
High-yield purification of an organic solvent-tolerant lipase from Pseudomonas sp. strain S5
8
ANALYTICAL BIOCHEMISTRY Analytical Biochemistry 341 (2005) 267–274 www.elsevier.com/locate/yabio 0003-2697/$ - see front matter 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.ab.2005.03.006 High-yield puriWcation of an organic solvent-tolerant lipase from Pseudomonas sp. strain S5 Raja Noor Zaliha R.A. Rahman ¤ , Syarul Nataqain Baharum, Mahiran Basri, Abu Bakar Salleh Enzyme and Microbial Technology Research, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia Received 2 December 2004 Available online 22 March 2005 Abstract An organic solvent-tolerant S5 lipase was puriWed by aYnity chromatography and anion exchange chromatography. The molecu- lar mass of the lipase was estimated to be 60 kDa with 387 puriWcation fold. The optimal temperature and pH were 45 °C and 9.0, respectively. The puriWed lipase was stable at 45 °C and pH 6–9. It exhibited the highest stability in the presence of various organic solvents such as n-dodecane, 1-pentanol, and toluene. Ca 2+ and Mg 2+ stimulated lipase activity, whereas EDTA had no eVect on its activity. The S5 lipase exhibited the highest activity in the presence of palm oil as a natural oil and triolein as a synthetic triglyceride. It showed random positional speciWcity on the thin-layer chromatography. 2005 Elsevier Inc. All rights reserved. Keywords: Organic solvent-tolerant lipase; Pseudomonas sp. strain S5; PuriWcation; Characterization Lipases (E.C. 3.1.1.3) are glycerol ester hydrolases that catalyze the hydrolysis of triaclyglycerols into fatty acid, partial acylglycerols, and glycerol. The world market for industrial enzymes has been estimated at approximately US $600 million, with lipases comprising approximately US $20 million [1]. The interest of lipases stems primar- ily from their ability to preferentially hydrolyze long/ short or saturated/unsaturated fatty acyl residues, but they also exhibit a positional speciWcity for either the 1(3) or 1,2(2,3) positions of a triacylglycerol molecule. In addition, the ability of enzyme being active in the presence of organic solvents has received much attention during the past two decades [2]. In an organic solvent environment, some enzymes have enhanced thermosta- bility and there is a possibility to perform reactions that are suppressed in water [3]. Substrates and products of lipase-catalyzed mecha- nisms are often insoluble in aqueous solution, and the enzyme is usually insoluble in organic solvents. Reactions catalyzed by lipases were usually carried out in organic- aqueous two-phase media that are favorable because the separation of enzyme from substrates or products is easy. However, in general, enzymes are easily denatured and their catalytic activities disappear in the presence of organic solvents, even if they are nearly water insoluble [4]. The bacterial genus Pseudomonas secretes a number of extracellular enzymes, which include lipases, in response to Xuctuating external nutrients. Interest in Pseudomonas lip- ases stems either from their potential usefulness in a variety of biotechnological applications or from their detrimental eVect on stored food products such as refrigerated milk [5]. We previously isolated from local source a benzene, tol- uene, ethyl-benzene, and xylene (BTEX) 1 degrader identi- Wed as Pseudomonas sp. strain S5, which secreted an organic solvent-tolerant lipase [6]. The lipase was very stable in the * Corresponding author. Fax: +60 3 8943 0913. E-mail address: [email protected] (R.N.Z.R.A. Rahman). 1 Abbreviations used: BTEX, benzene, toluene, ethyl-benzene, and xylene; FFA, free fatty acid; BSA, bovine serum albumin; Con A, con- canavalin A.
Transcript of High-yield purification of an organic solvent-tolerant lipase from Pseudomonas sp. strain S5

ANALYTICALBIOCHEMISTRY
Analytical Biochemistry 341 (2005) 267–274
www.elsevier.com/locate/yabio
High-yield puriWcation of an organic solvent-tolerant lipase from Pseudomonas sp. strain S5
Raja Noor Zaliha R.A. Rahman ¤, Syarul Nataqain Baharum, Mahiran Basri, Abu Bakar Salleh
Enzyme and Microbial Technology Research, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Received 2 December 2004Available online 22 March 2005
Abstract
An organic solvent-tolerant S5 lipase was puriWed by aYnity chromatography and anion exchange chromatography. The molecu-lar mass of the lipase was estimated to be 60 kDa with 387 puriWcation fold. The optimal temperature and pH were 45 °C and 9.0,respectively. The puriWed lipase was stable at 45 °C and pH 6–9. It exhibited the highest stability in the presence of various organicsolvents such as n-dodecane, 1-pentanol, and toluene. Ca2+ and Mg2+ stimulated lipase activity, whereas EDTA had no eVect on itsactivity. The S5 lipase exhibited the highest activity in the presence of palm oil as a natural oil and triolein as a synthetic triglyceride.It showed random positional speciWcity on the thin-layer chromatography. 2005 Elsevier Inc. All rights reserved.
Keywords: Organic solvent-tolerant lipase; Pseudomonas sp. strain S5; PuriWcation; Characterization
Lipases (E.C. 3.1.1.3) are glycerol ester hydrolases that enzyme is usually insoluble in organic solvents. Reactions
catalyze the hydrolysis of triaclyglycerols into fatty acid,partial acylglycerols, and glycerol. The world market forindustrial enzymes has been estimated at approximatelyUS $600 million, with lipases comprising approximatelyUS $20 million [1]. The interest of lipases stems primar-ily from their ability to preferentially hydrolyze long/short or saturated/unsaturated fatty acyl residues, butthey also exhibit a positional speciWcity for either the1(3) or 1,2(2,3) positions of a triacylglycerol molecule.In addition, the ability of enzyme being active in thepresence of organic solvents has received much attentionduring the past two decades [2]. In an organic solventenvironment, some enzymes have enhanced thermosta-bility and there is a possibility to perform reactions thatare suppressed in water [3].
Substrates and products of lipase-catalyzed mecha-nisms are often insoluble in aqueous solution, and the
* Corresponding author. Fax: +60 3 8943 0913.E-mail address: [email protected] (R.N.Z.R.A. Rahman).
0003-2697/$ - see front matter 2005 Elsevier Inc. All rights reserved.doi:10.1016/j.ab.2005.03.006
catalyzed by lipases were usually carried out in organic-aqueous two-phase media that are favorable because theseparation of enzyme from substrates or products is easy.However, in general, enzymes are easily denatured andtheir catalytic activities disappear in the presence oforganic solvents, even if they are nearly water insoluble [4].
The bacterial genus Pseudomonas secretes a number ofextracellular enzymes, which include lipases, in response toXuctuating external nutrients. Interest in Pseudomonas lip-ases stems either from their potential usefulness in a varietyof biotechnological applications or from their detrimentaleVect on stored food products such as refrigerated milk [5].
We previously isolated from local source a benzene, tol-uene, ethyl-benzene, and xylene (BTEX)1 degrader identi-Wed as Pseudomonas sp. strain S5, which secreted an organicsolvent-tolerant lipase [6]. The lipase was very stable in the
1 Abbreviations used: BTEX, benzene, toluene, ethyl-benzene, andxylene; FFA, free fatty acid; BSA, bovine serum albumin; Con A, con-canavalin A.

268 Organic solvent-tolerant lipase from Pseudomonas / R.N.Z.R.A. Rahman et al. / Anal. Biochem. 341 (2005) 267–274
presence of various organic solvents such as n-hexane,cyclohexane, toluene, and 1-octanol. In this article, wereport the puriWcation and characterization of organic sol-vent-tolerant lipase secreted by Pseudomonas sp. strain S5.
Materials and methods
Culture condition
Pseudomonas sp. strain S5, which secreted organic sol-vent-tolerant lipase, was used in this study [6].
Cultivation
The culture was grown at 37 °C for 48 h in themedium prepared by optimizing the previously usedmedium for lipase production [6]. In the cultivation pro-cedure, 6% of inoculum was inoculated into a 250 mlblue cap bottle containing 50 ml of the medium.
Preparation of culture supernatant
The culture supernatant was prepared by removingthe cells in the culture by centrifugation at 12,000g for20 min at 4 °C.
Measurement of lipase activity
Determination of liberated free fatty acid (FFA) wasmeasured by the modiWed method of Kwon and Rhee [7].Olive oil was used as a substrate. The reaction mixture,consisting of 1.0 ml of crude enzyme, 2.5 ml olive oil emul-sion, and 0.02 ml of 20 mmol CaCl2·2H2O, was incubatedfor 30 min with shaking at 200 rpm at 37 °C. Then thereaction was stopped by adding 1.0 ml of 6 N HCl and5.0 ml isooctane. The upper layer (4 ml) was pipetted outinto a test tube, and 1.0 ml cupric acetate pyridine wasadded. The FFA dissolved in isooctane was determinedby measuring the absorbency of isooctane solution at715 nm. Lipase activity was determined by measuring theamount of FFA from the standard curves of oleic acid.One unit of lipase activity is deWned as the amount ofenzyme that liberated 1�mol FFA in 1 min at 37 °C.
Determination of protein concentrations
The protein concentrations in this study were deter-mined by the method of Bradford [8] using bovine serumalbumin (BSA, Sigma Chemical, St. Louis, MO, USA) asa standard.
Con A–Sepharose column chromatography
The enzyme solution was loaded onto the column(1.6 cm diameter, 20.0 cm height) of concanavalin A
(Con A)–Sepharose (Amersham–Pharmacia Biotech,Uppsala, Sweden) preequilibrated with 25 mM Tris–HClbuVer (pH 7.5). The column was washed with 5£ of col-umn bed volume of the equilibration buVer. The proteinsbound to the aYnity gel were eluted by increasing theconcentration of methyl �-D-glucoside from 0.1 to 0.5 Mwith a step gradient in 100 ml of 25 mM Tris–HCl buVer(pH 7.5) and 100 ml of the same buVer containing methyl�-D-glucoside. Fractions (approximately 5 ml of each)that exhibited lipase activity in the gradient region of themethyl �-D-glucoside concentration were collected. Theenzyme solution was then dialyzed overnight againstbuVer containing methyl �-D-glucoside.
DEAE–Sephacel column chromatography
The enzyme solution from the previous column wasloaded onto a column (1.6 cm diameter, 10.0 cm height)of DEAE–Sephacel (Amersham–Pharmacia Biotech)preequilibrated with 25 mM Tris–HCl buVer (pH 7.5).The column was washed with 100 ml of the equilibrationbuVer. The proteins bound to the anion exchange gelwere eluted by increasing the concentration of NaCl in25 mM Tris–HCl buVer (pH 7.5) from 0.1 to 0.5 M with alinear gradient using 50 ml of 25 mM Tris–HCl buVer(pH 7.5) and 50 ml of the same buVer containing NaCl.Fractions (approximately 2 ml of each) that exhibitedlipase activity in the gradient region of the NaCl concen-tration were collected. The enzyme solution was dialyzedovernight against buVer containing NaCl.
Determination of the purity and molecular mass of puriWed S5 lipase by SDS–PAGE
The solution containing puriWed S5 lipase andsample buVer solution was heated in a boiling waterbath for 3 min. The sample was run in a stacking gelcontaining 4.35%(w/v) polyacrylamide and 0.3%(w/v)N,N-methylene-bis(acrylamide) and in a separating gelcontaining 14.8%(w/v) polyacrylamide and 0.2%(w/v)N, N-methylene-bis(acrylamide) under the conditionsdeveloped by Laemmli [9]. The protein was stained bysilver staining.
Organic solvent stability of the enzyme
The stability of the puriWed lipase in the organic sol-vent was investigated by the method as described byOgino et al. [10] with slight modiWcation. The puriWed S5lipase-containing 25 mM Tris–HCl (pH 7.5) was Wlter-sterilized with a cellulose acetate membrane Wlter (0.2 �mpore size). Then 1 ml of an organic solvent was added to3 ml of the Wltrate in a vial bottle, and the mixture wasincubated at 37 °C with shaking at 150 rpm for 2 h. Theremaining lipase activity was measured by the methodby Kwon and Rhee [7] as described above.

Organic solvent-tolerant lipase from Pseudomonas / R.N.Z.R.A. Rahman et al. / Anal. Biochem. 341 (2005) 267–274 269
EVect of pH and pH stability of the S5 lipase
The eVect of pH on the S5 lipase activity was investi-gated by the method using olive oil as a substrate. Vari-ous buVer systems (pH 4.0–12.0) at 25 mM were added toolive oil to perform olive oil emulsion. Then the enzymewas assayed at diVerent pH values at 37 °C for 30 minwith shaking at 200 rpm. For pH stability studies, 200�lof puriWed enzyme solution was mixed with 800 �l of25 mM buVer at speciWc pH. The mixture was incubatedat 37 °C for 30 min, and the residual activity was deter-mined. BuVer systems were used at a concentration of25 mM: glycine–HCl buVer (pH 4.0–5.0), sodium hydro-gen phosphate–NaOH buVer (pH 5.0–7.0), Tris–HClbuVer (pH 7.0–9.0), disodium hydrogen orthophos-phate–NaOH buVer (pH 9.0–11.0), and glycine–NaOHbuVer (pH 10.0–12.0).
EVect of temperature on activity and thermostability of S5 lipase
The eVect of temperature on lipase activity wasstudied by carrying out the enzyme reaction at diVer-ent temperatures in the range of 30–55 °C at pH 9.0using Tris–HCl buVer (25 mM). The reaction was car-ried out for 0 and 30 min. The thermostability oflipase was tested at various temperatures, rangingfrom 37 to 45 °C, for diVerent time intervals, rangingfrom 0 to 6 h. At each time interval, 500 �l sampleswere pippeted out and immediately frozen prior tobeing assayed.
Substrate speciWcity
To determine the eVect of substrates on puriWedlipase, the olive oil emulsion was substituted with thevarious substrates and was assayed with 1.0 ml enzymesolution at 37 °C for 30 min at pH 9.0.
EVect of metal ions and oxidizing, reducing, and chelating agents
The eVect of metal ions and oxidizing, reducing, andchelating agents was investigated by incubating theenzyme in a 1:1 ratio at pH 9 for 15 and 30 min at 37 °C.The solution of metal ions and oxidizing, reducing, andchelating agents was prepared at a concentration of1 mM.
Positional speciWcity
Mixtures containing 100 mM of triolein, 9 ml phos-phate buVer (pH 7.0), and 2 ml enzyme solution wereshaken at 200 rpm at 37 °C. At 0, 3, 6, 9, and 12 h, thereaction products were extracted with 5 ml n-hexane andanalyzed by thin-layer chromatography. A silica gel
plate (Silica gel 60, Merck, Darmstadt, Germany) wasdeveloped in a mixture of chloroform and acetone (96:4).Spots of the hydrolysis products from triolein werevisualized by iodine vapor. As standards, 1(3)-monoo-lein, 2-monoolein, 1,2(2,3)-diolein, triolein, and oleic acidpurchased from Sigma were used. The commercialenzymes Lipozyme and Novozyme were included in thisexperiment as a control because the positional speciWcityhas already been veriWed.
Results and discussion
PuriWcation of S5 lipase
The objective of enzyme puriWcation is to get rid of asmuch unwanted protein as possible while retaining theenzyme activity. Lipase was puriWed with Con A–Sepharose as the initial steps, followed by DEAE–Sephacel. When the enzyme was loaded onto Con A–Sepharose column chromatography, the absorbedenzyme was eluted out at a 250 mM concentration ofmethyl �-D-glucoside. At this stage, a high percentage ofrecovery of 89% protein with 30 puriWcation fold wasobtained. The enzyme was further puriWed with anionexchange column chromatography and eluted out byNaCl treatment at a concentration of 0.3 M. A singleband was obtained for S5 lipase, which was puriWed toapproximately 387.5-fold with a speciWc activity of1240 U/mg and an overall yield of 52%.
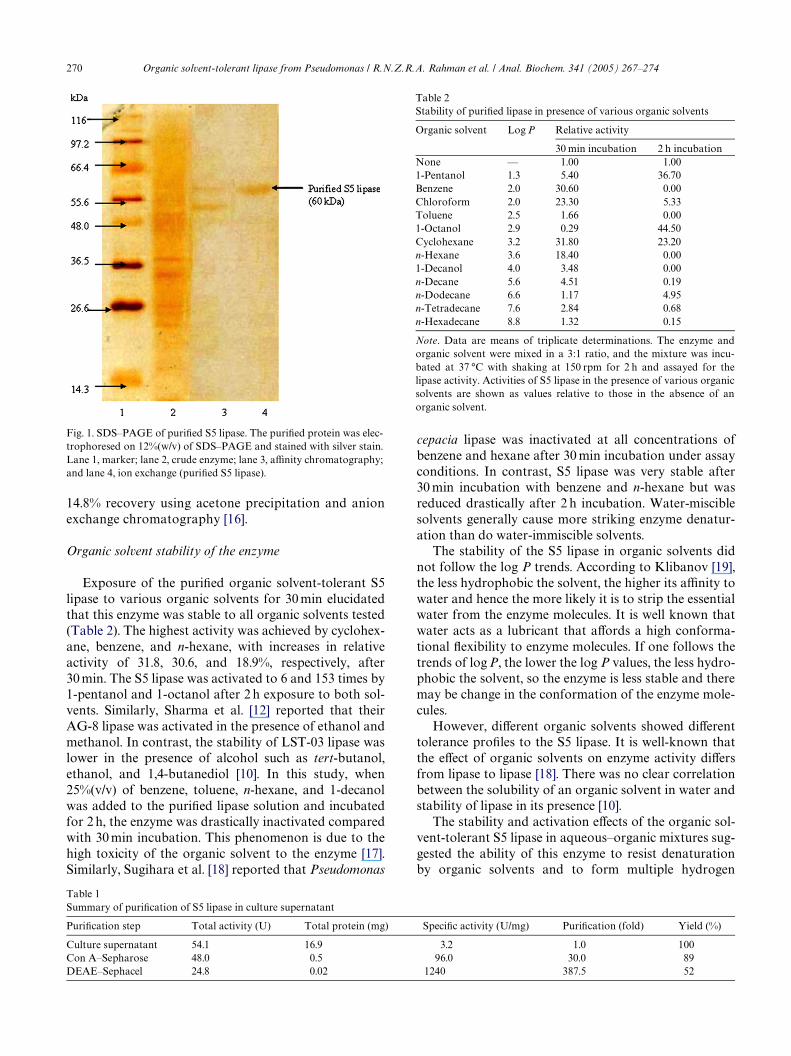
Homogeneity of the eluted peak was conWrmed bySDS–PAGE. The silver staining revealed a single proteinband with a molecular weight of approximately 60kDa(Fig. 1). The puriWcation results are summarized in Table 1.
So far, S5 lipase is the Wrst organic solvent-tolerantPseudomonas lipase to be puriWed using combinationof aYnity chromatography and anion exchange chro-matography. An organic solvent-stable lipase fromPseudomonas aeruginosa LST-03 was puriWed by ionexchange and hydrophobic interaction chromatogra-phy [10]. Other Pseudomonas lipases were puriWed byusing (NH4)2SO4 saturation as a step in partial puriW-
cation [11,12]. Lin et al. [13] puriWed an alkaline lipasefrom Pseudomonas pseudoalcaligenes F-111 by acetoneprecipitation, gel Wltration, and hydrophobic interac-tion followed by another gel Wltration. Compared withother puriWed Pseudomonas lipases, S5 lipase showed ahigher fold and recovery. According to Ogino et al.[10], LST-03 organic solvent-tolerant lipase was puri-Wed to 34.7-fold with an overall yield of 12.6%. Pseudo-monas lipase KWI-56 was puriWed to 13.9-fold byacetone precipitation and gel Wltration by HPLC with2.9% recovery [14]. Other Pseudomonas lipasesobtained 31-fold with 18% recovery and 140-fold with15% recovery [13,15]. Meanwhile, Pseudomonas mendo-cina PK-12CS lipase was puriWed to 240-fold with
270 Organic solvent-tolerant lipase from Pseudomonas / R.N.Z.R.A. Rahman et al. / Anal. Biochem. 341 (2005) 267–274
14.8% recovery using acetone precipitation and anionexchange chromatography [16].
Organic solvent stability of the enzyme
Exposure of the puriWed organic solvent-tolerant S5lipase to various organic solvents for 30 min elucidatedthat this enzyme was stable to all organic solvents tested(Table 2). The highest activity was achieved by cyclohex-ane, benzene, and n-hexane, with increases in relativeactivity of 31.8, 30.6, and 18.9%, respectively, after30 min. The S5 lipase was activated to 6 and 153 times by1-pentanol and 1-octanol after 2 h exposure to both sol-vents. Similarly, Sharma et al. [12] reported that theirAG-8 lipase was activated in the presence of ethanol andmethanol. In contrast, the stability of LST-03 lipase waslower in the presence of alcohol such as tert-butanol,ethanol, and 1,4-butanediol [10]. In this study, when25%(v/v) of benzene, toluene, n-hexane, and 1-decanolwas added to the puriWed lipase solution and incubatedfor 2 h, the enzyme was drastically inactivated comparedwith 30 min incubation. This phenomenon is due to thehigh toxicity of the organic solvent to the enzyme [17].Similarly, Sugihara et al. [18] reported that Pseudomonas
Fig. 1. SDS–PAGE of puriWed S5 lipase. The puriWed protein was elec-trophoresed on 12%(w/v) of SDS–PAGE and stained with silver stain.Lane 1, marker; lane 2, crude enzyme; lane 3, aYnity chromatography;and lane 4, ion exchange (puriWed S5 lipase).
Table 1
cepacia lipase was inactivated at all concentrations ofbenzene and hexane after 30 min incubation under assayconditions. In contrast, S5 lipase was very stable after30 min incubation with benzene and n-hexane but wasreduced drastically after 2 h incubation. Water-misciblesolvents generally cause more striking enzyme denatur-ation than do water-immiscible solvents.
The stability of the S5 lipase in organic solvents didnot follow the log P trends. According to Klibanov [19],the less hydrophobic the solvent, the higher its aYnity towater and hence the more likely it is to strip the essentialwater from the enzyme molecules. It is well known thatwater acts as a lubricant that aVords a high conforma-tional Xexibility to enzyme molecules. If one follows thetrends of log P, the lower the log P values, the less hydro-phobic the solvent, so the enzyme is less stable and theremay be change in the conformation of the enzyme mole-cules.
However, diVerent organic solvents showed diVerenttolerance proWles to the S5 lipase. It is well-known thatthe eVect of organic solvents on enzyme activity diVersfrom lipase to lipase [18]. There was no clear correlationbetween the solubility of an organic solvent in water andstability of lipase in its presence [10].
The stability and activation eVects of the organic sol-vent-tolerant S5 lipase in aqueous–organic mixtures sug-gested the ability of this enzyme to resist denaturationby organic solvents and to form multiple hydrogen
Table 2Stability of puriWed lipase in presence of various organic solvents
Note. Data are means of triplicate determinations. The enzyme andorganic solvent were mixed in a 3:1 ratio, and the mixture was incu-bated at 37 °C with shaking at 150 rpm for 2 h and assayed for thelipase activity. Activities of S5 lipase in the presence of various organicsolvents are shown as values relative to those in the absence of anorganic solvent.
Organic solvent Log P Relative activity
30 min incubation 2 h incubationNone — 1.00 1.001-Pentanol 1.3 5.40 36.70Benzene 2.0 30.60 0.00Chloroform 2.0 23.30 5.33Toluene 2.5 1.66 0.001-Octanol 2.9 0.29 44.50Cyclohexane 3.2 31.80 23.20n-Hexane 3.6 18.40 0.001-Decanol 4.0 3.48 0.00n-Decane 5.6 4.51 0.19n-Dodecane 6.6 1.17 4.95n-Tetradecane 7.6 2.84 0.68n-Hexadecane 8.8 1.32 0.15
Summary of puriWcation of S5 lipase in culture supernatant
PuriWcation step Total activity (U) Total protein (mg) SpeciWc activity (U/mg) PuriWcation (fold) Yield (%)
Culture supernatant 54.1 16.9 3.2 1.0 100Con A–Sepharose 48.0 0.5 96.0 30.0 89DEAE–Sephacel 24.8 0.02 1240 387.5 52

Organic solvent-tolerant lipase from Pseudomonas / R.N.Z.R.A. Rahman et al. / Anal. Biochem. 341 (2005) 267–274 271
bonds with water for structural Xexibility and conforma-tional mobility for optimal catalysis [20]. Therefore, theS5 lipase is naturally stable in organic solvent and holdsthe potential for use in organic synthesis and relatedapplications [21].
EVect of pH on activity and stability of the S5 lipase
The optimal pH for lipase activity was 9.0 (Fig. 2).The activity was reduced drastically at pH below 6.0 andlost 80% of the maximal activity at pH 10.0. Similarly,Gilbert et al. [15] reported that EF2 lipase had maximalactivity at pH 8.5–9.0. Other Pseudomonas lipases, suchas LST-03 [10] and F-111 [13], have their maximal activi-ties at pH values ranging from 6.0 to 10.0. On the otherhand, lipases from P. cepacia and Pseudomonas sp.KWI-56 showed optimal pH values at 5.5–6.5 and 5.5–7.0, respectively [14,18]. In a pH stability test, the S5lipase showed good stability after 30 min at pH 7.0–9.0;however, the activity was reduced drastically at pH 10.0–12.0 (Fig. 3). The enzyme retained 86 and 64% activity inpH 8.0 and 7.0, respectively, and the enzyme lost 80%activity in pH 6.0. The enzyme was also inhibited byacidic pH. In comparison, Jinwal et al. [16] reported thatPseudomonas lipase PK-12CS was very stable at a broadpH range (5.6–9.0) for 14 h at 37 °C. AG-8 lipase stillretained 90–100% activity at pH 7.0–10.0 for 24 h at25 °C [12]. LST-03 lipase was very stable at a pH rangeof 5.0–8.0 for 10 min at 30 °C [10].
EVect of temperature on activity and thermostability of S5 lipase
The lipase from Pseudomonas sp. S5 was consideredto be thermostable, as indicated by the optimal tempera-ture of the activity of pure lipase, that is, 45 °C (Fig. 4).The enzyme retained 86 and 52% of its maximum activ-ity at 37 and 50 °C, respectively. Similarly, Sharma et al.
Fig. 2. EVect of pH on lipase activity. Various buVer systems (pH 4.0–12.0) at 25 mM were added to olive oil as a substrate to perform oliveoil emulsion. Then the enzyme was assayed at diVerent pH values at37 °C for 30 min with shaking at 200 rpm. Note. The buVer systemsused were as follows: 25 mM glycine–HCl (�), sodium hydrogen phos-phate–NaOH (�), Tris–HCl (�), disodium hydrogen orthophos-phate–NaOH (�), and glycine–NaOH (�).
[12] reported that their Pseudomonas sp. AG-8 lipase hadoptimal activity at 45 °C. Organic solvent-stable LST-03lipase had maximal activity at 37 °C [10]. On the otherhand, Lin et al. [13] reported that P. pseudoalcaligenesF-111 lipase had an optimal temperature of 40 °C. Thethermal stability proWle of the lipase is shown in Fig. 5.The enzyme was stable below 45 °C for 30 min. Itretained approximately 80% of its initial activity at 50 °Cfor 30 min. The half-lives of the enzyme at 45 and 50 °Cwere 2 and 1 h, respectively. The enzyme lost all of theoriginal activity at 55 °C after 2 h incubation. In contrast,organic solvent-stable LST-03 lipase [10] showed a lowerstability (below 40 °C for 10 min) than did S5 lipase.
Substrate speciWcity
To assess substrate speciWcity, activity was assayedunder standard conditions. The lipase activity of Pseu-domonas sp. S5 against various triacylglycerols and nat-ural oils was investigated. The activities of the enzyme
Fig. 3. EVect of pH on lipase stability. PuriWed enzyme solution(200 �l) was mixed with 800 �l of 25 mM buVer at speciWc pH. Themixture was incubated at 37 °C for 30 min, and the residual activitywas determined. Note. The buVer systems used were as follows: 25 mMglycine–HCl (�), sodium hydrogen phosphate–NaOH (�), Tris–HCl(�), disodium hydrogen orthophosphate–NaOH (�), and glycine–NaOH (�).
Fig. 4. Temperature proWle of puriWed S5 lipase, The eVect of tempera-ture on lipase activity was studied by carrying out the enzyme reactionat diVerent temperatures in the range of 30–55 °C at pH 9.0 using Tris–HCl buVer (25 mM). The reaction was carried out for 0 and 30 min.

272 Organic solvent-tolerant lipase from Pseudomonas / R.N.Z.R.A. Rahman et al. / Anal. Biochem. 341 (2005) 267–274
against substrate are shown relative to the olive oil inTable 3. S5 lipase was found to have the highest activityagainst palm oil, followed by coconut oil (125%).According to Ogino et al. [10], enzyme had the highestactivity against coconut oil because the principal fattyacid component was lauric acid (12:0), which was morehydrolyzable by the lipase than was oleic acid (18:1), theprincipal fatty acid component of olive oil. Similarly,the puriWed lipases eYciently hydrolyzed various oilsand fats such as palm oil, soybean oil, and coconut oil[14,16].
In the oleochemical industry, enzymatic hydrolysis ofoils and fats is an energy-saving process compared withconventional high-temperature and high-pressure pro-cesses, and unsaturated fatty acids can be produced with-out oxidation [14]. Therefore, S5 lipase with the aboveproperties presents a suitable candidate for this purpose.
Fig. 5. Thermal stability of puriWed lipase. The enzyme solution wasincubated at 30 °C (�), 37 °C (�), 45 °C (�), 50 °C (�), and 55 °C (�)for 6 h. The incubated enzyme was assayed at diVerent time intervalsto check for the residual activity.
Table 3Hydrolytic activities of S5 lipase against various natural oils and tria-cylglycerols
Note. Data are means of triplicate determinations. The olive oil emul-sion was substituted with the various substrates and assayed with1.0 ml enzyme solution at 37 °C for 30 min at pH 9.0.
a The activities of the enzyme against substrates are shown as valuesrelative to those with olive oil.
Natural oil/triglycerol Relative activitya (%)
Olive oil (18:1) 100Coconut oil (12:0) 125Palm oil (16:0) 322Groundnut oil (18:1) 42Sesame oil (18:2) 50Soybean oil (18:3) 56Triacetin (C2) 50Tributyrin (C4) 58Tricaproin (C6) 65Tricaprylin (C8) 66Tricaprin (C10) 96Trilaurin (C12) 122Trimyristin (C14) 186Tripalmitin (C16) 217Triolein (C18) 247
S5 lipase was found to have the highest activityagainst triolein, which possessed longer carbon chainlengths. This result proved, in principle, that the lipase ispreferred to hydrolyze longer carbon chains of thesubstrates. In contrast, Ogino et al. [10] reported thatLST-03 lipase had the highest hydrolytic activity againsttricaproin (C6).
EVect of metal ions and oxidizing, reducing, and chelating agents
According to Voet et al. [22], nearly one-third of allknown enzymes require the presence of metal ions forcatalytic activity. This group of enzymes includes themetal enzymes, which contain tightly bound metal ioncofactors, most commonly transition metal ions such asFe2+, Fe3+, Cu2+, Mn2+, and Zn2+. Metal-activatedenzymes, in contrast, loosely bind metal ion from solu-tion, usually the alkali and alkaline earth metal ionsNa+, K+ Mg2+, and Ca2+.
The eVect of metal ions and oxidizing, reducing, andchelating agents was tested at 1 mM in 25 mM Tris–HClbuVer at pH 9.0 (Table 4). Lipase was completely inhib-ited by Fe3+, Ba2+, K+, Zn2+, and Cu2+, known as transi-tion metal ions, after 30 min incubation. Alkaline earthmetal ions, Mg2+ and Ca2+, were found to stimulate thelipase activity at 15 min incubation. However, prolongedincubation (30 min) led to decreases in enzyme activityof 52 and 95% for Mg2+ and Ca2+, respectively. This sug-gested that S5 lipase is a metal-activated enzyme. In thisgroup of enzymes, the ions often play a structural rolerather than a catalytic one. The ions bind to the enzymeand change the conformation of the protein to countergreater stability to the enzyme. But transition metal ionschange the conWrmation of the protein to less stable dueto ion toxicity.
Table 4EVect of various reagents on lipase activity
Note. Data are means of triplicate determinations. S5 lipase was incu-bated with diVerent reagents in a 1:1 ratio at pH 9.0 for 15 and 30 minat 37 °C before the residual activity was measured.
Reagent (1 mM) Relative activity (%)
15 min incubation 30 min incubation
No addition 100 100NaCl 48 53MgCl2 161 52CaCl2 114 95KCl 52 36BaCl2 45 25FeCl3 23 20CuCl2 32 25ZnSO4 23 20Ammonium persulfate 50 48KI 18 12Sodium citrate 19 15EDTA 100 992-Mercaptoethanol 25 15

Organic solvent-tolerant lipase from Pseudomonas / R.N.Z.R.A. Rahman et al. / Anal. Biochem. 341 (2005) 267–274 273
Similar results were reported by Sharma et al. [12]with their lipase from Pseudomonas sp. AG-8. Theydescribed that Ca2+ ions activated the enzyme, whereasFe3+ and Zn2+ strongly inhibited its activity. A possibleexplanation of this phenomenon is that Ca2+ has a spe-cial enzyme-activating eVect that it exerts by concentrat-ing at the fat–water interface. Therefore, calcium ionsmay carry out three distinct roles in lipase action:removal of fatty acids as insoluble Ca2+ salts in certaincases, direct enzyme activation resulting from concentra-tion at the fat–water interface, and stabilizing eVect onthe enzyme. Fe3+ and Zn2+, and especially Cu2+, werehighly toxic to lipase even at diluted solution [23].
The activity of the lipase from Pseudomonas sp. S5decreased with both oxidizing and reducing agents.Among the reducing agents, the enzyme retained 48%activity after 30 min exposure with ammonium persul-fate, whereas it retained 12% of the maximal activityafter 30 min exposure with potassium iodide. When thereducing agent 2-mercaptoethanol was added at a con-centration of 1 mM, enzyme activity dropped by 25 and15% after 15 and 30 min, respectively.
The enzyme activity was not aVected by 1 mM EDTA,indicating that the lipase from Pseudomonas sp. S5 wasnot a metalloenzyme. Lin et al. [13] also showed thatactivity of lipase produced from P. pseudoalcaligenesF-111 was not aVected by 1 mM EDTA.
Positional speciWcity against triacylglycerols
According to Farias et al. [24], lipase can be classiWedin terms of speciWcity toward the position of the acylgroup of triglycerides into two groups: 1,3-speciWc andnonspeciWc or random. The results of the thin-layerchromatography analysis of the hydrolysis products of
triolein are shown in Fig. 6. The enzyme cleaved not onlythe 1,3-positioned ester bonds but also the 2-positionedester bond of triolein. Thus, S5 lipase could hydrolyzethe ester bonds of triolein nonspeciWcally. Similarly,LST-03 organic solvent-tolerant lipase also hydrolyzedester bonds nonspeciWcally [10]. Similar Wndings werereported by Sigiura et al. [25] with their PseudomonasXuorescens lipase that cleaved ester bond nonspeciWcally.Lipozyme and Novozyme were included in this experi-ment as a control because the 1,3-positional speciWcityand nonspeciWc position, respectively, have already beenveriWed.
According to Gao et al. [26], Pseudomonas sp. strain2106 lipase shows 1,3-positional speciWcity. Sugihara etal. [18] reported that P. cepacia lipase cleaved all of theester bonds of triolein, with some preference for the 1,3-ester bonds. The same Wndings were reported by Men-cher and Alford [27] with their Pseudomonas fragi lipasethat hydrolyzed 1,3-ester bonds.
References
[1] M.V. Arbridge, W.H. Pitcher, Industrial enzymology: a looktowards the future, Trends Biotechnol. 7 (1989) 330–335.
[2] G.A. Sellek, J.B. Chaudhuri, Biocatalysis in organic media usingenzymes from extramophiles, Enzyme Microbiol. Technol. 25(1999) 471–482.
[3] B.C. Koops, H.M. Verheij, A.J. Slotboom, M.R. Egmond, EVect ofchemical modiWcation on the activity of lipases in organic sol-vents, Enzyme Microbiol. Technol. 25 (1999) 622–631.
[4] H. Ogino, K. Miyamoto, M. Yasuda, K. Ishimi, H. Ishikawa, Growthof organic solvent-tolerant Pseudomonas aeruginosa LST-03 in thepresence of various organic solvents and the production of lipolyticenzyme in the presence of cyclohexane, Biochem. Eng. 4 (1999) 1–6.
[5] R. Stead, Microbial lipases: their characteristics, role in food spoil-age, and industrial uses, J. Dairy Res. 53 (1986) 481–505.
Fig. 6. Thin-layer chromatogram of the hydrolysis products of triolein catalyzed by S5 lipase. Mixtures containing 100 mM of triolein, 9 ml phos-phate buVer (pH 7.0), and 2 ml enzyme solution were shaken at 200 rpm at 37 °C. At 0, 3, 6, 9, and 12 h, the reaction products were extracted with 5 mlof n-hexane and analyzed by thin-layer chromatography. Lane 1, triolein; lane 2, 1(3) monoolein; lane 3, 1(2) diolein; lane 4, 1(3) diolein; lane 5, oleicacid; lane 6, 0 h; lane 7, 3 h; lane 8, 6 h; lane 9, 9 h; lane 10, 12 h; lane 11, Lipozyme; lane 12, Novozyme; and lane 13, marker.

274 Organic solvent-tolerant lipase from Pseudomonas / R.N.Z.R.A. Rahman et al. / Anal. Biochem. 341 (2005) 267–274
[6] S.N. Baharum, A.B. Salleh, C.N.A. Razak, M. Basri, M.B.A. Rah-man, R.N.Z.R.A. Rahman, Organic solvent tolerant lipase byPseudomonas sp. strain S5: stability of enzyme in organic solventand physical factors aVecting its production, Ann. Microbiol. 53(2003) 75–83.
[7] D.Y. Kwon, J.S. Rhee, A simple and rapid colorimetric methodfor determination of free fatty acid for lipase assay, J. Am. OilChem. Soc. 63 (1986) 69–92.
[8] M.M. Bradford, A rapid and sensitive method for the quantitationof microgram quantities of protein using the principle of protein–dye binding, Anal. Biochem. 72 (1976) 248–254.
[9] U.K. Laemmli, Cleavage of structure proteins during assembly ofthe head of bacteriophage T4, Nature 277 (1970) 680–685.
[10] H. Ogino, S. Nakagawa, K. Shinya, T. Muto, N. Fujimura, M.Yasudo, H. Ishikawa, PuriWcation and characterization of organicsolvent tolerant lipase from organic solvent tolerant Pseudomonasaeruginosa LST-03, J. Biosci. Bioeng. 89 (2000) 451–457.
[11] C. Schuepp, S. Kermasha, M.C. Michalski, A. Marin, Production,partial puriWcation, and characterization of lipase from Pseudo-monas fragi CRDA 037, Proc. Biochem. 32 (1997) 225–232.
[12] A.K. Sharma, R.P. Tiwari, G.S. Hoondal, Properties of a thermo-stable and solvent stable extracellular lipase from a Pseudomonassp. AG-8, J. Basic Microbiol. 41 (2001) 363–366.
[13] S.F. Lin, C.M. Chiou, C.M. Yeh, Y.C. Tsai, PuriWcation and par-tial characterization of an alkaline lipase from Pseudomonaspseudoalcaligenes F-111, Appl. Environ. Microbiol. 62 (1996)1093–1095.
[14] T. Iizumi, K. Nakamura, T. Fukase, PuriWcation and characteriza-tion of a thermostable lipase from newly isolated Pseudomonas sp.KWI-56, Agric. Biol. Chem. 54 (1990) 1253–1258.
[15] E.J. Gilbert, A. Cornish, C.W. Jones, PuriWcation and properties ofextracellular lipase from Pseudomonas aeruginosa EF2, J. Gen.Microbiol. 137 (1991) 2223–2229.
[16] U.K. Jinwal, U. Roy, A.R. Chowdhury, A.P. Bhaduri, P.K. Roy,PuriWcation and characterization of an alkaline lipase from anewly isolated Pseudomonas mendocina PK-12CS and chemoselec-tive hydrolysis of fatty acid ester, Bioorg. Med. Chem. 11 (2003)1041–1046.
[17] J. Sikkema, D.J.A.M. Bont, B. Poolman, Intercalations of cyclichydrocarbons with biological membranes, J. Biol. Chem. 269(1994) 8022–8028.
[18] A. Sugihara, M. Ueshima, Y. Shimada, S. Tsunasawa, Y. Tomi-naga, PuriWcation and characterization of a novel thermostablelipase from Pseudomonas cepacia, J. Biochem. 112 (1992) 598–603.
[19] A.M. Klibanov, Enzymes that work in organic solvents, Chem-Tech. (1986) 354–359.
[20] A.M. Klibanov, Improving enzymes by using them in organic sol-vents, Nature 409 (2001) 241–246.
[21] R.P. Yadav, P. Agarwal, S.N. Upadhay, Microbial lipases: toolsfor drug discovery, J. Sci. Ind. Res. 59 (2000) 977–987.
[22] D. Voet, J.G. Voet, C.W. Pratt, Fundamentals of Biochemistry,John Wiley, New York, 1999.
[23] E.D. Wills, The relation of metals and –SH groups to the activityof pancreatic lipase, Biochim. Biophys. Acta 40 (1960) 481–490.
[24] R.N. Farias, M. Torres, R. Canela, Spectrophotometric determi-nation of the positional speciWcity of nonspeciWc and 1,3-speciWclipases, Anal. Biochem. 252 (1997) 186–189.
[25] M. Sigiura, M. Isobe, F. Hirata, T. Inigawa, Application of lipo-protein lipase for the assay of serum triglyceride, Chem. Pharm.Bull. (Tokyo) 23 (1975) 1417–1420.
[26] X.G. Gao, S.G. Cao, K.C. Zhang, Production, properties andapplication to nonaqueous enzymatic catalysis of lipase fromnewly isolated Pseudomonas strain, Enzyme Microbiol. Technol.27 (2000) 74–82.
[27] J.R. Mencher, J.A. Alford, PuriWcation and characterization of thelipase of Pseudomonas fragi, J. Gen. Microbiol. 48 (1967) 317–328.




















