
Human Immunodeficiency Virus Type 1 Escape from Cyclotriazadisulfonamide-Induced CD4-Targeted Entry...
7
JOURNAL OF VIROLOGY, Sept. 2009, p. 9577–9583 Vol. 83, No. 18 0022-538X/09/$08.000 doi:10.1128/JVI.00648-09 Copyright © 2009, American Society for Microbiology. All Rights Reserved. NOTES Human Immunodeficiency Virus Type 1 Escape from Cyclotriazadisulfonamide-Induced CD4-Targeted Entry Inhibition Is Associated with Increased Neutralizing Antibody Susceptibility Kurt Vermeire, 1 * Kristel Van Laethem, 1 Wouter Janssens, 2 Thomas W. Bell, 3 and Dominique Schols 1 Rega Institute for Medical Research, Katholieke Universiteit Leuven, Leuven, Belgium 1 ; Institute of Tropical Medicine, Antwerp, Belgium 2 ; and Department of Chemistry, University of Nevada, Reno, Nevada 3 Received 30 March 2009/Accepted 21 June 2009 Continuous specific downmodulation of CD4 receptor expression in T lymphocytes by the small molecule cyclotriazadisulfonamide (CADA) selected for the CADA-resistant human immunodeficiency virus type 1 (HIV-1) NL4.3 virus containing unique mutations in the C4 and V5 regions of gp120, likely stabilizing the CD4-binding conformation. The amino acid changes in Env were associated with decreased susceptibility to anti-CD4 monoclonal antibody treatment of the cells and with higher susceptibility of the virus to soluble CD4. In addition, the acquired ability of a CADA-resistant virus to infect cells with low CD4 expression was associated with an increased susceptibility of the virus to neutralizing antibodies from sera of several HIV- 1-infected patients. The introduction of highly active antiretroviral therapy has revolutionized the management of human immunodeficiency virus (HIV) infections. Recently, the CCR5 receptor antago- nist maraviroc was approved by the FDA for antiretroviral therapy of HIV and thus became the first approved drug that inhibits HIV by targeting a cellular receptor. In addition, clin- ical studies with the anti-CD4 monoclonal antibody (MAb) ibalizumab (formerly TNX-355) demonstrated antiviral activ- ity in vivo and proved the feasibility of targeting the CD4 receptor without CD4 T-cell depletion or serious adverse effects (20). Cyclotriazadisulfonamide (CADA) compounds represent a new class of small-molecule HIV entry inhibitors (40). We previously demonstrated that the lead compound CADA specif- ically decreases surface CD4 in several types of human CD4 cells (i.e., T lymphocytes, monocytes, and dendritic cells), pro- ducing broad-spectrum antiviral potency against infection by laboratory HIV strains and clinical isolates belonging to vari- ous clades, independent of tropism (37–39, 41). The compound does not directly interact with gp120 or cell surface CD4; it specifically decreases cellular biosynthesis of CD4 (unpub- lished data). CADA compounds are small-molecule alterna- tives to therapeutic strategies involving CD4 gene knockdown (29), CD4-specific designed ankyrin repeat proteins (34), and CD4-binding/downmodulating MAbs (20, 28). CADA was synthesized as described in detail elsewhere (5). Treatment of T cells with CADA resulted in a time-dependent reduction in cell surface CD4 expression (88% decrease, as shown by staining with anti-CD4 MAbs) (Fig. 1A). These re- sults were compared with results from treatment of the cells with RPA-T4, a MAb that binds to the extracellular NH 2 - terminal D1 region of CD4. RPA-T4 MAb treatment also resulted in a time-dependent decrease in the number of CD4 molecules at the cell surface (Fig. 1A). Flow cytometric staining for CD4 (with allophycocyanin [APC]-labeled SK3 antibody) showed a 60% CD4 downmodulation induced by RPA-T4 treatment, an observation that is in line with the receptor-downmodulating activity of other CD4-binding MAbs (12, 28). A major challenge in the treatment of HIV is the emergence of escape mutants that are no longer susceptible to drugs (27). We therefore investigated the profile of HIV type 1 (HIV-1) in vitro resistance to this viral entry inhibitor. By culturing the HIV-1 CXCR4-tropic (X4) molecular clone NL4.3 (HIV- 1 NL4.3 ) in MT-4 cells in increasing concentrations of CADA, we selected for a virus that showed reduced susceptibility to the compound (Fig. 1B). The virus that was isolated after 40 passages in cell culture became insensitive to the CD4 recep- tor-downmodulating effect of CADA (CADAres40) (Table 1). Infection of MT-4 cells by the escape mutant was inhibited by different reverse transcriptase, protease, and entry inhibitors in a manner similar to that for the wild-type (WT) counterpart subcultivated in parallel in control medium (WT40), as evi- denced by the comparable 50% effective concentrations (EC 50 s) (Table 1). However, for the anti-CD4 MAb RPA-T4, a significant decrease in activity against the CADA-resistant virus compared to that against the WT virus was noted (sixfold * Corresponding author. Mailing address: Rega Institute for Medi- cal Research, Katholieke Universiteit Leuven, Minderbroedersstraat 10, B-3000 Leuven, Belgium. Phone: 32-16-337341. Fax: 32-16-337340. E-mail: [email protected]. Published ahead of print on 1 July 2009. 9577
Transcript of Human Immunodeficiency Virus Type 1 Escape from Cyclotriazadisulfonamide-Induced CD4-Targeted Entry...

JOURNAL OF VIROLOGY, Sept. 2009, p. 9577–9583 Vol. 83, No. 180022-538X/09/$08.00�0 doi:10.1128/JVI.00648-09Copyright © 2009, American Society for Microbiology. All Rights Reserved.
NOTES
Human Immunodeficiency Virus Type 1 Escape fromCyclotriazadisulfonamide-Induced CD4-Targeted
Entry Inhibition Is Associated with IncreasedNeutralizing Antibody Susceptibility�
Kurt Vermeire,1* Kristel Van Laethem,1 Wouter Janssens,2 Thomas W. Bell,3 and Dominique Schols1
Rega Institute for Medical Research, Katholieke Universiteit Leuven, Leuven, Belgium1; Institute of Tropical Medicine, Antwerp,Belgium2; and Department of Chemistry, University of Nevada, Reno, Nevada3
Received 30 March 2009/Accepted 21 June 2009
Continuous specific downmodulation of CD4 receptor expression in T lymphocytes by the small moleculecyclotriazadisulfonamide (CADA) selected for the CADA-resistant human immunodeficiency virus type 1(HIV-1) NL4.3 virus containing unique mutations in the C4 and V5 regions of gp120, likely stabilizing theCD4-binding conformation. The amino acid changes in Env were associated with decreased susceptibility toanti-CD4 monoclonal antibody treatment of the cells and with higher susceptibility of the virus to soluble CD4.In addition, the acquired ability of a CADA-resistant virus to infect cells with low CD4 expression wasassociated with an increased susceptibility of the virus to neutralizing antibodies from sera of several HIV-1-infected patients.
The introduction of highly active antiretroviral therapy hasrevolutionized the management of human immunodeficiencyvirus (HIV) infections. Recently, the CCR5 receptor antago-nist maraviroc was approved by the FDA for antiretroviraltherapy of HIV and thus became the first approved drug thatinhibits HIV by targeting a cellular receptor. In addition, clin-ical studies with the anti-CD4 monoclonal antibody (MAb)ibalizumab (formerly TNX-355) demonstrated antiviral activ-ity in vivo and proved the feasibility of targeting the CD4receptor without CD4� T-cell depletion or serious adverseeffects (20).
Cyclotriazadisulfonamide (CADA) compounds represent anew class of small-molecule HIV entry inhibitors (40). Wepreviously demonstrated that the lead compound CADA specif-ically decreases surface CD4 in several types of human CD4�
cells (i.e., T lymphocytes, monocytes, and dendritic cells), pro-ducing broad-spectrum antiviral potency against infection bylaboratory HIV strains and clinical isolates belonging to vari-ous clades, independent of tropism (37–39, 41). The compounddoes not directly interact with gp120 or cell surface CD4; itspecifically decreases cellular biosynthesis of CD4 (unpub-lished data). CADA compounds are small-molecule alterna-tives to therapeutic strategies involving CD4 gene knockdown(29), CD4-specific designed ankyrin repeat proteins (34), andCD4-binding/downmodulating MAbs (20, 28).
CADA was synthesized as described in detail elsewhere (5).
Treatment of T cells with CADA resulted in a time-dependentreduction in cell surface CD4 expression (88% decrease, asshown by staining with anti-CD4 MAbs) (Fig. 1A). These re-sults were compared with results from treatment of the cellswith RPA-T4, a MAb that binds to the extracellular NH2-terminal D1 region of CD4. RPA-T4 MAb treatment alsoresulted in a time-dependent decrease in the number of CD4molecules at the cell surface (Fig. 1A). Flow cytometricstaining for CD4 (with allophycocyanin [APC]-labeled SK3antibody) showed a 60% CD4 downmodulation induced byRPA-T4 treatment, an observation that is in line with thereceptor-downmodulating activity of other CD4-binding MAbs(12, 28).
A major challenge in the treatment of HIV is the emergenceof escape mutants that are no longer susceptible to drugs (27).We therefore investigated the profile of HIV type 1 (HIV-1) invitro resistance to this viral entry inhibitor. By culturing theHIV-1 CXCR4-tropic (X4) molecular clone NL4.3 (HIV-1NL4.3) in MT-4 cells in increasing concentrations of CADA,we selected for a virus that showed reduced susceptibility tothe compound (Fig. 1B). The virus that was isolated after 40passages in cell culture became insensitive to the CD4 recep-tor-downmodulating effect of CADA (CADAres40) (Table 1).Infection of MT-4 cells by the escape mutant was inhibited bydifferent reverse transcriptase, protease, and entry inhibitors ina manner similar to that for the wild-type (WT) counterpartsubcultivated in parallel in control medium (WT40), as evi-denced by the comparable 50% effective concentrations(EC50s) (Table 1). However, for the anti-CD4 MAb RPA-T4,a significant decrease in activity against the CADA-resistantvirus compared to that against the WT virus was noted (sixfold
* Corresponding author. Mailing address: Rega Institute for Medi-cal Research, Katholieke Universiteit Leuven, Minderbroedersstraat10, B-3000 Leuven, Belgium. Phone: 32-16-337341. Fax: 32-16-337340.E-mail: [email protected].
� Published ahead of print on 1 July 2009.
9577

FIG. 1. (A) Time-dependent CD4 receptor downmodulation by CADA and anti-CD4 MAb RPA-T4. MT-4 cells were treated with CADA (10�g/ml) or the unlabeled anti-CD4 MAb RPA-T4 (10 �g/ml) for 30 min with cooling on ice (“30 min; 0°C”) or at 37°C for 4 h (“4 h; 37°C”), 24 h(“1 d; 37°C”), or 72 h (“3 d; 37°C”). At the different time points, cells were washed and stained for CD4 expression. Cell surface CD4 staining wasperformed by simultaneous administration of two anti-CD4 MAbs recognizing distinct epitopes of the receptor, phycoerythrin (PE)-labeledanti-CD4 MAb clone RPA-T4 (eBioscience, San Diego, CA) and APC-labeled anti-CD4 MAb clone SK3 (BD Biosciences, San Jose, CA). By this
9578 NOTES J. VIROL.

increase in EC50; P � 0.015 by Student’s t test). The partialcross-resistance and reduced susceptibility of the CADA-resis-tant virus to this MAb (Fig. 2B) are consistent with the mod-erate CD4-downmodulating activity of RPA-T4 and the abilityof the CADA-resistant virus to utilize lower levels of CD4.
Analysis of the genome of the CADA-resistant virus re-vealed only five amino acid mutations in the envelope glyco-protein gp120: Y7H, V68A, Q308H, S438R, and S463P (Fig.1C, CA40). After 20 passages of CADA selection, we observedtwo substitutions in the viral gp120: V68A and S438R (Fig. 1C,
CA20). However, these two mutations were responsible foronly a fourfold decrease in sensitivity to CADA. After 20additional passages, three extra mutations occurred: Y7H,Q308H, and S463P. The mutations Y7H and V68A lie withinthe N-terminal region of gp120, which is not involved in CD4binding (23). Substitution Q308H in the hypervariable loop V3of the CADA-resistant virus seems to occur frequently in otherin vitro-generated resistant viruses: we noted this mutation inNL4.3 virus resistant to the CXCR4 antagonist AMD3100 (7),SDF-1 (33), and the broadly neutralizing MAb 2G12 (19).Substitution S438R (C4 region of gp120) has been observed insome viruses that are resistant to BMS-378806 (correspondingmutation, S440R) (13, 25), a small-molecule compound thatinhibits CD4 attachment by binding to gp120 and altering itsconformation (16). Residue 438 lies outside both the CD4 andthe BMS-378806 binding site (6), so the S438R mutation mayhelp to stabilize the CD4-binding conformation of gp120. Thissubstitution is apparently responsible for the modest (fourfold)resistance of the 20-passage mutant, which also contains theirrelevant V68A mutation (Fig. 1C, CA20).
Substitutions Q308H and S463P apparently confer most ofthe (70-fold) CADA resistance of the 40-passage mutant (Ta-bles 1 and 2). The serine residue at position 463 (correspond-ing to 465 in the crystal structure of CD4-liganded gp120) (23)lies in the V5 loop linking two CD4-binding sequences (453 to457 and 467 to 472). Proline is unique in its ability to restrictprotein conformations, so the S463P mutation may furtherenhance CD4 binding by favoring the liganded conformationof gp120. This conformational mechanism explains the ob-served cross-resistance toward BMS-378806, which does notbind to CD4-liganded gp120 (16). The S463P mutation alsoaffects an N-glycosylation site sequon, resulting in the loss of apotential N-glycosylation site (Fig. 1C). Deletion of N glyco-sylations in gp120 has been reported to occur during develop-ment of HIV-1 resistance to other entry inhibitors, such asMAb 2G12 (19), the mannose-specific plant lectins (3), andseveral other carbohydrate-binding agents (1, 2, 4). In contrast,deletion of N glycosylation in gp120 sometimes results in anenhanced neutralizing capacity of antiviral agents, e.g., carbo-hydrate-binding agents, against 2G12-resistant virus (19). In
means, we could discriminate between competition for receptor binding (through steric hindrance) and downregulation of the complete receptor.The CADA compound does not bind cell surface CD4 but downmodulates the expression of the CD4 receptor, as shown by the similar effects ofboth anti-CD4 MAbs. The CD4-downmodulating effect of CADA was confirmed with a panel of six other anti-CD4 MAbs, all showing the removalof the CD4 receptors from the cell surface after 24 h of treatment (not shown). RPA-T4 antibody treatment of the cells resulted in a sterichindrance for PE-labeled RPA-T4 staining (detectable after 30 min treatment at 0°C) but also in more than 50% CD4 downregulation after 3 daysof treatment (determined with APC-labeled SK3). CD4 MAb binding is given as the percentage of stained nontreated control cells (based on themean fluorescence intensity values). Comparable results were also obtained with peripheral blood mononuclear cells (data not shown). The barsrepresent the means � standard errors of the means from three independent experiments. (B) Selection of CADA-resistant virus in MT-4 cells.Cells and HIV-1 X4NL4.3 virus were subcultivated in the presence of CADA every 4 days and microscopically examined for signs of virus replication.Following virus breakthrough, the supernatant was collected and stored and used to infect fresh cell cultures with a higher concentration of CADA.The procedure was repeated up to a concentration of 50 �g/ml of CADA (i.e., 40 passages; CADAres40; red line). In parallel, cells were infectedwith WT HIV-1 X4NL4.3 virus (WT1) and subcultivated in control medium without compound for 40 passages (WT40; blue dashed line). (C) gp120amino acid sequences of WT and CADA-resistant viruses. HIV-1 X4NL4.3 virus was selected as described for panel B. At passages 1, 20, and 40,the collected supernatant was subjected to genome sequencing. The figure shows the amino acid sequences of the original virus (WT01; black),the WT control virus after 40 passages in medium without compound (WT40; blue), and the CADA-resistant virus after 20 and 40 passages inmedium with CADA (CA20 and CA40, respectively; red). The variable regions are indicated as V1 to V5. The residues of gp120 involved in CD4binding (according to reference 23) are starred. The numbering of the amino acids is in comparison to a reference strain (GenBank accession no.AF324493). Five mutations in the gp120 genome of CADA-resistant virus were observed (Y7H, V68A, Q308H, S438R, and S463P), with the lossof the potential N-glycosylation site at position 461 in the V5 region.
TABLE 1. Profiles of WT40 and CADAres40 HIV-1NL4.3 virussusceptibility to antiviral agentsa
AgentEC50
bFold
resistancecFold
sensitivitydWT40 CADAres40
CADA (�g/ml) 1.2 � 0.2 �50 �42Zidovudine (nM) 11.0 � 1.7 6.4 � 1.4 2Tenofovir (�M) 4.1 � 0.9 3.2 � 0.8 1Delavirdine (ng/ml) 18.5 � 1.7 11.4 � 2.9 2Efavirenz (ng/ml) 1.1 � 0.2 1.0 � 0.3 1Saquinavir (ng/ml) 13.2 � 3.9 8.5 � 3.1 2Indinavir (ng/ml) 23.8 � 5.9 12.5 � 2.5 2Lectin GNAe
(ng/ml)140.8 � 57.7 100.8 � 51.4 1
Lectin HHAf
(ng/ml)148.4 � 60.8 106.0 � 43.7 1
Enfuvirtide (ng/ml) 56.0 � 10.8 62.3 � 10.2 1AMD3100 (ng/ml) 7.4 � 2.1 9.4 � 2.3 1DS-5000 (�g/ml) 0.81 � 0.18 0.70 � 0.23 1Anti-CD4 MAb
RPA-T4 (�g/ml)0.32 � 0.08 1.97 � 0.51 6
a WT NL4.3 virus was selected after 40 passages in control medium (WT40),and CADA-resistant virus was selected after 40 passages in CADA compound(CADAres40), as described in the legend for Fig. 1B. The antiviral activities ofthe compounds were tested using MT-4 cells infected with equal inputs of theseWT40 and CADAres40 viruses (50% tissue culture infective doses of 50). After5 days of infection, the supernatant was collected and tested for HIV-1 p24antigen content by enzyme-linked immunosorbent assay.
b Values are means � standard errors of the means from three to five differentexperiments. The units of measure are given in the first column.
c Resistance was calculated as follows: (EC50 CADAres40)/(EC50 WT40).Values shown in boldface type indicate significant differences as given in the text.
d Sensitivity was calculated as follows: (EC50 WT40)/(EC50 CADAres40).e GNA, Galanthus nivalis agglutinin.f HHA, Hippeastrum sp. hybrid agglutinin.
VOL. 83, 2009 NOTES 9579
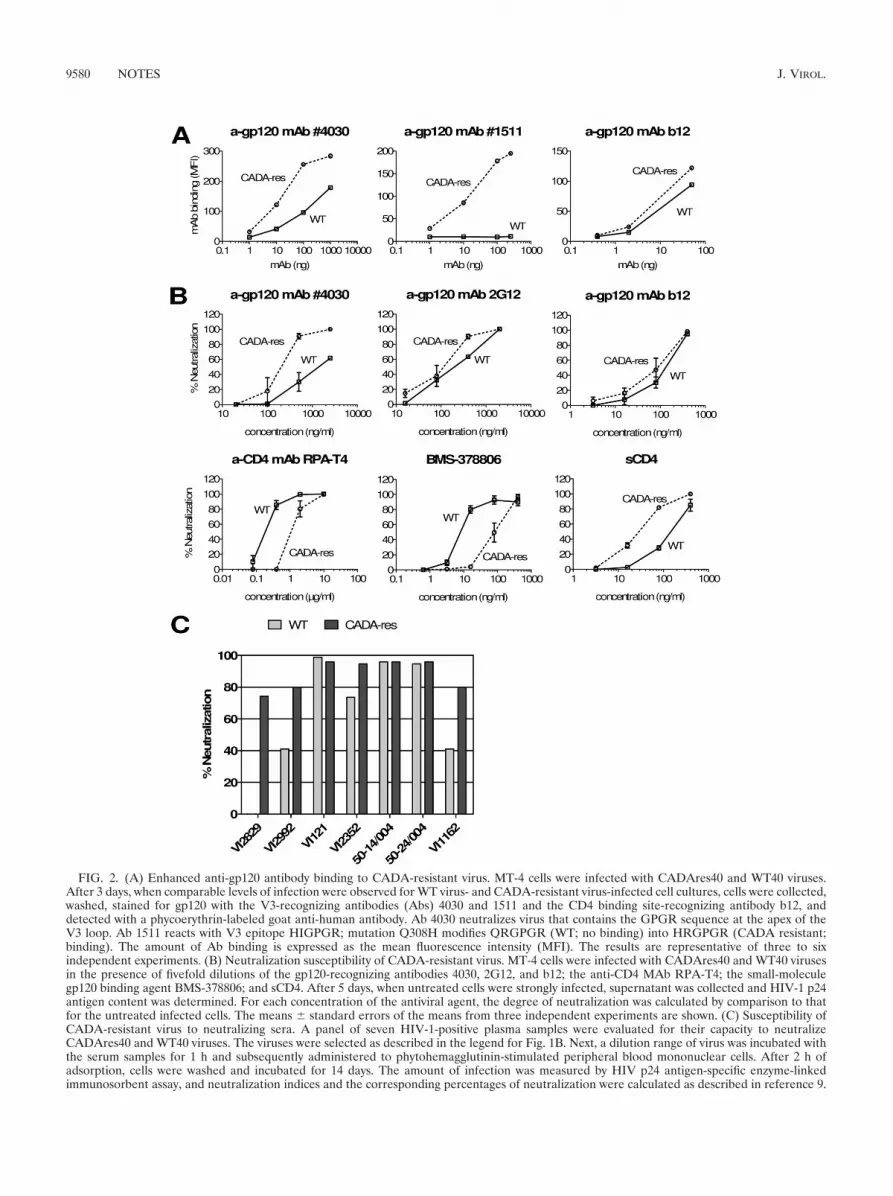
FIG. 2. (A) Enhanced anti-gp120 antibody binding to CADA-resistant virus. MT-4 cells were infected with CADAres40 and WT40 viruses.After 3 days, when comparable levels of infection were observed for WT virus- and CADA-resistant virus-infected cell cultures, cells were collected,washed, stained for gp120 with the V3-recognizing antibodies (Abs) 4030 and 1511 and the CD4 binding site-recognizing antibody b12, anddetected with a phycoerythrin-labeled goat anti-human antibody. Ab 4030 neutralizes virus that contains the GPGR sequence at the apex of theV3 loop. Ab 1511 reacts with V3 epitope HIGPGR; mutation Q308H modifies QRGPGR (WT; no binding) into HRGPGR (CADA resistant;binding). The amount of Ab binding is expressed as the mean fluorescence intensity (MFI). The results are representative of three to sixindependent experiments. (B) Neutralization susceptibility of CADA-resistant virus. MT-4 cells were infected with CADAres40 and WT40 virusesin the presence of fivefold dilutions of the gp120-recognizing antibodies 4030, 2G12, and b12; the anti-CD4 MAb RPA-T4; the small-moleculegp120 binding agent BMS-378806; and sCD4. After 5 days, when untreated cells were strongly infected, supernatant was collected and HIV-1 p24antigen content was determined. For each concentration of the antiviral agent, the degree of neutralization was calculated by comparison to thatfor the untreated infected cells. The means � standard errors of the means from three independent experiments are shown. (C) Susceptibility ofCADA-resistant virus to neutralizing sera. A panel of seven HIV-1-positive plasma samples were evaluated for their capacity to neutralizeCADAres40 and WT40 viruses. The viruses were selected as described in the legend for Fig. 1B. Next, a dilution range of virus was incubated withthe serum samples for 1 h and subsequently administered to phytohemagglutinin-stimulated peripheral blood mononuclear cells. After 2 h ofadsorption, cells were washed and incubated for 14 days. The amount of infection was measured by HIV p24 antigen-specific enzyme-linkedimmunosorbent assay, and neutralization indices and the corresponding percentages of neutralization were calculated as described in reference 9.
9580 NOTES J. VIROL.

this regard, we observed a slightly higher neutralizing potencyof MAb 2G12 against the CADA-resistant virus than againstthe WT virus (Fig. 2B). However, during in vitro resistanceselection N-glycosylation deletions in the viral envelope caneasily be tolerated and occur spontaneously.
Next, we wanted to verify if the viral envelope of the escapevirus was solely responsible for the decreased susceptibility toCADA. Based on chimeric virus technology (11), we per-formed the recombination of the env gene gp160 sequences ofthe CADAres40 and WT40 viruses into a proviral WT HIV-1clone (pNL4.3) from which the corresponding env gene wasdeleted (11). Both recombinant viruses proved to exert similarinfection efficiencies (not shown). When the pNL4.3 WT andCADA-resistant viruses were tested for susceptibility to vari-ous anti-HIV agents, we noted for the recombinant viruses asusceptibility profile that was comparable to that of the corre-sponding native strains. Replication of the pNL4.3 CADA-resistant mutant could not be prevented by CADA treatment(up to 50 �g/ml), indicating that the viral envelope was solelyresponsible for CADA resistance (Table 2). Again, the CADA-resistant virus proved to be significantly less susceptible thanthe WT virus to anti-CD4 MAb treatment of the cells (EC50swere 2.0 and 0.4 �g/ml, respectively; P � 0.021, Student’s ttest) (Table 2). In contrast, there was a tendency for solubleCD4 (sCD4) (35) to exert a higher neutralizing activity againstthe CADA-resistant virus (Fig. 2B): we measured a 4.3-foldincrease in potency compared to that for the WT virus (P �0.056, Student’s t test) (Table 2). The anti-gp120 MAb b12showed no difference in activity toward the CADA-resistant
virus versus the WT virus (Fig. 2A and B; Table 2). This isconsistent with reports that so-called CD4 binding site anti-bodies, such as b12, recognize conformations of gp120 differentfrom that recognized by CD4 and CD4-induced antibodies,which are induced by the CD4-liganded conformation of gp120(42).
Finally, we further explored the neutralizing susceptibility ofthe CADA escape mutant CADAres40 and found that severalanti-gp120 MAbs bind to the viral envelope of CADA-resistantvirus more efficiently than to that of WT virus (Fig. 2A). Inaddition, by means of neutralizing antibodies isolated fromHIV-1-infected individuals, as described elsewhere (9), weobserved that the CADA-resistant virus was markedly moreprone than the WT to the neutralizing capacities of thedifferent sera (Fig. 2C). Whereas the WT virus could beneutralized by only half of the serum samples, the CADA-resistant virus proved to be susceptible to virtually all of thetested sera (Fig. 2C).
Conformational flexibility and N glycosylation of gp120 arebelieved to be two important strategies used by HIV to evadethe immune system (8, 24). The observed mutations in CADA-resistant gp120 apparently stabilize the conformation of gp120and reduce glycosylation, both of which can enhance the abilityto elicit neutralizing antibodies (8, 24). Increased susceptibilityto neutralizing antibodies has also been reported for CD4-independent viruses (17), which could explain the rarity ofstrict CD4-independent HIV variants in vivo (10, 22). How-ever, these viruses still show higher infectivity and replicationability when CD4 is expressed on the cell surface. Here, CADAresistance is not associated with escape of CD4 receptor usage,as the resistant virus was not able to infect CD4-negativeU87.CXCR4� cells or other CD4-negative cells, such as theSupT1 clone B2 (not shown). A similar loyalty of HIV-1 to itsreceptor has also been reported for R5 viruses resistant toCCR5 antagonists and X4 escape mutants to CXCR4 inhibi-tors (26, 31, 33, 36).
In the HIV entry process, multiple CD4 and coreceptorbinding events are needed to efficiently activate a viral spike,consisting of three gp120 molecules noncovalently attached tothree subunits of gp41, thus implying that CD4 receptor andcoreceptor densities have a crucial role in the efficiency of HIVinfection. Accordingly, several studies have already demon-strated that (co)receptor density levels in individuals are ratelimiting for infection (21, 30) and that downregulation ofCCR5 expression, e.g., by the G1 cytostatic agent rapamycin,can be an interesting strategy for both the prevention and thetreatment of HIV infection (14, 15). In addition, not only couldinhibiting the expression of a cellular receptor have a directantiviral effect by preventing viral entry, but it is possible thatthis could force the virus to become less dependent on use ofits receptor by changing its viral envelope. Hypothetically,these escape mutants could then be eliminated more efficientlyby neutralizing antibodies, so that in this case development ofresistance to the drug might even be considered favorable,providing a new way of trapping the virus. Recently, and in linewith our results, it has been reported that sequence changes inthe envelope that confer resistance to other HIV entry inhib-itors can increase the accessibility of some neutralizing anti-body epitopes and ultimately make these viruses more prone toantibody neutralization (18, 31, 32).
TABLE 2. Profiles of recombinant pNL4.3 env WT40 andrecombinant env CADAres40 virus susceptibility to
antiviral agentsa
AgentEC50
bFold
resistancecFold
sensitivitydWT40 CADAres40
CADA (�g/ml) 0.7 � 0.1 �50 �71Tenofovir (�M) 1.9 � 0.9 1.3 � 0.4 1Lopinavir (ng/ml) 16.3 � 2.8 8.0 � 0.8 2Raltegravir (nM) 3.9 � 0.4 2.0 � 1.1 2Enfuvirtide (ng/ml) 59.0 � 18.8 36.7 � 11.3 2AMD3100 (ng/ml) 2.5 � 0.2 3.0 � 0.2 1Maraviroc (ng/ml) �1,000 �1,000Anti-CD4 MAb
RPA-T4 (�g/ml)0.36 � 0.08 1.98 � 0.54 6
Anti-gp120 MAbb12 (ng/ml)
109.1 � 15.6 77.0 � 24.8 1
sCD4 (ng/ml) 120.5 � 37.0 28.1 � 5.2 4CD4M33 (ng/ml) 45.3 � 18.6 19.6 � 11.3 2
a Recombination of gp160 sequences of CADAres40 and WT40 viruses into aproviral WT HIV-1 clone (pNL4.3) from which the corresponding env gene wasdeleted was performed. The amino acid sequences of the recombinants weredetermined (in comparison to that of pNL4.3 �GenBank accession no.AF324493�) and confirmed the presence of the CADA resistance mutations(as depicted in Fig. 1C).
b The antiviral activities of the compounds were tested using MT-4 cells in-fected with equal inputs of recombinant pNL4.3 env WT and CADA-resistantNL4.3 viruses (50% tissue culture infective dose of 50). After 5 days of infection,the supernatant was collected and tested for HIV-1 p24 antigen content byenzyme-linked immunosorbent assay. Values are means � standard errors of themeans from three to seven different experiments. The units of measure are givenin the first column.
c Resistance was calculated as follows: (EC50 CADAres40)/(EC50 WT40).Values shown in boldface type indicate significant differences.
d Sensitivity was calculated as follows: (EC50 WT40)/(EC50 CADAres40).
VOL. 83, 2009 NOTES 9581

In conclusion, CADA-resistant virus selected for mutationsin the viral envelope that support the infection of cells with lowCD4 expression. The amino acid substitutions were associatedwith decreased susceptibility to anti-CD4 MAb treatment andwith higher sensitivity to neutralizing antibodies from HIV-infected patients. These in vitro observations may have inter-esting implications for novel antiretroviral therapies and canopen new roads to the successful design of an HIV vaccine.
HIV-1 V3 MAbs 447-52D (4030) and 268-D IV (1511) were ob-tained through the AIDS Research and Reference Program, Divisionof AIDS, NIAID, NIH, from Susan Zolla-Pazner. The human immu-noglobulin G1 b12 antibody and the CD4 mimic CD4M33 were kindgifts from Dennis R. Burton and Loïc Martin, respectively. We thankSandra Claes, Eric Fonteyn, Becky Provinciael, and Yoeri Schrootenfor excellent technical assistance. We are grateful to Kris Covens forhelping with the pNL4.3 recombinant viruses and to Betty Willems forassisting with the neutralization experiments.
Kurt Vermeire is a postdoctoral researcher at the Fonds voorWetenschappelijk Onderzoek (F.W.O.)—Vlaanderen (Belgium).
This work was supported by the Centers of Excellence from theKatholieke Universiteit Leuven (EF/05/15), EMPRO 503558, Fonda-tion Dormeur, and F.W.O. (G-485-08).
REFERENCES
1. Balzarini, J., K. Van Laethem, D. Daelemans, S. Hatse, A. Bugatti, M.Rusnati, Y. Igarashi, T. Oki, and D. Schols. 2007. Pradimicin A, a carbohy-drate-binding nonpeptidic lead compound for treatment of infections withviruses with highly glycosylated envelopes, such as human immunodeficiencyvirus. J. Virol. 81:362–373.
2. Balzarini, J., K. Van Laethem, S. Hatse, M. Froeyen, W. Peumans, E. VanDamme, and D. Schols. 2005. Carbohydrate-binding agents cause deletionsof highly conserved glycosylation sites in HIV GP120: a new therapeuticconcept to hit the achilles heel of HIV. J. Biol. Chem. 280:41005–41014.
3. Balzarini, J., K. Van Laethem, S. Hatse, K. Vermeire, E. De Clercq, W.Peumans, E. Van Damme, A. M. Vandamme, A. Bolmstedt, and D. Schols.2004. Profile of resistance of human immunodeficiency virus to mannose-specific plant lectins. J. Virol. 78:10617–10627.
4. Balzarini, J., K. Van Laethem, W. J. Peumans, E. J. Van Damme, A. Bolm-stedt, F. Gago, and D. Schols. 2006. Mutational pathways, resistance profile,and side effects of cyanovirin relative to human immunodeficiency virus type1 strains with N-glycan deletions in their gp120 envelopes. J. Virol. 80:8411–8421.
5. Bell, T. W., S. Anugu, P. Bailey, V. J. Catalano, K. Dey, M. G. Drew, N. H.Duffy, Q. Jin, M. F. Samala, A. Sodoma, W. H. Welch, D. Schols, and K.Vermeire. 2006. Synthesis and structure-activity relationship studies of CD4down-modulating cyclotriazadisulfonamide (CADA) analogues. J. Med.Chem. 49:1291–1312.
6. Chen, B., E. M. Vogan, H. Gong, J. J. Skehel, D. C. Wiley, and S. C.Harrison. 2005. Structure of an unliganded simian immunodeficiency virusgp120 core. Nature 433:834–841.
7. De Vreese, K., V. Kofler-Mongold, C. Leutgeb, V. Weber, K. Vermeire, S.Schacht, J. Anne, E. De Clercq, R. Datema, and G. Werner. 1996. Themolecular target of bicyclams, potent inhibitors of human immunodeficiencyvirus replication. J. Virol. 70:689–696.
8. Dey, B., M. Pancera, K. Svehla, Y. Shu, S. H. Xiang, J. Vainshtein, Y. Li, J.Sodroski, P. D. Kwong, J. R. Mascola, and R. Wyatt. 2007. Characterizationof human immunodeficiency virus type 1 monomeric and trimeric gp120glycoproteins stabilized in the CD4-bound state: antigenicity, biophysics, andimmunogenicity. J. Virol. 81:5579–5593.
9. Donners, H., D. Davis, B. Willems, and G. van der Groen. 2003. Inter-subtype cross-neutralizing antibodies recognize epitopes on cell-associatedHIV-1 virions. J. Med. Virol. 69:173–181.
10. Edwards, T. G., T. L. Hoffman, F. Baribaud, S. Wyss, C. C. LaBranche, J.Romano, J. Adkinson, M. Sharron, J. A. Hoxie, and R. W. Doms. 2001.Relationships between CD4 independence, neutralization sensitivity, andexposure of a CD4-induced epitope in a human immunodeficiency virus type1 envelope protein. J. Virol. 75:5230–5239.
11. Fikkert, V., P. Cherepanov, K. Van Laethem, A. Hantson, B. Van Remoortel,C. Pannecouque, E. De Clercq, Z. Debyser, A. M. Vandamme, and M.Witvrouw. 2002. env chimeric virus technology for evaluating human immu-nodeficiency virus susceptibility to entry inhibitors. Antimicrob. Agents Che-mother. 46:3954–3962.
12. Fishwild, D. M., D. V. Hudson, U. Deshpande, and A. H. Kung. 1999.Differential effects of administration of a human anti-CD4 monoclonal an-tibody, HM6G, in nonhuman primates. Clin. Immunol. 92:138–152.
13. Guo, Q., H. T. Ho, I. Dicker, L. Fan, N. Zhou, J. Friborg, T. Wang, B. V.
McAuliffe, H. G. Wang, R. E. Rose, H. Fang, H. T. Scarnati, D. R. Langley,N. A. Meanwell, R. Abraham, R. J. Colonno, and P. F. Lin. 2003. Biochem-ical and genetic characterizations of a novel human immunodeficiency virustype 1 inhibitor that blocks gp120-CD4 interactions. J. Virol. 77:10528–10536.
14. Heredia, A., A. Amoroso, C. Davis, N. Le, E. Reardon, J. K. Dominique, E.Klingebiel, R. C. Gallo, and R. R. Redfield. 2003. Rapamycin causes down-regulation of CCR5 and accumulation of anti-HIV beta-chemokines: anapproach to suppress R5 strains of HIV-1. Proc. Natl. Acad. Sci. USA100:10411–10416.
15. Heredia, A., O. Latinovic, R. C. Gallo, G. Melikyan, M. Reitz, N. Le, andR. R. Redfield. 2008. Reduction of CCR5 with low-dose rapamycin enhancesthe antiviral activity of vicriviroc against both sensitive and drug-resistantHIV-1. Proc. Natl. Acad. Sci. USA 105:20476–20481.
16. Ho, H. T., L. Fan, B. Nowicka-Sans, B. McAuliffe, C. B. Li, G. Yamanaka, N.Zhou, H. Fang, I. Dicker, R. Dalterio, Y. F. Gong, T. Wang, Z. Yin, Y. Ueda,J. Matiskella, J. Kadow, P. Clapham, J. Robinson, R. Colonno, and P. F. Lin.2006. Envelope conformational changes induced by human immunodefi-ciency virus type 1 attachment inhibitors prevent CD4 binding and down-stream entry events. J. Virol. 80:4017–4025.
17. Hoffman, T. L., C. C. LaBranche, W. Zhang, G. Canziani, J. Robinson, I.Chaiken, J. A. Hoxie, and R. W. Doms. 1999. Stable exposure of the core-ceptor-binding site in a CD4-independent HIV-1 envelope protein. Proc.Natl. Acad. Sci. USA 96:6359–6364.
18. Hu, Q., N. Mahmood, and R. J. Shattock. 2007. High-mannose-specificdeglycosylation of HIV-1 gp120 induced by resistance to cyanovirin-N andthe impact on antibody neutralization. Virology 368:145–154.
19. Huskens, D., K. Van Laethem, K. Vermeire, J. Balzarini, and D. Schols.2007. Resistance of HIV-1 to the broadly HIV-1-neutralizing, anti-carbohy-drate antibody 2G12. Virology 360:294–304.
20. Jacobson, J. M., D. R. Kuritzkes, E. Godofsky, E. DeJesus, J. A. Larson, S. P.Weinheimer, and S. T. Lewis. 2009. Safety, pharmacokinetics, and antiret-roviral activity of multiple doses of ibalizumab (formerly TNX-355), ananti-CD4 monoclonal antibody, in human immunodeficiency virus type 1-in-fected adults. Antimicrob. Agents Chemother. 53:450–457.
21. Kabat, D., S. L. Kozak, K. Wehrly, and B. Chesebro. 1994. Differences inCD4 dependence for infectivity of laboratory-adapted and primary patientisolates of human immunodeficiency virus type 1. J. Virol. 68:2570–2577.
22. Kolchinsky, P., E. Kiprilov, and J. Sodroski. 2001. Increased neutralizationsensitivity of CD4-independent human immunodeficiency virus variants.J. Virol. 75:2041–2050.
23. Kwong, P. D., R. Wyatt, J. Robinson, R. W. Sweet, J. Sodroski, and W. A.Hendrickson. 1998. Structure of an HIV gp120 envelope glycoprotein incomplex with the CD4 receptor and a neutralizing human antibody. Nature393:648–659.
24. Li, Y., B. Cleveland, I. Klots, B. Travis, B. A. Richardson, D. Anderson, D.Montefiori, P. Polacino, and S. L. Hu. 2008. Removal of a single N-linkedglycan in human immunodeficiency virus type 1 gp120 results in an enhancedability to induce neutralizing antibody responses. J. Virol. 82:638–651.
25. Lin, P. F., W. Blair, T. Wang, T. Spicer, Q. Guo, N. Zhou, Y. F. Gong, H. G.Wang, R. Rose, G. Yamanaka, B. Robinson, C. B. Li, R. Fridell, C. Deminie,G. Demers, Z. Yang, L. Zadjura, N. Meanwell, and R. Colonno. 2003. Asmall molecule HIV-1 inhibitor that targets the HIV-1 envelope and inhibitsCD4 receptor binding. Proc. Natl. Acad. Sci. USA 100:11013–11018.
26. MacArthur, R. D., and R. M. Novak. 2008. Maraviroc: the first of a new classof antiretroviral agents. Clin. Infect. Dis. 47:236–241.
27. Martinez-Cajas, J. L., and M. A. Wainberg. 2008. Antiretroviral therapy:optimal sequencing of therapy to avoid resistance. Drugs 68:43–72.
28. Newman, R., K. Hariharan, M. Reff, D. R. Anderson, G. Braslawsky, D.Santoro, N. Hanna, P. J. Bugelski, M. Brigham-Burke, C. Crysler, R. C.Gagnon, P. Dal Monte, M. L. Doyle, P. C. Hensley, M. P. Reddy, R. W. Sweet,and A. Truneh. 2001. Modification of the Fc region of a primatized IgGantibody to human CD4 retains its ability to modulate CD4 receptors butdoes not deplete CD4� T cells in chimpanzees. Clin. Immunol. 98:164–174.
29. Novina, C. D., M. F. Murray, D. M. Dykxhoorn, P. J. Beresford, J. Riess,S. K. Lee, R. G. Collman, J. Lieberman, P. Shankar, and P. A. Sharp. 2002.siRNA-directed inhibition of HIV-1 infection. Nat. Med. 8:681–686.
30. Platt, E. J., K. Wehrly, S. E. Kuhmann, B. Chesebro, and D. Kabat. 1998.Effects of CCR5 and CD4 cell surface concentrations on infections by mac-rophagetropic isolates of human immunodeficiency virus type 1. J. Virol.72:2855–2864.
31. Pugach, P., T. J. Ketas, E. Michael, and J. P. Moore. 2008. Neutralizingantibody and anti-retroviral drug sensitivities of HIV-1 isolates resistant tosmall molecule CCR5 inhibitors. Virology 377:401–407.
32. Reeves, J. D., F. H. Lee, J. L. Miamidian, C. B. Jabara, M. M. Juntilla, andR. W. Doms. 2005. Enfuvirtide resistance mutations: impact on human im-munodeficiency virus envelope function, entry inhibitor sensitivity, and virusneutralization. J. Virol. 79:4991–4999.
33. Schols, D., J. A. Este, C. Cabrera, and E. De Clercq. 1998. T-cell-line-tropichuman immunodeficiency virus type 1 that is made resistant to stromalcell-derived factor 1� contains mutations in the envelope gp120 but does notshow a switch in coreceptor use. J. Virol. 72:4032–4037.
9582 NOTES J. VIROL.

34. Schweizer, A., P. Rusert, L. Berlinger, C. R. Ruprecht, A. Mann, S. Corthesy,S. G. Turville, M. Aravantinou, M. Fischer, M. Robbiani, P. Amstutz, and A.Trkola. 2008. CD4-specific designed ankyrin repeat proteins are novel potentHIV entry inhibitors with unique characteristics. PLoS Pathog. 4:e1000109.
35. Smith, D. H., R. A. Byrn, S. A. Marsters, T. Gregory, J. E. Groopman, andD. J. Capon. 1987. Blocking of HIV-1 infectivity by a soluble, secreted formof the CD4 antigen. Science 238:1704–1707.
36. Trkola, A., S. E. Kuhmann, J. M. Strizki, E. Maxwell, T. Ketas, T. Morgan,P. Pugach, S. Xu, L. Wojcik, J. Tagat, A. Palani, S. Shapiro, J. W. Clader, S.McCombie, G. R. Reyes, B. M. Baroudy, and J. P. Moore. 2002. HIV-1escape from a small molecule, CCR5-specific entry inhibitor does not involveCXCR4 use. Proc. Natl. Acad. Sci. USA 99:395–400.
37. Vermeire, K., J. Brouwers, Y. Van Herrewege, R. Le Grand, G. Vanham, P.Augustijns, T. W. Bell, and D. Schols. 2008. CADA, a potential anti-HIVmicrobicide that specifically targets the cellular CD4 receptor. Curr. HIVRes. 6:246–256.
38. Vermeire, K., K. Princen, S. Hatse, E. De Clercq, K. Dey, T. W. Bell, and D.
Schols. 2004. CADA, a novel CD4-targeted HIV inhibitor, is synergistic withvarious anti-HIV drugs in vitro. AIDS 18:2115–2125.
39. Vermeire, K., and D. Schols. 2003. Specific CD4 down-modulating com-pounds with potent anti-HIV activity. J. Leukoc. Biol. 74:667–675.
40. Vermeire, K., and D. Schols. 2005. Cyclotriazadisulfonamides: promisingnew CD4-targeted anti-HIV drugs. J. Antimicrob. Chemother. 56:270–272.
41. Vermeire, K., Y. Zhang, K. Princen, S. Hatse, M. F. Samala, K. Dey, H. J.Choi, Y. Ahn, A. Sodoma, R. Snoeck, G. Andrei, E. De Clercq, T. W. Bell, andD. Schols. 2002. CADA inhibits human immunodeficiency virus and humanherpesvirus 7 replication by down-modulation of the cellular CD4 receptor.Virology 302:342–353.
42. Xiang, S. H., P. D. Kwong, R. Gupta, C. D. Rizzuto, D. J. Casper, R. Wyatt,L. Wang, W. A. Hendrickson, M. L. Doyle, and J. Sodroski. 2002. Mutagenicstabilization and/or disruption of a CD4-bound state reveals distinct confor-mations of the human immunodeficiency virus type 1 gp120 envelope glyco-protein. J. Virol. 76:9888–9899.
VOL. 83, 2009 NOTES 9583




















