
Hypnotic induction decreases anterior default mode activity
8
Hypnotic induction decreases anterior default mode activity William J. McGeown a , Giuliana Mazzoni a , Annalena Venneri a,b , Irving Kirsch a, * a Department of Psychology, University of Hull, UK b Department of Neuroscience, University of Modena and Reggio Emilia, Italy article info Article history: Received 27 April 2009 Available online 25 September 2009 Keywords: Hypnosis Suggestion Trance State Default mode Consciousness Functional magnetic resonance imaging (fMRI) Neuroimaging Frontal cortex abstract The ‘default mode’ network refers to cortical areas that are active in the absence of goal- directed activity. In previous studies, decreased activity in the ‘default mode’ has always been associated with increased activation in task-relevant areas. We show that the induc- tion of hypnosis can reduce anterior default mode activity during rest without increasing activity in other cortical regions. We assessed brain activation patterns of high and low suggestible people while resting in the fMRI scanner and while engaged in visual tasks, in and out of hypnosis. High suggestible participants in hypnosis showed decreased brain activity in the anterior parts of the default mode circuit. In low suggestible people, hyp- notic induction produced no detectable changes in these regions, but instead deactivated areas involved in alertness. The findings indicate that hypnotic induction creates a distinc- tive and unique pattern of brain activation in highly suggestible subjects. Ó 2009 Elsevier Inc. All rights reserved. 1. Introduction Our purpose in designing this study was to establish whether the induction of hypnosis produces a unique hypnotic state (Lynn, Kirsch, & Hallquist, 2008) and, if so, to identify its neural correlates. There have been several brain imaging studies on hypnosis, but these have not contributed consistent results, and the neurobiological correlates of the hypnotic induction per se have not been reliably identified (Oakley, 2008). Inconsistency between findings might potentially be accounted for by methodological differences between the studies (Oakley, 2008). In many studies, the hypnotic induction has been confounded with the administration of specific hypnotic suggestions, so that brain activation following a hypnotic suggestion (i.e., a suggested change in experience given after the induction of hypnosis) is compared to activation without either the induction of hypnosis or the suggestion (Faymonville et al., 2000; Grond, Pawlik, Walter, Lesch, & Heiss, 1995; Maquet et al., 1999). The experimental design adopted in these brain imaging experiments does not allow a clear distinction between differences in brain activation that might arise from the induction of hypnosis and those due to task-related suggestion. A better strategy might be to hold suggestion and other task demands constant, so that the only difference would be the presence/absence of hypnotic induction. This strategy has been adopted in a few published studies (Egner, Jamieson, & Gru- zelier, 2005; Rainville, Hofbauer, Bushnell, Duncan, & Price, 2002; Rainville et al., 1999). Even in these studies, however, the design was such that the effects of hypnosis on brain physiology per se could not be determined. For example, participants in the Egner et al. (2005) study engaged in a Stroop task, and those in the Rainville et al. (1999, 2002) studies had their hand immersed in warm or painfully hot water. These studies reported modulation of activity in the anterior cingulate cortex due 1053-8100/$ - see front matter Ó 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.concog.2009.09.001 * Corresponding author. Address: Department of Psychology, University of Hull, Cottingham Road, Hull HU6 7RX, UK. Fax: +44 (0) 1482 465599. E-mail address: [email protected] (I. Kirsch). Consciousness and Cognition 18 (2009) 848–855 Contents lists available at ScienceDirect Consciousness and Cognition journal homepage: www.elsevier.com/locate/concog
-
Upload
strathclyde -
Category
Documents
-
view
0 -
download
0
Transcript of Hypnotic induction decreases anterior default mode activity

Consciousness and Cognition 18 (2009) 848–855
Contents lists available at ScienceDirect
Consciousness and Cognition
journal homepage: www.elsevier .com/locate /concog
Hypnotic induction decreases anterior default mode activity
William J. McGeown a, Giuliana Mazzoni a, Annalena Venneri a,b, Irving Kirsch a,*
a Department of Psychology, University of Hull, UKb Department of Neuroscience, University of Modena and Reggio Emilia, Italy
a r t i c l e i n f o a b s t r a c t
Article history:Received 27 April 2009Available online 25 September 2009
Keywords:HypnosisSuggestionTranceStateDefault modeConsciousnessFunctional magnetic resonance imaging(fMRI)NeuroimagingFrontal cortex
1053-8100/$ - see front matter � 2009 Elsevier Incdoi:10.1016/j.concog.2009.09.001
* Corresponding author. Address: Department ofE-mail address: [email protected] (I. Kirsch).
The ‘default mode’ network refers to cortical areas that are active in the absence of goal-directed activity. In previous studies, decreased activity in the ‘default mode’ has alwaysbeen associated with increased activation in task-relevant areas. We show that the induc-tion of hypnosis can reduce anterior default mode activity during rest without increasingactivity in other cortical regions. We assessed brain activation patterns of high and lowsuggestible people while resting in the fMRI scanner and while engaged in visual tasks,in and out of hypnosis. High suggestible participants in hypnosis showed decreased brainactivity in the anterior parts of the default mode circuit. In low suggestible people, hyp-notic induction produced no detectable changes in these regions, but instead deactivatedareas involved in alertness. The findings indicate that hypnotic induction creates a distinc-tive and unique pattern of brain activation in highly suggestible subjects.
� 2009 Elsevier Inc. All rights reserved.
1. Introduction
Our purpose in designing this study was to establish whether the induction of hypnosis produces a unique hypnotic state(Lynn, Kirsch, & Hallquist, 2008) and, if so, to identify its neural correlates. There have been several brain imaging studies onhypnosis, but these have not contributed consistent results, and the neurobiological correlates of the hypnotic induction perse have not been reliably identified (Oakley, 2008).
Inconsistency between findings might potentially be accounted for by methodological differences between the studies(Oakley, 2008). In many studies, the hypnotic induction has been confounded with the administration of specific hypnoticsuggestions, so that brain activation following a hypnotic suggestion (i.e., a suggested change in experience given after theinduction of hypnosis) is compared to activation without either the induction of hypnosis or the suggestion (Faymonvilleet al., 2000; Grond, Pawlik, Walter, Lesch, & Heiss, 1995; Maquet et al., 1999). The experimental design adopted in thesebrain imaging experiments does not allow a clear distinction between differences in brain activation that might arise fromthe induction of hypnosis and those due to task-related suggestion.
A better strategy might be to hold suggestion and other task demands constant, so that the only difference would be thepresence/absence of hypnotic induction. This strategy has been adopted in a few published studies (Egner, Jamieson, & Gru-zelier, 2005; Rainville, Hofbauer, Bushnell, Duncan, & Price, 2002; Rainville et al., 1999). Even in these studies, however, thedesign was such that the effects of hypnosis on brain physiology per se could not be determined. For example, participants inthe Egner et al. (2005) study engaged in a Stroop task, and those in the Rainville et al. (1999, 2002) studies had their handimmersed in warm or painfully hot water. These studies reported modulation of activity in the anterior cingulate cortex due
. All rights reserved.
Psychology, University of Hull, Cottingham Road, Hull HU6 7RX, UK. Fax: +44 (0) 1482 465599.

W.J. McGeown et al. / Consciousness and Cognition 18 (2009) 848–855 849
to hypnosis, but they included concomitant tasks (Stroop task/pain) which are also known to involve this area. The differ-ences in brain activity in hypnotic and non-hypnotic conditions might, therefore, be task specific. In addition, in most studiesthe participants were aware of the purpose of the experiment and what was required of them, which might have influencedbrain activity and produced changes that are not specific to hypnosis. Our alternative approach was to scan participants dur-ing rest periods following the induction procedure, while they were not performing any specific task and were unaware thatassessment had begun. This method minimizes the confounding effects of task, demand characteristics, and performanceexpectations.
The pattern of spontaneous physiological brain activity that is normally detectable during normal resting state is referredto as the ‘default mode’ network of brain function (Raichle et al., 2001). Areas collectively activated during the default modestate involve a set of midline brain structures, including the anterior cingulate, ventral and dorsal medial prefrontal cortex,posterior cingulate and precuneus (Fox & Raichle, 2007; Mason et al., 2007; Raichle et al., 2001). Oakley and Halligan (2009)have suggested that a deviation from the normal default mode activity might provide a neural signature of hypnosis. In thepresent study, we examined whether any changes occurred to the standard pattern of brain activity during rest after a hyp-notic induction.
A group of low suggestible participants (i.e. people who do not respond to hypnotic suggestions) were also included to seewhether similar alterations to the pattern of spontaneous physiological brain activity occurs in both high and low suggest-ible people. Changes that are not specific to hypnosis should be found in both groups, whereas changes that are specific tohypnosis should be found only in people who are responsive to hypnosis.
In this study, periods of scanning while participants were resting were alternated with periods of passive and active view-ing, which participants had been led to believe were the focus of the experiment. Brain activity during these conditions wasrecorded following a hypnotic induction and also without the induction of hypnosis. By comparing scans acquired duringrest periods between hypnosis/non hypnosis runs within subjects in the first level analysis, this experimental procedureminimized the influence of demand characteristics, concurrent tasks, hypnotic suggestions, and performance expectationson brain activity. Instructions were worded in a way that participants had no awareness that scans collected during theseresting periods would be used in data analyses. For the passive condition they had to perform a relatively undemanding pas-sive visual perception task (e.g. look at a complex colour or greyscale pattern); for the active condition they had to perform ademanding active visual hallucination task (e.g. draining colour from the colour pattern or adding colour to the greyscalepattern). The aim of this fMRI study was to discover if alterations in the pattern of spontaneous physiological brain activityduring rest occur in high and/or low suggestible participants once hypnosis is induced when compared to rest activity out ofhypnosis.
2. Method
2.1. Participants
Two hundred sixty three potential participants were screened for hypnotic suggestibility on a modified version of theCarleton University Responsiveness to Suggestion Scale (CURSS) (Comey & Kirsch, 1999; Spanos et al., 1983). The CURSSis a widely used group scale for assessing hypnotic suggestibility. It consists of a hypnotic induction followed by seven sug-gestions requesting ideomotor movements, movement inhibition, and alterations of perception and memory. Participantsreceive one point for each suggestion to which they respond behaviorally. The Comey and Kirsch (1999) modification ofthe CURSS has an extended hypnotic induction and introductory instructions that are similar to those of the individuallyadministered Stanford Hypnotic Susceptibility Scale: Form C (Weitzenhoffer & Hilgard, 1962), which is generally consideredthe gold standard for measuring hypnotic suggestibility. Screening was conducted by university faculty members withgroups of participants ranging from 1 to 60 individuals. Individuals who scored in the top (5–7) or bottom range (0–1) wereinvited for further participation.
In a second screening session, participants were asked individually, both in and out of hypnosis, to respond the perceptualalteration suggestions that they were told were the focus of the study. The hypnotic induction was taken from Kirsch, Lynn,and Rhue (1993) and consisted of suggestions for relaxation, pleasant visual imagery, and entry into a hypnotic state. Selfreport data of these participants’ responses to the perceptual alteration suggestions are reported elsewhere (Mazzoniet al., 2009). Out of these people, eleven highly suggestible participants who succeeded in responding to the perceptual alter-ation experiences (mean suggestibility rating 5.82 on the CURSS, SD 0.87, range 5–7) and seven low suggestible participants(mean suggestibility rating 0.29, SD 0.49, range 0–1) were included in the fMRI study.
2.2. Procedure
Participants were informed that the purpose of the study was to assess what happens in the brain when people respondto suggestions for perceptual alterations both in and out of hypnosis. They were told that while in the scanner, they wouldsee a screen with a fixation point on which they were to focus their eyes, that the fixation point would be followed by a pat-tern which they had to look at, and that this would then be followed by a signal to alter their perception of the pattern. Theywere also told that, as in the second screening session, this would be done in and out of hypnosis and that the purpose of the

850 W.J. McGeown et al. / Consciousness and Cognition 18 (2009) 848–855
study was to compare brain activation when responding to the perceptual alteration suggestion in and out of hypnosis. Thefocus of the present study, however, was on the eight 20 s intervals during which the fixation point was on the screen beforebeing replaced by the pattern. Given the instructions and the previous screening on the perceptual alteration suggestionsand since the experimenter who instructed the participants was also not aware of the true focus of the study at the timeof data collection, it seems unlikely that participants would be able to divine that these brief intervals were of any interest.After explaining the experimental procedure, informed consent was obtained for all participants.
Subsequent to the fMRI session, high suggestible participants were contacted via email and asked to think back abouttheir experiences and indicate how they differed in and out of hypnosis. All of the responses to this question referred to theirexperience of the suggestions. None referred to the rest periods or to their experience of hypnosis per se. This supports oursupposition that participants understood the perceptual alteration task, rather than the 20 s between-trial pauses (i.e., theresting state periods), to be the focus of the study. Hence, any influence of demand characteristics and performance expec-tations on the data reported here are most likely negligible.
2.3. fMRI methods
Echo planar T2* weighted MRI images were acquired on a 3T Philips Achieva MRI system (TR = 2 s, TE = 35 ms, flip an-gle a = 80�, voxel dimensions 1.88 � 1.88 � 3.00 mm, field of view 240 mm, matrix 128 � 128 � 30). Three hundred sev-enty six volumes of 30 contiguous slices were acquired in each run. Four runs were acquired, two in hypnosis and two outof hypnosis. Hypnosis was induced using an abbreviated version of the induction used during the second screening ses-sion. Each run was preceded by 30 s of dummy scans to allow the scanner to reach equilibrium. The experiment followeda block design, alternating the conditions of rest, passive task (view a colour or greyscale pattern), and active task (draincolour from a coloured pattern or add colour to a greyscale pattern). Colour and greyscale trials were counterbalancedacross runs. These conditions were repeated four times in each run. The hypnosis/no hypnosis conditions were counter-balanced across the participants with half being imaged under hypnosis first and the other half out of hypnosis first, toensure that order effects (participant or scanner related) would not affect the findings. The focus of the study was on thefMRI scans acquired over the repeated rest periods (in which participants had no task to perform and no specific require-ments). The total scanning time for rest in hypnosis was 320 s and 320 s out of hypnosis (16 epochs in and 16 epochs outof hypnosis for each participant). Suggestions and task instructions were kept constant for the hypnosis and no-hypnosisruns, thus ensuring that the effects of hypnotic induction on brain activation would not be confounded by these factors.This design enabled the within subject comparison of resting brain activation patterns, in and out of hypnosis. At the endof the fMRI session participants were also asked to indicate on a four point scale (normal state, relaxed, hypnotized, anddeeply hypnotized) (Hilgard & Tart, 1966) the level of hypnotic state they reached for both the hypnosis runs and the no-hypnosis runs.
2.4. fMRI data analysis
Imaging data were analyzed using Statistical Parametric Mapping (SPM5) image analysis software (Wellcome Centre forNeuroimaging, London, UK). All volumes from each subject were re-aligned after creating a mean as reference and re-slicedusing 4th Degree B-Spline interpolation methods to adjust for residual motion related signal changes. Images were spa-tially normalized to the standard EPI template available in SPM5 using non-linear estimation of parameters. Normalizedimages were then spatially smoothed with an 8 mm full width at half maximum isotropic Gaussian kernel to compensatefor any residual variability after spatial normalization. A boxcar waveform convolved with a synthetic hemodynamic re-sponse function (HRF) was used as the reference waveform for each condition. Image data were high-pass filtered witha set of discrete cosine basis functions with a cut-off period of 128 s. Head motion was less than 3 mm in all volunteersbut one, whose scans were not included in the analysis. Head motion was included as a regressor in the first level analysesfor the remaining participants. At first level, all four runs for each participant were entered into an analysis using the gen-eral linear model. For all conditions (rest, passive, active) contrasts were defined for each condition across scans, i.e. con-trasting each condition in the hypnosis runs directly with the same condition in the out of hypnosis runs and vice versa.For example, rest in hypnosis was compared to rest out of hypnosis and the converse comparison was also made. Six con-trast images were therefore generated for each participant. For the second level group analysis, random effect analyseswere carried out by entering each set of contrast images into one sample t-tests, one for each of the six sets of contrastimages. This was repeated for both the high and low suggestible participants. Group comparisons were also carried outfor each set of contrast images using an independent sample t-test. Height threshold of significance was set at p < .05 (cor-rected) for individual group analyses and p < .01 (corrected) for group comparisons. For some group analyses a more liberaluncorrected threshold level was used, primarily to see whether there were any significant clusters in the default modeareas described by previously published studies which were not detected at the chosen threshold level. When that wasthe case, only significant clusters in areas defined a priori based on published evidence which had an extent of at least250 voxels and survived small volume correction were taken into account. The type of correction used is detailed in thetables. Anatomical regions were identified using the Talairach Daemon Client (http://ric.uthscsa.edu/projects/tdc/) (Lancas-ter et al., 2000), following conversion of the Montreal Neurological Institute coordinates extracted from the SPM analysesinto Talairach coordinates.

W.J. McGeown et al. / Consciousness and Cognition 18 (2009) 848–855 851
3. Results
During data acquisition, one of the high suggestible participants had excessive movement and was, therefore, excludedfrom the analyses. This left the high suggestible group with 7 females and 3 males between the ages of 20 and 53 (mean25.00, SD 10.32). The low suggestible group had 5 females and 2 males between the ages of 20 and 35 (mean 26.86, SD 6.54).
3.1. Effects of hypnosis on the brain in the resting state
Immediately after the scanning session, participants were asked to rate their feelings of being hypnotized during both thehypnosis and no-hypnosis part of the session using a 4 point scale with the verbal anchors, normal awake state, very relaxed,mild hypnotic state, and deeply hypnotized. When out of hypnosis, both high and low suggestible people reported that theywere either in a normal awake state or very relaxed (mean 1.22, SD 0.44 and mean 1.29, SD 0.49 for highs and lows respec-tively). After hypnotic induction three highs reported being relaxed and the others reported being mildly or deeply hypno-tised (mean 3.00, SD 0.83), while lows rated themselves as being either relaxed or in a normal awake state (mean 1.57, SD0.53). Wilcoxon signed ranks tests revealed a significant difference between the state rating (before and after hypnosis) inthe high suggestible (Z = 2.68; p < .01), but not in the low suggestible (Z = 1.41; ns), indicating that hypnosis had a greatereffect on high than low suggestible people.
The areas which had significantly reduced levels of activation during hypnosis were identified by comparing the rest per-iod in the no hypnosis condition to the rest period in hypnosis. During hypnosis, high suggestible participants showed reduc-tions in brain activity in the anterior cingulate, medial and superior frontal gyri bilaterally, in addition to the left inferior andmiddle frontal gyri (see Table 1a, Fig. 1). The converse comparison, hypnosis rest versus no hypnosis rest, which should re-veal areas recruited significantly more after the hypnotic induction, found no significant differences.
The same analyses yielded a different pattern of changes in low suggestible participants when under hypnosis. These par-ticipants showed reductions related to the hypnotic induction during rest, in the posterior cingulate, thalamus, caudate nu-cleus and insula, bilaterally. Further decreases in activation were seen in the left inferior frontal gyrus, claustrum, lentiformnucleus and right subthalamic nucleus (see Table 1b, Fig. 1). As in the high suggestible participants, low suggestibles showedno areas which were significantly activated to higher levels after the hypnotic induction.
A direct comparison of high and low suggestible people showed that when under hypnosis high suggestible participantshad lower levels of activation than low suggestibles in the left inferior, middle, superior and medial frontal gyri (Table 1c, Fig1). No areas were present in which high suggestibles had significantly higher levels of activation than low suggestibles.
3.2. Effects of hypnosis on the brain during passive and active tasks
To determine if the hypnosis-induced decreases in prefrontal activation persisted when the participants engaged in tasks,the same comparisons were carried out on the periods when the participants performed the two undemanding visual tasks(passive viewing) and when they performed the two demanding hallucination tasks (active hallucination).
No significant group (high versus low suggestible and vice versa) differences in brain activity were found between scansacquired in and out of hypnosis during the passive viewing tasks. For the active tasks, direct group (high versus low suggest-ible and vice versa) comparisons showed that, when under hypnosis, the high suggestible participants had significantly low-er levels of activation in the right middle and superior frontal gyri and in the caudate nucleus than the low suggestibleparticipants (see Table 1d). High suggestible people also had significantly higher levels of activation in the right fusiformand lingual gyrus (see Table 1e).
To test whether the reductions that occurred in the high suggestible people when resting under hypnosis were constantacross conditions, the voxel that had the greatest reduction in activity (identified in the no hypnosis rest versus hypnosis restcomparison) was determined and the magnitude of the difference in signal in this voxel was then measured across the pas-sive and active tasks, in both high and low suggestible participants. Fig. 2 shows that in the high suggestible participants (butnot in the lows) differences between the hypnosis and no hypnosis periods appeared to decrease as task demands increased.
4. Discussion
This fMRI study of resting brain activity showed that brain activity decreased significantly in the anterior part of the ‘de-fault mode’ network (prefrontal cortex) in high suggestible participants when hypnotized. No areas had significant increasesin activation in this group when in hypnosis. Reduction of spontaneous brain activation in the dorsal and ventromedial pre-frontal cortex was not observed in low suggestible people after the hypnotic induction, nor was any area of increased acti-vation detected.
Normally, these modifications in the spontaneous physiological resting brain activity, which occurred only in high sug-gestible individuals after hypnotic induction, would be observable only when people engage in an externally orientated task.In that case, however, reduced activity in anterior default mode areas should also be accompanied by decreases in the pos-terior section of the default mode network, in addition to concurrent increases in activation in areas related to the task theyare attending to. To our knowledge, this study is the first to have observed a reduction in spontaneous default brain activity
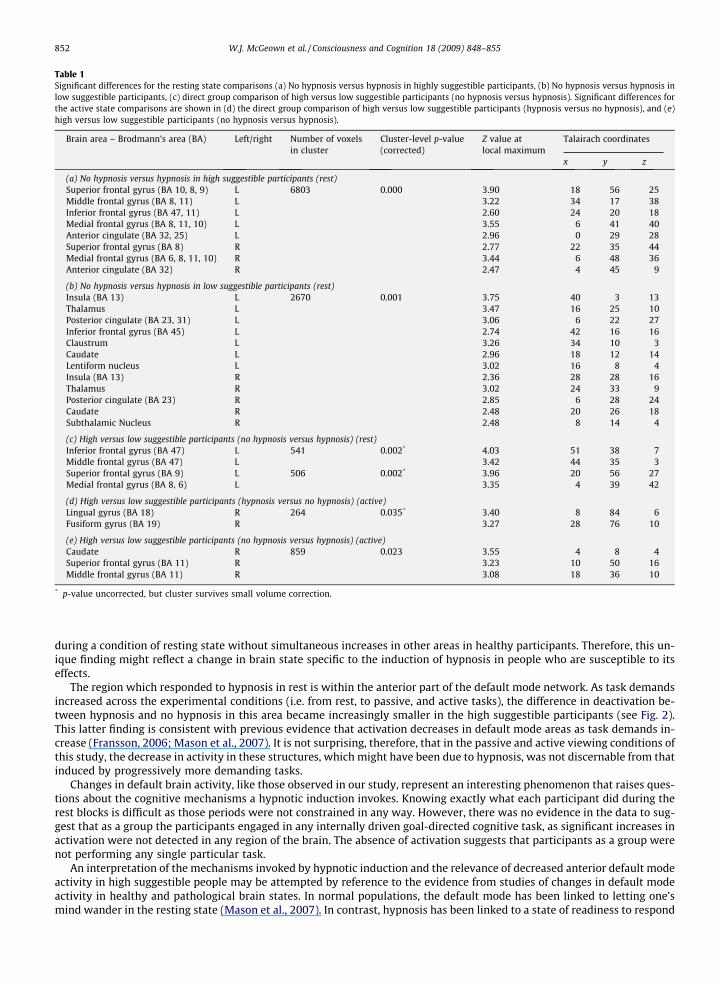
Table 1Significant differences for the resting state comparisons (a) No hypnosis versus hypnosis in highly suggestible participants, (b) No hypnosis versus hypnosis inlow suggestible participants, (c) direct group comparison of high versus low suggestible participants (no hypnosis versus hypnosis). Significant differences forthe active state comparisons are shown in (d) the direct group comparison of high versus low suggestible participants (hypnosis versus no hypnosis), and (e)high versus low suggestible participants (no hypnosis versus hypnosis).
Brain area – Brodmann’s area (BA) Left/right Number of voxelsin cluster
Cluster-level p-value(corrected)
Z value atlocal maximum
Talairach coordinates
x y z
(a) No hypnosis versus hypnosis in high suggestible participants (rest)Superior frontal gyrus (BA 10, 8, 9) L 6803 0.000 3.90 �18 56 25Middle frontal gyrus (BA 8, 11) L 3.22 �34 17 38Inferior frontal gyrus (BA 47, 11) L 2.60 �24 20 �18Medial frontal gyrus (BA 8, 11, 10) L 3.55 �6 41 40Anterior cingulate (BA 32, 25) L 2.96 0 29 28Superior frontal gyrus (BA 8) R 2.77 22 35 44Medial frontal gyrus (BA 6, 8, 11, 10) R 3.44 6 48 36Anterior cingulate (BA 32) R 2.47 4 45 9
(b) No hypnosis versus hypnosis in low suggestible participants (rest)Insula (BA 13) L 2670 0.001 3.75 �40 �3 13Thalamus L 3.47 �16 �25 10Posterior cingulate (BA 23, 31) L 3.06 �6 �22 27Inferior frontal gyrus (BA 45) L 2.74 �42 16 16Claustrum L 3.26 �34 �10 �3Caudate L 2.96 �18 12 14Lentiform nucleus L 3.02 �16 �8 4Insula (BA 13) R 2.36 28 �28 16Thalamus R 3.02 24 �33 9Posterior cingulate (BA 23) R 2.85 6 �28 24Caudate R 2.48 20 �26 18Subthalamic Nucleus R 2.48 8 �14 �4
(c) High versus low suggestible participants (no hypnosis versus hypnosis) (rest)Inferior frontal gyrus (BA 47) L 541 0.002* 4.03 �51 38 �7Middle frontal gyrus (BA 47) L 3.42 �44 35 �3Superior frontal gyrus (BA 9) L 506 0.002* 3.96 �20 56 27Medial frontal gyrus (BA 8, 6) L 3.35 �4 39 42
(d) High versus low suggestible participants (hypnosis versus no hypnosis) (active)Lingual gyrus (BA 18) R 264 0.035* 3.40 8 �84 �6Fusiform gyrus (BA 19) R 3.27 28 �76 �10
(e) High versus low suggestible participants (no hypnosis versus hypnosis) (active)Caudate R 859 0.023 3.55 4 8 �4Superior frontal gyrus (BA 11) R 3.23 10 50 �16Middle frontal gyrus (BA 11) R 3.08 18 36 �10
* p-value uncorrected, but cluster survives small volume correction.
852 W.J. McGeown et al. / Consciousness and Cognition 18 (2009) 848–855
during a condition of resting state without simultaneous increases in other areas in healthy participants. Therefore, this un-ique finding might reflect a change in brain state specific to the induction of hypnosis in people who are susceptible to itseffects.
The region which responded to hypnosis in rest is within the anterior part of the default mode network. As task demandsincreased across the experimental conditions (i.e. from rest, to passive, and active tasks), the difference in deactivation be-tween hypnosis and no hypnosis in this area became increasingly smaller in the high suggestible participants (see Fig. 2).This latter finding is consistent with previous evidence that activation decreases in default mode areas as task demands in-crease (Fransson, 2006; Mason et al., 2007). It is not surprising, therefore, that in the passive and active viewing conditions ofthis study, the decrease in activity in these structures, which might have been due to hypnosis, was not discernable from thatinduced by progressively more demanding tasks.
Changes in default brain activity, like those observed in our study, represent an interesting phenomenon that raises ques-tions about the cognitive mechanisms a hypnotic induction invokes. Knowing exactly what each participant did during therest blocks is difficult as those periods were not constrained in any way. However, there was no evidence in the data to sug-gest that as a group the participants engaged in any internally driven goal-directed cognitive task, as significant increases inactivation were not detected in any region of the brain. The absence of activation suggests that participants as a group werenot performing any single particular task.
An interpretation of the mechanisms invoked by hypnotic induction and the relevance of decreased anterior default modeactivity in high suggestible people may be attempted by reference to the evidence from studies of changes in default modeactivity in healthy and pathological brain states. In normal populations, the default mode has been linked to letting one’smind wander in the resting state (Mason et al., 2007). In contrast, hypnosis has been linked to a state of readiness to respond
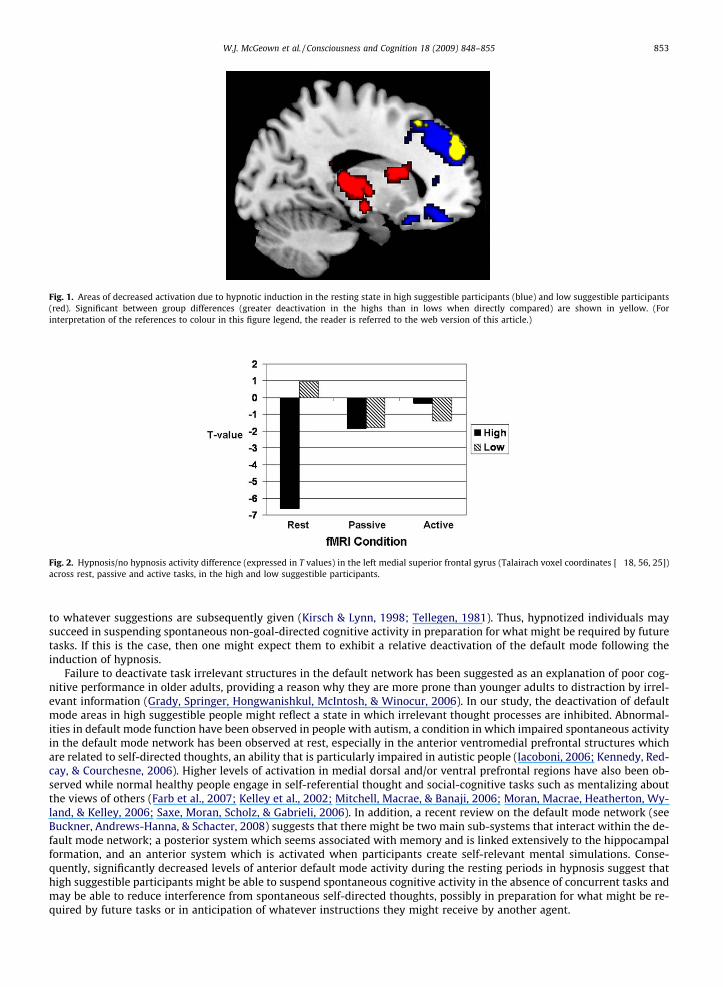
Fig. 1. Areas of decreased activation due to hypnotic induction in the resting state in high suggestible participants (blue) and low suggestible participants(red). Significant between group differences (greater deactivation in the highs than in lows when directly compared) are shown in yellow. (Forinterpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
Fig. 2. Hypnosis/no hypnosis activity difference (expressed in T values) in the left medial superior frontal gyrus (Talairach voxel coordinates [�18, 56, 25])across rest, passive and active tasks, in the high and low suggestible participants.
W.J. McGeown et al. / Consciousness and Cognition 18 (2009) 848–855 853
to whatever suggestions are subsequently given (Kirsch & Lynn, 1998; Tellegen, 1981). Thus, hypnotized individuals maysucceed in suspending spontaneous non-goal-directed cognitive activity in preparation for what might be required by futuretasks. If this is the case, then one might expect them to exhibit a relative deactivation of the default mode following theinduction of hypnosis.
Failure to deactivate task irrelevant structures in the default network has been suggested as an explanation of poor cog-nitive performance in older adults, providing a reason why they are more prone than younger adults to distraction by irrel-evant information (Grady, Springer, Hongwanishkul, McIntosh, & Winocur, 2006). In our study, the deactivation of defaultmode areas in high suggestible people might reflect a state in which irrelevant thought processes are inhibited. Abnormal-ities in default mode function have been observed in people with autism, a condition in which impaired spontaneous activityin the default mode network has been observed at rest, especially in the anterior ventromedial prefrontal structures whichare related to self-directed thoughts, an ability that is particularly impaired in autistic people (Iacoboni, 2006; Kennedy, Red-cay, & Courchesne, 2006). Higher levels of activation in medial dorsal and/or ventral prefrontal regions have also been ob-served while normal healthy people engage in self-referential thought and social-cognitive tasks such as mentalizing aboutthe views of others (Farb et al., 2007; Kelley et al., 2002; Mitchell, Macrae, & Banaji, 2006; Moran, Macrae, Heatherton, Wy-land, & Kelley, 2006; Saxe, Moran, Scholz, & Gabrieli, 2006). In addition, a recent review on the default mode network (seeBuckner, Andrews-Hanna, & Schacter, 2008) suggests that there might be two main sub-systems that interact within the de-fault mode network; a posterior system which seems associated with memory and is linked extensively to the hippocampalformation, and an anterior system which is activated when participants create self-relevant mental simulations. Conse-quently, significantly decreased levels of anterior default mode activity during the resting periods in hypnosis suggest thathigh suggestible participants might be able to suspend spontaneous cognitive activity in the absence of concurrent tasks andmay be able to reduce interference from spontaneous self-directed thoughts, possibly in preparation for what might be re-quired by future tasks or in anticipation of whatever instructions they might receive by another agent.

854 W.J. McGeown et al. / Consciousness and Cognition 18 (2009) 848–855
Decreased activity confined to the anterior cingulate cortex within the anterior default mode network (a region in whichdecreased activity at rest was seen in this study), has been previously reported in investigations of brain activity during hyp-nosis (Raz, Fan, & Posner, 2005), but the reported differences were detected while participants exhibited decreased suscep-tibility to the Stroop interference effect by interpreting visual words as nonsense strings of letters. The finding wasinterpreted as evidence that the suggestion given in hypnosis decreased activation in brain areas involved in conflict reso-lution (Oakley, 2008; Raz et al., 2005). This explanation cannot apply to the finding of decreases in the anterior cingulatecortex during rest obtained in the current study, as the effect of suggestion was excluded (i.e., it was the same for the hyp-nosis and no hypnosis condition).
The decreased activation in the anterior cingulate during hypnosis has been discussed in the present paper primarily interms of its role in the default mode network. However, activity in anterior cingulate cortex has also been detected in studiesof conflict resolution, error detection and inhibition (Braver, Barch, Gray, Molfese, & Snyder, 2001; Kiehl, Liddle, & Hopfinger,2000; Lutcke & Frahm, 2008; Pardo, Pardo, Janer, & Raichle, 1990). Nevertheless, our findings cannot be completely explainedin terms of these functions, given the extent of decrease observed in other parts of the anterior default mode network. In-stead, the extent of changes in these anterior regions during hypnosis suggests that modulation of a broader area is occur-ring, such as that involved in the self-referential anterior default mode processing system.
Low suggestible people differed from high suggestible individuals in the pattern of changes of rest brain activity duringhypnosis. Instead of deactivation in the default mode regions, after hypnotic induction, the low suggestible group showedlowering of activation in the thalamus. Previous evidence shows that changes in thalamic activity are associated withchanges in alertness (Sturm & Willmes, 2001) and activation in this area is reduced when people are under general anaes-thesia (White & Alkire, 2003). This finding implies that low suggestible individuals enter a state of decreased alertness, whichis associated with the thalamus. The effect of hypnotic induction on thalamic activity in the low suggestible participantsmight be due to the component of the induction which suggested relaxation. Furthermore, the absence of thalamic decreasesin the high suggestible participants when resting under hypnosis suggests that the induced state is not simply one of relax-ation or of a lowered level of alertness.
It is important to note that our findings do not resolve the altered state controversy. Although they are consistent withaltered state theories, they are also consistent with Wagstaff’s (1998) contention that the induced ‘state’ is an epiphenom-enon. According to Wagstaff, suggestible subjects respond to the suggestion to experience a hypnotic state, just like they doto any other suggestion, and this experience is bound to have neurological correlates. To support the altered state theory, onewould have to show a causal connection between the induced state and response to suggestions (Lynn, Kirsch, Knox, Fassler,& Lilienfeld, 2007). That is, it would need to be shown that reducing default mode activity prior to the administration of hyp-notic suggestions enhances responsiveness to those suggestions. Also, our study did not include any participants in the med-ium range of suggestibility. This is important to consider because differences between high and low suggestible individualscould be due to characteristics of low suggestible participants (e.g., low task involvement, anxiety about hypnosis, disinter-est) rather than ‘‘state alterations” in the highly suggestible subjects. Indeed, our finding that low suggestibles exhibitedsigns of decreased alertness is consistent with motivational differences across the suggestibility groups. Future studies onthe hypothesized hypnotic state could benefit from the inclusion of moderately suggestible participants.
In conclusion, by using a refined experimental design (and a direct within subject comparison of rest epochs across scans,contrasting hypnosis versus non hypnosis directly rather than rest versus another within scan task), this fMRI study hasshown that highly suggestible people suspend brain activity in the anterior system of the default mode network in rest fol-lowing hypnotic induction while this is not the case for low suggestible people.
Acknowledgments
The authors thank L. Nocetti, P. Nichelli, L. Foan and K. Roberts for their various contributions to this study. This study waspartially funded by grants from MIUR to AV and from the BBC to IK and GM.
References
Braver, T. S., Barch, D. M., Gray, J. R., Molfese, D. L., & Snyder, A. (2001). Anterior cingulate cortex and response conflict: Effects of frequency, inhibition anderrors. Cerebral Cortex, 11(9), 825–836.
Buckner, R. L., Andrews-Hanna, J. R., & Schacter, D. L. (2008). The brain’s default network: Anatomy, function, and relevance to disease. Annals of the NewYork Academy of Sciences, 1124, 1–38.
Comey, G., & Kirsch, I. (1999). Intentional and spontaneous imagery in hypnosis: The phenomenology of hypnotic responding. International Journal of Clinicaland Experimental Hypnosis, 47(1), 65–85.
Egner, T., Jamieson, G., & Gruzelier, J. (2005). Hypnosis decouples cognitive control from conflict monitoring processes of the frontal lobe. Neuroimage, 27(4),969–978.
Farb, N. A., Segal, Z. V., Mayberg, H., Bean, J., McKeon, D., Fatima, Z., et al (2007). Attending to the present: Mindfulness meditation reveals distinct neuralmodes of self-reference. Social Cognitive and Affective Neuroscience, 2(4), 313–322.
Faymonville, M. E., Laureys, S., Degueldre, C., DelFiore, G., Luxen, A., Franck, G., et al (2000). Neural mechanisms of antinociceptive effects of hypnosis.Anesthesiology, 92(5), 1257–1267.
Fox, M. D., & Raichle, M. E. (2007). Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nature ReviewsNeuroscience, 8(9), 700–711.
Fransson, P. (2006). How default is the default mode of brain function? Further evidence from intrinsic BOLD signal fluctuations. Neuropsychologia, 44(14),2836–2845.

W.J. McGeown et al. / Consciousness and Cognition 18 (2009) 848–855 855
Grady, C. L., Springer, M. V., Hongwanishkul, D., McIntosh, A. R., & Winocur, G. (2006). Age-related changes in brain activity across the adult lifespan. Journalof Cognitive Neuroscience, 18(2), 227–241.
Grond, M., Pawlik, G., Walter, H., Lesch, O. M., & Heiss, W. D. (1995). Hypnotic catalepsy-induced changes of regional cerebral glucose metabolism. PsychiatryResearch, 61(3), 173–179.
Hilgard, E. R., & Tart, C. T. (1966). Responsiveness to suggestions following waking and imagination instructions and following induction of hypnosis. Journalof Abnormal Psychology, 71, 196–208.
Iacoboni, M. (2006). Failure to deactivate in autism: The co-constitution of self and other. Trends in Cognitive Sciences, 10(10), 431–433.Kelley, W. M., Macrae, C. N., Wyland, C. L., Caglar, S., Inati, S., & Heatherton, T. F. (2002). Finding the self? An event-related fMRI study. Journal of Cognitive
Neuroscience, 14(5), 785–794.Kennedy, D. P., Redcay, E., & Courchesne, E. (2006). Failing to deactivate: Resting functional abnormalities in autism. Proceedings of the National Academy of
Sciences of the United States of America, 103(21), 8275–8280.Kiehl, K. A., Liddle, P. F., & Hopfinger, J. B. (2000). Error processing and the rostral anterior cingulate: An event-related fMRI study. Psychophysiology, 37(2),
216–223.Kirsch, I., & Lynn, S. J. (1998). Hypnotic involuntariness and the automaticity of everyday life. In I. Kirsch, A. Capafons, E. Cardena, & S. Amigo (Eds.), Clinical
hypnosis and self regulation: Cognitive behavioral perspectives. Washington, DC: American Psychological Association.Kirsch, I., Lynn, S. J., & Rhue, J. W. (1993). Introduction to clinical hypnosis. In J. W. Rhue, S. J. Lynn, & I. Kirsch (Eds.), Handbook of clinical hypnosis (pp. 3–22).
Washington, DC: American Psychological Association.Lancaster, J. L., Woldorff, M. G., Parsons, L. M., Liotti, M., Freitas, C. S., Rainey, L., et al (2000). Automated Talairach atlas labels for functional brain mapping.
Human Brain Mapping, 10(3), 120–131.Lutcke, H., & Frahm, J. (2008). Lateralized anterior cingulate function during error processing and conflict monitoring as revealed by high-resolution fMRI.
Cerebral Cortex, 18(3), 508–515.Lynn, S. J., Kirsch, I., & Hallquist, M. N. (2008). Social cognitive theories of hypnosis. In M. R. Nash & A. Barnier (Eds.), Oxford handbook of hypnosis
(pp. 111–139). Oxford: Oxford University Press.Lynn, S. J., Kirsch, I., Knox, J., Fassler, O., & Lilienfeld, S. O. (2007). Hypnosis and neuroscience. Implications for the altered state debate. In G. A. Jamieson (Ed.),
Hypnosis and conscious states. Oxford: Oxford University Press.Maquet, P., Faymonville, M. E., Degueldre, C., Delfiore, G., Franck, G., Luxen, A., et al (1999). Functional neuroanatomy of hypnotic state. Biological Psychiatry,
45(3), 327–333.Mason, M. F., Norton, M. I., Van Horn, J. D., Wegner, D. M., Grafton, S. T., & Macrae, C. N. (2007). Wandering minds: The default network and stimulus-
independent thought. Science, 315(5810), 393–395.Mazzoni, G., Rotriquenz, E., Carvalho, C., Vannucci, M., Roberts, K., & Kirsch, I. (2009). Suggested visual hallucinations in and out of hypnosis. Consciousness
and Cognition, 18, 494–499.Mitchell, J. P., Macrae, C. N., & Banaji, M. R. (2006). Dissociable medial prefrontal contributions to judgments of similar and dissimilar others. Neuron, 50(4),
655–663.Moran, J. M., Macrae, C. N., Heatherton, T. F., Wyland, C. L., & Kelley, W. M. (2006). Neuroanatomical evidence for distinct cognitive and affective components
of self. Journal of Cognitive Neuroscience, 18(9), 1586–1594.Oakley, D. A. (2008). Hypnosis, trance and suggestion: Evidence from neuroimaging. In M. R. Nash & A. Barnier (Eds.), Oxford handbook of hypnosis
(pp. 365–392). Oxford: Oxford University Press.Oakley, D. A., & Halligan, P. W. (2009). Hypnotic suggestion and cognitive neuroscience. Trends in Cognitive Sciences, 13(6), 264–270.Pardo, J. V., Pardo, P. J., Janer, K. W., & Raichle, M. E. (1990). The anterior cingulate cortex mediates processing selection in the Stroop attentional conflict
paradigm. Proceedings of the National Academy of Sciences of the United States of America, 87(1), 256–259.Raichle, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J., Gusnard, D. A., & Shulman, G. L. (2001). A default mode of brain function. Proceedings of the
National Academy of Sciences of the United States of America, 98(2), 676–682.Rainville, P., Hofbauer, R. K., Bushnell, M. C., Duncan, G. H., & Price, D. D. (2002). Hypnosis modulates activity in brain structures involved in the regulation of
consciousness. Journal of Cognitive Neuroscience, 14(6), 887–901.Rainville, P., Hofbauer, R. K., Paus, T., Duncan, G. H., Bushnell, M. C., & Price, D. D. (1999). Cerebral mechanisms of hypnotic induction and suggestion. Journal
of Cognitive Neuroscience, 11(1), 110–125.Raz, A., Fan, J., & Posner, M. I. (2005). Hypnotic suggestion reduces conflict in the human brain. Proceedings of the National Academy of Sciences of the United
States of America, 102(28), 9978–9983.Saxe, R., Moran, J. M., Scholz, J., & Gabrieli, J. (2006). Overlapping and non-overlapping brain regions for theory of mind and self reflection in individual
subjects. Social cognitive and affective neuroscience, 1(3), 229–234.Spanos, N. P., Radtke, H. L., Hodgins, D. C., Bertrand, L. D., Stam, H. J., & Dubreuil, D. L. (1983). The Carleton University responsiveness to suggestion scale:
Stability, reliability, and relationships with expectancy and ‘‘hypnotic experiences”. Psychological Reports, 53, 555–563.Sturm, W., & Willmes, K. (2001). On the functional neuroanatomy of intrinsic and phasic alertness. Neuroimage, 14(1 Pt 2), S76–84.Tellegen, A. (1981). Practicing the 2 Disciplines for relaxation and enlightenment – Comment on role of the feedback signal in electromyograph biofeedback
– the relevance of attention. Journal of Experimental Psychology-General, 110(2), 217–226.Wagstaff, G. F. (1998). The semantics and physiology of hypnosis as an altered state: Towards a definition of hypnosis. Contemporary Hypnosis, 15, 149–165.Weitzenhoffer, A. M., & Hilgard, E. R. (1962). Stanford hypnotic susceptibility scale: Form C. Palo Alto. CA: Consulting Psychologists Press.White, N. S., & Alkire, M. T. (2003). Impaired thalamocortical connectivity in humans during general-anesthetic-induced unconsciousness. Neuroimage, 19(2
Pt 1), 402–411.




















