
Identification and genetic characterisation of orthopaedic Staphylococcus isolates collected in...
6
Identification and genetic characterisation of orthopaedic Staphylococcus isolates collected in Italy by automated EcoRI ribotyping Agnese Andollina a , Alessandra De Cesare b , Giorgia Bertoni a , Liliana Modelli a , Gerardo Manfreda b, * a Research Laboratory of Clinical Pathology and Microbiology, Rizzoli Orthopaedic Institute, Bologna, Italy b Department of Food Science, Alma Mater Studiorum, University of Bologna, Via S. Giacomo 9, 40126 Bologna, Italy Received 19 January 2004; received in revised form 9 March 2004; accepted 22 March 2004 First published online 9 April 2004 Abstract The aim of this study was to evaluate the possibility to use automated EcoRI ribotyping to address, during the same analysis, both identification and genetic characterisation of 38 Staphylococcus aureus and 64 coagulase-negative staphylococci collected from surgical injuries. The ribotyping identification results confirmed those obtained using the API Staph Ò system for 96% of the isolates. All strains were successfully genotyped and the ribotyping discriminatory power, calculated using the Simpson’s index of dis- crimination, was very high for both groups of staphylococci tested. The same, as well as different biotypes, were identified among isolates with the identical ribotyping profile. Ó 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. Keywords: Staphylococcus; Automated ribotyping; Identification; Characterisation 1. Introduction Staphylococcus aureus and the coagulase-negative staphylococci (CNS) represent the 31.5% and 30.2% of the isolates collected from surgical injuries at the Rizzoli Orthopaedic Institutes (Bologna, Italy) during the year 2000. In the hospital routine work the staph- ylococci identification is often not completed by their genetic characterisation to strain level. In fact, many characterisation techniques are very laborious and produce unrepeatable results. However, the isolates characterisation might be very useful, from an epide- miological point of view, to differentiate sporadic and clonal strains, to trace their sources and to limit their diffusion. Among the earliest schemes for classification of staphylococci and micrococci there was that of Baird Parker, which remained the method of choice until that of Kloos and Schleifer, with the identification of several new Staphylococcus species [1]. More recently, several molec- ular approaches have been proposed for the identification of Staphylococcal taxa. These approaches fall into two categories: those based on the detection of species-specific sequences, that are not present in the chromosome of all staphylococci, and those based on the detection of se- quence variations in ubiquitous elements, such as rRNA and tRNA operons or chaperonin-encoding genes [2]. Staphylococcus aureus and CNS epidemiological studies have been traditionally performed by bacterio- phage typing [3,4], plasmid pattern analysis [5,6], whole- cell protein profile analysis [7], immunoblotting profile analysis [8]. More recently, many genotyping methods have been applied, such as pulsed-field gel electropho- resis (PFGE) [9], fluorescent amplified fragment length * Corresponding author. Tel.: +39-051-2096765/2096766; fax: +39- 051-2096764. E-mail address: [email protected] (G. Manfreda). 0378-1097/$22.00 Ó 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.femsle.2004.03.044 FEMS Microbiology Letters 234 (2004) 275–280 www.fems-microbiology.org
Transcript of Identification and genetic characterisation of orthopaedic Staphylococcus isolates collected in...

FEMS Microbiology Letters 234 (2004) 275–280
www.fems-microbiology.org
Identification and genetic characterisation of orthopaedicStaphylococcus isolates collected in Italy by
automated EcoRI ribotyping
Agnese Andollina a, Alessandra De Cesare b, Giorgia Bertoni a, Liliana Modelli a,Gerardo Manfreda b,*
a Research Laboratory of Clinical Pathology and Microbiology, Rizzoli Orthopaedic Institute, Bologna, Italyb Department of Food Science, Alma Mater Studiorum, University of Bologna, Via S. Giacomo 9, 40126 Bologna, Italy
Received 19 January 2004; received in revised form 9 March 2004; accepted 22 March 2004
First published online 9 April 2004
Abstract
The aim of this study was to evaluate the possibility to use automated EcoRI ribotyping to address, during the same analysis,
both identification and genetic characterisation of 38 Staphylococcus aureus and 64 coagulase-negative staphylococci collected from
surgical injuries. The ribotyping identification results confirmed those obtained using the API Staph� system for 96% of the isolates.
All strains were successfully genotyped and the ribotyping discriminatory power, calculated using the Simpson’s index of dis-
crimination, was very high for both groups of staphylococci tested. The same, as well as different biotypes, were identified among
isolates with the identical ribotyping profile.
� 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: Staphylococcus; Automated ribotyping; Identification; Characterisation
1. Introduction
Staphylococcus aureus and the coagulase-negative
staphylococci (CNS) represent the 31.5% and 30.2% ofthe isolates collected from surgical injuries at the
Rizzoli Orthopaedic Institutes (Bologna, Italy) during
the year 2000. In the hospital routine work the staph-
ylococci identification is often not completed by their
genetic characterisation to strain level. In fact, many
characterisation techniques are very laborious and
produce unrepeatable results. However, the isolates
characterisation might be very useful, from an epide-miological point of view, to differentiate sporadic and
clonal strains, to trace their sources and to limit their
diffusion.
* Corresponding author. Tel.: +39-051-2096765/2096766; fax: +39-
051-2096764.
E-mail address: [email protected] (G. Manfreda).
0378-1097/$22.00 � 2004 Federation of European Microbiological Societies
doi:10.1016/j.femsle.2004.03.044
Among the earliest schemes for classification of
staphylococci and micrococci there was that of Baird
Parker, which remained themethod of choice until that of
Kloos and Schleifer, with the identification of several newStaphylococcus species [1]. More recently, several molec-
ular approaches have been proposed for the identification
of Staphylococcal taxa. These approaches fall into two
categories: those based on the detection of species-specific
sequences, that are not present in the chromosome of all
staphylococci, and those based on the detection of se-
quence variations in ubiquitous elements, such as rRNA
and tRNA operons or chaperonin-encoding genes [2].Staphylococcus aureus and CNS epidemiological
studies have been traditionally performed by bacterio-
phage typing [3,4], plasmid pattern analysis [5,6], whole-
cell protein profile analysis [7], immunoblotting profile
analysis [8]. More recently, many genotyping methods
have been applied, such as pulsed-field gel electropho-
resis (PFGE) [9], fluorescent amplified fragment length
. Published by Elsevier B.V. All rights reserved.
276 A. Andollina et al. / FEMS Microbiology Letters 234 (2004) 275–280
polymorphism (FAFLP) [10], polymerase chain reaction
restriction fragment length polymorphism (PCR-RFLP)
[11] and multilocus sequence typing [12]. The major
limitations of most typing methods include a lack of
standardisation, a need for highly skilled technical staff,and significant hands-on time required for performance.
Due to its facility of use and speed of analysis, an au-
tomated genotyping technique, such as that represented
by automated ribotyping, may allow clinical laborato-
ries to apply molecular typing more broadly. Automated
ribotyping, which is possible using the RiboPrinter�
system (DuPont, Qualicon Inc.), is based on the analysis
of the genomic fragments generated by restriction di-gestion of the rRNA operons, highly conserved and
present in several copies on the bacterial chromosome
[13]. The most conserved regions of the operons allow
identification of a bacterium to genus and species level,
whereas variable and flanking regions permit discrimi-
nation between strains [14]. Manual ribotyping has been
described as a useful molecular technique for typing of
staphylococci [15–18]. In this study automated EcoRIribotyping was used to confirm the identification of 102
Staphylococcus isolates previously submitted to API
Staph�. Moreover, the discriminatory power of auto-
mated ribotyping as typing method was calculated and
the genetic relationships among strains were quantified
using cluster analysis.
2. Materials and methods
2.1. Bacterial isolates
A total of 102 Staphylococcus isolates, collected from
surgical injuries during three months (October–Decem-
ber) of the year 2000 at the Rizzoli Orthopaedic Insti-
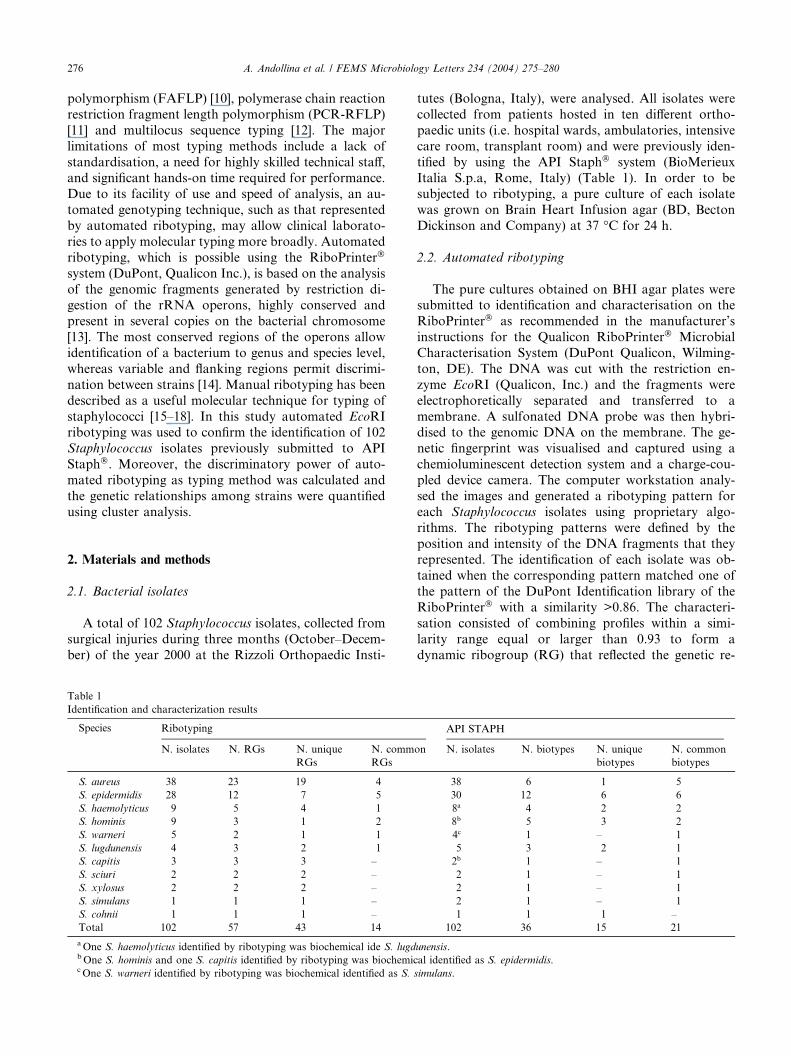
Table 1
Identification and characterization results
Species Ribotyping
N. isolates N. RGs N. unique
RGs
N. comm
RGs
S. aureus 38 23 19 4
S. epidermidis 28 12 7 5
S. haemolyticus 9 5 4 1
S. hominis 9 3 1 2
S. warneri 5 2 1 1
S. lugdunensis 4 3 2 1
S. capitis 3 3 3 –
S. sciuri 2 2 2 –
S. xylosus 2 2 2 –
S. simulans 1 1 1 –
S. cohnii 1 1 1 –
Total 102 57 43 14
aOne S. haemolyticus identified by ribotyping was biochemical ide S. lugdbOne S. hominis and one S. capitis identified by ribotyping was biochemiccOne S. warneri identified by ribotyping was biochemical identified as S.
tutes (Bologna, Italy), were analysed. All isolates were
collected from patients hosted in ten different ortho-
paedic units (i.e. hospital wards, ambulatories, intensive
care room, transplant room) and were previously iden-
tified by using the API Staph� system (BioMerieuxItalia S.p.a, Rome, Italy) (Table 1). In order to be
subjected to ribotyping, a pure culture of each isolate
was grown on Brain Heart Infusion agar (BD, Becton
Dickinson and Company) at 37 �C for 24 h.
2.2. Automated ribotyping
The pure cultures obtained on BHI agar plates weresubmitted to identification and characterisation on the
RiboPrinter� as recommended in the manufacturer’s
instructions for the Qualicon RiboPrinter� Microbial
Characterisation System (DuPont Qualicon, Wilming-
ton, DE). The DNA was cut with the restriction en-
zyme EcoRI (Qualicon, Inc.) and the fragments were
electrophoretically separated and transferred to a
membrane. A sulfonated DNA probe was then hybri-dised to the genomic DNA on the membrane. The ge-
netic fingerprint was visualised and captured using a
chemioluminescent detection system and a charge-cou-
pled device camera. The computer workstation analy-
sed the images and generated a ribotyping pattern for
each Staphylococcus isolates using proprietary algo-
rithms. The ribotyping patterns were defined by the
position and intensity of the DNA fragments that theyrepresented. The identification of each isolate was ob-
tained when the corresponding pattern matched one of
the pattern of the DuPont Identification library of the
RiboPrinter� with a similarity >0.86. The characteri-
sation consisted of combining profiles within a simi-
larity range equal or larger than 0.93 to form a
dynamic ribogroup (RG) that reflected the genetic re-
API STAPH
on N. isolates N. biotypes N. unique
biotypes
N. common
biotypes
38 6 1 5
30 12 6 6
8a 4 2 2
8b 5 3 2
4c 1 – 1
5 3 2 1
2b 1 – 1
2 1 – 1
2 1 – 1
2 1 – 1
1 1 1 –
102 36 15 21
unensis.
al identified as S. epidermidis.
simulans.

A. Andollina et al. / FEMS Microbiology Letters 234 (2004) 275–280 277
latedness of the isolates [14]. The profiles collected
during this research were labelled with alphanumeric
codes (i.e. 153-42S4), which defined the ribogroup of
the isolate examined.
2.3. Simpson’s index of discrimination (SID)
The suitability of EcoRI ribotyping and API Staph�
for differentiation of Staphylococcus isolates was deter-
mined using the Simpson’s numerical index, as described
by Hunter and Gaston [19]. This index represents the
probability that two randomly chosen isolates would be
distinguished by the technique used. As the numericalindex approaches the maximum value of one (repre-
senting 100% discriminatory ability) the higher the
probability that the technique will be able to discrimi-
nate between two unrelated isolates.
2.4. Cluster analysis
Automated EcoRI ribotyping profiles were importedinto the program named BioNumerics (version 2.0;
Applied Maths) and dendrograms were generated based
on the banding pattern similarities. Unweighted pair
group method with arithmetic averages (UPGMA) and
Pearson correlation coefficient were used.
Fig. 1. Staphylococcus aureus cluster analysis results.
3. Results
All isolates of this study set were successfully ribo-
typed. The ribotyping identification results confirmed
those obtained biochemically for all isolates but four
(Table 1). In particular, all S. aureus, S. epidermidis,
S. lugdunensis, S. sciuri, S. xylosus, S. simulans and
S. cohnii identified by ribotyping were biochemically
confirmed (Table 1). Moreover, one of nine isolatesidentified as S. haemolyticus by ribotyping was bio-
chemically classified as S. lugdunensis, biotype 6716152;
one of nine isolates identified as S. hominis was bio-
chemically classified as S. epidermidis, biotype 6306152;
one of three isolates identified as S. capitis was bio-
chemically classified as S. epidermidis, biotype 6306111;
one of five isolates identified as S. warneri was bio-
chemically classified as S. simulans, biotype 6430113.Among the 57 ribogroups identified, 75% were asso-
ciated with single isolates and labelled as unique,
whereas 25% were shared between two to ten isolates
and labelled as common (Table 1). Both common, as
well as unique ribotyping profiles, were characterised by
8–13 well defined non-overlapping bands, with a mo-
lecular weight ranging from less than one to more than
48 kbp. Among the 36 biotypes identified, 42% wereassociated with single isolates and 58% to two to six
isolates. The SID calculated for S. aureus was 0.94,
whereas the SID for CNS, referring for a group of iso-
lates which comprise several different species, was 0.96.
TheS. aureus ribotyping profileswere clustered in three
main groups (Fig. 1), containing 17, 12 and 9 strains, re-
spectively, with a similarity ranging from 52% to 98%. Asexpected, using the BioNumerics software the majority of
the isolates classified within the same ribogroup clustered
with a similarity equal or larger than 93%. However,
strains A-SR 06 and A-SR 89, as well as A-SR 42 and A-
SR 97, clustering with similarities of 93%, were assigned
to different ribogroups because they have a different
number of bands, resulting in different genetic profiles.
Otherwise, strain A-SR07, with ribogroup 153-42S4,clustered with a similarity of 90%with strainsA-SR71,A-
SR90, A-SR94, A-SR77, A-SR92, A-SR15, A-SR05 be-
longing to the same 153-42S4 ribogroup.
The CNS ribotyping profiles belonging to the same
species generally clustered together (Fig. 2). In particu-
lar, all S. haemolyticus strains were grouped with a
similarity ranging from 68% to 98%. The only exception
was represented by isolate H-SR 60 clustering along

278 A. Andollina et al. / FEMS Microbiology Letters 234 (2004) 275–280
with the S. hominis strains with a similarity ranging from
85% to 90%. The S. lugdunensis isolates were grouped
along with the S. cohnii labeled as CO-SR91 with a
Fig. 2. Coagulase-negative staphylococci cluster analysis results.
similarity ranging from 68% to 95%. The S. xylosus,
capitis and sciuri isolates clustered together, as well as
the S. epidermidis strains. However the isolates belong-
ing to this last species were grouped in two clusters with
a similarity lower than 30%. Finally the S. simulans
isolate clustered alone and did not showed any signifi-
cant similarity to the isolates of the other species. From
the biochemical point of view, strain H-SR60 showed
identical biotype to strains H-SR03, H-SR17 and H-
SR43 showing different ribogroups. Moreover, strains
E-SR04, E-SR112, E-SR18 and E-SR24 shared their
biotypes with strains of the same species classified in a
different cluster.
4. Discussion
The correct identification of Staphylococcus isolates,
as well as their characterisation to strain level, is crucial
to develop efficient epidemiological surveillance pro-
grams and to manage an outbreak investigation, inwhich a cluster of infections due to organisms of the
same species is identified. In particular, the strain typing
provides evidence of biological and genetic relatedness
among isolates, and allows to determine the presence of
clonal spread and sources of infection [20–22].
One of the aims of this research was to evaluate the
possibility to use the RiboPrinter� to confirm the
identification results of 102 Staphylococcus isolates ob-tained using the rapid biochemical test named API
Staph�. The ribotyping identification results confirmed
those obtained biochemically for all isolates but four. In
particular, two isolates biochemically identified as
S. epidermidis were identified as S. hominis and S. capitis
by ribotyping. Moreover, two isolates biochemically
identified as S. lugdunensis and S. simulans, respectively,
were classified as S. haemolyticus and S. warneri usingribotyping. The RiboPrinter� identification process is
based on the analysis of rRNA restriction fragments,
highly conserved within genus and species, whereas
biochemical tests are based on the expression of proteins
and enzymes environmentally conditioned. This differ-
ence might explain the lack of correspondence between
some identification results obtained using the two
methods. Otherwise, it seems significant that the groupof species identified by the two techniques was the same.
For the evaluation of genotyping techniques for bac-
teria characterisation, Struelens [23] proposed strict cri-
teria, such as high typeability, reproducibility, stability
and discriminatory power. All the isolates of this study
were successfully ribotyped (100% typeability) and the
ribotyping discriminatory power, calculated using the
SID, was very high for both S. aureus (0.94) and CNS(0.96). These results might be due to the presence of six
rRNAoperons [24] inStaphylococcus genome. In fact, the
ribotyping discriminatory power generally increases with

A. Andollina et al. / FEMS Microbiology Letters 234 (2004) 275–280 279
the number of rRNA operons, that varies from one, in
Rickettsia prowazekii and Mycoplasma pneumoniae to a
maximum of 15 in Clostridium paradoxum [25].
Due to its complete automation and standardisation,
the stability and reproducibility of the ribotyping patternsis very high [26]. In fact, the pattern of one isolate is always
the same also after one year period. Staphylococcus ri-
botyping usingHidIII and EcoRI, as restriction enzymes,
has been described from other authors [16,27]. They
found that EcoRI produces more bands and ribotypes
than HidIII. The results of this research confirm that
EcoRI is a good enzyme for the discrimination of different
genetic types, within the sameStaphylococcus species, andfor Staphylococcus identification. In fact, the ribogroup
designation obtained for all strains tested was always
linked to the Staphylococcus species.
The similarity among the ribotyping profiles collected
in this research was quantified by cluster analysis using
the BioNumerics software. The reason why some S.
aureus isolates (i.e. A-SR06 and A-SR89, A-SR42 and
A-SR97) showing similarities of 93% were classifiedwithin different ribogroups and isolates with the same
ribogroup (i.e. A-SR07 and A-SR05) clustered with
similarity <93% is related to the different algorithms
used in the RiboPrinter� and BioNumerics software to
perform the similarity analysis.
The S. aureus strains with ribogroup 153-56S4 shared
the same biotype (Table 2), whereas isolates with ribo-
groups 153-42S4, 153-47S3 and 153-51S4 were charac-terised by two biotypes that differed for only one
biochemical test (i.e. b-naphtyl-acid phosphate, N-ace-
tyl-glucosamine and potassium nitrate). The four
S. warneri strains in ribogroup 153-51S6 shared the
same biotype, as did the two S. epidermidis strains in
ribogroup 153-46S8 (Table 2). In contrast, all other
CNS isolates showed different biotypes for strains with
Table 2
Biotypes identified among isolates with the same ribogroup
Species Common RGs (N. isolates) B
S. aureus 153-42S4(8) 6
153-47S3(5) 6
153-51S4(3) 6
153-56S4(3) 6
S. epidermidis 153-46S6(10) 6
153-42S1(3) 6
153-56S6(4) 6
153-89S5(2) 6
153-46S8(2) 6
S. haemolyticus 153-46S7(5) 6
S. hominis 153-46S1(4) 6
153-51S1(4) 6
S. warneri 153-51S6(4) 6
S. lugdunensis 153-47S8(2) 6
a Biotypes 6712151 and 6306152 correspond to Staph lugdunensis and Stap
the same ribogroup. In particular, the biotypes of the
S. haemolyticus strains in ribogroup 153-46S7, the
S. hominis strains in ribogroups 153-46S1 and 153-51S1
as well as the S. lugdunensis strains in ribogroup 153-
47S8 differed by two or more biochemical reactions. Incontract, the differences in the biochemical profiles of
the S. epidermidis strains in ribogroups 153-46S6, 153-
42S1, 153-56S6 and 153-89S5 were limited to single re-
actions, such as production of the enzymes urease or
arginine dihydrolase (ADH) or production of a-naphtyl-acid phosphate (PAL). Finally, the same biotypes (i.e.,
S. aureus 6736153; S. epidermidis 6706112, 6706113,
6702112; S. hominis 6216153, 6212132) were identifiedfor isolates with different ribogroups.
If further studies will confirm that automated EcoRI
ribotyping is suitable to obtain both the identification as
well as the genetic characterisation of Staphylococcus
isolates during the same analysis, clinical laboratories
having to deal with large number of isolates might select
the RiboPrinter� as first identification and typing
method. The automated ribotyping analysis is per-formed in 8 h and the RiboPrinter� can process 32
isolates per day. API Staph� test, PCR-RFLP, PFGE,
AFLP, and DNA sequencing time to results are 1, 1, 3, 2
and 2 days, respectively [28]. The cost of the analysis of
each sample by automated ribotyping (i.e. 51 Euro) is
comparable with that obtained adding the cost of the
manual API Staph� test (i.e. 6.5 Euro for each sample)
to that of mostly used typing methods. It has been es-timated that the PCR-RFLP, PFGE, AFLP, and DNA
sequencing costs are about 11, 38, 20 and 40 Euro per
sample [28]. Moreover the automated ribotyping cost
decrease over the years in relation to the number of
isolates tested resulting lower than that of manual ri-
botyping and PFGE in laboratories analysing more than
2000 isolates per year [29].
iotypes for each ribotype (N. isolates)
732153(2); 6736153(6)
736113(2); 6736153(3)
736152(2); 6734152(1)
736151(3)
706112(5); 6706113(2); 6702112(1); 6702113(1); 6704113(1)
706110(1); 6706113(1); 6706132(1)
702112(2); 6706112(2)
706111(1); 6706113(1)
706113(2)
232150(3); 6610171(1); 6712151(1)a
216153(3); 6212132(1)
306152(1)a; 6212132(1); 6216152(1); 6216153(1)
332113(4)
712110(1); 6716152(1)
h epidermidis, respectively.

280 A. Andollina et al. / FEMS Microbiology Letters 234 (2004) 275–280
Acknowledgements
The authors would like to thank Dr. Ludwig Mo-
roder (APPA, Bolzano, Italy) for assistance in the
cluster analysis.
References
[1] Gemmell, C.G. and Dawson, J.E. (1982) Identification of coag-
ulase-negative Staphylococci with the API Staph system. J. Clin.
Microbiol. 16, 874–877.
[2] Chesneau, O., Morvan, A., Aubert, S. and El Solh, N. (2000) The
value of rRNA gene restriction site polymorphism analysis for
delineating taxa in the genus Staphylococcus. Int. J. Syst. Evol.
Microbiol. 50, 689–697.
[3] Luzar, M.A., Coles, G.A., Faller, B., Slingeneyer, A., DahDah,
G., Briat, C., Wone, C., Knefati, Y., Kessler, M. and Peluso, F.
(1990) Staphylococcus aureus nasal carriage and infection in
patients on continuous ambulatory peritoneal dialysis. New Engl.
J. Med. 322, 505–509.
[4] Yu, V.L., Goetz, A., Wagener, M., Smith, P.B., Rihs, J.D.,
Hanchett, J. and Zuraleff, J.J. (1986) Staphylococcus aureus nasal
carriage and infection in patients on haemodialysis: efficacy of
antibiotic prophylaxis. New Engl. J. Med. 315, 91–96.
[5] Archer, G.K. and Mayhall, C.G. (1983) Comparison of epidemi-
ological markers used in the investigation of an outbreak of
methicillin-resistant Staphylococcus aureus infections. J. Clin.
Microbiol. 18, 395–399.
[6] Collins, J.K., Smith, J.S. and Kelly, M.T. (1984) Comparison of
phage typing, plasmid mapping, and antibiotic resistant patterns
as epidemiologic markers in a nosocomial outbreak of methicillin
resistant Staphylococcus aureus infections. Diagn. Microbiol.
Infect. Dis. 2, 233–245.
[7] Costas, M., Cookson, B.D., Talsania, H.G. and Owen, R.J. (1989)
Numerical analysis of electrophoresis protein patterns of methi-
cillin resistant strains of Staphylococcus aureus. J. Clin. Microbiol.
27, 2574–2581.
[8] Gaston, M.S., Duff, P.S., Naidoo, J., Ellis, K., Roberts, J.I.S.,
Richardson, J.F., Marples, R.R. and Cooke, E.M. (1988) Eval-
uation of electrophoretic methods for typing methicillin resistant
Staphylococus aureus. J. Med. Microbiol. 26, 189–197.
[9] Wang, J.T., Chen, Y.C., Yang, T.L. and Chang, S.C. (2002)
Molecular epidemiology and antimicrobial susceptibility of meth-
icillin-resistant Staphylococcus aureus in Taiwan. Diagn. Micro-
biol. Infect. Dis. 42, 199–203.
[10] Grady, R., Blanc, D., Hauser, P. and Stanley, J. (2001) Genotyp-
ing of European isolates of methicillin-resistant Staphylococcus
aureus by fluorescent amplified-fragment length polymorphism
analysis (FAFLP) and pulsed field gel electrophoresis (PFGE)
typing. J. Med. Microbiol. 50, 588–593.
[11] Wichelhaus, T.A., Hunfeld, K.P., Boddinghaus, B., Kraiczy, P.,
Schafer, V. and Brade, V. (2001) Rapid molecular typing of
methicillin resistant Staphylococcus aureus by PCR-RFLP. Inf.
Contr. Hosp. Epidemiol. 22, 294–298.
[12] Grundmann, H., Hori, S., Enright, M.C., Webster, C., Tami, A.,
Feil, E.J. and Pitt, T. (2002) Determining of genetic structure of
the natural population of Staphylococcus aureus: a comparison of
multilocus sequence typing with pulsed field gel electrophoresis,
randomly amplified polymorphic DNA analysis and phage typing.
J. Clin. Microbiol. 40 (12), 4544–4546.
[13] Woese, C.R. (1987) Bacterial evolution. Microbiol. Rev. 51, 221–
271.
[14] Bruce, J. (1996) Automated system rapidly identifies and cha-
racterises micro-organisms in foods. Food Technol. 50, 77–78.
[15] El Sohl, N., De Buyser, M.L., Morvan, A., Grimont, F., Walcher-
Salesse, S., Aubert, S., Monzon-Moreno, C., Chesneau, O. and
Allignet, J. (1990) Use of Bacillus subtilis 16S rRNA genes as a
probe to identify species, subspecies and types in the genus
Staphylococcus. In: Molecular biology of staphylococci (Novick,
R.P., Ed.), pp. 585–593. VCH Publisher, Inc, New York.
[16] De Buyser, M.L., Morvan, A., Aubert, S., Dilasser, F. and El
Sohl, N. (1992) Evaluation of ribosomal RNA gene probe for
identification of species and subspecies within the genus Staphy-
lococcus. J. Gen. Microbiol. 138, 889–899.
[17] Izard, N.C., H€achler, Grehn, M. and Kayser, F.H. (1992)
Ribotyping of coagulase-negative staphylococci with special
emphasis on intraspecific typing of Staphylococcus epidermidis. J.
Clin. Microbiol. 30, 817–823.
[18] Hesselbarth, J. and Schwarz, S. (1995) Comparative ribotyping of
Staphylococcus intermedius from dogs, pigeons, horses and mink.
Vet. Microbiol. 45, 11–17.
[19] Hunter, P.R. and Gaston, M.A. (1988) Numerical index of
discriminatory ability of typing systems: an application of
Simpson’s index of diversity. J. Clin. Microbiol. 26, 2465–2466.
[20] Back, N.A., Linnemann, C.C., Pfaller, M.A., Staneck, J.L. and
Morthland, V. (1993) Recurrent epidemics caused by a single
strain of erythromycin-resistant Staphylococcus aureus: the im-
portance of molecular epidemiology. JAMA 270, 1329–1333.
[21] Diekema, D.J., Pfaller, M.A., Schmitz, F.J., Turnidge, J., Verhoef,
J., Bell, J., Fluit, A.C., Doern, G.V., Jones, R.N. and the Sentry
Participants Group (2000) Genetic relatedness of multi-drug
resistant, methicillin (oxacillin)-resistant Staphylococcus aureus
bloodstream isolates from SENTRY antimicrobial resistance
surveillance centres world-wide. Microbiol. Drug Resist. 6, 213–
221.
[22] Diekema, D.J., Pfaller, M.A., Schmitz, F.J., Smayevsky, J., Bell,
J., Jones, R.N., Beach, M. and the Sentry Participants Group
(2001) Survey of infections due to Staphylococcus species:
frequency of occurrence and antimicrobial susceptibility of
isolates collected in the United States, Canada, Latin America,
Europe, and the Western Pacific for the SENTRY antimicro-
bial surveillance program, 1997–1999. Clin. Infect. Dis. 32,
S114–S132.
[23] Struelens, M.J. (1996) Consensus guidelines for appropriate use
and evaluation of microbial epidemiologic typing systems. Clin.
Microbiol. Infect. 2, 2–11.
[24] Wada, A., Ohta, H., Kulthanan, K. and Hiramatsu, K. (1993)
Molecular cloning and mapping of 16S-23S rRNA gene complexes
of Staphylococcus aureus. J. Bacteriol. 175, 7483–7487.
[25] Klappenbach, J.A., Dunbar, J.M. and Schmidt, T.M. (2000)
rRNA operon copy number reflects ecological strategies of
bacteria. Appl. Environm. Microbiol. 66, 1328–1333.
[26] Hollis, R.J., Bruce, J.L., Fritschel, S.J. and Pfaller, M.A. (1999)
Comparative evaluation of an automated ribotyping instrument
versus pulsed field gel electrophoresis for epidemiological inves-
tigation of clinical isolates of bacteria. Diagn. Microbiol. Infect.
Dis. 34, 263–268.
[27] Thomson-Carter, F.M., Carter, P.E. and Pennington, T.H. (1989)
Differentiation of staphylococcal species and strains by ribosomal
RNA gene restriction patterns. J. Gen. Microbiol. 135, 2093–2097.
[28] Olive, D.M. and Bean, P. (1999) Principle and applications of
methods for DNA-based typing of microbial organisms. J. Clin.
Microbiol. 37, 1661–1669.
[29] Wiedmann, M., Weilmeier, D., Dineen, S.S., Ralyea, R. and Boor,
K.J. (2000) Molecular and phenotypic characterization of Pseu-
domonas spp. isolated from milk. Appl. Environ. Microbiol. 66,
2085–2095.




















