
Hypoxia-responsive growth factors upregulate periostin and osteopontin expression via distinct...
31
Hypoxia-Responsive Growth Factors Upregulate Periostin and Osteopontin Expression via Distinct Signaling Pathways in Rat Pulmonary Arterial Smooth Muscle Cells Peng Li, Suzanne Oparil, Wenguang Feng, Yiu-Fai Chen Vascular Biology and Hypertension Program Division of Cardiovascular Disease Department of Medicine Birmingham, Alabama 35294 Running Head: Regulation of Periostin Expression in Pulmonary VSMCs Word counts: Abastract: 250 Text: 5551 Figures: 8 Correspondence to: Yiu-Fai Chen, Ph.D. 1008 Zeigler Research Building University of Alabama at Birmingham UAB Station Birmingham, Alabama 35294 (205) 934-2580 (Phone) (205) 934-0424 (Fax) E-mail: [email protected] Articles in PresS. J Appl Physiol (April 30, 2004). 10.1152/japplphysiol.01311.2003 Copyright © 2004 by the American Physiological Society.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Hypoxia-responsive growth factors upregulate periostin and osteopontin expression via distinct...

Hypoxia-Responsive Growth Factors Upregulate Periostin and Osteopontin Expression
via Distinct Signaling Pathways in Rat Pulmonary Arterial Smooth Muscle Cells
Peng Li, Suzanne Oparil, Wenguang Feng, Yiu-Fai Chen
Vascular Biology and Hypertension Program
Division of Cardiovascular Disease
Department of Medicine
Birmingham, Alabama 35294
Running Head: Regulation of Periostin Expression in Pulmonary VSMCs
Word counts: Abastract: 250
Text: 5551
Figures: 8
Correspondence to: Yiu-Fai Chen, Ph.D.
1008 Zeigler Research Building
University of Alabama at Birmingham
UAB Station
Birmingham, Alabama 35294
(205) 934-2580 (Phone)
(205) 934-0424 (Fax)
E-mail: [email protected]
Articles in PresS. J Appl Physiol (April 30, 2004). 10.1152/japplphysiol.01311.2003
Copyright © 2004 by the American Physiological Society.

1
ABSTRACT
This study tested the hypothesis that expression of the novel adhesion molecule periostin
(PN) and osteopontin (OPN), are increased in lung and in isolated pulmonary arterial smooth
muscle cells (PASMCs) in response to the stress of hypoxia and explored the signaling pathways
involved. Adult male rats were exposed to 10% O2 for 2 wks; and growth-arrested rat PASMCs
were incubated under 1% O2 for 24 hrs. Hypoxia increased PN and OPN mRNA expression in
rat lung. In PASMCs, hypoxia increased PN, but not OPN expression. The hypoxia-responsive
growth factors fibroblast growth factor-1 (FGF-1) and angiotensin II (ANG II) caused dose- and
time-dependent increases in PN and OPN expression in PASMCs. FGF-1-induced PN
expression was blocked by the FGFR-1 antagonist PD166866 and by inhibitors of PI3K
(LY294002, Wortmannin), p70S6K (rapamycin), MEK1/2 (U0126, PD98059) or p38MAPK
(SB203580), but not of JNK (SP600125). ANG II-induced PN expression was blocked by the AT1
receptor antagonist losartan and by inhibitors of PI3K and MEK1/2. In contrast, FGF-1-induced
OPN expression was blocked by inhibitors of JNK or MEK1/2, but not of PI3K, p70S6K or
p38MAPK. Activation of p70S6K and p38MAPK by anisomycin robustly stimulated PN, but not
OPN expression. This study is the first to demonstrate that growth factor-induced expression of
PN in PASMCs is mediated through PI3K/p70S6K, Ras/MEK1/2 and Ras/p38MAPK signaling
pathways, while the expression of OPN is mediated through Ras/MEK1/2 and Ras/JNK signaling
pathways. These differences in signaling suggest that PN and OPN may play different roles in
pulmonary vascular remodeling under pathophysiologic conditions.
Keywords: Pulmonary vascular remodeling, Pulmonary arterial smooth muscle cells, Periostin,
Osteopontin, Hypoxia

2
INTRODUCTION
We have recently reported that expression of periostin (PN, a putative bone extracellular
matrix [ECM] protein previously named osteoblast-specific-factor 2, OSF-2) (5,6,10,18,23) is
greatly increased during remodelling of the cardiovascular system under stressful conditions, e.g.
pressure overload-induced cardiac hypertrophy and balloon-induced endoluminal injury of carotid
arteries (16,46). PN mRNA expression was increased 12- to 43-fold in hearts of wildtype and
atrial natriuretic peptide-deficient mice, respectively, with transverse aortic constriction
(TAC)-induced cardiac hypertrophy. This was the most dramatic response to TAC-induced
pressure overload of any of > 5,000 genes studied by microarray analysis (26).
Immunohistochemical analysis showed that PN is preferentially localized to the cardiac
fibroblasts, interstitium, and vascular smooth muscle cells (VSMCs) in these experimental
settings. Importantly, increased PN expression was associated with increased expression of
osteopontin (OPN), collagens, fibronectin, thrombospondin and MMPs (11,26), suggesting that
PN may participate in ECM remodelling in heart and blood vessels under stressful conditions (26).
The present study expanded these important findings to examine the regulation of PN and
OPN, in lungs of hypoxia-adapted rats and pulmonary arterial SMCs (PASMCs) exposed to
hypoxia or hypoxia-responsive growth factors in vitro. Based on our findings in the heart and our
previous experience with pulmonary vascular remodelling in the lung of hypoxia-adapted rats, we
postulated that PN and OPN expression would increase as part of a generalized ECM response to
hypoxic stress in the lung of adult rats and in PASMCs in vitro. Accordingly, this study tested the
primary hypothesis that PN and OPN are increased in lung and in isolated PASMCs in response
to normobaric hypoxia exposure. We further tested whether hypoxia-induced growth factors,
including the tyrosine kinase receptor-associated growth factor fibroblast growth factor-1 (FGF-1)

3
(13) and the G-protein-coupled receptor-associated growth factor angiotensin II (ANG II) (19)
stimulate increased expression of ECM molecules in PASMCs. Finally, we used selective kinase
inhibitors to delineate the signaling pathways involved in FGF-1 and ANG II-induced PN and OPN
expression in PASMCs.
MATERIALS AND METHODS
Animals and cell culture
Male Sprague-Dawley rats were obtained from Charles River Breeding Laboratories
(Wilmington, MA) at age 8 wks. All animals used in this study were cared for and handled
according to the NIH Guide for the Care and Use of Laboratory Animals. For in vivo chronic
hypoxia studies, rats were exposed to hypoxia (10% O2, 1 atm) in an 800-liter model 818GBB
Plexiglas glove box (Plas Labs, Lansing, MI) beginning at age 8-9 wk for a total of 2 wks as
previously described (3,4,21).
For in vitro studies, PASMC were initially grown in DMEM (GIBCO BRL, Rockville, MD)
with 10% heat-inactivated low endotoxin fetal bovine serum (FBS) (GIBCO BRL), 2 mM
glutamine, 100 U/ml penicillin, and 100 µg/ml streptomycin at 37oC in a humidified atmosphere
containing 95% air-5% CO2 and were passaged by harvesting with trypsin-EDTA and seeding in
6-cm cell culture dishes. Passaged cells were grown in DMEM with 10% FBS. PASMCs were
used for experiments between passage 4 and 8. To confirm the characteristics of SMCs in
culture, immunohistochemical staining of -actin was performed using specific -actin antibody
and peroxidase-labeled anti-IgM or anti-IgG antibodies.
Prior to each study, PASMCs were grown to 100% confluence and then made quiescent
by placing them in medium containing 0.1% FBS for 24 hrs. To examine the effects of hypoxia on
PN and OPN mRNA expression, PASMCs were transferred into an air-tight hypoxic chamber
(model 2710, Cell Culture Incubator, Queue Systems) containing 1%O2/5%CO2/94%N2 as

4
previously described (13,22). For the study of signal transduction pathways, PASMCs were
pretreated with selective inhibitors for 45 min before addition of growth factors (FGF-1 or ANG II)
to the medium and incubation for an additional 24 hrs. For treatment with FGF-1, heparin was
added to the medium at a final concentration of 5 U/ml, because it protects FGF-1 against
enzymatic digestion and promotes FGF receptor binding (28).
Immunohistochemical Analysis of Periostin in lung
At 2 wks after initiation of hypoxic exposure, the lungs from hypoxia adapted and air
control rats were fixed in the distended state by infusion of 10% buffered formalin into the
pulmonary artery and trachea at 100 and 25 mmHg pressure, respectively, for 1 min, and then
placed in a bath of 4% paraformaldhyde for 24 hrs, paraffin embedded, and sectioned for
immunohistochemical examination of PN as previously described (4,26).
RNA extraction and Northern blot analysis
Lungs from 2-wk hypoxia adapted or air control rats or cultured PASMCs were
homogenized, and total RNA was extracted using the TRIZOL total RNA isolation reagent
(GibcoBRL, Life Technologies). Northern analysis was performed using a 32P-labeled selective
cDNA probes for PN and OPN which had been generated in our laboratory by reverse
transcription (RT) followed by the DNA polymerase chain reaction (PCR) using lung RNA as the
template, as previously described (13). A 32P-labeled 18S rRNA-oligonucleotide
(5=-ACGGTATCTGATCGTCTTCGAACC-3=) was used as the control probe to normalize data.
Autoradiographic signals were scanned with an optical densitometer (Model GS-670 Imaging
Densitometer, Bio-Rad, Hercules, CA). To estimate steady-state specific mRNA levels, PN and
OPN mRNA/18S rRNA ratios were determined by dividing the absorbance corresponding to the
specific cDNA probe hybridization by the absorbance corresponding to the 18S rRNA probe
hybridization. The results were expressed as the ratios of specific mRNA to 18S rRNA.

5
Western blot analysis for Akt, GSK3, MEK1/2 and ERK1/2
To evaluate the effects of FGF-1 and ANG II on activities of the
phosphatidylinositol-3-kinase (PI3K)/Akt and the Ras/mitogen activated protein kinase
(MAPK)/extracellular signal-regulated kinase 1/2 (ERK1/2) pathways, as well as selectivity of the
PI3K inhibitor LY294002 and the ERK kinase 1/2 (MEK1/2) inhibitor U0126, Western blot analysis
for phosphorylated Akt and GSK3 (an Akt substrate), as well as phosphorylated MEK1/2 and
ERK1/2 (p44/p42 MAPK), was performed using the PhosphoPlus Akt Antibody Kit and
Phospho-ERK1/2 Pathway Sample Kit (Cell Signaling Technology Inc., Beverly, MD),
respectively, as previously described (13). PASMCs were made quiescent by placing them in
medium containing 0.1% FBS for 24 hrs. For the study of signal transduction pathways, PASMCs
were pretreated with LY294002 (10 uM) or U0126 (5 uM) for 45 min before addition of FGF-1 (10
ng/ml) or ANG II (100 nM) to the medium and incubation for an additional 15 min.
After treatment with FGF-1 or ANG II, cells were lysed by the addition of 0.5 ml of ice-cold
lysis buffer containing (mM) NaCl 50, NaF 50, sodium pyrophosphate 50, EDTA 5, EGTA 5,
Na3VO4 2, phenylmethylsulfonyl fluoride 0.5, and HEPES 10 at pH 7.4 along with 0.1% Triton
X-100 and 10 ug/ml leupeptin, followed by immediate freezing on ethanol/dry ice. The cell lysates
were thawed on ice, scraped, sonicated, and centrifuged at 14,000x g at 4oC for 30 min. Cell
lysates (25 µg protein) were subjected to electrophoresis on a 10% SDS-polyacrylamide gel and
transferred to nitrocellulose membranes. The membranes were blocked for 2 hrs in 1% casein
(Hammarsten-prepared), 0.05% Tween, and 0.05% azide in PBS. Western blot analysis was
performed using anti-phospho-AKT (Ser473), anti-phospho-GSK3, anti-phospho-MEK1/2
(Ser217/221), anti-phospho-ERK1/2 (Thr202/Tyr204)-specific primary antibodies and a
horseradish peroxidase-conjugated goat anti-rabbit IgG. Immune complexes were detected by

6
Phototop-HRP Western Detection Kit (Cell Signaling Technology Inc.). Autoradiograms exposed
in the linear range of film density were scanned using a densitometer (Model GS-670 Imaging
Densitometer, Bio-Rad).
Reagents
ANG II and the ANG II type 1 (AT-1) receptor antagonist losartan, human recombinant
FGF-1 and transcriptional inhibitor actinomycin D were purchased from Sigma Inc. (St. Louis,
MO). Selective rabbit anti-PN polyclonal antibody was provided by Drs. Lan Bo Chen and Meiru
Dai at Harvard Medical School (26). The FGF-1 receptor tyrosine kinase inhibitor PD-166866 was
provided by Parke-Davis Pharmaceutical Research, Division of Pfizer (17). The p38MAP and
p70S6 kinase activator anisomycin, the MEK1/2 inhibitors U0126 and PD98059, the PI3K
inhibitors LY-294002 and wortmannin, the p38MAP kinase inhibitor SB203580, the p70S6 kinase
inhibitor rapamycin, and the JNK (c-Jun NH2-terminal kinase) inhibitor SP600125 were purchased
from Calbiochem-Novabiochem Corp. (San Diego, CA).
Statistical analysis
Results were expressed as means ± SEM. Statistical analyses were carried out using the
SigmaStat package (Jandel Scientific Software, San Rafael, CA) on a PC. Statistical
comparisons of mRNA levels were performed with the one-way analysis of variance (ANOVA) or
unpaired t-test. If ANOVA results were significant, a post-hoc comparison among groups was
performed with the Newman-Keuls test. Differences were reported as significant if the P value
was < 0.05.
RESULTS
Effects of hypoxia on PN, and OPN expression in lungs and PASMCs
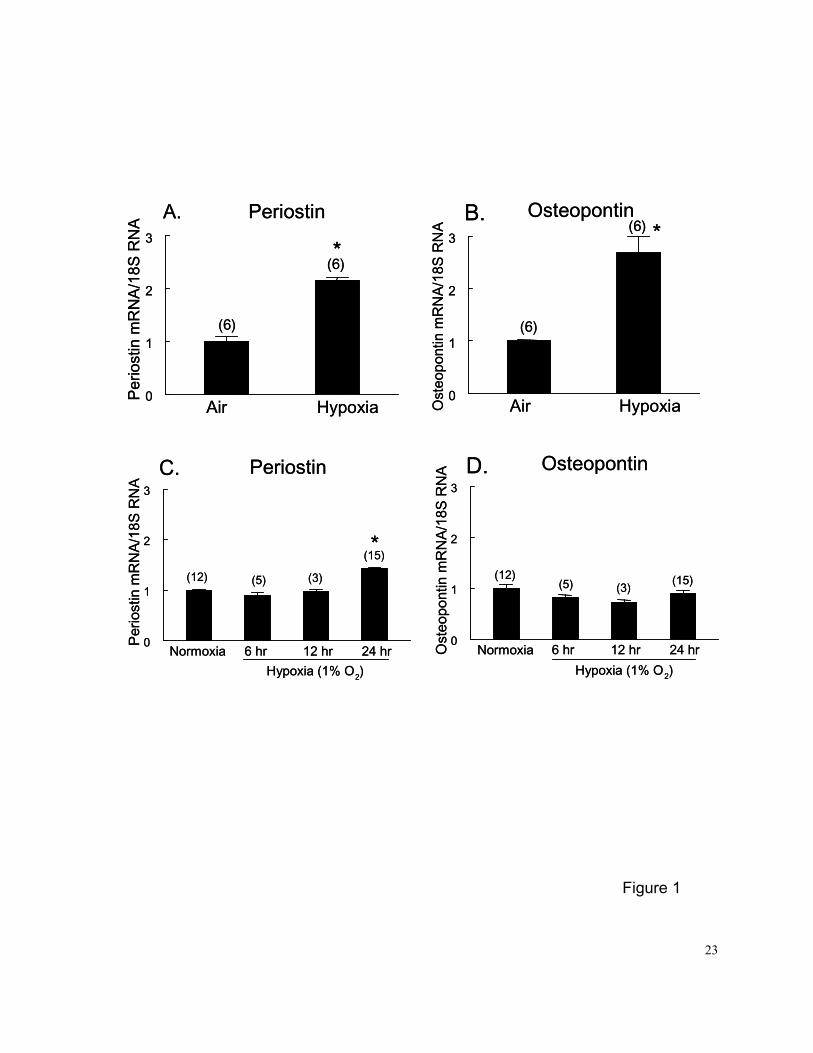
Chronic normobaric hypoxic exposure for 2 wks significantly increased steady state PN (a
2.2-fold increase) and OPN (a 2.7-fold increase) mRNA levels in rat lung (Figures 1A and 1B).

7
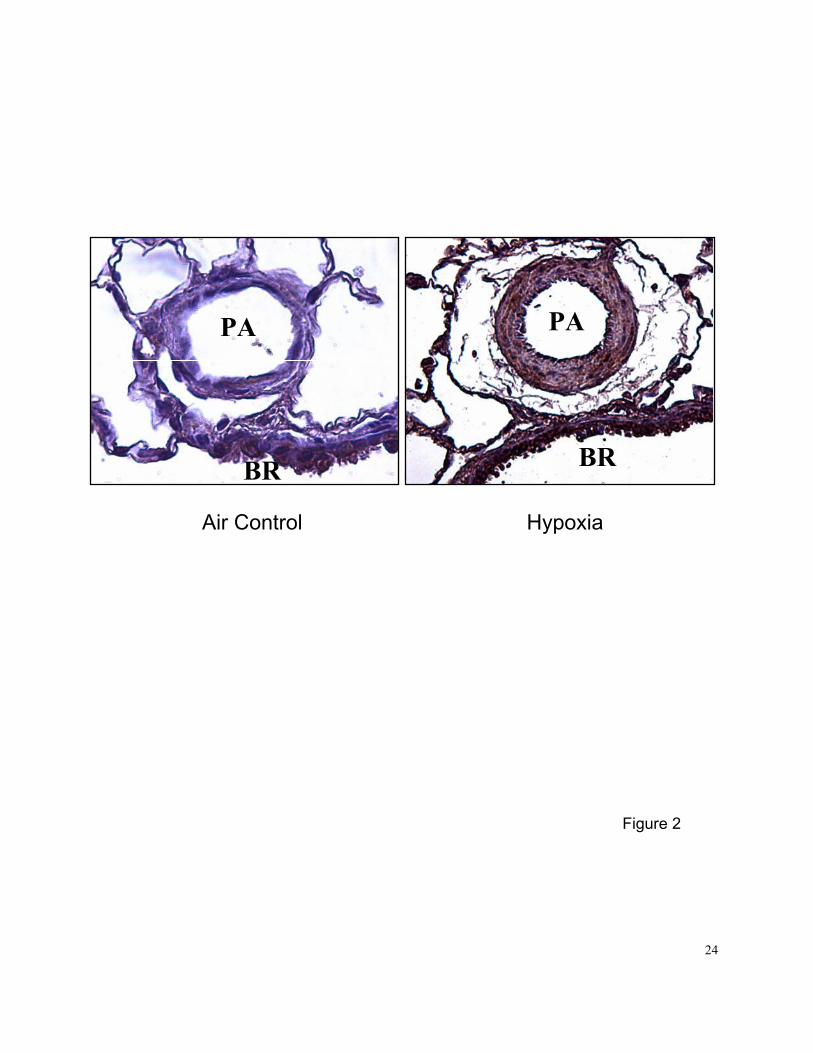
Immunohistochemical analysis showed that 2-wk exposure to normobaric hypoxia increased PN
expression in pulmonary artery and bronchi of hypoxia-adapted rats compared with air controls
(Figure 2). PN was mainly expressed (brown stain) in medial SMCs and bronchial epithelial cells.
Exposure of growth-arrested PASMCs to 1% O2 (pO2=20-30 mmHg in culture media) for 6, 12 and
24 hrs resulted in a slight but significant increase in PN (a 40% increase) mRNA levels at 24 hrs,
but had no effect on OPN (Figures 1C and 1D). The effect of hypoxia on PN mRNA expression in
PASMCs in vitro was much less robust than that in intact lung in vivo. No significant cell death
(assessed by trypan blue [0.4%] exclusion) was observed in PASMCs exposed to hypoxia or
normoxia. These results suggest that other mediators, such as hypoxia-responsive growth
factors, may be needed to increase expression of PN and OPN in isolated PASMCs in vitro.
Dose- and time- dependent effects of FGF-1 and ANG II on PN and OPN expression in
PASMCs
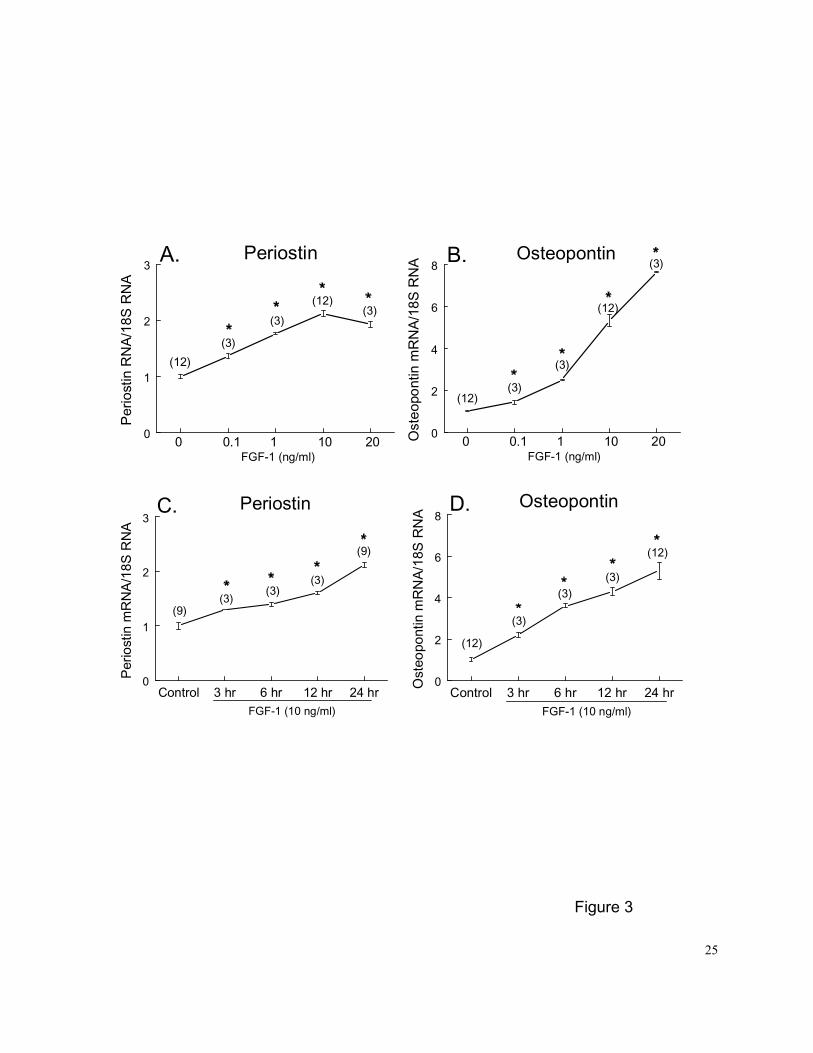
Quantitative Northern blot analysis demonstrated that PN and OPN mRNA expression
increased in a dose-dependent fashion (range 0.1-20 ng/ml) in PASMCs treated with FGF-1 for
24 hrs (Figures 3A and 3B). The threshold concentration was 0.1 ng/ml (~ 6.7 pM), and the
largest effect was observed at doses of 10 and 20 ng/ml for PN (a 2.1-fold increase) and OPN (a
7.6-fold increase), respectively. The time course for enhancement of PN and OPN expression
with FGF-1 was examined at a concentration of 10 ng/ml. Significant increases in PN and OPN
mRNA levels in PASMCs were observed after 3 hrs of FGF-1 treatment (Figures 3C and 3D). PN
and OPN mRNA levels increased progressively over the 24 hr period of FGF-1 exposure.
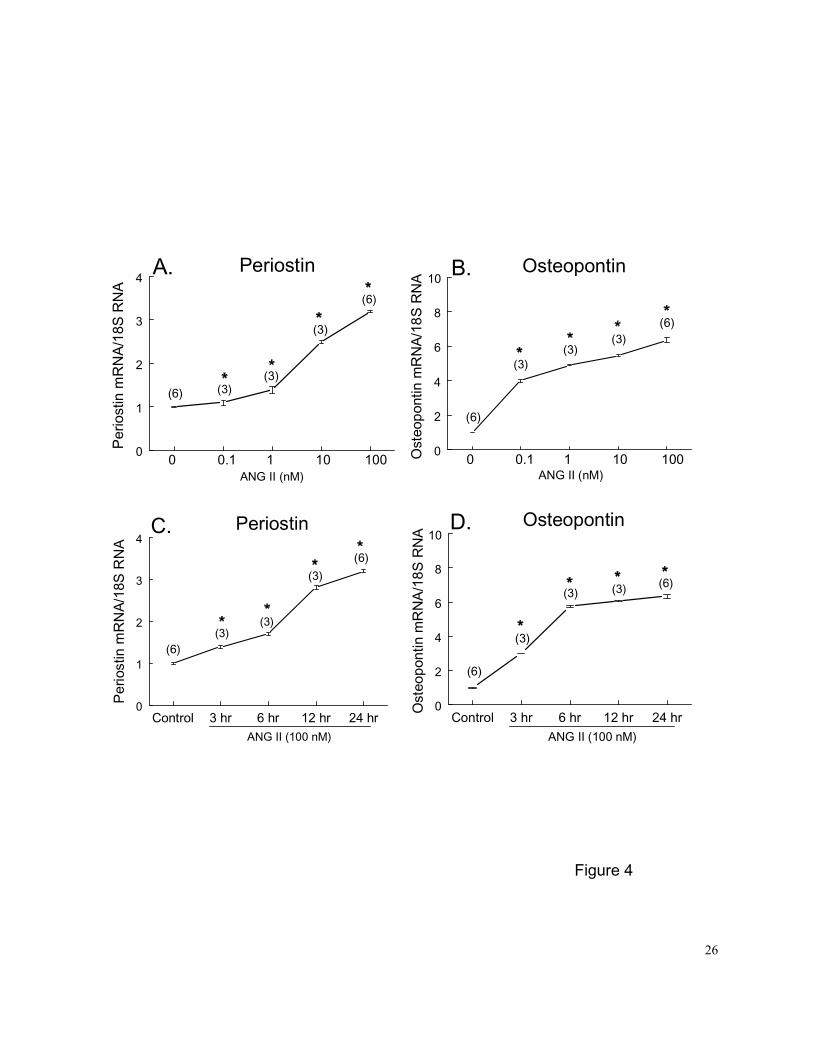
Similarly, PN and OPN mRNA expression increased in a dose-dependent fashion (range
0.1-100 nM, or 0.1-100 ng/ml) in PASMCs treated with ANG II for 24 hrs (Figures 4A and 4B). The
threshold concentration was 0.1 nM (or 0.1 ng/ml), and the largest effect (a 3.2-fold increase of
PN and a 6.5-fold increase of OPN) was observed at the maximal dose of 100 nM. The time

8
course for enhancement of PN and OPN gene expression with ANG II was examined at a
concentration of 100 nM. Significant increases in PN and OPN mRNA levels in PASMCs were
observed after 3 hrs of ANG II treatment (Figures 4C and 4D).
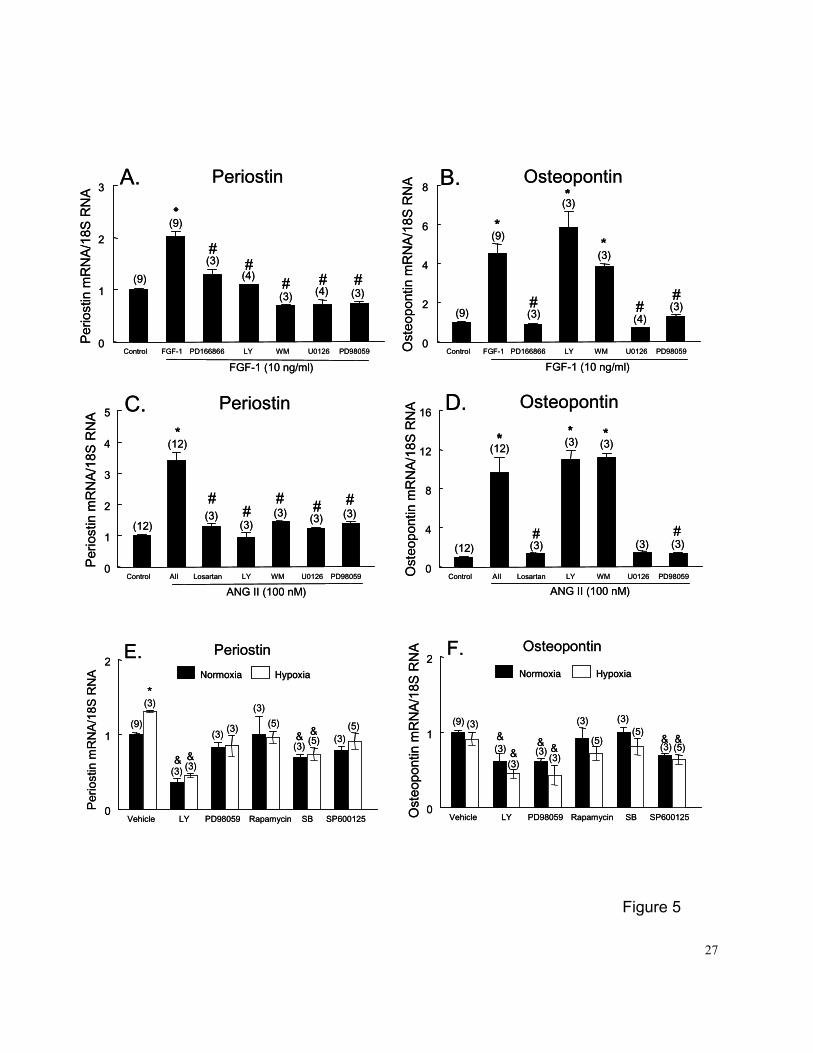
Effects of receptor inhibition on FGF-1 or ANG II-induced upregulation of PN and OPN
expression
PASMCs were pretreated with PD-166866 (5 µM), a potent and selective inhibitor of
FGF-1 receptor tyrosine kinase (17), or losartan (10 mM), a selective ANG II AT1 receptor
antagonist, for 45 min before FGF-1 (10 ng/ml) or ANG II (100 nM) was added to the medium for
an additional 24 hrs of incubation. PD-166866 or losartan completely blocked FGF-1 or ANG
II-induced upregulation of PN and OPN mRNA expression (Figures 5A-5D), indicating the
receptor dependence of growth factor stimulation. FGF-1-induced upregulation of PN and OPN
mRNA expression was also completely blocked by pretreatment of PASMCs with actinomycin D
(5 uM) (data not shown), indicating that transcription is essential to FGF-1-stimulated PN and
OPN mRNA expression.
Effects of signal transduction pathway inhibitors on FGF-1 or ANG II-induced upregulation
of PN and OPN expression
PASMCs were pre-treated with inhibitors for 45 min before FGF-1 (10 ng/ml) or ANG II
(100 nM) was added to the medium for an additional 24 hrs of incubation. LY294002 (5 uM) or
wortmannin (20 µM), PI3K inhibitors, as well as U0126 (5 uM) or PD98058 (20 uM), MEK1/2
inhibitors, completely blocked the FGF-1 or ANG II induced upregulation of PN mRNA expression
(Figures 5A and 5C). The MEK1/2 inhibitors U0126 (5 uM) or PD98059 (20 uM) also completely
blocked FGF-1 or ANG II-induced upregulation of OPN mRNA expression (Figures 5B and 5D).
In contrast, the PI3K inhibitors LY294002 (5 uM) or wortmannin (20 uM) did not alter the
stimulatory effect of FGF-1 or ANG II on OPN mRNA expression.

9
The effects of signaling pathway inhibitors on PN and OPN mRNA expression in PASMCs
exposed to normoxia (control) or hypoxia for 24 hrs are shown in Figures 5E and 5F. PASMCs
were pretreated with inhibitors for 45 min before exposing them to hypoxia (1% O2) for an
additional 24 hrs. Inhibition of PI3K (by LY294002) and p38MAP kinase (by SB203580)
decreased PN mRNA expression; and inhibition of PI3K (by LY294002), MEK1/2 (by PD98059)
and JNK (by SP600125) decreased OPN mRNA expression in PASMCs exposed to either
normoxia or hypoxia.
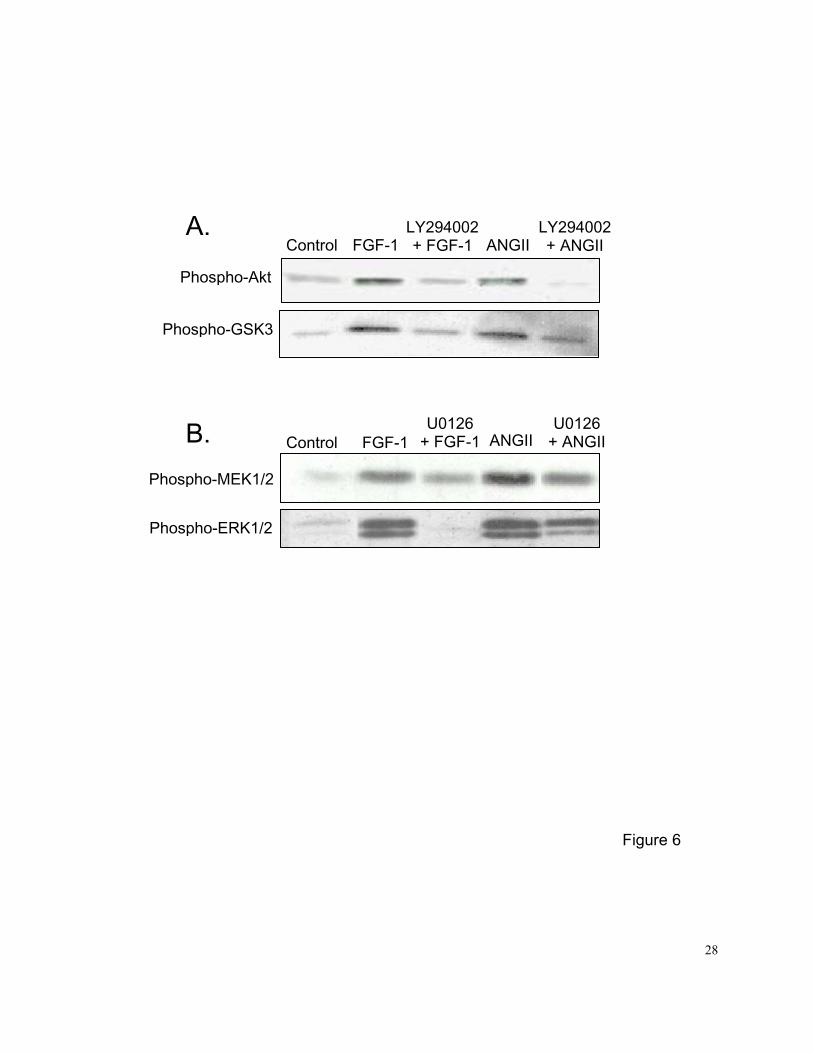
Western blot analysis confirmed that FGF-1 or ANG II significantly increased activation of
PI3K and MEK1/2 within 15 min in PASMCs. The FGF-1 or ANG II-stimulated phosphorylation of
Akt and GSK3 was blocked by pretreatment with the selective PI3K inhibitor LY294002 (Figure
6A), and the FGF-1 or ANG II-stimulated phosphorylation of ERK1/2 was blocked by pretreatment
with the selective MEK1/2 inhibitor U0126 (Figure 6B). LY294002 did not block FGF-1 or ANG
II-stimulated phosphorylation of ERK1/2 (data not shown) and U0126 did not block FGF-1 or ANG
II-stimulated phosphorylation of Akt (data not shown) or MEK1/2 (Figure 6B), indicating the
selectivity of these signaling pathway inhibitors.
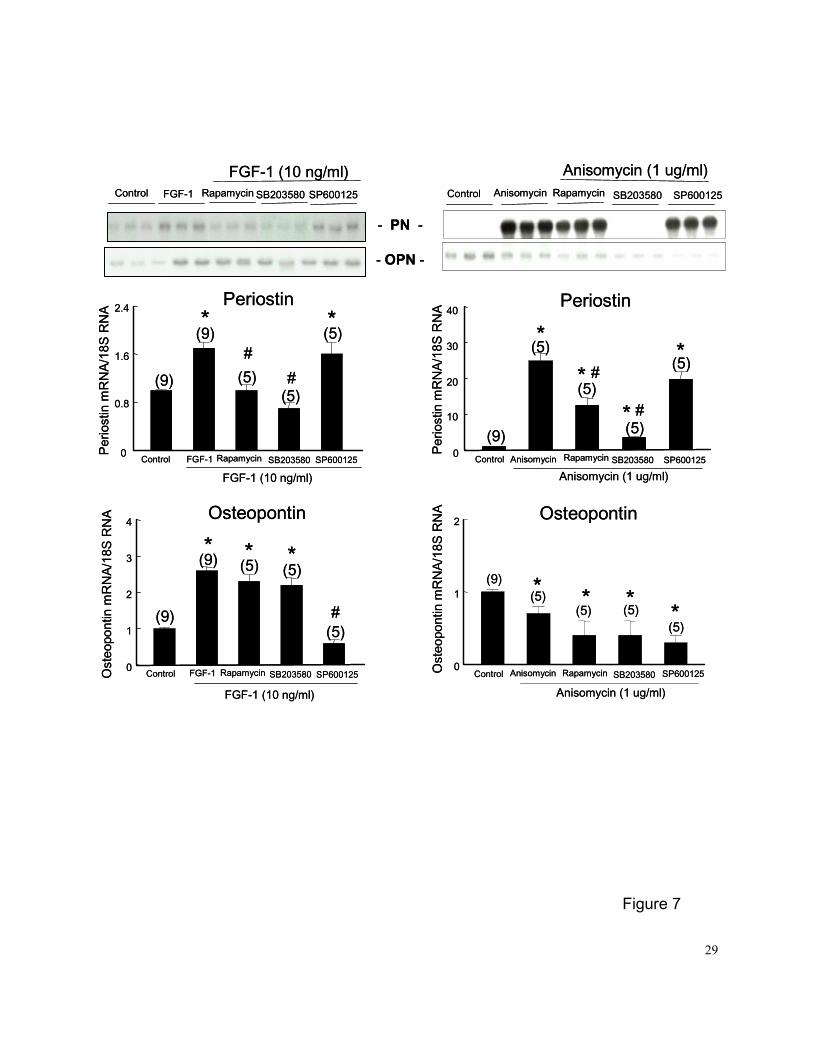
Effects of p70S6K and p38MAPK activation on PN and OPN expression in PASMCs
Quiescent PASMCs were treated with FGF-1 (10 ng/ml) or the p38MAPK and p70S6K
activator anisomycin (1 ug/ml) for 24 hrs with or without pretreatment (45 min) with selective
inhibitors of intracellular signaling, including rapamycin (100 nM, a p70S6K inhibitor), SB203580
(10 uM, a p38MAPK inhibitor), or SP600125 (20 uM, a JNK inhibitor). Both FGF-1 and
anisomysin induced PN expression were blocked by p70S6K and p38MAPK inhibitors, but not by
the JNK inhibitor (Figures 7A and 7B). In contrast, the FGF-1 induced OPN expression was
blocked by the JNK inhibitor, but not by p70S6K and p38 MAPK inhibitors (Figure 7C).
Anisomycin alone slightly but significantly decreased OPN expression in PASMCs, and

10
rapamycin, SB203580 and SP600125 did not alter the effects of anisomycin on OPN expression
in PASMCs (Figure 7D).
DISCUSSION
The present study is the first to demonstrate increased expression of PN in lung and
pulmonary arteries of rats adapted to hypoxia as part of a generalized ECM response to hypoxic
stress, which also included increased OPN mRNA expression. We have previously reported that
hypoxia induces acute pulmonary vasoconstriction, chronic pulmonary hypertension, and
pulmonary vascular remodeling characterized by medial hypertrophy and marked increased
medial-wall thickness of small pulmonary arteries (50-100 um) in the rat (3,4). The present results
demonstrate localization of hypoxia-induced PN expression to PASMCs of hypertrophic
pulmonary arteries. This finding, coupled with the observation that PN expression increases in
parallel with OPN in lung of hypoxia adapted rats, suggests an involvement of PN in ECM
formation and pulmonary vascular hypertrophy and remodeling in response to chronic hypoxia
exposure.
Another novel finding of present study is the demonstration that the hypoxia-responsive
growth factors FGF-1 and ANG II stimulate PN expression in PASMCs in vitro. The rapid
increases in steady-state PN mRNA levels observed in response to low concentrations of FGF-1
and ANG II suggest that these growth factors, in addition to hypoxia per se, could be endogenous
physiological stimulators of PN gene expression in PASMCs. The FGFs (at least isoforms FGF-1,
-2, and -7) and ANG II are expressed in lung and play important roles in diverse aspects of
pulmonary development and growth, including epithelial cell, VSMC, and myofibroblast
proliferation, differentiation, angiogenesis and vascular constriction (14,19). It has been
hypothesized that these growth factors may contribute to hyperproliferation of PASMCs and

11
muscularization of the pulmonary vasculature in hypoxia-adapted animals (14,19).
OPN is abundantly expressed in a range of lung diseases (16,25). Bronchial epithelial
cells and alveolar macrophages in normal lung express OPN (2), and its expression is greatly
amplified in epithelium, alveolar and interstitial macrophages, fibroblasts, and endothelial cells
and VSMCs in injured and diseased lungs (16). Microarray analysis (7) has demonstrated
increased OPN mRNA expression in the lungs of rats and mice exposed to chronic hypobaric
hypoxia for 1 and 3 wks. These data provide indirect evidence that OPN may play a role in the
pathogenesis of pulmonary vascular disease.
The current findings extend previous observations of the effects of hypoxia exposure on
OPN expression in vivo and in vitro by elucidating a role for hypoxia-responsive growth factors on
this process. While chronic hypoxia exposure per se increased OPN gene expression in lung of
hypoxia adapted rats, FGF-1 and ANG II, but not hypoxia per se, increased OPN mRNA
expression in PASMCs in vitro. This result is similar to our previous findings that ANP clearance
receptor (NPR-C) expression is downregulated in the lung of rats and mice exposed to hypoxia
but not in PASMCs cultured under hypoxic conditions (22). The absence of a direct effect of
hypoxia on OPN mRNA levels in PASMCs in vitro differs from the result of Sodhi et al. (20), who
found a biphasic increase in OPN mRNA and protein levels with exposure to 3% O2 (maximal at
2 hrs, no effect at 6 and 12 hrs, and slight increase at 24 hrs) in rat mesangial cells and aortic
SMCs in vitro. In the current study, we did not sample time points earlier than 6 hrs, but found no
effect of hypoxia (1% O2) on OPN expression in PASMCs at 6, 12 or 24 hrs. Apparent differences
between these results may relate to differences in tissues of origin of the SMCs (kidney/aorta vs.
lung), duration of hypoxia exposure or the severity of hypoxia. The observations that growth
factors time and dose-dependently induced increases in OPN mRNA expression in PASMCs in
vitro and that FGF-1, FGF-2 (13), and ANG II (19) expression increased in lung of rats adapted to

12
chronic hypoxia provide additional evidence for a functional role for hypoxia-responsive growth
factors in stimulating ECM formation and remodeling in the hypoxic lung.
FGF-1 binds to any of 4 receptor isoforms (FGFR-1, -2, -3, and -4), and ANG II binds to
at least 2 ANG receptors (AT-1 and AT-2). FGFR-1 and AT-1 are the predominant receptor
isoforms in adult VSMCs (1,27). PD 166866 is a potent (nanomolar range) and highly selective
small molecule inhibitor of FGFR-1-dependent signaling that does not perturb signals induced by
PDGFR, EGFR, Src, MEK, PKC, or CDK4 (17), and losartan is highly potent and selective
non-peptide AT-1 receptor antagonist. The present study demonstrated that PD166866 or
losartan effectively inhibited PASMC expression of PN and OPN mRNA following FGF-1 or ANG
II treatment. This result indicates that FGF-1 or ANG II stimulation of PN or OPN mRNA
expression in PASMCs in vitro is mediated in an FGFR-1 or AT-1-dependent manner.
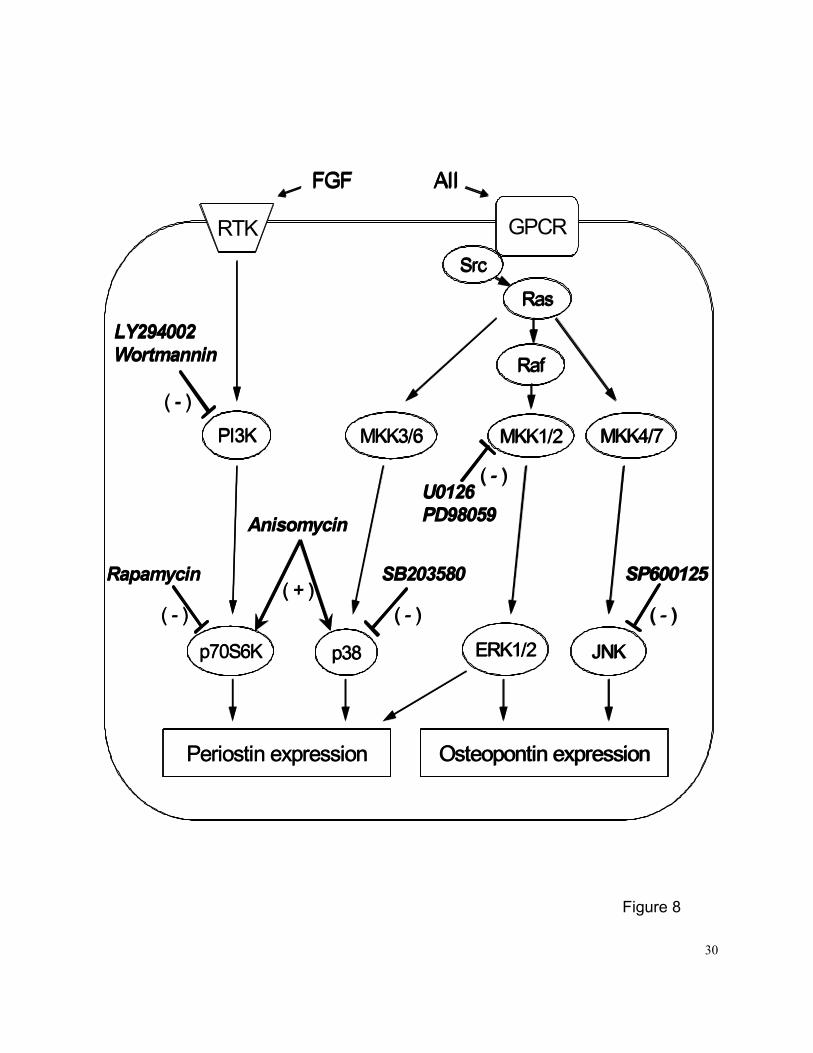
Multiple intracellular signaling pathways are activated by FGFs and ANG II (8,9) (Figure
8). Activation of the PI3K-mediated Akt and p706K, and Ras-mediated MEK1/2/ERK1/2,
p38MAPK, and JNK pathways are necessary for the mitogenic activity of FGFs and ANG II in
VSMCs (8,15,24). In this study, we investigated the involvement of PI3K-mediated and
Ras-mediated signaling pathways in the regulation of PN expression in PASMCs. Inhibition of
PI3K by LY294002 or wortmannin, p70S6K by rapamycin, MEK1/2 by U0128 or PD98059,
p38MAPK by SB203580, significantly inhibited FGF-1 or ANG II-induced PN mRNA expression.
However, inhibition of JNK by SP600125 had no effect on PN mRNA expression under FGF-1 or
anisomycin-treated conditions. These results suggest that the PI3K/p70S6K,
Ras/MEK1/2/ERK1/2 and Ras/p38MAPK pathways, but not the Ras/JNK pathway, are involved in
FGF-1 or ANG II-stimulated PN mRNA upregulation in PASMCs (Figure 8).
This study elucidates for the first time the signaling pathways by which FGF-1 and ANG
II stimulate OPN mRNA expression in PASMCs (Figure 8). We observed that FGF-1 and ANG II

13
potently and time- and dose-dependently stimulate expression of OPN mRNA in PASMCs in vitro
through the activation of the Ras/MEK1/2/ERK1/2 and Ras/JNK, but not the PI3K or p38MAPK
signaling pathways. These results are consistent with our previous observation that FGF
ligand-activated FGFR-1 plays a significant role in upregulation of OPN gene expression at the
transcriptional level via activation of FGFR-1 and subsequent activation of the
Src/Ras/MEK/MAPK pathway, but not the PI3K and p38MAPK pathways, in rat aortic SMCs in
vitro (12).
In summary, the present study provides the first evidence that the PN mRNA expression
is increased in lung of hypoxia-adapted animals and in cultured PASMCs exposed to hypoxia, and
that the hypoxia-responsive growth factors FGF-1 and ANG II play significant roles in upregulation
of PN and OPN expression at the transcriptional level via activation of FGFR-1 and AT-1
receptors, respectively. We also demonstrated that activation of the PI3K/Akt/p70S6K,
Ras/MEK1/2/ERK1/2 and Ras/p38MAPK signaling pathways, but not the Ras/JNK pathway, by
FGF-1 or ANG II increases PN gene expression in PASMCs in vitro. In contrast, the FGF-1 and
ANG II-stimulated OPN expression in PASMCs is mediated through activation of
Ras/MEK1/2/ERK1/2 and Ras/JNK, but not the PI3K/Akt/p70S6K or Ras/p38MAPK signaling
pathways. These differences in signaling suggest that PN and OPN may play different roles in
pulmonary vascular remodeling.

14
ACKNOWLEDGMENTS
This work was supported in part by National Heart, Lung, and Blood Institute Grants
HL-44195, HL-50147, HL-45990, HL-07457 and HL56046.

15
REFERENCES
1. Arendshorst WJ, Brannstrom K, Ruan X. Actions of angiotensin II on the renal
microvasculature. J Am Soc Nephrol 10 Suppl 11:S149-161, 1999.
2. Brown LF, Berse B, Van de Water L, Papadopoulos-Sergiou A, Perruzzi CA, Manseau EJ,
Dvorak HF, Senger DR. Expression and distribution of osteopontin in human tissues:
widespread association with luminal epithelial surfaces. Mol Biol Cell 3:1169-1180, 1992.
3. Chen SJ, Chen YF, Meng QC, Durand J, DiCarlo VS, Oparil S. Endothelin receptor
antagonist bosentan prevents and reverses hypoxia induced pulmonary hypertension in
rats. J Appl Physiol 79:2122-2131, 1995.
4. Chen SJ, Chen YF, Opgenorth TJ, Wessale JL, Meng QC, Durand J, DiCarlo VS, Oparil
S. The orally active nonpeptide endothelin A-receptor antagonist A-127722 prevents and
reverses hypoxia-induced pulmonary hypertension and pulmonary vascular remodeling in
Sprague-Dawley rats. J Cardiovasc Pharmacol 29:713-725, 1997.
5. Gillan L, Matei D, Fishman DA, Gerbin CS, Karlan BY, Chang DD. Periostin secreted by
epithelial ovarian carcinoma is a ligand for alpha(V)beta(3) and alpha(V)beta(5) integrins
and promotes cell motility. Cancer Res 62:5358-5364, 2002.
6. Horiuchi K, Amizuka N, Takeshita S, Takamatsu H, Katsuura, Ozawa H, Toyama, Y,
Bonewald LF, Kudo A. Identification and characterization of a novel protein, periostin, with
restricted expression to periosteum and periodontal ligament and increased expression by
transforming growth factor . J Bone Miner Res 14:1239-1249, 1999.
7. Hoshikawa Y, Nana-Sinkam P, Moore MD, Sotto-Santiago S, Phang T, Keith RL, Morris
KG, Kondo T, Tuder RM, Voelkel NF, Geraci MW. Hypoxia induces different genes in the
lungs of rats compared with mice. Physiol Genomics 12:209-219, 2003.
8. Inagami T, Eguchi S. Angiotensin II-mediated vascular smooth muscle cell growth

16
signaling. Braz J Med Biol Res 33:619-624, 2000.
9. Klint P, Claesson-Welsh L. Signal transduction by fibroblast growth factor receptors. Front
Biosci 4:D165-177, 1999.
10. Kruzynska-Frejtag A, Machnicki M, Rogers R, Markwald RR, Conway SJ. Periostin (an
osteoblast-specific factor) is expressed within the embryonic mouse heart during valve
formation. Mechanisms of Development 103:183-188, 2001.
11. Li G, Wang D, Chen YF, Oparil S. Vascular injury induces expression of periostin, a novel
vascular extracellular matrix protein, via the PI3K-kinase-MAP kinase pathway. J Am Coll
Cardiol 41:34A, 2003.
12. Li G, Oparil S, Kelpke SS, Chen YF, Thompson JA. Fibroblast growth factor receptor-1
signaling induces osteopontin expression and vascular smooth muscle cell-dependent
adventitial fibroblast migration in vitro. Circulation 106:854-859, 2002.
13. Li P, Oparil S, Sun JZ, Thompson JA, Chen YF. Fibroblast growth factor mediates
hypoxia-induced endothelin--A receptor expression in lung artery smooth muscle cells. J
Appl Physiol 95:643-651, 2003.
14. Matsui R, Brody JS, Yu Q. FGF-2 induces surfactant protein gene expression in foetal rat
lung epithelial cells through a MAPK-independent pathway. Cell Signal 11:221-228, 1999.
15. Miyamoto T, Leconte I, Swain JL, Fox JC. Autocrine FGF signaling is required for
vascular smooth muscle cell survival in vitro. J Cell Physiol 177:58-67, 1998.
16. O’Regan A. The role of osteopontin in lung disease. Cytokine & Growth Factor Reviews
14:479-488, 2003.
17. Panek RL, Lu GH, Dahring TK, Batley BL, Connolly C, Hamby JM, and Brown KJ. In vitro
biological characterization and antiangiogenic effects of PD166866, a selective inhibitor
of the FGF-1 receptor tyrosine kinase. J Pharmacol Exp Ther 286:569-577, 1998.

17
18. Sasaki H, Yu CY, Dai M, Tam C, Loda M, Auclair D, Chen LB, Elias A. Elevated serum
periostin levels in patients with bone metastases from breast but not lung cancer. Breast
Cancer Res Treat 77:245-252, 2003.
19. Shimoda LA, Sham JSK, Sylvester JT. Altered pulmonary vasoreactivity in the chronically
hypoxic lung. Physiol Res 49:549-560, 2000.
20. Sodhi CP, Phadke SA, Batlle D, Sahai A. Hypoxia stimulates osteopontin expression and
proliferation of cultured vascular smooth muscle cells: potentiation by high glucose.
Diabetes 50:1482-1490, 2001.
21. Sun JZ, Chen SJ, Li G, Chen YF. Hypoxia reduces atrial natriuretic peptide clearance
receptor gene expression in ANP knockout mice. Am J Physiol Lung Cell Mol Physiol
279:L511-L519, 2000.
22. Sun JZ, Oparil S, Lucchesi P, Thompson JA, and Chen YF. Tyrosine kinase receptor
activation inhibits NPR-C in lung arterial smooth muscle cells. Am J Physiol
281:L155-L163, 2001.
23. Takeshita S, Kikuno R, Tezuka KI, Amann E. Osteoblast-specific factor 2: cloning of a
putative bone adhesion protein with homology with the insect protein fasciclin I. Biochem
J 294:271-278, 1993.
24. Touyz RM, Schiffrin EL. Signal transduction mechanisms mediating the physiological and
pathophysiological actions of angiotensin II in vascular smooth muscle cells. Pharmacol
Rev 52:639-672, 2000.
25. Tuck AB, Hota C, Wilson SM, Chambers AF. Osteopontin-induced migration of human
mammary epithelial cells involves activation of EGF receptor and multiple signal
transduction pathways. Oncogene 22:1198-1205, 2003.
26. Wang D, Oparil S, Feng JA, Li P, Perry G, Chen LB, Dai M, John SW, Chen YF. Effects

18
of pressure overload on extracellular matrix expression in the heart of the atrial natriuretic
peptide-null mouse. Hypertension 42:88-95, 2003.
27. Xin X, Johnson AD, Scott-Burden T, Engler D, Casscells W. The predominant form of
fibroblast growth factor receptor expressed by proliferating human arterial smooth muscle
cells in culture is type I. Biochem Biophys Res Commun 204:557-564, 1994.
28. Zhou FY, Kan M, Owens RT, McKeehan WL, Thompson JA, Linhardt RJ, and Hook M.
Heparin-dependent fibroblast growth factor activates: effects of defined heparin
oligosaccharides. Eur J Cell Biol 73:71-80, 1997.

19
FIGURE LEGENDS
Figure 1. (A and B), effects of 2-wk hypoxic exposure (10% O2, 1 atm) on steady-state periostin
(PN) and osteopontin (OPN) mRNA expression in rat lung in vivo. Air controls were
rats exposed to room air during the same period. (C and D), effects of 6 to 24-hr
hypoxic exposure (1% O2) on steady-state PN and OPN mRNA expression in
pulmonary arterial smooth muscle cells (PASMCs) in vitro. Growth arrested PASMCs
were exposed to hypoxia for 6, 12, and 24 hr before being harvested. Normoxic
controls were PASMCs cultured in 0.1% FBS medium for 24-hr and exposed to 21%
O2 for an additional 24 hrs. Numbers in parentheses are the number of rats or plates
of PASMCs contributing data to each group. Northern blot analysis was carried out
with 15 ug of total RNA extracted from each lung or plates of PASMCs. The Northern
blot membrane was probed with PN and OPN cDNAs, and 18S rRNA oligonucleotide
sequentially. The mRNA data were normalized by 18S rRNA to correct for variations in
RNA loading. Results are means±SE. * p<0.05 vs. respective air or normoxia control
groups by unpaired t-test (in vivo study) or 1-way ANOVA (in vitro study).
Figure 2. Representative light micrographs of small pulmonary arteries (PA; ~100 um) from rats
exposed to chronic hypoxia (10% O2) for 2 wks or room air. Lung sections were
immunostained (brown stain) with selective rabbit anti-PN polyclonal antibody.
Sections were counterstained with hematoxylin and eosin. BR-bronchi.
Figure 3. Dose- and time-dependent effects of FGF-1 on regulation of steady-state PN (A and
C) and OPN (B and D) mRNA expression in PASMCs in vitro. Growth arrested
PASMCs were exposed to different concentrations of FGF-1 for 24 hr (A and B) or 10

20
ng/ml FGF-1 for 3, 6, 12 and 24 hrs (C and D) before being harvested. Controls were
PASMCs cultured in 0.1% FBS medium for 24-hr and treated with vehicle for an
additional 24 hrs. The mRNA loading was normalized by 18S rRNA. Numbers in
parentheses are the number of plates contributing data to each group. Results are
means±SE. * p<0.05 vs. control group by 1-way ANOVA.
Figure 4. Dose- and time- dependent effects of ANG II on regulation of steady-state PN (A and
C) and OPN (B and D) mRNA expression in PASMCs. Growth arrested PASMCs were
exposed to different concentrations of ANG II for 24 hr (A and B) or 100 nM ANG II for
3, 6, 12 and 24 hrs (C and D) before being harvested. Controls were PASMCs
cultured in 0.1% FBS medium for 24-hr and treated with vehicle for an additional 24
hrs. The mRNA loading was normalized by 18S rRNA. Numbers in parentheses are
the number of plates contributing data to each group. Results are means±SE. *
p<0.05 vs. control group by 1-way ANOVA.
Figure 5. (A and B), effects of the FGF-1 receptor antagonist PD166866 and signaling pathway
inhibitors on the upregulation of PN and OPN mRNA expression in PASMCs by
FGF-1; (C and D), effects of the AT-1 receptor antagonist losartan and signaling
pathway inhibitors on the upregulation of PN and OPN mRNA expression in PASMCs
by ANG II; and (E and F), effects of signaling pathway inhibitors on PN and OPN
mRNA expression in PASMCs exposed to normoxia or hypoxia (1% O2) for 24 hrs.
Controls (or vehicles) were growth-arrested PASMCs cultured in 0.1% FBS medium.
PASMCs were pretreated with PD166866 (5 uM) or losartan (10 mM) for 45 min
before exposing to FGF-1 or ANG II for 24 hr. Other experimental groups were

21
PASMCs pretreated with the PI3K inhibitors LY294002 (LY, 10 uM) or wortmannin (20
uM), as well as the MEK1/2 inhibitors U-0126 (5 uM) or PD98059 (20 uM), for 45 min
before adding FGF-1 (10 ng/ml) (A and B) or ANG II (100 nM) (C and D) to the medium
or before exposing to hypoxia (E and F). An additional 24-hr incubation was carried
out before harvesting for analysis. The mRNA loading was normalized by 18S RNA.
Numbers in parentheses are the number of plates contributing data to each group.
Results are mean±SE. * p<0.05 vs. respective control groups; # p<0.05 vs. respective
FGF-1 or ANG II groups; and & p<0.05 vs. respective vehicle control groups by 1-way
ANOVA. Rapamycin (100 nM, a p70S6K inhibitor), SB203580 (SB, 10 uM, a p38
MAPK inhibitor), and SP600125 (20 uM, a JNK inhibitor).
Figure 6. A: effects of the selective PI3K inhibitor LY294002 (10 uM) on FGF-1 or ANG
II-stimulated PI3K in PASMCs. Control was growth-arrested PASMCs cultured in 0.1%
FBS medium. Quiescent PASMCs were stimulated with FGF-1 (10 ng/ml) or ANG II
(100 nM) for 15 min without or with pretreatment with LY294002 (10 uM, 45 min). Cell
lysates (25 ug) were size fractionated by SDS-PAGE, and Western blot analysis was
performed with or without anti-phospho-Akt or anti-phospho-GSK3 specific antibodies.
LY294002 selectively blocked the FGF-1 or ANG II-induced phosphorylation of Akt
and GSK3.
B: effects of the selective MEK1/2 inhibitor U0126 (5 uM) on FGF-1 or ANG
II-stimulated MAP kinase in PASMCs. Quiescent PASMCs were stimulated with
FGF-1 (10 ng/ml) or ANG II (100 nM) for 15 min without or with pretreatment with
U0126 (5 uM, 45 min). Cell lysates (25 ug) were size fractionated by SDS-PAGE, and
Western blot analysis was performed with anti-phospho-MEK1/2 or

22
anti-phospho-ERK1/2 specific antibodies. U0126 selectively blocked the FGF-1 or
ANG II-induced phosphorylation of ERK1/2, but not the phosphorylation of MEK1/2.
Figure 7. PN (A and B) and OPN (C and D) mRNA expression in PASMCs treated with FGF-1
(10 ng/ml) or the p38MAPK and p70S6K activator anisomycin (1 ug/ml) for 24 hrs in
the presence or absence of selective inhibitors intracellular signaling, including
rapamycin (100 nM, a p70S6K inhibitor), SB203580 (10 uM, a p38 MAPK inhibitor), or
SP600125 (20 uM, a JNK inhibitor). Quiescent PASMCs were pretreated with
inhibitors for 45 min before adding FGF-1 or anisomycin. Controls were PASMCs
cultured in 0.1% FBS medium for 24-hr and treated with vehicle for an additional 24
hrs. Representative Northern blots are shown at top. The mRNA loading was
normalized by 18S rRNA. Numbers in parentheses are the number of plates
contributing data to each group. Results are means±SE. * p<0.05 vs. control group;
and # p<0.05 vs. respective FGF-1 or anisomycin groups by 1-way ANOVA.
Figure 8. A simplified schematic illustration of the specific action of various signaling pathway
inhibitors and the intracellular signaling pathways that mediate FGF-1 or
ANGII-induced PN and OPN expression in cultured PASMCs. RTK - receptor tyrosine
kinase, GPCR - G-protein coupled receptor.
23
Figure 1
Air Hypoxia0
1
2
3
(6)
Ost
eopo
ntin
mR
NA
/18S
RN
A (6) *OsteopontinB.
Air Hypoxia0
1
2
3
(6)
Per
iost
inm
RN
A/1
8S R
NA
(6)*
PeriostinA.
6 hr 12 hr 24 hr0
1
2
3
(12)
Per
iost
inm
RN
A/1
8S R
NA
(5) (3)
(15)*
PeriostinC.
Hypoxia (1% O2)
Normoxia 6 hr 12 hr 24 hr0
1
2
3
(12)
Ost
eopo
ntin
mR
NA
/18S
RN
A
(5) (3)
OsteopontinD.
Hypoxia (1% O2)
Normoxia
(15)
Air Hypoxia0
1
2
3
(6)
Ost
eopo
ntin
mR
NA
/18S
RN
A (6) *OsteopontinB.
Air Hypoxia0
1
2
3
(6)
Per
iost
inm
RN
A/1
8S R
NA
(6)*
PeriostinA.
6 hr 12 hr 24 hr0
1
2
3
(12)
Per
iost
inm
RN
A/1
8S R
NA
(5) (3)
(15)*
PeriostinC.
Hypoxia (1% O2)
Normoxia 6 hr 12 hr 24 hr0
1
2
3
(12)
Ost
eopo
ntin
mR
NA
/18S
RN
A
(5) (3)
OsteopontinD.
Hypoxia (1% O2)
Normoxia
(15)
24
Air Control Hypoxia
Figure 2
PA
BR
PA
BR
25
Figure 3
0 0.1 1 10 200
2
4
6
8
(12)O
steo
pont
in m
RN
A/1
8S R
NA
FGF-1 (ng/ml)
(3)
(3)
(12)
(3)OsteopontinB.
0 0.1 1 10 200
1
2
3
(12)
Per
iost
in R
NA
/18S
RN
A
FGF-1 (ng/ml)
(3)
(3)
(12)(3)
PeriostinA.
Control 3 hr 6 hr 12 hr 24 hr0
1
2
3
(9)
Per
iost
in m
RN
A/1
8S R
NA
FGF-1 (10 ng/ml)
(3)
(3)(3)
(9)
PeriostinC.
Control 3 hr 6 hr 12 hr 24 hr0
2
4
6
8
(12)
Ost
eopo
ntin
mR
NA
/18S
RN
A
FGF-1 (10 ng/ml)
(3)
(3)(3)
(12)
OsteopontinD.
26
Figure 4
0 0.1 1 10 1000
2
4
6
8
10
(6)O
steo
pont
in m
RN
A/1
8S R
NA
ANG II (nM)
(3)(3)
(3)(6)
OsteopontinB.
0 0.1 1 10 1000
1
2
3
4
(6)
Per
iost
in m
RN
A/1
8S R
NA
ANG II (nM)
(3)(3)
(3)
(6)
PeriostinA.
Control 3 hr 6 hr 12 hr 24 hr0
1
2
3
4
(6)
Per
iost
in m
RN
A/1
8S R
NA
ANG II (100 nM)
(3)
(3)
(3)
(6)
PeriostinC.
Control 3 hr 6 hr 12 hr 24 hr0
2
4
6
8
10
(6)
Ost
eopo
ntin
mR
NA
/18S
RN
A
ANG II (100 nM)
(3)
(3)(3)(6)
OsteopontinD.
27
Figure 5
Control FGF-1 PD166866 LY WM U0126 PD980590
1
2
3
(9)
Per
iost
inm
RN
A/1
8S R
NA
FGF-1 (10 ng/ml)
(9)
(4)
(3)
Periostin
(4)
## #
A.
(3)#
(3)#
Control FGF-1 PD166866 LY WM U0126 PD980590
2
4
6
8
(9)
Ost
eopo
ntin
mR
NA
/18S
RN
A
FGF-1 (10 ng/ml)
(9)
(3)
(3)
Osteopontin
(4)#
B.
(3)#
(3)#
Control AII Losartan LY WM U0126 PD980590
1
2
3
4
5
(12)
Per
iost
inm
RN
A/1
8S R
NA
ANG II (100 nM)
(12)
(3) (3)
Periostin
(3)## #
C.
(3)#
(3)
Control AII Losartan LY WM U0126 PD980590
4
8
12
16
(12)
Ost
eopo
ntin
mR
NA
/18S
RN
A
ANG II (100 nM)
(12)(3) (3)
Osteopontin
(3)
D.
(3)##
(3)
Vehicle LY PD98059 Rapamycin SB SP6001250
1
2
(9)
Per
iost
inm
RN
A/1
8S R
NA
(3)
(3)
(3)
(5)
E.
(5)
(3)&
*Normoxia Hypoxia
(3)(3) (5)
(3)(3)
Periostin
&
& &
Vehicle LY PD98059 Rapamycin SB SP6001250
1
2
(9)
Ost
eopo
ntin
mR
NA
/18S
RN
A
(3)
(3)
(3)(5)
F.
(5)(3)&
Normoxia Hypoxia
(3)(3)
(5)
(3)
(3)
Osteopontin
&& &
& &
#
Control FGF-1 PD166866 LY WM U0126 PD980590
1
2
3
(9)
Per
iost
inm
RN
A/1
8S R
NA
FGF-1 (10 ng/ml)
(9)
(4)
(3)
Periostin
(4)
## #
A.
(3)#
(3)#
Control FGF-1 PD166866 LY WM U0126 PD980590
2
4
6
8
(9)
Ost
eopo
ntin
mR
NA
/18S
RN
A
FGF-1 (10 ng/ml)
(9)
(3)
(3)
Osteopontin
(4)#
B.
(3)#
(3)#
Control AII Losartan LY WM U0126 PD980590
1
2
3
4
5
(12)
Per
iost
inm
RN
A/1
8S R
NA
ANG II (100 nM)
(12)
(3) (3)
Periostin
(3)## #
C.
(3)#
(3)
Control AII Losartan LY WM U0126 PD980590
4
8
12
16
(12)
Ost
eopo
ntin
mR
NA
/18S
RN
A
ANG II (100 nM)
(12)(3) (3)
Osteopontin
(3)
D.
(3)##
(3)
Vehicle LY PD98059 Rapamycin SB SP6001250
1
2
(9)
Per
iost
inm
RN
A/1
8S R
NA
(3)
(3)
(3)
(5)
E.
(5)
(3)&
*Normoxia Hypoxia
(3)(3) (5)
(3)(3)
Periostin
&
& &
Vehicle LY PD98059 Rapamycin SB SP6001250
1
2
(9)
Ost
eopo
ntin
mR
NA
/18S
RN
A
(3)
(3)
(3)(5)
F.
(5)(3)&
Normoxia Hypoxia
(3)(3)
(5)
(3)
(3)
Osteopontin
&& &
& &
#
28
Figure 6
Control FGF-1LY294002+ FGF-1 ANGII
LY294002+ ANGII
Phospho-Akt
Phospho-GSK3
Control FGF-1U0126
+ FGF-1 ANGIIU0126
+ ANGII
Phospho-ERK1/2
Phospho-MEK1/2
A.
B.
29
Figure 7
Control FGF-1 Rapamycin SB203580 SP6001250
1
2
3
4
(9)
Ost
eopo
ntin
mR
NA
/18S
RN
A
FGF-1 (10 ng/ml)
(9) (5) (5)
Osteopontin
(5)
*
#
* *
Control FGF-1 Rapamycin SB203580 SP6001250
0.8
1.6
2.4
(9)
Per
iost
inm
RN
A/1
8S R
NA
FGF-1 (10 ng/ml)
(9)
(5)(5)
Periostin
(5)#
#
* *
Control Anisomycin Rapamycin SB203580 SP6001250
1
2
(9)
Ost
eopo
ntin
mR
NA
/18S
RN
A
Anisomycin (1 ug/ml)
(5)(5) (5)
Osteopontin
(5)
* * **
Control FGF-1 Rapamycin SB203580 SP600125 Control Anisomycin Rapamycin SB203580 SP600125
Anisomycin (1 ug/ml)FGF-1 (10 ng/ml)
- OPN -
- PN -
Control Anisomycin Rapamycin SB203580 SP6001250
10
20
30
40
(9)P
erio
stin
mR
NA
/18S
RN
A
Anisomycin (1 ug/ml)
(5)
(5)
(5)
Periostin
(5)
*
*
*
*#
#
Control FGF-1 Rapamycin SB203580 SP6001250
1
2
3
4
(9)
Ost
eopo
ntin
mR
NA
/18S
RN
A
FGF-1 (10 ng/ml)
(9) (5) (5)
Osteopontin
(5)
*
#
* *
Control FGF-1 Rapamycin SB203580 SP6001250
0.8
1.6
2.4
(9)
Per
iost
inm
RN
A/1
8S R
NA
FGF-1 (10 ng/ml)
(9)
(5)(5)
Periostin
(5)#
#
* *
Control Anisomycin Rapamycin SB203580 SP6001250
1
2
(9)
Ost
eopo
ntin
mR
NA
/18S
RN
A
Anisomycin (1 ug/ml)
(5)(5) (5)
Osteopontin
(5)
* * **
Control FGF-1 Rapamycin SB203580 SP600125 Control Anisomycin Rapamycin SB203580 SP600125
Anisomycin (1 ug/ml)FGF-1 (10 ng/ml)
- OPN -
- PN -
Control Anisomycin Rapamycin SB203580 SP6001250
10
20
30
40
(9)P
erio
stin
mR
NA
/18S
RN
A
Anisomycin (1 ug/ml)
(5)
(5)
(5)
Periostin
(5)
*
*
*
*#
#
30
Figure 8
PI3K
GPCR
p70S6K
Src
Raf
MKK1/2
JNKERK1/2p38
Periostin expression Osteopontin expression
MKK3/6 MKK4/7
LY294002Wortmannin
Rapamycin
U0126 PD98059
SB203580 SP600125
( - )
Anisomycin
FGF AII
RTK
Ras
( - )( - )( + )
( - )
( - )
PI3K
GPCR
p70S6K
Src
Raf
MKK1/2
JNKERK1/2p38
Periostin expression Osteopontin expression
MKK3/6 MKK4/7
LY294002Wortmannin
Rapamycin
U0126 PD98059
SB203580 SP600125
( - )
Anisomycin
FGF AII
RTK
Ras
( - )( - )( + )
( - )
( - )




















