
Strategies for Generating Human Pluripotent Stem Cell ... - MDPI
Upload
independentCategory
view
2download
0
EMBRYONIC STEM CELLS/INDUCED PLURIPOTENT STEM CELLS
Human-Induced Pluripotent Stem Cells Form Functional Neurons
and Improve Recovery After Grafting in Stroke-Damaged Brain
KOICHI OKI,a,b
JEMAL TATARISHVILI,a,b
JAMES WOOD,b,c
PHILIPP KOCH,dSOMSAK WATTANANIT,
a,bYUTAKA MINE,
b,c
EMANUELA MONNI,a,b DANIEL TORNERO,a,b HENRIK AHLENIUS,a,b JULIA LADEWIG,d OLIVER BRUSTLE,d
OLLE LINDVALL,b,c ZAAL KOKAIAa,b
aLaboratory of Neural Stem Cell Biology and Therapy and cLaboratory of Neurogenesis and Cell Therapy,
University Hospital, Lund, Sweden; bLund Stem Cell Center, Lund, Sweden; dInstitute of Reconstructive
Neurobiology, Life & Brain Center, University of Bonn and Hertie Foundation, Bonn, Germany
Key Words. Stem cell • Stroke • Pluripotent • Human • Transplantation
ABSTRACT
Reprogramming of adult human somatic cells to inducedpluripotent stem cells (iPSCs) is a novel approach to pro-
duce patient-specific cells for autologous transplantation.Whether such cells survive long-term, differentiate to
functional neurons, and induce recovery in the stroke-injured brain are unclear. We have transplanted long-term self-renewing neuroepithelial-like stem cells, gener-
ated from adult human fibroblast-derived iPSCs, into thestroke-damaged mouse and rat striatum or cortex. Recov-
ery of forepaw movements was observed already at 1week after transplantation. Improvement was most likelynot due to neuronal replacement but was associated with
increased vascular endothelial growth factor levels, prob-
ably enhancing endogenous plasticity. Transplanted cellsstopped proliferating, could survive without forming
tumors for at least 4 months, and differentiated to mor-phologically mature neurons of different subtypes. Neu-
rons in intrastriatal grafts sent axonal projections to theglobus pallidus. Grafted cells exhibited electrophysiologi-cal properties of mature neurons and received synaptic
input from host neurons. Our study provides the first evi-dence that transplantation of human iPSC-derived cells is
a safe and efficient approach to promote recovery afterstroke and can be used to supply the injured brain withnew neurons for replacement. STEM CELLS 2012;30:1120–1133
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
Ischemic stroke is a leading cause of mortality and disability.Ongoing studies in animal models of stroke suggest that stemcell-based approaches could provide therapeutic benefit in aclinical setting. The observed improvements may, at leastpartly, be due to neuronal replacement [1]. After transplanta-tion into the stroke-damaged rodent brain, embryonic stemcell (ESC)-derived neural stem cells (NSCs) differentiatedinto neurons and improved some impaired functions [2–4].Electrophysiological recordings showed mature neuronal prop-erties of the grafted cells and signs of synaptic integrationinto host brain [5, 6]. Human fetal NSCs also gave rise tomature neurons after transplantation into the stroke-damagedrat brain [7–9]. Transplanted NSCs may promote recoverywithout differentiating to neurons through several other mech-
anisms, for example, modulation of inflammation [10–12],neuroprotection [12], stimulation of angiogenesis [11, 13, 14],and enhancing brain plasticity [15–17].
Somatic cells can be reprogrammed to pluripotent stemcells by introduction of transcription factors [18]. Theseinduced pluripotent stem cells (iPSCs) can be differentiated tospecific neuronal subtypes, for example, dopaminergic neu-rons [19, 20] and motor neurons [21, 22]. Autologous trans-plantation of neurons generated from iPSCs seems to be moreattractive in stroke therapy than in the treatment of chronicneurodegenerative disorders, in which patient-specific cellsmay exhibit increased susceptibility to the pathological proc-esses. Recently, mouse iPSCs, implanted into the stroke-dam-aged brain of mice, were reported to generate large numbersof neuroblasts and a few mature neurons but to form tumors[23]. When these cells were mixed with fibrin glue and weredelivered subdurally into the injured brain tissue in rats after
Author contributions: K.O., J.T., J.W., P.K., and S.W.: conception and design, collection and assembly of data, data analysisand interpretation, and manuscript writing; Y.M. and D.T.P.: collection and assembly of data and data analysis; E.M.: conception anddesign, collection and assembly of data, and data analysis; H.A. and J.L.: collection and assembly of data and data analysis andinterpretation; O.B.: conception and design, data interpretation, financial support, and manuscript writing; O.L. and Z.K.: conceptionand design, data analysis and interpretation, financial support, and manuscript writing. K.O., J.T., J.W., and P.K. contributed equally tothis article.
Correspondence: Zaal Kokaia, Ph.D., Lund Stem Cell Center, University Hospital BMC B10, SE-221 84 Lund, Sweden.Telephone: þ46-46-222-0276; Fax: þ46-46-222-0560; e-mail: [email protected] Received December 14, 2011; accepted forpublication March 22, 2012; first published online in STEM CELLS EXPRESS April 11, 2012. VC AlphaMed Press 1066-5099/2012/$30.00/0doi: 10.1002/stem.1104
STEM CELLS 2012;30:1120–1133 www.StemCells.com

stroke, there was a reduction of the infarct volume and somebehavioral improvement [24]. Finally, human fibroblast-derived iPSCs implanted into the striatum of stroke-damagedrats were found to improve short-term sensorimotor recovery,but whether any neurons were formed is unclear [25]. Anotherrecent study did not detect any effect of iPSCs on stroke-induced behavioral impairments at 4 weeks after transplanta-tion [26]. The long-term consequences after transplantation ofhuman iPSC-derived cells in the stroke-damaged brain, if theycan differentiate in vivo to morphologically and functionallymature neurons, establish connections to target areas, andimprove behavioral deficits resembling those in strokepatients, are currently unknown.
Here, we have transplanted long-term expandable neuro-epithelial-like stem (lt-NES) cells, generated from adult humanfibroblast-derived iPSCs, into the stroke-damaged mouse andrat brain. We show for the first time that grafted human iPSC-derived lt-NES cells generate neurons with mature morphologi-cal and electrophysiological properties in vivo, send axonalprojections throughout the host brain, receive synaptic inputfrom surrounding host neurons, and improve motor recovery inbehavioral tests relevant for human stroke.
MATERIALS AND METHODS
Generation of Human-iPSCs
For production of retroviral particles, PhoenixGP cells (ATTC#3514) were transfected with plasmids encoding for the viral glyco-protein VSV-G (Addgene 12259, Cambridge, Massachusetts, http://www.addgene.org) and the reprogramming factors (Oct4, Sox2,KLF4, and c-MYC (Addgene 17217-17220). Dermal fibroblastswere infected twice with ultracentrifuge-concentrated virus in Dul-becco’s modified Eagle’s medium (DMEM) high glucose media con-taining 10% fetal calf serum, 1% sodium pyruvate, 1% nonessentialamino acids, and 1% L-glutamine (all life technologies) in the pres-ence of 10 ng/ml FGF2. Cells were split into plates preseeded withmouse embryonic fibroblasts (MEFs). Medium was switched tohuman iPSC culture medium containing DMEM/F12 supplementedwith 20% knockout serum replacement, 0.1 mM nonessential aminoacids (all from life technologies), 1 mM L-glutamine, 0.1 mM b-mer-captoethanol, and 50 ng/ml zebrafish basic fibroblast growth factor(zbFGF) 6 days post-transduction and changed every other day. Fourweeks after transduction, colonies with human pluripotent stem cellmorphology were manually picked, carefully triturated, andexpanded to establish human iPSC lines.
Neural Differentiation of Human iPSCs
Human iPSCs were induced to differentiate as described [27].Briefly, iPSC colonies were detached from the MEF layer usingcollagenase (life technologies), and embryonic body (EB) forma-tion was induced by plating the colonies on nonadhesive plasticculture dishes in human iPSC media without zbFGF. Fifty percentof the media was changed every second day, and 5-day-old EBswere plated on tissue culture plates coated with 0.1 mg/mlpoly(L-ornithine) (Sigma, St. Louis, Missouri, http:www.sigma-aldrich.com). Neural rosette structures started to emerge about 1week after plating. Rosettes were carefully picked with a needle,and isolated clusters were transferred to a nonadhesive cultureplate in DMEM/F12, 2 mM L-glutamine, 1.6 g/l glucose, and N2supplement (1:100) (all life technologies). To establish lt-NEScell lines, floating rosettes were dissociated 2 days later usingtrypsin followed by inhibition of trypsin by trypsin inhibitor (lifetechnologies). Cells were centrifuged for 5 minutes at 300g andplated onto poly(L-ornithine)-and 10 lg/ml laminin (Sigma)-coated plates into the same media supplemented with 10 ng/mlFGF2, 10 ng/ml epidermal growth factor (EGF) (R&D systems,
Minneapolis, Minnesota, http://www.rndsystems.com), and B27(1 ll/ml, life technologies). These iPSC-derived lt-NES cellswere passaged at a ratio of 1:2-1:3 every second to third dayusing trypsin. All transplantation studies were performed with lt-NES cells passages 20-30.
Quantitative Reverse Transcription PolymeraseChain Reaction
Messenger RNA samples were isolated using mRNA extraction kit(Qiagen, Hilden, Germany, http://www.qiagen.com) according tosupplier’s instructions. Adult brain RNA served as positive control(Stratagen, http://www.stratagen.com). One microgram of totalRNA was used for reverse transcription (RT) with the iScriptcDNA synthesis kit (BioRad, Hercules, California, http://www3.bio-rad.com) following the manufacturer’s protocol. QuantitativeRT polymerase chain reaction (PCR) analyses were performed intriplicates on a Biorad iCycler using SYBRI-green detectionmethod. PCR products were assessed by dissociation curve and gelelectrophoresis. Data were normalized to 18S rRNA levels. Primersfor vascular endothelial growth factor (VEGF): forward: 50-cagc-tactgccgtccaatcg-30, reverse: 50-tcaccgcctcggttgt-30.
Middle Cerebral Artery Occlusion in Mice and Rats
All experimental procedures were approved by the Malm€o-LundEthical Committee. Stroke was induced using the intraluminal fil-ament model of middle cerebral artery occlusion (MCAO) inNude rats [28, 29] and C57bl6 mice [30, 31]. Briefly, the rightcommon carotid artery (CCA) and its proximal branches wereisolated. The CCA and external carotid artery (ECA) wereligated, and internal carotid artery (ICA) was temporarily clippedusing a metal microvessel clip. A nylon monofilament wasadvanced through the ICA until resistance was felt. Thirtyminutes after the start of occlusion, the nylon filament was care-fully removed, and ECA was ligated permanently.
Distal MCAO in Rats
Focal ischemic injury in cerebral cortex was induced by permanentocclusion of the distal cortical branch of MCA and transient (30minutes) occlusion of both CCAs in Nude rats, as described previ-ously [32]. Briefly, the temporal bone was exposed. A craniotomyof approximately 3 mm was made, the dura was carefully opened,and the cortical branch of MCA was ligated by suture. Both CCAswere isolated and temporarily ligated. At 30 minutes after the liga-tion, CCAs were released and surgical wounds were closed.
Transplantation
Intracerebral transplantation of human iPSC-derived lt-NES cellswas performed stereotaxically at 1 week and 48 hours after MCAOin mice and rats, respectively. At the day of transplantation, iPSC-derived lt-NES cells transduced with lentivirus carrying green fluo-rescent protein (GFP) and naıve cells for rats and mice, respec-tively, were resuspended to a final concentration of 100,000 cellsper microliter. Coordinates for transplantation sites and volume arepresented in Supporting Information Table S1. Tooth bar was set at�3.3 mm. Mice were given subcutaneous injections of 10 mg/kgCyclosporine A every other day after transplantation.
Retrograde Tracer Injection
At 9 weeks after transplantation or vehicle injection, all mice withMCAO lesions were injected iontophoretically with 2% Fluoro-Gold(FG; Biotium, Hayward, California, http://www.biotium.com), dis-solved in distilled water, into globus pallidus (GP) ipsilateral to thetransplant. A separate group of mice (n ¼ 5) was transplanted withGFP-labeled lt-NES cells into the intact striatum, injected with FGinto GP 2 weeks later, and sacrificed 3 weeks after transplantation,that is, 1 week after FG injection. Iontophoretic injections weremade using microcapillaries with tip diameters of 20–30 lm and 5lA positive current pulsed for 7 out of every 14 seconds during 7minutes using a constant current generator (Midgard; Stoelting,Wood Dale, Illinois, https://www.stoeltingco.com).
Oki, Tatarishvili, Wood et al. 1121
www.StemCells.com

Immunocytochemistry
Cells were fixed in 4% paraformaldehyde for 10–20 minutes atroom temperature, washed in phosphate buffered saline (PBS) andincubated in blocking solution containing 10% FCS and 0.25%Triton X-100 for 1 hour at room temperature. For detection ofGABA, 0,1% glutaraldehyde was added to the fixative. Mice weresacrificed at 10 weeks and rats were sacrificed at 2 weeks, 2 or 4months after transplantation, respectively. Animals were perfusedtranscardially with 4% paraformaldehyde. Coronal sections (30 lm)and cells were preincubated in blocking solution (5% normal serumand 0.25% Triton X-100 in 0.1 M potassium-buffered PBS).Primary antibodies (Supporting Information Table S2) were dilutedin the blocking solution and applied overnight at 4�C. Fluorophore-conjugated secondary antibodies (Molecular Probes, Eugene,Oregon, http://www.invitrogen. com or Jackson Laboratories, BarHarbor, Maine, http://www.jax. org) were diluted in blockingsolution and applied for 2 hours followed by three rinses inpotassium PBS.
Behavioral Tests
The staircase test was used to assess ‘‘side-specific’’ skilled fore-limb reaching and grasping abilities [33, 34] in a staircase appa-ratus (Campden Instruments, Loughborough, U.K., http://www.campden-inst.com). The corridor test, [35] adapted to mice[36], was used to assess sensorimotor impairment caused by stria-tal damage.
Cell Counting and VEGF Immunoreactivity
For quantification of colocalization of HuNuþ cells with differentmarkers, approximately 1,000 HuNuþ cells in each animal wereanalyzed. Double-labeled cells identified in an epifluorescence/light microscope were validated with a confocal microscope(Leica, Solms, Germany, http://www.leica-camera.com). Forquantification of surviving grafted cells and FGþ/HuNuþ cells inmice, all HuNuþ cells in all sections were counted. Total numberof HuNuþ and GFPþ cells in rats were counted stereologicallyby using C.A.S. T-Grid software (Olympus, Tokyo, Japan, http://www.olympus.com) and cell numbers were calculated accordingto the optical fractionator formula [37].
VEGF immunoreactivity was assessed in the astrocytes andblood vessels outside the graft in animals transplanted with lt-NEScells and in the corresponding areas of vehicle-injected animals.Similarly, VEGF immunoreactivity was measured in the graft andin the corresponding area in the vehicle-injected animals. To quan-tify VEGF immunoreactivity, images of different regions of interest(ROIs) in two representative sections from each brain double-stained for VEGF and glial fibrillary acidic protein (GFAP) wereacquired. The area of VEGF-immunoreactivity within GFAPþ andGFAP� cells and in grafted and corresponding vehicle-injectedregions was determined by image analysis using Cellsens Dimen-sion 2010 software (Olympus). In each section, areas of immunore-activity were identified using defined representative ranges ofthreshold for specific signal. Using these defined parameters, theimages of each ROI were analyzed by software, which calculatedthe total area covered by pixels/specific immunopositive signal. Thevalues corresponding to total fluorescence areas were averaged andexpressed as the mean VEGFþ area per animal.
Electrophysiology
Rats were anesthetized with isofluorane and decapitated. Brainswere rapidly removed and placed in ice-cold, gassed (95% O2, 5%CO2) solution (pH 7.2–7.4, 295–300 mosm), containing (in mM):75 sucrose, 67 NaCl, 26 NaHCO3, 1.25 NaH2PO4, 0.5 CaCl2, 7MgCl2, and 25 glucose. Coronal slices (300 lm), cut on a vibra-tome (Leica VT1200S), were placed in an incubation chamber withgassed (95% O2, 5% CO2) artificial cerebral spinal fluid (aCSF, pH7.2–7.4, 295–300 mosm) containing (in mM): 119 NaCl, 2.5 KCl,1.3 MgSO4, 2.5 CaCl2, 26 NaHCO3, 1.25 NaH2PO4, and 25 glu-cose. Slices were constantly perfused with aCSF (32�C–34�C). Re-
cording pipettes were filled with solution (pH 7.2–7.4, 295–300mosm) containing (in mM): 122.5 potassium gluconate, 12.5 KCl,10.0 KOH-HEPES, 0.2 KOH-EGTA, 2.0 Mg-ATP, 0.3 Na3-GTP,and 8.0 NaCl. For measurements of spontaneous inhibitory postsy-naptic currents recording pipettes contained (in mM): 135.0 CsCl,10.0 CsOH, 0.2 CsOH-EGTA, 2.0 Mg-ATP, 0.3 Na3-GTP, 8.0NaCl, and 5.0 lidocaine N-ethyl bromide (QX-314). Biocytin(0.5%; Sigma-Aldrich) was freshly dissolved in the pipette solution.Evoked currents were measured after electrical stimulation (two100 ls square-wave pulses of 150–300 lA timed 50 ms apart)delivered by a single stainless steel wire enclosed in a glass pipettefilled with aCSF (�2 MX resistance). Voltage-gated sodium chan-nels were blocked with 1 lM tetrodotoxin (TTX) (Tocris, Bristol,UK, http://www.tocris.com). Voltage-activated potassium channelswere blocked using tetraethylammonium (TEA) (Sigma). N-methyl-D-aspartic acid (NMDA) and a-amino-3-hydroxy-5-methyl-4-isoxa-zolepropionic (AMPA) receptors were blocked using 50 lM D-AP5and 5 lM 2,3-dihydroxy-6-nitro-7-sulfamoyl-benzo[f]quinoxaline-2,3-dione (NBQX), respectively. GABAA receptors were blockedusing 100 lM picrotoxin (PTX) (Tocris).
RESULTS
Generation and In Vitro Properties of lt-NES Cellsfrom Human iPSCs
Human iPSC lines were produced by standard retroviral trans-duction [38]. Selected clones were further propagated to estab-lish iPSC lines. The iPSC line used in this study showed mor-phologies indistinguishable from those of human ESCs (Fig.1A) and expressed the pluripotency-associated markers (Fig.1B–1D). Following transplantation into immunodeficient mice,the cells formed teratomas in vivo consisting of all three germlayers (Fig. 1E–1G) and had a normal karyotype (Fig. 1H).
A homogeneous population of iPSC-derived lt-NES cellswas established using our previously established protocol[27]. In the presence of the growth factors FGF2 and EGF,these cells can be propagated along multiple passages withoutlosing their stem cell characteristics or their differentiationpotential. Human iPSC-derived lt-NES cells (Fig. 1I) uni-formly expressed the NSC-associated markers Sox2 and nestin(Fig. 1J, 1K). They assembled into rosette-like patterns withexpression of the rosette-associated transcription factors PLZFand Dach1 and tight junction maker ZO-1 (Fig. 1K, 1L).Upon growth factor withdrawal and independent of passagenumber, human iPSC-derived lt-NES cells differentiated intoneurons (>70%) and astrocytes (Fig. 1M–1P).
Transplantation of Human iPSC-Derived lt-NESCells into the Stroke-Damaged Mouse StriatumImproves Recovery of Fine Forelimb MovementsIndependent of Long-Term Graft Survival
We first explored whether implantation of human iPSC-derived lt-NES cells could influence the behavioral recoveryafter stroke. Twenty-three mice subjected to MCAO were di-vided into two groups and transplanted into the striatum withhuman iPSC-derived lt-NES cells (n ¼ 12) or vehicle (n ¼11) at 1 week after the ischemic insult. A third group ofsham-operated animals (n ¼ 9) was given an intrastriatal ve-hicle injection. We evaluated fine forelimb movement usingthe staircase test at 2, 5, and 9 weeks after MCAO or shamsurgery, that is, 1, 4, and 8 weeks after transplantation. In thistest, the number of retrieved pellets reflects the ability tomove the forelimb, whereas the number of eaten pelletsdepends on the ability of both grasping and eating pellets,requiring fine and well-coordinated forelimb movements.
1122 Human iPSCs in Stroke-Damaged Brain

Vehicle-injected, stroke-affected mice showed impairment inboth parameters (Fig. 2A, 2B), whereas animals with humaniPSC grafts did not exhibit any deficits at any time point andperformed significantly better than vehicle-treated animals.Taken together, our data indicate that the human iPSC-derivedlt-NES cells gave rise to sustained improvement of fine fore-paw movement, probably starting already early after trans-plantation. In contrast, human iPSC-derived lt-NES cell trans-plantation did not reverse the deficit detected in the corridortest (Fig. 2C).
We next determined the location, survival, and prolifera-tive activity of the grafted cells. The MCAO caused a repro-ducible lesion of the dorsolateral part of the striatum in allmice (Fig. 3A) [39]. Surviving grafts were found in seven outof 12 animals (Fig. 3B). The vast majority of the human cellswere located in the striatum. A few HuNuþ cells were alsodetected in cerebral cortex and corpus callosum. The numberof HuNuþ cells in each graft was 9,378 6 2,517, that is,approximately 10% of the initial number of transplanted cells.
At 10 weeks after transplantation, only few HuNuþ cells(1.6% 6 0.9%) kept proliferating and were positive for Ki67(Fig. 3C–3E).
We observed no differences in the pattern or location ofthe stroke-induced damage between the human iPSC-derivedlt-NES cell- and vehicle-implanted groups. At 11 weeks afterthe insult, the volume of the injured striatum did not differbetween cell-transplanted and vehicle-injected groups (4.19 60.6 mm3 and 4.69 6 0.5 mm3, respectively). In five out of 12animals implanted with human iPSC-derived lt-NES cells, nosurviving grafts were detected 10 weeks after transplantation.We reanalyzed our behavioral data and compared the impair-ment and magnitude of recovery between the groups with andwithout surviving grafts. The two groups showed no signifi-cant difference in number of retrieved or eaten pellets or inthe corridor test (data not shown). These findings suggest thatlong-term survival of grafted cells is not necessary for thefunctional recovery induced by iPSC-derived lt-NES cellsduring the first 2 months after transplantation.
Figure 1. Generation of the human-induced pluripotent stem cell (iPSC)-derived long-term expandable neuroepithelial-like stem (lt-NES) cells.(A–H): Skin fibroblast-derived iPSC colonies (A) stain positive for the pluripotency-associated markers alkaline phosphatase (B), Tra1-60 (C),and Tra1-81 (D). Following transplantation into immunodeficient mice, the iPSCs form teratomas consisting of cell types of all three germ layerssuch as neuroectodermal rosettes (E), glandular structures (F), and cartilage (G). Single nucleotide polymorphism analysis revealed no significantkaryotypic abnormalities (F). For each chromosome, the B allele frequency (upper row) and log A ratio (lower row) are shown. Long-term self-renewing neuroepithelial stem cells generated from the iPSCs (I) express the NSC markers Sox2 (J) and nestin (K) and the rosette-associatedtranscription factors Dach1 (K) and PLZF (L). The tight junction protein ZO-1 is expressed apically in the rosette-like structures (L). Upongrowth factor withdrawal, iPSC-derived lt-NES cells differentiate to a major proportion of neurons (M), expressing beta III-tubulin (N) andMAP2 (O). A minor fraction of GFAP-positive astrocytes is also present in the cultures (N). Many of the neurons are immunoreactive for GABA(P). (A), (I), and (M) are phase-contrast images. Scale bars ¼ 50 lm (L); 100 lm (J, K, N); 200 lm (C, D, O, P). Abbreviation: GFAP, glialfibrillary acidic protein. [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
Oki, Tatarishvili, Wood et al. 1123
www.StemCells.com

Human iPSC-Derived lt-NES Cells Differentiate toNeurons and Establish Axonal Connections with theGP After Transplantation in Stroke-DamagedMouse Brain
We assessed the proportion of grafted cells expressing neuronalmarkers such as doublecortin (DCX, a marker for neuroblasts)and HuD (a marker for young and mature neurons) at 10 weeksafter transplantation (Fig. 3C–3E). The majority of grafted cellswere HuNuþ/HuDþ (78.5% 6 11.2%) and mainly located inthe center of the graft. A smaller portion of the cells wasHuNuþ/DCXþ (13.4% 6 4.7%), with preferential location at
the periphery of the graft. Thus, most of the grafted humaniPSC-derived lt-NES cells had differentiated to neurons.
We then examined whether neurons generated from humaniPSC-derived lt-NES cells send axonal projections to the appro-priate location in the host brain. We injected the retrogradetracer FG into the GP (Fig. 3F) ipsilateral to the graft (Fig. 4H)at 9 weeks after transplantation (1 week prior to sacrifice). Infive animals, FG staining was confined to GP (Fig. 4G). Inthese animals, a small fraction of HuNuþ cells in the striatalgrafts was stained with FG (1.16% 6 0.16%; Fig. 3I–3M).Axons were occasionally observed to extend from HuNuþ/FGþ cells (Fig. 3I). In addition, many FGþ cells were found inthe remaining intact striatum ipsilateral to the lesion, demon-strating the projection of axons from host striatal neurons to GP(Fig. 3I, 3J). In the control group in which the grafted lt-NEScells were allowed to differentiate only for 3 weeks, weobserved many FGþ cells throughout the whole striatum (Fig.3N). However, we detected no FG-labeled GFPþ lt-NES cellsin the transplant providing evidence that 3 weeks are not suffi-cient for the grafted neurons to extend their axons to GP. Ourfindings also provide further support for the idea that the FGlabeling observed in the main experiment is due to retrogradeaxonal transport of FG from the GP to the grafted lt-NES cellsin striatum and not due to passive diffusion. Our findings pro-vide evidence that the neurons generated from the human iPSC-derived lt-NES cells, implanted into the stroke-damaged mousestriatum, can extend their axons to the GP, which is the mainprojection area for striatal medium-sized spiny neurons.
Human iPSC-Derived lt-NES Cells InduceIncreased Vascular Endothelial Growth FactorImmunoreactivity in Blood Vessels Early AfterTransplantation in Stroke-Damaged Mouse Brain
In order to explore possible mechanisms underlying theobserved functional improvements, mice were transplantedinto striatum with either human iPSC-derived lt-NES cells (n¼ 5) or vehicle (n ¼ 6) at 1 week after stroke and sacrificed1 week later, that is, at the earliest time point when theenhanced recovery was observed. Stereological analysisrevealed no differences in the area of neurogenic subventricu-lar zone cells immunopositive to DCX and BrdU (data notshown). We also obtained no evidence that the grafted cellshad influenced the number of activated microglia (Iba1þ/ED1þ) or astrocytes (GFAPþ) in the striatum.
Figure 2.
Figure 2. Transplantation of human-induced pluripotent stem cell-derived long-term expandable neuroepithelial-like stem cells intostriatum of stroke-damaged mice induces behavioral improvement.(A–C): Comparisons between cell-implanted (‘‘cell’’; n ¼ 12) and ve-hicle-injected (‘‘vehicle’’; n ¼ 11) mice subjected to stroke and sham-operated, nontransplanted mice (‘‘sham’’; n ¼ 9) in performance inthe staircase (A, B) and corridor tests (C). Performance for the stair-case test was calculated as the number of pellets on the impaired sidedivided by the total number of pellets on both sides and expressed aspercentage of performance at baseline. Performance for the corridortest was calculated by dividing the number of contralateral retrievalswith the total number of retrievals from both sides. Both numbers ofeaten (A) and retrieved pellets (B) are higher in the cell group ascompared to the vehicle group, the performance in the stroke-dam-aged animals receiving cell grafts being similar to that in the sham-operated group. In contrast, cell grafts did not improve performancecompared to vehicle injections in the corridor test (C). Data aremeans 6 SEM. Repeated measures analysis of variance (ANOVA)between cell and vehicle group: (A) p ¼ .0080, (B) p ¼ .0437, and(C) p ¼ .9853 with *, p < .05; **, p < .01. Repeated measuresANOVA between vehicle and sham group: (A) p ¼ .0472, (B) p ¼.0193, and (C) p ¼ .0134 with *, p < .05.
1124 Human iPSCs in Stroke-Damaged Brain
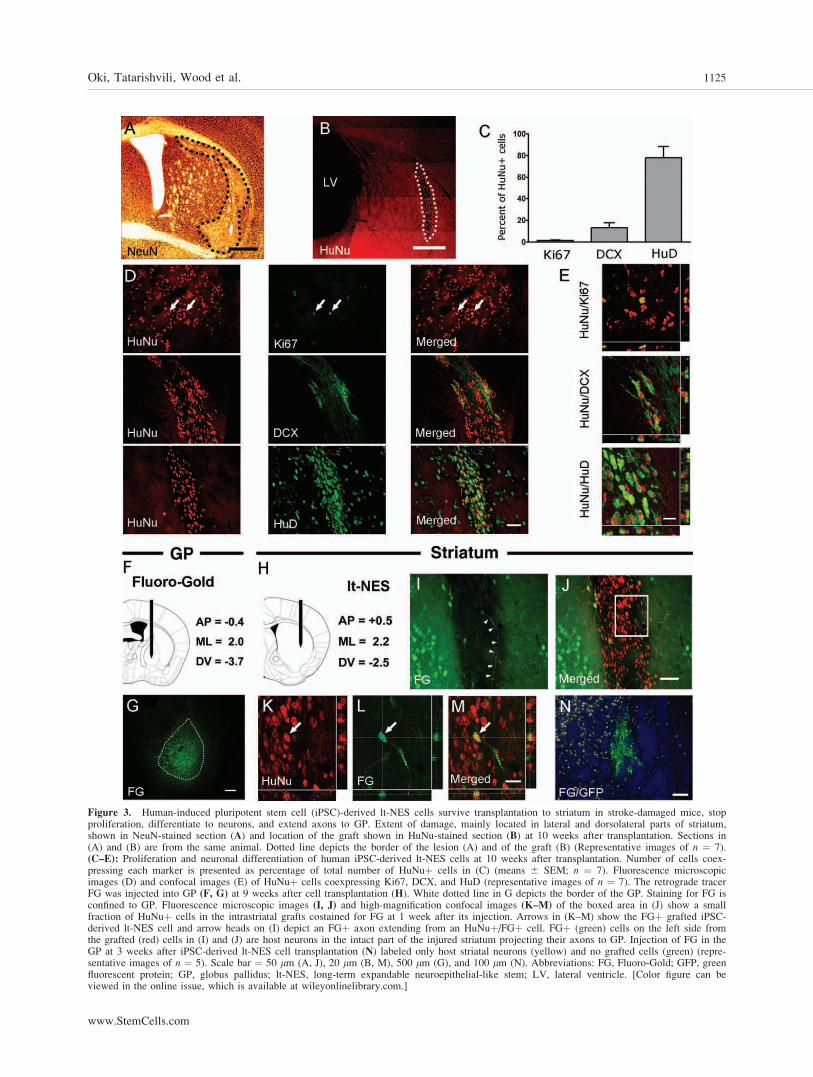
Figure 3. Human-induced pluripotent stem cell (iPSC)-derived lt-NES cells survive transplantation to striatum in stroke-damaged mice, stopproliferation, differentiate to neurons, and extend axons to GP. Extent of damage, mainly located in lateral and dorsolateral parts of striatum,shown in NeuN-stained section (A) and location of the graft shown in HuNu-stained section (B) at 10 weeks after transplantation. Sections in(A) and (B) are from the same animal. Dotted line depicts the border of the lesion (A) and of the graft (B) (Representative images of n ¼ 7).(C–E): Proliferation and neuronal differentiation of human iPSC-derived lt-NES cells at 10 weeks after transplantation. Number of cells coex-pressing each marker is presented as percentage of total number of HuNuþ cells in (C) (means 6 SEM; n ¼ 7). Fluorescence microscopicimages (D) and confocal images (E) of HuNuþ cells coexpressing Ki67, DCX, and HuD (representative images of n ¼ 7). The retrograde tracerFG was injected into GP (F, G) at 9 weeks after cell transplantation (H). White dotted line in G depicts the border of the GP. Staining for FG isconfined to GP. Fluorescence microscopic images (I, J) and high-magnification confocal images (K–M) of the boxed area in (J) show a smallfraction of HuNuþ cells in the intrastriatal grafts costained for FG at 1 week after its injection. Arrows in (K–M) show the FGþ grafted iPSC-derived lt-NES cell and arrow heads on (I) depict an FGþ axon extending from an HuNuþ/FGþ cell. FGþ (green) cells on the left side fromthe grafted (red) cells in (I) and (J) are host neurons in the intact part of the injured striatum projecting their axons to GP. Injection of FG in theGP at 3 weeks after iPSC-derived lt-NES cell transplantation (N) labeled only host striatal neurons (yellow) and no grafted cells (green) (repre-sentative images of n ¼ 5). Scale bar ¼ 50 lm (A, J), 20 lm (B, M), 500 lm (G), and 100 lm (N). Abbreviations: FG, Fluoro-Gold; GFP, greenfluorescent protein; GP, globus pallidus; lt-NES, long-term expandable neuroepithelial-like stem; LV, lateral ventricle. [Color figure can beviewed in the online issue, which is available at wileyonlinelibrary.com.]
Oki, Tatarishvili, Wood et al. 1125
www.StemCells.com

It has been reported that human fetal NSCs through secre-tion of VEGF can improve behavioral performance already at1 week after implantation and 2 weeks after stroke [11]. Wefound VEGF expression in the stroke-affected hemisphere ofboth lt-NES cell-transplanted and vehicle-injected animals,but the VEGF-immunoreactive areas were larger in animalsimplanted with lt-NES cells. Microscopic examinationrevealed that activated microglia did not express VEGF,which was confined to astrocytes and blood vessels (Fig. 4A–4F). VEGF was expressed both in the presumed astrocyticendfeet and the vessel wall (Fig. 4A–4F). The vast majorityof GFAPþ astrocytes were positive for VEGF (Fig. 4A–4C)with no difference between the groups (90.9% 6 4.3% and96.6% 6 2.8% in cell-grafted and vehicle-injected mice,respectively), and approximately 85% of VEGF immunoreac-
tivity outside the graft was found in GFAPþ cells. Computer-based, unbiased analysis of immunostained sections revealedthat in animals transplanted with lt-NES cells, the area ofVEGF immunoreactivity in astrocytes was more than twofoldhigher as compared to vehicle-injected animals (Fig. 4G). Thearea of VEGF immunoreactivity in CD31þ blood vessels wasonly approximately 10% of that in astrocytes, and lt-NES-transplanted animals did not show any significant differencein VEGF immunoreactivity compared to vehicle-injected ani-mals (Fig. 4G). Also many grafted lt-NES cells exhibitedVEGF immunoreactivity (Fig. 4K–4O). This is in agreementwith our in vitro data on VEGF gene expression in humaniPSC-derived lt-NES cells (Fig. 4H). VEGF immunoreactivityin the graft was much higher than in the corresponding areain vehicle-injected animals (Fig. 4G, 4I, 4J).
Figure 4. Human-induced pluripotent stem cell (iPSC)-derived lt-NES cell grafts in stroke-damaged mouse striatum contain VEGF-expressing cellsand induce increased VEGF immunoreactivity in host blood vessels and astrocytes. Fluorescence photomicrographs (A–F, I, J) and confocal images(K–O) of GFAPþ astrocytes (A, arrows), CD31þ vessels (D, arrowheads), and SC121þ grafted human iPSC-derived lt-NES cells (L, N) coexpress-ing VEGF (B, C, E, F, I, J, K, M and O), at 2 weeks after stroke and 1 week after transplantation. Area of VEGF immunoreactivity in the region ofinterest (n ¼ 4, in three representative sections) in stroke-damaged mouse brain after vehicle injection (n ¼ 4) or human iPSC-derived lt-NES celltransplantation (n ¼ 6) (G; Student’s unpaired t test, *, p < .05; means 6 SEM). Relative expression of VEGF mRNA in human iPSC-derived lt-NES cells, human fibroblasts, and adult brain (brain tissue from 66-year-old female) using quantitative reverse transcription polymerase chain reac-tion with expression in the adult brain set to 1 (H). (H; one-way ANOVA followed by Bonferroni post hoc test; *, p < .05; means 6 SEM). Meanexpression levels of four independent experiments run in technical triplicates. Scale bar ¼ 20 lm (A–C), 20 lm (D–F), and 50 lm (I, J). Abbrevia-tions: GFAP, glial fibrillary acidic protein; lt-NES, long-term expandable neuroepithelial-like stem; VEGF, vascular endothelial growth factor. [Colorfigure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
1126 Human iPSCs in Stroke-Damaged Brain

We then explored whether the increased VEGF immuno-reactivity observed at 2 weeks after transplantation of lt-NEScells had influenced vasculogenesis. Stereological estimationof vessel length density and measurement of CD31 immuno-reactivity were performed in CD31-stained sections of lt-NEScell-grafted and vehicle-injected mice at 9 weeks after stroke.At this time point, when the lt-NES grafted animals exhibitedsignificant behavioral improvement, we did not detect any dif-ferences between lt-NES cell-grafted and vehicle-injectedgroups either in vessel length density or in CD31 immunore-activity (data not shown).
Human iPSC-Derived lt-NES Cells Survive for 4Months and Differentiate into Neurons AfterTransplantation into Striatum ofT-Cell-Deficient Rats
We wanted to determine whether the human iPSC-derived lt-NES cells and their progeny could survive longer than 10
weeks, differentiate to specific neuronal subtypes, and exhibitthe electrophysiological properties of functional neurons. Wetherefore implanted the cells in Nude, T-cell-deficient adultrats in which xenotransplanted human cells can survive long-term without immunosuppression.
Rats were subjected to intrastriatal transplantation ofGFP-labeled human iPSC-derived lt-NES cells at 48 hours af-ter MCAO and were perfused 2 weeks (n ¼ 3), and 2 (n ¼ 8)and 4 (n ¼ 5) months thereafter. We could not detect humaniPSC-derived lt-NES cell grafts in two rats. The total numberof grafted cells was estimated at 64.8% 6 12.4% and 51.2%6 13.3% at 2 and 4 months after transplantation, respectively,as compared to the number of implanted cells. The prolifera-tive activity in the grafts (Fig. 5A–5D), quantified as the per-centage of HuNuþ cells coexpressing Ki67, was 40.2% 64.9% at 2 weeks after transplantation but dropped to 8.2 60.5 and 1.0% 6 0.2% at 2 and 4 months, respectively. In linewith this observation, we did not detect any overgrowth ofgrafted cells up to 4 months after transplantation.
Figure 5. Proliferative activity and number of neuroblasts generated from human-induced pluripotent stem cell-derived long-term expandable neuroepi-thelial-like stem cells decrease after transplantation into stroke-damaged rat striatum but maintain multipotency. Fluorescence photomicrographs (A–C, E–G, I–K) and confocal image (D, H, L) of HuNuþ cells coexpressing the mitotic marker Ki67 (arrows) at 2 weeks (A), 2 months (B), and 4 months (C)and neuroblast marker DCX at 2 (E–H) and 4 (I–L) months postgrafting, respectively (representative images of n ¼ 3, 6, 5 at 2 weeks, 2 months, and 4months, respectively). Confocal images with orthogonal reconstruction of grafted cells giving rise to a mature neuron (GFPþ/NeuNþ; M–O) or an astro-cyte (HuNuþ/GFAPþ; P–R) after intrastriatal transplantation. Confocal images of intrastriatally (S–U, X) or intracortically (V and W) implanted cells,showing GFP autofluorescence (S–V) or stained with the human-specific antibody SC121 (W, X) and exhibiting the morphology of mature neurons (S, V,W), neuroblasts (T, X), or an astrocyte (U) (representative images of n ¼ 5 in striatum and n ¼ 5 in cortex). Scale bar ¼ 50 lm (A–C), 10 lm (D, H,L), 20 lm (E–G, I–K), 10 lm (M–R), and 20 lm (S–X). Abbreviations: GFAP, glial fibrillary acidic protein; GFP, green fluorescent protein. [Color figurecan be viewed in the online issue, which is available at wileyonlinelibrary.com.]
Oki, Tatarishvili, Wood et al. 1127
www.StemCells.com

Microscopic examination of the sections revealed that at 2months after transplantation, the entire core of the graft andmost likely the majority of grafted cells were DCXþ (Fig.5E–5H). In contrast, at 4 months, only a few grafted cellsexhibited DCX immunoreactivity (Fig. 5I–5L). Because ofcross-reactivity between the HuNu and the mature neuronalmarker NeuN antibodies, the neuronal phenotype of graftedcells was determined using GFP and NeuN double-immuno-staining (Fig. 5M–5O). At 2 and 4 months after implantation,65.7% 6 3.9% and 72.9% 6 4.7% of GFPþ cells in striatum,respectively, coexpressed the mature neuronal marker NeuN.Only a fraction of grafted human iPSC-derived lt-NES cellswas GFAPþ astrocytes (3.8% 6 0.5% and 6.1% 6 1.5% at 2and 4 months after transplantation, respectively; Fig. 5P–5R).These cells were distributed mainly at the periphery of thegraft core.
Human iPSC-Derived Lt-NES Cells Grafted intoStriatum Differentiate to Neurons of DifferentPhenotype
To characterize the morphological properties of the graftedcells and their progeny, we used immunocytochemical stain-ing for GFP and for the human cytoplasmic marker SC121.The SC121 antibody labels >90% of human neural cells anddoes not cross-react with rat cells [7, 9]. Grafted GFPþ cellsthat exhibited clear morphological characteristics of migratingneuroblasts (Fig. 5T, 5X), mature neurons (Fig. 5S, 5V, 5W),or astrocytes (Fig. 5U) were more easily detectable in theareas outside the graft core. However, GFPþ cells with typi-cal neuronal morphology were often not immunopositive forNeuN.
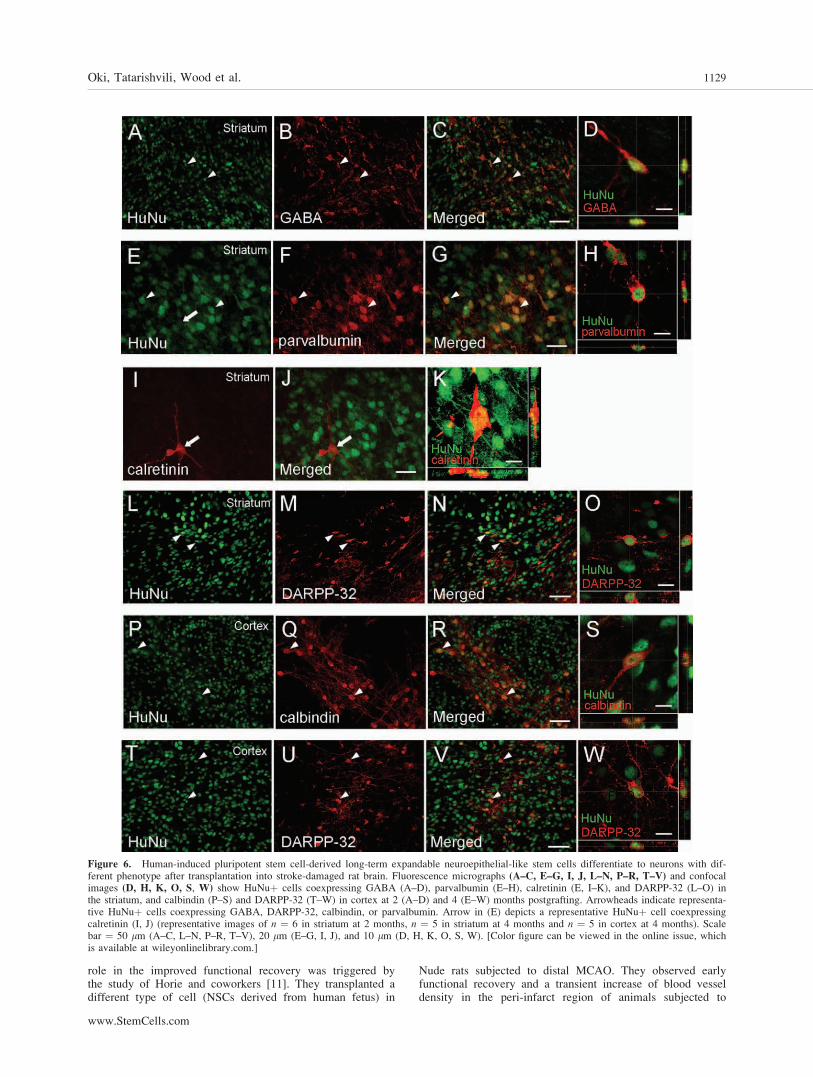
Many of the grafted cells showed GABA immunoreactiv-ity (Fig. 6A–6D; 21.9% 6 1.8% and 10.9% 6 0.6% at 2 and4 months, respectively). At 4 months after transplantation,grafted cells also stained for markers of GABAergic inter-neurons, parvalbumin, and calretinin (Fig. 6E–6K). Few(3.6% 6 0.7%) of the grafted cells were calretininþ (Fig. 6I–6K), whereas a substantial proportion was HuNuþ/parvalbuminþ (15.4% 6 2.6%) (Fig. 6E–6H). Interestingly,at 4 months, a small population (4.3% 6 1.3%) of theHuNuþ cells in the intrastriatal grafts expressed DARPP-32(Fig. 6L–6O), a specific marker of medium-sized spiny striatalprojection neurons.
Human iPSC-Derived lt-NES Cells Survive for4 Months and Differentiate to Neurons AfterTransplantation into Cerebral Cortex ofT-Cell-Deficient Rats
We analyzed the survival and neuronal differentiation ofhuman iPSC-derived lt-NES cells implanted into the cortex ofanimals subjected to the occlusion of the distal branchof MCA (dMCAO), which causes cortical damage. Seven outof 10 rats that survived these procedures were killed after2 (n ¼ 2) or 4 (n ¼ 5) months. The number of cells in thegrafts at 2 and 4 months after transplantation was estimated at80.8% 6 6.2% and 60.1% 6 10.6%, respectively, comparedto the number of implanted cells. The percentage of cellswith proliferative activity resembled that of the intrastriatalgrafts (7.2% 6 1.5% and 0.9% 6 0.2%, in 2- and 4-monthgroups, respectively). NeuN was expressed by 71.8% 6 8.6%of the GFPþ cells and 15.5% 6 2.7% of HuNuþ cells wasalso GABAþ at 2 months. Many of the grafted cells dis-played neuronal morphology as shown by GFP (Fig. 5V) andSC-121 (Fig. 5W) immunostainings. Although the percentageof GFPþ/NeuNþ cells remained high at 4 months after trans-plantation (77.2% 6 3.2%), the GABAþ cells in the grafts
decreased to 8.8% 6 0.8%. At this time point, we found pop-ulations of parvalbuminþ (11.9% 6 1%), calretininþ (2.0%6 0.5%), and calbindinþ cells (8.1% 6 0.8%; Fig. 6P–6S) inthe intracortical grafts. Also when transplanted into cerebralcortex, the iPSC-derived lt-NES cells differentiated to cellsexpressing DARPP-32 (Fig. 6T–6W). The percentage ofgrafted HuNuþ/DARPP-32þ cells (5.2% 6 1.5%) resembledthat observed in the intrastriatal grafts.
Neurons Generated from Grafted HumaniPSC-Derived lt-NES Are Functional andReceive Synaptic Input from Host Neurons
We finally studied the electrophysiological properties of cellsgenerated from iPSC-derived lt-NES cells at 5 months aftertransplantation into striatum or cerebral cortex of Nude ratsby performing whole-cell patch-clamp recordings in acutebrain slice preparations (Fig. 7A–7G). Grafted cells were clas-sified as neuronal or glial based on resting membrane poten-tial and input resistance (Supporting Information Table S3). Aproportion of the cells was functionally mature neurons witha resting membrane potential of approximately �50 mV and�58 mV in the striatum and cortex, respectively, the majoritygenerated action potentials in response to depolarizing currentinjection (Fig. 7H, 7M). In voltage-clamp configuration, depo-larizing voltage steps induced characteristic Naþ and Kþwhole-cell currents, which were sensitive to the voltage-gatedNaþ channel blocker, TTX, and the voltage-gated Kþ chan-nel blocker, TEA, respectively (Fig. 7I, 7N).
Spontaneous postsynaptic currents were frequentlyobserved in grafted cells in the striatum (Fig. 7J, 7O). Sponta-neous excitatory postsynaptic currents (sEPSCs) wererecorded in the presence of the GABAA receptor antagonist,PTX (Fig. 7K, 7Q), and addition of glutamate receptor antag-onists NBQX and D-AP5 blocked all postsynaptic currents(Fig. 7L), indicating that the cells had functional excitatorysynapses. Interestingly, spontaneous inhibitory postsynapticcurrents were not observed (n ¼ 5 cells).
In addition, we detected miniature postsynaptic currentsthat were action potential independent (not blocked by TTX)(Fig. 7P). In two of 10 grafted cells, excitatory, AMPA recep-tor-mediated currents could be evoked by stimulating a corti-cal region remote from the transplant (Fig. 7S). This findingsuggests that transplanted neurons generated from iPSC-derived lt-NES cells receive synaptic input from host neuronsand functionally integrate into host brain neural circuitries.
DISCUSSION
We show here that intrastriatal transplantation of lt-NEScells, derived from human iPSCs, gives rise to improved re-covery of a clinically relevant motor deficit. Although wefound that at 10 weeks the majority of cells in the survivinggrafts expressed neuronal markers, recovery was probablypromoted earlier, before any functioning neurons could havebeen developed from the grafted human iPSC-derived lt-NES cells. Most likely, this early improvement is due tomechanisms other than neuronal replacement. In support ofthis interpretation, the enhancement of behavioral recoveryin our experiment was observed irrespective of whether thegrafts were surviving at 10 weeks or not. Similarly, humaniPSCs improved performance in the cylinder test from 4days after intrastriatal implantation into stroke-damaged rats[25].
Our hypothesis that the increased levels of VEGF in thelt-NES cell-transplanted animals could have an important
1128 Human iPSCs in Stroke-Damaged Brain
role in the improved functional recovery was triggered bythe study of Horie and coworkers [11]. They transplanted adifferent type of cell (NSCs derived from human fetus) in
Nude rats subjected to distal MCAO. They observed earlyfunctional recovery and a transient increase of blood vesseldensity in the peri-infarct region of animals subjected to
Figure 6. Human-induced pluripotent stem cell-derived long-term expandable neuroepithelial-like stem cells differentiate to neurons with dif-ferent phenotype after transplantation into stroke-damaged rat brain. Fluorescence micrographs (A–C, E–G, I, J, L–N, P–R, T–V) and confocalimages (D, H, K, O, S, W) show HuNuþ cells coexpressing GABA (A–D), parvalbumin (E–H), calretinin (E, I–K), and DARPP-32 (L–O) inthe striatum, and calbindin (P–S) and DARPP-32 (T–W) in cortex at 2 (A–D) and 4 (E–W) months postgrafting. Arrowheads indicate representa-tive HuNuþ cells coexpressing GABA, DARPP-32, calbindin, or parvalbumin. Arrow in (E) depicts a representative HuNuþ cell coexpressingcalretinin (I, J) (representative images of n ¼ 6 in striatum at 2 months, n ¼ 5 in striatum at 4 months and n ¼ 5 in cortex at 4 months). Scalebar ¼ 50 lm (A–C, L–N, P–R, T–V), 20 lm (E–G, I, J), and 10 lm (D, H, K, O, S, W). [Color figure can be viewed in the online issue, whichis available at wileyonlinelibrary.com.]
Oki, Tatarishvili, Wood et al. 1129
www.StemCells.com

Figure 7. Human-induced pluripotent stem cell-derived long-term expandable neuroepithelial-like stem (lt-NES) cells have developed electro-physiological properties of functional neurons at 4.5–5.5 months after transplantation into intact rat striatum and cortex. Whole-cell configurationof GFP-expressing lt-NES cells (A–C). Photomicrograph of a grafted in the striatum (D) and cortex (F) and recorded cells (in the boxed area) la-beled with biocytin and GFP. Enlargement of boxed areas in (D) and (F) showing (arrows) grafted GFPþ cell filled with biocytin after recording(E, G). Patch-clamp recordings from the cells grafted in the striatum (H–L) and the cortex (M–S). Representative traces of membrane potentialresponses (H, M, striatum and cortex, respectively) to step injection of hyperpolarizing and depolarizing current (10 pA steps) showing actionpotentials that are blocked by TTX (right). Representative traces of whole-cell Naþ and Kþ currents (I, N), blocked using TTX and TEA (I),respectively, elicited by voltage steps from �70 mV to þ40 mV in 10 mV increments. Representative traces of spontaneous (J, O) and excitatory(K, Q) postsynaptic currents recorded in the presence of PTX, in voltage-clamp configuration at �70 mV. Absence of postsynaptic currents afteraddition of PTX and glutamate receptor antagonists, NBQX and D-AP5 (L) or only NBQX (R). Representative traces of miniature (P) postsynap-tic currents recorded in the presence of TTX, in voltage-clamp configuration at �70 mV. Postsynaptic AMPA-receptor-mediated currents (S) areevoked by paired-pulse electrical stimulation delivered from a stainless-steel electrode placed approximately 300 lm away from the transplant inthe cortex (three representative traces superimposed; * indicates stimulation artifact). Insets show respective traces on an expanded scale. A total25 cells were recorded from four rats (Supporting Information Table S3). Abbreviations: aCSF, artificial cerebral spinal fluid; Ctx, cortex; DIC,differential interference contrast; GFP, green fluorescent protein; LV, lateral ventricle; PTX, picrotoxin; Str, striatum; TTX, tetrodotoxin. [Colorfigure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
1130 Human iPSCs in Stroke-Damaged Brain

transplantation. Secretion of VEGF by the NSCs was foundto be important for the suppression of inflammation and neo-vascularization in the peri-infarct region probably underlyingthe improved recovery [11]. Our data provide no evidencethat the elevated VEGF immunoreactivity observed at 2weeks after stroke in lt-NES cell-grafted animals gives riseto a long-lasting increase of blood vessel length density.Therefore, it is conceivable that mechanisms other thanVEGF-induced vascularization and angiogenesis are responsi-ble for the long-term behavioral improvement observed inour lt-NES cell-grafted animals. We hypothesize that VEGFleads to early and long-lasting behavioral improvement byinfluencing brain plasticity in the postischemic phase [40].In line with such a role of VEGF, it was recently reportedthat conditional transient increase of VEGF expression for5–15 days leads to significant increase of long-term potentia-tion in the hippocampus through modulation of synapticplasticity in mature neurons [41]. In addition, VEGF hasbeen implicated as a mediator of human NSC-inducedeffects on dendritic sprouting, axonal plasticity, and axonaltransport [15]. It should be emphasized, although, thatincreased VEGF signaling is only one possible explanationfor the beneficial effects and other mechanisms, not exploredin this study, could be responsible for the observedimproved behavioral performance.
One can speculate that the VEGF immunoreactivityobserved in the host tissue is due to VEGF released from thegrafted lt-NES cells. However, the widespread distribution ofVEGF immunoreactivity far away from the transplant withoutany clear gradient, with very little if any extracellular appear-ance and with close association to astrocytes and blood ves-sels, makes this suggestion unlikely.
The studies with human iPSCs in stroke published sofar provided no evidence for the formation of neuroblastsor morphologically mature neurons [25, 26]. We found thatat 4 months after human iPSC-derived lt-NES cells hadbeen implanted into the striatum or cerebral cortex of T-cell-deficient rats in two different models of stroke, 73%and 77%, respectively, of grafted cells expressed themature neuronal marker NeuN. These data indicate thatgrafted iPSC-derived lt-NES cells can efficiently differenti-ate to mature neurons in vivo and survive long-term in thestroke-damaged brain.
The grafted cells exhibited characteristic neuronal mor-phologies and expressed markers providing evidence that theyhad differentiated to various subtypes of neurons expressingGABA, parvalbumin, calretinin, or calbindin. At 4 months,the grafts also contained a population of cells expressing thespecific striatal projection neuron marker DARPP-32 not onlyin the striatum but also in the cortex, where only weaklystained DARPP-32þ neurons are normally seen [42]. Two im-portant conclusions of relevance for cell replacement strat-egies can be drawn from our data. First, that the spontaneousneuronal differentiation of the grafted human iPSC-derived lt-NES cells in vivo is determined by intrinsic mechanisms andnot significantly affected by external cues. Second, that effec-tive neuronal replacement in different stroke-damaged brainregions will require directed in vitro differentiation of thehuman iPSC-derived lt-NES cells to precursors, which giverise to the lost types of neurons after transplantation.
Our electrophysiological data show, for the first time,that transplanted NSCs generated from reprogrammed adulthuman fibroblast-derived iPSCs can differentiate into func-tional neurons in vivo and receive proper connections fromthe host brain. Similarly, grafted human ESC-derived NSCshave previously been shown to differentiate to cells withelectrophysiological properties of neurons and synaptic
input from host neurons in a stroke-injured environment[6]. We found here that at about 5 months after transplan-tation into striatum or cortex, grafted cells exhibited theelectrophysiological properties of mature neurons. The find-ings of sEPSCs, which were blocked by glutamate receptorantagonists, and of AMPA-receptor-mediated currentsevoked by stimulating a cortical region remote from theintracortical graft, provide evidence that the grafted neu-rons had developed excitatory synaptic input and were, atleast partly, functionally integrated into host neuralcircuitries.
Our findings using the retrograde tracer FG indicated thatgrafted cells have capacity to establish axonal projection tothe GP. This brain region is a major projection area of striatalneurons, and following a stroke, there is extensive injury tothe striatopallidal system. Therefore, the ability of human-derived iPSCs to reestablish this system could potentially beimportant for restoration of function. Our data further illus-trate the remarkable capacity of neurons derived from xeno-grafted human iPSCs [19], ESCs [43], and fetal brain tissue[44] to extend specific, long axonal projections to their appro-priate target areas in the rodent brain.
Tumor formation and graft overgrowth [45–47] from plu-ripotent stem cells and their derivatives are major hurdlesfor the application of stem cell therapy in stroke and otherbrain diseases. In previous studies, human iPSCs showedhigh tumorigenicity after intracerebral implantation instroke-damaged brain [24]. Here, we used transplantation ofiPSCs-derived lt-NES cells into immunocompromised, T-cell-deficient rats subjected to stroke and a 4 months obser-vation period to ensure lack of rejection of the xenotrans-planted cells and to create optimum environment for assess-ing their tumorigenicity. No tumor formation or transplantovergrowth was detected in any animal grafted with iPSC-derived lt-NES cells. Our data suggest that predifferentiationof iPSCs and generation of long-term self-renewing neuralcell lines is an effective strategy for minimizing the risk fortumor formation.
CONCLUSION
Our experimental study provides evidence that transplantationof human fibroblast-derived iPSCs is a new, promisingapproach for promoting functional recovery after ischemicstroke. Most importantly, our findings demonstrate for the firsttime the potential of iPSCs for neuronal replacement in thestroke-damaged brain. The iPSC-derived grafts survived long-term and contained a high proportion of cells with morpho-logical and electrophysiological properties of neurons. Theseneurons received afferent inputs from the host brain andextended their axons to an appropriate target area. Before anyclinical application can be considered, the mechanisms regu-lating the differentiation of the iPSC-derived lt-NES cells intospecific neuron types in the stroke-damaged brain have to bemuch better understood and effectively controlled, and the be-havioral recovery induced by neuronal replacement has to beoptimized.
ACKNOWLEDGMENTS
We thank Dr. Vladimer Darsalia and Vivek Verma for their con-tribution in initial stages of the study, Dr. Andreas Toft Sørensenfor advice regarding electrophysiological recordings, Camilla
Oki, Tatarishvili, Wood et al. 1131
www.StemCells.com

Ekenstierna for technical assistance, and the Institute of HumanGenetics (LIFE & BRAIN Center) for single nucleotide poly-morphism analysis. This work was supported by the SwedishResearch Council, the Swedish Government funding for theStrategic Research Areas of Stem Cells and Regenerative Medi-cine (StemTherapy), AFA insurance, EU 7th Framework Pro-grams ‘‘European Stroke Network’’ (Grant number 201024),‘‘NeuroStemCell’’ (Grant number 22943), TargetBraIn (Grantnumber 279017), BMBF Grants 01GN0813 and 01GN1009B,
DFG Grant SFB-TR3 D2, and the Hertie Foundation. D.T.P. wassupported by grant from Instituto de Salud Carlos III (Spain).
DISCLOSURE OF POTENTIAL
CONFLICTS OF INTEREST
The authors indicate no potential conflicts of interest.
REFERENCES
1 Lindvall O, Kokaia Z. Stem cell research in stroke: How far from theclinic? Stroke 2011;42:2369–2375.
2 Daadi MM, Maag AL, Steinberg GK. Adherent self-renewable humanembryonic stem cell-derived neural stem cell line: Functional engraft-ment in experimental stroke model. PLoS One 2008;3:e1644.
3 Hicks AU, Lappalainen RS, Narkilahti S et al. Transplantation ofhuman embryonic stem cell-derived neural precursor cells andenriched environment after cortical stroke in rats: Cell survival andfunctional recovery. Eur J Neurosci 2009;29:562–574.
4 Ramos-Cabrer P, Justicia C, Wiedermann D et al. Stem cell mediationof functional recovery after stroke in the rat. PLoS One 2010;5:e12779.
5 Buhnemann C, Scholz A, Bernreuther C et al. Neuronal differentiationof transplanted embryonic stem cell-derived precursors in strokelesions of adult rats. Brain 2006;129:3238–3248.
6 Daadi MM, Li Z, Arac A et al. Molecular and magnetic resonanceimaging of human embryonic stem cell-derived neural stem cell graftsin ischemic rat brain. Mol Ther 2009;17:1282–1291.
7 Darsalia V, Kallur T, Kokaia Z. Survival, migration and neuronal dif-ferentiation of human fetal striatal and cortical neural stem cellsgrafted in stroke-damaged rat striatum. Eur J Neurosci 2007;26:605–614.
8 Darsalia V, Allison SJ, Cusulin C et al. Cell number and timing oftransplantation determine survival of human neural stem cell grafts instroke-damaged rat brain. J Cereb Blood Flow Metab 2011;31:235–242.
9 Kelly S, Bliss TM, Shah AK et al. Transplanted human fetal neuralstem cells survive, migrate, and differentiate in ischemic rat cerebralcortex. Proc Natl Acad Sci USA 2004;101:11839–11844.
10 Lee ST, Chu K, Jung KH et al. Anti-inflammatory mechanism ofintravascular neural stem cell transplantation in haemorrhagic stroke.Brain 2008;131:616–629.
11 Horie N, Pereira MP, Niizuma K et al. Transplanted stem cell-secretedvascular endothelial growth factor effects poststroke recovery, inflam-mation, and vascular repair. Stem Cells 2011;29:274–285.
12 Bacigaluppi M, Pluchino S, Jametti LP et al. Delayed post-ischaemicneuroprotection following systemic neural stem cell transplantationinvolves multiple mechanisms. Brain 2009;132:2239–2251.
13 Lee HJ, Kim KS, Park IH et al. Human neural stem cells over-expressing VEGF provide neuroprotection, angiogenesis and func-tional recovery in mouse stroke model. PLoS One 2007;2:e156.
14 Chen J, Zhang ZG, Li Y et al. Intravenous administration of humanbone marrow stromal cells induces angiogenesis in the ischemicboundary zone after stroke in rats. Circ Res 2003;92:692–699.
15 Andres RH, Horie N, Slikker W et al. Human neural stem cellsenhance structural plasticity and axonal transport in the ischaemicbrain. Brain 2011;134:1777–1789.
16 Daadi MM, Davis AS, Arac A et al. Human neural stem cell graftsmodify microglial response and enhance axonal sprouting in neonatalhypoxic-ischemic brain injury. Stroke 2010;41:516–523.
17 Liu Z, Li Y, Zhang X et al. Contralesional axonal remodeling of thecorticospinal system in adult rats after stroke and bone marrow stro-mal cell treatment. Stroke 2008;39:2571–2577.
18 Takahashi K, Yamanaka S. Induction of pluripotent stem cells frommouse embryonic and adult fibroblast cultures by defined factors. Cell2006;126:663–676.
19 Hargus G, Cooper O, Deleidi M et al. Differentiated Parkinsonpatient-derived induced pluripotent stem cells grow in the adult rodentbrain and reduce motor asymmetry in Parkinsonian rats. Proc NatlAcad Sci USA 2010;107:15921–15926.
20 Wernig M, Zhao JP, Pruszak J et al. Neurons derived from reprog-rammed fibroblasts functionally integrate into the fetal brain andimprove symptoms of rats with Parkinson’s disease. Proc Natl AcadSci USA 2008;105:5856–5861.
21 Ebert AD, Yu J, Rose FF,Jr. et al. Induced pluripotent stem cells froma spinal muscular atrophy patient. Nature 2009;457:277–280.
22 Dimos JT, Rodolfa KT, Niakan KK et al. Induced pluripotent stemcells generated from patients with ALS can be differentiated intomotor neurons. Science 2008;321:1218–1221.
23 Kawai H, Yamashita T, Ohta Y et al. Tridermal tumorigenesis ofinduced pluripotent stem cells transplanted in ischemic brain. J CerebBlood Flow Metab 2010;30:1487–1493.
24 Chen SJ, Chang CM, Tsai SK et al. Functional improvement of focalcerebral ischemia injury by subdural transplantation of induced pluri-potent stem cells with fibrin glue. Stem Cells Dev 2010;19:1757–1767.
25 Jiang M, Lv L, Ji H et al. Induction of pluripotent stem cells trans-plantation therapy for ischemic stroke. Mol Cell Biochem 2011;354:67–75.
26 Jensen MB, Yan H, Krishnaney-Davison R et al. Survival and differ-entiation of transplanted neural stem cells derived from humaninduced pluripotent stem cells in a rat stroke model. J Stroke Cerebro-vasc Dis 2011 [Epub ahead of print].
27 Koch P, Opitz T, Steinbeck JA et al. A rosette-type, self-renewinghuman ES cell-derived neural stem cell with potential for in vitroinstruction and synaptic integration. Proc Natl Acad Sci USA 2009;106:3225–3230.
28 Kokaia Z, Zhao Q, Kokaia M et al. Regulation of brain-derived neuro-trophic factor gene expression after transient middle cerebral arteryocclusion with and without brain damage. Exp Neurol 1995;136:73–88.
29 Zhao Q, Smith M-L, Siesj€o BK. The x-conopeptide SNX-111, an N-type calcium channel blocker, dramatically ameliorates brain damagedue to transient focal ischaemia. Acta Physiol Scand 1994;150:459–461.
30 Andsberg G, Kokaia Z, Lindvall O. Upregulation of p75 neurotrophinreceptor after stroke in mice does not contribute to differential vulner-ability of striatal neurons. Exp Neurol 2001;169:351–363.
31 Hara H, Huang PL, Panahian N et al. Reduced brain edema and in-farction volume in mice lacking the neuronal isoform of nitric oxidesynthase after transient MCA occlusion. J Cereb Blood Flow Metab1996;16:605–611.
32 Chen ST, Hsu CY, Hogan EL et al. A model of focal ischemic strokein the rat: Reproducible extensive cortical infarction. Stroke 1986;17:738–743.
33 Baird AL, Meldrum A, Dunnett SB. The staircase test of skilledreaching in mice. Brain Res Bull 2001;54:243–250.
34 Montoya CP, Campbell-Hope LJ, Pemberton KD et al. The ‘‘staircasetest’’: A measure of independent forelimb reaching and grasping abil-ities in rats. J Neurosci Methods 1991;36:219–228.
35 Dowd E, Monville C, Torres EM et al. The corridor task: A simpletest of lateralised response selection sensitive to unilateral dopaminedeafferentation and graft-derived dopamine replacement in the stria-tum. Brain Res Bull 2005;68:24–30.
36 Grealish S, Mattsson B, Draxler P et al. Characterisation of behaviou-ral and neurodegenerative changes induced by intranigral 6-hydroxy-dopamine lesions in a mouse model of Parkinson’s disease. Eur JNeurosci 2010;31:2266–2278.
37 West MJ. Stereological methods for estimating the total number ofneurons and synapses: issues of precision and bias. Trends Neurosci1999;22:51–61.
38 Falk A, Koch P, Kesavan J et al. Capture of neuroepithelial-like stemcells from pluripotent stem cells provides a versatile system for invitro production of human neurons. PLoS One 2012;7:e29597.
39 Andsberg G, Kokaia Z, Bjorklund A et al. Amelioration of ischaemia-induced neuronal death in the rat striatum by NGF-secreting neuralstem cells. Eur J Neurosci 1998;10:2026–2036.
40 Hermann DM, Zechariah A. Implications of vascular endothelialgrowth factor for postischemic neurovascular remodeling. J CerebBlood Flow Metab 2009;29:1620–1643.
41 Licht T, Goshen I, Avital A et al. Reversible modulations ofneuronal plasticity by VEGF. Proc Natl Acad Sci USA 2011;108:5081–5086.
42 Ouimet CC, Miller PE, Hemmings HC,Jr. et al. DARPP-32, a dopa-mine- and adenosine 30:50-monophosphate-regulated phosphoprotein
1132 Human iPSCs in Stroke-Damaged Brain

enriched in dopamine-innervated brain regions. III. Immunocytochemi-cal localization. J Neurosci 1984;4:111–124.
43 Steinbeck JA, Koch P, Derouiche A et al. Human embryonic stemcell-derived neurons establish region-specific, long-range projectionsin the adult brain. Cell Mol Life Sci 2012;69:461–470.
44 Wictorin K, Brundin P, Gustavii B et al. Reformation of long axonpathways in adult rat central nervous system by human forebrain neu-roblasts. Nature 1990;347:556–558.
45 Aubry L, Bugi A, Lefort N et al. Striatal progenitors derivedfrom human ES cells mature into DARPP32 neurons in vitro and in
quinolinic acid-lesioned rats. Proc Natl Acad Sci USA 2008;105:16707–16712.
46 Brederlau A, Correia AS, Anisimov SV et al. Transplantation ofhuman embryonic stem cell-derived cells to a rat model of Parkin-son’s disease: Effect of in vitro differentiation on graft survival andteratoma formation. Stem Cells 2006;24:1433–1440.
47 Tabar V, Panagiotakos G, Greenberg ED et al. Migration and differen-tiation of neural precursors derived from human embryonic stem cellsin the rat brain. Nat Biotechnol 2005;23:601–606.
See www.StemCells.com for supporting information available online.
Oki, Tatarishvili, Wood et al. 1133
Copyright © 2022 FDOKUMEN




















