
High affinity uptake and Ca++ -dependent release of glutamic acid in the developing cerebellum
9
Neuroscience Vol. 11, No. 4, pp. 867-875, 1984 Printed in Great Britain 0306-4522/84 $3.00 + 0.00 Pergamon Press Ltd 0 1984IBRO HIGH AFFINITY UPTAKE AND Ca2+-DEPENDENT RELEASE OF GLUTAMIC ACID IN THE DEVELOPING CEREBELLUM M. E. SANDOVAL*, C. A. TORNBR and L. MEDRANO Centro de Investigaciones en Fisiologia Celular, Universidad National Autonoma de Mexico, Mexico, D.F., Mexico Abstract-This study was undertaken in order to evaluate the postulated role of glutamic acid as the neurotransmitter for the parallel fibers of the cerebellar cortex. We studied the Ca*+-dependent release and the high affinity uptake of glutamic acid in the developing cerebellum. The Ca*+dependent release of glutamic acid from cerebellar molecular layer during development closely follows the time course of parallel fibers synaptogenesis. Little glutamic acid release was observed at 15 days, then it increased to the adult values at the 21st postnatal day. In the rat the bulk of synapses of the parallel fibers appear between the 15th and the 21st postnatal days, the time at which the nerve terminals of the climbing fibers, the other excitatory input to the Purkinje cells, are already developed. An enhanced Na+dependent, high affinity uptake of glutamic acid was observed in the developing cerebellum relative to the adult rat. That this higher accumulation of glutamic acid is not related to a releasable pool is suggested by the fact that an enhanced glutamic acid, Ca 2+-dependent release relative to the adult was not observed. These results support the view that glutamic acid is the transmitter for the cerebellar parallel fibers. The cerebellar cortex has been considered an inter- esting model for studying the neurotransmitters at specific synapses, because it has been extensively studied from an anatomical and physiological points of view.a,‘2*27 Biochemical and physiological studies suggest that Purkinje, basket and Golgi neurons utilize y-aminobutyric acid (GABA) as their neu- rotransmitter,34”8*P4,45 and there is increasing evidence that glutamic acid may be an excitatory transmitter in the cerebellum. Glutamic acid has an excitatory action on the Purkinje cells4 and both Ca*+-dependent release and high affinity uptake of glutamic acid have been described in several prepara- tions from cerebellar cortex.‘0,‘3*‘4~20*32+46 However it is unclear whether glutamic acid may serve as a trans- mitter at either parallel or climbing fiber nerve end- ings or both, although there is considerable evidence suggesting that glutamic acid may be the transmitter of the parallel fibers. After loss of granule cells, glutamic acid levels are reduced23*3’s4’ and there is a selective depletion of both high affinity uptake of aspartic and glutamic acid3’*& and Ca2+-dependent release of glutamic acid from cerebellar slices’3 and synaptosomes.32 Developmental studies in the newborn rat, on the other hand, showed that synaptogenic activity of the climbing and parallel fibers occurs at different post- *To whom correspondence should be addressed: Centro de Investigaciones en Fisiologia Celular, Universidad Na- cional Autonoma de Mexico, Apartado Postal 70-600, 04510 Mexico D. F., Mexico. Abbreviak_ms: EGTA, Ethyleneglycol-his-@-aminoethyl ether) N,N’-tetra-acetic acid; GABA, y-aminobutyric acid. natal days: synapse formation by the climbing fibers begins on the 3rd day after birth and mature, func- tional synapses have been observed about the 7th day, before the full development of the dendritic tree of the Purkinje cells.2*6*7*‘7,29,37 The maturation of parallel fibers starts later, the bulk of granule cells appear during the 2nd postnatal week and due to the time required for cell migration and the lag in the formation of the dendrites, few synapses appear before the 2nd week after birth and synaptogenesis is complete around the 32nd day.2q3,37*44 Synapses of the inhibitory interneurons develop during the 2nd post- natal week.3*37 Taking advantage of the specific time course of synapse formation in both parallel and climbing fibers, the developing cerebellum offers an excellent model to assess the role of glutamic acid as the neurotransmitter at its excitatory synapses. In the present work, we studied the high affinity uptake and the Ca*+-dependent release of glutamic acid, two of the major criteria in the identification of neu- rotransmitters, in the developing rat cerebellum. EXPERIMENTAL PROCEDURES Tissue preparation Wistar albino rats-15-32 days old and adults-were decapitated and the cerebellum removed. Crude syn- aptosomal fractions (P2) were prepared as follows: tissue samples were homogenized in 0.32 M sucrose and homoge- nates centrifuged at 158Og for 10min; the supematant was centrifuged at 18,OOOg for 15 min and the resulting pellet resuspended in the appropriate medium. Homogenates of the molecular layer were used in the release experiments. Sagittal sections (0.4-0.5 mm) of the cerebellum were obtained in a McIlwain tissue chopper. The slices were placed in cold 0.32 M sucrose and the molecular 867
Transcript of High affinity uptake and Ca++ -dependent release of glutamic acid in the developing cerebellum

Neuroscience Vol. 11, No. 4, pp. 867-875, 1984 Printed in Great Britain
0306-4522/84 $3.00 + 0.00 Pergamon Press Ltd
0 1984 IBRO
HIGH AFFINITY UPTAKE AND Ca2+-DEPENDENT RELEASE OF GLUTAMIC ACID IN THE
DEVELOPING CEREBELLUM
M. E. SANDOVAL*, C. A. TORNBR and L. MEDRANO Centro de Investigaciones en Fisiologia Celular, Universidad National Autonoma de Mexico, Mexico,
D.F., Mexico
Abstract-This study was undertaken in order to evaluate the postulated role of glutamic acid as the neurotransmitter for the parallel fibers of the cerebellar cortex. We studied the Ca*+-dependent release and the high affinity uptake of glutamic acid in the developing cerebellum. The Ca*+dependent release of glutamic acid from cerebellar molecular layer during development closely follows the time course of parallel fibers synaptogenesis. Little glutamic acid release was observed at 15 days, then it increased to the adult values at the 21st postnatal day. In the rat the bulk of synapses of the parallel fibers appear between the 15th and the 21 st postnatal days, the time at which the nerve terminals of the climbing fibers, the other excitatory input to the Purkinje cells, are already developed. An enhanced Na+dependent, high affinity uptake of glutamic acid was observed in the developing cerebellum relative to the adult rat. That this higher accumulation of glutamic acid is not related to a releasable pool is suggested by the fact that an enhanced glutamic acid, Ca 2+-dependent release relative to the adult was not observed.
These results support the view that glutamic acid is the transmitter for the cerebellar parallel fibers.
The cerebellar cortex has been considered an inter- esting model for studying the neurotransmitters at specific synapses, because it has been extensively studied from an anatomical and physiological points of view.a,‘2*27 Biochemical and physiological studies suggest that Purkinje, basket and Golgi neurons utilize y-aminobutyric acid (GABA) as their neu- rotransmitter,34”8*P4,45 and there is increasing evidence that glutamic acid may be an excitatory transmitter in the cerebellum. Glutamic acid has an excitatory action on the Purkinje cells4 and both Ca*+-dependent release and high affinity uptake of glutamic acid have been described in several prepara- tions from cerebellar cortex.‘0,‘3*‘4~20*32+46 However it is unclear whether glutamic acid may serve as a trans- mitter at either parallel or climbing fiber nerve end- ings or both, although there is considerable evidence suggesting that glutamic acid may be the transmitter of the parallel fibers. After loss of granule cells, glutamic acid levels are reduced23*3’s4’ and there is a selective depletion of both high affinity uptake of aspartic and glutamic acid3’*& and Ca2+-dependent release of glutamic acid from cerebellar slices’3 and synaptosomes.32
Developmental studies in the newborn rat, on the other hand, showed that synaptogenic activity of the climbing and parallel fibers occurs at different post-
*To whom correspondence should be addressed: Centro de Investigaciones en Fisiologia Celular, Universidad Na- cional Autonoma de Mexico, Apartado Postal 70-600, 04510 Mexico D. F., Mexico.
Abbreviak_ms: EGTA, Ethyleneglycol-his-@-aminoethyl ether) N,N’-tetra-acetic acid; GABA, y-aminobutyric acid.
natal days: synapse formation by the climbing fibers begins on the 3rd day after birth and mature, func- tional synapses have been observed about the 7th day, before the full development of the dendritic tree of the Purkinje cells.2*6*7*‘7,29,37 The maturation of parallel fibers starts later, the bulk of granule cells appear during the 2nd postnatal week and due to the time required for cell migration and the lag in the formation of the dendrites, few synapses appear before the 2nd week after birth and synaptogenesis is complete around the 32nd day.2q3,37*44 Synapses of the inhibitory interneurons develop during the 2nd post- natal week.3*37
Taking advantage of the specific time course of synapse formation in both parallel and climbing fibers, the developing cerebellum offers an excellent model to assess the role of glutamic acid as the neurotransmitter at its excitatory synapses. In the present work, we studied the high affinity uptake and the Ca*+-dependent release of glutamic acid, two of the major criteria in the identification of neu- rotransmitters, in the developing rat cerebellum.
EXPERIMENTAL PROCEDURES
Tissue preparation
Wistar albino rats-15-32 days old and adults-were decapitated and the cerebellum removed. Crude syn- aptosomal fractions (P2) were prepared as follows: tissue samples were homogenized in 0.32 M sucrose and homoge- nates centrifuged at 158Og for 10min; the supematant was centrifuged at 18,OOOg for 15 min and the resulting pellet resuspended in the appropriate medium.
Homogenates of the molecular layer were used in the release experiments. Sagittal sections (0.4-0.5 mm) of the cerebellum were obtained in a McIlwain tissue chopper. The slices were placed in cold 0.32 M sucrose and the molecular
867

layer was hand-dissected from each shcc under a dissecting microscope. Molecular layer strips dissected out from about nine cerebellar slices wcrc used for each expcrlment with adult rats: while for younger ammals I? I4 slices from two cerebella were employed. Molecular layer strips were kept in cold 0.32 M sucrose before use (about S&6Omin). Tissue samples are able to accumulate and release neu- rotransmitters under these experimental conditions.‘4.” Im- mediately before use. molecular layer strips were homoge- nized in 0.32 M sucrose and diluted in a Krebs-bicarbonate medium of the following composition, (mM): NaCI. 1 IS; KCI, 3: CaCI,, 2; NaH,PO,, 1.2; MgSO,, I.2 NaHCO,, 25; glucose, 10; pH 7.4 maintained with CO,-0, mixture (S”Q95pd).
Uptake experimenrs
We followed the method for transmitter uptake described elsewhere.32.y3 Briefly, P, fractions were resuspended in a Na’-free medium; (see below) and aliquots (about IO0 pg protein/ml) preincubated for 3 min at 37°C in either of two media: the Krebs-bicarbonate buffer described above or in a Nat-free medium; then aliquots of [“Cjclglutamic acid were added. After 2 min. aliquots of the incubation medium were filtered in a Millipore multifilter, using Unipore disc prefilters, and the tissue samples were washed, at room temperature, with 2ml of the Na+-free medium. The Na+-free medium used was of the following composition, (mM): choline chloride, 137; KC). 3; MgSO,, IS; KH$O,, 1.2; glucose, 10; HEPES, IO; pH 7.4 with Tris.
Radioactivity in the filters was determined in Tritosolts after tissue digestion in 2% sodium dodecyl sulphate. 20 mM ethylenediaminetetra-acetate pH 8.0.
Natdependent uptake was taken as [‘4C]glutamic acid accumulated in the Krebs-bicarbonate medium after sub- tracting the amino acid uptake measured in the Na.‘-free medium.
Eflux experiments
The procedure described for the study of the transmitter efflux from synaptosomal fractions was fo11owed.“~” Ali- quots of cerebellar molecular layer homogenates (about 200 fig protein/ml) were preincubated at 37°C in the Krebs- bicarbonate described above; after 3min. aliquots of the radiolabeled amino acids (0.8 p M final concentration) were added and incubation continued for Smin. One mililiter aliquots were then transferred into modified Swinex Milli- pore filter units placed in a vacuum box, as described earlier,2’J2~33 on Unipore disc prefilters. Af:er removal of the incubation medium by filtration, tissue samples received six 2 ml washes of CL+*‘-free, Krebs-bicarbonate medium con- taining 1 mM Ethylmeglycol-his-(B-aminoethyl ether) NJ’-tetra-acetic acid (EGTA). Each wash lasted for 30s and the wash solution was subsequently removed by vac- uum application; this treatment results in a constant base- line of radioactivity efflux. After the sixth wash, samples received a 30s pulse with 2ml of one of the following experimental solutions: (a) Ca*+-free, Krebs-bicarbonate medium containing 1 mM EGTA; (b) Ca" -free, Krebs- bicarbonate medium containing 1 mM EGTA and either 56mM KCI (NaCl reduced to 53 mM) or veratrine (60 pg/ml); (c) Krebs-bicarbonate medium containing 2 mM CaCI, and either 56 mM KC1 or veratrine (60 II dml). At the end df the experiment, radioactivity in the flit& w& deter- mined as described above while radioactivity in the filtrates collected during the application of the experimental pulse was measured in tritosoi.”
Results are expressed as the ratio of radioactivity released after the experimental pulse to the total radioactivity in the tissue before the pulse,’ as follows:
% efflux = -__ dpm filtrate
(dpm filtrate) + (dpm tissue) X loo.
Under these expcnmental con&ions. the iollo\rtng p- rametcrs can be determined: (a) baseline eRlux ammo XXI spontaneous efflux under non-stimulating condltlonz: (hr depolarization-induced efflux amino acid efflux evoked h! high K. or veratrme in the absence of external Ca’ anti (c) Ca’--dependent releascaminn acid ctIhrx Induced hy depolarization.
The metabolism of [‘%Jglutamic acid during the caper,- mental procedure was determined as follows: alter the experimental pulse. tissue samples received an osmotic shock with 2ml of cold distilled water and filtered after 2 min. With this treatment more than 90”; of radioactivlty in the tissue was released. Aliquots of the collected mLdia and the tissue lysata were acidified and desalted by using a cationic exchange column AGSOW x 8 and amino acids eluted with I M NH,OH; aiiquots from the eluate were dried under vacuum in a Savant centrifuge. Samples were resuspended in 0.1 N HCl and analyzed by thin layer chromatography with the following solvent system: 2-butanol:H,O (4:1 v/v) and then, ethanol:H,O:ethyl- eneglycol (7:3:1 v/v). The chromatography lane was scanned for radioactivity by removing 0.5 cm wide strips of silica from the plate and counting each sample in tritosol after elution with 0.1 N HCI.
Results showed that under the experimental conditions described above. 59.75 3.1. (n = 5). and 72.5 + 10.1. (n = 51, per cent of the total radroactivity remained as glutamic acid in the homogenates of molecular layer from I5 day old and adult cerebellum. respectively. The per- centage of unmetabolized [‘4C]glutamic acid in the collected media from control samples were: 57.9 + 14. (n = 4), for the 15 day-old cerebellum. and 54. I i 15. (n = 4). for the adult cerebellum. Similar values were obtained when un- metabolized [“CJglutamic acid was determined both in tissue samples and collected media from molecular layer homogenates depolarized for 30s. Protein was measured following the method described by Lowry pi ol.??
Histological conrrolv
Some strips of molecular layer dissected out from I8 day old and adult cerebellum, were fixed by immersion in phosphate buffered-44; formaldehyde for 24h at 4’C, washed overnight in the same phosphate buffer (0. I M. pH 7.2). embedded in paraffin, sectioned and stained with hematoxylin-eosin for examination under the tight micro- scope.
MaIeria1.s
[L-U-‘4C]glutamic acid (sp. act. 297 mCi/mmol), [)H]GABA (40.2 Ci/mmol) and [L-U-‘%Z]aspartic acid (225 mCi/mmoI). were purchased from New England Nuclear Corp. (Boston, MA). Veratrine was obtained from Sigma Chemical Co. (St. Louis. MO).
RESULTS
High afinity uptake of [14Clglutmic acid in the dayel- oping cerebellum
P2 fractions from cerebella obtained from IS to 32 days old and adult rats were used for studying (“Qlutamic acid uptake. Na’dependent, high affinity uptake of [‘*C$lutamic acid was observed at ail the ages studied. (Fig. 1). However, a clear different activity for glutamate accumulation was observed during ccrcbeIlar development. As shown in Fig. 1, P2 fractions from 15 day old cerebellum presented the highest activity for giutamic acid up- take. This activity changes dramaticalIy from the 15th to the 17th and 19th postnatal days. We found
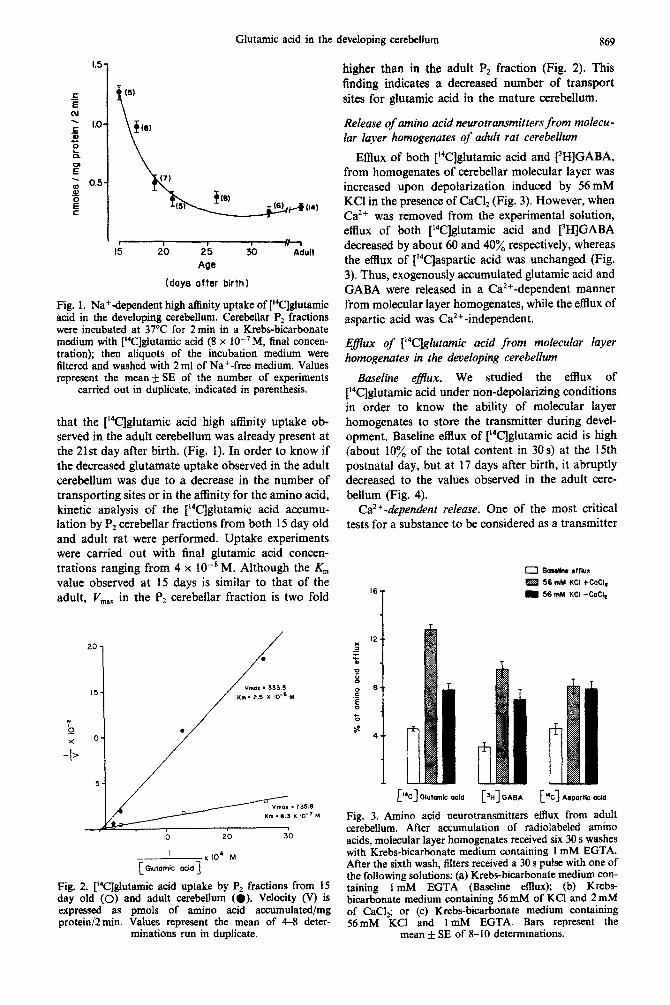
Glutamic acid in the developing cerebellum 869
higher than in the adult PZ fraction (Fig. 2). This finding indicates a decreased number of transport sites for glutamic acid in the mature cerebellum.
I , I I 1 I
I5 20 25 30 Adult
Aw
(days after birth)
Fig. 1. Na+dependent high affinity uptake of [i4C]glutamic acid in the developing cerebellum. Cerebellar P2 fractions were incubated at 37°C for 2min in a K~bs-bi~r~nate medium with [i4C]giutamic acid (8 x lo-’ M, final concen- tration); then aliquots of the incubation medium were filtered and washed with 2 ml of Na+-free medium. Values represent the mean &-SE of the number of experiments
carried out in duplicate, indicated in parenthesis.
that the [‘4C]glutamic acid high affinity uptake ob- served in the adult cerebellum was already present at the 21st day after birth. (Fig. 1). In order to know if the decreased glutamate uptake observed in the adult cerebellum was due to a decrease in the number of transporting sites or in the affinity for the amino acid, kinetic analysis of the [14C]~utamic acid accumu- lation by Pt cerebellar fractions from both 15 day old and adult rat were performed. Uptake experiments were carried out with final glutamic acid concen- trations ranging from 4 x IO-* M. Although the X;, value observed at 15 days is similar to that of the adult, V,,,,, in the P2 cerebellar fraction is two fold
-I>
c I YO’ M Glutomic acid
Fig. 2. [t4CJglutamic acid uptake by P2 fractions from 15 day old (0) and adult cerebellum (0). Velocity (V) is expressed as pmols of amino acid accumulated/mg protein/2min. Values represent the mean of 4-8 deter-
minations run in duplicate.
Release of amino acid neurotransmitters from molecu- lar layer homogenates of adult rat cerebellum
Efflux of both [‘4C]glutamic acid and rH]GABA, from homogenates of cerebellar molecular layer was increased upon depolarization induced by 56mM KC1 in the presence of CaCI, (Fig. 3). However, when Ca*+ was removed from the experimental solution, efflux of both [‘4C]glutamic acid and t3H]GABA decreased by about 60 and 40% respectively, whereas the efflux of [14C]aspartic acid was unchanged (Fig. 3). Thus, exogenously accumulated glutamic acid and GABA were released in a Ca2+-dependent manner from molecular layer homogenates, while the efflux of aspartic acid was Ca2+-independent.
EJGx of [‘4clglutumic acid from molecular layer homogenates in the developing cerebellum
Baseline e&x. We studied the efflux of ~‘4C]glutamic acid under non-depoI~zing conditions in order to know the ability of molecular layer homogenates to store the transmitter during devel- opment. Baseline efflux of [‘4C]glutamic acid is high (about 10% of the total content in 30s) at the 15th postnatal day, but at 17 days after birth, it abruptly decreased to the values observed in the adult cere- bellum (Fig. 4).
Ca2i-dependent release. One of the most critical tests for a substance to be considered as a transmitter
[“C]Gl”tomk Dcld
0 - rifftw m 56 mM KCI +CaCI.
- 56 mM KCI -CoCI.
[%] GABA [*C] AsDartk aO!d
Fig. 3. Amino acid neurotransmitte~ efflux from adult cerebellum. After accumulation of radiolabeled amino acids, molecular layer homogenates received six 30 s washes with Krebs-bicarbonate medium containing 1 mM EGTA. After the sixth wash, filters received a 30 s pulse with one of the following solutions: (a) K~bs-bicar~nate medium con- taining 1 GM EGTA -(Baseline efftux); (b) Krebs- bicarbonate medium containing 56mM of KC1 and 2mM of CaCl,; or (c) Krebs-bicar&nate medium containing 56mM KC1 and 1 mM EGTA. Bars represent the
mean f SE of 8-10 determinations.
1 , 15
I , 1 ”
20 25 30 Adult
Age
(days after birth)
Fig. 4. Baseline efllux of [“Cjglutamic acid in the devel- oping arcbcllum. After loading with [“Qktamic acid, molecular layer homogenates were treated as described in Fig. 3. Baseline effiux is shown. Data represent the mean + SE of the number of determinations indicated in
parenthesis.
candidate is to show that it is released from nerve
endings in a Ca” -dependent manner when a depolar- izing stimulus is applied. Therefore. we studied the Ca’+dependent, depolarization induced release of [‘Qlutamic acid from molecular layer homogenates in the developing cerebellum.
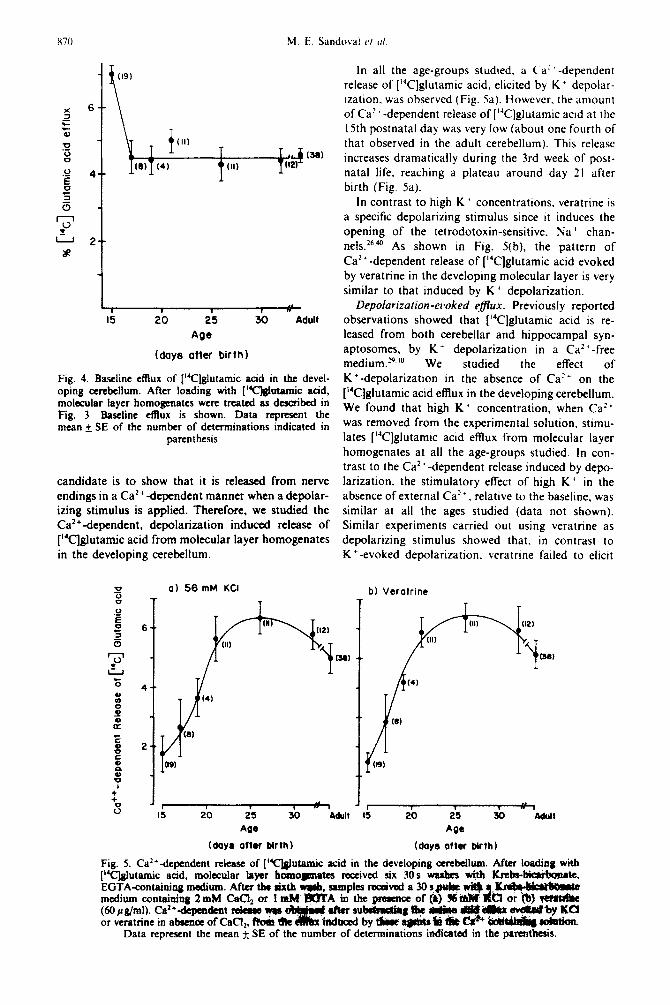
In all the age-groups studied. a C’a” -dependent
release of [“C]glutamic acid. elicited by K + depolar-
ization. was observed (Fig. Sa). However. the amount
of Ca? ’ -dependent release of [ “‘Clglutamic acid at the
15th postnatal day was very low (about one fourth of
that observed in the adult cerebellum). This release
increases dramatically during the 3rd week of posl-
natal life, reaching a plateau around day 21 after
birth (Fig. 5a).
In contrast to high K ’ concentrations. veratrine is
a specific depolarizing stimulus since it induces the
opening of the tetrodotoxin-sensitive. Na + chan-
nels.2”.“’ As shown in Fig. 5(b), the pattern of
Ca’+-dependent release of [“Clglutamic acid evoked
by veratrine in the developing molecular layer is very
similar to that induced by K’ depolarization.
Depolarization-eroked t$lux. Previously reported
observations showed that [‘4C]glutamic acid is re-
leased from both cerebellar and hippocampal syn-
aptosomes, by K- depolarization in a Ca’ ’ -free
medium.B.‘O We studied the effect of
K+-depolarization in the absence of Ca” on the
[“Clglutamic acid efflux in the developing cerebellum.
We found that high K ’ concentration, when Ca”
was removed from the experimental solution, stimu-
lates [“C]glutamic acid efflux from molecular layer
homogenates at all the age-groups studied. In con-
trast to the Ca2 + -dependent release induced by depo-
larization. the stimulatory effect of high K + in the
absence of external Ca”. relative to the baseline, was
similar at all the ages studied (data not shown).
Similar experiments carried out using veratrine as depolarizing stimulus showed that, in contrast lo
K +-evoked depolarization. veratrine failed to elicit
0 a) 56 mM KCI :: 1 T
Data represent the mean k SE of the number of determinations indicated in the parenthesis.
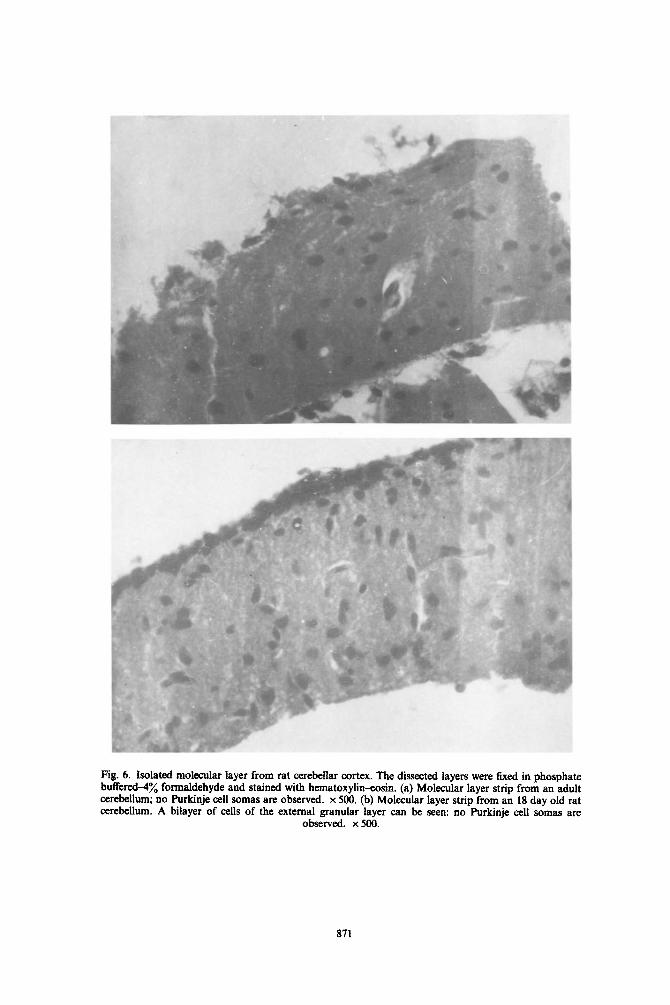
Ag. 6. Isolated molecular layer from rat cerebeilar cortex. The dissected Iayers were tixed in phosphate buffered+ formaldehyde and stained with hematoxylin-eosin. (a) Molecular layer strip from an adult cerebellum; no Purkinje cell somas are observed. x 500. (b) Molecular layer strip from an 18 day old rat cerebellum. A bilayer of cells of the external granular layer can be seen: no Purkinje cell somas are
observed. x 500.
871


Table 1. [‘WABA efeux from molaculiv layer of rat cerebeftum
Baseline Ca’+-dependent K+-evoked efflux release efflux
1 S-Days-old 3.34 * 0.34 2.37 f 0.32 2.60 f 0.44- (s) (81 (8)
Adult 3,I 1 f 0.35 3.03 1.0.50 3.80 f 0.70 (10) (9) GO)
After loading with [‘H]GABA, molecular layer homogenates were treated as described in Fig. 3. K+-evoked elllux was taken as the efftox of [‘HPABA induced by 56 mM KC1 in the absence of CaC&. Data represent the mean8 & SE of the number of dete~i~ationa indicated in parenthesis.
further [‘4C]glutamic acid efflux over the baseline efflux, when Ca2* was removed from the depolarizing solution (data not shown).
It has been described that the inhibitory neurons of the cerebellar cortex appear during the first week after birth and synapses of the steilate and basket cells are functional around the 12th postnatal day.2z*34;Jo Therefore, we studied the Ca2+dependent release of [3H]GABA, the putative inhibitory trans- mitter in these synapses,31*3sf11,4z in the molecular layer homogenates from 15 day old and adult cerebellum. Table 1 shows that Caz’-dependent release of PH]GABA evoked by 56mM KC1 is about 20% higher in the adult rat than that observed in the 15 day old cerebellar molecular layer. The baseline e&x and the K+-evoked f3H]GABA efflux were of the same magnitude in both 15 day old and adult molec- ular layer homogenates (Table 1).
Since ne~ochemical data obtained from the mo- lecular layer homogenates depends on the dissection of fragments of this tissue, a histological control was performed. As shown in Fig. 4(a), tissue fragments from adult cerebellar cortex employed in the present study only included elements of the molecular layer; however, in those obtained from an 18 day-old rat, a bilayer of cells corresponding to the external gran- ular layer could be ciearly identified (Fig. 6b). Most of the strips examined showed neither Purkinje cells nor internal granular layer.
DISCUSSION
This study shows that in the developing rat cere- bellum, high affinity uptake of glutamic acid precedes the development of the Ca*+-dependent release mech- anisms for this amino acid. [‘4Clglutamic acid is highly accumulated by a Na*-dependent, high affinity transport system by Pz fractions from 15 day old rat cerebellum. In the 3rd postnatal week, uptake decreased abruptly and remained constant until the adult age. Our data showed a decrease in the trans- porting sites rather than allinity for the neu- rotransmitter during maturation of the cerebellar cortex (Fig. 2). These results are in agreement with previous studies which indicate that the degree of GABA uptake in the developing rat brain and that of choline by chick embryo retina during development are greater in younger animals.30,3q Gliai structures
have heen reported as the principal sites of uptake of acidic amino acids in the newborn” and adult cere- bellum.10~23~* The accumulation of glutamic acid by perikarya, dendrites and axons of the putatively ~ut~a~r~~ granule cells is very to~.‘~*“~~ In the newborn rat, glial processes are first seen on the 7th postnatal day and become extensive about IO-12 days after birth; by the 15th day they cover the secondary and tertiary dendrites in the lower molec- ular layer. In general, when synapses are present on the processes of Purkinje cells, they are partially or completely ensheathed by glia, su~esting that at any region the establishment of synapses is soon followed by glial insulation. Thus it is likely that a change in the high affinity uptake of glutamic acid in the cerebellar cortex may reflect a change in the aceumu- lation of this amino acid by glial cells. The reduced glutamic acid uptake observed after the 3rd postnatal week may be due to the disappearance of transient synapses and its respective glial processes during development.’ On the other hand, it is possible that the enhanced glutamic acid uptake found at early ages may reflect activity of the cells of the external granular layer, which population drastically de- creases towards the 21st postnatal day,’ and no longer exists in the adult cerebeilar cortex (Fig. 6).
This work confirms previous observa- tions1a~13,14.20.28,3z showing that glutamic acid and GABA are released from cerehellar preparations in a way similar to the stimul~~~etion coupling system described for well known n~urotransmitte~.’ This release was not observed for [‘4Qspartic acid. These results are in agreement with the reported data of Foster and Roberts14 and Levi et al.” that showed Ca2+-dependent release of both endogenous glu- tamate and GABA from cerebellar slices, whereas the release of aspartic acid was Cat+-independent. How- ever, several reports indicate that the release of endogenous aspartic acid from cerebellar slices evoked by K+, decreased after calcium ions were removed from the depolarizing solution.‘S*42 Al- though this discrepancy has not been investigated, in this study we used exogenous [‘“cjaspartic acid that can be accumulated by glutamate@ nerve endings which do not release it. These observations strengthen the suggestion of a transmitter role for glutamic acid and GABA in the cerebellum, whereas the role of aspartic acid remains unclear.
In contrast to high affinity uptake, the CY+-dependent refease system for glutamic acid de- velops in a progressive fashion, similar to that ob- served for other transmitters in the nervous sys- tem.‘6,‘qS30*39 Since the amount of ~dioa~ti~ty that remained as glutamic acid in both molecular layer homo~nat~s and perfusates from 15 day dd and adult cerebellum was similar, it is unhkely that the time course of development of glutamic acid release may be due to the different rate of the amino acid metabolism by the tissue samples at different ages. Thus, we suggest that in the immature cerebellum the

x74 M, E. Sandoval 01 (I/.
recently accumulated glutamic acid is not stored in a functionally releasable pool.
Possible functional implicutions
This study provides further evidence to support the
idea that glutamic acid is the transmitter of the parallel fibers. The profile of Ca2+ -dependent release of [‘4C]glutamic acid from cerebellar molecular layer during development clearly follows the time course of parallel fibers synaptogenic activity. Glutamic acid release was poorly represented at IS days after birth, at which time few parallel fiber synapses are ob- Served;2.K” the climbing fiber synapses, the other excitatory input to the Purkinje cells, are already developed at this age.6.?17.29.17.44 Therefore, Ca’+-dependent release of glutamic acid is mainly related to the formation of synapses by the granule cells. The higher baseline efflux of glutamic acid observed at day I5 after birth, may represent a poor retention of the exogenous transmitter by immature glutamatergic synapses or may be due to glial efflux.
The Ca” -dependent release of [-‘H]GABA, the postulated neurotransmitter of inhibitory neurons of the cerebellar cortex,‘8,2’.‘5,‘“.‘X was similar in the molecular layer homogenates from I5 day old and
adult cerebellum. Thus, the protile of development of glutamic acid release observed during maturation of the cerebellar cortex may represent a specific pattern for maturation of glutamatergic synapses.
It has been reported that the formation of glutamic acid binding sites. that may correspond to the recep- tor for the transmitter. occurs approximately at the same time as the formation of the synapses between parallel fibers and the dendritic spines of the Purkinje cell~.~ The amount of specific binding sites increases during the 2nd and 3rd week of postnatal life,* at which time we observed the larger increment of glutamic acid release (Fig. 5). These results show a clear relationship between the synaptogenic activity of the cerebellar granule cells and the critical bio- chemical events that lead to a functional synapse: transmitter uptake and liberation as well as synthesis of specific postsynaptic receptors.
Acknowledgemenrs--This work was supported in part by CONACyT Grant PCCBNAL 790298..We are grateful to Drs R. Lara. H. Pasantes-Morales and R. Tapla for their critical comments on the manuscript, to Dr. A. Feria- Velasco and M. D. C. Beas-Zarate for their assistance in the histological studies and to Miss V. Diaz de Castro for secretarial assistance.
I.
2.
3 .
4.
5.
6. 7.
8.
9.
10.
Il.
12. 13.
14.
IS. 16.
17.
18.
19.
20.
REFERENCES
Altman J. (1972) Postnatal development of the cerbellar cortex in the rat: I. The external germinal layer and the transitional molecular layer. J. camp. Neural. 145, 353-398. Ahman J. (1972) Postnatal development of the cerebellar cortex in the rat: II. Phases in the maturation of Purkinje cells and the molecular layer. J. camp. Neural. 145, 399-464. Altman J. (1972~) Postnatal development of the cerebellar cortex in the rat: III. Maturation of the components of the granular layer. J. camp. Neural. l& 465-514. Chuio T.. Yamada Y. and Yamamoto C. (1975) Sensitivitv of Purkinie cell dendrites to nlutamic acid. Exol Brain Res. 23, i93-300.
r
Cotman C. W., Haycock J. W. and White W. F. (1976) Stimulus-secretion coupling processes in brain: analysis of noradrenaline and gamma-amino-butyric acid release. J. Physiol., Land. 254, 475-505. Crepe1 F. (1971) Maturation of the climbing fiber responses in the rat. Brain Res. 35, 272-276. Crepe1 F. (I 974) Excitatory and inhibitory processes acting upon cerebellar Purkinje cells during maturation in the rat: influence of hypothyroidism. Expi Brain Res. 20, 403-420. _ De&in J. C. (1974) Histoloaical evidence supwrtina the inferior olive as the maior source of cerebellar climbinn fibers in the rat. &in ies. 77, 365-384. . * -
Y
de Barry J.. Vincedon G. and Gombos G. (1980) High affinity glutamate binding during postnatal development of rat cerebellum. FE&S Lerr. 199, 175179. de Barry J., Langley 0. K., Vincedon G. and Gombos G. (1982) tglutamate and L-glutamine uptake in adult rat cerebellum: an autoradiographic study. Neuroscience 7, 1289-1297. East.M. J., Dutton G. R. and Currie N. D. (1980) Transport of GABA, /I-alanine and glutamine into perikarya of postnatal rat cerebellum. J. Neurochem. 34, 523-530. Eales J. C., Ito M. and Szentagothai J. (1967) The Cerebellum us a Neuronal Machine. Springer, New York. Flint R. S., Rea M. A. and McBride W. J. (1981) In vitro release of endogenous amino acids from granule cell-stellate cell- and climbing fiber-deficient cerebella. /. Neurochem. 37, 1425-1430. Foster A. C. and Roberts P. J. (1980) Endogenous amino acid release from rat cerebellum in oirro. J. Newochem. 35, 517-519. Fricke U. (1975) Tritosol: a new scintillation cocktail based on Triton X-100. Analyf. Eiochem. 63, 555-558. HolIytieId J. G., Raybom M. E., Satthy P. V. and Lam D. M. K. (1979) The emergence, locahzation and maturation of nemotransmitter systems during development of the retina in Xenopus latwis. I. Y-aminobutytic acid. J. camp. Neurol. 188, 587-598. Knowles W. D. and Phillips M. 1. (1980) Neurophysiologlycal and behavioral maturation of cerebellar function studied with tremorigenic drugs. Neuropharmacology 19, 745-756. Kurivama K.. Haber B.. Sisken B. and Roberts E. (1966) The y-aminobutyric acid system in the rabbit cerebellum. Proc: natn. Acad. Sci. (Wash.) SS, 846-852. Lam D. M. K., Fuog S-C. and Kong Y. C. (1980) Postnatal development of GABA-ergic neurons in the rabbit retina. J. camp. Neural. 193, 89-102. Levi G., GOKIOLI R. D., Gal10 V., Wilkin G. P. and B&us R. (1982) Putative acidic amino acid transmitters in the cerebellum I. Depolarization-induced release. Brain Res. 239, 42H45.

Glutamic acid in the developing cerebellum 875
21. Levy W. B., Redburn D. A. and Cotman C. W. (1973) Stim~us~upl~ secretion of y-~nobut~c acid from rat brain s~apto~m~. Science, N.Y. 181, 676678.
22. Lowry 0. H., Rosebrough N. J., Farr A. L. and Randall R. J. (1951) Protein rn~s~~ent with the Folin phenol agent. J. biol. Chem. 193, 265-275.
23. McBride W. J., Nadi N. S., Altman J. and Aprison M. H. (1976) Effects of selective doses of X-irradiation on the levels of several amino acids in the cerebellum of the rat. Neurochem. Res. I_, 141-152.
24. Mikoshiba K. and Changeux J. P. (1978) Morphological and biochemtcal studies on isolated molecular layers from bovine cerebellum. Bruin Res. 142, 487-504.
25. Minchin M. C. W. (1975) Factors influencing the efflux of [‘HIgamma-aminobutyric acid from satellite glial cells in cat sensory ganglia. J. Neurochem. 24, 571-577.
26. Ohta M., Narahashi T. and Keeler R. F. (1973) Effects of veratrium alcaloids on membrane potential and conductance on squid and crayfish giant axons. J. Pharmuc. exp. Ther. 184, 143-154.
27. Palay S. L. and Chan-Palay V. (1974) Cerebellar Cortex Cytology and Organization. Springer, Berlin. 28. Pearce B. R., Currie N, D., Beale R. and Dutton G. R. (1981) Potassium stimulated, calcium-dependent release of
[3H]GABA from neuron and glia enriched cultures of cells dissociated from rat cerebellum. Brain Res. Ws, 485-489. 29. Puro D. G. and Woodward D. J. (1977) Maturation of evoked climbinrr fibers innut to rat cerebellum Purkinie c&s.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40. 41.
42.
43.
44.
45.
46.
(I) EXpr Brain Res. 28, 85-100. ~ ‘ Redburn D. A., Broome D., Ferkany J. and Enna S. J. (1978) Development of rat brain uptake and ~lci~-d~d~t release of GABA. Brain Res. 152, 511-519. Rhode B. H., Rea M. A., Simon J. R. and McBride W. J. (1979) Effects of X-irradiation induced loss of cerebellar granule cells on the synaptosomal levels and the high affinity uptake of amino acids. J. Neurochem. 32, 1431-1435. Sandoval M. E. and Cotman C. W. (1978) Evaluation of glutamate as a neurotransmitter of cerebellar parallel fibers. Neuroscience 3, 199-206. Sandoval M. E., Horch P. and Cotman C. W. (1978) Evaluation of glutamate as a hippocampal neurotransmitter: glutamate uptake and release from synaptosomes. Brain Res. 142, 285-299. Schon F. and Iversen L. L. (1972) Selective accumulation of [3H]GABA by stellate cells in the rat cerebellar cortex in vivo. Brain Res. 42, 503-507. Sellstrom A. and Hamberger A. (1976) Gamma-aminobutyric acid release from neurons and glia. Acta physiof. stand. 98, 94-102. Sellstrom A. and Hamberger A. (1977) Potassium-stimulated y-aminobutyric acid release from neurons and glia. Brain Res. 119, 189-198. Shimono T., Nosaka S. and Sasaki K. (1976) El~~ophysiolo~cal study on the postnatal development of neural mechanisms in the rat cerebellar cortex. Brain Res. loS, 279-294. Stow-Ma~isen J. (1976) Distribution of the components of the GA3A system in neuronal tissue: Cerebellum and hippocampus-effects of axotomy. In GABA in Nervous System Function (eds Roberts E., Chase T. N. and Tower D. B.), pp. 149-168. Raven Press, New York. Thompson J. M. (1982) Increase of acetylcholine release from chick embryo retina during development. Deal Brain Res. 4, 259-264. Ulbricht W. (1969) The effect of veratridine on excitable membranes of nerves and muscles. Ergebn. Physiol. 61, 18-71. Valcana T., Hudson D. and Timiras P. S. (1972) Effects of the X-irradiation on the content of amino acids in the developing rat cerebellum. J. Neurochem. 19, 2229-2232. Wiklund L., Toggenburger G. and Cuenod M. (1982) Aspartate: possible neurotransmitter in cerebellar climing fibers. Science, N. Y. 216, 78-80. Wilkin G. P., Garthwaite J. and Balazs R. (1982) Putative acidic amino acid transmitters in the cerebellum. II. Electron microscopic localization of transport sites. Brain Res. 244, 69-80. Woodward D. J., Hoffer R. J., Siggins G. R. and Bloom F. E. (1971) The ontogenic development of synaptic junctions, synaptic activation and responsiveness to neurotrans~tter substances in rat cerebellar Purkinje cells. Brain Res. 34, 73-97. Wo~ward D. J., Hoffer R. J., Siggins G. R. and Oliver A. P. (1976) Inhibition of Purkinje cells in the frog cerebellum. II. Evidence for GABA as the inhibitory transmitter. Bruin Res. 33, 91-100. Young A. B., Oster-Granite M. L., Hemdon R. M. and Snyder S. H. (1974) Glutamic acid: selective depletion by viral induced granule cells loss in hamster cerebellum. Brain Res. 73, 1-13.
(Accepted 1 September 1983)




















