
Hydraulic and chemical signalling in the regulation of stomatal conductance and plant water use in...
15
Hydraulic and chemical signalling in the regulation of stomatal conductance and plant water use in field grapevines growing under deficit irrigation M. Lucília Rodrigues A,D , Tiago P. Santos A , Ana P. Rodrigues A,B , Claudia R. de Souza B , Carlos M. Lopes A , João P. Maroco B,C , João S. Pereira A and M. Manuela Chaves A,B A Instituto Superior de Agronomia, Universidade Técnica de Lisboa, Tapada da Ajuda, 1349-017 Lisboa, Portugal. B Laboratório de Ecofisiologia Molecular, Instituto de Tecnologia Química e Biológica, Apartado 127, 2780-901 Oeiras, Portugal. C Instituto Superior de Psicologia Aplicada. Rua Jardim do Tabaco, 34, 1149-047 Lisboa, Portugal. D Corresponding author. Email: [email protected] Abstract. Effects of irrigation strategies on stomata and plant water use were studied in field-grown grapevines (Vitis vinifera L.). We assessed the importance of root-derived chemical signals vs. hydraulic signalling in stomatal regulation. The experiment included two treatments with the same water added to the soil (50% ET c ) applied either to the whole root system (DI) or to half of the roots, alternating irrigation side every 15 days (PRD). Well-watered plants (FI) (100% ET c ) and non- irrigated grapevines (NI) were also studied. Partial stomata closure occurred in both PRD and DI plants. [ABA] of xylem sap remained constant during the day and was maintained throughout the season, with higher values in NI plants. Xylem sap pH was not affected by soil water availability. A positive correlation between y pd and maximum g s was found, indicating that grapevine stomata strongly respond to plant water status. In contrast, ABA did not explain stomatal control at veraison. At mid-ripening g s was significantly correlated with ABA, apparently interacting with the rise in xylem sap pH. Therefore, our data suggest that hydraulic feedback and feed-forward root-to-shoot chemical signalling mechanisms might be involved in the control of stomata in response to decreased soil water availability, hydraulic signals playing the dominant role. Additional keywords: partial root drying, sap flow, stomata, Vitis vinifera L., water relations, xylem ABA. Introduction Predicted scenarios of climate change over the coming decades include global warming and shifts in the amount, seasonality and distribution of precipitation (Schultz 2000). The foreseen increase in the frequency of heat waves in summer and a lengthening of the dry season, particularly intense in the Mediterranean area, will lead to extended and severe drought events, requiring increases in crop irrigation. Although water supplement in winegrapes has only recently been introduced in the Iberian Peninsula, irrigation will become common practice to ensure the long-term sustainability of viticulture. Irrigation in grapevines has to ensure normal physiological activity and an adequate balance between vegetative and reproductive development, while preserving yield quality. Deficit irrigation strategies are being used to achieve these aims, while allowing an increase in plant water-use efficiency. Among the various deficit irrigation techniques regulated deficit irrigation (RDI) is largely used in warm regions and involves a suspension or reduction in water supply during some stages of the growing season to improve fruit quality (McCarthy 1997; Possingham 2000). Although some RDI studies in grapevines have been successful (Bravdo and Naor 1996; McCarthy et al. 2002), others have shown a loss in yield; thus, decreasing the efficiency of the applied water (Wample and Smithyman 2002). Recently, another deficit irrigation technique, partial root zone drying (PRD), has gained importance in vineyard management (Dry and Loveys 1999; Loveys et al. 2004). PRD irrigation is based on split-root experiments (Zhang et al. 1987; Gowing et al. 1990; Sobeih et al. 2004) where only a part of the root system is watered while the other part is allowed to dry. In theory, the watered parts of the root system allow for the maintenance of the water status in the shoots, while the dried split roots will produce chemical signals that are transmitted to the shoots with potential inhibitory effects on vegetative growth and stomatal conductance. Stomatal closure in the absence of leaf water deficit supports the concept that roots are the primary sensors of plant drought, regulating stomatal aperture through root-sourced signals transported via the xylem. Early evidence was gathered from apple seedlings where the excision of dehydrated roots allowed for the reopening of stomata (Gowing et al. 1990). A great deal of evidence highlights the importance of ABA in this signalling (reviewed in Davies et al. 1994; Dodd et al. 1996; Wilkinson and Davies 2002). Nevertheless, other compounds, including precursors of ABA (Netting et al. 1997; Sauter et al. 2002), or low concentrations of cytokinins (Shashidhar et al. 1996; Stoll et al. 2000; Hansen and CSIRO PUBLISHING www.publish.csiro.au/journals/fpb Functional Plant Biology, 2008, 35, 565–579 ȑ CSIRO 2008 10.1071/FP08004 1445-4408/08/070565
Transcript of Hydraulic and chemical signalling in the regulation of stomatal conductance and plant water use in...

Hydraulic and chemical signalling in the regulationof stomatal conductance and plant water use in fieldgrapevines growing under deficit irrigation
M. Lucília RodriguesA,D, Tiago P. SantosA, Ana P. RodriguesA,B, Claudia R. de SouzaB,Carlos M. LopesA, João P. MarocoB,C, João S. PereiraA and M. Manuela ChavesA,B
AInstituto Superior de Agronomia, Universidade Técnica de Lisboa, Tapada da Ajuda,1349-017 Lisboa, Portugal.
BLaboratório de Ecofisiologia Molecular, Instituto de Tecnologia Química e Biológica,Apartado 127, 2780-901 Oeiras, Portugal.
CInstituto Superior de Psicologia Aplicada. Rua Jardim do Tabaco, 34, 1149-047 Lisboa, Portugal.DCorresponding author. Email: [email protected]
Abstract. Effects of irrigation strategies on stomata and plant water use were studied in field-grown grapevines (VitisviniferaL.).We assessed the importance of root-derived chemical signals vs. hydraulic signalling in stomatal regulation. Theexperiment included two treatments with the samewater added to the soil (50%ETc) applied either to the whole root system(DI) or to half of the roots, alternating irrigation side every 15 days (PRD). Well-watered plants (FI) (100% ETc) and non-irrigated grapevines (NI) were also studied. Partial stomata closure occurred in both PRD andDI plants. [ABA] of xylem sapremained constant during the day and wasmaintained throughout the season, with higher values in NI plants. Xylem sap pHwas not affected by soil water availability. A positive correlation between ypd and maximum gs was found, indicating thatgrapevine stomata strongly respond to plant water status. In contrast, ABA did not explain stomatal control at veraison. Atmid-ripening gs was significantly correlated with ABA, apparently interacting with the rise in xylem sap pH. Therefore, ourdata suggest that hydraulic feedback and feed-forward root-to-shoot chemical signalling mechanisms might be involved inthe control of stomata in response to decreased soil water availability, hydraulic signals playing the dominant role.
Additional keywords: partial root drying, sap flow, stomata, Vitis vinifera L., water relations, xylem ABA.
Introduction
Predicted scenarios of climate change over the coming decadesinclude global warming and shifts in the amount, seasonality anddistributionof precipitation (Schultz2000).The foreseen increasein the frequency of heatwaves in summer and a lengthening of thedry season, particularly intense in the Mediterranean area, willlead to extended and severe drought events, requiring increases incrop irrigation. Although water supplement in winegrapes hasonly recently been introduced in the Iberian Peninsula, irrigationwill become common practice to ensure the long-termsustainability of viticulture. Irrigation in grapevines has toensure normal physiological activity and an adequate balancebetween vegetative and reproductive development, whilepreserving yield quality. Deficit irrigation strategies are beingused to achieve these aims, while allowing an increase in plantwater-use efficiency.
Among the various deficit irrigation techniques regulateddeficit irrigation (RDI) is largely used in warm regions andinvolves a suspension or reduction in water supply duringsome stages of the growing season to improve fruit quality(McCarthy 1997; Possingham 2000). Although some RDIstudies in grapevines have been successful (Bravdo and Naor1996; McCarthy et al. 2002), others have shown a loss in yield;
thus, decreasing the efficiency of the applied water (Wample andSmithyman 2002). Recently, another deficit irrigation technique,partial root zonedrying (PRD), hasgained importance in vineyardmanagement (Dry and Loveys 1999; Loveys et al. 2004). PRDirrigation is based on split-root experiments (Zhang et al. 1987;Gowing et al. 1990; Sobeih et al. 2004) where only a part of theroot system is watered while the other part is allowed to dry. Intheory, the watered parts of the root system allow for themaintenance of the water status in the shoots, while the driedsplit rootswill produce chemical signals that are transmitted to theshoots with potential inhibitory effects on vegetative growth andstomatal conductance. Stomatal closure in the absence of leafwater deficit supports the concept that roots are the primarysensors of plant drought, regulating stomatal aperture throughroot-sourced signals transported via the xylem. Early evidencewas gathered from apple seedlings where the excision ofdehydrated roots allowed for the reopening of stomata(Gowing et al. 1990). A great deal of evidence highlights theimportance of ABA in this signalling (reviewed in Davies et al.1994; Dodd et al. 1996; Wilkinson and Davies 2002).Nevertheless, other compounds, including precursors of ABA(Netting et al. 1997; Sauter et al. 2002), or low concentrations ofcytokinins (Shashidhar et al. 1996; Stoll et al. 2000; Hansen and
CSIRO PUBLISHING
www.publish.csiro.au/journals/fpb Functional Plant Biology, 2008, 35, 565–579
� CSIRO 2008 10.1071/FP08004 1445-4408/08/070565

Dorffling 2003) or changes in the mineral composition or pH ofthe xylem (Wilkinson and Davies 1997; Hartung et al. 1998;Prokic et al. 2006; Jia andDavies 2007)might also be involved inthe regulation of water use at the leaf level.
Under controlled conditions, using plants with split roots(Gowing et al. 1990) or plants placed in a root pressure vesselto maintain a high plant water status (Gollan et al. 1986), straightchemical regulation of stomatal behaviour in drought-affectedplants was observed. However, when plants are growing underfield conditions they are slowly subjected to reductions in soilwater availability, and shoot water relations generally decreaseand interfere with plant physiological activity. Under thesecircumstances, both hydraulic and chemical signals arethought to co-regulate stomata and growth responses todrought (Davies et al. 1994; Dodd et al. 1996; Liu et al.2003). In fact, studies where stomatal conductance wasnegatively correlated with the ABA concentration in the xylemhave shown that the relationship is affected by the time of day(Correia et al. 1995; Tardieu et al. 1996) or by the intensity of thewater stress (Tardieu andDavies 1992;Correia andPereira 1995),suggesting an interaction with leaf water status. Nevertheless, therelative importance of the two types of signalling in stomatalcontrol is still a subject of discussion, with some studiessuggesting that hydraulic limitation rather than chemicalsignalling may dominate (Saliendra et al. 1995; Fuchs andLivingston 1996; Comstock 2002), depending on the speciesand/or experimental conditions. This will be the case in somewoody species where root-to-shoot signalling seems to be a lessefficient mechanism to control stomatal behaviour (Augé andMoore 2002) or when other abiotic stresses co-occur withdrought, as usually happens when plants are growing in theirnatural environment.
Our previous field studies using grapevine (Vitis vinifera L.)cv. ‘Trincadeira preta’ grown under different soil wateravailabilities have shown that differences in daily maximumleaf conductance were closely related to changes in ABAconcentration in the xylem sap (Correia et al. 1995). Recentwork in two grapevine varieties (cv. Grenache and cv. Syrah) hasprovided evidence of their contrasting stomatal sensitivity to soilwater stress,whichwasexplainedbydifferences in their hydraulicconductances (Schultz 2003). When comparing stomatalresponses to atmospheric moisture stress (which is the usualcofactor during water deficit) for these same varieties, Soaret al. (2006) also found that Grenache had a higher stomatalsensitivity compared with Syrah, and this was associated withhigher levels of ABA in the xylem sap of Grenache.
In the present study, we evaluated whether hydraulic factors,determined using leaf water potential and turgor, or chemicalsignals (ABA concentration and pH of the xylem sap) aredominant in the regulation of stomatal behaviour in twovarieties of field-grown grapevines subjected to differentirrigation regimes.
Materials and methodsPlant material and location
The experiment was carried out during the 2002 growing seasonin a commercial vineyard located 70 km south-east of Lisbon atthe Centro Experimental de Pegões (30�380N and 8�390W). The
climate is typicallyMediterranean, with hot and dry summers andmild winters. The average annual rainfall (1955–1984) is703.5mm, 57% of which falls in autumn and winter. Thegrowing season of 2002 (June until the beginning ofSeptember) was very dry, with rainfall of only 8.8mm; thetotal rainfall between January and September was 390mm(Santos et al. 2005). The soil is derived from podzols, mostlysandyandwith a clay-rich (lowpermeability) horizonat a depthof~1m. The grapevines studied, growing side by side, were Vitisvinifera L. cv. Castelão, an early-season red-wine variety and cv.Moscatel (syn. Muscat of Alexandria), a white variety; bothgrapevines were grafted onto 1103 Paulsen rootstock in 1996and 1997, respectively. The grapevines were spaced 2.5mbetween rows (oriented north–south) and 1.0m within rowsand trained on a bilateral Royat Cordon (16 buds per vine)system using two pairs of movable foliage wires for upwardsshoot positioning. The usual cultural practices of the region wereapplied to all treatments.
Irrigation treatments
Irrigation was applied by drip emitters, two per vine,independently controlled and positioned 0.30m from the vinetrunk, on both sides of the row. Thewater was supplied accordingto the crop evapotranspiration (ETc), estimated from the potentialevapotranspiration (ET0), calculated from Class A panevaporation and corrected with the crop coefficient (Kc), thatis, ETc = ET0�Kc. We used the most suitable Kc for ourconditions, which according to Prichard (1992) was 0.6 in Juneand0.7 in July andAugust. Each irrigation treatmentwas checkedby a time-clock-valve assembly to control the water supply.
The irrigation treatments were: fully irrigated (FI) (100% oftheETcwas suppliedbygiving50%toeach sideof the root systemat a rate of 4 L h�1 by dripper); deficit irrigated (DI) (50% of theETcwas supplied bygiving25% to each sideof the vine at a rate of2 L h�1 by dripper); partial root zone drying (PRD) (50% of theETc was supplied to only one side of the vine, with a 4 L h�1
dripper, while the other dripper was closed; the irrigated side wasalternated approximately every 15 days); non-irrigated (NI)(rainfed). Irrigation was applied twice per week from thebeginning of berry development (19 June) until 1week beforeharvest (3 September). The total amount of water supplied to theFI plants was 196.8mm (493 L per vine), and half of that volumewas applied to both the PRD and DI vines.
Physiological data for the present study were obtained fromplants belonging to two replicate plots from a Latin square withfour irrigation treatments and four replicates. Each treatment perplot had three rows with 15–20 grapevines in each row, and allmeasurements were made in the central row.
Plant water relations, soil water contentand stomatal conductance
Plant water relations parameters
Pre-dawn (ypd) and midday (ymd) leaf water potentials weremeasured weekly with a Scholander-type pressure chamber(Model 1000; PMS Instrument Company, Corvallis, OR,USA) from the beginning of berry development until harvest.On each sampling occasion, measurements were carried out onone fully expanded leaf from six replicate plants per treatment.
566 Functional Plant Biology M. L. Rodrigues et al.

The leaves were covered with plastic bags just after being cut tominimise water loss and were placed into the pressure chamberwhere the inside surface was lined with moist cotton. Soon aftermeasuringypd, leaf discs (0.7 cmdiameter) were taken from eachleaf, placed in Eppendorf vials and immediately plunged intoliquid nitrogen and stored at �80�C for later determination ofosmotic potential (yp). After thawing the samples for 10min atroom temperature,ypwasmeasured usingC52 chambers (2 h fortemperature and water vapour equilibration) connected to aWescor HR 33T dewpoint microvoltmeter (Wescor, Logan,UT, USA) operating in the dewpoint mode. Standard NaClsolutions were used for each chamber calibration, generating acalibration regression thatwasused to convert themVoutput toypinMPa. Leaf turgor (yp) was estimated as the difference betweenypd and yp. Leaf solute content (Ns) was calculated asNs =�yp (V/RT), where R is the universal gas constant, T isleaf temperature and V is the total volume of water in the leaftissue, which was determined from another set of discs punchedfrom the same leaves. Vwas estimated as the difference betweenthe fresh and dry weights, determined after drying the discs at70�C for 24 h. Relative leaf solute content (SC) was calculated asNs divided by the dry weight.
Midday stem water potential (ystem) was measured using thepressure chamber on the shoot of the same plants used for theymd
measurements. Leaves (six per treatment) were bagged with bothplastic sheet and aluminium foil for at least 2 h beforemeasurement to prevent leaf transpiration and to enable leafand stem water equilibrium (Choné et al. 2001). For ystem
measurements, basal shaded leaves located near the trunk wereselected to avoid overheating during the bagging period.
Soil water content
Soil moisture was monitored twice per week (before and aftereach irrigation) during the growing season until harvest using aDiviner 2000 capacitance probe (Sentek EnvironmentalTechnologies, Stepney, Australia). Water content in the soilprofile was determined down to a depth of 1.5m at 0.1-mintervals using access tubes located 0.1m from the plant row.For details on the soil moisture evolution throughout the growingseason see Santos et al. (2005, 2007).
Leaf stomatal conductance
Diurnal time-courses of leaf water potential, leaf conductanceto water vapour and concentration of ABA in the xylem wereconducted on sunny and clear days in two periods: at veraison(25 July in Castelão and 29 July inMoscatel) and at mid-ripening(22 August in Castelão and 26 August in Moscatel). These datescorresponded to 3 days after irrigation, and for PRD plants tothe day before the alternation of the watered side (25 July), thethird day after the alternation (29 July) and 1week (22August) or10 days (26 August) after that change.
Leaf stomatal conductance (gs) was measured with a steady-state porometer (LI-1600; Li-cor, Lincoln, NE, USA) on theabaxial surface (hypostomatous leaves) of four fully expandedleaves (each leaf from a different plant) per treatment. The sensorhead was held according to the natural position and the angle ofthe leaf. At each sampling time the air temperature, relativehumidity and PPFD were also measured with the porometer;
the porometer chamber was acclimated for ~30min to ambientconditions before the measurements were conducted.
Plant transpirationIn Castelão, plant water loss by transpiration was evaluated
by sapflowmeasurementsmade inAugust (post-veraison period)using a Dynamax Flow 32-AO system (Dynamax, Houston, TX,USA). The equipment uses the constant-power heat-balancemethod (Baker and van Bavel 1987), allowing for directmeasurement of mass flow through the xylem in a continuousway on undisturbed plants. Stem flow gauges, SGA 10 (designedfor stems measuring 8–10 cm in diameter), were installed in thebasal part of one main representative shoot of two vines pertreatment according to the guidelines from the manufacturer.Rough bark and dead tissue were carefully removed and a thinlayer of silicone-grease-based electrical insulating compoundwas spread on the branch at the point where the gauge wasinstalled. The gauges and the sections of stem above andbelow them were covered with aluminium foil to minimiseexternally induced temperature gradients. The power supply tothe gauges, which was 4.0V, was switched off from 2 h beforesunrise until 2 h after sunset to avoid overheating the stem. Thethermal conductivity of the stem was assumed to be0.42Wm�1 K�1 (value for woody plants). The conductancevalue of each gauge, Ksh, was first determined from the valuesin the night period, checked weekly and manually adjusted.
The signals generated by the gauges were recordedevery 60 s and stored as 15min average values in a CR10Xdata logger (Campbell Scientific, Logan,Utah, USA). A softwarefilter eliminated spurious flow calculations at times of low or highflow conditions. Sap flow data were expressed on a leaf areabasis (gm�2 h�1) bydividing themeasured sapfluxdensity by theshoot leaf area above the gauge, which was determined withan area meter (LI 3000; Li-Cor) at the end of the measurementperiod. Daily water use per plant (L vine�1 day�1) wasestimated by integrating the sap flow values from each 15-minperiod and using the total foliar area per plant, determinedaccording to Santos et al. (2005). The volumetric flux pergrapevine was converted to mmday�1 using an area of 2.5m2
of soil surface per plant.
ABA measurements in the xylem sapAt each sampling time (early morning, midday and afternoon) onthe reported days (25 and 29 July and 22 and 26 August), oncompletion of the gs measurements, leaves were detached andplaced in a pressure chamber with the inside surface lined withmoist cotton to collect xylem sap. The petiole cut surface wascleaned with distilled water and dried with blotting paper. Afterslowly applying pressure until the water potential was equalised,thefirst twodroplets of the xylem sap expressedwere discarded toavoid any solute contamination from the phloemandparenchymacells disrupted at the cut surface. Then, using an overpressure of0.4MPa above the balancing pressure for 60–120 s, sap wascollected with a syringe and placed into an Eppendorf vial andstored in the dark. After the pH of the sap was measured using amicroelectrode interfaced with a pH meter (RL 100; Russel,Hanwell, London, UK) the samples were immediately frozenin liquid nitrogen and stored at �80�C until analysis for ABAconcentration ([ABA]).ABAwasquantified byanELISAusing a
Hydraulic and chemical signalling in grapevines Functional Plant Biology 567
monoclonal antibody for ABA (Phytodetek; Agdia, Elkhart, IN,USA). All assays were made in duplicate or triplicate fromsamples of sap without any previous purification. Nointerference of the antibody with other substances in thegrapevine xylem sap was detected when previously checked(see Correia et al. 1995). The flux of ABA delivered to theleaves was estimated from the [ABA] in the xylem multipliedby the transpiration rate.
Statistical analysis
Statistical data analysiswas carried out using one-wayANOVAs.Least significant differences (P< 0.05) and regression analyseswere carried out using the STATISTICA software (version 5.0;StatSoft, Tulsa,OK,USA). Toquantify the relative importance ofypd and [ABA] in the xylem on stomatal responses, a forwardstepwise regression between gs (dependent variable) andypd and[ABA] (independent variables) was carried out using a 0.15critical F statistic for variable selection. The slope andintercepts of the linear regressions of gs v. ypd and gs v.[ABA] obtained at veraison and mid-ripening were comparedusing adjusted Student’s t-tests (see Zar 1999). Allmeasurementsshown are means� s.e.
Results
Plant water relations
The ypd of the FI grapevines remained constant throughoutthe growing season and close to �0.22MPa in both varieties,whereas the ypd of the NI plants decreased from July onwards,attaining values of �0.78MPa in Castelão and �0.64MPa inMoscatel by the end ofAugust (Table 1). Approximately 3weeksafter the first watering, on 11 July (Castelão) and 8 July(Moscatel), ypd were already significantly (P< 0.001) higherin the FI grapevines and lower in NI vines. The treatmentswith deficit irrigation showed intermediate values comparedwith those of the FI and NI plants. For Castelão, the ypd
of the PRD grapevines was significantly higher than thatobserved in the DI plants, with the exception of themeasurement taken on 22 August. However, the morefavourable plant water status of the PRD Moscatelgrapevines as compared with the DI vines was only apparenton 8 July. After the beginning of irrigation (19 June) the middayleaf water potentials of the FI grapevines were alwayssignificantly higher than those of the NI vines (Table 1). Inboth varieties, the PRD plants had ymd values close to thoseof the FI grapevines and usually with a tendency towards highervalues than those of the DI plants. Nevertheless, there wereno differences between the PRD and DI vines on most of thesampled dates.
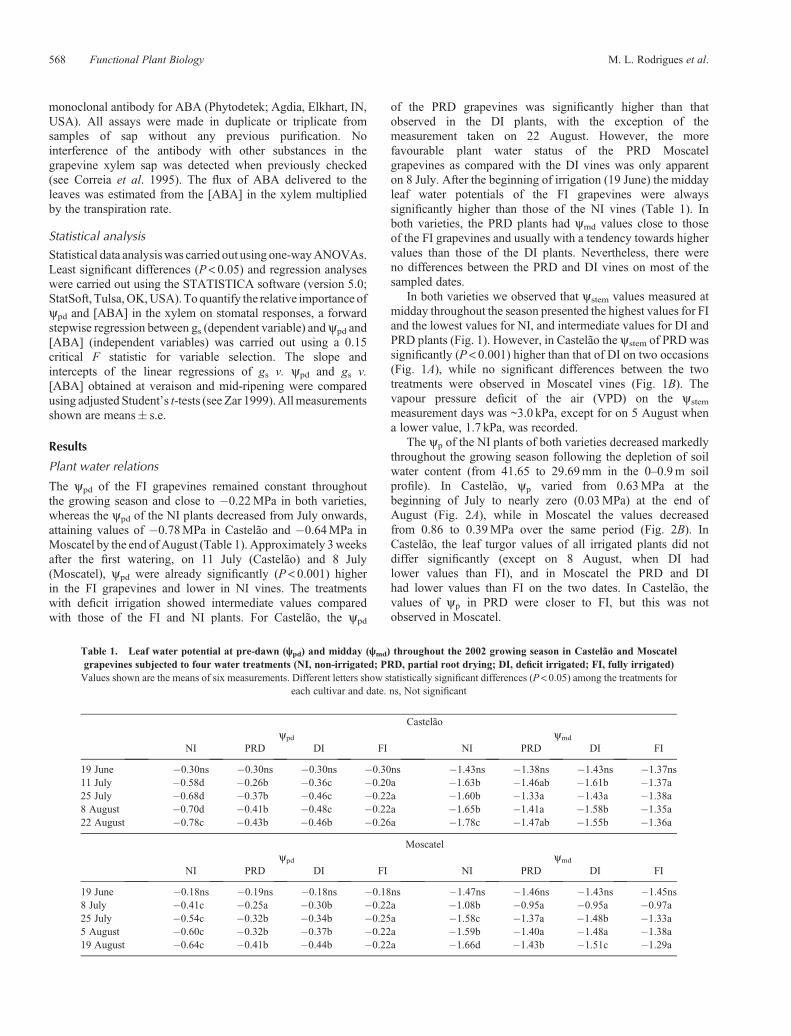
In both varieties we observed that ystem values measured atmidday throughout the season presented the highest values for FIand the lowest values for NI, and intermediate values for DI andPRD plants (Fig. 1). However, in Castelão the ystem of PRD wassignificantly (P < 0.001) higher than that of DI on two occasions(Fig. 1A), while no significant differences between the twotreatments were observed in Moscatel vines (Fig. 1B). Thevapour pressure deficit of the air (VPD) on the ystem
measurement days was ~3.0 kPa, except for on 5 August whena lower value, 1.7 kPa, was recorded.
The yp of the NI plants of both varieties decreased markedlythroughout the growing season following the depletion of soilwater content (from 41.65 to 29.69mm in the 0–0.9m soilprofile). In Castelão, yp varied from 0.63MPa at thebeginning of July to nearly zero (0.03MPa) at the end ofAugust (Fig. 2A), while in Moscatel the values decreasedfrom 0.86 to 0.39MPa over the same period (Fig. 2B). InCastelão, the leaf turgor values of all irrigated plants did notdiffer significantly (except on 8 August, when DI hadlower values than FI), and in Moscatel the PRD and DIhad lower values than FI on the two dates. In Castelão, thevalues of yp in PRD were closer to FI, but this was notobserved in Moscatel.
Table 1. Leaf water potential at pre-dawn (cpd) and midday (cmd) throughout the 2002 growing season in Castelão and Moscatelgrapevines subjected to four water treatments (NI, non-irrigated; PRD, partial root drying; DI, deficit irrigated; FI, fully irrigated)Values shown are the means of six measurements. Different letters show statistically significant differences (P< 0.05) among the treatments for
each cultivar and date. ns, Not significant
Castelãoypd ymd
NI PRD DI FI NI PRD DI FI
19 June �0.30ns �0.30ns �0.30ns �0.30ns �1.43ns �1.38ns �1.43ns �1.37ns11 July �0.58d �0.26b �0.36c �0.20a �1.63b �1.46ab �1.61b �1.37a25 July �0.68d �0.37b �0.46c �0.22a �1.60b �1.33a �1.43a �1.38a8 August �0.70d �0.41b �0.48c �0.22a �1.65b �1.41a �1.58b �1.35a22 August �0.78c �0.43b �0.46b �0.26a �1.78c �1.47ab �1.55b �1.36a
Moscatelypd ymd
NI PRD DI FI NI PRD DI FI
19 June �0.18ns �0.19ns �0.18ns �0.18ns �1.47ns �1.46ns �1.43ns �1.45ns8 July �0.41c �0.25a �0.30b �0.22a �1.08b �0.95a �0.95a �0.97a25 July �0.54c �0.32b �0.34b �0.25a �1.58c �1.37a �1.48b �1.33a5 August �0.60c �0.32b �0.37b �0.22a �1.59b �1.40a �1.48a �1.38a19 August �0.64c �0.41b �0.44b �0.22a �1.66d �1.43b �1.51c �1.29a
568 Functional Plant Biology M. L. Rodrigues et al.
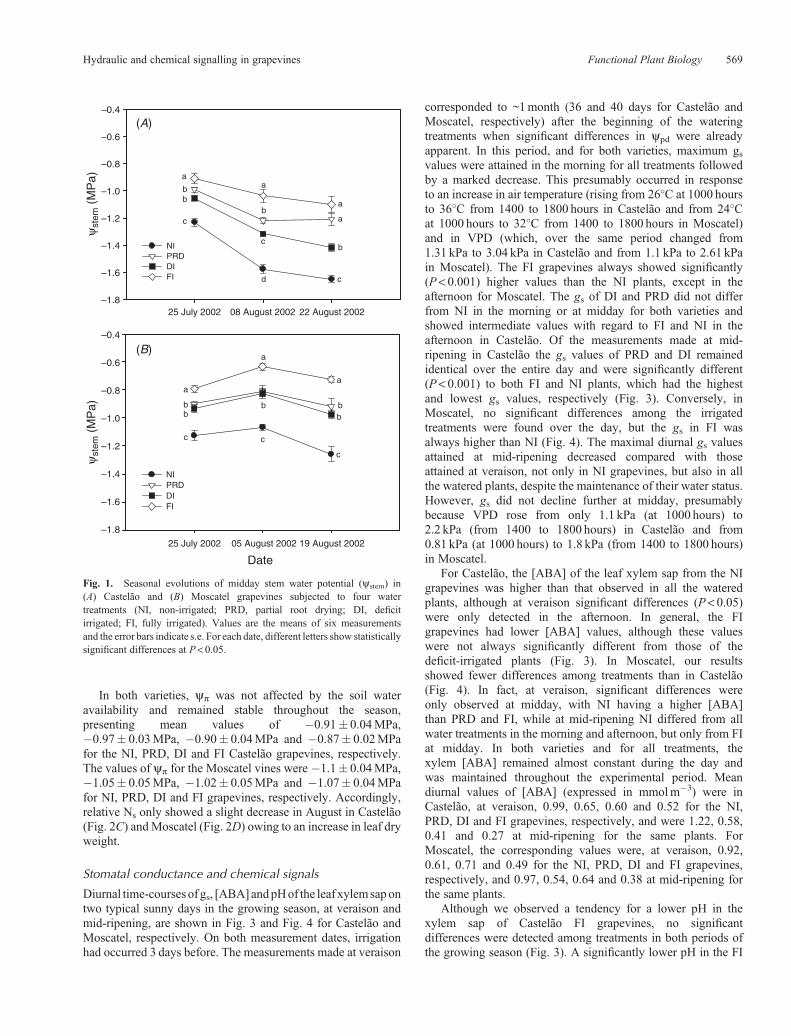
In both varieties, yp was not affected by the soil wateravailability and remained stable throughout the season,presenting mean values of �0.91� 0.04MPa,�0.97� 0.03MPa, �0.90� 0.04MPa and �0.87� 0.02MPafor the NI, PRD, DI and FI Castelão grapevines, respectively.The values of yp for the Moscatel vines were �1.1� 0.04MPa,�1.05� 0.05MPa, �1.02� 0.05MPa and �1.07� 0.04MPafor NI, PRD, DI and FI grapevines, respectively. Accordingly,relative Ns only showed a slight decrease in August in Castelão(Fig. 2C) andMoscatel (Fig. 2D) owing to an increase in leaf dryweight.
Stomatal conductance and chemical signals
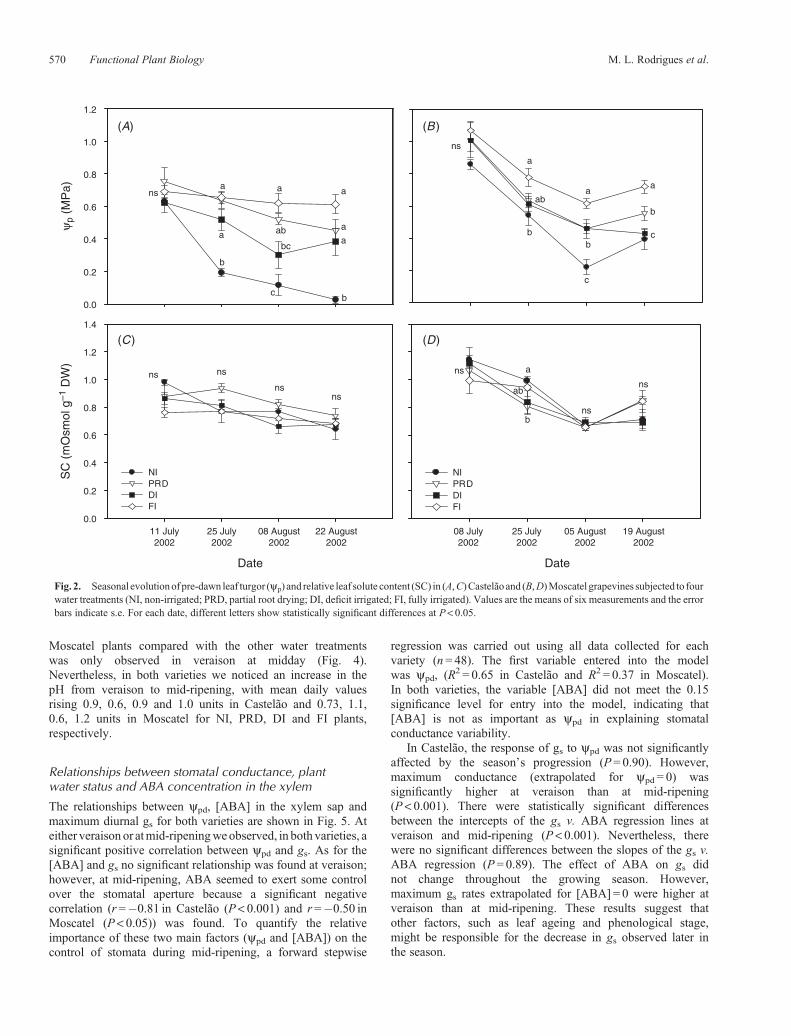
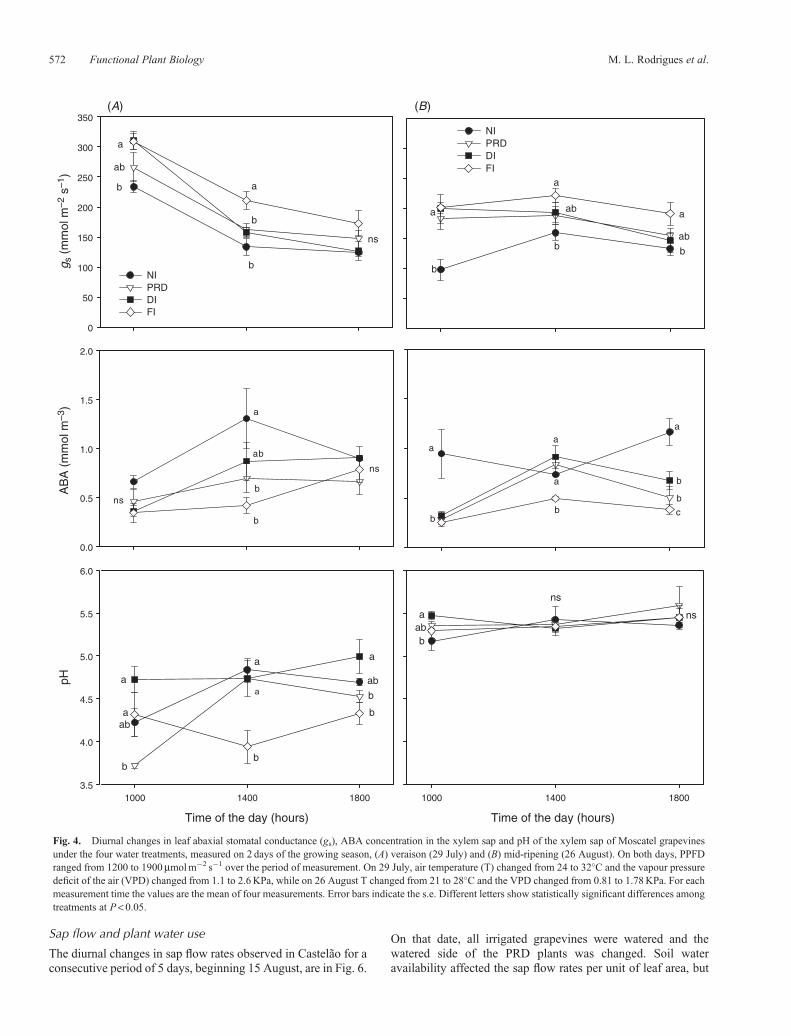
Diurnal time-courses of gs, [ABA] andpHof the leaf xylemsapontwo typical sunny days in the growing season, at veraison andmid-ripening, are shown in Fig. 3 and Fig. 4 for Castelão andMoscatel, respectively. On both measurement dates, irrigationhad occurred 3 days before. The measurements made at veraison
corresponded to ~1month (36 and 40 days for Castelão andMoscatel, respectively) after the beginning of the wateringtreatments when significant differences in ypd were alreadyapparent. In this period, and for both varieties, maximum gsvalues were attained in the morning for all treatments followedby a marked decrease. This presumably occurred in responseto an increase in air temperature (rising from 26�C at 1000 hoursto 36�C from 1400 to 1800 hours in Castelão and from 24�Cat 1000 hours to 32�C from 1400 to 1800 hours in Moscatel)and in VPD (which, over the same period changed from1.31 kPa to 3.04 kPa in Castelão and from 1.1 kPa to 2.61 kPain Moscatel). The FI grapevines always showed significantly(P< 0.001) higher values than the NI plants, except in theafternoon for Moscatel. The gs of DI and PRD did not differfrom NI in the morning or at midday for both varieties andshowed intermediate values with regard to FI and NI in theafternoon in Castelão. Of the measurements made at mid-ripening in Castelão the gs values of PRD and DI remainedidentical over the entire day and were significantly different(P< 0.001) to both FI and NI plants, which had the highestand lowest gs values, respectively (Fig. 3). Conversely, inMoscatel, no significant differences among the irrigatedtreatments were found over the day, but the gs in FI wasalways higher than NI (Fig. 4). The maximal diurnal gs valuesattained at mid-ripening decreased compared with thoseattained at veraison, not only in NI grapevines, but also in allthe watered plants, despite the maintenance of their water status.However, gs did not decline further at midday, presumablybecause VPD rose from only 1.1 kPa (at 1000 hours) to2.2 kPa (from 1400 to 1800 hours) in Castelão and from0.81 kPa (at 1000 hours) to 1.8 kPa (from 1400 to 1800 hours)in Moscatel.
For Castelão, the [ABA] of the leaf xylem sap from the NIgrapevines was higher than that observed in all the wateredplants, although at veraison significant differences (P< 0.05)were only detected in the afternoon. In general, the FIgrapevines had lower [ABA] values, although these valueswere not always significantly different from those of thedeficit-irrigated plants (Fig. 3). In Moscatel, our resultsshowed fewer differences among treatments than in Castelão(Fig. 4). In fact, at veraison, significant differences wereonly observed at midday, with NI having a higher [ABA]than PRD and FI, while at mid-ripening NI differed from allwater treatments in the morning and afternoon, but only from FIat midday. In both varieties and for all treatments, thexylem [ABA] remained almost constant during the day andwas maintained throughout the experimental period. Meandiurnal values of [ABA] (expressed in mmolm�3) were inCastelão, at veraison, 0.99, 0.65, 0.60 and 0.52 for the NI,PRD, DI and FI grapevines, respectively, and were 1.22, 0.58,0.41 and 0.27 at mid-ripening for the same plants. ForMoscatel, the corresponding values were, at veraison, 0.92,0.61, 0.71 and 0.49 for the NI, PRD, DI and FI grapevines,respectively, and 0.97, 0.54, 0.64 and 0.38 at mid-ripening forthe same plants.
Although we observed a tendency for a lower pH in thexylem sap of Castelão FI grapevines, no significantdifferences were detected among treatments in both periods ofthe growing season (Fig. 3). A significantly lower pH in the FI
–1.8
–1.6
–1.4
–1.2
–1.0
–0.8
–0.6
–0.4
–1.8
–1.6
–1.4
–1.2
–1.0
–0.8
–0.6
–0.4
NIPRDDIFI
aa
a
b
ba
b
cb
c
d c
25 July 2002 08 August 2002 22 August 2002
25 July 2002 05 August 2002 19 August 2002
ψst
em (
MP
a)ψ
stem
(M
Pa)
NIPRDDIFI
a
a
a
bb bb
c
b
c
b
c
Date
(B)
(A)
Fig. 1. Seasonal evolutions of midday stem water potential (ystem) in(A) Castelão and (B) Moscatel grapevines subjected to four watertreatments (NI, non-irrigated; PRD, partial root drying; DI, deficitirrigated; FI, fully irrigated). Values are the means of six measurementsand the error bars indicate s.e. For each date, different letters show statisticallysignificant differences at P< 0.05.
Hydraulic and chemical signalling in grapevines Functional Plant Biology 569
Moscatel plants compared with the other water treatmentswas only observed in veraison at midday (Fig. 4).Nevertheless, in both varieties we noticed an increase in thepH from veraison to mid-ripening, with mean daily valuesrising 0.9, 0.6, 0.9 and 1.0 units in Castelão and 0.73, 1.1,0.6, 1.2 units in Moscatel for NI, PRD, DI and FI plants,respectively.
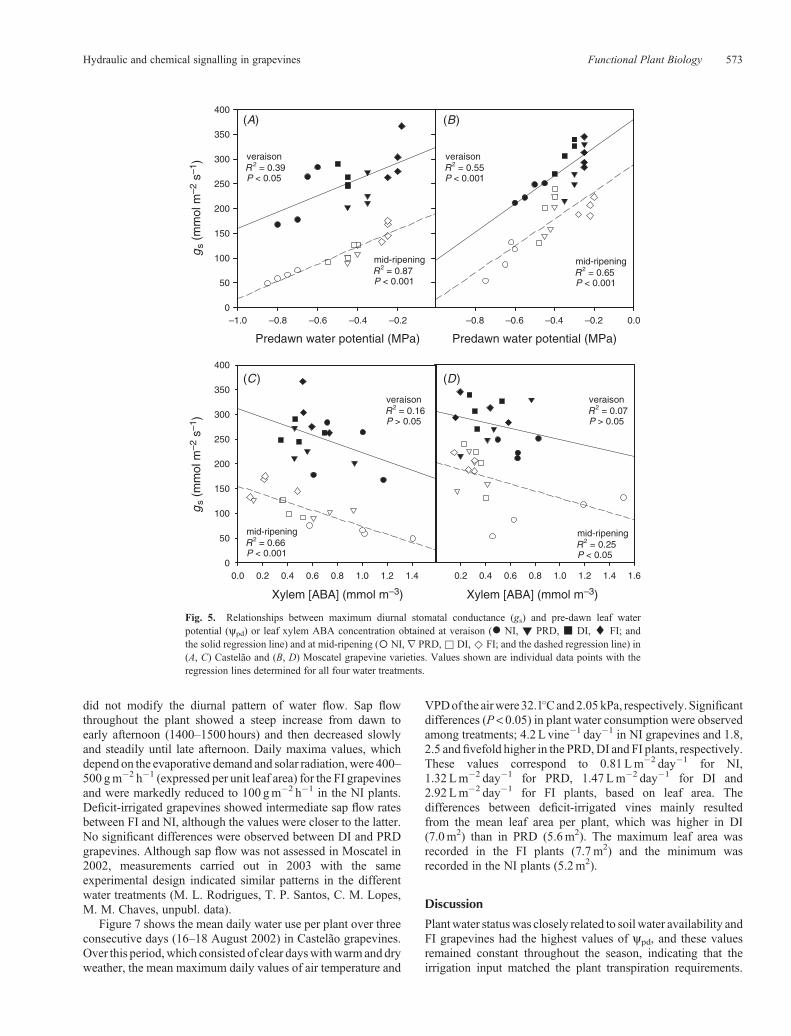
Relationships between stomatal conductance, plantwater status and ABA concentration in the xylem
The relationships between ypd, [ABA] in the xylem sap andmaximum diurnal gs for both varieties are shown in Fig. 5. Ateither veraison or atmid-ripeningweobserved, in both varieties, asignificant positive correlation between ypd and gs. As for the[ABA] and gs no significant relationship was found at veraison;however, at mid-ripening, ABA seemed to exert some controlover the stomatal aperture because a significant negativecorrelation (r=�0.81 in Castelão (P< 0.001) and r=�0.50 inMoscatel (P< 0.05)) was found. To quantify the relativeimportance of these two main factors (ypd and [ABA]) on thecontrol of stomata during mid-ripening, a forward stepwise
regression was carried out using all data collected for eachvariety (n= 48). The first variable entered into the modelwas ypd, (R
2 = 0.65 in Castelão and R2 = 0.37 in Moscatel).In both varieties, the variable [ABA] did not meet the 0.15significance level for entry into the model, indicating that[ABA] is not as important as ypd in explaining stomatalconductance variability.
In Castelão, the response of gs to ypd was not significantlyaffected by the season’s progression (P= 0.90). However,maximum conductance (extrapolated for ypd = 0) wassignificantly higher at veraison than at mid-ripening(P < 0.001). There were statistically significant differencesbetween the intercepts of the gs v. ABA regression lines atveraison and mid-ripening (P < 0.001). Nevertheless, therewere no significant differences between the slopes of the gs v.ABA regression (P = 0.89). The effect of ABA on gs didnot change throughout the growing season. However,maximum gs rates extrapolated for [ABA] = 0 were higher atveraison than at mid-ripening. These results suggest thatother factors, such as leaf ageing and phenological stage,might be responsible for the decrease in gs observed later inthe season.
NIPRDDIFI
Date
ns a
ns
nsab
b
11 July2002
25 July2002
08 August2002
22 August2002
08 July2002
25 July2002
05 August2002
19 August2002
SC
(m
Osm
ol g
–1 D
W)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
NIPRDDIFI
Date
ns ns
nsns
ns
a
aa
abb
bb
c
c
ψp
(MP
a) nsa a a
a
ab a
abc
b
cb
a
(A) (B )
(C) (D)
Fig. 2. Seasonal evolutionof pre-dawn leaf turgor (yp) and relative leaf solute content (SC) in (A,C)Castelão and (B,D)Moscatel grapevines subjected to fourwater treatments (NI, non-irrigated; PRD, partial root drying; DI, deficit irrigated; FI, fully irrigated). Values are the means of six measurements and the errorbars indicate s.e. For each date, different letters show statistically significant differences at P< 0.05.
570 Functional Plant Biology M. L. Rodrigues et al.
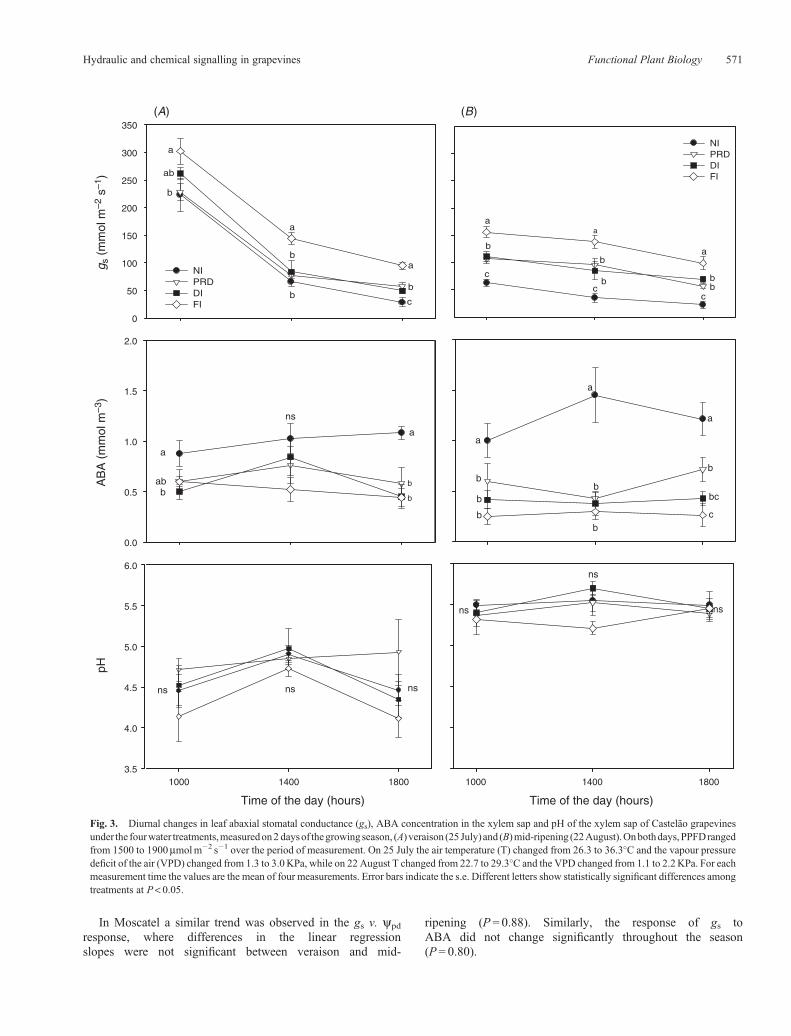
In Moscatel a similar trend was observed in the gs v. ypd
response, where differences in the linear regressionslopes were not significant between veraison and mid-
ripening (P = 0.88). Similarly, the response of gs toABA did not change significantly throughout the season(P= 0.80).
1000 1400 18001000 1400 1800
ns
ns
ns
a
b
b
b
a
b
b
a
b
bc
c
AB
A (
mm
ol m
–3)
0.0
0.5
1.0
1.5
2.0
a
abb
ns
a
b
b
NIPRDDIFI
a
b
c
a
b
bc
a
b
cb
g s (
mm
ol m
–2 s
–1)
0
50
100
150
200
250
300
350
NIPRDDIFI
a
ab
b
a
b
ba
b
c
Time of the day (hours) Time of the day (hours)
pH
3.5
4.0
4.5
5.0
5.5
6.0
ns ns ns
(A) (B)
Fig. 3. Diurnal changes in leaf abaxial stomatal conductance (gs), ABA concentration in the xylem sap and pH of the xylem sap of Castelão grapevinesunder the fourwater treatments,measuredon2 daysof thegrowingseason, (A) veraison (25 July)and (B)mid-ripening (22August).Onbothdays,PPFDrangedfrom 1500 to 1900mmolm�2 s�1 over the period of measurement. On 25 July the air temperature (T) changed from 26.3 to 36.3�C and the vapour pressuredeficit of the air (VPD) changed from 1.3 to 3.0KPa, while on 22 August T changed from 22.7 to 29.3�C and the VPD changed from 1.1 to 2.2KPa. For eachmeasurement time the values are the mean of four measurements. Error bars indicate the s.e. Different letters show statistically significant differences amongtreatments at P< 0.05.
Hydraulic and chemical signalling in grapevines Functional Plant Biology 571
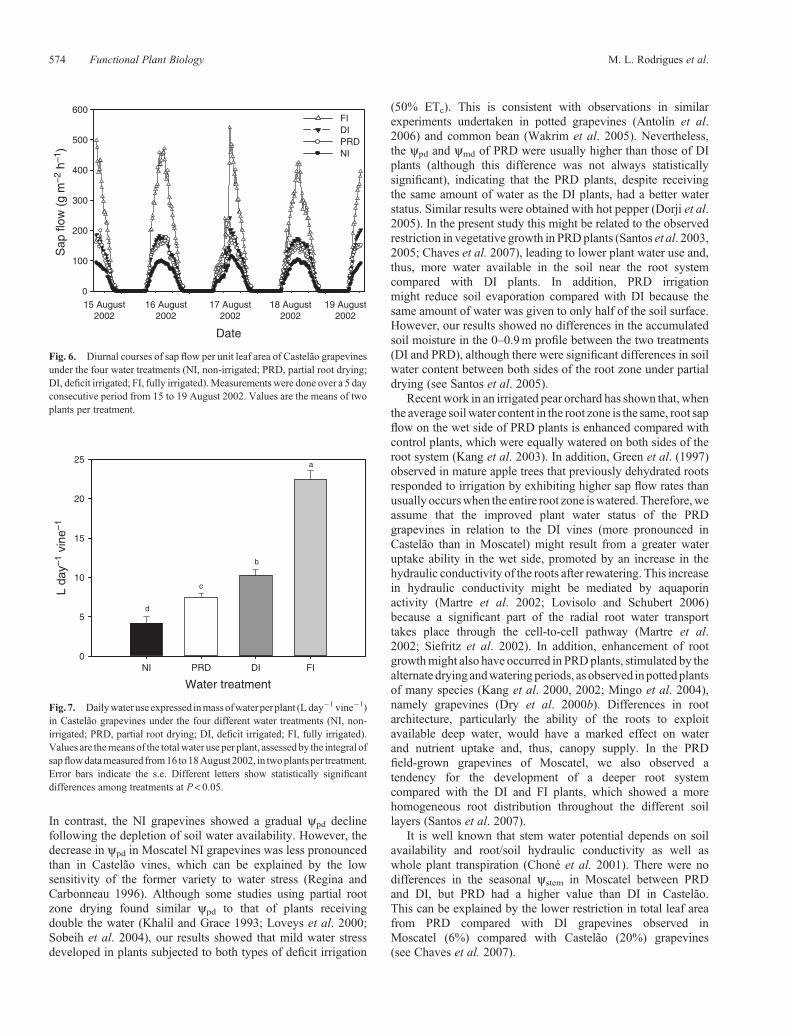
Sap flow and plant water use
The diurnal changes in sap flow rates observed in Castelão for aconsecutive period of 5 days, beginning 15 August, are in Fig. 6.
On that date, all irrigated grapevines were watered and thewatered side of the PRD plants was changed. Soil wateravailability affected the sap flow rates per unit of leaf area, but
ab
ns
ns
b
a
a
b
a
b
a
b
b
c
a
0.0
0.5
1.0
1.5
2.0
ns
b
ns
a
ab
b
NIPRDDIFI
a
b
a
a
ab
b b
ab
a
0
50
100
150
200
250
300
350
NIPRDDIFI
a
ab
b a
b
b
ns
3.5
4.0
4.5
5.0
5.5
6.0
a
b
a
ab
b
a
a
a
ab
b
b
AB
A (
mm
ol m
–3)
g s (
mm
ol m
–2 s
–1)
pH
1000 1400 1800
Time of the day (hours)
1000 1400 1800
Time of the day (hours)
(A) (B)
Fig. 4. Diurnal changes in leaf abaxial stomatal conductance (gs), ABA concentration in the xylem sap and pH of the xylem sap of Moscatel grapevinesunder the four water treatments, measured on 2 days of the growing season, (A) veraison (29 July) and (B) mid-ripening (26 August). On both days, PPFDranged from 1200 to 1900mmolm�2 s�1 over the period of measurement. On 29 July, air temperature (T) changed from 24 to 32�C and the vapour pressuredeficit of the air (VPD) changed from 1.1 to 2.6KPa, while on 26 August T changed from 21 to 28�C and the VPD changed from 0.81 to 1.78KPa. For eachmeasurement time the values are the mean of four measurements. Error bars indicate the s.e. Different letters show statistically significant differences amongtreatments at P< 0.05.
572 Functional Plant Biology M. L. Rodrigues et al.
did not modify the diurnal pattern of water flow. Sap flowthroughout the plant showed a steep increase from dawn toearly afternoon (1400–1500 hours) and then decreased slowlyand steadily until late afternoon. Daily maxima values, whichdepend on the evaporative demand and solar radiation,were 400–500 gm�2 h�1 (expressed per unit leaf area) for the FI grapevinesand were markedly reduced to 100 gm�2 h�1 in the NI plants.Deficit-irrigated grapevines showed intermediate sap flow ratesbetween FI and NI, although the values were closer to the latter.No significant differences were observed between DI and PRDgrapevines. Although sap flow was not assessed in Moscatel in2002, measurements carried out in 2003 with the sameexperimental design indicated similar patterns in the differentwater treatments (M. L. Rodrigues, T. P. Santos, C. M. Lopes,M. M. Chaves, unpubl. data).
Figure 7 shows the mean daily water use per plant over threeconsecutive days (16–18 August 2002) in Castelão grapevines.Over this period,which consisted of clear dayswithwarm anddryweather, the mean maximum daily values of air temperature and
VPDof the airwere32.1�Cand2.05 kPa, respectively. Significantdifferences (P < 0.05) in plant water consumption were observedamong treatments; 4.2 L vine�1 day�1 in NI grapevines and 1.8,2.5 andfivefold higher in the PRD,DI and FI plants, respectively.These values correspond to 0.81 Lm�2 day�1 for NI,1.32 Lm�2 day�1 for PRD, 1.47 Lm�2 day�1 for DI and2.92 Lm�2 day�1 for FI plants, based on leaf area. Thedifferences between deficit-irrigated vines mainly resultedfrom the mean leaf area per plant, which was higher in DI(7.0m2) than in PRD (5.6m2). The maximum leaf area wasrecorded in the FI plants (7.7m2) and the minimum wasrecorded in the NI plants (5.2m2).
Discussion
Plantwater statuswas closely related to soilwater availability andFI grapevines had the highest values of ypd, and these valuesremained constant throughout the season, indicating that theirrigation input matched the plant transpiration requirements.
Xylem [ABA] (mmol m–3)Xylem [ABA] (mmol m–3)
0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6
g s, m
mol
m–2
s–1
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.40
50
100
150
200
250
300
350
400
Predawn water potential (MPa) Predawn water potential (MPa)
–1.0 –0.8 –0.6 –0.4 –0.2
gs
(mm
ol m
–2 s
–1)
gs
(mm
ol m
–2 s
–1)
0
50
100
150
200
250
300
350
400
–0.8 –0.6 –0.4 –0.2 0.0
veraisonR 2 = 0.39P < 0.05
veraisonR2 = 0.55P < 0.001
mid-ripeningR2 = 0.87P < 0.001
veraisonR2 = 0.16P > 0.05
veraisonR2 = 0.07P > 0.05
mid-ripeningR2 = 0.66P < 0.001
mid-ripeningR2 = 0.25P < 0.05
mid-ripeningR2 = 0.65P < 0.001
(A) (B)
(C) (D)
Fig. 5. Relationships between maximum diurnal stomatal conductance (gs) and pre-dawn leaf waterpotential (ypd) or leaf xylem ABA concentration obtained at veraison (* NI, ! PRD, & DI, ¤ FI; andthe solid regression line) and at mid-ripening (� NI,r PRD,& DI,^ FI; and the dashed regression line) in(A, C) Castelão and (B, D) Moscatel grapevine varieties. Values shown are individual data points with theregression lines determined for all four water treatments.
Hydraulic and chemical signalling in grapevines Functional Plant Biology 573
In contrast, the NI grapevines showed a gradual ypd declinefollowing the depletion of soil water availability. However, thedecrease in ypd in Moscatel NI grapevines was less pronouncedthan in Castelão vines, which can be explained by the lowsensitivity of the former variety to water stress (Regina andCarbonneau 1996). Although some studies using partial rootzone drying found similar ypd to that of plants receivingdouble the water (Khalil and Grace 1993; Loveys et al. 2000;Sobeih et al. 2004), our results showed that mild water stressdeveloped in plants subjected to both types of deficit irrigation
(50% ETc). This is consistent with observations in similarexperiments undertaken in potted grapevines (Antolín et al.2006) and common bean (Wakrim et al. 2005). Nevertheless,the ypd and ymd of PRD were usually higher than those of DIplants (although this difference was not always statisticallysignificant), indicating that the PRD plants, despite receivingthe same amount of water as the DI plants, had a better waterstatus. Similar results were obtained with hot pepper (Dorji et al.2005). In the present study this might be related to the observedrestriction in vegetative growth in PRDplants (Santos et al. 2003,2005; Chaves et al. 2007), leading to lower plant water use and,thus, more water available in the soil near the root systemcompared with DI plants. In addition, PRD irrigationmight reduce soil evaporation compared with DI because thesame amount of water was given to only half of the soil surface.However, our results showed no differences in the accumulatedsoil moisture in the 0–0.9m profile between the two treatments(DI and PRD), although there were significant differences in soilwater content between both sides of the root zone under partialdrying (see Santos et al. 2005).
Recent work in an irrigated pear orchard has shown that, whenthe average soilwater content in the root zone is the same, root sapflow on the wet side of PRD plants is enhanced compared withcontrol plants, which were equally watered on both sides of theroot system (Kang et al. 2003). In addition, Green et al. (1997)observed in mature apple trees that previously dehydrated rootsresponded to irrigation by exhibiting higher sap flow rates thanusually occurswhen the entire root zone iswatered. Therefore,weassume that the improved plant water status of the PRDgrapevines in relation to the DI vines (more pronounced inCastelão than in Moscatel) might result from a greater wateruptake ability in the wet side, promoted by an increase in thehydraulic conductivity of the roots after rewatering. This increasein hydraulic conductivity might be mediated by aquaporinactivity (Martre et al. 2002; Lovisolo and Schubert 2006)because a significant part of the radial root water transporttakes place through the cell-to-cell pathway (Martre et al.2002; Siefritz et al. 2002). In addition, enhancement of rootgrowthmight also have occurred in PRDplants, stimulated by thealternate dryingandwateringperiods, as observed inpottedplantsof many species (Kang et al. 2000, 2002; Mingo et al. 2004),namely grapevines (Dry et al. 2000b). Differences in rootarchitecture, particularly the ability of the roots to exploitavailable deep water, would have a marked effect on waterand nutrient uptake and, thus, canopy supply. In the PRDfield-grown grapevines of Moscatel, we also observed atendency for the development of a deeper root systemcompared with the DI and FI plants, which showed a morehomogeneous root distribution throughout the different soillayers (Santos et al. 2007).
It is well known that stem water potential depends on soilavailability and root/soil hydraulic conductivity as well aswhole plant transpiration (Choné et al. 2001). There were nodifferences in the seasonal ystem in Moscatel between PRDand DI, but PRD had a higher value than DI in Castelão.This can be explained by the lower restriction in total leaf areafrom PRD compared with DI grapevines observed inMoscatel (6%) compared with Castelão (20%) grapevines(see Chaves et al. 2007).
Date
Sap
flow
(g
m–2
h–1
)
0
100
200
300
400
500
600FIDIPRDNI
15 August2002
16 August2002
17 August2002
18 August2002
19 August2002
Fig. 6. Diurnal courses of sap flow per unit leaf area of Castelão grapevinesunder the four water treatments (NI, non-irrigated; PRD, partial root drying;DI, deficit irrigated; FI, fully irrigated).Measurements were done over a 5 dayconsecutive period from 15 to 19 August 2002. Values are the means of twoplants per treatment.
Water treatment
NI PRD DI FI
L da
y–1
vine
–1
0
5
10
15
20
25
d
c
b
a
Fig. 7. Dailywateruse expressed inmassofwaterperplant (L day�1 vine�1)in Castelão grapevines under the four different water treatments (NI, non-irrigated; PRD, partial root drying; DI, deficit irrigated; FI, fully irrigated).Values are themeansof the totalwater use per plant, assessedby the integral ofsapflowdatameasured from16 to18August2002, in twoplantsper treatment.Error bars indicate the s.e. Different letters show statistically significantdifferences among treatments at P < 0.05.
574 Functional Plant Biology M. L. Rodrigues et al.

Although some grapevine varieties under water stress haveshown the ability for osmotic adjustment at both the leaf(Rodrigues et al. 1993; Patakas and Noitsakis 1999) and rootlevel (Düring and Dry 1995), in the present study leaf osmoticpotential and therefore the relative leaf solute content wasunchanged in response to decreased soil water availability(Fig. 2C, D). Thus, the differences in leaf turgor amongtreatments and varieties (Fig. 2) were solely determined by theleaf water potential values.
The diurnal pattern of water flow measured in Castelãowas independent of the plant water status (see Fig. 6) asobserved in other grapevine cultivars (Vitis vinifera, cv.Freisa) by Lovisolo et al. (2000). Our data also showed goodadjustment of grapevine sapflow to soil water availability, the sapflux density being markedly controlled by stomatal conductance(see Fig. 3 and also Souza et al. (2005)). Indeed, vine canopies areaerodynamically rough surfaces, presenting high boundary layerconductance and they are highly coupled to the atmosphere(Lu et al. 2003).
Water consumption per plant was reduced by 54, 67 and 80%in the DI, PRD and NI plants, respectively, compared with theFI grapevines. This was a consequence of the markedrestriction in stomatal conductance together with the decreasein total leaf area of PRD and NI plants. In addition, a reduction inshoot hydraulic conductivity in NI and deficit-irrigated plantsmight also have limited water transport. In fact, Lovisolo andSchubert (1998) observed in potted grapevines a decrease inxylem vessel diameter, even at moderate water stress levels(ymd =�0.60MPa) and the occurrence of partial embolismof the vessels under more severe drought conditions. AlthoughPRD and DI plants exhibited similar transpiration rates, theaforementioned larger restriction in canopy leaf area perplant observed in the former compared with the latter (Chaveset al. 2007) determined their lower plant water use.
For both varieties, similar stomatal behaviour in the PRD andDIgrapevineswasobserved in thepresent study (Figs3 and4) andalso in a detailed analysis of gas exchange over the wholegrowing season in this same experiment (Souza et al. 2005).Our results are in accordance with those obtained from a fieldexperiment in California with Sauvignon blanc grapevines(Gu et al. 2004), indicating that plants are responsive to theamount of irrigation independently of the way in which the wateris applied. In addition, Collins et al. (2005) in a Shiraz vineyard inAustralia with two irrigation levels showed no differences in gsmeasured over the season between PRD and DI plants for awatering regime corresponding to 0.9 ETc. However, at 0.45 ETc
irrigation, the gs of PRD grapevines was maintained, while adecline was noticed in DI plants. Conversely, some studieshave shown a marked decrease in gs in PRD plants relative toplants conventionally irrigated, despite comparable shoot waterstatus (Poni et al. 1992; Du et al. 2006). Therefore, plantresponses to PRD may be influenced not only by the amountof water given but also by the climate, soil type and variety(Loveys et al. 2004; Collins et al. 2005; Souza et al. 2005;Du et al. 2006).
In the present study, a decrease in gs measured in the morningwas observed in plants of both varieties and in allwater treatmentsbetween veraison and mid-ripening. As this occurred regardlessof the evaporative demand conditions (or changes in plant water
status in irrigated plants), the reason appears to be related to leafageing or to changes associated with the phenological stage(Wilson et al. 2000; Ruiz-Sánchez et al. 2007).
Most of the PRD studies carried out in potted plants haveimplicated root-sourced ABA in the regulation of stomataaperture, with PRD plants having more of that hormone in thesap xylem compared with plants irrigated in the whole rootsystem with the same amount of water (Liu et al. 2006).However, analysing the daily pattern of xylem sap [ABA]from PRD and deficit-irrigated field-grown CabernetSauvignon grapevines, Stoll et al. (2000) only detected anincrease in ABA in PRD plants at mid-morning coincidentwith a depression of gs (~20% in this period) compared withDI plants. It would be expected that, if a sustained root ABAsynthesis occurred in PRD grapevines, an increased [ABA]would be observed in the afternoon because of a decrease inthe transpiration stream.
Our results have indicated that grapevine stomata stronglyrespond to plant water status because a positive significantcorrelation between ypd and maximum gs was always foundthroughout the season for both varieties (see Fig. 5). However, nosignificant relationship was found between gs and [ABA]transported in the xylem in the measurements done atveraison. Nevertheless, we noticed a higher [ABA] in the leafxylem sap of NI grapevines compared with all the watered plants(more evident in Castelão than in Moscatel) (see Figs 3 and 4).This rise in ABA in NI plants can be root sourced, but might alsobe leaf originated because of an internal redistribution withrelease from the symplastic pool in the leaf or because of anincrease in its biosynthesis as a consequence of leaf turgor loss. Inthe latter case, ABA can be recirculated to the shoot via the xylemafter having been translocated down to the root in the phloem(Soar et al. 2004; Wilkinson 2004). The decline of gs in deficit-irrigated grapevines compared with FI plants appears to beindependent of the [ABA] circulating in the xylem sap.However, we cannot disregard that the different stomatalbehaviour, regardless of the ABA content delivered to theleaves ([ABA] in the xylem multiplied by the transpirationrate), might have been produced by changes incompartmentalisation and catabolism of ABA in themesophyll. The likely release of stored symplastic ABA and/orthe lower capacity to metabolise ABA in the deficit-irrigatedplants might have increased hormone access to the apoplastsurrounding guard cells, the place of ABA action on stomata,given the lack of cytosolic connections between those cells andthe subsidiary cells (Hartung and Slovik 1991; Trejo et al. 1993,1995; Hartung et al. 1998). Indeed, the apparent stomatalsensitivity to a given [ABA] has been shown to increase asleaf water potential declines (Tardieu and Davies 1992;Tardieu et al. 1993). This response appears to be implicated inthe decreased gs throughout the day, even in FI grapevines,despite the relatively constant diurnal content of [ABA] in thexylem stream (Figs 3 and 4), as was observed in a previous fieldgrapevine experiment with cv. Trincadeira Preta grapevines(Correia et al. 1995).
Alternating the root wet/dry sides in PRD plants is essential tomaintain the signalling effects on the shoot considering thetransient nature of the root ABA accumulation in dehydratedroots (Dry et al. 2000a; Loveys et al. 2004). Dodd et al. (2006)
Hydraulic and chemical signalling in grapevines Functional Plant Biology 575

using PRD tomato plants receiving the same amount of watershowed a transient increase in [ABA] in the xylem and ofstomatal closure in plants with alternate irrigation comparedwith those in which a dry side was maintained. In ourexperiment, no significant differences were apparent in thediurnal courses of sap xylem [ABA] between PRD and DICastelão plants made either at veraison or at mid-ripening,dates corresponding to the end or the middle of the soil-dryingcycle, respectively.The lackof a difference in [ABA] in thexylemsap between PRD and DI grapevines might indicate that thelowering of the root water status in the dry half of the root systemof PRD plants was not sufficiently strong to induce increasedsynthesis of that compound or the transient nature of it was notcaptured in our experiment. The first hypothesis might beexplained by water transfer from wet to drying roots byhydraulic redistribution (Caldwell et al. 1998). The dynamicsof water transfer by root systems have been studied using stableisotope tracing (Brookes et al. 2002; Smart et al. 2005) or sapflowmeasurements in roots (Sakuratani et al. 1999;Burgess andBleby2006) and have been observed to occur in many plant species,including grapevines (Stoll et al. 2000; Smart et al. 2005). Theseauthors, using deuterium-labelled water in an Americangrapevine hybrid (Vitis riparia�Vitis berlandieri) growing infield conditions, showed that water from roots with access to soilmoisture was laterally transferred across the vine trunk and intoroots on the opposite non-irrigated side. In addition, the deeperwetted zone of the soil profile in PRD plants suggests thepossibility of the occurrence of some hydraulic lift, with watermoving vertically upwards.
Xylem sap pH can modulate stomatal response to rootchemical signals produced in drying soils (Wilkinson andDavies 1997, 2002; Wilkinson 2004). Indeed, in acidic xylemsap, ABA is rapidly taken by the leaf and metabolised orpartitioned into alkaline compartments in the symplast of theleaf cells, away from the sites of action of the hormone on thestomata. Conversely, as the pH increases, the proportion ofionised ABA transported in the xylem sap rises and ismaintained longer in the leaf apoplast adjacent to the bindingsites on the guard cells, which have more control on stomatalbehaviour (Wilkinson 2004). This effect is particularlyimportant in grapevines because, in general, they have pHvalues close to the pKa of ABA (pH 4.8), as indicated by ourresults and also by the results of Stoll et al. (2000). Therefore, anincrease in xylem sap pH can induce stomatal closure evenwithout an increase in [ABA]. In grapevines, a higher sap pHwas shown inPRDplants comparedwith those inwhich thewholeroot systemwaswatered (Stoll et al. 2000). In addition, in tomato,an increased pH in PRD plants consistently occurred comparedwith plants receiving double the water (Sobeih et al. 2004).However, in common bean (Wakrim et al. 2005) and in ourexperiment, no significant differences in pH were induced bydifferent soil water availabilities, although FI grapevinesusually had slightly more acidic values (see Figs 3 and 4).Nevertheless, we noticed at mid-ripening a significant increasein xylem sap pH in all plants compared with the veraison values.This suggests that the rise in the xylem sap pH was at least partlyinvolved in stomatal control, enabling a highly significantnegative relationship between [ABA] and gs, which wasabsent in veraison (see Fig. 5).
In conclusion, the present study showed that mild water stressoccurred in deficit-irrigated grapevines (50% ETc), but PRDplants had a better (Castelão) or similar (Moscatel) waterstatus compared with DI vines, as assessed by pre-dawn andstemwater potentials. Stomata behaved similarly in the PRD andDI grapevines; in Castelão this was evident when evaluated bothat the leaf (stomatal conductance) and shoot level (sap flow).Furthermore, the absence of differences in [ABA] in the xylem orin the pH of xylem sap in those plants does not support thehypothesis that at least those root-sourced chemical signalsregulated stomatal conductance in partial root dryinggrapevines. This agrees with the idea that woody plants areless dependent on slow-moving root signals for short-termstomata regulation compared with herbaceous species(Saliendra et al. 1995; Fuchs and Livingston 1996).Nevertheless, the restriction effects observed in vegetativegrowth in PRD grapevines compared with DI vines are notexplained by hydraulic signals, but rather suggest the effect ofchemical signalling, probably involving a reduction of cytokinins(Kudoyarova et al. 2007) or an increase of ethylene (Sobeihet al. 2004).
Low responsiveness to soil water depletion of the varietiesand rootstock used in our experiment as well as the moderateirrigation used might have contributed to the similar stomatalbehaviour of PRD and DI plants. Loveys et al. (2004) reportedsimilar results in a field experiment with Cabernet Sauvignonapplying comparable watering rates to ours. These authorsonly found a significant depression in stomatal conductancein PRD compared with DI plants at high water irrigation rates(4–5L per plant per day).
A positive significant correlation between ypd and maximumgs was always found for both varieties, indicating that grapevinestomata strongly respond to plantwater status. Atmid-ripening gswas significantly correlated with ABA, apparently because of aninteractionwith rising xylem sap pH. Thiswas not the case earlierin the season (veraison), where ABA did not explain stomatalcontrol.
Consequently, our data suggest that both hydraulic feedbackand feed-forward root-to-shoot chemical signalling mechanismscan be involved in the control of grapevine stomata apertures inresponse to decreased soil water availability; however, hydraulicsignals appear to play the dominant role.
Acknowledgements
We are grateful to DRARO, namely the Centro Experimental de Pegões, forthe experimental vineyard facilities. We thank M. José Moreira and VictoriaPicado from ISA for their laboratory technical assistance. This research wasfinancially supported by the EU project IRRISPLIT (ICA-1999-10026).
References
Antolín MC, Ayari M, Sánchez-Díaz M (2006) Effects of partial rootzonedryingonyield, ripeningandberryABAinpottedTempranillo grapevineswith split roots. Australian Journal of Grape and Wine Research 12,13–20. doi: 10.1111/j.1755-0238.2006.tb00039.x
Augé RM,Moore JL (2002) Stomatal response to nonhydraulic root-to-shootcommunication of partial soil drying in relation to foliar dehydrationtolerance. Environmental and Experimental Botany 47, 217–229.doi: 10.1016/S0098-8472(01)00129-0
576 Functional Plant Biology M. L. Rodrigues et al.

Baker JM, van Bavel CHM (1987) Measurement of mass flow ofwater in stems of herbaceous plants. Plant, Cell & Environment 10,777–782.
BravdoB,NaorA (1996)Effect ofwater regimeonproductivity andquality offruit and wine. Acta Horticulturae 427, 15–26.
Brookes JR,MeinzerC,CoulombeR,Gregg J (2002)Hydraulic redistributionof soil water during summer drought in two contrasting Pacific Northwestconiferous forests. Tree Physiology 22, 1107–1117.
Burgess SSO, Bleby TM (2006) Redistribution of soil water by lateral rootsmediatedbystem tissues.JournalofExperimentalBotany57, 3283–3291.doi: 10.1093/jxb/erl085
CaldwellMM,Dawson TE, Richards JH (1998) Hydraulic lift: consequencesof water efflux from the roots of plants. Oecologia 113, 151–161.doi: 10.1007/s004420050363
Chaves MM, Santos TP, Souza CR, Ortuño MF, Rodrigues ML, Lopes CM,Maroco JP, Pereira JS (2007) Deficit irrigation in grapevine improveswater-use efficiency while controlling vigour and production quality.The Annals of Applied Biology 150, 237–252. doi: 10.1111/j.1744-7348.2006.00123.x
Choné X, Van Leeuwen C, Dubourdieu D, Gaudillère JP (2001) Stem waterpotential is a sensitive indicator of grapevine water status. Annalsof Botany 87, 477–483. doi: 10.1006/anbo.2000.1361
Collins M, Fuentes S, Barlow S (2005) Water-use of grapevines toPRD irrigation at two water levels. A case study in North-EasternVictoria. The Australian & New Zealand Grapegrower & Winemaker502, 41–45.
Comstock JP (2002) Hydraulic and chemical signalling in the control ofstomatal conductance and transpiration. Journal of Experimental Botany53, 195–200. doi: 10.1093/jexbot/53.367.195
Correia MJ, Pereira JS (1995) Absicisic acid in apoplastic sap can accountfor the restriction in leaf conductance of white lupins duringmoderate soil drying and after rewatering. Plant, Cell & Environment17, 845–852.
Correia MJ, Pereira JS, Chaves MM, Rodrigues ML, Pacheco CA (1995)ABA xylem concentrations determine maximum daily leaf conductanceof field-grown Vitis vinifera L. plants. Plant, Cell & Environment 18,511–521. doi: 10.1111/j.1365-3040.1995.tb00551.x
Davies WJ, Tardieu F, Trejo CL (1994) How do chemical signals work inplants that grow in drying soil. Plant Physiology 104, 309–314.
Dodd IC, Stikic R, Davies WJ (1996) Chemical regulation of gas exchangeand growth of plants in drying soil in the field. Journal of ExperimentalBotany 47, 1475–1490. doi: 10.1093/jxb/47.10.1475
Dodd IC, Theobald JC, BaconMA, DaviesWJ (2006) Alternation of wet anddry sides during partial rootzone drying irrigation alters root-to-shootsignalling of abscisic acid. Functional Plant Biology 33, 1081–1089.doi: 10.1071/FP06203
Dorji K, Behboudian MH, Zegbe-Domínguez JA (2005) Water relations,growth, yield, and fruit quality of hot pepper under deficit irrigation andpartial rootzone drying. Scientia Horticulturae 104, 137–149.doi: 10.1016/j.scienta.2004.08.015
Dry PR, Loveys BR (1999) Grapevine shoot growth and stomatalconductance are reduced when part of the root system is dried. Vitis38, 151–156.
Dry PR, Loveys BR, Düring H (2000a) Partial drying of the rootzone ofgrape. I. Transient changes in shoot growth and gas exchange. Vitis 39,3–7.
Dry PR, Loveys BR, Düring H (2000b) Partial drying of the rootzone ofgrape. II. Changes in the patterns of root development. Vitis 39, 9–12.
DuT,KangS,ZhangJ,LiF,HuX(2006)Yieldandphysiological responsesofcotton to partial root-zone irrigation in the oasis field of northwest China.Agricultural Water Management 84, 41–52. doi: 10.1016/j.agwat.2006.01.010
DüringH,DryPR (1995)Osmoregulation inwater-stressed roots—responsesof leaf conductance and photosynthesis. Vitis 34(1), 15–17.
FuchsEE,LivingstonNJ (1996)Hydraulic control of stomatal conductance inDouglasfir [Pseudotsugamenziesi (Mirb.) Franco] and alder [Alnus rubra(Bong)] seedlings. Plant, Cell & Environment 19, 1091–1098.doi: 10.1111/j.1365-3040.1996.tb00216.x
Gollan T, Passioura JB,MunnsR (1986) Soil-water status affects the stomatalconductance of fully turgid wheat and sunflower leaves. AustralianJournal of Plant Physiology 13, 459–464.
Gowing DJG, DaviesWJ, Jones HG (1990) A positive root-sourced signal asan indicator of soil drying in apple, Malus domestics Borkh. Journalof Experimental Botany 41, 1535–1540. doi: 10.1093/jxb/41.12.1535
Green SR, Clothier BE,McLeodDJ (1997) The response of sap flow in appleroots to localised irrigation. Agricultural Water Management 33, 63–78.doi: 10.1016/S0378-3774(96)01277-2
GuSL,DuGQ, ZoldoskeD,HakimA,CochranR, FugelsangK, JorgensenG(2004) Effects of irrigation amount on water relations, vegetativegrowth, yield and fruit composition of Sauvignon blanc grapevinesunder partial rootzone drying and conventional irrigation in theSan Joaquin Valley of California, USA. Journal of HorticulturalScience & Biotechnology 79(1), 26–33.
HansenH,DorfflingK (2003) Root-derived trans-zeatin riboside and abscisicacid in drought-stressed and rewatered sunflower plants: interaction in thecontrol of leaf diffusive resistance? Functional Plant Biology 30,365–375. doi: 10.1071/FP02223
Hartung W, Slovik S (1991) Physicochemical properties of plant growthregulators and plant tissues determine their distribution and redistribution:stomatal regulation by abscisic acid in leaves. The New Phytologist 119,361–382. doi: 10.1111/j.1469-8137.1991.tb00036.x
Hartung W, Wilkinson S, Davies WJ (1998) Factors that regulateabscisic acid concentrations at the primary site of action at the guardcell. Journal of Experimental Botany 49, 361–367. doi: 10.1093/jexbot/49.suppl_1.361
Jia W, Davies WJ (2007) Modification of leaf apoplastic pH in relation tostomatal sensitivity to root-sourced abscisic acid signals.PlantPhysiology143, 68–77. doi: 10.1104/pp.106.089110
Kang S, Liang Z, Pan Y, Shi P, Zhang J (2000) Alternate furrow irrigation formaize production in an arid area. Agricultural Water Management 45,267–274. doi: 10.1016/S0378-3774(00)00072-X
Kang S, Shi W, Cao H, Zhang J (2002) Alternate watering in soil verticalprofile improved water use efficiency of maize (Zea mays). Field CropsResearch 77, 31–41. doi: 10.1016/S0378-4290(02)00047-3
KangS,HuX, Jerie P,Zhang J (2003)The effects of partial rootzonedryingonroot, trunkflowandwater balance in an irrigatedpear (PyruscommunisL.)orchard. Journal of Hydrology (Amsterdam) 280, 192–206.
Khalil AM, Grace J (1993) Does xylem sap ABA control the stomatalbehaviour of water stressed Sycamore (Acer pseudoplatanus L.)seedlings? Journal of Experimental Botany 44, 1127–1134.doi: 10.1093/jxb/44.7.1127
Kudoyarova GR, Vysotskaya LB, Cherkozyanova A, Dodd IC (2007)Effect of partial rootzone drying on the concentration of zeatin-type cytokinins in tomato (Solanum lycopersicum L.) xylem sap andleaves. Journal of Experimental Botany 58, 161–168. doi: 10.1093/jxb/erl116
Liu F, JensenCR,AndersenMN (2003)Hydraulic and chemical signals in thecontrol of leaf expansion and stomatal conductance in soybean exposed todrought stress. Functional Plant Biology 30, 65–73. doi: 10.1071/FP02170
Liu F, Shahnazari A, Andersen MN, Jacobsen S, Jensen CR (2006)Physiological responses of potato (Solanum tuberosum L.) to partialroot-zone drying: ABA signalling, leaf gas exchange, and water useefficiency. Journal of Experimental Botany 57, 3727–3735.doi: 10.1093/jxb/erl131
LoveysBR,DryPR, StollM,McCarthyMG(2000)Usingplant physiology toimprove thewater use efficiencyof horticultural crops.ActaHorticulturae537, 187–197.
Hydraulic and chemical signalling in grapevines Functional Plant Biology 577

Loveys BR, Stoll M, DaviesWJ (2004) Physiological approaches to enhancewater use efficiency in agriculture: exploiting plant signalling in novelirrigation practice. In ‘Water use efficiency in plant biology’.(Ed MA Bacon) pp. 113–141. (Blackwell Publishing: Oxford)
Lovisolo C, Schubert A (1998) Effects of water stress on vessel size andxylem hydraulic conductivity in Vitis vinifera L. Journal of ExperimentalBotany 49, 693–700. doi: 10.1093/jexbot/49.321.693
Lovisolo C, Schubert A (2006) Mercury hinders recovery of shoot hydraulicconductivity during rehydration: evidence from a whole-plant approach.The New Phytologist 172(3), 469–478. doi: 10.1111/j.1469-8137.2006.01852.x
Lovisolo C, Schubert A, Peterlunger E, Ferraris S (2000) Sap flow and stemconductivity of potted water stressed grapevines. ActaHorticulturae 526,187–190.
Lu P, Yunusa IAM, Walker RR, Müller WJ (2003) Regulation of canopyconductance and transpiration and theirmodelling in irrigated grapevines.Functional Plant Biology 30, 689–698. doi: 10.1071/FP02181
Martre P, Morillon R, Barrieu F, North GB, Nobel PS, Chrispeels MJ (2002)Plasma membrane aquaporins play a significant role during recoveryfrom water deficits. Plant Physiology 130, 2101–2110. doi: 10.1104/pp.009019
McCarthy MG (1997) The effect of transient water deficit on berrydevelopment of cv. Shiraz (Vitis vinifera L.). Australian Journalof Grape and Wine Research 3, 102–108.
McCarthy MG, Loveys BR, Dry PR, Stoll M (2002) Regulated deficitirrigation and partial rootzone drying as irrigation managementtechniques for grapevines. In ‘Deficit irrigation practices. FAO waterreports, No. 22’. pp. 79–87 (FAO: Rome)
Mingo DM, Theobald JC, Bacon MA, Davies WJ, Dodd IC (2004)Biomass allocation in tomato (Lycopersicon esculentum) plantsgrown under partial rootzone drying: enhancement of root growth.Functional Plant Biology 31, 971–978. doi: 10.1071/FP04020
Netting AG, Windsor ML, Milborrow BV (1997) Endogenousbiosynthetic precursors of (+) abscisic acid. III. Incorporation of 2H2Ofrom 18O2 into precursors. Australian Journal of Plant Physiology 24,175–184.
Patakas A, Noitsakis B (1999) Osmotic adjustment and partitioning of turgorresponses to drought in grapevines leaves. American Journal of Enologyand Viticulture 50, 76–80.
Poni S, Tagliavini M, Neri D, Scudellari D, Toselli M (1992)Influence of root pruning and water stress on growth andphysiological factors of potted apple, grape, peach and pear trees.Scientia Horticulturae 52, 223–236. doi: 10.1016/0304-4238(92)90023-6
Possingham JV (2000) The influence of controlled water inputs on grapequality in warm regions. In ‘Proceedings of the XXVWorld Congress ofVine and Wine, Paris’, pp. 63–68. (OIV: Paris)
Prichard TL (1992) A volume balance approach to quality wine grapeirrigation. In ‘Viticultural practices’. (Eds MA Walker, WM Kliewer)pp. 12–23. (University of California: Davies)
Prokic L, Jovanovic Z, McAinsh MR, Vucinic Z, Stikic R (2006) Species-dependent changes in stomatal sensitivity to absicisic acid mediated byexternal pH. Journal of Experimental Botany 57(3), 675–683.doi: 10.1093/jxb/erj057
Regina MA, Carbonneau A (1996) Trocas gasosas em Vitis vinifera sobregime de estresse hídrico. I. Caracterizacão do comportamento varietal.Pesquisa Agropecuaria Brasileira 31, 869–876.
Rodrigues ML, Chaves MM, Wendler R, David MM, Quick P, Leegood R,StittM, Pereira JS (1993)Osmotic adjustment inwater stressed grapevineleaves in relation to carbon assimilation. Australian Journal of PlantPhysiology 20, 309–321.
Ruiz-Sánchez MC, Domingo R, Perez-Pastor A (2007) Daily variations inwater relations of apricot trees under different irrigation systems.BiologiaPlantarum 51(4), 735–740. doi: 10.1007/s10535-007-0150-5
Sakuratani T, Aoe T, Higuchi H (1999) Reverse flow in roots of Sesbaniarostrata measured using the constant power heat balance method. Plant,Cell & Environment 22, 1153–1160. doi: 10.1046/j.1365-3040.1999.00464.x
Saliendra NZ, Sperry JS, Comstock J (1995) Influence of leaf water statuson stomatal response to humidity, hydraulic conductance, and soildrought in Betula occidentalis. Planta 196, 357–366. doi: 10.1007/BF00201396
Santos T, Lopes C, Rodrigues ML, Souza CR, Silva JR, Maroco JP,Pereira JS, Chaves MM (2003) Partial rootzone drying effectson growth and fruit quality of field-grown grapevines (Vitisvinifera). Functional Plant Biology 30, 663–671. doi: 10.1071/FP02180
SantosT,LopesC,RodriguesML,SouzaCR,Silva JR,Maroco JP, Pereira JS,ChavesMM(2005)Effects of partial root-zonedrying irrigationon clustermicroclimate and fruit composition of Castelão field-grown grapevines.Vitis 44(3), 117–125.
SantosT,LopesC,RodriguesML,SouzaCR,Silva JR,Maroco JP, Pereira JS,Chaves MM (2007) Effects of deficit irrigation strategies on clustermicroclimate for improving fruit composition of Moscatel field-growngrapevines. Scientia Horticulturae 112, 321–330. doi: 10.1016/j.scienta.2007.01.006
Sauter A, Dietz KJ, Hartung W (2002) A possible stress physiological roleof abscisic acid conjugates in root-to-shoot signalling. Plant,Cell & Environment 25, 223–228. doi: 10.1046/j.1365-3040.2002.00747.x
SchultzHR (2000)Climate change and viticulture: a European perspective onclimatology, carbon dioxide and UV-B effects. Australian Journalof Grape and Wine Research 6, 2–12. doi: 10.1111/j.1755-0238.2000.tb00156.x
Schultz HR (2003) Differences in hydraulic architecture account for near-isohydric and anisohydric behaviour of two field-grown Vitis vinifera L.cultivars during drought. Plant, Cell & Environment 26, 1393–1405.doi: 10.1046/j.1365-3040.2003.01064.x
Shashidhar VR, Prasad TG, Sudharshan L (1996) Hormonal signals fromroots to shoots of sunflower (Helianthus annuusL.).Moderate soil dryingincreases delivery of absicisic acid and depresses delivery of cytokinins inthe xylem sap. Annals of Botany 78, 151–155. doi: 10.1006/anbo.1996.0107
Siefritz F, Tyree MT, Lovisolo C, Schubert A, Kaldenhoff R (2002) PIP1Plasmamembrane aquaporins in tobacco: from cellular effects to functionin plants. The Plant Cell 14, 869–876. doi: 10.1105/tpc.000901
Smart DR, Carlisle E, Goebel M, Núñez BA (2005) Transverse hydraulicredistribution by a grapevine. Plant, Cell & Environment 28, 157–166.doi: 10.1111/j.1365-3040.2004.01254.x
Soar CJ, Speirs J, Maffei SM, Loveys BR (2004) Gradients in stomatalconductance, xylem sap ABA and bulk leaf ABA along canes ofVitis vinifera cv. Shiraz: molecular and physiological studiesinvestigating their source. Functional Plant Biology 31, 659–669.doi: 10.1071/FP03238
SoarCJ, Speirs J,Maffei SM, PenroseAB,McCarthyMG,LoveysBR (2006)Grape vine varieties Shiraz andGrenache differ in their stomatal responseto VPD: apparent links with ABA physiology and gene expression in leaftissue. Australian Journal of Grape and Wine Research 12, 2–12.doi: 10.1111/j.1755-0238.2006.tb00038.x
Sobeih WY, Dodd IC, Bacon MA, Grierson D, Davies WJ (2004)Long-distance signals regulating stomatal conductance and leafgrowth in tomato (Lycopersicon esculentum) plants subjected to partialrootzone drying. Journal of Experimental Botany 55, 2353–2363.doi: 10.1093/jxb/erh204
Souza C, Maroco J, Santos T, Rodrigues ML, Lopes C, Pereira JS,Chaves MM (2005) Control of stomatal aperture and carbon uptake bydeficit irrigation in two grapevine cultivars. Agriculture Ecosystems &Environment 106, 261–274. doi: 10.1016/j.agee.2004.10.014
578 Functional Plant Biology M. L. Rodrigues et al.

Stoll M, Loveys B, Dry P (2000) Hormonal changes induced by partialrootzone drying of irrigated grapevine. Journal of Experimental Botany51, 1627–1634. doi: 10.1093/jexbot/51.350.1627
Tardieu F, DaviesWJ (1992) Stomatal responses to abscisic acid is a functionof current plant water status. Plant Physiology 98, 540–545.
TardieuF,ZhangJ,GowingDJG(1993)Stomatal control of both [ABA] in thexylemsapand leafwater status: a test of amodel for droughtedorABA-fedfield-grownmaize.Plant,Cell&Environment16, 413–420. doi: 10.1111/j.1365-3040.1993.tb00887.x
Tardieu F, Lafarge T, Simonneau T (1996) Stomatal control by fed orendogenous xylem ABA in sunflower interpretation of correlationsbetween leaf water potential and stomatal conductance in anisohydricspecies. Plant, Cell & Environment 19, 75–84. doi: 10.1111/j.1365-3040.1996.tb00228.x
Trejo CL, Davies WJ, Ruiz L (1993) Sensitivity of stomata to abscisic acid.An effect of the mesophyll. Plant Physiology 102, 497–502.
Trejo CL, Clephan AL, DaviesWJ (1995) How do stomata read abscisic acidsignals? Plant Physiology 109, 803–811.
Wakrim R, Wahbi S, Tahi H, Aganchich B, Serraj R (2005) Comparativeeffects of partial root drying (PRD) and regulated deficit irrigation (RDI)on water relations and water use efficiency in common bean (Phaseolusvulgaris L.). Agriculture, Ecosystems & Environment 106, 275–287.doi: 10.1016/j.agee.2004.10.019
Wample RL, Smithyman R (2002) Regulated deficit irrigation as a watermanagement strategy in Vitis vinifera production. In ‘Water reports, FAOpublication number 22’. pp. 89–101. (FAO: Rome)
Wilkinson S (2004) Water use efficiency and chemical signalling. In ‘Wateruse efficiency in plant biology’. (Ed MA Bacon) pp. 75–112. (BlackwellPublishing: Oxford)
Wilkinson S, Davies WJ (1997) Xylem sap pH increase: a drought signalreceived at the apoplastic face of the guard cell that involves thesuppression of saturable abscisic acid uptake by the epidermalsymplast. Plant Physiology 113, 559–573.
Wilkinson S, Davies WJ (2002) ABA-based chemical signalling: theco-ordination of responses to stress in plants. Plant, Cell &Environment 25, 195–210. doi: 10.1046/j.0016-8025.2001.00824.x
Wilson KB, Baldocchi DD, Hanson PJ (2000) Quantifying stomatal andnon-stomatal limitations to carbon assimilation resulting from leaf ageingand drought in mature deciduous tree species. Tree Physiology 20,787–797.
Zar J (1999) ‘Biostatistical Analysis, 4th edn.’ (Prentice-Hall: New Jersey)Zhang J, Schurr U, Davies WJ (1987) Control of stomatal behaviour by
abscisic acid, which apparently originates in the roots. Journalof Experimental Botany 38, 1174–1181. doi: 10.1093/jxb/38.7.1174
Manuscript received 9 January 2008, accepted 22 June 2008
Hydraulic and chemical signalling in grapevines Functional Plant Biology 579
http://www.publish.csiro.au/journals/fpb




















