
(2014): „‘Goldland Kolumbien‘ – ein Eldorado!?“. En: Amerindian Research 9/1, 31, pp. 20-30.
Upload
independentCategory
view
1download
0
HLA in Jaidukama: an Amerindian secluded Colombianpopulation with new haplotypes and Asian and Pacific-sharedalleles
J. Martinez-Laso • F. Montoya • C. Areces • J. Moscoso •
C. Silvera • D. Rey • C. Parga-Lozano • P. Gomez-Prieto •
M. Enriquez de Salamanca • A. Arnaiz-Villena
Received: 24 September 2010 / Accepted: 9 November 2010 / Published online: 26 November 2010
� Springer Science+Business Media B.V. 2010
Abstract America first inhabitants and peopling are still
debated. In order to increase knowledge about these
questions, we have aimed to detect HLA genes of an
Amerindian secluded community: Jaidukama, who lives in
North Colombia Equatorial forest. HLA genotyping and
extended haplotype calculations were carried out in 39
healthy individuals belonging to 13 families. HLA fre-
quencies were compared to other Amerindians and
worldwide populations by calculating genetic distances,
relatedness dendrograms and correspondence analyses.
Only four DRB1 alleles were found (*0404, *0407, *1402
and *1602); however a total of 17 Amerindian different
extended class I–class II HLA haplotypes were directly
counted from the family studies, nine of them were specific
of Jaidukamas. Some of the alleles or group of alleles
within an extended haplotype (i.e. DQB1–DRB1) were also
found in Asians and Pacific Islanders, further supporting
existence of Asian and Pacific gene flow with Amerindians
or a common founder effect. It is further supported that
HLA extended haplotypes vary faster than alleles in pop-
ulations. It is concluded that this unique model of Amer-
indian secluded families study suggests that rapid HLA
haplotype variation may be more important than allele
variation for survival (starting immune responses). This
work may also be useful for future transplant programs in
the area.
Keywords Jaidukama � Colombia � Amerindians � HLA �America peopling
Introduction
The first Amerindian natives are postulated to have come
from Asia through the Bering land bridge between 30,000
and 12,000 years before the present (BP). These conclu-
sions have been based on cultural, morphological and
genetic similarities between American and Asian popula-
tions. Both Siberia [1] and Mongolia [2, 3] have been put
forward as the most likely places of origin in Asia.
Greenberg first postulated the triple migration theory for
explaining the peopling of the Americas [4]: Amerindians
(most North and South American Indians; 12,000 years
BP), Na-Dene (Athabascans, Navajo, Apache; 8,000 years
BP) and Eskimo-Aleuts (6,000 years BP) [5]. Some studies
have or have not supported the hypothesis by Y Chr and
mtDNA markers [2, 3, 6–12]. However these conclusions
do not explain all linguistic and genetic findings [13–15].
Y Chr and mtDNA studies seem unable to definitively
clarify American people and Amerindian origins, mainly
The authors J. Martinez-Laso and F. Montoya contributed equally for
this work and the order of authorship is arbitrary.
J. Martinez-Laso
Unidad de Inmunoterapia Celular, Centro Nacional de
Microbiologıa, Instituto de Salud Carlos III, Madrid, Spain
F. Montoya
Section of Immunology, Corporacion para Investigaciones
Biologicas, Medellin, Colombia
C. Areces � J. Moscoso � C. Silvera � D. Rey �C. Parga-Lozano � P. Gomez-Prieto �M. Enriquez de Salamanca � A. Arnaiz-Villena
Department of Immunology, University Complutense,
The Madrid Regional Blood Center, Madrid, Spain
A. Arnaiz-Villena (&)
Departamento de Inmunologia, Facultad de Medicina,
Universidad Complutense, Pabellon 5, planta 4. Avda.
Complutense s/n, Madrid 28040, Spain
e-mail: [email protected]
URL: http://chopo.pntic.mec.es/biolmol/
123
Mol Biol Rep (2011) 38:3689–3701
DOI 10.1007/s11033-010-0483-6

because the studies have been gene—rather than popula-
tion—based. Their emphasis has been on variant genealo-
gies rather than on population frequencies studies. In this
regard HLA data may be more informative [16] because
maternal and paternal lineages and both frequencies (i.e.:
genetic distances, dendrograms and correspondence anal-
yses) and genealogies (quasi-specific HLA alleles and
haplotypes) may be studied for comparing populations.
Recently, new mtDNA analysis has suggested that all
mtDNA lineages must have been isolated in Asia before
entering the New World by at least 7,000–15,000 years.
They even suggest that this place must have been Beringia
[17]. Also, a dispersal of Amerindians coming from Asia
has been put forward through Coastal Pacific line [18]
based on all available archaeological, anthropological and
mtDNA and other genetic data. A Trans-Pacific route of
American peopling from Asia or Polynesia had been sug-
gested because HTLV-1 virus strains shared identical
sequences in Japan and in the northern coast of South
America [19] and some HLA alleles may have been
introduced by the same Trans-Pacific route [20]. Finally,
both genetic [21] and archaeological [22] evidence sug-
gests that a two-way Trans-Atlantic traffic occurred before
Columbus discovered America. Archaeologists in New
Mexico have also found tools used 20,000 years ago in
Spanish Solutrean culture [22]. Recently our own studies
by using autosomic HLA markers and combining geneal-
ogies particular markers and population gene frequencies
comparisons, show that Asian, Polinesian and Oceania
gene flow with Amerindians has been possible [15].
On the other hand, the first Colombian settlers arrived
about 40,000 years ago. El Abra site is one of the most
ancient and completely studied of human occupation
finding. (http://todacolombia.com/etapasprecolobinas.html)
(Fig. 1). Embera speaking Amerindians are postulated to
have come to Jaidukama ethnic group placement through
Panama or through Caribbean Islands Arch, following
the Orinocco River basin (http://todacolombia.com/
etapasprecolobinas.html; (Fig. 1). However the amount of
Panama Embera speakers makes it difficult the arrival
through other ways different to Panama. Jaidukama ethnic
group placement is presently close to Ituango city (Antio-
quia, Colombia). Only in the last years have been
Fig. 1 Jaidukama ethnic group placement is presently close to
Ituango city, Antioquia province, Colombia. Yellow colour: Areas
where Embera Katıo language is used. (Color figure online)
Fig. 2 Jaidukama’s dwellings are built on stakes and are separated by
extensive areas of forest vegetation. They form small concentration of
round type houses, usually inhabited by related people
3690 Mol Biol Rep (2011) 38:3689–3701
123

Jaidukama visited regularly and recently by urbanized I-
tuango city inhabitants. Jaidukama speak Embera-Katio
dialect (http://www.ituango-antioquia.gov.co/index.shtml).
This very much secluded Jaidumaka Amerindian popula-
tion belongs to the ethnic and linguistic Embera group,
which is spoken by some 30,000 Amerindians and exhibit a
scattered settlement pattern in Panama and Colombia
(todacolombia.com/etapasprecolobinas.html).
In the present work, we have studied the HLA gene
profile in the Embera-Katio speaking community of the
Jaidukama Amerindians and compared it with other North
and South American First Inhabitants and worldwide
populations. Our aims are: (1) To determine the HLA class
I (-A and -B) and class II (-DRB1, -DQA1 and -DQB1)
Jaidukama alleles by using indirect DNA sequencing, (2) to
establish real haplotypes by family studies and (3) to
compare the Jaidukama HLA profile with that of other First
American Natives and other worldwide populations in
order to clarify the much debated peopling of the Americas
and in order to establish future transplant and epidemiology
programs of HLA marker diseases.
Materials and methods
Thirty-nine healthy individuals belonging to 13 families
including the father, mother and one child were HLA class
I and class II typed. Whenever a parent was missing,
Table 1 Populations studied in the present work. A total of 12,202 chromosomes were analyzed and geographical locations are represented in
Fig. 1
Identification
number
Population n Reference Identification
number
Population N Reference
1 Mazatecan 89 Arnaiz-Villena et al. [32] 24 Koryaks 92 Grahovac et al. [60]
2 Seri 100 Petzl-Erler et al. [56] 25 Chukchi 59 Grahovac et al. [60]
3 Mixe 55 Petzl-Erler et al. [56] 26 Kets 22 Grahovac et al. [60]
4 Mixteco 103 Petzl-Erler et al. [56] 27 Evenks 35 Grahovac et al. [60]
5 Zapoteco 75 Petzl-Erler et al. [56] 28 Singapore-Chinese 71 Imanishi et al. [33, 34]
6 Mexican Mestizo 99 Vargas-Alarcon et al. [80] 29 Buyi 70 Imanishi et al. [33, 34]
7 Wayuu 112 Yunis et al. [54] 30 Manchu 50 Geng et al. [69]
8 Arhuaco 123 Yunis et al. [54] 31 Korean 100 Imanishi et al. [33, 34]
9 Kogi 67 Yunis et al. [54] 32 Japanese 493 Imanishi et al. [33, 34]
10 Arsario 20 Yunis et al. [54] 33 Khalk-Mongolian 202 Munkhbat et al. [59]
11 Cayapa 100 Titus-Trachtenberg et al. [55] 34 Tuvins 197 Martinez-Laso et al. [70]
12 Xavantes 74 Cerna et al. [20] 35 Khoton-Mongolian 85 Munkhbat et al. [59]
13 Guarani 32 Cerna et al. [20] 36 Germans 295 Imanishi et al. [33, 34]
14 Toba-Pilaga 19 Cerna et al. [20] 37 Sardinians 91 Imanishi et al. [33, 34]
15 Mataco- Wichi 49 Cerna et al. [20] 38 Italians 284 Imanishi et al. [33, 34]
16 Eastern-Toba 135 Cerna et al. [20] 39 French 179 Imanishi et al. [33, 34]
17 Jaidukama 39 Present study 40 Spaniards 176 Martinez-Laso et al. [27]
18 Aymara 101 Arnaiz-Villena et al. [66] 41 Spanish-Basques 80 Martinez-Laso et al. [27]
19 Eskimos 35 Grahovac et al. [60] 42 Algerians 102 Arnaiz-Villena et al. [71]
20 Athabascan 124 Monsalve et al. [58] 43 Berbers 98 Izaabel et al. [72]
21 Tlingit 53 Imanishi et al. [33, 34] 44 Morocans 98 Gomez-Casado et al. [73]
22 Nivkhs 32 Grahovac et al. [60] 45 West Samoa 102 Gao et al. [75]
23 Udegeys 23 Grahovac et al. [60] 46 Madang 65 Gao et al. [74, 75, 76]
47 Rabaul 60 Gao et al. [74, 75, 76] 54 Cape York 80 Gao et al. [74, 75, 76]
48 New Caledonia . 65 Gao et al. [74, 75, 76] 55 South African Blacks 59 Imanishi et al. [33, 34]
49 Fiji 57 Gao et al. [74, 75, 76] 56 North American
Blacks
447 Imanishi et al. [33, 34]
50 Papua New Guinean 57 Gao et al. [74, 75, 76] 57 Hottentot 65 Imanishi et al. [33, 34]
51 Central Desert 152 Lester et al. [77] 58 Bushmen 103 Imanishi et al. [33, 34]
52 Yuendumu 119 Lester et al. [77] 59 Ainu 50 Bannai et al. [61]
53 Kimberley 82 Gao et al. [74, 75, 76]
West Samoa From Central Polynesia, Madang Melanesians on the North New Guinea mainland Coast, Rabaul Melanesians in New Britain, NewCaledonia Melanesians from Noumea, Fiji From largest Fijian island; Central desert, Yuendumu, Kimberley, and Cape York: Australian aborigines
Mol Biol Rep (2011) 38:3689–3701 3691
123

samples were taken from two of the children in order to
construct the full parental HLA haplotypes.
The samples were extracted and rapidly transported by
helicopter to the CIB (Corporacion para Investigaciones
Biologicas) laboratories in Medellin. The Jaidukama set-
tlement is located in the northwestern part of the Antioquia
province (Fig. 1). There were 157 Jaidukama Amerindians
distributed among 27 families that inhabit 27 tambos (areas
separated from other families); these natives were in this
place probably since long time ago surrounded by a thick
forest. Blood from volunteers were taken in the frameshift
of a preventive and epidemiology work. Full consent was
given by Jaidukama volunteers. Their dwellings are char-
acteristically built on stakes and are separated by extensive
areas of forest vegetation that require several hours of walk
between dwellings. They form small concentrations of
three to four round-type houses according to parental ties
(Fig. 2). The main source of nourishment is constituted by
plantains, corn and beans. Women know only the local
dialect (a specific Embera-Katio one) and men are in
charge of communication between neighboring ethnic
groups, which leads to think there has not been much
mixing with other races since they establishment in site.
Each individual studied was born in the Antioquia forest or
other small urban areas, had a Jaidukama physical aspect
and their parents had been born in this area; they also spoke
the specific Embera-Katio dialect [23, 24]. The origin of all
other populations used for comparisons are detailed in
Table 1; a total of 12,202 chromosomes were studied. It
includes populations from different origins, Caucasoids,
Orientals, Negroids, Polynesians, Micronesians, Na-Dene,
Eskimos and Amerindians. Particularly, the Amerindians
includes groups from, Macro-Mixteco (Mixtecan and
Zapotecan), Macro-Maya (Mixe), Macro-Yuma (Seris),
Chibcha (Arsario, Kogi, Arhuaco and Cayapa), Arawak
(Wayu) and Ge Pano Caribe (Xavantes, Mataco and Toba)
linguistic families [23, 24] (Fig. 5).
HLA genotyping and DNA sequencing
Generic HLA class I (A and B) genotyping was done by a
PCR-SSO technique as previously described [25] with
slight modifications that yield a resolution equivalent to the
standard serology techniques (see Results and Discussion
and Table 2). Reverse dot-blot technique with the Auto-
mated Innolipa system (Innogenetics N.V., Zwijndrecht,
Belgium) were used when oligotyping yield ambiguous
results. High Resolution HLA class II (DRB1 DQA1, and
DQB1) was performed by using reverse dot-blot technique
with the Automated Innolipa system (Innogenetics N.V.,
Zwijndrecht, Belgium). HLA-A, -B, -DRB1, -DQA1, and -
DQB1 allele DNA sequencing was only done when indirect
DNA typing yielded ambiguous results as described in
detail by [26]. High resolution HLA-B typing was made by
sequencing technology as previously described [27–29].
Statistical analysis
Statistical analysis was performed with Arlequin vl.l soft-
ware kindly provided by [30]. In summary, this program
calculated HLA-A, -B, -DRB1, -DQA1 and -DQB1 allele
frequencies, Hardy–Weinberg equilibrium and the linkage
disequilibrium between two alleles at two different loci.
Their level of significance (P) for 2 9 2 comparisons was
determined as previously described [31]. In addition, the
most frequent complete haplotypes were tentatively
deduced from: (1) the 2, 3, and 4 HLA loci haplotype
frequencies [32]; (2) the previously described haplotypes in
other populations [33]; and (3) haplotypes if they appeared
in two or more individuals and the alternative haplotype
was well defined [33]. In order to compare phenotype and
haplotype HLA frequencies with other populations, the
reference tables of the 11th and 12th International HLA
Workshops were used [34, 35] (also see Table 1). Phylo-
genetic trees (dendrograms) were constructed with the
allelic frequencies by using the Neighbor-Joining (NJ)
method [36] with the genetic distances between popula-
tions [37], by using the software DISPAN which contain
the programs GNKDST and TREEVIEW [38, 39]. Corre-
spondence analysis in three dimensions and its bidimen-
sional representation was carried out by using the VISTA
v5.02 computer program [40] (http://forrest.psych.unc.edu).
Table 2 Allele frequencies in Jaidukama Amerindians (Only unre-
lated parents are considered)
Alleles Allele frequencies (%)
HLA-A
A*02 30.8
A*24 69.2
HLA-B
B*35 50.0
B*39 9.6
B*40 7.7
B*51 32.7
HLA-DRB1*
1602 23.1
0407 32.7
1402 28.8
0404 15.4
HLA-DQB1*
301 44.2
302 55.8
DRB1*0404, 0407, 1402 and 1604 are Amerindian alleles which are
also found in other American, Asian or Pacific populations [47]
3692 Mol Biol Rep (2011) 38:3689–3701
123

Table 3 HLA haplotypes distribution in Jaidukama Amerindian families (segregation analysis)
Extended HLA haplotypes
HLA-A HLA-B HLA-DRB1 HLA-DQA1 HLA-DQB1
Family 1
F A24 B40 DRB1*1602 DQA1*0501 DQB1*0301
A24 B35 DRB1*1402 DQA1*0501 DQB1*0301
M A24 B35(*3510) DRB1*1602 DQA1*0501 DQB1*0301
A24 B35(*3501) DRB1*1402 DQA1*0501 DQB1*0301
S A24 B35 DRB1*1602 DQA1*0501 DQB1*0301
A24 B35 DRB1*1402 DQA1*0501 DQB1*0301
Family 2
F A24 B39 DRB1*1402 DQA1*0501 DQB1*0301
A2 B40 DRB1*0407 DQA1*03 DQB1*0302
M A2 B40 DRB1*1402 DQA1*0501 DQB1*0301
A24 B35 DRB1*1402 DQA1*0501 DQB1*0301
S A2 B40 DRB1*0407 DQA1*03 DQB1*0302
A24 B35 DRB1*1402 DQA1*0501 DQB1*0301
Family 3
F A2 B35 DRB1*0404 DQA1*03 DQB1*0302
A24 B40 DRB1*0407 DQA1*03 DQB1*0302
M A24 B35(*3501) DRB1*0407 DQA1*03 DQB1*0302
A2 B51 DRB1*0407 DQA1*03 DQB1*0302
S A2 B35 DRB1*0404 DQA1*03 DQB1*0302
A2 B51 DRB1*0407 DQA1*03 DQB1*0302
Family 4
F A24 B35(*3510) DRB1*1602 DQA1*0501 DQB1*0301
A24 B40 DRB1*1402 DQA1*0501 DQB1*0301
M A24 B35 DRB1*1402 DQA1*0501 DQB1*0301
A24 B40 DRB1*1602 DQA1*0501 DQB1*0301
S A24 B40 DRB1*1602 DQA1*0501 DQB1*0301
A24 B40 DRB1*1402 DQA1*0501 DQB1*0301
Family 5
F A2 B40 DRB1*1402 DQA1*0501 DQB1*0301
A24 B35(*3504) DRB1*0404 DQA1*03 DQB1*0302
M A2 B40 DRB1*1402 DQA1*0501 DQB1*0301
A24 B35(*3510) DRB1*1602 DQA1*0501 DQB1*0301
S A24 B35 DRB1*1602 DQA1*0501 DQB1*0301
A24 B35 DRB1*0404 DQA1*03 DQB1*0302
Family 6
F A2 B51 DRB1*0404 DQA1*03 DQB1*0302
A2 B40 DRB1*0404 DQA1*03 DQB1*0302
M A24 B35(*3501) DRB1*0407 DQA1*03 DQB1*0302
A24 B35 DRB1*1402 DQA1*0501 DQB1*0301
S A2 B51 DRB1*0404 DQA1*03 DQB1*0302
A24 B35 DRB1*1402 DQA1*0501 DQB1*0301
Family 7
F A24 B35(*3504) DRB1*0404 DQA1*03 DQB1*0302
A2 B35 DRB1*0407 DQA1*03 DQB1*0302
M A24 B40 DRB1*1402 DQA1*0501 DQB1*0301
A2 B35 DRB1*0407 DQA1*03 DQB1*0302
Mol Biol Rep (2011) 38:3689–3701 3693
123

Table 3 continued
Extended HLA haplotypes
HLA-A HLA-B HLA-DRB1 HLA-DQA1 HLA-DQB1
S A2 B35 DRB1*0407 DQA1*03 DQB1*0302
A2 B35 DRB1*0407 DQA1*03 DQB1*0302
Family 8
F A24 B40 DRB1*0407 DQA1*03 DQB1*0302
A2 B35 DRB1*0407 DQA1*03 DQB1*0302
M A2 B35 DRB1*0407 DQA1*03 DQB1*0302
A24 B39 DRB1*0407 DQA1*03 DQB1*0302
S A2 B35 DRB1*0407 DQA1*03 DQB1*0302
A24 B40 DRB1*0407 DQA1*03 DQB1*0302
Family 9
F A2 B51 DRB1*0404 DQA1*03 DQB1*0302
A24 B35(*3504) DRB1*0404 DQA1*03 DQB1*0302
M A2 B40 DRB1*0404 DQA1*03 DQB1*0302
A24 B39(*3905) DRB1*1402 DQA1*050 DQB1*0301
S A24 B35(*3504) DRB1*0404 DQA1*03 DQB1*0302
A24 B39(*3905) DRB1*1402 DQA1*050 DQB1*0301
Family 10
F A24 B35(*3501) DRB1*0407 DQA1*03 DQB1*0302
A24 B35(*3504) DRB1*0404 DQA1*03 DQB1*0302
M A24 B35(*3510) DRB1*1602 DQA1*050 DQB1*0301
A24 B40 DRB1*1402 DQA1*050 DQB1*0301
S A24 B35 DRB1*0407 DQA1*03 DQB1*0302
A24 B40 DRB1*1402 DQA1*050 DQB1*0301
Family 11
F A24 B35(*3501) DRB1*0407 DQA1*03 DQB1*0302
A24 B35(*3504) DRB1*0404 DQA1*03 DQB1*0302
M A24 B40 DRB1*1402 DQA1*050 DQB1*0301
A2 B35 DRB1*0407 DQA1*03 DQB1*0302
S A24 B35 DRB1*0407 DQA1*03 DQB1*0302
A2 B35 DRB1*0407 DQA1*03 DQB1*0302
Family 12
F A2 B51 DRB1*0404 DQA1*03 DQB1*0302
A24 B40 DRB1*1402 DQA1*050 DQB1*0301
M A24 B35(*3510) DRB1*1602 DQA1*050 DQB1*0301
A24 B39(*3905) DRB1*1402 DQA1*050 DQB1*0301
S A24 B39 DRB1*1402 DQA1*050 DQB1*0301
A24 B40 DRB1*1402 DQA1*050 DQB1*0301
Family 13
F A24 B35(*3501) DRB1*0407 DQA1*03 DQB1*0302
A24 B39 DRB1*0407 DQA1*03 DQB1*0302
M A24 B40 DRB1*1602 DQA1*050 DQB1*0301
A24 B40 DRB1*0404 DQA1*03 DQB1*0302
S1 A24 B35(*3501) DRB1*0407 DQA1*03 DQB1*0302
A24 B40 DRB1*1602 DQA1*050 DQB1*0301
S2 A24 B40 DRB1*0404 DQA1*03 DQB1*0302
A24 B35 DRB1*0407 DQA1*03 DQB1*0302
* Deduced from HLA siblings typings
3694 Mol Biol Rep (2011) 38:3689–3701
123

Correspondence analysis consists of a geometric technique
that may be used for displaying a global view of the rela-
tionships among populations according to HLA (or other)
allele frequencies. This methodology is based on the allelic
frequency variance among populations (similarly to the
classical components methodology) and on the display of
statistical visualization of the differences.
Results and discussion
Characteristic HLA allele frequencies of the Jaidukama
Amerindian population compared to other populations
Table 2 shows the HLA allele frequencies found in the
Jaidukama population. Two different HLA-A (A2 and A24)
and four different HLA-B (B35, B39, B40 and B51) genetic
alleles were found in the Jaidukama ethnic group. HLA-B
subtypes were determined in a few number of individuals by
DNA sequencing due to the lack of sufficient DNA amount.
Alleles found were B*3501, B*3504, B*3510, B*3905,
B*4002 and B*4004 (see Table 3). These alleles have been
previously described in Amerindians [27, 28, 41–43] and
now are included in some extended haplotypes. Regarding
to the HLA class II alleles, four different DRB1 alleles were
found at similar frequencies (DRB 1*0404, *0407, *1602
and *1402) in the studied population, DRB 1*0404 allele
has been also found in Eastern Island [44–46]; all four
DRB1 alleles have been found in well defined Asian and/or
Pacific ethnic groups [47]. DQB1 allele frequencies reflect
the DRB1 locus allele distribution due to the strong linkage
disequilibrium between these two loci and confirmed by
family studies (see Table 3). Data reported in this study on
HLA class I and class II polymorphism are concordant with
those previously described in other ethnic Amerindian
groups [20, 48–53]. It is striking that B35 family has several
variants in such small relatively reduced group. If this is so,
it would mean that relative inbreeding might enhance gene
conversion events, and diversification.
Two types of analyses were done in order to compare
Jaidukama HLA frequencies with other world population
frequencies using unrelated parents (n = 26): (1) with
pooled DRB1 and DQB1 data; and (2) with DRB1 only. It
was impossible to do a study comparing HLA class I allele
frequencies or HLA class I and II conjointly due to the lack
of class I studies (particularly in high resolution) in many
Amerindians and other populations. The DRB1 study was
made to compare the Amerindian HLA population fre-
quencies with those of Polynesians, Melanesians and
Micronesians who lacked DQB1 analyses (Table l).
Fig. 3 Neighbor-Joining (NJ) dendrogram shows relationships
between Jaidukama and other Amerindians, Caucasoids, Na-Dene/
Eskimos and Orientals. Genetic distances between populations were
calculated from HLA-DRB1 and -DQB1 frequencies. Populations
used for study are: Mazatecos [57]; Seri [56]; Mixe [56]; Mixteco
[56]; Zapoteco [56]; Mexican Mestizo (Unpublished); Arhuaco [54];
Kogi [54]; Arsario [54]; Cayapa [55]; Xavante [20]; Guarani [56];
Toba-Pilaga [20]; Mataco-Wichi [20]; Eastern-Toba [20]; Jaidukama
(Present study); Eskimos [60]; Athabascans [58]; Tlingit [33]; Nivkhs
[60]; Udegeys [60]; Koryaks [60]; Chukchi [60]; Kets [60]; Evenks
[33]; Singapore-Chinese [33]; Buyi [33]; Manchu [69]; Coreans [33];
Japonese [33]; Khalk-Mongolians [59]; Tuvins [70]; Khoton-Mon-
golians [59]; Germans [33]; Sardinians [33]; Italians [33]; French
[33]; Spanish [32]; Spanish-Basque [32]; Algerians [71]; Moroccan-
Bereberes [72]; Moroccans [73]; Western Samoa [74]; Madang [75];
Rabaul [76]; New Caledonia [74]; Fidji [75]; Papua-Nueva Guinea
[76]; Central Desert; [77]; Yuendumu [77]; Kimberley [74]; Cape
York [75]; South American Negroids [33]; North American Negroids
[33]; Hottentots [33]; San (Bushmen) [33]; Ainu [61]; Zuni [33];
NYC Negroids [78]; Bubi [79]; Caucasoids [45]; Choco [45];
Providencia [45]; Senegalese [33]; Wayu (Unpublished); Afrocolom-
bians [65]; Colombian Mestizos
Mol Biol Rep (2011) 38:3689–3701 3695
123
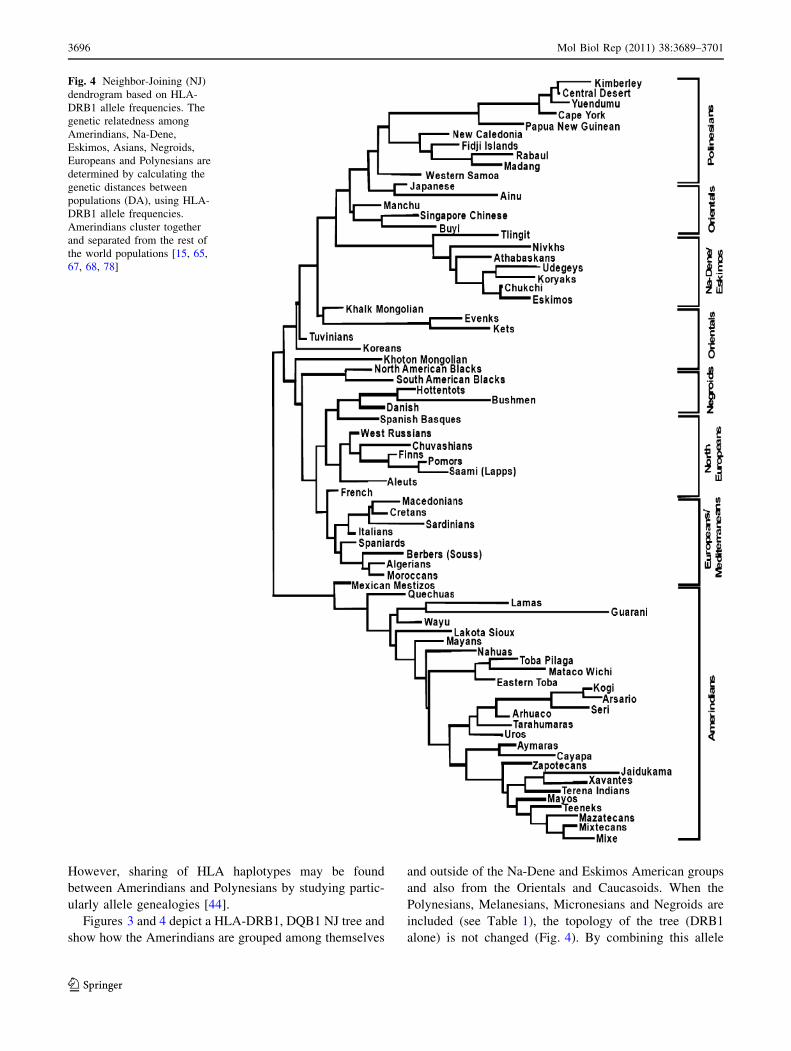
However, sharing of HLA haplotypes may be found
between Amerindians and Polynesians by studying partic-
ularly allele genealogies [44].
Figures 3 and 4 depict a HLA-DRB1, DQB1 NJ tree and
show how the Amerindians are grouped among themselves
and outside of the Na-Dene and Eskimos American groups
and also from the Orientals and Caucasoids. When the
Polynesians, Melanesians, Micronesians and Negroids are
included (see Table 1), the topology of the tree (DRB1
alone) is not changed (Fig. 4). By combining this allele
Fig. 4 Neighbor-Joining (NJ)
dendrogram based on HLA-
DRB1 allele frequencies. The
genetic relatedness among
Amerindians, Na-Dene,
Eskimos, Asians, Negroids,
Europeans and Polynesians are
determined by calculating the
genetic distances between
populations (DA), using HLA-
DRB1 allele frequencies.
Amerindians cluster together
and separated from the rest of
the world populations [15, 65,
67, 68, 78]
3696 Mol Biol Rep (2011) 38:3689–3701
123

frequency studies with particular quasi-specific Amerin-
dian alleles a gene flow may be detected between Amer-
indians and Pacific and other Asian people [15].
HLA-A, -B, -DRB1, DQA1 and DQB1 correspondence
analysis (Fig. 5) and extended haplotypes (Tables 3
and 4) in Jaidukama Amerindians
Correspondence analysis with DRB1 and DQB1 alleles
(Fig. 5) shows that Amerindians constitute a different
cluster from Na-Dene, Eskimo, Siberian, Asian and Cau-
casoid populations. This supports results in dendrograms
(Figs. 3 and 4). Phylogenetic trees and correspondence
analyses results support the hypothesis that Amerindians
have a unique genetic background but with a noticeable
gene flow between them and Asian and Pacific Islanders
[15].
Real extant (not calculated by statistic methods but by
family haplotype segregation) HLA haplotypes were
described for the first time in a small and secluded Amerin-
dian population, due to the possibility to study families:
inheritance pattern (see Tables 3 and 4). The HLA class II
haplotypes found are characteristics of the Amerindian pop-
ulations although some of them appear in non-Amerindian
populations: (a) DRB1*1602-DQA1*0501-DQB1*0301 is
exclusively found in Amerindians [20, 54–57]; (b)
DRB1*0404-DQA1*03011-DQB1*0302 is present in other
Amerindians [20, 55–57], in the Penutian Indians from
British Columbia [58], and in low frequency in Orientals
(Singapore-Chinese, Japanese, Khoton-Mongolian and
Khalk-Mongolian [33, 59]. This suggests a positive gene flow
between Orientals and Amerindians [15]; (c) DRB1*1402-
DQA1*0501-DQB 1*0301 present in other Amerindians [15,
20, 54–57], in the Na-Dene Indians, Eskimos and Siberians in
high frequency [58, 60] and is also found in low frequency in
Orientals [33, 59]; and (d) DRB1*0407-DQA1*03011-DQB
1*0302 present in other Amerindians [20, 54, 56, 57] and in
low frequency in NaDene Indians [58].
On the other hand, DRB1*1406-DQA1*0501-
DQB1*0301 haplotype has previously been described in
Amerindians and in Ainu Asians [20, 57, 61] and
DRB1*0802-DQA1*0401-DQB1*0402 haplotype previ-
ously described in Amerindians, Na-Dene Indians, Eskimos
and Ainu Asians, [20, 54–58, 61] are not present in the Ja-
idukama Amerindians.
Also, the lack of Caucasoid and Negroid haplotypes
reflects the absence of admixture of Jaidukama Amerindi-
ans with other ethnic groups.
The extended haplotypes found in Jaidukama Amerindi-
ans are shown in Table 4. The DRB1*0407-DQA1*03011-
DQB1*0302 class II haplotype was included in several
extended haplotypes with A2-B35, A24-B35, A2-B40, A24-
B40, A24-B39, and A2-B51. Haplotype DRB1*1602-
DQA1*0501-DQB1*0301 was found together with A24-
B40 and A24-B35. Haplotype DQA1*03011-DQB1*0302 is
found with A2-B40, A24-B40, A2-B35, A24-B35 and A2-
B51 and haplotype DRB1*1602-DQA1*0501-DQB1*0301
is observed with A24-B35 and A24-B40.
Some full extended haplotypes (including genetic HLA-A
and B typings) may be the same than these found in Lakota-
Fig. 5 Correspondence analysis
based on HLA-DRB1 and HLA-
DQB1 allele frequencies. The
analysis shows a global view of
the genetic relationships among
Amerindian, Na-Dene, Eskimo,
Asian, Negroid and European
populations according to HLA-
DRB1 and HLA-DQB1 allele
frequencies. These relationships
are calculated in n dimensions
and represented in two
dimensions. Colours represent
an approximate grouping of
populations [15, 65, 67, 68, 78].
(Color figure online)
Mol Biol Rep (2011) 38:3689–3701 3697
123

Sioux [62]. This highlights how the same haplotypes may be
found in North (Lakota-Sioux) and South American Amer-
indians (Jaidukama), probably pointing out the existence of a
founder effect for both North and South Amerindians. Also
the fact that these small Jaidukama group has probably
evolved by having a considerable amount of new specific
extended HLA haplotypes supports that haplotypes may vary
faster than alleles [15]. Data on HLA class I and class II
extended haplotypes studies in Amerindian populations are
now quite complete [15, 47]: 5 out of 17 haplotypes found in
Jaidukama Amerindians have not been previously described
as Amerindians; this is a striking finding. Full HLA haplo-
types may be more important for an efficient immune
response than alleles [63]. Finally, the sharing of this
secluded Jaidukama Amerindians of HLA alleles and DRB1-
DQB1 modules with Asian and Pacific Islanders supports
existence of a gene flow of Amerindians with outside-
America populations [15]. The study of this small and
secluded (until recently) Amerindian groups are not only of
anthropological and immunological interest, but also
important for future transplantation programs and HLA
linked diseases epidemiology studies.
Acknowledgments This work was supported in part by grants from
the Spanish Ministry of Health (FISS PI051039 and PI080838), and
three different Mutua Madrilena Automovilista grants. We are
grateful to the Jaidukama Indian Community, Organizacion Indi-
genista (OIA) and to Programa Aereo de Salud del Servicio Seccional
de Salud de Antioquia.
References
1. Crawford MH (1998) The origins of Native Americans: evidence
from anthropological genetics. Cambridge University Press,
Cambridge
2. Kolman CJ, Sambuughin N, Bermingham E (1996) Mitochon-
drial DNA analysis of Mongolian populations and implications
for the origin of New World founders. Genetics 142:1321–1334
3. Merriwheter DA, Hall WW, Vahlne A, Ferrell RE (1996) mtRNA
variation indicates Mongolia may have been the source for the
founding population for the New World. Am J Hum Genet
59:204–212
Table 4 Haplotype frequency of the extended haplotypes in Jaidukama Amerindians (Only unrelated parents are considered)
Extended HLA haplotypes Haplotype
frequencies (%)
Possible origin
HLA-A HLA-B HLA-DRB1 HLA-DQA1 HLA-DQB1
1. A24 B35(*3501) DRB1*0407 DQA1*03 DQBl*0302a 11.5 Amerindian
2. A2 B35 DRB1*0407 DQA1*03 DQBl*0302b 9.6 Amerindian
3. A24 B35(*3510) DRB1*1602 DQA1*0501 DQBl*0301c 9.6 Jaidukama
4. A24 B40 DRB1*1402 DQA1*0501 DQBl*0301d 9.6 Amerindian
5. A24 B35(*3504) DRB1*0404 DQA1*03 DQBl*0302c 9.6 Jaidukama
6. A24 B35 DRB1*1402 DQA1*0501 DQB 1*0301e 7.7 Amerindian
7. A24 B39(*3905) DRB1*1402 DQA1*0501 DQB1*0301c 5.8 Jaidukama
8. A24 B40 DRB1*1602 DQA1*0501 DQB 1*0301c 5.8 Jaidukama
9. A2 B40 DRB1*1402 DQA1*0501 DQB 1*0301f 5.8 Amerindian
10. A2 B51 DRB1*0404 DQA1*03 DQBl*0302c 5.8 Jaidukama
11. A24 B40 DRB1*0407 DQA1*03 DQBl*0302c 3.8 Jaidukama
12. A2 B40 DRB1*0404 DQA1*03 DQB1*0302c 3.8 Jaidukama
13. A24 B39 DRB1*0407 DQA1*03 DQBl*0302g 3.8 Amerindian
14. A24 B40 DRB1*0404 DQA1*03 DQB1*0302c 1.9 Jaidukama
15. A2 B40 DRB1*0407 DQA1*03 DQB1*0302h 1.9 Amerindian
16. A2 B51 DRB1*0407 DQA1*03 DQBl*0302i 1.9 Amerindian
17. A2 B35 DRB1*0404 DQA1*03 DQB1*0302c 1.9 Jaidukama
a This haplotype has also been found in Lakota-Sioux [62]b Haplotype also found in Mazatecos [57]c These haplotypes have only been found on in Jaidukama (present work)d This haplotype has also been found in Yucpa [64]e This haplotype has also been found in Uros [65] and Aymaras [66]f Haplotype also found in Tarahumara [67], Sioux [62] and Aymaras [65]g This haplotype has also been found in Mazatecos [51]h This haplotype is also found in Aymaras [66] and Lamas [68]i This haplotype has also been found in Sioux [62]
3698 Mol Biol Rep (2011) 38:3689–3701
123

4. Greenberg JH, Turner CG, Zegura SL (1986) The settlement of
the Americas: a comparison of the linguistic, dental and genetic
evidence. Curr Anthropol 27:477–498
5. Wallace DC, Torroni A (1992) American Indian prehistory as
written in the mitochondrial DNA: a review. Hum Biol
64:403–416
6. Cavalli-Sforza LL, Menozzi P, Piazza A (1994) The history and
geography of human genes. Princeton University Press, Princeton
7. Parham P, Ohta T (1996) Population biology of antigen presen-
tation by MHC class I molecules. Science 272:67–74
8. Horai S, Kondo R, Nakagawa-Hattori Y, Hayashi S, Sonoda S,
Tajima K (1993) Peopling of the Americas, founded by four
major lineages of mitochondrial DNA. Mol Biol Evol 10:23–47
9. Torroni A, Sukernik RI, Schurr TG, Starikovskaya YB, Cabell
MF, Crawford MH, Comuzzie Ag (1993) mtDNA variation of
aboriginal Siberians reveals distinct genetic affinities with Native
Americans. Am J Hum Genet 53:591–608
10. Santos FR, Rodriguez-Delfin L, Pena SDJ, Moore J, Weiss KM
(1996) North and South Amerindians may have the same major
founder Y chromosome haplotype. Am J Hum Genet
58:1369–1370
11. Karafet T, Zegura SL, Vuturo-Brady J, Posukh O, Osipova L,
Weibe V, Romero F (1997) Y chromosome markers and trans-
Bering Strait dispersals. Am J Phys Anthropol 102:301–314
12. Santos FR, Pandya A, Tyler-Smith C, Pena SD, Schanfield M,
Leonard WR, Osipova L, Crawford MH, Mitchell RJ (1999) The
central Siberian origin for native American Y chromosomes. Am
J Hum Genet 64:619–628
13. Bolnick A, Shook B, Campbell L, Goddard I (2004) Problematic
use of Greenberg’s linguistic classification of the Americas in
studies of Native American genetic variation. Am J Hum Genet
75:519–523
14. Wilson JAP (2008) A new perspective on later migrations: the
possible recent origin of some Native American haplotypes. Crit
Anthropol 28:267–278
15. Arnaiz-Villena A, Parga-Lozano C, Moreno E et al (2010) The
origin of Amerindians and the peopling of the Americas
according to HLA genes: admixture with Asian and Pacific
people. Curr Genomics 11(2):103–114
16. Uinuk-Ool TS, Takezaki N, Sukernik RI, Nagl S, Klein J (2002)
Origin and affinities of indigenous Siberian populations as
revealed by HLA class II gene frequencies. Hum Genet
110:209–226
17. Mulligan CJ, Kitchen A, Miyamoto MM. (2008) Updated three-
stage model for the peopling of the Americas. PLoS ONE p-3199
18. Goebel T, Waters MR, O’Rourke DH (2008) The late Pleistocene
dispersal of modern humans in the Americas. Science
319:1497–1502
19. Leon-S FE, Ariza-Deleon A, Leon-S ME, Ariza-C A (1996)
Peopling the Americas. Science 273:723–725
20. Cerna M, Falco M, Friedman H, Raimondi E, Maccagno A,
Fernandez-Vina M, Stastny P (1993) Differences in HLA class II
alleles of isolated South American Indian populations from Brazil
and Argentina. Hum Immunol 37:213–220
21. Bruges-Armas J, Martinez-Laso J, Martins B, Allende L, Gomez-
Casado E, Longas J, Varela P, Castro MJ, Arnaiz-Villena A
(1999) HLA in the Azores Archipelago: possible presence of
Mongoloid genes. Tissue Antigens 54:349–359
22. Holden C (1999) Were Spaniards among the first Americans?
Science 286:1467–1468
23. Ruhlen M (1987) A guide to the world’s languages. Volume I:
classification. Edward Arnold (ed). London, UK
24. Swadesh M (1959) Indian linguistic groups of Mexico. Escuela
Nacional de Antropologia e Historia, Mexico
25. Arguello R, Avakian H, Goldman JM, Madrigal JA (1996) A
novel method for simultaneous high resolution identification of
HLA-A, HLA-B, and HLA-Cw alleles. Proc Natl Acad Sci USA
93:10961–10965
26. Arnaiz-Villena A, Timon M, Corell A, Perez-Aciego P, Martin-
Villa JM, Regueiro JR (1992) Primary immunodeficiency caused
by mutations in the gene encoding the CD3-y subunit of the T-
lymphocyte receptor. N Eng J Med 327:529–533
27. Martinez-Laso J, Layrisse Z, Gomez-Casado E, Montoya F,
Guedez Y, Domınguez E, Varela P, Arnaiz-Villena A (1995) A
new HLA-B15 allele (B*1522) found in Bari- Motilones Amer-
indians in Venezuela: comparison of its intron 2 sequence with
those of B* 1501 and B*3504. Immunogenetics 43:108–109
28. Gomez-Casado E, Vargas-Alarcon G, Martinez-Laso J, Perez-
Bias M, Granados J, Layrisse Z, Montoya F, Varela P, Arnaiz-
Villena A (1997) Generation of the HLA-B35, -B5, -B16, and
B15 groups of alleles studied by intron 1 and 2 sequence analysis.
Immunogenetics 46:469–476
29. Gomez-Casado E, Vargas-Alarcon G, Martinez-Laso J, Granados
J, Varela P, Alegre R, Longas J, Gonzalez-Hevilla M, Martin-
Villa JM, Arnaiz-Villena A (1999) Tissue Antigens 53:153–160
30. Excoffier L, Slatkin M (1995) Maximum-likelihood estimation of
molecular haplotype frequencies in a diploid population. Mol
Biol Evol 12:921–927
31. Imanishi T, Akaza T, Kimura A, Tokunaga K, Gojorobi T (1992)
Estimation of allele and haplotype frequencies for HLA and
complement loci. In: Tsuji K, Aizawa M, Sasazuki T (eds) HLA
1991, Vol I. Oxford University Press, Oxford, pp 76–79
32. Arnaiz-Villena A, Martınez-Laso J, Gomez-Casado E (1997)
Relatedness among Basques, Portuguese, Spaniards, and Algerian
studied by HLA allelic frequencies and haplotypes. Immunoge-
netics 47:37–43
33. Imanishi T, Akaza T, Kimura A, Tokunaga K, Gojobori T
(1992a) Allele and haplotype frequencies for HLA and comple-
ment loci in various ethnic groups. In: Tsuji K, Aizawa M,
Sasazuki T (eds) HLA 1991, vol I. Oxford University Press,
Oxford, pp 1065–1220
34. Imanishi T, Wakisaka A, Gojorobi T (1992b) Genetic relation-
ships among various human populations indicated by MHC
polymorphisms. In: Tsuji K, Aizawa M, Sasazuki T (eds) HLA
1991. Vol I. Oxford University Press, Oxford, pp 627–632
35. Clayton J, Lonjou C (1997) Allele and haplotype frequencies for
HLA loci in various ethnic groups. In: Charron D (ed) Genetic
diversity of HLA, functional and medical implication, Vol I.
EDK, Paris, pp 665–820
36. Saitou N, Nei M (1987) The neighbor-joining method: a new
method for reconstructing phylogenetic tree. Mol Biol Evol 4:406
37. Nei M (1972) Genetic distances between populations. Am Nat
106:283
38. Nei M (1973) Analysis of gene diversity in subdivided popula-
tions. Proc Natl Acad Sci USA 70:3321–3323
39. Nei M, Tajima F, Tateno Y (1983) Accuracy of estimated phy-
logenetic trees from molecular data. J Mol Evol 19:153–170
40. Young FW, Bann CM (1996) A visual statistics system. In: Stine
RA, Fox J (eds) Statistical computing environments for social
researches. Sage, Thousand Oaks, pp 207–236
41. Gomez-Casado E, Montoya F, Martinez-Laso J, Bedoya CI,
Varela P, Diaz-Campos N, Arnaiz-Villena A (1995) A new HLA-
B35 allele (B*3510) found in isolated Jaidukama South American
Indians. Immunogenetics 42:231–232
42. Marcos CY, Fernandez-Vina MA, Lazaro AM, Nulf CJ, Rai-
mondi EH, Stastny P (1997) Novel HLA-B35 subtypes: putative
gene conversion events with donor sequences from alleles com-
mon in native Americans (HLA-B*4002 or B*4801). Hum
Immunol 53:148–155
43. Martinez-Arends A, Layrisse Z, Arguello R, Herrera F, Mon-
tagnani S, Matos M, Ross J, Dune P, Marsh SG, Madrigal JA
(1998) Characterization of the HLA class I genotypes of a
Mol Biol Rep (2011) 38:3689–3701 3699
123

Venezuelan Amerindian group by molecular methods. Tissue
Antigens 52:51–56
44. Thorsby E, Flam ST, Woldseth B, Dupuy BM, Sanchez-Mazas A,
Fernandez-Vina MA (2009) Further evidence of an Amerindian
contribution to the Polynesian gene pool on Easter Island. Tissue
Antigens 73:582–585
45. Trachtenberg EA, Keyeux G, Bernal J, Rhodas C, Erlich HA
(1996) Results of Expedicion Humana. I. Analysis of HLA class
II (DRB1-DQA1-DQB1-DPB1) alleles and DR-DQ haplotypes in
nine Amerindian populations from Colombia. Tissue Antigens
48:174–181
46. Tsuneto LT, Probst CM, Hutz MH, Salzano FM, Rodriguez-
Delfin LA, Zago MA, Hill K, Hurtado AM, Ribeiro-dos-Santos
AKC, Petzl-Erler ML (2003) HLA class II diversity in seven
Amerindian populations. Clues about the origins of the Ache.
Tissue Antigens 60:512–526
47. Middleton D, Menchaca L, Rood H, Komerofsky R (2003) New
allele frequency data base. Tissue Antigens 61:403–407
48. Sonoda S, Arce-Gomez B, Satz ML (1992) Ethnic report on
native Americans in South America and Mexico. In: Tsuji K,
Aizawa M, Sasazuki T (eds) HLA 1991. Vol. Oxford University
Press, Oxford, pp 685–688
49. Williams RC, Troup GM, Nelson JL (1992) Report of the North
American Indian anthropology section. In: Tsuji K, Aizawa M,
Sasazuki T (eds) HLA 1991. Vol 1. Oxford University Press,
Oxford, pp 683–685
50. Fernandez-Vina MA, Moraes ME, Gao X, Moraes JR, Salatier I,
Miller S, Tsai S, Sun YAJ, Layrisse Z, Gazit E, Stastny P (1991)
Alleles at four HLA class II loci determined by oligonucleotide
hybridization and their associations in five ethnic groups.
Immunogenetics 34:299–312
51. Erlich HA, Zeidler A, Chang J, Shaws S, Raffel LS, Klitz W,
Beshkoy Y (1993) HLA class II alleles and susceptibility and
resistance to insulin dependent diabetes mellitus in Mexican
American families. Nat Genet 3:358–364
52. Layrisse Z, Heinen D, Balbas O, Garcıa E, Stoikow Z (1988)
Unique HLA–DR/DQ association revealed by family studies in
Warao Amerindians. Haplotype and homozigosity frequencies.
Hum Immunol 23:45–57
53. Layrisse Z, Guedez Y, Domınguez E, Herrera F, Soto M, Balbas
O, Matos M, Alfonzo JC, Granados J, Scorza J (1995) Extended
HLA haplotypes among the Bari Amerindians of the Perija range.
Relationship to other tribes based on four-loci haplotype fre-
quencies. Hum Immunol 44:228–235
54. Yunis JJ, Ossa H, Salazar M, Delgado MB, Deulofeut R, De la
Hoz A, Bing DH, Ramos O, Yunis EJ (1994) Major histocom-
patibility complex class II alleles and haplotypes and blood
groups of four Amerindian tribes of northern Colombia. Hum
Immunol 41:248–258
55. Titus-Trachtenberg EA, Richards O, De Stefano GF, Erlich HA
(1994) Analysis of HLA class II haplotypes in the Cayapa Indians
of Ecuador: A novel DRB1 allele reveals evidence for convergent
evolution and balancing selection at position 86. Am J Hum
Genet 55:160–167
56. Petzl-Erler ML, Gorodezky C, Layrisse Z, Klitz W, Fainboim L,
Vullo C, Bodmer JG, Egea E, Navarrete C, Infante E, Alaez C,
Olivo A, Debaz H, Bautista N, De la Rosa G, Vazquez MN,
Navarro JL, Pujol MJ, Duran C, Schafhauser C, Faucz FR, Janzen
M, Maciag P, Boldt ABW, Souza PSA, Probst CM, Da Silva GF,
Makhatadze N, Domınguez E, Montagnani S, Matos M, Martinez
A, Herrera F, Hollenbach J, Thomson G, Pando M, Satz L,
Larriba J, Fernandez G, Pesoa SA, Borosky A, Garavito G, Angel
L, Brown J, Llop E (1997) Anthropology report for region Latin-
America: Amerindian and admixture populations. In: Charron D
(ed) Genetic diversity of HLA. Functional and medical implica-
tion, vol I. EDK, Sevres, pp 337–345
57. Arnaiz-Villena A, Vargas-Alarcon G, Granados J (2000) HLA
genes in Mexican Mazatecans, the peopling of the Americas and
the uniqueness of Amerindians. Tissue Antigens 56:405–416
58. Monsalve NV, Edin G, Devine DV (1998) Analysis of HLA class
I and class II in Na-Dene and Amerindian populations from
British Columbia, Canada. Hum Immunol 59:4855
59. Munkhbat B, Sato T, Hagihara M, Sato K, Kimura A, Munk-
htuvshin N, Tsuji K (1997) Molecular analysis of HLA poly-
morphism in Khoton-Mongolians. Tissue Antigens 50:124–134
60. Grahovac B, Sukernik RI, O’hUigin C (1998) Polymorphism of
the HLA class II loci in Siberian populations. Hum Genet
102:27–43
61. Bannai M, Tokunaga K, Imanishi T, Harihara S, Fujisawa K, Juji
T, Omoto K (1996) HLA class II alleles in Ainu living in Hidaka
district, Hokkaido, Northern Japan. Am J Phys Anhropol 101:1–9
62. Leffel MS, Fallin D, Hildebrand WH, Cavett JH, Iglehart BA,
Zachery AA (2004) HLA alleles and haplotypes among the
Lakota Sioux report of the ASHI minority workshops: part III.
Hum Immunol 65:78–89
63. Degli-Esposti MA, Leaver AL, Frank T, Christiansen FT, Witt
CS, Abraham LJ, Dawkins RL (1992) Ancestral haplotypes:
conserved population MHC haplotypes. Hum Immunol
34:242–252
64. Layrisse Z, Guedez Y, Domınguez E, Paz N, Montagnani S,
Matos M, Herrera F, Ogando V, Balbas O, Rodrıguez-Larralde A
(2001) Extended HLA haplotypes in a Carib Amerindian popu-
lation: the Yucpa of the Perija Range. Hum Immunol
62:992–1000
65. Arnaiz-Villena A, Gonzalez-Alcos V, Serrano-Vela JI, Reguera
R, Barbolla L, Parga-Lozano C, Gomez-Prieto P, Abd-El-Fatah-
Khalil S, Moscoso J (2009) HLA genes in Uros from Titikaka
Lake, Peru: origin and relationship with other Amerindians and
worldwide populations. Int J Immunogenet 36:159–167
66. Arnaiz-Villena A, Siles N, Moscoso J, Zamora J, Serrano-Vela JI,
Gomez-Casado E, Castro MJ, Martinez-Laso J (2005) Origin of
Aymaras from Bolivia and their relationship with other Amer-
indians according to HLA genes. Tissue Antigens 65:379–390
67. Garcıa-Ortiz E, Sandoval-Ramırez L, Rangel-Villalobos H,
Maldonado-Torres H, Cox S, Garcıa-Sepulveda CA, Figuera LE,
Marsh SGA, Little AM, Madrigal JA, Moscoso J, Arnaiz-Villena
A, Arguello JR (2006) High-resolution molecular characteriza-
tion of the HLA class I and class II in the Tarahumara Amerin-
dian population. Tissue Antigens 68:135–146
68. Moscoso J, Seclen S, Serrano-Vela JI, Villena A, Martinez-Laso
J, Zamora J, Moreno A, IRA-Cachafeiro J, Arnaiz-Villena A
(2006) HLA genes in Lamas Peruvian–Amazonian Amerindians.
Mol Immunol 43:1881–1889
69. Geng L, Imanishi T, Tokunaga K, Zhu D, Mizuki N, Xu S, Geng
Z, Gojobori T, Tsuji K, Inoko H (1995) Determination of HLA
class II alleles by genotyping in a Manchu population in the
northern part of China and its relationship with Han and Japanese
populations. Tissue Antigens 46:111–116
70. Martinez-Laso J, Sartakova M, Allende L, Konenkov V, Moscoso
J, Silvera-Redondo C, Pacho A, Trapaga J, Gomez-Casado E,
Arnaiz-Villena A (2001) HLA molecular markers in Tuvinians: a
population with both Oriental and Caucasoid characteristics. Ann
Hum Genet 65:245–261
71. Arnaiz-Villena A, Benmamar D, Alvarez M et al (1995) HLA
allele and haplotype frequencies in Algerians. Relatedness to
Spaniards and Basques. Hum Immunol 43:259–268
72. Izaabel H, Garchon HJ, Caillat-Zucman S et al (1998) HLA class
II DNA polymorphism in a Moroccan population from the Souss,
Agadir area. Tissue Antigens 51:106–110
73. Gomez-Casado E, del Moral P, Martinez-Laso J et al (2000) HLA
genes in Arabic-speaking Moroccans: close relatedness to Ber-
bers and Iberians. Tissue Antigens 55:239–249
3700 Mol Biol Rep (2011) 38:3689–3701
123

74. Gao X, Zimmet P, Serjeantson SW (1992a) HLA-DR, DQ
sequence polymorphisms in Polynesians, Micronesians and
Javanese. Hum Immunol 34:153–161
75. Gao X, Veale A, Serjeantson SW (1992b) HLA class II diversity
in Australian aborigines: unusual HLA-DRB1 alleles. Immuno-
genetics 36:333
76. Gao X, Bhatia K, Trent RJ, Serjeantson SW (1992c) HLA-DR,
DQ nucleotide sequence polymorphisms in five Melanesian
populations. Tissue Antigens 40:31–37
77. Lester S, Cassidy S, Humphreys I, Bennett G, Hurley CK, Bo-
ettcher B, McCluskey J (1995) Evolution in HLA-DRB1 and
major histocompatibility complex class II haplotypes of Australia
aborigines. Definition of a new DRB1 allele and distribution of
DRB1 gene frequencies. Hum Immunol 42:154–160
78. Just JJ, King MC, Thomson G, Klitz W (1997) African-American
HLA class II allele and haplotype diversity. Tissue Antigens
49:547–555
79. de Pablo R, Garcia-Pacheco JM, Vilches C et al (1997) HLA
class I and class II allele distribution in the Bubi population from
the island of Bioko (Equatorial Guinea). Tissue Antigens
50:593–601
80. Vargas-Alarcon G, Granados J, Rodrıguez-Perez JM, Parga C,
Perez-Hernandez N, Rey D, Zuniga J, Arnaiz-Villena A (2010)
Distribution of HLA class II alleles and haplotypes in Mexican
Mestizo population: comparison with other populations. Immunol
Invest 39(3):268–283
Mol Biol Rep (2011) 38:3689–3701 3701
123
Copyright © 2022 FDOKUMEN




















