et al., 1992). In the last few years, a number of genes coding for cytoskeletal proteins, tran- scription factors, enzymes, cell-adhesion molecules, neu- rotrophins and their receptors, or other proteins have been found to be under thyroid control in the rodent brain during the neonatal period (for review, see Bernal and Guadan ˜o-Ferraz, 1998). It is, however, clear that addi- tional thyroid hormone target genes must exist, and that their identification will increase our understanding of the molecular events underlying cerebral function and brain development. To identify new genes regulated by thyroid hormone in the brain, we have used the differential display PCR Received May 4, 1999; revised manuscript received August 3, 1999; accepted August 3, 1999. Address correspondence and reprint requests to Dr. A. Mun ˜oz at Instituto de Investigaciones Biome ´dicas “Alberto Sols,” Arturo Dup- erier 4, E-28029 Madrid, Spain. Abbreviations used: Cy, cyclophilin; MAG, myelin-associated gly- coprotein; P, postnatal day; PBS, phosphate-buffered saline; SSC, standard saline citrate; T3, 3,39,5-triiodothyronine; T4, thyroxine; Tom, translocase of the outer mitochondrial membrane; TPR, tetratricopep- tide repeat; TR, thyroid receptor. 2240 Journal of Neurochemistry Lippincott Williams & Wilkins, Inc., Philadelphia © 1999 International Society for Neurochemistry
Transcript of Identification of a Mammalian Homologue of the Fungal Tom70 Mitochondrial Precursor Protein Import...

Identification of a Mammalian Homologue of the Fungal Tom70Mitochondrial Precursor Protein Import Receptor as a Thyroid
Hormone-Regulated Gene in Specific Brain Regions
Manuel Alvarez-Dolado, *†Marı´a Gonza´lez-Moreno, *Alfonso Valencia, ‡Martin Zenke,Juan Bernal, and Alberto Mun˜oz
Instituto de Investigaciones Biome´dicas “Alberto Sols,” Consejo Superior de Investigaciones Cientı´ficas (CSIC)–UniversidadAutonoma de Madrid (UAM),*Centro Nacional de Biotecnologı´a, Consejo Superior de Investigaciones Cientı´ficas, Cantoblanco,
and †Centro de Astrobiologı´a, INTA, Madrid, Spain; and‡Max-Delbruck Center for Molecular Medicine, Berlin, Germany
Abstract: Thyroid hormone is an important regulator ofmammalian brain maturation. By differential display PCR,we isolated a cDNA clone (S2) that is specifically up-regulated in the striatum of neonatal hypothyroid rats. S2was identified as KIAA0719, the first human gene dis-tantly homologous to the fungal Tom70, which encodes amember of the translocase mitochondrial outer mem-brane complex involved in the import of preproteins intothe mitochondria. By northern and in situ hybridizationstudies, KIAA0719 was found to be up-regulated in thestriatum, nucleus accumbens, and discrete cortical layersof 15-day-old hypothyroid rats. In contrast, lower expres-sion was found in the olfactory tubercle, whereas nodifferences were detected in other brain regions. Signifi-cantly, treatment of hypothyroid animals with single in-jections of thyroxine restored the normal levels ofKIAA0719 expression. Moreover, treatment of control an-imals with thyroxine led to a reduced expression, dem-onstrating a negative hormonal regulation in vivo. Thus,KIAA0719 gene expression is regulated by thyroid hor-mone in the neonatal rat brain in a region-specific fash-ion. Given the role of the homologous Tom70 gene, thealteration of KIAA0719 expression may contribute to thechanges in mitochondrial morphology and physiologycaused by hypothyroidism in the developing rat brain.Key Words: Thyroid hormone —KIAA0719 gene —Mito-chondrial protein import receptor—Tom70—Striatum—Differential display.J. Neurochem. 73, 2240–2249 (1999).
Thyroid hormone [3,39,5-triiodothyronine (T3); thy-roxine (T4)] is a main regulator of growth, development,and homeostasis in higher organisms. Lack of adequatelevels of thyroid hormones during the fetal and neonatalperiods leads to mental deficiency in humans and multi-ple brain abnormalities in experimental animals, such asthe rat (Legrand, 1984, 1986; Porterfield and Hendrich,1993; Bernal and Nunez, 1995). These abnormalitiesinclude biochemical, metabolic, cellular, and behavioralalterations throughout the brain, including a reduction in
the mean size of neuronal cell bodies, a decreased den-sity and abnormal distribution of dendritic spines,changes in the number of particular cellular populationsand in synaptogenesis, reduced arborization and delocal-ization of cerebellar Purkinje cells, retarded and dimin-ished myelination, and migration defects (Schwartz andOppenheimer, 1997; Bernal and Guadan˜o-Ferraz, 1998).
Thyroid hormone acts by regulating target gene ex-pression through binding to its high-affinity receptor[thyroid receptor (TR)], which belongs to the superfam-ily of nuclear hormone receptors that function as ligand-regulated transcription factors (Sap et al., 1986; Wein-berger et al., 1986). The different TR isoforms are dis-tinctly expressed in a time- and region-specific fashion inthe developing brain, presumably regulating the tran-scription of a particular set of genes (Mellstro¨m et al.,1991; Bradley et al., 1992). In the last few years, anumber of genes coding for cytoskeletal proteins, tran-scription factors, enzymes, cell-adhesion molecules, neu-rotrophins and their receptors, or other proteins havebeen found to be under thyroid control in the rodent brainduring the neonatal period (for review, see Bernal andGuadan˜o-Ferraz, 1998). It is, however, clear that addi-tional thyroid hormone target genes must exist, and thattheir identification will increase our understanding of themolecular events underlying cerebral function and braindevelopment.
To identify new genes regulated by thyroid hormonein the brain, we have used the differential display PCR
Received May 4, 1999; revised manuscript received August 3, 1999;accepted August 3, 1999.
Address correspondence and reprint requests to Dr. A. Mun˜oz atInstituto de Investigaciones Biome´dicas “Alberto Sols,” Arturo Dup-erier 4, E-28029 Madrid, Spain.
Abbreviations used:Cy, cyclophilin; MAG, myelin-associated gly-coprotein; P, postnatal day; PBS, phosphate-buffered saline; SSC,standard saline citrate; T3, 3,39,5-triiodothyronine; T4, thyroxine; Tom,translocase of the outer mitochondrial membrane; TPR, tetratricopep-tide repeat; TR, thyroid receptor.
2240
Journal of NeurochemistryLippincott Williams & Wilkins, Inc., Philadelphia© 1999 International Society for Neurochemistry

technique (Liang and Pardee, 1992; Welsh et al., 1992;McClelland et al., 1995). This procedure has been shownpreviously to be useful to isolate genes differentiallyexpressed in the brain, including a thyroid-responsivecDNA expressed in the mature rat cerebrum (Dalal et al.,1994; Joseph et al., 1994; Douglass et al., 1995; Shahet al., 1997). We compared the pattern of PCR productsobtained from total RNA populations isolated from cer-ebellum, striatum, cortex, or hippocampus of hypothy-roid and euthyroid rats at postnatal day 15 (P15). Wechose P15 because at this developmental stage both theexpression and occupancy of TRs are maximal(Schwartz and Oppenheimer, 1997; Bernal andGuadan˜o-Ferraz, 1998). We report here the identificationof KIAA0719 gene, a mammalian homologue to a fungalcomponent (Tom70) of the translocase of the outer mi-tochondrial membrane (Tom) complex (Nagase et al.,1998) as a thyroid hormone-regulated gene in specificregions of the rat brain.
EXPERIMENTAL PROCEDURES
Materials2-Mercapto-1-methylimidazole and T4 were from Sigma
Chemical Co. Guanidinium isothiocyanate was from Fluka, andnylon membranes from Schleicher & Schuell. T7 and Sp6polymerases were purchased from Boehringer Mannheim. Am-pliTaq polymerase was from Perkin–Elmer. Radioisotopeswere from Amersham. The pCRII T/A cloning vector was fromInvitrogen (San Diego, CA, U.S.A.). All other chemicals werereagent grade (Boehringer Mannheim, Merck, or Sigma).
Induction of hypothyroidismWistar rats maintained in the animal facilities of our Instituto
de Investigaciones Biome´dicas were used. All efforts weremade to minimize animal suffering, to reduce the number ofanimals used, and to use alternatives to in vivo techniques. Themaintenance and handling of the animals were as recom-mended by the European Communities Council Directive ofNovember 24, 1986 (86/609/EEC). To induce neonatal hypo-thyroidism, 2-mercapto-1-methylimidazole (0.02%) was ad-ministered in the drinking water of the dams from the 9th dayafter conception and was continued until the animals werekilled. In addition, surgical thyroidectomy was performed at P5to ensure that the animals are hypothyroid during the entireneonatal period (Rodrı´guez-Pen˜a et al., 1993; Alvarez-Doladoet al., 1994, 1998). P0 animals were killed 8–12 h after birth.T4 was used for the in vivo hormonal treatments because itcrosses the blood–brain barrier more efficiently than T3 and isconverted to T3 in the brain (Dickson et al., 1987). T4 wasadministered as single daily intraperitoneal injections of 1.8mg/100 g of body weight starting 4 days before death. Ratswere killed 24 h after the last T4 injection. At least threeanimals were studied per experimental group to obtain repre-sentative values.
RNA extraction and northern analysisTotal RNA was obtained from pools of different brain re-
gions from a variable number of animals, depending on theweight of each region (four striata, two cortices, and threehippocampi or cerebella). Total RNA was prepared by theguanidinium isothiocyanate/phenol/chloroform procedure(Chomczynski and Sacchi, 1987). RNAs were fractionated in
formaldehyde–agarose gels and blotted onto nylon membranesfollowing standard techniques (Sambrook et al., 1989). Ascontrols for the amount and integrity of RNA, blots werestained in a 0.02% methylene blue solution made in 0.3Msodium acetate and rehybridized with a cyclophilin (Cy) cDNAprobe. To control the thyroid status, filters were also hybridizedwith a probe for myelin-associated glycoprotein (MAG), a genedown-regulated by hypothyroidism (Rodrı´guez-Pen˜a et al.,1993). Radioactive probes were prepared by the random prim-ing procedure (Feinberg and Vogelstein, 1983). Cy and MAGcDNA clones were a gift from Dr. J. G. Sutcliffe (ScrippsResearch Institute, San Diego, CA, U.S.A.). Autoradiogramswere analyzed using a La Cie scanner connected to a PowerMacintosh G3 computer using Adobe Photoshop 4.0LE andNIH Image programs.
Differential display PCRTotal RNA populations from the cerebellum, striatum, cor-
tex, or hippocampus of 15-day-old rats were retrotranscribed aspreviously described (Liang and Pardee, 1992). We usedNN(T)11 oligonucleotides as 39 primers. One tenth of eachretrotranscription reaction was subjected to PCR using Ampli-Taq in the presence of [a-33P]dATP and random decamers(Bauer et al., 1993) as 59 primers (1 pmol/ml for 59 decamersand 5 pmol/ml for 39 primers) as follows: 40 cycles of 30 s at94°C, 1 min at 40°C, 30 s at 72°C, and 5 min at 72°C forelongation at final concentrations of 1.5 mM MgCl2 and 20mMdNTPs mix in a Perkin–Elmer PCR system 2400 thermocycler.Radiolabeled amplified products were subjected to high-reso-lution polyacrylamide/urea gel electrophoresis as described(Liang and Pardee, 1992). Selected bands were excised fromthe gel, and DNA was eluted in buffer E (0.5M ammoniumacetate, 10 mM MgCl2, 1 mM EDTA, and 0.1% sodium dode-cyl sulfate) and precipitated. They were reamplified by standardPCR and subcloned into the pCRII T/A cloning vector. Bidi-rectional sequencing was done using an Applied BiosystemsAutomatic DNA Sequencer 373A by the fluorescentdideoxynucleotide terminator method according to Sanger et al.(1977). The sequence was confirmed further by isolating andcloning an independent cDNA from the same band correspond-ing to the cortex-derived RT-PCR RNA.
In situ hybridizationUnder profound pentobarbital anesthesia, normal and hypo-
thyroid rats of different ages were perfused through the heartwith cold 4% paraformaldehyde in 0.1M sodium phosphate(pH 7.4). The brains were removed quickly and cryoprotectedin 4% paraformaldehyde and 30% sucrose (wt/vol) in phos-phate-buffered saline (PBS) at 4°C. Subsequently, 25-mm coro-nal sections were cut using a cryostat. Nonradioactive in situhybridization on floating sections was performed as described(Gall and Isackson, 1989; Alvarez-Dolado et al., 1998). Thecomplete S2 cDNA clone and T7 and Sp6 RNA polymeraseswere used to obtain the antisense and sense riboprobes, respec-tively, using a digoxigenin RNA labeling kit (BoehringerMannheim). To develop the digoxigenin staining, we used thenucleic acid detection kit from Boehringer Mannheim. Sectionswere mounted onto slides, dehydrated by ethanol series (con-taining 0.3M ammonium acetate), coverslipped, and stored inthe dark at room temperature. For radioactive in situ hybrid-ization, 25-mm-thick coronal sections were thawed, washedwith PBS for 5 min, treated for an additional 10 min at roomtemperature under free-floating conditions with 0.1% TritonX-100, 0.2M hydrochloric acid, 0.25% acetic anhydride in 0.1M triethanolamine, and postfixed with 4% paraformaldehyde.
J. Neurochem., Vol. 73, No. 6, 1999
2241THYROID HORMONE REGULATES KIAA0719/Tom70 GENE

They were then preincubated in hybridization solution [0.6MNaCl, 20 mM piperazine-N,N9-bis(2-ethanesulfonic acid)(PIPES) sodium salt (pH 6.8), 10 mM EDTA, 50% formamide,0.2% sodium dodecyl sulfate, 53 Denhardt’s solution, 10%dextran sulfate, 50 mM dithiothreitol, 250 mg/ml shearedsalmon sperm DNA, and 250mg/ml yeast tRNA] for 3–5 h at55°C, and then incubated in the same solution containing the35S-UTP-labeled riboprobe (163 106 cpm/ml) overnight at55°C. Sections were washed consecutively once in 23 standardsaline citrate (SSC; 13 SSC5 0.15M NaCl, 0.015M sodiumcitrate) and 10 mM b-mercaptoethanol at room temperature for30 min, once in 53 TEN [50 mM Tris (pH 7.5), 5 mM EDTA,0.5 M NaCl] supplemented with 4mg/ml RNase at 37°C for1 h, twice in 0.53 SSC, 50% formamide, and 10 mM b-mer-captoethanol at 55°C for 1 h, once in 0.13 SSC and 10 mMb-mercaptoethanol at 68°C for 1 h, and finally twice in PBS atroom temperature for 5 min. Sections were mounted ontoslides, dehydrated by ethanol series (containing 0.3M ammo-nium acetate), and exposed for 1 week to Hyperfilmb-maxfilms (Amersham). For anatomical abbreviations, we followedthose used by Paxinos and Watson (1998).
RESULTS
To isolate new genes regulated by thyroid hormone inthe CNS, three independent differential display PCRreactions were performed using cDNA populations ret-rotranscribed from RNAs extracted from dissected brainregions (cortex, cerebellum, striatum, or hippocampus)of P15 control and hypothyroid rats. A set of NN(T)11oligonucleotides and random decamers were used as 39and 59 primers, respectively. PCR products were re-solved in polyacrylamide gels to search for differentiallyexpressed transcripts. One band of;300 bp was chosenfor further analysis based on its repetitive higher abun-dance in PCR reactions performed with striatal RNAfrom hypothyroid animals as compared with controls(Fig. 1A). To characterize this band, which was named
S2, the DNA was isolated from the gel, amplified, andsubcloned into the pCRII vector.
To confirm that the S2 cDNA clone corresponded to agene regulated by T3 in vivo, we used it as a molecularprobe to hybridize northern blots containing mRNA fromstriatum of control and hypothyroid P15 rats. Two RNAswere detected of;4.2 and 2.0 kb, which in agreementwith the differential display data were expressed athigher levels in hypothyroid than in control animals (Fig.1B, upper panel). To ascertain the thyroid status of theanimals and ensure equal loading of the gel, the filterswere rehybridized with probes for MAG, a gene down-regulated by hypothyroidism, and for the housekeepingCy gene (Fig. 1B, lower panel).
The S2 cDNA clone was sequenced and a homologysearch was performed by using BLASTP/nrdb95,BLASTN/nrnee, GeneQuiz, and Psi-Blast (Altschulet al., 1997; Andrade et al., 1999). This study clearlyshowed that the S2 clone was almost identical to afragment of the human KIAA0719 cDNA (Fig. 2A).KIAA0719 is a gene expressed in the brain and manyother tissues. Its full-length cDNA sequence has beendetermined recently in a study of new brain cDNAclones coding for large proteins in vitro (Nagase et al.,1998). KIAA0719 has been reported to be distantly ho-mologous (28.6% identity, 559 overlapping amino acidresidues) to theNeurospora crassamitochondrial pre-cursor protein import receptor (Nagase et al., 1998). Inline with this, we found 25–31% homology between S2and several members of the fungal Tom70 family of thetranslocase mitochondrial outer membrane complex (Fig.2B). This multisubunit complex is involved in the rec-ognition, unfolding, and translocation of preproteins intothe mitochondria (Neupert, 1997; Pfanner and Meijer,1997). KIAA0719 is the first characterized mammaliangene homologous to this family of Tom genes, which upto now only included fungal members. It is interestingthat homology affects several regions distributed throughthe whole sequence, and includes the transmembranedomain and a fragment corresponding to two tetratri-copeptide repeats (TPRs), a peptide sequence usuallyfound in the Tom family of receptors that is involved inprotein–protein interactions (Fig. 2B) (Das et al., 1998).
To confirm further the regulation of KIAA0719 geneby thyroid hormone, we next studied whether its expres-sion was affected by hormone treatment in vivo. Asshown in Fig. 3A, single injections of T4 during 4 dayswere able to restore the normal levels of the 4.2- and2.0-kb mRNAs in hypothyroid rats. In agreement with aninhibitory hormonal action on KIAA0719 expression, asimilar treatment with T4 of control animals led to areduction of the basal KIAA0719 RNA level (Fig. 3A).The effect of T4 treatment was also analyzed in thecerebellum, hippocampus, and cerebral cortex. Accord-ing to the PCR display data, KIAA0719 RNAs werefound to be expressed in these brain regions to verysimilar levels in both control and hypothyroid animals(Fig. 3B). Next, we analyzed whether the observed up-regulation of KIAA0719 RNA expression in the striatum
FIG. 1. Isolation of S2 as a thyroid hormone-regulated gene bydifferential display PCR. A: Differential display analysis of geneexpression in several brain regions of hypothyroid (H) and con-trol (C) rats. Cb, cerebellum; Hp, hippocampus; S, striatum; Cx,cortex. The S2 band is indicated. B: Northern blot analysis of S2expression in the striatum of control (C) and hypothyroid (H)15-day-old rats. The filter was rehybridized with the MAG and Cyprobes. Twenty micrograms of total RNA was loaded per lane.Sizes of the respective bands are indicated on the right.
J. Neurochem., Vol. 73, No. 6, 1999
2242 M. ALVAREZ-DOLADO ET AL.
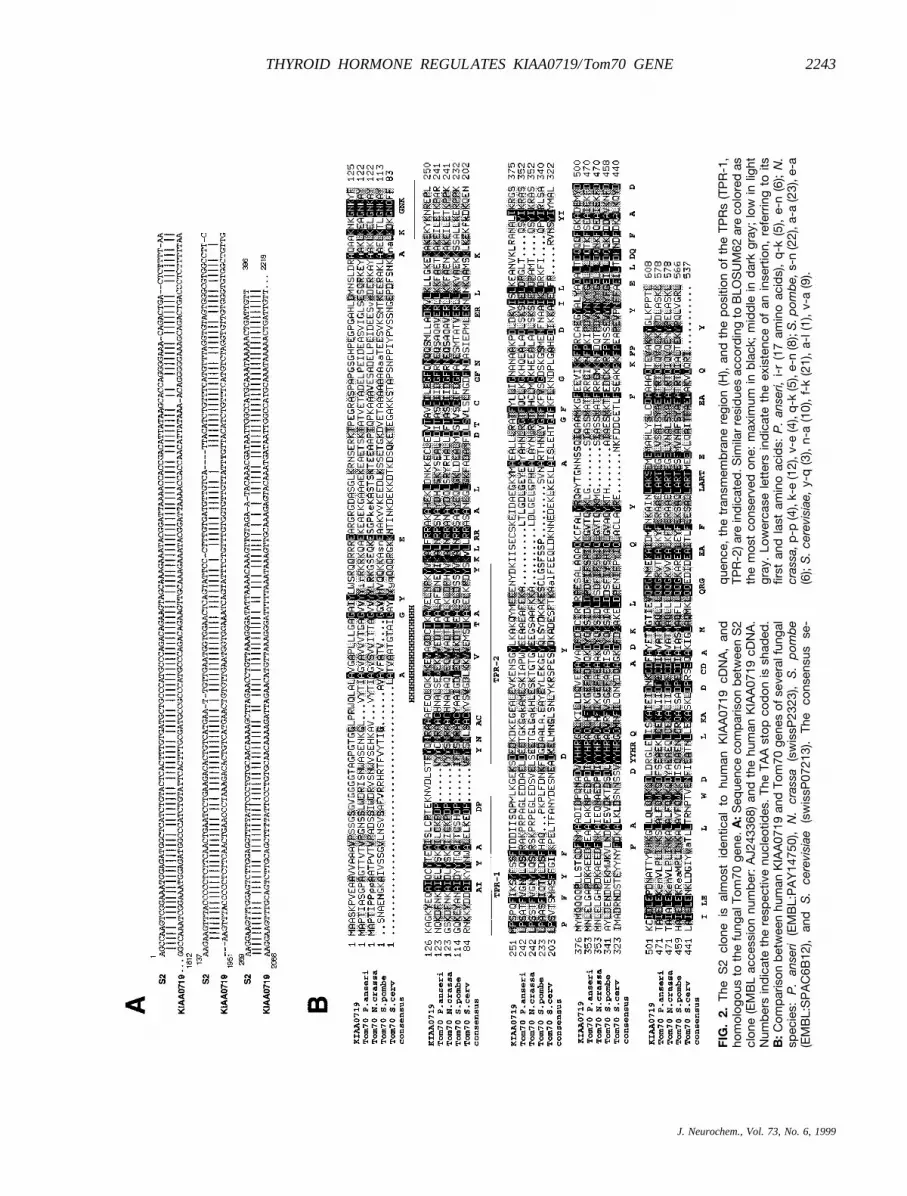
FIG
.2.
The
S2
clon
eis
alm
ost
iden
tical
tohu
man
KIA
A07
19cD
NA
,an
dho
mol
ogou
sto
the
fung
alTo
m70
gene
.A:S
eque
nce
com
par
ison
bet
wee
nS
2cl
one
(EM
BL
acce
ssio
nnu
mb
er:
AJ2
4336
8)an
dth
ehu
man
KIA
A07
19cD
NA
.N
umb
ers
ind
icat
eth
ere
spec
tive
nucl
eotid
es.
The
TAA
stop
cod
onis
shad
ed.
B:C
omp
aris
onb
etw
een
hum
anK
IAA
0719
and
Tom
70ge
nes
ofse
vera
lfun
gal
spec
ies:
P.
anse
ri(E
MB
L:P
AY
1475
0),
N.
cras
sa(s
wis
sP23
23),
S.
pom
be
(EM
BL:
SP
AC
6B12
),an
dS
.ce
revi
siae
(sw
issP
0721
3).
The
cons
ensu
sse
-
que
nce,
the
tran
smem
bra
nere
gion
(H),
and
the
pos
ition
ofth
eTP
Rs
(TP
R-1
,TP
R-2
)are
ind
icat
ed.S
imila
rre
sid
ues
acco
rdin
gto
BLO
SU
M62
are
colo
red
asth
em
ost
cons
erve
don
e:m
axim
umin
bla
ck;
mid
dle
ind
ark
gray
;lo
win
light
gray
.Lo
wer
case
lett
ers
ind
icat
eth
eex
iste
nce
ofan
inse
rtio
n,re
ferr
ing
toits
first
and
last
amin
oac
ids:
P.
anse
ri,i-
r(1
7am
ino
acid
s),
q-k
(5),
e-n
(6);
N.
cras
sa,p
-p(4
),k-
e(1
2),v
-e(4
),q
-k(5
),e-
n(8
);S
.pom
be,
s-n
(22)
,a-a
(23)
,e-a
(6);
S.
cere
visi
ae,
y-q
(3),
n-a
(10)
,f-
k(2
1),
a-l(
1),
v-a
(9).
J. Neurochem., Vol. 73, No. 6, 1999
2243THYROID HORMONE REGULATES KIAA0719/Tom70 GENE

of hypothyroid P15 rats occurs at other developmentalstages. As shown in Fig. 3C, this alteration was alsofound at P5. In contrast, no differences were found inadult animals (Fig. 3C). The results showed that theKIAA0719 gene is regulated in vivo by thyroid hormoneduring the early postnatal period in the striatum, but notin other brain regions.
Given the widespread expression of the KIAA0719gene in the organism (Nagase et al., 1998), we investi-gated whether thyroid hormone also regulates theKIAA0719 gene in tissues other than the brain. To thisend, we compared the levels of KIAA0719 RNA in theliver, kidney, heart, testis, lung, and eye of control andhypothyroid P15 rats. KIAA0719 expression in theseorgans was lower than in CNS. As shown in Fig. 4, nodifferences were detected in any nonneural tissue, aresult that reinforces the specificity of thyroid hormoneaction.
To analyze further the pattern of KIAA0719 expres-sion in the euthyroid and hypothyroid rat brain, weperformed in situ hybridization studies using first adigoxigenin-labeled S2 riboprobe. According to results
obtained by differential display PCR and northern blotassays, KIAA0719 RNA expression was detectedthroughout the brain of P15 animals (Figs. 5 and 6, anddata not shown). In euthyroid animals, the most intensesignal was found in the olfactory tubercle, particularly inlayer II, the medial forebrain bundle area surrounding theislands of Calleja, nucleus accumbens, globus pallidus,caudate putamen, and cortex (Fig. 5A, C, and E). Astrong signal was also found in the hippocampus (datanot shown). In the cortex, highest KIAA0719 expressionwas detected in pyramidal neurons of layer V and in thepiriform cortex (Fig. 5A and E). Lower expression wasfound in cortex layers II–IV and VI and was absent fromlayer I (Figs. 5A and 6A). Minor KIAA0719 expressionwas detected in thalamic and hypothalamic nuclei, andalso in Purkinje and granule cells of the cerebellum (datanot shown). Cell bodies were preferentially labeled, withan overall pattern that is compatible with a predominantexpression in neurons.
In line with the previous data, a clear increase inKIAA0719 gene expression was found in the striatum(caudate putamen and globus pallidus), piriform cortex,nucleus accumbens, and cortical layers II–IV and VI ofhypothyroid rats (Fig. 5B, D, and F). In the striatum, boththe number of labeled cell bodies and the intensity ofsignal were increased by hypothyroidism, whereas nohybridization was found in the fiber tracts (Fig. 5C andD). In the cortex, hypothyroidism led to a uniformly highKIAA0719 expression that caused the loss of the pre-dominant expression in layer V observed in euthyroidrats (compare Figs. 5A and 6A to Figs. 5B and 6B).Remarkably, in contrast, hypothyroidism caused a down-regulation of KIAA0719 expression in all layers of theolfactory tubercle, as well as in the medial forebrainbundle area (compare Fig. 5A and E with Fig. 5B and F).This result suggests that thyroid hormone action is mod-ulated by locally acting agent(s) or region-specific mol-ecule(s) in these areas. Hybridization with a sense S2riboprobe showed only background signal in the struc-tures studied (Fig. 5S). To confirm these data, we per-formed also in situ hybridization using a radioactive S2
FIG. 4. S2 RNA expression in nonneuronal tissues of P15 con-trol (open columns) and hypothyroid (filled columns) rats. Thelevel of S2 RNA expression (both bands) in different tissues wasquantified from a northern blot containing 20 mg of total RNA perline. Values were normalized to Cy RNA expression. E, eye; H,heart; K, kidney; L, liver; Lu, lung; T, testis; S, striatum.
FIG. 3. S2 is regulated in vivo by thyroid hormone in the stria-tum, but not in other brain regions. A: Northern blot (left) andquantification (right) of S2 expression in the striatum of P15control (C), T4-treated control (C 1 T4), hypothyroid (H), andT4-treated hypothyroid (H 1 T4) rats. Filters were rehybridizedwith probes for the MAG and Cy genes. Numbers indicate thesizes of the respective bands. Statistical significance: ** p, 0.01; *** p , 0.001. B: Northern blot analysis of S2 expressionin several brain areas of P15 control (C), T4-treated control (C1 T4), hypothyroid (H), and T4-treated hypothyroid (H 1 T4) rats.Cb, cerebellum; Hp, hippocampus; Cx, cortex. Twenty micro-grams of total RNA was loaded per lane. Numbers indicate thesizes of the two S2 RNAs. C: Northern analysis of S2 RNA in thestriatum of control (C) and hypothyroid (H) 5-day-old (P5) or adult(P60) rats.
J. Neurochem., Vol. 73, No. 6, 1999
2244 M. ALVAREZ-DOLADO ET AL.

riboprobe. By this method, we found the same differ-ences in the pattern of expression: up-regulation ofKIAA0719 RNA in the striatum and nucleus accumbens,down-regulation in the olfactory tubercle, and identicalalterations in the cerebral cortical layers of hypothyroidanimals (Fig. 6C–F). Only in the piriform cortex, thedifferences seem to be lost.
DISCUSSION
In this study, we describe the identification by differentialdisplay PCR of KIAA0719, a human homologue of thefungal Tom70, as a thyroid-regulated gene in the CNS. Bynorthern blotting, we found that hypothyroidism leads to atwo- to threefold increase in KIAA0719 RNA expression inthe striatum of P15 rats. This effect is region-specific,because no alterations were found in the cortex, cerebellum,or hippocampus. In addition, the fact that T4 treatmentrestores normal KIAA0719 expression in hypothyroid ani-mals and reduces also the physiological levels in control
rats strongly suggests a specific inhibitory role of the hor-mone in vivo. In situ hybridization studies confirmed anegative regulation of KIAA0719 expression by thyroidhormone in the striatum (caudate putamen, globus pallidus)and nucleus accumbens. Hypothyroid rats showed alsohigher KIAA0719 RNA expression in the subplate andcortical layers II–IV, but lower expression in the olfactorytubercle. These opposite changes probably compensated toproduce no differences in total cortical KIAA0719 RNAlevels when they were analyzed by northern blotting. Al-though both techniques, northern blotting and in situ hy-bridization, have limitations to measure RNA expression interms of distribution and quantification, respectively, andfurther studies will be required to address the complexquestion of site-specific regulation, our data strongly sug-gest that brain expression of KIAA0719 gene is underthyroid control in the rat.
The regulation of KIAA0719 gene expression appearsto be restricted to neural tissue during the early postnatal
FIG. 5. In situ hybridization analysis of S2expression in coronal sections of P15control (A, C, and E) and hypothyroid (B,D, and F) rats. A digoxigenin-labeled ribo-probe corresponding to full-length S2clone was used. A and B: Sections show-ing higher S2 RNA expression in severallayers of the cerebral cortex (Cx), caudateputamen (CPu), nucleus accumbens(Acb), and piriform cortex (Pir) of hypothy-roid (B) versus control (A) animals. In con-trast, lower expression was found in theolfactory tubercule (Tu) of hypothyroidrats. White arrows indicate S2 expressionin cortical layer V of control animals; ccindicates corpus callosum. C and D: Highmagnification showing higher number ofpositively stained cell bodies and alsostronger labeling in the striatum of hypo-thyroid (D) versus control (C) animals. Fi-ber tracts (ft) are not labeled. E and F:Detail of the local differences in S2 ex-pression in the piriform cortex (Pir), olfac-tory tubercle (Tu), and ventral pallidum(VP) between control (E) and hypothyroid(F) rats. S: Background signal was exam-ined by hybridizing control brain sectionswith a sense riboprobe. Scale bars: on A,1 mm for A and B; on C, 50 mm for C andD; on S, 400 mm for panels E, F, and S.
J. Neurochem., Vol. 73, No. 6, 1999
2245THYROID HORMONE REGULATES KIAA0719/Tom70 GENE

period, because no changes by hypothyroidism werefound in nonneural tissues or in brain at P30 and adultlife. Importantly, the first two postnatal weeks is theperiod of intense neuronal differentiation and maturationwhen the presence of thyroid hormone is required fornormal brain development in the rat (Schwartz and Op-penheimer, 1997; Bernal and Guadan˜o-Ferraz, 1998).With the exception of the neuronal RC3/neurogranin andTHRP/thyroid hormone-responsive protein genes, noother gene has been found to be regulated by thyroidhormone in the adult brain (In˜iguez et al., 1993; Shah etal., 1997). In line with these results, several examples ofcomplex regulation of gene expression by thyroid hor-mone have been described. Thus, distinct age- or region-dependent hormonal effects have been found for tenas-cin-C, RC3/neurogranin, type 2 iodothyronine deiodi-nase, or the brain lipocalin-type prostaglandin D2synthase (In˜iguez et al., 1993; Garcı´a-Ferna´ndez et al.,1997; Alvarez-Dolado et al., 1998; Guadan˜o-Ferraz etal., 1999). Together, these data indicate that thyroidhormone cooperates with local, transiently acting factors
to regulate the expression of multiple genes during braindevelopment in a precise and timely manner.
KIAA0719 shows greatest homology to the fungalTom70 gene, which codes for a component of Tom, amultisubunit machinery involved in the import of nucleus-encoded precursor proteins into the mitochondria, facil-itating their recognition, insertion, and translocation(Kaldi and Neupert, 1998; Kunkele et al., 1998).KIAA0719 is the first mammalian protein characterizedbelonging to the Tom family. Homology betweenKIAA0719 and Tom70 extends over the whole sequence,including the transmembrane domain and the TPR mo-tifs. These are domains consisting of a variable numberof degenerate tandem 34-amino acid repeats, each oneforming two amphipathic helices separated by a turnwhose hydrophobic faces function as targeting surfacesmediating protein–protein interactions (Lamb et al.,1995; Das et al., 1998). The degree of sequence homol-ogy found does not allow us to ascertain whether mam-malian KIAA0719 gene is orthologous to fungal Tom70,that is, whether their respective encoded proteins have
FIG. 6. Effect of hypothyroidism on S2 expres-sion in cerebral cortex and caudate putamen ofP15 rats. A and B: Digoxigenin in situ hybridiza-tion (carried out as in Fig. 5). Coronal brain sec-tions of control and hypothyroid rats show thealteration in the pattern of S2 expression, with auniformly stronger labeling and partial loss of thepredominant expression in layer V in hypothyroidrats. Cortical layers V and VIb or subplate (sp) areindicated. Note the higher S2 expression in thesubplate (sp) and caudate putamen (CPu) of hy-pothyroid animals. C–F: Radioactive in situ hy-bridization. Sections of control and hypothyroidanimals show the up-regulation of S2 expressionin the hypothyroid rat. Regions and layers arelabeled as in Fig. 5 and above. Scale bars: on A,50 mm for A and B; on C, 150 mm for panels Cand D; on E, 500 mm for E and F.
J. Neurochem., Vol. 73, No. 6, 1999
2246 M. ALVAREZ-DOLADO ET AL.

the same biological activity, or if alternativelyKIAA0719 is a member of the Tom family with relatedbut not identical function in mammals.
It is interesting that thyroid hormone has been reportedto play an important role in the regulation of mitochon-drial function in several tissues (Nelson, 1990). How-ever, brain mitochondria were considered for years asrefractory to thyroid hormone (Schwartz and Oppenhei-mer, 1978), and only recently brain mitochondrial alter-ations following thyroid manipulations were demon-strated (Battie and Verity, 1979; Katyare et al., 1994;Vega-Nunez et al., 1997). Several reports have describedthe regulation by thyroid hormone of the expression ofnucleus-encoded genes (Luciakova and Nelson, 1992;Izquierdo and Cuezva, 1993). In addition, the mitochon-drial genes coding for the NADH dehydrogenase subunit3, the cytochromec oxidase subunits I, II, and III, and the12S and 16S rRNAs have been reported to be induced bythyroid hormone (van Itallie, 1990; Iglesias et al., 1995;Vega-Nunez et al., 1995; Koibuchi et al., 1996). Sug-gesting a direct transcriptional regulation of some ofthese genes, TRs have been found inside the mitochon-dria (Ardail et al., 1993; Wrutnial et al., 1995), andbinding sites have been characterized in the mitochon-drial genome (Iglesias et al., 1995). Recently, thyroidhormone has been reported to modulate the mRNA/rRNA ratio by a direct regulation of the mitochondrialtranscription machinery (Enrı´quez et al., 1999). Ouridentification of a component of the complex involved inthe import into the mitochondria of precursor proteins asa thyroid hormone target gene shows an additional levelof control of mitochondrial function by the hormone.Moreover, it gives a substantial support to the descriptionby electron microscopy of structural and electricalchanges in the mitochondria from the cerebral cortex,striatum, and granule cells of the hippocampus (Vega-Nunez et al., 1997). Hypothyroidism causes alterations inphospholipid composition, membrane fluidity, and oxi-dative phosphorylation in rat brain mitochondria (Satavand Katyare, 1982; Bangur et al., 1995). Our data indi-cate that the effect of thyroid hormone on the phenotypeof brain mitochondria is mediated at least partiallythrough the regulation of the expression of theKIAA0719/Tom70 gene and, consequently, the controlof import of mitochondrial components. In line with ourdata, Craig et al. (1998) have found recently that thyroidhormone increases mitochondrial protein import withoutaffecting degradation in cardiac muscle cells. These au-thors described an increase by the hormone of the importof the matrix-localized precursor proteins malate dehy-drogenase and ornithine carbamoyltransferase, whereasthe outer membrane protein Bcl-2 was unaffected. Thiseffect is coincident with an elevation by thyroid hormoneof the content of outer membrane receptor Tom20 andthe matrix heat-shock protein mthsp70 in cardiac musclecells.
Transmembrane potential is essential for an ade-quate mitochondrial function, and it is likely thatalterations in the mitochondrial transport machinery
will affect this parameter. Intriguingly, a decrease inmitochondrial transmembrane potential has been de-scribed in hyperthyroid rat liver mitochondria but, incontrast, a similar decrease was reported in the hypo-thyroid rat brain (Gregory and Berry, 1991; Vega-Nunez et al., 1997). Thus, brain and liver mitochon-dria appear to respond distinctly to changes in thethyroid status, indicating tissue-specific effects of thy-roid hormone on this organule. These data are inagreement with our results showing a specific regula-tion of KIAA0719/Tom70 by thyroid hormone in stri-atum and other brain regions, whereas other tissues arenot affected. This specific regulation of KIAA0719gene may be due to tissue- and region-specific differ-ences in mitochondrial composition and/or regulation,or more probably are the result of a complex regula-tion by thyroid hormone and other unidentified factors.
In summary, our data indicate that the expression ofthe KIAA0719 gene is under thyroid control in specificareas of the brain during the early postnatal period.Further studies will establish the role of this effect on theprofound alterations caused by hypothyroidism on mito-chondrial morphology and physiology during CNS de-velopment.
REFERENCES
Altschul S., Madden T., Schaffer A., Zhang J., Zhang Z., Miller W.,and Lipman D. (1997) Gapped BLAST and PSI-BLAST: a newgeneration of protein database search programs.Nucleic AcidsRes.25, 3389–3402.
Alvarez-Dolado M., Iglesias T., Rodrı´guez-Pen˜a A., Bernal J., andMunoz A. (1994) Expression of neurotrophins and the trk familyof neurotrophin receptors in normal and hypothyroid rat brain.Mol. Brain Res.27, 249–257.
Alvarez-Dolado M., Gonza´lez-Sancho J. M., Bernal J., and Mun˜oz A.(1998) Developmental expression of tenascin-C is altered byhypothyroidism in the rat brain.Neuroscience84, 309–322.
Andrade M. A., Brown N., Leroy C., Hoersch S., de Daruvar A., ReichC., Franchini A., Tamames J., Valencia A., Ouzouzonis C., andSander C. (1999) Automated genome sequence analysis.Bioin-formatics15, 391–412.
Ardail D., Lerme F., Puymirat J., and Morel G. (1993) Evidence for thepresence ofa- andb-related T3 receptors in rat liver mitochon-dria. Eur. J. Cell Biol.62, 105–113.
Bangur C., Howland J., and Katyare S. (1995) Thyroid hormonetreatment alters phospholipid composition and membrane fluidityof rat brain mitochondria.Biochem. J.305,29–32.
Battie C. and Verity M. (1979) Membrane enzyme development innerve ending mitochondria during neonatal hypothyroidism.Dev.Neurosci.2, 139–148.
Bauer D., Muller H., Reich J., Riedel H., Warthoe P., and Strauss M.(1993) Identification of differentially expressed mRNA species byan improved display technique (DDRT-PCR).Nucleic Acids Res.21, 4272–4280.
Bernal J. and Guadan˜o-Ferraz A. (1998) Thyroid hormone and thedevelopment of the brain.Curr. Opin. Endocrinol. Diabetes5,296–302.
Bernal J. and Nunez J. (1995) Thyroid hormones and brain develop-ment.Eur. J. Endocrinol.133,390–398.
Bradley D. J., Towle H. C., and Young W. S. III (1992) Spatial andtemporal expression ofa and b thyroid hormone receptor mR-NAs, including b2-subtype, in the developing mammalian ner-vous system.J. Neurosci.12, 2288–2302.
J. Neurochem., Vol. 73, No. 6, 1999
2247THYROID HORMONE REGULATES KIAA0719/Tom70 GENE

Chomczynski P. and Sacchi N. (1987) Single step method of RNAisolation by acid guanidinium thiocyanate–phenol–chloroform ex-taction.Anal. Biochem.162,156–159.
Craig E., Chesley A., and Hood D. (1998) Thyroid hormone modifiesmitochondrial phenotype by increasing protein import withoutaltering degradation.Am. J. Physiol.275,1508–1515.
Dalal S. S., Welsh J., Tkachenko A., Ralph D., DiCicco-Bloom E.,Bordas L., McClelland M., and Chada K. (1994) Rapid isolationof tissue-specific and developmentally regulated brain cDNAsusing RNA arbitrarily primed PCR (RAP-PCR).J. Mol. Neurosci.5, 93–104.
Das A., Cohen P., and Barford D. (1998) The structure of thetetratricopeptide repeats of protein phosphatase 5: implicationsfor TPR-mediated protein–protein interactions.EMBO J. 17,1192–1199.
Dickson P., Aldred A., Menting J., Marley P., Sawyer W., and Schre-iber G. (1987) Thyroxine transport in choroid plexus.J. Biol.Chem.262,13907–13915.
Douglass J., McKinzie A., and Couceyro P. (1995) PCR differentialdisplay identifies a rat brain mRNA that is transcriptionallyregulated by cocaine and amphetamine.J. Neurosci.15, 2471–2481.
Enrıquez J. A., Ferna´ndez-Silva P., Garrido-Pe´rez N., Lopez-Pe´rezM. J., Perez-Martos A., and Montoya J. (1999) Direct regulationof mitochondrial RNA synthesis by thyroid hormone.Mol. Cell.Biol. 19, 657–670.
Feinberg P. and Vogelstein B. (1983) A technique for radiolabelingDNA restriction endonuclease fragments to high specific activity.Anal. Biochem.137,266–267.
Gall C. M. and Isackson P. J. (1989) Limbic seizures increase neuronalproduction of messenger RNA for nerve growth factor.Science245,758–761.
Garcıa-Ferna´ndez L. F., Rausell E., Urade Y., Hayaishi O., Bernal J.,and Munoz A. (1997) Hypothyroidism alters the expression ofprostaglandin D2 synthase/b-trace in specific areas of the devel-oping rat brain.Eur. J. Neurosci.9, 1544–1560.
Gregory R. and Berry M. (1991) The administration of triiodothyronineto rats results in a lowering of the mitochondrial membranepotential in isolated hepatocytes.Biochim. Biophys. Acta1133,89–94.
Guadan˜o-Ferraz A., Esca´mez M. J., Rausell E., and Bernal J. (1999)Expression of type 2 iodothyronine deiodinase in hypothyroid ratbrain indicates an important role of thyroid hormone in the de-velopment of specific primary sensory systems.J. Neurosci.19,3430–3439.
Iglesias T., Caubı´n J., Zaballos A., Bernal J., and Mun˜oz A. (1995)Identification of the mitochondrial NADH dehydrogenase sub-unit 3 (ND3) as a thyroid hormone regulated gene by wholegenome PCR analysis.Biochem. Biophys. Res. Commun.210,995–1000.
Iniguez M. A., Rodrı´guez-Pen˜a A., Ibarrola N., Aguilera M., Mun˜ozA., and Bernal J. (1993) Thyroid hormone regulation of RC3, abrain-specific gene encoding a protein kinase C substrate.Endo-crinology 133,467–473.
Izquierdo J. M. and Cuezva J. (1993) Thyroid hormones promotetranscriptional activation of the nuclear gene coding for mitochon-drial b-F1-ATPase in rat liver.FEBS Lett.323,109–112.
Joseph R., Dou D., and Tsang W. (1994) Molecular cloning of novelmRNA (neuronatina) that is highly expressed in neonatal mam-malian brain.Biochem. Biophys. Res. Commun.201, 1227–1234.
Kaldi K. and Neupert W. (1998) Protein translocation into mitochon-dria. Biofactors8, 221–224.
Katyare S., Bangur C., and Hovland J. (1994) Is respiratory activity inthe brain mitochondria responsive to thyroid hormone action? Acritical reevaluation.Biochem. J.302,857–860.
Koibuchi N., Matsuzaki S., Ichimura K., Ohtake H., and Yamaoka S.(1996) Ontogenic changes in the expression of cytochromecoxidase subunit I gene in the cerebellar cortex of the perinatalhypothyroid rat.Endocrinology137,5096–5108.
Kunkele K., Heins S., Dembowski M., Nargang F., Benz R., ThieffryM., Walz J., Lill R., Nussberger S., and Neupert W. (1998) The
preprotein translocation channel of the outer membrane of mito-chondria.Cell 93, 1009–1019.
Lamb J., Tugendreich S., and Hieter P. (1995) Tetratricopeptide repeatinteractions: to TPR or not to TPR?Trends Biochem. Sci.20,257–259.
Legrand J. (1984) Effects of thyroid hormones on central nervoussystem, inNeurobehavioural Teratology(Yanai J., ed), pp. 331–363. Elsevier Science Publishers, Amsterdam.
Legrand J. (1986) Thyroid hormone metabolism effects on growth anddevelopment, inThyroid Hormone Metabolism(Henneman G.,ed), pp. 503–534. Marcel Dekker Inc., New York.
Liang P. and Pardee A. B. (1992) Differential display of eukaryoticmessenger RNA by means of the polymerase chain reaction.Science257,967–971.
Luciakova K. and Nelson D. (1992) Transcript levels for nuclear-encoded mammalian mitochondrial respiratory-chain componentsare regulated by thyroid in an uncoordinated fashion.Eur. J. Bio-chem.207,247–251.
McClelland M., Mathieu-Daude F., and Welsh J. (1995) RNA finger-printing and differential display using arbitrarily primed PCR.Trends Genet.11, 242–246.
Mellstrom B., Naranjo J. R., Santos A., Gonza´lez A. M., and Bernal J.(1991) Independent expression of thea and b c-erbA genes indeveloping rat brain.Mol. Endocrinol.5, 1339–1350.
Nagase T., Ishikawa K.-I., Suyama M., Kikuno R., Miyajima N.,Tanaka A., Kotani H., Nomura N., and Ohara O. (1998) Predictionof the coding sequences of unidentified human genes. XI. Thecomplete sequences of 100 new cDNA clones from brain whichcode for large proteins in vitro.DNA Res.5, 277–286.
Nelson B. D. (1990) Thyroid hormone regulation of mitochondrialfunction. Comments on the mechanism of signal transduction.Biochim. Biophys. Acta1018,275–277.
Neupert W. (1997) Protein import into mitochondria.Annu. Rev. Bio-chem.66, 863–917.
Paxinos G. and Watson C. (1998)The Rat Brain In Stereotaxic Coor-dinates,4th edit. Academic Press, San Diego.
Pfanner N. and Meijer M. (1997) The Tom and Tim machine.Curr.Biol. 7, 100–103.
Porterfield S. and Hendrich C. (1993) The role of thyroid hormones inprenatal and neonatal neurological development. Current perspec-tives.Endocr. Rev.14, 94–106.
Rodrıguez-Pen˜a A., Ibarrola N., In˜iguez M. A., Munoz A., and BernalJ. (1993) Neonatal hypothyroidism affects the timely expressionof myelin-associated glycoprotein in the rat.J. Clin. Invest.91,812–817.
Sambrook J., Fritsch E. F., and Maniatis T. (1989)Molecular Cloning.A Laboratory Manual.Cold Spring Harbor Laboratory, ColdSpring Harbor, New York.
Sanger F., Nicklen S., and Coulson A. R. (1977) DNA sequencing withchain termination inhibitors.Proc. Natl. Acad. Sci. USA74,5463–5467.
Sap J., Mun˜oz A., Damm K., Goldberg Y., Ghysdael J., Leutz A., BeugH., and Vennstro¨m B. (1986) The c-erbA protein is a high affinityreceptor for thyroid hormone.Nature324,635–640.
Satav J. and Katyare S. (1982) Effect of experimental thyrotoxicosis onoxidative phosphorylation in rat liver, kidney and brain mitochon-dria. Mol. Cell. Endocrinol.28, 173–189.
Schwartz H. and Oppenheimer J. (1978) Ontogenesis of 3,5,39-triiodo-thyronine receptors in neonatal rat brain: dissociation betweenreceptor concentration and stimulation of oxygen consumption by3,5,39-triiodothyronine.Endocrinology103,943–948.
Schwartz H. and Oppenheimer J. (1997) Molecular basis of thyroidhormone dependent brain development.Endocr. Rev.18, 462–475.
Shah G., Li J., Schneiderjohn P., and Mooradian A. (1997) Cloning andcharacterization of a complementary DNA for a thyroid hormone-responsive protein in mature rat cerebral tissue.Biochem. J.327,617–623.
van Itallie C. (1990) Thyroid hormone and dexamethasone increase thelevels of a messenger ribonucleic acid for a mitochondriallyencoded subunit but not for a nuclear-encoded subunit of cyto-chromec oxidase.Endocrinology127,55–62.
J. Neurochem., Vol. 73, No. 6, 1999
2248 M. ALVAREZ-DOLADO ET AL.

Vega-Nunez E., Mene´ndez-Hurtado A., Garesse R., Santos A., andPerez-Castillo A. (1995) Thyroid hormone-regulated brain mito-chondrial genes revealed by differential cDNA cloning.J. Clin.Invest.96, 893–899.
Vega-Nunez E., Alvarez A., Mene´ndez-Hurtado A., Santos A., and Pe´rez-Castillo A. (1997) Neuronal mitochondrial morphology and trans-membrane potential are severely altered by hypothyroidism during ratbrain development.Endocrinology138,3771–3778.
Weinberger C., Thompson C. C., Ong E. S., Lebo R., Gruol D. J., and
Evans R. M. (1986) The c-erbA encodes a thyroid hormonereceptor.Nature324,641–646.
Welsh J., Chada K., Dalal S. S., Cheng R., Ralph D., and McClellandM. (1992) Arbitrarily primed PCR fingerprinting of RNA.NucleicAcids Res.20, 4965–4970.
Wrutnial C., Cassar-Malek I., Marchal S., Rascle A., Heusser S., Keller J.,Flechon J., Dauca M., Samarut J., Ghysdael J., and Cabello G. (1995)A 43-kDa protein related to c-erbAa1 is located in the mitochondrialmatrix of rat liver.J. Biol. Chem.270,16347–16354.
J. Neurochem., Vol. 73, No. 6, 1999
2249THYROID HORMONE REGULATES KIAA0719/Tom70 GENE




















