
Active multistage coarsening of actin networks driven by myosin motors
Upload
independentCategory
view
6download
0
Heterogeneity of myosin heavy-chain expression in fast-twitch fiber types of mature avian pectoralis muscle1
B.W.C. Rosser, M. Wick, D.M. Waldbillig, and E. Bandman
Abstract: The aims of this study are to investigate the diversity of myosin heavy-chain (MyHC) expression among avian fast- twitch fibers, and to test the hypothesis that dissimilar MyHC isoforms are found in each of the principal avian fast-twitch fiber types. MyHCs within the muscle fibers of the pectoralis of 31 species of bird are characterized using immunocytochemical methods. A library of 11 monoclonal antibodies previously produced against chicken MyHCs is used. The specificity of these antibodies for MyHCs in each of the muscles studied is confirmed by Western blots. The results show that avian fast-twitch glycolytic fibers and fast-twitch oxidative-gylcolytic fibers can contain different MyHCs. Among the species studied, there is also a conspicuous variety of MyHC isoforms expressed. In addition, the results suggest that two epitopes are restricted to chickens and closely allied gallinaceous birds. There are no apparent correlations between MyHC epitope and presupposed contractile properties. However, the presence of different isoforms in different fast-twitch fiber types suggests a correlation between isoform and contractile function.
Key words: muscle, fiber, myosin, avian.
RbumC : Le but de ce travail Ctait d'ktudier la diversit6 de l'expression de la chaine lourde de myosine dans les fibres rapides d'oiseaux et de verifier l'hypothkse que diffkrentes isoformes de la chaine lourde de myosine se trouvent dans chacun des principaux types de fibres rapides d'oiseaux. Les chaines lourdes de myosine des fibres de muscles pectoraux de 31 espkces d'oiseaux ont CtC caractCris6es par des mtthodes immunocytochimiques. Une banque de 11 anticorps monoclonaux dirigCs contre les chaines lourdes de myosine du poulet a tt6 utiliske. La sp6cificitC de ces anticorps envers les chaines lourdes de myosine dans chacun des muscles CtudiBs a CtC confirmBe par des transferts Western. Les rCsultats indiquent que les fibres rapides B glycolyse anabobie et les fibres rapides B glycolyse aCrobie d'oiseaux contiennent diffkrentes chaines lourdes de myosine. 11 y a une grande variCtC d'isoformes de chaine lourde de myosine exprimCes chez les espbces d'oiseaux CtudiCes. De plus, les rCsultats suggkrent que deux Cpitopes se trouvent seulement chez le poulet et les autres gallinaces trks apparent& au poulet. I1 n'y a pas de corrklation apparente entre un Cpitope de la chaine lourde de myosine et les propriBt6s contractiles prtsumCes. Cependant, la prCsence de diffkrentes isoformes dans diffkrents types de fibres rapides suggkre qu'il y a une corrtlation entre les isoformes et la capacitt de contraction.
Mots clis : muscle, fibre musculaire, myosine, oiseau.
[Traduit par la rkdaction]
Introduction
Myosin, a filamentous protein, is intimately involved in the contraction of all skeletal muscles (Sellers and Goodson 1995;
Received August 12, 1996. Revised October 16, 1996. Accepted October 23, 1996.
Abbreviations: MyHC, myosin heavy-chain; FOG, fast-twitch oxidative glycolytic; FG, fast-twitch glycolytic; SDS-PAGE, sodium dodecyl sulfate - polyacrylamide gel electrophoresis.
B.W.C. ~ o s s e 9 and D.M. Waldbillig. University of Saskatchewan, College of Medicine, Department of Anatomy and Cell Biology, Saskatoon, SK S7N 5E5, Canada. M. Wick and E. Bandman. University of California, Department of Food Sciences and Technology, Davis, CA 95616, U.S.A.
This paper is dedicated to Dr. John C. George, University of Guelph, in recognition of his pioneering studies of avian muscle fiber types and in honour of his seventy-fifth birthday. Author to whom all correspondence should be addressed.
Voet and Voet 1995). In muscle, each myosin molecule is a hexamer consisting of two heavy and four light chains. MyHCs, with molecular weights of approximately 200 000, contribute more to skeletal muscle mass than any other myo- fibrillar protein (Yates and Greaser 1983; Voet and Voet 1995). In vertebrates, MyHCs are encoded by a diverse multi- gene family (see Bandman et al. 1994) and there are many bio- chemically distinct but related MyHC isoforms (Bandman et al. 1990,1994).
MyHC expression has been studied in an impressive vari- ety of mammalian skeletal muscles and species (see Hoh 1992; Hamalainen and Pette 1993; Schiaffino and Reggiani 1994; Stal et al. 1994). By comparison, there are fewer studies of MyHCs in avian skeletal muscles. Moreover, studies of avian MyHCs have been confined to several muscles of the domestic chicken (Bandman et al. 1990; Williams and Dhoot 1992; Connold et al. 1993; Gauthier and Orofanos 1993; Maier 1994) and their close allies, the turkey (Maruyama et al. 1993; Cherel et al. 1994) and Japanese quail (Merrifield et al. 1989; Alway 1993).
Biochem. Cell Biol. 74: 715-728 (1996). Printed in Canada I Imprimd au Canada

Biochem. Cell Biol. Vol. 74, 1996
Muscle fibers, the contractile cells of skeletal muscle, are broadly classified as either fast twitch or slow, depending on their contractile properties (Rosser and George 1986; Will- iams and Dhoot 1992). Vertebrate skeletal muscles consist of varying proportions of different types of fibers (Peters 1989; Hoh 1992). In the earliest stages of development, fast-twitch and slow muscle fibers contain the same embryonic MyHC isofoms (Bandman et al. 1990, 1994; Russell et al. 1993; Schiaffino and Reggiani 1994). During subsequent stages, each type of fiber may contain a number of different MyHC isofoms (Bandman et al. 1990, 1994; Russell et al. 1993; Schiaffino and Reggiani 1994). In mature vertebrate muscles, fast-twitch and slow fibers contain different MyHC isofoms, although there may be uncommon transitional fiber types con- taining mixtures of slow and fast isoforms (Bandman et al. 1990, 1994; Schiaffino et al. 1990; Russell et al. 1993; Staron and Pette 1993). In mature mammalian muscles, three fast- twitch fiber types have been identified and each of these prin- cipal fiber types contains a characteristic MyHC isoform (Hoh 1991; Harnalainen and Pette 1993; Bottinelli et al. 1994; Schi- affino and Reggiani 1994). In avian muscle, while several dis- tinct isoforms can also appear sequentially within the same fast-twitch fiber during development (Bandman et al. 1990, 1994), different MyHCs have not been described among the principal fast-twitch fiber types of mature muscle (Menifield 1989; Williams and Dhoot 1992).
The purpose of this study is to explore the diversity of MyHC expression among avian fast-twitch fibers and to test the hypothesis that the principal avian fast-twitch fiber types differ from each other in their MyHC content. MyHCs in fibers of the pectoralis muscle of 3 1 species of bird are charac- terized using a library of monoclonal antibodies previously generated against known chicken MyHCs. The specificity of these antibodies for MyHCs in the muscles studied is verified by Western blots. Our results indicate that there is a pro- nounced diversity in the MyHC isofoms expressed and that different avian fast-twitch fiber types do contain distinct MyHCs.
Materials and methods
Fast-twitch fiber nomenclature In mature mammalian muscles, three fast-twitch fiber types have been identified: type 2A, type 2X (also called 2D), and type 2B (Hoh 1991; Hamalainen and Pette 1993; Schiaffino and Reggiani 1994). In this scheme, fiber type determined by MyHC content has been correlated with physiologic perfor- mance (Bottinelli et al. 1994). Metabolic criteria, however, have also been incorporated into fast-fiber terminology (see Nemeth 1990) and correlated with physiologic performance (Nemeth et al. 1991). The FOG and FG fibers of the metabolic nomenclature are, respectively, largely synonymous with type 2A and type 2B of the myosin-based scheme (see Nemeth 1990). It has been suggested that past discrepancies between these two schemes were due to the presence of type 2X fibers, of which earlier investigators were unaware (Schiaffino et al. 1990).
Avian fast-twitch fiber nomenclature has most often incor- porated metabolic criteria (George and Berger 1966; Rosser and George 1986; Kaplan and Goslow 1989; Lundgren and Kiessling 1988; Gaunt et al. 1990; Boesiger 1992; Torrella et
al. 1993; Tobalske 1996). Avian type FOG and FG fibers are, respectively, for the most part synonymous with type 2A and type 2B fibers (see Rosser and George 1986). To date, type 2X fibers have not been described in avian muscle. Therefore, in this study, we have chosen to continue to include metabolic criteria to describe fast-twitch fibers as either FG or FOG.
Avian FOG and FG fibers are also synonymous with, respectively, red and white fast-twitch fiber types (see George and Berger 1966; Rosser and George 1986). An intermediate fiber type has also been described. These intermediate fibers are between red and white fibers in their metabolic activities and morphologic characteristics (see George and Berger 1966; Rosser and George 1986). In mammalian muscles, a complete spectrum of overlapping metabolic activities has been described among fast-twitch fibers (see Nemeth 1990), and some of these fibers may represent transitional types contain- ing mixtures of different MyHCs (Staron and Pette 1993). The avian fast-twitch intermediate type merits attention because it has been described as a discrete population meeting defined functional demands in flight muscles (Parker and George 1975; Tobalske 1996). However, unless otherwise indicated, in this study for simplicity we refer to both avian intermediate and red fibers as FOG subtypes.
Experimental model The pectoralis muscle (M. pectoralis pars thoracicus) of birds extends from the sternum, clavicle, and ribs to the humerus (Raikow 1985). The pectoralis is the most massive muscle in the body of volant species (Hartman 1961). It provides almost all power for the downstroke of the wing in flight and assists in decelerating the wing during the upstroke (Biewener et al. 1992; Dial 1992).
The fiber type complement of the pectoralis muscle is gen- erally correlated with a species' mode of flight (George and Berger 1966; Rosser and George 1986; Rosser et al. 1994; Tobalske 1996). We have previously reported the proportion and distribution of fiber types in the pectoralis of some 44 spe- cies of birds (Rosser and George 1986; Rosser et al. 1987). Subsequently, additional observations have been published (Edman et al. 1988; Lundgren and Kiessling 1988; Kaplan and Goslow 1989; Gaunt et al. 1990; Boesiger 1992; Evans et al. 1992; Swain 1992; Torrella et al. 1993; Tobalske 1996). Invariably, in those birds that rely upon flapping flight, the pectoralis is composed almost exclusively (99-100%) of fast- twitch fiber types.
Slow fibers are found sparingly within the avian pectoralis muscle. They are present within a tiny narrow band, termed the deep red strip, which has been located in the deepest cra- nial region of the muscle in just four species: chicken, Japa- nese quail, double-crested cormorant, and red-tailed hawk (Rosser and George 1986; Rosser et al. 1987; Shear et al. 1988). Also, in certain species using soaring flight (such as vultures and pelicans), the pectoralis has an accessory deep belly consisting entirely of slow fibers (Rosser et al. 1994). In addition, flightless forms (such as the kiwi, emu, and ostrich) have comparatively diminutive pectoralis muscles, which contain a high proportion of slow fibers throughout (see Rosser and George 1985).
MyHC expression within the pectoralis muscle of the domestic chicken is fairly well understood (Bandman et al. 1990; Gauthier and Orfanos 1993). The muscle consists

Rosser et al. 717
Table 1. Specificity of antiMyHC monoclonal antibodies against chicken MyHCs.
Fast Slow Monoclonal antibody* Embryonic* * Neonatal Adult 1 2 Ventricular Apparent specificity
NA4 + + + + + + All myosins AG6 + + + - - - All fast myosins EB 165 + - + - - - Embryonic and adult fast AB8 - - + - - - Adult fast
2E9 - + - - - - Neonatal fast B 103 - + + - - - - Embryonic and neonatal fast NA1 - - - - + - Slow 2
NA2 - - + + - Slow 1 and 2 -
NA7 - - + - - Slow 1 NA8 - - - - + - Slow 2 H V l l - - - - - + Ventricular
*The preparation and specificity of these monoclonal antibodies against chicken MyHCs (myosin heavy chains) has been detailed elsewhere (Cemy and Bandman 1987; Bandman and Bennett 1988; Bourke et al. 1991).
**Embryonic fast refers to multiple isoforms; B103 labels embryonic 1 or 3 but not embryonic 2 (Moore et al. 1992).
almost entirely of fast fibers except for the deep red strip that comprises less than 1 % of the muscle and contains about one- third slow fibers (Shear et al. 1988). Throughout the muscle, excepting the red strip, embryonic MyHC isoforms are entirely replaced by a neonatal isoform at about 2 weeks after the birds hatch. This neonatal isoform is in turn wholly replaced by an adult isoform by about 4 months of age. A com- parable embryonic to neonatal to adult MyHC transformation also occurs in the pectoralis muscle of the Japanese quail (Merrifield et al. 1989) and the turkey (Maruyama et al. 1993). However, a different sequence of MyHC transformations takes place within the fast-twitch fibers located in the red strip (Shear et al. 1988), as well as within fast-twitch fibers of other muscles of the chicken (Bandman et al. 1990).
Species Studied The nomenclature for the 3 1 species studied is, unless other- wise indicated, from Sibley and Munroe (1990). The species studied were as follows: American crow (Corvus brachyrhyn- chos), American robin (Turdus migratorius), American white pelican (Pelecanus erythrorhynchos), American wigeon (Anas americana), belted kingfisher (Megaceryle alcyon), Boe- hemian waxwing (Bombycilla garrulus), common or island canary (Serinus canaria), domestic chicken (Gallus gallus; from Crawford 1990), domestic or rouen duck (Anas platy- rhynchos; from Crawford 1990), domestic or rock pigeon (Columba livia), great homed owl (Bubo virginianus), grey partridge (Perdix perdix), homed lark (Eremophila alpestris), house wren (Troglodytes aedon), Japanese quail (Coturnix japonica), least sandpiper (Calidris minutilla), long-eared owl (Asio otus), merlin (Falco columbarius), mulga parrot (Pse- photus varius), northern flicker (Colaptes auratus), northern harrier (Circus cyaneus), northern shoveller (Anas clypeata), ostrich (Struthio camelus), red-eyed vireo (Vireo olivaceus), red-winged blackbird (Agelaius phoeniceus), rufous-sided towhee (Pipilo erythrophthalmus), short-eared owl (Asiojlam- meus), sora rail (Porzana carolina), stilt sandpiper (Micro- palama himantopus), Swainson's hawk (Buteo swainsoni), and zebra finch (Taeniopygia guttata).
All birds were judged to be mature (adult) individuals, with the exceptions of the ostrich (2 months), house wren (newly fledged), and great homed owl, which were sexually immature (juveniles).
Specimens and tissue preparation Blocks of muscle were excised from the more superficial areas of either the right or left pectoralis muscle of each bird. Blocks were removed from a location near the keel of the sternum, about midway between the cranial and caudal edges of the muscle. As in our earlier work (Rosser and George 1986; Rosser et al. 1987), samples were obtained from the sternobra- chialis and not the thoracobrachialis portion (Dial 1992) of each muscle. Each block was about 0.6 x 0.6 x 3 cm and was cut so that its long axis ran in the same direction as the muscle fibers. In the smallest birds studied, a block could contain most of the muscle. Blocks of muscle were coated with Tis- sue-Tek O.C.T. compound (Miles Inc., Elkhart, Ind.), quick frozen in 2-methylbutane cooled to - 160°C by liquid nitro- gen (Dubowitz 1985), and then stored at -80°C. From tissue blocks representative of each bird, sections of 4-6 p (1 p. = 1 p,m) thickness were cut in a cryostat maintained at -20°C. Two to three sections were picked up on each microscope slide coated with gelatin-chrome alum. An extensive series of serial sections was obtained from each muscle.
Histochemistry Avian fast-twitch and slow fibers are readily differentiated on the basis of their myosin ATPase activity (Rosser and George 1986). Avian fast-twitch fibers are further divided into FG and FOG types by the biochemical pathways used to produce the energy for contraction (Rosser and George 1986). A his- tochemical stain for a mitochondria1 oxidative enzyme is com- monly used for this purpose (Rosser and George 1986). Predictably, mainly as a result of the greater density of mito- chondria in oxidative fibers, the smaller diameter fibers stain dark and the larger fibers stain light (George and Berger 1966). These fibers are then classified, respectively, as FOG and FG. For the purposes of this study, the FOG fibers can
71 8 Biochem. Cell Biol. Vol. 74, 1996
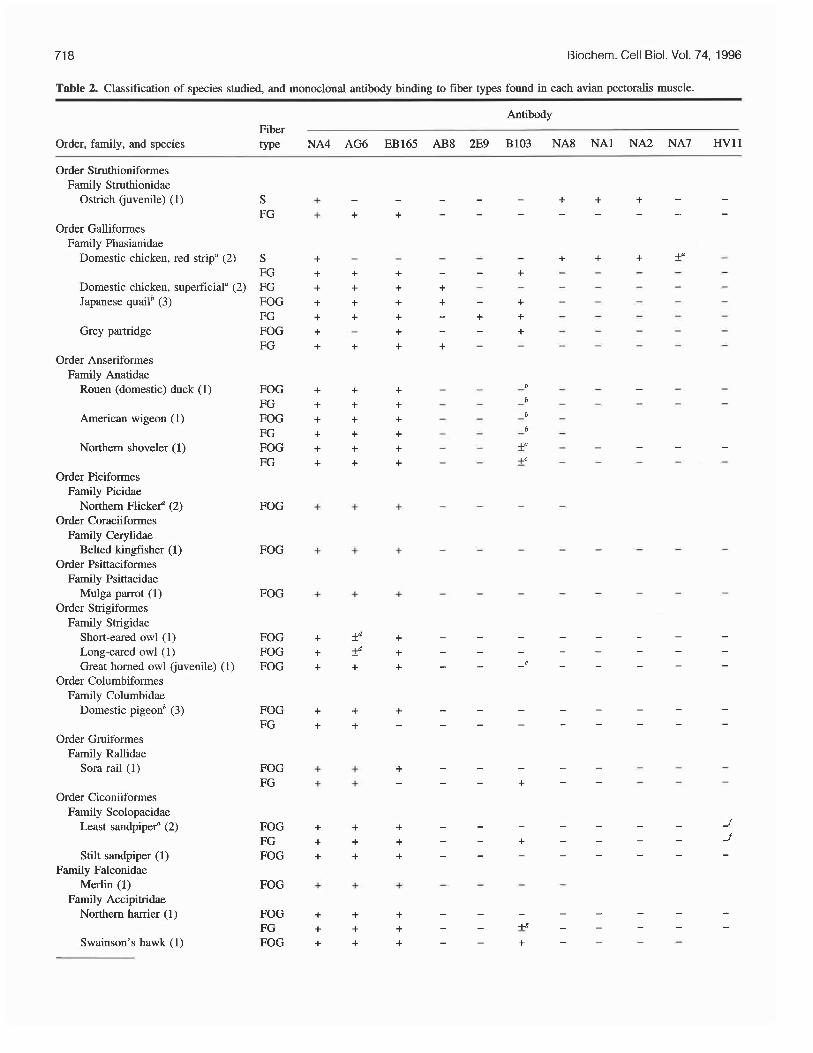
Table 2. Classification of species studied, and monoclonal antibody binding to fiber types found in each avian pectoralis muscle.
Antibody Fiber type NA4 AG6 EB165 AB8 2E9 B103 NA8 NA1 NA2 NA7 HVll Order, family, and species
Order Struthioniformes Family Struthionidae
Ostrich (juvenile) (1)
Order Galliformes Family Phasianidae
Domestic chicken, red strip" (2) S + - - FG + + + FG + + + FOG + + + FG + + + FOG + - + FG + + +
Domestic chicken, superficiala (2) Japanese quailb (3)
Grey partridge
Order Anseriformes Family Anatidae
Rouen (domestic) duck (1) FOG + + + FG + + + FOG + + + FG + + + FOG + + + FG + + +
American wigeon (1)
Northern shoveler (1)
Order Piciformes Family Picidae
Northern Flickef (2) Order Coraciiformes
Family Cerylidae Belted kingfisher (1)
Order Psittaciformes Family Psittacidae
Mulga parrot (1) Order Strigiformes
Family Strigidae Short-eared owl (1) Long-eared owl (1) Great homed owl (juvenile) (1)
Order Columbiformes Family Columbidae
Domestic pigeonb (3)
FOG + + +
FOG + + +
FOG + -C -t
FOG + fd + FOG + P + FOG + + +
FOG FG
Order Gruiformes Family Rallidae
Sora rail (1) FOG FG
Order Ciconiiformes Family Scolopacidae
Least sandpipef (2) FOG FG FOG Stilt sandpiper (1)
Family Falconidae Merlin (1)
Family Accipitridae Northern harrier (1)
FOG
FOG FG FOG Swainson's hawk (1)
Rosser et al. 719
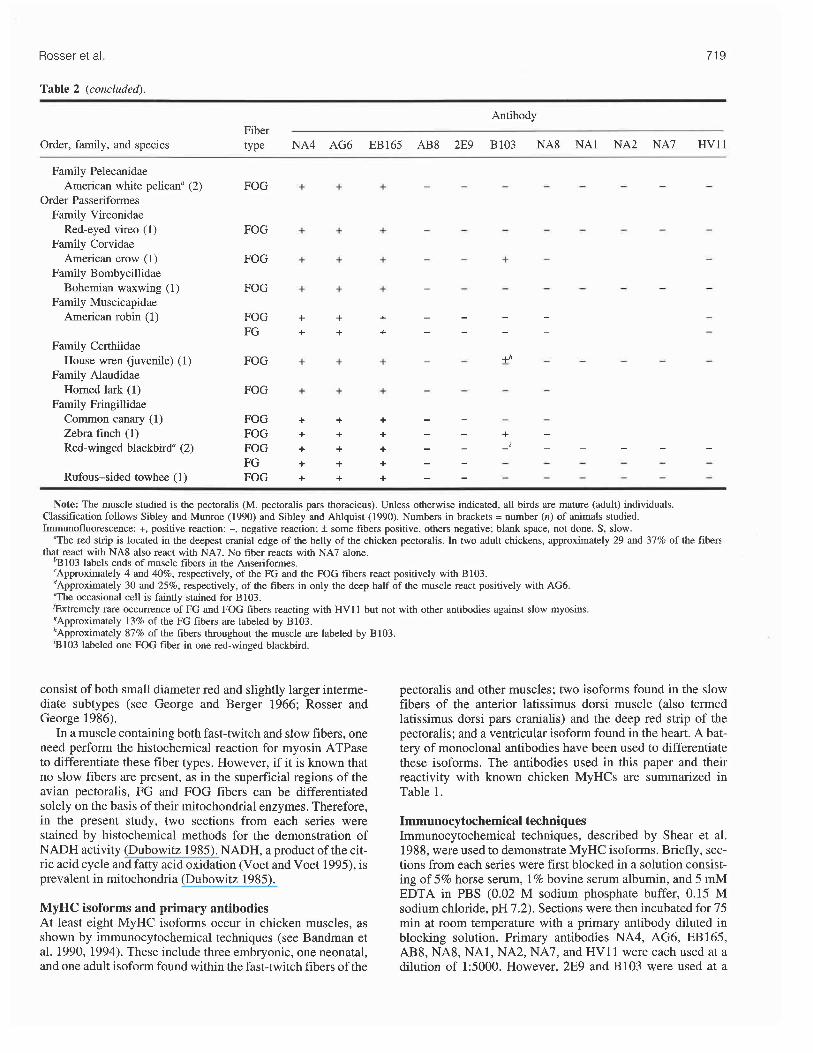
Table 2 (concluded).
Antibody Fiber
Order, family, and species type NA4 AG6 EB165 AB8 2E9 B103 NA8 NA1 NA2 NA7 HVll
Family Pelecanidae American white pelicana (2)
Order Passeriformes Family Vireonidae
Red-eyed vireo (1) Family Corvidae
American crow (1) Family Bombycillidae
Bohemian waxwing (1) Family Muscicapidae
American robin (1)
FOG
FOG
FOG
FOG
FOG FG
Family Certhiidae House wren (juvenile) (1)
Family Alaudidae Homed lark (1)
Family Fringillidae Common canary (1) Zebra finch (1) Red-winged blackbird" (2)
FOG
FOG
FOG FOG FOG FG FOG Rufous-sided towhee (1)
Note: The muscle studied is the pectoralis (M. pectoralis pars thoracicus). Unless otherwise indicated, all birds are mature (adult) individuals. Classification follows Sibley and Munroe (1990) and Sibley and Ahlquist (1990). Numbers in brackets = number (n) of animals studied. Immunofluorescence: +, positive reaction; -, negative reaction; f some fibers positive, others negative; blank space, not done. S, slow.
"The red strip is located in the deepest cranial edge of the belly of the chicken pectoralis. In two adult chickens, approximately 29 and 37% of the that react with NA8 also react with NA7. No fiber reacts with NA7 alone.
%103 labels ends of muscle fibers in the Anseriformes. 'Approximately 4 and 40%, respectively, of the FG and the FOG fibers react positively with B103. dApproximately 30 and 25%, respectively, of the fibers in only the deep half of the muscle react positively with AG6. 'The occasional cell is faintly stained for B103. Gxtremely rare occurrence of FG and FOG fibers reacting with HVI 1 but not with other antibodies against slow myosins. gApproximately 13% of the FG fibers are labeled by B103. hApproximately 87% of the fibers throughout the muscle are labeled by B103. 3103 labeled one FOG fiber in one red-winged blackbird.
fibers
consist of both small diameter red and slightly larger interme- diate subtypes (see George and Berger 1966; Rosser and George 1986).
In a muscle containing both fast-twitch and slow fibers, one need perform the histochemical reaction for myosin ATPase to differentiate these fiber types. However, if it is known that no slow fibers are present, as in the superficial regions of the avian pectoralis, FG and FOG fibers can be differentiated solely on the basis of their mitochondria1 enzymes. Therefore, in the present study, two sections from each series were stained by histochemical methods for the demonstration of NADH activity (Dubowitz 1985). NADH, aproduct of the cit- ric acid cycle and fatty acid oxidation (Voet and Voet 1995), is prevalent in mitochondria (Dubowitz 1985).
pectoralis and other muscles; two isoforms found in the slow fibers of the anterior latissimus dorsi muscle (also termed latissimus dorsi pars cranialis) and the deep red strip of the pectoralis; and a ventricular isoform found in the heart. A bat- tery of monoclonal antibodies have been used to differentiate these isoforms. The antibodies used in this paper and their reactivity with known chicken MyHCs are summarized in Table 1.
Immunocytochemical techniques Immunocytochemical techniques, described by Shear et al. 1988, were used to demonstrate MyHC isoforms. Briefly, sec- tions from each series were first blocked in a solution consist- ing of 5% horse serum, 1% bovine serum albumin, and 5 mM EDTA in PBS (0.02 M sodium phosphate buffer, 0.15 M sodium chloride, pH 7.2). Sections were then incubated for 75 min at room temperature with a primary antibody diluted in blocking solution. Primary antibodies NA4, AG6, EB165, AB8, NA8, NA1, NA2, NA7, and HVl1 were each used at a dilution of 1:5000. However, 2E9 and B103 were used at a
MyHC isoforms and primary antibodies At least eight MyHC isoforms occur in chicken muscles, as shown by immunocytochemical techniques (see Bandman et al. 1990, 1994). These include three embryonic, one neonatal, and one adult isoform found within the fast-twitch fibers of the
720 Biochem. Cell Biol. Vol. 74. 1996
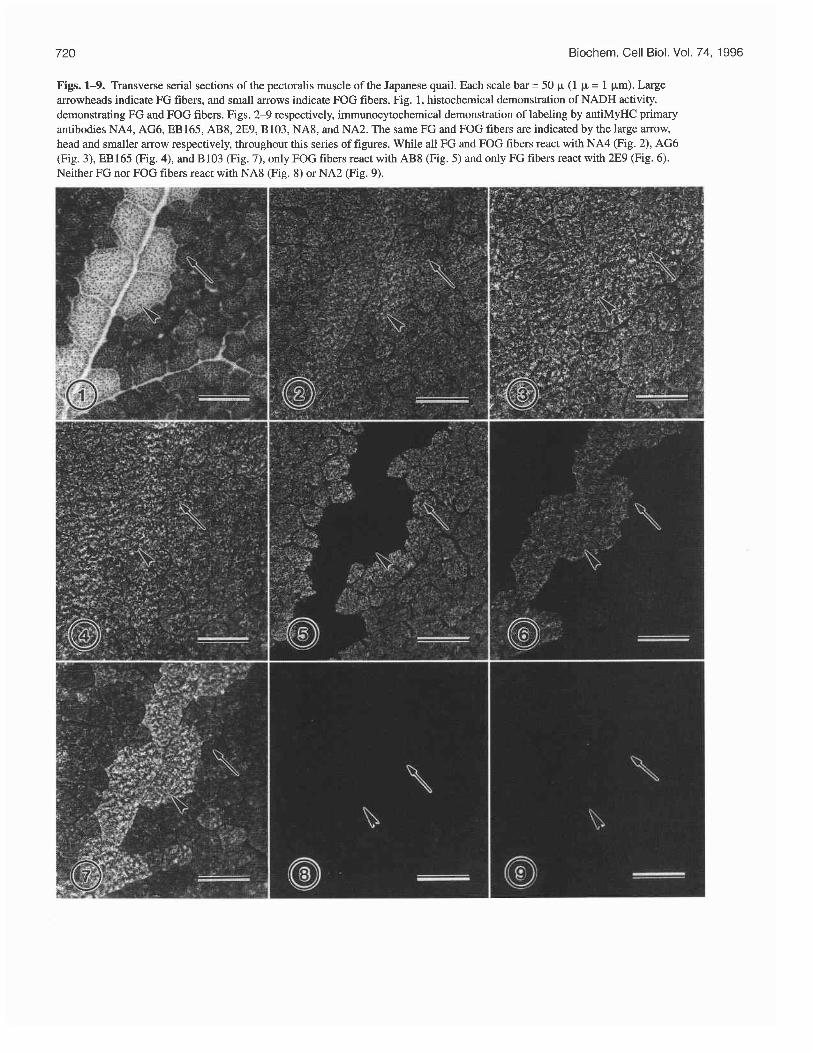
Figs. 1-9. Transverse serial sections of the pectoralis muscle of the Japanese quail. Each scale bar = 50 p (1 p = 1 pm) Large arrowheads indicate FG fibers, and small arrows indicate FOG fibers. Fig. 1, histochemical demonstration of NADH activity, demonstrating FG and FOG fibers. Figs. 2-9 respectively, immunocytochemical demonstration of labeling by antiMyHC primary antibodies NA4, AG6, EB165, AB8,2E9, B103, NA8, and NA2. The same FG and FOG fibers are indicated by the large arrow, head and smaller arrow respectively, throughout this series of figures. While all FG and FOG fibers react with NA4 (Fig. 2), AG6 (Fig. 3), EB165 (Fig. 4), and B103 (Fig. 7), only FOG fibers react with AB8 (Fig. 5) and only FG fibers react with 2E9 (Fig. 6). Neither FG nor FOG fibers react with NA8 (Fig. 8) or NA2 (Fig. 9).
Rosser et al
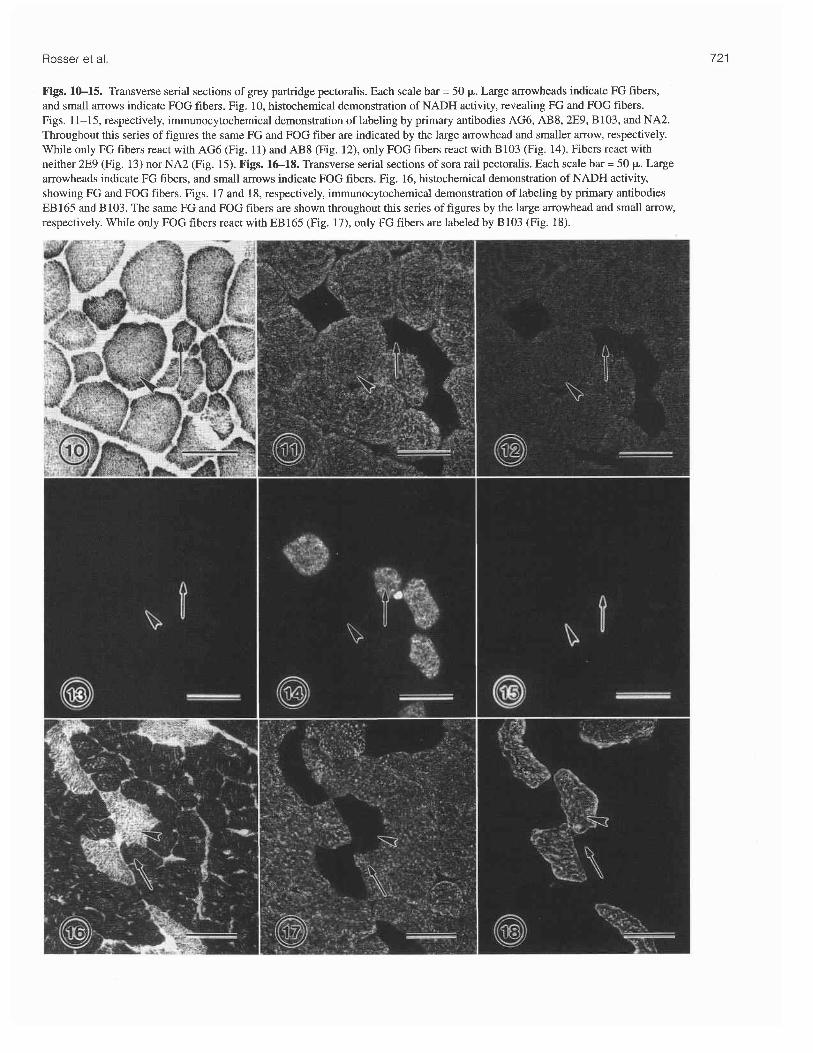
Figs. 1&15. Transverse serial sections of grey partridge pectoralis. Each scale bar = 50 p. Large arrowheads indicate FG fibers, and small arrows indicate FOG fibers. Fig. 10, histochemical demonstration of NADH activity, revealing FG and FOG fibers. Figs. 11-15, respectively, immunocytochemical demonstration of labeling by primary antibodies AG6, AB8, 2E9, B103, and NA2. Throughout this series of figures the same FG and FOG fiber are indicated by the large arrowhead and smaller arrow, respectively. While only FG fibers react with AG6 (Fig. 11) and AB8 (Fig. 12), only FOG fibers react with B103 (Fig. 14). Fibers react with neither 2E9 (Fig. 13) nor NA2 (Fig. 15). Figs. 16-18. Transverse serial sections of sora rail pectoralis. Each scale bar = 50 p. Large arrowheads indicate FG fibers, and small arrows indicate FOG fibers. Fig. 16, histochemical demonstration of NADH activity, showing FG and FOG fibers. Figs. 17 and 18, respectively, immunocytochem~cal demonstration of labeling by primary antibodies EB 165 and B103. The same FG and FOG fibers are shown throughout this series of figures by the large arrowhead and small arrow, respectively. While only FOG fibers react with EB165 (Fig. 17), only FG fibers are labeled by B103 (Fig. 18).

Biochem. Cell Biol. Vol. 74, 1996
Figs. 19-20. Transverse serial sections of long-eared owl pectoralis. Each scale bar = 50 k. Small arrows indicate FOG fibers. Fig. 19, histochemical demonstration of NADH activity, revealing FOG fibers only. Fig. 20, immunocytochemical demonstration of labeling by primary antibody AG6. The same two FOG fibers are indicated by the small arrows in these figures. While the FOG fiber on the right is labeled by AG6, the FOG fiber on the left is not. Only a minority of the fibers react with AG6. Fig. 21. Transverse section of great homed owl (juvenile) pectoralis muscle. Scale bar = 50 p. All of these FOG fibers (determined by NADH histochemistry, not shown) are labeled by the primary antibody AG6.
dilution of 1 :2000. Primary antibody dilutions were deter- mined empirically and on the basis of their known reactivity with appropriate control muscles. Sections were then rinsed twice, 5 min for each rinse, with PBS. Binding of the primary antibodies was then visualized by incubating the sections for 30 min at room temperature with a fluoresceinated horse anti- mouse IgG secondary antibody (Vector Laboratories, Burlin- game, Calif.) used at a dilution of 1:64 in PBS. Sections were then rinsed twice, 5 min for each rinse, in PBS and subse- quently fixed for 3 min in 4% buffered formalin. Sections were again rinsed twice, 5 min for each rinse, in PBS and then mounted in citifluor mountant (Marivac Ltd., N.S.).
Western blots SDS-PAGE and Western blot analyses were done to deter- mine whether the antibody reactivity demonstrated by our immunocytochemical techniques was, in fact, a result of anti- body affinity for MyHCs. For each species, a piece of pecto- ralis muscle (-0.5 cm3) was excised from the frozen tissue blocks embedded in O.C.T. compound. Excess O.C.T. com- pound was cut away at -20°C using arazor blade, and myosin was extracted from each piece of muscle (described by Band- man et al. 1982). The myosin samples were analyzed by SDS- PAGE on 10% polyacrylamide gels and electrophoretically transferred to nitrocellulose (see Bandman 1985). The nitro- cellulose blots were then incubated with 5% nonfat milk pow- der - 3% Tween-20 in PBS (PBS - TM) for 30 min at 37°C. Subsequently, for 60 min at room temperature with agitation, each blot was incubated with one of the following monoclonal antibodies: NA4, EB165, AG6, or B103. The myosin extracted from each pectoralis was normalized to give relatively similar appearances when reacted with NA4 at 1 : 100 000. The antibodies were diluted in PBS-TM as fol- lows: NA4 (1 : 100 OOO), EB 165 (1 : 25 OOO), AG6 (1 : 50 000) and B103 (1 : 100 000). In addition, myosin from the gallina- ceous birds (chicken, Japanese quail and grey partridge) was also similarly blotted and incubated with AB8 (1:5000) or 2E9
(1:2500). Other antibodies were not done because either our immunocytochemical results demonstrated that they did not react with the pectoralis samples or their reactivity with chicken muscle had been published elsewhere. The blots were then washed three times in PBS-TM over 15 min and subse- quently incubated with horseradish-peroxidase-conjugated goat antimouse IgG (Cappel Laboratories, Pa., 1:1000 in PBS-TM) for 60 rnin at room temperature with agitation. The blots were then washed three times in a 0.1 M Tris buffer (pH 7.4) over 15 min. The blots were developed in 0.1 M Tris (pH 7.4) containing diaminobenzidine (0.2 mg/mL) - H,O, (0.024%).
Results
Histochemistry and immunocytochemistry The histochemical and immunocytochemical results for the 3 1 species studied are summarized in Table 2, where the birds are arranged phylogenetically according to Sibley and Ahlquist (1990) and Sibley and Munroe (1990). Figures 1-21 illustrate the histochemical and irnmunocytochemical profiles of those pectoralis muscles representative of the overall results.
Fast-twitch fibers were readily differentiated as either FG or FOG by their staining for NADH activity. In concurrence with earlier works (George and Berger 1966; Rosser and George 1986), FOG fibers had comparatively smaller diame- ters and were darkly stained, and FG fibers had larger diame- ters and were lightly stained (Figs. 1, 10, 16, and 19). Unambiguous FG and FOG fiber types were found in the pec- toralis muscle of 11 species, including Japanese quail, grey partridge, rouen (domestic) duck, American wigeon, northern shoveler, domestic pigeon, sora rail, least sandpiper, northern harrier, American robin, and the red-winged blackbird. The pectoralis of the ostrich and the chicken each consisted chiefly of FG fibers (see following paragraph). The remaining 18 spe- cies had a pectoralis containing FOG fibers only. The FOG fibers could be discerned as red and intermediate subtypes in 9
Rosser et al. 723
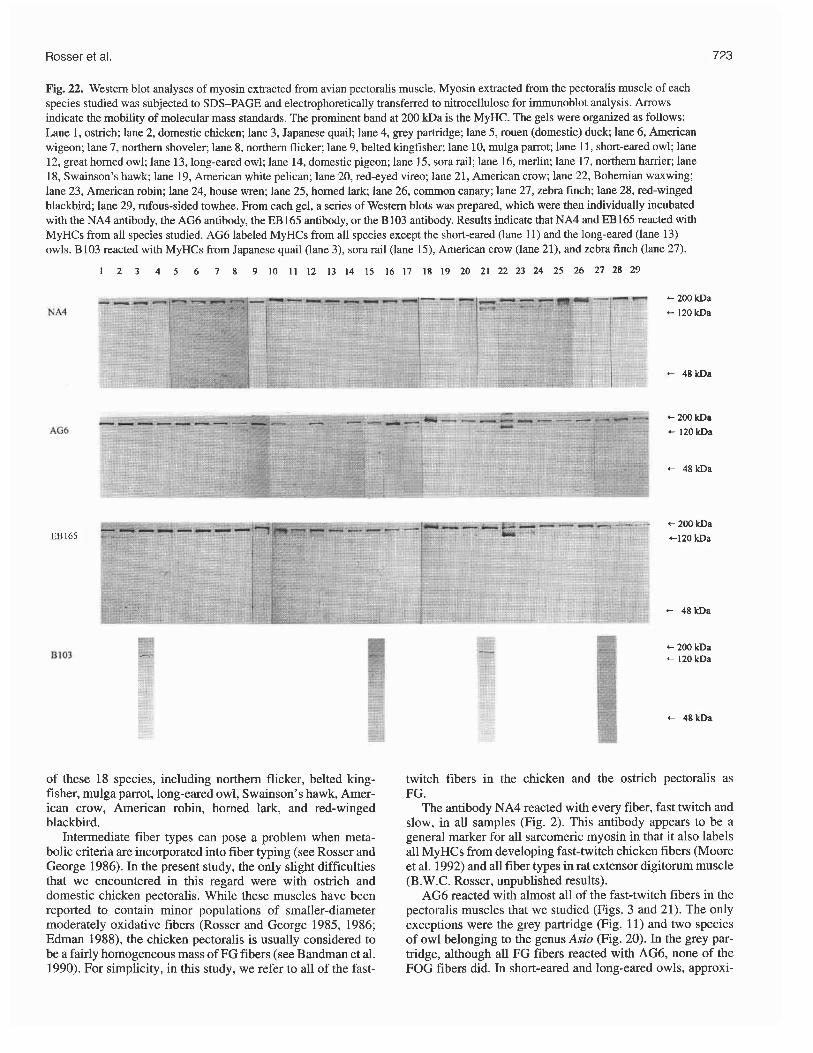
Fig. 22. Western blot analyses of myosin extracted from avian pectoralis muscle. Myosin extracted from the pectoralis muscle of each species studied was subjected to SDS-PAGE and electrophoretically transferred to nitrocellulose for immunoblot analysis. Arrows indicate the mobility of molecular mass standards. The prominent band at 200 kDa is the MyHC. The gels were organized as follows: Lane 1, ostrich; lane 2, domestic chicken; lane 3, Japanese quail; lane 4, grey partridge; lane 5, rouen (domestic) duck; lane 6, American wigeon; lane 7, northern shoveler; lane 8, northern flicker; lane 9, belted kingfisher; lane 10, mulga parrot; lane 11, short-eared owl; lane 12, great homed owl; lane 13, long-eared owl; lane 14, domestic pigeon; lane 15, sora rail; lane 16, merlin; lane 17, northern harrier; lane 18, Swainson's hawk; lane 19, American white pelican; lane 20, red-eyed vireo; lane 21, American crow; lane 22, Bohemian waxwing; lane 23, American robin; lane 24, house wren; lane 25, homed lark; lane 26, common canary; lane 27, zebra finch; lane 28, red-winged blackbird; lane 29, rufous-sided towhee. From each gel, a series of Western blots was prepared, which were then individually incubated with the NA4 antibody, the AG6 antibody, the EB165 antibody, or the B103 antibody. Results indicate that NA4 and EB165 reacted with MyHCs from all species studied. AG6 labeled MyHCs from all species except the short-eared (lane 11) and the long-eared (lane 13) owls. B103 reacted with MyHCs from Japanese quail (lane 3), sora rail (lane 15), American crow (lane 21), and zebra finch (lane 27).
of these 18 species, including northern flicker, belted king- fisher, mulga parrot, long-eared owl, Swainson's hawk, Amer- ican crow, American robin, homed lark, and red-winged blackbird.
Intermediate fiber types can pose a problem when meta- bolic criteria are incorporated into fiber typing (see Rosser and George 1986). In the present study, the only slight difficulties that we encountered in this regard were with ostrich and domestic chicken pectoralis. While these muscles have been reported to contain minor populations of smaller-diameter moderately oxidative fibers (Rosser and George 1985, 1986; Edman 1988), the chicken pectoralis is usually considered to be a fairly homogeneous mass of FG fibers (see Bandman et al. 1990). For simplicity, in this study, we refer to all of the fast-
twitch fibers in the chicken and the ostrich pectoralis as FG.
The antibody NA4 reacted with every fiber, fast twitch and slow, in all samples (Fig. 2). This antibody appears to be a general marker for all sarcomeric myosin in that it also labels all MyHCs from developing fast-twitch chicken fibers (Moore et al. 1992) and all fiber types in rat extensor digitorum muscle (B.W.C. Rosser, unpublished results).
AG6 reacted with almost all of the fast-twitch fibers in the pectoralis muscles that we studied (Figs. 3 and 21). The only exceptions were the grey partridge (Fig. 11) and two species of owl belonging to the genus Asio (Fig. 20). In the grey par- tridge, although all FG fibers reacted with AG6, none of the FOG fibers did. In short-eared and long-eared owls, approxi-

Biochem. Cell Biol. Vol. 74, 1996
Fig. 23. Western blot analyses of myosin extracted from pectoralis muscle of galliforms. Myosin extracted was subjected to SDS-PAGE and electrophoretically transferred as in Fig. 22. The gels were organized as follows: lane 2, domestic chicken, lane 3, Japanese quail; and lane 4, grey partridge. From each gel, Western blots were incubated with either the AB8 antibody or the 2E9 antibody. Results indicate that AB8 reacted with MyHCs from the pectoralis of each of the three galliforms studied. 2E9 labelled MyHCs from Japanese quail pectoralis only.
mately 30 and 25%, respectively, of the fibers in only the deep half of the pectoralis reacted with AG6. This antibody did not label any of the slow fibers that we studied.
Similarly, EB165 reacted with almost all of the fast-twitch fibers that we studied (Fig. 4). The only exceptions were the FG fibers in the pectoralis muscles of both sora rail (Fig. 17) and pigeon. Nevertheless, the FOG fibers of these two species were labeled by this antibody (Fig. 17).
AB8 and 2E9 were confined in their reactivity to the order Galliformes. Throughout the bulk of the chicken pectoralis, all fast-twitch fibers are labeled with only AB8 in the adult and by only 2E9 in the neonate (Cerny and Bandman 1987). In the red strip, however, the fast-twitch fibers of the adult react with nei- ther 2E9 nor AB8 (Shear et al. 1988). In Japanese quail and grey partridge, the muscle contained both FG and FOG fibers (Figs. 1 and 10). In mature Japanese quail, the FOG fibers were labeled by AB8 and the FG fibers by 2E9 (Figs. 5 and 6). In adult grey partridge, however, only the FG fibers were labeled by AB8 and neither fiber type reacted with 2E9 (Figs. 12 and 13).
B103 reacted with the pectoralis of several phylogeneti- cally diverse species. Previous work had shown that this anti- body recognizes embryonic and neonatal MyHC isoforms in developing chicken pectoralis (Bandman and Bennett 1988). In this study, we also observed B103 reacting with the fast- twitch fibers in the red strip of mature chickens. In the other Galliforms, we found that B 103 labeled both the FOG and the FG fibers in the adult Japanese quail pectoralis (Fig. 7) but only the FOG and not the FG fibers in the grey partridge (Fig. 14). In addition, respectively 4 and 40% of the FG and FOG fibers of the northern shoveler pectoralis were labeled by B103. This antibody also reacted with the occasional fiber (FOG) in the fledgling great horned owl; all FG but no FOG fibers of both the sora rail (Fig. 18) and the least sandpiper; 13% of the FG fibers in the northern harrier; all fibers (FOG) in Swainson's hawk, American crow, and zebra finch; 87% of the fibers (FOG) in the juvenile house wren; and a single FOG fiber in one of two red-winged blackbirds.
B 103 also labeled the ends of muscle fibers in the pectora- lis of the Anseriforms studied. In these birds, this antibody normally labels the pectoralis of neonates (E. Bandman, unpublished observations). Similarly, the tapered ends of the fast-twitch fibers in the chicken pectoralis were reported to contain both neonatal and adult MyHCs (Rosser et al. 1995).
Antibodies against slow MyHCs (NA1, NA2, NA7, and NA8) reacted with only two of our samples: ostrich pectoralis and the deep red strip of the chicken pectoralis. ATPase fiber typing has shown that the ostrich pectoralis contains from 13 to 39% slow muscle fibers (Rosser and George 1985). Simi- larly, ATPase and immunocytochernical techniques have shown that the red strip of the chicken pectoralis normally contains about one third slow fibers (see Shear et al. 1988). Antibodies against slow MyHCs did not react with pectoralis from any other species that we studied (Figs. 8, 9, and 15). Conversely, of those antibodies labeling fast-twitch fibers, only NA4 was colocalized with the antibodies against slow fibers.
HV11, which normally reacts with ventricular MyHC, also reacts with certain embryonic and regenerating avian skeletal muscle fibers (Hartley et al. 1991). It is expressed in all differ- entiating avian fibers and is considered to be a definitive marker for the activation of the myogenic program (Bourke et al. 1991). In the present study, its reactivity was restricted to just several FG and FOG fibers in the pectoralis of the least sandpiper.
Western blots Western blots for 29 of the 31 species studied are presented in Figs. 22 and 23. Westerns were not done for the two sandpip- ers. These blots not only confirmed our immunocytochemical findings, but also showed that the chicken monoclonal anti- bodies were recognizing MyHCs in the pectoralis of each of the other species studied. Almost all myosin extracts yielded a well-defined MyHC band, which indicated that the MyHCs had not degraded. The most noteworthy exception to this was that of the Bohemian waxwing pectoralis, in which there appeared to be some degradation of the MyHCs, as shown by the presence of several bands on each gel.
The monoclonal antibodies NA4 and EB165 each labeled MyHCs from all species studied (Fig. 22). AG6 labeled MyHCs from all species except the long- and the

Rosser et al.
short-eared owls (Fig. 22). B103 reacted with MyHCs from Japanese quail, sora rail, American crow, and zebra finch (Fig. 22). AB8 labeled MyHCs in the pectoralis of the domestic chicken (see also Cerny and Bandman 1987), Japa- nese quail, and grey partridge (Fig. 23). 2E9 reacted with MyHCs from the Japanese quail pectoralis (Fig. 23), as well as neonatal chicken pectoralis (not shown; see Cerny and Bandman 1987).
Certain of our immunocytochemical results were not veri- fied by the Westerns: AG6 in the long- and the short-eared owls and B 103 in grey partridge, northern shoveler, and north- ern harrier. In each of these instances, however, our immuno- cytochemical results indicated that the antibody reacted with a very minor proportion of the total fiber population. It seems reasonable to suggest, therefore, that the Western blots were not sensitive enough to detect the very low concentrations of labeled MyHCs in each of these muscles.
Discussion
Fiber type complement The belly of the pectoralis muscle consists almost entirely (99-100%) of fast-twitch fibers in those birds relying mainly on flapping flight (see Experimental model in Materials and methods). This observation is corroborated by our results showing that antibodies against slow MyHCs labeled only known slow fibers in the ostrich pectoralis and chicken red strip. These antibodies also labeled the slow fibers in the accessory deep belly of the pectoralis, found in soaring birds (Rosser et al. 1994).
FOG fibers are capable of sustained rapid contraction, and FG fibers of a more powerful but fast-fatiguing contrac- tion (Peters 1989). FOG fibers predominate in the pectoralis of all birds capable of migratory flight, and FG fibers are in the majority in the pectoralis of those relatively few species capable of only short-duration explosive flapping flight (see Rosser and George 1986). Although most smaller bodied migratory birds (such as sparrows, warblers, and humming- birds) have a pectoralis consisting entirely of FOG fibers, the pectoralis of larger bodied birds (such as pigeons, ducks, and herons) usually contains a minority of FG fibers (see Rosser and George 1986). Presumably these FG fibers assist in providing an increase in power during certain activities such as take-off (Rosser and George 1986; Welsford et al. 1991). In certain species, the intermediate subtype of the FOG fibers may perform functions not unlike those of the FG fibers of larger species (Parker and George 1975; Tobal- ske 1996).
The fiber type complement of the pectoralis muscle in the birds of this study largely concurs with published observations on 14 species, including ostrich, domestic chicken, Japanese quail, domestic duck, northern flicker, belted kingfisher, great homed owl, domestic pigeon, least sandpiper, northern harrier, white pelican, American crow, American robin, and red- winged blackbird (George and Berger 1966; Rosser and George 1985, 1986; Rosser et al. 1987, 1994; Tobalske 1996). For the remaining 17 species studied, the fiber type comple- ment of the pectoralis has not been previously reported. For these species, however, a comparable fiber type complement has been shown in at least one related species in the same genus, family, or order.
Differences in MyHC content between avian FG and FOG fibers
Differences in MyHC content were not found between FG and FOG fibers in leg muscles of adult chickens (Williams and Dhoot 1992; Maier 1994). Similarly, earlier works did not demonstrate any difference between these two fiber types in the pectoralis muscles of Japanese quail (Memfield et al. 1989) or other Galliformes (Bandman et al. 1990; Maruyama et al. 1993). Fast-twitch fibers in the latissimus dorsi muscles of adult Japanese quail (Alway 1993) and turkey (Cherel et al. 1994) were also reported to be homogeneous in their MyHC content. However, Bandman and Bennett (1988) used a bat- tery of monoclonal antibodies against chicken MyHCs to show that adult chicken gastrocnemius and posterior latissi- mus dorsi muscles contained, respectively, three and two dif- ferent populations of fast-twitch fibers. Nevertheless, whether these fibers were either type FG or FOG was not determined.
We find clear differences in MyHC content between avian FG and FOG fibers. This was the case in 5 of the 11 species having both fiber types, including Japanese quail, grey partridge, domestic pigeon, sora rail, and least sand- piper. Using monoclonal antibodies generated against chicken MyHCs, in each of these species we were able to differentiate all FG and FOG fibers as typed by their NADH activity. A small proportion (13%) of FG fibers in a sixth species, northern harrier, could be differentiated from FOG fibers in the same muscle. We could not discern any differ- ences in MyHC content between FG and FOG fibers in the remaining 5 species, including domestic duck, American wigeon, northern shoveler, American robin, and red-winged blackbird. This does not necessarily mean that in each of these species the MyHCs contained in the FG and the FOG fibers were indistinguishable. Rather, it could indicate that the antibodies that we used were unable to detect any differ- ences. In addition, we did not identify differences in MyHC content between the red and intermediate FOG subtypes in nine of the species studied.
Epitopes and isoforms The monoclonal antibodies used in this study were originally generated against specific MyHC isoforms from various developmental stages and muscles of the chicken, and each antibody has been shown to recognize one or more isoforms (see Table 1). In fact, each monoclonal antibody recognizes only a small specific epitope (amino acid sequence) at a pre- cise location on each MyHC molecule (Moore et al. 1992). The same epitope may appear on several different MyHC iso- forms and, thus, the antibody against that epitope will recog- nize each isoform containing the epitope. None of the antibodies that we use has been shown to recognize more than one epitope.
The question then arises: is a monoclonal antibody that rec- ognizes a specific epitope in chicken recognizing the identical amino acid sequence when it labels fibers from another spe- cies? The answer is that we do not know. However, for the purposes of this paper, this is not crucial. We identify pro- nounced heterogeneity of MyHCs among the fast-twitch fibers of the avian pectoralis muscle, and any interpretation of our data will support this. For example, the antibody AB8 labels (see Table 2) almost all fast-twitch fibers in adult chicken pectoralis, only FG but not FOG fibers in grey par-

Biochern. Cell Biol. Vol. 74, 1996
Table 3. Epitopes contained in fast-twitch MyHC isofonns in chicken vectoralis muscle.
Isofom Epitopes
Embryonic 1 and 3 Embryonic 2 Neonatal Adult
Note: Each epitope is named for the monoclonal antibody that labels it.
Table 4. Four new MyCHs in the avian pectoralis muscle and their epitopes.
MyHC Epitopes
Owl (FOG) Partridgelowl (FOG) Pigeon (FG) Rail (FG)
tridge, and only FOG fibers but not FG fibers in the Japanese quail. The simplest interpretation of these observations is that there are two different MyHC isoforms; one with and one without the AB8 epitope. Thus, we demonstrate heterogeneity of MyHCs. On the other hand, if the antibody is recognizing more than one epitope, at least three MyHC isoforms would be present: that with the AB8-1 epitope, that with the AB8-2 epitope, and that with neither epitope. Using this interpreta- tion, we demonstrate an even greater heterogeneity. Therefore, for the purposes of this paper, whether each antibody is recog- nizing the same epitope in each species does not alter our over- all conclusion of MyHC heterogeneity.
How many avian fast-twitch MyHCs? Bandman and colleagues (1990, 1994) have identified at least three embryonic, one neonatal, and one adult fast-twitch MyHC isoform in chicken pectoralis muscle (Table 3).
The most conservative interpretation of our results is that in which each antibody recognizes the identical epitope in each species, and it is possible for all mature avian pectoralis fibers of a particular type to contain more than one MyHC isoform. Even with this cautious interpretation, we can discern at least four new MyHCs in the avian pectoralis muscle (Table 4).
It would be an overstatement to claim that we have shown four new avian isoforms because, the same isofom can pre- sumably vary slightly from species to species. We have, how- ever, demonstrated at least four new structural varieties of avian MyHCs.
Evolution of MyHCs The first birds appeared some 150 x lo6 years ago (Sibley and Ahlquist 1990; Feduccia 1995), and the radiation of birds into the modem orders occurred about 65 x lo6 years ago (Feduc- cia 1995). It is thought that the chicken myosin heavy-chain gene family has expanded within the last 70 x lo6 years (Moore et al. 1993). In this study, the monoclonal antibodies 2E9 and AB8 labeled MyHCs in only those birds belonging to the order Galliformes, including chicken, Japanese quail, and
grey partridge. These antibodies also labeled MyHCs in the turkey (Mamyama et al. 1993). All four of these species are close relatives within the family Phasianidae. Within class Aves, Anserifoms (ducks and geese) diverged from the same stock as Galliforms (Sibley and Ahlquist 1990). Yet, Anseri- form MyHCs are labeled by neither 2E9 nor AB8. It seems reasonable to postulate, therefore, that these antibodies are labeling epitopes that appeared since the emergence of the order Galliformes. Studies of other Galliform families could provide further insight into the evolution of avian MyHCs.
Functional considerations Different MyHC isoforms have each been correlated with con- tractile properties of fast-twitch fibers in the avian pectoralis (Reiser et al. 1988) and mammalian muscle (Bottinelli et al. 1994). Other factors, however, such as myosin light-chain composition (Bottinelli et al. 1994; VanBuren et al. 1994) and metabolic criteria (Nemeth et al. 1991) have also been corre- lated with physiologic properties of muscle fibers. While FG and FOG fibers each have distinctive contractile properties, in our study there appears to be no general correlations between monoclonal antibody labeling and fiber type. There are sev- eral possible explanations for this apparent lack of correlation. The particular epitope labeled by an antibody may not be cru- cial in determining the contractile properties of the MyHC molecule. Conversely, the antibody could be recognizing dif- ferent epitopes in different species. Also, as indicated in the preceding, there are several other determinants of muscle fiber function. It has been suggested that the diversity of contractile function in birds may be less dependent on MyHC isoform composition than that in mammals (Bandman et al. 1994). However, the presence of different MyHCs in avian FOG and FG fiber types does suggests a correlation between MyHC content and contractile function.
Acknowledgements The authors thank the many associates and colleagues in Saskatoon and at the University of Saskatchewan who pro- vided the specimens. The following individuals are acknowl- edged for their cooperation and generosity: Ms. Claire Bullaro and Mr. Ken King of Saskatoon; Drs. Gary Bortolotti and Karen Wiebe of the Department of Biology; Dr. Helene Phili- bert of the Department of Veterinary Pathology; Drs. Jocelyn Hudon and Lynn W. Oliphant and Mr. Pat Thompson of the Department of Veterinary Anatomy; Dr. Hank Classen and Mr. Robert Gonda of the Department of Animal and Poultry Science; and Dr. Cheri L. Gratto-Trevor of the Canadian Wildlife Service. This study was supported by operating grants awarded to B.W.C.R. by the Saskatchewan Health Research Board and by the Natural Sciences and Engineering Research Council of Canada.
References Alway, S.E. 1993. Stretch induces non-uniform isomyosin expres-
sion in the quail anterior latissimus dorsi muscle. Anat. Rec. 237: 1-7.
Bandman, E. 1985. Continued expression of neonatal myosin heavy chain in adult dystrophic skeletal muscle. Science (Washington, D.C.), 227: 780-782.
Bandman, E., and Bennett, T. 1988. Diversity of fast myosin heavy chain expression during development of gastrocnemius, bicep

Rosser et ai
brachii and posterior latissimus dorsi muscles in normal and dys- trophic chickens. Dev. Biol. 130: 22C23 1.
Bandman, E., Matsuda, R., and Strohman, R.C. 1982. Developmen- tal appearance of myosin heavy and light chain isoforms in vivo and in vitro in chicken skeletal muscle. Dev. Biol. 93: 508-518.
Bandman, E., Bourke, D.L., and Wick, M. 1990. Regulation of myo- sin heavy chain expression during development, maturation, and regeneration in avian muscles: the role of myogenic and non- myogenic factors. In The dynamic state of muscle fibers. Edited by D. Pette. Walter de Gruyter, Berlin. pp. 127-138.
Bandman, E., Moore, L.A., Arrizubieta, M.J., Tidyman, W.E., Her- man, L., and Wick, M. 1994. The evolution of the chicken sarco- meric myosin heavy chain multigene family. Soc. Gen. Physiol. Ser. 49: 129-139.
Biewener, A.A., Dial, K.P., and Goslow, G.E., Jr. 1992. Pectoralis muscle force and power output during flight in the starling. J. Exp. Biol. 164: 1-18.
Boesiger, B. 1992. Histologie, immunocytologie, histochimie et innervation des fibres musculaires du muscle pectoralis major et du muscle supracoracoideus de Excalfactoria chinensis chinensis (L.). Acta Anat. 145: 35-43.
Bottinelli, R., Betto, R., Schiaffino, S., and Reggiani, C. 1994. Unloaded shortening velocity and myosin heavy chain and alkali light chain isoform composition in rat skeletal muscle fibers. J. Physiol. (London), 478: 341-349.
Bourke, D.L., Wylie, S.R., Wick, M., and Bandman, E. 1991. Differ- entiating skeletal muscle cells initially express a ventricular myo- sin heavy chain. BAM, 1: 369-379.
Cemy L.C., and Bandman, E. 1987. Expression of myosin heavy chain isofoms in regenerating myotubes of innervated and den- ervated chicken pectoral muscle. Dev. Biol. 119: 350-362.
Cherel, Y., Hurtel, M., Gardahaut, M.-F., Merly, F., Magras-Resch, C., Fontaine-Pems, J., and Wyers, M. 1994. Comparison of post- natal development of anterior latissimus dorsi (ALD) muscle in heavy- and light-weight strains of turkey (Meleagris gallopavo). Growth Dev. Aging, 58: 157-165.
Connold, A.L., Kamel-Reid, S., Vrbova, G., and Zak, R. 1993. Inac- tivity induces muscle hypertrophy and redistribution of myosin isozymes in chicken anterior latissimus dorsi muscle. Pflugers Arch. 423: 34-40.
Crawford, R.D. 1990. Origin and history of poultry species. In Poul- try breeding and genetics. Edited by R.D. Crawford. Elsevier, N.Y. pp. 1 4 1 .
Dial, K.P. 1992. Activity patterns of the wing muscles of the pigeon (Columba livia) during different modes of flight. J. Exp. Zool. 262: 357-373.
Dubowitz, V. 1985. Muscle biopsy: a practical approach. Bailliere- Tindall, Philadelphia, Pa.
Edman, A.-C., Lexell, J., Sjostrom, M., and Squire, J.M. 1988. Structural diversity in muscle fibres of chicken breast. Cell Tis- sue Res. 251: 281-289.
Evans, P.R., Davidson, N.C., Uttley, J.D., and Evans, R.D. 1992. Premigratory hypertrophy of flight muscles: an ultrastructural study. Omis. Scand. 23: 238-243.
Feduccia, A. 1995. Explosive evolution in tertiary birds and mam- mals. Science (Washington, D.C.), 267: 637-638.
Gaunt, A.S., Hikida, R.S., Jehl, J.R., Jr., and Fenbert, L. 1990. Rapid atrophy and hypertrophy of an avian flight muscle. Auk, 107: 649459.
Gauthier, G.F., and Orfanos, G. 1993. Developmental transitions in the myosin patterns of two fast muscles. J. Musc. Res. Cell Mot. 14: 99-109.
George, J.C., and Berger, A.J. 1966. Avian myology. Academic Press, N.Y.
Hamalainen, N., and Pette, D. 1993. The histochemical profiles of fast fiber types IIB, IID, and IIA in skeletal muscles of mouse, rat and rabbit. J. Histochem. Cytochem. 41: 733-743.
Hartley, R.S., Bandman, E., and Yablonka-Reuveni, Z. 1991. Myo- blasts from fetal and adult skeletal muscle regulate myosin expression differently. Dev. Biol. 148: 249-260.
Hartman, F.A. 1961. Locomotor mechanisms of birds. Smithson. Misc. Coll. 143: 1-91.
Hoh, J.F.Y. 1991. Myogenic regulation of mammalian skeletal mus- cle fibres. News Physiol. Sci. 6: 1 4 .
Hoh, J.F.Y. 1992. Muscle fiber types and function. Curr. Opin. Rheumatol. 4: 801-808.
Kaplan, S.R., and Goslow, G.E., Jr. 1989. Neuromuscular organiza- tion of the pectoralis (pars thoracicus) of the pigeon (Columba livia): implications for motor control. Anat. Rec. 224: 426-430.
Lundgren, B.O., and Kiessling, K.-H. 1988. Comparative aspects of fibre types, areas, and capillary supply in the pectoralis muscle of some passerine birds with differing migratory behaviour. J. Comp. Physiol. B, 158: 165-173.
Maier, A. 1994. Type and regional diversity in the distribution of myosin heavy chains in chicken intrafusal muscle fibers. Anat. Rec. 240: 507-515.
Maruyama, K., Kanemaki, N., Potts, W., and May, J.D. 1993. Body and muscle growth of domestic turkeys (Meleagris gallopavo) and expression of myosin heavy chain isoforms in breast muscle. Growth Dev. Aging, 57: 31-43.
Merrifield, P.A., Sutherland, W.M., Litvin, J., and Konigsberg, I.R. 1989. Temporal and tissue-specific expression of myosin heavy chain isoforms in developing and adult avian muscle. Dev. Genet. 10: 372-385.
Moore, L.A., Arrizubieta, M.J., Tidyman, W.E., Herman, L.A., and Bandman, E. 1992. Analysis of the chicken fast myosin heavy chain family. Localization of isoform-specific antibody epitopes and regions of divergence. J. Mol. Biol. 225: 1143-1151.
Moore, L.A., Tidyman, W.E., Arrizubieta, M.J., and Bandrnan, E. 1993. The evolutionary relationship of avian and mammalian myosin heavy chain genes. J. Mol. Evol. 36: 21-30.
Nemeth, P.M. 1990. Metabolic fiber types and influences on their transformation. In The segmental motor system. Edited by M.D. Binder and L.M. Mendell. Oxford University Press, Oxford, U.K. pp. 258-277.
Nemeth, P.M., Rosser, B.W.C., and Wilkinson, R.S. 1991. Metabolic and contractile uniformity of isolated motor unit fibers of snake muscle. J. Physiol. (London), 434: 41-55.
Parker, G.H., and George, J.C. 1975. Effect of in vivo cold-exposure on intracellular glycogen reserves in the starling type avian pec- toralis. Life Sci. 15: 1415-1423.
Peters, S.E. 1989. Structure and function in vertebrate skeletal mus- cle. Am. Zool. 29: 221-234.
Raikow, R.J. 1985. Locomotor systems. In Form and function of birds. Vol. 3. Edited by A.S. King and J. McLelland. Academic Press, N.Y. pp. 57-146.
Reiser, P.J., Graham, J.M., Greaser, M.L., and Moss, R.L. 1988. Avian pectoralis muscle (PM): molecular and functional diver- sity at the single cell level. Biophysical J. 53: 587a.
Rosser, B.W.C., and George, J.C. 1985. Histochemical characteriza- tion and distribution of fiber types in the pectoralis muscle of the ostrich (Struthio camellus) and emu (Dromaius novaeholland- iae). Acta Zool. 66: 191-198.
Rosser, B.W.C., and George, J.C. 1986. The avian pectoralis: his- tochemical characterization and distribution of muscle fiber types. Can. J. Zool. 64: 1174-1185.
Rosser, B.W.C., George, J.C., and Frombach, S.K. 1987. Archi- tecture of the pectoralis muscle of the Japanese quail (Coturnix japonica): histochemical and ultrastructural charac- terization, and distribution of muscle fiber types. Can. J. ZOO^. 65: 63-71.
Rosser, B.W.C., Waldbillig, D.M., Wick, M., and Bandman, E. 1994. Muscle fiber types in the pectoralis of the white pelican, a soar- ing bird. Acta Zool. 75: 329-336.

Biochem. Cell Biol. Vol. 74, 1996
Rosser, B.W.C., Waldbillig, D.M., Lovo, S.D., Armstrong, J.D., and Bandman, E. 1995. Myosin heavy chain expression within the tapered ends of skeletal muscle fibers. Anat. Rec. 242: 462-470.
Russell, S.D., Cambon, N.A., and Whalen, R.G. 1993. Two types of neonatal-to-adult fast myosin heavy chain transitions in rat hind- limb muscle fibers. Dev. Biol. 157: 359-370.
Schiaffino, S., and Reggiani, C. 1994. Myosin isoforms in mamma- lian skeletal muscle. J. Appl. Physiol. 77: 493-501.
Schiaffino, S., Gorza, L., Ausoni, S., Bottinelli, R., Reggiani, C., Larson, L., Edstrom, L., Gundersen, K., and Lomo, T. 1990. Muscle fiber types expressing different myosin heavy chain iso- forms. Their functional properties and adaptive capacity. In The dynamic state of muscle fibers. Edited by D. Pette. Walter de Gruyter, Berlin, pp. 329-341.
Sellers, J.R., and Goodson, H.V. 1995. Motor proteins 2: myosin. Protein Profile, 2(12): 1323-1423.
Shear, C.R., Bandman, E., and Rosser, B.W.C. 1988. Myosin heavy chain expression during development and following denervation of fast fibers in the red strip of the chicken pectoralis. Dev. Biol. 127: 326337.
Sibley, C.G., and Ahlquist, J.E. 1990. Phylogeny and classification of birds: a study in molecular evolution. Yale University Press, New Haven, Conn.
Sibley, C.G., and Monroe, B.L., Jr. 1990. Distribution and taxonomy of birds of the world. Yale University Press, New Haven, Conn.
Stal, P., Eriksson, P.-O., Schiaffino, S., Butler-Browne, G. S., and Thornell, L.-E. 1994. Differences in myosin composition between human oro-facial, masticatory and limb muscles: enzyme-imunohisto- and bio-chemical studies. J. Musc. Res. Cell Motil. 15: 5 17-534.
Staron, R.S., and Pette, D. 1993. The continuum of pure and hybrid myosin heavy chain-based fibre types in rat skeletal muscle. His- tochemistry, 100: 149-153.
Swain, S.D. 1992. Flight muscle catabolism during overnight fasting in a passerine bird Eremophila alpestris. J. Comp. Physiol. B, 162: 383-392.
Tobalske, B.W. 1996. Scaling of muscle composition, wing mor- phology, and intermittent flight behavior in woodpeckers. Auk, 113: 151-177.
Torrella, J.R., Fouces, V., Palomeque, J., and Viscor, G. 1993. Inner- vation distribution pattern, nerve ending structure, and fiber types in pigeon skeletal muscle. Anat. Rec. 237: 178-186.
VanBuren, P., Guillermina, S.W., Harris, D.E., Trybus, K.M., War- shaw, D.M., and Lowey, S. 1994. The essential light chain is required for full force production by skeletal muscle myosin. Proc. Natl. Acad. Sci. U.S.A. 91: 12 403 - 12 407.
Voet, D., and Voet, E. 1995. Biochemistry. John Wiley & Sons, Inc., N.Y.
Welsford, I.G., Meyers, R.A., Wilson, D.S., Satterlie, R.A. and Gos- low, G.E., Jr. 1991. Neuromuscular organization for "wing" con- trol in a mollusc (Clione limacina) and a bird (Colurnba livia): parallels in design. Am. Zool. 31: 670-679.
Williams, K., and Dhoot, G.K. 1992. Heterogeneity and distribution of fast myosin heavy chains in some adult vertebrate skeletal muscles. Histochemistry, 97: 361-370.
Yates, L.D., and Greaser, M.L. 1983. Quantitative determination of myosin and actin in rabbit skeletal muscle. J. Mol. Biol. 168: 123-141.
Copyright © 2022 FDOKUMEN




















