
Identification, isolation and characterization of new impurity in rabeprazole sodium
7
JOURNAL OF BACTERIOLOGY, 0021-9193/97/$04.0010 Oct. 1997, p. 6441–6447 Vol. 179, No. 20 Copyright © 1997, American Society for Microbiology Identification, Isolation, and Characterization of the 42-Kilodalton Major Outer Membrane Protein (MompA) from Treponema pectinovorum ATCC 33768 STEPHEN G. WALKER, 1 J. L. EBERSOLE, 2 AND S. C. HOLT 1 * Department of Microbiology 1 and Periodontics, 2 University of Texas Health Science Center at San Antonio, San Antonio, Texas 78284 Received 22 May 1997/Accepted 12 August 1997 The major protein present in the isolated outer membrane of Treponema pectinovorum ATCC 33768, MompA, was identified, purified, and characterized. Immuno-gold electron microscopy, using anti-MompA serum, and cell fractionation experiments confirmed the localization of MompA to the outer membrane. MompA was determined by sodium dodecyl sulfate-polyacrylamide gel electrophoresis to have a molecular mass of 42 kDa when heat denatured, whereas native MompA formed a number of detergent-stable forms with molecular masses of 71, 76, and 83 kDa. A temperature of 60°C was required to convert the native protein to the 42-kDa form. A number of detergents and chemical agents that are capable of breaking ionic and hydrogen bonds of proteins did not convert native MompA to the 42-kDa species. The native forms of the protein were resistant to the combined action of proteinase K, trypsin, and chymotrypsin, whereas the 42-kDa form of MompA was not. The N-terminal amino acid sequence of MompA was determined to be DVTVNINSRVRPVLYTT, and database searches did not identify any homology with known protein sequences. Amino acid compositional analysis showed the protein to be rich in proline and glycine, with these amino acids accounting for 28 and 13%, respectively, of the total amino acids. Antiserum raised against the major outer membrane protein of T. denticola GM-1 and ATCC 35405 did not cross-react with MompA, and antiserum raised against MompA did not react with any cellular components of Treponema denticola, Treponema vincentii, or Treponema socranskii.A major outer membrane protein similar in molecular mass to MompA was identified in eight clinical isolates of T. pectinovorum. The major outer membrane protein produced by four of the clinical isolates reacted strongly, by Western blotting, with anti-MompA serum, whereas proteins of the other strains did not. The periodontal pocket represents a complex ecosystem which undergoes a series of cyclic alterations in its microbial ecology. During health, the number of anaerobic bacteria is low, with gram-positive cocci and rods predominating. During disease progression, there is a dramatic increase in the gram- negative bacterial load such that they come to represent the major portion of the resident microbiota. Oral treponemes are also present in small numbers during periodontal health and increase to levels .50% of the total bacterial population dur- ing severe periodontal disease (23) and have been implicated as important members of the periodontopathic microbiota (2, 16, 22). Over 30 oral treponeme species have been identified in the periodontal pocket by using molecular DNA probes; how- ever, most remain uncultivable (6, 41). To date, there are five named (36–38, 49) and three unnamed (31) species of cultiva- ble oral treponemes. Of these, Treponema denticola has been the most intensely studied, and a number of potential virulence factors have been identified for this microorganism (1, 7, 13, 17, 32, 33, 42, 47, 50). One of these putative virulence factors is the major outer membrane protein. This protein has been termed the major surface antigen (9, 13), major cell envelope protein antigen (19), major surface protein (11), and major outer sheath protein (10, 48). Due to confusion generated by the use of numerous names for this protein and the name for the membrane in which it resides, we will use the terms major surface protein (Msp) and outer membrane (OM). The Msps of T. denticola strains are heterogeneous with respect to molecular mass, antigenicity, and primary DNA sequence. Strains of T. denticola produce a Msp of either approximately 64 or 58 kDa (10, 48). The 58-kDa Msp has been estimated in some publications to have a molecular mass of 53 kDa (9, 13, 19), and although DNA sequence analysis confirmed the protein to be 58 kDa (10), the 53-kDa estima- tion still persists in the literature (11). Antibody raised against the 64-kDa Msp of T. denticola GM-1 did not recognize the 58-kDa Msp produced by T. denticola ATCC 35404 (48), while antibody raised against the 58-kDa Msp of T. denticola ATCC 35405 did not recognize the 68-kDa Msp produced by T. den- ticola OTK, weakly recognized the 58-kDa Msp of T. denticola ATCC 33520, and strongly recognized the Msp produced by T. denticola ATCC 35404 (11). DNA sequence analysis by Fenno et al. (11) determined that the genes encoding the Msps of T. denticola ATCC 33520 and T. denticola ATCC OTK had 94.6 and 50.6% identity to that of T. denticola 35405. Despite the immunological and genetic heterogeneity ob- served in the Msps of T. denticola strains, these proteins have been noted to share many structural and functional character- istics. Upon extraction from the OM, the Msps form a deter- gent-stable, proteinase-resistant, high-molecular-mass species that is converted to a lower-molecular-mass proteinase-sensi- tive species upon heat treatment (13, 48). The Msps appear to have multiple functions that may contribute to pathogenicity, including treponeme-host cell interactions (26, 47), binding to host cell surface proteins (13), and acting as a porin-like pro- tein (9, 46). Studies by Moore et al. (27, 28) have demonstrated the association of Treponema pectinovorum with periodontal dis- * Corresponding author. Mailing address: Department of Microbi- ology, The University of Texas Health Science Center at San Antonio, 7703 Floyd Curl Dr., San Antonio, TX 78284-7758. Phone: (210) 567- 3594. Fax: (210) 567-6612. E-mail: [email protected]. 6441
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Identification, isolation and characterization of new impurity in rabeprazole sodium

JOURNAL OF BACTERIOLOGY,0021-9193/97/$04.0010
Oct. 1997, p. 6441–6447 Vol. 179, No. 20
Copyright © 1997, American Society for Microbiology
Identification, Isolation, and Characterization of the42-Kilodalton Major Outer Membrane Protein (MompA)
from Treponema pectinovorum ATCC 33768STEPHEN G. WALKER,1 J. L. EBERSOLE,2 AND S. C. HOLT1*
Department of Microbiology1 and Periodontics,2 University of Texas Health Science Center at San Antonio,San Antonio, Texas 78284
Received 22 May 1997/Accepted 12 August 1997
The major protein present in the isolated outer membrane of Treponema pectinovorum ATCC 33768, MompA,was identified, purified, and characterized. Immuno-gold electron microscopy, using anti-MompA serum, andcell fractionation experiments confirmed the localization of MompA to the outer membrane. MompA wasdetermined by sodium dodecyl sulfate-polyacrylamide gel electrophoresis to have a molecular mass of 42 kDawhen heat denatured, whereas native MompA formed a number of detergent-stable forms with molecularmasses of 71, 76, and 83 kDa. A temperature of 60°C was required to convert the native protein to the 42-kDaform. A number of detergents and chemical agents that are capable of breaking ionic and hydrogen bonds ofproteins did not convert native MompA to the 42-kDa species. The native forms of the protein were resistantto the combined action of proteinase K, trypsin, and chymotrypsin, whereas the 42-kDa form of MompA wasnot. The N-terminal amino acid sequence of MompA was determined to be DVTVNINSRVRPVLYTT, anddatabase searches did not identify any homology with known protein sequences. Amino acid compositionalanalysis showed the protein to be rich in proline and glycine, with these amino acids accounting for 28 and 13%,respectively, of the total amino acids. Antiserum raised against the major outer membrane protein of T.denticola GM-1 and ATCC 35405 did not cross-react with MompA, and antiserum raised against MompA didnot react with any cellular components of Treponema denticola, Treponema vincentii, or Treponema socranskii. Amajor outer membrane protein similar in molecular mass to MompA was identified in eight clinical isolatesof T. pectinovorum. The major outer membrane protein produced by four of the clinical isolates reactedstrongly, by Western blotting, with anti-MompA serum, whereas proteins of the other strains did not.
The periodontal pocket represents a complex ecosystemwhich undergoes a series of cyclic alterations in its microbialecology. During health, the number of anaerobic bacteria islow, with gram-positive cocci and rods predominating. Duringdisease progression, there is a dramatic increase in the gram-negative bacterial load such that they come to represent themajor portion of the resident microbiota. Oral treponemes arealso present in small numbers during periodontal health andincrease to levels .50% of the total bacterial population dur-ing severe periodontal disease (23) and have been implicatedas important members of the periodontopathic microbiota (2,16, 22). Over 30 oral treponeme species have been identified inthe periodontal pocket by using molecular DNA probes; how-ever, most remain uncultivable (6, 41). To date, there are fivenamed (36–38, 49) and three unnamed (31) species of cultiva-ble oral treponemes. Of these, Treponema denticola has beenthe most intensely studied, and a number of potential virulencefactors have been identified for this microorganism (1, 7, 13,17, 32, 33, 42, 47, 50). One of these putative virulence factorsis the major outer membrane protein. This protein has beentermed the major surface antigen (9, 13), major cell envelopeprotein antigen (19), major surface protein (11), and majorouter sheath protein (10, 48). Due to confusion generated bythe use of numerous names for this protein and the name forthe membrane in which it resides, we will use the terms majorsurface protein (Msp) and outer membrane (OM).
The Msps of T. denticola strains are heterogeneous withrespect to molecular mass, antigenicity, and primary DNAsequence. Strains of T. denticola produce a Msp of eitherapproximately 64 or 58 kDa (10, 48). The 58-kDa Msp hasbeen estimated in some publications to have a molecular massof 53 kDa (9, 13, 19), and although DNA sequence analysisconfirmed the protein to be 58 kDa (10), the 53-kDa estima-tion still persists in the literature (11). Antibody raised againstthe 64-kDa Msp of T. denticola GM-1 did not recognize the58-kDa Msp produced by T. denticola ATCC 35404 (48), whileantibody raised against the 58-kDa Msp of T. denticola ATCC35405 did not recognize the 68-kDa Msp produced by T. den-ticola OTK, weakly recognized the 58-kDa Msp of T. denticolaATCC 33520, and strongly recognized the Msp produced by T.denticola ATCC 35404 (11). DNA sequence analysis by Fennoet al. (11) determined that the genes encoding the Msps of T.denticola ATCC 33520 and T. denticola ATCC OTK had 94.6and 50.6% identity to that of T. denticola 35405.
Despite the immunological and genetic heterogeneity ob-served in the Msps of T. denticola strains, these proteins havebeen noted to share many structural and functional character-istics. Upon extraction from the OM, the Msps form a deter-gent-stable, proteinase-resistant, high-molecular-mass speciesthat is converted to a lower-molecular-mass proteinase-sensi-tive species upon heat treatment (13, 48). The Msps appear tohave multiple functions that may contribute to pathogenicity,including treponeme-host cell interactions (26, 47), binding tohost cell surface proteins (13), and acting as a porin-like pro-tein (9, 46).
Studies by Moore et al. (27, 28) have demonstrated theassociation of Treponema pectinovorum with periodontal dis-
* Corresponding author. Mailing address: Department of Microbi-ology, The University of Texas Health Science Center at San Antonio,7703 Floyd Curl Dr., San Antonio, TX 78284-7758. Phone: (210) 567-3594. Fax: (210) 567-6612. E-mail: [email protected].
6441

ease and gingivitis. T. pectinovorum was more frequently re-covered in disease than in health and was the only species ofcultivable oral treponemes which increased in frequency dur-ing active periodontitis (28). Ongoing studies in our laboratoryhave identified T. pectinovorum as the most common cultivableoral treponeme in a subgroup of human immunodeficiencyvirus (HIV)-infected patients with destructive periodontal dis-ease (44). The presence of T. pectinovorum as a significantmember of the subgingival microbiota of HIV-infected peri-odontitis subjects and its presence in lower numbers in HIV-seronegative subjects suggest that T. pectinovorum is a majorcontributor to the progression of destructive periodontal dis-ease in HIV-infected individuals. Our laboratory is currentlyanalyzing T. pectinovorum for the presence of putative viru-lence factors.
This report details the identification, purification, and initialcharacterization of the major OM protein, MompA, from T.pectinovorum ATCC 33768. MompA was found to form deter-gent-stable, proteinase-resistant high-molecular-mass formsthat could be converted by heating to a proteinase-sensitivelower-molecular-mass species which are also characteristics incommon with the Msps produced by T. denticola strains. How-ever, although MompA was localized to the OM, it did notappear to be exposed on the cell surface and was distinct fromthe Msp of T. denticola with respect to antigenicity, molecularweight, amino acid composition, and N-terminal amino acidsequence.
MATERIALS AND METHODS
Chemicals. Unless otherwise stated, all chemicals and reagents were from theSigma Chemical Company, St. Louis, Mo.
Bacterial strains and growth conditions. The following strains of T. pectino-vorum were used in this study: ATCC 33768 (37), P2, P3, P5, P8 (45), X1, S1,M1A, and M2A. X1 and S1 were isolated from dental plaque from HIV-infectedsubjects undergoing periodontal treatment at the University of California at SanFrancisco (Gary Armitage, Division of Periodontology, University of Californiaat San Francisco). M1A and M2A were isolated from the plaque of a nonhumanprimate, Macaca fascicularis, which was undergoing experimental ligature-in-duced periodontitis at the University of Texas Health Science Center at SanAntonio. T. pectinovorum ATCC 33768, P2, P3, P5, and P8 were obtained fromthe laboratory collection of E. Peter Greenberg (University of Iowa, Iowa City).All T. pectinovorum strains used in this study are considered to be isolates of T.pectinovorum ATCC 33768 as determined by 16S rRNA sequencing (31).
T. denticola GM-1 (35), ATCC 35405, ATCC 35404, and ATCC 33520 werefrom our laboratory collection. Treponema socranskii subsp. socranskii ATCC35536 and Treponema vincentii ATCC 35580 were obtained from the collectionof E. Peter Greenberg.
Oral treponemes were incubated in an anaerobic chamber (Coy LaboratoryProducts, Ann Arbor, Mich.) in an atmosphere of 85% N2–10% H2–5% CO2 at37°C, with the humidity maintained between 30 and 40%. T. pectinovorum strainswere routinely maintained by daily transfer in GM-1 broth (4) without glucose orrabbit serum but supplemented with 0.3% pectin. For isolation and purificationof MompA, the cells were grown in GM-1 broth containing 2% (vol/vol) heat-inactivated rabbit serum, 150 mM HEPES buffer, 0.6% (wt/vol) pectin, and0.75% (wt/vol) yeast extract (Becton Dickinson and Company, Cockeysville,Md.). Before autoclaving, the pH was adjusted to 7.5 with 10 N KOH. All otheroral treponeme species were cultured in a modification of NOS medium (5, 21).NOS was modified by increasing the concentrations of yeast extract, volatile fattyacid mix, and heat-inactivated rabbit serum to 0.75, 0.5, and 5%, respectively, andby the addition of 0.2% ammonium sulfate, 0.2% L-ascorbic acid, and 0.1%pyruvic acid. For cultivation of T. socranskii, the serum was omitted. This mod-ified NOS medium supported better growth than the standard formulation, asjudged by an increased final optical density and a reduction in the numbers ofround bodies present in late-exponential-phase cultures.
For long-term storage of strains, dimethyl sulfoxide (American Type CultureCollection, Rockville, Md.) was added to mid-logarithmic-phase cultures to afinal concentration of 5% (vol/vol), and the cultures were frozen at 270°C.
Cell fractionation. T. pectinovorum strains were grown to late exponentialphase (optical density at 660 nm of 0.15 to 0.4), depending on the strain andmedium used, and then cooled rapidly to 4°C in an ice-water bath. Unlessotherwise indicated, all manipulations were carried out at 4°C. The cells werewashed twice with phosphate-buffered saline (PBS; 137 mM NaCl, 2.5 mMKH2PO4, 7 mM K2HPO4 [pH 7.2]) by centrifugation (5,000 3 g for 10 min) andresuspended in 0.05 M Tris-HCl (pH 7.2) containing 0.02 M MgCl2, DNase I (2mg/ml), and 2 mM (each) of the protease inhibitors phenylmethylsulfonyl fluo-
ride and N-p-tosyl-L-lysine chloromethylketone. This cell suspension was used toprepare cell envelopes or OM. For preparation of cell envelopes, the cells werebroken by sonication and then unbroken cells were removed by centrifugation at5,000 3 g for 10 min. The membranes were pelleted by ultracentrifugation at25,000 3 g for 30 min and then washed once with 0.05 M Tris-HCl. The OM wasprepared by the method of Masuda and Kawata (25). Briefly, washed cells weresuspended in buffer containing MgCl2, DNase, and proteinase inhibitors andthen subjected to 40 cycles of freeze-thaw (liquid N2-water, 20°C). The samplewas then centrifuged at 5,000 3 g for 10 min to remove the cytoplasmic cylinders,and the supernatant, containing the OM, was recovered and pelleted by centrif-ugation at 25,000 3 g for 30 min. The OM was resuspended in Tris-HCl bufferand washed twice by ultracentrifugation and resuspension. Finally, contaminat-ing flagella were dissolved by suspending the OM in 0.1 M sodium acetate-HClbuffer (pH 3.0) for 2 h at room temperature, then the OM was pelleted bycentrifugation and resuspended in Tris-HCl buffer (pH 7.2).
Electrophoresis. Discontinuous sodium dodecyl sulfate (SDS)-polyacrylamidegel electrophoresis (PAGE) was performed as described by Laemmli (20). Fol-lowing electrophoresis, gels were stained with Coomassie brilliant blue R250(Bio-Rad Laboratories, Hercules, Calif.) or by silver staining (Bio-Rad). Proteinloading was estimated by the method of Markwell et al. (24). Bio-Rad low-range-molecular-weight standards were used to estimate the molecular masses of trepo-neme proteins. Preparative electrophoresis using a model 491 Prep Cell (Bio-Rad) was performed as described by the manufacturer.
Western blotting. Following SDS-PAGE, gels were transferred to Immo-bilon-P membranes (Millipore Corporation, Bedford, Mass.), using a bufferconsisting of 10 mM 3-(cyclohexylamino)-1-propanesulfonic acid (CAPS), 0.05%DL-dithiothreitol, and 1% methanol (pH adjusted to 10.5 with 10 N NaOH) (29).The polyacrylamide gel was not equilibrated in the transfer buffer prior toelectroblotting. After transfer, the polyacrylamide gel and a duplicate membranewere stained with Coomassie brilliant blue R250 to assess the efficiency ofprotein transfer to the membrane. The membrane, used for reaction with theantibody, was blocked with 1% bovine serum albumin dissolved in 100 mMTris-HCl (pH 7.5)–0.9% NaCl–0.1% polyoxyethylene sorbitan monolaurate(TTBS). Primary antibody and goat anti-rabbit immunoglobulin G (IgG) conju-gated to alkaline phosphatase secondary antibody were diluted in TTBS toconcentrations of 1/5,000 and 1/3,000 respectively. Rabbit antiserum to the re-combinant Msp of T. denticola ATCC 35405 was a gift from J. C. Fenno and B. C.McBride (10). Antiserum to whole cells of T. denticola ATCC 35404 and to theMsp of T. denticola GM-1 was a gift from A. Weinberg (University of Washing-ton, Seattle). The blots were developed with a 5-bromo-4-chloro-3-indolylphos-phate–nitroblue tetrazolium system as specified by the manufacturer. Bio-RadKaleidoscope prestained molecular weight standards were used to estimate themolecular masses of reactive proteins.
Antiserum to purified MompA. Purified MompA (see below) was mixed 50:50with Titer Max adjuvant (CytRx Corporation, Norcross, Ga.) to give a finalconcentration of 100 mg of protein in a final volume of 400 ml. A female NewZealand White rabbit was injected subcutaneously in both shoulders and quad-riceps with 100 ml per site. After 4 weeks, the rabbit was given a booster dose of25 mg of MompA, mixed 50:50 with the adjuvant in a total volume of 80 ml, bytwo intramuscular injections in each hind quadriceps. Antibody titers were de-termined by enzyme-linked immunosorbent assay (8).
Extraction of the MompA from OM with detergents and chemicals. Tenmicroliters of OM, containing 60 mg of protein, was incubated with 1% (wt/vol)detergent in 20 mM Tris-HCl (pH 8.0) in a final volume of 20 ml for 30 min at22°C. The detergents used were Zwittergent 3-14 (Calbiochem Corporation, LaJolla, Calif.), n-octyl-b-D-glucopyranoside, n-octylpolyoxyethylene (Bachem Cal-ifornia Inc., Torrance, Calif.), SDS (Bio-Rad), Triton X-100, Nonidet P-40,3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate (CHAPS), andN,N-bis-(3-D-gluconamidopropyl)-cholamide (BIG CHAPS; Calbiochem). Thesample was then pelleted in an Eppendorf microcentrifuge for 10 min at 14,000rpm to separate soluble and insoluble material. Both the soluble (20 ml ofsupernatant) and insoluble (pellet) fractions were mixed with 20 ml of SDS-PAGE loading buffer, heated at 100°C for 10 min, and analyzed by SDS-PAGE.
OM was extracted as described above, but the detergents were replaced witheither 0.1 M Na2CO3 (pH 11) (EM Science, Gibbstown, N.J.), 0.6 M NaCl (EMScience), 1.6 M urea (Bio-Rad), or 0.1 N NaOH (EM Science). Followingincubation at 37°C, 30 min, soluble and insoluble material were separated bycentrifugation and analyzed by SDS-PAGE.
Chemical treatment of Zwittergent 3-14-extracted MompA. The effects ofselected chemicals on the integrity of native MompA were studied. Twelvemicrograms of Zwittergent 3-14-soluble OM, consisting of 95% MompA, wassuspended in Tris-HCl (pH 8.0) containing one of the chemical agents in a finalvolume of 15 ml. The chemicals, at their final concentrations in the assay buffer,were 1% 2-mercaptoethanol, 8 M urea, 25 mM EDTA, 25 mM EGTA, 100 mMNaCl, 25 mM MgCl2, and 25 mM CaCl2. After incubation for 30 min at roomtemperature, SDS-PAGE loading buffer was added and samples were analyzedby SDS-PAGE without heating.
Heat modification of MompA. Samples of OM (20 mg of protein) or Zwitter-gent 3-14-soluble OM fraction (12 mg) were suspended in 17 ml of SDS-PAGEloading buffer, and a sample was incubated at room temperature and 40, 50, 60,70, 80, and 100°C for 5 min and then allowed to cool to room temperature.Samples were then analyzed by SDS-PAGE.
6442 WALKER ET AL. J. BACTERIOL.

Proteinase treatment of Zwittergent 3-14-extracted MompA. Four samples of12 mg of Zwittergent 3-14-soluble OM was suspended in Tris-HCl (pH 8.0) to afinal volume of 10 ml. Two of the samples were heated at 100°C for 5 min andthen allowed to cool to room temperature. Five microliters of a proteinasecocktail was added to one of the heat-treated and one of the nonheated samples,and 5 ml of Tris-HCl (pH 8.0) was added to the remaining two samples. Theaddition of the proteinase cocktail resulted in a 50-mg/ml final concentration(each) for a-chymotrypsin, b-chymotrypsin, proteinase K, and trypsin. Sampleswere incubated at 37°C for 30 min, cooled to room temperature, mixed withSDS-PAGE loading buffer, and analyzed by SDS-PAGE without boiling prior toelectrophoresis.
Identification of heat-modifiable OM proteins. Heat-modifiable OM proteinswere identified by the two-dimensional SDS-PAGE method of Hindahl andIglewski (14).
Purification of MompA. OM was extracted with 1% (wt/vol) Zwittergent 3-14,and the solubilized native high-molecular-mass species of MompA was purifiedfrom low-molecular-mass contaminants by preparative SDS-PAGE on a 12%gel. The recovered native MompA was concentrated in a Centricon 30 microcon-centrator (Amicon, Beverly, Mass.), mixed with SDS-PAGE loading buffer, andheated to 100°C for 10 min to convert the native form to the lower-molecular-mass heat-modified form. The heated sample was subjected to a second round ofpreparative SDS-PAGE on a 7.5% gel, and pure MompA was recovered andconcentrated as described above.
Amino acid analysis and N-terminal protein sequencing of MompA. PurifiedMompA was electrophoresed and blotted onto an Immobilon-P membrane (Mil-lipore) as described above except that piperazine diacrylamide (Bio-Rad) wassubstituted for N,N9-methylenebisacrylamide in the polyacrylamide gel. The pro-tein was visualized by staining with Ponceau S and excised from the membrane.Internal sequence was obtained by Lys-C (WAKO, Richmond, Va.) proteinasetreatment of gel-purified MompA, and the peptides were isolated by high-pressure liquid chromatography (HPLC). N-terminal amino acid sequencing anddetermination of amino acid composition were done by standard methods at theUniversity of Texas Health Science Center Institutional Protein Facility as de-scribed previously (48).
Electron microscopy. Whole cells and OM of T. pectinovorum ATCC 33768were examined by negative-stain electron microscopy, as described previously(43), for the presence of a regular surface structure as has been observed in T.denticola strains (9). Whole cells and OM from T. denticola GM-1 were used aspositive controls. Immunoelectron microscopy of whole mounts and thin-sec-tioned T. pectinovorum ATCC 33768 was conducted as described by Weinbergand Holt (48). The IgG fraction of the preimmune and monospecific polyclonalrabbit serum raised against MompA (see above) was used at a dilution of 1:100.Anti-rabbit IgG–gold complex (5-nm gold; E-Y Laboratories, Inc., San Mateo,Calif.) was used as the secondary antibody. Samples were viewed in a JEOL 1200transmission electron microscope operating at 60 kV.
RESULTSIdentification and purification of MompA. Figure 1 shows
that T. pectinovorum ATCC 36768 produces a major heat-modifiable protein of 42 kDa (compare lanes 2 and 3). Thisprotein was segregated to the membrane fraction of sonicatedcells (compare lanes 4 and 5) and represents the major proteindetected in the OM (lanes 6 and 7). The 42-kDa major OM
protein of T. pectinovorum ATCC 33768 was designatedMompA. Analysis of the OM by two-dimensional SDS-PAGEalso identified the presence of two minor heat-modifiable pro-teins with estimated molecular masses of 31 and 98 kDa (datanot shown). MompA was solubilized from the OM with Zwit-tergent 3-14 (lane 8 and 9), and SDS-PAGE of the detergent-soluble MompA, using a 7.5% gel, clearly resolved the nativeforms of the protein into three prominent bands with esti-mated molecular masses of 71, 76, and 83 kDa (data notshown).
The heat-modifiable nature of MompA was exploited topurify it from the Zwittergent 3-14-soluble OM fraction. Theunheated Zwittergent 3-14-soluble OM fraction was subjectedto preparative SDS-PAGE to recover the native form ofMompA. This procedure removed contaminating proteins withthe same electrophoretic mobility as the heat-modifiedMompA. The isolated native forms of MompA were thensolubilized at 100°C and subjected to a second round of pre-parative SDS-PAGE, and the heat-modifiable form of MompAwas recovered. This second preparative SDS-PAGE purifica-tion step removed contaminating proteins with electrophoreticmobilities similar to those of the native forms of MompA. Thepurified heat-modified MompA is shown in lane 10 of Fig. 1.SDS-PAGE, followed by silver staining, of the purifiedMompA did not reveal the presence of contaminating proteinsor a lipopolysaccharide-like molecule that is also found in theOM of T. pectinovorum (data not shown).
Effects of select detergents, chemicals, and proteinases onMompA. One percent Nonidet P-40, n-octyl-b-D-glucopyrano-side, SDS, and n-octylpolyoxyethylene extracted the nativeform of MompA from the OM with the same efficiency asZwittergent 3-14, while treatment with CHAPS, BIG CHAPS,or Triton X-100 did not solubilize MompA. The SDS-PAGEbanding patterns of MompA extracted with the different de-tergents gave identical protein banding patterns (data notshown). The OM was also exposed to 0.1% Na2CO3 (pH 11),1.6 M urea, 0.6% NaCl, and 0.1 N NaOH to determine if theywere capable of removing MompA. With the exception of theNaOH, which solubilized the OM, none of these chemicalsremoved MompA from the OM (data not shown). The Zwit-tergent 3-14-soluble native MompA was exposed to severalchemicals often used to cleave noncovalent protein bonds todetermine if any were capable of converting the native forms tothe 42-kDa form. We found that 8 M urea, 25 mM EDTA,EGTA, MgCl2, CaCl2 or 100 mM NaCl had no effect on thenative structure of MompA (data not shown).
The native forms of MompA were found to be resistant toseveral proteinases which were capable of digesting the 42-kDaspecies (Fig. 2). Untreated and heat-treated (100°C) Zwitter-gent 3-14-soluble MompA was incubated in the presence of aproteinase cocktail containing trypsin, chymotrypsin, and pro-teinase K and then analyzed by SDS-PAGE (Fig. 2, lanes 4 and5, respectively). The native forms of MompA were resistant tothe action of the proteinases, whereas the 42-kDa form wascompletely digested. Lanes 1 and 2 of Fig. 2 are controls thatdid not receive proteinase treatment.
Determining the transition temperature for the conversionof the native MompA to the 42-kDa form. Figure 3 demon-strates the conversion of the native form of MompA to the42-kDa form by incubation of the OM, in SDS-PAGE samplebuffer, at various temperatures for 5 min prior to analysis bySDS-PAGE. The native forms were completely converted tothe 42-kDa form by incubation at 60°C (lane 5), while only asmall amount of the protein was converted by incubation at50°C (lane 4), indicating that the transition temperature isbetween 50 and 60°C. Zwittergent 3-14-solubilized MompA
FIG. 1. SDS-PAGE illustrating the purification of MompA from T. pectino-vorum ATCC 33768. Lanes: 1, protein standards; 2 and 3, unheated and heatedwhole-cell lysates, respectively; 4, heated soluble fraction from sonicated wholecells; 5, heated whole membrane fraction from sonicated whole cells; 6 and 7,unheated and heated OM, respectively; 8 and 9, unheated and heated Zwitter-gent 3-14-soluble OM fraction, respectively; 10, purified MompA. O and Mdenote the running positions of native and heat-modified MompA, respectively.Molecular masses of the protein standards, in kilodaltons, are shown at the left.The 10% polyacrylamide gel was stained with Coomassie brilliant blue R250.Samples loaded in lanes 2 to 9 contained 10 mg of protein, and the sample loadedin lane 10 contained 4 mg of protein.
VOL. 179, 1997 T. PECTINOVORUM 42-kDa OUTER MEMBRANE PROTEIN 6443
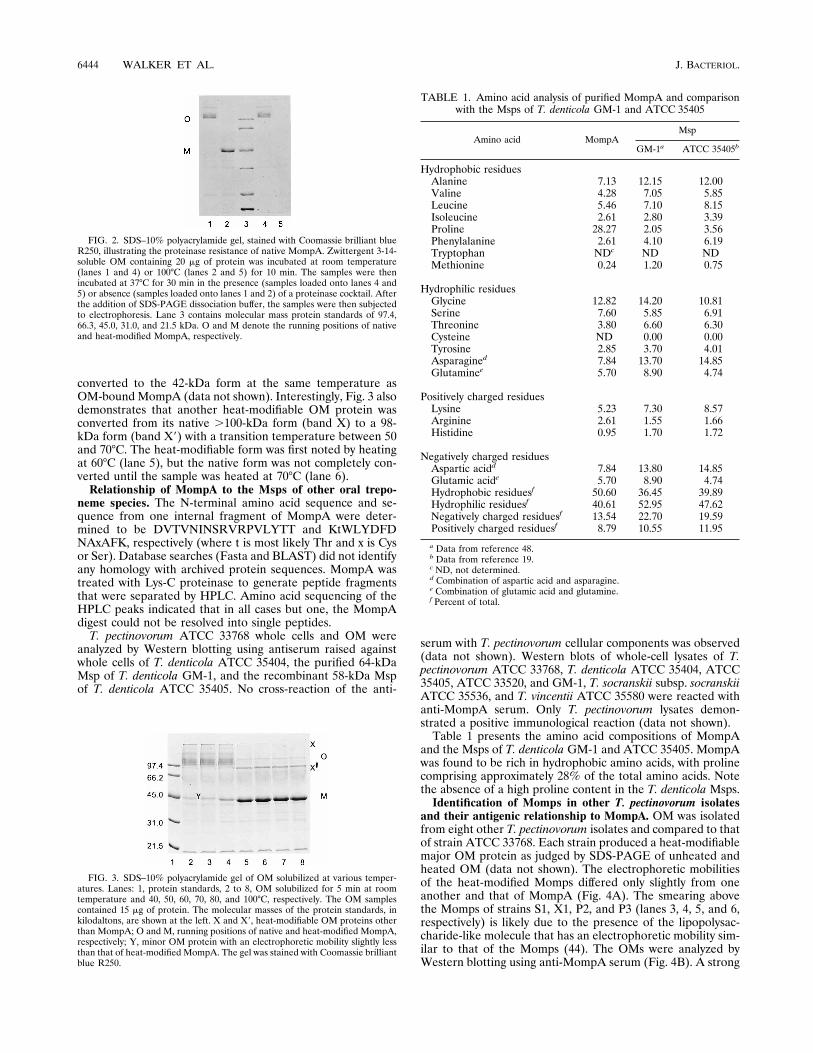
converted to the 42-kDa form at the same temperature asOM-bound MompA (data not shown). Interestingly, Fig. 3 alsodemonstrates that another heat-modifiable OM protein wasconverted from its native .100-kDa form (band X) to a 98-kDa form (band X9) with a transition temperature between 50and 70°C. The heat-modifiable form was first noted by heatingat 60°C (lane 5), but the native form was not completely con-verted until the sample was heated at 70°C (lane 6).
Relationship of MompA to the Msps of other oral trepo-neme species. The N-terminal amino acid sequence and se-quence from one internal fragment of MompA were deter-mined to be DVTVNINSRVRPVLYTT and KtWLYDFDNAxAFK, respectively (where t is most likely Thr and x is Cysor Ser). Database searches (Fasta and BLAST) did not identifyany homology with archived protein sequences. MompA wastreated with Lys-C proteinase to generate peptide fragmentsthat were separated by HPLC. Amino acid sequencing of theHPLC peaks indicated that in all cases but one, the MompAdigest could not be resolved into single peptides.
T. pectinovorum ATCC 33768 whole cells and OM wereanalyzed by Western blotting using antiserum raised againstwhole cells of T. denticola ATCC 35404, the purified 64-kDaMsp of T. denticola GM-1, and the recombinant 58-kDa Mspof T. denticola ATCC 35405. No cross-reaction of the anti-
serum with T. pectinovorum cellular components was observed(data not shown). Western blots of whole-cell lysates of T.pectinovorum ATCC 33768, T. denticola ATCC 35404, ATCC35405, ATCC 33520, and GM-1, T. socranskii subsp. socranskiiATCC 35536, and T. vincentii ATCC 35580 were reacted withanti-MompA serum. Only T. pectinovorum lysates demon-strated a positive immunological reaction (data not shown).
Table 1 presents the amino acid compositions of MompAand the Msps of T. denticola GM-1 and ATCC 35405. MompAwas found to be rich in hydrophobic amino acids, with prolinecomprising approximately 28% of the total amino acids. Notethe absence of a high proline content in the T. denticola Msps.
Identification of Momps in other T. pectinovorum isolatesand their antigenic relationship to MompA. OM was isolatedfrom eight other T. pectinovorum isolates and compared to thatof strain ATCC 33768. Each strain produced a heat-modifiablemajor OM protein as judged by SDS-PAGE of unheated andheated OM (data not shown). The electrophoretic mobilitiesof the heat-modified Momps differed only slightly from oneanother and that of MompA (Fig. 4A). The smearing abovethe Momps of strains S1, X1, P2, and P3 (lanes 3, 4, 5, and 6,respectively) is likely due to the presence of the lipopolysac-charide-like molecule that has an electrophoretic mobility sim-ilar to that of the Momps (44). The OMs were analyzed byWestern blotting using anti-MompA serum (Fig. 4B). A strong
FIG. 2. SDS–10% polyacrylamide gel, stained with Coomassie brilliant blueR250, illustrating the proteinase resistance of native MompA. Zwittergent 3-14-soluble OM containing 20 mg of protein was incubated at room temperature(lanes 1 and 4) or 100°C (lanes 2 and 5) for 10 min. The samples were thenincubated at 37°C for 30 min in the presence (samples loaded onto lanes 4 and5) or absence (samples loaded onto lanes 1 and 2) of a proteinase cocktail. Afterthe addition of SDS-PAGE dissociation buffer, the samples were then subjectedto electrophoresis. Lane 3 contains molecular mass protein standards of 97.4,66.3, 45.0, 31.0, and 21.5 kDa. O and M denote the running positions of nativeand heat-modified MompA, respectively.
FIG. 3. SDS–10% polyacrylamide gel of OM solubilized at various temper-atures. Lanes: 1, protein standards, 2 to 8, OM solubilized for 5 min at roomtemperature and 40, 50, 60, 70, 80, and 100°C, respectively. The OM samplescontained 15 mg of protein. The molecular masses of the protein standards, inkilodaltons, are shown at the left. X and X9, heat-modifiable OM proteins otherthan MompA; O and M, running positions of native and heat-modified MompA,respectively; Y, minor OM protein with an electrophoretic mobility slightly lessthan that of heat-modified MompA. The gel was stained with Coomassie brilliantblue R250.
TABLE 1. Amino acid analysis of purified MompA and comparisonwith the Msps of T. denticola GM-1 and ATCC 35405
Amino acid MompAMsp
GM-1a ATCC 35405b
Hydrophobic residuesAlanine 7.13 12.15 12.00Valine 4.28 7.05 5.85Leucine 5.46 7.10 8.15Isoleucine 2.61 2.80 3.39Proline 28.27 2.05 3.56Phenylalanine 2.61 4.10 6.19Tryptophan NDc ND NDMethionine 0.24 1.20 0.75
Hydrophilic residuesGlycine 12.82 14.20 10.81Serine 7.60 5.85 6.91Threonine 3.80 6.60 6.30Cysteine ND 0.00 0.00Tyrosine 2.85 3.70 4.01Asparagined 7.84 13.70 14.85Glutaminee 5.70 8.90 4.74
Positively charged residuesLysine 5.23 7.30 8.57Arginine 2.61 1.55 1.66Histidine 0.95 1.70 1.72
Negatively charged residuesAspartic acidd 7.84 13.80 14.85Glutamic acide 5.70 8.90 4.74Hydrophobic residuesf 50.60 36.45 39.89Hydrophilic residuesf 40.61 52.95 47.62Negatively charged residuesf 13.54 22.70 19.59Positively charged residuesf 8.79 10.55 11.95
a Data from reference 48.b Data from reference 19.c ND, not determined.d Combination of aspartic acid and asparagine.e Combination of glutamic acid and glutamine.f Percent of total.
6444 WALKER ET AL. J. BACTERIOL.

positive reaction was noted for T. pectinovorum ATCC 33768,P2, P5, P8, and M1A. Strains S1, X1, and P3 gave a muchweaker response. In the OM sample from M2A, note that theantibody did not react with the major OM protein but did reactstrongly with a minor OM protein with an electrophoreticmobility slightly less than that of the major OM protein (com-pare lanes 10 of Fig. 4A and B). The minor antibody-positivebands below the most prominent immunoreactive band in Fig.4B are most likely due to breakdown products of the Mompsthat occurred during purification of the OM. These minorimmunoreactive bands were absent when whole-cell lysateswere analyzed by Western blotting using anti-MompA serum(data not shown). When unheated OM or whole-cell lysateswere analyzed by Western blotting using anti-MompA serum,a positive immunoreaction was noted for the native species ofMomps (data not shown).
Electron microscopy. Negative-stain electron microscopy ofT. pectinovorum ATCC 33768 whole cells and isolated OM didnot reveal any regular structured array pattern such as thatobserved on whole cells and isolated OM from T. denticolastrains (data not shown). Immuno-gold electron microscopywas used to determine if MompA was exposed on the surfaceof the OM. No gold label was detected when glutaraldehyde-fixed cells were used as whole mounts and processed for im-munoelectron microscopy (not shown). However, when thesecells were embedded and thin sectioned prior to being immu-nologically probed with anti-MompA serum, labeling wasnoted and the gold was predominantly associated with the OM(Fig. 5). No gold labeling was noted when preimmune IgGfraction was used in place of the immune IgG fraction (notshown).
DISCUSSION
A major heat-modifiable protein, MompA, from T. pectino-vorum ATCC 33768 was identified, purified, and characterized.Cellular fractionation procedures and immunoelectron micros-
copy indicate that the protein is present in the OM. Eightadditional strains of T. pectinovorum were examined and foundto produce heat-modifiable Momps with approximately thesame heat-modified molecular mass as MompA. Western blot-ting using anti-MompA serum showed that the Momps fromfour strains reacted with the same intensity as MompA,whereas three had a reduced response. The Momp of T. pecti-novorum M2A did not react with the anti-MompA serum, buta minor OM protein did react with the antiserum. This findingsuggests that at least two serotypes of Momps exist in T. pecti-novorum strains.
MompA was not extracted from the OM with reagents whichare commonly used to remove peripheral membrane proteins(i.e., 0.1% Na2CO3, 1.6 M urea, and 0.6 M NaCl), suggestingthat MompA is intimately associated with or within the OM(18, 40). Several detergents, however, were capable of extract-ing MompA as a number of high-molecular-mass species.These detergents are commonly used to extract porin proteinsin their native functional form from the OMs of a number ofbacterial species (3, 12). The native forms of MompA werestable in the presence of detergents, proteinase, and chemicalsthat are capable of disrupting noncovalent protein bonds.MompA would convert to a putative monomeric 42-kDa formonly by heat treatment. These characteristics of MompA arealso properties common to classical porin proteins (30). Al-though MompA share characteristics with porin proteins, thehigh proline content of MompA argues against it having porinfunction.
The physical characteristics of MompA, discussed above, arealso characteristics of the Msps of T. denticola ATCC 35405and GM-1 (13, 48). MompA and the Msps of T. denticolastrains form detergent- and proteinase-resistant high-molecu-lar-mass species that are converted to a monomeric proteinase-sensitive form by heating (13, 48). However, the transitiontemperature for MompA is less than that for the Msps. West-ern blotting experiments determined that MompA and theMsps were not immunologically related, and the N-terminaland internal peptide fragments of MompA had no homologywith the deduced amino acid sequences of the Msps from T.denticola ATCC 35405, ATCC 33520, and OTK or sequencedpeptide fragments from T. denticola GM-1 (10, 11, 48). Re-cently, Fenno et al. (11) reported the presence of a majorheat-modifiable protein in detergent extracts of whole cells ofT. pectinovorum ATCC 33768 which was most likely MompA.In that study, it was also noted that antiserum to the Msp of T.denticola ATCC 35405 did not react with the T. pectinovorumprotein and that PCR primers which amplified an Msp genefrom T. denticola ATCC 35405, ATCC 35404, ATCC 33520,and OTK and T. vincentii ATCC 35580 did not amplify anyproduct from T. pectinovorum chromosomal DNA. Thus, it isapparent that MompA and the Msps of T. denticola strains arenot genetically related.
Cell fractionation and thin-section immunoelectron micros-copy localized MompA to the OM, but we were unable todemonstrate that the protein is cell surface exposed by usingwhole-mount immunoelectron microscopy. Thus, MompA maybe buried within the OM, or if portions of the protein aresurfaced exposed, they are either physically masked or notreactive with the anti-MompA serum. We have recently iden-tified and isolated a 3-deoxy-D-manno-ocutulosonic acid con-taining lipopolysaccharide-like molecule from the OM of T.pectinovorum (44) whose homogeneous-length O antigencould, in theory, mask the OM surface.
The Msps of T. denticola strains have been shown to besurface exposed, and the surface of the OM is known to pos-sess a regularly structured array when examined by negative-
FIG. 4. (A) SDS–10% polyacrylamide gel of OM from T. pectinovorumstrains stained with Coomassie brilliant blue R250. Lanes: 1, protein standards;2 to 10, OM containing 10 mg of protein from strains ATCC 33768, S1, X1, P2,P3, P5, P8, M1A, and M2A, respectively. Molecular masses of the proteinstandards, in kilodaltons, are shown at the left. (B) Western blot of the gel shownin panel A, reacted with anti-MompA serum. Molecular masses of the prestainedprotein standards, in kilodaltons, are shown at the left.
VOL. 179, 1997 T. PECTINOVORUM 42-kDa OUTER MEMBRANE PROTEIN 6445
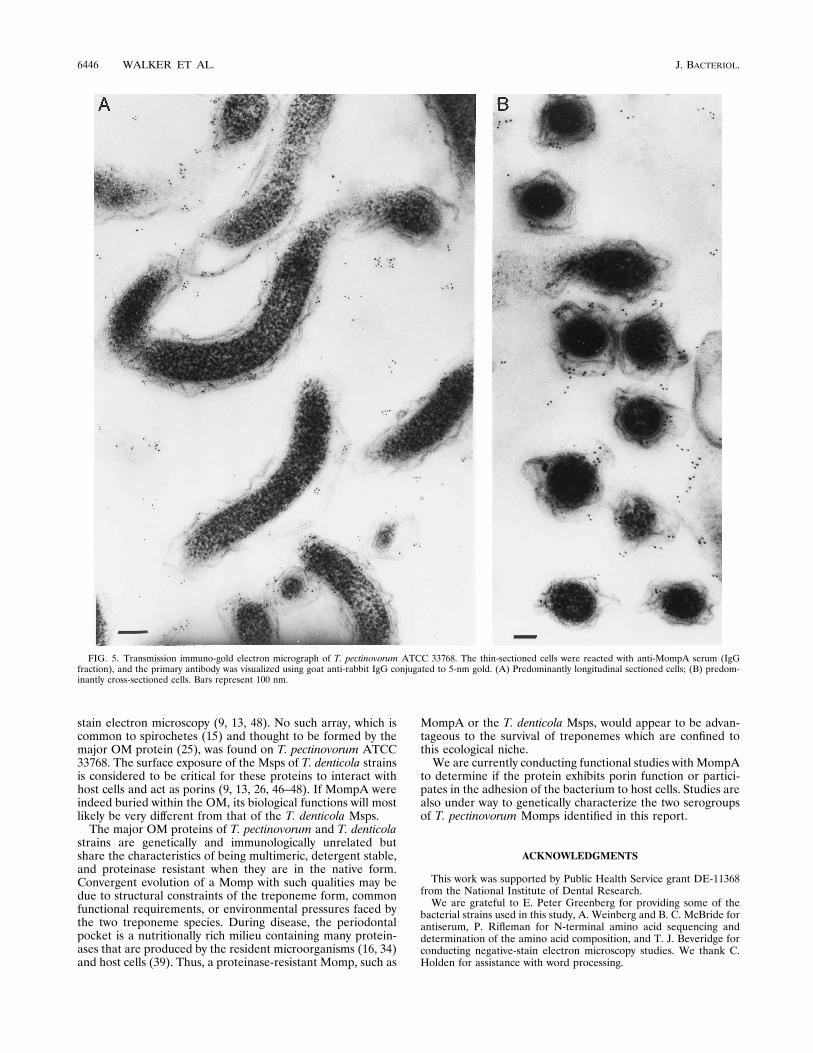
stain electron microscopy (9, 13, 48). No such array, which iscommon to spirochetes (15) and thought to be formed by themajor OM protein (25), was found on T. pectinovorum ATCC33768. The surface exposure of the Msps of T. denticola strainsis considered to be critical for these proteins to interact withhost cells and act as porins (9, 13, 26, 46–48). If MompA wereindeed buried within the OM, its biological functions will mostlikely be very different from that of the T. denticola Msps.
The major OM proteins of T. pectinovorum and T. denticolastrains are genetically and immunologically unrelated butshare the characteristics of being multimeric, detergent stable,and proteinase resistant when they are in the native form.Convergent evolution of a Momp with such qualities may bedue to structural constraints of the treponeme form, commonfunctional requirements, or environmental pressures faced bythe two treponeme species. During disease, the periodontalpocket is a nutritionally rich milieu containing many protein-ases that are produced by the resident microorganisms (16, 34)and host cells (39). Thus, a proteinase-resistant Momp, such as
MompA or the T. denticola Msps, would appear to be advan-tageous to the survival of treponemes which are confined tothis ecological niche.
We are currently conducting functional studies with MompAto determine if the protein exhibits porin function or partici-pates in the adhesion of the bacterium to host cells. Studies arealso under way to genetically characterize the two serogroupsof T. pectinovorum Momps identified in this report.
ACKNOWLEDGMENTS
This work was supported by Public Health Service grant DE-11368from the National Institute of Dental Research.
We are grateful to E. Peter Greenberg for providing some of thebacterial strains used in this study, A. Weinberg and B. C. McBride forantiserum, P. Rifleman for N-terminal amino acid sequencing anddetermination of the amino acid composition, and T. J. Beveridge forconducting negative-stain electron microscopy studies. We thank C.Holden for assistance with word processing.
FIG. 5. Transmission immuno-gold electron micrograph of T. pectinovorum ATCC 33768. The thin-sectioned cells were reacted with anti-MompA serum (IgGfraction), and the primary antibody was visualized using goat anti-rabbit IgG conjugated to 5-nm gold. (A) Predominantly longitudinal sectioned cells; (B) predom-inantly cross-sectioned cells. Bars represent 100 nm.
6446 WALKER ET AL. J. BACTERIOL.

REFERENCES
1. Arakawa, S., and H. K. Kuramitsu. 1994. Cloning and sequence analysis ofa chymotrypsin-like protease from Treponema denticola. Infect. Immun. 62:3424–3433.
2. Armitage, G. C., W. R. Dickinson, R. S. Jenderseck, S. M. Levine, and D. W.Chambers. 1982. Relationship between the percentage of subgingival spiro-chetes and the severity of periodontal disease. J. Periodontol. 53:550–556.
3. Armstrong, S. K., T. R. Parr, Jr., C. D. Parker, and R. E. W. Hancock. 1986.Bordetella pertussis major outer membrane porin protein forms small, anion-selective channels in lipid bilayer membranes. J. Bacteriol. 166:212–216.
4. Blakemore, R. P., and E. Canale-Parola. 1976. Arginine catabolism by Trepo-nema denticola. J. Bacteriol. 128:616–622.
5. Chan, E. C. S., R. Siboo, L. Z. G. Touyz, Y.-S. Qiu, and A. Klitorinos. 1993.A successful method for quantifying viable oral anaerobic spirochetes. OralMicrobiol. Immunol. 8:80–83.
6. Choi, B. K., B. J. Paster, F. E. Dewhirst, and U. B. Gobel. 1994. Diversity ofcultivable and uncultivable oral spirochetes from a patient with severe de-structive periodontitis. Infect. Immun. 62:1889–1895.
7. Chu, L., and S. C. Holt. 1994. Purification and characterization of a 45 kDahemolysin from Treponema denticola ATCC 35404. Microb. Pathog. 16:197–212.
8. Ebersole, J. L., E. E. Frey, M. A. Taubman, and E. J. Smith. 1980. An ELISAfor measuring serum antibodies to Actinobacillus actinomycetemocomitans. J.Periodontal Res. 15:621–632.
9. Egli, C., W. K. Leung, K. H. Muller, R. E. W. Hancock, and B. C. McBride.1993. Pore-forming properties of the major 53-kilodalton surface antigenfrom the outer sheath of Treponema denticola. Infect. Immun. 61:1694–1699.
10. Fenno, J. C., K.-H. Muller, and B. C. McBride. 1996. Sequence analysis,expression, and binding activity of recombinant major outer sheath protein(Msp) of Treponema denticola. J. Bacteriol. 178:2489–2497.
11. Fenno, J. C., G. W. K. Wong, P. M. Hannam, K.-H. Muller, W. K. Leung andB. C. McBride. 1997. Conservation of msp, the gene encoding the majorouter membrane protein of oral Treponema spp. J. Bacteriol. 179:1082–1089.
12. Garavito, R. M., and J. P. Rosenbusch. 1986. Isolation and crystallization ofbacterial porin. Methods Enzymol. 125:309–328.
13. Haapasalo, M., K. H. Muller, V.-J. Uitto, W. K. Leung, and B. C. McBride.1992. Characterization, cloning, and binding properties of the major 53-kilodalton Treponema denticola surface antigen. Infect. Immun. 60:2058–2065.
14. Hindahl, M. S., and B. H. Iglewski. 1984. Isolation and characterization ofthe Legionella pneumophila outer membrane. J. Bacteriol. 159:107–113.
15. Holt, S. C. 1978. Anatomy and chemistry of spirochetes. Microbiol. Rev.42:114–160.
16. Holt, S. C., and T. E. Bramanti. 1991. Factors in virulence expression andtheir role in periodontal disease pathogenesis. Crit. Rev. Oral Biol. Med.2:177–281.
17. Ishihara, K., and H. K. Kuramitsu. 1995. Cloning and expression of aneutral phosphatase gene from Treponema denticola. Infect. Immun. 63:1147–1152.
18. Ito, K., and Y. Akiyama. 1991. In vivo analysis of integration of membraneproteins in Escherichia coli. Mol. Microbiol. 5:2243–2253.
19. Kokeguchi, S., M. Miyamoto, K. Kato, I. Tanimoto, H. Kurihara, and Y.Murayama. 1994. Isolation and characterization of a 53 kDa major cellenvelope protein antigen from Treponema denticola ATCC 35405. J. Peri-odontal Res. 29:70–78.
20. Laemmli, U. K. 1970. Cleavage of the structural proteins during the assemblyof the head of bacteriophage T4. Nature (London) 227:680–685.
21. Leschine, S. B., and E. Canale-Parola. 1980. Rifampin as a selective agentfor isolation of oral spirochetes. J. Clin. Microbiol. 6:792–795.
22. Listgarten, M. A., and S. Levin. 1981. Positive correlations between theproportions of subgingival spirochetes and motile bacteria and susceptibilityof human subjects to periodontal deterioration. J. Clin. Periodontol. 8:122–138.
23. Loesche, W. L., and B. E. Laughon. 1982. Role of spirochetes in periodontaldisease, p. 62–75. In R. J. Genco and S. E. Mergenhagen (ed.), Host-parasiteinteractions in periodontal disease. American Society for Microbiology,Washington, D.C.
24. Markwell, M. A. K., S. M. Haas, L. L. Bieber, and N. E. Tolbert. 1978. Amodification of the Lowry procedure to simplify protein determination inmembrane and lipoprotein samples. Anal. Biochem. 87:206–210.
25. Masuda, K., and T. Kawata. 1982. Isolation, properties, and reassembly ofouter sheath carrying a polygonal array from an oral treponeme. J. Bacteriol.150:1405–1413.
26. Mathers, D. A., W. K. Leung, J. C. Fenno, Y. Hong, and B. C. McBride. 1996.Major surface protein complex of Treponema denticola depolarizes and in-duces ion channels in HeLa cell membranes. Infect. Immun. 64:2904–2910.
27. Moore, L. V., W. E. C. Moore, E. P. Cato, R. M. Smibert, J. A. Burmeister,A. M. Best, and R. R. Ranney. 1987. Bacteriology of human gingivitis. J.Dent. Res. 66:989–995.
28. Moore, W. E. C., L. V. Moore, R. R. Ranney, R. M. Smibert, J. A. Burmeister,and H. A. Schenkein. 1991. The microflora of periodontal sites showingactive destructive progression. J. Clin. Periodontol. 18:729–739.
29. Moos, M. 1992. Isolation of proteins for microsequence analysis, p. 8.7.1–8.7.12. In J. E. Coligan, A. M. Kruisbeek, D. H. Margulies, E. M. Shevach,and W. Strober (ed.), Current protocols in immunology. Greene PublishingAssociates and John Wiley & Sons, New York, N.Y.
30. Nikaido, H. 1992. Porins and specific channels of bacterial outer membranes.Mol. Microbiol. 6:435–442.
31. Paster, B. J. Personal communication.32. Rosen, G., R. Naor, S. Kutner, and M. N. Sela. 1994. Characterization of
fibrinolytic activities of Treponema denticola. Infect. Immun. 62:1749–1754.33. Rosen, G., R. Naor, E. Rahamim, R. Yishai, and M. N. Sela. 1995. Proteases
of Treponema denticola outer sheath and extracellular vesicles. Infect. Im-mun. 63:3973–3979.
34. Schmidt, E. F., W. A. Bretz, R. A. Hutchinson, and W. J. Loesche. 1988.Correlation of the hydrolysis of benzoyl-arginine naphthylamide (BANA) byplaque with clinical parameters and subgingival levels of spirochetes inperiodontal patients. J. Dent. Res. 67:1505–1509.
35. Sela, M. N., A. Weinberg, R. Borinsky, S. C. Holt, and T. Dishon. 1988.Inhibition of superoxide production in human polymorphonuclear leuko-cytes by oral treponemal factors. Infect. Immun. 56:589–594.
36. Smibert, R. M. 1984. Genus III. Treponema Schaudinn 1905, 1728AL, p.49–57. In N. R. Kreig and J. G. Holt (ed.), Bergey’s manual of systematicbacteriology, vol. 1. The Williams & Wilkins Co., Baltimore, Md.
37. Smibert, R. M., and J. A. Burmeister. 1983. Treponema pectinovorum sp. nov.isolated from humans with periodontitis. Int. J. Syst. Bacteriol. 33:852–856.
38. Smibert, R. M., J. L. Johnson, and R. R. Ranney. 1984. Treponema socranskiisp. nov., Treponema socranskii subsp. socranskii sub. sp. nov., Treponemasocranskii subsp. buccale subsp. nov., and Treponema socranskii subsp. pare-dis subs. nov. isolated from the human periodontia. Int. J. Syst. Bacteriol.34:457–462.
39. Sorsa, T., P.-L. Ding, T. Ingman, T. Salo, U. Westerlund, M. Haapasalo, H.Tschesche and Y. T. Konttinen. 1995. Cellular source, activation and inhi-bition of dental plaque collagenase. J. Clin. Periodontol. 22:709–717.
40. Stader, J., and T. J. Silhavy. 1988. A progenitor of the outer membraneLamB trimer. J. Bacteriol. 170:1973–1974.
41. Tzellas, N., B. J. Paster, B. E. Coleman, and F. E. Dewhirst. 1996. Not yetcultivable spirochetes from ANUG sites. J. Dent. Res. 75:319, abstr. 2411.
42. Uitto, V.-J., Y. M. Pan, W. K. Leung, H. Larjava, R. P. Ellen, B. B. Finlay,and B. C. McBride. 1995. Cytopathic effects of Treponema denticola chymo-trypsin-like proteinase on migrating and stratified epithelial cells. Infect.Immun. 63:3401–3410.
43. Walker, S. G., C. A. Flemming, F. G. Ferris, T. J. Beveridge, and G. W.Bailey. 1989. Physicochemical interaction of Escherichia coli envelopes andBacillus subtilis cell walls with two clays and ability of the composite toimmobilize heavy metals from solution. Appl. Environ. Microbiol. 42:315–335.
44. Walker, S. G., and S. C. Holt. Unpublished data.45. Weber, F. H., and E. Canale-Parola. 1984. Pectinolytic enzymes of oral
spirochetes from humans. Appl. Environ. Microbiol. 48:61–67.46. Weinberg, A. Porin-like activities of the 64-kDa major surface protein (Msp)
of Treponema denticola. Submitted for publication.47. Weinberg, A., and S. C. Holt. 1990. Interaction of Treponema denticola TD-4,
GM-1, and MS25 with human gingival fibroblasts. Infect. Immun. 58:1720–1729.
48. Weinberg, A., and S. C. Holt. 1991. Chemical and biological activities of a64-kilodalton outer sheath protein from Treponema denticola strains. J. Bac-teriol. 173:6935–6947.
49. Wyss, C., B. K. Choi, P. Schupbach, B. Guggenheim, and U. B. Gobel. 1996.Treponema maltophilum sp. nov., a small oral spirochete isolated from hu-man periodontal lesions. Int. J. Syst. Bacteriol. 46:745–752.
50. Yotis, W. W., V. K. Sharma, C. Gopalsami, S. Chegini, J. McNulty, K.Hoerman, J. Keene, Jr., and L. G. Simonson. 1991. Biochemical propertiesof the outer membrane of Treponema denticola. J. Clin. Microbiol. 29:1397–1406.
VOL. 179, 1997 T. PECTINOVORUM 42-kDa OUTER MEMBRANE PROTEIN 6447




















