
Characterization of a thermostable lipase showing loss of secondary structure at ambient temperature

ORIGINAL PAPER
A unique thermostable and organic solvent tolerant lipasefrom newly isolated Aneurinibacillus thermoaerophilusstrain HZ: physical factor studies
Malihe Masomian • Raja Noor Zaliha Raja Abd Rahman •
Abu Bakar Salleh • Mahiran Basri
Received: 26 April 2008 / Accepted: 9 February 2010
� Springer Science+Business Media B.V. 2010
Abstract A newly isolated thermophilic bacterium,
Aneurinibacillus thermoaerophilus strain HZ, from a hot
spring recreational area (Sungai Kelah, Malaysia), showed
an extracellular lipase activity. It was identified based on
16S rRNA sequencing, where phylogenetic analysis
revealed its homology to Aneurinibacillus thermoaerophi-
lus. The strain produced a lipase that was stable in various
organic solvents such as dimethyl sulfoxide, toluene,
p-xylene, and hexane. In order to increase lipase produc-
tion, optimization of physical factors which affected the
growth and lipase production was studied. The optimal
growth was obtained at 50�C and pH 8.0; while the max-
imal lipase production was achieved in the logarithmic
decline phase at 60�C and pH 7.5 with 7% starting inoc-
ulum and 150 rev/min shaking rate for 48 h incubation.
Keywords Aneurinibacillus thermoaerophilus
strain HZ � Thermostable lipase �Optimization of physical factors � Organic solvent tolerant
Introduction
Lipases (triacylglycerol acyl hydrolases (EC 3.1.1.3) are
lipolytic enzymes that catalyse both the hydrolysis and the
synthesis of esters at an oil–water interface (Haba et al.
2000). Many microorganisms, including plants, animals,
bacteria, yeast, and fungi, have been determined to secrete
lipases during their growth in hydrophobic substrates
(Holker et al. 2004). However, only microbial lipases are
commercially significant for their potential use in indus-
tries because of their unique catalytic performance and
versatility of catalyse numerous different reactions. They
are widely diversified in their enzymatic properties and
substrate specificities (Gupta et al. 2004). Lipases are the
choice of biocatalysts as they show unique chemo-, regio-,
enantioselectivites, which enable the production of novel
drugs, agrochemicals and fine products (Gotor-Fernandez
et al. 2006; kamini et al. 2000; Saxena et al. 2003). Many
lipases, due to their ability to perform both hydrolytic and
synthetic reactions, find immense applications in industries
like foods, detergents, pharmaceuticals, leather, textile and
dairy (Saxena et al. 2003; Hasan et al. 2006; Burkert et al.
2004).
Each application requires unique properties with respect
to specificity, stability, temperature, and pH dependence or
ability to catalyse synthetic ester reactions in organic sol-
vents. Meanwhile, thermostable enzymes are particularly
attractive for industrial applications because of their high
activities at the elevated temperatures and stabilities in
organic solvents (Niehaus et al. 1999; Pennisi 1997).
Substrates and products of lipase are often insoluble in
aqueous solutions, and the enzyme is usually insoluble in
organic solvents. Some reactions catalysed by lipase can be
carried out in organic aqueous two-phase media because the
separation of enzyme from substrates or products becomes
M. Masomian � R. N. Z. R. A. Rahman (&)
Enzyme and Microbial Technology Research, Department
of Microbiology, Faculty of Biotechnology and Biomolecular
Science, Universiti Putra Malaysia, 43400 Serdang, Selangor,
Malaysia
e-mail: [email protected]
A. B. Salleh
Enzyme and Microbial Technology Research, Department
of Biochemistry, Faculty of Biotechnology and Biomolecular
Science, Universiti Putra Malaysia, 43400 Serdang, Selangor,
Malaysia
M. Basri
Enzyme and Microbial Technology Research, Department
of Chemistry, Faculty of Science, Universiti Putra Malaysia,
43400 Serdang, Selangor, Malaysia
123
World J Microbiol Biotechnol
DOI 10.1007/s11274-010-0347-1

easy. However, in general, enzymes are easily inactivated in
the presence of organic solvents, even if they are almost
water-insoluble (Ogino et al. 1999). Accordingly, organic
solvent–stable microbial lipases which are of enormous
biotechnological potential possess the following character-
istics: (1) require no cofactors, (2) remain active in organic
solvents, (3) react on a broad substrate specificity and
(4) exhibit a high enantioselectivity (Bornscheuer 2002).
Pursuant to the above mentioned statements, screening
of thermophilic microorganisms with lipolytic activities
can facilitate the discovery of novel lipases for industrial
purposes that are stable and function optimally at extreme
temperature. Therefore, we conducted an extensive
screening and isolated a new thermophilic lipase-producing
organism, identified as Aneurinibacillus thermoaerophilus
strain HZ that is both organic solvent-tolerant and extre-
mely thermophilic. Here, we report the isolation and
physical optimization of the culture to produce high yield
of thermostable and organic solvent-tolerant lipase.
Materials and methods
Sample collection and enrichment culture
Samples were collected from hot spring in Sungai Kelah (up
to 100�C), Malaysia. The enrichment of samples was per-
formed at 55�C with shaking (150 rev/min) in a medium
containing peptone (0.5% w/v), yeast extract (0.5% w/v),
NaCl (0.05% w/v), CaCl2 (0.005% w/v), olive oil (1.0% v/v),
pH 8.5 and M2 (tryptone (3.0% w/v), yeast extract (1.0%
w/v), NaCl (0.05% w/v), glucose (0.5% w/v), pH 7.5 for
72 h.
Screening and isolation of lipolytic thermophile
bacteria
Screening for lipase-producing isolates was done qualita-
tively on tributyrin agar plates. Isolates that produced
lipolytic enzyme formed clearing zones around the colo-
nies on the plates. These positive isolates were then tested
for their lipase production on Rhodamine B agar plates and
triolein agar plates at 55�C for 24–48 h. Lipase production
on Rhodamine B agar plate was monitored with UV light at
350 nm. Orange fluorescent halos were shown around the
colonies of lipase producing strains and lipase producers
produced blue zone on triolein agar.
Production medium and growth conditions
A single colony of each lipase producer from a nutrient
agar slant was inoculated in 10 ml tryptone soy broth
medium and incubated at 55�C overnight in water bath
shaker. Two ml were inoculated in 50 ml of production
medium containing yeast extract (0.75% w/v), peptone
(0.75% w/v), olive oil (1.0% v/v), gum arabic (1.0% w/v),
NaCl (0.25% w/v), CaCl2.2H2O (0.001% w/v), pH 7.5 for
72 h at 55�C with shaking.
Lipase assay
Lipase activity was measured by a modified method of
Kwon and Rhee (1986). Ten ml of the culture were taken
out and centrifuged at 10,000g for 10 min at 4�C to sedi-
ment bacteria cell. The supernatant was assayed for lipase
activity. The reaction mixture consisting of 1.0 ml of crude
enzyme, 2.5 ml olive oil emulsion (1:1 ratio of olive oil
and phosphate buffer [K2HPO4 (50 mM), at pH 7.0)] and
0.02 ml of 20 mM CaCl2�2H2O, was incubated for 30 min
with shaking at 200 rev/min at 55�C. Then the reaction was
stopped by adding 1.0 ml of 6 M HCl and 5.0 ml isooc-
tane. The upper layer (4 ml) was pipetted out into a test
tube, and 1.0 ml cupric acetate pyridine [Kupfer(II)-Acetat
monohydrate (5%; pH 6.1), pH adjusted by adding pyri-
dine] was added. The free fatty acids dissolved in isooctane
were determined by measuring the absorbance of isooctane
solution at 715 nm. Each sample was assayed three times
and the average value was taken. One unit (U) of lipase
activity is defined as the rate of fatty acid formation per
min (standard assay condition). Standard deviation was
determined by Microsoft Excel program.
Identification of the bacteria
Aneurinibacillus thermoaerophilus strain HZ was identi-
fied via 16S rDNA sequencing. The genomic DNA was
extracted using DNeasy Tissue Kit (Qiagen, Germany)
according to the manufacture’s instructions and was used
as a template to perform Polymerase Chain Reaction (PCR)
amplification for 16S rDNA identification. The universal
primers were used: 16S-F (50-GAG TTT GAT CCT GGC
TCA G-30) and 16S-R (50-CGG CTA CCT TGT TAC GAC
TT-30). The reaction mixture was initially heated for 4 min
at 94�C, followed by 30 cycles PCR of 94�C 1 min, 58�C
1 min and 72�C 1 min; then, one cycle of 7 min at 72�C
and held at 4�C. The primers amplified a 1,502 bp PCR
product. The amplified products were examined by elec-
trophoresis and extracted by using QIAquick Gel Extrac-
tion Kit (Qiagen, Germany) according to the manufacture’s
instructions. Sequencing of purified product was down by
First Base Laboratories Sdn Bhd (Shah Alam, Selangor,
Malaysia), using BigDye� Terminator v3.1 cycle
sequencing kit chemistry. The complete sequence was
submitted to GenBank at NCBI. DNA homology search
in the GenBank database (http://www.ncbi.nih.gov) was
World J Microbiol Biotechnol
123

performed. A phylogenetic tree was constructed based on
comparison of the 16S rDNA sequence of the isolate with
other strains of Aneurinibacillus spp. All sequences were
aligned with CLUSTALW 1.75 from Biology WorkBench
database at website (http://workbench.sdsc.edu; Thompson
et al. 1994). The tree was constructed in GeneBee service
which involves bootstrap analysis (http://www.genebee.
msn.su). 16S rDNA sequences of other Aneurinibacillus
spp. were obtained from the GenBank database (http://
www.ncbi.nih.gov).
Optimal temperature
The effect of temperature of the crude lipase was investi-
gated by assaying at temperatures ranging from 45 to 70�C.
Lipase activity was determined using olive oil as substrate.
Effect of physical factors on bacterial growth and lipase
production
To investigate the influence of physical factors on the
growth and extracellular lipase production by Aneuriniba-
cillus thermoaerophilus strain HZ, an overnight bacterial
inoculum (5%; OD600 = 0.5) was inoculated into basal
medium and incubated for 48 h. Five parameters (medium
volume, pH, temperature, inoculum size and agitation)
were studied for their influences in lipase production and
bacterial growth. Lipase assay was performed after 48 h of
incubation by modified method of Kwon and Rhee (1986)
as described previously. The bacterial growth was esti-
mated by the number of c.f.u. on the nutrient agar plates.
Effect of medium volume
The effect of the medium volume was investigated by
cultivating the bacterium on different volumes of medium
50, 100, 150 and 200 ml in 500-ml blue cap bottles.
Effect of temperature
The ability of A. thermoaerophilus strain HZ to grow and
produce lipase at elevated temperatures (50–70�C) was
studied. Separate cultures were incubated at 50, 55, 60, 65
and 70�C for 48 h with agitation at 150 rev/min.
Effect of pH
The effect of pH was studied by adjusting the media (with
1.0 M of NaOH or HCl) to different pH values from 4 to 9
at 0.5 unit intervals. The media were autoclaved, cooled
and inoculated with an overnight culture of A. thermo-
aerophilus strain HZ.
Effect of inoculum size
The effect of inoculum size (OD600 = 0.5) was investi-
gated by using different inoculum sizes ranging from 1 to
9% at interval of 2% (v/v).
Effect of agitation
The effect of agitation was carried out by cultivating the
bacterium under different agitation rates, 0, 50, 100, 150,
and 200 rev/min.
Organic solvent stability of crude enzyme
A. thermoaerophilus strain HZ was cultured aerobically in
the absence of organic solvents at 55�C for 48 h. The
culture medium was centrifuged at 10,000g for 10 min at
4�C and the supernatant was used as the crude enzyme.
Various organic solvents were tested at the concentration
of 25% (v/v) by addition of 1 ml of organic solvent to 3 ml
of crude enzyme. The reaction mixture was preincubated
for 30 min at 37�C under shaking condition (150 rev/min)
to insure the continuous mixing of the enzyme and solvent.
The enzyme stability was expressed as the remaining
activity assayed according to the method of Kwon and
Rhee (1986) relative to the control value. For control,
phosphate buffer pH 7.0 was added instead of solvent. The
organic solvents used were dimethyl sulfoxide (-1.3),
benzene (2.0), toluene (2.5), 1-octanol (2.9), hexane (3.6),
dodecanol (5.0), decane (5.6), 1-dodecene (6.6), n-te-
tradecane (7.6), hexadecane (8.8).
Statistical analysis
For statistical analysis, a standard deviation for each
experimental result was calculated using the Excel Spread-
sheet available in the Microsoft Excel.
Nucleotide sequence accession number
The nucleotide sequence determined in this study has been
deposited in the Gene Bank database (http://www.
ncbi.nih.gov) under accession number DQ890194.
Results and discussion
Bacterial strain identification and phylogenetic tree
analysis
The 16S rRNA (*1,500 bp) contained several regions of
highly conserved sequence useful for obtaining proper
sequence alignments and on the other hand contained
World J Microbiol Biotechnol
123

sufficient sequence variability in other regions of the
molecule to serve as excellent chronometers. The differ-
ences of sequence in the hypervariable regions reflect the
strain variations. Comparisons of the sequences between
different species suggested a relatively earlier or later time
in which they shared a common ancestor (Woese 1998).
The polymerase chain reaction (PCR) technique was
used to amplify the rRNA gene using universal primers and
it amplified the whole region of the rRNA gene which is
1,502 bp. The amplified product was examined by elec-
trophoresis and then was purified and sequenced (Fig. 1).
Comparison of the 16S rRNA gene of the new isolate
with the GenBank database (http://www.ncbi.nih.gov)
showed that it is closely related to A. thermoaerophilus
strain L420-91, DSM 10154 (X94196) and A. thermo-
aerophilus strain: DSM 10154T (AB112726). The phylo-
genetic tree analysis of new isolate was compared with
other Bacillus spp. sequences and it showed high homology
with A. thermoaerophilus strains such as L420-91, DSM
10154 (X94196), DSM 10154T (AB112726), ATCC 12990,
DSM 10155 (X94197) and Bacillus sp. HC5 (AF252327).
The new isolate was identified as Aneurinibacillus ther-
moaerophilus strain HZ. The analysis result indicated that it
is phylogenetically distant from A. thermoaerophilus strain
ATCC 12990, DSM 10155 (X94197). In addition, there
have not yet been any reports on lipase from A. thermo-
aerophilus presented. Phylogenetic relationship of closely
related Aneurinibacillus spp. is shown in Fig. 2.
Effect of different assay temperatures on crude
HZ lipase
On the basis of relatively higher lipase activity detected for
A. thermoaerophilus strain HZ, the effect of temperature on
crude HZ lipase activity was examined from 45 to 70�C. As
shown in Fig. 3, the crude enzyme manifested its maximal
activity at 65�C with olive oil as substrate. Although lipase
activity was strongly dependent on assay temperature, a
further increase in temperature to 65�C did not follow this
trend because, most likely, the flexibility of enzyme
increased and hence loosely bound to substrate. This resul-
ted in decreasing its turnover number. The crude enzyme of
strain HZ was fairly active at higher temperature compared
to lipases from others Bacillus spp. (Sharma et al. 2002;
Li and Zhang 2005), and B. coagulans BTS-3 (0.75 U/ml;
Kumar et al. 2005) which all exhibited maximal activity at
55�C. Kim et al. (2000) reported 0.2 U/ml of crude lipase
activity at 50�C by B. sterarothermophilus L1.
The effect of medium volume on the production
of lipase
The medium volume may have a great effect on the
microorganism growth and enzyme production. While a
large volume contains more oxygen, food and nutrients, the
air space in the container is decreased. The more air space
provides more oxygen that can dissolve slowly in the
medium, so the more bacteria are grown.
The effect of medium volume on lipase production and
bacterial growth by A. thermoaerophilus strain HZ is
shown in Fig. 4. The highest bacterial growth and lipase
production were obtained in 50 ml medium. This volume
with its air space supplied more oxygen for the maximum
lipase production and growth. Lower bacterial growth
occurred in 200 ml production volume with 5% reduction
when compared to the maximum and lipase production
decreased to 50% of the maximum. Lower lipase produc-
tion in 200 ml production volume could be due to the less
air space and consequent poorer aeration.
The highest lipase production in 50 ml was caused
because of the lower volume provided a bigger air space
and also more oxygen that slowly would be dissolved into
the medium. Also, Kumar et al. (2005) reported that higher
lipase production occurred in 50 ml of medium by Bacillus
coagulans BTS-3.
Fig. 1 16S rDNA gene (1,500 bp) of isolate A10 gene amplified via
PCR. Lane 1–3: PCR product of amplified 16S gene. Lane 4 Marker
GeneRuler DNA 1 kb
World J Microbiol Biotechnol
123
The effect of temperature on the production of lipase
The culture was incubated at different temperatures rang-
ing from 50 to 70�C to investigate the optimum
temperature for bacterial growth and lipase production.
Growth and lipase production by A. thermoaerophilus
strain HZ was detected at all temperatures tested (Fig. 5).
Generally, the optimum temperature for lipase production
Fig. 2 Rooted phylogenetic
tree showing the relationship of
A. thermoaerophilus strain HZ
to other Bacillus spp.
Fig. 3 Effect of temperature on crude HZ lipase. Lipase activity was
determined by using olive oil as the substrate with incubation at
various temperatures, i.e.: 45, 50, 55, 60, 65 and 70�C
0
0.05
0.1
0.15
0.2
0.25
0.3
50 100 150 200
Medium volume (mL)
Lip
ase
acti
vity
(U
/mL
)
4.35
4.4
4.45
4.5
4.55
4.6
4.65
4.7
4.75
Log
CF
U/m
L
Lipase activity Colony count
Fig. 4 Effect of medium volume on growth and lipase production by
A. thermoaerophilus strain HZ. Different volumes of production
medium were incubated in 500 ml blue cap bottles at 60�C under 150
rev/min shaking rate for 48 h
World J Microbiol Biotechnol
123

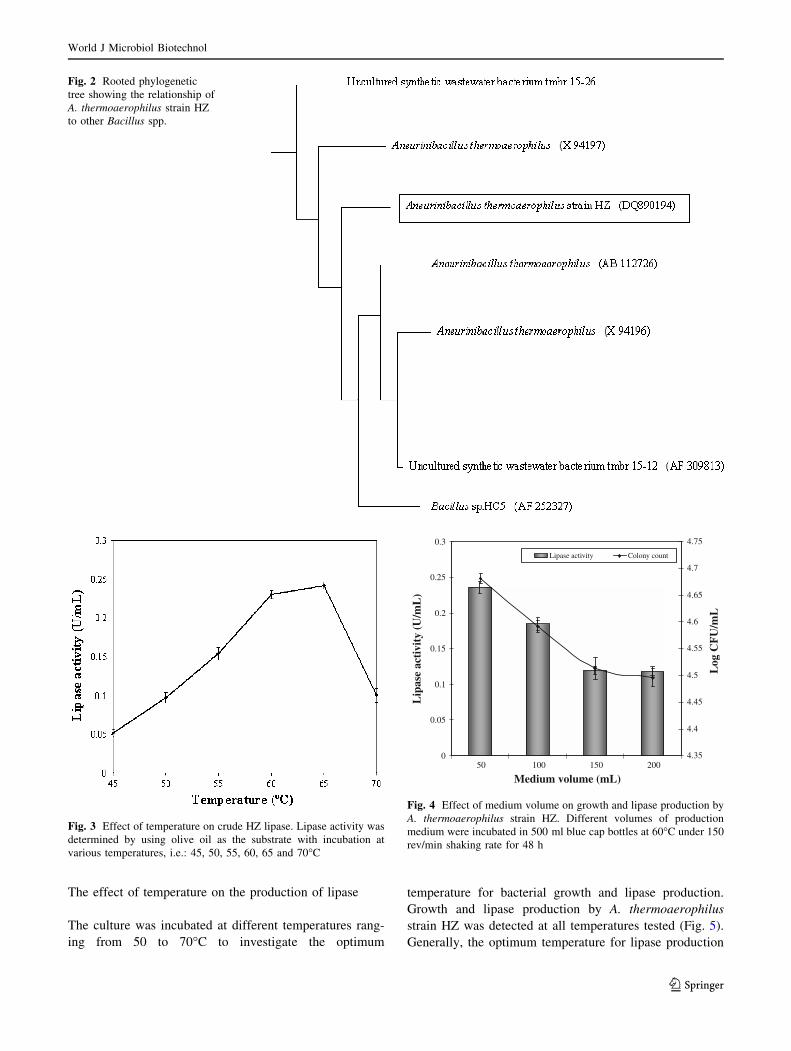
is different from that for optimum growth. The optimum
temperature for growth of A. thermoaerophilus strain HZ
was 50�C, but the optimum temperature for intracellular
lipase production was 60�C.
As the growth temperature increased more than 60�C,
the bacterial growth and lipase production by A. thermo-
aerophilus strain HZ decreased. The low lipase production
at 65�C can have been due to limited amount of total
dissolved oxygen in the medium. Oxygen has a low solu-
bility in aqueous media which further decreases with rising
temperature (Jacques 1977). In addition, increase in tem-
perature more than 60�C in A. thermoaerophilus strain HZ,
may possibly lead to denaturation of enzyme. In case of
extracellular enzymes, temperature influences their secre-
tion, possibility by changing the physical properties of the
cell membrane (Rahman et al. 2005a). Thus, too low and
too high temperatures are not suitable for lipase production
by A. thermoaerophilus strain HZ. The optimum temper-
ature for lipase production of the A. thermoaerophilus
strain HZ was higher than Bacillus coagulans BTS-3 that
exhibited maximum production at 55�C (Kumar et al.
2005). However, thermostable lipase from Geobacillus sp.
TW1 was reported to exhibit higher temperature of above
65�C (Li and Zhang 2005).
The effect of pH on the production of lipase
As microbial metabolism usually changes the pH of the
medium, some form of pH control is necessary in culture
systems.
Production medium of various pH values, ranging from
pH 4.0 to pH 9.0 were tested to determine the optimum pH
for lipase production and bacterial growth. As shown in
Fig. 6, very low lipase production was detected at acidic
(pH4.0 and 5.0) and alkaline conditions (pH 9.0). Maxi-
mum lipase production by A. thermoaerophilus strain HZ
was obtained at pH of 7.5 (0.26 U/ml) while maximum
bacterial growth was observed at pH 8.0. At pH 7.5, the
bacterial growth was almost the same with pH 8.0 but the
lipase production at pH 8.0 was only 71% of the maximum.
In acidic media from pH 4.0–6.5, bacterial growth was the
same but lipase production detected at pH 6.0 was only
70% of the maximum. The bacterial growth at alkaline pH
values (8.5 and 9.0) was lower compared with that in acidic
media and in pH 9.0 growth was greatly reduced.
The decrease in pH might have been due to the pro-
duction of acidic metabolites (Moon and Parulekar 1991).
In view of the close relationship between lipase synthesis
and use of the medium compounds, pH variation during
fermentation may reflect the kinetic changes in the lipase
production, such as the onset of production.
The effect of agitation on the production of lipase
Agitation has variable effect on lipase production by dif-
ferent organisms. It has been indicated as a key factor
influencing lipolytic enzyme production by mesophilic
microorganism (Sharma et al. 2002; Chen et al. 1999). The
relevance of this variable is even more remarkable when
thermophilic strains are considered, due to the low solu-
bility of gases at the high temperatures utilized throughout
the cultures. Although themophilic bacteria have adapted
to survive at low gas concentrations in their natural habi-
tats, rather high values of specific oxygen up-take rate have
0
0.05
0.1
0.15
0.2
0.25
50 55 60 65 70
Temperature (°C)
Lip
ase
acti
vity
(U
/mL
)
2
2.5
3
3.5
4
4.5
5
5.5
6
Log
CF
U/m
L
Lipase activityColony count
Fig. 5 Effect of temperature on growth and lipase production by
A. thermoaerophilus strain HZ. Cultures contain 50 ml of production
medium were incubated in different temperatures in water bath shaker
under 150 rev/min shaking rate for 48 h
0
0.05
0.1
0.15
0.2
0.25
0.3
4 5 6 6.5 7 7.5 8 8.5 9
pH
Lip
ase
acti
vity
(U
/mL
)
2
2.5
3
3.5
4
4.5
5
5.5
Log
CF
U/m
L
Lipase activityColony count
Fig. 6 Effect of pH on lipase production and growth by A.thermoaerophilus strain HZ. Cultures with different pHs from 4 to
9 at 0.5 unit intervals were incubated at 60�C in water bath shaker at
60�C under 150 rev/min shaking rate for 48 h
World J Microbiol Biotechnol
123
been reported in some cases (Belo et al. 2000). The use of
pressurized or gas lift fermentors has been proposed as
means to provide an adequate oxygen concentration in the
culture medium. The latter is also useful to minimise shear
stress, to which some extreme thermophiles appear to be
extremely sensitive (Belo et al. 2000; Holst et al. 1997).
The effect of agitation on bacterial growth and lipase
production was studied under static condition, 50, 100, 150
and 200 rev/min. As demonstrated in Fig. 7, the highest
lipase production was obtained when agitated at 150 rev/
min. The static condition to 50 rev/min shaking enhanced
bacterial growth by A. thermoaerophilus strain HZ while
lipase production was only 11% and 13% of the maximum,
respectively. Shaking at 100 rev/min supported HZ lipase
production at 77% compared to that at 150 rev/min, but its
bacterial growth was 84% at static condition and decreased
to 83% at 150 rev/min. Higher shaking rate reduces the
bacterial growth because during respiration, hydrogen
atoms may be combined with oxygen forming hydrogen
peroxide which is lethal to the cell. Too much aeration
facilitates hydrogen peroxide production to the certain
level where enzyme that is produced by the bacterial is not
enough to breakdown the hydrogen peroxide and thus leads
to cell death (Lee 2002).
Chander et al. (1980) opined that increased enzyme
secretion by Aspergillus wentii in liquid media when agi-
tated may be due to the greater availability of dissolved
oxygen for its growth and lipase production. Kumar et al.
(2005) reported 1.16 U/ml of lipase production by B. co-
agulans BTS-3 under 170 rev/min agitation of culture
medium. In contrast, Alford and Smith (1965) reported that
Geotrichum candidum cultured without shaking produced
more lipase than with shaking. This maybe due to
liberation of a proteolytic enzyme by G. candidum which
resulted in hydrolysis of lipase (Lawrence et al. 1967).
Rahman et al. (2006) found significant lipase production by
Pseudomonas sp. strain S5 under static condition. In con-
clusion, the general increase in lipase production with
agitation rate is probably due to improved oxygenation of
the medium.
The effect of inoculum size on the production of lipase
The total dissolved oxygen can be affected by using dif-
ferent inoculum size and hence influence the lipase
production.
Inoculating production medium with various inoculum
sizes of A. thermoaerophilus strain HZ affected the bac-
terial growth and lipase production. As observed in Fig. 8,
enzyme production and bacterial growth increased with the
increase of inoculum size with an optimum at 7% (v/v;
0.24 U/ml) after which the level of enzyme production and
bacterial count both decreased. At low inoculum size, the
nutrient and oxygen levels were sufficient for the growth of
bacteria and, therefore, enhanced lipase production.
A. thermoaerophilus strain HZ only produces high lipase
at the end of the stationary phase (data not shown). Lipase
production and bacterial growth were decreased to 84 and
83% when 9% inoculum size was applied to the medium,
respectively. This may be because the nutrients begin to be
used up by the large population of the bacterium before it is
physiologically ready to start producing lipase. On the
other hand, the low lipase production by the 1% (v/v)
0
0.05
0.1
0.15
0.2
0.25
0.3
0 50 100 150 200
Agitation (rpm)
Lip
ase
acti
vity
(U
/mL
)
2
2.5
3
3.5
4
4.5
5
5.5
6
Log
CF
U/m
L
Lipase activityColony count
Fig. 7 Effect of agitation on lipase production and growth by A.thermoaerophilus strain HZ. Cultures contain 50 ml of production
medium with initial pH 7.5 were incubated at 60�C with different
shaking rate at 60�C for 48 h
0
0.05
0.1
0.15
0.2
0.25
0.3
1 3 5 7 9
Inoculum size (%)
Lip
ase
acti
vity
(U
/mL
)
2
2.5
3
3.5
4
4.5
5
5.5
Log
CF
U/m
L
Lipase activityColony count
Fig. 8 Effect of inoculum size on lipase production and growth by
A. thermoaerophilus strain HZ. Cultures contain 50 ml of production
medium were inoculated with different percentage of inoculum size
and incubated at 60�C under 150 rev/min shaking rate for 48 h
World J Microbiol Biotechnol
123

inoculum is probably due to insufficient bacterium to
produce much enzyme. ul-Haq et al. (2002) stated that the
optimum population of microorganism will produce the
high amount of enzyme. However, high inoculum sizes
might not necessarily give higher enzyme yield or cell
growth. In addition, greater inoculum size can lead to an
impaired nutrient supply and thereby decreasing bacterial
growth and production capacity (Kopp 1987). Previous
studies have identified different optimum inoculum sizes
1% (v/v; Kambourova et al. 2003), 3% (v/v; Gupta et al.
2007), 6% (v/v; Rahman et al. 2005b) and 10% (v/v; Jasvir
et al. 1999) for lipase production of different enzymes. So,
there is no specific inoculum size for maximum lipase
production.
Organic solvent stability of crude enzyme
Substrates of the lipase are often insoluble in aqueous
solutions, and organic solvents or organic-aqueous two-
phase media are favorable for these reactions. However,
enzymes are generally not stable in the presence of organic
solvents and are apt to denature (Ogino et al. 1994). It is
well known that effect of organic solvents on lipase activity
differs from lipase to lipase (Sugihara et al. 1991).
Recently many efforts have been made to discover a new
organic solvent tolerant lipase that could create new
applications.
Organic solvents of various log P values from -1.3 to
8.8 were chosen to study the effect of solvent on the
hydrolysis of oil. Log P value, the partition coefficient
between water and 1-octanol, is often used as an indicator
of solvent polarity. Table 1 shows the crude lipase from
A. thermoaerophilus strain HZ that was stable in most of
the organic solvents with log P values of -1.3 to 3.6 and
the enzyme activity was more than 90% of that in the
absence of an organic solvent. The higher lipase activity
was exhibited by HZ lipase in present of dimethyl sulf-
oxide (Log P -1.3). In addition, toluene (Log P 2.5),
p-xylene (log P 3.1), and hexane (Log P 3.6) slightly
enhanced the crude lipase activity of A. thermoaerophilus
strain HZ. However, the HZ lipase exhibited less than 40%
decrease of its activity in the presence of dodecanol (log P
5.0), n-tetradecane (log P 7.6) and hexadecane (log P 8.8).
In the presence of decanol (Log P 4.0) and decane (log P
5.6), slight loss of activity was observed.
Hun et al. (2003) reported that 80% activity of lipase
from B. sphaericus strain 205y was lost in the presence of
dimethyl sulfoxide. In contrast, the most activity of the
crude HZ lipase was noticed in the presence of hydrophilic
solvents (-1.3 \ log P \ 3.6). However, the HZ lipase
activity was observed to be decreased by water-immiscible
organic solvents with log P [ 4. No lipase activity was
detected from lipase of B. sphaericus strain 205y in the
presence of hexadecane, which has a high log P value of
8.8 (Hun et al. 2003). From the results of this study, no
specific styles of lipase activities with respect to the
polarity of organic solvents were seen. Some organic sol-
vent tolerant lipases, including the HZ lipase, still retain
their activity even if few molecules of water remain; pre-
sumably just sufficient to stabilize the conformation of the
active site. Therefore, their stability in various organic
solvents may be different from other organic solvent tol-
erant lipase.
Acknowledgment This work was supported by the Sciencefund
Grant No. 02-01-04-SF0212 from the Malaysian Ministry of Science/
Technology and Innovation (MOSTI).
References
Alford JA, Smith JL (1965) Production of microbial lipase for the
study of triglyceride structure. J Am Oil Chem Soc 42(12):1038–
1040
Belo I, Pinheiro R, Mota M (2000) Response of the thermophile
Thermus sp. RQ-1 to hyperbaric air in batch and fed-batch
cultivation. Appl Microbiol Biotechnol 53:517–524
Bornscheuer UT (2002) Microbial carboxyl esterase: classification,
properties and application in biocatalysis. FEMS Microbiol Rev
26:73–81
Burkert JFM, Maugeri F, Rodrigues MI (2004) Optimization of
extracellular lipase production by Geotrichum sp. using factorial
design. Bioresour Technol 91(1):77–84
Chander H, Batish VK, Sannabhadti SS, Srinivasan RA (1980)
Factors affecting lipase production in Aspergillus wentii. J Food
Sci 45:598–600
Chen JY, Wen CM, Chen TL (1999) Effect of oxygen transfer on
lipase production by Acinetobacter radoresistens. Biotechnol
Bioeng 62:311–316
Gotor-Fernandez V, Brieva R, Gotor V (2006) Lipases: useful
biocatalysts for the preparation of pharmaceuticals. J Mol Catal
B Enzym 40(3–4):111–120
Table 1 Effect of organic solvents on stability of the crude lipase
from A. thermoaerophilus strain HZ .25% (v/v) of organic solvents
were added to the cell-free supernatant and incubated at 65�C with
200 rev/min shaking for 30 min
Organic solvent Log P Remaining lipase
activity (%)
SD
Dimethyl sulfoxide -1.3 123 0.005
Benzene 2.0 93.8 0.004
Toluene 2.5 106.9 0.01
p-Xylene 3.1 101.01 0.02
Hexane 3.6 106.4 0.03
Decanol 4.0 83.9 0.02
Dodecanol 5.0 61.1 0
Decane 5.6 91.2 0.03
n-Tetradecane 7.6 64.9 0.04
Hexadecane 8.8 72.9 0.06
No solvent – 100 0.04
World J Microbiol Biotechnol
123

Gupta R, Gupta N, Rathi P (2004) Bacterial lipases: an overview of
production, purification and biochemical properties. Appl
Microbiol Biotechnol 64(6):763–781
Gupta N, Sahai V, Gupta R (2007) Alkaline lipase from a novel strain
Burkholderia multivorans: statistical medium optimization and
production in a bioreactor. Process Biochem 42(4):518–526
Haba E, Bresco O, Ferrer C, Marque’s A, Busquets M, Manresa A
(2000) Isolation of lipase-secreting bacteria by deploying used
frying oil as selective substrate. Enzyme Microb Technol
26(1):40–44
Hasan F, Shah AA, Hameed A (2006) Industrial applications of
microbial lipases. Enzyme Microb Technol 39(2):235–251
Holker U, Hofer M, Lenz J (2004) Biotechnological advantages of
laboratoryscale solid-state fermentation with fungi. Appl Micro-
biol Biotechnol 64:175–186
Holst O, Manelius A, Krahe M, Markl H, Raven N, Sharp R (1997)
Thermophiles and fermentation technology. Comp Biochem
Physiol 118(3):415–422
Hun CJ, Rahman RNZA, Salleh AB, Basri M (2003) A newly isolated
organic solvent tolerant Bacillus sphaericus 205y producing
organic solvent-stable lipase. Biochem Eng J 15(2):147–151
Jacques R (1977) Culture media. In: Moss MO, Smith JE (eds)
Industrial application of microbiology. Surrey University Press,
London, pp 27–58
Jasvir S, Gill N, Devasahayam G, Sahoo DK (1999) Studies on
alkaline protease produced by Bacillus sp.NG312. Appl Bio-
chem Biotechnol 76(1):57–63
Kambourova M, Kirilova N, Mandeva R, Derekova A (2003)
Purification and properties of thermostable lipase from a
thermophilic Bacillus stearothermophilus MC 7. J Mol Catal B
Enzym 22:307–313
Kamini NR, Fujii T, Kurosu T, Lefuji H (2000) Production,
purification and characterization of an extracellular lipase from
the yeast, Cryptococcus sp. S-2. Process Biochem 36(4):317–324
Kim MH, Kim HK, Lee JK, Park SY, Oh TK (2000) Thermostable
lipase of Bacillus sterarothermophilus: high-level production,
purification and calcium-dependent thermostability. Biosci Bio-
technol Biochem 64(2):280–286
Kopp B (1987) Long-term alkaloid production by immobilized cells
of Claviceps purpurea. Meth Enzymol 136:317–329
Kumar S, Kikon K, Upadhyay A, Kanwar SS, Gupta R (2005)
Production, purification, and characterization of lipase from
thermophilic and alkaliphilic Bacillus coagulans BTS-3. Protein
Expr Purif 41:38–44
Kwon DY, Rhee JS (1986) A simple and rapid colorimetric method
for determination of free fatty acids for lipase assay. J Am Oil
Chem Soc 63(1):89–92
Lawrence RC, Fryer TF, Reiter B (1967) The production and
characterization of lipases from a micrococcus and a pseudo-
monad. J Gen Microbiol 48(3):401–418
Lee PG (2002) Production, purification, characterization and
expression of the organic solvent tolerant protease gene. Ph.D.
thesis, Universiti Putra Malaysia, Malaysia
Li H, Zhang X (2005) Characterization of thermostable lipase from
thermophilic Geobacillus sp. TW1. Protein Expr Purif 42:153–159
Moon SH, Parulekar SJ (1991) A parametric study of protease
production in batch and fed-batch culture of Bacillus firmus.
Biotechnol Bioeng 37(5):467–483
Niehaus F, Bertoldo C, Kahler M, Antranikian G (1999) Extremo-
philes as a source of novel enzymes for industrial application.
Appl Microbiol Biotechnol 51:711–729
Ogino H, Miyamoto K, Ishikawa H (1994) Organic solvent-tolerant
bacterium which secretes organic-solvent-stable lipolytic
enzyme. Appl Environ Microbiol 60:3884–3886
Ogino H, Miyamoto K, Yasuda M, Ishimi K, Ishikawa H (1999)
Growth of organic solvent-tolerant Pseudomonas aeruginosaLST-03 in the presence of various organic solvents and
production of lipolytic enzyme in the presence of cyclohexane.
Biochem Eng J 4:1–6
Pennisi E (1997) Biotechnology: in industry, extremophiles begin to
make their mark. Science 276:705–706
Rahman RNZA, Geok LP, Basri M, Salleh AB (2005a) Physical
factors affecting the production of organic solvent-tolerant
protease by Pseudomonas aeroginosa strain K. Bioresource
Technol 96(4):429–436
Rahman RNZRA, Baharum SN, Basri M, Salleh AB (2005b) High-
yield purification of an organic solvent-tolerant lipase from
Pseudomonas sp. strain S5. Anal Biochem 341:267–274
Rahman RNZRA, Baharum SN, Salleh AB, Basri M (2006) S5 lipase:
an organic solvent tolerant enzyme. J Microbiol 44(6):583–590Saxena RK, Davidson WS, Sheoran A, Giri B (2003) Purification and
charactrization of an alkaline thermostable lipase from Asper-gillus carneus. Process Biochem 39(2):239–247
Sharma R, Soni SK, Vohra RM, Gupta LK, Gupta JK (2002)
Purification and characterization of a thermostable alkaline
lipase from a new thermophilic Bacillus sp. RSJ-1. Process
Biochem 37(10):1075–1084
Sugihara A, Tani T, Tominaga Y (1991) Purification and character-
ization of a novel thermostable lipase from Bacillus sp.
J Biochem 109(2):211–215
Thompson JD, Higgins DG, Gibson TJ (1994) CLUST AL W:
important the sensitivity of progressive multiple sequence
alignment through sequence weighting, position-specific gap
penalties and weight matrix choice. Nucleic Acids Res
22(22):4673–4680
ul-Haq I, Idrees S, Rajoka MI (2002) Production of lipases by
Rhizopus oligosporous by solid-state fermentation. Process
Biochem 37(6):637–641
Woese C (1998) The universal ancestor. Proc Natl Acad Sci USA
95(12):6854–6859
World J Microbiol Biotechnol
123
Copyright © 2022 FDOKUMEN




![EVcc`c Wc`^ 2W hZ]] SV UVR]e+ 45D - Daily Pioneer](https://static.fdokumen.com/doc/165x107/632551385c2c3bbfa803227c/evccc-wc-2w-hz-sv-uvre-45d-daily-pioneer.jpg)















