
Herbs on the soil moisture gradient (in Russian)
274
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Herbs on the soil moisture gradient (in Russian)

С . Н . Ш е р е м е т ь е в 10Щ
Т Р А В Ы Н А Г Р А Д И Е Н Т Е
^ В Л А Ж Н О С Т И П О Ч В Ы
Москва • 2005

РОССИЙСКАЯ АКАДЕМИЯ НАУК БОТАНИЧЕСКИЙ ИНСТИТУТ имени В. Л. КОМАРОВА
С. Н. Шереметьев
ТРАВЫ НА ГРАДИЕНТЕ ВЛАЖНОСТИ ПОЧВЫ
(ВОДНЫЙ ОБМЕН И СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ
ОРГАНИЗАЦИЯ)
Товарищество научных изданий КМ К Москва • 2005

УДК 581.1
Шереметьев С. Н. Травы на градиенте влажности почвы (водный обмен и структурно-функциональная организация). — М.: Товарищество научных изданий КМК. 2005. — 271 с. В книге обобщены многолетние исследования водного режима большого числа видов травянистых растений на широком градиенте влажности почвы. Подробно проанализированы изменения интенсивности транспирации, дефицита водного насыщения, содержания воды, осмотического давления клеточного сока листьев каждого вида. Показаны закономерности изменений средневидовых величин элементов водного режима на градиенте влажности почвы. Для 24 видов дан обзор структурно-функциональной организации растений. Рассматриваются соотнощение тканей и свободных пространств в листьях, динамика парциальных объемов на градиенте влажности почвы, межвидовые структурные и структурно-функциональные корреляции. В заключении обсуждаются возможные тренды экологической эволюции травянистых растений. Книга рассчитана на специалистов в области экологической физиологии растений.
Табл. 42. Рис. 160. Библиогр. 219 назв.
Исследования поддержаны грантами Российского фонда фундаментальных исследований (95-04-11195 и 98-04-49814)
Издание осуществлено при финансовой поддержке Российского фонда фундаментальных исследований согласно проекту 05-04-62003
Р И
5-87317-244-7 © С. Н. Шереметьев, 2005 © Т-во научных изданий КМК, издание 2005

ВВЕДЕНИЕ
Полевые исследования водного режима проводятся в широком спектре экологических условий. В литературе можно найти характерные величины показателей водообмена для доминантов сообществ практически всех климатических зон. Несмотря на такое обилие фактов, обычно обсуждаются лишь феноменологические различия у растений разных сообществ или экотопов. Отсутствуют как количественные, так и качественные обобщения полевых наблюдений водного режима на достаточно широких градиентах факторов среды. Между тем, именно на этой основе можно ставить вопросы о причинах и приспособительном значении лабильности показателей водообмена, их характерных величинах в разных экотопах на градиентах факторов среды, определять экологическую принадлежность видов — то есть решать задачи собственно экологической физиологии растений.
В этой книге будет сделана попытка в некоторой степени восполнить существующие пробелы. В общем виде цели исследования сводятся к а) изучению функциональных и структурных элементов растений на градиенте влажности почвы; б) выявлению взаимосвязей между этими элементами; в) исследованию на этой основе свойств организованности (целостности) систем, составленных из изученных элементов.
Для достижения этих целей нам нужно рассмотреть, как изменяются на градиенте влажности почвы основные показатели водообмена: интенсивность транспирации, дефицит водного насыщения, содержание воды, осмотическое давление клеточного сока.
Исследование межвидовых структурных и структурно-функциональных корреляций на градиентах факторов среды представляет несомненный интерес для выяснения ряда вопросов экологической анатомии и физиологии растений. Межвидовые структурные корреляции могут указать на характер компартментации потоков и объем пулов запасания в листьях растений, согласованность их изменений при изменении условий произрастания.
Ряд показателей водообмена, судя по всему, имеет определенную структурную основу. Это можно выяснить, исследуя структурно-функциональные корреляции. Широко известно, например, что плотность сложения мезофилла является признаком ксероморфной организации растений, она увеличивается в направлении от гумидных

4 С.Н. Шереметьев. Травы на градиенте влажности почвы
к аридным местообитаниям. Очевидно, что в этом же направлении уменьшаются интенсивность транспирации и оводненность листьев. Судя по всему, должны существовать экологические ряды видов, на материале которых проявляются межвидовые корреляции структурных и функциональных признаков у растений как в пределах эко-топа, так и на градиенте факторов среды. Эти корреляции могут помочь выявить иерархию и характер влияния тех или иных структурных признаков на функциональные. Само наличие таких корреляций наводит на мысль о существовании параллельной структурно-функциональной изменчивости у неродственных таксонов.
При обзоре структурно-функциональной организации растений мы рассмотрим а) соотношение тканей и свободных пространств в листьях однодольных и двудольных растений; б) динамику парциальных объемов тканей и свободных пространств в листьях на градиенте влажности почвы; в) структурные корреляции в листьях растений; г) структурно-функциональные корреляции. В заключении мы обсудим возможные тренды экологической эволюции травянистых растений.
Автор выражает глубокую признательность Ю. Б . Гамалею, Н. Н. Слемневу Р. В. Камелину за поддержку, обсуждения, критические замечания, сотрудникам лабораторий аридной зоны и экологической физиологии БИН РАН, благодарен ныне покойным Н. Н. Измайловой, В. М. Свешниковой и Ю. В. Титову. Эта работа была бы невозможна без содействия руководства Советско-Монгольской (ныне Российско-Монгольской) комплексной биологической экспедиции — П. Б. Виппера и П. Д. Гунина, а также без помощи руководителей Хоперского природного заповедника — А. И. Зо-бова и Н. А. Карпова. Особую благодарность хочу выразить Т. Б. Шереметьевой и геоботанику Хоперского заповедника Е. С. Нескрябиной.
МАТЕРИАЛ И МЕТОДИКА
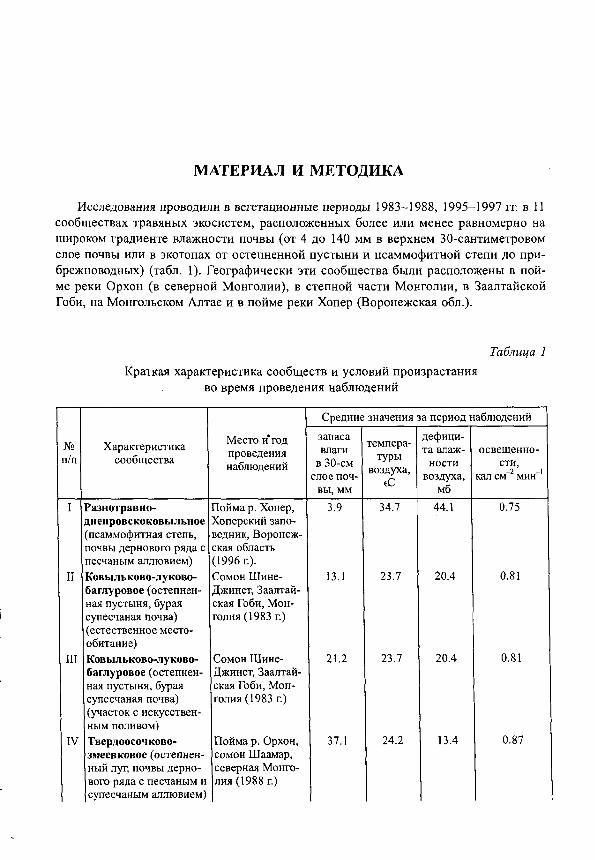
Исследования проводили в вегетационные периоды 1983-1988, 1995-1997 гг. в 11 сообществах травяных экосистем, расположенных более или менее равномерно на щироком градиенте влажности почвы (от 4 до 140 мм в верхнем 30-сантиметровом слое почвы или в экотопах от остепненной пустыни и псаммофитной степи до при-брежноводных) (табл. 1). Географически эти сообщества были расположены в пойме реки Орхон (в северной Монголии), в степной части Монголии, в Заалтайской Гоби, на Монгольском Алтае и в пойме реки Хопер (Воронежская обл.).
Таблица 1 Краткая характеристика сообществ и условий произрастания
во время проведения наблюдений
Средние значения за период наблюдений
№ Характеристика Место и год проведения наблюдений
запаса влаги темпера дефици
та влаж освещенноп/п сообщества
Место и год проведения наблюдений в 30-см туры
воздуха, (С
ности сти, -2 - I слое поч
туры воздуха,
(С воздуха. кал см мин вы, мм
туры воздуха,
(С мб I Разнотравно-
днепровскоковыльное (псаммофитная степь, почвы дернового ряда с песчаным аллювием)
Пойма р. Хопер, Хоперский заповедник. Воронежская область (1996 г).
3.9 34.7 44.1 0.75
II Ковыльково-луково-баглуровое (остепнен-ная пустыня, бурая супесчаная почва) (естественное местообитание)
Сомон Шине-Джинст, Заалтай-ская Гоби, Монголия (1983 г.)
13.1 23.7 20.4 0.81
III Ковыльково-луково-баглуровое (остепнен-ная пустыня, бурая супесчаная почва) (участок с искусственным поливом)
Сомон Шине-Джинст, Заалтай-ская Гоби, Монголия (1983 г.)
21.2 23.7 20.4 0.81
IV Твердоосочково-змеевковое (остепнен-ный луг, почвы дернового ряда с песчаным и супесчаным аллювием)
Пойма р. Орхон, сомон Шаамар, северная Монголия (1988 г)
37.1 24.2 13.4 0.87

6 С.Н. Шереметьев. Травы на градиенте влажности почвы
Окончание таблицы 1
Средние значения за период наблюдений
№ п/п
Характеристика сообщества
Место и год проведения наблюдений
запаса влаги
в 30-см
температуры
воздуха, еС
дефицита влаж
ности освещенно
сти, -2 -1 слое поч
температуры
воздуха, еС воздуха. кал см мин
вы, мм
температуры
воздуха, еС
мб V Ковыльно-
разнотравно-вострецовое (настоящая степь, лугово-кащтановая остепнен-ная почва)
Сомон Туменцогт, восточная Монголия (1984-1985 г.)
50.3 21.1 11.6 0.77
VI Разнотравно-осоковое (среднепоемный настоящий луг, почвы лугового ряда с суглинистым аллювием)
Пойма р. Хопер, Хоперский заповедник. Воронежская область (1997 г.)
55.9 26.1 19.1 0.78
VII Злаково-осоковое (долгопоемный настоящий луг, почвы болотного ряда с глинистым и иловатым аллювием)
Пойма р. Орхон, Сомон Шаамар, северная Монголия (1986 г)
99.6 21.8 8.6 1.24
УШ Осоково-разнотравно-злаковое (долгопоемный настоящий луг, почвы лугового ряда с суглинистым аллювием)
Пойма р. Орхон, Сомон Шаамар, северная Монголия (1987 г.)
104.0 22.3 14.2 0.84
IX Крупноразнотравно-дернистоосоковое (заболоченный луг, почвы болотного ряда с глинистым и иловатым аллювием)
Пойма р. Хопер, Хоперский заповедник. Воронежская область (1995 г.)
111.5 25.3 18.4 0.71
X Злаково-богаторазнотравное (заболоченный луг, почвы лугового ряда с суглинистым аллювием)
Пойма р. Орхон, Сомон Шаамар, северная Монголия (1987 г.)
123.5 22.6 13.7 1.31
XI Остроосоковое (участок прибрежноводной растительности, песчаная почва прибрежной зоны озера)
Пойма р. Хопер, Хоперский заповедник. Воронежская область (1996 г.)
143.9 21.6 11.6 0.26

МАТЕРИАЛ И МЕТОДИКА 7
Объектами наблюдений были представители 50 видов растений, из которых подавляющее больщинство — поликарпические травы, и два — полукустарнички {АпаЬа81з Ьгеу1/оИа С.А. Меу. и АПетша /гщШа Поскольку некоторые виды встречались не в одном, а в разных сообществах, то число объектов наблюдений было несколько больше, чем видов, а именно — 59 (табл. 2). В фитоценотическом отношении все они являлись доминантами или содоминантами своих сообществ.
Для удобства анализа градиент влажности почвы был разделен на три примерно равные части: аридную с запасами влаги от О до 50 мм, среднюю — с запасами влаги от 50 до 100 мм и гумидную — с запасами воды от 100 до 150 мм.
Таблица 2 Список изученных видов и их краткая характеристика
№ Эко-участ № ло-ка по объ Вид гиче- Описание вида табл. екта ский
1 тип I 1 АПетша аишпаса
1асц. К Многолетнее травянистое корнеотпрысковое расте
ние; корневая система проникает на глубину свыше 1 м (Шалыт, 1950; Зозулин, 1959). Высота растений до 30 см. Стебли и листья с шелковисто-беловато-шерстисгым опушение.м. Листья многократно рассечены. Доли листьев мелкие, узкие. Цветет в июле -августе. Растет преимущественно на степных склонах, остепненных полянах (Цвелев, 1988).
2 Сагех со1сЫса Оау
К Многолетнее травянистое длиннокорневищное растение высотой 10-50 см (Алексеев, 1996). Корни могут проникать на глубину 30 см (Байтулин, 1979), а горизонтальное (осевое) корневище растет на глубине 10-15 см (Абачев, 1990). Побеги одиночно или редкими пучками расположены на корневище. Листья двускладчатые, узкие и тонкие. Цветет в июне -июле. Встречается на слабо задернованных песках, пойменных песчаных лугах (Цвелев, 1988).
3 СаНит ги1кетсит ШЫ.
К Многолетнее травянистое стержнекорневое короткокорневищное растение (Заугольнова, 1974), высотой 25^0 см. Листья с листовидными прилистниками линейные, нитевидные по 8-10 в муговке. Корневая система глубокая (проникает глубже 1 м). Цветет в середине июня - середине июля. Растет на остепненных полянах, сухих лугах, склонах (Цвелев, 1988).
4 811ра Ьогуз1кетса К1ок. ех Ргокиё.
К Многолетний плотнодерновинный злак высотой 30-80 см с глубокой (от 1 м и глубже) кистекорневой системой (Титов, Печенюк, 1990). Листовые пластинки до 3 мм шириной, очень жесткие, гладкие Цветет в конце мая - начале июня. Растет на песчаных склонах, в песчаной степи (Цвелев, 1988).

8 С.Н. Шереметьев. Травы на градиенте влажности почвы
Продолжение таблицы 2
№ Эко-)'част- № ло-капо объ Вид гиче- Описание вида табл. екта ский
1 тип 5 Тг1/оИит агуепхе Ъ. К Однолетнее стержнекорневое растение с вертикаль
ным главным корнем, проходящим сквозь гумусовый горизонт и достигающим глубины 40-60 см (Шалыт, 1950). Высота особей 5-30 см. Тонкие стебли прямостоячие, разветвленные; стебель и листья опушенные (Губанов и др., 1990). Цветет в июне. Растет на сухих лугах, на степных склонах, в песчаной степи (Цвелев, 1988).
6 Уеготса 5р1са1а Ь. К Многолетнее травянистое корневищно-кустовое (Серебрякова, Кагарлицкая, 1972; Еленевский, 1978) растение высотой 10-20 см. Принадлежит к группе ксерофильных видов данного рода (Серебрякова, Кагарлицкая, 1972). Опушено короткими простыми волосками, вверху железистыми. Цветет в июне -июле. Растет на слабо задернованных песках, пойменных песчаных лугах (Цвелев, 1988).
11 7 АШит ро1угк12ит Тигс2. ех К.е§е1
С Плотнодерновинный лук с корневой системой, проникающей в глубину до 30 см и с радиусом распространения корней 30-40 см. Произрастает, главным образом, в микропонижениях остепненной пустыни. Корневая система не очень глубока, но чрезвычайна чувствительна даже к небольшим осадкам (Попова, 1977). В сухие годы луки покоятся, позднелетне-осенние осадки вызывают их бурный рост (Санжид, Федорова, 1983).
8 АпаЪа815 Ьгеу1/оИа СК Полукустарничек, афилльный суккулент, корневая С. А. Меу. система которого простирается в глубину до 100-
140 см. Является одним из основных эдификаторов не только остепненной, но и настоящей и крайне-аридной пустынь (Рачковская, Федорова, 1983). Фенологическое развитие растений сильно зависит от запасов влаги в почве. Период вегетации юве-нильных и имматурных особей в крайне засушливые годы может сокращаться до одного месяца; в сухие годы летом наблюдается сильное подсыхание и опадение части вегетативных побегов, осенняя окраска у значительной части молодых особей (Санжид, Федорова, 1983). Помимо этого известно, что в побегах растений имеется большой объем водоносной паренхимы, которая может функционировать как резервуар, абсорбирующий воду в ночное время и отдающий ее мезофиллу в течение дня (Гамалей, 1984).

МАТЕРИАЛ И МЕТОДИКА 9
Продолжение таблицы 2
№ Эко-участ № ло-ка по объ Вид гиче- Описание вида табл.
1 екта ский
тип 9 8Ира §1агео5а
Р. 8т1т. МК Плотнодерновинный злак со склероморфными листьями
фестукоидного типа (Гамалей, 1983), имеющий хорошо развитую корневую систему до 80 см глубиной. Обладает отчетливым аридным габитусом (Гамалей, 1984), однако, в отличие от АпаЬаз1з Ьгег1/оНа, редко встречается в настоящей пустьше и отсушгвует в крайнеарид-ной пустьше. В остепненной пустьше произрастает в микропонижениях (Рачковская, Федорова, 1983).
III 10 АШит ро1угЫгит С См. № 7. I I АпаЬа81з ЬгеУ1/оИа СК См. № 8. 12 8Ира §1агеоза МК См. № 9.
IV 13 Сагех ^игшхси1а С. А. Меу.
К Многолетнее растение с тонкими шнуровидными подземными побегами, образующими на концах маленькие густые дерновинки (Юнатов, 1954). Шнуровидное корневище находится на глубине 1-2 см от поверхности почвы (Федоскин, 1979). Корневая система проникает в глубину до 60 см. Этот вид широко распространен в степных районах Монголии, часто и обильно произрастает по равнинам восточной части этой страны, заходит в северную Гоби. Встречаясь в очень разнообразных экологических условиях, предпочитает сухие местообитания, обычен в горньк и равнинных степях (Юнатов, 1954),
14 С1еШо§епе5 здиаг-гоза (Тпп.) Кеп§
К Рыхлокустовой дерновинный злак с корневой системой, проникающей до глубины 30 см. Является одним из широко распространенных видов северной и центральной Монголии. Тесно связан с зоной степей, где часто является эдификатором главнейших степных формаций: змеевко-тырсовых, полынно-змеевко-тыр-совых, злаково-карагановых (Юнатов, 1954).
15 Ьеутш сМпепз18 (Тгш.) Т2Уе1.
К Многолетний длиннокорневищный злак с корневой системой, проникающей в глубину до 150 см (Полюш-кин, Горшкова, 1979). Взрослые растения имеют отходящие от корневищ корни, пронизывающие густой сеткой все горизонты почвы (Горшкова, 1966). Маньч-журско-даурско-монгольский вид (Лавренко, 1978). Он относится к числу наиболее распространенных злаков Монголии, очень обилен в восточной части страны и является, наряду с ковьшем-волосатиком, основным эдификатором степей (Юнатов, 1954). Растет на солонцах, солонцеватых лугах, песках, галечниках речных долин и на склонах сопок и гор (Чойжамц, 1978). Изредка встречается в Гобийской части Монголии, где обитает исключительно по увлажненным низинам (Юнатов, 1954). Наиболее благоприятные условия находит на широких днищах межгорных долин, на межо-земистых, слабозасоленных почвах (Буевич, 1983).

10 С.Н. Шереметьев. Травы на градиенте влажности почвы
Продолжение таблицы 2
№ Эко-участ № ло-ка по объ Вид гиче- Описание вида табл. екта ский
1 тип 16 РоШИИа Ы/игса Ь. МК Невысокий многолетник с восходящими, одревесне
вающими у основания побегами. Имеет стержнекор-невую систему, доходящую до глубины 60 см. Этот вид обладает широкой экологической амплитудой, встречается в травостоях горных степей и отмечается в составе сухостепных и пустынностепных группировок (Юнатов, 1954)
V 17 Аг1ет1з1а /гщШа МК Полукустарничек высотой 10-30 см, с придаточной корневой системой, проникающей в глубину до 80-100 см (Полюшкин, Горшкова, 1979). З.В. Карамышева (1969) отмечает, что он по типу ареала является казах-станско-монгольско-сибирско-североамериканским видом. В Монголии это наиболее широко распространенный вид ПОЛЬШИ, встречается в пределах всех лесостепных и степных районов. Входит в состав тырсовьк, змеевко-тырсовых и вострецово-тырсовых сухих степей, более обильно разрастаясь на щебнистых почвах (Юнатов, 1954).
18 /га сИскогота Ра11. С Многолетнее травянистое короткокорневищное растение. Система придаточных корней функционирует в течение 5-7 лет, обеспечивая достаточное водоснабжение растений. Встречается на Дальнем Востоке, в Японии и Китае (Горшкова, 1966). В Монголии растет в песчаных степях и на береговых песках в Хэнтейском, Монголо-Даурском, Прихинганском и Восточномонгольском ботанико-географических районах (Грубов, 1982).
19 Ьеутш сктеппз МК С м № 15. 20 РоШИНа 1апасеИ[о-
Иа ты. ех ЗсЫесЫ.
МК Многолетнее травянистое длиннокорневищное растение с корнями, проникающими в глубину до 150-200 см (Горшкова, 1966). Произрастает в степях, на остепненных лугах, среди кустарников в предгорьях и низкогорьях (Губанов, Ганболд, 1983).
21 РиЫаИНа Шгсгат-поун Кгу1. е1 5ег§.
МК Многолетнее травянистое растение, имеющее хорошо развитую корневую систему длинностержневого типа с глубиной проникновения корней до 70-100 см (Полюшкин, Горшкова, 1979). Этот вид особенно хорошо распространен в петрофитных степях (Буевич, 1983) восточной Монголии: в Прихубсугулье, Восточном Хангае, Хэнтее (Юнатов, 1954).
22 8Ира кгу1оуИ КовЬеу.
к Плотнодерновинный злак высотой 30 см, корни которого проникают в глубину до 80 см (Чойжамц, 1978). Развитая корневая система состоит из многочисленных придаточных корней, густо пронизывающих почвенные слои (Буевич, 1983). Вегетирует

МАТЕРИАЛ И МЕТОДИКА 11
Продолжение таблицы 2
№ Эко-участ № ло-ка по объ Вид гиче- Описание вида табл.
1 екта ский
тип 22 Зйра кгу1оуЦ
ЯовЬеу. К с мая по сентябрь. Цветет в августе. Е.М. Лавренко
(1978) отмечал, что это восточносибирско-централь-ноазиатский вид, распространенный от Восточного Тибета и Восточного Тянь-Шаня с некоторыми перерывами до Центральной Якутии. В степной зоне Монголии является одним из ценозообразователей змеевково-тырсовых, вострецово-тырсовых и полын-но-тырсовых степей (Юнатов, 1954).
VI 23 Са1ата^гозИ5 ер1§еюз (С.) КоЛ
К Многолетнее длиннокорневищное растение с кистекорневой пучковатой системой гл^'биной до 200 см. Стебель высотой 80-150 см. Цветет во второй половине июня. Растет преимущественно на песчаных (хорошо дренированных) почвах, но встречается и на заболоченных лугах (Губанов и др., 1990). Типичен для молодых сосновых культур и лесных опушек надлуговой террасы, откуда он и заходит на участки псаммофитно-степной растительности.
24 Сагех те1апо81аскуа В1еЬ. ех ты.
МК Многолетнее травянистое короткокорневищное растение с кистекорневой системой глубиной до 60 см. Цветет в июне. Растет преимущественно во влажных местообитаниях, на лугах, лесных полянах, по берегам водоемов.
25 СаНит гШИетсит ты.
МК Многолетнее травянистое длиннокорневищное растение высотой 25^0 см. Листья с листовидными прилистниками линейные, нитевидные по 8-10 в мутовке. Имеет многоосевую стержнекорневую систему с максимумом ветвления в средней и нижней частях глубиной до 200 см. Цветет в середине июня - начале июля. Растет на остепненных полянах, сухих лугах, степных склонах.
26 Ьугкгит угг^аШт Ь. к м Многолетнее травянистое растение с толстым деревянистым корнем и прямостоячим четырехгранным простым или ветвистым стеблем высотой 30-100 см. Цветет в июле-августе. Преимущественно растет в сырых местах: на влажных заливных лугах, травяных болотах, по берегам водоемов.
27 Зап^шзогЬа о01с(-паНз Ъ.
к м Многолетнее короткокорневищное растение с кистекорневой универсальной системой глубиной до 120-150 см. Стебли высотой 100-150 см. Цветет в июле -августе. Преобладает на лугах с влажнолуговым увлажнением, реже растет на суходольных лугах, в луговых степях. Благодаря хорошо развитой корневой системе, проникающей в глубину до 1 м, кровохлебка хорошо переносит засухи, способна расти на бедных почвах (Губанов и др., 1990).

12 С.Н. Шереметьев. Травы на градиенте влажности почвы
Продолжение таблицы 2
№ Эко-участ № ло-ка по объ Вид гиче- Описание вида табл. екта ский
1 тип 28 ТапасеШт уи1§аге
Ь. М Многолетнее травянистое растение с ползучим, раз
ветвленным корневищем. Стебель высотой 30-100 см, прямой, разветвленный вверху, обильно облиственный. Цветет в июле - октябре. Часто растет в виде отдельных скоплений в травостое на сухих и умеренно влажных участках. Выносит краткосрочное затопление и небольшое заиление (Губанов и др., 1990).
VII 29 А1оресигиз агип-Шпасеиз Ро1г.
МК Многолетний рыхлокустовой злак до 60-80 см высотой (Юнатов, 1954). Корневая система проникает до глубины 60 см. Встречается по влажным и сырым солончаковым лугам, берегам рек и озер, прибрежным галечникам в Хангае, Хэнтее и Монгольском Алтае (Юнатов, 1954; Грубов, 1982).
30 Сагех а1Него(1е5 8ргеп§.
МК Многолетнее растение до 50-60 см высотой, образующее рыхлые дерновины. Корневая система простирается до глубины 60 см. Произрастает на сырых и мокрых лугах, дренируемых проточными водами. Чаще всего встречается в Хангае, Хэнтее и степной полосе восточной Монголии (Юнатов, 1954).
31 Сагех ШИорИИа Тигс2.
МК Многолетнее летнезеленое растение с корневой системой, проникающей в глубину до 60 см. Произрастает на сырых лугах, по заболоченным берегам рек, в прибрежных зарослях кустарников в Хангае, Хэнтее, Монгольском Алтае (Грубов, 1982).
32 Е1у(п§1а гереп! (Ь.) Ксузк!
МК Ползучекорневищный многолетний зтак высотой до 100 см. Корневая система проникает до глубины 60 см, Произрастает преимущественно по долинам рек в северной Монголии, изредка заходя к югу по горньм хребтам Гобийского Алтая (Юнатов, 1954). Имеет довольно широкую экологическую амплитуду и легко приспосабливается к различным условиям обитания: к влажным и сухим участкам, к почвам легкого и тяжелого механического состава, Нередко, особенно в поймах рек, образует чистые заросли, но чаще доминирует в травяном покрове вместе с другими злаками (Губанов и др., 1990).
33 / т зЛтса Ь. с Многолетнее растение до 70 см высотой, образующее дерновины (Юнатов, 1954). Корневая система может доходить до глубины 60 см. Произрастает по берегам рек, на сырых заболоченных лугах, кочковатьк болотах (Грубов, 1982). К почвам нетребователен, но предпочитает богатые гумусом, достаточно увлажненные с нейтральной или слабокислой реакцией. Может образовывать чистые заросли или доминировать в травяном покрове, но чаще не играет ведущей роли в сложении злаковых и разнотравных лугов (Губанов др., 1990).

МАТЕРИАЛ И МЕТОДИКА 13
Продолжение таблицы 2
№ Эко-участ № ло-ка по объ Вид гиче- Описание вида табл. екта ский
1 тип 34 РИаЬгоШез
агипсИпасеа (Ь.) КаиясЬеП.
КМ Корневищный злак высотой до 70 см с глубиной проникновения корневой системы до 60 см. Отмечен только по заливным лугам при слиянии рек Орхона и Селенги (Юнатов, 1954). Часто встречается как субдоминант травяно-осоковых заболоченных лугов и болот, выдерживает затопление, но хорошо переносит и засуху (Губанов и др., 1990).
35 Ро1у^опит зШпсит Ьахт.
М Невысокое (до 20-30 см) многолетнее растение, корневая система которого может доходить до глубины 60 см. Произрастает почти по всей Монголии на солончаках и солончаковатых лугах, заходит на галечники и засоленные низинные пески (Юнатов, 1954).
VIII 36 А^гозйз тощкоИса ЯозЬеу.
МК Многолетний злак до 60-70 см высотой, образующий рыхлые дерновинки. Корневая система проникает до глубины 30 см. Произрастает по сырым и влажным лугам низкого уровня в поймах рек, по днищам падей и окраинам озер почти во всех районах северной Монголии, всегда в условиях достаточного почвенного питания. Даже умеренную засоленность почвы переносит плохо (Юнатов, 1954).
37 Апетопе (ИскоЬота Ь.
М Многолетнее растение высотой до 30 см. Корневая система имеет глубину проникновения до 30 см. Произрастает на заливных и сырых лесных лугах, в прибрежных ивняках (Грубов, 1982).
38 Сагех шНегоЛез МК См. № 30. 39 Роа ап§изН/оИа Ь. к Многолетний рыхлодерновинный злак, очень часто с
многочисленными корневищами и вегетативными побегами. Корневая система проникает до 30 см. Произрастает в небольшом количестве во всех районах северной Монголии. Приурочен к влажным луговым почвам, изредка встречается в лесах и чаще по луговым степям, прилесным лугам (Юнатов, 1954).
40 ТНаНс1гит зтр1ех Ь.
к м Многолетнее растение 60-70 см высотой с корневой системой до 30 см. Растет довольно обильно по разнотравным луговым степям, прилесным лугам (Юнатов, 1954).
41 У1с1а сгасса Ь. м Корневищное многолетнее растение с многочисленными лазящими стеблями. Глубина проникновения корней до 60 см. Произрастает в пределах лесостепной и степной зон по сухим и влажным лугам, лесным опушкам (Юнатов, 1954).
IX 42 Сагех саезрНоза Ь. МК Многолетнее короткокорневищное растение, образуто-щее плотные дерновины и кочки. Корневая система мочковатая, достигающая (в основной своей массе) 30-40 см. Обычно растет по сырым лугам. Доминирует при сыролуговом и болотно-луговом увлажнении.
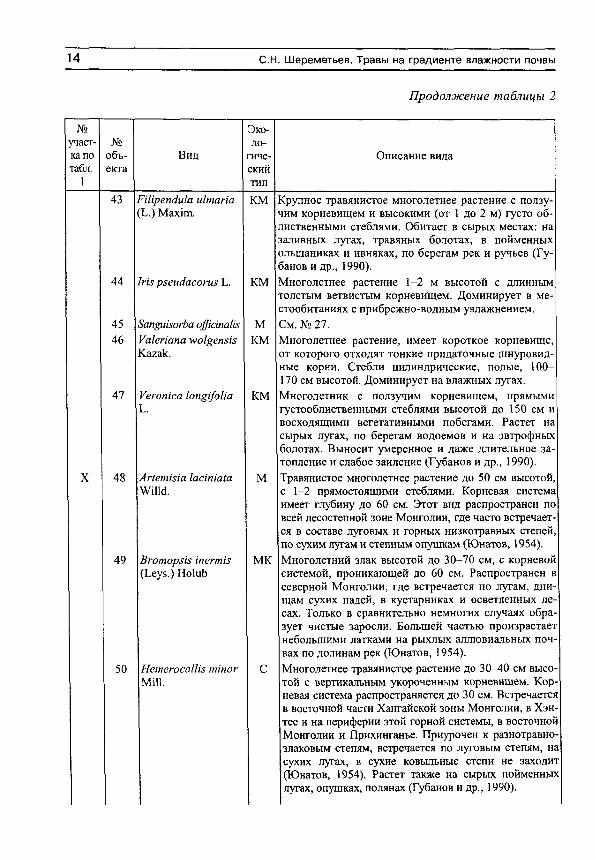
14 С.Н. Шереметьев. Травы на градиенте влажности почвы
Продолжение таблицы 2
№ Эко-участ № ло-ка по объ Вид гиче- Описание вида табл. екта ский
1 тип 43 РШрепМа и1тапа
(Ь.) Мах1пг КМ Крупное травянистое многолетнее растение с ползу
чим корневищем и высокими (от 1 до 2 м) густо облиственными стеблями. Обитает в сырых местах: на заливных лугах, травяных болотах, в пойменных ольшаниках и ивняках, по берегам рек и ручьев (Губанов и др., 1990).
44 / т рхеис1асоп15 Ъ. КМ Многолетнее растение 1-2 м высотой с длинным толстым ветвистым корневищем. Доминирует в местообитаниях с прибрежно-водным увлажнением.
45 8ап§и1зогЬа а(р1апаИ5 М См. № 27. 46 Уакпапа м>о1^епш
Кагак. КМ Многолетнее растение, имеет короткое корневище,
от которого отходят тонкие придаточные шнуровид-ные корни. Стебли цилиндрические, полые, 100-170 см высотой. Доминирует на влажных лугах.
47 Уеготса 1оп§1/оИа к м Многолетник с ползучим корневищем, прямыми густооблиственными стеблями высотой до 150 см и восходящими вегетативными побегами. Растет на сырых лугах, по берегам водоемов и на эвтрофных болотах. Выносит умеренное и даже длительное затопление и слабое заиление (Губанов и др., 1990).
X 48 Аг1ет1з1а 1аста1а тм.
м Травянистое многолетнее растение до 50 см высотой, с 1-2 прямостоящими стеблями. Корневая система имеет глубину до 60 см. Этот вид распространен по всей лесостепной зоне Монголии, где часто встречается в составе луговых и горных низкотравных степей, по сухим лугам и степным опушкам (Юнатов, 1954).
49 Вготорзк тегтЬ (Ьеуз.) Но1иЬ
МК Многолетний злак высотой до 30-70 см, с корневой системой, проникающей до 60 см. Распространен в северной Монголии, где встречается по лугам, днищам сухих падей, в кустарниках и осветленных лесах. Только в сравнительно немногих случаях образует чистые заросли. Большей частью произрастает небольшими латками на рыхлых аллювиальных почвах по долинам рек (Юнатов, 1954).
50 НетегосаШз ттог МШ.
с Многолетнее травянистое растение до 30^0 см высотой с вертикальным укороченным корневищем. Корневая система распространяется до 30 см. Встречается в восточной части Хангайской зоны Монголии, в Хэнтее и на периферии этой горной системы, в восточной Монголии и Прихинганье. Приурочен к разнотравно-злаковым степям, встречается по луговым степям, на сухих лугах, в сухие ковыльные степи не заходит (Юнатов, 1954). Растет также на сырых пойменных лугах, опушках, полянах (Губанов и др., 1990).

МАТЕРИАЛ И МЕТОДИКА 15
Окончание таблицы 2
№ Эко-участ № ло-ка по объ Вид гиче- Описание вида табл.
1 екта ский
тип 51 Ьеутиз сктепзЫ МК См.№ 15. 52 Ро1епШ1а апзегта Ъ. М Многолетнее стелющееся растение с тонкими ползу
чими, укореняющимися в узлах, стеблями. Корневая система достигает глубины 30 см. Встречается по всей северной лесостепной и степной зонам Монголии, также заходит в пустынно-степную зону. Этот вид приурочен к сырым луговым почвам, но не чрезмерно заболоченным. Сильно разрастается (до преобладания в травостое) в результате интенсивного выпаса (Юнатов, 1954).
53 8ап^13огЬа о]1}1стаШ М См. № 27. XI 54 АИзта р1ап1а§о-
ациаНса Ь. Гд Многолетнее короткокорневищное растение с при
даточными корнями. При обсыхании местообитаний (также как и другие исследованные гидрофиты) переходит в наземную форму. Цветет в июле. Растет на болотах и отмелях, по берегам водоемов (Цвелев, 1988).
55 ВШотиз итЬеИаШз Ь,
Гд Многолетнее короткокорневищное растение. Цветет в конце мая. Растет по берегам водоемов, на болотах (Цвелев, 1988).
56 Сагех асиШ Ь. Гг Многолетнее длиннокорневищное растение (Алексеев, 1996) с весенним сроком цветения. Образует пучки побегов (при застойном увлажнении — кочки) на ветвистом ползучем корневище. Растет на болотах и болотистых лугах, по берегам водоемов, часто длительно заливаемых (Губанов и др., 1990).
57 Ьусориз еигораеиз Ь. Гг Многолетнее корневищное растение. Цветет с конца июня по сентябрь. Растет на болотах, болотистых лугах, по берегам водоемов (Цвелев, 1988).
58 8а§Шапа за§Ш1[оИа Ь.
Гд Многолетний гидрофит. При переходе в наземную форму формирует толстые корневища. Цветет в июне -августе. Растет на болотах, по берегам водоемов (Цвелев, 1988).
59 8раг§апшт егесШт Ъ.
Гд Многолетний длиннокорневищный гидрофит. Цветет в июле - августе. Растет на болотах, по берегам водоемов (Цвелев, 1988).
Примечание: экологический тип по отношению к почвенному увлажнению: Гд — гидрофит, Гг — гигрофит, М — мезофит, КМ — ксеромезофит, МК — мезоксерофит, К — ксерофит, С — суккулент, СК — суккулент с коронарным синдромом (по: Шереметьев, 1991 с изменениями).

16 С.Н. Шереметьев. Травы на градиенте влажности почвы
Распределение экологических типов растений (по отношению к условиям почвенного увлажнения) в разных частях градиента влажности почвы неодинаково. В аридной части градиента присутствуют только мезоксерофиты, ксерофиты и суккуленты, с преимущественным преобладанием ксерофитов. В средней части градиента можно наблюдать представителей всех экологических типов (за исключением, разумеется, гигрофитов и гидрофитов). В этих экологических условиях преобладание получают мезоксерофиты, а также ксеромезофиты и мезофиты. В гумидной части градиента можно встретить представителей всех без исключения экологических типов.
Показатели водообмена каждого объекта наблюдений были подробно изучены в дневной и сезонной динамике. Параллельно вели наблюдения за температурой и влажностью воздуха и почвы, освещенностью, скоростью ветра.
Интенсивность транспирации исследовали методом быстрого взвешивания, дефицит водного насыщения — методом насыщения, оводненность листьев — весовым методом, осмотическое давление — криоскопическим.
Интенсивность транспирации и содержание воды в листьях растений пересчитывали на сухую массу навесок. Однако для совместимости с результатами других авторов в таблице 7.2 приведены значения скорости испарения воды, рассчитанные в иных единицах измерения.
При необходимости пересчёта данных на сырую массу навесок можно воспользоваться следующими простыми соотношениями:
Ж С , = ^ ^ (2)
где ТК^ и ТК^ — интенсивность транспирации (ТК — Тгап8р1га110п Ка1е) в пересчете на сырую (индекс /[гезЬ]) и сухую (индекс ^[гу]) массу навесок (размерность — ^воды •Гсуx.̂ иассы - 1 " ' , гдс I — единицз времсни), \УС^тл }УС^— содержание воды (\Уа1ег Соп1еп{) в навесках в пересчете на их сырую и сухую массы (размерность — ^воды • ^^.массы )• Если В ИСХОДНЫХ данных }УС^ выражено в процентах, то в формулах (1) и (2) нужно заменить }УС^ на УУС^^^^^ЮО . Если ЖС^ нужно выразить в процентах, то ^^С .̂,,)= Щ- 100.
Были подробно изучены структурные характеристики листьев (парциальные объемы эпидермиса, губчатой и столбчатой паренхимы, паренхимной обкладки пучков, проводящей ткани, водоносной паренхимы, межклеточных пространств столбчатой и губчатой паренхимы, воздушных полостей) у 24 видов растений, произрастающих в Хоперском заповеднике (сообщества под номерами I , V I , IX и X I в табл. 1, 2). Измерения площадей поперечного сечения и парциальных объемов клеток и тканей проводили в 4-7-кратной повторности с использованием программы иТН8С8А 1та§е Тоо1 (Ь11р://тасогЬ.и1Ь5С5а.е(1и/с11§/11с1е5С.Ь1т1).
Данные обрабатывали с помощью стандартных статистических методов. Множественный регрессионный анализ проводили с помощью программы «81ерКе§ге5810п

МАТЕРИАЛ И МЕТОДИКА 17
V. 3.2». Эта программа позволяет проводить симметризующие преобразования переменных, оценивать вес влияния каждой переменной на отклик и подбирать наиболее адекватную реальным данным структуру уравнений (см.: Шереметьев, Жаргалсайхан, 1990). Дневную динамику элементов водообмена обрабатывали с помощью программы «Со51пог V. 2.4» (ее описание помещено в раздел «Определения»). Эти и другие программы для статистической обработки данных без ограничений доступны по адресу: Ы1р://ехсе151а1;.пе\ута11.ги. Для анализа рядов распределения показателей водного режима был использован квантильный анализ. Квантили порядка 0.1 и 0.9 (нижний и верхний децили), отсекающие по 10% наименьших и наибольших значений, рассматриваются как типичный минимум и максимум признака. Разность между верхним и нижним децилями составляет амплитуду типичных колебаний (см. Васильев и др., 1973; Васильев и др., 1982). Мы не использовали квантильный анализ для исключения крайних значений рядов распределений (т. н. «выскакивающих» значений, — они не исключались совсем). Поэтому иногда среднедневные величины показателей водного обмена могут быть ниже типичных минимумов (или выше типичных максимумов).
Коэффициенты парной (г^) и множественной (Я^) детерминации, приводимые в работе, статистически достоверны (если это специально не оговорено) на доверительном уровне Р = 0.95. На рисунках, в тексте и таблицах имеются следующие обозначения: п — число наблюдений, х — средние и 8х — стандартные отклонения, V — коэффициент вариации, ИТ — интенсивность транспирации, ДВН — дефицит водного насыщения, НСВ — насыщающее содержание воды, СВ — содержание воды в листьях растений, ОД — осмотическое давление клеточного сока, 1С — индекс сложности организации водного режима растений. Размерность интенсивности транспирации (г„„йы '^с^массы - ч " ' ) на рисункзх обозначсна ДЛЯ краткости как «г/(г-ч)», размерность содержания воды (а также насыщающего содержания воды) ( г ^ О Й М '^сух.массы ) в листьях — как «г/г».

ОПРЕДЕЛЕНИЯ
Сложность организации водного режима растений
Очевидно, что средний уровень любого элемента водного режима для каждого вида определяется как внешними факторами (прежде всего запасами доступной почвенной влаги), так и его морфологическими, анатомическими и физиологическими особенностями. Глубина и объем корневой системы, структура ассимиляционного аппарата, тип проводящей системы, способность к функциональной регуляции водного режима (соотношение различных элементов функционирования), — все эти и другие свойства растений определяют конкретное количественное выражение параметров водообмена, другими словами — их конституционный уровень. Оценками конституционных уровней элементов водного режима являются оценки их средних значений за достаточно продолжительный период времени (при увеличении количества наблюдений эти оценки точнее описывают средние значения). Сезонные колебания показателей водообмена возле этих конституционных уровней могут быть описаны более простыми связями, поскольку определяются (в большинстве случаев) быстрыми изменениями влажности воздуха и почвы и (или), опосредованно, изменениями других составляющих водного режима растений. Влияние на эту динамику онтогенетических структурно-морфологических изменений, как несравненно более медленных и имеющих вполне определенный вектор, должно быть пренебрежительно мало.
Поэтому можно сказать, что если (1) конституционный уровень какого-либо показателя водного режима интегрирует ответ растения как целого на сложившуюся экотопическую и ценотическую обстановку, то (2) флюктуации возле этого уровня связаны, прежде всего, с потенциальными возможностями растения приводить в соответствие с динамикой факторов среды свою динамику функционирования. В первом случае связи сложны и многообразны, и часто с трудом поддаются интерпретации на небольшом (в пределах экотопа) градиенте факторов среды. Во втором случае можно выявить непосредственную зависимость показателя функционирования от факторов среды или его связь с другими элементами водообмена, что может свидетельствовать о степени выраженности регуляторных процессов, приводящих к образованию обратных связей между этими элементами.
Это можно сделать, исследуя коэффициенты детерминации между элементами водного режима растений. Коэффициенты детерминации не имеют самодовлеющего значения, а важны для выявления сложности организации водного режима растений. Термину «сложность» в нашем понимании семантически наиболее близок термин «комплексность».
Сезонные ходы показателей водного режима не представляют значительного интереса сами по себе. Их анализ необходим для выявления взаимосвязей между эле-

ОПРЕДЕЛЕНИЯ 19
ментами водообмена и их зависимостей от факторов среды. Из этих зависимостей складывается показатель, который мы назвали индексом сложности организации. Сложность организации водного режима какого-либо вида травянистого растения рассчитывается нами как средний коэффициент детерминации для всех обнаруженных связей между элементами водного режима этого вида, а также между этими элементами и факторами среды и отражает следующую смысловую основу. Принято считать, что организованность какой-либо функциональной системы можно оценивать с помощью тех связей, которые накладывают определенные ограничения на реализацию ее элементов (Василевич, 1983). Чем больше связей между элементами системы и чем эти связи сильнее, тем более сложно она организована. Распад всех связей или их слабая выраженность означает, по нашему мнению, что система перешла в такой режим функционирования, который диктуется ее конституционными свойствами, и она не испытывает заметного внешнего воздействия.
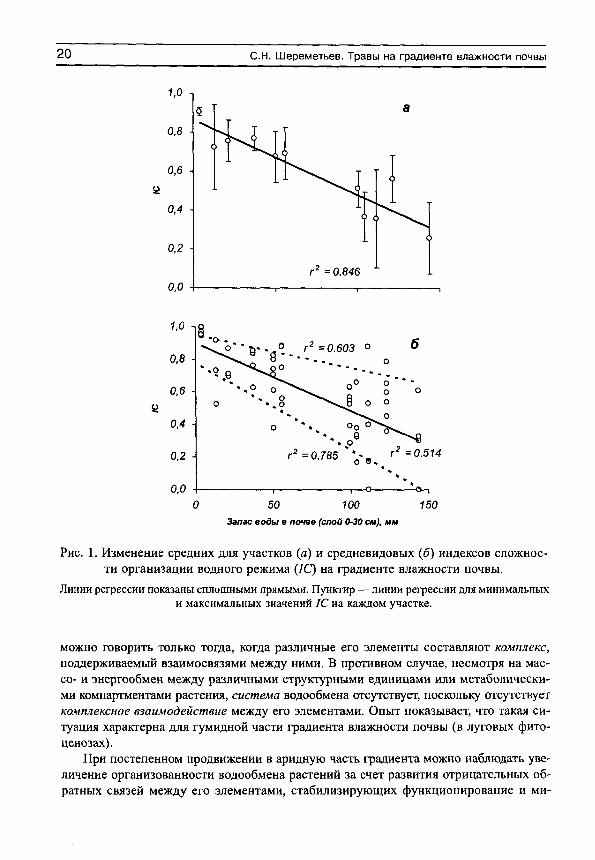
Основным свойством этого показателя является его увеличение (приближение к единице) при усилении водного стресса, и уменьшение (приближение к нулю) при оптимальных условиях почвенного водоснабжения. Фактические данные показывают, что организованность водообмена минимальна в гумидной части градиента влажности почвы и максимальна — в аридной (рис. 1, а). Более подробный анализ (рис. 1, б) показывает, что в гумидной части градиента влажности почвы могут встречаться виды как с высокими индексами организованности водообмена, так и с низкими. Это может быть обусловлено не столько воздействием факторов среды, сколько участием растений в межвидовых конкурентных взаимоотношениях. Можно предположить, что при сильном конкурентном прессе организованность водообмена возрастает, а при слабом — остается на низком уровне. По мере аридизации условий произрастания индексы организованности увеличиваются у всех видов, а диапазон их разброса сужается (рис. 1, б).
Из определения видно, что этот показатель может изменяться от О до 1, хотя, совершенно очевидно, нельзя ожидать достижения его верхней границы. По мере приближения к ней мы будем наблюдать усиление организованности водообмена за счет возникновения и развития связей между его элементами. Чем сильнее выражены эти связи, и чем большее число элементов они объединяют, тем более сложно (комплексно) организована система водообмена.
Поскольку растение функционирует как единое целое и процессы водообмена, фотосинтеза и дыхания протекают взаимосвязано, то можно сказать, что организованность водообмена является отражением организованности функционирования целого растения. Эта организованность развивается за счет возникновения отрицательных или положительных обратных связей между элементами системы. Поэтому часто невозможно выделить ведущий или ведомый элемент в такой системе: наличие обратных связей подразумевает взаимное угнетение или усиление взагшодействующих элементов.
Когда индекс сложности организации водного режима приближается или равен нулю, то мы имеем дело с полностью автономной реализацией каждого элемента функционирования: эта реализация задается только конституционными свойствами растений. Система функционирования если и существует, то только потенциально, поскольку существует структурная основа для ее развития. О системе водообмена
20 С.Н. Шереметьев. Травы на градиенте влажности почвы
1,0 -,
0,8 -
0,6 •
0,4 •
0,2
0,0 г'' = 0.846
1,0 -|
0,8 -
0,6 -
0,4 •
0,2 •
0,0 •
^ о ' - ^ . о г'=0.603 о ^ ° " * • * - о
„о о -° о о
о 50 100 150 Запас воды в почве (слой 0-30 си), мм
Рис. 1. Изменение средних для участков (я) и средневидовых (б) индексов сложности организации водного режима (/С) на градиенте влажности почвы.
Линии регрессии показаны сплошными прямыми. Пунктир — линии регрессии для минимальных и максимальных значений 1С на каждом участке.
можно говорить только тогда, когда различные его элементы составляют комплекс, поддерживаемый взаимосвязями между ними. В противном случае, несмотря на мас-со- и энергообмен между различными структурными единицами или метаболическими компартментами растения, система водообмена отсутствует, поскольку отсутствует комплексное взаимодействие между его элементами. Опыт показывает, что такая ситуация характерна для гумидной части градиента влажности почвы (в луговых фито-ценозах).
При постепенном продвижении в аридную часть градиента можно наблюдать увеличение организованности водообмена растений за счет развития отрицательных обратных связей между его элементами, стабилизирующих функционирование и ми-

ОПРЕДЕЛЕНИЯ 21
нимизирующих потери воды на испарение. Причины более тесной взаимозависимости элементов водного режима в условиях существенного ограничения ресурсов почвенной влаги следует искать, по всей видимости, как в структурных особенностях растений, так и в изменениях термодинамических показателей воды в них: несмотря на то, что при водном стрессе депрессия химического потенциала воды углубляется, а ее активность' снижается, коэффициент активности воды^ увеличивается за счет более быстрого снижения концентрации воды в растении по сравнению с ее активностью (Зялалов, 1984). Создается впечатление, что в условиях водного стресса растениям энергетически выгодно быстрое формирование водного дефицита, опережающее уменьшение активности воды. В этом случае развитие положительных обратных связей может способствовать увеличению коэффициента активности воды, за счет быстрого увеличения дефицита водного насыщения.
Насыщающее содержание воды
Обычно дефицит водного насыщения определяют как недонасыщенность ассимилирующих органов водой по формуле:
IV -IV / (1)
где И^50 — дефицит водного насыщения, безразмерная величина (при умножении правой части уравнения (1) на 100 дефицит насыщения выражается в процентах),
— масса листьев после полного насыщения водой, мг, — свежая масса листьев, мг, — сухая масса листьев, мг.
Поскольку относительное содержание воды (КЖС) в листьях равно:
ШС = -^ О )
то
\У80 = \-ШС. (3)
' Активность воды а „ — это концентрация воды в системе, учитывающая влияние всех ее компонентов («скорректированная» концентрация или термодинамически эффективная). Под системой понимается раствор каких-либо веществ, растительная клетка, лист или целое растение (КоЬе1, 1991). Для неидеальных растворов а „ = У^N^, где — коэффициент активности воды, Л^^ — молярная доля воды. Активность воды используется для определения химического потенциала системы: ц^-1х1= КТ\па„ , где — химические потенциалы раствора и чистой воды соответственно, К — универсальная газовая постоянная, Т — абсолютная температура (см., например, Слейчер, 1970, NоЬе1, 1991). Активность чистой воды равна 1 (химический потенциал чистой воды равен нулю). Эта величина безразмерная. Чем меньше активность воды в системе, тем ниже будет химический потенциал этой системы. ^ Для неидеальных растворов коэффициент активности воды равен отношению активности воды к молярной доле воды в растворе: у^^, =а„/М^ .

22 С.Н. Шереметьев. Травы на градиенте влажности почвы
При сравнении значений водного дефицита с параллельными данными по анатомической структуре листьев невозможно выявить, в какие их ткани поступает вода в процессе насыщения. Это ясно видно из выражения (1), в котором насыщающее количество воды относится к полному ее количеству после насыщения. Более правильным является расчет насыщающего количества воды отнесенного к сухой массе листьев. Мы назвали этот показатель насыщающим содержанием воды {'^VС) и определили как:
-1 Размерность этого показателя — г^^^^ • т'^ух.массы • Для того чтобы показать связь этого параметра с другими элементами водного
режима, необходимо сделать некоторые предварительные замечания. Определение содержания воды в листьях через выражение (2) кажется нам крайне
неудовлетворительным, хотя этот показатель широко используется в практике исследований водного режима растений. Считается, что он преодолевает проблему изменений сухой массы листьев в течение дня и/или сезона (см., например, Тигпег, 1981, р. 341). Однако он создает, на наш взгляд, еще большие проблемы, поскольку амплитуда дневных/сезонных колебаний количества воды в листьях (знаменатель формулы (2)), значительно превосходит такую амплитуду сухой массы листьев. Кроме того, нельзя сравнивать по параметру (2) виды, у которых конституционные уровни воды (обусловленные анатомической структурой ассимилирующих органов) различаются. Например, совершенно ясно, что, сравнивая по относительному содержанию воды склерофиты и суккуленты, мы неизбежно придем к неправильным выводам. Поэтому, мы считаем, что наиболее адекватной оценкой содержания воды в свежей навеске (И^Ср листьев является:
IV г-IV, у. }УСг = — (5)
Тогда суммирование (4) и (5) приведет к определению полного содержания воды {^VС^ в листьях (когда тургор максимален, а водный потенциал равен нулю):
же, =ЖСг+ЖС, =
Простые преобразования показывают, что:
ЖС^ = }УС,хШС.
Таким образом, видно, что насыщающее содержание воды является показателем, интегрированным в систему других показателей водного режима растений и его можно применять как одну из характеристик водообмена. Однако для анализа недостат-

ОПРЕДЕЛЕНИЯ 23
ка насыщения листьев водой на всем протяжении градиента влажности почвы мы использовали дефицит водного насыщения (1) как наиболее привычный и широко отраженный в литературе элемент водного режима. В то же время исследование такого показателя, как насыщающее содержание воды (4) было необходимо для того, чтобы попытаться выявить ткани, в которые может попадать вода в процессе насыщения ею листьев.
Показатели экологического сходства видов
Экологическое сходство видов оценивали по формуле (Джиллер, 1988):
^(со^+(о])/2 '
где Ру — безразмерная величина, характеризующая степень экологического сходства между видами г и у (считается, что если 1р| < 0.5, то сходство между видами велико, если |р | > 1, то сходство выражено слабо, если |р| > 3 , то сходство между видами не выражено), 4̂ , - — разность средних значений элементов водного режима, (0^,со^ — стандартные отклонения.
Очевидно, что эта формула пригодна для оценки экологического сходства видов, как по отдельным показателям, так и по их совокупности. Последнее, на наш взгляд, более предпочтительно, поскольку дает интегральную характеристику «сходства — различия» совместно произрастающих видов. Для оценки экологического сходства по совокупности изученных признаков (интенсивности транспирации, дефицита водного насыщения, содержания воды в листьях растений и осмотического давления клеточного сока) были рассчитаны средние оценки для вида по оценкам р,у для каждого показателя водного режима (эти оценки приводятся в матрицах экологического сходства в тексте книги). При изменении влажности почвы и воздуха интенсивность транспирации и содержание воды, с одной стороны, и дефицит водного насыщения в листьях растений и осмотическое давление, с другой, изменяются, как правило, в противоположных направлениях. Поэтому при вычислении оценки знак р,-, для дефицита водного насыщения и осмотического давления изменяли на противоположный. В противном случае эта оценка стремилась бы к нулю, что сделало бы невозможным выявление различий между видами.
В результате построены матрицы, в которых отражены индексы экологического сходства, как между отдельными видами сообщества, так и каждого вида с остальными. В этих таблицах виды ранжированы по возрастанию величины Р . По определению те из них, которые расположены в верхней части матриц, характеризуются наименьшими значениями интенсивности транспирации и оводненности (и показателями их варьирования) и наибольшими значениями (и показателями варьирования) дефицита водного насыщения и осмотического давления. Виды с противоположными свойствами водного режима занимают нижнюю часть матриц. Сочетание первой группы свойств характерно для более ксерофитных видов, сочета1ше последней группы характерно для более мезофитных растений. Другими словами, в верхней части

24 С.Н. Шереметьев. Травы на градиенте влажности почвы
матриц расположены относительно более ксерофитные виды, а в нижней — относительно более мезофитные по совокупности показателей водного обмена растений (подчеркнем относительность определения в данном контексте ксерофитности и мезофитности).
Описание косинор-анализа
Для анализа дневной динамики элементов водного режима был использован коси-нор-анализ. Его суть состоит в аппроксимации исходных данных синусоидами методом наименьших квадратов. Для этого была написана программа «Со51пог» для Ехсе! 2000/ХР по алгоритму, предложенному И. П. Емельяновым (1976, 1986). Существуют и другие алгоритмы косинор-анализа (На1Ьег§, 1969; Ые18оп е1 а1., 1979; В1п§Ьат е! а1., 1982; Багриновский и др., 1973 и др.). Одно из достоинств алгоритма И. П. Емельянова состоит в том, что измерения (служащие входной информацией для косинор-анализа) могут быть получены через разные интервалы времени и в разные часы суток (но не менее трех). Каждый массив наблюдений за один день будем называть хронограммой (в противоположность этому расчетные кривые — синусоидами). Тогда входной информацией для анализа суточных и ультрадианных ритмов транспирации будет массив хронограмм (ансамбль реализаций), полученных в течение вегетационного периода для каждого вида растения (см. табл. 3). Выходной информацией косинор-анализа являются основные параметры ритмов: мезор, т. е. величина среднего уровня синусоиды (Ь), амплитуда синусоиды (А) и акрофаза, т. е. время наступления максимума функции (ф) (рис. 2, а). Эти данные рассчитываются как для отдельных синусоид, так и для средней синусоиды (дальнейшее изложение будет опираться на анализ именно средних суточных и ультрадианных синусоид, причем в расчет будут браться, как правило, только те данные, которые находятся в пределах области интерполяции). Кроме того, рассчитывается и ряд дополнительных показателей: х, у, г, х^, у^, 8^, 5^, 6, 5, а, Ь, в (см. примечание к табл. 3) и собственно синусоиды. Среди этих показателей находятся и данные для построения эллипса ошибок (рис. 2, б).
Построение эллипса ошибок необходимо, в частности, для определения достоверности существования ритмов на принятом доверительном уровне (в нашей работе для суточных ритмов был принят уровень 0.95, для ультрадианных — 0.90), определения доверительных интервалов акрофаз с помощью касательных к эллипсу, опущенных из начала координат на циферблат (рис. 2, б). Наименьших затрат времени (порядка минут) для построения доверительных эллипсов можно добиться с помощью программы «СогеЮга\у», в которой легко и очень точно позиционируются любые графические объекты.
С положением акрофаз связана форма дневных ритмов транспирации (рис. 2, в). Если максимум транспирации наблюдается после полудня, то кривые имеют более или менее правильную куполообразную форму без заметной асимметрии. По мере смещения максимумов транспирации на ранние утренние часы все в большей степени проявляется правосторонняя асимметрия кривых транспирации, а течение большей части дня наблюдается ее депрессия. Эта депрессия может быть связана с недостаточным водоснабжением растений из-за низкой влажности почвы или конкуренции с другими видами сообщества (Шереметьев, 2002).

ОПРЕДЕЛЕНИЯ 25
8 9 10 11 12 13 14 15 16 17 18 19 20
Часы суток
Рис. 2.
а — основные характеристики дневных ритмов транспирации (на примере РоСепНИа апзегта, злаково-богаторазнотравное сообщество): мезор (Ь), амплитуда (А) и акрофаза (ф). Сплошная
линия — дневной ход транспирации, пунктир — расчетная синусоида. 6 — эллипс ошибок средней синусоиды для интенсивности транспирации РоШйИа апзегта на 24-часовом циферблате, ф̂ ^̂ ,̂ ф̂ ^̂ — доверительный интервал для ф; 2а, 2Ь — удвоенные большая и малая полуоси эллипса; 9 — угол наклона большой оси эллипса к оси абсцисс. Кружками обозначены параметры синусоид (в полярных координатах — модуль и фаза, которые равны амплитуде и акрофазе соответствующей синусоиды) для каждого дня наблюдений
(см. табл. 3). Остальные обозначения те же, что и на рис. 2, а. в — связь между положением акрофаз и формой дневных ритмов транспирации. 1 — Е1у1п^1а герепз, злаково-осоковое сообщество (ф = 13.8 ч), 2 — Зап^шзогЬа о^/^стаНз, крупноразнот-равно-дернистоосоковое сообщество (ф = 12.0 ч), 3 — 1у1Нгит у1г§а1ит, кровохлебковое сообщество (ф = 10.8 ч), 4 — ОаИит гШНетсит, разнотравно-днепровскоковыльное сообщество
(ф = 6.4 ч).

26 С.Н. Шереметьев. Травы на градиенте влажности почвы
Таблица 3 Пример исходных данных (РоСепННа апзегта, злаково-богаторазнотравное
сообщество) интенсивности транспирации (г^^^^г^ ^ „„с^ '̂ч) и расчета 24-часовых синусоид с помощью программы «Сохтог» (см. рис. 2, б)
Дни наблюдений Средняя Время Исходные хронограммы хроно
16.06.87 23.06.87 02.07.87 07.07.87 14.07.87 30.07.87 09.08.87 13.08.87 19.08.87 грамма
7.4 1.600 2,084 1,782 2,018 1,979 1,758 1,775 1,443 1,416 0.984 1.684 8.4 1.793 2,129 1,126 1,888 2,277 2,152 3,363 1,586 1,723 1,302 1.934 9.4 1.694 2,120 1,504 2,456 3,024 2,200 3,338 1.675 1,706 1.200 2.092
10.4 2.513 2,815 2,450 2,483 3,714 3,025 2,990 1,506 2,655 1,372 2.552 11.4 2.524 3,878 2,275 3,672 2.865 3,500 3,049 1.774 2,147 1.208 2.689 12.4 3.285 2,882 2,335 4,070 2,700 3,038 2,459 2,722 1,739 1,655 2.689 13.4 2.843 3,023 2,125 2.601 2,269 4.535 2.561 3.192 2.132 1,299 2.658 14.4 2.057 4,435 2,005 2,925 3,541 4,494 2,763 2,206 1,606 1,263 2.730 15.4 2.840 3,859 2,578 4,178 4,221 1,482 3.051 2.655 1,497 1,288 2.765 16.4 2.899 4,293 2,032 3,960 2,600 1,695 1,647 1,805 1,641 0,970 2.354 17.4 0.987 3,012 1,867 3,689 3,446 1,083 1,841 1.555 1,772 1,549 2.080 18.4 0.757 1,146 1,587 2,517 2,297 0,893 1,137 1,296 1,297 0,717 1.364
Параметры синусоид для каждого дня наблюдений X -2.004 -1,961 -0,838 -1,032 -0,824 -3,052 -1,369 -1.344 -0,661 -0,452
У -0,368 -0,895 -0,348 -0,917 -0,467 -0,285 0,261 -0,469 0,010 -0,058 к 0,850 1.623 1,400 2,261 2,330 0,551 1,687 1.047 1,369 0,945 А 2,038 2,155 0,907 1.381 0,948 3,065 1,394 1,424 0,661 0.456
9 12,7 13.6 13,5 14,8 14,0 12,4 11,3 13.3 11,9 12,5
6 0,443 0,652 0,284 0.528 0,572 0,669 0,423 0,371 0,276 0,197 Средняя
Время Расчетные синусоиды для каждого дня наблюаений. Период Т = 24 ч синусоида
7 1,014 1.265 1,280 1,642 2,092 1,065 2,293 0.942 1.550 1,066 1.415
8 1,534 1,827 1,517 1.982 2,337 1,829 2,597 1,313 1,708 1,121 1.777
9 2,007 2.376 1,746 2,342 2,582 2,507 2,839 1.666 1,843 1,223 2.113
10 2,402 2,873 1,952 2.696 2,810 3,051 3,003 1,977 1.946 1,307 2.402
11 2,691 3.285 2,119 3.020 3,005 3,425 3,077 2.224 2,010 1,366 2.622
12 2,854 3,583 2,238 3,293 3,154 3,603 3,056 2,391 2,030 1,397 2.760
13 2,881 3.748 2,299 3.495 3,247 3,572 2,942 2.467 2,005 1,396 2.805
14 2,770 3,768 2,300 3,613 3,277 3,336 2,742 2,446 1,936 1,365 2.755
15 2,527 3.642 2,239 3.639 3,243 2,910 2,470 2.329 1,829 1,305 2.613
16 2,171 3,378 2,120 3,571 3,146 2,324 2,146 2,125 1,691 1,221 2.389
17 1,724 2.995 1,953 3.414 2.994 1,616 1.789 1,848 1,530 1,117 2.098
18 1,218 2,518 1,748 3,178 2,797 0,836 1,426 1,516 1,359 1,002 1.760
19 0,687 1.980 1,519 2.880 2.567 0,036 1.081 1,152 1,188 0,883 1.397
Параметры средней синусоиды
-1,354 у, = -0,354 Н,= 1,406 Л = 1,399 9с = 12,976 <5с = 0.131
= 0,250 0,117 г = 0.186 Данные для построения эллипса ошибок
а = 0,798 Ь = 0,365 в = 6,329

ОПРЕДЕЛЕНИЯ 27
Часы суток цдсы суток
Рис. 3. Примеры дневных ритмов транспирации некоторых видов растений в разных условиях произрастания:
а) Ро1еп1И1а (апасеИ/оИа (ритм с периодом 24 ч), ковыльно-разнотравно-вострецовое сообщество, б) Сагех (1ипи8си1а (ритмы с периодами 24 и 4 ч), твердоосочково-змеевковое сообщество, в) Ьеутиз сктепз1з (ритмы с периодами 24 и 7.1 ч), ковыльно-разнотравно-вострецовое сообщество, г) 5ап§ш5огЬа о^/ьстаИв (ритм с периодом 3.1 ч), кровохлебковое сообщество.
Пунктиром показаны синусоиды с периодом 24 часа.
Примечание к таблице Ъ: х,у — координаты точки в прямоугольной системе координат, по которым определяют модуль и фазу (равные амплитуде и акрофазе) в полярной системе координат; к — мезор (величина среднего уровня 24-часовой синусоиды) (г„,̂ Уг ,̂,,̂ ,„„„'ч); Л — амплитуда (г,̂ „/г^^^^^_^^ч); ф — акрофаза (ч); 5— среднеквадратическое отклонение синусоиды от соответствующей хронограммы (те же параметры с индексом «с» относятся к средней синусоиде); 5̂ , 5^ — ощибки х^ а у/, г — коэффициент корреляции между х а у; а, Ь — большая и малая полуоси эллипса; в — угол наклона большой оси эллипса к оси абсцисс (в градусах).

28 С.Н. Шереметьев. Травы на градиенте влажности почвы
На рис. 3 приведены примеры средних хронограмм и соответствующих им средних суточных синусоид для некоторых видов растений с различными формами дневных ритмов транспирации. Анализ показал, что эти ритмы различаются не только визуально, но и по своим количественным характеристикам: 1) статистически достоверен только суточный ритм транспирации (рис. 3, а), 2) одновременно существуют статистически достоверные ритмы с периодами 24 и 4 ч (рис. 3, б), 3) 24 и 7 ч (рис. 3, в), 4) суточный ритм отсутствует (т. е. его существование невозможно доказать на принятом доверительном уровне и с имеющимся набором исходных данных, хотя объективно он может реализоваться), но ярко выражена гармоника с периодом 3 ч (рис. 3, г). Эти четыре формы кривых можно назвать характерными, поскольку именно ими (или их вариациями) исчерпываются почти все формы как индивидуальных, так и средних хронограмм, имеющихся в нашем распоряжении (не только транспирации, но и дефицита водного насыщения и оводненности листьев, в последнем случае с той разницей, что в дневные часы наблюдаются не акрофазы, а батифазы — время, в которое функция минимальна).
Наличие или отсутствие суточных или ультрадианных ритмов, как отмечено выше, определялось графически. Если какая-либо часть эллипса попадает на начало системы координат, то соответствующий ритм следует считать статистически недостоверным на принятом доверительном уровне. Напротив, существование ритмов считается доказанным, если эллипс не перекрывает начало системы координат. Например, можно показать, что у всех видов разнотравно-днепровскоковыльного сообщества не удалось выявить суточную периодичность транспирации (рис. 4, а). У видов злаково-осокового сообщества суточные ритмы испарения воды выявляются с высокой степенью достоверности (рис. 4, б). В чем причина таких различий в реализации процесса транспирации мы попытаемся разобраться в соответствующих разделах книги. Здесь же необходимо отметить следующее.
Рис. 4. Примеры доверительных эллипсов суточных ритмов транспирации для видов разнотравно-днепровскоковыльного (а) и злаково-осокового (б) сообществ.

ВВЕДЕНИЕ 29
Отсутствие статистически значимых суточных ритмов может быть вызвано тремя причинами. Первая причина вызвана просто тем, что амплитуды составляющих среднюю хронограмму исходных хронограмм невелики в силу, например, напряженности факторов среды и периодичность процесса в целом не поддается определению (именно эта ситуация отражена на рис. 4, а). Вторая причина обусловлена ярко выраженной ультрадианной периодичностью (при высоком среднем уровне транспирации), которая маскирует суточную периодичность. Именно по этой причине мы не смогли обнаружить суточные ритмы у нескольких видов растений. Третья причина состоит в том, что в исходных индивидуальных хронограммах ритмы прослеживаются достаточно отчетливо, но все они в течение сезона наблюдений смещаются, так что в средней хронограмме (и синусоиде) они нивелируются.
С другой стороны, у тех видов, для которых выявлены суточные ритмы транспирации, индивидуальные (для каждого дня наблюдений) хронограммы должны достаточно хорощо совпадать, не очень сильно смещаясь относительно друг друга в течение сезона вегетации, иметь достаточно высокую интенсивность процесса транспирации, так что в целом можно выявить компактные эллипсы ошибок, свидетельствующие о наличии суточной периодичности (рис. 4, б).
Показатели варьирования признаков
Важным моментом исследования водообмена растений является анализ динамики показателей вариации, поскольку считается, что размеры амплитуд признаков можно рассматривать как одно из выражений физиологической адаптации. Существует мнение, что «успех существования и доминирования ряда видов в засушливых ... условиях произрастания зависит наряду с другими обстоятельствами от способности в широких пределах изменять интенсивность главных элементов водного баланса растений» (Свешникова, 1979: с. 176-177; Свешникова, 1993). При этом под амплитудой понимается разность между максимальными и минимальными величинами какого-либо признака (или квантилями того или иного порядка — например, децилями или квартилями, принятыми за границы «типичных» изменений признака) (Свешникова, 1979; Бобровская, 1991).
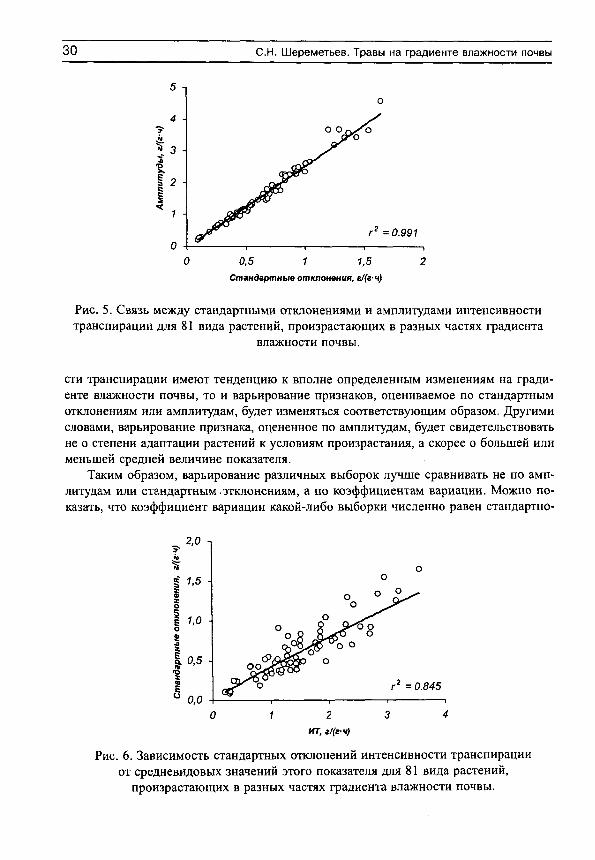
Рассмотрим сначала взаимосвязь между обычной статистической мерой варьирования признака (стандартным отклонением) и амплитудой (рассчитываемой нами как разность между верхним и нижним децилями) для имеющихся в нашем распоряжении выборок, например, по интенсивности транспирации (рис. 5, г^= 0.991). Оказывается, что эти меры варьирования настолько тесно связаны друг с другом, что в статистическом отношении являются эквивалентными. Поэтому, поскольку нельзя сравнивать варьирование выборок с различающимися средними значениями по стандартным отклонениям (см., например, Лакин, 1980; Зайцев, 1984), постольку нельзя сравнивать их и по амплитудам.
Для этих целей статистически адекватным является использование стандартных отклонений нормированных средними значениями, т. е. коэффициентов вариации. Это следует также из того, что стандартные отклонения (а, следовательно, и амплитуды) каждой выборки достаточно тесно связаны с их средними значениями (рис. 6). Но так как средневидовые (а также средние для участков) оценки показателей интенсивно-
30 С.Н. Шереметьев. Травы на градиенте влажности почвы
5 т
о
О 0.991
О 0,5 1 1,5
Стандартные отклонения, г/(г-ч) 2
Рис. 5. Связь между стандартными отклонениями и амплитудами интенсивности транспирации для 81 вида растений, произрастающих в разных частях градиента
влажности почвы.
сти транспирации имеют тенденцию к вполне определенным изменениям на градиенте влажности почвы, то и варьирование признаков, оцениваемое по стандартным отклонениям или амплитудам, будет изменяться соответствующим образом. Другими словами, варьирование признака, оцененное по амплитудам, будет свидетельствовать не о степени адаптации растений к условиям произрастания, а скорее о большей или меньшей средней величине показателя.
Таким образом, варьирование различных выборок лучше сравнивать не по амплитудам или стандартным отклонениям, а по коэффициентам вариации. Можно показать, что коэффициент вариации какой-либо выборки численно равен стандартно
го -,
о
0.845 0,0
О 1 2 3 4 ИТ, г/(гч)
Рис. 6. Зависимость стандартных отклонений интенсивности транспирации от средневидовых значений этого показателя для 81 вида растений,
произрастающих в разных частях градиента влажности почвы.

ВВЕДЕНИЕ 31
му отклонению той же выборки, но с удаленным средним. Предположим, что у нас имеется некоторая выборка X со средним X, стандартным отклонением 8^ и наблюдениями 1...П. Коэффициент вариации этой выборки равен
X (6)
Видоизменим исходную выборку таким образом, чтобы каждая варианта (2.) новой (стандартизованной) выборки (2) была равна
Тогда среднее {2) стандартизованной выборки будет равно
^ 1 42: х^-х X
= 0
а стандартное отклонение (5)
Поскольку среднее преобразованной выборки равно нулю, то:
5 . = х'
(7)
Идентичность правых частей уравнений (6) и (7) показывает, что К = 5.. Таким образом, сравнение варьирования выборок по коэффициентам вариации равносильно сравнению тех же выборок, но с удаленным средним, по их стандартным отклонениям. Заметим также, что коэффициента вариации ряда 7, не существует, поскольку его среднее равно нулю.

Ч А С Т Ь I
СЕЗОННАЯ ДИНАМИКА ВОДНОГО РЕЖИМА ДОМИНАНТОВ И ЭДИФИКАТОРОВ
СООБЩЕСТВ В РАЗНЫХ ЧАСТЯХ ГРАДИЕНТА ВЛАЖНОСТИ ПОЧВЫ
ГЛАВА 1. АРИДНАЯ ЧАСТЬ ГРАДИЕНТА
1.1. Разнотравно-днепровскоковыльное сообщество
Разнотравно-днепровскоковьшьное сообщество расположено на пологом песчаном склоне надпойменной террасы и относится к группе типов весьма краткопоемных лугов, занимающих экотопы с отметками выше 600-700 см над меженью р. Хопер (Титов, Печенюк, 1990). Почва здесь аллювиальная дерновая песчаная среднегуму-сированная с хорошо выраженным иллювиально-железистым горизонтом на глубине 60-100 см. Травостой редкий, проективное покрытие не более 60-65%. Средняя высота травостоя 40-50 см. Растительность сообщества разнообразна — насчитывается свыше 30 видов. В травостое содоминируют ЗНра ЬогузЛетса К1ок. ех Ргокис!., А§горугоп 1аугепсоапит Ргокий., Аг1ет1з1а тагзскаШапа 8ргеп§., А. аш(паса ^ас^., Роа ап^изИ/оИа Т., 5Иепе Ьогуз1Нетса (Огип.) ХУакегк. Массово встречаются Сагех со1сЫса } . Оау, С. зирта \\'аЫепЬ., РезШса \а1ез1аса ОаисИп, АШит зркаегосагрит. В наборе видов обычны такие виды, как Уеготса зргсаШ Ь., Егузтит <И//изит ЕЬгЬ., Ро(епШ1а аг§еп1еа Т., 8ес1ит асге Ь., 5. ШерНшт Ь., СаНит гШкетсит Весной массово встречаются эфемеры: Уеготса уегпа Ь., Уеготса М1епп СгаШг, Тп/оИит агуепзе Ь., МуозоИз з(пс(а Ыпк ех Коет . Объектами наблюдений в этом сообществе были выбраны следующие виды: Аг1ет1з1а аизШаса, Сагех со1сМса, ОаИит гШИет-сит, 5Нра ЬогузСНетса, Тп/оИит ап>епзе, Уеготса зргсаШ.
Условия произрастания во время проведения наблюдений в этом сообществе были близки к критическим: средний запас влаги в почве составлял 3.9 мм в верхнем (0-30 см) слое почвы, а максимальное значение дефицита влажности воздуха достигало 66 мб.

Глава 1. АРИДНАЯ ЧАСТЬ ГРАДИЕНТА 33
Исходные данные
Интенсивность транспирации в среднем для всех изученных видов была очень низка (0.286 г̂ ^^ ,̂ -г'^^^д^^.^, ч " ' : табл. 1.1.1). Это было обусловлено очень суровыми условиями произрастания во время проведения наблюдений. Эти условия были даже более суровыми, чем те, которые мы наблюдали в остепненной пустыне Заал-тайской Гоби МНР. Можно отметить, что средний уровень испарения воды растениями данного сообщества был самым низким на градиенте влажности почвы. Поэтому и варьирование этого признака было достаточно умеренным (в обычных условиях процессы, имеющие ярко выраженную суточную периодичность варьируют в очень широких пределах). Коэффициенты вариации интенсивности транспирации для всех видов находились в диапазоне 30-56%.
Колебания среднедневных значений скорости испарения воды возле средних уровней происходили в тесной связи с дефицитом водного насыщения у Аг1ет1з1а аш1паса, Сагех сокЫса и 8Нра Ьогу51кетса (коэффициенты детерминации равны 0.948^0.996). Зависимости среднедневных величин транспирации от оводненности листьев обнаружены у Сагех сокЫса ( г = 0.986) и ЗНра Ьогуз1кетса (г^ = 0.993). Воздействие факторов среды на испарение воды ограничивалось влиянием влажности почвы у СаИит гШкетсит (г^ = 0.928) и температуры воздуха у Уеготса зргсаШ {г' = 0.920). Во всех остальных случаях не обнаружено сколько-нибудь заметного воздействия факторов среды на транспирацию. Сухость воздуха и почвы оказывали столь сильное угнетающее воздействие на этот процесс, что какие-либо их изменения уже не воспринимались растениями.
Дефицит водного насыщения в листьях растений, в силу указанных обстоятельств, был очень высок. В среднем он составлял 61.9% (табл. 1.1.1), что указывает на критический уровень условий произрастания для жизнедеятельности растений в данный вегетационный период. Так, Тп/оНит агуепзе к середине июля практически закончил вегетацию из-за сильной засухи, а Уеготса зркаШ и АПетта аш1паса были близки к такому состоянию. Это нашло отражение в самых высоких значениях водного дефицита в листьях растений этих видов. В наилучшей степени засухе противостояли Сагех сокЫса и 5Нра ЬогузЛетса и имели относительно самые низкие величины дефицита водного насыщения. Можно отметить низкие величины варьирования этого признака. Коэффициенты вариации находились в пределах от 5 (у Уеготса зрка(а) до 21% (у Сагех сокЫса и Зйра Ьогуз1кепка).
Выявлены зависимости дефицита водного насыщения от оводненности листьев у всех видов (за исключением Уегопка зрка(а) (Н = 0.940^0.999). Также обнаружено, что водный дефицит связан со скоростью испарения воды (Н = 0.996-^0.999). Все зависимости отрицательны, за исключением корреляций между водным дефицитом и транспирацией у СаНит гШИепкит и Уегопка зркаШ, которые описываются полиномом второй степени. Не обнаружено воздействия факторов среды на дефицит водного насыщения.
Содержание воды в листьях растений данного сообщества составляло в среднем 0.812 г_̂ „дУг̂ у̂ „а;,;,„ (табл. 1.1.1). Средневидовые величины оводненности листьев находились в узком диапазоне 0.63-0.99 г̂ „дУг̂ у, „̂ сш- Столь сильное обезвоживание листьев ставило растения на грань выживания. Особенно страдали от такого обезвожи-
2 - 2353
34 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ.
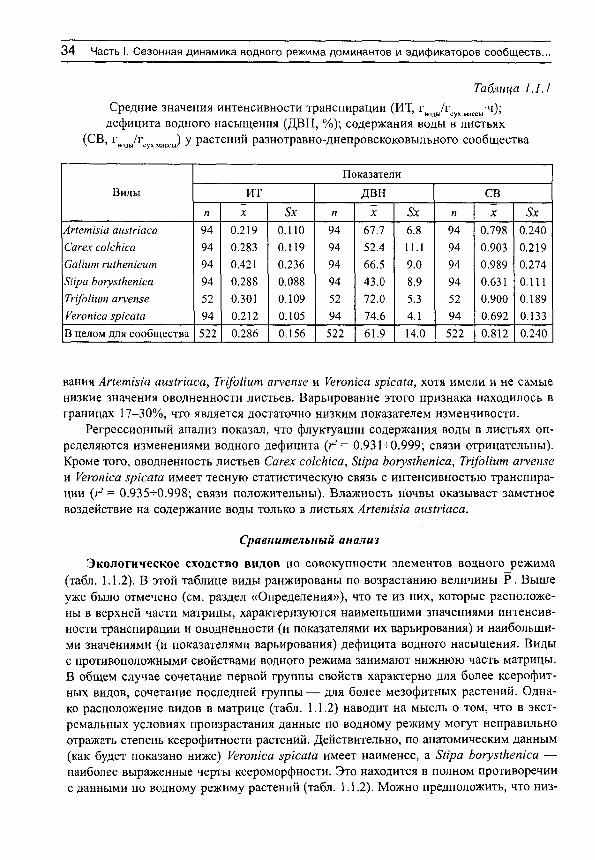
Таблица 1.1.1 Средние значения интенсивности транспирации (ИТ, Г|̂ „дУг̂ у,̂ „ас;,„ч);
дефицита водного насыщения (ДВН, % ) ; содержания воды в листьях (СБ, г̂ „д„/г̂ .у„ „^^^^ у растений разнотравно-днепровскоковыльного сообщества
Виды Показатели
Виды ИТ ДВН СВ Виды
п X 8х п X & п X Апетта ашШаса 94 0.219 0.110 94 61.1 6.8 94 0.798 0.240 Сагех сокЫса 94 0.283 0.119 94 52.4 11.1 94 0.903 0.219 СаИит гШкетсит 94 0.421 0.236 94 66.5 9.0 94 0.989 0.274 8Ира Ьогу51кеп1са 94 0.288 0.088 94 43.0 8.9 94 0.631 0,111 Тп/оИит агуепзе 52 0.301 0.109 52 72.0 5.3 52 0.900 0.189 Уеготса зркаШ 94 0.212 0.105 94 74.6 4.1 94 0.692 0.133 В целом для сообщества 522 0.286 0.156 522 61.9 14.0 522 0.812 0.240
вания АПетша аи$1паса, Тп/оНит агуете и Уегопка зркаШ, хотя имели и не самые низкие значения оводненности листьев. Варьирование этого признака находилось в границах 17-30%, что является достаточно низким показателем изменчивости.
Регрессионный анализ показал, что флуктуации содержания воды в листьях определяются изменениями водного дефицита (И = 0.93 КО.999; связи отрицательны). Кроме того, оводненность листьев Сагех сокМса, 8Ира Ьогу51кепка, Тп/оНит агуепзе и Уегопка зркаШ имеет тесную статистическую связь с интенсивностью транспирации (г^ = 0.935-^0.998; связи положительны). Влажность почвы оказывает заметное воздействие на содержание воды только в листьях АПетгзга аш(паса.
Сравнительный анализ
Экологическое сходство видов по совокупности элементов водного режима (табл. 1.1.2). В этой таблице виды ранжированы по возрастанию величины Р . Выше уже было отмечено (см. раздел «Определения»), что те из них, которые расположены в верхней части матрицы, характеризуются наименьшими значениями интенсивности транспирации и оводненности (и показателями их варьирования) и наибольшими значениями (и показателями варьирования) дефицита водного насыщения. Виды с противоположными свойствами водного режима занимают нижнюю часть матрицы. В общем случае сочетание первой группы свойств характерно для более ксерофит-ных видов, сочетание последней группы — для более мезофитных растений. Однако расположение видов в матрице (табл. 1.1.2) наводит на мысль о том, что в экстремальных условиях произрастания данные по водному режиму могут неправильно отражать степень ксерофитности растений. Действительно, по анатомическим данным (как будет показано ниже) Уегопка зркаШ имеет наименее, а 8Ира ЬогузЛепка — наиболее выраженные черты ксероморфности. Это находится в полном противоречии с данными по водному режиму растений (табл. 1.1.2). Можно предположить, что низ-
Глава 1. АРИДНАЯ ЧАСТЬ ГРАДИЕНТА 35
кие величины интенсивности транспирации и содержания воды, а также очень высокие значения водного дефицита в листьях растений связаны не с высокой степенью ксероморфности, а скорее с завяданием растений в условиях сильного водного стресса, с начинающейся потерей способности к функциональной регуляции водообмена. В наибольшей степени это относится к Тп/оНит агуепзе и Уеготса зркаШ, и в несколько меньшей — к АПетгзга аш1г1аса. Что касается 8Нра ЬогузЛетса, то он занял свое место наименее ксерофитного вида (табл. 1.1.2) в первую очередь из-за очень низкого (по сравнению с другими растениями) дефицита водного насыщения (табл. 1.1.1).
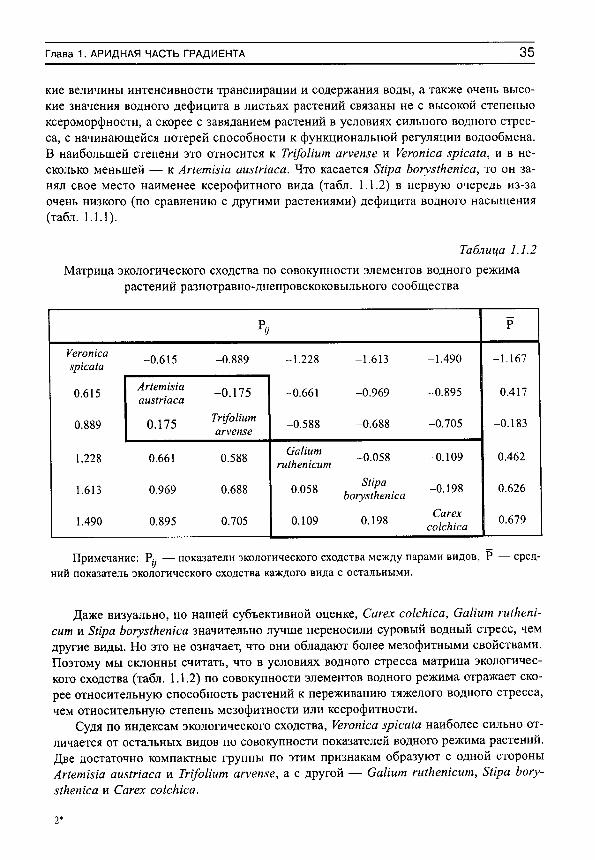
Таблица 1.1.2 Матрица экологического сходства по совокупности элементов водного режима
растений разнотравно-днепровскоковыльного сообщества
Р
Уеготса 5р1са1а -0.615 -0.889 -1.228 -1.613 -1.490 -1.167
0.615 Аг1етта аиз1паса -0.175 -0.661 -0.969 -0.895 -0.417
0.889 0.175 Тп/оИит ап'впзе -0.588 -0.688 -0.705 -0.183
1.228 0.661 0.588 СаНит Шкетсит -0.058 -0.109 0.462
1.613 0.969 0.688 0.058 8Ира Ьогуз1кетса -0.198 0.626
1.490 0.895 0.705 0.109 0.198 Сагех сокЫса 0.679
Примечание: Р^ — показатели экологического сходства между парами видов, Р — средний показатель экологического сходства каждого вида с остальными.
Даже визуально, по нашей субъективной оценке, Сагех соккка, СаИит гШкетсит и Бйра Ьогуз1кепка значительно лучше переносили суровый водный стресс, чем другие виды. Но это не означает, что они обладают более мезофитными свойствами. Поэтому мы склонны считать, что в условиях водного стресса матрица экологического сходства (табл. 1.1.2) по совокупности элементов водного режима отражает скорее относительную способность растений к переживанию тяжелого водного стресса, чем относительную степень мезофитности или ксерофитности.
Судя по индексам экологического сходства, Уегопка вркаШ наиболее сильно отличается от остальных видов по совокупности показателей водного режима растений. Две достаточно компактные группы по этим признакам образуют с одной стороны АПетша аш1паса и Тп/оНит агувте, а с другой — СаНит гШкепкит, 8Ира Ъогу-з1кепка и Сагех соккка.

36 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ..
Сложность организации водного режима растений. Напомним, что основным свойством этого показателя является его увеличение (приближение к единице) при усилении водного стресса, и уменьшение (приближение к нулю) при оптимальных условиях почвенного водоснабжения.
В исследованном сообществе средний для всех видов индекс сложности организации водного режима был очень высок и составил 0.969, т. е. для каждого вида (кроме Тп/оНит аггепзе, для которого индекс не рассчитывался) характерны очень высокие (близкие к единице) индексы организованности водообмена растений. Это дает повод предположить, что у растений в значительной степени была развита функциональная регуляция водного режима в ответ на водный стресс. Однако, несмотря на значительное развитие такой регуляции, Аг1ет181а аиз1паса, Тп/оНит агуепзе и Уеготса зр1са1а переносили водный стресс значительно хуже, чем остальные виды.
Растения разнотравно-днепровскоковыльного сообщества во время проведения наблюдений испытывали сильный водный стресс, что нашло отражение в высоких индексах организованности водообмена. Особенно это касается Сагех сокЫса и 8Ира ЬогузЛепка, для которых оценки этих индексов приближались к максимально возможной величине. Это свидетельствует, на наш взгляд, об усилении функциональной регуляции водного режима, которая в данных условиях позволяет сохранить жизнеспособность при минимально доступных ресурсах среды. Одним из аспектов такой регуляции может быть формирование отрицательных обратных связей между элементами функционирующей системы. Такие связи должны стабилизировать водообмен на таком уровне, который бы не являлся разрушительным для растений. Поэтому элементы водного режима в стрессовых условиях слабо связаны с факторами среды и сильно взаимосвязаны друг с другом. Однако в системе функциональной регуляции сохраняются все же положительные связи, которые позволяют отслеживать изменения в экологической обстановке, т. е. имеют, возможно, определенное сигнальное значение. Такими связями могут быть отмеченные выше сочетания транспирации и оводненности листьев. При увеличении количества доступной в почве влаги любой из этих элементов выйдет на более высокий уровень, что неизбежно повлечет за собой изменения в других элементах функциональной системы.
Морфолого-анатомическая характеристика
АНетша аив1паса имеет листья с шелковисто-беловато-шерстистым опушением. Отличается рыхлым изолатеральным мезофиллом, достаточно большим объемом межклетников, однослойной палисадной паренхимой с довольно крупными вытянутой формы клетками, трехслойной относительно плотной губчатой паренхимой, что говорит о скорее мезофитном строении листа. Но в тоже время, наличие моторных клеток, довольно толстой кутикулы, опушения, и различие между относительно мелко-клетной верхней и более крупноклетной нижней эпидермой являются явно ксеромор-фными признаками. Устьица расположены с нижней стороны листа и довольно часто, что в свою очередь также является признаком ксероморфности. Количественные соотношения тканей листа приведены в таблице 1.1.3.
Сагех соккка — растение, имеющее узкие листья с явными признаками ксероморфности. Характерным анатомическим признаком является довольно плотный го-

Глава 1. АРИДНАЯ ЧАСТЬ ГРАДИЕНТА 37
Таблица 1.1.3
Парциальный объем тканей и свободных пространств листьев растений разнотравно-днепровскоковыльного сообщества
Виды Э Пр ПО Пг Пс П Мг Мс М АПетша аизШаса 14.5 7.2 7.1 13.6 33.8 47.3 7.1 16.8 23.8 Сагех со1сЫса 15.7 13.3 5.5 40.9 0 40.9 8.5 0 8.5 ОаНит гиЛетсит 17.1 10.1 3.9 16.2 26.0 42.3 11.3 15.4 26.7 8Ира ЪогуННетса 7.4 13.9 3.7 32.0 0 32.0 2.6 0 2.6 Тп/оИит агуете 28.8 8.6 4.6 23.2 17.4 40.6 9.3 3.4 12.7 Уеготса зр1саш 12.7 5.4 7.4 24.5 37.2 61.7 7.2 5.6 12.7
Примечание: Э — эпидермис, Пр — проводящие ткани, ПО — паренхимная обкладка пучков, Пг — губчатая паренхима, Пс — столбчатая паренхима, П — паренхима, Мг — межклетники губчатой паренхимы, Мс — межклетники столбчатой паренхимы, М — межоетники.
могенный мезофилл, состоящий из мелких клеток, имеются воздушные полости. Сверху и снизу проводящих пучков представлены тяжи механических тканей. Эпи-дермальные клетки с обеих сторон листа одинаковых размеров, с довольно толстыми клеточными оболочками.
СаИит ги1кетсит имеет линейные нитевидные листья, по 8-10 в мутовке. С внутренней (адаксиальной) стороны листа отмечено заметное опушение, представленное одноклеточными простыми волосками что, наряду с наличием относительно толстой кутикулы и более крупных клеток эпидермы с абаксиальной стороны листа по сравнению с внутренней — является признаком ксероморфизма. Этому виду присуще наличие типичного дорзовентрального мезофилла. Палисадная паренхима 2-3-слойная, клетки довольно крупные, вытянутой формы. Губчатая паренхима рыхлая, только в районе главного пучка более плотно сложена, 4-5-слойная, изодиаметричес-кая, с крупными межклетниками. Устьица с внутренней стороны листа довольно крупные, слегка приподняты над поверхностью, подустьичные полости крупные.
8Ира Ьогуз1Нетса имеет очень жесткие, гладкие, узкие листья. Мезофилл — плот-ногомогенный, клетки мелкие, слегка вытянутой формы, плотно сомкнутые, с утолщенными стенками. Межклетников мало. Основную часть однослойной эпидермы составляют мелкие клетки с довольно толстой кутикулой, особенно на абаксиальной стороне. На этой же стороне листа имеются относительно мелкие моторные клетки. Листья сильно склерофицированные, волокна собраны в продольные тяжи, расположенные между пучками и эпидермой. Мелкие пучки связаны склеренхимой только с абаксиальной эпидермой.
Тп/оНит агуепзе — однолетнее растение с тонкими, прямостоячими, разветвленными, покрытыми волосками стеблями. Мезофилл — дорзовентральный. Палисадная паренхима однослойная, губчатая — довольно плотная, в основном изодиаметричес-кая (иногда можно заметить клетки вытянутые в горизонтальном направлении), многослойная. Верхняя и нижняя эпидерма по размеру клеток почти не различается, крупноклетная с тонким слоем кутикулы. Устьица не крупные. Имеется довольно сильное опушение.

38 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ.
Таблица 1.1.4 Матрица анатомического сходства по совокупности показателей анатомической
структуры листьев растений разнотравно-днепровскоковыльного сообщества
Р
8Ира Ьогу51кетса -0.307 -1.230 -1.781 -2.756 -3.358 -1.886
0.307 Сагех сокЫса -0.591 -1.169 -1.287 -3.084 -0.929
1.230 0.591 Уеготса зркаШ -0.593 -1.149 -1.118 -0.208
1.781 1.169 0.593 Тп/оНит ап>ете
-1.502 -1.604 0.087
2.756 1.287 1.149 1.502 Аг1ет1з1а ашШаса -0.093 1.320
3.358 3.084 1.118 1.604 0.093 СаИит гШкетсит 1.851
Рис. 1.1.1. Зависимость между индексами экологического сходства (по совокупности элементов водного режима) и парциальными объёмами проводящих тканей (а)
и паренхимы (б). Здесь и на рис. 1.1.2-1.1.8 точки — средневидовые значения показателей.
Уегопка $рка1а — растение с узкими, супротивно расположенными листьями. Характерным анатомическим признаком является довольно плотный дорзовентральный мезофилл, 3-слойная палисадная паренхима. Клетки вытянутой формы, не крупные. Губчатая паренхима достаточно плотная, клетки средней величины, межклетники мелкие. Клетки верхней эпидермы несколько меньших размеров, чем нижней. Одним из признаков ксероморфности является опушение короткими простыми волосками.

Глава 1. АРИДНАЯ ЧАСТЬ ГРАДИЕНТА 39
Для выявления количественных оценок сходства или различия видов по совокупности показателей анатомической структуры листьев построена матрица анатомического сходства листьев растений (табл. 1.1.4).
Относительно более ксероморфные виды характеризуются большим объемом эпидермиса, склеренхимы, паренхимы, проводящей ткани, и меньшим объемом межклетников. Они расположены в верхней части табл. 1.1.4. Это Сагех сокЫса, 8Нра Ьогуз-(кепка и, судя по всему, Уеготса зркаШ. Относительно более мезоморфной структурой листьев обладают Тп/оНит агуете, АШтша аш(паса и СаНит гШкепкит. Наибольшим сходством по соотношению тканей в листьях показывают две пары видов: Сагех соккка — 8ира ЬогузЛетса и Аг1ети1а аиз1паса — СаНит гигкепкит (табл. 1.1.4). Также можно отметить определенное сходство между 5Нра Ъогуз1кепка и Уегопка зркаШ, и между Уегопка зркаШ и Тп/оНит атуепзе. Но из этого не возникает такого же сходства по соотношению показателей водного режима (за исключением пары видов Сагех соккка — 8Нра Ьогуз(кепка) (табл. 1.1.2). Сравнение индексов экологического сходства видов по совокупности показателей водного режима и анатомической структуры показывает их полную несогласованность (табл. 1.1.2 и 1.1.4). Это, судя по всему, связано с тем, что показатели водного режима растений отражали не экологический статус видов, а степень критичности условий произрастания для процессов жизнедеятельности растений.
Вместе с тем, можно показать, что индекс экологического сходства в очень сильной степени зависит от парциальных объемов проводящих тканей (рис. 1.1.1, а) и паренхи-
^ мы и столбчатой паренхимы (а), меж-0 5 10 15 20 25 30 клетников столбчатой паренхимы (б).
Парциальный объём иежкпетников, % МСЖКЛСТНИКОВ ( в ) .

40 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ...
Парциальный объём проводящих тканей, % Парциальный объём проводящих тканей и паренхимной обкладки пучков, %
Рис. 1.1.3. Зависимости между парциальными объёмами столбчатой паренхимы и проводящих тканей (а), суммарным объёмом проводящих тканей и паренхимной
обкладки пучков (б).
70
60
I 50
I 40 •
30 г' =0.812
70 п
60 •
50 •
40
- , , , 30 11 13 15 3
=0.750
Парциальный объём проводящих тканей, % Парциальный объем паренхимной обкладки пучков, Уо
Рис. 1.1.4. Связи между парциальными объёмами паренхимы и проводящих тканей (а), паренхимной обкладки пучков (б).
мы (рис. 1.1.1, б) в листьях растений. Виды, которые лучше переносили засуху, имели в листьях относительно больший объем проводящих тканей, но меньший — паренхимы. Последнее обстоятельство связано с тем, что парциальные объемы проводящих тканей и паренхимы (особенно столбчатой) коррелируют между собой отрицательно.
Анализ соотношения тканей у разных видов показал наличие очень тесных статистических связей между некоторыми из них.
Губчатая паренхима находится в обратном соотношении со столбчатой паренхимой (рис. 1.1.2, а). То есть, если у вида достаточно хорошо развита губчатая паренхима, то относительное количество столбчатой паренхимы уменьшается. Точно так же, парциальные объемы губчатой паренхимы взаимосвязаны с парциальными объемами межклетников столбчатой паренхимы (рис. 1.1.2, б) и общими объемами меж-
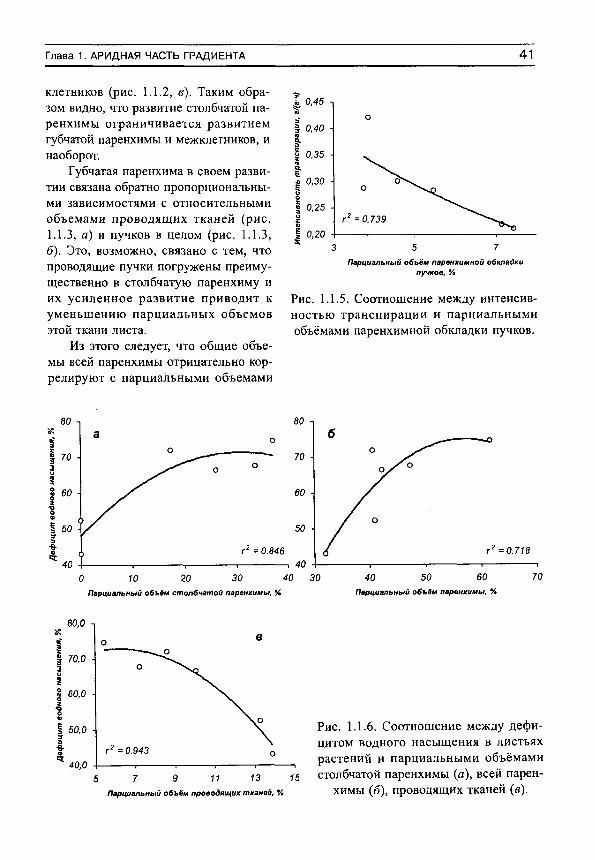
Глава 1. АРИДНАЯ ЧАСТЬ ГРАДИЕНТА 41
клетников (рис. 1.1.2, в). Таким образом видно, что развитие столбчатой паренхимы ограничивается развитием губчатой паренхимы и межклетников, и наоборот.
Губчатая паренхима в своем развитии связана обратно пропорциональными зависимостями с относительными объемами проводящих тканей (рис. 1.1.3, а) и пучков в целом (рис. 1.1.3, б). Это, возможно, связано с тем, что проводящие пучки погружены преимущественно в столбчатую паренхиму и их усиленное развитие приводит к уменьщению парциальных объемов этой ткани листа.
Из этого следует, что общие объемы всей паренхимы отрицательно коррелируют с парциальными объемами
I 0,45 ^
I 0,40
I 5 0,35 а I 0,30 и 2 I 0,25
0,20 =0.739
Парциальный объём паренхимной обкладки пучков, %
Рис. 1.1.5. Соотношение между интенсивностью транспирации и парциальными объёмами паренхимной обкладки пучков.
80 п 80 л
о 10 20 30 40 30 Парциальный объём столбчатой паренхимы, %
=0.718
40 50 60 70 Парциальный объём паренхимы, *А
= 0.943
5 7 9 11 13 15 Парциальный объём проводящих тканей, %
Рис. 1.1.6. Соотношение между дефицитом водного насыщения в листьях растений и парциальными объёмами столбчатой паренхимы (а), всей парен
химы (б), проводящих тканей (в).
42 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ...
1.0 1,0
0,6 0,6 5 10 15 20 25 30 2 4 6 8 10 12
Парциальный объём межклетников губчатой паренхимы, % Парциальный объем эпидермиса, %
Рис. 1.1.7. Соотношение между содержанием воды в листьях растений и парциальными объёмами эпидермиса (а) и межклетников губчатой паренхимы (б).
проводящих тканей (рис. 1.1.4, а). Но обкладки пучков развиваются тем заметнее, чем больше парциальный объем всей паренхимы в листьях растений (рис. 1.1.4, б).
Других корреляций между тканями (и свободными пространствами) в листьях растений не обнаружено.
Можно отметить некоторые корреляции между показателями водного режима растений и анатомической структурой листьев.
Интенсивность транспирации в достаточно заметной степени зависит от парциальных объемов паренхимных обкладок пучков (рис. 1.1.5). Исходя из этой зависимости, можно предположить, что обкладка пучков ограничивает скорость испарения воды. Возможно, это связано с тем, что проникновение воды из ксилемы в мезофилл и межклетники затрудняется лучше развитыми обкладками пучков. Так или иначе, но можно констатировать факт, что в данном местообитании растения с большим объемом обкладок имеют самую низкую интенсивность транспирации.
Дефицит водного насыщения ассоциируется с парциальными объемами как столбчатой паренхимы (рис. 1.1.6, а), так и всей паренхимы в целом (рис. 1.1.6, б). Достаточно уверенно можно предположить, что вода в процессе насыщения листьев попадает не в межклетные пространства, а, преимущественно, в столбчатую паренхиму и, отчасти, в губчатую. Поскольку объем паренхимы отрицательно коррелирует с объемом проводящих тканей, то возникает отрицательная зависимость водного дефицита от парциальных объемов проводящих тканей (рис. 1.1.6, в).
Содержание воды в листьях растений связано с парциальными объемами эпидермиса (рис. 1.1.7, а) и межклетников губчатой паренхимы (рис. 1.1.7, б). В первом случае зависимость кажется не очень тесной (г^ = 0.633). Однако ее в значительной степени «портит» Тп/оНит агуете, вид, у которого при самом большом парциальном объеме эпидермиса (табл. 1.1.3) содержание воды было не самым высоким (табл. 1.1.1), поскольку растения этого вида находились на грани полного увядания и не могли восстановить свои запасы воды. Без учета этого вида зависимость между содержанием воды и парциальным объемом эпидермиса значительно усиливается (г^ = 0.871) и неоднозначно свидетельствует, что у данных видов эпидермис является од-
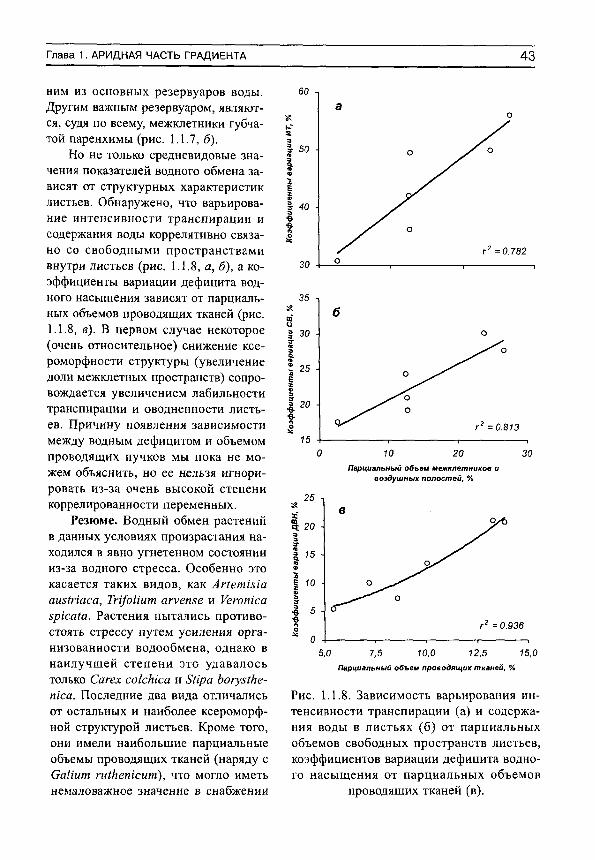
Глава 1. АРИДНАЯ ЧАСТЬ ГРАДИЕНТА 43
ним из основных резервуаров воды. Другим важным резервуаром, являются, судя по всему, межклетники губчатой паренхимы (рис. 1.1.7, б).
Но не только средневидовые значения показателей водного обмена зависят от структурных характеристик листьев. Обнаружено, что варьирование интенсивности транспирации и содержания воды коррелятивно связано со свободными пространствами внутри листьев (рис. 1.1.8, а, б), а коэффициенты вариации дефицита водного насыщения зависят от парциальных объемов проводящих тканей (рис. 1.1.8, в). В первом случае некоторое (очень относительное) снижение ксероморфности структуры (увеличение доли межклетных пространств) сопровождается увеличением лабильности транспирации и оводненности листьев. Причину появления зависимости между водным дефицитом и объемом проводящих пучков мы пока не можем объяснить, но ее нельзя игнорировать из-за очень высокой степени коррелированности переменных.
Резюме. Водный обмен растений в данных условиях произрастания находился в явно угнетенном состоянии из-за водного стресса. Особенно это касается таких видов, как Аг1ет1з1а аш1паса, Тп/оНит агуепзе и Уегопка зркаШ. Растения пытались противостоять стрессу путем усиления организованности водообмена, однако в наилучшей степени это удавалось только Сагех сокЫса и 5йра Ьогуз(ке-пка. Последние два вида отличались от остальных и наиболее ксероморф-ной структурой листьев. Кроме того, они имели наибольшие парциальные объемы проводящих тканей (наряду с СаНит гШНепкит), что могло иметь немаловажное значение в снабжении
60 п
50
4 40 •
30
35 п
8 I 30
I I | 2 0
I 15
25
I 20
= V 3 I 10 0) I 4 5
^^ =0.782
=0.813
10 20 30 Парциальный объем межклетников и
воздушных полостей, %
г' =0.936
5,0 7,5 10,0 12,5 15.0 Парциальный объем проводящих тканей, %
Рис. 1.1.8. Зависимость варьирования интенсивности транспирации (а) и содержания воды в листьях (б) от парциальных объемов свободных пространств листьев, коэффициентов вариации дефицита водного насыщения от парциальных объемов
проводящих тканей (в).

44 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ.
листьев водой. Выявлены взаимосвязи между различными тканями (и свободными пространствами листьев). Вместе с тем обнаружено, что развитие определенных тканей (или межклеточных пространств) влияет и на состояние водного режима растений. Интенсивность транспирации зависит от относительного объема обкладки пучков, дефицит водного насыщения связан с объемом столбчатой паренхимы, а оводненность листьев коррелирует с относительным количеством эпидермиса и межклетников губчатой паренхимы. Степень варьирование показателей водного обмена также зависит от ряда структурных характеристик листьев.
1.2. Ковыльково-луково-баглуровое сообщество
Наблюдения за водным режимом растений в данном сообществе были проведены в 1983 г на участке остепненной пустыни Заалтайской Гоби Монголии. Остепнен-ные пустыни (пустыни со злаками и луками) распространены в амплитуде высот от 1600 до 1850 м и занимают южный бэль гор Шинэ-Джинст (система гор Гобийского Алтая) (Рачковская, Гунин, 1980). Сообщества формации баглура {АпаЬаш Ьгеуг/оИа) являются наиболее характерными зональными типами пустынь (Рачковская, Гунин, 1980; Федорова, 1980). Ковыльково-луково-баглуровые сообщества распространены преимущественно на межсайровых пространствах. Доминируют полукустарничек АпаЬазгз Ьгеу1/оИа, плотнодерновинные злаки 8Нра §1агеоза и 5. §оЫса КозЬеу., луки АШит ро1угМ2ит и А. топ§оНсит Ке§е1. Растительные сообщества разрежены — проективное покрытие 3-5%. Почва на участке проведения наблюдений палево-бурая, супесчаная, подстилаемая песчано-щебнистыми пролювиальными отложениями (Федорова, 1980).
Объектами наблюдений были выбраны доминанты сообщества: АШит ро1угЫ2ит, АпаЬазк Ьгеу1/оИа и 5Ира §1агеоза. Исследования проводили в двух вариантах — в естественном местообитании (контроль) и на участке с поливом. Опыт с поливом начался в 1980 на трансекте размером 1.5x20 м (Слемнев и др., 1983). В 1983 г. полив проводился 8 (50 мм), 14 (40 мм), 22 (30 мм) и 29 (30 мм) июня, 5 (10 мм) и 16 (30 мм) июля (Слемнев, Цоож, 1988). Запасы влаги на этих вариантах различались достаточно заметно. В первом случае они составляли в среднем за период наблюдений 13 мм (в слое почвы 0-30 см), во втором — 21 мм (табл. 1). Кроме того, после первого дня работы выпало очень большое количество осадков (38 мм в течение двух дней), что вызвало заметную интенсификацию жизнедеятельности растений, особенно АШит ро1угЫ7ит (началось бурное отрастание новых побегов) и, в меньшей степени, ЗНра §1агеоза в естественном местообитании.
Исходные данные
Интенсивность транспирации растений АШит ро1угШгит в естественном местообитании в начале наблюдений составляла в среднем за день 0.44 г^д^^ • г^^^^^д^сы " ч ' • После выпадения осадков она резко возросла до 2, а в последующие дни до 2-6 г,„<)ы •^с^.массы - ч ' ' , а затем постепенно снижалась до 0.75 г,„аы • г сух.массы • ^^'^
(рис. 1.2.1, а). Средняя за период наблюдений величина этого показателя была равна 1-44 г , „й„ •^сух.массы "ч"' (табл. 1.2.1). Типичные значения находились в пределах 0.4-

Глава 1. АРИДНАЯ ЧАСТЬ ГРАДИЕНТА 45
2-8 г,„аы - ^сух.массы ч а коэффициент вариации был равен 75%. Сезонная динамика скорости испарения воды растениями этого вида находилась в тесной связи с изменениями влажности почвы и оводненности листьев {К^ = 0.98). Судя по частным Р-критериям, наибольшее влияние на транспирацию АШит ро1угк1гит в данных условиях произрастания оказывала влажность почвы (Р = 154) и значительно меньшее (Р = 36) — оводненность листьев. Зависимость такова, что с увеличением влажности почвы и листьев происходит увеличение интенсивности транспирации.
На участке с поливом сезонный ход транспирации АШит ро1угЫ2ит был несколько иным (рис. 1.2.1, а). Максимальная среднедневная величина скорости испарения воды (3.54 Гвойь, • ^'сух.массы ' ч " ' ) была зафиксиро-вана в самом начале проведения измерений. Выпадение осадков не оказало заметного влияния на этот процесс, и транспирация осталась примерно на том же уровне. Постепенно скорость транспирации выровнялась с таковой в естественном местообитании, и дальнейшие ее изменения происходили одинаково в обоих экото-пах. Поэтому средняя величина интенсивности транспирации на участке с поливом была ненамного выше, чем в естественном местообитании (табл. 1.2.1). Коэффициент вариации показателя бьш так же высок (68%), а типичные значения находились в пределах 0.57-4.0 Г,„йыТ;;!хл.ассь, - Ч ' ' -
Расчеты выявили зависимость сезонной динамики транспирации от влажности почвы (И = 0.785). При увеличении запасов воды в почве экспоненциально увеличивается скорость испарения воды.
0,5 -1 , , . , г 25.06.83 09.07.83 23.07.83 06.08.83 20.08.83 03.09.83
Дни сезона
Рис. 1.2.1. Сезонные ходы интенсивности транспирации АШит ро1угМ2ит {а), АпаЬазгз Ьгеу1/оИа (б) и 8Ира ^1агеоза (в) в естественном местообитании (1) и на поливном
участке (2).

46 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ.
Таблица 1.2.1
Средние значения интенсивности транспирации (ИТ, г̂ ^̂ ы '^'сух.массы "ч" ' ) , дефицита водного насыщения (ДВН, % ) , содержания воды (СВ, г^„з„ - "г'сух.массы )
осмотического давления клеточного сока (ОД, МПа) видов ковыльково-луково-баглурового сообщества
Вид ИТ ДВН СВ ОД Вид
п X 8х п X 8х п X 8х п X
Естественное местообитание АШит \11 1.338 1.000 177 19.5 4.6 \11 4.510 0.935 18 1.65 0.20 ро1угЫ2ит АпаЬаш 179 0.359 0.243 179 33.8 8.4 179 2.096 0.387 17 5.24 0.91 ЬгеУ1/оНа 8Ира §1агеош 176 1.249 0.316 174 26.8 8.4 176 1.043 0.185 17 3.46 1.11 В целом для 532 0.979 0.762 530 26.7 9.4 532 2.551 1.566 52 3.42 1.69 сообщества
Участок с поливом АШит 180 1.895 1.439 180 16.9 3.4 180 4.505 0.463 ро1угк1гит АпаЬаш 180 0.642 0.423 180 25.0 8.5 180 2.795 0.536 ЬгеуфИа 8Нра §1агеоза 172 1.510 0.680 167 24.7 7.3 172 1.155 0.213 — — — В целом для сообщества
532 1.347 1.089 527 22.1 7.7 532 2.843 1.430 — — —
Интенсивность транспирации растений АпаЬаш ЬгеуфИа (в естественном местообитании) в течение всего периода наблюдений бьша на очень низком уровне. Максимальная среднедневная величина этого показателя не превышала 0.71 г^^^^ -^сух..нассы • ч~ ' , а среднее за сезон значение было равно 0.36 г^^^^, сух.массы ' ч ' (табл. 1.2.1). После выпадения большого количества осадков скорость испарения воды увеличивалась достаточно медленно, пока не достигла максимума через две недели (рис. 1.2.1, б). Типичные значения этого признака находились в пределах 0.12-0.70 г^д^^, • т~1,х.массы ' ч~ ' , коэффициент вариации составил 66%. Сезонная динамика транспирации была связана с осмотическим давлением клеточного сока таким образом, что по мере увеличения среднедневных значений осмотического давления скорость испарения воды снижалась (Я = 0.887). Совершенно не проявилась статистическая зависимость транспирации от влажности почвы. Это бьшо, судя по всему, результатом замедленной реакции растения на изменения условий почвенного водоснабжения. Лаг-период между максимумами транспирации и влажности почвы составлял 12-14 дней.
На поливном участке интенсивность транспирации АпаЬази Ъгеуг/оЫа бьша в среднем почти вдвое выше, чем на контроле (табл. 1.2.1). Несколько увеличился и размах типичных колебаний, который находился в диапазоне 0.22-1.23 т^^^^, '^сух.массы 'Ч~' • Однако коэффициент вариации остался на том же уровне (66%). После выпадения осадков скорость испарения воды АпаЬазги Ьгеу1/оНа на поливном участке резко сни-

Глава 1. АРИДНАЯ ЧАСТЬ ГРАДИЕНТА 47
зилась, продемонстрировав реакцию сходную с реакцией на стрессовое воздействие (рис. 1.2.1, б). В дальнейшем изменения этого показателя были такими же, как и на контроле, с небольшой разницей в абсолютных значениях. Множественный регрессионный анализ выявил зависимость сезонной динамики транспирации от содержания воды в побегах растений и дефицита влажности воздуха {К^ = 0.943). Преобладающее влияние на скорость транспирации оказывала оводненность побегов (частный Р-критерий равен 75) и значительно меньшее — влажность воздуха (Р = 10.6). Эту зависимость можно описать следующим образом: увеличение влажности побегов и дефицита влажности воздуха вызывают увеличение среднедневных величин интенсивности транспирации.
Транспирация растений 8Ира ^(агеоза в естественном местообитании была на сравнительно высоком уровне — 1.25 т^^^^, - ^^ух.массы ' ч ' ' • Амплитуда колебаний признака находилась в довольно узком диапазоне между типичными минимумом (0-52 г,„й„ • г ; ^ ! ^ - ч " ' ) и максимумом (1.98 г.^^ь, тёухл.ассы ' ч" ' ) • Это подтверждается небольшим коэффициентом вариации, который был равен 25%. Сезонный ход транспирации (рис. 1.2.1, в) характеризуется ее довольно монотонным увеличением в течение 12-14 дней после выпадения осадков, и последующим достаточно резким снижением среднедневных величин. Множественный регрессионный анализ показал, что только дефицит водного насыщения листьев оказывает не очень сильное, но заметное влияние на этот процесс (И = 0.671, Р = 12.2): увеличение дефицита воды в листьях приводит к сокращению ее испарения растениями в сезонной динамике.
На участке с поливом транспирация 8Нра §1агео$а была незначительно выше, чем в естественном местообитании и составляла в среднем 1.51 г̂ ^^ ,̂ - т'^хмассы ч ' (табл. 1.2.1). Однако варьирование признака было более выраженным. Типичные минимум и максимум были равны 0.77 и 2.41 г^о^^ '^1:1хмассы " ч " ' , а коэффициент вариации находился на уровне 45%. Выпавшие в начале периода наблюдений осадки не вызвали заметного увеличения интенсивности транспирации (рис. 1.2.1, в ) . Она поддерживалась на одном уровне в течение двух недель, а затем стала изменяться так же, как и у растений в естественном местообитании. Анализ показал, что сезонный ход транспирации растений 8Нра §1агео5а на поливном участке изменялся в соответствии с изменениями влажности почвы {г^ = 0.723, Р = 15.7).
Дефицит водного насыщения в листьях растений АШит роЬгМгит в среднем за период наблюдений был на сравнительно невысоком уровне 19.5% (табл. 1.2.1). Типичные величины водного дефицита изменялись от 13.9 до 25.5%, а коэффициент вариации был равен 24%. После выпадения осадков дефицит изменялся незначительно, и лишь по прошествии 4-5 дней началось его заметное снижение (рис. 1.2.2, а). Не выявлены зависимости сезонных изменений водного дефицита от внешних или внутренних (других элементов водного режима) факторов.
На участке с поливом дефицит водного насыщения в листьях АШит ро1угЫ2ит был несколько ниже, чем на контроле, что было особенно заметно в первой половине периода наблюдений (рис. 1.2.2, а). В среднем он был равен 16.9% (табл. 1.2.1), а разность между типичными максимумом (21%) и минимумом (13.4%) составляла всего 7.6%. Коэффициент вариации был также низким (20%). Сезонные изменения водного дефицита были достаточно хорошо согласованы с изменениями влажности

48 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ.
24 п
15 -I , , , , г 25.06.83 09.07.83 23.07.83 06.08.83 20.08.83 03.09.83
Дни сезона
Рис. 1.2.2. Сезонные ходы дефицита водного насыщения АШит ро1угЫгит (а), АпаЬа-313 ЬгеУ1/оИа (б) и ЗИра §1агеоза (в) в естественном местообитании (1) и на поливном
участке (2).
почвы = 0.635, Р = 10.4): дефицит увеличивался по мере уменьщения запасов воды в почве.
У растений АпаЬазгз Ьгеуг/оИа (в естественном местообитании) дефицит водного насыщения в афилльных побегах достигал в среднем 34% (табл. 1.2.1). Изменения наблюдались внутри диапазона типичных значений 22.4-43.7%. Подвижность признака была незначительной ( К = 25%). Его изменения в сезоне (рис. 1.2.2, б) определялись изменениями осмотического давления клеточного сока и оводненности побегов {К' = 0.963, Р = 64): увеличение осмотического давления и уменьшение содержания воды в них сопровождались ростом дефицита водного насыщения.
На поливном участке водный дефицит в побегах АпаЬазгз Ьгеу1/оНа был значительно ниже, чем на контроле и составлял в среднем 25% (табл. 1.2.1, рис. 1.2.2, б). Типичные минимум и максимум признака были равны соответственно 15.2 и 36.8%, а коэффициент вариации (34%.) показал, что подвижность признака была несколько выше, чем на контроле. Множественный регрессионный анализ выявил зависимость водного дефицита только от оводненности побегов (Н = 0.817, Р = 27), которая засвидетельствовала увеличение дефицита при уменьшении содержания воды в побегах.
Листья 5Ира %1агеоза в начале проведения наблюдений на контроле испытывали значительный недостаток воды в листьях (39%). Однако после выпадения осадков водный дефицит резко снизился (рис. 1.2.2, в), а в среднем составил 26.8% (табл. 1.2.1). Амплитуда типичных колебаний находилась в диапазоне 18-38.2%, а подвижность признака (которую мы оценива-

Глава 1. АРИДНАЯ ЧАСТЬ ГРАДИЕНТА 49
ем с помощью коэффициентов вариации) была на уровне 31%. Сезонная динамика недонасыщенности листьев водой определялась изменениями их оводненности (И = 0.784, Р = 26). Также как и во всех остальных случаях, увеличение содержания воды сопровождалось снижением дефицита насыщения, и наоборот.
Водный дефицит в листьях 8Ира ^1агео5а на поливном участке был в среднем (24.7%) незначительно ниже, чем на контроле. Существенные различия наблюдались лишь в день наблюдений до выпадения осадков (рис. 1.2.2, в). Типичные значения находились в диапазоне 16.7-33.9%, а коэффициент вариации (29.6%) был несколько ниже, чем на контроле. Была выявлена взаимосвязь этого признака только с оводненностью листьев (г^ = § 0.701, Р = 14).
Содержание воды в листьях АШит ро1угН12ит в естественном местообитании в начале наблюдений было очень высоким (3.52 г^^^^ •'^'с1х.массы ) ,
а после выпадения осадков в течение двух недель увеличилось почти вдвое (до 5.84
^воды '^сух.массы
) (рис. 1.2,3, а). Такие существенные изменения оводненности листьев связаны, судя по всему, с наличием подземных многолетних запасающих органов, большим количеством водоносной паренхимы в листьях, но главным образом с резкой интенсификацией процессов жизнедеятельности, началом бурного отрастания новых побегов после быстрого и значительного увеличения запасов воды в почве. Средняя величина оводненности листьев была равна 4.51 ^еоды-'^',^.массы (тзбл. 1.2.1). Колебания признака наблюдались между 3.4 и 5.7 г^^^^ •г''с1х.массы , а его подвижность была на уровне 21%.
0,8 4- , , , , г 25.06.83 09.07.ВЗ 23.07.83 06.08.83 20.0883 03.09.83
Дни сезона
Рис. 1.2.3. Сезонные ходы содержания воды в листьях АШит ро1угЫ2ит (а) , АпаЬазк Ьгеуг/оИа (б) и 5Ира ^Ьгеоза (в) в естественном местообитании (1) и на поливном
участке (2).

50 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ.
В сезонном ходе оводненности листьев обнаружилась лишь очень слабая корреляция с влажностью почвы (И = 0.554), что было связано, возможно, с постоянным отрастанием новых побегов после выпадения осадков в начальный период наблюдений.
На поливном участке сезонные изменения содержания воды в листьях АШит ро1угЫгит не были такими резкими, как на контроле (рис. 1.2.3, а). Это могло быть обусловлено тем, что здесь растения были подготовлены к повышенной влажности почвы, и массового отрастания новых побегов после выпадения осадков не происходило. Средняя величина показателя была примерно такой же (4.505 Гдо<)„ г̂ ^̂ ), как и на контроле (табл. 1.2.1). Амплитуда колебаний типичных значений оводненности была заметно уже, чем на контроле и находилась между значениями 3.9 и 5-1 ^воды '^'сух.массы • Это опрсдблило И низкую ПОДВИЖНОСТЬ (10%) признзка на участке с поливом. Сезонная динамика содержания воды была обусловлена изменениями влажности почвы (Н = 0.661, Р = 11.7).
Оводненность побегов АпаЬазгз Ьгеу1/оИа в естественном местообитании была сравнительно высокой (в среднем 2.096 г^^^^ - ^сух.массы ) , но заметно ниже, чем листьев АШит ро1угЫ2ит (табл. 1.2.1). После выпадения осадков содержание воды увеличивалось медленно от 2.0 до 2.7 г̂ ^̂ ^̂ , -т'^^^^^^^^^ в течение двух недель (рис. 1.2.3, б). Типичные же значения признака лежали в границах 1.67-2.67 г^р^ ,̂ '^сух.массы > ^
коэффициент вариации составил всего 18.5%. В сезонной динамике оводненность побегов была взаимосвязана только с изменениями дефицита водного насыщения (Н = 0.898, Р = 53), но не с влажностью почвы, поскольку наблюдалось примерно двухнедельное отставание оводненности от изменений этого фактора среды.
На поливном участке содержание воды в побегах АпаЬаш Ьгеуг/оИа было равно в среднем 2.8 ^воды '^сух.массы (табл. 1.2.1). Осадки вызвали незначительное увеличение оводненности (рис. 1.2.3, б). Диапазон типичных значений показателя был несколько шире (2.1-3.5 г^д^^ 'Г у̂х.диссы )> чем на контроле, но подвижность находилась на том же уровне ( К = 19.2%). Изменения оводненности побегов в этом экотопе были тесно связаны с динамикой влажности почвы (г^ = 0.831, Р = 30).
Содержание воды в листьях 5Нра §1агео5а в естественном местообитании находилось на самом низком уровне — 1.0 г^^^,, •т~1х.массы (табл. 1.2.1), что объясняется ксе-роморфной структурой растений и низким запасом воды в почве. Осадки вызвали очень небольшое увеличение среднедневных значений оводненности листьев (рис. 1.2.3, в). Типичные значения находились в очень узком диапазоне — 0.81-1.29 ^тды '^сух.массы >
а подвижность признака характеризуется низким коэффициентом вариации (17.7%). Изменения содержания воды в листьях растений в сезоне находились в тесной связи с изменениями дефицита водного насыщения (И = 0,815, Р = 26).
На поливном участке оводненность листьев 8йра §1агеоза была лишь незначительно выше, чем на контроле и составила в среднем 1.16 г^^^у - ^сух.массы (табл. 1.2.1). Амплитуда типичных колебаний находилась в пределах 0.91-1.44 ^воды '^сух.массы , ^
подвижность на уровне 18.4%. Как и в естественном местообитании, оводненность листьев находилась в тесной корреляции с дефицитом водного насыщения (Н = 0.701).
Осмотическое давление клеточного сока растений определялось только у растений в естественном местообитании. В листьях АШит ро1угЫгит концентрация осмотически активных веществ была самой низкой. Средняя за период наблюдений вели-
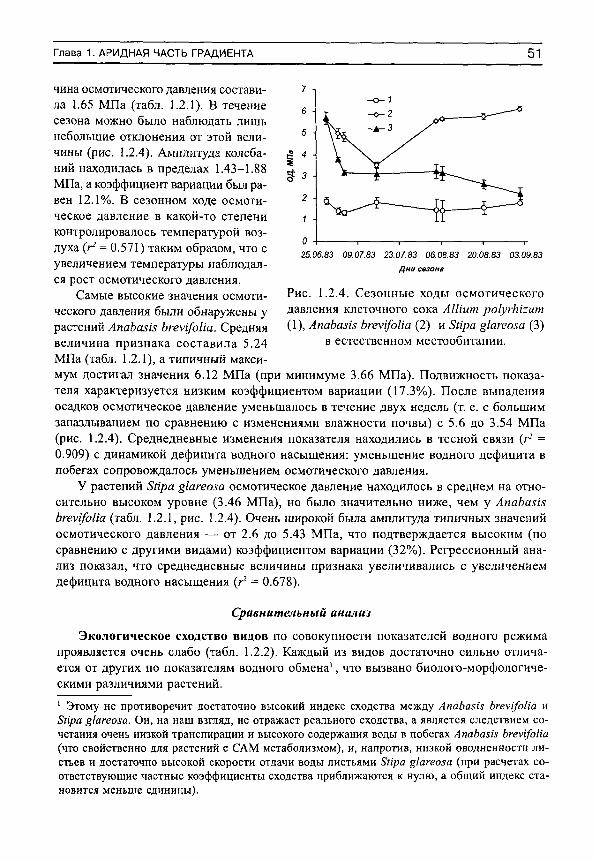
Глава 1. АРИДНАЯ ЧАСТЬ ГРАДИЕНТА 51
чина осмотического давления составила 1.65 МПа (табл. 1.2.1). В течение сезона можно было наблюдать лишь небольшие отклонения от этой величины (рис. 1.2.4). Амплитуда колебаний находилась в пределах 1.43-1.88 МПа, а коэффициент вариации был равен 12.1%. В сезонном ходе осмотическое давление в какой-то степени контролировалось температурой воздуха (г^ = 0.571) таким образом, что с увеличением температуры наблюдался рост осмотического давления.
Самые высокие значения осмотического давления были обнаружены у растений АпаЬаш Ьгегг/оИа. Средняя величина признака составила 5.24 МПа (табл. 1.2.1), а типичный максимум достигал значения 6.12 МПа (при минимуме 3.66 МПа). Подвижность показателя характеризуется низким коэффициентом вариации (17.3%). После выпадения осадков осмотическое давление уменьшалось в течение двух недель (т. е. с большим запаздыванием по сравнению с изменениями влажности почвы) с 5.6 до 3.54 МПа (рис. 1.2.4). Среднедневные изменения показателя находились в тесной связи (г^ = 0.909) с динамикой дефицита водного насыщения: уменьшение водного дефицита в побегах сопровождалось уменьшением осмотического давления.
У растений ЗНра §1агеоза осмотическое давление находилось в среднем на относительно высоком уровне (3.46 МПа), но было значительно ниже, чем у АпаЬазк Ьгеуг/оИа (табл. 1.2.1, рис. 1.2.4). Очень широкой была амплитуда типичных значений осмотического давления — от 2.6 до 5.43 МПа, что подтверждается высоким (по сравнению с другими видами) коэффициентом вариации (32%). Регрессионный анализ показал, что среднедневные величины признака увеличивались с увеличением дефицита водного насыщения (т^ = 0.678).
Сравнительный анализ
Экологическое сходство видов по совокупности показателей водного режима проявляется очень слабо (табл. 1.2.2). Каждый из видов достаточно сильно отличается от других по показателям водного обмена', что вызвано биолого-морфологическими различиями растений.
' Этому не противоречит достаточно высокий индекс сходства между АпаЬаз{.ч Ьгеу1/оИа и Зйра §1агео5а. Он, на наш взгляд, не отражает реального сходства, а является следствием сочетания очень низкой транспирации и высокого содержания воды в побегах АпаЬазьз Ьгеу1/оИа (что свойственно для растений с САМ метаболизмом), и, напротив, низкой оводненности листьев и достаточно высокой скорости отдачи воды листьями ЗИра §1агеоха (при расчетах соответствующие частные коэффициенты сходства приближаются к нулю, а общий индекс становится меньше единицы).
I 4 ^
2
1 -
25.06.83 09.07.83 23.07.83 06.08.83 20.08.83 03.0983 Дни сезона
Рис. 1.2.4. Сезонные ходы осмотического давления клеточного сока АШит ро1угЫгит (1), АпаЬази Ьгеу1/оНа (2) и 8Нра §1агеоза (3)
в естественном местообитании.
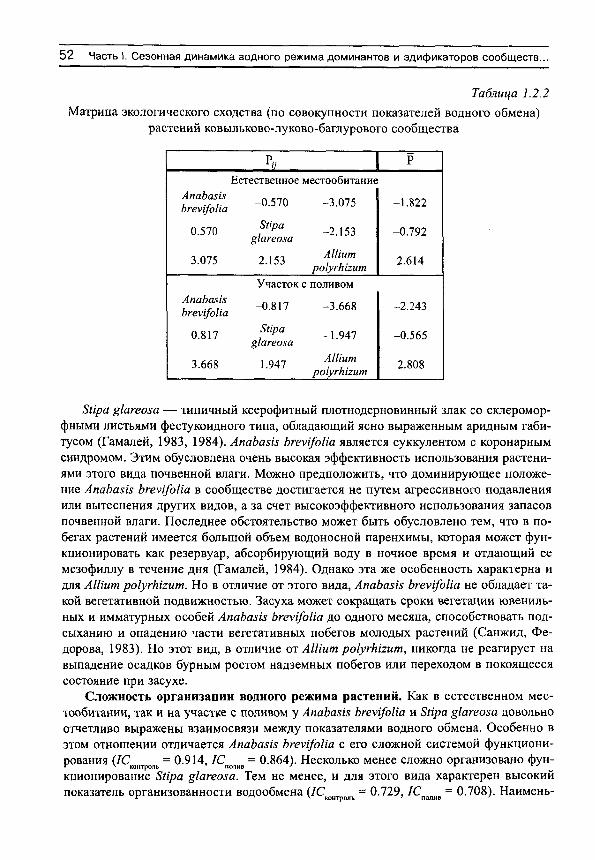
52 Часть I, Сезонная динамика водного режима доминантов и эдификаторов сообществ..
Таблица 1.2.2 Матрица экологического сходства (по совокупности показателей водного обмена)
растений ковыльково-луково-баглурового сообщества
Р. Естественное местообитание
АпаЬаш ЬгеуфНа -0.570 -3.075 -1.822
0.570 8Ира §1агео5а -2.153 -0.792
3.075 2.153 АШит ро1угк12ит 2.614
Участок с поливом АпаЪазЫ ЬгеУ1/оНа -0.817 -3.668 -2.243
0.817 8Ира §1агео5а -1.947 -0.565
3.668 1.947 АШит ро1угЫ2ит 2.808
8ира §1агеоха — типичный ксерофитный плотнодерновинный злак со склеромор-фными листьями фестукоидного типа, обладающий ясно выраженным аридным габитусом (Гамалей, 1983, 1984). АпаЬаз^з Ьгеуг/оНа является суккулентом с коронарным синдромом. Этим обусловлена очень высокая эффективность использования растениями этого вида почвенной влаги. Можно предположить, что доминирующее положение АпаЬаш Ьгеуг/оИа в сообществе достигается не путем агрессивного подавления или вытеснения других видов, а за счет высокоэффективного использования запасов почвенной влаги. Последнее обстоятельство может быть обусловлено тем, что в побегах растений имеется большой объем водоносной паренхимы, которая может функционировать как резервуар, абсорбирующий воду в ночное время и отдающий ее мезофиллу в течение дня (Гамалей, 1984). Однако эта же особенность характерна и для АШит ро1угМ2ит. Но в отличие от этого вида, АпаЬази Ьгеуг/оИа не обладает такой вегетативной подвижностью. Засуха может сокращать сроки вегетации ювениль-ных и имматурных особей АпаЬаш Ьгеу1/оНа до одного месяца, способствовать подсыханию и опадению части вегетативных побегов молодых растений (Санжид, Федорова, 1983). Но этот вид, в отличие от АШит ро1угЫ2ит, никогда не реагирует на выпадение осадков бурным ростом надземных побегов или переходом в покоящееся состояние при засухе.
Сложность организации водного режима растений. Как в естественном местообитании, так и на участке с поливом у АпаЬаш Ъгеух/оИа и ЗНра §1агеоза довольно отчетливо выражены взаимосвязи между показателями водного обмена. Особенно в этом отношении отличается АпаЬазгз Ьгеу1/оНа с его сложной системой функционирования (/С^„^^^ = 0.914, /С_,^„, = 0.864). Несколько менее сложно организовано функционирование 8Ира §1агеоза. Тем не менее, и для этого вида характерен высокий показатель организованности водообмена (/С^^ = 0.729,1С^^^^^ = 0.708). Наимень-

Глава 1. АРИДНАЯ ЧАСТЬ ГРАДИЕНТА 53
шая сложность организации наблюдается у АШит ро1угЫгит {1С^^^^ = 0.526, /С_^^„, = 0.694). Для этого вида характерно почти полное отсутствие связей между показателями водного режима, однако факторы среды (преимущественно влажность почвы и воздуха) вносят существенные ограничения в варьирование этих показателей
Низкий показатель организованности водообмена У 4 / / ш ш ро1угЫ2ит (особенно на контроле) связан с биологическими особенностями этого вида. Стратегия этого вида заключается, судя по всему, не в сохранении однолетних частей растений в экстремальных условиях произрастания путем выработки механизмов авторегуляции водного режима, а в том, чтобы сохранить жизнеспособными многолетние запасающие органы (к сходному выводу пришли ранее Н. И. Бобровская и Т. А. Попова, 1978, изучавшие эколого-биологические особенности АШит ро1угШгит в условиях Северной Гоби). Растения АШит ро1угЫ2ит, произрастающие, главным образом, в микропонижениях остепненной пустыни, имеют корневую систему, которая чрезвычайно чувствительна даже к небольшим осадкам (Попова, 1977). После выпадения осадков начинается бурное отрастание побегов, причем их оводненность поддерживается в этот период на очень высоком уровне, в чем существенную роль могут играть как большой объем водоносной паренхимы в листьях, так и многолетние запасающие органы растений. Для таких молодых, хорошо снабженных водой листьев, зависимости элементов водного режима от факторов среды (или взаимосвязи этих элементов) не имеют, возможно, существенного значения для развития особей.
У растений АпаЬазгз Ъгеут/оНа и Зйра §1агеоза, напротив, сильно выражены взаимосвязи между показателями водообмена (особенно на контроле), что отражает усиление функциональной регуляции в ответ на неблагоприятные факторы среды. При этом наблюдается 12-14-дневная задержка реакции ряда элементов на изменяющиеся условия почвенного водоснабжения. Измерения скорости фотосинтеза АпаЬаз1з Ьгеу1/оИа (Слемнев, Мокроносов, 1988) также показали 10-дневную задержку реакции этого процесса на изменения влажности почвы. Подобная замедленная реакция на изменения условий среды может быть обусловлена, в частности, тесными взаимосвязями элементов функционирования, что поддерживает их на некоторых, близких к оптимальным, уровнях.
Резюме. Показатели водного режима исследованных видов ковыльково-луково-баглурового сообщества в естественном местообитании и на поливном участке достаточно сильно различались в начальный период проведения наблюдений (рис. 1.2.1-1.2.3). Однако выпавшие осадки в большинстве случаев выровняли величины элементов водообмена в течение двух-трех недель (это было обусловлено также тем, что последний полив был проведен 16 июля). Реакция растений на осадки на контрольном и поливном участках была часто неодинакова. Если на контроле осадки вызвали увеличение скорости испарения воды растениями (рис. 1.2.1), то на «поливе» интенсивность транспирации либо снижалась, либо оставалась неизменной. На поливном участке после выпадения осадков очень мало изменялись дефицит водного насыщения и оводненность листьев (за исключением 8Ира §1агеоза) (рис. 1.2.2, 1.2.3). Заметные различия между растениями на «контроле» и «поливе» сохранились в течение всего периода наблюдений только у АпаЬазгз Ьгеуь/оИа по водному дефициту и содержанию воды в листьях (рис. 1.2.2, 1.2.3). Осмотическое давление клеточного сока АШит ро1угНггит оставалось примерно на одном уровне в течение всего периода наблюде-

54 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ...
НИИ (рис. 1.24). Выпавшие осадки снизили этот показатель почти вдвое у АпаЪазгз Ъгеуг/оЫа и 8Нра §1агеоза. У первого вида осмотическое давление к концу вегетации вернулось на первоначальный уровень, а у второго — постепенно снижалось. Экологическое сходство между видами (по совокупности показателей водообмена) выражено слабо, что обусловлено биолого-морфологическими свойствами растений. Организованность водообмена хорошо проявилась у АпаЬаш Ьгеу1/оНа и 8Ира §1агео5а, и, в меньшей степени, у АШит ро1угЫ2ит.
1.3. Твердоосочково-змеевковое сообщество
Наблюдения за водным режимом растений данного сообщества проводили в 1988 г на луговом стационаре Советско-Монгольской комплексной биологической экспедиции (Монголия, сомон Шаамар) в пойме р. Орхон. Участок располагался на хорошо дренируемом выровненном прирусловом валу протоки р. Орхон. Почва на этом участке аллювиальная дерновая остепняющаяся маломощная на грубом галечном аллювии. Объектами наблюдений были доминанты сообщества Сагех с1ипи5си1а, С1еШо§епе5 здиаггоза, Ьеутш сНтепш и Ро1епШ1а Ы/игса. Травостой редкий, проективное покрытие не более 40-50%. Дополнительные сведения по водному режиму этих видов можно найти в ряде работ, где приводятся среднесезонные величины показателей и/или амплитуды их варьирования (Горшкова, 1966; Копытова, Горшкова, 1977; Горшкова, Зверева, 1982; Горшкова и др., 1982; Измайлова, 1986; Якимова, Павлова, 1989).
Исходные данные
Интенсивность транспирации. Скорость испарения воды с поверхности листьев бьша в среднем достаточно близкой у всех видов (табл. 1.3.1). Только в отдельные дни можно было наблюдать относительно более высокую скорость транспирации у Ро1еп1И1а Ы/игса, и более низкую — у Ьеутиз сЫпепзгз (рис. 1.3.1). Варьирование признака было небольшим и достаточно близким у всех видов (33-39%). Типичные
Таблица ЬЗ.1
Средние значения интенсивности транспирации (ИТ, г^^^^, -т'у^^иассы ' ч ' ) , дефицита водного насыщения (ДВН, % ) , содержания воды (СВ, г„„а„ - ^сух.массы )
осмотического давления клеточного сока (ОД, МПа) видов твердоосочково-змеевкового сообщества
Вид ИТ ДВН СВ ОД
Вид Вид п X 8х п X 8х п X & п X &
Сагех (1игшзси1а 230 0.922 0.362 228 22.2 5.3 230 1.475 0.137 27 2.67 0.42 СкШо^епез 228 0.900 0.285 228 14.7 5.0 228 1.506 0.162 27 1.95 0.32 5^иа^^оза Ьеутиз сМпепз1з 230 0.742 0.259 228 10.2 4.6 230 1.673 0.106 27 2.14 0.32 РоШИИа Ы/игса 228 1.015 0.335 228 19.5 6.0 228 1.574 0.181 27 2.39 0.29 В целом для сообщества
916 0.894 0.327 912 16.6 6.9 916 1.557 0.167 108 2.29 0.43

Глава 1. АРИДНАЯ ЧАСТЬ ГРАДИЕНТА 55
1,5 п
1,3
1,1 -
^ 0,9
0,7
0,5 -
0,3 15.06.88 29.06.88 13.07.88
Дни сезона
27.07.88 10.08.88
Рис. 1.3.1. Сезонный ход интенсивности транспирации видов твердоосочково-змеевкового
сообщества. Здесь и на рис. 1.3.2-1.3.4: 1 — Сагех с1ипизси1а, 2 — С1е1з1о§епез здиаггоза, 3 — Ьеутиз сЫпепзгз,
4 — Ро1еп1И1а Ы/игса.
минимумы показателя находились на уровне 0.43 (Ьеутиз сктепзгз) — 0.63 (Ро1епШ1а Ы/игса) т,„^„ • г;̂ !̂ ч " ' , типичные максимумы — на уровне 1.11 (Ьеутиз сНтепзгз) — 1.47 (Сагех с1игшзси1а) г,,^„ • г;^!,.^„„^ • ч " ' .
Множественный регрессионный анализ показал, что сезонные изменения интенсивности транспирации определялись влажностью почвы у Сагех (1ипизси1а (И = 0.844), С1е1з1о^епез здиаггоза (г^ = 0.815) и Ьеутиз сЫпеп-313 (г^ = 0.810). В то же время скорость испарения воды с поверхности листьев Ро1епШ1а Ы/игса бьша связана только с дефицитом водного насыщения (г^ = 0.643). Можно показать зависимость транспирации этого вида также и от осмотического давления, но она проявляется лишь на более низком доверительном уровне (Р = 0.9).
Дефицит водного насыщения. Виды достаточно отчетливо разделились на три категории: с низким водным дефицитом (в среднем 10.2%, Ьеутиз сктепз1з), средним (14.7%, С1е1з1о§епез здиаггоза) и высоким (19.5-22.2%, Сагех с1ипизси1а, Ро1еп-ИИа Ы/игса) (табл. 1.3.1, рис. 1.3.2). Это может быть в целом обусловлено скоростью испарения воды растениями: для видов с большей интенсивностью транспирации свойственны и большие в среднем величины водного насыщения, хотя в сезонных ходах непосредственная взаимосвязь между этими признаками была обнаружена только для РошИПа Ы/игса. Варьирование дефицита насыщения было невысоким и составляло для всех видов 24-45%. Типичные минимумы были на уровне 5.7-16.3%, а типичные максимумы — на уровне 16.2-28.7%. Несмотря на такой разброс значений величины амплитуд водного дефицита у всех видов были близки и изменялись от 12.3% у С1е1з1о§епез здиаггоза до 16% у РогепйИа Ы/игса.
Фактором среды, определяющим сезонную динамику дефицита водно-
30 п
25
20
15
10
15.06.88 29.06.88 13.07.88 Дни сезона
27.07.88 10.08.88
Рис. 1.3.2. Сезонный ход дефицита водного насыщения видов твердоосочково-змеевкового
сообщества.

56 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ..
го насыщения Сагех <1ипи5си1а (г^ = 0.683) и Ьеутиз сШпепзгз (при совместном влиянии влажности воздуха и оводненности листьев = 0.872), была влажность воздуха. Сезонные изменения водного дефицита СШз1о^епез здиаггоза были подвержены также и влиянию влажности почвы (Я' = 0.873). Эти зависимости таковы, что при увеличении влажности воздуха и почвы (а в случае Ьеутиз сктепзгз, и содержания воды в листьях) дефицит водного насыщения снижался. Только у Ро1епШ1а Ы/игса недо-насыщенность листьев водой не зависела от факторов среды, но была взаимосвязана с интенсивностью транспирации (г^ = 0.643).
Содержание воды в листьях было достаточно высоким и сравнительно близким у всех видов (1.48-1.67 т^д^^, •I'сух.массы ^ табл. 1.3.1), хотя в отдельные дни можно было наблюдать статистически значимые различия (рис. 1.3.3). Варьирование признака было очень низким, составляя 6.3-11.5%, а амплитуды типичных колебаний не превышали ни у одного вида 0.4 г^^ы '^сух.массы • Типичные минимумы находились на уровне 1.33-1.54 г^д^^ •г'сух.массы > а типичные максимумы — на уровне 1.64-
1.80 ^еоды '^сух.массы •
Сезонная динамика оводненности листьев Сагех с1иг1изси1а была подвержена влиянию влажности почвы (Р,,,„ = 52.7) и воздуха (Р^^^^ = 6.1) {К^ = 0.913). Судя по частным Р-критериям, влажность почвы в наибольшей степени влияет на содержания воды в листьях Сагех с1игшзси1а. Оводненность листьев СЫз1о§епез здиаггоза изменялась в сезоне под влиянием влажности воздуха (И = 0.797) таким образом, что увеличение сухости воздуха сопровождалось снижением содержания воды в листьях. У растений Ьеутиз сЫпепз1з изменения влажности листьев были связаны только с изменениями дефицита водного насыщения (И = 0.645). Колебания содержания воды в листьях Ро1еп1И1а Ы/игса за
висели, с одной стороны, от температуры воздуха, с другой — от осмотического давления клеточного сока [К^ = 0.684).
Осмотическое давление клеточного сока. Наибольшее осмотическое давление было обнаружено у Сагех с1ипизси1а (в среднем за период наблюдений 2.67 МПа) и РогепйИа Ы/игса (2.39 МПа). Несколько меньшее — у СШз1о§епез здиаггоза (1.96 МПа) и Ьеутиз сЫпепз1з (2.14 МПа) (табл. 1.3.1). Однако надо заметить, что эти различия в достаточной степени проявлялись лишь в отдельные дни наблюдений и в наилучшей степени были выражены между Сагех с1ипизси1а и СШз1о§епез здиаггоза (рис. 1.3.4). Подвижность осмотического давления была невысокой у всех видов и состав-
15.06.88 29.06.88 13.07.88 27.07.88 10.08.88 Дни сезона
Рис. 1.3.3. Сезонный ход содержания воды в листьях видов твердоосочково-змеевкового
сообщества.
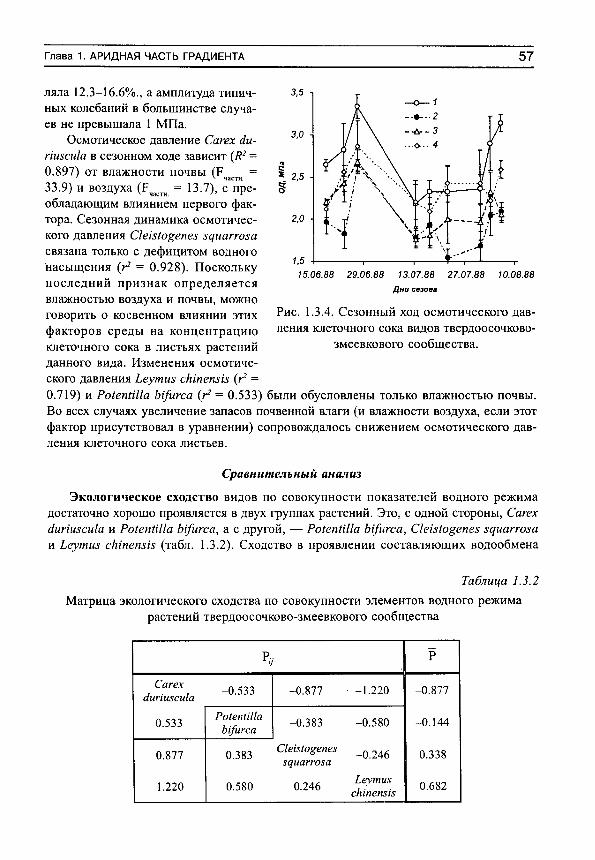
Глава 1. АРИДНАЯ ЧАСТЬ ГРАДИЕНТА 57
ляла 12.3-16.6%., а амплитуда типичных колебаний в большинстве случаев не превышала 1 МПа.
Осмотическое давление Сагех Ли-пшси1а в сезонном ходе зависит {К' = 0.897) от влажности почвы (Р =
^ ^ част 33.9) и воздуха (Р̂ ^̂ _̂̂ = 13.7), с преобладающим влиянием первого фактора. Сезонная динамика осмотического давления С1еШо§епез здиаггоза связана только с дефицитом водного насыщения (г^ = 0.928). Поскольку последний признак определяется влажностью воздуха и почвы, можно говорить о косвенном влиянии этих факторов среды на концентрацию клеточного сока в листьях растений данного вида. Изменения осмотического давления Ьеутиз сЫпепзгз (г^ = 0.719) и Ро1еп1И1а Ы/игса (И = 0.533) были обусловлены только влажностью почвы. Во всех случаях увеличение запасов почвенной влаги (и влажности воздуха, если этот фактор присутствовал в уравнении) сопровождалось снижением осмотического давления клеточного сока листьев.
Сравнительный анализ
Экологическое сходство видов по совокупности показателей водного режима достаточно хорошо проявляется в двух группах растений. Это, с одной стороны, Сагех с1игшзси1а и Ро1епИ11а Ы/игса, а с другой, — Ро1еп1И1а Ы/игса, С1е(з{о§епез здиаггоза и Ьеутиз сЫпепзгз (табл. 1.3.2). Сходство в проявлении составляющих водообмена
Таблица ЬЗ.2 Матрица экологического сходства по совокупности элементов водного режима
растений твердоосочково-змеевкового сообщества
Р
Сагех <1ипизси1а -0.533 -0.877 -1.220 -0.877
0.533 РоШНИа Ы/игса -0.383 -0.580 -0.144
0.877 0.383 С1еШо^епез здиаггоза -0.246 0.338
1.220 0.580 0.246 Ьеутиз сЫпепз1з 0.682
15.06.88 29.06.88 13.07.88 27.07.88 10.08.88 Дни сезоеа
Рис. 1.3.4. Сезонный ход осмотического давления клеточного сока видов твердоосочково-
змеевкового сообщества.

58 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ.
между такими разными во всех отношениях (габитуальном, биолого-морфологическом, систематическом) видами обусловлено, судя по всему, давлением среды.
Запас воды в слое почвы 0-30 см составлял в среднем за период наблюдений 37 мм (табл. 1). Однако особенность почвы, на которой произрастали эти виды, состоит в том, что горизонты, расположенные ниже 20 см и сложенные из песка (20-115 см) и гальки (115 см и ниже) прерывают связь верхних горизонтов с грунтовыми водами (находившимися в данный сезон вегетации на глубине 2.2-2.5 м). Выпадающие осадки не изменяли значительно и на долгое время запасы влаги в корнео-битаемом слое, поскольку из верхнего 5-ти сантиметрового горизонта вода быстро испарялась, а влага нижележащих слоев просачивалась до грунтовых вод. С другой стороны, капиллярная кайма грунтовых вод не доходила до корнеобитаемого слоя. Поэтому, например, запас воды в слое почвы 20-30 см был практически постоянен в течение большей части периода вегетации и составлял всего 2—4 мм, а растения использовали влагу, как можно предположить, преимущественно из самого верхнего слоя почвы.
Кроме того, для данного местообитания характерно обильное выпадение росы в течение всего периода вегетации, что в значительной степени определяет влажность приземного слоя воздуха в предрассветные и ранние утренние часы. Колебания влажности этого слоя, как показывают данные, во многих случаях определяют динамику элементов водного режима. Это может свидетельствовать о поглощении (в предрассветные часы) влаги, содержащейся в воздухе и использовании ее растениями, наряду с почвенной, в течение дня.
Из этого следует, что для всех видов ресурсы среды были примерно одинаковыми, а их недостаток не мог компенсироваться различиями в строении корневой системы. Поэтому отмеченные выше сходства могли быть обусловлены подавляющим воздействием недостатка этих ресурсов, а отличия — разной способностью растений в их использовании. Проявляющие относительно менее ксерофитные свойства СШзЮ^епез здиаггоза и Ьеутиз сктепз1з, судя по всему, более эффективно использовали доступную им влагу. Относительно менее успешными в этом отношении показали себя Сагех с1ипизси1а и РоШНИа Ы/игса, для которых характерен несколько более ксерофитный характер показателей водообмена.
Сложность организации водного режима растений. У всех видов индексы этого показателя были высокими. Минимальный индекс (0.626) был обнаружен для РоШИПа Ы/игса, максимальный (0.853) — для С1е1з1о§епез здиаггоза. У каждого вида обнаруживаются свои особенности в характере взаимосвязей между элементами водного режима и их зависимостей от факторов среды. У Сагех Лигшзси1а (индекс 0.834) взаимосвязи между элементами не найдены совсем, но выявлены очень тесные зависимости показателей водообмена от влажности почвы и воздуха. Функциональная регуляция водного режима не играет заметной роли в реализации его сезонной динамики, которая почти полностью ограничивается факторами среды. Почти то же самое характерно и для Ьеутиз сЫпепз1з (0.762), за тем исключением, что оводненность листьев растений взаимосвязана с дефицитом водного насыщения и не зависит от факторов среды. Достаточно тесные взаимосвязи между показателями водообмена вьивлены у С1е1з1о§епез здиаггоза и Ро1епШ1а Ы/игса, но эти показатели, вместе с тем, в своей сезонной динамике, контролируются влажностью воздуха и почвы или тем-

Глава 1. АРИДНАЯ ЧАСТЬ ГРАДИЕНТА 59
пературой воздуха. На примере этих видов прослеживается плавный переход от полного отсутствия функциональной регуляции водообмена у Сагех с1ипшси1а до заметного его развития у РоШННа Ы/игса. Экологическое сходство, отмеченное между этими двумя видами, возникает, очевидно, благодаря разным средствам. С одной стороны, это исключительно ограничивающее потенции растений воздействие факторов среды, с другой — в какой-то степени и самоограничения, обусловленные отрицательными обратными связями между элементами функционирования. Все это, возможно, наложило отпечаток и на подвижность водного режима растений. В среднем для каждого вида она небольшая и оценивается коэффициентами вариации 22-25%.
Резюме. Величины интенсивности транспирации и содержания воды в листьях всех видов были достаточно близкими и находились на уровнях, характерных для аридной части градиента. Сагех (1ипи$си1а и Ро1епИПа Ы/игса отличались более высокими значениями дефицита водного насыщения и осмотического давления клеточного сока. Это определило их относительно более ксерофитную организацию, по сравнению с С1еШо§епе5! здиаггоза и Ьеутиз сЫпепзгз. На примере видов твердоосочково-змеевкового сообщества прослеживается плавный переход от полного отсутствия функциональной регуляции водообмена у Сагех йипизси1а до заметного его развития у Ро{еп1И1а Ы/игса. Выявленное экологическое сходство между видами достигается разными средствами: в одних случаях — ограничивающим потенциальные возможности растений влиянием факторов среды, в других — развитием отрицательных обратных связей между показателями функционирования.

60 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ..
ГЛАВА 2. СРЕДНЯЯ ЧАСТЬ ГРАДИЕНТА
2.1. Ковыльно-разнотравно-вострецовое сообщество
Наблюдения в ковыльно-разнотравно-вострецовом сообществе проводили в 1984-1985 гг. на восточно-степном стационаре Советско-Монгольской комплексной биологической экспедиции (сомон Туменцогт). Участок, на котором произрастало сообщество, располагался в верхней части плоской аккумулятивной равнины на лугово-каш-тановых остепненных почвах. Район стационара, по ботанико-географическому районированию А. А. Юнатова (1950), относится к Даурско-Монгольской степной провинции Центральноазиатской степной области. Объектами наблюдений были доминанты и эдификаторы сообщества — Апетша /п§Ша, Мз (ИсНо1ота, Ьеутш сНтет1з, Ро1еп(И1а 1апасеН/оНа, РиЬаИИа (игсгаптоуН, ЗИра кгу1оуИ.
Дополнительные сведения по водному обмену указанных видов, произрастающих в разных местообитаниях, можно найти в ряде работ (Горшкова, Буркова, 1955, 1962; Свешникова, 1963, 1988; Горшкова, 1966, 1970, 1971; Бобровская, 1968, 1988; Бей-деман, Паутова, 1969; Паутова, 1969; Свешникова, Бобровская, 1969; Паутова, Копытова, 1970; Свешникова и др., 1971; Чойжамц, 1974, 1978; Измайлова, 1977, 1978, 1986; Копытова, Горшкова, 1977; Касьянова, Погодаева, 1979; Банникова и др., 1982; Горшкова, Зверева, 1982; Горшкова и др., 1982; 12та)1оуа, 1985).
8Ира кгу1оуИ является наиболее конкурентоспособным видом (как будет показано ниже) ковыльно-разнотравно-вострецового сообщества и принимает заметное участие в его сложении. В наиболее благоприятные годы этот вид составляет до 30% от общей массы травостоя, но наиболее часто от 5 до 10%. Вторым по относительной конкурентоспособности видом этого сообщества является Ьеутиз сЫпепзгз, несмотря на то, что его доля участия может достигать 25-^0%. В формировании надземной фитомассы сообщества принимает заметное участие Аг(ет1з1а/п^Ша (до 10%), а также / г а сИскоЮта, Ро1епШ1а 1апасеИ/оНа и РиЬаИИа (игсгаптоуИ (в среднем по 2-4%).
Условия произрастания в 1984-1985 гг были благоприятными для развития растений. В 1984 г. выпало 357 мм осадков, в 1985 — 292 мм, что превышает среднюю многолетнюю норму (по данным метеостанции Туменцогт). Вместе с тем условия произрастания в 1984 г. были несколько более напряженными, чем в 1985 г. Это определялось тем, что за июнь - июль 1984 г. выпало 149 мм осадков против 203 мм за этот же период в 1985 г В первый год наблюдений влажность почвы (в слое 0-30 см) была несколько ниже, чем во второй, а дефицит влажности воздуха достигал 41 мб. В 1985 г этот показатель не превышал 21 мб.
Исходные данные
Интенсивность транспирации. По скорости испарения воды растения данного сообщества можно условно разделить на две группы — с высокой и низкой скоростью транспирации. Наиболее интенсивно испаряли воду АПетгзга /п§1^а (1.527 ^еоды -^'с^х-массы-ч'^)' Ро1еп(Ша ШпасеИ/оНа (1.862 т,„^„ •г^з!,.^,,,^ • ч " ' ) и РиЬаИПа Шгсгаптоуп (1.510 Г ^ ^ Й Ы '^'^ух.массы 'Ч ' ) (табл. 2.1.1). Значительно меньшая интенсивность транспирации зафиксирована у 8Ира кгу1оУп (0.708 г̂ оаы '''^'с1х.массы ч " ' ) и

Глава 2. СРЕДНЯЯ ЧАСТЬ ГРАДИЕНТА 61
Таблица 2.1.1
Средние значения интенсивности транспирации (ИТ, г̂ ^^ы •^'сух.массы ч ' ' ) , дефицита водного насыщения (ДВН, % ) , содержания воды (СВ, г^^^^ • г^Д.^^^^^ )
осмотического давления клеточного сока (ОД, МПа) видов ковьшьно-разнотравно-вострецового сообщества
Вид ИТ ДВН СВ ОД Вид
п X 8х п X 8х п X 8х п X 5х АНетша /п§1с1а 440 1.527 0.831 438 28.2 10.4 440 2.083 0.483 51 2.02 0.74 1пз сИскоЮта 437 1.307 0.629 437 11.4 6.3 437 4.724 0.645 51 1.54 0.20 Ьеутиз сктепз1з 442 1.078 0.468 441 12.8 6.3 442 1.458 0.180 51 2.48 0.63 РоСепИПа 416 1.862 0.869 416 23.4 6.5 416 1.793 0.237 48 2.257 0.40 ШпасеИ/оИа РиЬшШа 440 1.510 0.756 437 14.3 6.2 440 2.044 0.350 51 2.56 0.62 1игс2атпоуН ЗНра кгу1оуИ 442 0.708 0.328 442 23.4 6.8 442 0.949 0.108 51 2.63 0.62 В целом для сообщества
2617 1.326 0.765 2611 18.9 9.6 2617 2.174 1.265 303 2.25 0.68
Ьеутиз сктепз1з (1.078 г^о^ы '^сух.массы 'Ч ' ) • Растения Мз сИско(ота испаряли воду с достаточно умеренной скоростью (1.307 г^^^^ '^сух.массы "ч" ' ) - Типичные минимумы признака в обеих группах были достаточно близки. Для первой группы видов они составляли 0.456 (Аг1ет(з1а /гщШа) — 0.698 {Ро1епИ11а ШпасеИ/оИа), для второй — 0.387 {8Ира кгу1оуЦ) — 0.584 {Мз сИско1ота) г^д^^ сух массы ' ч" ' • Типичные максимумы различались сильнее. У более интенсивно испарявших воду видов они бьши на уровне 2.441 (РШзаИИа Шгсгатпоуп) — 2.968 (Ро(епШ1а (апасеИ/оИа) т^д^^, •г~^^з,д^^,„ - ч " ' . У растений с более низкой скоростью транспирации эти максимумы находились между 1.090 (8Ира кгу1оуН) — 2.136 {Мз (Иско1ота) г^о^ы •^~с1х.массы ' ч " ' • Подвижность интенсивности транспирации, тем не менее, была практически одинаковой у всех видов (оценки коэффициентов вариации равны 43.4-54.4%).
В группе видов с высокими значениями интенсивности транспирации {Аг1ет1з1а /гщ1йа, Ро1епШ1а (апасей/оНа, РиЬаИНа (игсгаптоуп) сезонные ходы процесса были очень близки, различаясь только в отдельные дни наблюдений (рис. 2.1.1, а). У видов с более низкой скоростью испарения воды {Мз сИсНо1ота, Ьеутиз сЫпепз1з, 8Ира кгуЬуп) различия в сезонной динамике проявлялись чаще (рис. 2.1.1, б). Рисунок сезонного хода был одинаков у всех видов, но колебания транспирации у Ьеутиз сМпепзгз и ЗНра кгу1оуЦ были более сглаженными, без резких колебаний (рис. 2.1.1).
Множественный регрессионный анализ показал, что сезонная динамика транспирации Мз сИско1ота определяется только влажностью почвы. Коэффициент детерминации не очень велик (0.592), но статистически достоверен. Для Аг1ет1з1а /гщШа и Ро1еп1И1а (апасеИ/оНа характерна зависимость транспирации от влажности почвы и оводненности листьев (соответствующие коэффициенты детерминации равны 0.839 и 0.732). Сезонная динамика процесса у Ьеутиз сктепзгз (К^ = 0.907) и ЗИра кгу1оуИ {К^ = 0.762) зависит не только от этих двух факторов, но также и от освещенности.
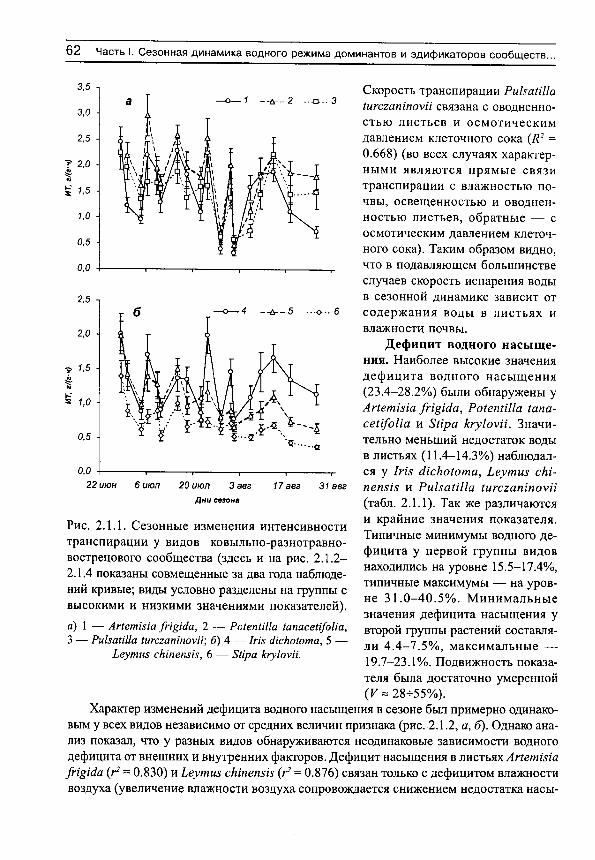
62 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ,
3,5
3,0
2,5
2,0
1.5
1,0
0,5
0,0
....... 3
..,о.. 6
Скорость транспирации РиЬаИИа (игсгатпоуп связана с оводненностью листьев и осмотическим давлением клеточного сока {К^ = 0.668) (во всех случаях характерными являются прямые связи транспирации с влажностью почвы, освещенностью и оводненностью листьев, обратные — с осмотическим давлением клеточного сока). Таким образом видно, что в подавляющем большинстве случаев скорость испарения воды в сезонной динамике зависит от содержания воды в листьях и влажности почвы.
Дефицит водного насыщения. Наиболее высокие значения дефицита водного насыщения (23.4-28.2%) были обнаружены у Аг1етша /гщШа, Ро1еп(И1а Шпа-сеН/оНа и ЗИра кгу1оуЦ. Значительно меньший недостаток воды в листьях (11.4-14.3%) наблюдался у Мз сИсИоЮта, Ьеутш сЫ-пеп$1з и РиЬаИИа (игсгаптоуИ (табл. 2.1.1). Так же различаются и крайние значения показателя. Типичные минимумы водного дефицита у первой группы видов находились на уровне 15.5-17.4%, типичные максимумы — на уровне 31.0-40.5%. Минимальные значения дефицита насыщения у второй группы растений составляли 4.4-7.5%, максимальные — 19.7-23.1%. Подвижность показателя была достаточно умеренной (К =28-55%).
Характер изменений дефицита водного насыщения в сезоне бьш примерно одинаковым у всех видов независимо от средних величин признака (рис. 2.1.2, а, б). Однако анализ показал, что у разных видов обнаруживаются неодинаковые зависимости водного дефицита от внешних и внутренних факторов. Дефицит насыщения в листьях АПетша /п^Ша (Н = 0.830) и Ьеутш сНтет1з (г^ = 0.876) связан только с дефицитом влажности воздуха (увеличение влажности воздуха сопровождается снижением недостатка насы-
2.5 1
2,0 -
•? 1,5
•а « 1,0 -
0,5
0,0 22июн биюл 20июл Заве 17 авг 31 авг
Дна сезона
Рис. 2.1.1. Сезонные изменения интенсивности транспирации у видов ковыльно-разнотравно-вострецового сообщества (здесь и на рис. 2.1.2-2.1.4 показаны совмещенные за два года наблюдений кривые; виды условно разделены на группы с высокими и низкими значениями показателей).
а) 1 — АПетша /п§Ша, 2 — РоШИИа 1апасеИ/оНа, 3 — РиЬаННа ШгсгатпоуИ; б) 4 — 1пз сИскоЮта, 5 —
Ьеутш сктепз13, 6 — Зйра кгу1оуН.

Глава 2. СРЕДНЯЯ ЧАСТЬ ГРАДИЕНТА 63
щения листьев водой) . С е з о н н а я динамика д е ф и ц и т а водного насыщения двух видов определяется не только в л а ж н о с т ь ю воздуха, но и другими элементами водного режима. Для Ро1епШ1а (апасеа/оНа [К^ = 0.741) это оводненность лист ь е в , а д л я 8Ира кгу1оуЦ (Я^ = 0.558) — интенсивность транспирации (увеличение скорости испарения воды и ее содержания в листьях сопровождается снижением д е ф и ц и т а н а с ы щ е н и я ) . Д л я 1пз сИсИоЮта в ы я в л е н а д о с т а т о ч н о тесная (обратная) связь (г^ = 0.696) водного д е ф и ц и т а с в л а ж н о с т ь ю почвы. На водный д е ф и ц и т 5Ира кгу1оуЦ к р о м е в л а ж н о с т и п о ч в ы оказывает заметное п р я м о е влиян и е о с в е щ е н н о с т ь {К^ = 0.848). Этот анализ показывает значительное влияние условий увлажнения экотопа (либо в л а ж н о с т и почвы, либо влажности воздуха) на сезонную динамику д е ф и ц и т а водного насыщения у всех исследованных видов.
Содержание воды в листьях р а с т е н и й . О ч е н ь в ы с о к а я оводн е н н о с т ь л и с т ь е в (4 .724 г,оды •^сух.массы) была выявлсна у суккулента / т сИс1го1ота. Значительно меньшее количество воды содержат листья АПетша /гщШа (2.083 г,„аы •^'сух.массы)кРи1^а(И1а Шгкгаптот (2.044 г,^^^ • г^^!, ).
60 п
50 -
40 -
30
20
10
35
30 -
25 -
55 20 -
Ч 15 -
10 -
5 -
0 -22июн 6 июл 20 июп 3 авг Павг 31 авг
Дни сезона
Рис . 2.1.2. С е з о н н ы е и з м е н е н и я д е ф и ц и т а водного н а с ы щ е н и я у в и д о в к о в ы л ь н о - р а з н о т р а в н о -
вострецового со о бщ ест ва .
а) 1 — Аг1ет1з1а/п§Ша, 2 — Ро1еп1Ша 1апасеН/оИа, 3 — 8ира кгу1оуИ; 6) 4 — 1г13 сИсЬоЮта, 5 — Ьеутиз
сЫпетгз, 6 — РиЬаНИа ШгсштпоуИ.
сух.массы . Группу в и д о в с н а и м е н ь ш е й о в о д н е н н о с т ь ю листьев представляют Ьеутт сЫпети (1.458 г^„з„ •г;^!^ „„„„ ) , РошНПа ШпасефПа (1.793 г^„^„ - ^сух.массы (0.949 г .
) и 5Нра кгу1огИ
воды ^^суx.,тссы ) (табл. 2.1.1). Как видно , / т сИско1ота и 8Ира кгу1оуИ представляют собой крайние ч л е н ы этого ряда . У первого из них т и п и ч н ы е значения показателя н а х о д и л и с ь в г р а н и ц а х 0.828-1.073 г^ О Й Ы ''^сух.массы, У второго — м е ж д у 3.979 и 5.546 г^^^^ •г^ '̂̂ .кассы • У о с т а л ь н ы х видов т и п и ч н ы е м и н и м у м ы о во дненно с ти листьев оц ен и ваю тся ч и с л а м и 1.248^1.648 г^о^^ •г'^х..тссы > а т и п и ч н ы е м а к с и м у мы — 1.679-2.691 г,ой„ т сух.массы . П о д в и ж н о с т ь признака низкая {V~ 11.4-^23.2%),
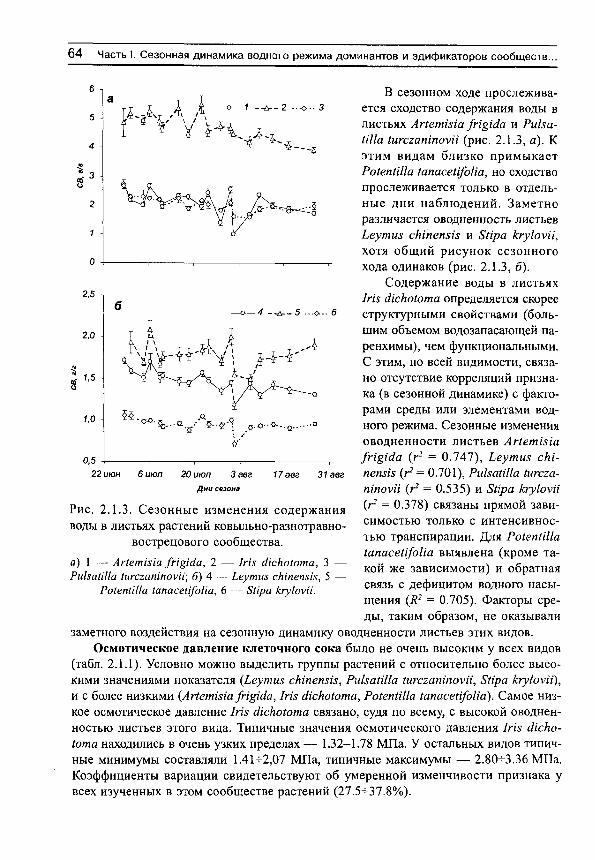
64 Часть 1. Сезонная динамика водного режима доминантов и эдификаторов сообществ..
3"
А -1 - ^ - 2 . . .о. . 3
5..
в с е з о н н о м ходе п р о с л е ж и в а ется сходство с о д е р ж а н и я воды в листьях АПетШа /п§Ша и РиЬа-иИа Ыгсгатпоуп (рис . 2.1.3, а). К э т и м в и д а м б л и з к о п р и м ы к а е т Ро1еп(И1а ШпасеИ/оНа, но сходство п р о с л е ж и в а е т с я только в отдельн ы е д н и н а б л ю д е н и й . З а м е т н о различается оводненность листьев Ьеутш сНшепзгз и 8Ира кгу1оуп, хотя о б щ и й р и с у н о к с е з о н н о г о хода одинаков (рис . 2.1.3, б).
С о д е р ж а н и е в о д ы в л и с т ь я х 1пз (ИскоЮта определяется скорее с т р у к т у р н ы м и с в о й с т в а м и (больш и м объемом водозапасающей паренхимы) , чем функциональными. С этим, по всей видимости , связано отсутствие корреляций признака (в сезонной динамике) с фактор а м и среды или э л е м е н т а м и водного режима. Сезонные изменения о в о д н е н н о с т и л и с т ь е в Аг1ет1з1а ^гщШа (И = 0.747), Ьеутш сЫ-пепш (И = 0.701), Ри1шШ1а (игсга-тпоуп (г^ = 0.535) и БИра кгу1оуп {И = 0.378) связаны п р я м о й завис и м о с т ь ю только с и н т е н с и в н о с т ь ю т р а н с п и р а ц и и . Д л я РоШНИа ШпасеН/оИа в ы я в л е н а ( к р о м е такой ж е з а в и с и м о с т и ) и о б р а т н а я связь с д е ф и ц и т о м водного н а с ы щ е н и я {К^ = 0.705). Факторы сред ы , т а к и м образом, не оказывали
заметного воздействия на сезонную динамику оводненности листьев этих видов.
О с м о т и ч е с к о е д а в л е н и е клеточного сока было не очень в ы с о к и м у всех видов (табл. 2.1.1). Условно м о ж н о выделить группы р а с т е н и й с о т но сит ель но более в ы с о кими значениями показателя {Ьеутш сМпепзи, РиЬаИИа (игсгашпоуп, 8Ира кгу1оуЦ), и с более низкими {Аг(етша/гщШа, (ИскоЮта, Ро1епШ1а (апасеН/оНа). Самое низкое осмотическое давление 1п$ сИско1ота связано, судя по всему, с высокой оводненностью листьев этого вида . Т и п и ч н ы е значения о смо т ическо г о д а в л е н и я Мз (ИскоЮта находились в очень узких пределах — 1.32-1.78 МПа . У остальных видов типичные минимумы составляли 1.41+2,07 М П а , типичные м а к с и м у м ы — 2.80+3.36 М П а . К о э ф ф и ц и е н т ы в а р и а ц и и свидетельствуют об у м е р е н н о й и з м е н ч и в о с т и п р и з н а к а у всех изученных в этом сообществе растений (27.5+37.8%).
2,5
2,0
1,5
1,0
0,5 22июн 6 июп 20 июл 3 авг
Дни сезона 17 авг 31 авг
Р и с . 2.1.3. С е з о н н ы е и з м е н е н и я с о д е р ж а н и я воды в листьях растений ковыльно-разнотравно-
в о с т р е ц о в о г о сообщества .
а) 1 — Аг1ет1з1а /гщШа, 2 — 1пз (ИсИо1ота, 3 — РиЬаИНа Шгсгаптоуп; б) 4 — Ьеутиз сктепш, 5 —
Ро1еп1И1а ШпасеИ/оИа, 6 — 5Нра кгу!оуЦ.
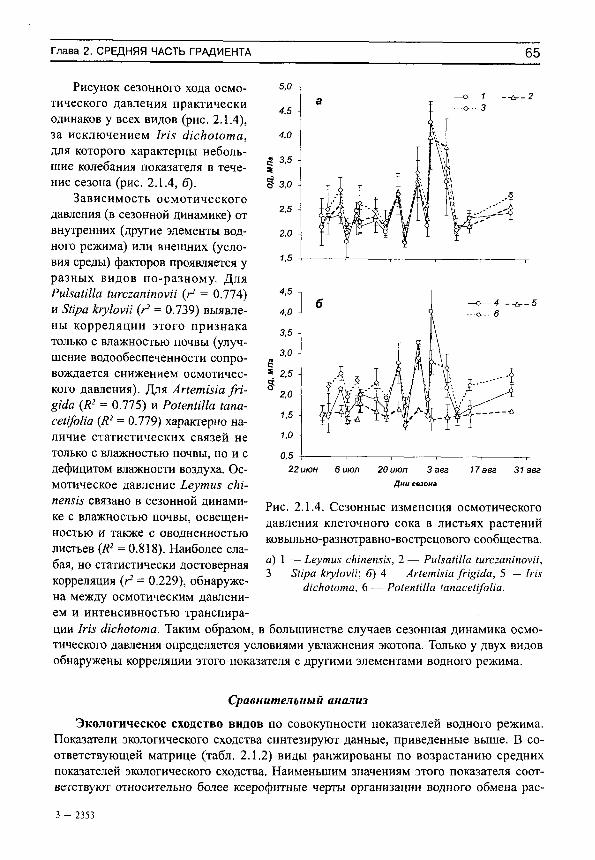
Глава 2. СРЕДНЯЯ ЧАСТЬ ГРАДИЕНТА 65
Рисунок сезонного хода осмотического д а в л е н и я п р а к т и ч е с к и одинаков у всех видов (рис. 2.1.4), за и с к л ю ч е н и е м 1г18 сИсИоЮта, для которого характерны неболь ш и е колебания показателя в течение сезона (рис. 2.1.4, б).
З а в и с и м о с т ь о с м о т и ч е с к о г о давления (в сезонной динамике) от внутренних (другие элементы водного режима) ил и в н е ш н и х (условия среды) факторов проявляется у р а з н ы х в и д о в п о - р а з н о м у . Д л я РиЬаНИа ШгсгатпоуИ (И = 0.774) и 8Ира кгу1оуп (г^ = 0.739) выявлен ы к о р р е л я ц и и э т о г о п р и з н а к а только с влажностью почвы (улучшение водообеспеченности сопровождается с н и ж е н и е м осмотичес кого давления) . Д л я Аг(ет1з1а /п-§Ша (К^ = 0.775) и Ро1епШ1а Шпа-сеИ/оНа (К^ = 0.779) характерно нал и ч и е с т а т и с т и ч е с к и х с в я з е й не только с влажностью почвы, но и с дефицитом влажности воздуха. Осмотическое д а в л е н и е Ьеутш сЫ-пепз1з связано в сезонной д и н а м и ке с в л а ж н о с т ь ю почвы, освещенностью и также с о в о д н е н н о с т ь ю листьев (Л^ = 0.818). Наиболее слабая, но статистически достоверная корреляция (г^ = 0.229), обнаружена между о с м о т и ч е с к и м давлением и и н т е н с и в н о с т ь ю т р а н с п и р а ции Мз сИсНоЮта. Таким образом, в большинстве случаев сезонная д и н а м и к а осмотического давления определяется условиями увлажнения экотопа. Только у двух видов обнаружены корреляции этого показателя с другими элементами водного режима .
4,5
4,0
3,5
3,0
2,5
2,0
1,5
1,0
0,5 22 июн 6 июл 20 июп 3 авг
Дни сезона 17 авг 31 авг
Рис . 2.1.4. С е з о н н ы е и з м е н е н и я осмотического д а в л е н и я к л е т о ч н о г о со ка в л и с т ь я х р а с т е н и й ковыльно-разнотравно-вострецового сообщества.
й) 1 — Ьеутиз сЫпет'13, 2 — РиЬаНИа Шгсгаптоуп, 3 — 8Ира кгу1оуИ; б) 4 — Апетта /п^Ша, 5 — 1г1з
сИскоСота, 6 — Ро1еп1И1а (апасеН/оИа.
Сравнительный анализ
Э к о л о г и ч е с к о е сходство видов по совокупности показателей водного р е ж и м а . Показатели экологического сходства синтезируют данные , приведенные в ы ш е . В соответст вую щей м а т р и ц е (табл . 2.1.2) в и д ы р а н ж и р о в а н ы п о в о з р а с т а н и ю с р е д н и х показателей экологического сходства. Н а и м е н ь ш и м значениям этого показателя соответствуют относительно более ксерофитные черты организации водного обмена рас -
3 - 2353
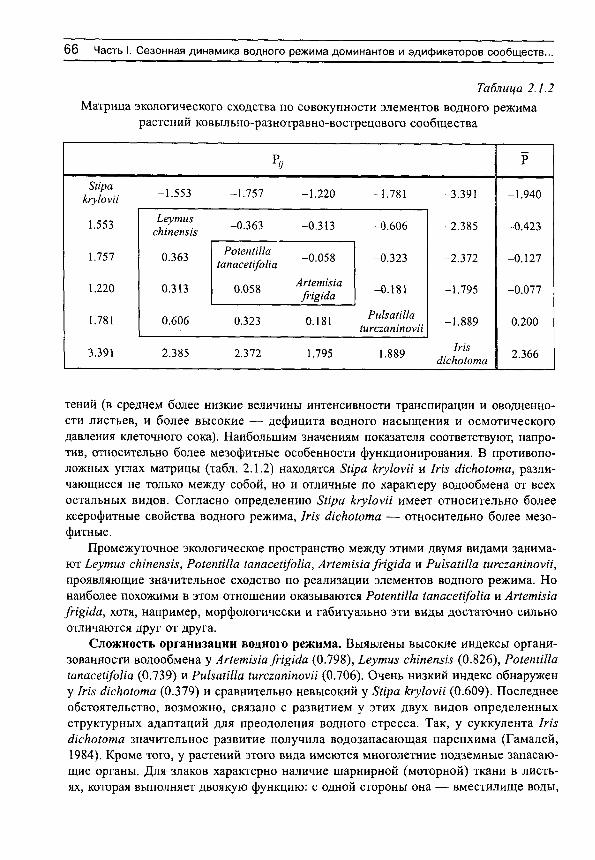
66 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ...
Таблица 2.1.2
Матрица экологического сходства по совокупности элементов водного р е ж и м а растений ковыльно-разнотравно-вострецового с о о б щ е с т в а
Р
8Ира кгу1оуЦ -1.553 -1.757 -1.220 -1.781 -3.391 -1.940
1.553 Ьеутиз сктепзьз -0.363 -0.313 -0.606 -2.385 -0.423
1.757 0.363 Ро1епШ1а (апасеИ/оНа -0.058 -0.323 -2.372 -0.127
1.220 0.313 0.058 АПетша -0.181 -1.795 -0.077
1.781 0.606 0.323 0.181 РиЬаННа 1игс2атпоуп -1.889 0.200
3.391 2.385 2.372 1,795 1.889 1пз
сИско1ота 2.366
тений (в среднем более низкие величины интенсивности транспирации и оводненности л и с т ь е в , и б о л е е в ы с о к и е — д е ф и ц и т а в о д н о г о н а с ы щ е н и я и о с м о т и ч е с к о г о давления клеточного сока). Наибольшим значениям показателя соответствуют, напротив, относительно более мезофитные особенности функционирования . В противопол о ж н ы х углах м а т р и ц ы (табл. 2.1.2) находятся 8Ира кгу1оуИ и Мз (ИсНо1ота, р а з л и чающиеся не только м е ж д у собой, но и отличные по характеру водообмена от всех о с т а л ь н ы х в и д о в . С о г л а с н о о п р е д е л е н и ю 5Нра кгу1оуИ и м е е т о т н о с и т е л ь н о более ксерофитные свойства водного режима , 1пз сИскоЮта — о т но сит ель но более мезо фитные .
П р о м е ж у т о ч н о е экологическое пространство м е ж д у э т и м и дву мя видами занимают Ьеутиз сктепз1з, Ро1епШ1а ШпасеИ/оИа, Аг1ет1з1а/гщЫа и РиЬаННа ШгсгатпоуИ, проявляющие значительное сходство по р е а л и з а ц и и э лемент о в водного р е ж и м а . Н о наиболее похожими в этом отношении оказываются РоШИПа ШпасеИ/оНа и Апет1з1а /гщШа, хотя, н а п р и м е р , м о р ф о л о г и ч е с к и и габитуально эти в и д ы д о с т а т о ч н о силь но отличаются друг от друга .
С л о ж н о с т ь о р г а н и з а ц и и водного режима . В ы я в л е н ы в ы с о к и е индексы организованности водообмена у Аг1ет1з1а/гщШа (0.798), Ьеутиз сктепз1з (0.826), РоШИИа (апасеИ/оИа (0.739) и РиЬаИПа (игсгатпоуп (0.706). О ч е н ь низкий индекс о бнар у жен у 1пз сИско1ота (0.379) и сравнительно невысок ий у ЗИра кгу1оуИ (0.609). П о с л е д н е е обстоятельство , в о з м о ж н о , с в я з а н о с р а з в и т и е м у этих д в у х в и д о в о п р е д е л е н н ы х с т р у к т у р н ы х а д а п т а ц и и д л я п р е о д о л е н и я водного с т р е с с а . Так , у с у к к у л е н т а 1пз сИско1ота з н а ч и т е л ь н о е р а з в и т и е п о л у ч и л а в о д о з а п а с а ю щ а я п а р е н х и м а ( Г а м а л е й , 1984). Кроме того , у р а с т е н и й этого вида и м е ю т с я м н о г о л е т н и е п о д з е м н ы е з а п а с а ю щие органы. Д л я злаков характерно н а л и ч и е ш а р н и р н о й ( м о т о р н о й ) ткани в листь ях, которая выполняет д в о я к у ю функцию: с одной стороны она — в м е с т и л и щ е воды.

Глава 2. СРЕДНЯЯ ЧАСТЬ ГРАДИЕНТА 67
С другой — способствует свертыванию листьев при их о без во живании , чем достига ется эфф ект и вн ое с о к р а щ е н и е т р а н с п и р и р у ю щ е й п о в е р х н о с т и (Гамалей , 1984). Это свойственно, впрочем, не только 5Ира кгу1оуИ, но и Ьеутиз сЫпепзи, с его в ы с о к и м индексом орган и зованности . М о ж н о предположить , что в некоторых случаях структурная р е г у л я ц и я водного о б м е н а может д о п о л н я т ь ф у н к ц и о н а л ь н у ю , а в некотор ы х —• ослаблять ее.
Взаимосвязи м е ж д у элементами водного р е ж и м а почти у всех видов ограничиваются корреляциями м е ж д у о в о д н е н н о с т ь ю листьев и и н т е н с и в н о с т ь ю т р а н с п и р а ц и и . В некоторых случаях выявляются и другие з ависимо ст и . Однако степень организованности водного р е ж и м а определяется не только в н у т р е н н и м и , но и в н е ш н и м и связями. С этой точки зрения практически все виды, за и с к л ю ч е н и е м Мз <ИсНо1ота, демонстри рую т достаточно в ы с о к у ю (для д а н н ы х у с л о в и й пр о из р аст ания ) с л о ж н о с т ь организации водного обмена .
Э л е м е н т ы водного р е ж и м а п о д в е р ж е н ы з а м е т н о м у в л и я н и ю ф а к т о р о в среды и реакция их в б о л ь ш и н с т в е случаев адекватна и з м е н е н и я м у с л о в и й п р о и з р а с т а н и я . В а ж н е й ш и м и из этих факторов являются в л а ж н о с т ь почвы и воздуха . Но в некотор ы х случаях з амет н ое влияние на с е з о н н у ю д и н а м и к у элементов во до о бмена оказывает о с в е щ е н н о с т ь . Так, у Ьеутиз сЫпепзгз и ЗИра кгу1оуЦ о с в е щ е н н о с т ь влияет на сезонную динамику интенсивности транспирации. Еще Н. А. М а к с и м о в (1952) указывал на важное влияние света на процессы водообмена у растений. В . Г. Реуцкий (1984) показал в ф а к т о р о с т а т н о м э к с п е р и м е н т е с у щ е с т в о в а н и е с в е т о з а в и с и м о г о потока у растений . Сходные д а н н ы е содержатся в работе Н. Н. П о п о в а (1971). П о н а ш и м (не-
Таблица 2.1.3
Оп ределен и е относительной конкурентоспособности растений ковыльно-разнотравно-вострецового с о о б щ е с т в а
Вид
Обилие за последний год
(% от общей массы травостоя)
Сумма обилий за последние 5 лет
(% от общей массы травостоя)
Разность обилий (последний год минус первый
год)
Сумма мест
( I М) Место КСА Вид
величина место величина место величина место
Сумма мест
( I М) Место КСА
АПет1з1а 5.6 4 47.6 3 1.6 3 10 4 0.44
1пз 5.8 3 17.7 4 2.9 2 9 3 0.50 (ИскоЮта Ьеутиз 28.3 1 153.8 1 -5.2 6 8 2 0.56 сЫпепз1з РоШИИа 0.7 6 6.9 6 -0.5 4 16 6 0.11 ШпасеН/оНа РиЫиИа 1.3 5 11.6 5 -2.0 5 15 5 0.17 (игсгатпоуп 5Ира кгу1оуЦ 15.8 2 47.6 2 9.3 1 5 1 0.72
Примечание: КСА = 1 - X М / X М т а х , где X М т а х — максимально возможная сумма мест (в данном примере 18).
3»

68 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ..
опубликованным) д а н н ы м , п о л у ч е н н ы м в лаборат о р ных условиях , у Уша заНуа скорость ксилемного тока резко увеличивалась после включения света в течение 2-5 минут при н е и з м е н н о й температуре воздуха. Ф а к т ы свидетельствуют о том, что и в е с тественных у с л о в и я х произрастания свет оказывает (в некоторых с л у ч а я х ) з а м е т н о е влияние на некоторые элементы водного р е ж и м а растений .
Относительная конкурентоспособность видов. В нащем распоряжении имеются данные по д и н а м и к е живой надземной фитомассы видов ковыльно-разнотравно-вост-
рецо во г о с о о б щ е с т в а за 1982-1986 гг. ( Ш е р е м е т ь е в , Ж а р г а л с а й х а н , 1990). Это позволило определить относительн у ю к о н к у р е н т о с п о с о б н о с т ь р а с т е н и й по методу В . Д . Л о п а т и н а (1973). Ход о п р е д е л е н и я э т о г о и н д е к с а {КСА — ге1а11уе с о т р е 1 1 и у е аЫИ1у) п о к а з а н в табл . 2.1.3, из которой видно , что наиболее ко нку р ент о спо со бным видом я в ляется 8Ира кгу1оуИ. Затем, в порядке у м е н ь ш е н и я о т н о с и т е л ь н о й конкурент о с п о с о б н о с т и , в и д ы р а с п о л а г а ю т с я с л е д у ю щ и м образом: Ьеутш сИтешй, Мз (ИскоЮта, АПетша /гщШа, Ри1-заНПа гигсгаптоуп, РаепИИа 1апа-сей/оИа.
Анализ выя вил очень т е с н у ю связь (г^ = 0.836) м е ж д у о т н о с и т е л ь н о й конкурентоспособность растений и интенс и в н о с т ь ю т р а н с п и р а ц и и (рис . 2.1.5).
Из этой з а в и с и м о с т и следует, что чем ниже скорость и с п а р е н и я воды, тем в ы ш е относительная к о н к у р е н т о с п о с о б н о с т ь вида. К сходному выво ду на о с н о в а н и и сво их многолетних наблюдений п р и ш л а Н. Н. Измайлова (1986). Этот факт возвращает нас к взглядам Ш и м п е р а ( З с Ы т р е г , 1898, цит. по : Генкель, 1969) о том , что к с е р о ф и т ы переносят засуху благодаря малой и н т е н с и в н о с т и т р а н с п и р а ц и и . Э т а с п о с о б н о с т ь .
Таблица 2.1.4
О п р е д е л е н и е э ф ф е к т и в н о с т и использования п о ч в е н н о й влаги видами ковыльно-разнотравно-вострецового с о о б щ е с т в а ( средние за 1984-1985 гг.)
Вид А Б Эф = А/Б
Аг1ет1Б1а /п^Ша 8.1 7.9 1.02 1п8 (Иско1ота 4.3 3.6 1.18 Ьеутиз сЫпепзгз 20.7 29.0 0.71 Ро1епШ1а 1апасеИ/оНа 1.8 1.8 0.97 РиЬайИа Шгсгаптоуп 2.6 2.4 1.08 8Ира кгу1оуИ 4.8 7.7 0.62
0,8 -I
0,6
8 0.4 .
0.2 -
г' = 0.836 0.0
0.5 1.0 1.5 ИТ, г/(г-ч)
2,0
Рис. 2.1.5. Связь м е ж д у индексами относительной к о н к у р е н т о с п о с о б н о с т и (КСА) и и н т е н с и в н о с т ь ю т р а н с п и р а ц и и для видов к о в ы л ь н о - р а з н о т р а в н о - в о с т р е ц о в о г о
сообщества .
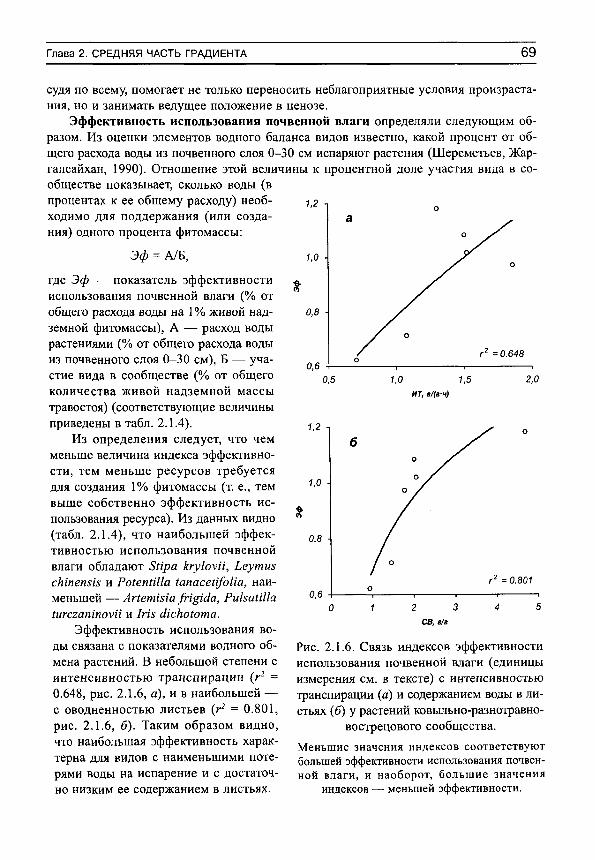
Глава 2. СРЕДНЯЯ ЧАСТЬ ГРАДИЕНТА 69
судя по всему, помогает не только переносить неблагоприятные у сло вия произрастания, но и занимать ведущее положение в ценозе .
Э ф ф е к т и в н о с т ь и с п о л ь з о в а н и я п о ч в е н н о й влаги о п р е д е л я л и с л е д у ю щ и м образом. И з о ц е н к и элементов водного баланса видов известно , какой п р о ц е н т от общего расхода воды из почвенного слоя 0-30 см испаряют растения (Шереметьев , Жаргалсайхан, 1990). О т н о ш е н и е этой в е л и ч и н ы к процентной д о л е у ч а с т и я вида в соо б щ е с т в е показывает , сколько воды (в процентах к ее о б щ е м у расходу) необ- 12 х о д и м о д л я п о д д е р ж а н и я (или создания) одного процента фитомассы:
Эф = А/Б ,
где Эф — п о к а з а т е л ь э ф ф е к т и в н о с т и использования п о ч в е н н о й влаги (% от общего расхода воды на 1 % живой надземной фитомассы) , А — расход воды растениями (% от общего расхода воды из почвенного слоя 0-30 см) , Б — участие вида в с о о б щ е с т в е ( % от общего к о л и ч е с т в а ж и в о й н а д з е м н о й м а с с ы травостоя) (соответствующие величины приведены в табл . 2.1.4).
Из о п р е д е л е н и я следует , что ч е м меньше величина индекса эффективности , т е м м е н ь ш е р е с у р с о в т р е б у е т с я для создания 1 % ф и т о м а с с ы (т. е., тем в ы ш е с о б с т в е н н о э ф ф е к т и в н о с т ь и с пользования ресурса) . Из данных видно (табл . 2.1.4), ч т о н а и б о л ь ш е й э ф ф е к т и в н о с т ь ю и с п о л ь з о в а н и я п о ч в е н н о й влаги о б л а д а ю т 8Ира кгу1оуп, Ьеутш сктепз1з и Ро1епй11а ШпасеИ/оИа, наименьшей — АПетта /гщШа, РиЬаЧИа Шгсгаптоуп и / т сИсНоЮта.
Э ф ф е к т и в н о с т ь использования вод ы связана с показателями водного обмена растений. В небольшой степени с и н т е н с и в н о с т ь ю т р а н с п и р а ц и и (г^ = 0.648, рис . 2.1.6, а), и в наибольшей — с о в о д н е н н о с т ь ю л и с т ь е в (Н = 0.801, р и с . 2.1.6, б ) . Т а к и м о б р а з о м в и д н о , что н а и б о л ь ш а я э ф ф е к т и в н о с т ь характерна для видов с н а и м е н ь ш и м и потерями воды на и сп арение и с достаточно низким ее с о д е р ж а н и е м в листьях .
1,0
0,8 •
0,6 = 0.648
0,5 1,0 1,5 2,0 ИТ, г/(гч)
1,2 п
1,0
0,8
0,6 г' = 0.801
0 1 2 3 4 5 СВ. г/г
Рис. 2.1.6. Связь индексов э ффект ивно ст и использования п о ч в е н н о й влаги ( единицы измерения см. в тексте) с и н т е н с и в н о с т ь ю транспирации (а) и содержанием воды в листьях (б) у растений ковьшьно-разнотравно-
вострецового со о бщ ест ва .
Меньшие значения индексов соответствуют большей эффективности использования почвенной влаги, и наоборот , большие значения
индексов — меньшей эффективности.
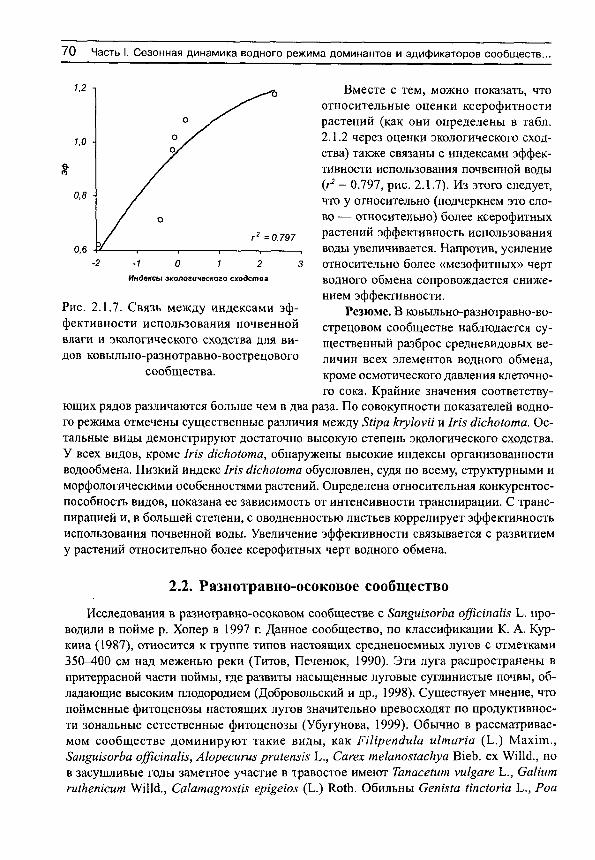
70 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ.
В м е с т е с тем , м о ж н о показать , что о т н о с и т е л ь н ы е о ц е н к и к с е р о ф и т н о с т и р а с т е н и й (как они о п р е д е л е н ы в табл . 2.1.2 через оценки экологического сходства) также связаны с индексами эффективности использования почвенной воды (Н = 0.797, р и с . 2.1.7). И з этого следует, что у относительно (подчеркнем это слово — относительно) более ксерофитных растений э ф ф е к т и в н о с т ь использования воды увеличивается. Напротив, усиление относительно более « м е з о ф и т н ы х » черт водного обмена сопровождается снижением эффективности .
Резюме. В ковыльно-разнотравно-во-стрецовом с о о б щ е с т в е наблюдается сущест венны й р а з б р о с средневидовых велич ин всех элемент о в водного обмена , кроме осмотического давления клеточного сока. К р а й н и е значения соответству
ю щ и х рядов различаются больше чем в два раза. По совокупности показателей водного р е ж и м а отмечены существенные различия между ЗИра кгуЬуп и / т сИскоЮта. О с тальные в и д ы д е м о н с т р и р у ю т достаточно в ы с о к у ю степень экологического сходства. У всех видов , кроме Мз сИскоЮта, обнаружены высокие и н д е к с ы организованности водообмена. Н и з к и й индекс 1пз сИскоЮта обусловлен, судя по всему, с т р у кт у р ны ми и морфологическими особенностями растений. Определена относительная конкурентоспособность видов , показана ее зависимость от интенсивности т р а н с п и р а ц и и . С транс-пирацией и, в б о л ь ш е й степени , с оводненностью листьев коррелирует эффект ивно ст ь использования почвенной воды. Увеличение эффективности связывается с развитием у растений относительно более ксерофитных черт водного обмена .
2.2. Разнотравно-осоковое сообщество
Исследования в разнотравно-осоковом сообществе с 8ап§ш5огЬа о01стаИз Ь . пр о водили в п ой ме р . Хопер в 1997 г Данное сообщество , по к л а с с и ф и к а ц и и К. А. Кур-кина (1987), относится к группе т и п о в настоящих с р е д н е п о е м н ы х лугов с отметками 350-400 см н а д м е ж е н ь ю р е к и (Титов, Печенюк , 1990). Э т и луга р а с п р о с т р а н е н ы в притеррасной части поймы, где развиты насыщенные луговые суглинистые почвы, обладающие высоким плодородием (Добровольский и др., 1998). Существует мнение, что пойменные фитоценозы настоящих лугов значительно превосходят п о продуктивнос ти зональные естественные фитоценозы (Убугунова, 1999). О б ы ч н о в рассматривае м о м с о о б щ е с т в е д о м и н и р у ю т т а к и е в и д ы , к а к ртрепс1и1а и1таг1а (Ь . ) М а х 1 т . , Зап^шзогЪа о^стаНз, А1оресигиз рга1епз1з Ь . , Сагех те1апоз1аскуа В1еЬ. ех ХУНЫ., но в засушливые годы заметное участие в травостое и м е ю т ТапасеШт уи1§аге Ь . , СаНит гШкепкит ШЫ., Са1ата^тзИз ерщеюз (Ь.) НоШ. О б и л ь н ы СетзШ Ипс1опа Ь . , Роа
1,0 •
0,8
0,6 г' ^0.797
- 2 - 1 0 1 2 Индексы экологического сходства
Р и с . 2.1.7. С в я з ь м е ж д у и н д е к с а м и э ф ф е к т и в н о с т и и с п о л ь з о в а н и я п о ч в е н н о й влаги и экологического сходства для видов ковыльн о-разнотравно-вострецового
сообщества .
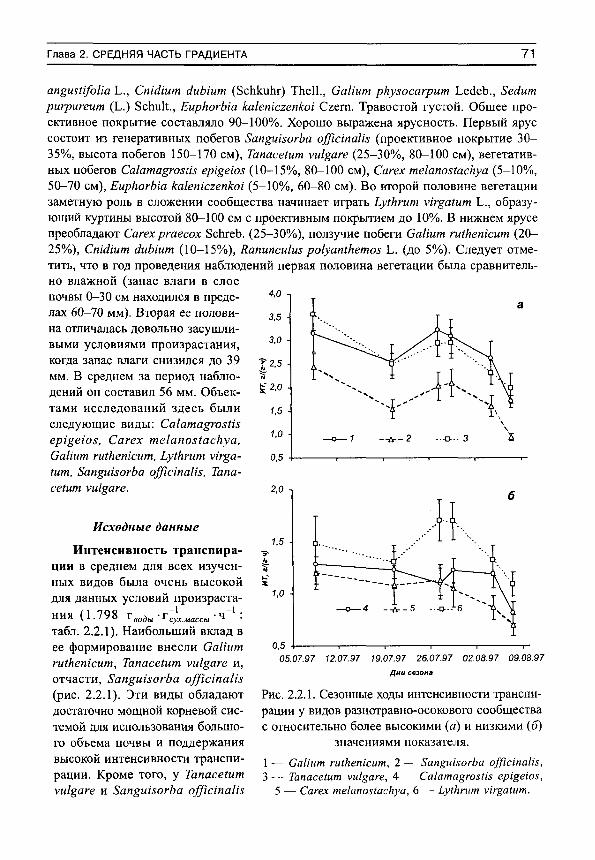
Глава 2. СРЕДНЯЯ ЧАСТЬ ГРАДИЕНТА 71
ап§тИ/оИа ^., СпШит с1иЫит (8сЬкиЬг) ТЬе11., ОаНит ркузосагрит ЬейеЬ. , 5ес1ит ригригеит (Ь.) ЗсЬиН., ЕиркогЫа ШепкгепШ С г е т . Травостой густой . О б щ е е проективное покрытие составляло 90-100%. Хорощо выражена ярусность . П е р в ы й ярус состоит из генеративных побегов 8ап§ш5огЬа о^стаНз (проективное покрытие 3 0 -35%, высота побегов 150-170 см) , ТапасеШт уи1§аге (25-30%, 80-100 см) , вегетативных побегов Са1ата§гозИз ерщеюз (10-15%о, 80-100 см), Сагех теЬпозШскуа (5-10%), 50-70 см) , ЕиркогЫа кактсгепШ (5-10%), 60-80 см) . В о второй половине вегетации заметную роль в сложении сообщества начинает играть Ьу1кгит у1щаШт Ь . , образующий куртины высотой 80-100 см с проективным покрытием д о 10%. В нижнем ярусе преобладают Сагех ргаесох 8сЬгеЬ. (25-30%)), ползучие побеги СаИит ги1кетсит ( 20 -25%)), СпШшт (1иЫит (10-15%)), КапипсиЫз ро1уап1кетоз Ь . (до 5%). Следует отметить , что в год проведения наблюдений первая половина вегетации была сравнительно в л а ж н о й ( запас влаги в с л о е почвы 0-30 см находился в пределах 60-70 мм) . Вторая ее половина отличалась довольно засушливыми у с л о в и я м и п р о и з р а с т а н и я , когда запас влаги снизился до 39 мм. В среднем за период наблюдений он составил 56 мм. Объект а м и и с с л е д о в а н и й з д е с ь б ы л и с л е д у ю щ и е в и д ы : Са1ата^гозНз ерг^еюз, Сагех те1апоз1аскуа, ОаНит ги1кетсит, ЬуЛгит у1г§а-Шт, Зап^шзогЬа о^рстаИз, ТапасеШт Уи1§аге.
4,0
3,5
3,0
"а ^-2,0 -I
1,5
1,0
0,5
2,0
Исходные данные
И н т е н с и в н о с т ь т р а н с п и р а ции в среднем д л я всех изученных в и д о в б ы л а о ч е н ь в ы с о к о й для д а н н ы х у с л о в и й произраста н и я (1 .798 г,„аыт;;!^.л,ассы-ч" ' : табл. 2.2.1). Н а и б о л ь ш и й вклад в ее ф о р м и р о в а н и е в н е с л и СаИит гШкетсит, ТапасеШт уи1§аге и, о т ч а с т и , 8ап§шзогЬа о//1стаНз (рис. 2.2.1). Э т и в и д ы о б л а д а ю т достаточно мощной корневой системой для использования большого объема почвы и п о д д е р ж а н и я высокой интенсивности транспир а ц и и . К р о м е того , у ТапасеШт уи1§аге и 5ап§шзогЬа о/рсшаИз
1,5
I 1,0
0,5 05.07.97 12.07.97 19.07.97 26.07.97 02.08.97 09.08.97
Дни сезона
Рис. 2.2.1. Сезонные ходы интенсивности транспирации у видов разнотравно-осокового сообщества с относительно более высокими (а) и низкими (б)
значениями показателя .
1 — ОаИит гШкетсит, 2 — 8ап§шзогЬа о/ДсшаИз, 3 — ТапасеШт уи1§аге, 4 — Са1ата§гозИз ер1^еюз,
5 — Сагех те1апоз1аскуа, 6 — ЬуЛгит \чг^аШт.

72 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ..
Таблица 2.2.1
Средние значения интенсивности т р а н с п и р а ц и и (ИТ, г^^^^, •т~1^,„ассы - ч " ' ) ; д е ф и ц и т а водного н а с ы щ е н и я ( Д В Н , % ) ; содержания воды в листьях ( С В , г^^^^ •'^~с1х..массы )
у растений разнотравно-осокового с о о б щ е с т в а
Виды Показатели
Виды ИТ ДВН СВ Виды
п X 8х п X 8х п X Са1ата^говИз ер1§еюз 120 1.139 0.368 120 15.8 6.5 120 1.504 0.110 Сагех те1апоз1аскуа 120 1.003 0.376 120 21.6 3.7 120 1.507 0.109 ОаНит тгНетсит 120 2.726 0.916 120 21.3 5.3 120 2.331 0,310 Ьу1кгит ргг^аШт 120 1.432 0.464 120 20.6 4.3 120 1.624 0.171 5ап§ш50гЬа о//1стаНз 120 1.778 0.724 120 10.6 5,0 120 1.797 0.242 ТапасеШт уи1§аге 120 2.711 0.831 120 14.1 3.9 120 2.289 0,252 В целом для сообщества 720 1.798 0.951 720 17.3 6.4 720 1.842 0,405
нет заметных структурных приспособлений для ограничения потерь воды, что также сказывается на скорости ее испарения . Однако столь высокие затраты влаги на испарение привели к тому, что уже в конце июля, когда запасы воды в почве стали постепенно снижаться , э ти виды начали терять листовую массу из-за в ы с ы х а н и я листьев . Интенсивность транспирации также заметно снизилась (на 1.4-1.6 г ^ а ы •'<^~с1х.массы ' Ч ^ ' за период н а б л ю д е н и й ) , тогда как у Са1ата§гозаз ерщеюз, Сагех те1апоз(аскуа и 1у(кгит угг^аШт с н и ж е н и е б ы л о незначительным (на 0.4-0.5 г^о^ы '^с1х массы " ч " ' ) -
Т и п и ч н ы е значения признака находились между 1.6 и 3.89 г^^^^ '^^ух.массы ч ' у ОаИит гШкетсит, и между 1.68 и 3.76 г ^ ^ ^ '"^ сух массы ' ч ' У ТапасеШт уи1§аге. В то же время границы т и п и ч н ы х колебаний интенсивности транспирации у СаЬта^гозНз ер1§еюз и Сагех теШпозШскуа составляли 0.65-1.63 и 0.59-1.49 г^о^^ '^сух.массы ' ч ' с о о т в е т с т в е н н о . Г р а н и ч н ы е з н а ч е н и я п р и з н а к а у Ьу1кгит У1г§аШт ( 0 .87 -2 .02 '^воды '^сух.массы ' ч ' ) сближали Э Т О Т В И Д С растсниями, менее интенсивно испарявшими воду Напротив, 8ап§шзогЪа о^1стаНз по этому признаку (0.90-2.83 г^^^^ •^~с1х.массы ' ч ~ ' ) была более похожа на СаИит гШкетсит и ТапасеШт уи1§аге.
Подвижность интенсивности транспирации у видов разнотравно-осокового сообщества была достаточно умеренной — коэффициенты вариации составляли 30.6-40.7%.
Исследование д и н а м и к и интенсивности т р а н с п и р а ц и и в т е ч е н и е сезона с помощ ь ю множественного регрессионного анализа показало , что у ряда в и д о в изменения данного элемента водного р е ж и м а зависит только от запасов влаги в почве . 8ап§шзог-Ьа о/^стаИз и ТапасеШт уи1^аге продемонстрировали очень в ы с о к у ю степень сопряженности скорости и с п а р е н и я воды от влагообеспеченности экотопа ( к о э ф ф и ц и е н т ы детерминации находились в диапазоне 0.884-0.977). Д л я Ьу1кгит угг^аШт в ы я в л е н а не столь тесная , но статистически достоверная связь (И = 0.658) м е ж д у с к о р о с т ь ю испарения воды и в л а ж н о с т ь ю почвы. Д л я Сагех те1апозШскуа характерна з ависимость т р а н с п и р а ц и и не только от влажности почвы (Р , ,^^= 105.9), но и от д е ф и ц и т а
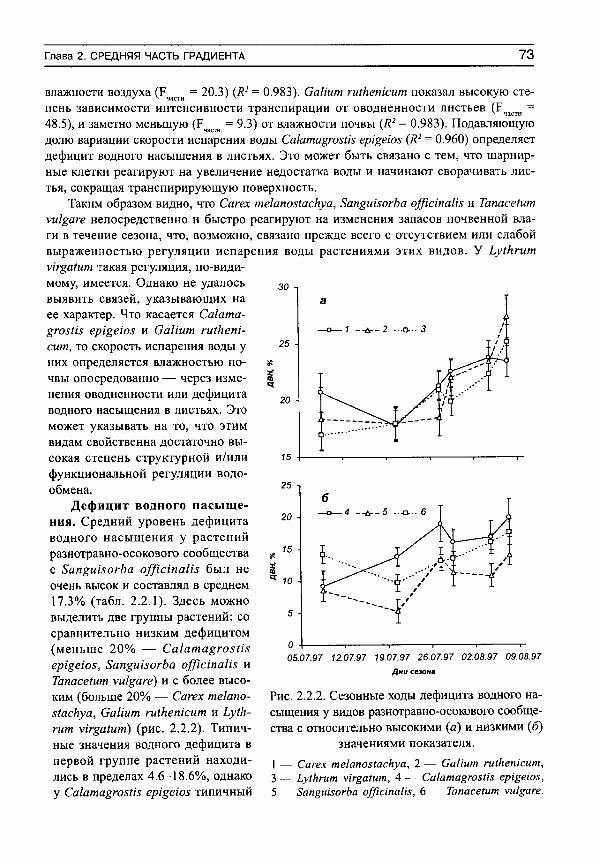
Глава 2. СРЕДНЯЯ ЧАСТЬ ГРАДИЕНТА 73
25
5"
20 -
влажности воздуха (Р,,̂ .̂̂ „ = 20.3) (Я^ = 0.983). СаИит гшНепкит показал высокую степень з а в и с и м о с т и и н т е н с и в н о с т и т р а н с п и р а ц и и от о в о д н е н н о с т и л и с т ь е в (Р,,̂ ,̂̂ „ = 48.5), и заметно меньшую (Р^^^^ = 9.3) от влажности почвы (К^ = 0.983). П о д а в л я ю щ у ю долю вариации скорости испарения воды СаЬта^гозНз ерщею$ {К' = 0.960) определяет д е ф и ц и т водного н а с ы щ е н и я в листьях . Это может быть связано с т е м , что ш а р н и р ные клетки р е а г и р у ю т на увеличение недостатка воды и н а ч и н а ю т сворачивать лис тья, сокращая т р а н с п и р и р у ю щ у ю поверхность .
Таким образом видно, что Сагех те1апо8Шскуа, 8ап§ш80гЬа о/^1стаИз и ТапасеШт уи1§аге непосредственно и быстро р е а г и р у ю т на изменения запасов п о ч в е н н о й влаги в течение сезона, что, возможно , связано прежде всего с о т с у т с т в и е м или слабой в ы р а ж е н н о с т ь ю р е г у л я ц и и и с п а р е н и я в о д ы р а с т е н и я м и э т и х в и д о в . У Ьугкгит У1г§аШт такая регуляция , по-видимому, имеется . О д н а к о не удалось в ы я в и т ь связей , у к а з ы в а ю щ и х на ее характер. Что касается Са1ата-§гозН8 ерщеюз и ОаНит гШкетсит, то скорость испарения воды у них определяется в л а ж н о с т ь ю почвы опосредованно — через изменения оводненности или д е ф и ц и т а водного насыщения в листьях. Это м о ж е т у к а з ы в а т ь на то , что э т и м видам свойственна достаточно высокая с т е п е н ь с т р у к т у р н о й и /или ф у н к ц и о н а л ь н о й р е г у л я ц и и водообмена.
Д е ф и ц и т в о д н о г о н а с ы щ е ния . С р е д н и й у р о в е н ь д е ф и ц и т а в о д н о г о н а с ы щ е н и я у р а с т е н и й разнотравно-осокового сообщества с 8ап^ш$огЬа о/^гсгпаНз б ы л не очень высок и составлял в среднем 17.3% (табл . 2.2.1). З д е с ь м о ж н о выделить д в е г р у п п ы растений ; со с р а в н и т е л ь н о н и з к и м д е ф и ц и т о м ( м е н ь ш е 2 0 % — Са1ата^гозНз ер1^ею5, Зап^шзогЬа о^сшаНз и ТапасеШт уи1§аге) и с более высоким (больше 20%) •— Сагех те1апо-хШскуа, ОаНит пикетсит и Ьу1к-гит у1г§аШт) (рис . 2.2.2). Т и п и ч ные значения водного д е ф и ц и т а в п е р в о й г р у п п е р а с т е н и й н а х о д и лись в пределах 4.6-18.6%), однако у Са1ата§го5Н8 ерщеюз т и п и ч н ы й
15
25 -,
20
15
10
О 05.07.97 12.07.97 19.07.97 26.07.97 02.08.97 09.08.97
Дни сезона
Рис. 2.2.2. Сезонные ходы дефицита водного насыщения у видов разнотравно-осокового сообщества с относительно высокими (а) и низкими (б)
значениями показателя .
1 — Сагех те1апоз1аскуа, 2 — ОаНит Шкетсит, 3 — Ьугкгит У1г§аШт, 4 — Са1ата§гозНз ер1^еюз, 5 — 8ап^т$огЬа о^юпаНз, 6 — ТапасеШт уи1§аге.
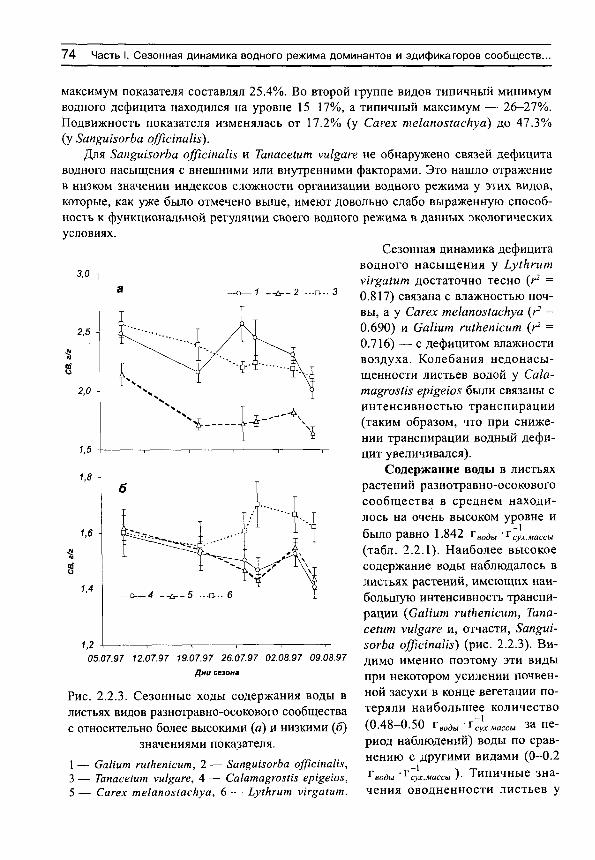
74 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ..
максимум показателя составлял 25.4%. Во второй группе в и д о в т и п и ч н ы й м и н и м у м водного д е ф и ц и т а находился на уровне 15-17%, а т и п и ч н ы й м а к с и м у м — 26-27%. П о д в и ж н о с т ь п о к а з а т е л я и з м е н я л а с ь от 17.2%) (у Сагех те1апо81аскуа) до 47.3%) (у 8ап§шзогЬа о^гсгпаНз).
Для 8ап^ш50гЬа о^стаИз и ТапасеШт \и1^аге не о б н а р у ж е н о связей д е ф и ц и т а водного н а с ы щ е н и я с в н е щ н и м и или внутренними факторами. Это н а ш л о отражение в низком значении индексов сложности организации водного р е ж и м а у этих видов , которые, как уже б ы л о отмечено в ы ш е , имеют довольно слабо в ы р а ж е н н у ю с п о с о б ность к ф у н к ц и о н а л ь н о й регуляции своего водного р е ж и м а в д а н н ы х экологических условиях.
Сезонная д и н а м и к а д е ф и ц и т а в о д н о г о н а с ы щ е н и я у 1у1Нгит угг^аШт д о с т а т о ч н о т е с н о (Н = 0.817) связана с в л а ж н о с т ь ю почвы , а у Сагех те1апо5Шскуа ( г = 0.690) и СаНит гШкетсит {г' = 0.716) — с д е ф и ц и т о м влажности в о з д у х а . К о л е б а н и я н е д о н а с ы -щ е н н о с т и л и с т ь е в водой у Са1а-та^гозНз ерщеюз б ы л и связаны с и н т е н с и в н о с т ь ю т р а н с п и р а ц и и ( т а к и м о б р а з о м , что п р и с н и ж е нии т р а н с п и р а ц и и в о д н ы й д е ф и цит увеличивался ) .
С о д е р ж а н и е в о д ы в листьях р а с т е н и й р а з н о т р а в н о - о с о к о в о г о с о о б щ е с т в а в с р е д н е м н а х о д и -
3,0 п
2,5 -
-1 --^-2 . . .о-. 3
8
2,0 -
1.5
1.8
1,6 -
1.* -
ЛОСЬ на очень вы со ко м у р о в н е и ,-1 сух.массы
1,2 05.07.97 12.07.97 19.07.97 26.07.97 02.08.97 09.08.97
Дни сезона
Рис . 2.2.3. С е з о н н ы е ходы с о д е р ж а н и я воды в листьях видов разнотравно-осокового сообщества с относительно более высокими (а) и низкими (б)
значениями показателя ,
1 — СаИит гШкетсит, 2 — 5ап§шзогЬа о/^гсшаНз, 3 — ТапасеШт уи1§аге, 4 — Са1ата^гозИз ерг^еюз, 5 — Сагех те1апоз1аскуа, 6 — Ьу1кгит У1г§а1ит.
б ы л о р а в н о 1.842 г^^^^ г ( т а б л . 2.2.1). Н а и б о л е е в ы с о к о е с о д е р ж а н и е воды н а б л ю д а л о с ь в л и с т ь я х р а с т е н и й , и м е ю щ и х наиб о л ь ш у ю интенсивность транспир а ц и и {СаНит гШкетсит, ТапасеШт уи1§аге и, отчасти , 8ап§ш-зогЬа о^рстаНз) (рис . 2.2.3). В и д и м о и м е н н о п о э т о м у эти в и д ы при некотором у с и л е н и и почвенной засухи в конце вегетации по т е р я л и н а и б о л ь ш е е к о л и ч е с т в о (0.48-0.50 г,„аы •Гсухл.^ссы за пер ио д н а б л ю д е н и й ) в о д ы по сравн е н и ю с д р у г и м и в и д а м и (0-0.2 ^воды •^'^х.массы )• Т и п и ч н ы е Знач е н и я о в о д н е н н о с т и л и с т ь е в у

Глава 2. СРЕДНЯЯ ЧАСТЬ ГРАДИЕНТА 75
этих р а с т е н и й н а б л ю д а л и с ь в пределах 1.592-2.707 г,,^^^, •^'^.массы • В и д ы с более низким содержанием воды в листьях {Са1ата§гозНз ерщеюз, Сагех те1апоз1аскуа и Ьу(Игит у1г^а1ит) отличались и по размаху т и п и ч н ы х колебаний показателя (1.344-1.855 г^о^^, -т'ух.массы )• П о д в и ж н о с т ь содержания воды у всех р а с т е н и й была на низком уровне (У= 7.2-^13.5%).
Анализ флюктуации оводненности листьев в течение сезона показал , что у одних видов они реализуются в связи с динамикой и н т е н с и в н о с т и т р а н с п и р а ц и и {Са1ата-§говНз ерщею5, СаНит гШкетсит, 8ап§шзогЬа о^сгпаНз, с о о т в е т с т в у ю щ и е коэффициенты дет ерми н ац и и р а в н ы 0.724, 0.930 и 0.673). Д и н а м и к а содержания воды в листьях Ьу1кгит у1г§аШт с в я з а н а с д и н а м и к о й т е м п е р а т у р ы в о з д у х а (И = 0.774), а ТапасеШт уи1§аге — с и з м е н е н и я м и в л а ж н о с т и п о ч в ы (г^ = 0.702). Д л я Сагех те1апоБШскуа не удалось обнаружить статистически значимых связей с какими-либо из внешних или внутренних факторов .
Сравнительный анализ
Э к о л о г и ч е с к о е сходство в и д о в по с о в о к у п н о с т и показателей водного р е ж и м а (табл. 2.2.2). Н а п о м н и м , что виды, расположенные в верхней части м а т р и ц ы , характеризуются н а и м е н ь ш и м и значениями интенсивности т р а н с п и р а ц и и и оводненности и наибольшими значениями д е ф и ц и т а водного насыщения . В и д ы с противоположными свойствами водного р е ж и м а занимают н и ж н ю ю часть матрицы. Сочетание первой группы свойств характерно для более ксерофитных видов, сочетание последней группы характерно д л я более мезофитных растений. Д р у г и м и словами , в верхней части матрицы расположены относительно более ксерофитные виды, а в нижней — относи-
Таблица 2.2.2
М а т р и ц а экологического сходства по совокупности элементов водного р е ж и м а р а с т е н и й разнотравно-осокового с о о б щ е с т в а
Р
Сагех те1апо$1аскуа -0.357 -0.523 -1.345 -1.517 -2.160 -1.180
0.357 Са1ата^го8йз ер1§еюз -0.169 -0.892 -1.226 -1.702 -0.726
0.523 0.169 Ьу1кгит у1г^а1ит
-0.882 -1.112 -1.640 -0.588
1.345 0.892 0.882 8ап§и1зогЬа оЛГ1С1паН5 -0.250 -0.604 0.453
1.517 1.226 1.112 0.250 СаНит
пикетсит -0.343 0.752
2.160 1.702 1.640 0.604 0.343 ТапасеШт
уи1§аге 1.290
Примечание: — показатели экологического сходства между парами видов, Р — средний показатель экологического сходства каждого вида с остальными.

76 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ.
1.5
1.0 I 0,0 I -0,5
1-1,0
-1.5
=0.946
10 20 30 40 Парциальный объем меитлетников. %
Р и с . 2.2.4. С в я з ь м е ж д у п а р ц и а л ь н ы м и объемами м е ж к л е т н и к о в и индексами экологического сходства у видов разнотравно-
осокового сообщества .
тельно более м е з о ф и т н ы е по совокупности показателей водного обмена рас т е н и й . Э т о п о д т в е р ж д а е т с я в ы с о к о й степенью коррелированности Р и парциального объема м е ж к л е т н и к о в у видов данного сообщества (рис. 2.2.4; И = 0.946): в и д ы с б о л ь ш и м и з н а ч е н и я м и Р характеризуются менее плотным мезофиллом, и наоборот. К р о м е того, зас луживает внимания факт тесной статистической связи проективного п о к р ы тия видов и Р (рис. 2.2.5; И = 0.829). И з этого следует, что н а и б о л ь ш е е п р о ективное п о к р ы т и е и м е ю т относительно более м е з о ф и т н ы е растения .
В матрице экологического сходства отчетливо в ы д е л я ю т с я две группы вид о в : о т н о с и т е л ь н о более к с е р о ф и т н а я ' (Сагех те1апо81асНуа, Са1ата§тзНз ер1§еюз и Ьу(Нгит у1г§а(ит) и м е н е е к с е р о ф и т н а я {8ап^ш80гЬа 0^р1с1паИ8, СаНит гшИетсит и ТапасеШт уи1§а-ге) (табл. 2.2.2). Известно , что Са1ата-^гозНз ерщеюв, Сагех те1апозШскуа и Ьу1Игит угщаШт обычно произрастают в более у в л а ж н е н н ы х экотопах: на песч а н ы х откосах в о д о е м о в или на л у г а х с и з б ы т о ч н ы м у в л а ж н е н и е м . В о б о и х случаях растения этих видов могут периодически испытывать з а м е т н ы й водн ы й стресс в результате очень низкого или , напротив, очень высокого стояния г р у н т о в ы х вод . Э т о и могло п р и д а т ь этим видам относительно более ксероф и т н ы е ф у н к ц и о н а л ь н ы е ч е р т ы п о с р а в н е н и ю с в и д а м и в т о р о й г р у п п ы .
В противном случае придется признать, что, составляя з а м е т н у ю часть травостоя д а н ного сообщества , в и д ы первой группы развили эти черты в большей степени, чем более с в о й с т в е н н ы е д а н н о м у э к о т о п у Зап^шзогЬа о//1с1паН5, ОаНит гШкетсит и ТапасеШт уи1§аге. В результате м ы и м е е м достаточно у н и к а л ь н у ю с и т у а ц и ю , когда в сообществе , п р о и з р а с т а ю щ е м в средней части г р а д и е н т а в л а ж н о с т и п о ч в ы , д о м и нируют относительно более мезофитные по своим ф у н к ц и о н а л ь н ы м свойствам виды.
35
30
^ 25 I I 15 I 'О
5
О
-1,5 -1.0 -0.5 0.0 0.5 1.0 1,5 Индекс экологического сходства
Рис. 2.2.5. Связь между индексами экологического сходства и проективным покрытием видов р а з н о т р а в н о - о с о к о в о г о сообщества .
=0.829
' Еще раз подчеркнем, что в данном контексте определения ксерофитности или мезофитно-сти имеют относительный характер — растениям не присваивается соответствующий статус.

Глава 2. СРЕДНЯЯ ЧАСТЬ ГРАДИЕНТА 77
Это м о ж е т б ы т ь с л е д с т в и е м б о л е е в ы с о к о й к о н к у р е н т о с п о с о б н о с т и Зап^шзогЪа о^стаНз, СаНит гшНетсит и ТапасеШт уи1§аге по с р а в н е н и ю с Сагех теЬпозШ-скуа, СаЬта^гозНз ерщеюз и 1у1кгит у1г§аШт.
С л о ж н о с т ь о р г а н и з а ц и и водного р е ж и м а р а с т е н и й . О ч е в и д н о , что не все растения с п о с о б н ы к ф у н к ц и о н а л ь н о й регуляции своего водного р е ж и м а . В д а н н о м сообществе м и н и м а л ь н ы е индексы сложности организации водного р е ж и м а наблюдали у Сагех те1апоз1аскуа (0.559), Зап^шзогЬа о/ДстаНз (0.519) и ТапасеШт уи1§аге (0.560), а максимальные — у Са1ата§гозНз ерщеюз (0.881), СаНит Шкетсит (0.876) и Ьу1кгит у1г§аШт (0.750).
Слабо выраженная способность к функциональной регуляции может компенсироваться с т р у к т у р н ы м и п р и с п о с о б л е н и я м и к с н и ж е н и ю потерь воды р а с т е н и я м и при недостатке ее в почве, как у Сагех те1апозШскуа в данном сообществе . Растения этого вида могут эффективно сокращать т р а н с п и р и р у ю щ у ю поверхность , достаточно плотно смыкая п олови н ки листьев . В о з м о ж н ы варианты сочетания с т р у к т у р н о й и функциональной регуляции , как у Са1ата§гозНз ерщеюз, л и с т ь я которого с к р у ч и в а ю т с я при усилении засухи.
Морфолого-анатани ческая характеристика
Са1ата^гозНз ерщеюз — многолетнее травянистое растение с л и н е й н ы м и , довольно ш и р о к и м и л и с т ь я м и . М е з о ф и л л гомогенный, д о в о л ь н о р ы х л ы й . Т я ж и механиче ской ткани находятся с верхней и н и ж н е й сторон пучка ; и м е ю т с я д о в о л ь н о к р у п н ы е моторные клетки. Клетки верхней и нижней эпидермы одинаковы по размеру. Устьица некрупные.
Сагех те1апозШскуа — многолетник с л и н е й н ы м и , д о в о л ь н о у з кими л и с т ь я м и , состоящи ми из п ластинки и влагалища . М е з о ф и л л гомогенный с очень слабо выраж е н н ы м с л о е м п а л и с а д н ы х клеток . К л е т к и м е з о ф и л л а р а с п о л о ж е н ы о т н о с и т е л ь н о рыхло, имеются воздушные полости. Верхние клетки эпидермы заметно крупнее нижних. Тяжи механических тканей представлены сверху и снизу п р о в о д я щ и х пучков.
СаНит ги1кетсит и ТапасеШт ш1§аге — многолетние т р а в я н и с т ы е р аст ения . У ТапасеШт уи1§аге л и с т ь я п е р и с т ы е с л и н е й н ы м и з у б ч а т ы м и д о л я м и . У СаИит гШкетсит листья с л и с т о в и д н ы м и прилистниками л и н е й н ы е , н и т е в и д н ы е по 8-10 в мутовке . Х а р а к т е р н ы м анатомическим признаком этих р а с т е н и й является д о в о л ь н о р ы х л ы й м е з о ф и л л , и золатеральный у ТапасеШт уи1^аге и д о р з и в е н т р а л ь н ы й с п р и знаками изолатеральности у центрального пучка у СаНит Шкетсит. П а л и с а д н а я паренхима 1-2-слойная, клетки некрупные, вытянутой формы. Клетки верхней и нижней э п и д е р м ы у ТапасеШт уи1§аге одинаковые , а у ОаНит ги1кетсит верхние крупнее н и ж н и х , с н и ж н е й с т о р о н ы л и с т а п р и с у т с т в у ю т во ло ски . У ТапасеШт уи1§аге довольно крупные п о д у с т ь и ч н ы е полости .
Ьу1кгит угг^аШт, 8ап^13огЬа о^рсшаНз — многолетние т р авя нист ые растения с довольно крупными м е з о м о р ф н ы м и листьями, которые и м е ю т дорзивентральное строение с 1-2-слойной палисадной паренхимой. Клетки довольно крупные, вытянутой формы. Губчатая паренхима рыхлая , изодиаметрическая с относительно кр у пными межклетниками. Клетки верхней эпидермы значительно крупнее, чем нижней . Устьица не крупные, расположены с нижней стороны листа. Листья имеют довольно тонкую кутикулу.

78 Часть 1. Сезонная динамика водного режима доминантов и эдификаторов сообществ..
Таблица 2.2.3
П а р ц и а л ь н ы й объем тканей и свободных пространств л и с т ь е в растений разнотравно-осокового с о о б щ е с т в а
Виды Э Пр ПО Пг Пс П Мг Мс М Са1ата§гозйз ер1§ею$ 22.3 5.1 1.3 43.9 0 43.9 17.9 0 17.9 Сагех те1апозШсНуа 15.4 14.6 3.2 32.7 0 32,7 2.4 0 2.4 СаНит ги1кетсит 23.5 3.1 3.0 12.8 25.5 38.3 17.4 14.8 32.4 1у1кгит у1г§а1ит 27.5 1.2 4.2 14.1 30.6 44.7 11.8 10,6 22.4 8ап§ш50гЬа о]]1стаИз 21.7 3.9 5.6 14.7 26.6 41.3 17.6 9.9 27.5 ТапасеШт уи1§аге 11.3 2.9 5.1 14.1 27.2 41.2 11.0 28.4 39.4
Примечание: Э — эпидермис, Пр — проводящие ткани, ПО — паренхимная обкладка пуч-- ков, Пг — губчатая паренхима, Пс — столбчатая паренхима, П — паренхима, Мг — межклет
ники губчатой паренхимы, Мс - межклетники столбчатой паренхимы, М — межклетники.
Анализируя количественные соотношения тканей и свободных пространств в л и стьях и зуч ен н ых в и д о в (табл. 2.2.3), можно отметить л и ш ь о т д е л ь н ы е корреляции . Так, достаточно хорошо проявляется связь м е ж д у п а р ц и а л ь н ы м и о бъемами межклет ников и п р о в о д я щ и х тканей (рис. 2.2.6, а; И = 0.90). У видов с о т н о с и т е л ь н о больш и м количеством м е ж к л е т н и к о в проводящие ткани р а з в и т ы о т н о с и т е л ь н о м е н ь ш е . Кроме того, при у в е л и ч е н и и п а р ц и а л ь н ы х объемов п а р е н х и м ы о т н о с и т е л ь н о е у час тие проводящих тканей и обкладок пучков т а к ж е снижается (рис . 2.2.6, б; И = 0.726).
Для выявления количественных оценок сходства или различия видов по совокупности показателей анатомической структуры листьев п о с т р о е н а матрица анатомического сходства листьев р а с т е н и й (табл. 2.2.4).
о 10 20 30 40 30 35 40 45 Парциальный объем иежклвтникоа, % Парциальный объем паренхимы, %
Рис. 2.2.6. С о о т н о ш е н и е м е ж д у п а р ц и а л ь н ы м и о бъемами п р о в о д я щ и х тканей и м е ж клетников {а) и м е ж д у п а р ц и а л ь н ы м и о б ъ е м а м и п у ч к о в и п а р е н х и м ы (б) у в и д о в
разнотравно-осокового со о бщ ест ва .

Глава 2. СРЕДНЯЯ ЧАСТЬ ГРАДИЕНТА 79
Таблица 2.2.4
Матрица анатомического сходства по совокупности показателей анатомической структуры листьев растений разнотравно-осокового со о бщ ест ва
Р
Сагех те1апо51аскуа -1.964 -1.162 -1.523 -3.010 -3.130 -2.158
1.964 Са1ата^о8И5 ер1§ею8 -0.834 -1.271 -2.237 -2.898 -1.056
1.162 0.834 1у1кгит \1г^а1ит -0.637 -0.672 -2.098 -0.282
1.523 1.271 0.637 8ап§шзогЬа о/Т'сшаНз -0.331 -1.468 0.326
3.010 2.237 0.672 0.331 ОаНит Шкетсит -1.590 0.932
3.130 2.898 2.098 1.468 1.590 ТапасеШт ш1^аге 2.237
Относительно более ксероморфные в и д ы характеризуются большим объемом эпидермиса, склеренхимы, паренхимы, проводящей ткани, и м е н ь ш и м объемом межклетников . О н и р а с п о л о ж е н ы в в е р х н е й части табл . 2.2.4. Это Сагех те1апо81аскуа и Са1ата§го8Нз ерщеюз. О т н о с и т е л ь н о более м е з о м о р ф н о й структурой листьев обладают Ьу1кгит У1г§а(ит, Зап^шзогЬа о^стаНз, СаНит гШкетсит и ТапасеШт уи1§аге. Первые три из этих видов проявляют и заметное сходство по относительному составу тканей и м е ж к л е т н и к о в в листьях (табл. 2.2.4).
Сравнение индексов экологического и анатомического сходства показывает их полное с о в п а д е н и е (рис. 2.2.7; И = 0.936). Н е з а в и с и м ы е д а н н ы е по водному о б м е н у р а с т е н и й и анатомическому с т р о е н и ю их листьев одинаково отражают экологический статус видов относительно друг друга . О т н о с и т е л ь н о более м е з о м о р ф н а я с т р у к т у р а листьев ассоциируется с относительно более м е з о ф и т н ы м и ч е р т а м и водного обмена, и н аоборот Поэтому проективное покрытие растений коррелирует не т о л ь к о с и н д е к с а м и э к о л о г и ч е с к о г о (рис. 2.2.5), но и анатомического сходства (рис. 2.2.8, = 0.715). Эти результаты н е о д н о з н а ч н о у к а з ы в а ю т на то , что участие растений в сложении сооб-
0 -
-2
-3 = 0.936
-1,5 -1,0 -0.5 0.0 0.5 1.0 Индекс экологического сходства
1,5
Рис. 2.2.7. С о о т н о ш е н и е между индексами экологического и анатомического сходства у видов разнотравно-осокового сообщества.
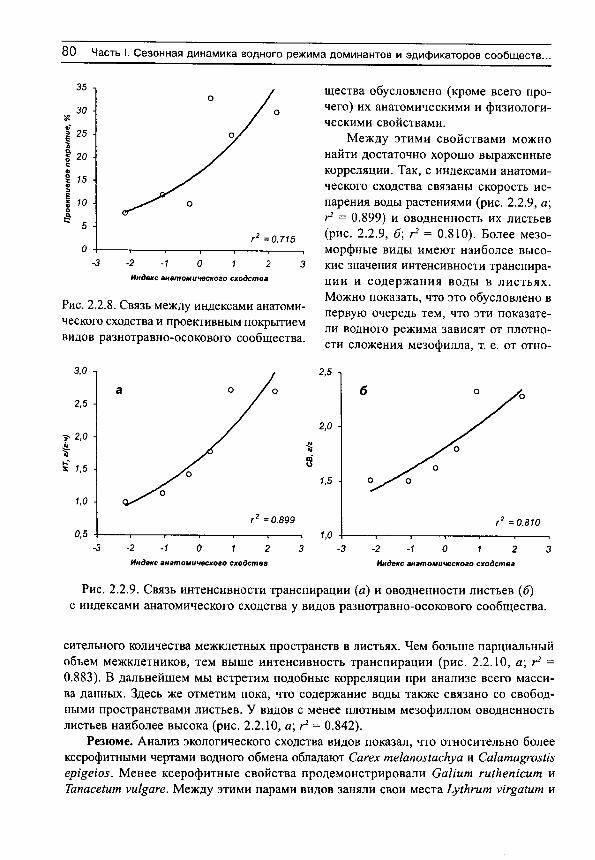
80 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ..
35 -,
30 -I 20 -I I *5 • 1 I 10
О
г' =0.715
- 3 - 2 - 1 0 1 2 3 Индекс анатомического сходства
Рис. 2.2.8. Связь между индексами анатомического сходства и проективным покрытием видов разнотравно-осокового сообщества .
3,0 -I
2,5
^ 2,0 ^
I 1,5 А
1,0
0,5
щества обусловлено (кроме всего пр о чего) их а н а т о м и ч е с к и м и и физиологич е с к и м и свойствами.
М е ж д у э т и м и с в о й с т в а м и м о ж н о найти достаточно х о р о ш о в ы р а ж е н н ы е корреляции. Так, с индексами анатомического сходства связаны скорость испарения воды растениями (рис. 2.2.9, а;
= 0.899) и о в о д н е н н о с т ь их листьев (рис . 2.2.9, б; 1^ = 0.810). Бо лее мезо м о р ф н ы е в и д ы и м е ю т н а и б о л е е в ы с о кие значения интенсивности транспирац и и и с о д е р ж а н и я в о д ы в л и с т ь я х . М о ж н о показать , что это обусловлено в первую очередь тем , что эти показатели водного р е ж и м а зависят от плотности сложения мезофилла , т. е. от отно-
2,5 п
2,0 -
8"
=0.899
1,5
1,0 - 3 - 2 - 1 0 1 2
Индекс анатомического сходства
=0.810
-2 -1 Индекс аиа
О 1 2 гичесхого сходства
Рис. 2.2.9. Связь интенсивности т р а н с п и р а ц и и (а) и о в о д н е н н о с т и л и с т ь е в (5) с индексами анатомического сходства у видов разнотравно-осокового со о бщ ест ва .
сительного количества м е ж к л е т н ы х пространств в листьях. Чем больше пар циаль ны й объем м е ж к л е т н и к о в , т е м в ы ш е и н т е н с и в н о с т ь т р а н с п и р а ц и и ( р и с . 2.2.10, а; Н = 0.883). В д а л ь н е й ш е м м ы в с т р е т и м подобные корреляции при анализ е всего массива дан н ых . Здесь же о т м е т и м пока, что содержание воды т а к ж е связано со свободными пространствами листьев . У видов с менее п л о т н ы м м е з о ф и л л о м о в о д н е н н о с т ь листьев наиболее высока (рис . 2.2.10, а; И = 0.842).
Резюме. А н а л и з экологического сходства видов показал , что о т но сит ель но более ксерофитными чертами водного обмена обладают Сагех те1апоз1асНуа и СаШта^озНз ерщеюз. М е н е е к с е р о ф и т н ы е с в о й с т в а п р о д е м о н с т р и р о в а л и СаНит гШкепюит и ТапасеШт уи1^аге. М е ж д у этими парами видов заняли свои места ЬугЬгит У1г§аШт и
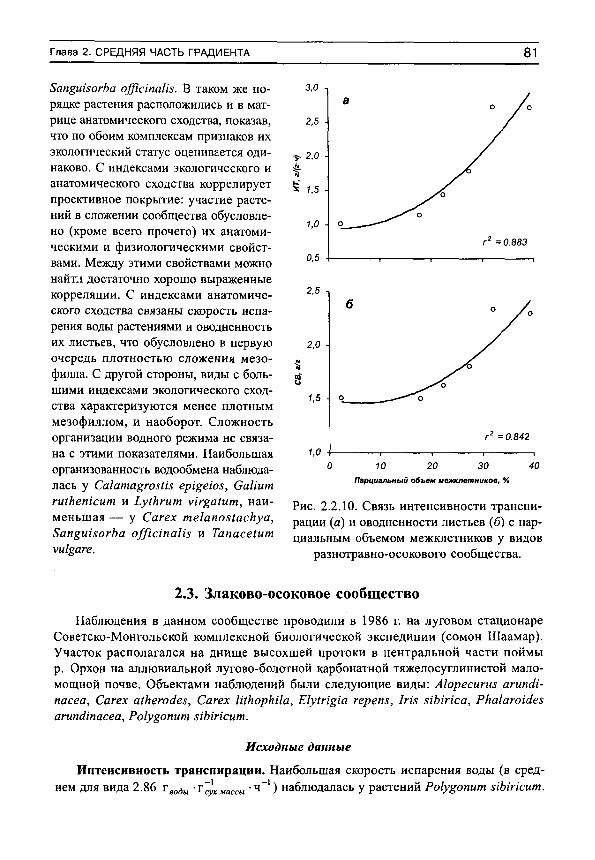
Глава 2. СРЕДНЯЯ ЧАСТЬ ГРАДИЕНТА 81
8ап§ш$огЬа о^гстаИз. В таком же по рядке растения расположились и в матрице анатомического сходства, показав, что по обоим комплексам признаков их экологический статус оценивается одинаково. С индексами экологического и анатомического сходства коррелирует проективное покрытие : участие растений в сложении сообщества обусловлено (кроме всего прочего) их анатомическими и ф и з и о л о г и ч е с к и м и свойствами. М е ж д у этими свойствами м о ж н о найти достаточно хорошо в ы р а ж е н н ы е корреляции . С и н д е к с а м и а н а т о м и ч е ского сходства связаны скорость испарения воды растениями и оводненность их листьев, что обусловлено в первую о ч е р е д ь п л о т н о с т ь ю с л о ж е н и я м е з о филла. С другой стороны, виды с больш и м и индексами экологического сходства х а р а к т е р и з у ю т с я менее п л о т н ы м м е з о ф и л л о м , и н аоборот . С л о ж н о с т ь организации водного р е ж и м а не связана с этими показателями. Наибольшая организованность водообмена наблюдалась у Са1ата^го5Н8 ерщеюз, ОаНит гШкетсит и Ьу1кгит угг^аШт, н а и м е н ь ш а я — у Сагех те1апоз1аскуа, 8ап^Ш80гЬа о^/гстаИз и ТапасеШт ш1^аге.
2,5
2,0
8" 1,5 -
1.0
г' = 0.842
10 20 30 40 Парциальный объем межклетников, %
Рис . 2.2.10. Связь и н т е н с и в н о с т и т р а н с п и рации (а) и оводненности листьев (б) с парц и а л ь н ы м объемом м е ж к л е т н и к о в у видов
разнотравно-осокового со о бщ ест ва .
2.3. Злаково-осоковое сообщество
Н а б л ю д е н и я в д а н н о м с о о б щ е с т в е проводили в 1986 г. на л у г о в о м с т а ц и о н а р е С о в е т с к о - М о н г о л ь с к о й к о м п л е к с н о й б и о л о г и ч е с к о й э к с п е д и ц и и ( с о м о н Ш а а м а р ) . У ч а с т о к р а с п о л а г а л с я н а д н и щ е в ы с о х ш е й п р о т о к и в ц е н т р а л ь н о й ч а с т и п о й м ы р . Орхон на а л л ю в и а л ь н о й лугово-болотной карбонатной т я ж е л о с у г л и н и с т о й малом о щ н о й почве . О б ъ е к т а м и н а б л ю д е н и й б ы л и с л е д у ю щ и е в и д ы : А1оресигш агипсИпасеа, Сагех а(кегос1ез, Сагех ШкоркИа, Е1у1гщ1а герепз, Мз згЫпса, РкаЫгоЫез агипсИпасеа, Ро1у§опит згЫпсит.
Исходные данные
И н т е н с и в н о с т ь т р а н с п и р а ц и и . Н а и б о л ь ш а я скорость и с п а р е н и я воды (в сред
нем для вида 2.86 т^^^^ - ^сух.массы " ч " ' ) наблюдалась у р а с т е н и й Ро1у§опит зЛтсит.

82 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ..
Таблица 2.3.1
Средние значения интенсивности транспирации (ИТ, г ^ ^ ^ ''''Сух.массы ' ) . д е ф и ц и т а водного насыщения (ДВН, % ) , содержания воды (СВ, г^„^^, •^~1х.массы) осмотического
давления клеточного сока (ОД, М П а ) видов злаково-осокового сообщества
Вид ИТ ДВН СВ ОД
Вид п X 8х п X 8х п X 8х п X 8х
А1оресигт агипсИпасеиз
280 1.697 0.595 280 1.2 2.1 280 2,394 0.393 36 1.61 0.32
Сагех а1кего<1е8 284 1.184 0.347 284 9.2 2.4 284 2.037 0.188 39 2.15 0.47 Сагех ШНорИИа 284 1.344 0.380 284 15.8 3.0 284 1.931 0.186 39 2.28 0.41 Е1у1гщ1а герепз 242 1.489 0.476 242 14.1 5.1 242 1.872 0.420 33 2.13 0.37 1п5 зШпса 284 1.285 0.553 284 14.5 4.9 284 2.767 0.362 39 2.03 0.31 РкаЬгоЫез агипсИпасеа
286 1.558 0.490 286 6.9 1.8 286 2.500 0.344 39 1.75 0.37
Ро1у§опит зШпсит
284 2.860 1.336 284 7.1 2.3 284 3.379 0.976 39 1.27 0.40
В целом для сообщества
1944 1.634 0.861 1944 10.6 4.9 1944 2.423 0.690 264 1.89 0.51
Она была значительно в ы щ е средней для участка интенсивности т р а н с п и р а ц и и (табл. 2.3.1). У о с т а л ь н ы х в и д о в средние значения показателя б ы л и д о с т а т о ч н о близкими , хотя м о ж н о отметить , что осоки и 1п5 зШпса испаряли воду с н а и м е н ь ш е й скоростью. Транспирация АЬресигиз агипсИпасеиз и Рка1аго1с1ез агипсИпасеа бьша несколько в ы ш е (рис . 2.3.1). С а м ы й н и з к и й т и п и ч н ы й м и н и м у м показателя о б н а р у ж е н у Мз зШпса (0.584 г^^^^ сух.массы " ч " ' ) , у остальных видов т и п и ч н ы е м и н и м у м ы находились на у р о в н е 0.76-1.14 г^д^^ - ^сух.массы • Т и п и ч н ы й м а к с и м у м т р а н с п и р а ц и и Ро1у§опит зШпсит был очень высок (4.57 г^о^^ • г~^^^д^^.„ • ч ~ ' ) , что значительно пре вышает найденные значения для других видов (1.61-2.50 г^о^ы '^~с/х.массы ч " ' ) . Вариация признака б ы л а достаточно умеренной и составляла 28-47%.
Сезонная динамика интенсивности транспирации у трех видов определяется полностью или ч а с т и ч н о факторами среды. Д л я Сагех ШкоркНа характерна з а в и с и м о с т ь транспирации от в л а ж н о с т и воздуха и почвы (Л^ = 0.831), для Мз зШпса — осве щенности , в л а ж н о с т и почвы , температуры воздуха (а т а к ж е о смо т ическо г о давления клеточного сока) (К^ = 0.933), для АЬресигиз агипсИпасеиз — от в л а ж н о с т и воздуха (а также от содержания воды в листьях) (Л^ = 0.850). У растений Сагех а{кегос1ез (г^ = 0.510), РНаЬгоШез агипсИпасеа (И = 0.514) и Ро1у§опит згЫггсит (И = 0.573) п р о с л е живается слабо выраженная связь сезонной д и н а м и к и скорости испарения в о д ы с из менениями оводненности листьев , а у растений Е1у1гщ1а герепз — с д е ф и ц и т о м водного н а с ы щ е н и я (г^ = 0.650).
Д е ф и ц и т водного н а с ы щ е н и я . В среднем для всех в и д о в д е ф и ц и т водного насыщения был достаточно низким — 10.6%о (табл. 2.3.1). Однако м о ж н о выделить виды с более в ы с о к и м и з н а ч е н и я м и п о к а з а т е л я {Сагех ШкоркИа, Е1у{гщ1а герепз, 1пз згЫпса), и с б о л е е н и з к и м и {А1оресигиз агипсИпасеиз, Рка1аго1с1ез агипйтасеа.
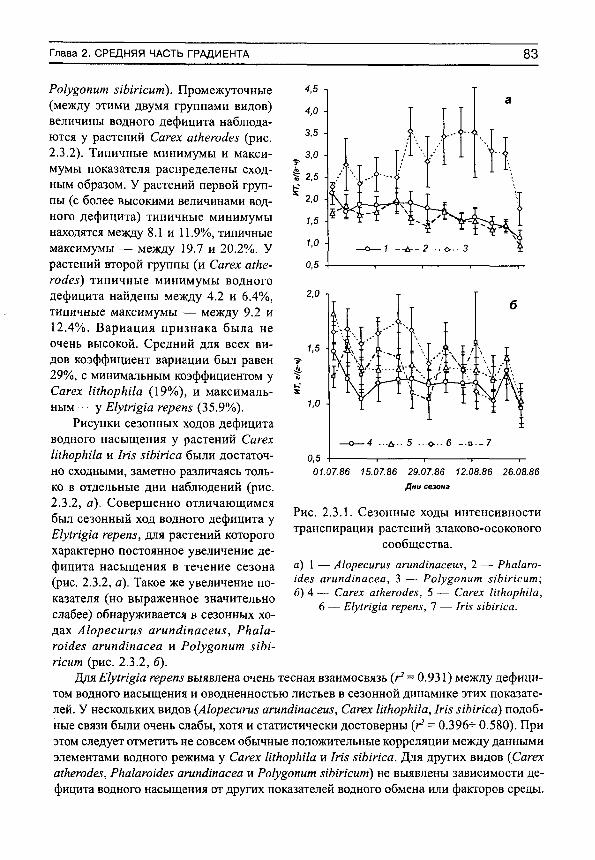
Глава 2. СРЕДНЯЯ ЧАСТЬ ГРАДИЕНТА 83
4,5
4,0
3,5
3,0 •5-
# 2 , 5
^'2,0
1,5
1,0
0,5
Ро1у§опит зШпсит). П р о м е ж у т о ч н ы е (между э т и м и двумя группами видов) величины водного д е ф и ц и т а наблюдаются у растений Сагех а1кего(1е5 (рис . 2.3.2). Т и п и ч н ы е м и н и м у м ы и максим у м ы показателя р а с п р е д е л е н ы сходным образом. У растений первой группы (с более высокими величинами водного д е ф и ц и т а ) т и п и ч н ы е м и н и м у м ы находятся между 8.1 и 11.9%, типичные максимумы — м е ж д у 19.7 и 20.2%. У растений второй г р у п п ы (и Сагех а1Не-гоЛез) т и п и ч н ы е м и н и м у м ы в о д н о г о д е ф и ц и т а н а й д е н ы м е ж д у 4.2 и 6.4%), т и п и ч н ы е м а к с и м у м ы — м е ж д у 9.2 и 12.4%о. В а р и а ц и я п р и з н а к а б ы л а не очень высокой . С р е д н и й д л я всех видов к о э ф ф и ц и е н т вариации б ы л равен 29%, с минимальным коэффициентом у Сагех ШЬорНИа (19%)), и м а к с и м а л ь ным — у Е1у(гщ1а герепз (35.9%)).
Рисунки се зонных ходов д е ф и ц и т а водного н а с ы щ е н и я у р а с т е н и й Сагех ШкоркИа и 1пз зШпса были достаточно сходными, заметно различаясь только в отдельные д н и н а б л ю д е н и й (рис . 2.3.2, а). С о в е р ш е н н о о т л и ч а ю щ и м с я был се зонный ход водного д е ф и ц и т а у Е1у(гщ1а герепз, для р а с т е н и й которого характерно постоянное увеличение де ф и ц и т а н а с ы щ е н и я в т е ч е н и е с е з о н а (рис. 2.3.2, а). Такое же увеличение показателя (но в ы р а ж е н н о е значительно слабее) обнаруживается в сезонных ход а х А1оресигиз агипсИпасеиз, Рка1а-гоИез агипйтасеа и Ро1у^опит зШпсит (рис. 2.3.2, б).
Для Е1у(п§1а герепз выявлена очень тесная взаимосвязь {г' = 0.931) м е ж д у д е ф и ц и том водного насыщения и оводненностью листьев в сезонной д и н а м и к е этих показателей. У нескольких видов (ЛЬресигиз агипсИпасеиз, Сагех ШкоркНа, Мз зШпса) подобные связи б ы л и очень слабы, хотя и статистически д о с т о в е р н ы (Н = 0.396^ 0.580). П р и этом следует отметить не совсем о б ы ч н ы е положительные корреляции м е ж д у д а н н ы м и элементами водного р е ж и м а у Сагех ШкоркИа и Мз згЫпса. Д л я д р у г и х видов {Сагех а(кегос1ез, РкаЬгоШез агипсИпасеа и Ро1у§опит з1Ыпсит) не выявлены зависимости дефицита водного насыщения от других показателей водного обмена или факторов среды.
01.07.86 15.07.86 29.07.86 Дни сезона
12.08.86 26.08.86
Рис . 2.3.1. С е з о н н ы е ходы и н т е н с и в н о с т и т р а н с п и р а ц и и р а с т е н и й злаково-осокового
сообщества .
а) 1 — АЬресигиз агипсИпасеш, 2 — РкаЬгоШез агипсИпасеа, 3 — Ро1у§опит згЫггсит; б) 4 — Сагех а1кегос1ез, 5 — Сагех И(коркИа,
6 — Е1у1п^1а герепз, 1 — Ыз зШпса.
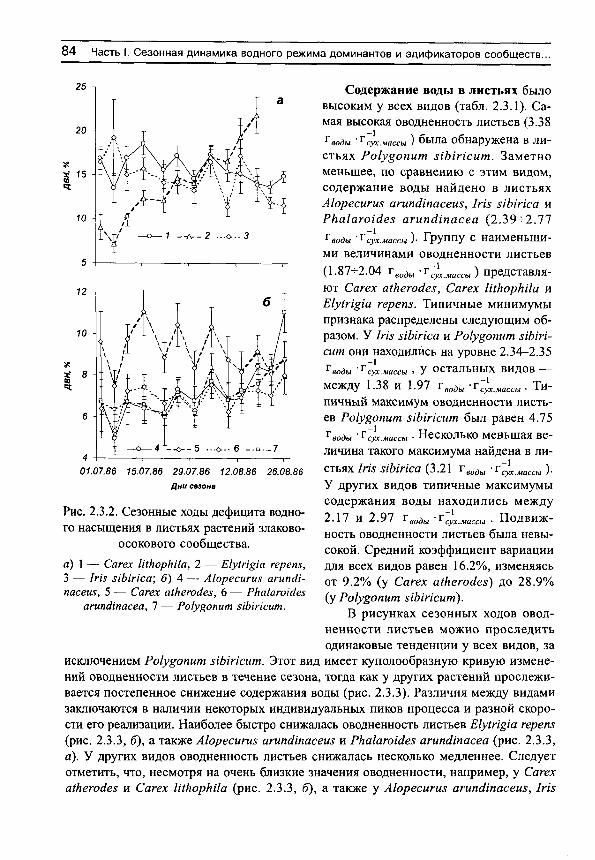
84 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ..
25 1
20 -
»5 1^5
10 -
С о д е р ж а н и е в о д ы в л и с т ь я х было в ы с о к и м у всех в и д о в (табл. 2.3.1). Са мая высокая оводненность листьев (3.38 ^воды сух.массы ) б ы л а обнаружсна в л и с т ь я х Ро1у§опит з1Ыг1сит. З а м е т н о м е н ь ш е е , по с р а в н е н и ю с этим видом, с о д е р ж а н и е в о д ы н а й д е н о в л и с т ь я х АЬресигш агипсИпасеш, / г к в1Ыпса и РНа1аго1йез агипсИпасеа (2.39-^2.77
^воды ' ^сух.массы
). Группу С н а и м е н ь ш и ми в е л и ч и н а м и о в о д н е н н о с т и л и с т ь е в
, - 1 (1.87^2.04 г , „ , „ т - , ) представля-
12
10
ю т Сагех а11гегос1ез, Сагех ШНоркИа и Е1у1п§1а герепз. Т и п и ч н ы е м и н и м у м ы признака р а с п р е д е л е н ы с л е д у ю щ и м образом. У Мз згЫпса и Ро1у§опит зШпсит они находились на уровне 2.34-2.35
воды ' ̂ сух.массы У о с т а л ь н ы х в и д о в — -1 сух.массы Ти-между 1.38 и 1.97 т^^^^, - г
п и ч н ы й максимум о в о д н е н н о с т и листь ев Ро1у§опит зШпсит б ы л р авен 4.75
_<^4 - 4 ^ - 5 ..-о-. 6 - . и . - 7
. . - 1 ' воды ^ сух.массы
01.07.86 15.07.86 29.07.86 12.08.86 Дни сезона
. Несколько м е н ь ш а я величина такого максиму ма найдена в ли-
„ - 1 26.08.86 стьях Мз з1Ыпса (3.21 г,„^„ т сух.массы
Рис. 2.3.2. Сезонные ходы дефицита водного насыщения в листьях растений злаково-
осокового сообщества .
)• У д р у г и х видов т и п и ч н ы е м а к с и м у м ы с о д е р ж а н и я в о д ы н а х о д и л и с ь м е ж д у 2.17 и 2.97 г,„й„ т -1
сух.массы П о д в и ж -
а) 1 — Сагех ШкоркИа, 2 — Е1у1гщ1а герепз, 3 — 1пз зШпса; б) Л — Л1оресигиз агипсИпасеиз, 5 — Сагех а1кегос1ез, 6 — Рка1агоШез
агипсИпасеа, 1 — Ро1у§опит зШпсит.
ность оводненности листьев была невысокой. С р е д н и й к о э ф ф и ц и е н т вар иации для всех в и д о в равен 16.2%, из меня я сь от 9.2%. (у Сагех а1кегос1ез) до 28.9%. (у Ро1у§опит зШпсит).
В р и с у н к а х с е з о н н ы х ходов о в о д н е н н о с т и л и с т ь е в м о ж н о п р о с л е д и т ь одинаковые т е н д е н ц и и у всех видов , за
исключением Ро1у§опит зШпсит. Этот вид имеет к у п о л о о б р а з н у ю к р и в у ю из мене ний оводненности л и с т ь е в в течение сезона , тогда как у д р у г и х р а с т е н и й п р о с л е ж и вается п о с т е п е н н о е с н и ж е н и е содержания воды (рис . 2.3.3). Р а з л и ч и я м е ж д у в и д а м и заключаются в н а л и ч и и некоторых индивидуальны х пиков п р о ц е с с а и разной скорости его реализации. Наиболее быстро снижалась оводненность листьев Е1у1гщ1а герепз (рис. 2.3.3, б), а т а к ж е АШресигиз агипсИпасеиз и РкаШгоШез агипсИпасеа (рис . 2.3.3, а). У других видов оводненность листьев с н и ж а л а с ь несколько медленнее . С л е д у е т отметить, что, несмотря на очень близкие значения оводненности , н а п р и м е р , у Сагех а1кегос1ез и Сагех ШкоркИа ( рис . 2.3.3, б), а т а к ж е у АШресигиз атпсИпасеиз, Мз
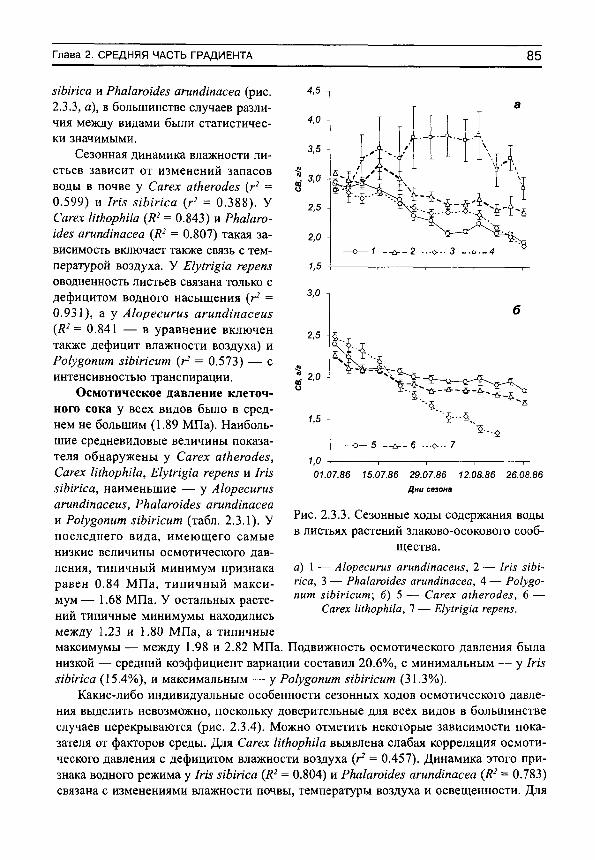
Глава 2. СРЕДНЯЯ ЧАСТЬ ГРАДИЕНТА 85
зШпса и РНаЬгоМез агипсИпасеа (рис. 2.3.3, а), в большинстве случаев различия между в и д а м и б ы л и статистически значимыми.
Сезонная динамика влажности листьев з а в и с и т от и з м е н е н и й з а п а с о в воды в почве у Сагех а1НегоИез (Н = 0.599) и Мз зШпса (Н = 0.388). У Сагех ШкоркИа (К' = 0.843) и РкаЬгоШез агипсИпасеа {К^ = 0.807) такая зависимость включает также связь с температурой воздуха. У Е1у1гщш герепз оводненность листьев связана только с д е ф и ц и т о м водного н а с ы щ е н и я (г^ = 0.931), а у АЬресигиз агипсИпасеиз {К^ = 0.841 — в у р а в н е н и е в к л ю ч е н также д е ф и ц и т в л а ж н о с т и воздуха) и Ро1у^опит зШпсит (г^ = 0.573) — с интенсивностью транспирации .
О с м о т и ч е с к о е д а в л е н и е клеточного сока у всех в и д о в б ы л о в среднем не большим (1.89 М П а ) . Наибольшие средневидовые в е л и ч и н ы показателя о б н а р у ж е н ы у Сагех а1кегос1ез, Сагех ШкорЫЬ, Е1у1гщш герепз и Мз зШпса, н а и м е н ь ш и е — у АЬресигиз атпсИпасеиз, РкаЬгоШез агипсИпасеа и Ро1у§опит зШпсит (табл. 2.3.1). У п о с л е д н е г о в и д а , и м е ю щ е г о с а м ы е низкие в е л и ч и н ы осмотического дав ления , т и п и ч н ы й м и н и м у м п р и з н а к а р а в е н 0.84 М П а , т и п и ч н ы й м а к с и мум — 1.68 М П а . У о с т а л ь н ы х растений т и п и ч н ы е м и н и м у м ы находились м е ж д у 1.23 и 1.80 М П а , а т и п и ч н ы е
м а к с и м у м ы — м е ж д у 1.98 и 2.82 М П а . П о д в и ж н о с т ь осмотического д а в л е н и я б ы л а низкой — средний к о э ф ф и ц и е н т вариации составил 20.6%, с м и н и м а л ь н ы м — у Мз зШпса (15.4%.), и м а к с и м а л ь н ы м — у Ро1у§опит зШпсит (31.3%.).
Какие-либо и н д и в и д у а л ь н ы е о с о б е н н о с т и се зонных ходов о с м о т и ч е с к о г о давления выделить н е в о з м о ж н о , поскольку доверительные д л я всех в и д о в в б о л ь ш и н с т в е случаев перекрываются (рис . 2.3.4). М о ж н о отметить некоторые з а в и с и м о с т и показателя от факторов среды. Д л я Сагех ШкорЫЬ в ы я в л е н а слабая корреляция о с м о т и ческого давления с д е ф и ц и т о м в л а ж н о с т и воздуха (г^ = 0.457). Д и н а м и к а этого признака водного р е ж и м а у Мз зШпса {К' = 0.804) и РкаЬгоШез атпс1тасеа (Я^ = 0.783) связана с изменениями влажности почвы, температуры воздуха и о с в е щ е н н о с т и . Д л я
15.07.86 29.07.86 Дни сезона
12.08.86 26.08.86
Рис. 2.3.3. Сезонные ходы содержания воды в листьях растений злаково-осокового сооб
щества .
а) 1 — АЬресигиз агипсИпасеиз, 2 — Мз згЫпса, 3 — РкаЬгоШез агипсИпасеа, 4 — Ро1у§о-пит згЫпсит; б) 5 — Сагех а1кегос1ез, 6 —
Сагех ШкорЫЬ, 7 — Е1у1п§1а герепз.
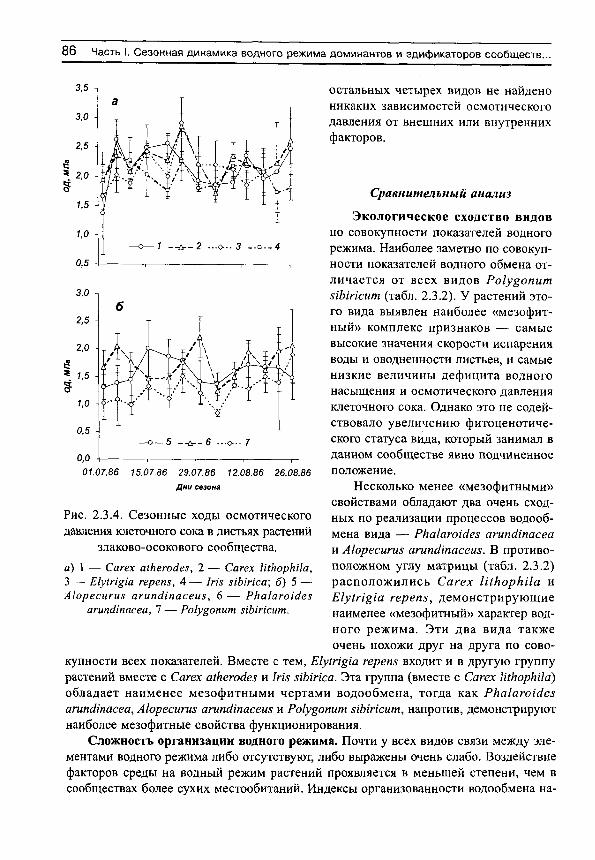
86 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ..
3,5 п
3,0 -
2,5 -
2.0 -
1,5
1.0
0,5
о с т а л ь н ы х четырех в и д о в не н а й д е н о никаких з а в и с и м о с т е й о смо т ическо г о давления от в н е ш н и х или в н у т р е н н и х факторов .
1 - - Л - - 2 . . .о . . 3 --а.-4
3,0 -,
2,5
2,0
1,5 -
1,0
0.5 --5 6 . - -0- . 7
0,0 01.07.86 15.07.86 12.08.86 26.08.86
Сравнительный анализ
Э к о л о г и ч е с к о е с х о д с т в о в и д о в по совокупности показателей водного режима. Наиболее заметно по совокупности показателей водного о бмена отл и ч а е т с я о т в с е х в и д о в Ро1у§опит зШпсит (табл. 2.3.2). У растений этого вида в ы я в л е н н а и б о л е е « м е з о ф и т -н ы й » к о м п л е к с п р и з н а к о в — с а м ы е высокие значения скорости испарения воды и оводненности листьев , и самые н и з к и е в е л и ч и н ы д е ф и ц и т а в о д н о г о н а с ы щ е н и я и о смо т ическо г о д а в л е н и я клеточного сока. Однако это не содейс т в о в а л о у в е л и ч е н и ю ф и т о ц е н о т и ч е -ского статуса вида, который занимал в д а н н о м с о о б щ е с т в е я в н о п о д ч и н е н н о е положение .
Несколько м е н е е « м е з о ф и т н ы м и » свойствами о б л а д а ю т два очень сходн ы х по р е а л и з а ц и и п р о ц е с с о в водообмена вида — РкаЬгоШез агипсИпасеа и АЬреситз атпсИпасеиз. В противоп о л о ж н о м углу м а т р и ц ы (табл . 2.3.2) р а с п о л о ж и л и с ь Сагех ИЛоркИа и Е1у(г1§1а герепз, д е м о н с т р и р у ю щ и е наименее « м е з о ф и т н ы й » характер водн о г о р е ж и м а . Э т и д в а в и д а т а к ж е очень похожи д р у г на друга по сово
купности всех показателей . В м е с т е с тем, Е1у1гщш герепз входит и в д р у г у ю г р у ппу растений вместе с Сагех а1кегос1ез и Мз зШпса. Эта группа (вместе с Сагех ШкорЫЬ) о б л а д а е т н а и м е н е е м е з о ф и т н ы м и ч е р т а м и в о д о о б м е н а , т о г д а к а к РкаЬго1с1ез агипсИпасеа, АЬреситз атпс1тасеиз и Ро1у§опит зШпсит, напротив, демонстрируют наиболее м е з о ф и т н ы е свойства функционирования .
С л о ж н о с т ь организации водного режима . Почти у всех видов связи м е ж д у элементами водного р е ж и м а либо отсутствуют, либо выражены очень слабо . Воздействие факторов среды на водный р е ж и м растений проявляется в м е н ь ш е й степени , чем в сообществах более сухих местообитаний . Индексы организованности водообмена на-
29.07.86 Дни сезона
Рис. 2.3.4. С е з о н н ы е ходы о с м о т и ч е с к о г о давления клеточного сока в листьях растений
злаково-осокового сообщества .
а) 1 — Сагех а1кегос1е5, 1 — Сагех ШкоркИа, 3 — Е1у1п§1а герепз, 4 — 1пз зШпса; б) 5 — АЬресигиз агипсИпасеиз, 6 — РкаЬго1с1ез
агипсИпасеа, 1 — Ро1у§опит зШпсит.

Глава 2. СРЕДНЯЯ ЧАСТЬ ГРАДИЕНТА 87
Таблица 2.3.2
Матрица экологического сходства по совокупности элементов водного р е ж и м а растений злаково-осокового с о о б щ е с т в а
Р
Сагех ШкоркИа -0.238 -0.710 -0.948 -1.869 -1.835 -2.335 -1.323
0.238 Е1у1гщ1а герепз -0.241 -0.525 -1.175 -1.238 -1.850 -0.798
0.710 0.241 Сагех а1кегос1ез -0.417 -1.141 -1.101 -1.633 -0.557
0.948 0.525 0.417 зШпса -0.663 -0.749 -1.610 -0.189
1.869 1.175 1.141 0.663 РкаЬгоШез агипсИпасеа -0.043 -0.912 0.649
1.835 1.238 1.101 0.749 0.043 А1оресип13 агипсИпасеиз -0.872 0.682
2.335 1.850 1.633 1.610 0.912 0.872 Ро1у§опит зЛтсит 1.535
Примечание: — показатели экологического сходства между парами видов, — средний показатель экологического сходства каждого вида с остальными.
ходятся между 0.287 (у Ро1у§опит зШтсит) и 0.630 (у 1ш зШтса). Средний для всех видов индекс равен 0.511, что может свидетельствовать о том , что в более в л а ж н ы х условиях п рои зраст ания сложность организации водного р е ж и м а р а с т е н и й снижает ся, т е. ослабляются связи между элементами , воздействие факт о р о в с р е д ы на водный обмен растений уменьшается . Это, на наш взгляд, является отражением того, что условия п р о и з р а с т а н и я п р и б л и ж а ю т с я к о п т и м а л ь н ы м . В т а к и х у с л о в и я х в о д н ы й стресс может возникать в течение сезона вегетации л и ш ь э п и з о д и ч е с к и , вода практически не является л и м и т и р у ю щ и м фактором. Поэтому сезонный ход показателей водообмена во м н о г и х случаях реализуется с л у ч а й н ы м образом возле некоторого , определяемого конституционными свойствами растений, уровня. Это приводит не только к ослаблению з а в и с и м о с т е й от факторов среды, но и к о п р е д е л е н н о й автономности элементов водного р е ж и м а друг от друга. В д а н н о м со о бщ ест ве м ы находим один из первых п р и м е р о в у м е н ь ш е н и я водного стресса и п р и б л и ж е н и я у с л о в и й произраста ния к о п т и м а л ь н ы м , в результате чего ф у н к ц и о н а л ь н а я с л о ж н о с т ь с и с т е м ы водного обмена снижается .
Резюме. По совокупности показателей водного р е ж и м а о т н о с и т е л ь н о н а и м е н е е « м е з о ф и т н ы м » комплексом этих показателей ( н и з к и м и з н а ч е н и я м и и н т е н с и в н о с т и транспирации и оводненности листьев , высокими в е л и ч и н а м и д е ф и ц и т а водного насыщения и осмотического давления клеточного сока) о б л а д а ю т д о м и н а н т ы злаково-осокового сообщества Сагех а(кегос1ез, Сагех ШкоркИа, а т акже Е1у(п§ш герепз и Мз

88 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ...
пЪМса. О т н о с и т е л ь н о более « м е з о ф и т н ы й » характер водного о бмена показал Ро1у-2опит зШпсит, который, впрочем, з анимает в с о о б щ е с т в е п о д ч и н е н н о е положение . Содоминанты АЬресигш агипсИпасеш и РНаЬгоЬез агипсИпасеа, о б л а д а ю щ и е очень сходным характером водообмена , з анимают промежу т о чно е положение м е ж д у этими двумя к р а й н и м и п о з и ц и я м и . Н и з к и е и н д е к с ы о р г а н и з о в а н н о с т и водного о б м е н а у большинства в и д о в натолкнули на предположение , что в д а н н о м с о о б щ е с т в е м ы находим один из п е р в ы х п р и м е р о в у м е н ь ш е н и я водного с т р е с с а и п р и б л и ж е н и я у с л о вий произрастания к о п т и м а л ь н ы м , в результате чего ф у н к ц и о н а л ь н а я с л о ж н о с т ь системы водного о б м е н а снижается .

Глава 3. ГУМИДНАЯ ЧАСТЬ ГРАДИЕНТА 89
ГЛАВА 3. ГУМИДНАЯ ЧАСТЬ ГРАДИЕНТА
3.1. Осоково-разнотравно-злаковое сообщество
Наблюдения в ковыльно-разнотравно-вострецовом с о о б щ е с т в е проводили в 1987 г. на луговом стационаре Советско-Монгольской комплексной б и о л о г и ч е с к о й экспедиции (сомон Ш а а м а р ) . Участок располагался в притеррасной части п о й м ы р . Орхон на аллювиальной луговой темноцветной карбонатной с р е д н е м о щ н о й тяжелосуглинистой на грубо п е с ч а н о м а л л ю в и и почве . О б ъ е к т а м и н а б л ю д е н и й б ы л и с л е д у ю щ и е виды: А§го$йз топ§коИса, Апетопе сИсНо1ота, Сагех а(кего(1ез, Роа ап§изН/оИа, ТкаИс1гит 8шр1ех, Уша сгасса. В литературе м о ж н о найти д о п о л н и т е л ь н у ю и н ф о р мацию по двум из перечисленных видов : А^гозИз топфоИса (Касьянова , Погодаева , 1979) и Уша сгасса (Алексеенко , 1976).
Исходные данные
Интенсивность транспирации. Средняя для всех видов этого сообщества величина скорости испарения воды была равна 1.63 г^ О Й М ''^~с1х.массы ' ч~' (табл. 3.1.1). Однако разброс средневидовых значений показателя возле этой величины был достаточно велик. Наименьшая интенсивность транспирации была обнаружена у растений Роа ап^изИ/оНа (0-80 г,„йь, •^с1х.массы - ч " ' ) , наибольшая — у Уша сгасса (2.25 г.^^ы •^~с''ух.массы-'^'^)-
В целом м о ж н о отметить , что в и д ы двудольных испаряли воду несколько более интенсивно, чем виды однодольных. Это проявилось и в т и п и ч н ы х максимумах признака . У Апетопе сИско(ота, ТНаНс1гит зтр1ех и Уша сгасса они находились м е ж д у 2 .80-3.46 г^о^ы •г~^^.^д^.^„ - ч " ' , у А^гозИз топ§коИса и Сагех а1кего(1ез б ы л и р а в н ы соответственно 1.96 и 1.94 г^о^ы '^~с1х.массы ' ч ~ ' , а у Роа ап§изИ/оНа т и п и ч н ы й максимум
Таблица 3.1.1
Средние значения интенсивности т р а н с п и р а ц и и (ИТ, г^д^^ '^'сух.массы " ч " ' ) , дефицита водного н а с ы щ е н и я ( Д В Н , % ) , содержания воды ( С В , г^^^^, - г'с^х.массы )>
осмотического давления клеточного сока (ОД, М П а ) в и д о в осоково-разнотравно-злакового сообщества
Вид ИТ ДВН СВ ОД
Вид п X 8х п X 8х п X 8х п X 8х
А^гозНз 239 1.446 0.386 239 11.5 2.6 239 2.734 0.302 30 1.63 0.47 топ§коНса Апетопе 240 2.112 0.773 240 14.4 3,7 240 3.212 0.460 30 2.15 0.47 сИско1ота Сагех а1кегос1е5 240 1.296 0.451 240 11.6 2.8 240 2.028 0.175 30 1.80 0.55 Роа ап^шИ/оИа 240 0.804 0.187 240 11.7 2.2 240 1.665 0.095 30 1.67 0.74 ТкаНс1гит зтр1ех 239 1.862 0.792 239 16.1 3.2 239 2.399 0.261 30 1.85 0.36 УШа сгасса 240 2.254 0.836 238 16.0 4.4 240 2.799 0.464 30 1.52 0.44
В целом для сообщества
1438 1.629 0.797 1436 13.5 3.8 1438 2.473 0.606 180 1.77 0.55

90 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ.
3,5
3,0 -
^2,5
! X 2,0
1.5
1.0
2.5
2.0
I « 1.5
1.0 •
б ы л „ - 1
р а в е н в с е г о 1.05 г сух.массы ^ • Т и п и ч н ы е м и н и м у
м ы показателя имели меньший разброс и находились (для всех видов) м е ж д у
^сух.массы
0.59 -1
1.26
^ - - • - 1 '
ч • . П о д в и ж н о с т ь и н т е н с и в н о с т и т р а н с п и р а ц и и б ы л а сравнительно невысокой. Средний к о э ф ф и ц и е н т в а р и а ц и и с о с т а в и л 33.5%, и з м е н я я с ь от 23.3%) (у Роа
ап^шП/оИа) д о 42.5%) (у ПаПс1-гит зтр1ех).
В р и с у н к а х с е з о н н ы х х о д о в и н т е н с и в н о с т и т р а н с п и р а ц и и вид о в д в у д о л ь н ы х (рис . 3 .1 .1 , а) и Сагех а1Негос1е5! ( р и с . 3 . 1 . 1 , б) прослеживается о б щ а я тенденция с н и ж е н и я с к о р о с т и и с п а р е н и я воды от начала к концу п е р и о д а н а б л ю д е н и й , н е с м о т р я на значит е л ь н ы е к о л е б а н и я п о к а з а т е л я . Д л я А^гозНз топ^коИса и Роа ап^изН/оИа (рис . 3 .1 .1 , б) характ е р н ы и з м е н е н и я п р о ц е с с а возле н е к о т о р ы х с р е д н и х у р о в н е й без заметного их снижения в течение сезона .
В ы я в л е н ы з а в и с и м о с т и с е з о н н о й д и н а м и к и и н т е н с и в н о с т и т р а н с п и р а ц и и от изменений освещенности и осмотического давления клеточного сока для Апетопе сИско1ота {К^ = 0 .648) , Роа ап^и-
зй/оНа {К^ = 0 .666) и ТкаНагит зтркх {К^ = 0.670). Э т и зависимости таковы, что при увеличении о с в е щ е н н о с т и скорость транспирации повышается , а при увеличении ос мотического давления — снижается . Д л я Сагех а1кегоЛез п р о я в и л а с ь прямая связь процесса с дефицитом влажности воздуха и н е д о н а с ы щ е н н о с т ь ю листьев водой {К^ = 0.885) . И н т е н с и в н о с т ь т р а н с п и р а ц и и Уша сгасса з ависела только от о в о д н е н н о с т и листьев ( И = 0 .465) , а А§гозИз топ§коИса — от скорости ветра {г' = 0 .534) .
Д е ф и ц и т водного н а с ы щ е н и я в листьях растений был сравнительно н е в ы с о к и м у всех видов — средневидовые величины показателя находились м е ж д у 11.5 и 16.1%о (табл. 3.1.1). Диапазон т и п и ч н ы х колебаний был также не очень ш и р о к и м . Т и п и ч н ы е м и н и м у м ы п р и з н а к а б ы л и на у р о в н е 8.1-12.1%), а т и п и ч н ы е м а к с и м у м ы — 1 4 . 6 -21.5%о. П о д в и ж н о с т ь водного д е ф и ц и т а оценивается к о э ф ф и ц и е н т о м вариации , р а в ным (в среднем) 23%, с м и н и м а л ь н ы м его значением у Роа ап§изИ/оИа (18 .5%) и
0,5 09.06.87 23.06.87 07.07.87 21.07.87 04.08.87 18.08.87
Дни сезона
Рис. 3.1.1. С езон н ые ходы интенсивности транспирации растений осоково-разнотравно-злакового
сообщества .
а) 1 — Апетопе сИско1ота, 2 — Тка1Шгит зтркх, 3 — Кгсш сгасса; б) 4 — А§гозИз топ§коИса, 5 —
Сагех а1кегос1ез, 6 — Роа ап^изИ/оНа.
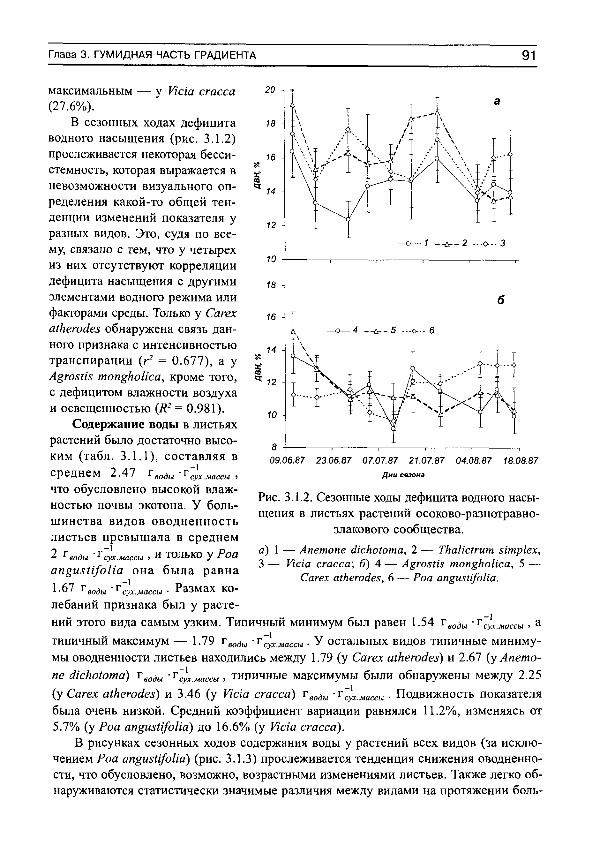
Глава 3. ГУМИДНАЯ ЧАСТЬ ГРАДИЕНТА 91
максимальным — у Уша сгасса (27.6%).
В се зонных ходах д е ф и ц и т а водного н а с ы щ е н и я (рис . 3.1.2) прослеживается некоторая бессистемность , которая выражается в невозможности визуального опр е д е л е н и я какой-то о б щ е й т е н денции и з м е н е н и й показателя у разных видов . Это , судя по всему, связано с тем, что у четырех из них отсутствуют к о р р е л я ц и и дефицита н а с ы щ е н и я с д р у г и м и элементами водного р е ж и м а или факторами среды. Только у Сагех аШегоёез о б н а р у ж е н а связь д а н ного признака с и н т е н с и в н о с т ь ю т р а н с п и р а ц и и (г^ = 0.677), а у А^гозШ топ^НоИса, кроме того , с д е ф и ц и т о м влажности воздуха и о с в е щ е н н о с т ь ю {К^ = 0.981).
Содержание воды в листьях растений было достаточно высок и м ( т а б л . 3.1.1), с о с т а в л я я в с р е д н е м 2.47 -т'^^^^^^ , что обусловлено в ы с о к о й влажн о с т ь ю почвы экотопа . У б о л ь -щ и н с т в а в и д о в о в о д н е н н о с т ь л и с т ь е в п р е в ы ш а л а в с р е д н е м 2 Г в о й ы •^'^.массы , и только у Р О Й
ап§изИ/оИа о н а б ы л а р а в н а _-1
1-67 г , „ й „ т сух.массы Р а з м а х ко
го п
18
16
14 -
12
10
18
16 -
1 -^-2 ...о.. 3
14 -
12
10
-4 5 . .-о-. 6
8 09.06.87 23.06.87 07.07.87 21.07.87
Дни сезона 04.08.87 18.08.87
Рис. 3.1.2. Сезонные ходы дефицита водного насыщения в листьях растений осоково-разнотравно-
злакового сообщества .
а) 1 — Апетопе (ЛсНо1ота, 2 — ТкаНс1гит зтр1ех, 3 — У1с1а сгасса; б) А — Л^гозНз топ^коИса, 5 —
Сагех агкегоёез, 6 — Роа ап§из11/оНа.
л е б а н и й п р и з н а к а б ы л у р а с т е
ний этого вида с а м ы м узким. Т и п и ч н ы й м и н и м у м был равен 1.54 г„„^„ - ^сух.массы, а „ - 1
т и п и ч н ы й максимум — 1.79 г^^^^ - г -1 сух.массы . У о с т а л ь н ы х в и д о в т и п и ч н ы е м и н и м у
мы оводненности листьев находились между 1.79 (у Сагех а1кегос1ез) и 2.67 (у Апето-, - 1
пе сИсИо1ота) ^еоды ''^сух.массы > т и п и ч н ы е м а к с и м у м ы были о б н а р у ж е н ы м е ж д у 2.25 (у Сагех а1кегоИез) и 3.46 (у Исга сгасса) ^^оды '^сух.массы • П о д в и ж н о с т ь показателя была очень низкой. С р е д н и й коэффициент вариации р а в н я л с я 11.2%о, и з м е н я я с ь от 5.7% (у Роа ап§изИ/о1га) д о 16.6%) (у Уша сгасса).
В рисунках с е з о н н ы х ходов содержания воды у р а с т е н и й всех в и д о в (за и с к л ю чением Роа ап^шН/оИа) (рис. 3.1.3) прослеживается тенденция с н и ж е н и я оводненности, что обусловлено , возможно, возрастными изменениями листьев . Также легко об наруживаются статистически значимые различия м е ж д у видами н а протяжении боль-
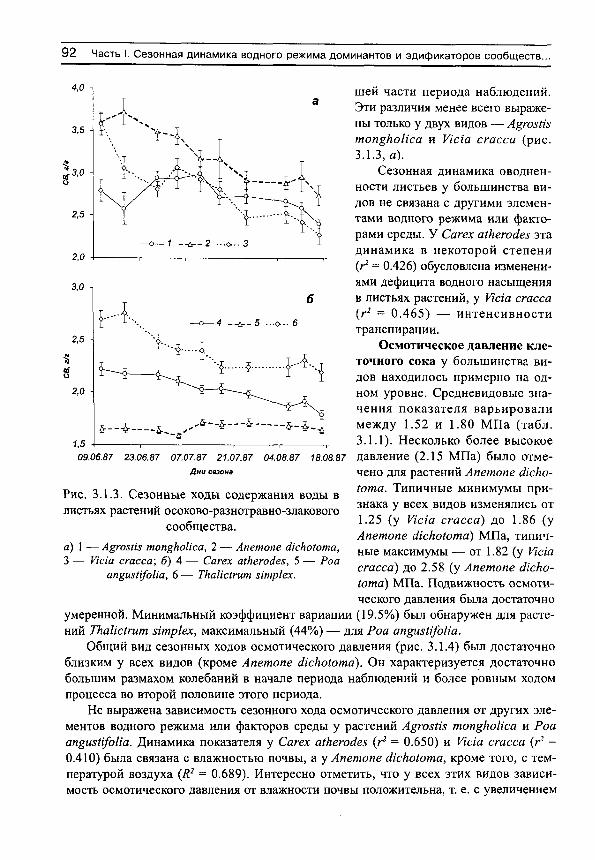
92 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ.
4,0 -,
3,5 -
3,0
2,5 -
2,0
3,0 -I
2,5
8" 2,0
1,5
щ е й ч а с т и п е р и о д а н а б л ю д е н и й . Эти различия менее всего выражены только у двух видов — А§гозШ топфоИса и УЫа сгасса ( р и с . 3.1.3, а).
С е з о н н а я д и н а м и к а о в о д н е н ности л и с т ь е в у б о л ь ш и н с т в а видов не связана с д р у г и м и э л е м е н т а м и водного р е ж и м а или фактор а м и среды. У Сагех аЛегоИез эта д и н а м и к а в н е к о т о р о й с т е п е н и (г^ = 0.426) обусловлена изменениями д е ф и ц и т а водного насы щ ения в листьях растений , у Уша сгасса {г^ = 0 .465) — и н т е н с и в н о с т и т р а н с п и р а ц и и .
О с м о т и ч е с к о е д а в л е н и е клеточного сока у б о л ь ш и н с т в а вид о в н а х о д и л о с ь п р и м е р н о на одном у р о в н е . С р е д н е в и д о в ы е знач е н и я п о к а з а т е л я в а р ь и р о в а л и м е ж д у 1.52 и 1.80 М П а ( т а б л . 3.1.1). Н е с к о л ь к о б о л е е в ы с о к о е д а в л е н и е (2.15 М П а ) б ы л о отмечено для растений Апетопе сИскоЮта. Т и п и ч н ы е м и н и м у м ы п р и знака у всех видов из меня лись от 1.25 (у УЫа сгасса) д о 1.86 (у Апетопе сИско(ота) М П а , т и п и ч ные максимумы — от 1.82 (у Исга сгасса) до 2.58 (у Апетопе сИскоЮта) М П а . П о д в и ж н о с т ь о с м о т и ческого давления бьша достаточно
умеренной. М и н и м а л ь н ы й коэффициент вариации (19.5%) был о бнар у жен для расте ний ТкаИс1тт ятр1ех, максимальный (44%) — для Роа ап§и$И/оИа.
О б щ и й вид с е з о н н ы х ходов осмотического давления (рис . 3.1.4) б ы л до ст ат о чно близким у всех видов (кроме Апетопе (ИскоЮта). О н х ар акт ер из у ет ся д о с т а т о ч н о б о л ь ш и м размахом колебаний в начале периода наблюдений и более р о в н ы м ходом процесса во второй половине этого периода .
Не выражена зависимость сезонного хода осмотического д а в л е н и я от др у г их элементов водного р е ж и м а или факторов среды у р а с т е н и й А^гозИз топ§коНса и Роа ап^шИ/оНа. Д и н а м и к а показателя у Сагех а1кегос1е$ {г^ = 0.650) и Угсга сгасса (г^ = 0.410) б ы л а связана с в л а ж н о с т ь ю почвы, а у Апетопе (ИскоЮта, кроме того , с т е м пературой воздуха = 0.689). И н т е р е с н о отметить , что у всех этих в и д о в з ависимость осмотического давления от влажности почвы положительна, т. е. с увеличением
09.06.87 23.06.87 07.07.87 21.07.87 Дни сезона
04.08.87 18.08.87
Рис . 3.1.3. С е з о н н ы е х о д ы с о д е р ж а н и я в о д ы в листьях растений осоково-разнотравно-злакового
сообщества .
а) 1 — А^гозИз топ^коИса, 1 — Апетопе (ИскоЮта, 3 — Уша сгасса; б) 4 — Сагех а(кего(1ез, 5 — Роа
ап§изИ/ока, 6 — ТкаИс1гит 51тр1ех.
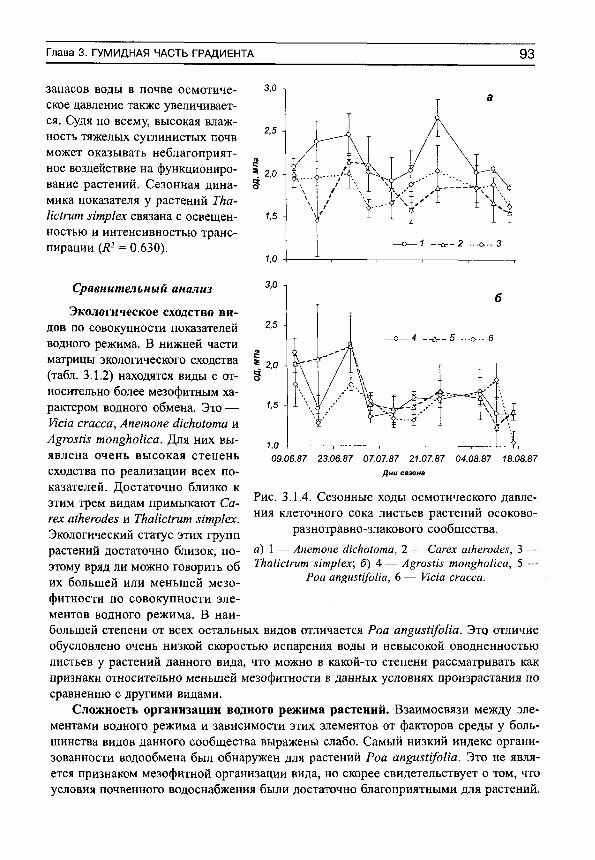
Глава 3. ГУМИДНАЯ ЧАСТЬ ГРАДИЕНТА 93
запасов воды в почве о с м о т и ч е ское давление также увеличивается. Судя по всему, высокая влажность тяжелых суглинистых почв м о ж е т о к а з ы в а т ь н е б л а г о п р и я т ное воздействие на функционирование р а с т е н и й . С е з о н н а я дина мика показателя у растений ТНа-Ис1гит 5тр1ех связана с освещенностью и и н т е н с и в н о с т ь ю транс пирации {К^ = 0.630).
3,0 .
2,5 -
* 2,0
1,5
1,0 1 - ^ - 2 . . .о-. 3
4 - -Д--5 . . .0- . 6
09.06.87 23.06.87 07.0787 210787 Дни сезона
04.08.87 18.08.87
Сравнительный анализ
Экологическое сходство в и дов по совокупности показателей водного режима. В нижней части матрицы экологического сходства (табл. 3.1.2) находятся виды с относительно более мезофитным ха-раюгером водного обмена. Это — ИЫа сгасса, Апетопе (ИскоЮта и А^гозНз топфоИса. Для них выя в л е н а о ч е н ь в ы с о к а я с т е п е н ь сходства по р е а л и з а ц и и всех показателей . Д о с т а т о ч н о близко к этим трем в и д а м п р и м ы к а ю т Сагех агкего(1е8 и ТкаИс1гит зтр1ех. Экологический статус этих групп растений достаточно близок, по этому вряд ли м о ж н о говорить об их б о л ь ш е й или м е н ь ш е й м е з о -ф и т н о с т и по с о в о к у п н о с т и э л е ментов водного р е ж и м а . В н а и большей степени от всех о с т а л ь н ы х видов отличается Роа ап§шИ/оНа. Это отличие обусловлено очень низкой с к о р о с т ь ю испарения воды и невысокой о в о д н е н н о с т ь ю листьев у растений д а н н о г о вида , что м о ж н о в какой-то степени рассматривать как признаки относительно м е н ь ш е й мезофитности в д а н н ы х условиях произрастания по сравнению с другими видами .
С л о ж н о с т ь о р г а н и з а ц и и водного р е ж и м а р а с т е н и й . В з а и м о с в я з и м е ж д у элементами водного р е ж и м а и з ависимости этих элементов от факторов с р е д ы у большинства видов д а н н о г о с о о б щ е с т в а в ы р а ж е н ы слабо. С а м ы й низкий индекс организованности водообмена б ы л обнаружен для растений Роа ап§шИ/оНа. Это не является признаком м е з о ф и т н о й организации вида, но скорее свидетельствует о том, что условия почвенного водоснабжения были достаточно б л а г о п р и я т н ы м и д л я растений .
Рис . 3.1.4. С е з о н н ы е ходы осмотического давления клеточного сока л и с т ь е в р а с т е н и й осоково-
разнотравно-злакового сообщества .
а) 1 — Апетопе (Иско1ота, 2 — Сагех а1кегос1е5, 3 — ТкаИс1гит 8тр1ех; 6) А — А^гозИв топфоИса, 5 —
Роа ап^изИ/оИа, 6 — Уша сгасса.

94 Часть I. Сезонная динамика водного режима доминантов и эдификаторов сообществ...
Таблица 3.1.2
М а т р и ц а экологического сходства по совокупности элементов водного р е ж и м а р а с т е н и й осоково-разнотравно-злакового с о о б щ е с т в а
Р
Роа ап§шИ/оИа -0.961 -0.915 -1.197 -1.326 -1.760 -1.232
0.961 Сагех а1Иегос1ез -0.238 -0.749 -0.788 -0.896 -0.342
0.915 0.238 ТкаИс1гит зтр1ех -0.597 -0.567 -0.654 -0.133
1.197 0.749 0.597 сгасса -0.069 -0.101 0.475
1.326 0.788 0.567 0.069 Апетопе (ИсНо1ота -0.074 0.535
1.76 0.896 0.654 0.101 0.074 Л^говИз топ§НоИса 0.697
Примечание: Р̂-̂- — показатели экологического сходства между парами видов, Р — средний показатель экологического сходства каждого вида с остальными.
Н е с м о т р я на это они д е м о н с т р и р о в а л и черты, ко нст ит у цио нно с в о й с т в е н н ы е ксерофитам — очень н и з к у ю и н т е н с и в н о с т ь т р а н с п и р а ц и и и с р а в н и т е л ь н о н е в ы с о к у ю оводненность листьев . М о ж н о предположить , по аналогии с близ кими в и д а м и , что у Роа ап^шИ/оИа очень п л о т н ы й м е з о ф и л л (с м а л ы м о бъемо м м е ж к л е т н и к о в ) и при л ю б ы х условиях почвенного водоснабжения р а с т е н и я не с м о г у т п р е в ы с и т ь д о с т у п ные им в е л и ч и н ы указанных элементов водного режима .
У А^ГОБИЗ топфоИса, Апетопе (ИсНо1ота, ТЬаИс1гит 5тр1ех и Исга сгасса эти индексы находились м е ж д у 0.325-0.379. Достаточно вы со кий индекс организованности (0.660) б ы л обнаружен только для растений Сагех а1Негос1ез, что , вероятно , м о ж но рассматривать как признак недостаточно благоприятных у с л о в и й п р о и з р а с т а н и я для этого вида в д а н н о м экотопе. Однако средний для всех и с с л е д о в а н н ы х видов индекс был равен 0.367. Это значительно ниже, чем в с о о б щ е с т в а х с м е н ь ш и м и запасами воды в почве . О ч е в и д н о , что, в общем, усло вия п р о и з р а с т а н и я р а с т е н и й о с о ково-разнотравно-злакового сообщества м о ж н о рассматривать как до ст ат о чно благоприятные . Некоторые п р и з н а к и указывают на т о , что не с н и ж е н и е , а у в е л и ч е н и е запасов воды в почве экотопа вызывают реакции (в о сно вно м о с м о т и ч е с к о г о давления клеточного сока) , в какой-то степени сходные с р е а к ц и я м и на водный ст р есс . Одна ко избыточное увлажнение тяжелосуглинистой почвы местообитания не было ни чрезмерным, ни д л и т е л ь н ы м и заметных р е а к ц и й ф у н к ц и о н и р о в а н и я н а с т р е с с обнаружено не было .
Резюме. Условия произрастания осоково-разнотравно-злакового сообщества б ы л и достаточно б л а г о п р и я т н ы м и для большинства видов . У всех видо в , за и с к л ю ч е н и е м

Глава 3. Г У М И Д Н А Я Ч А С Т Ь Г Р А Д И Е Н Т А 95
Роа ап§шИ/оНа обнаружены высокие значения интенсивности транспирации и оводненности листьев. Все виды характеризуются низкими величинами дефицита водного насыщения и осмотического давления клеточного сока. Относительно более мезо-фитный характер водного обмена проявляют К/сш сгасса, Апетопе сИско1ота и А^гозНз топ^ИоИса. Достаточно близко к этим трем видам примыкают Сагех а1кегос1ез и ТкаНс1гит зтр1ех. В наибольшей степени от всех остальных растений отличается Роа ап^шИ/оИа. Средний для всех исследованных видов индекс организованности водообмена был равен 0.367. Это значительно ниже, чем в экотопах с меньшими запасами воды в почве, что также является признаком адекватности условий произрастания для изученных в этом сообществе растений.
3.2. Крупноразнотравно-дернистоосоковое сообщество
Наблюдения в крупноразнотравно-дернистоосоковом сообществе проводили в 1995 г. Это сообщество расположено в притеррасной части поймы р. Хопер. Произрастающие здесь фитоценозы относятся к группе типов долгопоемных лугов и занимают экотопы с отметкой 270-290 см над меженью р. Хопер (Титов, Печенюк, 1990). Год наблюдений был достаточно засушливым: выпало лишь небольшое количество осадков (около 80 мм) за 3 летних месяца, а дефицит влажности воздуха достигал в дневные часы 40 мб. Средний за период наших наблюдений запас влаги в верхнем (0-30 см) горизонте почвы составлял 111.5 мм. Почва здесь иловато-глинистая, мощность гумусового горизонта 80-100 см, ниже расположен глеевый горизонт с линзами песка. Травостой густой. Общее проективное покрытие составляет 90-100%. Высота травостоя 100-170 см. Если ранее в данном сообществе было отмечено доминирование 5ап§шзогЬа о^Г^с^па^^з, Ьа1куп15 рга1епз13 Ь., РШрепс1и1а иШаИа, А1ореситз рга1епз1з Ь., Сагех ргаесох 8сЬгеЬ., Е1у1п§1а герет, СаИит ркузосагрит ЬейеЬ., Роа (гтаНз Ь. (Титов, Печенюк, 1990), то в год наших наблюдений основу травостоя составлял Сагех саезрКоза, образующий крупные (до 50-70 см в диаметре) кочки. На долю этого вида приходилось до 50% проективного покрытия. Содоминантами были виды, занимавшие межкочечное пространство: РШрепс1и1а иЫапа (15-20%), Уакпапа УV0^§епз^з (20-25%), 8ап§ш50гЬа о^/гстаИз (10%), СаИит ркузосагрит (10%>), а также Уеготса 1оп§1/оИа и Мз рзеийасогиз. Последний вид, хотя и не имел заметного проективного покрытия, образовывал мощные дернины, имел высокие (до 110-130 см) побеги и оказывал заметное влияние на структуру травостоя. Поэтому объектами своих исследований в данном сообществе мы выбрали Сагех саезрИоза, РШрепс1и1а и1тапа, Мз рзеи^асогиз, 8ап§шзогЬа о^гсгпаИз, Уа1епапа м>о1§епз1з и Уеготса 1оп^1/оИа.
Исходные данные
Интенсивность транспирации. У исследованных видов ИТ была, против ожидания, не очень высока. Исключение составляли 8ап§шзогЬа оЛГ1стаНз и Уеготса 1оп§1/оНа, которые в среднем испаряли воду с достаточно высокой скоростью — 1.854 и 2.056 Гдойу •гё^!x.^ассы ' Ч " ' соответственно (табл. 3.2.1). Наименьшую среднюю величину ИТ наблюдали у видов, занимающих ведущее положение в крупноразнотрав-

96 Ч а с т ь I. С е з о н н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в .
Таблица 3.2. ]
Средние значения интенсивности транспирации (ИТ, г^^^^ ' ̂ ёух.массы ' ч~ ' ) , дефицита водного насыщения (ДВН, % ) , содержания воды в листьях (СБ, г^^^^ •^'^ух.массы )
у растений крупноразнотравно-дернистоосокового сообщества
Показатели Виды ИТ ДВН СБ
п X п X 8х п X 5х
Сагех саезрИош 224 1.339 0.471 224 23.4 5.6 224 1.979 0.230 Ртрепс1и1а и1тапа 224 1.436 0.535 224 29.1 7.9 224 1.642 0.236 1п5 рзеи<1асоп15 226 1.443 0.455 226 23.4 10.4 226 2.007 0.446 5ап§ш50гЬа оЛГ1с1паИз 228 2.056 0.750 228 11.8 3.9 228 2.092 0.277 Уакпапа м>о1§ет15 228 1.222 0.450 226 15.8 3.8 228 3.721 0.495 Уеготса Ьп^г/оНа 226 1.854 0.680 226 18.2 4.2 226 2.390 0.293 В целом для сообщества 1356 1.559 0.641 1354 20.3 8.6 1356 2.458 0.770
но-дернистоосоковом сообществе: Сагех саезрИоза (1.339 г^о^ы '^сух.мтсы " ч ' ' ) и Уакпапа у/о1§епз13 (1.222 т^^^^ -^'сух.массы "ч" ')- Типичные минимумы признака у этих двух видов были равны соответственно 1.25 и 1.12 г^^^^ '^'с^х.мжсы " ч " ' , а типичные максимумы — 3.0 и 2.64 т^^^^ •^~с1х.массы - ч " ' . У остальных видов типичные минимумы находились на уровне 0.75-0.89 т^^^^ • ^сух.массы - ч " ' , а типичные максимумы — в пределах 1.73-2.20 Г^^ЙЫ '^'сух.массы ' ч ' • Подвижность транспирации была достаточно умеренной — коэффициенты вариации изменялись от 31.6 (у Мз рзеис1асотз) до 37.2% (у ртреп(Ли1а иЫапа).
В сезонных ходах интенсивности транспирации (рис. 3.2.1) трудно уловить общие для всех видов закономерности. Сезонная динамика показателя у Сагех саезрИоза в небольшой степени определяется дефицитом влажности воздуха (Н = 0.533). Для РШрепёи1а и1тапа обнаружена не очень сильная связь процесса с изменениями влажности почвы (у = 0.516). Растения Мз (ИсНо1ота реагируют на динамику освещенности и температуры воздуха (Л^ = 0.881), что характерно и для Уеготса 1оп§{/оНа (К^ = 0.782) с той разницей, что транспирация растений этого вида определяется также и оводненностью листьев. Транспирация Уа1епапа У[>о1§епз13 взаимосвязана только с дефицитом водного насыщения листьев (И = 0.598), а транспирация 8ап§шзогЬа оЛГгстаНз не зависит от других элементов водного режима или факторов среды.
Дефицит водного насыщения в листьях растений составлял в среднем 20.3%). Наибольшие средние величины этого показателя были у РШрепс1и1а иЫапа (29.1%), Сагех саезрИоза и Мз рзеийасогиз (23.4%) (табл. 3.2.1). Минимальная величина наблюдалась у Зап^тзогЬа о/^1стаНз (11.8%). Уа1епапа \уо1§епз13 и Уеготса 1оп^1/оИа по средним значениям водного дефицита занимали промежуточное положение. У первых трех видов обнаружен наибольший разброс значений дефицита водного насыщения. Границы типичных колебаний признака у Сагех саезрИоза находились в пределах 16.5-30.7 , у РШрепМа иЫапа — 19.1-39.8%, у Мз рзеис1асотз — 11.8-39.7%.
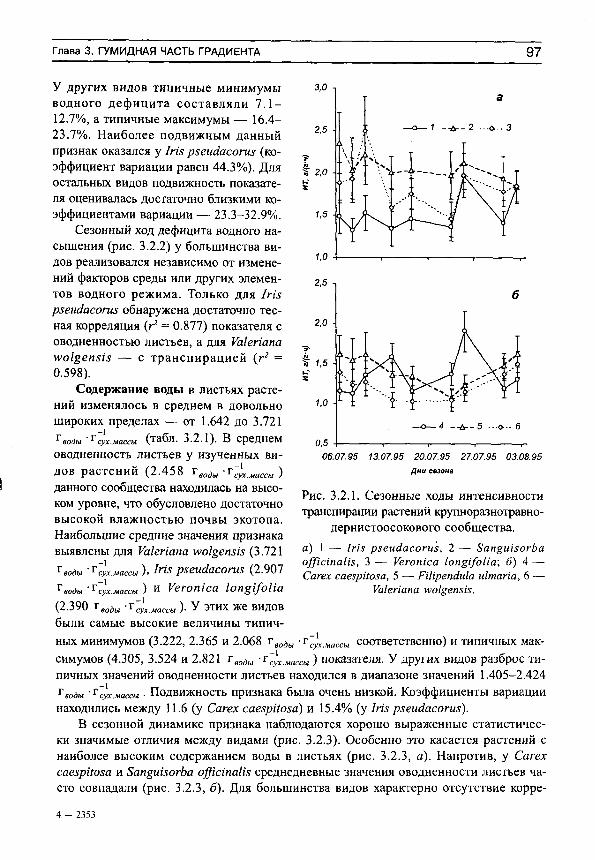
Глава 3. Г У М И Д Н А Я Ч А С Т Ь Г Р А Д И Е Н Т А 97
У других видов типичные минимумы водного д е ф и ц и т а составляли 7 . 1 -12.7%, а типичные максимумы — 16.4-23.7%. Наиболее подвижным данный признак оказался у Мз рзеЫасотз (коэффициент вариации равен 44.3%). Для остальных видов подвижность показателя оценивалась достаточно близкими коэффициентами вариации — 23.3-32.9%о.
Сезонный ход дефицита водного насыщения (рис. 3.2.2) у больщинства видов реализовался независимо от изменений факторов среды или других элементов водного режима. Только для 1пз рзеис{асогиз обнаружена достаточно тесная корреляция (/^ = 0.877) показателя с оводненностью листьев, а для Уа1епапа м/о1ёепз1з — с транспирацией (г^ = 0.598).
Содержание воды в листьях растений изменялось в среднем в довольно широких пределах — от 1.642 до 3.721 ^воды - ^'^.массы (табл. 3.2.1). в среднем оводненность листьев у изученных видов растений (2.458 г^^^^-г -1
сух.массы ) данного сообщества находилась на высоком уровне, что обусловлено достаточно высокой влажностью почвы экотопа. Наибольшие средние значения признака выявлены для Уакпапа •л1о1§епз1з (3.721 ^воды •^'с1х.массы ) , 1пз рзеис1асотз (2.907
•г„1, „,„,„) и Уеготса 1оп§1/оНа ). У этих же видов
воды (2.390 г , „ , „ . г сух.массы
1 - . д . _ 2 . . . о - . 3
06.07.95 13.07.95 20.07.95 Дни сезона
27.07.95 03.08.95
Рис. 3.2.1. Сезонные ходы интенсивности транспирации растений крупноразнотравно-
дернистоосокового сообщества.
а) 1 — 1г15 рзеиЛасогиз, 1 — 5ап%т8огЪа о^^'ШпаНв, 3 — Уеготса 1оп§1/оИа; б) 4 — Сагех саезркоза, 5 — РШрепМа иЫапа, 6 —
Уакпапа м;о1^епз1з.
были самые высокие величины типичных минимумов (3.222, 2.365 и 2.068 т^оды симумов (4.305, 3.524 и 2.821 г^^^ы -1
сух.массы
,-1
• сух.массы соответственно) и типичных мак-) показателя. У других видов разброс ти
пичных значений оводненности листьев находился в диапазоне значений 1.405-2.424 ^воды '^'сух.массы • ПОДВИЖНОСТЬ признака была очень низкой. Коэффициенты вариации находились между 11.6 (у Сагех саезрИоза) и 15.4%) (у Мз рзеис1асотз).
В сезонной динамике признака наблюдаются хорошо выраженные статистически значимые отличия между видами (рис. 3.2.3). Особенно это касается растений с наиболее высоким содержанием воды в листьях (рис. 3.2.3, а). Напротив, у Сагех саезрИоза и 8ап§шзогЬа о^!(1стаНз среднедневные значения оводненности листьев часто совпадали (рис. 3.2.3, б). Для большинства видов характерно отсутствие корре-
4 - 2353

98 Часть I. С е з о н н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в .
ляций данного показателя с внешними или внутренними факторами. Однако отмечена взаимосвязь оводненности листьев с дефицитом водного насыщения у 1пз рзеис1асоги5 {г' = 0.877) и с освещенностью, в л а ж н о с т ь ю воздуха и транспирацией у Уеготса 1оп§1/оНа (Л- = 0.802).
Сравнительный анализ
Экологическое сходство видов по совокупности показателей водного режима. В нижней части матрицы экологического сходства (табл. 3.2.2), как было отмечено выше, располагаются виды с относительно более мезофитным сочетанием признаков водного обмена. Это, имеющие достаточно высокое сходство по реализации всех признаков, 8ап§шзогЪа о^стаНз и Уа1епапа и^о/-§епз1з. Очень близко к этим видам примыкают Мз рзеи(1асотз и Уеготса 1оп-81/оНа. Признаком такой близкого сходства является то, что Зап^шзогЬа о^с1-паНз входит как в первую, так и во вторую группы растений (табл. 3.2.2). Наименее мезофитный комплекс признаков (прежде всего за счет высоких значений водного дефицита и низких величин оводненности листьев) отмечен у РШ-репс1и1а иЫапа и Сагех саезрИоза. Последний вид, занимавший доминирующее положение в сообществе, испытывал, судя по всему, определенные трудности с почвенным водоснабжением.
Действительно, год наблюдений был достаточно засушливым. Однако для большинства видов в данном сообществе эта ситуация не была экстремальной, так как недостаток воды в почве восполнялся за счет внутрипочвенного подтока воды, и поэтому запас влаги в верхнем горизонте почвы не опускался ниже 100 мм. Сагех саезрИоза образует массивные кочки 40-50 см в высоту и 50-70 см в диаметре. В них сосредоточена практически вся мочковатая корневая система растений. После окончания весеннего паводка водоснабжение Сагех саезрИоза резко ухудшается, поскольку корни лишь в небольшом количестве достигают почвы. Доминирование в сообществе данного вида обусловлено скорее морфологическими особенностями растений, высо-
06.07.95 03.08.95
Рис. 3.2.2. Сезонные ходы дефицита водного насыщения в листьях растений крупноразнотравно-дернистоосокового
сообщества.
а) 1 — Сагех саезрИоза, 2 — Р1ирепс1и1а и1тапа, 3 — Мз рзеис^асо^из•, б) 4 — Зап^ш-зогЬа о//гсшаИз, 5 — Уакпапа •л'о1§епз1з, 6 —
Уеготса 1оп§1/оИа.
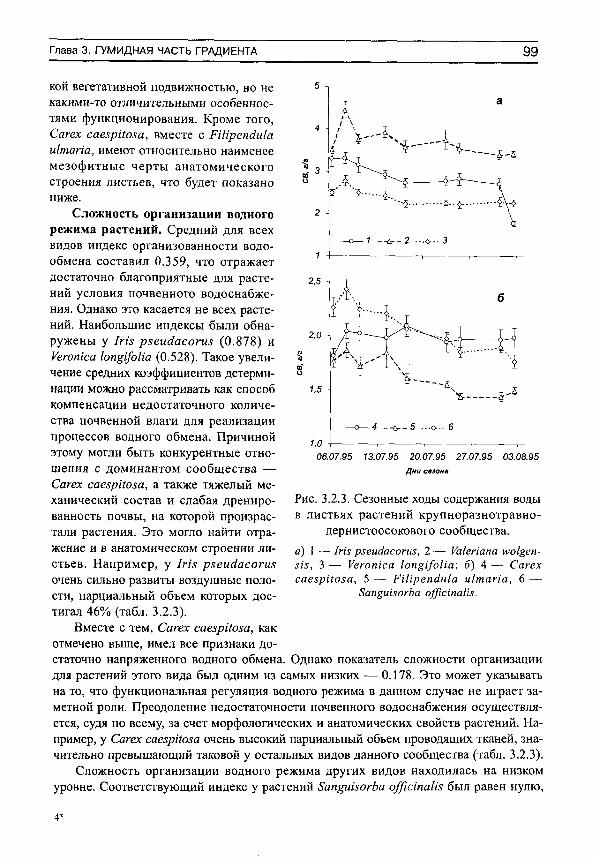
Глава 3. Г У М И Д Н А Я Ч А С Т Ь Г Р А Д И Е Н Т А 99
кои вегетативной подвижностью, но не какими-то отличительными особенностями функционирования. Кроме того, Сагех саезрИоза, вместе с РШрепйи1а иЫапа, имеют относительно наименее мезофитные черты анатомического строения листьев, что будет показано ниже.
Сложность организации водного режима растений. Средний для всех видов индекс организованности водообмена составил 0.359, что отражает достаточно благоприятные для растений условия почвенного водоснабжения. Однако это касается не всех растений. Наибольшие индексы были обнаружены у 1г1з рзеис(асогиз (0.878) и Уеготса 1оп§1/оНа (0.528). Такое увеличение средних коэффициентов детерминации можно рассматривать как способ компенсации недостаточного количества почвенной влаги для реализации процессов водного обмена. Причиной этому могли быть конкурентные отношения с доминантом сообщества — Сагех саезрИоза, а также тяжелый механический состав и слабая дрениро-ванность почвы, на которой произрастали растения. Это могло найти отражение и в анатомическом строении листьев. Например, у 1пз рзеи^асогиз очень сильно развиты воздушные полости, парциальный объем которых достигал 46% (табл. 3.2.3).
Вместе с тем, Сагех саезрИоза, как отмечено выше, имел все признаки достаточно напряженного водного обмена. Однако показатель сложности организации для растений этого вида был одним из самых низких — 0.178. Это может указывать на то, что функциональная регуляция водного режима в данном случае не играет заметной роли. Преодоление недостаточности почвенного водоснабжения осуществляется, судя по всему, за счет морфологических и анатомических свойств растений. Например, у Сагех саезрИоза очень высокий парциальный объем проводящих тканей, значительно превышающий таковой у остальных видов данного сообщества (табл. 3.2.3).
Сложность организации водного режима других видов находилась на низком уровне. Соответствующий индекс у растений 8ап§шзогЬа о^стаИз был равен нулю,
03.08.95
Рис. 3.2.3. Сезонные ходы содержания воды в листьях растений крупноразнотравно-
дернистоосокового сообщества.
а) 1 — / т рзеиёасогиз, 2 — Уа1епапа м'о1§еп-513, 3 — Уеготса 1оп§1/оИа; б) А — Сагех саезрИоза, 5 — ртреп(1и[а и1таг1а, 6 —
8ап§шзогЬа о]]1стаИз.
4*
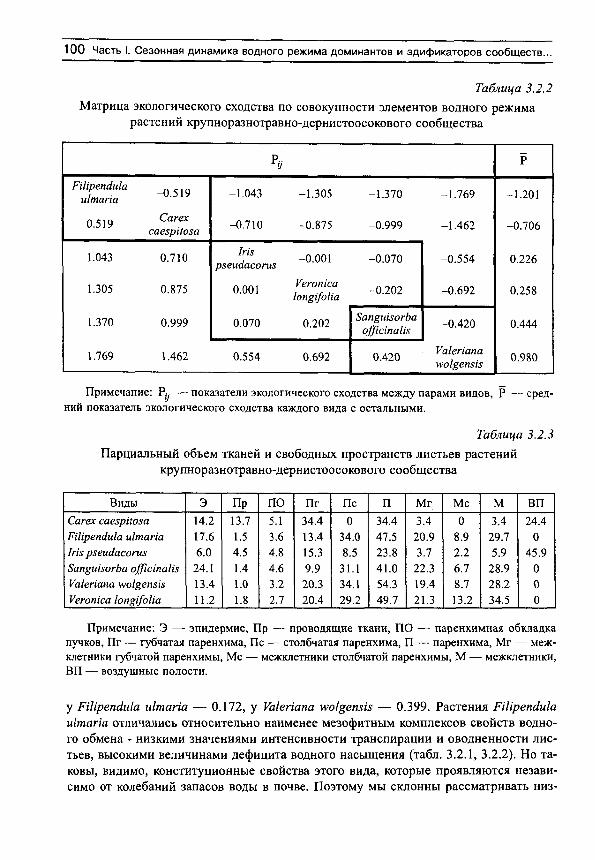
100 Часть I. С е з о н н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в . . .
Таблица 3.2.2 Матрица экологического сходства по совокупности элементов водного режима
растений крупноразнотравно-дернистоосокового сообщества
Р
РШреп<1и1а и1тапа -0.519 -1.043 -1.305 -1.370 -1.769 -1.201
0.519 Сагех саезрИоза -0.710 -0.875 -0.999 -1.462 -0.706
1.043 0.710 Мз рзеибасогиз -0.001 -0.070 -0.554 0.226
1.305 0.875 0.001 Уеготса 1оп§1/оИа -0.202 -0.692 0.258
1.370 0.999 0.070 0.202 Зап^шзогЬа о^Г'сшаИз -0.420 0.444
1.769 1.462 0.554 0.692 0.420 Уакпапа м/о1§епх13 0.980
Примечание: Р^ — показатели экологического сходства между парами видов, Р — средний показатель экологического сходства каждого вида с остальными.
Таблица 3.2.3
Парциальный объем тканей и свободных пространств листьев растений крупноразнотравно-дернистоосокового сообщества
Виды Э Пр ПО Пг Пс П Мг Мс М ВП Сагех саезрИоза 14.2 13.7 5.1 34.4 0 34.4 3.4 0 3.4 24.4 РШреп(1и1а и1тапа 17.6 1.5 3.6 13.4 34.0 47.5 20.9 8.9 29.7 0 Мз рзеи^асотз 6.0 4.5 4.8 15.3 8.5 23.8 3.7 2.2 5.9 45.9 8ап^1зогЬа оЛГ1стаИз 24.1 1.4 4.6 9.9 31.1 41.0 22.3 6.7 28.9 0 Уакпапа м'о1^епз1з 13.4 1.0 3.2 20.3 34.1 54.3 19.4 8.7 28.2 0 Уеготса 1оп§1/оИа 11.2 1.8 2.7 20.4 29.2 49.7 21.3 13.2 34.5 0
Примечание: Э — эпидермис, Пр — проводящие ткани, ПО — паренхимная обкладка пучков, Пг — губчатая паренхима, Пс — столбчатая паренхима, П — паренхима, Мг — межклетники губчатой паренхимы, Мс — межклетники столбчатой паренхимы, М — межклетники, ВП — воздушные полости.
у РШрепЛи1а иЫапа — 0.172, у Уакпапа ^о1§епв1в — 0.399. Растения РШрепс1и1а иЫапа отличались относительно наименее мезофитным комплексов свойств водного обмена - низкими значениями интенсивности транспирации и оводненности листьев, высокими величинами дефицита водного насыщения (табл. 3.2.1, 3.2.2). Но таковы, видимо, конституционные свойства этого вида, которые проявляются независимо от колебаний запасов воды в почве. Поэтому мы склонны рассматривать низ-

Глава 3. Г У М И Д Н А Я Ч А С Т Ь Г Р А Д И Е Н Т А 101
кие индексы организованности водообмена у всех трех видов как отражение снижения функциональной регуляции в ответ на благоприятные условия почвенного водоснабжения,
Морфолого-анатомическая характеристика
8ап§шзогЪа о^стаНз, РШрепЛиШ и1тапа, Уа1епапа ко1§епз13, Уеготса 1оп§1/оИа — это многолетние травы с мезоморфными довольно крупными, рассеченными или сложными листьями, имеющими пластинку и черешок. Характерным анатомическим признаком этих растений является довольно рыхлый дорсивентральный мезофилл. Палисадная паренхима варьирует от 1-слойной у Уа1епапа \Vо^§епз^з и 8ап-^шзогЪа о//1С1паИз до 2-слойной у Ртрепс1и1а иЫапа и Уеготса 1оп§1-/оИа. Клетки довольно крупные, вытянутой формы. Губчатая паренхима рыхлая, довольно крупноклетная, в основном изодиаметрическая с крупными межклетниками. Верхняя и нижняя эпидерма по размеру клеток почти не различаются (относительно крупно-клетные). Устьица некрупные.
Сагех саезрИоза и 1г1з рзеис1асо-тз — многолетники с линейными, довольно широкими листьями {Сагех саезрИоза — около 4 мм шириной, Мз рзеис1асогиз — около 2 см), состоящими из пластинки и влагалища. Мезо-
Рис. 3.2.5. Корреляции между парциальными объемами проводящих тканей и столбчатой паренхимы (а), межклетников (б) в листьях растений крупноразнотравно-дернистоосокового
сообщества.
60 .
Парциальный объем эпидермиса, %
Рис. 3.2.4. Корреляция между парциальными объемами эпидермиса и свободных пространств в листьях растений крупноразнот-
равно-дернистоосокового сообщества.
о -) 1 1 1
о 5 10 15 Парциальный объем проводящих тканей, %

102 Часть 1. С е з о н н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в .
35 -, 5?
о -I , , 1 ,
о 10 20 30 40 Парциальный объем столбчатой паренхимы, %
Рис. 3.2.6. Корреляция между парциальными объемами столбчатой паренхимы и межклетников в листьях растений крупнораз-нотравно-дернистоосокового сообщества.
6 -I
§ 3
5 -
20 30 40 50 60 Парциальный объем паренхимы, %
филл изолатеральный у / г и р5еи<1асо-гиз и гомогенный у Сагех саезрИоза, со слабо выраженным слоем палисадных клеток. Все клетки мезофилла расположены рыхло. Межклетники довольно крупные, имеются воздухоносные полости. Эпидерма 1-слойная, с довольно крупными клетками в верхнем слое, с утолщенными наружными стенками. У Сагех саезрИоза довольно толстая кутикула, тяжи механических тканей представлены сверху и снизу проводящих пучков, в то время как у Мз рзеи-(1асогиз механические ткани развиты очень слабо. Опушение отсутствует.
Анализ соотношения тканей и свободных пространств в листьях растений (табл. 3.2.3) позволил выявить некоторые межвидовые корреляции.
Парциальные объемы эпидермиса и свободных пространств (межклетников и воздушных полостей) имеют достаточно тесную статистическую связь (рис. 3.2.4). У видов с относительно более развитым эпидермисом межклетные пространства имеют меньший объем; уменьшение парциального объема эпидермиса сопровождается увеличением доли свободных пространств в листьях растений.
Парциальные объемы проводящих тканей коррелируют с парциальными объемами столбчатой п а р е н х и м ы (рис. 3.2.5, а) и межклетников (рис. 3.2.5, б). Виды с относительно слабо развитыми проводящими пучками име-
Рис. 3.2.7. Корреляции между парциальными объемами паренхимы и обкладок пучков (а), межклетников (б) в листьях растений крупноразнотравно-
дернистоосокового сообщества.

Глава 3. Г У М И Д Н А Я Ч А С Т Ь Г Р А Д И Е Н Т А 103
ют более развитую столбчатую паренхиму и менее плотный (с большим количеством межклетников) мезофилл. По мере увеличения доли участия проводящих тканей в сложении листьев парциальные объемы столбчатой паренхимы и межклетников снижаются.
Парциальные объемы столбчатой паренхимы и межклетников (как следует уже из рис. 3.2.5) тесно взаимосвязаны (рис. 3.2.6). Виды с малыми объемами столбчатой паренхимы имеют и относительно небольшое количество межклетников. Усиление развития столбчатой паренхимы сопровождается и увеличением объема межклетников.
Парциальные объемы всей паренхимы (хлоренхимы) отрицательно коррелируют с парциальными объемами обкладок пучков (рис. 3.2.7, я): по мере увеличения обкладок пучков относительное участие паренхимы в сложении листьев снижается. Хло-ренхима, так же как и столбчатая паренхима, положительно коррелирует с межклетниками (рис. 3.2.7, б). Увеличение относительной доли участия столбчатой паренхимы и хлоренхимы в целом сопровождается увеличением парциальных объемов межклетников.
Для выявления количественных оценок сходства или различия видов по совокупности показателей анатомической структуры листьев построена матрица анатомического сходства листьев растений (табл. 3.2.4).
Относительно более ксероморфные виды характеризуются большим объемом эпидермиса, склеренхимы, паренхимы, проводящей ткани, и меньшим объемом межклетников. В данном случае это один вид, расположенный в верхней части табл. 3.2.4, — Сагех саезрИоза. В противоположном углу матрицы анатомического сходства находится вид с относительно более мезоморфными особенностями строения листьев — 1пз рзеи^асогиз. Оба эти вида, Сагех саезрИоза и Мз рзеис1асогиз, не только сильно отличаются друг от друга, но и от всех других, расположенных между ними достаточно компактной группой (табл. 3.2.4).
Таблица 3.2.4
Матрица анатомического сходства по совокупности показателей анатомической структуры листьев растений крупноразнотравно-дернистоосокового сообщества
Р
Сагех саезрИоза -0.996 -1.063 -1.599 -1.942 -3.959 -1.912
0.996 РШрепс1и1а иЫапа -0.020 -0.304 -0.446 -3.402 -0.635
1.063 0.020 8ап§ш50гЬа оЛГ1стаНз -0.354 -0.396 -3.120 -0.557
1.599 0.304 0.354 Уакпапа м!о1§епз1з -0.177 -3.671 -0.318
1.942 0.446 0.396 0.177 Уеготса 1оп^1/оИа -1.971 0.198
3.959 3.402 3.120 3.671 1.971 Мз рзеис1асогиз 3.225
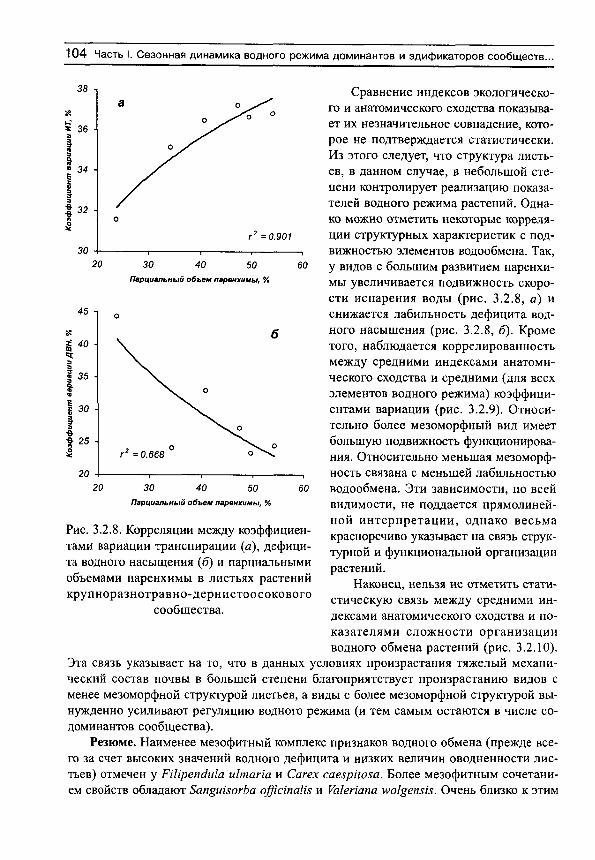
104 Часть I. С е з о н н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в . .
38 п
зе
34
32
30 г =0.901
20 30 40 50 60 Парциальный объем паренхимы, *
Сравнение индексов экологического и анатомического сходства показывает их незначительное совпадение, которое не подтверждается статистически. Из этого следует, что структура листьев, в данном случае, в небольшой степени контролирует реализацию показателей водного режима растений. Однако можно отметить некоторые корреляции структурных характеристик с подвижностью элементов водообмена. Так, у видов с большим развитием паренхимы увеличивается подвижность скорости испарения воды (рис. 3.2.8, а) и снижается лабильность дефицита водного насыщения (рис. 3.2.8, б). Кроме того, наблюдается коррелированность между средними индексами анатомического сходства и средними (для всех элементов водного режима) коэффициентами вариации (рис. 3.2.9). Относительно более мезоморфный вид имеет большую подвижность функционирования. Относительно меньшая мезоморф-ность связана с меньшей лабильностью водообмена. Эти зависимости, по всей видимости, не поддается прямолинейной интерпретации , однако весьма красноречиво указывает на связь структурной и функциональной организации растений.
Наконец, нельзя не отметить статистическую связь между средними индексами анатомического сходства и показателями сложности организации водного обмена растений (рис. 3.2.10).
Эта связь указывает на то, что в данных условиях произрастания тяжелый механический состав почвы в большей степени благоприятствует произрастанию видов с менее мезоморфной структурой листьев, а виды с более мезоморфной структурой вынужденно усиливают регуляцию водного режима (и тем самым остаются в числе со-доминантов сообщества).
Резюме. Наименее мезофитный комплекс признаков водного обмена (прежде всего за счет высоких значений водного дефицита и низких величин оводненности листьев) отмечен у ЕШрепМа иЫапа и Сагех саезрИоза. Более мезофитным сочетанием свойств обладают 8ап§шзогЪа оЛПстаИз и Уа1епапа м!о1^епз1з. Очень близко к этим
45
г? "О -
ч
I 35 •
I 30
I •а |-
20
= 0.668
20 30 40 50 Парциальный объем паренхимы, %
60
Рис. 3.2.8. Корреляции между коэффициентами вариации транспирации (а), дефицита водного насыщения (б) и парциальными объемами паренхимы в листьях растений крупноразнотравно-дернистоосокового
сообщества.
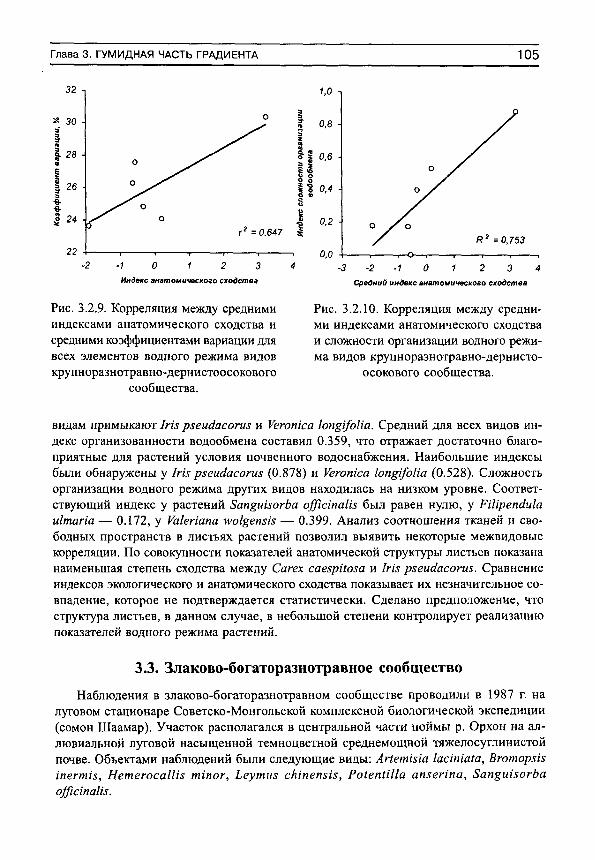
Глава 3 . Г У М И Д Н А Я Ч А С Т Ь Г Р А Д И Е Н Т А 105
32 п
« 30
22 -1 , , , , , , - 2 - 1 0 1 2 3 4
Индекс анатоиическоео сходства
Рис. 3.2.9. Корреляция между средними индексами анатомического сходства и средними коэффициентами вариации для всех элементов водного режима видов крупноразнотравно-дернистоосокового
сообщества.
Средний индекс анатомического сходства
Рис. 3.2.10. Корреляция между средними индексами анатомического сходства и сложности организации водного режима видов крупноразнотравно-дернисто-
осокового сообщества.
г' = 0.647
1,0
1
I 0.8
| | 0,6
I I 0,4 0 «
1 0,2
видам примыкают 1пз р5еи(1асоги5 и Уеготса 1оп^1/оНа. Средний для всех видов индекс организованности водообмена составил 0.359, что отражает достаточно благоприятные для растений условия почвенного водоснабжения. Наибольщие индексы были обнаружены у 1т рзеи^асогш (0.878) и Уеготса 1оп§1/оИа (0.528). Сложность организации водного режима других видов находилась на низком уровне. Соответствующий индекс у растений 8ап^тюгЪа о^стаНз был равен нулю, у РШрепМа и1тапа — 0.172, у Уакпапа м>о1§епв1з — 0.399. Анализ соотношения тканей и свободных пространств в листьях растений позволил выявить некоторые межвидовые корреляции. По совокупности показателей анатомической структуры листьев показана наименьшая степень сходства между Сагех саезрИоза и Мз рзеи^асогиз. Сравнение индексов экологического и анатомического сходства показывает их незначительное совпадение, которое не подтверждается статистически. Сделано предположение, что структура листьев, в данном случае, в небольшой степени контролирует реализацию показателей водного режима растений.
3.3. Злаково-богаторазнотравное сообщество
Наблюдения в злаково-богаторазнотравном сообществе проводили в 1987 г на луговом стационаре Советско-Монгольской комплексной биологической экспедиции (сомон Шаамар). Участок располагался в центральной части поймы р. Орхон на аллювиальной луговой насыщенной темноцветной среднемощнои тяжелосуглинистой почве. Объектами наблюдений были следующие виды: Апет1з1а 1асша1а, Вготорз1з тегтгз, НетегосаШз ттог, Ьеутиз ск1пепзгз, РогепИИа апзегта, Зап^шзогЬа о/]1ста1гз.

106 Часть I. С е з о н н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в .
Исходные данные
Интенсивность транспирации растений сообщества была достаточно высока и составляла в среднем 2.12 Г ^ „ Й „ -^'с^хмассы ' ч ' ' (табл. 3.3.1). Отчетливо выделяются группы видов с более высокой {Апетша 1аста1а, РоШНИа апзегта и 8ап§шзогЬа о^стаИз) и низкой скоростью испарения воды {Вготорзгз тегтгз, НетегосаШз ттог и Ьеутиз сктепзгз) (табл. 3.3.1, рис. 3.3.1). Самый широкий размах колебаний признака наблюдался у Апетгзга 1ас1та1а (типичный минимум был равен 1.58, типичный мак-
-1
симум — 5.16 Гдд̂ у ''^сух.массы )• ̂ растсний Ро1еп(И1а апзегта и Бап^шзогЬа о^1С1паИз диапазон типичных изменений показателя находился между 1.21 и 3.73
. _-1
' воды ̂ сух.массы ч ' . Менее всего была подвержена колебаниям интенсивность транспирации Ьеутиз сктепзгз (типичный минимум — 0.77, типичный максимум — 1.72
_-1 ч ' ) и Вготор$1з Ыегт1з (0.89-2.06 ^ воды
5 -I
-1 .-1 •
2 -
1 - ^ _ 2 . . . о . . 3
3,0
2.5
•3-2,0
1.0
0.5 14.06.87 28,06.87 12.07.87 26.07.87 09.08.87 23.08.87
Дни сезона
Рис. 3.3.1. Сезонные ходы интенсивности транспирации растений злаково-богаторазнотравного
сообщества.
а) 1 — Апетша 1ас1та1а, 2 — Ро1еп1И1а апзегта, 3 — 5ап^1зогЬа о/^1стаНз; б ) 4 — Вготорз/з тегтгз, 5 —
НетегосаШз ттог, 6 — Ьеутиз сИтепзьз.
^воды • ^с'ух.массы ' ^ ' )• ДоСТаТОЧНО широким был размах типичных колебаний у НетегосаШз ттог (1.04-2.69 г , „ , „ т ; ; , „ - ч - ' ) . Однако коэффициенты вариации, с помощью которых мы оценивали подвижность элементов водного обмена, бьши близки у всех видов, показывая достаточно умеренную лабильность признака (32.2-42.5%).
В сезонной динамике (рис. 3.3.1) у большинства видов наблюдались согласованные изменения скорости испарения воды. Это, видимо, обусловлено сходными комплексами корреляций, выявленными почти для всех видов, кроме 5ап§и1зогЬа оЛ[1стаНз. Интенсивность транспирации Ьеутиз сктепзгз положительно коррелировала с оводненностью листьев (И = 0.894). Для Вго-торзгз тегтгз {К^ = 0.982), НетегосаШз ттог {К' = 0.875) и Ро1еп-аПа апзегта {К^ = 0.894) отмечена зависимость сезонной динамики транспирации от оводненности листьев и дефицита влажности воздуха. Скорость испарения воды растений Аг1ет1з1а 1ас1та1а была связана только с дефицитом влажности воздуха (г^ = 0.405).

Глава 3. Г У М И Д Н А Я Ч А С Т Ь Г Р А Д И Е Н Т А 107
Таблица 3.3.1
Средние значения интенсивности транспирации (ИТ, г̂ ^^ ,̂ -г^^^^^^.^, - ч " ' ) , дефицита водного насыщения (ДВН, % ) , содержания воды (СВ, г^^^ ,̂ '^сух.массы
осмотического давления клеточного сока (ОД, МПа) видов злаково-богаторазнотравного сообщества
Вид ИТ ДВН СВ ОД
Вид п X 8х п X & п п X 8х
АПетша 1ас1таШ 238 3.216 1.368 234 19.9 6.7 238 3.203 0.565 24 1.51 0.26 Вготорз18 тегтгз 168 1.454 0.485 138 8.6 2.4 168 2.044 0.246 21 1.78 0.41 НетегосаШз ттог 232 1.799 0.643 232 9.7 1.9 232 3.316 0.406 24 1.67 0.41 Ьеутиз сМпепзхз 238 1.185 0.381 238 8.8 2.3 238 1.805 0.180 24 2.12 0.58 Ро1епШ1а апзегта 238 2.305 0.946 238 18.4 4.4 238 2.044 0.219 24 1.79 0.58 5ап^1зогЬа 238 2.553 0.926 238 15.0 4.3 238 2.369 0.383 24 2.06 0.38 о(11стаНз В целом для сообщества
1352 2.119 1.113 1348 13.6 6.1 1352 2.481 0.695 141 1.82 0.49
Дефицит водного насыщения в листьях растений был сравнительно невысоким у всех видов — в среднем 13.6% (табл. 3.3.1). Однако отчетливо выделяются группы с относительно более высокими {Аг1ет151а 1ас1та1а, Ро1еп1И1а апзегта, 8ап§шзогЬа о^стаШ) и низкими (Вготорз15 тегтгз, НетегосаШз ттог, Ьеутиз сЫпепзгз) значениями показателя. У растений второй группы размах колебаний типичных значений признака был очень узким и находился между 5.5 и 11.9%. Виды с более высоким водным дефицитом в листьях продемонстрировали и более широкий диапазон, внутри которого находились типичные значения показателя. Он составил 9.5-29.6%. Но такие различия в диапазонах варьирования не могут служить мерой подвижности признака (см. раздел «Определения»). Оценки коэффициентов вариации показывают, что лабильность дефицита водного насыщения бьша примерно одинаковой у всех растений (с некоторыми исключениями). Минимальная подвижность показателя была обнаружена у НетегосаШз ттог (19.3%), максимальная — у АПетгзга 1ас1та(а (33.6%>). У остальных видов подвижность дефицита насыщения листьев была практически неразличимой — 23.7-28.6%).
Рисунки сезонных ходов дефицита водного насыщения (рис. 3.3.2) были очень сходными у всех видов. В большинстве случаев они очень хорошо повторяли друг друга. У растений с более высокими значениями показателя можно было обнаружить статистически значимые различия между некоторыми среднедневными величинами (рис. 3.3.2, а). У другой группы видов таких различий практически не наблюдалось (рис. 3.3.2, б).
Сезонная динамика дефицита водного насыщения Вготорзгз тегтгз (И = 0.762) и НетегосаШз ттог (г^ = 0.775) бьша связана с влажностью воздуха. Для Аг1ет1з1а 1ас1-таШ (К' = 0.720) и 8ап§шзогЬа о//1стаНз {К^ = 0.815) выявлены, кроме того, корреляции и с оводненностью листьев. Водный дефицит Ро1еп1И1а апзегта коррелировал

108 Часть I. С е з о н н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в . .
30
25
К 20
15
10 -1 _ ^ - 2 ...о--- 3
и 1
12 -
10 •
только с оводненностью листьев = 0.529). Для Ьеутш сЫпепзгз
никаких корреляций не выявлено. Содержание воды в листьях
растений было высоким и составило в среднем 2.48 ,оды-'^~с1х.л,ассы (табл. 3.3.1). Наибольшее содержание воды было обнаружено в листьях НетегосаШз ттог и Аг1ет1-31а 1асш1а(а. Типичные значения признака у этих видов находились между 2.5 и 3.9 г,„^„-г^^^^^^^^ . Наименьшая оводненность листьев наблюдалась у Ьеутш сктепз13 с диапазоном типичных значений между 1.6 и 2.0 Г , „ ^ „ •^'с^^.массы • Для других видов (с промежуточными средневидовыми величинами показателя) размах колебаний составил 1.7-2.8 воды '^'^^'х.массы • Однако подвижность оводненности листьев была очень низкой, а коэффициенты вариации мало отличались у разных видов (10.0-17.6%).
В сезонных ходах обнаруживается общая тенденция снижения оводненности листьев по мере их старения. Наибольшую выраженность эта тенденция имеет у растений Бап^шзогЬа о/^гстаНз (рис, 3.3.3, а), наименьшую — у Ьеутиз сНтепзгз (рис. 3.3.3, б).
Сезонная динамика содержания воды в листьях Аг1ет1з1а 1асШа(а {К^ = 0.670) и Зап^шзогЪа о^а^стаНз (К^ = 0.609) связана с изменениями дефицита водного насыщения, а также температуры воздуха. Оводненность листьев Вготорзгз тегтгз (Н = 0.864), Ьеутиз сЫпепзгз (Н = 0.811) и Ро1еп1Ша апзегта (Н = 0.726) была взаимосвязана только с интенсивностью транспирации, а НетегосаШз ттог (К^ = 0.876), кроме того, с температурой воздуха.
Осмотическое давление клеточного сока у всех видов различалось мало, изменяясь в среднем от 1.51 {у Аг1ет1з1а 1асШа1а) до 2.12 МПа {у Ьеутиз сШпепзгз) (табл. 3.3.1). Несколько отличались диапазоны варьирования признака. У Ьеутш сЫпепзгз типичный минимум осмотического давления бьш равен 1.68, а типичный максимум — 2.63 МПа. Соответствующие величины для 8ап§шзогЪа о/]1стаНз составили 1.84 и
74.06.87 28.06.87 12.07.87 26.07.87 09.08.87 23.08.87 Дни сезона
Рис. 3.3.2. Сезонные ходы дефицита водного насыщения в листьях растений злаково-богато
разнотравного сообщества.
а) 1 — Аг1ет1з1а 1ас1та1а, 2 — Ро1епШ1а апзегта, 3 — Зап^шзогЬа о//1стаИз; б) 4 — Вготорзгз тегтгз, 5 — НетегосаШз ттог, 6 — Ьеутиз сНгпепзгз.

Глава 3. ГУМИДНАЯ ЧАСТЬ ГРАДИЕНТА 109
2.21 МПа. У остальных видов диапазон типичных колебаний показателя находился между 1.38 (у АПетта 1асша(а) и 1.98 МПа (у Ро1еп1г11а апзегта). Подвижность признака бьша умеренной и примерно одинаковой у всех растений: коэффициенты вариации изменялись от 22.9 (у НетегосаШз ттог) до 27.8% (у Аг1ет1з1а 1ас1таш).
В сезонной динамике осмотического давления (рис. 3.3.4) обнаружены корреляции этого показателя с температурой воздуха у НетегосаШз ттог (г^ = 0.622) и с освещенностью у Ьеутиз сЫ-пепзгз (г^ = 0.761). Для остальных видов не удалось обнаружить статистические связи этого признака с другими элементами водного режима или факторами среды.
4,5
4.0
3,5
3,0
2,5 -
2,0 -
1,5
2,5
2,3
2,1
-1 . ^ - 2 . . -с. . . 3 •-5..5-..-5
1.9
1,7
1.5 14.06.87 28.06.87 12.07.87 26.07.87 09.08.87 23.08.87
Днни сезона
Рис. 3.3.3. Сезонные ходы содержания воды в листьях растений злаково-богаторазнотравного
сообщества.
а) 1 — Аг1ет1з1а 1аст1а1а, 2 — НетегосаШз ттог, 3 — 5ап§шзогЬа о^рстаНз; б) 4 — Вготорзгз тегтгз,
5 — Ьеутиз сНтепзгз, 6 — Ро1еШШа апзегта.
Сравнительный анализ
Экологическое сходство видов по совокупности элементов водного режима. Напомним, что анализ экологического сходства видов позволяет выявить группы видов со сходной реализацией признаков, а также ранжировать растения по степени их относительной мезофитности.
В нижней части матрицы экологического сходства (табл. 3.3.2) расположены Апетгзга 1аста1а и НетегосаШз ттог, проявляющие высокую степень сходства в реализации совокупности элементов водного обмен. Эти виды характеризуются и относительно более мезофитным комплексом признаков по сравнению с другими растениями (в основном за счет высокой оводненности листьев и низкого осмотического давления, а также высокой интенсивности транспирации у Апет1з1а 1асШа(а). В противоположном ушу матрицы расположен Ьеутиз сЫпепзгз, с относительно менее мезофитным комплексом признаков водного режима. Этот вид имеет высокое экологическое сходство с Ро1еп1Ша апзегта, который в свою очередь входит в группу видов с промежуточным набором признаков (вместе с Бап^шзогЬа о^ГгстаИз и Вготорзгз тегтгз). Однако и различия Ьеутиз сМпепзгз с последними двумя видами не очень

110 Часть I. С е з о н н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в .
3,0
2,5
2,0 -
1,5 -
1.0
. . .5
-1 - ^ - 2 . - - о - . 3
2,2 -,
2,0
1.8
' .6
1,4
1,2 -.4 5 б
сильны и при некотором ослаблении критерия экологического сходства все четыре вида могут быть объединены в одну группу. Таким образом, в данных условиях произрастания мы можем выделить две группы; первую, относительно более мезофитную, составляют АНетша 1ас1та1а и НетегосаШз ттог, вторую, несколько менее мезофитную, — Ьеутиз сНтепз1з, Ро(еп1И1а апзегта, 8ап§шзогЬа о^стаИз и Вготорзгз гпегтгз.
Сложность организации водного режима растений. Средний индекс организованности водообмена для растений данного сообщества составил 0.563. Наименьшие индексы были выявлены для 8ап-^тзогЬа о/ДстаНз (0.356) и Аг1е-т1з1а ШсШаШ' (0.449), наибольшие — для Вготорз1з тегтгз (0.652) и НетегосаШз ттог Для Ьеутиз сЫпепзгз и Ро1епШ1а апзегта соответствующие индексы были равны 0.596 и 0.537. Все эти индексы достаточно высоки для данных условий произрастания и в среднем превышают индексы, обнаруженные в двух предыдущих сообществах (осоково-разнотрав-но-злаковом и крупноразнотравно-дернистоосоковом) с меньшими запасами почвенной влаги.
Можно было бы предположить, что тяжелый механический состав почвы данного (злаково-богаторазнотравного) сообщества мешает нормальному водообмену растений, однако отсутствие зависимостей элементов водного режима от влажности почвы (в сезонной динамике) свидетельствует, что это не так. Кроме того, средние величины показателей вполне соответствуют условиям произрастания. Остается предположить, что увеличение запасов воды в почве, сопровождающееся увеличением плотности сообществ, благоприятствует возникновению и процветанию популяций с конкурентными и/или конкурентно-рудеральными стратегиями, что связано с достаточным количеством ресурсов (Бигон и др., 1989) и слабой напряженностью экотопического отбора. Не исключено, что усиление конкурентных отношений между растениями может способствовать некоторому росту организованности водного обмена.
1.0
14.08.87 28.06.87 12.07.87 26.07.87
Дни сезона
09.08.87 23.08.87
Рис. 3.3.4. Сезонные ходы осмотического давления клеточного сока в листьях растений злаково-
богаторазнотравного сообщества.
а) 1 — Ьеутиз сктепзгз, 2 — Ро1еп1И1а апзегта, 3 — 8ап^и13огЬа о//1с1паИз\) 4 — Аг1ет1з1а ЬстгаШ, 5 — Вготорзгз тегтгз, 6 — НетегосаШз
ттог.

Глава 3. Г У М И Д Н А Я Ч А С Т Ь Г Р А Д И Е Н Т А 111
Таблица 3.3.2 Матрица экологического сходства по совокупности элементов водного режима
растений злаково-богаторазнотравного сообщества
Р
Ьеутиз сктепз18 -0.138 -0.532 -0.622 -1.121 -1.609 -0.805
0.138 Рошииа апзегта -0.385 -0.420 -0.959 -1.525 -0.630
0.532 0.385 8ап^шзогЬа о//1стаИз -0.016 -0.779 -1.012 -0.178
0.622 0.420 0.016 Вготорз1з тегт1з -0.727 -1.037 -0.141
1.121 0.959 0.779 0.727 Аг1ет1з1а 1ас1та1а -0.129 0.692
1.609 1.525 1.012 1.037 0.129 НетегосаШз ттог 1.063
Примечание: Р .̂ — показатели экологического сходства между парами видов, Р — средний показатель экологического сходства каждого вида с остальными.
Резюме. В данных условиях произрастания можно выделить две группы растений: первую, относительно более мезофитную, составляют Аг1ет181а ШсгтаШ и НетегосаШз ттог, вторую, несколько менее мезофитную, — Ьеутиз сНтепз1з, Ро1еп(И1а апзегта, 8ап§и1зогЬа о^)1стаИз и Вготорзгз тегтгз. Обнаружены достаточно высокие для данных условий произрастания индексы организованности водообмена. В среднем они превышают индексы, обнаруженные в двух сообществах (осоково-раз-нотравно-злаковом и крупноразнотравно-дернистоосоковом) с меньшими запасами почвенной влаги. Сделано предположение, что усиление конкурентных отношений между растениями может способствовать некоторому росту организованности водного обмена.
3.4. Остроосоковое сообщество
Остроосоковое сообщество расположено на границе притеррасной части поймы р. Хопер (Хоперский заповедник, Воронежская область) с надпойменной террасой. Сырые остроосоковые сообщества произрастают в прибрежной зоне на пологом склоне обсохшего озера Калмычок (0-30 см от уровня воды) и занимают экотопы с отметкой ниже 300 см над меженью р. Хопер на оглееных песчаных почвах (Титов, Печенюк, 1990). Травостой густой — общее проективное покрытие составляет 85-100%. Средняя высота травостоя — 90-120 см. Сагех асиШ — доминант сообщества, дающий до 60-70% проективного покрытия. Массово встречается А§гозИз з1о1от/ега Ь., из наиболее распространенных — Сагех рзеис1осуреп13 ^., Ьусориз еигораеиз. У са-

112 Часть I. С е з о н н а я д и н а м и к а водного р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в . .
мого уреза воды выделяются группировки гидрофитов: 8раг§апшт егесШт, 8а^Ша-па ш^иН/оНа, АНзта р1ап(а^о-адиааса, ВШотиз итЬеНаШз. Объектами исследования здесь были гигрофиты и гидрофиты АНзта р1ап1а§:о-адиаИса, ВШотиз итЬеИаШз, Сагех асиШ, Ьусориз еигораеиз, 8а§Шапа за§ИИ/оИа, 8раг§апшт егесШт. Корневые системы всех исследованных растений достигали грунтовых вод, которые находились на глубине 10-20 см. Однако увлажнение экотопа не было застойным, о чем свидетельствовало отсутствие кочек Сагех асиШ. Средний запас влаги в верхнем слое почвы составлял 144 мм и практически не изменялся в течение периода наблюдений.
Исходные данные
Интенсивность транснирации в среднем для сообщества составляла 2.977 ^воды ''^^ух.массы "ч"' (табл. 3.4.1). Такос интенсивное испарение воды было обусловлено оптимальной увлажненностью экотопа. Несмотря на это, между видами наблюдались большие различия. АНзта р1ап1а§о-адиаНса, Ьусориз еигораеиз и 8а§Шапа за§ИН/оНа (рис. 3.4.1, а) испаряли воду значительно более интенсивно, чем ВиЮтиз итЪеПаШз, Сагех асШа, 8раг§атит егесШт (рис. 3.4.1, б). Это обусловлено, в первую очередь, различиями в объеме межклетников у перечисленных видов. У видов с большим парциальным объемом межклетников наблюдалась более интенсивная транспирация, и наоборот. Типичные минимумы признака у АНзта р1ап1а§о-адиаНса и 8а§Шапа за§ИН/оНа находились на уровне 2.74-2.86 Тд„^^ • г'^ух.массы • ч~ ' , а типичные максимумы составляли 5.35 и 5.24 г^^^^ - ^сух.массы ' ч ' - Типичный максимум интенсивности транспирации Ьусориз еигораеиз был также высок (4.65 т^^^^ -г^^х.массы ) .
а типичный минимум был примерно таким же, как и у остальных видов (1.28-1.61 ^воды '^сух массы • ч~' )• ПОДВИЖНОСТЬ признака была достаточно умеренной: коэффициенты вариации находились между 22.8 (у 8а^Шапа за§НН/оНа) и 39.5% (у Ьусориз еигораеиз).
Таблица 3.4.1
Средние значения интенсивности транспирации (ИТ, г^о^ы '^сух.массы 'ч~'). дефицита водного насыщения (ДВН, % ) , содержания воды в листьях (СВ, т^^^^ - ^сух.массы)
у растений остроосокового сообщества
Виды
Показатели
Виды ИТ ДВН СВ
п X 8х п X 8х п X
АНзта р1ата^о- 138 3.978 1.008 138 2.7 1.8 138 4.981 0.563 адиаИса Вшотиз итЬеИаШз 118 2.407 0.698 118 12.5 6.7 118 6.609 0.816 Сагех асиШ 138 1.951 0.491 138 12.4 4.2 138 2.142 0.235
Ьусориз еигораеиз 138 3.167 1.252 138 10.2 2.9 138 3.895 0.519
8а§Шапа за^Шг/оНа 138 4.078 0.930 138 2.5 1.7 138 5.968 0.596
8раг§апшт егесШт 138 2.198 0.676 138 15.2 4.6 138 4.889 0.466
В целом для сообщества
808 2.977 1.221 808 9.2 6.3 808 4.701 1.536

Глава 3. Г У М И Д Н А Я Ч А С Т Ь Г Р А Д И Е Н Т А 113
1 4
1 - - Д . - 2 • о - - 3
В сезонной динамике (рис. 3.4.1) интенсивность транспирации За^Шапа за^Иа/оНа была тесно связана (К' = 0.902) с изменениями влажности воздуха и дефицита водного насыщения в листьях растений. У остальных видов сезонные флуктуации скорости испарения воды возле средних уровней происходили, судя по всему, случайно: не удалось выявить связи этого показателя с другими элементами водного режима или факторами среды. Однако изменение какого-то фактора (предположительно снижение температуры воздуха) оказало заметное угнетающее влияние на скорость испарения воды у нескольких видов в отдельные дни наблюдений (рис. 3.4.1,6).
Дефицит водного насыщения в листьях растений был достаточно высоким для данных условий произрастания (табл. 3.4.1). Причем, наибольшие значения этого показателя (12-15%) отмечены у наименее интенсивно испарявших воду видов. Самые низкие средние величины водного дефицита (2.5-2.7%) выявлены для двух видов — АНзта р1апСа^о-адиа-Нса и За^Шапа за^НН/оИа. У этих же видов наблюдался диапазон самых низких типичных значений признака (0.4-5.0%). У остальных видов типичные минимумы изменялись от 5.5 (у ВШотиз итЬеИаШз) до 9.9% (у 8раг§атит егесШт), а типичные максимумы — от 14
{у Ьусориз еигораеиз) до 23.3%) (у ВШотиз итЬеНаШз). Подвижность признака была высока у АНзта р1апШ§о-адиаНса (коэффициент вариации равен 65.7%), Вшотиз итЬеНаШз (53.3%)) и За^Шапа за§Шг/оИа (69.6%). Для других видов подвижность дефицита водного насыщения оценивалась умеренными значениями коэффициентов вариации (28.3-34.4%).
В сезонной динамике колебания водного дефицита (рис. 3.4.2) были связаны с факторами среды у АНзта р1апШ§о-адиаНса (с дефицитом влажности воздуха и освещенностью; 0.981), ВШотиз итЬеПаШз (с освещенностью; Н = 0.856) и 5а§Шапа за^гШ/оНа (с дефицитом влажности воздуха и интенсивностью транспирации; = 0.932). Для остальных видов не выявлено связи колебаний данного показателя водного режима с какими-либо внутренними или внешними факторами.
22.07.96 29.07.96
Дни сезоне
05.08.96
Рис. 3.4.1. Сезонные ходы интенсивности транспирации растений остроосокового
сообщества:
а) 1 — АИзта р1ап1а§о-адиа11са, 2 — Ьусориз еигораеиз, 3 — 8а§Шапа за^ИИ/оНа; б) 4 — ВиЮтиз итЬеПаШз, 5 — Сагех асиШ, 6 —
8раг§ап1ит егес/ит.
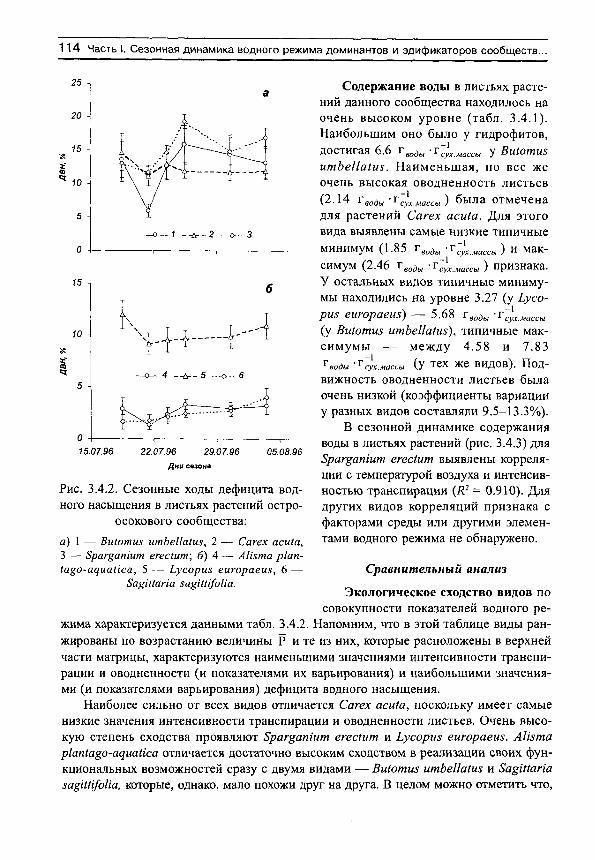
114 Часть I. С е з о н н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в .
25
20
15
10
5 -
• 1 2
15
10
-4 - - ^ - 5
О 15.07.96 22.07.96 29.07.96
Дни сезона
05.08.96
Рис. 3.4.2. Сезонные ходы дефицита водного насыщения в листьях растений остро
осокового сообщества:
а) 1 — Вшотт итЬеПаШз, 1 — Сагех асШа, 3 — 8раг§ашит егесШт; б) А — АНзта р1ап-(а^о-адиайса, 5 — Ьусориз еигораеиз, 6 —
8а^Шапа за^Шг/оИа.
Содержание воды в листьях растений данного сообщества находилось на очень высоком уровне (табл. 3.4.1). Наибольшим оно было у гидрофитов, достигая 6.6 г^„йы - г" ' сух.массы У ВиЮтив итЬеИаШз. Наименьшая , но все же очень высокая оводненность листьев (2.14 г , о й „ •г^з!^^^^^^) была отмечена для растений Сагех асиШ. Для этого вида выявлены самые низкие типичные
,-1 минимум (1.85 г^„д„ .г;: симум (2.46 г,„^„ г -1
сух.массы
.ссы) И мак-) признака.
симумы — Т воды ' Т сух.масс
У остальных видов типичные минимумы находились на уровне 3.27 (у Ьусориз еигораеиз) — 5.68 г.^^ь, '^с^ух.массы
(у Вшотиз итЬеНаШз), типичные мак-между 4.58 и 7.83 (у тех же видов). Под
вижность оводненности листьев была очень низкой (коэффициенты вариации у разных видов составляли 9.5-13.3%).
В сезонной динамике содержания воды в листьях растений (рис. 3.4.3) для 8раг§атит егесШт выявлены корреляции с температурой воздуха и интенсивностью транспирации {К^ = 0.910). Для других видов корреляций признака с факторами среды или другими элементами водного режима не обнаружено.
Сравнительный анализ
Экологическое сходство видов по совокупности показателей водного ре
жима характеризуется данными табл. 3.4.2. Напомним, что в этой таблице виды ранжированы по возрастанию величины Р и те из них, которые расположены в верхней части матрицы, характеризуются наименьшими значениями интенсивности транспирации и оводненности (и показателями их варьирования) и наибольшими значениями (и показателями варьирования) дефицита водного насыщения.
Наиболее сильно от всех видов отличается Сагех асиШ, поскольку имеет самые низкие значения интенсивности транспирации и оводненности листьев. Очень высокую степень сходства проявляют 8раг§апшт егесШт и Ьусориз еигораеиз. АНзта рШта^о-адиайса отличается достаточно высоким сходством в реализации своих функциональных возможностей сразу с двумя видами — ВиЮтиз итЪеИаШз и 8а§Шапа за§Ш{/оИа, которые, однако, мало похожи друг на друга. В целом можно отметить что.
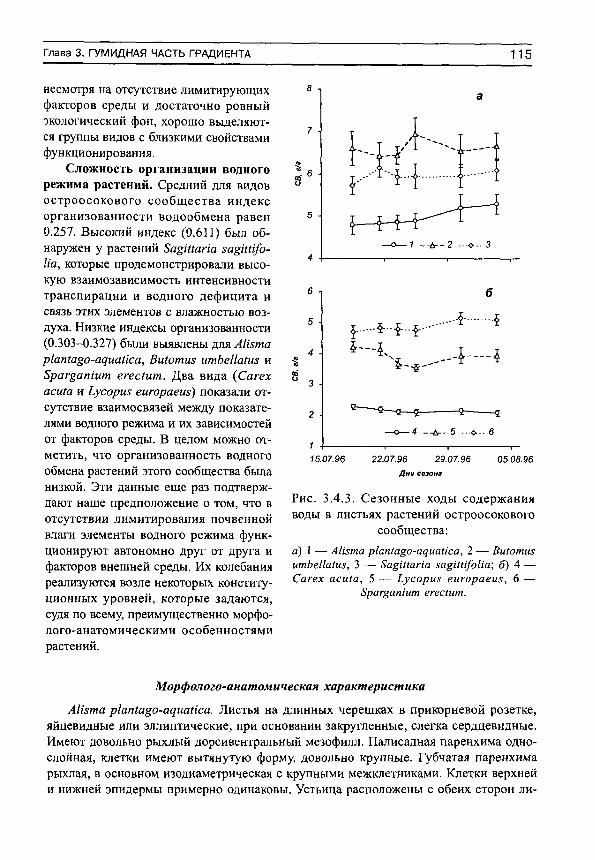
Глава 3, Г У М И Д Н А Я Ч А С Т Ь Г Р А Д И Е Н Т А 115
несмотря на отсутствие лимитирующих фаЕсгоров среды и достаточно ровный экологический фон, хорошо выделяются группы видов с близкими свойствами функционирования.
Сложность организации водного режима растений. Средний для видов остроосокового сообщества индекс организованности водообмена равен 0.257. Высокий индекс (0.611) был обнаружен у растений За^Шапа зафН/о-Иа, которые продемонстрировали высокую взаимозависимость интенсивности транспирации и водного дефицита и связь этих элементов с влажностью воздуха. Низкие индексы организованности (0.303-0.327) были выявлены для АНзта р1ап1а^о-адиаНса, ВШотиз итЬеНаШз и Зрагёатит егесШт. Два вида {Сагех асиШ и Ьусориз еигораеиз) показали отсутствие взаимосвязей между показателями водного режима и их зависимостей от факторов среды. В целом можно отметить, что организованность водного обмена растений этого сообщества была низкой. Эти данные еще раз подтверждают наше предположение о том, что в отсутствии лимитирования почвенной влаги элементы водного режима функционируют автономно друг от друга и факторов внешней среды. Их колебания реализуются возле некоторых конституционных уровней, которые задаются, судя по всему, преимущественно морфо-лого-анатомическими особенностями растений.
8 1
7 •
5 •
.1 . ^ - 2 - -о-.. 3
8"
{ - ^ ч - ^ -
б
......А 5
.4 - - Д - 5 6
15.07,96 22.07.96 29,0796
Дни сезона
05.08.96
Рис. 3.4.3. Сезонные ходы содержания воды в листьях растений остроосокового
сообщества:
а) 1 — АНзта р1ап(а§о-адиаНса, 2 — ВиСотиз итЬеИаШз, 3 — 8а§Шапа за§Ш1/оНа; б) 4 — Сагех аси(а, 5 — Ьусориз еигораеиз, 6 —
8раг§апшт егесШт.
Морфолого-анатомическая характеристика
АНзта р1апШ§о-адиаНса. Листья на длинных черешках в прикорневой розетке, яйцевидные или эллиптические, при основании закругленные, слегка сердцевидные. Имеют довольно рыхлый дорсивентральный мезофилл. Палисадная паренхима однослойная, клетки имеют вытянутую форму, довольно крупные. Губчатая паренхима рыхлая, в основном изодиаметрическая с крупными межклетниками. Клетки верхней и нижней эпидермы примерно одинаковы. Устьица расположены с обеих сторон ли-

116 Часть I. С е з о н н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в . .
Таблица 3.4.2 Матрица экологического сходства по совокупности элементов водного режима
растений остроосокового сообщества
Р
Сагех асиШ -1.806 -1.559 -2.042 -3.029 -3.589 -2.405
1.806 Зраг^апшт егесШт -0.066 -0.841 -1.462 -1.997 -0.512
1.559 0.066 Ьусориз еигораеиз -0.690 -1.465 -1.943 -0.495
2.042 0.841 0.690 ВШотиз итЬеИаШз -0.376 -0.798 0.480
3.029 1.462 1.465 0.376 АНзта
р1ап1а§о-адиаИса
-0.479 1.171
3.589 1.997 1.943 0.798 0.479 8а§Шапа за§1(Н/оИа 1.761
Примечание: Р .̂ — показатели экологического сходства между парами видов, Р — средний показатель экологического сходства каждого вида с остальными.
ста с крупными подустьичными полостями. В мезофилле листа имеются млечные ходы.
ВШотиз итЪеНаШз. Все листья прикорневые, с влагалищем, снизу трехгранные, выще — линейные. Мезофилл гомогенный, расположенный непосредственно под эпидермой, состоит из 4-6 слоев. Центральную часть листа занимают воздухоносные полости, разделенные однослойными тяжами паренхимных клеток. Сосудистые пучки расположены в мезофилле и в местах соединения тяжей. В районе сосудистых пучков в мезофилле находятся мелкие млечники. Клетки эпидермы мелкие, мало отличаются от мезофилла.
Сагех асШа. Листья жесткие, до 9 мм шириной. Мезофилл гомогенный. Клетки мезофилла расположены относительно плотно. Имеются крупные воздухоносные полости. Эпидерма однослойная, с довольно крупными клетками с нижней стороны. Тяжи механических тканей представлены с обеих сторон проводящего пучка, но не всегда образуют сплошные перемычки, пересекающие всю толщу листа.
Ьусориз еигораеиз. Листья с черешками, продолговатые, самые нижние рассечены на продолговатые доли. Имеется рыхлый дорсивентральный мезофилл. Палисадная паренхима однослойная, клетки имеют вытянутую форму, крупные. Губчатая паренхима рыхлая, в основном изодиаметрическая, с крупными межклетниками. Клетки верхней эпидермы заметно крупнее нижних. Устьица расположены с нижней стороны листьев.

Глава 3. ГУМИДНАЯ ЧАСТЬ ГРАДИЕНТА 117
Таблица 3.4.3 Парциальный объем тканей и свободных пространств листьев растений
остроосокового сообщества
Виды Э Пр ПО Пг Пс П Мг Мс М ВП АНзта р1ап1а§о- 15.1 1.1 3.1 26.3 17.0 43.2 27.7 9.0 36.8 0 адиаНса Вшотиз итЬеНаШз 3.0 8.3 8.1 8.3 0 8.3 2.6 0 2.6 61.1 Сагех асиШ 9.4 9.7 4.2 25.2 0 25.2 3.1 0 3.1 39.5 Ьусориз еигораеиз 24.5 0.7 3.4 10.8 24.1 34.9 21.6 14.9 36.5 0 За^Шапа за^НИ^оНа 18.6 0.8 2.3 15.2 18.4 33.6 26.9 16.3 43.3 0 5раг§атит егесШт 4.4 2.1 2.4 31.4 0 31.4 10.5 0 10.5 31.1
Примечание: Э — эпидермис, Пр — проводящие ткани, ПО — паренхимная обкладка пучков, Пг — губчатая паренхима, Пс — столбчатая паренхима, П — паренхима, Мг — межклетники губчатой паренхимы, Мс — межклетники столбчатой паренхимы, М — межклетники, ВП — воздушные полости.
8а§Шапа за§ИИ/оНа. Воздушные листья стреловидные или треугольно-стреловидные, острые, с расходящимися и примерно равными остальной части пластинки лопастями. Плавающие листья — эллиптические; погруженные — лентовидные, около 1 см шириной. Палисадная паренхима двухслойная, рыхлая, клетки имеют вытянутую форму. Эпидерма крупноклетная, клетки верхней эпидермы несколько крупнее нижних. Устьица расположены с обеих сторон, с большими подустьичными полостями. В мезофилле имеются млечные ходы.
Зраг^атит егесШт. Листья трехгранные, снизу по всей длине с сильно выдающимся острым килем. Мезофилл гомогенный, рыхлый, представлен клетками разной формы и размеров. Имеются довольно крупные воздухоносные полости. Сосудистые пучки расположены в верхней и нижней частях продольных перемычек, разделяющих воздухоносные полости. Механическая ткань представлена со стороны эпидермы от сосудистых пучков и отдельными тяжами в мезофилле. На границе перемычек и воздухоносных полостей расположены крупные тонкостенные безхлорофильные клетки неправильной формы. В районе перемычек имеются млечные ходы. Клетки эпидермы мелкие.
В листьях ряда видов {ВиЮтиз итЬеНаШз, Сагех асиШ и Зраг^апшт егесШт) имеются хорошо развитые воздушные полости (табл. 3.4.3). В данном случае точнее было бы назвать эти образования не воздушными полостями, а воздушными каналами, поскольку они хорошо отграничены от мезофилла и продолжаются на всем протяжении листьев. Развитие этих каналов оказывает сильное влияние на соотношение тканей и, вследствие этого, на одно из важнейших проявлений водного обмена растений — интенсивность транспирации.
Можно отметить, что у видов с хорошо развитыми воздушными полостями заметно (в несколько раз) уменьшается парциальный объем эпидермиса, достигая минимальных величин у ВиЮтиз итЬеИаШз и Зраг§апшт егесШт (рис. 3.4.4, а; табл. 3.4.3). Также в несколько раз уменьшается парциальный объем паренхимы (рис. 3.4.4, б),

118 Часть I. С е з о н н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в .
50 п
О 20 40 60 Парциальный объем воздушных полостей, %
г' = 0.939
20 40 60 80 Парциальный объем воздушных полостей, %
Рис. 3.4.4. Межвидовые соотношения парциальных объемов воздушных полостей с парциальными объемами эпидермиса (а), паренхимы (б), межклетников (в), пучков (проводящих тканей + обкладок) (г) в листьях растений остроосокового сообщества.
50 п
о 5 10 15 20 25 30 Парциальный объем эпидермиса, %
Рис. 3.4.5. Межвидовые соотношения между парциальными объемами эпидермиса и межклетников в листьях растений остроосокового сообщества.

Глава 3. Г У М И Д Н А Я Ч А С Т Ь Г Р А Д И Е Н Т А 119
50 п
40 -
30 -
I 20 -
10 -
2 4 6 8
Парциальный объем проводящих тканей, %
10
50
40
30
20 -
мезофилл становится очень плотным (особенно у ВиЮтиз итЬеПаШз и Сагех асиШ) (рис. 3.4.4, в; табл. 3.4.3), однако увеличивается относительное участие в сложении листьев проводящих пучков (рис. 3.4.4, г). Развитие воздушных полостей (каналов) создает видимость достаточно «ксероморфной» организации мезофилла (особенно если судить по плотности его сложения), однако большие размеры клеток, в целом листьев, их высокая оводненность свидетельствуют о значительной степени относительности такой оценки. Тем не менее, при составлении матрицы анатомического сходства (табл. 3.4.4), мы учитывали этот элемент строения листьев как усиливающий «ксероморф-ную» организацию листьев.
Анализ соотношений других элементов анатомической структуры листьев выявил достаточно тесную положительную корреляцию между парциальными объемами эпидермиса и межклетников (рис. 3.4.5). Увеличение относительной доли участия в сложении листьев проводящих тканей и пучков в целом (проводящих тканей и их обкладок) сопровождается уменьшением парциальных объемов межклетников (рис. 3.4.6, а) и паренхимы (рис. 3.4.6, б). При этом увеличение плотности мезофилла сопровождается существенным (почти в два раза) снижением скорости испарения воды (рис. 3.4.7, а), а также увеличением дефицита водного насыщения в листьях растений (рис. 3.4.7, б). Наконец, нельзя не отметить достаточно тесную статистическую связь парциальных объемов межклетников (для ВиЮтиз итЬеНаШз — сумма парциальных объемов межклетников и воздушных полостей) с оводненностью листьев (рис. 3.4.8).
Сравнение видов по совокупности показателей анатомической структуры листьев (эпидермиса, паренхимы, проводящих тканей, а также межклетников и воздушных полостей) (табл. 3.4.4) показывает, что Сагех асиШ и ВиЮтиз итЬеНаШз, расположенные в верхней части матрицы, заметно отличаются друг от друга и от всех остальных видов. Можно было бы говорить об их менее «мезоморфной» организации по сравнению с другими видами, однако это вряд ли уместно для растений, произрас-
I 10 - =0.763
10 15 20
Парциальный объем пучков, %
Рис. 3.4.6. Межвидовые соотношения парциальных объемов проводящих тканей и межклетников (а), пучков (проводящих тканей + обкладок пучков) и паренхимы (б) в листьях
растений остроосокового сообщества.

120 Часть I. С е з о н н а я д и н а м и к а водного р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в .
4,5
4,0
3,5 -
# 3,0 -
2,5 •
2,0 •
1,5
= 0.874
10 20 30 40 50
16
12
2"
4 •
=0.688
тающих в условиях с практически неограниченными запасами почвенной влаги. Отметим только, что основное отличие этих видов от других определяется очень плотным сложением мезофилла и развитием, з а н и м а ю щ и х большой объем, воздушных полостей.
Что касается индексов сходства для видов, расположенных в центре матрицы (табл. 3.4.4), то мы должны отметить, что при очень больших различиях в строении листьев (как в этом случае) данный метод расчетов, судя по всему, «не работает» и возникают ошибочные, не отражающие реального соотношения тканей листа, индексы. По отдельным показателям, а именно по соотношению парциальных объемов проводящих тканей и паренхимы можно найти определенное сходство между Ьусорш еигораеиз и Ба§Шапа за^НИ-/оИа. По соотношению количества межклетников выявляется сходство между АНзта р1ап1а^о-адиаИса и Ьусориз еигораеиз. Дальнейших примеров сходства привести невозможно. Поэтому нужно признать ограниченность метода расчета матриц сходства только исходными данными с небольшим разбросом величин. Если значения какого-либо признака различаются примерно на порядок, то нужно с большой осторожностью подходить к анализу полученных результатов.
Резюме. Для видов остроосокового сообщества характерны высокие значения интенсивности транспирации и оводненности листьев. Относительно невысокие значения этих показателей обнаружены только у растений Сагех асШа. Дефицит водного насыщения в листьях был высоким для данных условий произрастания у большинства видов. Только у АИзта р1ап1а§о-адиаИса и За^Шапа за§г1И/оИа наблюдались близкие к нулю величины этого признака. Организованность водного обмена растений этого сообщества была низкой. Эти данные еще раз подтверждают наше предположение о том, что в отсутствии лимитирования почвенной влаги в большинстве случаев элементы водного режима функционируют автономно друг от друга и факторов внешней среды. Отмечено, что, несмотря на отсутствие лимитирующих факторов среды и достаточно ровный экологический фон, хорошо выделяются группы
10 20 30 40 50
Парциальный объам межклетников, %
Рис. 3.4.7. Межвидовые соотношения между парциальным объемом межклетников и показателями водного режима (а — интенсивность транспирации, б — дефицит водного насыщения) в листьях растений
остроосокового сообщества.

Глава 3. Г У М И Д Н А Я ЧАСТЬ Г Р А Д И Е Н Т А 121
Таблица 3.4.4 Матрица анатомического сходства по совокупности показателей анатомической
структуры листьев растений остроосокового сообщества
Р
Сагех асиШ -1.580 -3.953 -5.760 -6.675 -6.123 -4.818
1.580 Вшотиз итЬеНаШз -0.990 -2.211 -5.586 -2.389 -1.919
3.953 0.990 8раг§атит егесШт -0.336 -2.295 -1.965 0.069
5.760 2.211 0.336 АНзта
р1апШ§о-адиаНса
-0.163 -1.152 1.399
6.675 5.586 2.295 0.163 Ьусориз еигораеиз
-0.711 2.802
6.123 2.389 1.965 1.152 0.711 8а^Шапа за§Ш1/оИа 2.468
видов с близкими свойствами функционирования. Анализ анатомической структуры листьев показал, что у видов с хорошо развитыми воздушными полостями заметно уменьшается парциальный объем эпидермиса, в несколько раз уменьшается парциальный объем паренхимы, мезофилл становится очень плотным, увеличивается относительное участие в сложении листьев проводящих пучков. Выявлена корреляция между парциальными объемами эпидермиса и межклетников. Увеличение относительной доли участия проводящих тканей и пучков в целом коррелирует с уменьшением парциальных объемов межклетников и паренхимы. Увеличение плотности мезофилла сопровождается существенным снижением скорости испарения воды. Показана статистическая связь парциальных объемов межклетников с оводненностью листьев.
о 10 20 30 40 50 60 70 Парциальный объем межклетников, %
Рис. 3.4.8. Корреляция между парциальным объемом межклетников (для ВиЮтиз итЪе1-1а1из — сумма парциальных объемов межклетников и воздушных полостей) и оводненностью листьев растений остроосокового
сообщества.

Ч А С Т Ь II
ДНЕВНАЯ ДИНАМИКА ВОДНОГО РЕЖИМА ДОМИНАНТОВ И ЭДИФИКАТОРОВ
СООБЩЕСТВ В РАЗНЫХ ЧАСТЯХ ГРАДИЕНТА ВЛАЖНОСТИ ПОЧВЫ
Вопросу о дневной динамике элементов водного режима растений в литературе уделяется большое внимание. Например, форма дневных колебаний интенсивности транспирации рядом исследователей связывается с экологической приуроченностью растений, уровнем увлажнения экотопа или, в более широком плане, со степенью оптимальности условий произрастания. Очевидно, что этот вопрос можно рассматривать с двух сторон, которые тесно связаны друг с другом. С одной стороны, дневные колебания интересуют исследователей тем, что они в той или иной степени отражают ход основных факторов среды (освещенности, температуры и влажности воздуха), свидетельствуя при этом об адаптационных возможностях растений. С другой стороны, многие авторы рассматривают собственно форму дневных колебаний как свидетельство того или иного состояния (напряженности или сбалансированности) водного режима в определенных условиях произрастания. Иными словами, подход к вопросу о характере дневных колебаний направлен на оценку состояния водного режима растений в конкретной экологической обстановке.
Здесь следует сказать о степени взаимосвязи этих двух сторон одного явления. Можно предположить, что форма дневных колебаний элементов водного режима во многом обусловлена функциональным состоянием всего растения, которое определяется многообразными физиологическими изменениями в ответ на изменения факторов среды. В таком случае степень зависимости дневных колебаний элементов водного режима от колебаний факторов среды должна также, в конечном итоге, определяться функциональным состоянием растения как целого. Но нельзя исключить и обратного синхронизирующего влияния факторов среды на дневные ритмы элементов водного режима.
Рассмотрим некоторые литературные данные о зависимости дневных колебаний интенсивности транспирации (поскольку чаще всего именно этот показатель являет-

123
ся предметом изучения данного вопроса) от дневного хода основных факторов среды. По данным Л. А. Иванова с соавторами (1953), зависимость интенсивности транспирации от температуры в большей степени проявляется в экотопах с высокой влажностью почвы. Ян Баочжень (1960), изучая водный режим эдификаторов сухих степей Центрального Казахстана, установил, что дневная динамика интенсивности транспирации всех изученных видов зависит от многих экологических факторов, особенно влажности и температуры воздуха. К такому же выводу пришел Ж. Ж. Жатканба-ев (1961), который изучал водный режим эдификаторов сообществ пустынных степей Центрального Казахстана. В. М. Свешникова (1962) показала высокую степень зависимости интенсивности транспирации от температуры у растений высокогорных пустынь Памира.
Однако, по данным К. Р. Витко (1963), в гырнецовой дубраве не наблюдается четкой связи между дневным ходом транспирации у изученных видов и изменением метеорологических факторов, что, по ее мнению, обусловлено низким содержанием доступной влаги в почве. А. А. Горшкова (1970), изучая водный режим растений Забайкалья, пришла к выводу, что дневные колебания транспирации часто не согласуются с ходом метеорологических факторов, а определяются внутренней регулировкой отдачи воды растениями. Это, по мнению автора, свидетельствует о том, что степные растения (даже в сезоны с максимальным количеством осадков для данной местности) испытывают значительные затруднения в водоснабжении. В субальпийском и субнивальном поясах Большого Кавказа, по данным Г. Ш. Нахуцришвили (1972), обычно наблюдается корреляция между дневным ходом транспирации и факторов среды, хотя имели место и некоторые несоответствия. По мнению А. П. Савинкина (1973), связь между дневным ходом транспирации и метеофакторов обусловлена увлажнением экотопа. При достаточном увлажнении эта связь выражается в параллельном ходе транспирации и метеофакторов; при недостаточном увлажнении, напротив, такая синхронность утрачивается. Л. Д. Копытова (1974) также указывает на то, что при недостаточном увлажнении дневные колебания транспирации у растений в степях Забайкалья часто не согласуются с изменениями метеорологических факторов. Л. Н. Алексеенко (1976) на основании собственных и литературных данных поддерживает мысль о том, что в условиях ограниченного водоснабжения корреляционная зависимость между интенсивностью транспирации и факторами среды ниже, чем при оптимальном водоснабжении. Г. Сабиров (1982), изучавший водный режим растений Устюрта, показал, что почти у всех видов при резко выраженном недостатке влаги в почве и воздухе нарушается связь дневного хода транспирации с метеорологическими факторами. По мнению Т. К. Горышиной (1979) при обильном и достаточном водоснабжении ход транспирации в целом следует дневным изменениям метеорологических факторов. При недостаточном водоснабжении эта связь нарушается, что может быть вызвано усилением внутренней регуляторной деятельности растений (Иванов и др., 1953; Горшкова, 1971; Нахуцришвили, 1972; Алексеенко, 1976).
Что касается формы дневных колебаний, то для растений гумидной зоны характерна, как правило, одновершинная кривая с максимумом в околополуденное время. У растений аридных районов обычно наблюдаются многовершинные кривые или кривые с максимумом в ранние утренние часы (подробный литературный обзор этих данных содержится в работах: Свешникова, 1962, 1979; Алексеенко, 1976). Однако

124 Часть II. Д н е в н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в . . .
Л. Н. Алексеенко (1976) отмечает, что для луговых растений характерны самые разные формы кривых интенсивности транспирации. То же отмечает и Г. Ш. Нахуцрищ-вили (1972). В условиях гумидного климата Большого Кавказа у 23% исследованных видов он наблюдал одновершинные, у 55% — двух-, а у 22% — трехвершинные кривые дневного хода транспирации. Сходные данные содержатся в работе Н. И. Тулаш-вили (1976).
На основании собственных, а также многочисленных литературных данных Г. Ш. Нахуцришвили (1972) приходит к выводу, что нельзя определенно сказать, какого типа кривая транспирации характерна для различных климатических областей. Этот автор полагает, что двухвершинность и многовершинность кривых транспирации у изучавшихся им видов растений свидетельствует не о нарушении типичного хода транспирации, а об активном регулировании водного режима этими видами.
Некоторая неопределенность в обсуждении этих вопросов заставила нас в более широком объеме, чем это делалось ранее, применить количественные методы для исследования дневной динамики элементов водного режима растений. Была поставлена задача выяснить, от чего зависит: 1) положение акрофаз (время, в которое функция принимает максимальные значения) в дневных ритмах транспирации; 2) форма дневных колебаний транспирации, водного дефицита и оводненности листьев; 3) сила связи дневных ритмов этих элементов водного режима с факторами среды.
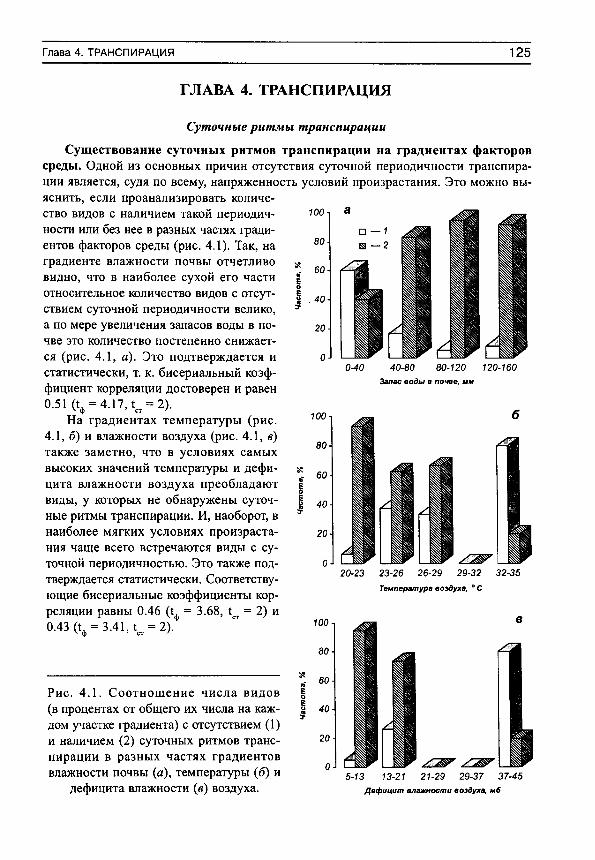
Глава 4. Т Р А Н С П И Р А Ц И Я 125
ГЛАВА 4. ТРАНСПИРАЦИЯ
100
80
60
40
20
Суточные ритмы транспирации
Существование суточных ритмов транспирации на градиентах факторов среды. Одной из основных причин отсутствия суточной периодичности транспирации является, судя по всему, напряженность условий произрастания. Это можно выяснить, если проанализировать количество видов с наличием такой периодичности или без нее в разных частях градиентов факторов среды (рис. 4.1). Так, на градиенте влажности почвы отчетливо видно, что в наиболее сухой его части относительное количество видов с отсутствием суточной периодичности велико, а по мере увеличения запасов воды в почве это количество постепенно снижается (рис. 4.1, а). Это подтверждается и статистически, т. к. бисериальный коэффициент корреляции достоверен и равен 0.51(1^ = 4.17,1^ = 2).
На градиентах температуры (рис. 4.1, б) и влажности воздуха (рис. 4.1, в) также заметно, что в условиях самых высоких значений температуры и дефицита влажности воздуха преобладают виды, у которых не обнаружены суточные ритмы транспирации. И, наоборот, в наиболее мягких условиях произрастания чаще всего встречаются виды с суточной периодичностью. Это также подтверждается статистически. Соответствующие бисериальные коэффициенты корреляции равны 0.46 (1ф = 3.68, I = 2) и 0.43(1 =3.41,1 = 2 ) .
0-»0 40-80 80-120 120-160 Запас воды в почве, мм
100
80
60
40
20
20-23 23-26 26-29 29-32
Температура воздуха, ** С
32-35
100
Рис. 4 .1 . Соотношение числа видов (в процентах от общего их числа на каждом участке градиента) с отсутствием (1) и наличием (2) суточных ритмов транспирации в разных частях градиентов влажности почвы (а), температуры (б) и
дефицита влажности (в) воздуха.
а-
80
60
40
20
5-13 13-21 21-29 29-37 37-45 Дефицит влажности воздуха, мб
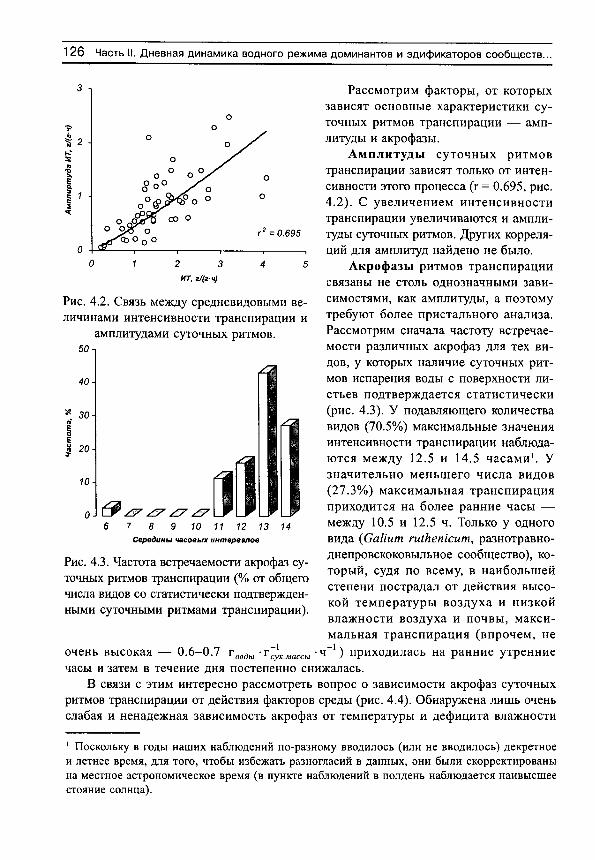
126 Часть II. Д н е в н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в . .
Рис. 4.2. Связь между средневидовыми величинами интенсивности транспирации и
амплитудами суточных ритмов. 50 л
40-
20
10
0-1 ^ ^ ^ ^ ^ 6 7 8 9 10 11 12 13
Середины часовых интервалов
14
Рис. 4.3. Частота встречаемости акрофаз суточных ритмов транспирации (% от общего числа видов со статистически подтвержденными суточными ритмами транспирации).
очень высокая — 0.6-0.7 г . „-1
Рассмотрим факторы, от которых зависят основные характеристики суточных ритмов транспирации — амплитуды и акрофазы.
Амплитуды суточных ритмов транспирации зависят только от интенсивности этого процесса (г= 0.695, рис. 4.2). С увеличением интенсивности транспирации увеличиваются и амплитуды суточных ритмов. Других корреляций для амплитуд найдено не было.
Акрофазы ритмов транспирации связаны не столь однозначными зависимостями, как амплитуды, а поэтому требуют более пристального анализа. Рассмотрим сначала частоту встречаемости различных акрофаз для тех видов, у которых наличие суточных ритмов испарения воды с поверхности листьев подтверждается статистически (рис. 4.3). У подавляющего количества видов (70.5%) максимальные значения интенсивности транспирации наблюдаются между 12.5 и 14.5 часами ' . У значительно меньшего числа видов (27.3%) максимальная транспирация приходится на более ранние часы — между 10.5 и 12.5 ч. Только у одного вида {СаИит гШкетсит, разнотравно-днепровскоковыльное сообщество), который, судя по всему, в наибольшей степени пострадал от действия высокой температуры воздуха и низкой влажности воздуха и почвы, максимальная транспирация (впрочем, не
ч""') приходилась на ранние утренние ^ воды ^ сух.массы
часы и затем в течение дня постепенно снижалась. В связи с этим интересно рассмотреть вопрос о зависимости акрофаз суточных
ритмов транспирации от действия факторов среды (рис. 4.4). Обнаружена лишь очень слабая и ненадежная зависимость акрофаз от температуры и дефицита влажности
' Поскольку в годы наших наблюдений по-разному вводилось (или не вводилось) декретное и летнее время, для того, чтобы избежать разногласий в данных, они были скорректированы на местное астрономическое время (в пункте наблюдений в полдень наблюдается наивысшее стояние солнца).
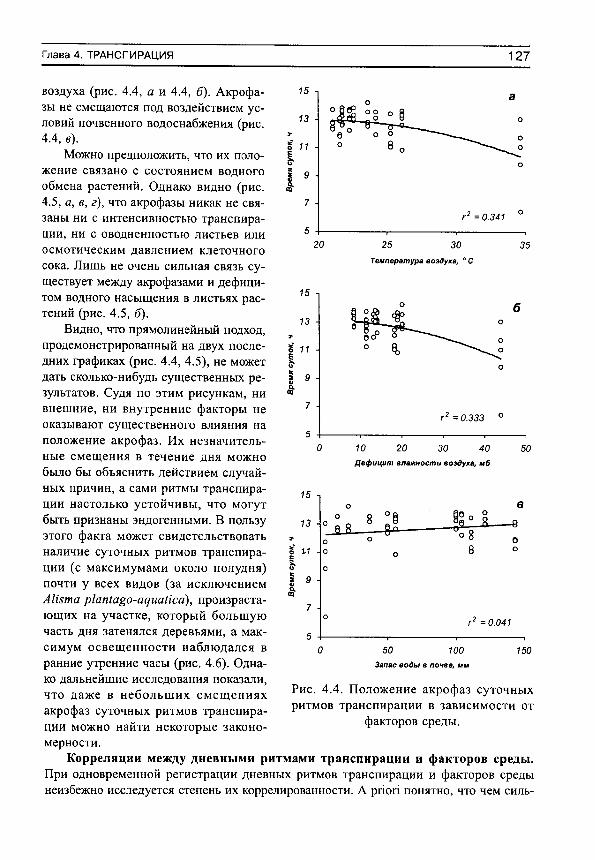
Глава 4 . Т Р А Н С П И Р А Ц И Я 127
15 1
13
8" 11
г' =0.341
20 25 30
Температура воздуха, ° С
15 1
13 -
о
—*в-
8" 11 •
1 00° о
° ^
7 •
.ч -
= 0.333
10 20 30 40
Дефицит влажности воздуха, мб
15
13 Р о 8
35
воздуха (рис. 4.4, а и 4.4, б). Акрофазы не смещаются под воздействием условий почвенного водоснабжения (рис. 4.4, в).
Можно предположить, что их положение связано с состоянием водного обмена растений. Однако видно (рис. 4.5, а, в, г), что акрофазы никак не связаны ни с интенсивностью транспирации, ни с оводненностью листьев или осмотическим давлением клеточного сока. Лищь не очень сильная связь существует между акрофазами и дефицитом водного насыщения в листьях растений (рис. 4.5, б).
Видно, что прямолинейный подход, продемонстрированный на двух последних графиках (рис. 4.4, 4.5), не может дать сколько-нибудь существенных результатов. Судя по этим рисункам, ни внешние, ни внутренние факторы не оказывают существенного влияния на положение акрофаз. Их незначительные смещения в течение дня можно было бы объяснить действием случайных причин, а сами ритмы транспирации настолько устойчивы, что могут быть признаны эндогенными. В пользу этого факта может свидетельствовать наличие суточных ритмов транспирации (с максимумами около полудня) почти у всех видов (за исключением АНзта р1ап1а§о-адиаНса), произрастающих на участке, который большую часть дня затенялся деревьями, а максимум освещенности наблюдался в ранние утренние часы (рис. 4.6). Однако дальнейшие исследования показали, что даже в небольших смещениях акрофаз суточных ритмов транспирации можно найти некоторые закономерности.
Корреляции между дневными ритмами транспирации и факторов среды. При одновременной регистрации дневных ритмов транспирации и факторов среды неизбежно исследуется степень их коррелированности. А рпоп понятно, что чем силь-
50
=0.041
50 100 150
Запас воды в почве, мм
Рис. 4.4. Положение акрофаз суточных ритмов транспирации в зависимости от
факторов среды.

1 28 Часть II. Д н е в н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в . .
2 ИТ, г/(гч)
г' = 0.052
15
13
11
9
7
5
о о
оо
^^ =0.022
4
СВ, г/г
9
7 •
5
О
= 0.365
20 40
ДВН, %
60 80
15
13
11
9
7
5 =0.030
ОД, МПа
Рис. 4.5. Положение акрофаз суточных ритмов транспирации в зависимости от средневидовых величин водного режима растений.
5 т -г 50
3 -•
2 -•
12 13 14 15
Время суток, ч
Рис. 4.6. Усредненные дневные ходы освещенности (7) и интенсивности транспирации видов остроосокового
сообщества.
1 — АНзта р1ап1а§о-адиаИса, 2 — ВШотиз итЬеНаШз, 3 — Сагех асШа, 4 — Ьусориз еигораеиз, 5 — 8а§Шапа за^шуоНа,
6 — 8раг§апшт егесШт.

Глава 4. Т Р А Н С П И Р А Ц И Я 129
нее коррелированны ритмы транспирации и факторов среды, тем меньше они различаются по форме и больше совпадают по времени. Поэтому возникает вопрос о влиянии этой коррелированности на положение акрофаз ритмов транспирации.
У большинства видов интенсивность транспирации тесно коррелированна с факторами среды (рис. 4.7). Достоверные коэффициенты корреляции с температурой воздуха проявляются у 57% видов, с дефицитом влажности воздуха — у 53%, с освещенностью — у 67%). Таким образом, у большинства растений транспирация в своей дневной динамике достаточно тесно связана с тем или иным фактором среды (только 8 видов демонстрируют отсутствие такой связи). Можно предположить, что сила такой связи влияет на положение акрофаз транспирации как синхронизирующий фактор. Оказалось, что чем сильнее корреляция интенсивности транспирации с температурой и дефицитом влажности воздуха, тем позже наступает дневной максимум транспирации (рис. 4.8, а, б). Влияние дневных ритмов освещенности на ритмы транспирации иное. При отрицательных фазовых сдвигах между этими ритмами (т. е. когда акрофазы транспирации наблюдались позже акрофаз освещенности в соответствии с выражением Аф = Фф^^^р - Ф^ранспираци») акрофазы транспирации наступали тем раньше (ближе к полудню), чем сильнее этот процесс был коррелирован с освещенностью (рис. 4.8, в-2). При положительных фазовых сдвигах между ритмами транспирации и освещенности акрофазы транспирации также стремились к полудню, но в этом случае процесс смещения акрофаз транспирации был менее определенным (рис. 4.8, в-1), чем в первом случае (рис. 4.8, в-2).
Таким образом видно, что дневные ритмы факторов среды являются мощными синхронизирующими агентами для интенсивности транспирации. Причем действие их направлено в разные стороны. Температура и дефицит влажности воздуха смещают акрофазы транспирации на более поздние (от полудня) часы дня, а освещенность «подтягивает» их к полудню (рис. 4.8). Но, судя по всему, разные виды не одинаково реагируют на факторы среды. Для одних более сильным «синхронизатором» является температура (и/или дефицит влажности) воздуха, для других — освещенность.
Это можно отчетливо видеть на рисунке, где одновременно показано положение акрофаз транспирации и факторов среды для всех изученных видов растений (рис. 4.9). Акрофазы транспирации у большинства видов находятся в достаточно узком (примерно 3 ч) коридоре между акрофазами температуры воздуха и освещенности. По положению акрофаз транспирации видно, что для одних видов более мощным синхронизирующим агентом является температура воздуха, для других — освещен-
40-.
-0,7 -0,5 -0,3 -0,1 0,1 0,3 0,5 0,7 0,9 Середины интервалов коэффициентов
корреляции
Рис. 4.7. Гистограмма распределения коэффициентов корреляции между дневными ритмами транспирации и
факторами среды:
1 — температурой воздуха, 2 — дефицитом влажности воздуха, 3 — освещенностью.
5 - 2353

130 Часть II. Д н е в н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в .
15 п
13 •
11 •
ность, а некоторые виды занимают некоторое среднее положение, находясь под одинаково сильным влиянием как температуры воздуха, так и освещенности.
Несколько видов имеют акрофазы транспирации, наступающие раньше акрофаз факторов среды (рис. 4.9). Особенно сильно это проявляется в самом засушливом (во время проведения наблюдений) экотопе, в котором произрастало разнотравно-днеп-ровскоковыльное сообщество. Выше было показано (рис. 2, в, раздел «Определения»), что такое раннее наступление максимумов интенсивности транспирации связано с де
прессией этого процесса на протяжении большей части дня. Поэтому можно предположить, что раннее наступление акрофаз транспирации (раньше акрофаз освещенности) является признаком недостаточно благоприятных условий произрастания, возникающих либо в результате плохого водоснабжения растений, либо из-за напряженных конкурентных отношений между видами сообщества.
Это явление было отмечено ранее другими исследователями. В. М. Свешникова (1979) пишет, что если растения находятся в неблагоприятных условиях водоснабжения, то активная транспирация приурочивается к утренним часам и дневной ход имеет вид одновершинной, резко ниспадающей кривой. При оптимальных условиях водообеспеченности максимальная интенсивность транспи-
=0.814
-1,0 -0,5 0,0 0,5 1,0
15 1
13 -
11 •
9 •
7 •
5
=0.763
-1,0
16 1
14 •
I 12 •
I 'О
8
6
-0,5 0,0 0,5 1.0
Г = 0.456
0,2 0,4 0,6 0,8 Коэффициенты корреляции
1,0
Рис. 4.8. Положение акрофаз суточных ритмов транспирации в зависимости от коррелированности этих ритмов с ритмами температуры (а), дефицита влажности (б) воздуха и освещенности (в: 1 — при положительных фазовых сдвигах между ритмами транспирации и освещенности, 2 — при отрицательных фазовых сдвигах; не приведены данные для видов остроосокового сообщества, поскольку максимум освещенности там приходился на 9 часов утра из-за затенения
участка деревьями).
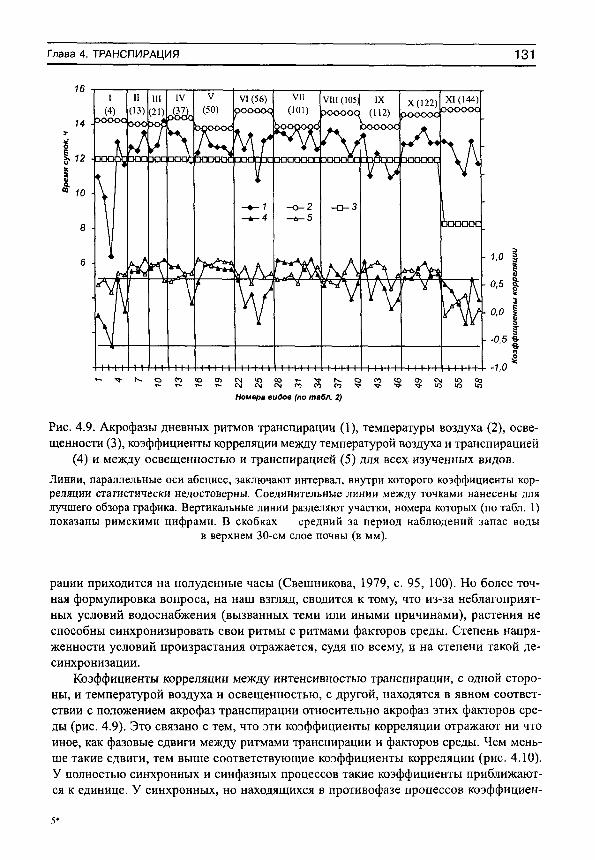
Глава 4. Т Р А Н С П И Р А Ц И Я 131
16
» - ' Ч - К . О < 0 < О 0 > С ^ 1 О С 0 г - Ч - Г ^ . О С 0 < О О > Г М Ю 0 0
Номера видов (по табл. 2)
Рис. 4.9. Аьфофазы дневных ритмов транспирации (1), температуры воздуха (2), освещенности (3), коэффициенты корреляции между температурой воздуха и транспирацией
(4) и между освещенностью и транспирацией (5) для всех изученных видов.
Линии, параллельные оси абсцисс, заключают интервал, внутри которого коэффициенты корреляции статистически недостоверны. Соединительные линии между точками нанесены для лучшего обзора графика. Вертикальные линии разделяют участки, номера которых (по табл. 1) показаны римскими цифрами. В скобках — средний за период наблюдений запас воды
в верхнем 30-см слое почвы (в мм).
рации приходится на полуденные часы (Свешникова, 1979, с. 95, 100). Но более точная формулировка вопроса, на наш взгляд, сводится к тому, что из-за неблагоприятных условий водоснабжения (вызванных теми или иными причинами), растения не способны синхронизировать свои ритмы с ритмами факторов среды. Степень напряженности условий произрастания отражается, судя по всему, и на степени такой де-синхронизации.
Коэффициенты корреляции между интенсивностью транспирации, с одной стороны, и температурой воздуха и освещенностью, с другой, находятся в явном соответствии с положением акрофаз транспирации относительно акрофаз этих факторов среды (рис. 4.9). Это связано с тем, что эти коэффициенты корреляции отражают ни что иное, как фазовые сдвиги между ритмами транспирации и факторов среды. Чем меньше такие сдвиги, тем выше соответствующие коэффициенты корреляции (рис. 4.10). У полностью синхронных и синфазных процессов такие коэффициенты приближаются к единице. У синхронных, но находящихся в противофазе процессов коэффициен-
5*
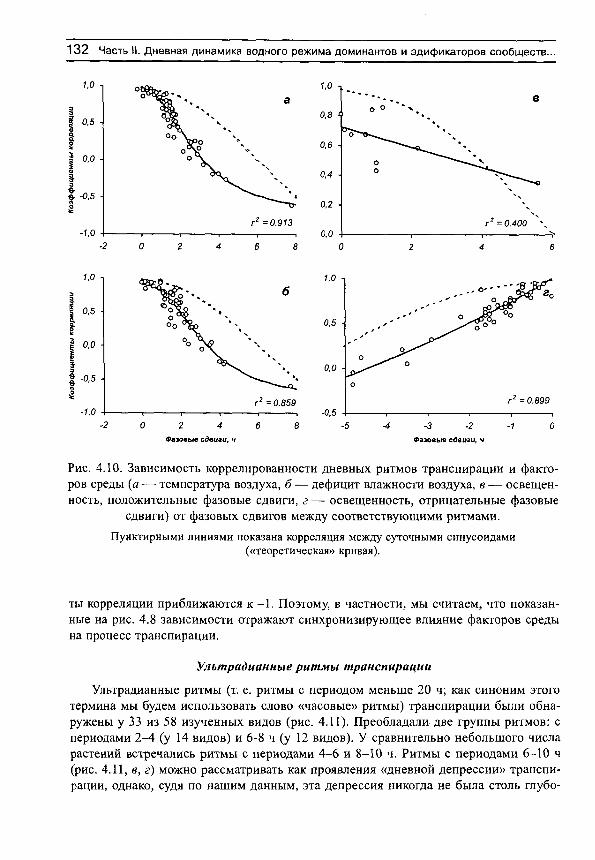
132 Ч а с т ь II. Д н е в н а я д и н а м и к а в о д н о г о р е ж и м а д о м и н а н т о в и э д и ф и к а т о р о в с о о б щ е с т в . .
-2 0 2 4 6 8 -5 -4 -3 -2 -1 0 Фазовые сдвиги, ч Фазовые сдвиги, ч
Рис. 4.10. Зависимость коррелированности дневных ритмов транспирации и факторов среды (а — температура воздуха, б — дефицит влажности воздуха, в — освещенность, положительные фазовые сдвиги, г — освещенность, отрицательные фазовые
сдвиги) от фазовых сдвигов между соответствующими ритмами.
Пунктирными линиями показана корреляция между суточными синусоидами («теоретическая» кривая).
ты корреляции приближаются к - 1 . Поэтому, в частности, мы считаем, что показанные на рис. 4.8 зависимости отражают синхронизирующее влияние факторов среды на процесс транспирации.
Улыпрадианные ритмы транспирации
Ультрадианные ритмы (т. е. ритмы с периодом меньше 20 ч; как синоним этого термина мы будем использовать слово «часовые» ритмы) транспирации были обнаружены у 33 из 58 изученных видов (рис. 4.11). Преобладали две группы ритмов: с периодами 2-4 (у 14 видов) и 6-8 ч (у 12 видов). У сравнительно небольшого числа растений встречались ритмы с периодами 4-6 и 8-10 ч. Ритмы с периодами 6-10 ч (рис. 4.11, в, г) можно рассматривать как проявления «дневной депрессии» транспирации, однако, судя по нашим данным, эта депрессия никогда не была столь глубо-
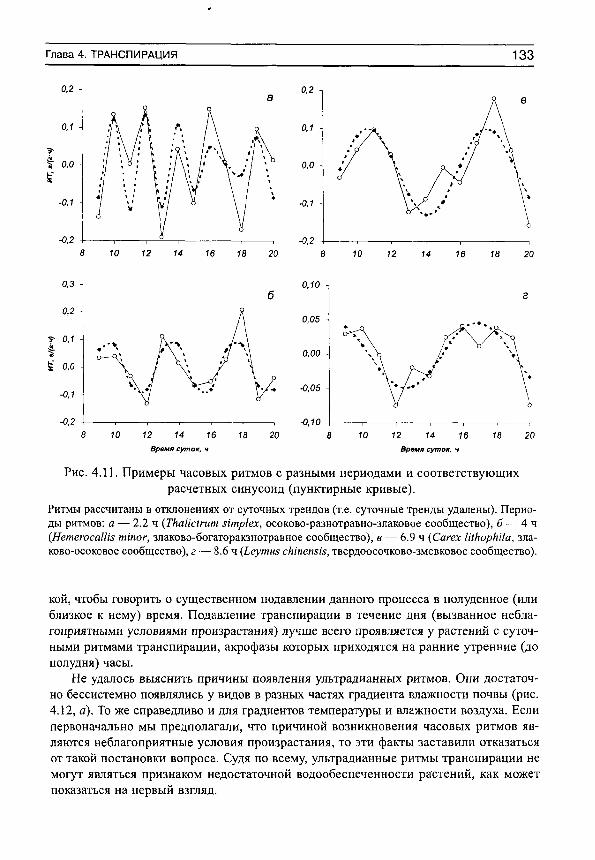
Глава 4. ТРАНСПИРАЦИЯ 133
0,2 -, 0,2 -|
8 10 12 14 16 18 20 8 10 12 14 16 18 20
-0,2 -1 , , , , , , -0,10 -\ ^ , , , , б 10 12 14 16 18 20 8 10 12 14 16 18 20
Время суток, ч Время суток, ч
Рис. 4.11. П р и м е р ы часовых р и т м о в с р а з н ы м и периодами и с о о т в е т с т в у ю щ и х расчетных синусоид (пунетирные кривые) .
Ритмы рассчитаны в отклонениях от суточных трендов (т.е. суточные тренды удалены). Периоды ритмов: а — 2.2 ч {ТкаНс1гит зтрЫх, осоково-разнотравно-злаковое сообщество), б— 4 ч (НетегосаШз ттог, злаково-богаторакзнотравное сообщество), в — 6.9 ч {Сагех ШкоркИа, зла-ково-осоковое сообщество), г — 8.6 ч {Ьеутиз сктепз1з, твердоосочково-змевковое сообщество).
кой, чтобы говорить о существенном подавлении данного процесса в полуденное (или близкое к нему) время . Подавление транспирации в течение дня (вызванное неблагоприятными условиями произрастания) л у ч ш е всего проявляется у растений с суточными ритмами т ран спирации , акрофазы которых приходятся на р а н н и е утренние (до полудня) часы.
Не удалось выяснить п р и ч и н ы появления ультрадианных р и т м о в . О н и достаточно бессистемно появлялись у видов в разных частях градиента влажности почвы (рис. 4.12, а). То же справедливо и для градиентов температуры и влажности воздуха. Если первоначально м ы предполагали , что причиной возникновения часовых р и т м о в являются неблагоприятные условия произрастания , то эти факты заставили отказаться от такой постановки вопроса . Судя по всему, ультрадианные р и т м ы т р а н с п и р а ц и и не могут являться признаком недостаточной водообеспеченности р а с т е н и й , как м о ж е т показаться на первый взгляд.
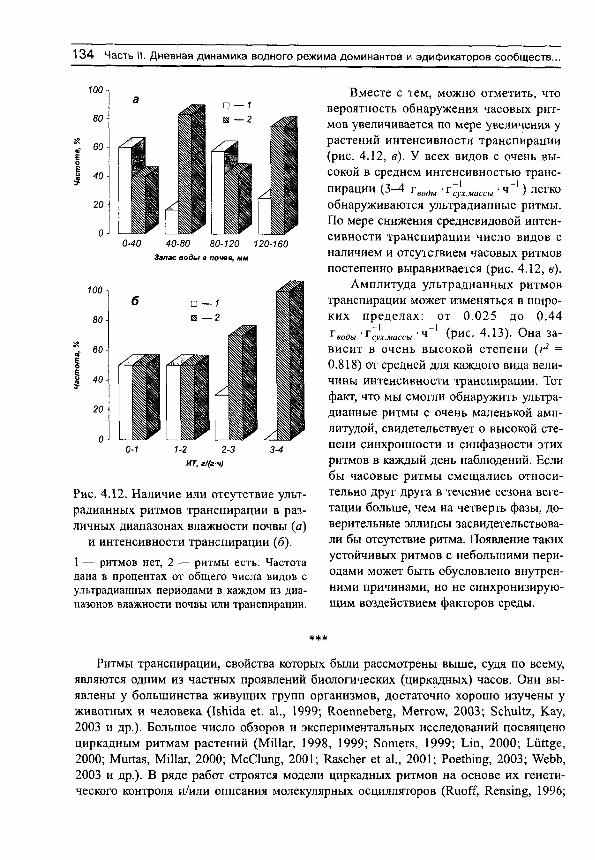
134 Часть И. Дневная динамика водного режима доминантов и эдификаторов сообществ.
100
ИТ, г/(гч)
Рис . 4.12. Н а л и ч и е или отсутствие ультрадианных р и т м о в т р а н с п и р а ц и и в раз личных диапазонах в л а ж н о с т и почвы (а)
и интенсивности т р а н с п и р а ц и и (б).
1 — ритмов нет, 2 — ритмы есть. Частота дана в процентах от общего числа видов с ультрадианных периодами в каждом из диапазонов влажности почвы или транспирации.
В м е с т е с т е м , м о ж н о отметить , что вероятность о б н а р у ж е н и я часовых ритмов увеличивается по мере увеличения у р а с т е н и й и н т е н с и в н о с т и т р а н с п и р а ц и и (рис . 4.12, в ) . У всех видов с очень высокой в среднем и н т е н с и в н о с т ь ю трансп и р а ц и и (3-4 г,„й„ •г:у,,^ассы-^'^) легко о б н а р у ж и в а ю т с я у л ь т р а д и а н н ы е р и т м ы . П о мере снижения средневидовой интенс и в н о с т и т р а н с п и р а ц и и ч и с л о в и д о в с наличием и отсутствием часовых ритмов постепенно выравнивается (рис. 4.12, в).
А м п л и т у д а у л ь т р а д и а н н ы х р и т м о в транспирации может изменяться в ш ир о к и х п р е д е л а х : о т 0.025 д о 0.44
^воды-^сух.массы-'^'^ ( р и с 4.13). О н а за
в и с и т в о ч е н ь в ы с о к о й с т е п е н и ( г ' = 0.818) от средней для каждого вида велич и н ы и н т е н с и в н о с т и т р а н с п и р а ц и и . Тот факт, что м ы смогли о б н а р у ж и т ь ультрад и а н н ы е р и т м ы с очень маленькой амплитудой, свидетельствует о высокой степени с и н х р о н н о с т и и с и н ф а з н о с т и этих ритмов в каждый день наблюдений. Если б ы ч а с о в ы е р и т м ы с м е щ а л и с ь о т н о с и тельно друг друга в т е ч е н и е сезона вегетации больше , чем на четверть фазы, доверительные эллипсы засвидетельствовали бы отсутствие ритма. Появление таких устойчивых р и т м о в с н е б о л ь ш и м и периодами может быть обусловлено внутренними пр ичинами , но не с и н х р о н и з и р у ю щ и м воздействием факторов среды.
***
Ритмы транспирации , свойства которых бьши рассмотрены в ы ш е , судя по всему, являются одним из ч а с т н ы х проявлений биологических (циркадных) часов . Они выявлены у б о л ь ш и н с т в а ж и в у щ и х групп организ мо в , д о с т а т о ч н о х о р о ш о и з у ч е н ы у ж и в о т н ы х и ч е л о в е к а ( Ь Ы й а е1. а1., 1999; К.оеппеЬег§, Меггочу, 2003; З с Ь и к г , Кау , 2003 и др.) . Большое число обзоров и экспериментальных исследований п о с в я щ е н о ц и р к а д н ы м р и т м а м р а с т е н и й (МШаг, 1998, 1999; З о т е г з , 1999; Ып, 2000; ЬйИ^е , 2000; Мийая , МШаг, 2000; МсС1ипд, 2001; КазсЬег е1 а1., 2001; Рое1Ып§, 2003; \УеЬЬ, 2003 и др.) . В р я д е работ строятся модели циркадных р и т м о в на основе их генетического контроля и/или описания молекулярных осцилляторов (Киой', Кеиз ш ^ , 1996;

Глава 4. ТРАНСПИРАЦИЯ 135
0,4 -
^-- 0,3 •
ч 0,1 •
0,0 = 0.818
2 ИТ, г/(гч)
Рис. 4.13. З а в и с и м о с т ь амплитуды ультрадианных ритмов от средневидовых величин
интенсивности транспирации .
ЗЫпЬго!, ЗсагЬгоиеЬ, 1999; Ь е т а е1 а1., 0,5 т 2000). П р е д п р и н и м а ю т с я п о п ы т к и с о здания новой категории моделей динамики популяций, в которой комбинирую т с я у р а в н е н и я Л о т к и - В о л ь т е р р а и уравнения биологических осцилляторов (Ва1(1о, 2002). П о с л е д н е е о б с т о я т е л ь ство подчеркивает значимость предпол о ж е н и я о т о м , ч т о ц и р к а д н ы е ч а с ы , прогнозируя и з м е н е н и я с р е д ы и коорд и н и р у я ф и з и о л о г и ч е с к и е п р о ц е с с ы , увеличивают приспособленность растений , их в ы ж и в а е м о с т ь и, в к о н е ч н о м итоге , р е п р о д у к т и в н ы й у с п е х (\УеЬЬ, 2003).
Циркадн ые ч а с ы являются сигнальной системой, с в я з ы в а ю щ е й изменения среды и физиологические п р о ц е с с ы организма ( 8 о т е г 5 , 1999). О д н и м из о с н о в н ы х периодических факторов среды, н а с т р а и в а ю щ и х эту систему, является свет. Э н е р г и я света непосредственно контролирует р е а к ц и ю (движения) устьиц п у т е м воздействия на рецепторные с и с т е м ы в их з а м ы к а ю щ и х клетках (Каххег, Карреп , 1997). Таким образом, потери воды растением , связанные с д в и ж е н и е м у с т ь и ц , н е и з б е ж н о д о л ж н ы иметь суточную периодичность . Существует предположение, что циркадный контроль движения устьиц увеличивает эффективность использования воды ( о т н о ш е н и е ассимиляции к испарению) растениями (\УеЬЬ, 2003). У растений циркадные часы, стимулируют открытие у с т ь и ц утром, в течение ранней части фотопериода , когда осве щенность высока, а температура воздуха е щ е не достигла своего максиму ма . В послеполуденное время , когда м о ж н о ожидать в ы с о к у ю температуру воздуха и высокие потери воды из-за т р а н с п и р а ц и и , ц и р к а д н ы е ч а с ы а к т и в и з и р у ю т з а к р ы т и е у с т ь и ц (имеется в виду не л а в и н о о б р а з н ы й , а п о с т е п е н н ы й , м е д л е н н ы й пр о цесс ) . Это приведет к у в е л и ч е н и ю фотосинтеза при ограничении т р а н с п и р а ц и и и, следовательно , к увеличению эффект и вности использования воды.
Комплексное воздействие о с в е щ е н н о с т и и темпер ат у р ы воздуха на р и т м ы транспирации хорошо в и д н о на рис . 4.8 и 4.10. А к р о ф а з ы суточных р и т м о в т р а н с п и р а ц и и сдвигаются под воздействием р и т м о в температуры воздуха и о с в е щ е н н о с т и («затягиваются» этими факторами среды) . П р и ч е м их действие имеет п р о т и в о п о л о ж н о е направление: о с в е щ е н н о с т ь сдвигает р и т м ы т р а н с п и р а ц и и ближе к п о л у д н ю (рис . 4.8, а), а температура воздуха — к более позднему времени (рис . 4.8, в). М о ж н о предположить вслед за А. А. К. \ \ 'еЬЬ (2003), что о с в е щ е н н о с т ь оказывает с т и м у л и р у ю щ е е влияние на р и т м ы т р а н с п и р а ц и и в первую половину дня , а т е м п е р а т у р а воздуха — угнетающее после полудня.
Можно показать, что разные в и д ы по-разному реаг ир у ю т н а комплексное воздействие факторов среды. Р и т м ы т р а н с п и р а ц и и некоторых видов (причем часто в одних и тех же экотопах) в б о л ь ш е й степени «затягиваются» р и т м а м и т е м п е р а т у р ы воздуха (их акрофазы почти совпадают с акрофазами температуры) , тогда как р и т м ы дру-

136 Часть II. Дневная динамика водного режима доминантов и эдификаторов сообществ...
гих растений преимущественно следуют за ритмами освещенности (рис. 4.9). Это, как можно предположить , является отражением специфических особенностей временной организации ф ун кционирования , характерных для р а з н ы х видов растений .
П одавляю щее большинство изученных растений поддерживает акрофазы ритмов транспирации в очень узком диапазоне значений — всего 2-3 часа (рис . 4.9), почти точно м е ж д у а к р о ф а з а м и р и т м о в температуры воздуха и о с в е щ е н н о с т и . Это может свидетельствовать как о в ы с о к о й э ф ф е к т и в н о с т и с и н х р о н и з а ц и и ф а к т о р а м и среды ритмов т ран сп и рации , так и об эндогенности этих р ит мо в , ра звившихся и поддерживающихся в течение длительного периода эволюции растений .
У некоторых видов акрофазы ритмов транспирации наблюдались р ань ш е акрофаз освещенности (рис . 4.9). Это может свидетельствовать , как у же б ы л о в ы ш е отмечено, о неблагоприятных (по тем или иным причинам) для этих растений условиях произрастания. В этом случае растения не могут н о р м а л ь н о реализовать р и т м ы т р анс пирации , а с и н х р о н и з а ц и я этих р и т м о в с р и т м а м и о с в е щ е н н о с т и я в н о затруднена (рис. 4.8, в-\, рис . 4.10, в ) . Т а к и м о б р а з о м понятно , что н е б л а г о п р и я т н ы е у сло вия произрастания оказывают отрицательное влияние не только на и н т е н с и в н о с т ь функционирования , но и на его в р е м е н н ю организацию.
Высокочастотные (ультрадианные) р и т м ы т р а н с п и р а ц и и ч а щ е наблюдаются у растений с наиболее в ы с о к и м и з н а ч е н и я м и скорости и с п а р е н и я воды с п о в е р х н о с т и листьев (рис . 4.12). А рпог ! мы предполагали , что появление высокочастотных ритмов является от ражением ухудшения экологической обстановки для р а с т е н и й . Однако факты з а с т а в л я ю т сделать п р я м о п р о т и в о п о л о ж н ы й в ы в о д — в ы с о к о ч а с т о т н ы е р и т м ы являются р е а л и з а ц и е й нормального ф у н к ц и о н и р о в а н и я и п р и у х у д ш е н и и условий произрастания не проявляются . Судя по всему, эти р и т м ы т а к ж е являются эндогенным п роц ессом и подавляются низкой в л а ж н о с т ь ю почвы и/или высокой температурой воздуха.
Резюме . П о к а з а н а в о з м о ж н о с т ь в ы я в л е н и я с у т о ч н ы х и у л ь т р а д и а н н ы х р и т м о в транспирации с п о м о щ ь ю п р о г р а м м ы « С о х т о г 2.4» (Ьир://ехсе151а1.пе\ута11.ги). П р о демонстрированы характерные ф о р м ы кривых, свойственные большинству изученных видов. Н а гради ентах температуры и в л а ж н о с т и воздуха о б н а р у ж е н о , что в условиях самых высоки х значений температуры и д е ф и ц и т а в л а ж н о с т и воздуха преобладают виды, у которых отсутствуют суточные р и т м ы т р а н с п и р а ц и и . И, наоборот, в наиболее мягких у с л о в и я х произрастания ч а щ е всего вст р ечаю т ся в и д ы с суточной периодичностью. У п о д а в л я ю щ е г о большинства видов (71%) м а к с и м а л ь н ы е значения и н т е н с и в н о с т и т р а н с п и р а ц и и н а б л ю д а ю т с я м е ж д у 12 и 15 ч а с а м и . У з начит ель но меньшего количества растений (27%) максимальная т р а н с п и р а ц и я приходится на более ранние часы — м е ж д у 10.5 и 12.5 ч. А к р о ф а з ы т р а н с п и р а ц и и не с м е щ а ю т с я под воздействием а б с о л ю т н ы х величин у с л о в и й почвенного в о д о с н а б ж е н и я или других факторов среды. П о л о ж е н и е акрофаз т р а н с п и р а ц и и не связано и с состоянием водного обмена растений . Однако сам процесс т р а н с п и р а ц и и достаточно т е с н о связан с тем или и н ы м ф актором среды. Ч е м сильнее корреляция и н т е н с и в н о с т и т р анспир а ции с т емп ерат урой и д е ф и ц и т о м влажности воздуха, т е м позже наступает дневно й максимум т р а н с п и р а ц и и . Усиление коррелированности п р о ц е с с а т р а н с п и р а ц и и с осв е щ е н н о с т ь ю с м е щ а е т а к р о ф а з ы испарения воды к полудню. А к р о ф а з ы транспирации у большинства видов находятся в достаточно узком (примерно 3 ч) коридоре меж-

Глава 4. ТРАНСПИРАЦИЯ 137
ду акрофазами температуры воздуха и освещенности . П о п о л о ж е н и ю акрофаз транспирации видно , что для одних видов более м о щ н ы м с и н х р о н и з и р у ю щ и м агентом яв ляется температура воздуха, для других — о с в е щ е н н о с т ь , а некоторые в и д ы занимают некоторое среднее положение, находясь под одинаково сильным влиянием как температуры воздуха, т а к и освещенности . Раннее наступление акр о фаз т р а н с п и р а ц и и (раньше акрофаз освещенности) является признаком недостаточно благоприятных условий произрастания , в о з н и к а ю щ и х л и б о в результате плохого водоснабжения растений, либо из-за н а п р я ж е н н ы х конкурентных о т н о ш е н и й м е ж д у в и д а м и со о бщ ест ва . Ультрадианные р и т м ы транспирации были о б н а р у ж е н ы у 33 из 58 и з у ч е н н ы х видов . Преобладали две группы р и т м о в : с периодами 2-4 (у 14 видов) и 6-8 ч (у 12 видов) . У сравнительно н е б о л ь ш о г о числа растений встречались р и т м ы с п е р и о д а м и 4-6 и 8-10 ч. Не удалось выяснить п р и ч и н ы появления этих р и т м о в . О н и до ст ат о чно бессистемно появлялись у в и д о в в ра зных частях градиентов факт о р о в среды. П р е д п о ложено, что высокочастотные р и т м ы являются реализацией нормального функционирования и при ухудшении у с л о в и й произрастания не проявляются .
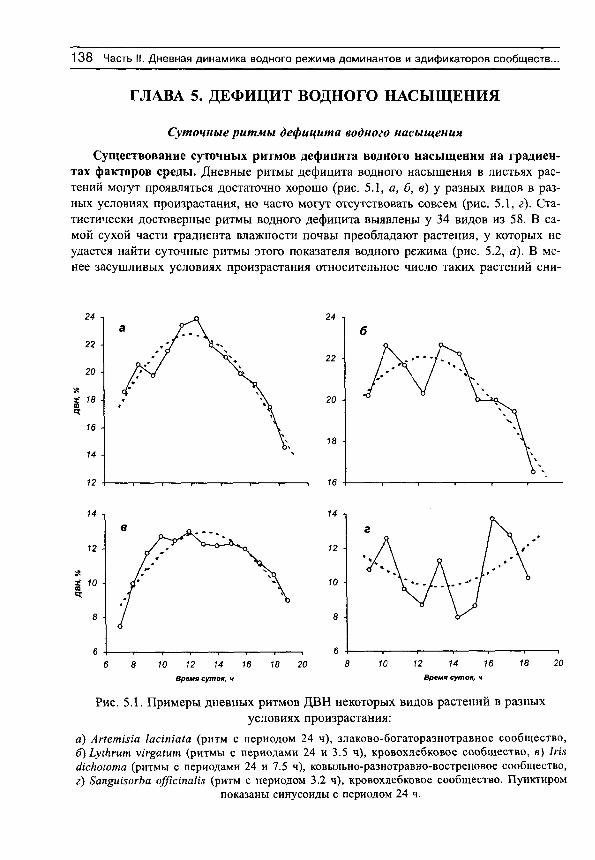
138 Часть II. Дневная динамика водного режима доминантов и эдификаторов сообществ..
ГЛАВА 5. ДЕФИЦИТ ВОДНОГО НАСЫЩЕНИЯ
Суточные ритмы дефицита водного насыщения
Существование суточных ритмов дефицита водного н а с ы щ е н и я на градиентах факторов среды. Д н е в н ы е р и т м ы д е ф и ц и т а водного н а с ы щ е н и я в листьях растений могут проявляться достаточно хорошо (рис. 5.1, а, б, в) у разных видов в разных условиях произрастания , но часто могут отсутствовать совсем (рис. 5.1, г). Статистически достоверные р и т м ы водного дефицита выявлены у 34 видов из 58. В самой сухой части градиента в л а ж н о с т и почвы пр ео бладаю т р а с т е н и я , у которых не удается найти суточные р и т м ы этого показателя водного р е ж и м а (рис. 5.2, а). В менее з асушливых условиях произрастания относительное число т а к и х растений сни-
24
22
20
16
14 -
12
14
12
5" ' °
8 -
10 12 14 Время суток, ч
16 18 20
24 -,
22 •
20
18
16
14
12
10
10 12 14 16 Время суток, ч
18 20
Рис . 5.1. П р и м е р ы д н е в н ы х р и т м о в Д В Н некоторых в и д о в р а с т е н и й в р а з н ы х условиях произрастания :
а) АНетша 1ас1та1а (ритм с периодом 24 ч), злаково-богаторазнотравное сообщество, б) Ьу(Нгит у/г^оГит (ритмы с периодами 24 и 3.5 ч), кровохлебковое сообщество, в) Мз (ИсНоЮта (ритмы с периодами 24 и 7.5 ч), ковыльно-разнотравно-вострецовое сообщество, г) 8ап§и150гЬа о^гстаНз (ритм с периодом 3.2 ч), кровохлебковое сообщество. Пунктиром
показаны синусоиды с периодом 24 ч.

Глава 5. ДЕФИЦИТ ВОДНОГО НАСЫЩЕНИЯ 139
0-40 40-80 80-120 120-160 Запас воды в почве, мм
23-26 26-29 29-32 32-35 Теипература воздуха, оС
Р и с . 5.2. С о о т н о ш е н и е ч и с л а в и д о в (в процентах от общего их числа на каждом интервале градиента) с отсутствием (1) или н а л и ч и е м (2) с у т о ч н ы х р и т м о в дефицита водного н а с ы щ е н и я в р а з н ы х частях градиентов в л а ж н о с т и п о ч в ы (а)
и температуры воздуха (б) .
30
25
г °
• 15
^ 10
5
О 2-4 4-6 6-8 8-10 Интервалы амплитуд ДВН, %
Р и с . 5.3. Г и с т о г р а м м а р а с п р е д е л е н и я а м п л и т у д с у т о ч н ы х р и т м о в д е ф и ц и т а
водного н а с ы щ е н и я .
10 1
8 •
I" ^ б
I 4
=0.196
10 20 ДВН, У.
30 40
Рис . 5.4. Связь м е ж д у амплит у дами деф и ц и т а водного н а с ы щ е н и я и средневи-д о в ы м и з н а ч е н и я м и показателя у видов с выявленной суточной периодичностью.
жается, и преобладают виды с хорошо в ы р а ж е н н ы м и суточными р и т м а м и процесса (рис. 5.2, а). В наиболее у в л а ж н е н н ы х экотопах отно сит ель но е количество видов с отсутствием или наличием таких ритмов выравнивается . На градиенте температуры воздуха отчетливо проявляется тенденция уменьшения числа в и д о в с н а л и ч и е м суточной периодичности водного д е ф и ц и т а по мере увеличения среднедневных температур воздуха (рис. 5.2, б). Эта тенденция подтверждается статистически (бисериаль-ный коэффициент корреляции равен 0.52, 1^ = 4.3, 1^ = 2). Это м о ж е т указывать на то, что сочетание высокой температуры воздуха с низкой в л а ж н о с т ь ю почвы приводит к подавлению суточной периодичности дефицита водного насы щ ения . Наиболее часто эта периодичность проявляется в достаточно г у м и д н ы х условиях произрастания с относительно н е в ы с о к и м и среднедневными т е м п е р а т у р а м и воздуха.
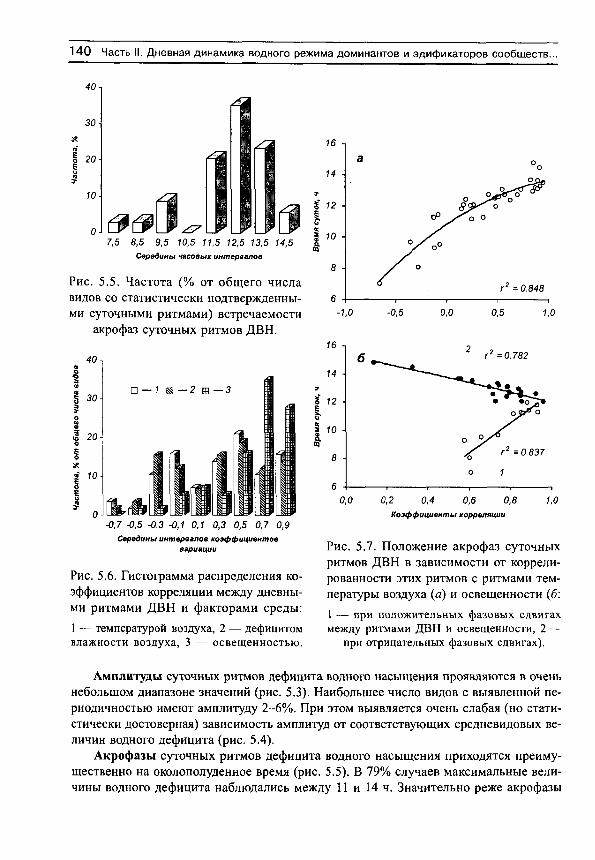
140 Часть II. Дневная динамика водного режима доминантов и эдификаторов сообществ.
40
30
20
10
7,5 8,5 9,5 70,5 11,5 12,5 13,5 14,5 Середины часовых интервалов
Р и с . 5.5. Ч а с т о т а ( % от о б щ е г о ч и с л а видов со статистически подтвержденными суточными р и т м а м и ) встречаемости
акрофаз суточных р и т м о в Д В Н .
40
5 30
20
10
16
14 -
8" 12 •
I к
I 8
б
•0,7 -0,5 -0,3 -0,1 0,1 0,3 0,5 0,7 0,9 Середины интервалов коэффициентов
вариации
Рис. 5.6. Гистограмма распределения коэффициентов корреляции между дневными р и т м а м и Д В Н и ф а к т о р а м и с р е д ы :
I — температурой воздуха, 2 — дефицитом влажности воздуха, 3 — освещенностью.
-1,0
16 т
14
10
8 -
= 0.848
-0,5 0,0 0.5 1,0
=0.782
г' = 0.837
0,0 0,2 0,4 0,6 0,8 Коэффициенты корреляции
1,0
Рис . 5.7. П о л о ж е н и е а к р о ф а з с у т о ч н ы х ритмов Д В Н в з а в и с и м о с т и от коррелированности этих р и т м о в с р и т м а м и температуры воздуха (а) и о свещ енно ст и (б:
1 — при положительных фазовых сдвигах между ритмами ДВН и освещенности, 2 —
при отрицательных фазовых сдвигах).
Амплитуды суточных р и т м о в дефицита водного н а с ы щ е н и я проявляются в очень небольшом диапазоне значений (рис. 5.3). Н а и б о л ь ш е е число видов с в ы я в л е н н о й периодичностью и м е ю т амплитуду 2-6%. П р и этом выявляется очень слабая (но статистически достоверная) зависимость амплитуд от соответствующих средневидовых величин водного д е ф и ц и т а (рис . 5.4).
А к р о ф а з ы с у т о ч н ы х р и т м о в д е ф и ц и т а водного н а с ы щ е н и я приходятся преимущественно на околополуденное время (рис. 5.5). В 79% случаев м а к с и м а л ь н ы е величины водного д е ф и ц и т а наблюдались между 11 и 14 ч. З н а ч и т е л ь н о р е ж е акр о фаз ы

Глава 5. ДЕФИЦИТ ВОДНОГО НАСЫЩЕНИЯ 141
о К;
5 -X
о 1 1 1 1 1 1 1 1 1 1 1 1 1 т т т т г г т т т т 1 1 1 1 1 1 1 1 1 1
Номера видов по табл. 2
Рис. 5.8. Акрофазы дневных ритмов Д В Н (1 — суточные р и т м ы выявлены, 2 — суточных ритмов нет) , температуры воздуха (3) и освещенности (4) для всех изученных видов.
Соединительные линии между точками нанесены для лучшего обзора графика.
ритмов этого показателя наблюдались в утренние часы, и л и ш ь в отдельных случаях после 14 ч.
Д н е в н ы е р и т м ы д е ф и ц и т а водного н а с ы щ е н и я у м н о г и х в и д о в к о р р е л и р у ю т с дневными р и т м а м и факторов среды. Наиболее часто (у 63% видов) о бнар у живаю т ся статистически достоверные корреляции д н е в н ы х ходов водного д е ф и ц и т а и освещенности (рис. 5.6). В 30% случаев в дневной д и н а м и к е проявляются согласованные изменения этого показателя водного р е ж и м а с т емперат у р о й и д е ф и ц и т о м в л а ж н о с т и воздуха. У подавляющего большинства видов (83%), таким образом, выявлены подобные корреляции с одним или несколькими факторами среды.
Чем сильнее корреляция дневных ходов элемента водного р е ж и м а и фактора среды, тем м е н ь ш е фазовый сдвиг м е ж д у этими ходами. Э т и м о бу сло влено с и н х р о н и зирующее влияние у с л о в и й произрастания на водный о б м е н растений . Температура воздуха «затягивает» д н е в н ы е р и т м ы водного д е ф и ц и т а т а к и м образом, что акрофазы этих р и т м о в с м е щ а ю т с я с у т р е н н и х на околополуденные ч а с ы (рис . 5.7, а). Осве щенность воздействует на р и т м ы водного д е ф и ц и т а так , что а к р о ф а з ы этих р и т м о в перемещаются ближе к п о л у д н ю (рис . 5.7, б). Это наблюдается как в случае отрицательных фазовых сдвигов м е ж д у р и т м а м и водного д е ф и ц и т а и о с в е щ е н н о с т и , т а к и в случае положительных фазовых сдвигов. Однако эффективность синх р о низ ир у ю щ его воздействия факторов на р и т м ы д е ф и ц и т а водного н а с ы щ е н и я з амет но м е н ь ш е , чем на р и т м ы т р а н с п и р а ц и и (ср . рис . 4.9 и 5.8). Только у 20 в и д о в (со статистичес ки п о д т в е р ж д е н н ы м и с у т о ч н ы м и р и т м а м и ) а к р о ф а з ы водного д е ф и ц и т а находятся внутри «коридора» акрофаз температуры воздуха и о с в е щ е н н о с т и . Вне этого «коридора» находятся акрофазы водного д е ф и ц и т а 14 видов . Эти р а з л и ч и я м е ж д у т р анс -пирацией и д е ф и ц и т о м водного н а с ы щ е н и я могут быть о б у с л о в л е н ы т е м , что р и т м ы транспирации в большей степени контролируются внешними факторами, а р и т м ы водного дефицита — в н у т р е н н и м и .
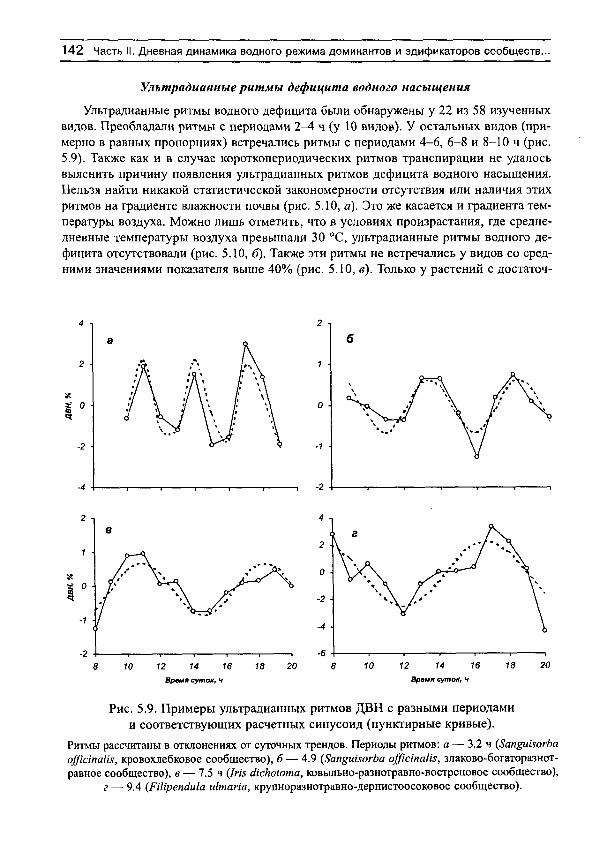
142 Часть II. Дневная динамика водного режима доминантов и эдификаторов сообществ..
Ультрадианные ритмы дефицита водного насыщения
Ультрадианные р и т м ы водного дефицита были обнаружены у 22 из 58 изученных видов. Преобладали р и т м ы с периодами 2-4 ч (у 10 видов) . У о с т а л ь н ы х видов (примерно в р а в н ы х пропорциях) встречались р и т м ы с периодами 4-6 , 6-8 и 8-10 ч (рис. 5.9). Также как и в случае короткопериодических р и т м о в т р а н с п и р а ц и и не удалось выяснить п р и ч и н у появления ультрадианных р и т м о в д е ф и ц и т а водного н а с ы щ е н и я . Нельзя найти никакой статистической закономерности отсутствия или н а л и ч и я этих ритмов на градиенте влажности почвы (рис. 5.10, а). Это же касается и градиента температуры воздуха. М о ж н о л и ш ь отметить , что в условиях произрастания , где среднедневные т е м п е р а т у р ы воздуха п р е в ы ш а л и 30 °С, ультрадианные р и т м ы водного де фицита отсутствовали (рис. 5.10, б). Также эти р и т м ы не встречались у видов со средними значениями показателя в ы ш е 40% (рис. 5.10, в). Только у р а с т е н и й с достаточ-
4 т 2
-2 -1 , , , , , , -6 -I , , , , . . 8 10 12 14 16 18 20 8 10 12 14 16 18 20
Время суток, ч Время суток, ч
Рис . 5.9. П р и м е р ы ультрадианных р и т м о в Д В Н с р а з н ы м и п е р и о д а м и и соот ветствующих расчетных синусо ид ( п у н к т и р н ы е к р и в ы е ) .
Ритмы рассчитаны в отклонениях от суточных трендов. Периоды ритмов: а — 3.2 ч {5ап^150гЬа о//1стаНз, кровохлебковое сообщество), б — 4.9 {8ап§ш50гЬа о^юшаИз, злаково-богаторазнотравное сообщество), в — 7.5 ч ( / т сИсНо1ота, ковыльно-разнотравно-вострецовое сообщество),
г — 9.4 (РШрепМа и1тапа, крупноразнотравно-дернистоосоковое сообщество).

Глава 5. ДЕФИЦИТ ВОДНОГО НАСЫЩЕНИЯ 143
НО умеренным развитием водного дефицита могли быть о б н а р у ж е н ы короткоперио-дические ритмы процесса . Это может служить указанием на то , что ультрадианные р и т м ы являются п р е и м у щ е с т в е н н о атрибутом нормального функционирования растений.
Резюме. С т ат и стически д о с т о в е р н ы е суточные р и т м ы водного д е ф и ц и т а выявлены у 34 видов из 58. На градиенте температуры воздуха отчетливо проявляется тенденция у м е н ь ш е н и я числа видов с наличием суточной п е р и о д и ч н о с т и водного дефицита по мере увеличения среднедневных температур воздуха . Это может указывать на то , что сочетание высокой температуры в о з д у х а с н и з к о й в л а ж н о с т ь ю почвы п р и в о д и т к п о д а в л е н и ю суточной п е р и о д и ч н о с т и д е ф и ц и т а водного н а с ы щения . А к р о ф а з ы суточных ритмов д е ф и цита водного н а с ы щ е н и я приходятся преимущественно на околополуденное время. В 79% случаев м а к с и м а л ь н ы е в е л и ч и н ы водного д е ф и ц и т а н аблюдались между 11 и 14 ч. Д н е в н ы е р и т м ы д е ф и ц и т а водного насыщения у многих видов коррелируют с д н е в н ы м и р и т м а м и факторов среды. Ч е м си льн ее к о р р е л я ц и я д н е в н ы х ходов элемента водного р е ж и м а и фактора сред ы , т е м м е н ь ш е ф а з о в ы й с д в и г м е ж д у этими ходами. Температура воздуха «затягивает» дневные р и т м ы водного дефицита таким образом, что акрофазы этих ритмов смещаются с у т р е н н и х н а околополуденные часы. О с в е щ е н н о с т ь воздействует на ритмы водного д е ф и ц и т а так, что акрофазы этих р и т м о в п е р е м е щ а ю т с я б л и ж е к полудню. Ультрадианные р и т м ы водного д е ф и ц и т а б ы л и о б н а р у ж е н ы у 22 из 58 изученных в и д о в . П р е о б л а д а л и р и т м ы с периодами 2-4 ч. Н е в ы я в л е н ы п р и ч и н ы появления ультрадианных ритмов , однако факты указывают на то, что эти р и т м ы появляются п р е и м у щ е с т в е н н о у н о р м а л ь н о ф у н к ц и о н и р у ю щ и х растений .
20-23 23-26 26-29 29-32 32-35 Температура воздуха, " С
0-20 20-^0 40-60 60-80 ДВН, %
Рис. 5.10. Н аличие или отсутствие ультрадианных р и т м о в Д В Н в р аз личны х диапазонах влажности почвы (а), температуры воздуха (б) , д е ф и ц и т а водного
н а с ы щ е н и я в листьях растений (в) .
Частота дана в процентах от общего числа видов в каждом из диапазонов. 1 — ритмов
нет, 2 — ритмы есть.

144 Часть II. Дневная динамика водного режима доминантов и эдификаторов сообществ..
ГЛАВА 6. СОДЕРЖАНИЕ ВОДЫ В ЛИСТЬЯХ
Суточные ритмы содержания воды в листьях
С у щ е с т в о в а н и е с у т о ч н ы х р и т м о в с о д е р ж а н и я в о д ы в л и с т ь я х р а с т е н и й на градиентах ф а к т о р о в среды. Статистически достоверные суточные р и т м ы оводненности листьев б ы л и о б н а р у ж е н ы у 30 из 58 изученных видов . Х а р а к т е р н о й особенностью этих р и т м о в (в отличие от ритмов т р а н с п и р а ц и и и д е ф и ц и т а водного насыщения) является то , что в дневное время наблюдается не м а к с и м у м (акрофаза) , а минимум (батифаза) ф у н к ц и и (рис. 6.1). Поэтому, п р и м е н и т е л ь н о к р и т м а м оводненности листьев , м ы будем анализировать не акрофазы, а батифазы процесса .
На градиенте в л а ж н о с т и почвы число видов с с у т о ч н ы м и р и т м а м и с о д е р ж а н и я воды в листьях п р и м е р н о р а в н о числу видов , не и м е ю щ и х т а к и х р и т м о в . Только в
'.7 п 2,0 -I
12 14 Время суток, ч
12 14 Время суток, ч
Рис . 6.1. П р и м е р ы д н е в н ы х ритмов содержания воды в л и с т ь я х некоторых видов р а с т е н и й в ра зных условиях произрастания :
а) С1еШо§епе5 вдиаггоза (ритм с периодом 24 ч), твердоосочково-змеевковое, б) РоШНИа ШпасеИ/оИа (ритмы с периодами 24 и 3.0 ч), ковыльно-разнотравно-вострецовое, в) Сагех ШНорИИа (ритмы с периодами 24 и 7.2 ч), злаково-осоковое, г) Уегомса 1оп§1/оИа (ритм с периодом 2.2 ч), крупноразнотравно-дернистоосоковое. Пунктиром показаны синусоиды
с периодом 24 ч.

Глава 6. СОДЕРЖАНИЕ ВОДЫ В ЛИСТЬЯХ 145
самой г у м и д н о и части г р а д и е н т а резко снижается относительная доля растений с в ы я в л е н н ы м и р и т м а м и о в о д н е н н о с т и листьев (рис . 6.2, а). На градиенте т е м пературы воздуха нет закономерностей в появлении и исчезновении р и т м о в (рис. 6.2, б). Однако м о ж н о заметить , что чем ниже средняя д л я вида в е л и ч и н а оводненности листьев , т ем л у ч ш е проявляется р и т м и ч н о с т ь показателя (рис. 6.2, в). У видов с с а м ы м и высокими значениями содержания воды в л и с т ь я х р и т м ы проявляются наиболее редко , изменения показателя в течение дня незначительны.
Вместе с тем, а м п л и т у д ы ритмов содержания воды вообще не велики и чаще всего не п р е в ы ш а ю т 0.3 г^„^„ - г'^у^.массы
(рис. 6.3, т а к ж е см. рис . 6.1). В е л и ч и н а этих амплитуд связана только с амплитуд а м и с у т о ч н ы х р и т м о в т р а н с п и р а ц и и (рис. 6.4). Это связь не очень сильна , но статистически достоверна . Отсюда выявляется тенденция — чем л у ч ш е выражен ритм транспирации, тем ч а щ е м о ж н о обнаружить суточные р и т м ы оводненности листьев .
Б а т и ф а з ы р и т м о в с о д е р ж а н и я воды чаще всего о б н а р у ж и в а ю т с я в послепо луденное время — м е ж д у 14 и 17 ч (рис. 6.5). Это происходит в 77% случаев , т.е. чаще всего м и н и м а л ь н а я о в о д н е н н о с т ь листьев имеет место через несколько часов после наступления акрофаз транспирации , когда и н т е н с и в н о с т ь последнего п р о ц е с с а у ж е с н и ж а е т с я . Э т о м о ж е т быть обусловлено значительно большей инерционностью изменений оводненности листьев по сравнению с транспираци-ей. Однако очень высокая интенсивность т р а н с п и р а ц и и « п о д т я г и в а е т » б а т и ф а з ы ритмов оводненности б л и ж е к полудню (рис. 6.6, а). Напротив , е с л и транспирация не о ч е н ь в ы с о к а , т о и м и н и м у м ы о в о д н е н н о с т и л и с т ь е в н а б л ю д а ю т с я в более п о з д н е е в р е м я д н я ( р и с . 6.6, а).
80
60
40
20
80
60 5«
Е 40
20
0-40 40-80 80-120 120-160 Запас воды в почве, мм
700 1 б
80
60
ш 40-
' • ' § 1
20- 1 0-
20-23 Температура воздуха, " С
0.6—2.6 4.6—6.6
Рис. 6.2. С о о т н о ш е н и е числа видов (в процентах от общего их числа в к а ж д о м интервале) с отсутствием (1) или н а л и ч и е м (2) суточных р и т м о в с о д е р ж а н и я воды в разны х частях градиентов в л а ж н о с т и по чвы (а), т емпер ат у р ы воздуха (б) и в разных интервалах оводненности листьев (в).

146 Часть II. Дневная динамика водного режима доминантов и эдификаторов сообществ.
Р и с . 6.3. Г и с т о г р а м м а р а с п р е д е л е н и я амплитуд суточных р и т м о в содержания
воды в листьях растений . 4 0 -
30
20
10
Рис . 6.4. С о о т н о ш е н и е м е ж д у а м п л и т у д а м и суточных р и т м о в т р а н с п и р а ц и и и
оводненности листьев .
73,5 74,5 75,5 76,5 77,5 78,5 Середины часовых интервалов
Рис. 6.5. Частота (в процентах от общего числа видов со статистически подтвержденными ритмами) встречаемости батифаз суточных ритмов содержания воды.
79 т
77
75
о а
о о
г' =0.410 13
19
17
15
13 г' =0.370^
1 2 ИТ, в/(гч)
2 СВ. г/а
Рис . 6.6. Связь м е ж д у батифазами суточных р и т м о в с о д е р ж а н и я воды в л и с т ь я х р а с т е н и й и показателями водного обмена .
Глава 6. СОДЕРЖАНИЕ ВОДЫ В ЛИСТЬЯХ 147
Рис. 6.7. Гистограмма распределения коэффициентов корреляции между дневными ритмами содержания воды и факторами
среды:
1 — температурой воздуха, 2 — дефицитом влажности воздуха, 3 — освещенностью.
Р и с . 6.8. П о л о ж е н и е а к р о ф а з ст ат ист и чески подтвержденных суточных ритмов с о д е р ж а н и я воды в з а в и с и м о с т и от корр е л и р о в а н н о с т и этих р и т м о в с р и т м а м и
т емпер ат у р ы воздуха.
Рис. 6.9. Батифазы дневных р и т м о в содержания воды (1 — суточные р и т м ы выявлены, 2 — суточных ритмов нет) , акрофазы температуры воздуха (3) и освещенности (4).
Соединительные линии между точками нанесены для лучщего обзора графика.

148 Часть II. Дневная динамика водного режима доминантов и эдификаторов сообществ.
Поскольку существует очень тесная статистическая связь (И = 0.750) м е ж д у средне-видовыми величинами интенсивности транспирации и содержания воды, то такое «затягивание» ритмов оводненности хорошо прослеживается и на фоне р а з н ы х значений количества воды в л и с т ь я х растений (рис. 6.6, б).
Видно, что на форму ритмов оводненности (т. е. амплитуды и положение батифаз) оказывает заметное влияние развитие процесса транспирации в течение д н я (рис. 6.4 и 6.6, а]. Однако д н е в н ы е ходы содержания воды в листьях р а с т е н и й часто коррелируют с д н е в н ы м и ходами температуры и д е ф и ц и т а в л а ж н о с т и воздуха (рис . 6.7). В 69% случаев о б н а р у ж и в а ю т с я статистически досто вер ные (отрицательные) корреляции м е ж д у о в о д н е н н о с т ь ю листьев и температурой воздуха, и несколько р еже (66%) между этим показателем водного р е ж и м а и в л а ж н о с т ь ю воздуха . Однако с и н х р о н и зирую щее воздействие этих факторов среды на р и т м ы о в о д н е н н о с т и листьев очень невелико. М о ж н о отметить только как тенденцию смещение батифаз этих ритмов бли-
0,04 -,
12 14 16 Врвия суток, ч
12 14 16 Время суток, ч
Рис. 6.10. П р и м е р ы ультрадианных р и т м о в с о д е р ж а н и я воды с р а з н ы м и п е р и о д а м и и соответствующих расчетных синусоид (пунктирные к р и в ы е ) .
Ритмы рассчитаны в отклонениях от суточных трендов. Периоды ритмов: а— 2.8 ч {ТНа1Шгит 5тр1ех, осоково-разнотравно-злаковое сообщество), б — 5.5 {Ьеутиз сНтепш, злаково-богаторазнотравное сообщество), в — 7.2 ч {Сагех ШкорНИа, злаково-осоковое сообщество), г — 9.2
{Сагех саезрИоза, крупноразнотравно-дернистоосоковое сообщество).
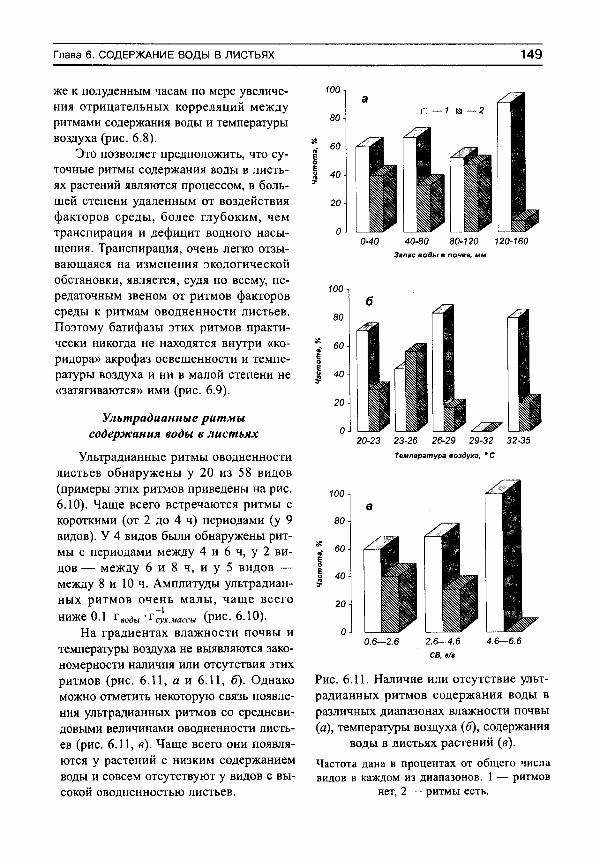
Глава 6. СОДЕРЖАНИЕ ВОДЫ В ЛИСТЬЯХ 149
же к полуденным часам по мере увеличения о т р и ц а т е л ь н ы х к о р р е л я ц и й м е ж д у ритмами содержания воды и температуры воздуха (рис . 6.8).
Это позволяет предположить , что суточные р и т м ы содержания воды в листь ях растений являются процессом, в большей степени у д а л е н н ы м от воздействия ф а к т о р о в с р е д ы , б о л е е г л у б о к и м , ч е м трансп и рац и я и д е ф и ц и т водного н а с ы щения . Транспирация , очень легко отзыв а ю щ а я с я на и з м е н е н и я э к о л о г и ч е с к о й обстановки , является , судя по всему, пе редаточным звеном от р и т м о в факторов среды к р и т м а м о в о д н е н н о с т и л и с т ь е в . Поэтому батифазы этих р и т м о в практически никогда не находятся внутри «коридора» акрофаз о с в е щ е н н о с т и и т е м п е ратуры воздуха и ни в м а л о й степени не «затягиваются» ими (рис . 6.9).
Ультрадианные ритмы содержания воды в листьях
Ультрадианные р и т м ы оводненности л и с т ь е в о б н а р у ж е н ы у 20 из 58 в и д о в (примеры этих ритмов приведены на рис. 6.10). Ч а щ е всего в с т р е ч а ю т с я р и т м ы с короткими (от 2 до 4 ч) п е р и о д а м и (у 9 видов). У 4 видов были о б н а р у ж е н ы ритмы с периодами м е ж д у 4 и 6 ч, у 2 вид о в — м е ж д у 6 и 8 ч, и у 5 в и д о в — между 8 и 10 ч. А м п л и т у д ы ультрадианн ы х р и т м о в о ч е н ь м а л ы , ч а щ е в с е г о ниже 0.1 г,<,йы т^ухл-ассы (рис . 6.10).
Н а г р а д и е н т а х в л а ж н о с т и п о ч в ы и температуры воздуха не выявляются закономерности наличия или отсутствия этих р и т м о в (рис . 6.11, а и 6.11, б). О д н а к о можно отметить некоторую связь появления ультрадианных р и т м о в со средневи-довыми величинами оводненности листьев (рис. 6.11, в) . Ч а щ е всего они появляются у р а с т е н и й с низким с о д е р ж а н и е м воды и совсем отсутствуют у видов с высокой о в о д н е н н о с т ь ю листьев .
20-23 23-26 26-29 29-32 32-35 Температура воздуха, " С
0 . 6 - 2 . 6 2.6—4.6 4.6—6.6 ее, г/г
Рис . 6.11. Н а л и ч и е или отсутствие ультр а д и а н н ы х р и т м о в с о д е р ж а н и я в о д ы в различных диапазонах в л а ж н о с т и почвы (а), температуры воздуха (б), содержания
воды в л и с т ь я х р а с т е н и й (в).
Частота дана в процентах от общего числа видов в каждом из диапазонов. 1 — ритмов
нет, 2 — ритмы есть.

150 Часть II. Дневная динамика водного режима доминантов и эдификаторов сообществ...
Резюме. Статистически достоверные суточные р и т м ы оводненности листьев бьши обнаружены у 30 из 58 и з у ч е н н ы х видов . Х а р а к т е р н о й о с о б е н н о с т ь ю этих р и т м о в является то, что в д н е в н о е время наблюдается м и н и м у м (батифаза ) ф у н к ц и и . Суточная р и т м и ч н о с т ь ч а щ е проявляется у видов с невысоким с о д е р ж а н и е м воды в листьях . В е л и ч и н а а м п л и т у д р и т м о в связана только с а м п л и т у д а м и с у т о ч н ы х р и т м о в транспирации. Батифазы р и т м о в содержания воды чаще всего обнаруживаются в послеполуденное время — м е ж д у 14 и 17 ч. Н а ф о р м у суточных р и т м о в оводненности оказывает з амет н ое влияние р а з в и т и е п р о ц е с с а т р а н с п и р а ц и и в т е ч е н и е дня . С и н х р о н и з и р у ю щ е е воздействие факторов среды на эти р и т м ы очень невелико . Сделано предположение , что суточные р и т м ы содержания воды в листьях р а с т е н и й являются процессом, в б о л ь ш е й степени , чем р и т м ы т р а н с п и р а ц и и и д е ф и ц и т а водного насыщения , удален н ым от воздействия факторов среды. Т р а н с п и р а ц и я может б ы т ь передаточным звеном от р и т м о в факторов среды к р и т м а м о в о д н е н н о с т и листьев . Ультрадианные р и т м ы о в о д н е н н о с т и листьев о б н а р у ж е н ы у 20 из 58 в и д о в . Ч а щ е всего они появляются у р а с т е н и й с низким содержанием воды.

Ч А С Т Ь 111
ЗАКОНОМЕРНОСТИ СТРУКТУРНО-ФУНКЦИОНАЛЬНОЙ ОРГАНИЗАЦИИ РАСТЕНИЙ
НА ГРАДИЕНТЕ ВЛАЖНОСТИ ПОЧВЫ
в п р е д ы д у щ и х главах м ы рассматривали о с о б е н н о с т и се зонной и дневной динамики показателей водного р е ж и м а каждого исследованного вида. В этой части будут исследованы закономерности изменения с р е д н е в и д о в ы х в е л и ч и н водного обмена на градиенте в л а ж н о с т и почвы, а также их связи с анатомической структурой листьев . Здесь и с п о л ь з о в а н ы д о п о л н и т е л ь н ы е д а н н ы е ( с о б р а н н ы е в п я т и с о о б щ е с т в а х ) , не вошедшие в п р е д ы д у щ и е разделы из-за сравнительно небольшого числа наблюдений.
ГЛАВА 7. ИНТЕНСИВНОСТЬ ТРАНСПИРАЦИИ
Полевые исследования интенсивности транспирации проводятся в широком спектре экологических условий . В литературе м о ж н о найти характерные в е л и ч и н ы этого показателя водообмена для д о м и н а н т о в с о о б щ е с т в практически всех к л и м а т и ч е с к и х зон (Бейдеман, 1983). В д а н н о й главе м ы попытаемся в некоторой степени дополнить с у щ е с т в у ю щ и е д а н н ы е . С этой ц е л ь ю будет 1) д а н а статистическая характеристика интенсивности транспирации ряда видов на градиенте влажности почвы, 2) проведен анализ закономерностей и з м е н е н и я средних показателей и их варьирования н а этом градиенте .
Статистический анализ интенсивности транспирации у видов однодольных и двудольных растений
Д а н н ы е с о б р а н ы в 16 сообществах , р а с п о л о ж е н н ы х в р а з н ы х частях г р а д и е н т а влажности почвы (табл. 7.1). Статистические характеристики средневидовых величин интенсивности т р а н с п и р а ц и и показаны в таблице 7.2.
Весь массив исходных данных интенсивности транспирации имеет среднее значение
X = 1.538 г ,„йь, • г;1.л«.ссы • ч " ' и стандартное отклонение = 1.013 г^^^ы сух.массы'"^'^

152 Часть III. Закономерности структурно-функциональной организации растений..
Таблица 7.1
Краткая характеристика сообществ и условий пр о из р аст ания во время проведения наблюдений
№ Характеристика Место и год Средние значения за период наблюдений п/п сообщества проведения
наблюдений запаса темпера дефици освещенпроведения
наблюдений влаги в туры та влаж ности. 30-см воздуха. ности калсм"^
слое поч °С воздуха. мин"' вы, мм мб
1 Разнотравно-днеп-ровскоковыльное (псаммофитная степь, почвы дернового ряда с песчаным аллювием)
Пойма р. Хопер, Хоперский заповедник. Воронежская область (1996 г.).
3.9 34.7 44.1 0.75
2 Ковыльково-луково-баглуровое (остепнен-ная пустыня, бурая супесчаная почва) (естественное местообитание)
Сомон Шине-Джинст, Заалтай-ская Гоби, Монголия (1983 г.)
13.1 23.7 20.4 0.81
3 Псаммофитно-степное злаковое (псаммофитная степь, почвы дернового ряда с песчаным аллювием)
Пойма р. Хопер, Хоперский заповедник. Воронежская область (1982 п)
13.6 22.9 14.6
4 Сообщество крио-фитных подушечни-ков (высокогорная мелкодерновиннозла-ковая степь, горная степная почва)
Сомон Мунх-Хайрхан, Монгольский Алтай (1991 п)
14.0 17.1 13.6
5 Кобрезиевое сообщество (высокогорная мелкодерновиннозла-ковая степь, горная степная почва)
Сомон Мунх-Хайрхан, Монгольский Алтай (1991 п)
20.9 17.2 13.9
6 Ковыльково-луково-баглуровое (остепнен-ная пустыня, бурая супесчаная почва) (участок с искусственным поливом)
Сомон Шине-Джинст, Заалтай-ская Гоби, Монголия (1983 г.)
21.2 23.7 20.4 0.81
7 Твердоосочково-змеевковое (остепнен-ный луг, почвы дернового ряда с песчаным и супесчаным аллювием)
Пойма р. Орхон, сомон Шаамар, северная Монголия (1988 г.)
37.1 24.2 13.4 0.87
8 Ковыльно-разно- Сомон Туменцогт, 50.3 21.1 11.6 0.77 травно-вострецовое восточная Монго(настоящая степь, лу- лия (1984-1985 г.) гово-каштановая ос-тепненная почвы)

Глава 7. ИНТЕНСИВНОСТЬ ТРАНСПИРАЦИИ 153
Окончание таблицы 7.1
№ Характеристика Место и год Средние значения за период наблюдений п/п сообщества проведения
наблюдений запаса темпера дефици освещенпроведения
наблюдений влаги в туры та влаж ности. 30-см воздуха, ности кал-см"^
слое поч °С воздуха. мин ' вы, мм мб
9 Кровохлебковое (среднепоемный настоящий луг, почвы лугового ряда с суглинистым аллювием)
Пойма р. Хопер, Хоперский заповедник. Воронежская область (1997 г )
55.9 26.1 19.1 0.78
10 Крупноразнотравное (среднепоемный настоящий луг, почвы лугового ряда с супесчаным и суглинистым аллювием)
Пойма р. Хопер, Хоперский заповедник. Воронежская область (1982 г.)
66.8 18.8 8.8
11 Крупноразнотравное Пойма р. Хопер, 93.0 20.5 9.8 — с осокой лисьей Хоперский запо(долгопоемный на ведник. Воронежстоящий луг, почвы ская область лугового ряда с сугли (1982 п) нистым аллювием)
12 Злаково-осоковое (долгопоемный настоящий луг, почвы болотного ряда с глинистым и иловатым аллювием)
Пойма р. Орхон, Сомон Шаамар, северная Монголия (1986 г )
99.6 21.8 8.6 1.24
13 Осоково-разнотравно-злаковое (долгопоемный настоящий луг, почвы лугового ряда с суглинистым аллювием)
Пойма р. Орхон, Сомон Шаамар, северная Монголия (1987 г.)
104.0 22.3 14.2 0.84
14 Крупноразнотрав-но-дернистоосоковое (заболоченный луг, почвы болотного ряда с глинистым и иловатым аллювием)
Пойма р. Хопер, Хоперский заповедник. Воронежская область (1995 г.)
111.5 25.3 18.4 0.71
15 Злаково-богаторазнотравное (заболоченный луг, почвы лугового ряда с суглинистым аллювием)
Пойма р. Орхон, Сомон Шаамар, северная Монголия (1987 г )
123.5 22.6 13.7 1.31
16 ОстроосокоБое (участок прибрежноводной растительности, песчаная почва прибрежной зоны озера)
Пойма р. Хопер, Хоперский заповедник, Воронежская область (1996 г.)
143.9 21.6 11.6 0.26

154 Часть III. Закономерности структурно-функциональной организации растений..
(п = 13837). Ряд распределения имеет положительну ю а с и м м е т р и ю ' с модой , р а в н о й 1 -25 г^о^ы • г^у^^^^^^ • ч " ' , а 90% его з н а ч е н и й (вариант ) н а х о д я т с я в д и а п а з о н е 0 -2-9 ^еоды'^с^хмассы'ч'^ ( о с т а л ь н ы с 10% в а р и а н т р а с п р е д е л е н ы в д и а п а з о н е 2 . 9 -'^•^ ^еоды '^сух.массы ' ) • М о ж н о сказать, ЧТО типичные значения интенсивности трансп и р а ц и и т р а в я н и с т ы х р а с т е н и й н а г р а д и е н т е в л а ж н о с т и п о ч в ы не п р е в ы ш а ю т
^воды '^сух.массы
Таблица 7.2
Средние значения и показатели вариации интенсивности т р а н с п и р а ц и и {ИТ), в ы р а ж е н н о й в г,„<,„ •т~1^,^^,ассы ' Ч ' ' (столбец х ] ) , а т а к ж е в д р у г и х е д и н и ц а х
( м м о л ь т ~ ^ ^ ^ , , д ^ ^ „ - м и н " ' — с т о л б е ц Х 2 , м м о л ь - м " ^ - с " ' — с т о л б е ц х з )
№ участка по табл.
7.1
№ объек
та Вид
ИТ № участка по табл.
7.1
№ объек
та Вид
п х\ ^х У,% Х2 ХЗ
1 1 Апетша аиз1г1аса 94 0.219 0.110 50.0 0.203 0.422 2 Сагех со1сЫса 94 0.283 0.119 42.1 0.262 0.518 3 ОаИит ги1Нетсит 94 0.421 0.236 56.1 0.389 0.656 4 8Нра Ъогуз1кетса 94 0.288 0.088 30.7 0.266 0.681 5 Тп/оИит апгете 52 0.301 0.109 36.2 0.278 0.609 6 Уеготса зркаШ 94 0.212 0.105 49.5 0.196 0.436
Суммы и средние для сообщества 522 0.286 0.156 54.4 0.265 0.549 2 7 АШит ро1угк12ит \11 1.338 1.000 74.7 1.238 0.742
8 АпаЬаз13 Ьгеуг/оИа 179 0.359 0.243 67.7 0.332 0.341 9 5Ира ^1агеоза 173 1.249 0.316 25.3 1.155 2.377
Суммы и средние для сообщества 532 0.979 0.762 77.8 0.906 1.079
' Для того, чтобы принять или отвергнуть нулевую гипотезу обычно используют такие статистики, как I , Р- или хи-квадрат. Все эти статистики имеют нормальное распределение и требуют, чтобы анализируемые переменные сами были нормально распределены в совокупности. Однако опыт показывает, что очень часто данные полевых ботанических исследований уклоняются от нормального закона распределения. В настоящее время считается, что во многих случаях все же можно использовать тесты, основанные на предположении нормальности, если объем выборки достаточно велик. Эта возможность основана на чрезвычайно важном принципе, позволяющем понять популярность тестов, основанных на нормальности. А именно, при возрастании объема выборки (п > 30), форма выборочного распределения (т.е. распределение выборочной статистики критерия) приближается к нормальной, даже если распределение исследуемых переменных не является нормальным. Этот принцип называется центральной предельной теоремой. Кроме того, исследования с помощью методов Монте-Карло интенсивно использовались для того, чтобы оценить, насколько тесты, основанные на предположении нормальности, чувствительны к различным нарушениям предположений нормальности. Общий вывод этих исследований состоит в том, что последствия нарушения предположения нормальности менее фатальны, чем первоначально предполагалось (81а18ой, 2001).
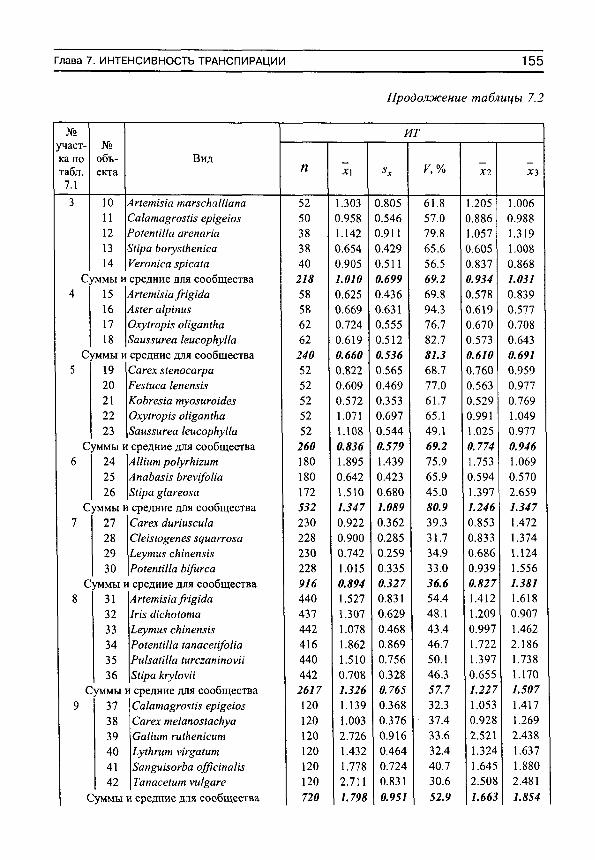
Глава 7. ИНТЕНСИВНОСТЬ ТРАНСПИРАЦИИ 155
Продолжение таблицы 7.2
№ участка по табл.
7.1
№ объекта
ИТ № участка по табл.
7.1
№ объекта
Вид п XI У,% Х2 хз
3 10 Апетша тагзсИаШапа 52 1.303 0.805 61.8 1.205 1.006 11 Са1ата§го5Из ер1^еюз 50 0.958 0.546 57.0 0.886 0.988 12 Ро1епШ1а агепапа 38 1.142 0.911 79.8 1.057 1.319 13 8ира ЬогузЛетса 38 0.654 0.429 65.6 0.605 1.008 14 Уеготса зр1саШ 40 0.905 0.511 56.5 0.837 0.868
С: 1^мы и средние для сообщества 218 1.010 0.699 69.2 0.934 1.031 4 15 Аг1етша /п^Ша 58 0.625 0.436 69.8 0.578 0.839
16 Аз(ег а1ртш 58 0.669 0.631 94.3 0.619 0.577 17 Оху1гор1$ оН§ап1На 62 0.724 0.555 76.7 0.670 0.708 18 8аиззигеа кисорНуИа 62 0.619 0.512 82.7 0.573 0.643
рты и средние для сообщества 240 0.660 0.536 81.3 0.610 0.691 5 19 Сагех з1епосагра 52 0.822 0.565 68.7 0.760 0.959
20 РезШса 1епепз13 52 0.609 0.469 77.0 0.563 0.977 21 КоЪгез1а туозигоШез 52 0.572 0.353 61.7 0.529 0.769 22 Оху1гор1з о1щап(На 52 1.071 0.697 65.1 0.991 1.049 23 8аиззигеа 1еисорНу11а 52 1.108 0.544 49.1 1.025 0.977
С уммы и средние для сообщества 260 0.836 0.579 69.2 0.774 0.946 6 24 АШит ро/угкггит 180 1.895 1.439 75.9 1.753 1.069
25 АпаЬаз1з Ьгвуг/оИа 180 0.642 0.423 65.9 0.594 0.570 26 8ира ^1агеоза 172 1.510 0.680 45.0 1.397 2.659
Суммы и средние для сообщества 532 1.347 1.089 80.9 1.246 1.347 7 27 Сагех (1ипизси1а 230 0.922 0.362 39.3 0.853 1.472
28 С1еШо^епез здиаггоза 228 0.900 0.285 31.7 0.833 1.374 29 Ьеутиз сИтепз1з 230 0.742 0.259 34.9 0.686 1.124 30 РоЬепИИа Ы/игса 228 1.015 0.335 33.0 0.939 1.556
С уммы и средние для сообщества 916 0.894 0.327 36.6 0.827 1.381 8 31 Аг1ет1з1а /п^Ша 440 1.527 0.831 54.4 1.412 1.618
32 Мз сИсНоЮта 437 1.307 0.629 48.1 1.209 0.907 33 Ьеутиз сНтепз1з 442 1.078 0.468 43.4 0.997 1.462 34 Ро1еп1И1а 1апасеИ/оИа 416 1.862 0.869 46.7 1.722 2.186 35 Ри1заШ1а ШгсгатпоуИ 440 1.510 0.756 50.1 1.397 1.738 36 8ира кгу1оуН 442 0.708 0.328 46.3 0.655 1.170
С уммы и средние для сообщества 2617 1.326 0.765 57.7 1.227 1.507 9 37 Са1ата^гозИ5 ерщеюз 120 1.139 0.368 32.3 1.053 1.417
38 Сагех те1апоз1асНуа 120 1.003 0.376 37.4 0.928 1.269 39 СаНит гиЛетсит 120 2.726 0.916 33.6 2.521 2.438 40 Ьу1кгит у1г^а1ит 120 1.432 0.464 32.4 1.324 1.637 41 8ап§шзогЬа о^ТшпаНз 120 1.778 0.724 40.7 1.645 1.880 42 ТапасеШт уи1§аге 120 2.711 0.831 30.6 2.508 2.481
Суммы и средние для сообщества 720 1.798 0.951 52.9 /.663 1.854

156 Часть III. Закономерности структурно-функциональной организации растений..
Продолжение таблицы 7.2
№ участка по табл.
7.1
объекта
ИТ № участка по табл.
7.1
объекта Вид
п х\ У,% Х2 хз
10 43 Сагех те1апозШскуа 34 0.786 0.424 53.9 0.727 0.868 44 РШрепс1и1а и1тапа 34 1.870 0.947 50.6 1.730 1.707 45 СаИит гиЫоШез 34 1.410 0.714 50.6 1.304 1.100 46 5ап§ш5огЬа о^ТюшаНз 34 2.331 1.292 55.4 2.156 1.839
С; {мыы и средние для сообщества 136 1.599 1.061 66.3 1.479 1.379 И 47 Сагех уи1рша 50 1.118 0.516 46.1 1.034 1.110
48 аЫит тсапит 50 2.971 1.542 51.9 2.748 1.655 49 ОаИит шЬюМез 50 2.434 1.199 49.3 2.251 2.133 50 Уеготса 1оп^1/оИа 50 1.958 1.039 53.0 1.812 1.615 51 Ута сгасса 50 3.570 1.643 46.0 3.302 2.931
С ут̂ 1мы и средние для сообщества 250 2.410 1.502 62.3 2.229 1.889 12 52 А1оресигиз агипсИпасеиз 280 1.697 0.595 35.1 1.570 1.889
53 Сагех а1кего(1е5 284 1.184 0.347 29.3 1.095 1.509 54 Сагех ШкоркИа 284 1.344 0.380 28.3 1.243 1.724 55 Е1у1п§1а герепз (Ъ.) 242 1.489 0.476 32.0 1.377 1.882 56 /то з1Ыпса 284 1.285 0.553 43.0 1.189 1.323 57 Рка1аго1с1ез атпсИпасеа 286 1.558 0.490 31.5 1.441 1.703 58 Ро1у§опит зШпсит 284 2.860 1.336 46.7 2.646 2.352
С уммы и средние для сообщества 1944 1.634 0.861 52.7 1.511 1.767 13 59 А^гозйз топ^коИса 239 1.446 0,386 26.7 1.338 1.414
60 Апетопе сИско1ота 240 2.112 0.773 36.6 1.954 1.865 61 Сагех агкего^ез 240 1.296 0.451 34.8 1.199 1.607 62 Роа ап^изИ/оИа 240 0.804 0.187 23.3 0.744 1.158 63 ТкаНс1гит зтр1ех 239 1.862 0,792 42.5 1.722 1.990 64 УШа сгасса 240 2.254 0.836 37.1 2.085 2.233
С уммы и средние для сообщества 1438 1.629 0.797 48.9 1.507 1.711 14 65 Сагех саезрИоза 224 1.339 0.471 35.1 1.239 1.394
66 РШрепМа и1тапа 224 1.436 0.535 37.2 1.329 1.657 67 1п! рзеис/асогиз 226 1.443 0.455 31.6 1.334 1.118 68 8ап^18огЬа о]]1стаШ 228 2.056 0.750 36.5 1.901 2.048 69 Уа1епапа ^01^6/1315 228 1.222 0.450 36.8 1.131 0.786 70 Уеготса 1оп§фИа 226 1.854 0.680 36.7 1.715 1.610
Суммы и средние для сообщества 1356 1.559 0.641 41.1 1.442 1.435 15 71 Аг1етша 1ас1та1а 238 3.216 1.368 42.5 2.975 2.958
72 ВготорзЫ тегтгз 168 1.454 0.485 33.4 1.345 1.842 73 НетегосаШз ттог 232 1.799 0.643 35.7 1.664 1.650 74 Ьеутиз сктепз1з 238 1.185 0.381 32.2 1.096 1.654
75 Ро(епШ1а апзегта 238 2.305 0.946 41.0 2.132 2.903 76 8ап§шзогЪа оЛГкшаНз 238 2.553 0.926 36.3 2.362 2.855
Суммы и средние для сообщества 1352 2.119 1.113 52.5 1.960 2.338

Глава 7. ИНТЕНСИВНОСТЬ ТРАНСПИРАЦИИ 157
Окончание таблицы 7.2
№ участка по табл. 7.1
№ объекта Вид
ИТ № участка по табл. 7.1
№ объекта Вид
п х\ У,% XI хз
16 11 АНзта р1ап1а§о-адиаИса 138 3.978 1.008 25.3 3.679 2.033 78 ВШотш итЬеИШт 118 2.407 0.698 29.0 2.227 0.918 79 Сагех асШа 138 1.951 0.491 25.2 1.805 1.677 80 Ьусориз еигораеиз 138 3.167 1.252 39.5 2,929 1.801 81 8а^Шапа ш^Ий/оИа 138 4.078 0.930 22.8 3.772 1.846 82 Зраг^атит егесШт 138 2.198 0.676 30.8 2.034 1.078
Суммы и средние для сообщества 808 2.977 1.221 41.0 2.754 1.575
Оптимальные для транспирации условия температуры и дефицита влажности воздуха были найдены методом огибающей кривой. Этот метод щироко использовался (ТгапдиИИш, 1954; Вознесенский, 1977; Кайбияйнен, 1984; Слемнев, 1988 и др.) для нахождения оптимумов процессов газообмена при влиянии на них каких-либо внешних или внутренних факторов. При этом точки, расположенные ниже огибающей кривой на диаграмме «фактор-процесс», будут указывать на то, что лимитирующими являются какие-либо другие, кроме исследуемого, факторы (Кайбияйнен, 1984). В экологии водного обмена растений данный метод, насколько нам известно, не применялся. Однако при наличии большого числа наблюдений он может дать интересную информацию о том, при каких условиях температуры и дефицита влажности воздуха могут наблюдаться максимальные значения скорости испарения воды и, следовательно, максимальное открытие устьиц.
Огибающая кривая показывает влияние температуры (рис. 7.1, а) и дефицита влажности воздуха (рис. 7.1, б) на скорость испарения воды и степень открытости устьиц в отсутствии лимитирования другими факторами среды. Оптимальные диапазоны температуры (20-28 °С) и дефицита влажности (5-24 мб) воздуха были определены на уровне 90% от максимальной величины интенсивности транспирации. С другой стороны, поскольку нижние и верхние децили температуры (16.5 и 29.2 °С) и дефицита влажности воздуха (4.6 и 27.6 мб) очень близки к верхним и нижним границам соответствующих оптимумов, то можно констатировать, что подавляющее число наблюдений (80%) проведено при достаточно благоприятных условиях этих факторов среды. Что касается освещенности, то огибающая кривая интенсивности транспирации в зависимости от этого фактора среды практически параллельна оси абсцисс, и поэтому никакого оптимума выделить невозможно.
Исследованный нами набор видов неоднороден по таксономическому и экологическому составу, анатомической структуре листьев. Это объясняется большим разнообразием условий произрастания на градиенте влажности почвы. В связи с этим, для проведения более детального анализа, вся выборка была разделена на две примерно равные по количеству видов группы однодольных и двудольных растений. В каждой
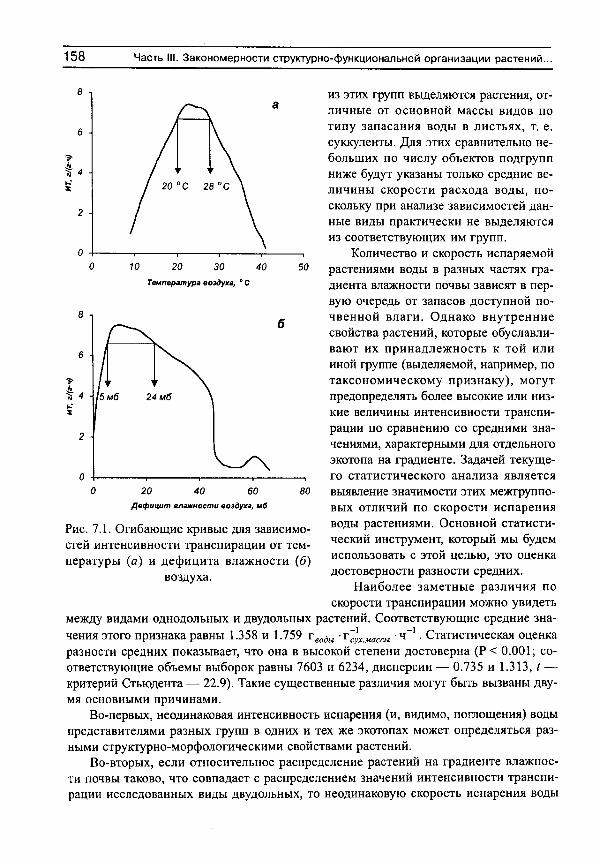
158 Часть III. Закономерности структурно-функциональной организации растений.
8 п
6 -
4 •
10 20 30 40 50
Температура воздуха, ° С
ИЗ ЭТИХ групп выделяются растения, отличные от основной массы видов по типу запасания воды в листьях, т. е. суккуленты. Для этих сравнительно небольших по числу объектов подгрупп ниже будут указаны только средние величины скорости расхода воды, поскольку при анализе зависимостей данные виды практически не выделяются из соответствующих им групп.
Количество и скорость испаряемой растениями воды в разных частях градиента влажности почвы зависят в первую очередь от запасов доступной почвенной влаги. Однако внутренние свойства растений, которые обуславливают их принадлежность к той или иной группе (выделяемой, например, по таксономическому признаку), могут предопределять более высокие или низкие величины интенсивности транспирации по сравнению со средними значениями, характерными для отдельного экотопа на градиенте. Задачей текущего статистического анализа является выявление значимости этих межгрупповых отличий по скорости испарения воды растениями. Основной статистический инструмент, который мы будем использовать с этой целью, это оценка достоверности разности средних.
Наиболее заметные различия по скорости транспирации можно увидеть
между видами однодольных и двудольных растений. Соответствующие средние значения этого признака равны 1.358 и 1.759 г^^^^, -^с^х.массы "ч"' • Статистическая оценка разности средних показывает, что она в высокой степени достоверна ( Р < 0.001; соответствующие объемы выборок равны 7603 и 6234, дисперсии — 0.735 и 1.313,; — критерий Стьюдента— 22.9). Такие существенные различия могут быть вызваны двумя основными причинами.
Во-первых, неодинаковая интенсивность испарения (и, видимо, поглощения) воды представителями разных групп в одних и тех же экотопах может определяться разными структурно-морфологическими свойствами растений.
Во-вторых, если относительное распределение растений на градиенте влажности почвы таково, что совпадает с распределением значений интенсивности транспирации исследованных виды двудольных, то неодинаковую скорость испарения воды
20 40 60
Дефицит влажности воздуха, мб
Рис. 7.1. Огибающие кривые для зависимостей интенсивности транспирации от температуры (а) и дефицита влажности (б)
воздуха.

Глава 7. ИНТЕНСИВНОСТЬ ТРАНСПИРАЦИИ 159
0-50 50-100 100-150 ' 0-50 50-100 100-150 Запас воды в почве, мм Запас воды в почве, мм
Рис. 7.2. Относительное количество исследованных видов однодольных и двудольных растений (в п р о ц е н т а х от о б щ е г о числа видов к а ж д о й категории) в р а з н ы х частях градиента в л а ж н о с т и п о ч в ы (а) и средние значения интенсивности т р а н с п и р а ц и и (б)
для этих групп растений .
1 — однодольные, 2 — двудольные.
видами двудольных и однодольных м о ж н о объяснить и м е н н о э т и м р а с п р е д е л е н и е м . Однако фактическое распределение растений на градиенте влажности почвы опровергает это предположение , поскольку в и д ы двудольных в о т н о с и т е л ь н о б о л ь ш е м количестве были исследованы в аридных экотопах, чем в гумидных , а в и д ы однодольных, напротив, — в г у м и д н ы х , по с р а в н е н и ю с а р и д н ы м и (рис . 7.2, а).
Следовательно , распределение растений не оказывало заметного влияния на статистические различия по скорости испарения воды между д а н н ы м и группами , а причины неодинаковой утилизации почвенной влаги нужно искать в структурно-функциональных и/или м о р ф о л о г и ч е с к и х свойствах растений .
Это тем более вероятно, что в местообитаниях с запасами влаги от 50 д о 150 м м интенсивность т р а н с п и р а ц и и двудольных растений была с у щ е с т в е н н о в ы ш е , чем однодольных (рис . 7.2, б) (оценки разностей средних д о с т о в е р н ы п р и Р < 0.001). Эти различия отчетливо проявляются , несмотря на то , что скорость и с п а р е н и я воды однодольными г и д р о ф и т а м и АИзта р1ап1а§о-адиаНса, ВШотш итЬеИаШз, За^Шапа $а§ии/оИа и Зращатит егесЫт была очень высокой (на у р о в н е д в у д о л ь н ы х растений и выше) .
В наиболее засушливых условиях произрастания (с запасами влаги в почве д о 50 мм) интенсивность т р а н с п и р а ц и и двудольных растений , наоборот, б ы л а несколько ниже , чем однодольных (рис. 7.2, б) (оценки разности средних д о с т о в е р н ы при Р < 0.001). При улучшении условий почвенного водоснабжения (на среднем участке градиента с запасами влаги от 50 д о 100 мм) скорость испарения воды д в у д о л ь н ы м и р а с т е н и я м и увеличилась более , чем в 2 раза (в среднем с 0.887 д о 2.156 г^о^^ • г~^^^дссы • ч ~ ' ) и затем (в гумидной части градиента с запасами влаги от 100 д о 150 м м ) возросла л и ш ь незначительно (до 2.224 т^„^^ •'^с^х.массы ч ~ ' ) - Для видов однодольных картина иная (рис. 7.2, б): интенсивность транспирации незначительно увеличилась в средней части
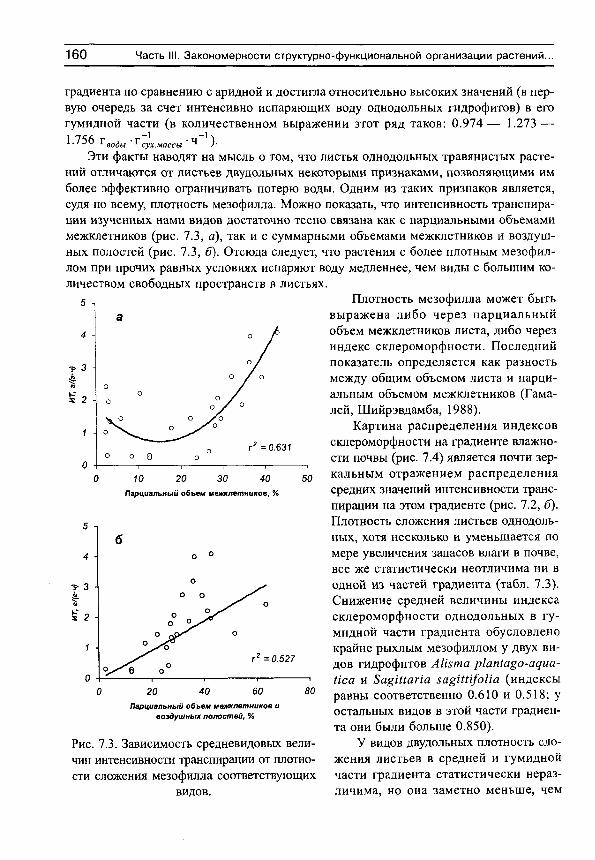
160 Часть III. Закономерности структурно-функциональной организации растений...
градиента по сравнению с аридной и достигла относительно высоких значений (в первую очередь за счет и н т е н с и в н о и с п а р я ю щ и х воду однодольных гидрофитов) в его г у м и д н о й ч а с т и (в к о л и ч е с т в е н н о м в ы р а ж е н и и этот р я д т а к о в : 0.974 — 1.273 —
^•"^^^ ^воды сух.массы ' ^ ) •
Эти факты наводят на м ы с л ь о том, что листья однодольных т р а в я н и с т ы х растений отличаются от листьев двудольных некоторыми п р и з н а к а м и , п о з в о л я ю щ и м и им более э ф ф е к т и в н о ограничивать потерю воды. О д н и м из т а к и х признаков является , судя по всему, плотность мезофилла . М о ж н о показать, что интенсивность транспирации изученных н а м и видов достаточно т е с н о связана как с п а р ц и а л ь н ы м и о б ъ е м а м и межклетников (рис. 7.3, а), так и с с у м м а р н ы м и о бъемами м е ж к л е т н и к о в и воздушных полостей (рис . 7.3, б). О т с ю д а следует, что растения с более п л о т н ы м м е з о ф и л лом при прочих р а в н ы х условиях испаряют воду медленнее , ч е м в и д ы с большим количеством свободных пространств в листьях .
П л о т н о с т ь м е з о ф и л л а м о ж е т б ы т ь в ы р а ж е н а л и б о ч е р е з п а р ц и а л ь н ы й объем межклет нико в листа , л и б о через и н д е к с с к л е р о м о р ф н о с т и . П о с л е д н и й показатель о п р е д е л я е т с я как р а з н о с т ь м е ж д у о б щ и м о бъемо м листа и парциа л ь н ы м объемом м е ж к л е т н и к о в (Гамалей , Ш и й р э в д а м б а , 1988).
К а р т и н а р а с п р е д е л е н и я и н д е к с о в склероморфности на градиенте влажности почвы (рис. 7.4) является почти зерк а л ь н ы м о т р а ж е н и е м р а с п р е д е л е н и я средних значений интенсивности транспирации на этом градиенте (рис. 7.2, б). Плотность сложения листьев однодольн ы х , хотя несколько и у м е н ь ш а е т с я по мере увеличения запасов влаги в почве, все же статистически н е о т л и ч и м а ни в одной из ч а с т е й г р а д и е н т а (табл . 7.3). С н и ж е н и е с р е д н е й в е л и ч и н ы и н д е к с а с к л е р о м о р ф н о с т и о д н о д о л ь н ы х в гум и д н о й ч а с т и г р а д и е н т а о б у с л о в л е н о крайне р ы х л ы м м е з о ф и л л о м у двух вид о в г и д р о ф и т о в АНзта р1ап1а§о-адиа-Нса и 5а§И1аг1а за^ИН/оНа ( и н д е к с ы р а в н ы соответственно 0.610 и 0.518; у остальных видов в этой части градиента они б ы л и б о л ь ш е 0.850).
У видов двудольных плотность слож е н и я л и с т ь е в в с р е д н е й и г у м и д н о й части г р а д и е н т а с т а т и с т и ч е с к и нераз л и ч и м а , но о н а з а м е т н о м е н ь ш е , чем
а
о 20 40 60 Парциальный объем межклетников и
воздушных полостей, %
Рис. 7.3. Зависимость средневидовых величин интенсивности транспирации от плотности сложения мезофилла соответствующих
видов.

Глава 7. ИНТЕНСИВНОСТЬ ТРАНСПИРАЦИИ 161
в экотопах с запасами влаги в почве д о 50 мм (табл. 7.3, рис . 7.4). Но во всех местообитаниях у однодольных расте ний листья более плотные , чем у двудольных (табл. 7.3).
О т м е ч е н н о е в ы ш е р а с п р е д е л е н и е и н д е к с о в с к л е р о м о р ф н о с т и д в у д о л ь ных растений на градиенте влажности почвы является подтверждением давно установленного факта , согласно котор о м у к с е р о м о р ф н а я о р г а н и з а ц и я л и с тьев с в я з а н а с б о л е е п л о т н ы м м е з о ф и л л о м , а м е з о м о р ф н а я — с б о л е е р ы х л ы м его сложением (Василевская , 1979). О б этом же с в и д е т е л ь с т в у е т и связь скорости испарения воды с количеством м е ж к л е т н и к о в в листьях рас тений (рис . 7.2, а). Это х а р а к т е р н о в основном для видов двудольных растений. Д л я однодольных р а с т е н и й связь скорости т р а н с п о р т а в о д ы с п л о т н о с т ь ю м е з о ф и л л а п о д т в е р ж д а е т с я только д в у м я точками (рис . 7.5, а), при и с к л ю ч е н и и которых она становится статистически недостоверной (поэтому м ы с к л о н н ы рассматривать д а н н ы й график только как отражение тенденции) . В и д ы двудольных и м е ю т достаточно т е с н у ю связь интенсивности транспирации с показателем плотности м е з о ф и л л а (рис. 7.5, б).
Таблица 7.3
Матрица разностей средних значений индексов с к л е р о м о р ф н о с т и у однодольных и двудольных растений на градиенте влажности почвы с у каз анием у р о в н е й
статистической з н а ч и м о с т и (при отсутствии статистически з н а ч и м ы х р аз личий ячейки оставлены незаполненными)
Однодольные (0-50 мм)
-0.074 (Р < 0.05)
-0.173 ( Р < 0.001)
-0.228 ( Р < 0.001)
0.074 (Р < 0.05)
Двудольные (0-50 мм)
0.079 (Р < 0.05)
-0.099 (Р < 0.05)
-0.154 ( Р < 0.001)
-0.079 (Р < 0.05)
Однодольные (50-100 мм)
-0.178 (Р<0 .0 1 )
-0.233 ( Р < 0.001)
0.173 ( Р < 0.001)
0.099 (Р < 0.05)
0.178 (Р<0 .01 )
Двудольные (50-100 мм)
0.134 (Р < 0.05)
-0.134 (Р < 0.05)
Однодольные (100-150 мм)
-0.189 ( Р < 0.001)
0.228 ( Р < 0.001)
0.154 ( Р < 0.001)
0.233 ( Р < 0.001)
0.189 ( Р < 0.001)
Двудольные (100-150 мм)
6 - 2353
0-50 50-100 100-150 Запас воды в почве, мм
Рис . 7.4. И н д е к с ы с к л е р о м о р ф н о с т и иссле дованных видов однодольных и двудольных
растений на градиенте в л а ж н о с т и почвы.
1 — однодольные, 2 — двудольные.

162 Часть III. Закономерности структурно-функциональной организации растений.
5 1
4 •
3 -
2 •
Статистическое описание интенсивности транспирации у видов однодольных и двудольных суккулентов
К числу однодольных суккулентов относятся с л е д у ю щ и е виды: АШит ро1угЫгит, НетегосаШз ттог, Мз (Иско1ота, 1пз рзеис1асогиз, / т зШпса, а т а к ж е ВиЮтиз итЪеНаШз (с в о д о н о с н о й обкладкой пучков) и 8раг§атит егес1ит ( и м е ю щ и й специализированную в о д о н о с н у ю ткань) . В группе двудольных р а с т е н и й имеется только один суккулент (с коронарной обкладкой пучков) — АпаЬазп Ь^еV^/о^^а.
Широко р а с п р о с т р а н е н о мнение , что суккуленты испар я ю т воду гораздо медленнее н е с у к к у л е н т н ы х р а с т е н и й (склерофитов) . Это , в и д и м о , с в я з а н о с тем, что при пересчете интенсивности транспирации на сырую массу навесок получаются занижен
н ы е в е л и ч и н ы этого п о к а з а т е л я из-за высокой в л а ж н о с т и л и с т ь е в суккулентов . Однако н а ш и д а н н ы е показывают, что интенсивность т р а н с п и р а ц и и однод о л ь н ы х с у к к у л е н т о в з а м е т н о в ы ш е , чем у о дно до ль ных с к л е р о ф и т о в (табл. 7.4). Это м о ж н о п р о д е м о н с т р и р о в а т ь с п о м о щ ь ю такого показателя , как время обмена запасов воды ( В О З В ) , который является о т н о ш е н и е м содержания воды к интенсивности т р а н с п и р а ц и и и измер я е т с я в е д и н и ц а х в р е м е н и (в н а ш е м с л у ч а е — в ч а с а х ) . Д л я о д н о д о л ь н ы х с к л е р о ф и т о в э т о т п о к а з а т е л ь р а в е н 1.8 ч, в то время как д л я о д н о д о л ь н ы х с у к к у л е н т о в он с о с т а в л я е т 3.5 ч. И з этого следует , ч т о , н е с м о т р я на более н и з к у ю и н т е н с и в н о с т ь т р а н с п и р а ц и и , запасы воды у склерофитов обновляются вдвое быстрее , чем у суккулентов .
В то же в р е м я , нельзя не отметить к р а й н е н и з к у ю с к о р о с т ь и с п а р е н и я воды с поверхности афилльных побегов АпаЬаз1з Ьгеуг/оИа, которая м о ж е т б ы т ь с в я з а н а как с а н а т о м и ч е с к и м и , т а к и ф у н к ц и о н а л ь н ы м и о с о б е н н о с т я м и этого вида. У суккулентов (особенно у видов с к о р о н а р н ы м с и н д р о м о м , с п о с о б н ы х к С ^ - С А М - ф о т о с и н т е з у , п о д о б н о АпаЪаз1з Ьгеу1/оИа) р е з е р в и р о в а н и е воды во внутренних тканях способствует более прочному ее удержанию и лучш е м у контролю н а д ее расходом (Гамалей , 1988).
= 0.624
10 20 30 40 50
4 п
3 -
'а 2
1 • 2 _ = 0.676
10 20 30 40 50 Парциальный объем межклетников, %
Рис. 7.5. З а в и с и м о с т ь с р е д н е в и д о в ы х значений и н т е н с и в н о с т и т р а н с п и р а ц и и однодольных (а) и двудольных (б) растений от п л о т н о с т и м е з о ф и л л а с о о т в е т с т в у ю щ и х
видов .

Глава 7. ИНТЕНСИВНОСТЬ ТРАНСПИРАЦИИ 163
Таблица 7.4
Средние в е л и ч и н ы интенсивности т р а н с п и р а ц и и (г^^^^ •^~с1х.массы ч " ' ) у р а з н ы х групп растений
Группа И X
Однодольные склерофиты 5811 1.288 0.846 Однодольные суккуленты 1792 1.588 0.853 Двудольные склерофиты 5875 1.833 1.125 Двудольный суккулент 359 0.501 0.372
Динамики средневидовых значе
Динамика средневидовых значений скорости и с п а р е н и я воды р а с т е н и я м и на г р а д и е н т е в л а ж н о с т и п о ч в ы обусловлена тем , что ни один из у ч т е н н ы х факторов среды (а и м е н н о — температ у р а и в л а ж н о с т ь в о з д у х а , о с в е щ е н ность), как показал статистический анализ, не влияют столь сильно на средние у р о в н и этого п о к а з а т е л я в о д о о б м е н а , как влажность п о ч в ы (рис. 7.6, а). О д нако видно , что р а з б р о с ы средних значений довольно велики. Ш и р и н а зоны р а з б р о с а д а н н ы х у в е л и ч и в а е т с я от аридной к г у м и д н о й ч а с т и градиента . Э т о х о р о ш о з а м е т н о п р и с р а в н е н и и трендов м и н и м а л ь н ы х и максимальных д л я к а ж д о г о с о о б щ е с т в а с р е д н е в и д о вых значений и н т е н с и в н о с т и транспирации (рис. 7,6, б).
Разность м е ж д у у к а з а н н ы м и трендами м и н и м а л ь н а в с а м о й сухой части градиента и макси мальна в его гумидной части. При этом т р е н д ы плавно изм е н я ю т с я вдоль г р а д и е н т а в л а ж н о с т и почвы. Скорость изменения м и н и м а л ь ных средневидовых значений интенсивн о с т и т р а н с п и р а ц и и п о ч т и в 3 р а з а меньше таковой для максимальных значений на всем п р о т я ж е н и и градиента . Поскольку и н т е н с и в н о с т ь т р а н с п и р а ции являет ся р е с у р с о з а в и с и м ы м про ц е с с о м ( е г о р е а л и з а ц и я в в ы с о к о й степени определяется запасами доступ-
й интенсивности транспирации
5 1 а
4 - 8 о
О 4 , . . о 50 100 150
Запас воды в почве, им
Рис. 7.6. Тренды средневидовых (а), миним а л ь н ы х (б — н и ж н я я л и н и я ) и м а к с и м а л ь н ы х (б — в е р х н я я линия ) д л я к а ж д о го с о о б щ е с т в а з н а ч е н и й и н т е н с и в н о с т и
т р а н с п и р а ц и и .
6*

164 Часть III. Закономерности структурно-функциональной организации растений.
ной почвенной влаги) , м о ж н о предположить , что р есу р сная ( экологическая) емкость экотопов плавно увеличивается вдоль градиента (в сторону п о в ы ш е н и я ) влажности почвы.
Вместе с тем, нельзя не отметить, что различия в скорости испарения воды между видами однодольных и двудольных определяют и различия в д и н а м и к е этого показа-
0 50 100 150 о 1 2 3 Запас воды в почве, мм а/(г-ч)
Рис. 7.7. Тренды средневидовых (а) значений и н т е н с и в н о с т и т р а н с п и р а ц и и о д н о дольных (а — 1) и двудольных (а — 2) растений ( п у н к т и р н ы м и л и н и я м и показаны доверительные интервалы), а также тренды разностей (б) между максимальными и минимальными для каждого сообщества сред-невидовыми значениями для всей совокупности д а н н ы х ( б — 1), д л я о д н о д о л ь н ы х
(б — 2) и двудольных (б — 3) растений.
Р и с . 7.8. З а в и с и м о с т ь м и н и м а л ь н ы х (нижние линии) и максимальных (верхние линии) средневидовых величин интенсивности транспирации от средних в кааодом сообществе значений этого показателя для однодольных (а) и двудольных
(б) растений .

Глава 7. ИНТЕНСИВНОСТЬ ТРАНСПИРАЦИИ 165
теля на градиенте . Однодольные растения н а большей части градиента испаряют воду примерно с одной и т о й же скоростью, л и ш ь в наиболее у в л а ж н е н н ы х экотопах скорость этого процесса резко возрастает (рис. 7.7, а - 1 ; = 0.652). Интенсивность транспирации двудольных, напротив, постоянно увеличивается на всем протяжении градиента (рис. 7.7, а -2 ; = 0.676). Однако разности м е ж д у м а к с и м а л ь н ы м и и м и н и м а л ь ными на каждом участке средневидовыми значениями д л я обеих г р у п п р а с т е н и й почти не различаются (рис. 7.7, 6-2, 3). Они в несколько раз меньше , чем разность межд у м и н и м а л ь н ы м и и м а к с и м а л ь н ы м и с р е д н е в и д о в ы м и в е л и ч и н а м и и н т е н с и в н о с т и транспирации, взятая без разделения всей совокупности объектов на группы (рис. 7.7, 6-1). Если принять , что р а с ш и р е н и е зоны р а з б р о с а д а н н ы х вдоль градиента влажности почвы отражает у в е л и ч е н и е экологической емкости экотопов , то нельзя не признать, что наличие в с о о б щ е с т в а х как однодольных, так и двудольных растений способствует наиболее п о л н о й утилизации ресурсов среды.
Увеличение зон ы р а з б р о с а средневидовых величин в наиболее у в л а ж н е н н ы х экотопах является о т р а ж е н и е м тесной , почти ф у н к ц и о н а л ь н о й з а в и с и м о с т и м и н и м а л ь н ы х и м а к с и м а л ь н ы х з н а ч е н и й от с р е д н и х (для экотопа ) у р о в н е й и н т е н с и в н о с т и транспирации (рис . 7.8; все к о э ф ф и ц и е н т ы д е т е р м и н а ц и и б о л ь ш е 0.94).
Динамика показателей вариации интенсивности транспирации
Обратимся к рассмотрению коэффициентов вариации выборок на градиенте влажности почвы (рис. 7.9). К о э ф ф и ц и е н т ы вариации интенсивности т р а н с п и р а ц и и имеют тенденцию к у в е л и ч е н и ю по мере уменьшения запасов влаги в почве (рис. 7.9, а, 6). Поэтому в определенном с м ы с л е может быть справедливо у т в е р ж д е н и е В . М . Свеш никовой (1979) об адаптивном значении этой тенденции (при корректной оценке лабильности к о э ф ф и ц и е н т а м и вариации) . С о статистической точки зрения это означает просто то, что по мере продвижения против градиента в л а ж н о с т и почвы (в сторону уменьшения запасов влаги в почве) величины стандартных отклонений приближаются к средним значениям (при отсутствии заметных различий м е ж д у видами однодольных и двудольных растений) .
Однако с у щ е с т в е н н ы м моментом этой тенденции является не просто у в е л и ч е н и е коэффициентов вариации против градиента в л а ж н о с т и почвы, а п о с т е п е н н о е р а с ш и рение зоны, в которой встречаются эти к о э ф ф и ц и е н т ы (рис . 7.9, в, г). В г у м и д н о й части градиента варьирование интенсивности т р а н с п и р а ц и и находится на одном (достаточно низком) уровне . По мере у м е н ь ш е н и я запасов влаги в почве диапазон , в котором наблюдаются с р е д н е в и д о в ы е к о э ф ф и ц и е н т ы вариации , р а с ш и р я е т с я .
***
И н т е н с и в н о с т ь т р а н с п и р а ц и и однодольных и двудольных р а с т е н и й р а з л и ч а е т с я на большей части градиента в л а ж н о с т и почвы. Это связано , судя по всему с различиями между э т и м и г р у п п а м и видов по плотности сложения м е з о ф и л л а . М е н ь ш а я скорость и с п а р е н и я в о д ы п р е д с т а в и т е л я м и г р у п п ы о д н о д о л ь н ы х п р е д о п р е д е л е н а меньшим п арц и альн ым объемом межклетников у этих растений . В то же время плотность сложения м е з о ф и л л а однодольных растений на градиенте п р а к т и ч е с к и не из меняется . Однако на этом о с н о в а н и и нельзя сделать з а к л ю ч е н и е о н е и з м е н н о й сте-
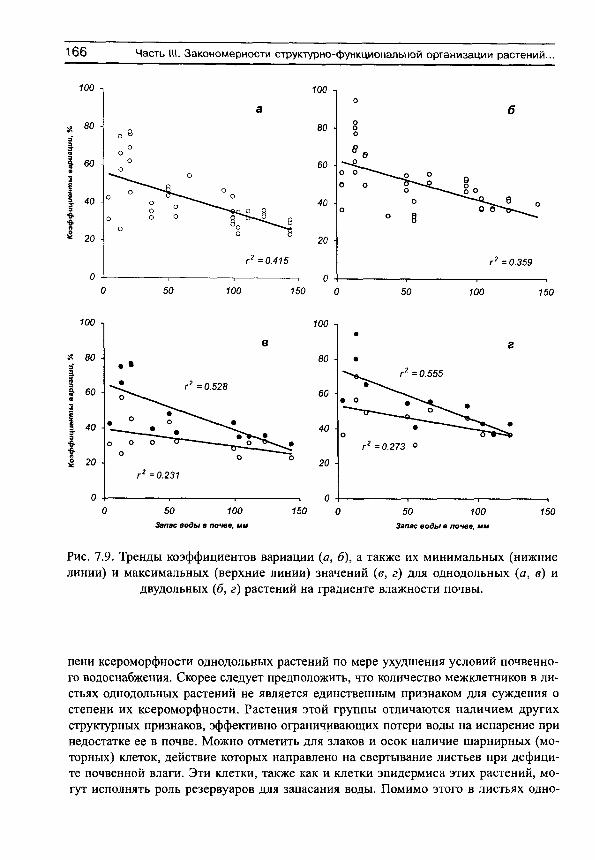
166 Часть III. Закономерности структурно-функциональной организации растений.
Рис. 7.9. Тренды к о э ф ф и ц и е н т о в вариации (а, б), а т а к ж е их м и н и м а л ь н ы х ( н и ж н и е линии) и м а к с и м а л ь н ы х ( в е р х н и е л и н и и ) значений {в, г) д л я о д н о д о л ь н ы х (а, в) и
двудольных (б, г) растений на градиенте в л а ж н о с т и п о ч в ы .
пени ксероморфности однодольных растений п о мере ухудшения у с л о в и й почвенного водоснабжения. Скорее следует предположить , что количество м е ж к л е т н и к о в в листьях однодольных р а с т е н и й не является е д и н с т в е н н ы м признаком для с у ж д е н и я о степени их к с е р о м о р ф н о с т и . Р а с т е н и я э т о й г р у п п ы о т л и ч а ю т с я н а л и ч и е м д р у г и х структурных признаков , эффективно о г р а н и ч и в а ю ш и х потери воды на испарение при недостатке ее в почве . М о ж н о отметить д л я злаков и о с о к н а л и ч и е ш а р н и р н ы х (моторных) клеток, д е й с т в и е которых направлено на свертывание листьев при д е ф и ц и те почвенной влаги . Эти клетки , т акже как и клетки э п и д е р м и с а этих р а с т е н и й , м о гут исполнять роль резервуаров для запасания воды. П о м и м о этого в л и с т ь я х одно-

Глава 7. ИНТЕНСИВНОСТЬ ТРАНСПИРАЦИИ 167
дольных часто достаточно х о р о ш о р а з в и т ы и с п е ц и а л и з и р о в а н н ы е в о д о з а п а с а ю щ и е ткани ' .
Поэтому, несмотря на то , что мы не м о ж е м дать с р а в н и т е л ь н у ю оценку степени ксероморфности двудольных и однодольных растений на о с н о в а н и и д а н н ы х о плотности мезофилла , мы все же можем сделать предположение , что именно этот признак оказывает влияние на наблюдаемые различия в скорости испарения воды между этими группами видов.
Разброс средневидовых величин интенсивности транспирации на градиенте влажности п о ч в ы и м е е т « к л и н о в и д н ы й » характер . Узкая часть « к л и н а » р а с п о л о ж е н а в аридной части градиента , ш и р о к а я — в г у м и д н о й (рис . 7.6). Сходство в и д о в в сообществах по р е а л и з а ц и и п р о ц е с с а испарения (и , видимо , п о г л о щ е н и я ) воды изменяется с изменением положения фитоценозов на градиенте в л а ж н о с т и почвы. В наиболее суровых условиях произрастания, с минимальным количеством доступной почвенной влаги п р о и з р а с т а ю т в и д ы с очень п о х о ж и м и з н а ч е н и я м и по каз ат елей водного режима. По мере продвижения вдоль градиента в сторону увеличения в л а ж н о с т и почвы сходство видов постоянно снижается , достигая м и н и м у м а в с а м ы х г у мидных экотопах.
Для того ч т о б ы обсудить этот и н т е р е с н ы й и в а ж н ы й э к о ф и з и о л о г и ч е с к и й факт сделаем р я д д о с т а т о ч н о с х е м а т и ч н ы х д о п у щ е н и й об и з м е н е н и я х э к о л о г и ч е с к о й и фитоценотической обстановок на градиенте влажности почвы.
1. Экологи ч еская е м к о с т ь экотопов у в е л и ч и в а е т с я вдоль г р а д и е н т а в л а ж н о с т и почвы. Поскольку, как уже было отмечено , т р а н с п и р а ц и я является в в ы с о к о й степени р е с у р с о з а в и с и м ы м п р о ц е с с о м (т. е. зависит от количества д о с т у п н о й почвенной влаги), экологическая емкость в данном случае связана, п р е ж д е всего, с количеством воды в почве .
2. Ч и с л о видов (ценопопуляций) в каждом с о о б щ е с т в е и м е е т т е н д е н ц и ю увеличиваться пропорционально увеличению экологической емкости экотопов. Это в какой-то степени может отражать т е н д е н ц и ю увеличения количества экониш в более гумидных местообитаниях . П о о п р е д е л е н и ю Т. А. Работнова (1973) количество н и ш в растительных сообществах соответствует количеству видов . Однако в связи с неоднозначностью самой проблемы экологической н и ш и у растений (Василевич , 1983) м ы не будем проводить прямого сопоставления емкости экотопа и количества экониш в нем. Вместе с т е м о т м е т и м , ч т о п л о т н о с т ь ц е н о п о п у л я ц и й и н а с ы щ е н н о с т ь с о о б щ е с т в видами увеличивается , а п р о с т р а н с т в е н н о е р а з о б щ е н и е ц е н о п о п у л я ц и й в ф и т о ц е н о -
' Кроме того, между однодольными и двудольными растениями существуют и другие существенные различия. Листья однодольных имеют поляризованный (интеркалярный) рост из точечного источника клеток в основании листа. Таким образом, наиболее старые клетки находятся в верхней части листовых пластинок. С другой стороны, листья двудольных имеют много точек роста, мозаично расположенных на пластинке, а клоны новых клеток образуются на протяжении всего периода роста и развития листьев. В связи с этим однодольные и двудольные растения различаются и по характеру распределения устьиц. У двудольных устьица распределены на листовой пластинке достаточно равномерно (или случайно). Молодые листья однодольных совсем не имеют устьиц. На развитых листьях этой группы растений устьица сгруппированы в линейные цепочки, внутри которых они чередуются с обычными эпидермаль-ными клетками (Сгох(1а1е, 1998, 2000; МаЛш, 01оуег, 1998; Нетапйег е1 а1„ 1999).

168 Часть III. Закономерности структурно-функциональной организации растений.
зах уменьшается вдоль градиента в л а ж н о с т и почвы. Так, в ковыльково-луково-баглу-ровом с о о б щ е с т в е о б щ е е п р о е к т и в н о е покрытие составляло 3-5% (запас н а д з е м н о й фитомассы изменялся в р а з н ы е по у с л о в и я м у в л а ж н е н и я годы от 13 до 30 гм"^) (Казанцева, 1986), в то в р е м я как в п с а м м о ф и т н о - с т е п н ы х с о о б щ е с т в а х — около 60% ( п л о т н о с т ь п о б е г о в с о с т а в л я л а 1200 экз .м"^, з а п а с н а д з е м н о й ф и т о м а с с ы — 8 0 -190 гм"^, видовая н а с ы щ е н н о с т ь — с в ы ш е 50 видов на 100 м^), на о с т е п н е н н ы х луг а х — 60-80% ( п л о т н о с т ь п о б е г о в — 1500 экз.-м'^, запас н а д з е м н о й ф и т о м а с с ы — 180-270 гм"^, видовая н а с ы щ е н н о с т ь — 30-35 видов на 100 м^), на н а с т о я щ и х и заболоченных п о й м е н н ы х л у г а х — 80-90% (плотность побегов — 1100-2000 э к з . м ^ запас н а д з е м н о й ф и т о м а с с ы — 140-610 г м ^ видовая н а с ы щ е н н о с т ь — с в ы ш е 6 0 -80 видов на 100 м^) (Титов, Печенюк , 1990).
3. Экотопический отбор, наиболее строгий в аридной части градиента , слабо проявляется в его гумидной части, где преимущественно произрастают долгопоемные заболоченные и с р е д н е п о е м н ы е настоящие луга (Титов , П е ч е н ю к , 1990). С о о б щ е с т в а этих типов лугов являются наименее у с т о й ч и в ы м и и с л а б о о р г а н и з о в а н н ы м и системами: р а з н о г о д и ч н ы е флуктуации урожая и обилия видов по массе в ы р а ж е н ы в них достаточно резко (Титов , П е ч е н ю к , 1990).
Экотопический отбор , судя по всему, не связан с к о н к у р е н т н ы м и о т н о ш е н и я м и между растениями и является тем первым грубым фильтром, который отсеивает виды, экологическая норма которых не соответствует условиям конкретного местообитания , не позволяя им давать всходы. В аридных экосистемах он р а з р е ш а е т возникать популяциям, и м е ю щ и м по классификации Д. Р. О п т е (1979), толерантную стратегию, что отражает н а и б о л ь ш у ю строгость этого отбора ' . В средней и г у м и д н о й частях градиента возникают п о п у л я ц и и с конкурентными и/или ко нку р ент но -р у дер аль ными стратегиями, что связано с достаточным количеством р есу р со в (Бигон и др . , 1989) и слабой н а п р я ж е н н о с т ь ю экотопического отбора.
' Этому не противоречит широкое распространение в аридных экосистемах растений с С^-син-дромом. С^-фотосинтез является дополнительным к стандартному С,-пути и увеличивает эффективность фиксации СО^. С^-растения используют в мезофилле фосфоенолпируват-карбок-силазу (РЕРС) для ассимиляции атмосферной углекислоты 3-углеродным фосфоенолпирува-том, в результате чего образуется 4-углеродные соединения. Они транспортируется затем в клетки обкладок пучков, где подвергаются декарбоксилированию, что создает внутри клеток относительно высокую концентрацию углекислоты. Эта углекислота присоединяется к рибу-лоздифосфату (КиВР) посредством ИиВР-карбоксилазы (К.иВ18СО) уже в цикле Кельвина (Ке11о§, 1998). Таким образом, С^-растения могли иметь повышенную конкурентоспособность в периоды геологической истории с низкой концентрацией углекислоты и высокой температурой воздуха (МсАШкгег е1 а1., 1998; Мопвоп, 1999, цит по: Ьоот18, А т Л о г , 1999; \Vа^с ,̂ 81га1п, 1999; Е(1\Vа̂ (18 е1 а1., 2001). Возникновение С^-фотосинтеза относят ко времени 15 миллионов лет назад (Ке11о§, 2001), а его широкое распространение, повлекшее за собой глобальные экосистемные перестройки, имело место 7-5 миллионов лет назад (ЕЫепп§ег, СегИп§, 1995). С^-растения, особенно злаки, широко распространены в степных экосистемах Северной и Южной Америк, Африки, Австралии (Ке11о§, 2001; Ковалев, 2000) Однако эти растения, с их ярко выраженными адаптивными возможностями и очень высокими показателями фотосинтеза и роста (Гамалей и др., 1992), не могут, по-видимому, создавать конкурентные популяции, поскольку плотность ценопопуляций этих растений (из-за сильного пресса среды) несравнима с плотностью любой ценопопуляций более гумидных экосистем.

Глава 7. ИНТЕНСИВНОСТЬ ТРАНСПИРАЦИИ 169
20 а1 Н 2 шз
04 о б @б
о 0-50 50-УОО 100-150
Запас воды в почве, им
Рис. 7.10. Распределение экологических типов растений на градиенте влажности почвы.
1 — гигрофиты и гидрофиты, 2 — мезофиты, 3 — ксеромезофиты, 4 — мезоксерофиты, 5 — ксерофиты, 6 — суккуленты и суккуленты с коронарным синдромом.
И л л ю с т р а ц и е й и з м е н е н и я н а п р я ж е н н о с т и экотопического отбора н а г р а д и е н т е влажности почвы м о ж е т служить , по н а ш е м у м н е н и ю , р а с п р е д е л е н и е экологических типов р а с т е н и й (по о т н о ш е н и ю к у с л о в и я м почвенно г о у в л а ж н е н и я ) в р а з н ы х его частях (рис. 7.10). В а р и д н о й части градиента присутствуют только м е з о к с е р о ф и т ы , ксерофиты и суккуленты, с п р е и м у щ е с т в е н н ы м п р е о б л а д а н и е м ксер о фит о в . В средней части градиента м о ж н о наблюдать представителей всех экологических т и п о в (за исключением, разумеется , г и г р о ф и т о в и гидрофитов ) . В этих экологических условиях преобладание получают мезоксерофиты, а также ксеромезофиты и мезофиты. В гумидной части градиента м о ж н о встретить представителей всех без и с к л ю ч е н и я экологических типов . Отсутствие р я д а экологических групп в ар идно й и средней частях градиента м о ж н о рассматривать скорее как результат д е й с т в и я экотопического отбора, чем конкурентных о т н о ш е н и й м е ж д у р а с т е н и я м и . В г у м и д н о й части г р а д и е н т а снижение доли у ч а с т и я мезоксерофитов , ксерофитов и суккулентов обусловлено , по нашему м н е н и ю , м е ж в и д о в о й конкуренцией при отсутствии или слабо й в ы р а ж е н н о сти экотопического отбора (однако м о ж н о представить , что в м е с т о о б и т а н и я х , свойственных г и д р о ф и т а м , действует с т р о ж а й ш и й экотопический отбор; в этом о т н о ш е нии наиболее а р и д н ы е и г у м и д н ы е экотопы п о д о б н ы д р у г другу ) .
Таким образом, в т р а в я н ы х сообществах а р и д н ы х экосистем о т н о с и т е л ь н о в ы с о кое сходство реализации процесса транспирации у разных растений может быть обусловлено ж е с т к и м э к о т о п и ч е с к и м о т б о р о м (через который проходит н а б о р видо в , в наибольшей степени соответствующий данным условиям произрастания) и слабой выраженн ост ью конкурентных о т н о ш е н и й м е ж д у ними . П о пу ля ции , и м е ю щ и е толерантные ж и з н е н н ы е стратегии , испаряют такое количество воды, которое д о с т у п н о им в этих м е с т о о б и т а н и я х . П о с л е в ы п а д е н и я осадков или в о п ы т а х с по ливо м скорость и с п а р е н и я в о д ы м о ж е т в о з р а с т а т ь в 1.5-3 ра за (даже у такого вида , как АпаЪазгз Ьге\1/оИа). Д р у г и м и словами , ф у н к ц и о н а л ь н ы й потенциал этих р а с т е н и й может быть значительно в ы ш е наблюдаемого , однако о г р а н и ч е н н ы е и о д и н а к о в ы е д л я всех о с о -

170 Часть III. Закономерности структурно-функциональной организации растений...
бей (в силу отсутствия или слабой конкуренции) р е с у р с ы воды в почве выравнивают средние уровни интенсивности транспирации . В и д о с п е ц и ф и ч н о с т ь по этому признаку практически отсутствует.
В г у м и д н ы х м е с т о о б и т а н и я х , несмотря на то, что з апасы воды в почве неогран и ч е н н ы (или д о с т а т о ч н о о б и л ь н ы ) , в одном и т о м же с о о б щ е с т в е с о с у щ е с т в у ю т виды как с высокой , т а к и с низкой скоростью испарения воды (рис . 7.6) при достаточно слабом экотопическом отборе , н о при более н а п р я ж е н н ы х ко нку р ент ных отношениях м е ж д у в и д а м и . Д л и т е л ь н о е и у с п е ш н о е с о с у щ е с т в о в а н и е в ф и т о ц е н о з е ценопопуляций с конкурентными стратегиями, использующих один и тот же ресурс, возможно только при разделении и м и экологических н и ш (как в п р о с т р а н с т в е , так и во времени) . Н а б л ю д а е м о е разнообразие средних уровней и с п а р е н и я в о д ы в г у м и д н ы х экотопах, судя п о всему, и является отражением такой д и ф ф е р е н ц и а ц и и . Поскольку в результате р а з д е л е н и я н и ш в и д ы сохраняют до .минирующее п о л о ж е н и е в сообществах, то и сп ользуемые ими запасы почвенной влаги д о л ж н ы отвечать их конституц и о н н ы м п от ребностям . П о э т о м у можно предположить , что у в е л и ч е н и е разнообразия средних у р о в н е й скорости испарения воды на градиенте в л а ж н о с т и почвы является следствием у в е л и ч е н и я разнообразия в и д о с п е ц и ф и ч е с к и х р е а к ц и й растений на изменения экологической обстановки . Но поскольку одни и те же в и д ы в разных экотопах легко и з м е н я ю т эти средние уровни , то м ы с к л о н н ы р а с с м а т р и в а т ь п о д о б н у ю видоспецифичность как в высокой степени относительную, не д о с т и г ш у ю уровня определенной функциональной специализации. Накопленный фактический материал показал , что р а з л и ч и я м е ж д у в и д а м и на г р а д и е н т е в л а ж н о с т и п о ч в ы н о с я т скорее «структурноспецифический» характер (что связано с принадлежностью к таксонам такого ранга, как однодольные и двудольные) .
Как уже у п о м и н а л о с ь в ы ш е , р а з б р о с к о э ф ф и ц и е н т о в в а р и а ц и и и н т е н с и в н о с т и транспирации на градиенте влажности почвы, также как и разброс средних значений, имеет « к л и н о в и д н ы й » характер (рис. 7.9). Различие состоит в том , что узкая часть «клина» находится в гумидной части градиента , а широкая — в аридной . В г у мидных экотопах, как видно из представленных данных, все коэффициенты вариации находятся на одном у р о в н е . В н а и б о л е е с у х и х местообитаниях могут встречаться как очень низкие к о э ф ф и ц и е н т ы вариации , так и очень высокие . Это может означать , что в ус ловиях с о граниченными запасами почвенной влаги, где диапазон экологических н и ш по определению очень узок , растения избегают м е ж в и д о в о й к о н к у р е н ц и и за воду не только за счет д и ф ф е р е н ц и а ц и и экониш (следует учитывать з амет но е пространственное р а з о б щ е н и е м е ж д у особями) , но и за счет р а з н ы х д и а п а з о н о в ф у н к ц и о н а л ь н о й изменчивости , о п р е д е л я е м ы х структурой . О д н и в и д ы и з м е н я ю т ско р о ст ь испарения воды вслед за каждым изменением экологической обстановки, другие довольствуются теми н е б о л ь ш и м и в о з м о ж н о с т я м и , которые предоставляет и м экотоп д л я поддержания жизнедеятельности . О б этом же свидетельствует зависимость к о э ф ф и ц и е н т о в вариации от с р е д н е в и д о в ы х значений интенсивности т р а н с п и р а ц и и . У в и д о в с низкими в среднем значениями скорости испарения воды зона р а з б р о с а коэффициентов вариации наиболее широка , в противоположност ь видам с в ы с о к и м и в е л и ч и н а м и интенсивности т р а н с п и р а ц и и .
В г у м и д н ы х экотопах п о д в и ж н о с т ь и н т е н с и в н о с т и т р а н с п и р а ц и и находится в очень узком диапазоне значений , несмотря на б о л ь ш и е р а з л и ч и я в средних уровнях

Глава 7. ИНТЕНСИВНОСТЬ ТРАНСПИРАЦИИ 171
процесса т р а н с п и р а ц и и . Это может быть связано только с т е м , что в д н е в н о й и сезонной д и н а м и к е этот показатель в малой степени коррелирует с и з м е н е н и я м и факторов среды, что является частным случаем низкой с л о ж н о с т и о р г а н и з а ц и и водного режима растений в местообитаниях с в ы с о к и м и запасами в л а ж н о с т и почвы.
Резюме. Статистический анализ интенсивности транспирации на градиенте влажности почвы показал следующее . Весь массив исходных д а н н ы х интенсивности транспирации имеет среднее значение х = 1.538 Гд^^^ -^сух.массы ' ч " ' • Т и п и ч н ы е значения интенсивности т р а н с п и р а ц и и травянистых растений на градиенте в л а ж н о с т и почвы не превышают 3 т^^^,^ -^сухмассы • • Оптимальные для транспирации диапазоны температуры и д е ф и ц и т а в л а ж н о с т и воздуха составляют соответственно 20-28 °С и 5 -24 мб. Наиболее з аметные различия по скорости т р а н с п и р а ц и и м о ж н о у видет ь между видами однодольных и двудольных растений . С о о т в е т с т в у ю щ и е средние значения этого признака р а в н ы 1.358 и 1.759 г^„^„ - Гсух.массы ' ч " ' • П о к а з а н о , что р а с т е н и я с более п л о т н ы м м е з о ф и л л о м при прочих р а в н ы х условиях и с п а р я ю т воду медленнее , чем виды с б о л ь ш и м количеством межклетников в листьях . В о всех местообитаниях у однодольных р а с т е н и й листья более плотные , чем у двудольных , что, п о - в и д и м о му, и определяет различия в скорости испарения воды этими г р у ппами растений . Однодольные суккуленты и с п а р я ю т воду значительно и н т е н с и в н е е , чем однодольные склерофиты. Однако запасы воды у склерофитов о б н о в л я ю т с я вдвое б ы с т р е е , чем у суккулентов. Ш и р и н а зоны разброса средневидовых значений увеличивается от аридной к гумидной части градиента. Это иллюстрируется сравнением трендов минимальных и макси мальн ых для каждого сообщества с р е д н е в и д о в ы х значений инт енсивно сти транспирации . П р и исследовании к о э ф ф и ц и е н т о в в а р и а ц и и в ы я в л е н о постепенное р а с ш и р е н и е зоны, в которой встречаются эти к о э ф ф и ц и е н т ы против градиента влажности почвы. В г у м и д н о й части градиента варьирование и н т е н с и в н о с т и т р анс пирации находится на одинаковом для всех растений уровне . П о мере у м е н ь ш е н и я запасов влаги в почве диапазон , в котором наблюдаются с р е д н е в и д о в ы е к о э ф ф и ц и е н ты вариации, расширяется . Сделаны следующие предположения . В травяных сообществах аридных экосистем относительно высокое сходство реализации процесса транспирации у р а з н ы х растений может быть обусловлено ж е с т к и м экотопическим отбором и слабой в ы р а ж е н н о с т ь ю конкурентных о т н о ш е н и й м е ж д у н и м и . Н а б л ю д а е м о е разнообразие средних у р о в н е й испарения воды в г у м и д н ы х экотопах я вля ет ся отражением д и ф ф е р е н ц и а ц и и э к о н и ш . Различия м е ж д у в и д а м и на г р а д и е н т е в л а ж н о с т и почвы носят « с т р у к т у р н о с п е ц и ф и ч е с к и й » характер , что связано с п р и н а д л е ж н о с т ь ю к таксонам такого высокого ранга , как однодольные и двудольные . В у с л о в и я х с ограниченными запасами почвенной влаги растения избегают м е ж в и д о в о й конкуренции за воду не только за счет д и ф ф е р е н ц и а ц и и экониш, но и за счет р а з н о й л а б и л ь н о с т и в использовании одного и того же р е с у р с а среды. Низкая п о д в и ж н о с т ь и н т е н с и в н о сти т р а н с п и р а ц и и в г у м и д н ы х экотопах связана с низкой с л о ж н о с т ь ю о р г а н и з а ц и и водного р е ж и м а растений .

172 Часть III. Закономерности структурно-функциональной организации растений..
Г Л А В А 8. Д Е Ф И Ц И Т В О Д Н О Г О Н А С Ы Щ Е Н И Я
Д е ф и ц и т водного н а с ы щ е н и я как с а м о с т о я т е л ь н ы й показатель водного обмена растений исследовался м н о г и м и отечественными авторами в р а з н ы х экологических условиях (Горшкова, 1971; Алексеенко, 1976; Банникова и др. . 1982; Измайлова , 1978, 1986; Свешникова , 1979, 1988, 1993; Горшкова, Зверева, 1982; Горшкова и др . , 1982; Бобровская , 1991, 1994 и д р . ) ' .
В листьях растений практически всегда наблюдается в ы р а ж е н н ы й в той или иной степени д е ф и ц и т н а с ы щ е н и я водой. Известно, что целый ряд факторов среды способствует в о з н и к н о в е н и ю и р а з в и т и ю водного д е ф и ц и т а . Это низкая температура , плохая аэрация корнеобитаемого слоя, его засоление, недостаток почвенной влаги, тяжелый гранулометрический состав почвы. Увеличение д е ф и ц и т а н а с ы щ е н и я при усилении н а п р я ж е н н о с т и ф а к т о р о в с р е д ы может стать п р и ч и н о й н а р у ш е н и я п р о ц е с с о в фотосинтеза из-за с н и ж е н и я поглощения СО^, процессов р о с т а и р а з в и т и я (Слейчер , 1970, Ва811(1е е ! а1., 1993) В м е с т е с тем это приводит к п о я в л е н и ю комплекса регуля-торных процессов на у р о в н е экспрессии генов, к о м п е н с и р у ю щ и х негативные последствия недостаточного н а с ы щ е н и я листьев водой (Вгау, 1993).
П р и н я т о считать , что водный д е ф и ц и т отражает степень н а п р я ж е н н о с т и водного режима растений (см.: Бобровская, 1991). Однако, несмотря на б о л ь ш у ю важность этого параметра в экологии водообмена и заметное число публикаций на эту тему, в литературе нет о п и с а н и й его изменений на сколько-нибудь з н а ч и т е л ь н о м градиенте влажности почвы. В о с п о л н я я в какой-то степени этот пробел , м ы п о с т а в и л и цель: 1) дать статистическое описание большого массива данных, с о б р а н н ы х в широком спектре экологических условий , 2) показать закономерности и з м е н е н и й этого показателя на градиенте в л а ж н о с т и почвы.
Статистический анализ дефицита водного насыщения у видов однодольных и двудольных растений
Д а н н ы е с о б р а н ы в 14 с о о б щ е с т в а х , р а с п о л о ж е н н ы х в р а з н ы х частях градиента влажности почвы (см. в табл . 7.1 сообщества за и с к л ю ч е н и е м 4 и 5). Статистические
' В работах зарубежных исследователей этот показатель используется при моделировании водного обмена растений ( Т Ь о т к у , 1996; ТЬот1еу, СаппеИ, 1997), при анализе (совместно с водным потенциалом) кривых "давление-объем" (Тугее, Н а т т е 1 , 1972; СЬеип^ е1 а1., 1976; Титег, 1981; Тугее, ШсЫег, 1981, 1982; Тугее, 1аг^18, 1982; Оа1е, ЗШсЬГГе, 1986; Ко1с1е е1 а1., 1989). Кривые "давление-объем" являются одним из наиболее широко используемых инструментов для получения информации о водном статусе растений (ВескеИ, 1997). Их анализ позволяет рассчитать ряд дополнительных характеристик водного режима: осмотический потенциал при полном насыщении, осмотический потенциал при полной потере тургора, относительное содержание воды в точке потери тургора, объем фракции апопластической (и, следовательно, симпластической) воды при полном насыщении, объемный модуль эластичности клеточных стенок. Вместе с тем, существует достаточно обоснованное мнение о некорректности определений терминов «водный потенциал» и «осмотическое давление» и значения этих параметров в транспорте воды по растению (Мелещенко, 2001). Показана также ограниченность объемного модуля эластичности из-за его сильных изменений в зависимости от тургора клетки (Мелещенко, 1997).

Глава 8. ДЕФИЦИТ ВОДНОГО НАСЫЩЕНИЯ 173
характеристики с р е д н е в и д о в ы х величин дефицита водного н а с ы щ е н и я показаны в таблице 8.1.
Весь массив исходных д а н н ы х де фицита водного насыщения имеет среднее значение х = 17.8% и стандартное отклонение = 12.5%) (и = 13314). Ряд распределения имеет п о л о ж и т е л ь н у ю асимметрию с модой, равной 12.5%), а 90%) его значений (вариант) находятся в диапазоне 0-30.1%) (остальные 10%) в а р и а н т р а с п р е д е л е н ы в д и а п а з о н е 30.1-86.5%). Е с л и п р и н я т ь в е р х н и й дециль ряда распределения за границу типичных изменений признака, то можно с к а з а т ь , ч т о т и п и ч н ы е з н а ч е н и я дефицита водного н асыщения травянистых растений на градиенте влажнос ти почвы не п р е в ы ш а ю т 30%.
С у щ е с т в е н н ы е различия по д а н н о м у признаку о б н а р у ж е н ы м е ж д у в и д а м и однодольных и двудольных растений . Соответствующие средние значения р а в н ы 15.0 и 21.4%. Статистическая оценка разности средних показывает , что она в высокой степени достоверна ( Р < 0.001; объемы выборок р а в н ы соответственно 7439 и 5875, дисперсии — 89.2 и 217.8, Г-критерий Стьюдента — 28.6). В е р х н и е децили рядов распределения водного д е ф и ц и т а однодольных и двудольных растений р а в н ы 26.0 и 35.3%.
В наи большей степени р а з л и ч и я м е ж д у этими группами р а с т е н и й проявляются в аридной части градиента (рис. 8.1). Здесь (в диапазоне запасов влаги в почве 0-50 мм) средняя в е л и ч и н а д е ф и ц и т а в о д н о г о н а с ы щ е н и я у в и д о в о д н о д о л ь н ы х р а в н а 21 .1%, у видов д в у д о л ь н ы х — 33.7%). При п р о д в и ж е н и и вдоль градиента в л а ж н о с т и п о ч в ы п р о и с х о д и т р е з к о е ( п о ч т и в 2 раза ) с н и ж е н и е д е ф и ц и т а н а с ы щ е н и я , м е ж групповые различия сглаживаются (не п р е в ы ш а я 3-5%), хотя и о с т а ю т с я статистически д о с т о в е р н ы м и .
Зависимость дефицита водного насыщения от факторов среды
М н о ж е с т в е н н ы й р е г р е с с и о н н ы й анализ показал , что о с н о в н ы м и ф а к т о р а м и сред ы , воздей ст вую щи ми на д е ф и ц и т водного н а с ы щ е н и я в листьях р а с т е н и й , является влажность п о ч в ы и воздуха . П о с к о л ь к у ш и р и н а р а з б р о с а с р е д н е в и д о в ы х значений водного д е ф и ц и т а достаточно узка и не изменяется с у щ е с т в е н н о на г р адиент ах этих факторов среды (рис. 8.2), целесообразно провести анализ 1) средних у р о в н е й (средних для каждого участка значений) , 2) с р е д н е в и д о в ы х значений всего массива дан ных, 3) с р е д н е в и д о в ы х значений этого показателя у о дно до ль ны х и д в у д о л ь н ы х р ас тений при и змен ен и и условий у в л а ж н е н и я экотопов.
Д и н а м и к а с р е д н и х у р о в н е й д е ф и ц и т а в о д н о г о н а с ы щ е н и я н а г р а д и е н т а х влажност и п о ч в ы и воздуха . П о д средними у р о в н я м и какого-либо показателя мы
40 -,
0-50 50-^00 ГОО-150 Запас воды в почве, мм
Рис. 8.1. Средние значения д е ф и ц и т а водного н а с ы щ е н и я в л и с т ь я х о дно до ль ны х (1) и д в у д о л ь н ы х (2) р а с т е н и й в р а з н ы х частях
градиента в л а ж н о с т и почвы.
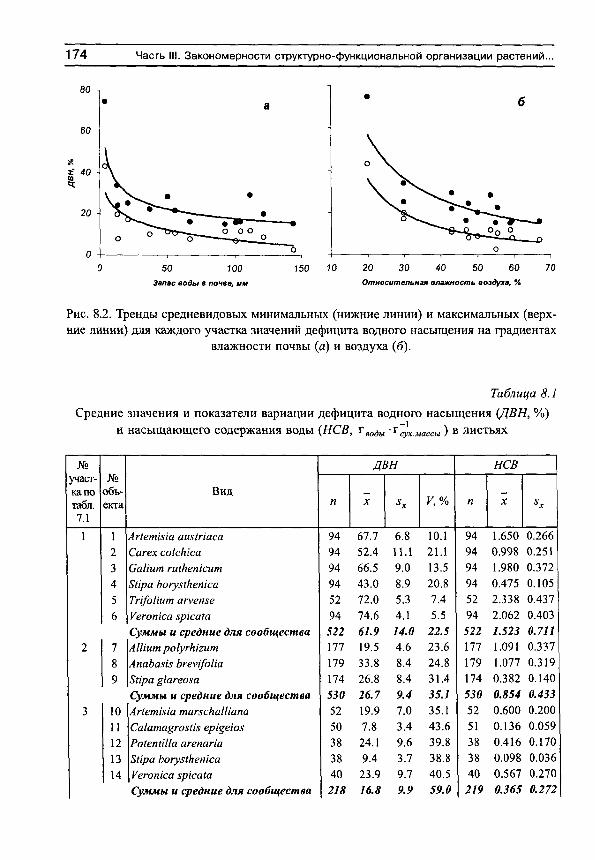
174 Часть III. Закономерности структурно-функциональной организации растений.
Рис. 8.2. Тренды средневидовых минимальных (нижние линии) и максимальных (верхние линии) для каждого участка значений дефицита водного насыщения на градиентах
влажности почвы (а) и воздуха (б) .
Таблица 8.1
Средние значения и показатели вариации д е ф и ц и т а водного н а с ы щ е н и я (ДВН, %)
и н а с ы щ а ю щ е г о с о д е р ж а н и я воды {НСВ, ^„оды '^'сух.массы) в л и с т ь я х
№ участка по табл. 7.1
№ обь-екта
Вид
ДВН НСВ № участка по табл. 7.1
№ обь-екта
Вид п X К, % п X ^х
1 1 Аг1етта аиз1паса 94 67.7 6.8 10.1 94 1.650 0.266 2 Сагех со1сЫса 94 52.4 11.1 21.1 94 0.998 0.251 3 ОаИит гиЛетсит 94 66.5 9.0 13.5 94 1.980 0.372
4 8йра Ъогуикетса 94 43.0 8.9 20.8 94 0.475 0.105 5 Тп/оНит апепзе 52 72.0 5.3 7.4 52 2.338 0.437
6 Уеготса 5р\са1а 94 74.6 4.1 5.5 94 2.062 0.403 Суммы и средние для сообщества 522 61.9 14.0 22.5 522 1.523 0.711
2 7 АШит ро1угЫ2ит 177 19.5 4.6 23.6 177 1.091 0.337 8 АпаЬаш Ьгеу1/оИа 179 33.8 8.4 24.8 179 1.077 0.319
9 5Нра ^1агеова 174 26.8 8.4 31.4 174 0.382 0.140 Суммы и средние для сообщества 530 26.7 9.4 35.1 530 0.854 0.433
3 10 Аг1етта тагзскаШапа 52 19.9 7.0 35.1 52 0.600 0.200
11 Са1ата^гозИз ерщеюз 50 7.8 3.4 43.6 51 0.136 0.059
12 Ро1еп1И1а агепапа 38 24.1 9.6 39.8 38 0.416 0.170
13 8йра ЬогухЛетса 38 9.4 3.7 38.8 38 0.098 0.036
14 Уегопка 5р1са(а 40 23.9 9.7 40.5 40 0.567 0.270
Суммы и средние для сообщества 218 16.8 9.9 59.0 219 0.365 0.272

Глава 8. ДЕФИЦИТ ВОДНОГО НАСЫЩЕНИЯ 175
Продолжение таблицы 8.1
№ участка по табл. 7.1
№ объекта
ДВН НСВ № участка по табл. 7.1
№ объекта
Вид п X К,% п X ^х
6 15 АШит ро1уг1г12ит 180 16.9 3.4 20.1 180 0.916 0.209 16 АпаЬаш Ьгеу1/оНа 180 25.0 8.5 34.2 180 0.916 0.300 17 8Ира §1агеоха 167 24.7 7.3 29.6 167 0.376 0.124
Суммы и средние для сообщества 527 22.1 7.7 35.0 527 0.745 0.337 7 18 Сагех (1игшзси1а 228 11.1 5.3 23.9 230 0.423 0.115
19 С1еШо§епез здиаггош 228 14.7 5.0 33.8 228 0.260 0.095 20 Ьеутиз сЫпепш 228 10.2 4.6 44.7 229 0.192 0.092 21 Ро1еп1И1а Ы/игса 228 19.5 6.0 30.5 228 0.390 0.149
Суммы и средние для сообщества 912 16.6 7.0 41.8 915 0.317 0.148 8 22 АПетша /п^Ша 438 28.2 10.4 36.8 438 0.795 0.263
23 Мз (ИсНо1ота 437 11.4 6.3 55.4 442 0.611 0.364 24 Ьеутиз сНтепш 441 12.8 6.3 49.0 441 0.219 0.115 25 Ро1еп1И1а 1апасе11/оНа 416 23.4 6.5 27.7 416 0.553 0.183 26 РикайИа гигсгаптоун 437 14.3 6.2 43.1 438 0.346 0.171 27 8йра кгу1оуИ 442 23.4 6.8 28.8 442 0.293 0.096
Суммы и средние для сообщества 2611 18.9 9.6 50.9 2617 0.468 0.298 9 28 СаЬта^гозйз ерщеюз 120 15.8 6.5 41.2 120 0.289 0.132
29 Сагех теШпозШсНуа 120 21.6 3.7 17.2 120 0.418 0.092 30 СаНит шкетсит 120 21.3 5.3 25.0 120 0.632 0.176 31 Ьу1кгит У1г§а1ит 120 20.6 4.3 20.8 120 0.428 0.121 32 8ап^13огЪа о^ПсшаНз 120 10.6 5.0 47.3 120 0.218 0.116 33 ТапасеШт уи1§аге 120 14.1 3.9 28.0 120 0.374 0.106
Суммы и средние для сообщества 720 17.3 6.4 36.9 720 0.393 0.181 10 34 Сагех те1апо51аскуа 34 16.1 6.3 38.8 34 0.289 0.113
35 РШрепс1и1а иШапа 34 13.3 4.9 36.9 34 0.296 0.114 36 СаИит гиЫоШез 32 15.7 6.6 42.0 32 0.456 0.226
37 8ап^шзогЬа о^^^с^па^^з 34 7.8 3.2 41.1 34 0.200 0.095
Суммы и средние для сообщества 134 13.2 6.3 47.8 134 0.308 0.170
11 38 Сагех уи1рта 50 12.5 4.1 32.6 50 0.261 0.109
39 агзшт тсапит 50 13.5 4.0 29.8 50 0.621 0.211
40 СаНит гиЫоШез 50 12.6 3.8 29.9 50 0.330 0.122
41 Уеготса 1оп§1/оИа 50 11.5 3.9 33.6 50 0.293 0.109
42 Уша сгасса 50 14.9 4.3 28.9 49 0.402 0.115
Суммы и средние для сообщества 250 13.0 4.1 31.8 249 0.381 0.189 12 43 А1оресигиз агигиНпасеиз 280 1.Ъ 2.1 28.5 280 0.186 0.048
44 Сагех а1кего(1ез 284 9.2 2.4 26.3 284 0.209 0.062
45 Сагех ШкоркИа 284 15.8 3.0 19.0 284 0.367 0.095
46 Е1у1гщ1а герепз 242 14.1 5.1 35.9 242 0.291 0.082
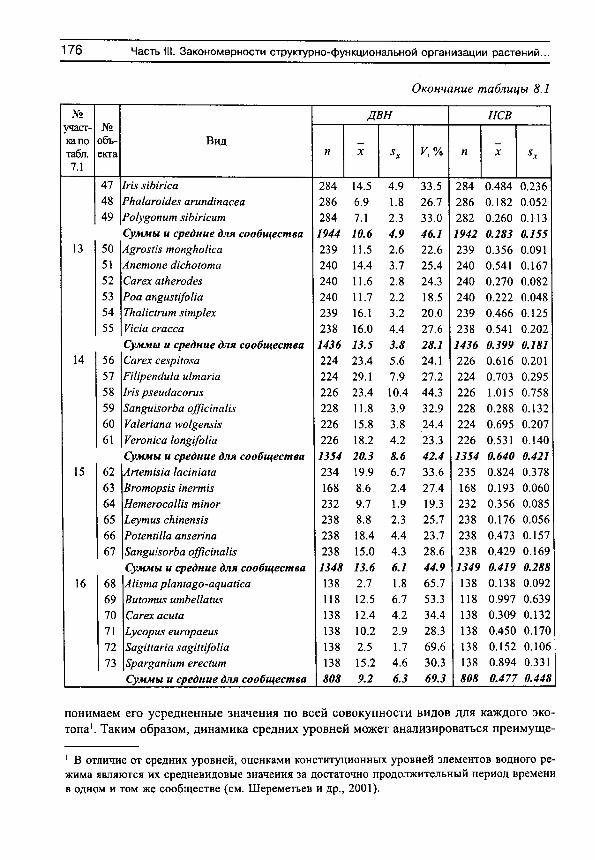
176 Часть III. Закономерности структурно-функциональной организации растений.
Окончание таблицы 8.1
№ №
ДВН НСВ участ № ка по обь- Вид _ табл. 7.1
екта п X У,% п X ^х
47 1пз 81Ыпса 284 14.5 4.9 33.5 284 0.484 0.236 48 РНа1агоШе5 агипсИпасеа 286 6.9 1.8 26.7 286 0.182 0.052 49 Ро1у^опит згЫпсит 284 7.1 2.3 33.0 282 0.260 0.113
Суммы и средние для сообщества 1944 10.6 4.9 46.1 1942 0.283 0.155 13 50 А^гозйз топ^НоИса 239 11.5 2.6 22.6 239 0.356 0.091
51 Апетопе еИско1ота 240 14.4 3.7 25.4 240 0.541 0.167 52 Сагех а1Негос1ез 240 11.6 2.8 24.3 240 0.270 0.082 53 Роа ап§изН/оИа 240 11.7 2.2 18.5 240 0.222 0.048 54 ТНаИс1гит зтр1ех 239 16.1 3.2 20.0 239 0.466 0.125 55 УШа сгасса 238 16.0 4.4 27.6 238 0.541 0.202
Суммы и средние для сообщества 1436 13.5 3.8 28.1 1436 0.399 0.181 14 56 Сагех сезрНоза 224 23.4 5.6 24.1 226 0.616 0.201
57 РШреп4и1а и1тапа 224 29.1 7.9 27.2 224 0.703 0.295 58 Мз рзеи<1асогиз 226 23.4 10.4 44.3 226 1.015 0.758 59 5ап^13огЬа о^]Г1апаИз 228 11.8 3.9 32.9 228 0.288 0.132 60 Уа1епапа м'о1^епз1з 226 15.8 3.8 24.4 224 0.695 0.207 61 Уегопка 1оп§1/оИа 226 18.2 4.2 23.3 226 0.531 0.140
Суммы и средние для сообщества 1354 20.3 8.6 42.4 1354 0.640 0.421 15 62 Аг1ет181а 1асШа1а 234 19.9 6.7 33.6 235 0.824 0.378
63 Вготорз13 тегтгз 168 8.6 2.4 27.4 168 0.193 0.060 64 НетегосаШз ттог 232 9.7 1.9 19.3 232 0.356 0.085 65 Ьеутиз скшепзгз 238 8.8 2.3 25.7 238 0.176 0.056 66 Ро1еп1И1а апзеппа 238 18.4 4.4 23.7 238 0.473 0.157 67 8ап§шзогЬа о^1стаНз 238 15.0 4.3 28.6 238 0.429 0.169
Суммы и средние для сообщества 1348 13.6 6.1 44.9 1349 0.419 0.288 16 68 АИзта р1ап1а§о-ациаИса 138 2.7 1.8 65.7 138 0.138 0.092
69 ВШотиз итЬеНаШз 118 12.5 6.7 53.3 118 0.997 0.639 70 Сагех асШа 138 12.4 4.2 34.4 138 0.309 0.132 71 Ьусориз еигораеиз 138 10.2 2.9 28.3 138 0.450 0.170 72 8а§ШаНа за§Ш1/оНа 138 2.5 1.7 69.6 138 0.152 0.106 73 Зраг^атит егесШт 138 15.2 4.6 30.3 138 0.894 0.331
Суммы и средние для сообщества 1 808 9.2 6.3 69.3 808 0.477 0.448
понимаем его у с р е д н е н н ы е значения по всей с о в о к у п н о с т и в и д о в д л я каждого экотопа ' . Таким образом, д и н а м и к а средних у р о в н е й может анализироваться п р е и м у щ е -
' В отличие от средних уровней, оценками конституционных уровней элементов водного режима являются их средневидовые значения за достаточно продолжительный период времени в одном и том же сообществе (см. Шереметьев и др., 2001).

Глава 8. ДЕФИЦИТ ВОДНОГО НАСЫЩЕНИЯ 177
70 1
О 50 100 150 10 30 50 70 Запас воды в почве, мм Относительная влажность воздуха, %
Рис . 8.3. Д и н а м и к а с р е д н и х у р о в н е й д е ф и ц и т а в о д н о г о н а с ы щ е н и я н а г р а д и е н т а х в л а ж н о с т и почвы {а) и воздуха (б).
ственно на градиентах факторов среды. А н а л и з из менений с р е д н и х у р о в н е й различных параметров водного р е ж и м а является надежной основой предсказания их наиболее вероятных значений (в пределах точности регрессионных моделей , о п и с ы в а ю щ и х динамику показателей на градиентах факторов среды) при р а з н ы х условиях произрастания .
Средние уровни д е ф и ц и т а водного н а с ы щ е н и я т е с н о связаны с в л а ж н о с т ь ю почвы и воздуха (рис . 8.3). Н а и б о л е е отчетливо эти связи проявляются в сухой части градиента в л а ж н о с т и почвы с запасами влаги менее 50 м м (в в е р х н е м 30-сантиметровом слое почвы) (рис . 8.3, в ) и в условиях с п о н и ж е н н о й (менее 40-50%) относительной в л а ж н о с т ь ю воздуха (рис. 8.3, б ) .
М н о ж е с т в е н н ы й р е г р е с с и о н н ы й анализ позволил выявить , что с р е д н и е у р о в н и водного д е ф и ц и т а и з м е н я ю т с я на градиентах в л а ж н о с т и почвы и воздуха в соответствии с в ы р а ж е н и е м :
У = а - н 6 е х р { с ( ^ 2 , - ь е 2 2 ) } , (6)
где
2^ ^-Н-кЩ-Х^),
У— д е ф и ц и т водного н а с ы щ е н и я , %>; — запас влаги (в слое п о ч в ы 0-30 см) , м м ; Х^ — относительная в л а ж н о с т ь воздуха, %; а (7.31), Ъ (8.808), с (0.806). с! (-0.475), е (-0.539), / ( - 1 . 4 6 3 ) , § (0.022), Н (-11.125), к (3.06), / (87.0) — к о э ф ф и ц и е н т ы . Коэффициент множественной детерминации равен 0.800, / " -критерий — 22.0 при степенях свободы 2 и 11, частные 7^-критерии для в л а ж н о с т и п о ч в ы и воздуха р а в н ы соответственно 8.6 и 11.1. Среднее относительное отклонение модели от ф а к т и ч е с к и х

178 Часть III. Закономерности структурно-функциональной организации растений.
данных составляет 14.5%. Средняя квадратичная ошибка аппроксимации равна 18.8%). Из этих д а н н ы х видно , что регрессионная модель удовлетворительно о п и с ы в а е т д и намику д е ф и ц и т а водного н а с ы щ е н и я на градиентах в л а ж н о с т и п о ч в ы и воздуха.
Д и н а м и к а с р е д н е в и д о в ы х з н а ч е н и й д е ф и ц и т а водного н а с ы щ е н и я на г ра д иентах в л а ж н о с т и п о ч в ы и воздуха. Ш и р и н а зоны р а з б р о с а с р е д н е в и д о в ы х значений д е ф и ц и т а водного н а с ы щ е н и я на градиентах влажности п о ч в ы и воздуха остается почти н е и з м е н н о й на всем их протяжении за и с к л ю ч е н и е м н а и б о л е е з а с у ш л и вых экотопов, где она несколько увеличивается (рис. 8.2, 8.4). О б р а щ а е т на себя внимание тот факт, что на большей части этих градиентов т р е н д ы с р е д н е в и д о в ы х значений почти п а р а л л е л ь н ы оси абсцисс , что свидетельствует о незначительном влия-
50 100 150 10 20 30 40 50 60 70 Запас воды в почве, мм Относительная влажность воздуха, %
Рис . 8.4. З а в и с и м о с т ь средневидовых значений д е ф и ц и т а водного н а с ы щ е н и я от в л а ж н о с т и почвы (а) и воздуха (б) .
Рис . 8.5. Поверхн ость , о п и с ы в а ю щ а я изменения с р е д н е в и д о в ы х значений д е ф и ц и т а водного н а с ы щ е н и я на градиентах факторов среды.

Глава 8. ДЕФИЦИТ ВОДНОГО НАСЫЩЕНИЯ 179
НИИ факторов среды в этих условиях произрастания на д е ф и ц и т водного н а с ы щ е н и я . Это отчетливо в и д н о на объемном графике ; р е з к и й и з н а ч и т е л ь н ы й по дъем д е ф и ц и та водного н а с ы щ е н и я наблюдается только в с а м ы х сухих м е с т о о б и т а н и я х (рис. 8.5). Н и ж е будет показано , что разброс средневидовых значений на г р а д и е н т а х влажно с ти почвы и воздуха может определяться не только в н е ш н и м и у с л о в и я м и или случайными причинами, но также анатомической структурой листьев растений и их принадл е ж н о с т ь ю к разным таксонам (двудольных и однодольных) .
Д и н а м и к а с р е д н е в и д о в ы х значений д е ф и ц и т а водного н а с ы щ е н и я однодольн ы х н двудольных р а с т е н и й на г р а д и е н т а х в л а ж н о с т и п о ч в ы и воздуха . О д н о д о л ь н ы е и двудольные р а с т е н и я заметно различаются п о сво ей р е а к ц и и н а изменение условий у в л а ж н е н и я экотопов. М н о ж е с т в е н н ы й р е г р е с с и о н н ы й а н а л и з показал , что з ависимость с р е д н е в и д о в ы х значений реального водного д е ф и ц и т а от влажно с ти почвы и в о з д у х а у о д н о д о л ь н ы х р а с т е н и й в ы р а ж е н а н е д о с т а т о ч н о о т ч е т л и в о
=0.210
20 40 60
= 0.902
50 100 Запас воды в почве, мм
80
150 О 20 40 60 80 Относительная влажность воздуха,'/.
Рис. 8.6. З а в и с и м о с т ь с р е д н е в и д о в ы х значений н а с ы щ а ю щ е г о с о д е р ж а н и я воды от влажности почвы (а , в) и воздуха (б, г) для видов о дно до ль ны х (а, б ) и двудольных
(в, г) растений.

180 Часть III. Закономерности структурно-функциональной организации растений..
(К^ = 0.400, !<'== 11.3, частные /"-критерии для влажности почвы и воздуха близки друг другу и р а в н ы соответственно 5.9 и 4.9). В то же время такая з а в и с и м о с т ь , найденная для двудольных растений , проявляется значительно сильнее {К^ = 0.702, Р= 39.0). При этом следует отметить для этой зависимости значительно большее влияние влажности воздуха (по с р а в н е н и ю с в л а ж н о с т ь ю почвы) на д е ф и ц и т водного н а с ы щ е н и я (соответствующие ч а с т н ы е / - к р и т е р и и р а в н ы 33.1 и 9.5).
Более отчетливо р а з л и ч и я м е ж д у однодольными и двудольными р а с т е н и я м и проявляются при ан али зе н а с ы щ а ю щ е г о содержания воды в з а в и с и м о с т и от в л а ж н о с т и почвы и воздуха (рис . 8.6). И з этих д а н н ы х видно , что с р е д н е в и д о в ы е в е л и ч и н ы нас ы щ а ю щ е г о содержания воды у видов однодольных практически не зависят от влажности почвы и воздуха (рис. 8.6, а, б). В то же время, н а с ы щ а ю щ е е со дер жание воды у двудольных растений достаточно тесно связано с этими факт о р ами с р е д ы (рис . 8.6, в, г).
Рис. 8.7. И з м е н е н и я к о э ф ф и ц и е н т о в вариации на градиентах в л а ж н о с т и п о ч в ы (а ) и воздуха (б) и т р е н д ы (в, г) м и н и м а л ь н ы х ( н и ж н и е линии) и м а к с и м а л ь н ы х (верхние
линии) для каждого участка коэффициентов вариации на этих градиентах .

Глава 8. ДЕФИЦИТ ВОДНОГО НАСЫЩЕНИЯ 181
Динамика показателей вариации дефицита водного насыщения
В а ж н ы м моментом исследования водообмена растений является анализ д и н а м и к и показателей вариации, поскольку считается, что размеры амплитуд признаков мо жно рассматривать как одно из в ы р а ж е н и й физиологической адаптации . И с с л е д у я изменения коэффициентов вариации д е ф и ц и т а водного н а с ы щ е н и я м о ж н о отметить л и ш ь слабо в ы р а ж е н н у ю т е н д е н ц и ю у в е л и ч е н и я этих к о э ф ф и ц и е н т о в вдоль г р а д и е н т о в влажности почвы и воздуха (рис. 8.7, а, б). На градиенте влажности воздуха (рис. 8.7, б) эта тенденция выражена несколько более отчетливо , чем на градиенте в л а ж н о с т и почвы (рис. 8.7, а), где она статистически недостоверна. Из вида этих графиков можно предположить, что ширина зоны разброса коэффициентов вариации также несколько увеличивается вдоль этих градиентов . Однако т р е н д ы м и н и м а л ь н ы х и максималь ных для каждого экотопа к о э ф ф и ц и е н т о в вариации п р а к т и ч е с к и п а р а л л е л ь н ы друг д р у г у на в с е м п р о т я ж е н и и г р а д и е н т о в в л а ж н о с т и п о ч в ы ( р и с . 8.7, в) и в о з д у х а (рис. 8.7, г).
Межвидовые структурно-функциональные корреляции
Анализ м е ж в и д о в ы х структурно-функциональных корреляций показал , что существуют некоторые тренды в изменениях насыщающего содержания воды и ряда структурных характеристик листьев . Отметим , что эти т р е н д ы не выявляются на всем массиве данных, но только при его разделении на группы однодольных и двудольных растений.
Н а с ы щ а ю щ е е с о д е р ж а н и е воды и п а р ц и а л ь н ы й объем э п и д е р м и с а . Н а л и ч и е достоверной корреляции между этими показателями указывает на то, что эпидермис может являться в некоторой степени препятствием для д и ф ф у з и и воды в м е з о ф и л л в процессе н а с ы щ е н и я листьев . Д л я видов однодольных растений в ы я в л е н а доста точно тесная корреляция м е ж д у п а р ц и а л ь н ы м объемом эпидермиса и н а с ы щ а ю щ и м содержанием воды (рис . 8.8, а). Э т а связь показывает, что у видов со сравнительно небольшим (меньше 7%) парциальным объемом эпидермиса {ВШотш итЬеПа(ш, Мз рзеиЛасогиз, 8раг§апшт егесШт) наблюдаются наиболее в ы с о к и е значения насыщающего содержания воды. М е н ь ш е всего насыщают ся водой листья АИзта р1ап1а§о-адиаНса, Са1ата§гозИз ерщеюз, Сагех асШа, С. сезрИоза, С. те1апоз1асНуа, 8а§ШаИа за^ИИ/оНа, 8Ира ЬогузЛетса, для которых о б н а р у ж е н ы д о с т а т о ч н о большие п а р ц и а л ь н ы е о б ъ е м ы э п и д е р м и с а . И з этого р я д а выделяется Сагех сокШса, в л и с т ь я которого , н е с м о т р я н а б о л ь ш о й п а р ц и а л ь н ы й о б ъ е м э п и д е р м и с а (13.8%), д и ф ф у н д и р о в а л о в п р о ц е с с е н а с ы щ е н и я с р а в н и т е л ь н о б о л ь ш о е к о л и ч е с т в о в о д ы
(0.998 г • г сух.массы ) •
у видов двудольных растений парциальный объем эпидермиса не связан с насыщаю щ и м содержанием воды (рис. 8.8, г). Отсутствие такой зависимости , возможно, свидетельствует о том, что эпидермис не является основным фактором, о г р аничиваю щ им диффузию воды, и она достаточно свободно проникает внутрь листа (отметим, что средние скорости насыщения водой листьев составляют 0.03-0.06 т^^^^ • г~у^^^,„^^^ • ч ~ ' , что в 30-40 раз меньше характерных величин интенсивности транспирации) . Сравним рис. 8.6 и 8.8, а, г. В и д н о , что для однодольных растений характерна ср авнит ель но тесная зависимость н а с ы щ а ю щ е г о содержания воды от парциального обьема эпидермиса при
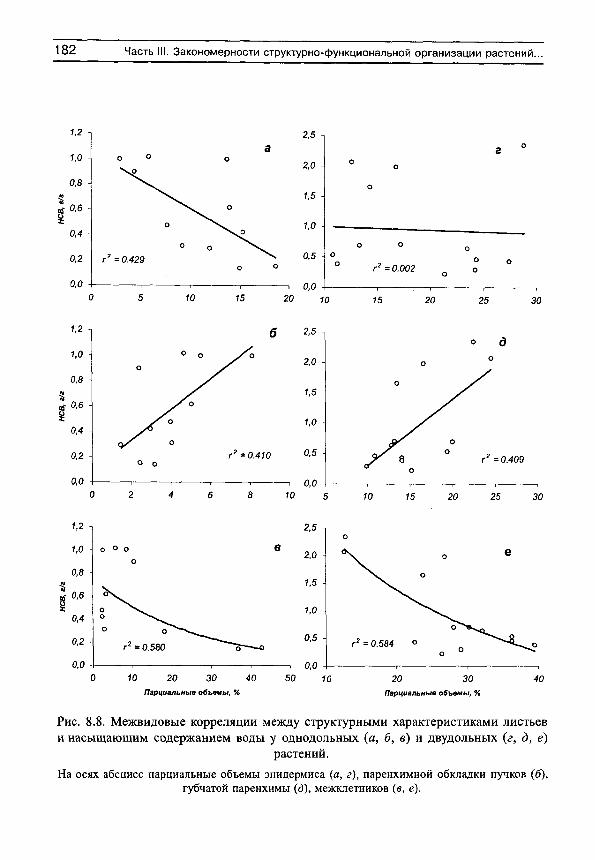
182 Часть III. Закономерности структурно-функциональной организации растений.
1,2 -|
1.0 -
0.8 -
0.6
0,4 -
0.2
0.0
г' = 0.429
10 15
2.5
2,0 -
1.5
1.0
0,5
0.0
20 10
г'=0.002 о о
15 20 25 30
10 20 30 40
Парциальные объемы, %
10 20 30
Парциальные объемы. %
Рис. 8.8. М е ж в и д о в ы е корреляции м е ж д у с т р у к т у р н ы м и х а р а к т е р и с т и к а м и листьев и н а с ы щ а ю щ и м с о д е р ж а н и е м в о д ы у о д н о д о л ь н ы х (а, б, в) и д в у д о л ь н ы х (г, д, е)
растений .
На осях абсцисс парциальные объемы эпидермиса {а, г), паренхимной обкладки пучков (б), губчатой паренхимы (д), межклетников (в, е).

Глава 8. ДЕФИЦИТ ВОДНОГО НАСЫЩЕНИЯ 183
почти полном отсутствии связи этого показателя водного р е ж и м а с факторами среды. Для двудольных наблюдается противоположная картина — тесная зависимость насыщающего содержания воды от факторов среды при отсутствии связи м е ж д у этим показателем и п арц и альным объемом эпидермиса .
Н а с ы щ а ю щ е е с о д е р ж а н и е в о д ы и п а р ц и а л ь н ы й о б ъ е м т к а н е й м е з о ф и л л а . Однодольные и двудольные растения и м е ю т корреляции н а с ы щ а ю щ е г о с о д е р ж а н и я воды с р а з н ы м и т канями мезофилла .
Д л я однодольных растений обнаружена связь м е ж д у п а р ц и а л ь н ы м о б ъ е м о м паренхимн ой обкладки пучков и этим показателем водного р е ж и м а (рис . 8.8, б). Ч е м больше (в о т н о с и т е л ь н о м в ы р а ж е н и и ) развита п а р е н х и м н а я обкладка , т е м б о л ь ш е н а с ы щ а ю щ е е содержание воды. М о ж н о предположить , что вода, п о с т у п а ю щ а я в листья в процессе н а с ы щ е н и я , концентрируется п р е и м у щ е с т в е н н о в клетках пар енх им-ных обкладок пучков . Это тем более вероятно, что ни с какими другими т к а н я м и мезофилла корреляции н а с ы щ а ю щ е г о содержания воды о б н а р у ж е н о не было . У видов с наибольшим р а з в и т и е м паренхимной обкладки {Ви1оти8 итЬеНшш, Сагех сокЫса и 1пз рзеи(1асоги$) наблюдается наибольшее н а с ы щ е н и е листьев водой . Такие виды, как АИзта р1ап(а§о-адиаИса, Са1ата§гозНз ерщеюз, 8а§Шапа за^ИИ/оНа и Сагех те1апоз1асНуа и м е ю т н а и м е н ь ш и е величины н а с ы щ а ю щ е г о с о д е р ж а н и я воды и парциальных объемов п а р е н х и м н о й обкладки межклетников .
В м е с т е с т е м нельзя не отметить , что р а з в и т и е п а р е н х и м н о й о бкладки пучков у однодольных растений связано с плотностью губчатой п а р е н х и м ы (рис . 8.9, а). У растений с наименее плотным сложением губчатой паренхимы (с б о л ь ш и м количеством межклетных пространств в этой ткани) наблюдается н а и м е н ь ш е е развитие паренхим-ных обкладок. Увеличение межклеточного пространства губчатой паренхимы, судя по всему, может л и м и т и р о в а т ь развитие п а р е н х и м н ы х о б к л а д о к (это характерно и для видов двудольных растений) . С другой стороны, плотность м е з о ф и л л а (при совокупном учете м е ж к л е т н ы х пространств и воздушных полостей) у однодольных растений очень тесно связана с в л а ж н о с т ь ю почвы (рис. 8.9, б) . П о э т о м у п о н я т н о , что насыщ а ю щ е е содерж ан и е воды отрицательно коррелирует с п а р ц и а л ь н ы м о бъемо м м е ж клетников (рис . 8.8, в): вдоль г р а д и е н т а в л а ж н о с т и п о ч в ы и в о з д у х а у м е н ь ш а е т с я плотность м е з о ф и л л а (увеличивается парциальный объем м е ж к л е т н и к о в ) и, соответственно, у м е н ь ш а е т с я р а з в и т и е околопучковой п а р е н х и м ы .
Для двудольных растений характерно наличие достаточно т е с н о й коррелятивной связи между п а р ц и а л ь н ы м объемом губчатой п а р е н х и м ы и н а с ы щ а ю щ и м содержанием воды (рис . 8.8, д). Поскольку не было о б н а р у ж е н о корреляций этого показателя водного р е ж и м а с д р у г и м и т к а н я м и мезофилла , то м о ж н о п р е д п о л о ж и т ь , что у двудольных растений вода в п р о ц е с с е н а с ы щ е н и я листьев попадает п р е и м у щ е с т в е н н о в губчатую паренхиму . П а р ц и а л ь н ы й объем этой ткани о т р и ц а т е л ь н о ко р р елир у ет с объемом меж клет н иков (рис . 8.9, в) . В то же время , п а р ц и а л ь н ы й о б ъ е м м е ж к л е т н и ков, особенно м е ж к л е т н и к о в губчатой п а р е н х и м ы , очень т е с н о связан с в л а ж н о с т ь ю почвы (рис. 8.9, г). Н а с ы щ а ю щ е е содержание воды отрицательно коррелирует с парциальным объемом межклетников (рис. 8.8, е) . Этот факт, судя п о всему, обусловлен тем, что вдоль градиента в л а ж н о с т и почвы и воздуха у в и д о в д в у д о л ь н ы х р а с т е н и й можн о н аблю дат ь у м е н ь ш е н и е п л о т н о с т и с л о ж е н и я м е з о ф и л л а и, с о о т в е т с т в е н н о , уменьшение п а р ц и а л ь н ы х объемов губчатой п а р е н х и м ы , и наоборот.
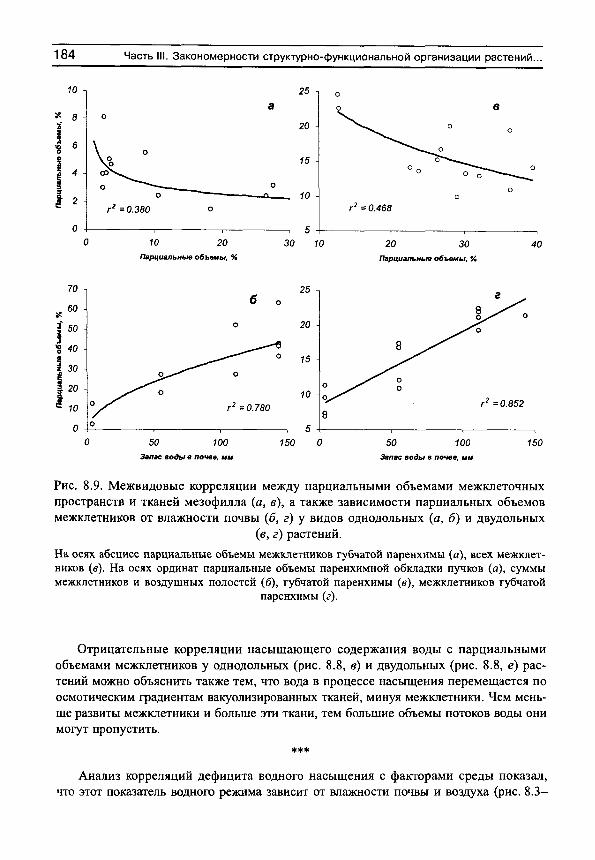
184 Часть III. Закономерности структурно-функциональной организации растений..
Парциальные объемы, % Парциальные объемы, •/.
О 50 100 150 О 50 100 150 Запас воды в почве, мм Запас воды в почве, мм
Рис. 8.9. М е ж в и д о в ы е к о р р е л я ц и и м е ж д у п а р ц и а л ь н ы м и о б ъ е м а м и м е ж к л е т о ч н ы х пространств и тканей м е з о ф и л л а (а, в), а т а к ж е з а в и с и м о с т и п а р ц и а л ь н ы х о б ъ е м о в межклетников от в л а ж н о с т и почвы (б, г) у видов о дно до ль ны х (а, б ) и дву до ль ных
(в, г) растений .
На осях абсцисс парциальные объемы межклетников губчатой паренхимы (а), всех межклетников (в). На осях ординат парциальные объемы паренхимной обкладки пучков (а), суммы межклетников и воздушных полостей (б), губчатой паренхимы (в), межклетников губчатой
паренхимы (г).
О т р и ц а т е л ь н ы е к о р р е л я ц и и н а с ы щ а ю щ е г о с о д е р ж а н и я в о д ы с п а р ц и а л ь н ы м и объемами м е ж к л е т н и к о в у однодольных (рис . 8.8, в) и двудольных (рис . 8.8, е) р ас тений мож н о объяснить также тем , что вода в процессе н а с ы щ е н и я п е р е м е щ а е т с я п о осмотическим градиентам вакуолизированных тканей, минуя межклетники . Чем меньш е развиты межклетники и больше эти ткани, т ем большие объемы потоков воды они могут пропустить .
***
А н а л и з корреляций д е ф и ц и т а водного н а с ы щ е н и я с ф а к т о р а м и с р е д ы показал , что этот показатель водного режима зависит от влажности почвы и воздуха (рис. 8.3-

Глава 8. ДЕФИЦИТ ВОДНОГО НАСЫЩЕНИЯ 185
8.5). Его связь с в л а ж н о с т ь ю почвы понятна: в условиях с п о н и ж е н н ы м и запасами доступной почвенной влаги дисбаланс между поступлением и расходом этого ресурса в растениях увеличивается, в результате чего растет и степень недонасыщенности листьев водой. Наоборот, при достаточном количестве воды в почве ее поступление в растения н езн ач и тельно отстает от расхода , а д е ф и ц и т водного н а с ы щ е н и я снижается .
Связь этого показателя с в л а ж н о с т ь ю воздуха (которая пр о я вля ет ся несколько сильнее , ч е м связь с в л а ж н о с т ь ю почвы) позволяет признать , что л и с т ь я р а с т е н и й могут восполнять недостаток воды за счет ее поглощения не только из по чвы , но и из окружающей их а т м о с ф е р ы ' . Действительно , уменьщение д е ф и ц и т а водного насыщения вдоль градиента в л а ж н о с т и воздуха м о ж н о объяснить , в частности , и поглощением листьями атмосферной влаги. Особенно хорощо это проявляется при раздельном анализе поведения однодольных и двудольных растений на градиентах факторов среды.
В ы ш е м ы отмечали, что зависимость средневидовых значений реального водного д е ф и ц и т а от в л а ж н о с т и почвы и воздуха у однодольных р а с т е н и й в ы р а ж е н а не достаточно отчетливо {К^ = 0.400, Г= 11.3, частные / - к р и т е р и и для в л а ж н о с т и почвы и воздуха близки друг другу и равны соответственно 5.9 и 4.9). В то же время такая з а в и с и м о с т ь , н а й д е н н а я для двудольных р а с т е н и й , п р о я в л я е т с я з н а ч и т е л ь н о сильн ее (К^ = 0.702, / = 39.0). П р и этом с л е д у е т о т м е т и т ь д л я этой з а в и с и м о с т и значительно б о л ь ш е е влияние в л а ж н о с т и воздуха (по с р а в н е н и ю с в л а ж н о с т ь ю по чвы) на д е ф и ц и т водного н а с ы щ е н и я ( соответствующие ч а с т н ы е / - к р и т е р и и р а в н ы 33.1 и 9.5).
Разбивка всей выборки на две группы по определенной таксономической принадл е ж н о с т и р а с т е н и й показывает их р а з н у ю р е а к ц и ю на и з м е н е н и я ф а к т о р о в среды. Д е ф и ц и т водного н а с ы щ е н и я и н а с ы щ а ю щ е е содержание в о д ы (рис . 8.6) однодольных растений в м е н ь ш е й степени связаны с в л а ж н о с т ь ю п о ч в ы и воздуха , чем двудольных. К р о м е того, п о с л е д н и е не в одинаковой степени и с п о л ь з у ю т п о ч в е н н у ю и а т м о с ф е р н у ю влагу для в о с п о л н е н и я недостатка воды в л и с т ь я х (это видно из приведенных в ы ш е ч а с т н ы х / - к р и т е р и е в ) . Судя по всему, у м е н ь ш е н и е водного д е ф и ц и та вдоль градиента влажности воздуха у двудольных растений происходит за счет преимущественного использования атмосферной влаги. Это может быть обусловлено тем, что размеры клеток э п и д е р м и с а у видов однодольных о г р аничиваю т д и ф ф у з и ю воды в листья (рис . 8.8, а). В то ж е время , проникновение воды в листья дву до ль ных р ас тений не связано с п а р ц и а л ь н ы м объемом эпидермиса (рис. 8.8, г).
Коли ч ест вен н ые р а з л и ч и я по средним в е л и ч и н а м д е ф и ц и т а водного н а с ы щ е н и я между однодольными и двудольными растениями (рис. 8.1) могут б ы т ь о б у с л о в л е н ы тем, что у однодольных растений вода в процессе н а с ы щ е н и я п о с т у п а е т в пар енх им-н у ю обкладку пучков (рис . 8.8, б), а у двудольных р а с т е н и й — в губчатую п а р е н х и -
' В ночное и предрассветное время возможны условия, когда градиент давления пара направлен в сторону листа и может происходить поглощение воды в парообразном состоянии листьями растений (Слейчер, 1970). Подробный анализ этого вопроса и библиографические ссылки можно найти также в работах Л. Н. Бабушкина (1976) и А. А. Зялалова (1984).

186 Часть III. Закономерности структурно-функциональной организации растений..
му ' (рис. 8.8, д). М о ж н о показать , что п а р ц и а л ь н ы й о б ъ е м губчатой п а р е н х и м ы в среднем п р и м е р н о в 5 раз больше парциального объема п а р е н х и м н о й обкладки пучков (21.7:4.3 соответственно) . При этом развитие п а р е н х и м н о й о б к л а д к и пучков (у однодольных) и губчатой п а р е н х и м ы (у д в у д о л ь н ы х ) с о п р о в о ж д а е т с я и з м е н е н и е м плотности м е з о ф и л л а (рис . 8.9): в листьях с р ы х л ы м м е з о ф и л л о м эти ткани развиваются слабее, и наоборот, наибольшее развитие они получают в л и с т ь я х с более плотной тканевой структурой . Но степень с к л е р о м о р ф н о с т и листьев очень тесно связана с запасами влаги в почве: вдоль градиента влажности почвы плотность с л о ж е н и я мезофилла у м е н ь ш а е т с я (рис . 8.9, б, г).
О т с ю д а в и д н о н е т о л ь к о прямое , но и косвенное в л и я н и е у с л о в и й по чвенно г о водоснабжения на формирование дефицита водного н а с ы щ е н и я , опосредованное анатомической структурой листьев . Развитие водного д е ф и ц и т а н а г р а д и е н т е в л а ж н о с ти почвы, по-видимому, является зеркальным отражением процесса н а с ы щ е н и я . Если пулами н а с ы щ е н и я однодольных растений являются клетки п а р е н х и м н о й обкладки пучков , а двудольн ых — губчатая паренхима , то и потери воды при ф о р м и р о в а н и и водного д е ф и ц и т а д о л ж н ы происходить п р е и м у щ е с т в е н н о из э т и х тканей . В р а з н ы х частях г р а д и е н т а в л а ж н о с т и п о ч в ы м о ж н о наблюдат ь р а з н о е с о о т н о ш е н и е т к а н е й мезофилла и соответственно разное развитие тканей , в ы с т у п а ю щ и х в р о л и о с м о т и чески активных пулов н а с ы щ е н и я водой. Это создает определенные п р е д п о с ы л к и для формирования водного д е ф и ц и т а . Если эти предпосылки находятся в соответствии с условиями увлажнения экотопов, то они будут реализованы. То есть , п е р в и ч н ы м фактором для ф о р м и р о в а н и я водного д е ф и ц и т а является , судя по всему, анатомическое строение мезофилла , которое опосредует воздействие условий у в л а ж н е н и я экотопов на этот п р о ц е с с водного р е ж и м а растений . В то же время , в к р а й н е а р и д н ы х м е с т о обитаниях развитие водного д е ф и ц и т а может идти как л а в и н о о б р а з н ы й п р о ц е с с во всех тканях листьев и г л а в н ы м фактором этого процесса д о л ж н ы б ы т ь у с л о в и я недостаточного почвенного водоснабжения .
Резюме. Статистический анализ дефицита водного насыщ ения на градиенте влажности почвы показал с л е д у ю щ е е . В е с ь массив исходных д а н н ы х д е ф и ц и т а водного насыщения имеет среднее значение х = 17.8%. Типичные значения д е ф и ц и т а водного насыщения травянистых растений на градиенте влажности почвы не п р е в ы ш а ю т 30%. Наиболее заметные различия по средним значениям дефицита водного насы щ ения выявлены между в и д а м и однодольных и двудольных растений . С о о т в е т с т в у ю щ и е средние значения этого признака р а в н ы 15.0 и 21.4%. С р е д н и е у р о в н и д е ф и ц и т а водного насыщения т е с н о связаны с влажностью почвы и воздуха. Н аибо лее отчетливо эти связи проявляются в сухой ч а с т и градиента в л а ж н о с т и п о ч в ы с з а п а с а м и влаги м е нее 50 м м и в у с л о в и я х с п о н и ж е н н о й (менее 40-50%) о т н о с и т е л ь н о й в л а ж н о с т ь ю
' Это объясняется различиями формы роста — интеркалярного у однодольных и апикального у двудольных и связанной с этим неадекватностью их фотосинтетических органов (осевых у однодольных и латеральных у двудольных) (Гамалей, 20026). Ткани, выступающие в роли пулов насыщения и формирования флоэмного потока, детерминирующего рост, не могут быть одинаковыми у однодольных и двудольных растений по тем же причинам: у однодольных это обкладка пучков, по сути и по форме осевая паренхима, у двудольных — губка, типично листовая паренхима (Гамалей, устное сообщение).

Глава 8. ДЕФИЦИТ ВОДНОГО НАСЫЩЕНИЯ 187
воздуха. Однодольные и двудольные растения заметно различаются по своей реакции на изменение у с л о в и й увлажнения экотопов, что, вероятно связано с р а з н о й ф о р м о й роста (интеркалярной у однодольных и апикальной у двудольных) . С р е д н е в и д о в ы е величины н а с ы щ а ю щ е г о содержания воды у видов о дно до ль ных п р а к т и ч е с к и не зависят от влажности п о ч в ы и воздуха. В то же время , н а с ы щ а ю щ е е со дер жание воды у двудольных р а с т е н и й достаточно тесно связано с э т и м и ф а к т о р а м и среды. Д л я видов однодольных растений выявлена достаточно тесная корреляция между парциальным объемом э п и д е р м и с а и н а с ы щ а ю щ и м содержанием воды. Э т а связь показывает , что у видов с н е б о л ь ш и м п а р ц и а л ь н ы м объемом э п и д е р м и с а н а б л ю д а ю т с я наиболее высокие значения н а с ы щ а ю щ е г о с о д е р ж а н и я воды, и наоборот . У в и д о в двудольных растений п а р ц и а л ь н ы й объем э п и д е р м и с а не связан с н а с ы щ а ю щ и м с о д е р ж а н и е м воды. Это м о ж е т свидетельствовать о том, что э п и д е р м и с д л я этой г р у п п ы р а с т е н и й не бьш о с н о в н ы м фактором, о г р а н и ч и в а ю щ и м д и ф ф у з и ю в о д ы в листья . Д л я однод о л ь н ы х р а с т е н и й о б н а р у ж е н а п о л о ж и т е л ь н а я связь м е ж д у п а р ц и а л ь н ы м о бъемо м паренхи мн ой обкладки пучков и н а с ы щ а ю щ и м с о д е р ж а н и е м воды. Д л я двудольных растений характерно наличие достаточно тесной положительной связи м е ж д у парциальным объемом губчатой п а р е н х и м ы и этим показателем водного р е ж и м а . С д е л а н ы предположения: а) листья растений могут восполнять недостаток воды за счет ее поглощения не только из почвы, но и из воздуха, что в н а и б о л ь ш е й ст епени характерно для двудольных растений; б) количественные различия п о средним в е л и ч и н а м де фицита водного н а с ы щ е н и я м е ж д у однодольными и д в у д о л ь н ы м и р а с т е н и я м и могут быть обусловлены т е м , что вода в процессе н а с ы щ е н и я по ст у пает в пулы, которые значительно р а з л и ч а ю т с я по своим возможностям запасания : у о д н о д о л ь н ы х расте ний — в п а р е н х и м н у ю о б к л а д к у пучков , а у двудольных — в г у бчат у ю пар енх иму ; в) изменения в развитии паренхимной обкладки пучков и губчатой п а р е н х и м ы сопровождаются и з м е н е н и я м и плотности мезофилла : в листьях с р ы х л ы м м е з о ф и л л о м эти ткани развиваются слабее , и наоборот, наибольшее развитие они п о л у ч а ю т в листь ях с более плотной тканевой структурой; г) поскольку степень с к л е р о м о р ф н о с т и листьев очень т е с н о связана с в л а ж н о с т ь ю почвы, она оказывает не только п р я м о е , но и косвенное влияние на ф о р м и р о в а н и е д е ф и ц и т а водного н а с ы щ е н и я , о по ср едо ван ное анатомической структурой листьев .

188 Часть III. Закономерности структурно-функциональндй организации растений..
ГЛАВА 9. СОДЕРЖАНИЕ ВОДЫ
Содержание воды в листьях является часто используемым показателем для характерист и ки с о с т о я н и я водного р е ж и м а р а с т е н и й в р а з н ы х у с л о в и я х п р о и з р а с т а н и я (Горшкова, 1970, 1971; Алексеенко , 1976; Свешникова , 1979; Бо бр о вская 1985, 1991 и др.) . С о д е р ж а н и е в о д ы определяется обычно т р е м я с п о с о б а м и как о т н о ш е н и е количества воды в навеске к: 1) сырой массе ткани, 2) сухой массе ткани , 3) массе воды в полностью н а с ы щ е н н о й водой ткани (Слейчер, 1970; Ко1с1е е! а!., 1989). Если в отечественной литературе для оценки оводненности листьев п р и м е н я ю т пр еиму щ ест вен но первый и, несколько реже , второй способы, то в з арубежных публикациях , за редкими и с к л ю ч е н и я м и (см. , например , О г а Ь а т , МоЬе1, 1999; Неггега е1 а1., 2000; КЬап е1 а1., 2000; М1§аЬ1с1, ЕШаак, 2001), используют третий (т. е. определяют «относитель ное содерж ан и е воды») . Содержание воды в листьях является о д н и м из в а ж н ы х параметров водообмена при проведении сравнительных исследований , поскольку д о с таточно хорошо отражает условия почвенного водоснабжения и о б щ е е состояние растений в этих условиях . В м е с т е с тем, существует мнение об ограниченности использования этого показателя в качестве самостоятельного признака , поскольку в совершенно н е с х о д н ы х экологических условиях можно встретить р а с т е н и я , о в о д н е н н ы е практически одинаково (Бобровская , 1991). М ы п о п ы т а е м с я разобраться в этом вопросе , исследовав о с о б е н н о с т и изменений оводненности л и с т ь е в у б о л ь ш о й г р у п п ы растений на ш и р о к о м градиенте влажности почвы.
Статистический анализ содержания воды в листьях у видов однодольных и двудольных растений
Д а н н ы е с о б р а н ы в 16 с о о б щ е с т в а х , р а с п о л о ж е н н ы х в р а з н ы х частях г р а д и е н т а влажности почвы (см. табл . 7.1). Статистические характеристики с р е д н е в и д о в ы х величин содержания воды показаны в таблице 9.1.
Весь массив исходных д а н н ы х содержания воды в листьях растений имеет среднее значение х =2.355 г^^^,, •г'у^,;^^^^^ и стандартное отклонение в^ = 1.172 г^^,,,, •т'^1^,„а^сы (п = 13837). Ряд р а с п р е д е л е н и я имеет п о л о ж и т е л ь н у ю а с и м м е т р и ю с модой, р а в н о й 2.0 г^ойу •^'^ух.'массы , а 90% его з н а ч е н и й ( в а р и а н т ) н а х о д я т с я в д и а п а з о н е 0 .932 -4.878 г^ойи •'1^сух.массы ( квантили порядков 0.05 и 095), 5% в ариант в ы ш е к в а н т и л я 0.95 распределены в д и а п а з о н е 4.878-9.167 г ,̂̂ ^ ,̂ '^^^.массы • М о ж н о сказать , что т и п и ч н ы е значения с о д е р ж а н и я воды в листьях травянистых р а с т е н и й на г р а д и е н т е в л а ж н о с т и почвы не п р е в ы ш а ю т 5 г^^^^, •т^'^,массы •
Однако это касается всего массива исходных данных. Если провести более детальное исследование , ра збив выборку на группы однодольных и двудольных растений и выделив из этих г р у п п в и д ы со с п о с о б н о с т ь ю накапливать воду в с п е ц и а л и з и р о в а н ных тканях ( суккуленты ' ) , то картина статистического о п и с а н и я заметно изменится .
' К числу однодольных суккулентов относятся следующие виды: АШит ро1угЫ2ит, Нете-госаШз ттог, Мз сИскоЮта, Мз рзеис1асоги5, Мз зШпса, а также ВиЮтиз итЬеИаШз (с водоносной обкладкой пучков) и 5раг§апшт егесШт (имеющий специализированную водоносную ткань). В группе двудольных растений имеется только один суккулент (с коронарной обкладкой пучков) — АпаЪаз1з Ьгеу1/оНа.

Глава 9. СОДЕРЖАНИЕ ВОДЫ 189
Таблица 9.1
Средние значения и показатели вариации содержания воды (СБ), выраженного
в ^воды ' ^сух.массы (СТОлбСЦ XI ), а ТВКЖС В ММОЛЬ ' 2^ ^,^^^1^ (столбец хг) и процентах от сырой массы листьев (столбец хг)
№ участка по
табл. 7.1
№ объекта
СВ № участка по
табл. 7.1
№ объекта
Вид п XI ^ х , У,Уо Х2 хъ
1 1 АПетша аизШаса 94 0.798 0.240 30.1 44.3 АА.А 2 Сагех сокЫса 94 0.903 0.219 24.3 50.1 47.4 3 СаИит Шкепкит 94 0.989 0.274 27.7 54.9 49.7 4 8Нра ЪогузХкепка 94 0.631 0.111 17.5 35.0 38.7 5 Тп/оИит ап/епзе 52 0.900 0.189 21.0 50.0 47.7 6 Уегопка зркаШ 94 0.692 0.133 19.2 38.4 40.9
Суммы и средние для сообщества 522 0.812 0.240 29.5 45.1 44.8 2 7 АШит ро1угк1гит 177 4.510 0.935 20.7 250.2 81.9
8 АпаЪазгз ЬгеуфИа 179 2.096 0.387 18.5 116.3 67.7 9 8Ира §1агеоза 176 1.043 0.185 17.7 57.9 51.1
Суммы и средние для сообщества 532 2.551 1.566 61.4 141.5 71.8 3 10 АПетша тагзскаИшпа 52 2.367 0.336 14.2 131.4 70.3
И Са1ата^гоз113 ерщеюз 50 1.627 0.201 12.4 90.3 61.9 12 РокпНИа агепапа 38 1.345 0.410 30.5 74.6 57.4 13 8йра Ьогуз1кепка 38 0.958 0.165 17.2 53.2 48.9 14 Уегопка зркаШ 40 1.772 0.245 13.8 98.3 63.9
Суммы и средние для сообщества 218 1.664 0.553 33.2 92.4 62.5 4 15 Аггет1з1а /гщ1^а 58 1.474 0.206 14.0 81.8 59.6
16 Аз(ег а1ртиз 58 2.128 0.372 17.5 118.1 68.0 17 Оху1гор1з оИ^аШка 62 1.922 0.122 6.3 106.7 65.8 18 Заиззигеа кисоркуИа 62 1.689 0.183 10.8 93.7 62.8
Суммы и средние для сообщества 240 1.803 0.338 18.8 100.1 64.3 5 19 Сагех зкпосагра 52 1.398 0.088 6.3 77.6 58.3
20 РезШса кпепзгз 52 0.997 0.098 9.8 55.3 49.9 21 КоЬгезьа туозигоШез 52 1.284 0.068 5.3 71.2 56.2 22 Оху1гор13 оН§ап1ка 52 2.083 0.169 8.1 115.6 67.6 23 8аиззигеа кисоркуПа 52 2.104 0.189 9.0 116.8 67.8
Суммы и средние для сообщества 260 1.573 0.464 29.5 87.3 61.1 6 24 АШит ро1угк{2ит 180 4.505 0.463 10.3 250.0 81.8
25 АпаЬазгз ЬгеуфИа 180 2.795 0.536 19.2 155.1 73.6 26 8Нра §1агеоза 172 1.155 0.213 18.4 64.1 53.6
Суммы и средние для сообщества 532 2.843 1.430 50.3 157.8 74.0 7 27 Сагех (1игЫзси1а 230 1.475 0.137 9.3 81.8 59.6
28 СШз1о^епез здиаггоза 228 1.506 0.162 10.8 83.6 60.1 29 Ьеутиз сктепзгз 230 1.673 0.106 6.3 92.8 62.6 30 РокпНИа Ы/игса 228 1.574 0.181 11.5 87.3 61.1
Суммы и средние для сообщества 916 1.557 0.167 10.7 86.4 60.9

190 Часть III. Закономерности структурно-функциональной организации растений.
Продолжение таблицы 9.1
№ участка по
табл. 7.1
№ объекта
СВ № участка по
табл. 7.1
№ объекта
Вид п х\ \ У,% Х2 хз
Суммы и средние для сообщества 916 1.557 0.167 10.7 86.4 60.9 8 31 Анетта /п§Ша 440 2.083 0.483 23.2 115.6 67.6
32 /га <аско1ота 437 4.724 0.645 13.7 262.1 82.5 33 Ьеутиз сЫпепз1з 442 1.458 0.180 12.3 80.9 59.3 34 Ро1еп1И1а (апасеН/оИа 416 1.793 0.237 13.2 99.5 64.2 35 РиЬаИИа (игааптоун 440 2.044 0.350 17.1 113.4 67.1 36 8йра кгу1оуИ 442 0.949 0.108 11.4 52.7 48.7
Суммы и средние для сообщества 2617 2.174 /.2Й5 58.2 120.6 68.5 9 37 Са1ата§гозН5 ерг^еюз 120 1.504 0.110 7.3 83.4 60.1
38 Сагех те1апоз1асИуа 120 1.507 0.109 7.2 83.6 60.1 39 СаИит гигНетсит 120 2.331 0.310 13.3 129.3 70.0 40 Ьу1Ьгит У1г§а1ит 120 1.624 0.171 10.5 90.1 61.9 41 Зап^шзогЬа о/рсшаНз 120 1.797 0.242 13.5 99.7 64.3 42 ТапасеШт уи1§аге 120 2.289 0.252 11.0 127.0 69.6
Суммы и средние для сообщества 720 1.842 0.405 22.0 102.2 64.8 10 43 Сагех те!апоШскуа 34 1.503 0.153 10.2 83.4 60.1
44 РШрепс1и1а и1тапа 34 1.919 0.206 10.7 106.5 65.7 45 СаНит гиЫоШез 34 2.394 0.390 16.3 132.8 70.5 46 8ап§шзогЬа о^рС1паИз 34 2.326 0.194 8.3 129.1 69.9
Суммы и средние для сообщества 136 2.036 0.437 21.5 113.0 67.1 11 47 Сагех уи1рта 50 Х.11Л 0.251 14.1 98.5 64.0
48 аЫит тсапит 50 3.968 0.440 11.1 220.2 79.9 49 СаНит гиЫоШез 50 2.250 0.273 12.1 124.8 69.2 50 Уеготса Ьп^г/оНа 50 2.223 0.164 7.4 123.4 69.0 51 УШа сгасса 50 2.324 0.385 16.6 128.9 69.9
Суммы и средние для сообщества 250 2.508 0.820 32.7 139.2 77.5 12 52 АЬресигиз атпсНпасеиз 280 2.394 0.393 16.4 132.8 70.5
53 Сагех а1кегос1ез 284 2.037 0.188 9.2 113.0 67.1 54 Сагех ШкоркИа 284 1.931 0.186 9.6 107.2 65.9 55 Е1у1п§1а герепз 242 1.872 0.420 22.4 103.9 65.2 56 1пз зШпса 284 2.767 0.362 13.1 153.5 73.5 57 РкаШгоШез агипсИпасеа 286 2.500 0.344 13.8 138.7 71.4 58 Ро1у§опит зШпсит 284 3.379 0.976 28.9 187.5 77.2
Суммы и средние для сообщества 1944 2.423 0.690 28.5 134.5 70.8 13 59 А^гозйз топ^коИса 239 2.734 0.302 11.0 151.7 73.2
60 Апетопе сИско1ота 240 3.212 0.460 14.3 178.2 76.3 61 Сагех а1Иегос1ез 240 2.028 0.175 8.6 112.5 67.0 62 Роа ап^изИ/оНа 240 1.665 0.095 5.7 92.4 62.5 63 ТкаИстгт зтр1ех 239 2.399 0.261 10.9 133.1 70.6 64 УШа сгасса 240 2.799 0.464 16.6 155.3 73.7
Суммы и средние для сообщества 1438 2.473 0.606 24.5 137.2 71.2

Глава 9. СОДЕРЖАНИЕ ВОДЫ 191
Окончание таблицы 9.1
№ участка по
табл. 7.1
№ объекта
Вид СВ № уча
стка по табл. 7.1
№ объекта
Вид п XI У,Уо Х2 хз
14 65 Сагех сезрИоза 224 1.979 0.230 И .6 109.8 66.4 66 РШрепМа иЫапа 224 1.642 0.236 14.4 91.1 62.2 67 1пз рзеи(1асогиз 226 2.907 0.446 15.4 161.3 74.4 68 5ап§и1зогЬа о//1стаНз 228 2.092 0.277 13.3 116.1 67.7 69 Уа1епапа \уо1^еп813 228 3.721 0.495 13.3 206.5 78.8 70 Уеготса 1оп§1/оНа 226 2.390 0.293 12.2 132.6 70.5
Суммы и средние для сообщества 1356 2.458 0.770 31.3 136.4 71.1 15 71 Аг1ет1з1а 1асШа1а 238 3.203 0.565 17.6 177.7 76.2
72 Вготорш тегт1з 168 2.044 0.246 12.0 113.4 67.1 73 НетегосаШз ттог 232 3.316 0.406 12.2 184.0 76.8 74 Ьеутиз сМпепзьз 238 1.805 0.180 10.0 100.2 64.3 75 Ро1ешИ1а апзеппа 238 2.044 0.219 10.7 113.4 67.1 76 Зап^шзогЪа о(/1стаНз 238 2.369 0.383 16.2 131.5 70.3
Суммы и средние для сообщества 1352 2.481 0.695 28.0 757.7 71.3 16 77 АИзта р1ап1а§о-адиаИса 138 4.981 0.563 11.3 276.4 83.3
78 ВШотиз итЬеИаШз 118 6.609 0.816 12.3 366.7 86.9 79 Сагех асиШ 138 2.142 0.235 11.0 118.8 68.2 80 Ьусориз еигораеиз 138 3.895 0.519 13.3 216.2 79.6 81 8а§Шапа за^кИ/оИа 138 5.968 0.596 10.0 331.2 85.6 82 8раг§атит егесЫт 138 4.889 0.466 9.5 271.3 83.0
Суммы и средние для сообщества 808 4.701 1.536 32.7 260.9 82.5
Рассмотрим несуккулентные в и д ы (склерофиты) однодольных и двудольных растений. Содержание воды в листьях однодольных склерофитов заметно м е н ь ш е , чем в листьях двудольных (табл. 9.2). Статистическая оценка р а з н о с т и средних показывает, что она в высокой степени достоверна ( Р < 0.001; соответствующие объемы выборок равны 5811 и 5875, д и с п е р с и и — 0.950 и 0.683, ^-критерий С т ь ю д е н т а — 23.2). Д о с товерные статистические различия по оводненности листьев м е ж д у этими г р у п п а м и растений наблюдаются на всем протяжении градиента в л а ж н о с т и п о ч в ы (рис. 9 .1 , а). Эти различия наиболее велики в аридной и средней частях градиента , в гумидной его части они становятся заметно меньше. Верхние пределы типичных изменений признака также ниже у однодольных склерофитов по сравнению с двудольными. У однодольных склерофитов т и п и ч н ы е значения (между квантилями порядков 0.05 и 0.95) содержания воды в листьях на всем градиенте влажности почвы заключены в границах 0 .9-3.3 г,„^„ -^сух-массы , У двудольных — В прсделах 1.0-3.8
^воды '^сух.массы •
Среднее содержание воды в листьях однодольных суккулентов значительно в ы ш е такового в листьях склерофитов (табл. 9.2). Высокая оводненность листьев однодольных суккулентов наблюдается в л ю б ы х частях градиента в л а ж н о с т и почвы, п р и ч е м в аридной части градиента она даже выше , чем в средней и гумидной его частях (рис. 9.1, б). Количество воды в а ф и л л ь н ы х побегах АпаЬазгз Ьгвуг/оИа (двудольного сук-
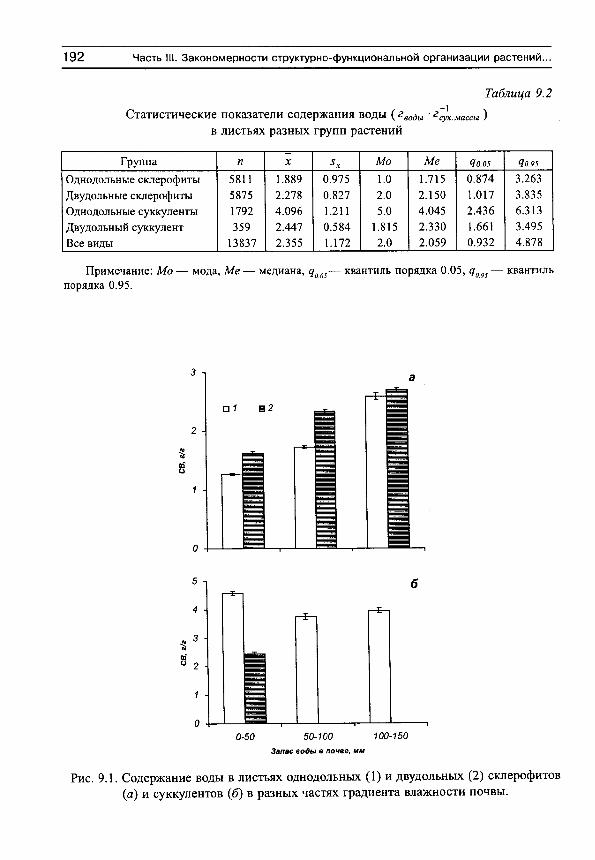
192 Часть III. Закономерности структурно-функциональной организации растений..
Таблица 9.2
Статистические показатели содержания воды ( г ^ ^ ^ • г^^^^^^ ) в листьях разных групп растений
Группа 71 X Мо Ме Я 0.05 Я 0.95
Однодольные склерофиты 5811 1.889 0.975 1.0 1.715 0.874 3.263 Двудольные склерофиты 5875 2.278 0.827 2.0 2.150 1.017 3.835 Однодольные суккуленты 1792 4.096 1.211 5.0 4.045 2.436 6.313 Двудольный суккулент 359 2.447 0.584 1.815 2.330 1.661 3.495 Все виды 13837 2.355 1.172 2.0 2.059 0.932 4.878
Примечание: Мо — мода, Ме — медиана, ^г^^^— квантиль порядка 0.05, — квантиль порядка 0.95.
0-50 50-100 100-150 Запас воды в почве, мм
Рис. 9 .1. Содержание воды в листьях однодольных (1) и двудольных (2) склерофитов (а) и суккулентов (б) в ра зных частях градиента в л а ж н о с т и почвы.

Глава 9. СОДЕРЖАНИЕ ВОДЫ 193
кулента с коронарным синдромом) несколько больше, ч е м у с к л е р о ф и т о в , однако заметно меньше, чем у однодольных суккулентов (табл. 9.2, рис . 9.1, б). В целом мо жно констатировать очень высокое содержание воды в а с с и м и л и р у ю щ и х о р г а н а х суккулентов в л ю б ы х частях градиента влажности п о ч в ы ' . Это м о ж е т б ы т ь связано как с анатомическими, так и функциональными особенностями этих растений . У суккулентов (особенно у видов с коронарным синдромом, с п о с о б н ы х к С ^ - С А М - ф о т о с и н т е -зу, подобно АпаЪа518 Ьгеу1/оНа) резервирование воды во в н у т р е н н и х т к а н я х способствует более прочному ее у д е р ж а н и ю и л у ч ш е м у контролю над ее расходом (Гамалей, 1988).
Связь количества воды и сухой массы в листьях с их площадью
Исследование з а в и с и м о с т и п л о щ а д и листьев от количества воды и сухой массы в них представляет н е с о м н е н н ы й интерес , н е с м о т р я н а то , что этот в о п р о с , на первый взгляд, прямо не связан с водообменом растений . Однако при д е т а л ь н о м его исследовании м о ж н о в ы я в и т ь т р е н д ы изменений пл о т но ст и л и с т ь е в и удельного количества воды в них на градиенте влажности почвы. П о м и м о этого д а н н ы й вопрос имеет и важное самостоятельное значения д л я аналитического о п р е д е л е н и я п л о щ а д и листьев по их в е с о в ы м параметрам .
С этой ц е л ь ю м ы исследовали выборку, с о с т о я щ у ю из в и д о в , п р о и з р а с т а ю щ и х в сообществах под номерами (в табл. 7.1 и 9.1) 1, 9, 14 и 16 (кроме ВиЮтш итЬеНаШз, вида, у которого очень обьемные , треугольные в поперечном сечении листья) . В ы б о р ка состояла из 156 точек (5 -10 -кратная повторность д л я каждого вида) . Б ы л и определены площади листьев , их сырая и абсолютно сухая массы, количество воды в них.
Корреляционный анализ выявил достаточно т е с н у ю зависимость п л о щ а д и листьев от количества сухой массы (рис . 9.2, а) . Более т е с н а я связь с у щ е с т в у е т м е ж д у пло щ а д ь ю листьев и количеством в них воды (рис. 9.2, б). М н о ж е с т в е н н ы й р е г р е с с и о н ный анализ подтвердил н а л и ч и е этих корреляций {К^ =0.856, 7^= 455.3 при степенях свободы 2 и 153) и показал , что наибольшее влияние на п л о щ а д ь л и с т ь е в оказывает количество с о д е р ж а щ е й с я в них воды (частный / " -критерий равен 200.1), тогда как сухая масса листьев обнаруживает значительно м е н ь ш е е воздействие на этот геометрический параметр ( ч а с т н ы й /" -критерий равен 17.8) (отметим, что р а з б и в к а в ы б о р ки на однодольные и двудольные существенно не изменяет результатов р е г р е с с и о н ного анализа) . П о результатам этого анализа б ы л а п о с т р о е н а р е г р е с с и о н н а я модель (которую м ы использовали для расчета площадей листьев однодольных и двудольных растений и связанных показателей, таких как удельная п л о щ а д ь и удельная масса л и стьев):
у = а ехр(^>[с 2 , + ^ 2 ]) + ^ >
' Это создает определенные трудности при анализе связей содержания воды с факторами среды, а также с другими показателями водного режима и анатомической структуры листьев растений. Поэтому при анализе таких связей использовались данные только по однодольным и двудольным склерофитам (в подписях к рисункам будет отмечено, когда анализ относится именно к этим группам растений).
7 - 2353
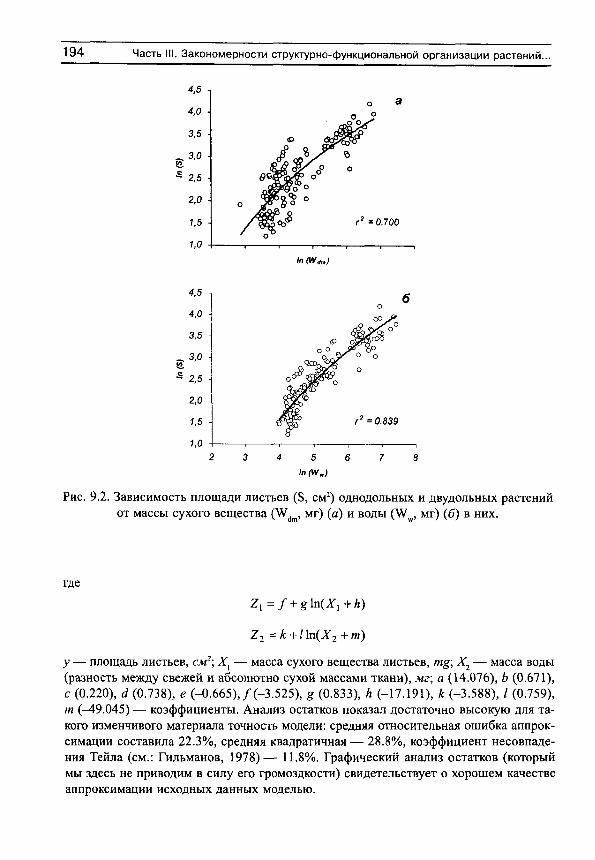
194 Часть III. Закономерности структурно-функциональной организации растений.
4,5 -,
4,0 •
3,5 • ©
_ 3,0 •
^ 2,5 -
2,0 - о 5" 5 °
1,5 -
1,0 -о
о° °
= 0.700
4,5
4,0
3,5
3,0 -
^ 2,5 -
2,0 -
Л5
1,0
г' = 0839
5
Рис. 9.2. З ави си мость п л о щ а д и листьев (8, см^) однодольных и дву до ль ных растений от массы сухого вещества (\\'^^, мг) (а) и воды мг) (б) в них .
где
2 , = / + ^ 1 п ( Х , + / г )
^2 =А: + /1п(Л '2 + '« )
у — площадь листьев , см^; — масса сухого вещества листьев , т§; — масса воды (разность м е ж д у свежей и абсолютно сухой массами ткани) , мг; а (14.076), Ь (0.671), с (0.220), а (0.738), е ( - 0 . 6 6 5 ) , / ( - 3 . 5 2 5 ) , ^ (0.833), И (-17.191), к (-3.588), / (0.759), т ( ^ 9 . 0 4 5 ) — к о э ф ф и ц и е н т ы . А н а л и з остатков показал до ст ат о чно в ы с о к у ю для такого изменчивого материала точность модели: средняя относительная о ш ибка аппроксимации сост ави ла 22.3%, средняя квадратичная — 28.8%, к о э ф ф и ц и е н т н е с о в п а д е ния Тейла (см.: Гильманов , 1978)— 11.8%. Г р а ф и ч е с к и й анализ остатков (который мы здесь не приводим в силу его громоздкости) свидетельствует о х о р о ш е м качестве аппроксимации исходных д а н н ы х моделью.

Глава 9. СОДЕРЖАНИЕ ВОДЫ 195
Рассчитав по этой модели п л о щ а д и листьев для всей вы бо р ки исходных д а н н ы х (представленной в табл. 9.1), м ы можем определить с л е д у ю щ и е п а р а м е т р ы :
ЗЬО = ОМ18,
5Ш= И'М/З,
8ЬР=310 + 8Ш= {ОМ+\УМ)13,
51А = 5-т/ом = \00/8ЬО,
где 8Ю — удельная масса сухого вещества листьев (8рес1Гю Ьеа^ 'Вгу [ т а з з ] ) , мг -см'^; 5Ш— удельная масса воды в листьях (8рес1йс Ьеа1'\Уа1ег [ т а з з ] ) , мг-см~^; 81Р — удельная масса свежих листьев (8рес1Г1с ЬеаГРгекЬ [так»] ) , мг • см~^; 81А — удельная площадь листьев (8рес1Йс ЬеаГ Агеа), л ш ^ • мг~^, ОМ— сухая масса листьев, мг; \УМ— масса воды в листьях , мг; 8 — площадь листьев , см^.
Удельная масса сухого вещества (в н а ш и х обозначениях 8Ш, а в и н о с т р а н н о й и современ н ой от еч ественной литературе она обозначается как 8ЬМ— З р е с Ш с ЬеаГ Мазз) есть не что иное , как показатель р ы х л о с т и сложения м е з о ф и л л а ' . Ч е м в ы ш е 810 (количество сухой массы на единицу площади) , т ем более плотно сложены листья, чем он ниже, т ем больше рыхлость мезофилла . Действительно , фактические данные показывают, что 810 имеет т е с н у ю отрицательную к о р р е л я ц и ю с суммой парциальных о б ъ е м о в межклетников и воздушных полостей^ у о дно до ль ных растений (рис. 9.3, а) и п а р ц и а л ь н ы х объемов межклетников (особенно м е ж к л е т н и к о в губчатой п а р е н х и м ы ) у двудольных (рис. 9.3, г). Это то же самое , что п о л о ж и т е л ь н ы е корреляции 8ЬО с с у м м о й всех тканей листа . Однако м о ж н о найти т е с н ы е статистичес кие связи 8ЬО с п а р ц и а л ь н ы м и объемами отдельных, наиболее л и г н и ф и ц и р о в а н н ы х и наименее оводненных , тканей мезофилла . Т а к и м и т к а н я м и оказались п р о в о д я щ а я у однодольных (рис. 9.3, б) и двудольных (рис 9.3, д), губчатая паренхима у однодольных (рис. 9.3, в) и паренхимная обкладка пучков у двудольных (рис . 9.3, еУ.
Поскольку 8ЬО отрицательно коррелирует с удельной массой воды {8ЫУ) в лис тьях р а с т е н и й (рис . 9.4), то м о ж н о ожидать , что п о с л е д н и й п а р а м е т р также м о ж е т являться показателем р ы х л о с т и сложения мезофилла . И з а н а л и з а материала видно , что 8ЬИ^ тесно коррелирует с суммой парциальных объемов межклетников и воздушных полостей у однодольных растений (рис. 9.5, а) и п а р ц и а л ь н ы м и о бъемами м е ж клетников губчатой п а р е н х и м ы у двудольных (рис . 9.5, б). Ч е м б о л ь ш е р а з в и т ы соответствующие свободные пространства в листьях однодольных и двудольных растений, т ем б о л ь ш е удельная масса воды. М о ж н о предпо ло жит ь , что по крайней мер е
' Ранее в отечественной литературе этот показатель назывался удельной поверхностной плотностью листьев (УППЛ). ^ Для однодольных (в отличие от двудольных) часто характерно наличие достаточно больших объемов воздушных полостей в мезофилле. ' Интересно отметить, что насыщающее содержание воды положительно коррелирует с паренхимной обкладкой пучков (но не с губчатой паренхимой) у однодольных и с губчатой паренхимой (но не с паренхимной обкладкой пучков) у двудольных. Это может служить косвенным подтверждением того, что эти ткани (губчатая паренхима у однодольных и паренхимная обкладка пучков у двудольных) наименее оводнены и наиболее лигнифицированы.
7*
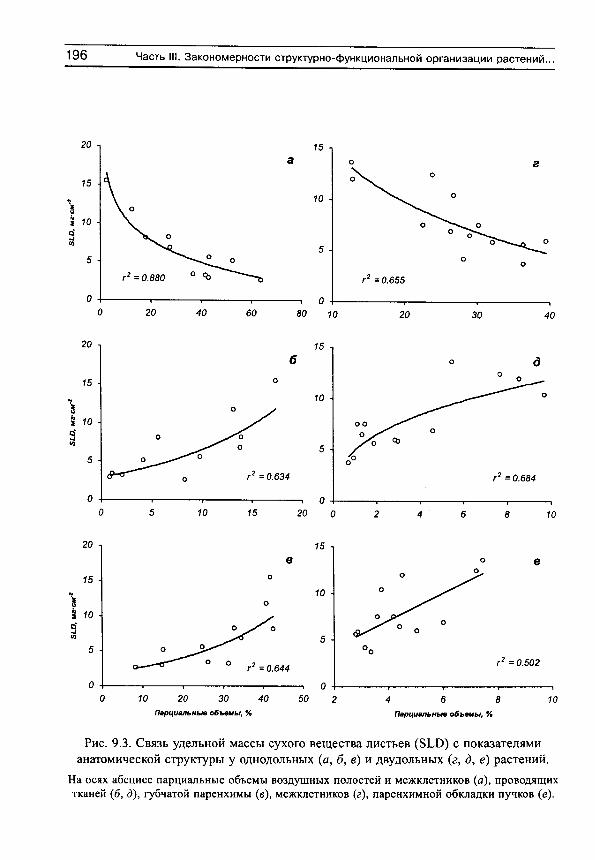
196 Часть III. Закономерности структурно-функциональной организации растений.
О -I , , , , , О -I , . , 1
о 10 20 30 40 50 2 4 6 8 10 Парциальные объемы, % Парциальмьш объемы, %
Рис. 9.3. Связь удельной массы сухого вещества листьев ( 8 Ь В ) с показателями анатомической структуры у однодольных (а, б, в) и двудольных (г, д, е) растений .
На осях абсцисс парциальные объемы воздушных полостей и межклетников (а), проводящих тканей (б, д), губчатой паренхимы (в), межклетников (г), паренхимной обкладки пучков (е).

Глава 9. СОДЕРЖАНИЕ ВОДЫ 197
Рис . 9.4. Связь удельной массы сухого в е щ е с т в а ( 8 Ь О ) с у д е л ь н о й м а с с о й воды ( 8 ^ \ V ) в л и с т ь я х однодольных и
двудольных растений.
67% (что соответствует д о л е объясненной вариации д л я двух связей на рис . 9.5) свободной апопластнои воды в л и стьях растений локализуется в их межклетных пространствах ' , а объем симп-ластной воды, т а к и м образом, не может п р е в ы ш а т ь 33% от ее о б щ е г о количества (в свободном состоянии) .
Рассмотрим изменения 8ЬО, 8ЫУ и 8ЬР на градиенте влажности почвы (рис. 9.6) . Удельная м а с с а сухого в е щ е с т в а листьев {810) уменьшается вдоль градиента в л а ж н о с т и почвы. М а к с и м а л ь н а я скорость и з м е н е н и й этого п о к а з а т е л я наблюдается в аридной части градиента (в среднем по л и н и и регрессии н а учас-
20 ^
~ 75
10
г' = 0.674
10 15 20 25 Парциальные объемы, !
Р и с . 9.5. С в я з ь у д е л ь н о й м а с с ы в о д ы (8Ь\ \ ' ) в листьях однодольных (а) и двудольны х (б) растений с п а р ц и а л ь н ы м и о б ъ е м а м и м е ж к л е т о ч н ы х п р о с т р а н с т в .
На осях абсцисс парциальные объемы воздушных полостей и межклетников (а), меж
клетников губчатой паренхимы (б).
тке градиента от 4 д о 50 м м эта «скорость» составляет 0.114 мгслГ'^ - мм''). В средней и гумидной частях градиента «скорость» изменений 810 н е з н а ч и т е л ь н а и р а в н а соответственно 0.009 и 0.004 мг-слГ'^ -мм'^, так что о н а практически н е о т л и ч и м а от нуля. Разброс р е а л ь н ы х значений может быть достаточно б о л ь ш и м (рис . 9.6), однако тренд не обн аруж и вает сколько-нибудь значительных и з м е н е н и й 8ЬП (т. е. по сути удельной п лот н ост и м е з о ф и л л а ) на большей части г р а д и е н т а в л а ж н о с т и почвы.
' М. I. Саппу (1995), например, относит к апопластнои воде не только воду клеточных стенок, но и всего межклеточного пространства.

198 Часть III. Закономерности структурно-функциональной организации растений.
30 1
Запас воды в почве, им Запас воды в почве, мм
Рис . 9.6. З а в и с и м о с т ь удельной массы сухого вещест ва ( 8 Ь В ) (1), воды (ЗЬХ^') (2), с в е ж и х н а в е с о к ( 8 Ь Р ) (3) л и с т ь е в однодольных и двудольных склерофитов
от в л а ж н о с т и почвы.
Рис. 9.7. Зависимость удельной площади листьев ( 8 Ь А ) однодольных и двудольных склерофитов от в л а ж н о с т и почвы.
З е р к а л ь н ы м о т р а ж е н и е м п р е д ы д у щ е й связи является и з м е н е н и е удельного количества воды (ЗЬЩ на г р а д и е н т е влажности почвы. Н аибо лее б ы с т р о 5 / , и з м е н я е т ся в аридной части градиента , а в средней и гумидно й его частях скорость из мене ний этого показателя з а м е т н о снижается (рис . 9.6).
Компенсирующие друг друга (комплементарные) изменения 8ЬО и 5/, приводят к тому, что удельная свежая масса листьев (8ЬР) остается п р а к т и ч е с к и н е и з м е н н о й на всем п р о т я ж е н и и градиента в л а ж н о с т и почвы (рис. 9.6). Д р у г и м и с л о в а м и , е сли при д в и ж е н и и вдоль градиента в л а ж н о с т и почвы м о ж н о наблюдать т е н д е н ц и ю к уве личению свежей массы листьев , то в удельном в ы р а ж е н и и (на е д и н и ц у п л о щ а д и ) эта закономерность не проявляется .
Известно, что площадь листьев увеличивается от аридных к г у м и д н ы м местообитаниям. Однако удельная (по о т н о ш е н и ю к единице сухой массы) л и с т о в а я п л о щ а д ь {8ЬА) в ари дн ой и средней частях градиента почти не изменяется , но резко возрастает в его гумидной части (рис. 9.7).
Динамики содержания воды в листьях растений
М н о ж е с т в е н н ы й р е г р е с с и о н н ы й анализ показал , что о с н о в н ы м фактором среды, воздействующим н а содержание воды в листьях растений, является в л а ж н о с т ь почвы. Рассмотрим, как и з м е н я ю т с я р а з л и ч н ы е средние в е л и ч и н ы на г р а д и е н т е в л а ж н о с т и

Глава 9. СОДЕРЖАНИЕ ВОДЫ 199
почвы, с этой целью целесообразно провести анализ 1) средних уровней (средних для каждого сообщества значений) , 2) средневидовых значений всего массива д а н н ы х , 3) средневидовых значений этого показателя у однодольных и двудольных растений при изменении условий увлажнения экотопов.
Д и н а м и к а с р е д н и х у р о в н е й с о д е р ж а н и я воды на г р а д и е н т е в л а ж н о с т и почвы. Средние у р о в н и оводненности листьев тесно связаны с в л а ж н о с т ь ю почвы (рис. 9.8, а). В и д н о , что на б о л ь ш е й части градиента эти средние в е л и ч и н ы уве личиваются сравнительно монотонно . Они резко возрастают в самой гумидной его части, где произрастают сообщества гидрофитов и гигрофитов . И з менения средних уровней содержания воды в листьях растений на градиенте влажности почвы можно описать уравнением:
5 п
4 •
1 •
у = а + Ь е х р ( - х / с ) ,
У- содержание воды в листьях расте-„-1
НИИ, г^„^у •Тсух.л.ассы ; х — запас воды в верхнем (0-30 см) слое п о ч в ы , мм; а (1.211), Ь (0.140), с (-^8.3) — коэффициенты. К о э ф ф и ц и е н т м н о ж е с т в е н н о й корреляции равен 0.846, /^-критерий — 35.7, стандартная ошибка уравнения —
0.338 Г„„й„ •Г'сух.массы • При ИСПОЛЬЗОВЗ-
нии ЭТОГО у р а в н е н и я , судя п о всему, м о ж н о п о л у ч и т ь д о с т а т о ч н о т о ч н ы е оценки среднего для экотопа содержания воды в ЛИСТЬЯХ р а с т е н и й по известным величинам в л а ж н о с т и почвы.
7 п
е -
5 •
8"
= 0,846
б о
о — о
=0536
Рис. 9.8. Д и н а м и к а с о д е р ж а н и я воды на градиенте в л а ж н о с т и почвы:
а) средних уровней, б) средневидовых значений и трендов минимальных (нижняя линия) и максимальных (верхняя линия) для каждого участка средневидовых величин, в) средневидовых значений у однодольных (1) и двудольных (2) склерофитов (прерывистые линии — доверительные
уровни).
7 1
О -I ^ 1 1
О 50 700 750 Запас воды в почве, мм

200 Часть III. Закономерности структурно-функциональной организации растений..
Д и н а м и к а средневидовых значений содержания в о д ы в листь ях на градиенте влаж ност и п о ч в ы . Ш и р и н а зоны разброса средневидовых значений оводненности листьев увеличивается от аридной части градиента к г у м и д н о й (рис . 9.8, б). В наиболее сухой части градиента у многих в и д о в наблюдаются д о с т а т о ч н о близкие величины содержания воды. П о мере продвижения вдоль г р а д и е н т а в л а ж н о с т и по чвы (в сторону ее увеличения) сходство между видами (по средним значениям оводненности листьев) уменьшается , достигая минимума в самых у в л а ж н е н н ы х экотопах. Увеличение м е ж в и д о в о й дифференциации по данному параметру водообмена может отражать, на н а ш взгляд, д и ф ф е р е н ц и а ц и ю растений по экологическим н и ш а м . П р и этом в самых гуми дных экотопах можно обнаружить такие же средневидовые значения содержания воды в листьях растений, как и в самых аридных (рис . 9.8, б). Скорость изменения (рассчитанная по линиям регрессии — рис . 9.8, б; для трендов мин и м а л ь н ы х з н а ч е н и й = 0.657, д л я т р е н д о в м а к с и м а л ь н ы х в е л и ч и н И = 0.752) п о градиенту м и н и м а л ь н ы х для каждого сообщества средневидовых значений оводненности листьев составляет 0.009 мг-см~'^ -мм"^, тогда как такая скорость для максимальных значений почти в 3 раза больше (0.024 мг-см'^ -мм'^). Этим подтверждается мнение , что во м н о г и х случаях невозможно найти р а з л и ч и я м е ж д у видами из с о о б щ е с т в с с а м ы м и р а з н ы м и у с л о в и я м и п о ч в е н н о г о у в л а ж н е н и я . С и т у а ц и я н е сколько у л у ч ш и т с я , если принимать во внимание , по крайней мере , т а к с о н о м и ч е с кую п ри н адлеж н ость растений , т. е. их принадлежность к классам однодольных или двудольных.
Д и н а м и к а с р е д н е в и д о в ы х з н а ч е н и й о в о д н е н н о с т и л и с т ь е в о д н о д о л ь н ы х и двудольных р а с т е н и й на градиенте влажности почвы. О д н о д о л ь н ы е и двудольные растения з амет н о р а з л и ч а ю т с я по содержанию воды на изменения у с л о в и й увлажнения экотопов. У о д н о д о л ь н ы х растений линия регрессии и з м е н е н и й в л а ж н о с т и лис тьев (рис. 9.8, в, экспоненциальная регрессия , = 0.675) на протяжении б о л ь ш е й части градиента располагалась ниже такой линии для однодольных (рис. 9.8, в, степенная регрессия , г' = 0.534). Только в сообществе п р и б р е ж н о в о д н ы х г и д р о ф и т о в произошло резкое увеличение оводненности листьев у однодольных (которые и представляли эту экологи ч ескую группу растений) . В то ж е в р е м я и н т е р е с н о о т мет ит ь , что у двудольных, п о с л е некоторого увеличения содержания воды в л и с т ь я х в ар идно й части градиента , л и н и я р е г р е с с и и демонстрирует л и ш ь н е з н а ч и т е л ь н ы й п о д ъ е м этого показателя при п р о д в и ж е н и и вдоль градиента влажности почвы вплоть д о самых гумидных экотопов.
Динамика показателей вариации содержания воды в листьях
С р е д н е в и д о в ы е к о э ф ф и ц и е н т ы в а р и а ц и и , п р и п о м о щ и к о т о р ы х м ы о ц е н и в а л и подвижность с о д е р ж а н и я воды в листьях растений , и м е ю т л и ш ь с л а б у ю т е н д е н ц и ю к увеличению против градиента влажности почвы (рис. 9.9, а) (связь коэффициентов вариации с другими факторами среды отсутствует п о л н о с т ь ю ) . Н е с м о т р я на то , что коэффициент корреляции статистически достоверен , эта т е н д е н ц и я в о с н о в н о м подтверждается только рядом точек в самой сухой части градиента , где наблюдалось наибольшее варьирование признака . П о э т о м у м о ж н о сказать, что л а б и л ь н о с т ь оводненности листьев остается в среднем практически неизменной на всем протяжении гра-
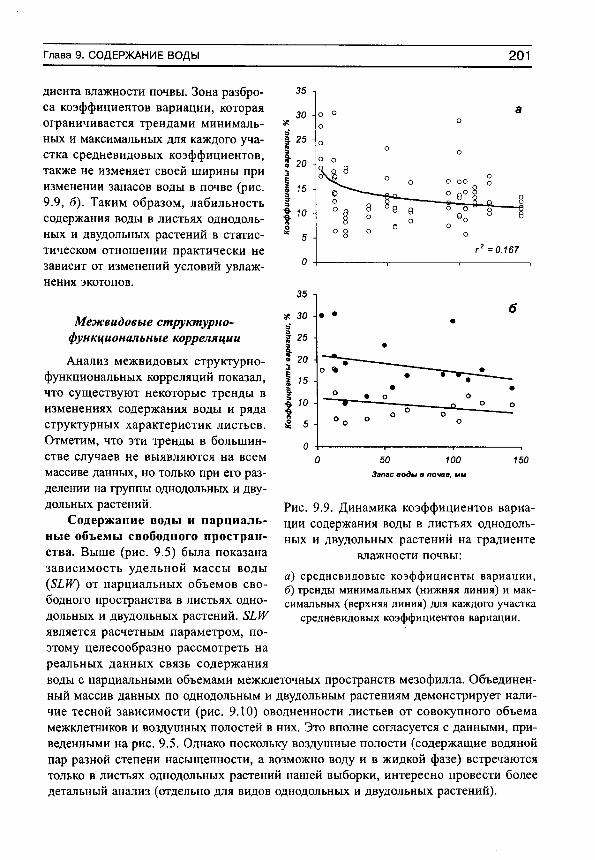
Глава 9. СОДЕРЖАНИЕ ВОДЫ 201
35 -,
30 -
25 -
20 -
15
10
=0.167
35
5? 30
| 2 5 э • 20
!•= 4 10
1 5
диента влажности почвы. Зона разброса к о э ф ф и ц и е н т о в в а р и а ц и и , которая о г р а н и ч и в а е т с я т р е н д а м и м и н и м а л ь ных и максимальных для каждого участка с р е д н е в и д о в ы х к о э ф ф и ц и е н т о в , также не изменяет своей ш и р и н ы при изменении запасов воды в почве (рис. 9.9, б). Т а к и м о б р а з о м , л а б и л ь н о с т ь содержания воды в листьях однодольных и двудольных растений в статист и ч е с к о м о т н о ш е н и и п р а к т и ч е с к и не зависит от и з м е н е н и й у с л о в и й увлажнения экотопов.
Межвидовые структурно-функциональные корреляции
А н а л и з м е ж в и д о в ы х с т р у к т у р н о -ф у н к ц и о н а л ь н ы х корреляций показал , что с у щ е с т в у ю т н е к о т о р ы е т р е н д ы в и з м е н е н и я х с о д е р ж а н и я в о д ы и р я д а с т р у к т у р н ы х х а р а к т е р и с т и к л и с т ь е в . О т м е т и м , что эти т р е н д ы в б о л ь ш и н стве с л у ч а е в не в ы я в л я ю т с я на в с е м массиве данных, но только при его разделении на группы однодольных и двудольных растений .
С о д е р ж а н и е в о д ы и п а р ц и а л ь н ы е о б ъ е м ы с в о б о д н о г о п р о с т р а н ства. В ы ш е (рис . 9.5) б ы л а п о к а з а н а з а в и с и м о с т ь у д е л ь н о й м а с с ы в о д ы (8ЫУ) от п а р ц и а л ь н ы х о б ъ е м о в с в о бодного пространства в л и с т ь я х однодольных и двудольных растений . является р а с ч е т н ы м п а р а м е т р о м , п о этому ц е л е с о о б р а з н о р а с с м о т р е т ь н а р е а л ь н ы х д а н н ы х с в я з ь с о д е р ж а н и я
воды с п а р ц и а л ь н ы м и объемами межклеточных пространств мез о филла . О бъединенн ы й массив д а н н ы х по однодольным и двудольным р а с т е н и я м д е м о н с т р и р у е т наличие т е с н о й з а в и с и м о с т и (рис . 9.10) о в о д н е н н о с т и л и с т ь е в от с о в о к у п н о г о о б ъ е м а межклетников и воздушных полостей в них. Это вполне согласуется с д а н н ы м и , приведенными на р и с . 9.5. Однако поскольку в о з д у ш н ы е п о л о с т и ( с о д е р ж а щ и е водяной пар разной степени н а с ы щ е н н о с т и , а возможно воду и в ж и д к о й фазе) встречаются только в л и с т ь я х однодольных растений нашей в ы б о р к и , и н т е р е с н о провести более детальный анализ (отдельно для видов однодольных и двудольных р аст ений) .
50 100 150 Запас воды в почве, мм
Рис . 9.9. Д и н а м и к а к о э ф ф и ц и е н т о в вариации содержания воды в л и с т ь я х однодольн ы х и д в у д о л ь н ы х р а с т е н и й на г р а д и е н т е
в л а ж н о с т и почвы:
а) средневидовые коэффициенты вариации, б) тренды минимальных (нижняя линия) и максимальных (верхняя линия) для каждого участка
средневидовых коэффициентов вариации.
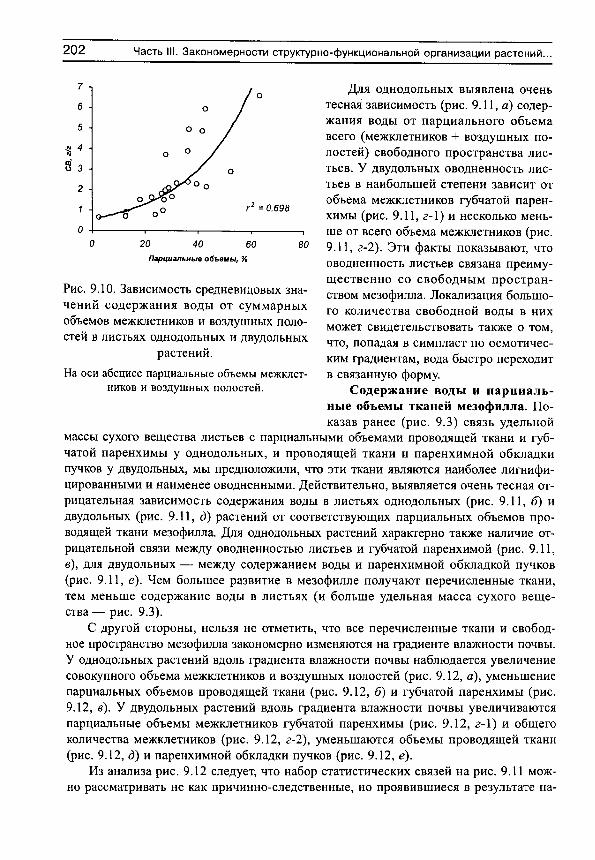
202 Часть III. Закономерности структурно-функциональной организации растений...
Д л я о д н о д о л ь н ы х в ы я в л е н а о чень тесная зависимость (рис. 9.11, а ) содерж а н и я в о д ы от п а р ц и а л ь н о г о о б ъ е м а всего (межклетников -ь в о з д у ш н ы х полостей ) сво бо дно г о п р о с т р а н с т в а л и с тьев . У двудольных о в о д н е н н о с т ь лис тьев в н а и б о л ь ш е й ст епени зависит от объема м е ж к л е т н и к о в губчатой паренх и м ы (рис. 9.11, г-1) и несколько меньше от всего объема межклетников (рис. 9.11, г-2). Э т и ф а к т ы показывают , что оводненность л и с т ь е в связана преимущ е с т в е н н о с о с в о б о д н ы м п р о с т р а н ством мезофилла. Локализация большого к о л и ч е с т в а с в о б о д н о й в о д ы в н и х может свидетельствовать т а к ж е о том, что, попадая в с и м п л а с т по о с м о т и ч е с ким градиентам, вода быстро переходит в связанную форму.
С о д е р ж а н и е в о д ы и п а р ц и а л ь н ы е о б ъ е м ы т к а н е й м е з о ф и л л а . П о казав р а н е е ( р и с . 9.3) свя з ь у д е л ь н о й
массы сухого вещества листьев с парциальными объемами проводящей ткани и губчатой п а р е н х и м ы у о д н о д о л ь н ы х , и п р о в о д я щ е й т к а н и и п а р е н х и м н о й о б к л а д к и пучков у двудольных, мы предположили, что эти ткани являются наиболее лигнифи-ц и р о в а н н ы м и и н а и м е н е е о в о д н е н н ы м и . Действительно , в ы я в л я е т с я очень т е с н а я отрицательная з а в и с и м о с т ь содержания воды в л и с т ь я х о д н о д о л ь н ы х (рис . 9.11, б) и двудольных (рис. 9.11, д) растений от соответствующих п а р ц и а л ь н ы х объемов пр о водящей ткани мезофилла. Для однодольных растений характерно также наличие отрицательной связи между оводненностью листьев и губчатой п а р е н х и м о й (рис. 9.11, в), для двудольных — м е ж д у с о д е р ж а н и е м воды и п а р е н х и м н о й о б к л а д к о й пучков (рис. 9.11, е) . Ч е м большее развитие в м е з о ф и л л е п о л у ч а ю т п е р е ч и с л е н н ы е ткани , тем м е н ь ш е с о д е р ж а н и е в о д ы в л и с т ь я х (и б о л ь ш е у д е л ь н а я м а с с а су х о г о в е щ е ства — рис . 9.3).
С другой стороны, нельзя не отметить , что все п е р е ч и с л е н н ы е т к а н и и свободное пространство мезофилла закономерно изменяются на градиенте влажности почвы. У однодольных растений вдоль градиента влажности почвы наблюдается увеличение совокупного объема межклетников и воздушных полостей (рис . 9.12, а ) , у м е н ь ш е н и е парциальных объемов проводящей ткани (рис . 9.12, б) и губчатой п а р е н х и м ы (рис. 9.12, в). У двудольных растений вдоль градиента в л а ж н о с т и п о ч в ы у в е л и ч и в а ю т с я парциальные о б ъ е м ы м е ж к л е т н и к о в губчатой п а р е н х и м ы (рис . 9.12, г -1) и общего количества м е ж к л е т н и к о в (рис . 9.12, г-2) , у м е н ь ш а ю т с я о б ъ е м ы п р о в о д я щ е й ткани (рис. 9.12, д) и п а р е н х и м н о й обкладки пучков (рис. 9.12, е).
Из анализа рис . 9.12 следует, что набор статистических связей н а р и с . 9.11 можно рассматривать не как причинно-следственные , н о п р о я в и в ш и е с я в результате па-
Парциальные объемы, %
Рис. 9.10. Зависимость средневидовых знач е н и й с о д е р ж а н и я в о д ы от с у м м а р н ы х объемов межклетников и воздушных полостей в листьях однодольных и двудольных
р а с т е н и й .
На оси абсцисс парциальные объемы межклетников и воздушных полостей.

Глава 9. СОДЕРЖАНИЕ ВОДЫ 203
О 20 40 60 80 О 10 20 30 40
7 5 п
О -I , , , , , О -I , , , О 10 20 30 40 50 2 4 6 8
Парциальные объемы, % Парциальные объемы, %
Рис. 9.11. З а в и с и м о с т ь содержания воды от парциаль ных объемов тканей м е з о ф и л ла и свободных пространств в листьях однодольных (а, б, в) и дву до ль ны х (г, д, е)
растений .
На осях абсцисс парциальные объемы межклетников и воздушных полостей (а), проводящих тканей (5, д), губчатой паренхимы (в), межклетников губчатой паренхимы (г-1), межклетников
(г-2), паренхимной обкладки пучков (е).
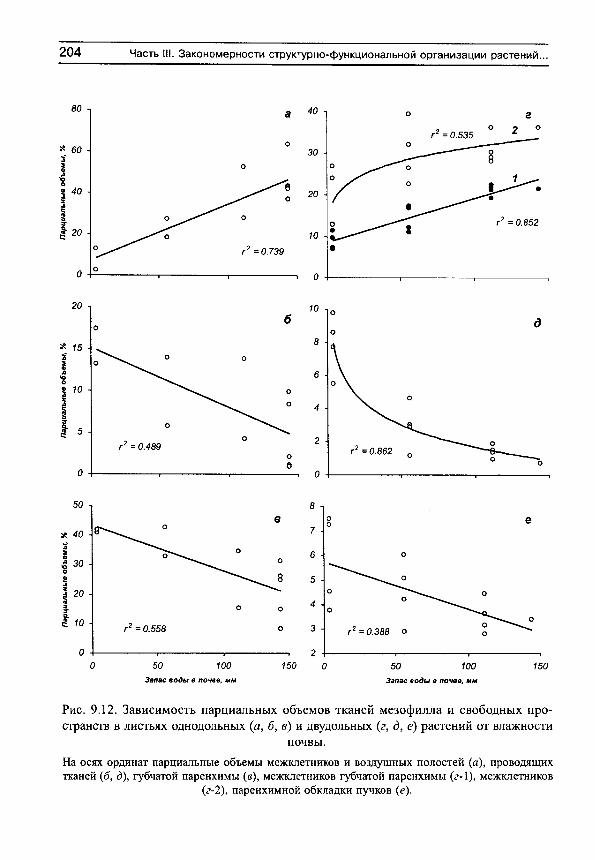
204 Часть III. Закономерности структурно-функциональной организации растений.
а "О
Запас воды в почве, им Запас воды в почве, мм
Рис . 9.12. З а в и с и м о с т ь п а р ц и а л ь н ы х объемов т к а н е й м е з о ф и л л а и с в о б о д н ы х пр о странств в листьях однодольных (а, б, в) и двудольных (г, д, ё) растений от влажности
почвы.
На осях ординат парциальные объемы межклетников и воздушных полостей (а), проводящих тканей (б, д), губчатой паренхимы (в), межклетников губчатой паренхимы (г-1), межклетников
(г-2), паренхимной обкладки пучков (е).

Глава 9. СОДЕРЖАНИЕ ВОДЫ 205
раллельных м о д и ф и к а ц и й структуры листьев и их о в о д н е н н о с т и на г р а д и е н т е влажности почвы. П р е д п о л о ж и м , однако, что связи, по каз анные на р и с . 9.11, не являются п р и ч и н н о - с л е д с т в е н н ы м и и не отражают р е а л ь н ы х с т р у к т у р н о - ф у н к ц и о н а л ь н ы х в заимоот н ошен и й . Тогда все изменения водообмена и о с о б е н н о с т е й анатомического строения листьев просто п а р а л л е л ь н ы изменениям условий почвенного водоснабжения и никак не взаимосвязаны. Но могут ли достаточно глубокие структурные перестройки находиться вне связи с функциональными? М ы предполагаем, что очень мала в е р о я т н о с т ь с о х р а н е н и я с т р у к т у р н о - ф у н к ц и о н а л ь н о г о п а р а л л е л и з м а на г р а д и е н т е влажности почвы большой (практически максимальной) п р о т я ж е н н о с т и без наличия структурно-функциональных взаимосвязей . Вследствие этого м ы с к л о н н ы рассматривать статистические связи, показанные на рис . 9.11, как п р и ч и н н о - с л е д с т в е н н ы е .
Содержание воды в листьях однодольных склер о фит о в н а п р о т я ж е н и и б о л ь ш е й части градиента в л а ж н о с т и почвы заметно м е н ь ш е , ч е м в л и с т ь я х двудольных склерофитов (рис. 9 .1 , а, 9.8, в). Это , судя по всему, связано с несколько большей скле-р о м о р ф н о с т ь ю листьев однодольных растений , у которых в среднем удельная масса сухого в е щ е с т в а (по ф а к т и ч е с к и м д а н н ы м ) несколько б о л ь ш е , ч е м у д в у д о л ь н ы х (9.5 мг-см''^ у однодольных против 7.3 мг-см'''' у дву до ль ны х ; р а з н о с т ь средних, оцененная по ^-критерию Стьюдента , достоверна при Р < 0.01). К р о м е того, удельная масса сухого вещества связана с парциальными объемами тканей, которые у однодольных растений р а з в и т ы б о л ь ш е , чем у двудольных. У о д н о д о л ь н ы х это п р о в о д я щ а я ткань и губчатая п а р е н х и м а (рис. 9.3, б, в), у двудольных — п р о в о д я щ а я ткань и паренхимная обкладка пучков (рис . 9.3, д, е).
В м е с т е с тем анализ показал , что содержание воды в л и с т ь я х р а с т е н и й связано п р е и м у щ е с т в е н н о с о с в о б о д н ы м и , м е ж к л е т о ч н ы м и п р о с т р а н с т в а м и (рис . 9.10). П о добная связь о в о д н е н н о с т и листьев с их п л о т н о с т ь ю б ы л а о б н а р у ж е н а для 14 видов злаков в работе Е. Оагп1ег и О. ЬаигеЩ (1994). Это неоднозначно свидетельствует о том, что вода (в п арообразном состоянии) л о к а л и з о в а н а в о с н о в н о й сво ей массе (до 67-70% свободной воды) в межклеточных пространствах листьев . С а м а во з мо жно ст ь поддержания достаточно интенсивной т р а н с п и р а ц и и м о ж е т зависеть от количества воды в межклетниках . Так, А. А . Зялалов (1984) отмечает, что при по ниженно й влажности межклетников падает проводимость , затормаживается п о с т у п л е н и е воды к ус тьицам и околоустьичным клеткам и, следовательно, снижается интенсивность транспирации (с другой стороны, локализация большого количества свободной воды в них может свидетельствовать также о том, что, попадая в симпласт по о с м о т и ч е с к и м градиентам, вода б ы с т р о переходит в с в я з а н н у ю фо р му ) . Л о кализ ация б о л ь ш о г о количества воды в межклетниках , т а к и м , образом, д о л ж н а приводить к очень т е с н о й взаи м о з а в и с и м о с т и м е ж д у о в о д н е н н о с т ь ю л и с т ь е в и т р а н с п и р а ц и е й в п о д а в л я ю щ е м большинстве случаев . К р о м е того , такая локализация зависит от п л о т н о с т и с л о ж е н и я а с с и м и л и р у ю щ и х органов . П о э т о м у р а з л и ч и я м е ж д у о д н о д о л ь н ы м и и д в у д о л ь н ы м и как п о с о д е р ж а н и ю воды в листьях , т а к и п о скорости ее и с п а р е н и я следует искать в разной с к л е р о м о р ф н о с т и листьев этих групп растений .
И з анализа связей, п о к а з а н н ы х на рис . 9.11, м о ж н о сделать до ст ат о чно о б о с н о ванное предположение о том, что наименее оводнены и н а и б о л е е л и г н и ф и ц и р о в а н ы у однодольных растений — проводящая ткань и губчатая паренхима , у двудольных — проводящая ткань и п а р е н х и м н а я обкладка (т. е. весь пучок) . Н а и м е н ь ш а я оводнен-

206 Часть III. Закономерности структурно-функциональной организации растений..
ность п р о в о д я щ е й ткани н е означает, что в н и х не с о д е р ж и т с я вода. К а к известно проводящая ткань неоднородна и состоит из ф л о э м ы и к с и л е м ы . В клетках фло э мы вода вполне может находиться преимущественно в связанной форме , а в клетках ксил е м ы (из которых, собственно , и поступает вода в м е з о ф и л л ) необязательно д о л ж н о быть б о л ь ш о е количество воды, но достаточно высокая скорость ее п е р е м е щ е н и я , которая с п о с о б н а п о д д е р ж и в а т ь в ы с о к у ю о в о д н е н н о с т ь т к а н е й и с в о б о д н о г о пр о странства мезофилла .
Из клеток к с и л е м ы п р о в о д я щ и х пучков у одно до ль ных р а с т е н и й вода поступает (кроме межклетников и воздушных полостей) в паренхимную обкладку пучков и столбчатую п а р е н х и м у (которая , в п р о ч е м , у д а н н о й г р у п п ы р а с т е н и й н а ш е й в ы б о р к и встречается достаточно редко) . У видов двудольных вода д о л ж н а поступать (кроме межклетников) в ткани губчатой и столбчатой п а р е н х и м ы . О т су т ст вие д о с т о в е р н ы х корреляций м е ж д у о в о д н е н н о с т ь ю листьев и п а р ц и а л ь н ы м и о б ъ е м а м и этих тканей может б ы т ь в ы з в а н о т е м , что вода в них достаточно бы ст р о переходит в связанную форму и не поддается о п р е д е л е н и ю .
Разброс средневидовых величин содержания воды на градиенте влажности почвы имеет « к л и н о в и д н ы й » характер . Узкая часть «клина» р а с п о л о ж е н а в ар идно й части градиента , ш и р о к а я — в г у м и д н о й (рис . 9.8, б ) . Сходство в и д о в в с о о б щ е с т в а х п о оводненности листьев изменяется с изменением положения фитоценозов на градиенте влажности почвы. В н а и б о л е е суровых условиях произрастания , с м и н и м а л ь н ы м количеством д о с т у п н о й почвенной влаги произрастают в и д ы с о чень п о х о ж и м и значениями этого показателя водного р е ж и м а . П о мер е п р о д в и ж е н и я вдоль г р а д и е н т а в сторону у в е л и ч е н и я в л а ж н о с т и почвы сходство видов п о с т о я н н о с н и ж а е т с я , д о с т и гая м и н и м у м а в с а м ы х г у м и д н ы х экотопах. Это п р е д о п р е д е л я е т то , что на протяжении практически всего градиента влажности почвы м о ж н о о б н а р у ж и т ь в и д ы с очень похожими значениями о в о д н е н н о с т и листьев .
Также как и в случае с и н т е н с и в н о с т ь ю т р а н с п и р а ц и и , в т р а в я н ы х с о о б щ е с т в а х аридных экосистем относительно высокое сходство с о д е р ж а н и я воды у р а з н ы х р ас тений может быть обусловлено жестким экотопическим отбором (через который проходит набор ви дов , в н а и б о л ь ш е й степени соответствующий д а н н ы м у с л о в и я м п р о израстания) и к р и т и ч е с к и м недостатком воды в почве ( в о з м о ж н о , т а к ж е , и с л а б о й выраженностью конкурентных отношений между н и м и ' ) . П о пу ля ции , и м е ю щ и е толерантные ж и зн ен н ые стратегии, накапливают такое количество воды, которое доступно им в этих м е с т о о б и т а н и я х и в соответствии со с в о и м и с т р у к т у р н ы м и о с о б е н н о с т я ми. Суккуленты ( о с о б е н н о АШит ро1угЫ2ит, табл . 9.1) и м е ю т очень в ы с о к у ю оводненность своих а с с и м и л и р у ю щ и х органов . У с к л е р о ф и т о в , однако , о г р а н и ч е н н ы е и одинаковые д л я всех особей ресурсы воды в почве в ы р а в н и в а ю т с р е д н и е уровни с о д е р ж а н и я воды. В и д о с п е ц и ф и ч н о с т ь среди них по э т о му п р и з н а к у не проявляется или практически отсутствует.
' Так, ]. Н. СоппеИ (1980, цит. по: Джиллер, 1988) полагает, что в очень суровых условиях среды численность популяций падает ниже тех уровней, при которых они конкурируют. В таких ситуациях межвидовая конкуренция уже по определению не может быть серьезной силой (Бигон и др., 1989). С другой стороны, в обзоре N. Роу/1ег (1986) приводятся доказательства наличия конкуренции в сообществах аридных и семиаридных экосистем.

Глава 9. СОДЕРЖАНИЕ ВОДЫ 207
В г у м и д н ы х местообитаниях , несмотря на то что запасы в о д ы в почве неограниченны (или достаточно обильны) , в одном и том же сообществе с о с у щ е с т в у ю т в и д ы как с высокой , т а к и с низкой о в о д н е н н о с т ь ю листьев (рис . 9.8, б) п р и достаточно слабом экотопическом отборе, но при более н а п р я ж е н н ы х к о н к у р е н т н ы х о т н о ш е н и ях между видами . Длительное и успешное сосуществование в фитоценозе ценопопу-ляций с к о н к у р е н т н ы м и стратегиями, и с п о л ь з у ю щ и х один и тот же р е с у р с , возможно только при разделении ими экологических н и ш (как в пространстве , так и во времени) . Наблюдаемое разнообразие средних значений оводненности листьев в гумидных экотопах, судя по всему, и является отражением такой дифференциации . Поскольку в результате разделения н и ш виды сохраняют д о м и н и р у ю щ е е положение в сообществах , то используемые и м и запасы почвенной влаги д о л ж н ы отвечать их конституционным потребностям. Поэтому можно предположить , что увеличение разнообразия средних величин содержания воды на градиенте в л а ж н о с т и почвы является следствием увеличения разнообразия видоспецифических реакций растений на изменения экологической обстановки . К р о м е того, накопленный ф а к т и ч е с к и й материал показал, что различия между видами на градиенте влажности почвы носят «структурноспеци-фический» характер (что связано с п р и н а д л е ж н о с т ь ю к таксонам такого ранга , как однодольные и двудольные) (рис . 9.8, в).
Резюме. Статистический анализ оводненности листьев т р а в я н и с т ы х растений на градиенте в л а ж н о с т и почвы показал следующее . В е с ь м а с с и в и с х о д н ы х д а н н ы х содержания воды в листьях растений имеет среднее значение х = 2.355 г^^^^, '^сух.массы •
Типичные значения этого показателя на градиенте в л а ж н о с т и п о ч в ы не п р е в ы ш а ю т 5 ^воды '^сух.массы • Наиболсе замстныс различия по оводненности листьев можно увидеть между видами однодольных и двудольных склерофитов . Соответствующие средние значения этого признака р а в н ы 1.889 и 2.278 г^^^^ '^сух.массы • Т и п и ч н ы е значения оводненности л и с т ь е в для однодольных с к л е р о ф и т о в находятся в пределах 0 .9-
3.3 Г^„йы •^~с1к.массы , д л я ДВуДОЛЬНЫХ — МСЖДу 1.0-3.8 Г«„йы •^'с1х.массы • ОдНОДОЛЬНЫС
суккуленты демонстрируют более широкий размах колебаний этого показателя — 2.4-6.3 ^воды '^сух.массы » тогда как двудольный суккулснт с к о р о н а р н ы м с и н д р о м о м (Апа-Ъаш ЬгеУ1/оИа) б о л ь ш е похож в этом о т н о ш е н и и на двудольные с к л е р о ф и т ы — 1.7-3-5 ^еоды '^су^.массы • П о к а з а н а т е с н а я з а в и с и м о с т ь п л о щ а д и л и с т ь е в от к о л и ч е с т в а воды и сухой м а с с ы в них . Рассчитаны показатели удельной м а с с ы сухого вещества {510), удельной м а с с ы воды {5ЬЩ, удельной м а с с ы с в е ж и х листьев {8ЬР), удельной площади листьев {8ЬА). В ы я в л е н ы корреляции 8ЬВ с п а р ц и а л ь н ы м и о б ъ е м а м и про водящей ткани у однодольных и двудольных, губчатой п а р е н х и м ы у однодольных и паренхимной обкладкой пучков у двудольных. Средние для каждого экотопа у р о в н и оводненности листьев тесно связаны с в л а ж н о с т ь ю почвы. Д и н а м и к а средневидовых значений оводненности листьев на градиенте влажности почвы у видов однодольных и двудольных растений заметно различается . Ш и р и н а з о н ы р а з б р о с а с р е д н е в и д о в ы х величин увеличивается от аридной к гумидной части градиента . Это и л л ю с т р и р у е т ся сравнением трендов м и н и м а л ь н ы х и максимальных для каждого сообщества средневидовых значений содержания воды. Изучение структурно-функциональных корреляций показало , что оводненность листьев связана п р е и м у щ е с т в е н н о со с в о б о д н ы м пространством мезофилла . Сделаны следующие предположения : а) наименее оводне-

208 Часть III. Закономерности структурно-функциональной организации растений.
ны и наиболее л и г н и ф и ц и р о в а н ы у однодольных р а с т е н и й — п р о в о д я щ а я ткань и губчатая паренхима, у двудольных — проводящая ткань и паренхимная обкладка пучков; б) из клеток ксилемы проводящих пучков у однодольных растений вода преимущественно поступает в паренхимную обкладку пучков и столбчатую паренхиму. У видов двудольных вода д о л ж н а поступать в ткани губчатой и столбчатой п а р е н х и м ы ; в) в травяных сообществах аридных экосистем относительно высокое сходство оводненности листьев у склерофитов может быть обусловлено жестким экотопическим отбором и недостатком воды в почве . Н а б л ю д а е м о е р аз но о бр аз ие средних величин содержания воды в г у м и д н ы х экотопах является отражением д и ф ф е р е н ц и а ц и и экониш; г) различия м е ж д у в и д а м и на градиенте влажности почвы н о с я т « с т р у к т у р н о с п е ц и -фический» харакгер, что связано с принадлежностью к таксонам такого высокого ранга, как однодольные и двудольные .

Глава 10. ОСМОТИЧЕСКОЕ ДАВЛЕНИЕ КЛЕТОЧНОГО СОКА 209
ГЛАВА 10. ОСМОТИЧЕСКОЕ ДАВЛЕНИЕ КЛЕТОЧНОГО СОКА
Осмотическое давление является одним из важнейших показателей водного р ежима растений . Его исследованию в полевых у с л о в и я х у д и к о р а с т у щ и х р а с т е н и й придается особое значение. В о б о б щ а ю щ и х сводках Г. Вальтера (1969, 1974, 1975) можно найти в е л и ч и н ы осмотического давления для т и п и ч н ы х представителей с о о б щ е с т в разных к л и м а т и ч е с к и х зон. Б о л ь ш о й вклад в изучение о смо т ическо г о д а в л е н и я внесла В . М . С в е ш н и к о в а (1956а , 19566, 1962, 1979). И с с л е д о в а н и ю этого показателя уделялось вн и ман и е в целом ряде работ (Бобровская , 1968; Свешникова , Бобровская , 1969; Г о р ш к о в а , 1971; И з м а й л о в а , 1977, 1978; Б а н н и к о в а и д р . , 1982; МесИпа, Ргапс18С0, 1994; 2о15, М Ш е г , 1994; 1оЬп5оп е1 а1., 1996; ЫШоп, КоЬе1 , 1999 и др . ) . Осмотическое д а в л е н и е как важная с о с т а в л я ю щ а я водного по т енциала рассматривается в о б з о р н ы х и теоретических работах (Слейчер , 1970; Н о б е л , 1973; М е л е щ е н к о , 2001; ЫоЬе1, 1983, 1991; Воуег, 1985; 81еис11е, 1989; З Н а г а т а т , МайЬауагао, 1997). М ы попыт аемся д о п о л н и т ь с у щ е с т в у ю щ у ю в литературе и н ф о р м а ц и ю к о л и ч е с т в е н н ы м описанием изменений осмотического давления клеточного сока листьев у ряда видов на градиенте в л а ж н о с т и почвы.
Д а н н ы е с о б р а н ы в 9 с о о б щ е с т в а х , р а с п о л о ж е н н ы х в р а з н ы х частях г р а д и е н т а влажности почвы (см. табл . 7 .1 , сообщества под номерами 2, 3, 7, 8, 10-13, 15). Исходя из з акономерностей (которые будут описаны н и ж е ) о п р е д е л е н ы р а с ч е т н ы е значения осмотического давления у видов сообществ 1, 6, 9, 14 и 16 (по табл . 7.1). Статистические характеристики с р е д н е в и д о в ы х величин осмотического давления показаны в таблице 10.1.
Статистический анализ осмотического давления клеточного сока листьев у видов однодольных и двудольных растений
Весь массив исходных д а н н ы х осмотического д а в л е н и я и м е е т среднее значение X = 2.08 М П а и стандартное отклонение = 0.71 М П а (« = 1188). Ряд р аспр еделе ния имеет п о л о ж и т е л ь н у ю а с и м м е т р и ю с модой, р а в н о й 1.68 М П а , а 90% его значений (вариант) находятся в диапазоне 1.32-3.26 М П а (квантили порядков 0.05 и 095) (5% вариант в ы ш е квантиля 0.95 распределены в д и а п а з о н е 3.26-6.16 М П а ) .
Однако это касается всего массива исходных данных. Если провести более детальное исследование , разбив выборку на группы однодольных и двудольных растений и выделив из этих групп в и д ы со с п о с о б н о с т ь ю накапливать воду в с п е ц и а л и з и р о в а н ных тканях (суккуленты) , то картина статистического описания заметно изменится .
Рассмот ри м н е с у к к у л е н т н ы е в и д ы (склерофиты) однодольных и двудольных растений. Статистические п а р а м е т р ы осмотического д а в л е н и я у этих групп р а с т е н и й настолько близки (табл. 10.2), что эти две выборк и м о ж н о объединить в одну. С другой стороны, у однодольных суккулентов осмотическое д а в л е н и е несколько м е н ь ш е , чем у склерофитов , а у двудольного суккулента {АпаЬазгз Ьгегг/оИа) с С^-САМ метаболизмом (Гамалей , Ш и й р э в д а м б а , 1988) о н о значительно (более чем в 2 раза ) пре вышает средние величины у других групп растений (табл. 10.2). Это м о ж е т б ы т ь связано с тем , что у растений, фотосинтезирующих по С^-САМ типу осмотически актив-
8 - 2353
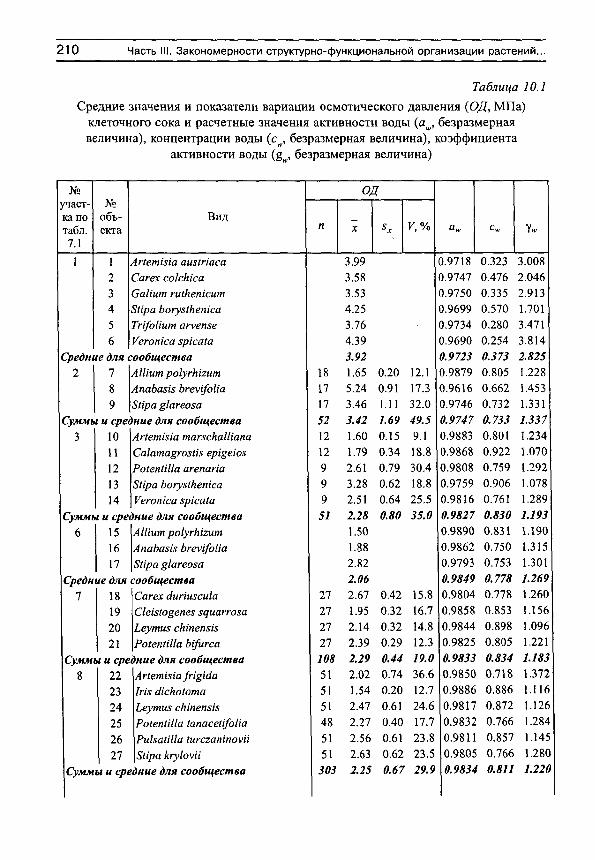
210 Часть III. Закономерности структурно-функциональной организации растений.
Таблица 10.1
Средние значения и показатели вариации осмотического д а в л е н и я (ОД, М П а ) клеточного сока и расчетные значения активности воды (а^, безразмерная величина) , концентрации воды (с^, безразмерная величина) , к о э ф ф и ц и е н т а
активности воды (§^, безразмерная величина)
участка по табл. 7.1
№ объекта
ОД
с^
участка по табл. 7.1
№ объекта
Вид п X с^
1 1 АПетша аих1паса 3.99 0.9718 0.323 3.008 2 Сагех сокЫса 3.58 0.9747 0.476 2.046 3 ОаИит Шкетсит 3.53 0.9750 0.335 2.913 4 ЗНра Ъогу51кеп1са 4.25 0.9699 0.570 1.701 5 Тп/оНит ап/епзе 3.76 0.9734 0.280 3,471 6 Уеготса 5р1саШ 4.39 0.9690 0.254 3.814
Средние для сообщества 3.92 0.9723 0.373 2.825 2 7 АШит ро1угЫ2ит 18 1.65 0.20 12.1 0.9879 0.805 1.228
8 АпаЬаш Ьгеу1/оИа 17 5.24 0.91 17.3 0.9616 0.662 1.453 9 8Ира §1агеоза 17 3.46 1.11 32.0 0.9746 0.732 1.331
Суммы и средние для сообщества 52 3.42 1.69 49.5 0.9747 0.733 1.337
3 10 АПет131а тапскаШапа 12 1.60 0.15 9.1 0.9883 0.801 1.234 И СаЬта^гозНз ерг^еюз 12 1.79 0.34 18.8 0.9868 0.922 1.070 12 РоШИИа агепапа 9 2.61 0.79 30.4 0.9808 0.759 1.292
13 8Ира ЬогузЛетса 9 3.28 0.62 18.8 0.9759 0.906 1.078 14 Уеготса зргсаШ 9 2.51 0.64 25.5 0.9816 0.761 1.289
Суммы и средние для сообщества 51 2.28 0.80 35.0 0.9827 0.830 1.193
6 15 АШит ро1угН12ит 1.50 0.9890 0.831 1.190
16 АпаЬаш Ьгеуь/оНа 1.88 0.9862 0.750 1.315
17 8Ира §1агеоза 2.82 0.9793 0.753 1.301
Средние для сообщества 2.06 0.9849 0.778 1.269
7 18 Сагех (1игш5си1а 11 2.67 0.42 15.8 0.9804 0.778 1.260
19 С1еШо§епе8 щиаггоза 11 1.95 0.32 16.7 0.9858 0.853 1.156
20 Ьеутих сЫпепш 11 2.14 0.32 14.8 0.9844 0.898 1.096
21 РоШИИа Ы/игса 11 2.39 0.29 12.3 0.9825 0.805 1.221
Суммы и средние для сообщества 108 2.29 0.44 19.0 0.9833 0.834 1.183
8 22 АПетша /п§Ша 51 2.02 0.74 36.6 0.9850 0.718 1.372
23 1пз сИскоЮта 51 1.54 0.20 12.7 0.9886 0.886 1.116
24 Ьеутиз сктеп81з 51 2.47 0.61 24.6 0.9817 0.872 1.126
25 РоШИПа ШпасеИ/оНа 48 2.27 0.40 17.7 0.9832 0.766 1.284
26 РиЫйИа (игсгапшоуИ 51 2.56 0.61 23.8 0.9811 0.857 1.145
27 8Ира кгуЫИ 51 2.63 0.62 23.5 0.9805 0.766 1.280
Суммы и средние для сообщества 303 2.25 0.67 29.9 0.9834 0.811 1.220

Глава 10. ОСМОТИЧЕСКОЕ ДАВЛЕНИЕ КЛЕТОЧНОГО СОКА 211
Продолжение таблицы 10.1
№ участка по табл.
7.1
№ объекта
ОД
а„, с„
№ участка по табл.
7.1
№ объекта
В и д п X У,% а„, с„
9 28 Са1ата§говйз ерщею8 2.33 0.9830 0.842 1.168 29 Сагех те1апо8ШсНуа 2.43 0.9823 0.784 1.253 30 СаИит шНетсит 1.99 0.9855 0.787 1.252 31 ЬуЛгит У1г§а1ит 2.33 0.9831 0.794 1.238 32 2.04 0.9852 0.894 1.102 33 ТапасеШт \т1§аге 1.90 0.9862 0.859 1.148
Средние для сообщества 2.17 0.9842 0.827 1.193 10 34 Сагех те1апо51аскуа 9 2.55 0.29 11.5 0.9810 0.839 1.170
35 РШрепМа и1тапа 9 2.25 0.24 10.6 0.9832 0.867 1.134
36 СаИит гиЫоШез 9 1.87 0.15 8.3 0.9861 0.843 1.170
37 8ап^ш50гЬа о//1ста1Ы 8 1.89 0.33 17.3 0.9860 0.922 1.069
Суммы и средние для сообщества 35 2.15 0.38 17.7 0.9841 0.868 1.136 11 38 Сагех т1рта 12 2.07 0.24 11,6 0.9847 0.875 1.126
39 аЫит тсапит 12 1.56 0.08 5.1 0.9885 0.865 1.143
40 СаНит гиЫоШез 12 1.67 0.14 8.1 0.9877 0.874 1.130
41 Уеготса 1оп§1/оИа 9 1.98 0.26 13.0 0.9853 0.885 1.113
42 У1с1а сгасса 12 1.75 0.33 18.7 0.9871 0.851 1.160
Суммы и средние для сообщества 57 1.79 0.29 16.3 0.9866 0.870 1.134
12 43 А1оресиги5 агипсИпасеш 36 1.61 0.32 19.6 0.9881 0.927 1.066
44 Сагех а{кегос1е8 39 2.15 0.47 21.8 0.9841 0.908 1.084
45 Сагех ШНорНИа 39 2.28 0.41 17.9 0.9832 0.842 1.168
46 Е1у1п§1а герепз 33 2.13 0.37 17.1 0.9843 0.859 1.146
47 1пз зШпса 39 2.03 0.31 15.4 0.9850 0.855 1.152
48 Рка1агоШ$ агипсИпасеа 39 1.75 0.37 21.3 0.9871 0.931 1.060
49 Ро1у§опит зЛтсит 39 1.27 0.40 31.3 0.9906 0.929 1.066
Суммы и средние для сообщества 264 1.89 0.51 26.8 0.9861 0.893 1.106
13 50 А^гозйз топ^НоНса 30 1.64 0.34 20.6 0.9880 0.885 1.116
51 Апетопе сИсНо1ота 30 2.15 0.29 13.4 0.9842 0.856 1.150
52 Сагех агНегос1е5 30 1.81 0.29 16.2 0.9867 0.884 1.116
53 Роа ап^511/оИа 30 1.68 0.38 22.9 0.9876 0.883 1.119
54 ТНаИс1гит !тр1ех 30 1.85 0.20 10.6 0.9864 0.839 1.176
55 УШа сгасса 30 1.52 0.29 19.0 0.9888 0.840 1.177
Суммы и средние для сообщества ш 1.77 0.36 20.3 0.9869 0.865 1.142
14 56 Сагех сеБрИоза 2.16 0.9840 0.766 1.285
57 РШрепМа иЫапа 2.43 0.9821 0.709 1.385
58 1пз рзеийасогиз 1.83 0.9865 0.766 1.288
59 8ап§ш5огЬа о^1с1паИз 1.93 0.9857 0.882 1.117
60 Уа1епапа м/о1§еп51з 1.59 0.9883 0.842 1.173
61 Уеготса 1оп§1/оИа 1.93 0.9858 0.818 1.205
Средние для сообщества 1.99 0.9854 0.797 1.242
8*
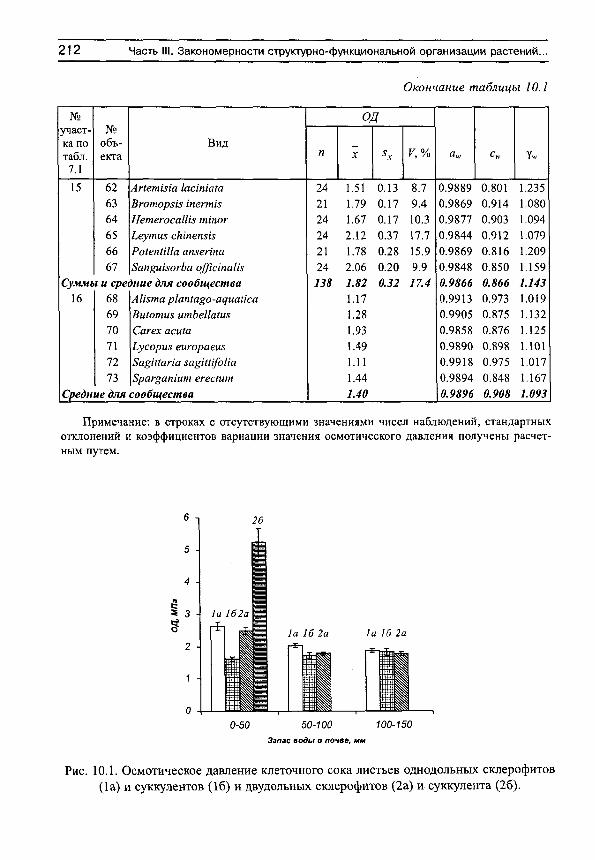
212 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й . .
Окончание таблицы 10.1
№ участка по табл.
7.1
№ объекта
В и д
ОД
а„ с„ У«
№ участка по табл.
7.1
№ объекта
В и д п X Г,% а„ с„ У«
15 62 Аг1етша 1асШа1а 24 1.51 0.13 8.7 0.9889 0.801 1.235
63 Вготорзгх тегтЫ 21 1.79 0.17 9.4 0.9869 0.914 1.080
64 НетегосаШз ттог 24 1.67 0.17 10.3 0.9877 0.903 1.094
65 Ьеутиз сМпепш 24 2.12 0.37 17.7 0.9844 0.912 1.079
66 Ро1еп1И1а апзегта 21 1.78 0.28 15.9 0.9869 0.816 1.209
67 8ап^13огЬа о^рютаИз 24 2.06 0.20 9.9 0.9848 0.850 1.159
Суммы и средние для сообщества 138 1.82 0.32 17.4 0.9866 0.866 1.143 16 68 АШта р1ап1а§о-адиаИса 1.17 0.9913 0.973 1.019
69 Вшотиз итЪеИаШз 1.28 0.9905 0.875 1.132
70 Сагех асШа 1.93 0.9858 0.876 1.125
71 Ьусориз еигораеиз 1.49 0.9890 0.898 1.101
72 8а^Шапа за^НИ/оИа 1.11 0.9918 0.975 1.017
73 8раг§апшпг егесШт 1.44 0.9894 0.848 1.167
Средние для сообщества 1.40 0.9896 0.908 1.093
Примечание: в строках с отсутствующими значениями чисел наблюдений , стандартных отклонений и коэффициентов вариации значения осмотического давления получены расчетным путем.
Р и с . 10.1. О с м о т и ч е с к о е д а в л е н и е к л е т о ч н о г о с о к а л и с т ь е в о д н о д о л ь н ы х с к л е р о ф и т о в
( 1 а ) и с у к к у л е н т о в (16) и д в у д о л ь н ы х с к л е р о ф и т о в ( 2 а ) и с у к к у л е н т а (26) .
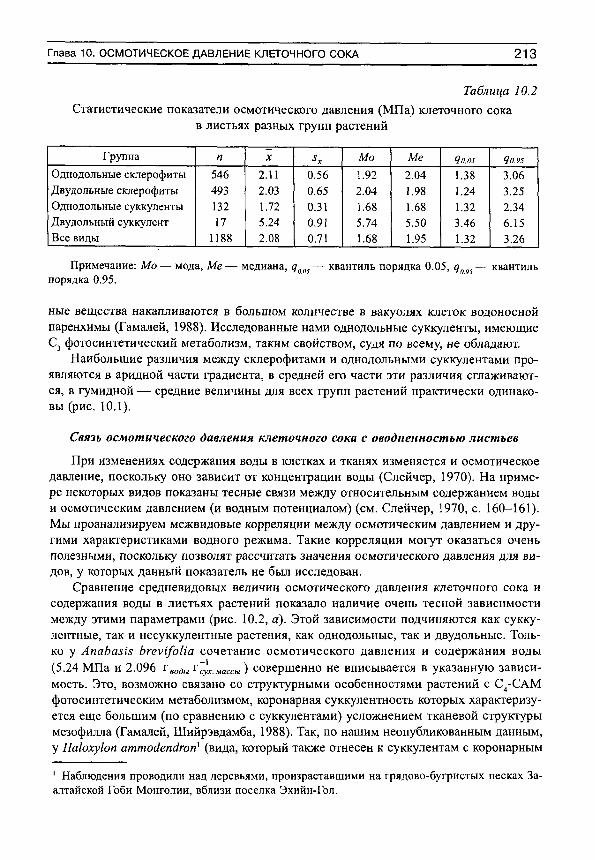
Глава 10. ОСМОТИЧЕСКОЕ ДАВЛЕНИЕ КЛЕТОЧНОГО СОКА 213
Таблица 10.2
Статистические показатели осмотического давления ( М П а ) клеточного сока в листьях р а з н ы х групп р а с т е н и й
Грзшпа п X Мо Ме Яо.о} Я 0.95 Однодольные склерофиты 546 2.11 0.56 1.92 2.04 1.38 3.06 Двудольные склерофиты 493 2.03 0.65 2.04 1.98 1.24 3.25 Однодольные суккуленты 132 1.72 0.31 1.68 1.68 1.32 2.34 Двудольный суккулент 17 5.24 0.91 5.74 5.50 3.46 6.15 В с е виды 1188 2.08 0.71 1.68 1.95 1.32 3.26
Примечание: Мо— мода, Ме— медиана, д^^^^— квантиль порядка 0.05, д^^^— квантиль порядка 0.95.
н ы е вещества накапливаются в большом количестве в вакуолях клеток в о д о н о с н о й паренхимы (Гамалей, 1988). Исследованные нами однодольные суккуленты, и м е ю щ и е Сз ф о т о с и н т е т и ч е с к и й метаболизм, таким свойством, судя п о всему, н е обладают.
Н а и б о л ь ш и е различия м е ж д у с к л е р о ф и т а м и и о д н о д о л ь н ы м и суккулентами проявляются в аридной части градиента , в средней его части эти р а з л и ч и я сглаживаются, в гумидной — средние в е л и ч и н ы для всех групп р а с т е н и й п р а к т и ч е с к и одинаковы (рис, 10.1).
Связь осмотического давления клеточного сока с оводненностью листьев
П р и изменениях содержания воды в клетках и тканях изменяется и осмотическое давление , поскольку оно зависит от концентрации воды (Слейчер , 1970). Н а п р и м е ре некоторых видов показаны тесные связи м е ж д у относительным содержанием воды и осмотическим давлением (и водным потенциалом) (см. С л е й ч е р , 1970, с. 160-161). М ы проанализируем м е ж в и д о в ы е корреляции м е ж д у о с м о т и ч е с к и м давлением и другими х а р а к т е р и с т и к а м и водного р е ж и м а . Такие к о р р е л я ц и и м о г у т оказаться очень полезными, поскольку позволят рассчитать значения осмотического давления для видов, у которых д а н н ы й показатель не был исследован .
С р а в н е н и е с р е д н е в и д о в ы х величин о с м о т и ч е с к о г о д а в л е н и я клеточного сока и содержания воды в л и с т ь я х растений показало н а л и ч и е очень т е с н о й з а в и с и м о с т и между этими п а р а м е т р а м и (рис . 10.2, а). Этой з а в и с и м о с т и п о д ч и н я ю т с я как сукку-лентные , т а к и несуккулентные растения , как однодольные , т а к и двудольные . Только у АпаЬа$1з ЬгеУ1/оИа с о ч е т а н и е о с м о т и ч е с к о г о д а в л е н и я и с о д е р ж а н и я в о д ы (5.24 М П а и 2.096 г^^йы ^сух.массы) с о в е р ш е н н о не вписывается в у к а з а н н у ю зависимость . Это , возможно связано со с т р у к т у р н ы м и о с о б е н н о с т я м и р а с т е н и й с С^-САМ фотосинтетическим метаболизмом, коронарная суккулентность которых характеризуется еще б о л ь ш и м (по с р а в н е н и ю с суккулентами) у с л о ж н е н и е м тканевой структуры мезофилла (Гамалей, Шийрэвдамба , 1988). Так, по н а ш и м неопубликованным данным, у На1оху1оп атто(1епс1гоп^ (вида, который также отнесен к суккулентам с коронарным
' Наблюдения проводили над деревьями, произраставшими на грядово-бугристых песках За-алтайской Гоби Монголии, вблизи поселка Эхийн-Гол.

214 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й .
2 •
г' =0.687
2 3
СВ, г/г
г' =0.175
10 20 ДВН, У.
30
Р и с . 10.2. З а в и с и м о с т ь о с м о т и ч е с к о г о давления клеточного сока от содержания воды (а) и д е ф и ц и т а водного н а с ы щ е н и я
в листьях растений (б).
с и н д р о м о м : Г а м а л е й , Ш и й р э в д а м б а , 1988) с о ч е т а н и е о с м о т и ч е с к о г о давле н и я и с о д е р ж а н и я в о д ы (4.18 М П а и 3.342 Г д о ^ у ^'^х.массы п р и соответствую-щих числах наблюдений 36 и 276) сходно с п о д о б н ы м с о ч е т а н и е м у АпаЬазгз ЪгеуфИа. Это сочетание характеризуется п о в ы ш е н н о й к о н ц е н т р а ц и е й в о д ы при очень в ы с о к о м о с м о т и ч е с к о м д а в лении клеточного сока. Если бы АпаЬа-з1в Ьгеу1/оНа и На1оху1оп атто(1еп(}гоп п о д ч и н я л и с ь о п и с а н н о й з а в и с и м о с т и (рис. 10.2, а), то осмотическое давление в и х а с с и м и л и р у ю щ и х о р г а н а х ( п р и у к а з а н н ы х в ы ш е значениях оводненности) бьшо бы примерно в 2.5 раза меньше .
К р о м е связи м е ж д у о с м о т и ч е с к и м д а в л е н и е м и с о д е р ж а н и е м воды в л и с тьях о б н а р у ж е н а т а к ж е корреляция ос мотического д а в л е н и я клеточного сока с д е ф и ц и т о м водного н а с ы щ е н и я . Э т а корреляция в ы р а ж е н а слабо (рис . 10.2, б ) , о д н а к о с т а т и с т и ч е с к и д о с т о в е р н а . М н о ж е с т в е н н ы й р е г р е с с и о н н ы й анализ п о к а з а л , ч т о э т а п е р е м е н н а я м о ж е т быть сохранена в у р а в н е н и и регрессии:
где
2 , = / + ^ 1 п ( Х , + Л )
^2 =к + 1\п{Х2+т)
у — осмотическое давление клеточного сока, М П а ; X , — содержание воды в листьях растений, ^еоды ^~с1х.массы \ — Дсфицит ВОДНОГО н а с ы щ е н и я , % ; а (1.248), Ь (0.332), с (-0.734), с1 (0.231), е (0.724), / (-2.037), § (2.785), Н (-9.969), к (-5.816), / (2.333), т (-1.601) — к о э ф ф и ц и е н т ы . К о э ф ф и ц и е н т множественной д е т е р м и н а ц и и = 0.669. Фактический Р-критерий равен 42.4, частные Р-критерии с о с т а в л я ю т 64.8 для содержани я воды и 6.4 для д е ф и ц и т а водного н а с ы щ е н и я . А н а л и з остатков показал д о с таточно высокую д л я такого изменчивого материала точность м о д е л и : средняя относительная о ш и б к а а п п р о к с и м а ц и и составила 9.5%, средняя к в а д р а т и ч н а я — 11.7%, коэффициент н е с о в п а д е н и я Тейла (см.: Гильманов , 1978 )— 6.0%. Г р а ф и ч е с к и й анализ остатков (который м ы здесь не приводим в силу его громоздкости) свидетельствует о хорошем качестве аппроксимации исходных д а н н ы х м о д е л ь ю .

Глава 10. О С М О Т И Ч Е С К О Е Д А В Л Е Н И Е К Л Е Т О Ч Н О Г О С О К А 215
М ы использовали эту модель д л я расчета н е д о с т а ю щ и х у р я д а в и д о в растений значений осмотического давления по известным в е л и ч и н а м с о д е р ж а н и я воды и дефицита водного насыщения (табл. 10.1; в строках с расчетными значениями отсутствуют данные по числу наблюдений и варьированию признака) . В д а л ь н е й ш е м в анализ включены как р е а л ь н ы е , т а к и расчетные величины осмотического давления (на рис . 10.3 они п о м е ч е н ы ч е р н ы м и точками) .
Динамика осмотического давления клеточного сока
М н о ж е с т в е н н ы й р е г р е с с и о н н ы й анализ показал , что о с н о в н ы м факто ром среды, в о з д е й с т в у ю щ и м на о с м о т и ч е с к о е д а в л е н и е к л е т о ч н о г о сока в листьях растений , является в л а ж н о с т ь п о ч в ы . Р а с с м о т р и м , к а к и з м е н я ю т с я различные средние в е л и ч и н ы на градиенте в л а ж н о с т и п о ч в ы . С этой ц е л ь ю ц е л е с о о б р а з н о п р о в е с т и а н а л и з 1) средних уровней (средних для каждого сообщест ва значений) , 2) средневидов ы х значений всего массива данных .
Д и н а м и к а с р е д н и х у р о в н е й осмот и ч е с к о г о д а в л е н и я н а г р а д и е н т е в л а ж н о с т и п о ч в ы . С р е д н и е у р о в н и осмотического давления тесно связаны с в л а ж н о с т ь ю почвы (рис . 10.3, а; коэффициент детерминации для совокупн о с т и ф а к т и ч е с к и х и р а с ч е т н ы х д а н ных равен 0.875, только д л я фактичес ких — = 0.735). Зависимость средних д л я экот оп ов в е л и ч и н о с м о т и ч е с к о г о д а в л е н и я от в л а ж н о с т и п о ч в ы м о ж н о описать п р о с т ы м у р а в н е н и е м :
У = а-х'',
где у — осмотическое давление , М П а , X — запас в о д ы в в е р х н е м (0 -30 см) слое почвы, мм; а (4.823), Ь (-0.206) — коэффициенты. С п о м о щ ь ю этого урав нения легко и достаточно точно можно о п р е д е л я т ь в е л и ч и н ы о с м о т и ч е с к о г о давления , характ ерн ые д л я р а з н ы х условий почвенного увлажнения .
Из ан али за л и н и и р е г р е с с и и (рис . 10.3, а) видно , что на б о л ь ш е м протя-
а
I 1 О I V
Т Г
О 5 -
\ 1\
50 100 150
Запас воды в почве, ми
Рис. 10.3. Динамика осмотического давления на градиенте в л а ж н о с т и почвы:
а) средних уровней, б) средневидовых значений и трендов минимальных (нижняя линия) и максимальных (верхняя линия) для каждого участка средневидовых величин. Белыми символами показаны фактические данные (сплошные линии р е г р е с с и и ) , ч е р н ы м и — р а с ч е т н ы е значения. Прерывистые линии регрессии даны для совок у п н о с т и ф а к т и ч е с к и х и р а с ч е т н ы х величин
осмотического давления.
4.0 -1
3.5 -
5 3.0 •
^ 2 . 5 .
2.0 -

216 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й .
жении градиента в л а ж н о с т и почвы (в его гумидно й и средней частях ) о с м о т и ч е с к о е давление изменяется н е з н а ч и т е л ь н о — от 1.73 М П а (в точке с з а п а с а м и воды в почве 144 м м ) д о 2.15 М П а (в т о ч к е с з а п а с а м и в о д ы 50 м м ) . С к о р о с т ь и з м е н е н и й о с м о т и ч е с к о г о д а в л е н и я н а э т о м п р о м е ж у т к е г р а д и е н т а с о с т а в л я е т п р и м е р н о 0.004 М П а • м м ' . В аридной части градиента эта скорость у в е л и ч и в а е т с я более ч е м в 7 раз и равна п р и м е р н о 0.032 М П а • м м '. Таким образом, в точке г р а д и е н т а с зап а с а м и в о д ы 4 м м д о с т и г а е т с я з н а ч и т е л ь н а я в е л и ч и н а о с м о т и ч е с к о г о д а в л е н и я 3.6 МП а . Отсюда видно , что скорость реакции растений на изменения условий увлажнения экотопа значительно в ы ш е в аридной части градиента п о с р а в н е н и ю с более обесп еч ен н ыми водой его частями.
Д и н а м и к а с р е д н е в и д о в ы х з н а ч е н и й о с м о т и ч е с к о г о д а в л е н и я н а г р а д и е н т е в л а ж н о с т и п о ч в ы . Ш и р и н а зоны р а з б р о с а с р е д н е в и д о в ы х значений осмотического давления в средней и гумидной частях градиента влажности почвы изменяется незначительно . Это в и д н о из анализа трендов м а к с и м а л ь н ы х и м и н и м а л ь н ы х для к а ж д о го участка с р е д н е в и д о в ы х в е л и ч и н этого показателя водного р е ж и м а (рис . 10.3, б) . Разность м е ж д у у к а з а н н ы м и т р е н д а м и изменяется (против г р а д и е н т а в л а ж н о с т и по чвы) от 0.75 д о 0.93 М П а . В с а м о й сухой точке градиента эта р аз но ст ь составляет 2.15 М П а , т.е. увеличивается более чем в 2 раза. Отсюда видно , что разнообразие ви-доспецифических реакций осмотического давления клеточного сока на изменения ус ловий почвенного в о д о с н а б ж е н и я определенно в ы ш е в а р и д н о й части г р а д и е н т а по сравнению со средней и гумидной его частями.
Это наблюдалось , н е с м о т р я на то, что в аридной части г р а д и е н т а б ы л и исследованы растения только трех экологических групп: м е з о к с е р о ф и т ы , к с е р о ф и т ы и суккуленты. В средней части градиента в л а ж н о с т и почвы и з у ч а л и с ь пр едст авит ели мезофитов, ксеромезофитов , мезоксерофитов , ксеро фит о в и суккулентов , а в г у м и д н о й его части, кроме этих групп также и в и д ы гидрофитов и г и г р о ф и т о в (рис . 7.10). Видно, что растения ксерофитной организации в аридной части градиента демонстрируют более ш и р о к и й спектр в и д о с п е ц и ф и ч е с к и х р е а к ц и й о с м о т и ч е с к о г о д а в л е н и я на изменения у в л а ж н е н н о с т и экотопов, чем виды м е з о ф и т н о й о р г а н и з а ц и и в более влажных м е с т о о б и т а н и я х . У некоторых видов в а р и д н о й ч а с т и г р а д и е н т а {АШит ро1у-гЫгит, Аг1ет1з1а тагзскаШапа, Са1ата§го$из ерщеюз) о смотическое давление может оставаться на оч ень низком у р о в н е , с р а в н и м о м с т а к о в ы м у р а с т е н и й г у м и д н ы х экотопов. У других видов в засушливых условиях произрастания осмотическое давление может п о в ы ш а т ь с я до 3—4.5 М П а и более (рис . 10.3, б) .
Такие различия могут быть связаны с неодинаковым состоянием воды в растениях (ее концентрации и активности , см . раздел « Т е р м о д и н а м и ч е с к и й аспект водного ре жима») и, как следствие этого, неодинаковым значением о с м о т и ч е с к о й регуляции водообмена . У р а с т е н и й с низким о с м о т и ч е с к и м д а в л е н и е м такая р е г у л я ц и я , как можно предположить , и грает менее значительную роль в о б щ е й р ег у ля ции водообмена , чем у растений с в ы с о к и м о с м о т и ч е с к и м д а в л е н и е м . Р а с т е н и я с н и з к и м о смо т иче ским давлением в а р и д н ы х экотопах и с п о л ь з у ю т д л я а к т и в н о й ж и з н е д е я т е л ь н о с т и наиболее влажные периоды и концентрация воды в них может поддерживаться на до статочно высоком у р о в н е . Так , с р е д н и е в е л и ч и н ы о в о д н е н н о с т и л и с т ь е в у АШит ро1угЫ2ит, АПетша тагзскаШапа, Са1ата§го5Ш ерщеюз с о с т а в л я л и соответственно 4.510, 2.367 и 1.627 г,^^„ ̂ ~с^х.массы • Это высокие показатели для д а н н ы х местооби-

Глава 10. О С М О Т И Ч Е С К О Е Д А В Л Е Н И Е К Л Е Т О Ч Н О Г О С О К А 217
таний. Другими словами, значение осмотической регуляции водообмена растений долж н о усиливаться п о мере снижения концентрации воды в тканях растений .
С д р у г о й с т о р о н ы , с у к к у л е н т ы с к о р о н а р н ы м с и н д р о м о м ( такие , как АпаЬазгв Ь^еV^/Ы^а и На1оху1оп атто(1епс1гоп) имеют, в и д и м о , с о в е р ш е н н о н е о б ы ч н о е сочетание высокой оводненности тканей и высокого осмотического давления . К такому соч е т а н и ю может привести не только тот или иной с п о с о б ф у н к ц и о н а л ь н о й регуляции водного режима , но и определенное усложнение анатомической структуры ассимилир у ю щ и х органов (Гамалей , 1988; Гамалей , Ш и й р э в д а м б а , 1988).
Динамика показателей вариации осмотического давления клеточного сока в листьях
С р е д н е в и д о в ы е к о э ф ф и ц и е н т ы вариации , при п о м о щ и которых м ы оцен и в а л и п о д в и ж н о с т ь о с м о т и ч е с к о г о давления клеточного сока в листьях растений, и м е ю т л и ш ь очень слабую тенд е н ц и ю к у в е л и ч е н и ю против градиента влажности почвы (рис. 10.4, а), котор а я не п о д т в е р ж д а е т с я с т а т и с т и ч е с к и (связь коэффициентов вариации с другими факторами среды также отсутствует ) . П о э т о м у м о ж н о с к а з а т ь , ч т о л а бильность осмотического давления остается в среднем практически неизменн о й н а в с е м п р о т я ж е н и и г р а д и е н т а влажности почвы. З о н а р а з б р о с а коэффициентов вариации, которая ограничивается трендами м и н и м а л ь н ы х и максимальных для каждого участка средневид о в ы х к о э ф ф и ц и е н т о в , т а к ж е п р а к т и чески не и з м е н я е т с в о е й ш и р и н ы при изменении запасов в о д ы в почве (рис . 10.4, б ) . Т а к и м о б р а з о м , л а б и л ь н о с т ь о с м о т и ч е с к о г о д а в л е н и я к л е т о ч н о г о сока в листьях о д н о д о л ь н ы х и двудольных растений в статистическом отношении практически не з а в и с и т от изменений у с л о в и й увлажнения экотопов.
Межвидовые структурно-функциональные корреляции
Р е з у л ь т а т ы , п р и в е д е н н ы е в э т о м разделе, являются в значительной степени г и п о т е т и ч н ы м и , п о с к о л ь к у в н е м
40 л
Запас воды в лочвв, ии
Рис. 10.4. Динамика коэффициентов вариац и и о с м о т и ч е с к о г о д а в л е н и я к л е т о ч н о г о с о к а н а г р а д и е н т е в л а ж н о с т и п о ч в ы : а) с р е д н е в и д о в ы е к о э ф ф и ц и е н т ы в а р и а ц и и ; б) т р е н д ы м и н и м а л ь н ы х ( н и ж н я я линия) и м а к с и м а л ь н ы х (верхняя л и н и я ) для каждого участка с р е д н е в и д о в ы х к о э ф ф и
циентов вариации .

218 Ч а с т ь III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й . . .
сравниваются р е а л ь н ы е д а н н ы е п о анатомической структуре л и с т ь е в с р а с ч е т н ы м и з н а ч е н и я м и о с м о т и ч е с к о г о д а в л е н и я . Тем не м е н е е , н а л и ч и е д о с т а т о ч н о т е с н ы х , логически о б о с н о в а н н ы х м е ж в и д о в ы х с т р у к т у р н о - ф у н к ц и о н а л ь н ы х корреляций позволяет п р е д п о л о ж и т ь , что с у щ е с т в у ю т н е к о т о р ы е з а к о н о м е р н о с т и в и з м е н е н и я х осмотического давления клеточного сока и ряда структурных характеристик листьев . Отметим, что эти закономерности в большинстве случаев не выявляются на всем массиве д а н н ы х , но только при его разделении на г р у п п ы о д н о д о л ь н ы х и двудольных растений.
Осмотическое д а в л е н и е клеточного сока и п а р ц и а л ь н ы е о б ъ е м ы проводящих тканей . В ы я в л е н ы достаточно т е с н ы е статистические связи м е ж д у п а р ц и а л ь н ы м и объемами проводящих тканей (без учета паренхимной обкладки пучков) и осмотическим давлением клеточного сока у однодольных (рис. 10.5, а) и двудольных (рис . 10.5, г) растений . У видов с наиболее высоким п а р ц и а л ь н ы м о бъемо м п р о в о д я щ е й ткани (среди о д н о д о л ь н ы х это Сагех сезрИоза, Сагех со1сЫса, Сагех те1апоз1асИуа, 8Нра Ьогуз1кетса, с р е д и д в у д о л ь н ы х — Аг1ет131а аш1паса, Тп/оНит агуепзе, СаИит гШНетсит) наблюдаются и более высокие величины осмотического давления . Для видов со слабо р а з в и т ы м и пучками (среди однодольных это АНзта р1ап{а§о-адиаИса, 8а§Шагга 8аё1П1ЙИа, Зращатит егесШт, среди д в у д о л ь н ы х — Ьусориз еигораеиз, 1у(кгит V^^§а^ит, 8ап§шзогЬа о^стаИз, Уакпапа •^о1§епз1з) х арактерно низкое осмотическое давление .
Эти связи м о г у т б ы т ь обусловлены наличием значительного количества осмотически а к т и в н ы х в е щ е с т в во флоэме и ксилеме п р о в о д я щ и х пучков . В о ф л о э м е , куда загружаются раст воримые продукты ассимиляции, д о л ж н о создаваться высокое осмотическое давление . В ксилеме , я в л я ю щ е й с я проводником в о д ы и р а с т в о р е н н ы х в ней мин еральн ых в е щ е с т в , т а к ж е может поддерживаться до ст ат о чно высокое о смо т ичес кое давление , хотя, в о з м о ж н о , и не в такой степени , как во ф л о э м е . В целом проводящие пучки представляются естественными о с м о т и ч е с к и м и к о м п а р т м е н т а м и листа. Поэтому м о ж н о предположить , что описанные в ы ш е статистические связи (рис . 10.5, а, г) о т р а ж а ю т р е а л ь н ы е корреляции м е ж д у в е л и ч и н а м и о с м о т и ч е с к о г о давления и п а р ц и а л ь н ы м и о б ъ е м а м и проводящих тканей.
О с м о т и ч е с к о е д а в л е н и е клеточного сока и п а р ц и а л ь н ы е о б ъ е м ы губчатой п а р е н х и м ы и п а р е н х и м н о й о б к л а д к и пучков. Д л я о д н о д о л ь н ы х р а с т е н и й выявлена т е с н а я ст ат и ст ическая связь (рис . 10.5, б) п а р ц и а л ь н ы х о б ъ е м о в губчатой паренх и м ы с о с м о т и ч е с к и м давлением клеточного сока. У видов о д н о д о л ь н ы х с наиболее развитой губчатой п а р е н х и м о й могут наблюдаться и наиболее в ы с о к и е в е л и ч и н ы осмотического давления .
В отличие от однодольных, для двудольных р а с т е н и й х а р а к т е р н а статистическая связь (рис. 10.5, д -1) м е ж д у п а р ц и а л ь н ы м и объемами п а р е н х и м н ы х о б к л а д о к пучков и этим показателем водного режима. С учетом обсуждавшейся в ы ш е связи м е ж д у осмотическим давлением и парциальным объемом проводящей ткани (рис . 10.5, г), можно сказать, что весь пучок (включая флоэму, ксилему и п а р е н х и м н у ю обкладку) двудольных р а с т е н и й является зоной высокого осмо т ическо г о д а в л е н и я (рис . 10.5, д-1). Кроме того, д л я этой группы видов не обнаружено статистических связей осмотического давления с д р у г и м и т к а н я м и мезофилла (в част но ст и , с х л о р е н х и м о й ) . М о ж н о поэтому п р е д п о л о ж и т ь , что у двудольных р а с т е н и й р а с т в о р и м ы е (т. е. о с м о т и ч е с к и
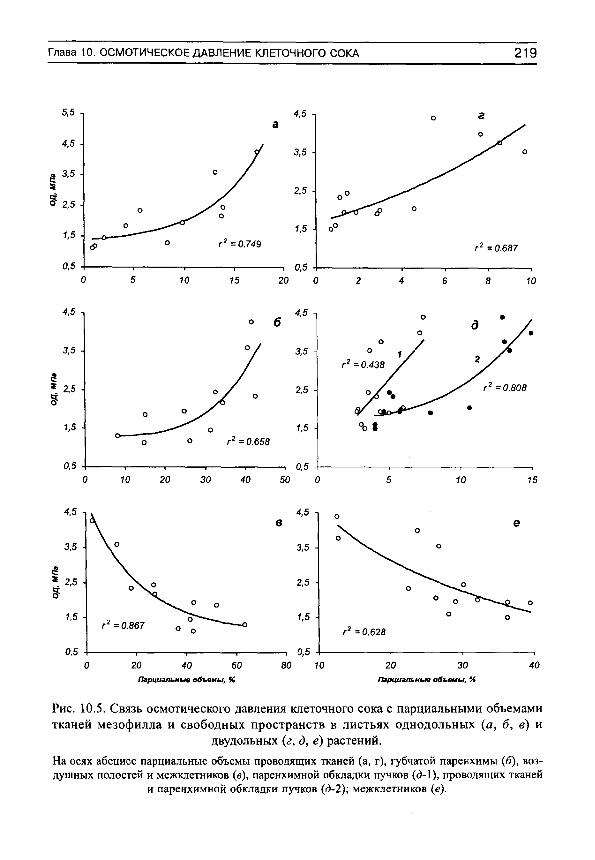
Глава 10 . О С М О Т И Ч Е С К О Е Д А В Л Е Н И Е К Л Е Т О Ч Н О Г О С О К А 219
5,5 -,
4,5 -
3,5 -
2,5 •
1,5
0,5
4,5 -I
3,5 •
2,5
,̂5 г' =0.749
10 15 —, 0,5
20 О
г' =0.687
8 10
4,5
3,5
5
1,5
0,5
о б 4,5 -,
2,5 -
-, 0,5 10 20 30 40 50 О 10 15
20 40 60 Парциальные объемы, %
г' = 0.628
—, 0,5 80 10 20 30 40
Парциальные объемы, %
Рис. 10.5. Связь осмотического давления клеточного сока с п а р ц и а л ь н ы м и объемами т к а н е й м е з о ф и л л а и с в о б о д н ы х п р о с т р а н с т в в л и с т ь я х о д н о д о л ь н ы х ( а , б, в) и
двудольных (г, д, ё) растений .
На осях абсцисс парциальные объемы проводящих тканей (а, г), губчатой паренхимы (б) , воздушных полостей и межклетников (в), паренхимной обкладки пучков (Э-1), проводящих тканей
и паренхимной обкладки пучков {д-2); межклетников (е).

220 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й .
активные) вещест ва л о к а л и з у ю т с я п р е и м у щ е с т в е н н о в п р о в о д я щ и х пучках и их обкладках. Это м о ж е т свидетельствовать о том, что количество образовавшихся в х л о -ренхиме ассимилятов примерно равно количеству ассимилятов, загружаемому во флоэму пучков.
О с м о т и ч е с к о е д а в л е н и е клеточного сока и п л о т н о с т ь с л о ж е н и я мезофилла . Как для однодольных, так и для двудольных растений выявлены статистические связи осмотического д а в л е н и я клеточного сока с плотно ст ь ю с л о ж е н и я м е з о ф и л л а . М е р о й плотности сложения м е з о ф и л л а у однодольных служит с у м м а п а р ц и а л ь н ы х объемов межклетников и воздушных полостей, у двудольных — парциальные объемы межклетников. Ч е м б о л ь ш е с в о б о д н ы х п р о с т р а н с т в в л и с т ь я х р а с т е н и й , т е м м е н е е плотно сложен м е з о ф и л л , и наоборот . П л о т н о с т ь м е з о ф и л л а о т р ажает у с л о в и я почвенного водоснабжения и соответственно расположение вида на градиенте в л а ж н о с т и почвы. Видимо поэтому в листьях однодольных (рис. 10.5, в) и двудольных (рис . 10.5, е) растений с наиболее п л о т н ы м сложением мезофилла наблюдаются максимальные велич и н ы осмотического д а в л е н и я клеточного сока, а в л и с т ь я х р а с т е н и й с р ы х л ы м мезофиллом — м и н и м а л ь н ы е .
Термодинамический аспект водного режима растений
О смот и ч еское д а в л е н и е является в а ж н е й ш и м компонентом водного потенциала растения:
Ч ' = / ' - 7 Г - Х ,
где \)/ — водный потенциал , Р — гидростатическое (тургорное) д а в л е н и е , л — осмотическое давление , т — м а т р и ч н о е давление . В и д н о , что п о в ы ш е н и е осмотического давления способствует п о н и ж е н и ю водного потенциала .
О смот и ч еское д а в л е н и е м о ж н о выразить через активность воды:
КТ п = - ^ \ п а „ , (1)
где К — у н и в е р с а л ь н а я газовая п о с т о я н н а я , Т — т е м п е р а т у р а п о ш к а л е Кельвина , у„ — п а р ц и а л ь н ы й м о л я р н ы й о б ъ е м воды, — активность воды.
А к т и в н о с т ь в о д ы отражает к о н ц е н т р а ц и ю воды в растворе . П р и с у т с т в и е растворенных вещест в в водном растворе у м е н ь ш а е т активность воды. К о н ц е н т р а ц и я воды становится м е н ь ш е в результате з а м е щ е н и я молекул воды м о л е к у л а м и растворенного вещества . Т а к и м образом , активность воды и о с м о т и ч е с к о е д а в л е н и е изменяются в п р о т и в о п о л о ж н ы х н а п р а в л е н и я х . А к т и в н о с т ь чистой в о д ы р а в н а е д и н и ц е , хотя в общем случае а„ р а в н а у„М„, где у„ — к о э ф ф и ц и е н т а к т и в н о с т и воды, Л^̂ ,, — ее мольная доля (Нобел , 1973).
Поскольку мольная доля воды в растворе влияет на у п р у г о с т ь водяного пара над раствором о с м о т и ч е с к о е давление можно выразить через с о о т н о ш е н и е :
КТ. р

Глава 10. О С М О Т И Ч Е С К О Е Д А В Л Е Н И Е К Л Е Т О Ч Н О Г О С О К А 221
где р — парциальное давление водяного пара над раствором, р^ — давление пара чистой воды (Слейчер , 1970; Нобел , 1973; Зялалов , 1984 и др.) .
В данном контексте нас интересует определение активности воды. Это достаточно просто сделать, используя формулу (1). Однако в связи с тем, что существует мнение о непостоянстве п а р ц и а л ь н о г о молярного объема воды (см. а р г у м е н т а ц и ю и обзор этого вопроса в работе А. А. Зялалова, 1984)' д л я определения активности воды было использовано с о о т н о ш е н и е :
Ро
Здесь м о ж н о найти из таблиц (см. , например , Гороновский и др . , 1974, с. 775). Однако значительно п р о щ е сделать это, используя у р а в н е н и е (в инт ер вале температур от О до 50 °С) :
Г 2 V _ а + сх + ех
^~ 1 + Ьх + с{х^ '
где у — давление пара над чистой водой ( р^), Па, х — температура , °С, а (24.716632), Ь (-0.007965), с (0.701536), с1 (0.000031), е (0.006253) — к о э ф ф и ц и е н т ы .
П а р ц и а л ь н о е д а в л е н и е в о д я н о г о п а р а н а д р а с т в о р о м м о ж н о н а й т и , испо ль з у я уравнение (248) из р а б о т ы Н. Э д л е ф с е н а и А. А н д е р с о н а (1966):
1 . 0 0 2 Г П - 2 - 1 0 ' ^ П ^ ; Р = Ро- 10
4.5487-1-26.3
Г 373^ 3 -1- 1-26.3
Ро 1 т)
где р — парциальное давление водяного пара над раствором, атм. (для перевода в Па полученное значение н у ж н о умножить на 101325), П — осмотическое давление , атм, Т— температура по шкале Кельвина .
Теперь у нас есть все д а н н ы е д л я определения а к т и в н о с т и воды ( см . табл . 10.1). Однако значительный интерес м о ж е т представлять также о п р е д е л е н и е ко э ффициента активности воды. К о э ф ф и ц и е н т активности можно рассматривать как поправочный фактор, оц ен и ваю щи й отклонения от идеального поведения , поскольку термодинамическая акт и вн ост ь в е щ е с т в а м е н ь ш е его к о н ц е н т р а ц и и ( Н о б е л , 1973). А к т и в н о с т ь
' В своей монографии Р. МоЬе! (1991), тем не менее, пишет, что на величину парциального молярного объема воды хотя и могут влиять различные растворенные вещества, для разбавленных растворов она близка к 18.0x10' м' моль ' (18.0 см' моль ' ) . Эту величину он и использовал для расчетов в своей книге. Отметим также, что если пересчитать данные столбца
в таблице 10.1 согласно формуле (1) пУ „ = - Д 7 1 п а ^ , и затем полученные величины поделить на значения осмотического давления из этой таблицы, то во всех строках обнаружится искомая величина (после приведения к соответствующей размерности) 18.2-18.4 см' м о л ь ' .

222 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й .
воды с п о м о щ ь ю к о э ф ф и ц и е н т а активности с о о т н о с и т с я с ее к о н ц е н т р а ц и е й (с„): и̂» - У»>'^№ (Нобел , 1973). А. А. Зялалов (1984) пр едло жил за е д и н и ч н у ю концентра
цию воды в р а с т и т е л ь н ы х объектах принять их оводненность при полном н а с ы щ е н и и и п о с л е д у ю щ и е значения о в о д н е н н о с т и с о о т н о с и т ь с исходной . Тогда, по м н е н и ю этого автора, концентрация будет равна :
^ _ Д о л я воды при созданной (текущей) активности
^ Доля воды при полном н а с ы щ е н и и
Это о п р е д е л е н и е представляет собой ни что иное , как определение относитель ного с о д е р ж а н и я воды. Таким образом, к о н ц е н т р а ц и ю воды м ы м о ж е м выразить через известный показатель (относительное содержание воды в ы р а ж е н н о е не в процент а х , а в в и д е б е з р а з м е р н о й в е л и ч и н ы ) и о п р е д е л и т ь к о э ф ф и ц и е н т а к т и в н о с т и у„ =а^/с„ : (результаты вычислений см. в табл. 10.1).
Рассмотрим взаимосвязь этих показателей — активности , концентрации и коэфф и ц и е н т а а к т и в н о с т и в о д ы (рис . 10.6). К о н ц е н т р а ц и я и а к т и в н о с т ь в о д ы с в я з а н ы простой л и н е й н о й з а в и с и м о с т ь ю : согласно о п р е д е л е н и ю у в е л и ч е н и е ко нцент р ации воды с о п р о в о ж д а е т с я у в е л и ч е н и е м ее активности (рис . 10.6, а). АпаЬазгз Ьгеу1/оНа не подчиняется этой з ависимости — при достаточно высокой ко нцент р ации воды в побегах этого р а с т е н и я ее активность очень низка (рис . 10.6, а). М о ж н о предполож и т ь , что с у к к у л е н т ы с к о р о н а р н ы м с и н д р о м о м о б р а з у ю т д р у г у ю п о с л е д о в а т е л ь н о с т ь с б о л е е н и з к и м у р о в н е м а к т и в н о с т и воды, что я в л я е т с я , судя п о всему, их специфической чертой .
Концентрация и коэффициент активности воды связаны по о п р е д е л е н и ю функциональной обратно п р о п о р ц и о н а л ь н о й з а в и с и м о с т ь ю (рис . 10.6, б).
Взаимосвязь коэффициента активности и активности воды (рис . 10.6, в) достаточно удовлетворительно описывается полиномом третьей степени . В и д н о , что при величинах а к т и в н о с т и воды б о л ь ш е 0.98 коэффициент а к т и в н о с т и м а л о отличается от единицы. Если активность воды снижается ниже указанного порога , то происходит быстрое увеличение коэффициента активности воды. Такое увеличение коэффициента активности является результатом более медленного падения активности по сравнению с концентрацией и свидетельствует о том, что п о мер е о б е з в о ж и в а н и я о с л а б е в а ю т взаимодействия , п о н и ж а ю щ и е химический потенциал воды (Зялалов , 1984). Возможно поэтому в у с л о в и я х сильной почвенной засухи растениям э н е р г е т и ч е с к и выгодно быстрое ф о р м и р о в а н и е (в о п р е д е л е н н ы х пределах , не в ы х о д я щ и х за р а м к и продолжения жизнедеятельности) д е ф и ц и т а водного н а с ы щ е н и я , о п е р е ж а ю щ е е у м е н ь ш е н и е активности в о д ы . С другой с т о р о н ы , резкое у в е л и ч е н и е к о э ф ф и ц и е н т а а к т и в н о с т и воды может с л у ж и т ь индикатором крайне неблагоприятных у с л о в и й пр о из р аст ания . Однако такой вид , как АпаЬазЬ ЬгеУ1/оИа и в этом случае не в п и с ы в а е т с я в указанную зависимость (рис . 10.6 в).
Изменения активности, концентрации и коэффициента активности воды н а градиенте влажности почвы (рис . 10.7) характеризуются т е м , что на б о л ь ш е й части градиента эти в е л и ч и н ы (с т о й или иной ш и р и н о й зоны р а з б р о с а значений) практически параллельны оси абсцисс, т. е. не зависят от условий почвенного водоснабжения. Толь-

Глава 10. ОСМОТИЧЕСКОЕ ДАВЛЕНИЕ КЛЕТОЧНОГО СОКА 223
0,960 0,970 0,980 Активность воды
0,990 50 100
Запас воды в почве, ии
Рис. 10.6. Взаимосвязи активности, концентрации и коэффициента активности
воды.
Ч е р н ы м и с и м в о л а м и п о м е ч е н ы з н а ч е н и я для АпаЬазгз Ьгеух/оНа.
Р и с . 10.7. Д и н а м и к а а к т и в н о с т и , концентрации и к о э ф ф и ц и е н т а активности в о д ы на г р а д и е н т е в л а ж н о с т и п о ч в ы .
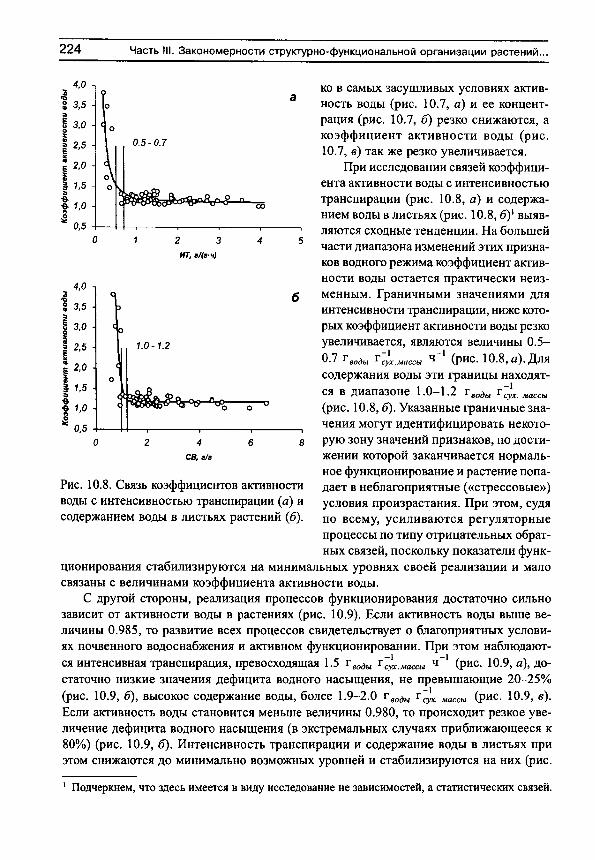
224 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й . .
4,0
3,5
3,0
2,5
2,0
1,5
1,0
0,5
0.5-0.7
-00
ИТ, 1/(гч)
1.0-1.2
КО В самых засушливых условиях активность воды (рис . 10.7, а ) и ее концентрация (рис. 10.7, б) резко снижаются , а к о э ф ф и ц и е н т а к т и в н о с т и в о д ы ( р и с . 10.7, в) так же резко увеличивается .
При исследовании связей коэффициента активности воды с и н т е н с и в н о с т ь ю транспирации (рис . 10.8, а) и содержанием воды в листьях (рис . 10.8,5) ' выявляются сходные т е н д е н ц и и . Н а б о л ь ш е й части диапазона изменений этих признаков водного р е ж и м а коэффициент активности воды остается п р а к т и ч е с к и неизм е н н ы м . Г р а н и ч н ы м и з н а ч е н и я м и д л я интенсивности транспирации, ниже которых коэффициент активности воды резко увеличивается, являются величины 0.5-0.7 г,„^„ г;^:с.массы ч " ' (рис. 1 0 . 8 , а ) . Д л я содержания воды эти г р а н и ц ы находятся в д и а п а з о н е 1.0-1.2 г„„^^ г'^^ ^^^^^ (рис . 10.8, б) . Указанные г р а н и ч н ы е значения м о г у т и д е н т и ф и ц и р о в а т ь некотор у ю зону значений признаков , по д о с т и жении которой з аканчивает ся н о р м а л ь ное фу нкцио нир о вание и растение попадает в н е б л а г о п р и я т н ы е («стрессовые») условия п р о и з р а с т а н и я . П р и этом, судя по всему , у с и л и в а ю т с я р е г у л я т о р н ы е п р о ц е с с ы по типу отрицательных обратных связей, поскольку показатели функ
ционирования с т а б и л и з и р у ю т с я на м и н и м а л ь н ы х у р о в н я х с в о е й р е а л и з а ц и и и мало связаны с в е л и ч и н а м и коэффициента активности воды.
С другой стороны, реализация процессов ф у н к ц и о н и р о в а н и я достаточно сильно зависит от активности воды в растениях (рис . 10.9). Если активность воды в ы ш е величины 0.985, то р а з в и т и е всех процессов свидетельствует о благоприятных условиях почвенного водоснабжения и активном функционировании . П р и этом наблюдают-
4
СВ, г/г
Рис. 10.8. Связь коэффициентов активности воды с интенсивностью транспирации (а) и содержанием воды в листьях растений (б).
ся интенсивная транспирация , превосходящая 1.5 ^^оды ^ -1 сух. массы ч-' (рис. 10.9, а), до
статочно низкие значения д е ф и ц и т а водного н а с ы щ е н и я , не п р е в ы ш а ю щ и е 20-25% (рис. 10.9, б) , высокое содержание воды, более 1.9-2.0 г^^^^ г ^ ^ ^^^^^ (рис. 10.9, в). Если активность воды становится м е н ь ш е величины 0.980, то происходит резкое увеличение д е ф и ц и т а водного н а с ы щ е н и я (в экстремальных случаях п р и б л и ж а ю щ е е с я к 80%) (рис. 10.9, б ) . И н т е н с и в н о с т ь т р а н с п и р а ц и и и с о д е р ж а н и е воды в листьях при этом снижаются д о м и н и м а л ь н о возможных уровней и стабилизируются на них (рис.
' Подчеркнем, что здесь имеется в виду исследование не зависимостей, а статистических связей.
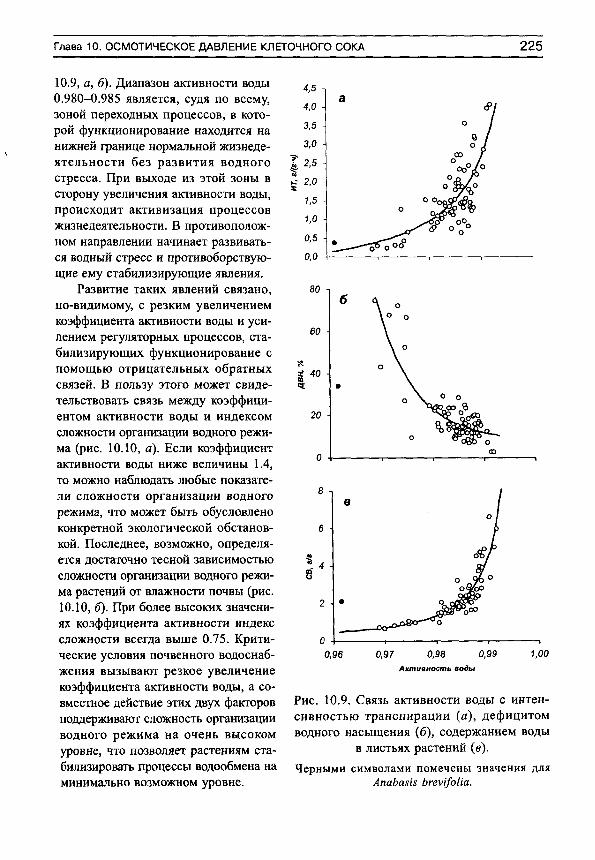
Глава 10. ОСМОТИЧЕСКОЕ ДАВЛЕНИЕ КЛЕТОЧНОГО СОКА 225
10.9, а, б). Диапазон активности воды 0.980-0.985 является , судя по всему, зоной переходных п р о ц е с с о в , в которой ф у н к ц и о н и р о в а н и е находится н а нижней границе нормальной жизнедея т е л ь н о с т и б е з р а з в и т и я в о д н о г о стресса . П р и выходе из этой з о н ы в сторону увеличения активности воды, п р о и с х о д и т а к т и в и з а ц и я п р о ц е с с о в жизнедеятельности . В противоположном направлении начинает развиваться водный стресс и противоборствующ и е ему с т а б и л и з и р у ю щ и е явления .
Развитие т а к и х я в л е н и й связано , по-видимому, с р е з к и м у в е л и ч е н и е м коэффициента активности воды и усилением регуляторных процессов , стаб и л и з и р у ю щ и х ф у н к ц и о н и р о в а н и е с п о м о щ ь ю о т р и ц а т е л ь н ы х о б р а т н ы х связей. В пользу этого м о ж е т свиде тельствовать связь м е ж д у коэффицие н т о м а к т и в н о с т и в о д ы и и н д е к с о м сложности организации водного режима (рис. 10.10, а). Е с л и коэффициент активности воды ниже величины 1.4, то можно наблюдать л ю б ы е показатели с л о ж н о с т и о р г а н и з а ц и и в о д н о г о режима, что может б ы т ь обусловлено конкретной экологической о б с т а н о в кой. Последнее , возможно, определяется достаточно тесной зависимостью сложности организации водного режима растений от влажности почвы (рис. 10.10, б). При более высоких значениях к о э ф ф и ц и е н т а а к т и в н о с т и индекс сложности всегда в ы ш е 0.75. К р и т и ческие условия почвенного водоснабж е н и я в ы з ы в а ю т р е з к о е у в е л и ч е н и е коэффициента активности воды, а совместное действие этих двух факторов поддерживают сложность организации в о д н о г о р е ж и м а н а о ч е н ь в ы с о к о м уровне, что позволяет растениям стабилизировать процессы водообмена на минимально возможном уровне .
4,5 1
4,0 -
3,5
3,0
I 2,5
^-2,0
^5
1,0 -
0,5 -
0,0
80 1
60
40
20
О 4
0,96 0,97 0,98 0,99 Активность воды
1,00
Р и с . 10.9. Связь а к т и в н о с т и в о д ы с и н т е н с и в н о с т ь ю т р а н с п и р а ц и и (а), д е ф и ц и т о м водного н а с ы щ е н и я (б) , с о д е р ж а н и е м воды
в листьях р а с т е н и й (в).
Черными символами помечены значения для АпаЬа513 Ъгем1/оИа.
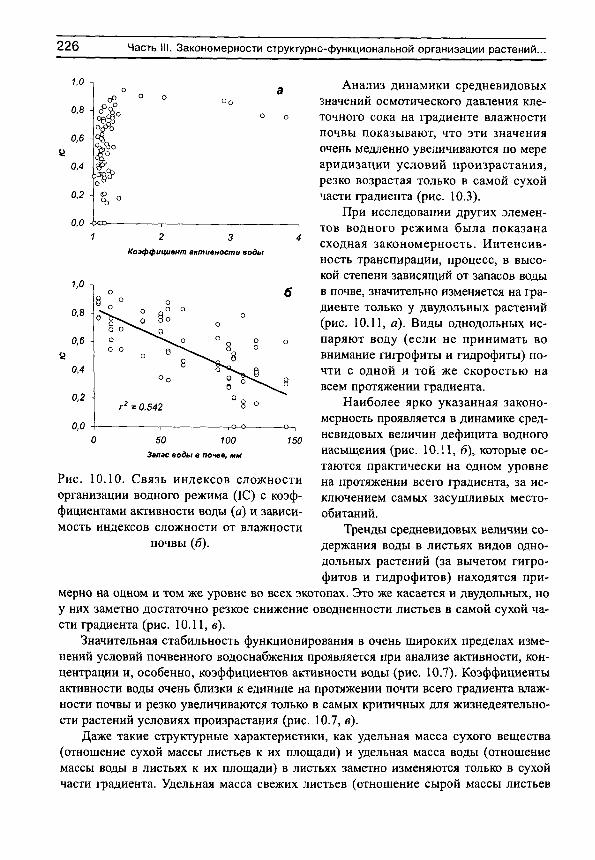
226 Ч а с т ь III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й . .
Оо
2 Иоэффицие 1 активности воды
1,0 -,
0,8 -
о
8 о °
8 ^ 0,6 - о
0 о
0,4 -
0,2 -г
0,0 -
= 0.542
А н а л и з д и н а м и к и с р е д н е в и д о в ы х значений о смо т ическо г о давления клеточного сока на г р а д и е н т е в л а ж н о с т и п о ч в ы п о к а з ы в а ю т , ч т о э т и з н а ч е н и я очень медленно увеличиваются по мере а р и д и з а ц и и у с л о в и й п р о и з р а с т а н и я , резко возрастая т о л ь к о в с а м о й сухой части градиента (рис . 10.3).
П р и и с с л е д о в а н и и д р у г и х э л е м е н т о в в о д н о г о р е ж и м а б ы л а п о к а з а н а с х о д н а я з а к о н о м е р н о с т ь . И н т е н с и в н о с т ь т р а н с п и р а ц и и , п р о ц е с с , в в ы с о кой степени зависящий от запасов воды в почве, значительно изменяется на град и е н т е только у д в у д о л ь н ы х р а с т е н и й (рис. 10.11, а). В и д ы о дно до ль ны х ис п а р я ю т в о д у ( е с л и н е п р и н и м а т ь во внимание г и г р о ф и т ы и г и д р о ф и т ы ) поч т и с о д н о й и т о й ж е с к о р о с т ь ю н а всем протяжении градиента .
Н а и б о л е е я р к о у к а з а н н а я з а к о н о мерность проявляется в д и н а м и к е средн е в и д о в ы х в е л и ч и н д е ф и ц и т а водного н а с ы щ е н и я (рис . 10.11, б), которые о с т а ю т с я п р а к т и ч е с к и на о д н о м у р о в н е на протяжении всего градиента , за иск л ю ч е н и е м с а м ы х з а с у ш л и в ы х м е с т о обитаний .
Тренды средневидовых величин сод е р ж а н и я воды в л и с т ь я х в и д о в однод о л ь н ы х р а с т е н и й (за в ы ч е т о м г и г р о ф и т о в и г и д р о ф и т о в ) н а х о д я т с я п р и
мерно на одном и т о м же уровне во всех экотопах. Это же касается и двудольных, н о у них заметно д о с т а т о ч н о резкое с н и ж е н и е оводненности л и с т ь е в в с а м о й сухой части градиента (рис . 10.11, в).
Значительная с т а б и л ь н о с т ь функционирования в очень ш и р о к и х пр еделах изменений условий почвенного водоснабжения проявляется при анализе активности , концентрации и, о с о б е н н о , к о э ф ф и ц и е н т о в активности в о д ы (рис . 10.7). К о э ф ф и ц и е н т ы активности воды очень близки к единице на протяжении почти всего градиента влажности почвы и резко увеличиваются только в самых к р и т и ч н ы х для ж и з н е д е я т е л ь н о сти растений у с л о в и я х произрастания (рис. 10.7, в).
Д а ж е т а к и е с т р у к т у р н ы е характеристики , как удельная м а с с а сухого в е щ е с т в а (отношение сухой массы листьев к их площади) и удельная масса воды (отношение массы воды в листьях к их площади) в листьях заметно изменяются только в сухой части градиента . Удельная масса свежих листьев ( о т н о ш е н и е с ы р о й м а с с ы листьев
50 100 750 Запас воды в почве, мм
Р и с . 10.10. С в я з ь и н д е к с о в с л о ж н о с т и организации водного р е ж и м а (1С) с коэффициентами активности воды (а) и зависимость и н д е к с о в с л о ж н о с т и от в л а ж н о с т и
п о ч в ы (б) .

Глава 10. ОСМОТИЧЕСКОЕ ДАВЛЕНИЕ КЛЕТОЧНОГО СОКА 227
к' « 2
К ИХ п л о щ а д и ) с о в е р ш е н н о не з а в и с и т о т и з м е н е н и й з а п а с о в в о д ы в п о ч в е (рис. 9.6).
Эти факты заставляют нас предположить , что почти на всем п р о т я ж е н и и градиента влажности почвы нет местообитаний, которые принято называть «стрессовыми» . Экологические условия этих местообитаний , как правило , соответствуют физиологич е с к и м н о р м а м р е а к ц и й р а с т е н и й -д о м и н а н т о в и э д и ф и к а т о р о в с о о б - 5 ществ . В и д ы в каж дом местообита нии п о л н о с т ь ю соответствуют эко- ^ логической и фитоценотической обстановке. Изменения интенсивности
^ 3 или степени проявления п р о ц е с с о в ф у н к ц и о н и р о в а н и я , с и с т е м н о с т и (сложности организации , ф о р м и р о вания и р а з в и т и я регуляторных яв лений, взаимосвязи функций) в направлении аридизации условий произрастания отражают скорее тенденции э в о л ю ц и о н н ы х и з м е н е н и й рас тений по мере освоения и м и н о в ы х экологических н и ш. Д р у г и м и словами, при перемещении против градиента влажности почвы м ы в и д и м не усиление водного стресса , а отражен и е э в о л ю ц и о н н о г о у с л о ж н е н и я структурно-функциональной организ а ц и и р а с т е н и й , н а п р а в л е н н о г о н а расширение экспансии видов во все более аридные экотопы.
В н у т р и э к о т о п и ч е с к а я и з м е н ч и вость п р и з н а к о в , н а п р о т и в , м о ж е т отражать степень д а в л е н и я у с л о в и й п р о и з р а с т а н и я на ф у н к ц и о н и р о в а -
а
8 •
. 2 • • <*
80 1
60
| 4 М
20
о о
7 -,
6
Рис . 10.11. Т р е н д ы с р е д н е в и д о в ы х (а) значений и н т е н с и в н о с т и т р а н с п и р а ц и и о д н о д о л ь н ы х (а -1 ) и двудольных (а-2) растений, (б) д е ф и ц и та водного н а с ы щ е н и я , (в) содержан и я в о д ы у о д н о д о л ь н ы х ( в - 1 ) и
двудольных (в-2) склерофитов .
Пунктирными линиями показаны доверительные интервалы.
•3 8 - 3 .
2 •
50 100
Запас воды в почве, им
в о .
150

228 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й .
ние растений . Значительное отклонение этих условий от о п т и м а л ь н ы х м о ж е т быть причиной и з м е н ч и в о с т и признаков водообмена в очень ш и р о к и х г р аницах . Эти границы могут быть ш и р е , чем диапазон соответствующих с р е д н е в и д о в ы х величин на протяжении б о л ь ш е й части градиента влажности почвы. Д л я п р и м е р а приведем данные по ковыльно-разнотравно-вострецовому сообществу (участок 8 в табл. 7 .1 , 10.1). Среднедневные значения интенсивности транспирации у о дно до ль ны х и двудольных склерофитов изменялись от 0.3 до 3.0 г^^^ ^^^.масш , д е ф и ц и т а водного н а с ы щ е ния — от 5 до 55%, содержания воды — от 0.7 д о 2.7 г,„^у т~^^ ^^^^, о с м о т и ч е с к о го давления клеточного с о к а — от 1.2 д о 4.3 М П а . Если о п р е д е л и т ь в о д н ы й стресс как ф у н к ц и о н и р о в а н и е с очень низкими показателями и н т е н с и в н о с т и т р а н с п и р а ц и и и оводненности листьев , и очень высокими — д е ф и ц и т а водного н а с ы щ е н и я и о с м о тического давления при экстремальных условиях почвенного водоснабжения (как недостаточного, так и избыточного) , то можно сказать, что время от времени в течение вегетации р а с т е н и я м о г у т и с п ы т ы в а т ь значительное д а в л е н и е этих э к с т р е м а л ь н ы х условий, в ы з ы в а ю щ е е состояние водного стресса .
Т а к и м образом, в пределах одного экотопа м о ж н о наблюдать т а к и е о б р а т и м ы е состояния функционирования , которые свидетельствуют о водном стрессе у растений. В противоположность этому, необратимые изменения функционирования на градиенте влажности почвы отражают э в о л ю ц и о н н ы е изменения видо в , н а п р а в л е н н ы е на освоение и м и н о в ы х экологических н и ш .
Резюме . С т а т и с т и ч е с к и й анализ осмотического д а в л е н и я клеточного сока в л и с тьях т р а в я н и с т ы х р а с т е н и й н а градиенте влажности п о ч в ы показал с л е д у ю щ е е . В е с ь м а с с и в и с х о д н ы х д а н н ы х о с м о т и ч е с к о г о д а в л е н и я и м е е т с р е д н е е з н а ч е н и е X = 2.08 М П а . Т и п и ч н ы е значения этого показателя н а г р а д и е н т е в л а ж н о с т и почвы не п р е в ы ш а ю т 3.26 М П а . Статистические п а р а м е т р ы о с м о т и ч е с к о г о д а в л е н и я у видов однодольных и двудольных склерофитов практически не р а з л и ч а ю т с я . Соответствующие средние значения этого признака р а в н ы 2.11 и 2.03 М П а . Т и п и ч н ы е значения осмотического давления для однодольных с к л е р о ф и т о в находятся в пределах 1.38-3.06 М П а , для д в у д о л ь н ы х — м е ж д у 1.24-3.25 М П а . П о к а з а н а т е с н а я связь о с мотического давления клеточного сока с оводненностью листьев . П р едло жена модель для расчета н е д о с т а ю щ и х значений осмотического давления по из вест ны м величинам содержания воды и д е ф и ц и т а водного насыщения . Средние для каждого экотопа уровни осмотического д а в л е н и я т е с н о связаны с в л а ж н о с т ь ю почвы. Ш и р и н а з о ны раз броса с р е д н е в и д о в ы х величин незначительно увеличивается от г у м и д н о й к ар идно й части градиента . Э т о и л л ю с т р и р у е т с я с р а в н е н и е м т р е н д о в м и н и м а л ь н ы х и максимальных д л я каждого сообщества средневидовых значений о смо т ическо г о давления . Изучение с т р у к т у р н о - ф у н к ц и о н а л ь н ы х корреляций показало , ч т о о с м о т и ч е с к о е дав ление связано у однодольных растений с проводящей т к а н ь ю и губчатой паренхимой , у двудольных — с п р о в о д я щ е й т к а н ь ю и п а р е н х и м н о й обкладкой пучков . О п р е д е л е ны величины активности и коэффициента активности воды у изученных растений. Показана д и н а м и к а этих показателей на градиенте в л а ж н о с т и почвы. С д е л а н о предположение , что диапазон активности воды 0.980-0.985 является , судя п о всему, зоной переходных процессов , в которой ф у н к ц и о н и р о в а н и е находится на н и ж н е й г р а н и ц е нормальной ж и з н е д е я т е л ь н о с т и без развития водного стресса . П р и выходе из этой

Глава 10. ОСМОТИЧЕСКОЕ ДАВЛЕНИЕ КЛЕТОЧНОГО СОКА 229
зоны в сторону увеличения активности воды, происходит активизация процессов жизн е д е я т е л ь н о с т и . В п р о т и в о п о л о ж н о м н а п р а в л е н и и н а ч и н а е т р а з в и в а т ь с я в о д н ы й стресс и п рот и воборствующие ему с т а б и л и з и р у ю щ и е явления . Это связано , по-видимому, с резким увеличением коэффициента активности воды и у с и л е н и е м регуляторных процессов , с т абилизирующих функционирование с п о м о щ ь ю отрицательных обратных связей . С д е л а н о предположение , что почти на всем п р о т я ж е н и и г р а д и е н т а влажности почвы нет местообитаний, которые принято называть «стрессовыми» . Экологические условия этих местообитаний , как правило , соответствуют физ ио ло г ичес ким нормам р е а к ц и й растений-доминантов и эдификаторов с о о б щ е с т в . В и д ы в каждом местообитании п о л н о с т ь ю соответствуют экологической и фитоценотической обстановке . Внутриэкотопическая изменчивость признаков , напротив , может отражать степень давления у с л о в и й произрастания на ф у н к ц и о н и р о в а н и е растений . Таким образом, в пределах одного экотопа можно наблюдать такие о б р а т и м ы е состояния функционирования , которые свидетельствуют о водном с т р е с с е у растений . В противоположность этому, необратимые изменения функционирования на градиенте в л а ж н о сти почвы отражают эволюционные изменения видов, направленные на освоение и м и новых экологических н и ш .

230 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й .
ГЛАВА 11. МЕЖВИДОВЫЕ СТРУКТУРНО-ФУНКЦИОНАЛЬНЫЕ КОРРЕЛЯЦИИ'
Исследование м е ж в и д о в ы х структурных и структурно-функциональных корреляций на градиентах факторов среды представляет н е с о м н е н н ы й и н т е р е с д л я в ы я с н е ния р я д а в о п р о с о в экологической анатомии и ф и з и о л о г и и р а с т е н и й . М е ж в и д о в ы е структурные корреляции могут указать на характер компартментации потоков и объем пулов запасания в л и с т ь я х растений , согласованность их и з м е н е н и й при и з м е н е н и и условий п р о и з р а с т а н и я . С у щ е с т в е н н ы й вклад в этом н а п р а в л е н и и б ы л сделан в работе Б. Р. Васильева (1988).
Ряд показателей водообмена , судя по всему, имеет о п р е д е л е н н у ю с т р у к т у р н у ю основу. Э т о м о ж н о в ы я с н и т ь , исследуя с т р у к т у р н о - ф у н к ц и о н а л ь н ы е к о р р е л я ц и и . Широко известно , например , что плотность сложения мезофилла является признаком ксероморфной организации растений , она увеличивается в направлении от гумидных к аридным м е с т о о б и т а н и я м (8Ь1еМ5, 1950; Василевская , 1979).
Очевидно , что в этом же направлении уменьшаются интенсивность транспирации и оводненность листьев . Судя по всему, д о л ж н ы существовать экологические р я д ы видов , на м а т е р и а л е которых проявляются м е ж в и д о в ы е корреляции с т р у к т у р н ы х и ф у н к ц и о н а л ь н ы х признаков у растений как в пределах экотопа, т а к и на градиенте факторов среды. Эти корреляции могут помочь выявить и е р а р х и ю и характер влияния тех или и н ы х структурных признаков на функциональные .
С а м о н а л и ч и е т а к и х корреляций наводит на м ы с л ь о с у щ е с т в о в а н и и п а р а л л е л ь ной с т р у к т у р н о - ф у н к ц и о н а л ь н о й изменчивости у н е р о д с т в е н н ы х таксонов . И м е е т с я большой р я д работ, в котором рассматриваются структурные и ф у н к ц и о н а л ь н ы е характеристики и их взаимосвязи в листьях травянистых или д р е в е с н ы х растений в раз н ы х э к о л о г и ч е с к и х у с л о в и я х (Когпег е1 а1. , 1989; Оат1ег, Ьаигеп1 , 1994; 8т1Йг, МсС1еап, 1989; АЬгашз е! а1., 1994; З т И Ь е! а1., 1998; Аскег1у, Ке1сЬ, 1999;Сат1ег е ! а1., 1999; Руапкоу е1 а1., 1999; Ога1аш, ВошЬеШ, 2000 и др.) . М ы п о п ы т а л и с ь выявить такие корреляции в листьях травянистых растений, п р о и з р а с т а ю щ и х в р а з н ы х частях широкого градиента в л а ж н о с т и почвы.
Д а н н ы е с о б р а н ы в вегетационные периоды 1995-1997 гг. в 4 с о о б щ е с т в а х травяных экосистем, расположенных более или менее равномерно н а широком градиенте влажности почвы (табл. 7.1, с о о б щ е с т в а под номерами 1, 9, 14, 16) в п о й м е р . Хо-пер (Хоперский государственный природный заповедник, В о р о н е ж с к а я область) .
Соотношение тканей и свободных пространств в листьях однодольных и двудольных растений
Анализ д а н н ы х по анатомической структуре листьев (табл. 11.1) показал , что существуют статистически значимые различия м е ж д у в и д а м и однодольных и двудольных растений почти по всем т и п а м тканей и ёмкостям запасания листьев , за и с к л ю чением п а р е н х и м н ы х обкладок пучков (рис. 11.1). У о д н о д о л ь н ы х о т н о с и т е л ь н о б о лее развиты п р о в о д я щ а я ткань и губчатая п а р е н х и м а ( склеренхима и в о з д у ш н ы е п о -
Раздел написан совместно с Ю . В. Гамалеем.

Таб
лица
11
.1
Ср
едн
ие
знач
ени
я и
по
каз
ател
и в
ари
аци
и п
арц
иал
ьны
х о
бъ
емо
в тк
аней
и с
во
бо
дн
ых
пр
ост
ран
ств
ли
стье
в
№
уча
№
Э С
кл
П
ПО
П
г П
с М
г М
с В
П
стка
по
та
бл.
7.1
объ
Ви
д п
стка
по
та
бл.
7.1
екта
X
X ^х
X
X X
^х
X ^х
X
^х
X X
^х
1 1
Анет
та
аизМ
аса
1асц
. 5
14.5
1.
3 0
1.1
4.0
7.1
1.8
13.6
1.
8 33
.8
3.9
7.1
1.9
16.8
3.
4 0
2 Са
гех
сокМ
са
}. О
ау
6 15
.7
4.3
12.0
1.
9 13
.3
5.5
5.5
1.4
40.9
4.
0 0
8.5
5.6
0 4.
0 4.
8
3 Са
Иит
гш
кет
сит
"Л̂
ПЫ.
4 17
.1
1.5
0 10
.1
1.8
3.9
0.6
16.2
3.
3 26
.0
1.4
11.3
2.
1 15
.4
1.2
0
4 8И
ра
Ьог
у51к
етса
К1о
к. е
х Рг
окис
!. 3
7.4
0.5
20.1
4.
1 13
.9
3.1
3.7
1.2
32.0
4.
1 0
2.6
0.9
0 0
5 Тп
/оН
ит
агуе
пзе
Ь.
4 28
.8
4.6
0 8.
6 1.
1 4.
6 1.
5 23
.2
7.4
17.4
7.
8 9.
3 4.
6 3.
4 1.
3 0
6 Уе
гот
са
зр1с
а1а
Ь.
4 12
.7
1.0
0 5.
4 2.
5 7.
4 2.
5 24
.5
4.1
37.2
3.
1 7.
2 3.
2 5.
6 4.
5 0
9 7
Са1а
та§
гозИ
з ер
щею
з 5
22.3
4.
1 8.
9 5.
2 5.
1 3.
1 1.
3 0.
9 43
.9
6.6
0 17
.9
2.9
0 0
8 Са
гех
те1
апоз
(асН
уа
В\е
Ъ.
4 15
.4
1.9
8.1
3.1
14.6
4.
8 3.
2 1.
4 32
.7
1.8
0 2.
4 1.
0 0
23.6
7.
7 ех
ХУ
НИ
.
9 О
аИит
т
скет
сит
5
23.5
2.
1 0
3.1
0.7
3.0
0.7
12.8
3.
4 25
.5
2.2
17.4
3.
5 14
.8
2.1
0
10
Ьугк
гит
У1
г§а1
ит
Ь.
5 27
.5
3.3
0 1.
2 0.
5 4.
2 1.
1 14
.1
2.4
30.6
4.
3 11
.8
5.5
10.6
2.
0 0
П
8ап^
1зог
Ьа
оЛС
ктаН
з Ь
. 5
21.7
3.
0 0
3.9
5.0
5.6
3.2
14.7
5.
5 26
.6
1.8
17.6
7.
1 9.
9 2.
5 0
12
Тапа
сеШ
т
уи1^
аге
Ь.
5 11
.3
1.3
0 2.
9 1.
8 5.
1 2.
6 14
.1
1.3
27.2
2.
6 11
.0
5.1
28.4
3.
7 0
14
13
Саге
х са
езрИ
оза
Ь.
5 14
.2
1.9
4.8
2.4
13.7
1.
8 5.
1 1.
5 34
.4
4.0
0 3.
4 2.
7 0
24.4
2.
1
14
Ртре
п<1и
1а
и1т
апа
5 17
.6
2.2
0 1.
5 0.
9 3.
6 1.
1 13
.4
2.1
34.0
3.
5 20
.9
3.6
8.9
0
15
1пз
рзеи
<1а
соги
з Ь
. 5
6.0
0.9
2.1
2.0
4.5
2.8
4.8
1.6
15.3
2.
5 8.
5 1.
2 3.
7 1.
2 2.
2 0.
6 45
.9
9.7
16
Зап^
шзо
гЬа
о//1
стаи
з 5
24.1
3.
2 0
1.4
0.9
4.6
2.3
9.9
2.0
31.1
3.
1 22
.3
5,0
6.7
0.5
0
17
Уа1е
папа
•н
>о1§
епз1
з Каг
ак.
5 13
.4
0.4
0 1.
0 0.
5 3.
2 1.
3 20
.3
3.8
34.1
6.
2 19
.4
5.5
8.7
3.8
0
18
Уего
тса
Ь
п^У
оИа
5 11
.2
2,5
0 1.
8 0.
3 2.
7 0.
7 20
.4
5.7
29.2
2.
6 21
.3
3.7
13.2
6.
5 0
16
19
АИзт
а р1
ап1а
§о-а
диаИ
са
Ь.
5 15
.1
1.0
0 1.
1 0.
4 3.
1 1.
4 26
.3
2.1
16.9
2.
9 27
.7
1.4
9,1
2.5
0
20
Вшот
из
итЬе
ИаШ
з Ь
. 5
3.0
0,6
0 8.
3 1.
6 8.
1 1.
1 8.
3 1.
3 0
2.6
1.8
0 61
.1
3.5

Око
нчан
ие
таб
лицы
11
.1
№
уча
стка
по
та
бл.
7.1
№
объ
ек
та
Ви
д
Э
Скл
П
П
О
Пг
Пс
Мг
Мс
ВП
№
уч
аст
ка
по
табл
. 7.
1
№
объ
ек
та
Ви
д
X X
X X
X X
^х
X X
^х
X •У
х
21
22
23
24
Саг
ех а
сиШ
Ь.
Ьус
ориз
еи
гора
еиз
Ь.
5а§Ш
апа
за§И
Н/о
Иа
Ь.
Зраг
^ат
ит
егес
Шт
Ь.
5 5 5 5
9.4
1.3
24.5
4.
6
18.6
2.
6
4.4
0.6
8.8
0.7
0 0 4.7
1.5
9.7
0.8
0.7
0.3
0.8
0.5
2.1
0.5
4.2
0.9
3.4
1.7
2.3
1.3
2.4
0.2
25.2
3.
7
10.8
2.
4
15.2
3.
3
31.4
4.
6
0
24.1
3.
6
18.4
3.
6
0
3.1
1.1
21.6
3.
4
26.9
4.
6
10.5
2.
5
0
14.9
3.
4
16.3
2.
6
0
39.5
4.
5
0 0
31.1
7.
2
При
меч
ани
е:
п —
чи
сло
пов
торн
осте
й,
х —
ср
едн
ие,
—
ст
анда
ртн
ые
отк
лон
ени
я,
Э —
эп
иде
рм
ис,
С
кл —
ск
лере
нхи
ма,
П
—
пров
одящ
ая т
кань
, П
О —
пар
енхи
мна
я об
клад
ка п
учко
в, П
г —
губ
чата
я па
ренх
има,
Пс
— с
толб
чата
я па
ренх
има,
Мг
— м
ежкл
етни
ки г
уб
чато
й п
арен
хим
ы,
Мс
— м
ежк
летн
ик
и ст
олбч
атой
пар
енхи
мы
, В
П —
воз
душ
ны
е п
олос
ти.

Глава 11. М Е Ж В И Д О В Ы Е С Т Р У К Т У Р Н О - Ф У Н К Ц И О Н А Л Ь Н Ы Е К О Р Р Е Л Я Ц И И 233
35 л
30 -
П 25 •
I о 20 -I ё ^5 -
5 -
О -
а1
Э Скл Пр ПО Пг Пс Мг Мс ВП
Рис. 11.1. Средние п а р ц и а л ь н ы е объемы тканей и свободных п р о с т р а н с т в в листьях однодольных (1) и двудольных (2) растений .
Э — эпидермис, Скл — склеренхима, Пр — проводящая ткань, П О — паренхимные обкладки пучков, Пг — губчатая паренхима, Пс — столбчатая паренхима, М г — межклетники губчатой
паренхимы, М с — межклетники столбчатой паренхимы, В П — воздушные полости.
лости встречались только у р а с т е н и й этой группы) . Эпидер мис , столбчатая паренхима, м е ж к л е т н и к и (губчатой и столбчатой п а р е н х и м ы ) з а н и м а ю т о т н о с и т е л ь н о больший объем в листьях двудольных растений.
Динамика парциальных объемов тканей и свободных пространств в листьях растений на градиенте влажности почвы
Различия м е ж д у однодольными и двудольными растениями о бнар у живаю т ся также п р и и с с л е д о в а н и и и з м е н е н и й п а р ц и а л ь н ы х о б ъ е м о в т к а н е й и с в о б о д н ы х п р о странств вдоль экологических рядов их видов на градиенте в л а ж н о с т и почвы.
У однодольных растений о б н а р у ж е н ы зависим о ст и изменений п а р ц и а л ь н ы х объемов от в л а ж н о с т и п о ч в ы для проводящей ткани (рис . 11.2, а), с к л е р е н х и м ы (рис . 11.2, в) , губчатой п а р е н х и м ы (рис . 11.2, г) и межклет нико в и в о з д у ш н ы х п о л о с т е й (рис. 11.2, е). Д л я всех тканей корреляции отрицательны, т. е. по мере аридизации у с ловий произрастания п а р ц и а л ь н ы е объемы проводящей ткани, с к л е р е н х и м ы и губчатой п а р е н х и м ы увеличиваются . Компенсация увеличения п а р ц и а л ь н ы х о б ъ е м о в тканей против градиента влажности почвы происходит за счет относительного у м е н ь ш е ния с в о б о д н ы х п р о с т р а н с т в в листьях (увеличения их п л о т н о с т и ) . О т м е т и м очень высокую степень корреляции относительных объемов склеренхимы и свободных пространств в листьях с в л а ж н о с т ь ю почвы. П р и д в и ж е н и и о д н о д о л ь н ы х р а с т е н и й из гумид н ых экотопов в а р и д н ы е происходит не только у п л о т н е н и е л и с т ь е в , но и д о с таточно ярко выраж енная их лигнификация .

234 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й .

Глава 11. М Е Ж В И Д О В Ы Е С Т Р У К Т У Р Н О - Ф У Н К Ц И О Н А Л Ь Н Ы Е К О Р Р Е Л Я Ц И И 235
Из всех тканей листьев двудольных растений только проводящая ткань (рис. 11.2, а) и п а р е н х и м н ы е о б к л а д к и пучков (рис. 11.2, б) и зменятся согласованно с изменениями в л а ж н о с т и почвы. В сухих экотопах относит ель но е количество этих тканей больше по с р а в н е н и ю с в л а ж н ы м и м е с т о о б и т а н и я м и ( с к л е р и ф и к а ц и я с т р у к т у р ы ) . Уплотнение листьев против градиента влажности почвы происходит за счет относительного у м е н ь ш е н и я количества межклетников губчатой п а р е н х и м ы (рис . 11.2, д) и, в меньшей степени, за счет уменьшения общего количества межклет нико в (рис. 11.2, е) в листьях двудольных растений .
Следует отметить отсутствие коррелятивных связей парциальных объемов эпидермиса , столбчатой п а р е н х и м ы , в целом х л о р е н х и м ы ( со во ку пно ст и губчатой и столбчатой п а р е н х и м ы за вычетом клеток , не с о д е р ж а щ и х х л о р о ф и л л ) с в л а ж н о с т ь ю почвы и у однодольных, и у двудольных растений .
Структурные корреляции в листьях растений
Э п и д е р м и с . Как для однодольных, так и для д ву до ль ны х р а с т е н и й о б н а р у ж е н ы корреляции п а р ц и а л ь н ы х объемов э п и д е р м и с а и х л о р е н х и м ы (рис . 11.3, а). Д л я однодольных эта связь п о л о ж и т е л ь н а (рис . 11.3, а -1) , для двудольных — отрицательна (рис. 11.3, а-2) . Такое значительное различие м е ж д у этими г р у п п а м и в и д о в обусловлено , судя по всему, р а з н ы м и п а р ц и а л ь н ы м и о б ъ е м а м и х л о р е н х и м ы . У двудольных растений эти объемы заметно больше, чем у однодольных и при у в е л и ч е н и и долевого участия эпидермиса в листьях уменьшаются . У однодольных напротив — относительное увеличение эпидермиса сопровождается увеличением доли х л о р е н х и м ы в листьях растений. В целом для обеих групп корреляция м е ж д у п а р ц и а л ь н ы м и о бъемами эпидермиса и х л о р е н х и м ы удовлетворительно описывается п о л и н о м о м второй степени (рис. 11.3, а -3) . Это означает, что у в е л и ч е н и е объема э п и д е р м и с а до 15-20% сопровождается у в е л и ч е н и е м объема х л о р е н х и м ы , а при б о л ь ш и х объемах э п и д е р м а л ь н о й ткани участие х л о р е н х и м ы в сложении листьев у м е н ь ш а е т с я .
Д л я одн одольн ых р а с т е н и й выявлена коррелятивная связь м е ж д у п а р ц и а л ь н ы м и объемами э п и д е р м и с а и в о з д у ш н ы х полостей (рис . 11.3, б). П р и у в е л и ч е н и и до ли эпидермальной ткани участие в о з д у ш н ы х п о л о с т е й в сло жении л и с т а у м е н ь ш а е т с я . Эта корреляция обусловлена наличием очень тесной связи м е ж д у парциальными обье-мами воздушных полостей и хлоренхимы в листьях однодольных растений (рис. 11.6, б). Поскольку о б ъ е м ы х л о р е н х и м ы и э п и д е р м и с а в з а и м о с в я з а н ы (рис . 11.3, а -1) , то и объемы воздушн ых полостей и эпидермиса у однодольных оказываются з ависимыми друг от друга (рис. 11.3, б) . Здесь м ы видим первые признаки того , что парциаль -
Рис. 11.2. И з м е н е н и я парциальных объемов тканей и свободных п р о с т р а н с т в в листь ях однодольных (светлые символы, сплошные линии регрессии) и двудольных (черные символы, п р е р ы в и с т ы е л и н и и регрессии) р а с т е н и й на градиенте в л а ж н о с т и почвы.
На осях ординат парциальные объемы проводящих тканей (а) , обкладок пучков (б) , склеренхимы (в), губчатой паренхимы (г), межклетников губчатой паренхимы (д), воздушных полостей
и межклетников (е-1), межклетников (е-2).

236 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й . .
= 0.575
10 20 30
70
I I 50 ° I 1 § I ^ 30
ные объемы всех т к а н е й и с в о б о д н ы х п р о с т р а н с т в в л и с т ь я х о д н о д о л ь н ы х растений оказываются в той или иной степени з а в и с и м ы м и друг от друга. И з м е н е н и е д о л и у ч а с т и я к а к о й - л и б о одной т к а н и и л и м е ж к л е т о ч н и к о в н е и з бежно ведет к к о р р е л я т и в н ы м из мене н и я м п а р ц и а л ь н ы х о б ъ е м о в д р у г и х структур мезофилла .
С к л е р е н х и м а . Э т а т к а н ь встречалась в листьях только о дно до ль ных растений. Б ы л и о б н а р у ж е н ы корреляции с к л е р е н х и м ы с п р о в о д я щ е й т к а н ь ю ' ( р и с . 11.4, а), г у б ч а т о й п а р е н х и м о й (рис. 11.4, б) и с в о б о д н ы м и п р о с т р а н ствами (суммой в о з д у ш н ы х полостей и межклетников) в л и с т ь я х (рис . 11.4, в).
Связь с к л е р е н х и м ы с п р о в о д я щ е й т к а н ь ю и губчатой п а р е н х и м о й выра жена не очень отчетливо , однако пр о с л е ж и в а е т с я о б щ а я т е н д е н ц и я б о л е е или менее согласованного у в е л и ч е н и я п а р ц и а л ь н ы х о б ъ е м о в э т и х т к а н е й с у в е л и ч е н и е м до ли у ч а с т и я с к л е р е н х и м ы в с л о ж е н и и л и с т ь е в . Н а и б о л е е сильно проявляется взаимосвязь м е ж д у свободными п р о с т р а н с т в а м и листьев и склеренхимой. Увеличение относительных о б ъ е м о в с к л е р е н х и м ы с о п р о в о ж дается у м е н ь ш е н и е м до ли участия свободных п р о с т р а н с т в в с л о ж е н и и м е з о филла (т. е. его у п л о т н е н и е м ) , и наоборот. Это м о ж е т б ы т ь о бу сло влено тем , что свободные пространства и склеренх и м а в л и с т ь я х о д н о д о л ь н ы х на гради
енте в л а ж н о с т и почвы и з м е н я ю т с я в противоположных н а п р а в л е н и я х (рис . 11.2, а; 11.2, е). Н о не и с к л ю ч е н о и п р я м о е взаимодействие м е ж д у н и м и : р а з в и т и е склер ен х и м ы в а р и д н ы х экотопах препятствует у в е л и ч е н и ю с в о б о д н ы х п р о с т р а н с т в , а раз витие свободных п р о с т р а н с т в во в л а ж н ы х местообитаниях ограничивает у в е л и ч е н и е относительного объема с к л е р е н х и м ы .
С в о б о д н ы е п р о с т р а н с т в а в л и с т ь я х . О д н о д о л ь н ы е и д в у д о л ь н ы е р а с т е н и я в своей совокупности п о д ч и н я ю т с я общей т е н д е н ц и и . Э т а т е н д е н ц и я з а к л ю ч а е т с я в
20
10
О
=0.491 о
.—о 1 о 9 5 10 15
Парциальный объем эпидермиса, %
Рис. 11.3. К о р р е л я ц и и п а р ц и а л ь н ы х объемов эпидермиса с парциальными объемами хлорен хи мы (а) и в о з д у ш н ы х полостей (б) в листьях однодольных (светлые символы) и двудольных (черные символы) растений.
Цифрами обозначены линии регрессии для однодольных (1), двудольных (2), однодольных и
двудольных (3) растений.
' Здесь и в дальнейшем п о д корреляциями м е ж д у какими-либо структурами листьев мы будем иметь в виду корреляции м е ж д у парциальными объемами этих структур.

Глава 11 . М Е Ж В И Д О В Ы Е С Т Р У К Т У Р Н О - Ф У Н К Ц И О Н А Л Ь Н Ы Е К О Р Р Е Л Я Ц И И 237
уменьшении парциального обьема межклетников губчатой паренхимы с увеличением доли участия проводящей ткани (рис. 11.5, а), а т а к ж е с у м м а р н о й д о л и п р о в о д я щ е й т к а н и и о б к л а д о к пучков (рис. 11.5, б). Это может б ы т ь связано с тем, что п р о в о д я щ и е пучки, как правило, расположены в губчатой паренхиме и их большее развитие в сухих м е стообитаниях ограничивает увеличение п а р ц и а л ь н ы х о б ъ е м о в м е ж к л е т н и к о в губчатой паренхимы.
У м е н ь ш е н и е м е ж к л е т н ы х п р о с т ранств коррелятивно связано с увеличен и е м п а р е н х и м н ы х т к а н е й в л и с т ь я х о д н о д о л ь н ы х и д в у д о л ь н ы х р а с т е н и й . Так, уменьшение совокупности воздушных полостей и меж клетников у однодольных растений сопровождается увел и ч е н и е м г у б ч а т о й п а р е н х и м ы ( р и с . 11.6, а -1 ) . У м е н ь ш е н и е п а р ц и а л ь н ы х объемов м е ж к л е т н и к о в в листьях двудольных т а к ж е связано с увеличением доли участия губчатой паренхимы (рис. 11.6, а-2) . Кроме того, для однодольных растений о б н а р у ж е н а очень тесная отрицательная зависимость между парциа л ь н ы м и о б ъ е м а м и в о з д у ш н ы х полостей и х л о р е н х и м ы (рис . 11.6, б) .
Х л о р е н х и м а . В ы ш е уже б ы л и рас смотрены корреляции между эпидермисом и х л о р е н х и м о й у о д н о д о л ь н ы х и д в у д о л ь н ы х р а с т е н и й ( р и с . 11.3, а ) , с к л е р е н х и м о й и губчатой п а р е н х и м о й (рис. 11.4, б), м е ж к л е т н ы м и простран ствами и п а р е н х и м о й ( р и с . 11.6, а - 1 ; 11.6, б) у однодольных, губчатой паренхимой и межклетниками (рис. 11.6, а-2) у двудольных. К р о м е того, можно у в и д е т ь н а л и ч и е н е о ч е н ь т е с н о й ( Н = 0.487) отрицательной связи м е ж д у сов о к у п н о с т ь ю в с е х с в о б о д н ы х п р о странств и х л о р е н х и м о й в листьях однодольных растений . И н т е р е с н о отметить, что не было обнаружено корреля-
20 п
10 15 20 25
50 п
10 15 20 25
70 п
г' = 0.824
О 5 10 15 20 Парциальный объем склеренхимы, %
Р и с . 11.4. К о р р е л я ц и и п а р ц и а л ь н ы х объемов с к л е р е н х и м ы с п а р ц и а л ь н ы м и объем а м и проводящей ткани (Пр) (а) , губчатой п а р е н х и м ы (Пг) (б) , в о з д у ш н ы х полостей и м е ж к л е т н и к о в ( В П и М ) (в) в л и с т ь я х
однодольных растений .
238 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й .
30 1
О 5 10 15 Парциальные объемы Пр, %
30 1
О -I , , , , О 5 10 15 20
Парциальные объемы Пр и ПО, %
Р и с . 11.5. К о р р е л я ц и и п а р ц и а л ь н ы х объемов п р о в о д я щ и х тканей (Пр) и об к л а д о к п у ч к о в ( П О ) с п а р ц и а л ь н ы м и о б ъ е м а м и м е ж к л е т н и к о в губчатой п а ренхимы (Мг) в листьях однодольных и
двудольных р а с т е н и й .
о 10 20 30 40 50
Парциальные объемы, %
Р и с . 11.6. К о р р е л я ц и и м е ж д у п а р ц и а л ь н ы м и о б ъ е м а м и с в о б о д н ы х п р о странств и губчатой п а р е н х и м ы (Пг) в листьях однодольных ( 1 , светлые символы) и двудольных (2, ч е р н ы е символы) р а с т е н и й ( а ) , и м е ж д у п а р ц и а л ь н ы м и о б ъ е м а м и х л о р е н х и м ы (Хл) и в о з д у ш н ы х п о л о с т е й в л и с т ь я х о д н о д о л ь н ы х
р а с т е н и й (б).
На осях абсцисс парциальные объемы возд у ш н ы х п о л о с т е й и м е ж к л е т н и к о в ( а - 1 ) , межклетников (а-2), воздушных полостей (б).

Глава 11. М Е Ж В И Д О В Ы Е С Т Р У К Т У Р Н О - Ф У Н К Ц И О Н А Л Ь Н Ы Е К О Р Р Е Л Я Ц И И 239
тивных связей столбчатой п а р е н х и м ы с д р у г и м и структурами листа . Э т а т к а н ь раз вивается, судя по всему, достаточно автономно как у однодольных (у которых, впр о чем, она обнаруживается у небольшого числа видов) , так и у двудольных р а с т е н и й .
Обобщение структурных корреляций
Все структурные корреляции, о которых было упомянуто в ы ш е , о б о б щ е н ы на схемах отдельно для однодольных (рис. 11.7, а) и двудольных (рис. 11.7, б) растений. Эти схемы иллюстрируют значительные различия между д а н н ы м и группами видов по степени коррелированности объемов тканей и свободных п р о с т р а н с т в в л и с т ь я х . У однодольных растений п р а к т и ч е с к и все структуры листьев о б ъ е д и н е н ы в одну коррел я ц и о н н у ю систему. И з м е н е н и я в л ю б о м из звеньев этой с и с т е м ы повлекут за собой неизбежные т р а н с ф о р м а ц и и во всех остальных . Н а п р и м е р , обитая в более з а с у ш л и вых условиях почвенного водоснабжения , однодольные ф о р м и р у ю т м е н ь ш е е количество межклетных пространств в листьях. С о п р я ж е н н о с этим меняются соотношения парциальных объемов всех остальных листовых структур: увеличатся относительные обьемы склеренхимы и проводящих тканей (вместе с обкладками пучков) , увеличится количество х л о р е н х и м ы , изменится удельный объем эпидермиса .
Двудольные растения , в противоположность однодольным, д е м о н с т р и р у ю т больш у ю а в т о н о м н о с т ь п а р ц и а л ь н ы х объемов тканей листа . У них с о х р а н я е т с я тесная связь м е ж д у м е ж к л е т н и к а м и и п р о в о д я щ и м и тканями, а т а к ж е м е ж д у м е ж к л е т н и к а ми и губчатой п а р е н х и м о й . Х л о р е н х и м а в целом и, в какой-то степени , эпидермис , развиваются достаточно самостоятельно . П р и ухудшении у с л о в и й почвенного водоснабжения у двудольных растений , т а к ж е как и у однодольных , п р о и з о й д е т некоторое у м е н ь ш е н и е доли межклетников , особенно м е ж к л е т н и к о в губчатой п а р е н х и м ы . В соответствии с э т и м изменится относительное количество в л и с т ь я х п р о в о д я щ и х тканей и губчатой п а р е н х и м ы . П р и этом с о в е р ш е н н о необязательно п р о и з о й д у т изменения п а р ц и а л ь н ы х объемов х л о р е н х и м ы в целом и э п и д е р м и с а .
Такие з н а ч и т е л ь н ы е р а з л и ч и я м е ж д у однодольными и д в у д о л ь н ы м и р а с т е н и я м и могут б ы т ь обусловлены ф у н д а м е н т а л ь н ы м и р а з л и ч и я м и в р о с т е и х листьев . Листья однодольных и м е ю т поляризованный (интеркалярный) рост из точечного источника клеток в о с н о в а н и и листа . Таким образом, наиболее ст ар ые к л е т к и находятся в верхней части листовых пластинок. Следовательно, большая часть листа однодольных находится в сформированном состоянии и если претерпевает изменения с возрастом, то только путем растяжения клеток. Относительные объемы тканей закладываются в одной зоне р о с т а (в начале развития листа) и диктую т ся , судя п о всему, к о н к р е т н ы м и условиями произрастания . С о о т н о ш е н и я п а р ц и а л ь н ы х о б ъ е м о в т к а н е й и с в о б о д н ы х пространств практически не изменяются по мере увеличения возраста листьев . В этом отношении рост листьев однодольных более сходен с ростом о с е в ы х органов — стеблей и корней.
Листья двудольных и м е ю т много точек роста , м о з аично р а с п о л о ж е н н ы х на пластинке, а клоны н о в ы х клеток образуются на протяжении всего периода р о с т а и развития листьев . Естественно, что в этой модели роста изначальная коррелированность парциальных объемов тканей может не сохраняться . Н а и б о л ь ш е е значение пр ио бр ет аю т свободные пространства и проводящие ткани листьев двудольных, парциальные объемы которых изменяются строго коррелированно с условиями почвенного водоснабжения .

240 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й .
Рис . 11.7. С х е м ы струьстурных корреляций в листьях однодольных (а) и двудольных
(б) растений .
Сильные связи (коэффициенты детерминации больше 0.6) показаны сплошными линиями, слабые (коэффициенты детерминации меньше 0.6) — прерывистыми. Э — эпидермис, Скл — склеренхима, Пг — губчатая паренхима, Пс — столбчатая паренхима, П — проводящая ткань, П О — паренхимная обкладка пучков, М — межклетники, М г — межклетники губчатой паренхимы,
М с — межклетники столбчатой паренхимы, В П — воздушные полости.

Глава 11 . М Е Ж В И Д О В Ы Е С Т Р У К Т У Р Н О - Ф У Н К Ц И О Н А Л Ь Н Ы Е К О Р Р Е Л Я Ц И И 241
Структурно-функциональные корреляции в листьях растений
Интенсивность т р а н с п и р а ц и и . Скорость испарения воды с поверхности листьев однодольных и двудольных растений связана с п л о т н о с т ь ю сложения мезофилла . В о всех случаях интенсивность транспирации в ы ш е у тех видов , у которых наблюдалось относительно большее количество межклеточных пространств в листьях . Так, для однодольных р а с т е н и й в ы я в л я ю т с я корреляции и н т е н с и в н о с т и т р а н с п и р а ц и и с парциальными объемами межклетников (рис. 11.8, а-1) , а также суммарного объема воздушных полостей и межклетников (И = 0.685). Для двудольных т а к ж е характерна тесная зависимость интенсивности транспирации от суммарного объема межклетников в листьях растений (рис . 11.8, а -2) .
Поскольку межклеточные пространства коррелятивно связаны с р а з л и ч н ы м и тканями листьев обеих групп растений , то выявляются о т р ицат ель ны е корреляции скорости транспирации с проводящей т к а н ь ю (рис. 11.8, 6-1) и с к л е р е н х и м о й (рис. 11.8,
о 10 20 30 40 50 о 5 10 15
Рис. 11.8. К о р р е л я ц и и между и н т е н с и в н о с т ь ю т р а н с п и р а ц и и и п а р ц и а л ь н ы м и объемами тканей и с в о б о д н ы х пространств в листьях однодольных ( 1 , светлые с и м в о л ы )
и двудольных (2, черные символы) растений .
На осях абсцисс парциальные объемы межклетников (а) , проводящих тканей (б) , склеренхимы (в), губчатой паренхимы (г).
9 - 2353
242 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й .
1.2
1,0
0,8
0,6
0,4
0,2
0,0
1,2 -1
1,0 -
0,8 -
0,6 -
0,4
0,2
0,0
1,2 п
1.0
0,8
0.6
0,4
0,2
0,0
2.5
о ° о
г' = 0.583
10 20 30 40 Парциальные объемы, %
50 10 20 30 40 Парциальные объемы, %
Рис. 11.9. М е ж в и д о в ы е корреляции м е ж д у с т р у к т у р н ы м и х ар акт ер ист иками листьев и н а с ы щ а ю щ и м с о д е р ж а н и е м воды ( Н С В ) у од но до ль ны х (а, б, в) и двудольных (г,
д, ё) растений .
На осях абсцисс парциальные объемы эпидермиса (а , г), паренхимной обкладки пучков (6), губчатой паренхимы (Э), межклетников (в, ё).

Глава 11 . М Е Ж В И Д О В Ы Е С Т Р У К Т У Р Н О - Ф У Н К Ц И О Н А Л Ь Н Ы Е К О Р Р Е Л Я Ц И И 243
в) у однодольных, а т а к ж е с проводящей т к а н ь ю (рис . 11.8, 6-2) и губчатой паренхимой (рис. 11.8, г) — у двудольных. Однако эти связи вряд ли и м е ю т с у щ е с т в е н н о е и непосредственное влияние на скорость испарения воды.
В о д н ы й дефицит . Д л я оценки водного д е ф и ц и т а в л и с т ь я х р а с т е н и й м ы использовали н а с ы щ а ю щ е е с о д е р ж а н и е воды ( о т н о ш е н и е к о л и ч е с т в а в о д ы , п о п а в ш е г о в листья в процессе н а с ы щ е н и я , к их сухой массе) . Для видов о д н о д о л ь н ы х растений выявлена достаточно тесная корреляция между п а р ц и а л ь н ы м о бъемо м э п и д е р м и с а и н а с ы щ а ю щ и м с о д е р ж а н и е м воды (рис . 11.9, а). Эта связь показывает , что у видов со сравнительно н е б о л ь ш и м ( м е н ь ш е 7%) п а р ц и а л ь н ы м о б ъ е м о м э п и д е р м и с а наблюдаются наиболее в ы с о к и е значения н а с ы щ а ю щ е г о содержания воды. У в и д о в двудольных растений парциальный объем эпидермиса не связан с н а с ы щ а ю щ и м содержанием воды (рис. 11.9, г). О тсутствие такой зависимости , возможно , свидетельствует о том, что эпидермис не является единственной тканью, накапливающей воду, она достаточно свободно проникает и в ткани мезофилла .
Для однодольных р а с т е н и й обнаружена связь м е ж д у п а р ц и а л ь н ы м о бъемо м паренхимн ой обкладки пучков и этим показателем водного р е ж и м а (рис . 11.9, б). Ч е м больше (в относительном выражении) развита паренхимная обкладка , т е м больше нас ы щ а ю щ е е содержание воды. М о ж н о предположить , что вода, п о с т у п а ю щ а я в листья в процессе н а с ы щ е н и я , концентрируется п р е и м у щ е с т в е н н о в клетках п а р е н х и м н ы х обкладок пучков . Это т е м более вероятно, что ни с какими д р у г и м и т к а н я м и м е з о филла корреляции н а с ы щ а ю щ е г о содержания воды обнаружено не бы ло .
Для двудольных р а с т е н и й характерно наличие коррелятивной связи м е ж д у пар ц и а л ь н ы м о б ъ е м о м губчатой п а р е н х и м ы и н а с ы щ а ю щ и м с о д е р ж а н и е м в о д ы (рис . 11.9, д). Поскольку не б ы л о обнаружено корреляций этого показателя водного р е ж и ма с другими т к а н я м и мезофилла , то можно предположить , что у дву до ль ны х р аст е ний вода в п р о ц е с с е н а с ы щ е н и я листьев попадает п р е и м у щ е с т в е н н о в губчатую паренхиму, которая и играет р о л ь основного ее накопителя .
П о м и м о этого о б н а р у ж е н а отрицательная корреляция н а с ы щ а ю щ е г о с о д е р ж а н и я воды с парциальными объемами межклетников как для однодольных (рис . 11.9,в), так и для двудольных (рис. 11.9, е) растений . Отрицатель ны е к о р р е л я ц и и н а с ы щ а ю щ е г о содержания воды с парциальными объемами межклетников мо жно объяснить тем, что вода в процессе н а с ы щ е н и я п е р е м е щ а е т с я по о с м о т и ч е с к и м г р а д и е н т а м вакуолизи-рованных тканей , минуя м е ж к л е т н и к и . Ч е м м е н ь ш е р а з в и т ы м е ж к л е т н и к и и б о л ь ш е эти ткани , тем большие о б ъ е м ы потоков воды они могут пропустить .
С о д е р ж а н и е воды в л и с т ь я х . Д л я однодольных в ы я в л е н а очень т е с н а я зависимость (рис. 11.10, а) содержания воды от парциального объема всего (межклетников + воздушных полостей) свободного пространства листьев . У двудольных оводненность листьев в наибольшей степени зависит от объема межклет нико в губчатой п а р е н х и м ы (рис. 11.10, г-1) и несколько м е н ь ш е от всего объема м е ж к л е т н и к о в (рис . 11.10, г-2). Эти факты показывают, что оводненность листьев связана п р е и м у щ е с т в е н н о со свободным пространством мезофилла . К р о м е того, выявляется очень тесная отрицательная з ависимость содержания воды в листьях однодольных (рис . 11.10, б) и двудольных (рис . 11.10, д) растений от соответствующих п а р ц и а л ь н ы х о б ъ е м о в п р о в о д я щ е й ткани мезофилла . Д л я однодольных растений характерно т а к ж е н а л и ч и е отрицатель ной связи м е ж д у о в о д н е н н о с т ь ю листьев и губчатой п а р е н х и м о й (рис . 11.10, в) , для
9*

244 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й , .
7
6
5
2
1
О
г^= 0.759
4 -
3 •
2 •
О 20 40 60 80 О 10 20 30 40
б 5 1
4
3
2
1
—. О 20 О
г' = 0.684
8 10
7 п
6
5
ИЗ
^^ = 0.648
о о
10 20 30 40 Парциальные объемы, %
5 т
4
3 •
2
1 = 0.473
- , О 50 2 4 б
Парциальные объемы, %
Рис. 11.10. З ави симость содержания воды от п а р ц и а л ь н ы х о б ъ е м о в т к а н е й м е з о ф и л ла и свободных пространств в листьях однодольных (а, б, в) и дву до ль ны х (г, д, е)
растений .
На осях абсцисс парциальные объемы межклетников и воздушных полостей (а), проводящих тканей (б, д), губчатой паренхимы (в), межклетников губчатой паренхимы (г-1), межклетников
(г-2), паренхимной обкладки пучков (е).

Глава 11. М Е Ж В И Д О В Ы Е С Т Р У К Т У Р Н О - Ф У Н К Ц И О Н А Л Ь Н Ы Е К О Р Р Е Л Я Ц И И 245
5,5 -,
4,5 •
3,5
2,5 •
1,5
0,5
4,5 -,
3,5 •
2,5 •
1,5 Г =0.749
0,5
г' =0.687
10 15 20 О 8 10
4,5 ^
3,5 -
« 2,5
1.5
0,5
4,5 -,
0,5 10 20 30 40 50 О 10 15
4,5 -I
20 40 60 Парциальные объемы, %
0,5
г' = 0.628
80 10 20 30 40 Парциальные объемы, %
Рис. 11.11. Связь осмотического давления клеточного сока с п а р ц и а л ь н ы м и объемами тканей м е з о ф и л л а и с в о б о д н ы х пространств в листьях о д н о д о л ь н ы х (а, б, в) и
двудольных (г, д, е) растений .
На осях абсцисс парциальные объемы проводящих тканей (а, г), губчатой паренхимы (б), воздушных полостей и межклетников (в), паренхимной обкладки пучков проводящих тканей
и паренхимной обкладки пучков (Э-2), межклетников (е).

246 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й .
двуд ольн ых — м е ж д у с о д е р ж а н и е м воды и п а р е н х и м н о й о б к л а д к о й п у ч к о в (рис . 11.10, е). Чем большее развитие в м е з о ф и л л е п о л у ч а ю т п е р е ч и с л е н н ы е ткани , т е м меньше содержание воды в листьях (и больше удельная масса сухого вещества ) .
О с м о т и ч е с к о е д а в л е н и е клеточного сока. В ы я в л е н ы достаточно т е с н ы е статистические связи м е ж д у п а р ц и а л ь н ы м и объемами п р о в о д я щ и х тканей (без учета паренхимной обкладки пучков) и осмотическим давлением клеточного сока у однодольных (рис. 11.11, а) и двудольных (рис. 11.11, г) растений . У видов с наиболее в ы с о ким п арц и альн ым объемом проводящей ткани наблюдаются и более высокие величины осмотического давления . Для видов со слабо р аз вит ыми пучками характерно низкое осмотическое давление .
Для одн одольных р а с т е н и й выявлена тесная статистическая связь (рис . 11.11, б) п а р ц и а л ь н ы х о б ъ е м о в губчатой п а р е н х и м ы с о с м о т и ч е с к и м д а в л е н и е м клеточного сока. У видов однодольных с наиболее развитой губчатой п а р е н х и м о й могут наблюдаться и наиболее в ы с о к и е величины осмотического давления . В отличие от однодольных, для двудольных р а с т е н и й характерна статистическая связь (рис . 11.11, (3-1) между п а р ц и а л ь н ы м и объемами паренхимных обкладок пучков и этим показателем водного режима .
С учетом о б с у ж д а в ш е й с я в ы ш е связи м е ж д у о с м о т и ч е с к и м д а в л е н и е м и парциальным объемом п р о в о д я щ е й ткани (рис. 11.11, г), м о ж н о сказать , что весь пучок (включая флоэму, ксилему и п а р е н х и м н у ю обкладку) двудольных р а с т е н и й является зоной высокого осмотического давления (рис. 11.11, д-2). К р о м е того, для этой группы видов не о б н а р у ж е н о статистических связей осмотического д а в л е н и я с д р у г и м и тканями мезофилла (в частности , с хлоренхимой) . М о ж н о поэтому предположить , что у двудольных р а с т е н и й р а с т в о р и м ы е (т. е. о с м о т и ч е с к и а к т и в н ы е ) в е щ е с т в а локализуются п р е и м у щ е с т в е н н о в проводящих пучках и их обкладках .
Как для однодольных, так и для двудольных растений выявлены статистические связи осмотического д а в л е н и я клеточного сока с п л о т н о с т ь ю с л о ж е н и я мезофилла . М е р о й п лот н ост и с л о ж е н и я м е з о ф и л л а у однодольных с л у ж и т с у м м а п а р ц и а л ь н ы х объемов межклетников и воздушных полостей, у двудольных — п а р ц и а л ь н ы е объемы межклетников . Чем больше свободных пространств в листьях растений , тем менее п лот н о с л о ж е н м е з о ф и л л , и наоборот. П л о т н о с т ь м е з о ф и л л а о т р а ж а е т степень реализации р а с т я ж е н и я листьев , она зависит только от в о д о с н а б ж е н и я . В условиях водного д е ф и ц и т а реализация растяжения очень низкая, вследствие чего наблюдается почти полное отсутствие межклетников и плотный мелко клет ный мезофилл . Это связано с р а с п о л о ж е н и е м видов на градиенте в л а ж н о с т и п о ч в ы . В и д и м о поэтому в листьях однодольных (рис. 11.11, в) и двудольных (рис . 11.11, е) р а с т е н и й с наиболее плотным с л о ж е н и е м м е з о ф и л л а наблюдаются м а к с и м а л ь н ы е в е л и ч и н ы осмотического д а в л е н и я клеточного сока , а в листьях р а с т е н и й с р ы х л ы м м е з о ф и л л о м — минимальные .
Обобщение структурно-функциональных корреляций
Н е с м о т р я на з аметные различия м е ж д у системами с т р у к т у р н ы х корреляций однодольных и двудольных растений (рис. 11.7), с т р у к т у р н о - ф у н к ц и о н а л ь н ы е корреляции у этих групп видов и м е ю т много общего (рис . 11.12).

Глава 11 . М Е Ж В И Д О В Ы Е С Т Р У К Т У Р Н О - Ф У Н К Ц И О Н А Л Ь Н Ы Е К О Р Р Е Л Я Ц И И 247
И н т е н с и в н о с т ь т р а н с п и р а ц и и и содержание воды у обеих г р у п п р а с т е н и й связаны в основном со свободными пространствами листьев . Это неоднозначно свидетельствует о том, что вода (не в жидкой фазе) л о к а л и з о в а н а в о с н о в н о й своей массе (до 67-70% свободной воды) в межклеточных пространствах . С а м а во з мо жно ст ь поддержания достаточно и н т е н с и в н о й т р а н с п и р а ц и и может з ависеть от количества паров воды в межклетниках . Так, А. А. Зялалов (1984) отмечает, что при по ниженно й влажности межклетников падает проводимость , затормаживается п о с т у п л е н и е воды к ус тьицам и околоустьичным клеткам и, следовательно, снижается интенсивность транспирации (с другой стороны, локализация большого количества свободной воды в них может свидетельствовать также о том, что попадая в симпласт по о с м о т и ч е с к и м градиентам , вода б ы с т р о переходит в связанную форму) . Л о к а л и з а ц и я большого количества воды в межклетниках , т а к и м , образом, должна приводить к очень т е с н о й взаи м о з а в и с и м о с т и м е ж д у о в о д н е н н о с т ь ю л и с т ь е в и т р а н с п и р а ц и е й в п о д а в л я ю щ е м большинстве случаев . Кроме того , такая локализация зависит от пло т но ст и сложения а с с и м и л и р у ю щ и х органов . Поэтому различия м е ж д у о д н о д о л ь н ы м и и д в у д о л ь н ы м и как по с о д е р ж а н и ю воды в листьях , так и по скорости ее и с п а р е н и я следует искать в разной степени с к л е р о м о р ф н о с т и листьев этих г р у п п растений .
Развитие водного д е ф и ц и т а на градиенте в л а ж н о с т и почвы, по-видимому, является з е р к а л ь н ы м от ражением п р о ц е с с а н а с ы щ е н и я . Е с л и пулами н а с ы щ е н и я однодольных р а с т е н и й являются клетки п а р е н х и м н о й о б к л а д к и пучков (рис . 11.9, б) , а д в у д о л ь н ы х — губчатая п а р е н х и м а (рис . 11.9, д), то и потери воды при фо р мир о ва нии водного д е ф и ц и т а д о л ж н ы происходить п р е и м у щ е с т в е н н о из этих тканей , вслед за первичной ее потерей из апопласта .
В разных частях градиента влажности почвы м о жно наблюдать разное соотношение тканей мезофилла и соответственно разное развитие тканей, выступающих в роли осмотически акт и вн ых пулов н а с ы щ е н и я водой. Это создает о п р е д е л е н н ы е предпосылки для ф о р м и р о в а н и я водного д е ф и ц и т а . Если эти п р е д п о с ы л к и находятся в соответствии с у с л о в и я м и у в л а ж н е н и я экотопов , то они будут р е а л и з о в а н ы . То есть , п е р в и ч н ы м фактором для ф о р м и р о в а н и я водного д е ф и ц и т а является , судя по всему, анатомическое строение мезофилла , которое опосредует воздействие условий увлажнения экотопов н а этот п р о ц е с с водного р е ж и м а р а с т е н и й , а т а к ж е р а с п р е д е л е н и е возникающих потоков м е ж д у симпластом и апопластом.
В то ж е время , в к р а й н е а р и д н ы х м е с т о о б и т а н и я х р а з в и т и е водного д е ф и ц и т а может идти как л а в и н о о б р а з н ы й п р о ц е с с во всех тканях л и с т ь е в и главным факт о ром этого процесса д о л ж н ы быть условия недостаточного почвенного водоснабжения. П о м и м о этого, количественные р а з л и ч и я по средним в е л и ч и н а м д е ф и ц и т а водного насыщения между однодольными и двудольными растениями могут быть обусловлены тем, что вода в процессе н а с ы щ е н и я поступает в пулы, которые значительно р а з л и чаются по своим в о з м о ж н о с т я м запасания : у однодольных р а с т е н и й — в п а р е н х и м ную обкладку пучков ( п а р ц и а л ь н ы е о б ъ е м ы 1-8%), а у дву до ль ны х — в губчатую паренхиму (парциальные о б ъ е м ы 10-25%).
Р а з л и ч и я м е ж д у однодольными и двудольными р а с т е н и я м и пр о я вля ю т ся в том, что разные ткани являются з о н а м и повышенного осмотического давления . Д л я однодольных р а с т е н и й это губчатая п а р е н х и м а (рис . 11.12, а), д л я д в у д о л ь н ы х — весь пучок (включая п а р е н х и м н ы е обкладки) . М о ж н о поэтому п р е д п о л о ж и т ь , что у дву-
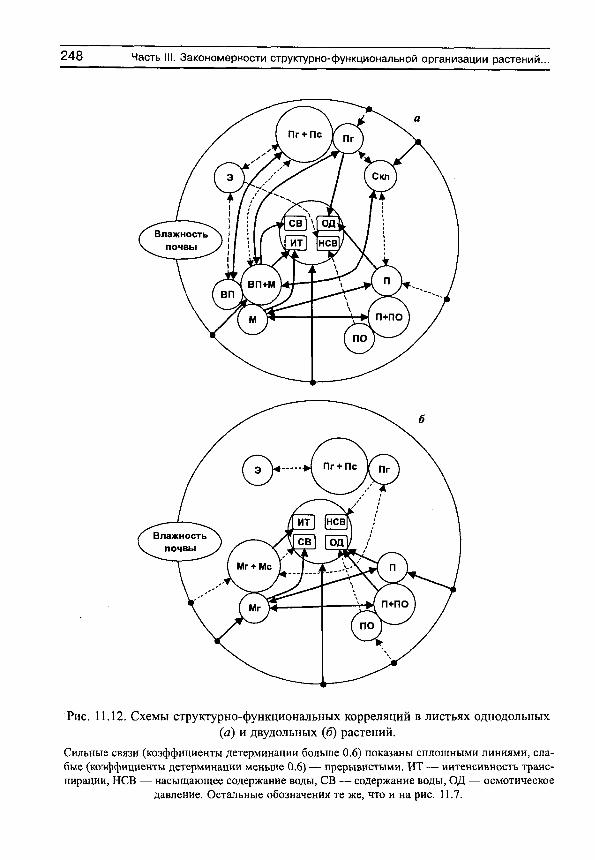
248 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й .
Рис. 11.12. Схемы структурно-функциональных корреляций в л и с т ь я х однодольных (а) и двудольных (б) растений .
Сильные связи (коэффициенты детерминации больше 0.6) показаны сплошными линиями, слабые (коэффициенты детерминации меньше 0.6) — прерывистыми. ИТ — интенсивность транспирации, НСВ — насыщающее содержание воды, СВ — содержание воды, ОД — осмотическое
давление. Остальные обозначения те же, что и на рис. 11.7.

Глава 11 . М Е Ж В И Д О В Ы Е С Т Р У К Т У Р Н О - Ф У Н К Ц И О Н А Л Ь Н Ы Е К О Р Р Е Л Я Ц И И 249
дольных р а с т е н и й р а с т в о р и м ы е (т. е. осмотически активные) в е щ е с т в а л о к а л и з у ю т ся преимущественно в проводящих пучках и их обкладках. Это может свидетельствовать о том, что количество образовавшихся в хлоренхиме ассимилятов примерно равно количеству ассимилятов , з агружаемому во флоэму пучков . У о дно до ль ны х расте ний, возможно , происходит некоторая задержка оттока и накопление ассимиля т о в в губчатой паренхиме , что и проявляется в соответствующей корреляции .
***
Полученные в ходе сравнительных исследований д а н н ы е обнаружили су щ ест вен ные различия состава тканей, структурной организ ации водных потоков и показате лей водного обмен а в п е р в у ю очередь д л я г р у п п однодольных и двудольных р аст е ний. Это провоцирует на более глубокий анализ р а з л и ч и й их ж и з н е н н ы х ф о р м . О ч е видно, что все изученные признаки связаны с особенностями морфогенеза этих групп растений. О д н о д о л ь н ы м свойственны интеркалярные по п о л о ж е н и ю м е р и с т е м ы , ин-теркалярный по этой п р и ч и н е р о с т н а д з е м н ы х органов, их о с е в о е (не латеральное на осях) заложение , и горизонтальный характер онтогенетического развития особей (горизонтальная модель с м е н ы метамеров ) . А п и к а л ь н ы й р о с т и р а з в и т и е двудольных, вытекающие из апикального положения меристем, со пр я жены с латеральным заложением листьев и побегов , с вертикальной м о д е л ь ю с м е н ы метамеров .
Листья однодольных не являются такими же, как листья двудольных, латеральными органами , о н и не г о м о л о г и ч н ы по п р о и с х о ж д е н и ю и не и д е н т и ч н ы по структуре . Сходство только ф у н к ц и о н а л ь н о е : в п а р е н х и м н ы х тканях тех и др у г их органов с и н тезируется хлорофилл , и они становятся в ходе развития ф о т о с и н т е з и р у ю щ и м и органами , хотя для одн одольных скорее о с е в ы м и , а д л я двудольных — бо ко выми . Базир у ю щ и й с я на р а з н о м положении меристем характер развития и с м е н ы фу нкцио нир у ю щ и х органов для них п р и н ц и п и а л ь н о различен . Р а з л и ч н а и о б щ а я схема развития транспортной системы, структура которой наиболее точно отражает с п е ц и ф и к у ж и з ненной формы.
Структ урн ые о с о б е н н о с т и т р а н с п о р т н о й системы у однодольных и двудольных растений , связанные с р а з н ы м характером ф о р м и р о в а н и я и с м е н ы их л и с т ь е в , уже обсуждались (Гамалей , 2002а , 20026). Реже о б с у ж д а е м ы м и и м е н е е п о н я т ы м и представляются структурные и ф у н к ц и о н а л ь н ы е последствия , в ы т е к а ю щ и е из о с о б е н н о стей морфогенеза о д н о д о л ь н ы х и двудольных.
Модель д и ф ф е р е н ц и а ц и и клеток и тканей дет ер минир о вана схемой р аспр еделе ния т р а н з и т н ы х д л я клеток т р а н с п о р т н ы х потоков , у п р а в л я ю щ и х д и ф ф е р е н ц и р о ванной э к с п р е с с и е й их г е н о м о в ( К о с Ь , 1996; Г а м а л е й , 1997, 2 0 0 2 а , 20026, 2004; Лутова и др . , 2000). С л е д о в а т е л ь н о , р а з л и ч и я о р г а н и з а ц и и т р а н с п о р т н о й с и с т е м ы сходных ф у н к ц и о н а л ь н о , но не гомологичных органов однодольных и двудольных, б а з и р у ю щ и е с я н а р а з н о м м о р ф о г е н е з е ( р а з н ы х м о д е л я х ф о р м и р о в а н и я и с м е н ы метамеров) , п р е д п о л а г а ю т появление еще большего спектра р а з л и ч и й гистогенеза , которые о к а з ы в а ю т с я в т о р и ч н ы м и , п р о и з в о д н ы м и о т н о с и т е л ь н о р а з л и ч и й м о р ф о генеза.
О с о б е н н о с т и гистогенеза в с в о ю очередь ведут к е щ е более д е т а л ь н ы м р а з л и ч и ям структуры и состава тканей , т и п о в специализ ации клеток , р а с п р е д е л е н и я пулов запасания воды и осмотиков , п р я м ы х и косвенных признаков о р г а н и з а ц и и водного

250 Ч а с т ь III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й .
обмена , которые и б ы л и о б н а р у ж е н ы в ходе п р о в е д е н н ы х и с с л е д о в а н и й . Э т о дает повод в е р н у т ь с я е щ е р а з к о б с у ж д е н и ю о с о б е н н о с т е й м о р ф о г е н е з а , г и с т о г е н е з а , структуры и ф у н к ц и о н и р о в а н и я проводящей системы и д р у г и х тканей о дно до ль ны х и двудольных растений в свете изложенных данных .
В ходе с р а в н и т е л ь н ы х исследований состава тканей установлен б о л ь ш и й парциальный объем х л о р е н х и м ы в листьях двудольных (рис. 11.1). О д н о в р е м е н н о у них найдена от ри ц ат ельная связь м е ж д у объемами м е з о ф и л л а и э п и д е р м ы , что вполне укладывается в и з в е с т н у ю модель р о с т а и особенно р а с т я ж е н и я л и с т ь е в двудольных . На эти факты р а н ь ш е как будто не о б р а щ а л о с ь в н и м а н и я , по крайней мер е нам не известны л и т е р а т у р н ы е д а н н ы е такого рода . М о ж н о ли и н т е р п р е т и р о в а т ь б о л ь ш и й объем х л о р е н х и м ы в л и с т ь я х двудольных как показатель их более высокой специализации как фотосинтетических органов? По-видимому, да . Эта т е н д е н ц и я прослеживается и вдоль ф и л о г е н е т и ч е с к и х л и н и й двудольных от исходных к более продвинут ы м таксонам .
В отличие от двудольных , для однодольных найдена отрицательная связь м е ж д у объемами э п и д е р м ы и межклетников . Это объяснимо о с е в ы м (не л а т е р а л ь н ы м ) нап р а в л е н и е м р а с т я ж е н и я ф о т о с и н т е т и ч е с к и х о р г а н о в , п р и к о т о р о м в с е т к а н е в ы е объемные с о о т н о ш е н и я оказываются более жестко з а в и с и м ы м и д р у г от друга , чем в латерально раст ягивающихся листьях двудольных.
Значительно б о л ь ш и й о б ъ е м о п о р н ы х элементов с к л е р е н х и м ы в л и с т ь я х однодольных растений свидетельствует о том, что их листовые органы сочетают фотосинт е т и ч е с к у ю ф у н к ц и ю не только с т р а н с п о р т н о й , но и с о п о р н о й . Э т о е с т е с т в е н н о вытекает из их осевой природы. П о л и ф у н к ц и о н а л ь н о с т ь тоже показатель более низ кой специализации . Развитие с к л е р е н х и м ы меняет с о о т н о ш е н и е всех о с т а л ь н ы х тканей не в направлении о п т и м и з а ц и и в е д у щ е й функции . На п р и м е р е о дно до ль ных это очевидно. В листьях двудольных отдельные склереиды могут встречаться внутри пучков, но никогда не о ф о р м л е н ы в тканевые т я ж и и не в л и я ю т сколько-нибудь с у щ е ственно на т кан евой состав .
Среди связей ф у н к ц и о н а л ь н ы х показателей водного о б м е н а с м о д е л ь ю развития и т к а н е в ы м с о с т а в о м зрелых а с с и м и л и р у ю щ и х органов о д н о д о л ь н ы х и двудольных обращает в н и м а н и е п о л о ж и т е л ь н а я связь и н т е н с и в н о с т и т р а н с п и р а ц и и с о б ъ е м о м межклетников у обеих групп изученных растений . Э т а связь понятна : м е ж к л е т н и к и и ф о р м и р у ю т с я в ходе развития листа вследствие водозапасания , в о з м о ж н о г о только в условиях достаточного или скорее даже избыточного водообеспечения , в противном случае , вакуолеобразование и растяжение л и с т а о ст аю т ся не р е а л и з о в а н н ы м и в полной мере . Структура , возникающая в таких условиях , известна как ксероморфная , не сочетаемая с высокими интенсивностями транспирации . Водозапасание , сопровождаемое р а с т я ж е н и е м клеток — в значительной степени ф и з и ч е с к и й п р о ц е с с , поэтому п р и н ц и п и а л ь н ы х р а з л и ч и й м е ж д у однодольными и д в у д о л ь н ы м и в этом о т но ш ении быть не может.
Н а с ы щ а ю щ е е содержание воды является одним из показателей, характеризующих водный д е ф и ц и т в листьях растений . Развитие водного д е ф и ц и т а н а градиенте влажности почвы, по-видимому, является з е р к а л ь н ы м отражением п р о ц е с с о в н а с ы щ е н и я , наблюдаемых в экспериментах . Если пулами н а с ы щ е н и я о д н о д о л ь н ы х р а с т е н и й я в ляются клетки п а р е н х и м н о й обкладки пучков, а двудольных — губчатая пар енх има .

Глава 11 . М Е Ж В И Д О В Ы Е С Т Р У К Т У Р Н О - Ф У Н К Ц И О Н А Л Ь Н Ы Е К О Р Р Е Л Я Ц И И 251
то и потери воды при формировании водного д е ф и ц и т а д о л ж н ы происходить преимущественно из этих тканей .
В разных частях градиента влажности почвы м о ж н о наблюдать разное соотношение тканей мезофилла и соответственно разное развитие тканей, в ы с т у п а ю щ и х в роли о с м о т и ч е с к и активных пулов н а с ы щ е н и я водой. Э т о создает о п р е д е л е н н ы е предпосылки для ф о р м и р о в а н и я водного д е ф и ц и т а . Если эти п р е д п о с ы л к и находятся в соответствии с у с л о в и я м и у в л а ж н е н и я экотопов , то они будут р е а л и з о в а н ы . То есть , п е р в и ч н ы м фактором для ф о р м и р о в а н и я водного д е ф и ц и т а является , судя по всему, анатомическое строение мезофилла , которое опосредует воздействие у с л о в и й увлажнения экотопов на этот процесс водного р е ж и м а р а с т е н и й . В то ж е время , в крайне аридных м е с т о о б и т а н и я х развитие водного д е ф и ц и т а м о ж е т идти как л а в и н о о б р а з ный процесс во всех тканях листьев и главным фактором этого процесса должны быть условия недостаточного почвенного водоснабжения .
Содержание воды в фотосинтезирующих органах тех и других растений коррелирует с объемом м е ж к л е т н о г о п р о с т р а н с т в а и со с т е п е н ь ю в а к у о л и з а ц и и клеток . У двудольных к наиболее вакуолизированным относятся клетки губчатой паренхимы или ее межпучковые фрагменты, названные паравейнальной пар енх имо й . У однодольных губчатой п а р е н х и м ы нет или о н а слабо в ы р а ж е н а в силу осевого ст р о ения фотосинтетических органов , поэтому связь ее развития с с о д е р ж а н и е м воды у них и не обнаружена, а если и обнаружена , то отрицательная .
Показатели осмотического давления у обеих групп растений связаны прежде всего с объемом п р о в о д я щ и х тканей . Это естественно , они отличаются на п о р я д о к более высок и ми к о н ц е н т р а ц и я м и з а г р у ж а е м ы х во флоэму и л и р а з г р у ж а е м ы х из к с и л е м ы осмотиков о т н о с и т е л ь н о их концентраций в п а р е н х и м н ы х т каня х . П о с р е д н и к а м и в процессах загрузки ф л о э м ы и разгрузки ксилемы, о с у щ е с т в л я ю щ и м и транзит, ч а щ е всего становятся клетки п а р е н х и м н о й обкладки пучков или вся губчатая паренхима . У двудольных такая их специализация более развита , отсюда и н а б л ю д а е м ы е связи (рис. 11.11) развития этих тканей с о с м о т и ч е с к и м потенциалом листа .
Связь осмотического давления клеточного сока с пло т но ст ь ю сложения мез о филла (объемом межклетников) , наблюдаемая на материале всех изученных групп расте ний, объясняется тем , что и тот и другой показатели напрямую зависят от количества доступной р а с т е н и я м воды. П л о т н о с т ь м е з о ф и л л а о т р ажает степень р а с т я ж е н и я л и стьев вследствие вакуолизации их клеток, и зависит она только от водоснабжения . В условиях водного д е ф и ц и т а реализация вакуолеобразования и растяжения низкая , как следствие отсутствие м е ж к л е т н и к о в , п л о т н ы й мелко клет ны й м е з о ф и л л , в ы с о к и й о с мотический потенциал , г а р а н т и р у ю щ и й р е а л и з а ц и ю р а с т я ж е н и я как только у с л о в и я водоснабжения это позволят.
Резюме . Р а с с м о т р е н н ы е признаки свидетельствуют о том, что с т р у к т у р н о - ф у н к ц и о н а л ь н ы е р а з л и ч и я м е ж д у о д н о д о л ь н ы м и и д в у д о л ь н ы м и б а з и р у ю т с я на р а з н о й модели развития их листьев . Л и с т ь я однодольных по характеру р а з в и т и я э в о л ю ц и о нировали как осевые органы интеркалярного развития , листья двудольных — как бо лее с п е ц и а л и з и р о в а н н ы е л а т е р а л ь н ы е органы, в к л ю ч е н н ы е в а п и к а л ь н у ю с и с т е м у развития побега. П о этой причине чем более продвинутые таксоны сравниваются , т е м контрастнее различия , и, вероятно , менее правомочно само сравнение . С к л а д ы в а е т ся о б щ е е впечатление в пользу более высокой э в о л ю ц и о н н о й с п е ц и а л и з а ц и и л и с т ь -

252 Часть III. З а к о н о м е р н о с т и с т р у к т у р н о - ф у н к ц и о н а л ь н о й о р г а н и з а ц и и р а с т е н и й .
ев двудольных, радикально отличающихся по характеру р а з в и т и я от о с е в ы х органов . А н а л о г и ч н ы е п о ф у н к ц и о н а л ь н о м у н а з н а ч е н и ю , н о не г о м о л о г и ч н ы е и м о р г а н ы однодольных в ходе эволюции приобретали листоподобную структурно-функциональную организацию, но все же остались по характеру развития о с е в ы м и органами. Этим и о б ъ я с н я ю т с я в с е н а б л ю д а е м ы е р а з л и ч и я состава т к а н е й и п о к а з а т е л е й водного обмена.

ЗАКЛЮЧЕНИЕ
Начиная это исследование более двадцати лет назад , м ы предполагали , что закономерности изменений средневидовых величин элементов водного о бмена вдоль градиента влажности почвы будут очень просты. М о ж н о б ы л о ожидать , что эти величины будут изменяться в строгом соответствии с и з м е н е н и я м и в л а ж н о с т и почвы. Однако это оказалось не совсем так .
Ш и р и н а зоны разброса средневидовых значений дефицита водного насы щ ения на градиенте в л а ж н о с т и п о ч в ы остается почти неизменной на всем его п р о т я ж е н и и за исключением наиболее з а с у ш л и в ы х экотопов , где она несколько у в е л и ч и в а е т с я . Н а большей части градиента т р е н д ы с р е д н е в и д о в ы х значений почти п а р а л л е л ь н ы оси абсцисс, что свидетельствует о незначительном влиянии в л а ж н о с т и почвы в этих условиях п р о и з р а с т а н и я н а д е ф и ц и т водного н а с ы щ е н и я . С у щ е с т в е н н о е у в е л и ч е н и е водного дефицита , соответствующее изменениям влажности почвы, происходит только в аридной части градиента .
Анализ д и н а м и к и с р е д н е в и д о в ы х значений осмотического д а в л е н и я клеточного сока на градиенте также показывает , что эти значения очень м е д л е н н о увеличиваются по мере аридизации условий произрастания , резко возрастая только в самой сухой части градиента .
Р а з б р о с с р е д н е в и д о в ы х в е л и ч и н и н т е н с и в н о с т и т р а н с п и р а ц и и на г р а д и е н т е влажности почвы имеет «клиновидный» характер. Узкая часть «клина» расположена в аридной части градиента , широкая — в гумидной . Сходство видов в сообществах по р е а л и з а ц и и п р о ц е с с а и с п а р е н и я (и , в и д и м о , п о г л о щ е н и я ) в о д ы и з м е н я е т с я с и з м е н е н и е м п о л о ж е н и я ф и т о ц е н о з о в н а г р а д и е н т е в л а ж н о с т и п о ч в ы . В н а и б о л е е суровых условиях произрастания, с минимальным количеством доступной почвенной влаги п р о и з р а с т а ю т в и д ы с очень п о х о ж и м и з н а ч е н и я м и п о к а з а т е л е й водного ре жима. По мере продвижения вдоль градиента в сторону увеличения влажности почвы сходство в и д о в п о с т о я н н о с н и ж а е т с я , д о с т и г а я м и н и м у м а в с а м ы х г у м и д н ы х экотопах.
В травяных сообществах аридных экосистем относительно высокое сходство реализации процесса транспирации у разных растений может быть обусловлено ж е с т к и м экотопическим отбором (через который проходит набор видов , в н а и б о л ь ш е й степени соответствующий д а н н ы м условиям произрастания) и слабой в ы р а ж е н н о с т ь ю конкурентных о т н о ш е н и й м е ж д у н и м и . Популяции , и м е ю щ и е т о л е р а н т н ы е ж и з н е н н ы е стратегии, и сп аряю т такое количество воды, которое д о с т у п н о и м в этих м е с т о о б и таниях. Кроме того растения подвергаются таким с т р у к т у р н ы м и /или ф у н к ц и о н а л ь ным перестройкам, которые позволяют и м эффективно сокращать т р а н с п и р а ц и о н н ы е потери воды. О н и могут переживать засушливые условия в безлиственном состоянии, сокращать п о в е р х н о с т ь и с п а р е н и я путем плотного свертывания лист ь ев , у м е н ь ш а т ь размеры л и с т ь е в и увеличивать их плотность , создавать с у щ е с т в е н н ы е запасы воды

254 С . Н . Ш е р е м е т ь е в . Травы на г р а д и е н т е в л а ж н о с т и почвы
в л и с т ь я х или побегах , усиливать о с м о т и ч е с к у ю р е г у л я ц и ю в о д о о б м е н а , и з м е н я т ь адекватно у с л о в и я м термодинамическое состояние воды.
В г у м и д н ы х м е с т о о б и т а н и я х , н е с м о т р я на то , что з апасы в о д ы в почве неогра н и ч е н н ы (или д о с т а т о ч н о о б и л ь н ы ) , в одном и том же с о о б щ е с т в е с о с у щ е с т в у ю т виды как с высокой , т а к и с низкой скоростью испарения воды при достаточно слабом экотопическом отборе , но при более н а п р я ж е н н ы х к о н к у р е н т н ы х о т н о ш е н и я х между в и д а м и . Д л и т е л ь н о е и у с п е ш н о е сосуществование в фитоценозе ценопопуля -ций с кон курен т н ыми стратегиями, и с п о л ь з у ю щ и х один и тот же р е с у р с , в о з м о ж н о только при р а з д е л е н и и и м и экологических н и ш (как в пр о ст р анст ве , так и во времени). Наблюдаемое разнообразие средних уровней испарения воды в г у м и д н ы х экотопах, судя по всему, и является отражением такой д и ф ф е р е н ц и а ц и и . Поскольку в ре зультате разделения н и ш в и д ы сохраняют д о м и н и р у ю щ е е п о л о ж е н и е в сообществах , то используемые и м и запасы почвенной влаги д о л ж н ы отвечать их к о н с т и т у ц и о н н ы м потребностям . П о э т о м у м о ж н о предположить , что у в е л и ч е н и е р а з н о о б р а з и я средневидовых в е л и ч и н скорости т р а н с п и р а ц и и на градиенте в л а ж н о с т и п о ч в ы является следствием у в е л и ч е н и я разнообразия видоспецифических реакций растений н а изменения ценотической обстановки .
О в о д н е н н о с т ь л и с т ь е в и м е е т такой же « к л и н о в и д н ы й » р а з б р о с с р е д н е в и д о в ы х величин н а г р а д и е н т е в л а ж н о с т и почвы, как и и н т е н с и в н о с т ь т р а н с п и р а ц и и . Узкая часть «клина» расположена в аридной части градиента, широкая — в гумидной . Сходство видов в с о о б щ е с т в а х по оводненности листьев и з м е н я е т с я с и з м е н е н и е м положения ф и т о ц е н о з о в на градиенте в л а ж н о с т и почвы. В н а и б о л е е с у р о в ы х у с л о в и я х произрастания , с м и н и м а л ь н ы м количеством д о с т у п н о й п о ч в е н н о й влаги произрастают в и д ы с очень похожими значениями этого показателя водного р е ж и м а . П о мере продвижения вдоль г р а д и е н т а в сторону увеличения в л а ж н о с т и почвы сходство видов постоянно с н и ж а е т с я , достигая м и н и м у м а в с а м ы х г у м и д н ы х экотопах (т. е. уве личивается р а з н о о б р а з и е , разброс средневидовых значений) . Это предопределяет то, что на протяжении практически всего градиента влажности почвы м о ж н о обнаружить виды с очень п о х о ж и м и значениями оводненности листьев .
Также как и в случае с и н т е н с и в н о с т ь ю т р а н с п и р а ц и и , в т р а в я н ы х с о о б щ е с т в а х аридных экосистем относительно высокое сходство с о д е р ж а н и я воды у р а з н ы х р ас тений может б ы т ь обусловлено ж е с т к и м экотопическим о т б о р о м и к р и т и ч е с к и м недостатком воды в почве (возможно, также , и слабой в ы р а ж е н н о с т ь ю конкурентных отношений м е ж д у н и м и ) . П о п у л я ц и и , и м е ю щ и е т о л е р а н т н ы е ж и з н е н н ы е стратегии, накапливают такое количество воды, которое д о с т у п н о и м в этих м е с т о о б и т а н и я х и в соответствии со своими структурными особенностями. Суккуленты и м е ю т очень высокую о в о д н е н н о с т ь своих а с с и м и л и р у ю щ и х о р г а н о в . У с к л е р о ф и т о в , однако, ограниченные и одинаковые д л я всех особей р е с у р с ы воды в почве в ы р а в н и в а ю т средние уровни с о д е р ж а н и я воды. В и д о с п е ц и ф и ч н о с т ь среди них п о этому признаку не проявляется или практически отсутствует.
В гуми дн ых м е с т о о б и т а н и я х , несмотря на то , что з апасы воды в почве неогран и ч е н н ы (или д о с т а т о ч н о о б и л ь н ы ) , в одном и том же с о о б щ е с т в е с о с у щ е с т в у ю т виды, как с высокой , т а к и с низкой о в о д н е н н о с т ь ю л и с т ь е в п р и до ст ат о чно слабом экотопическом отборе , но при более н а п р я ж е н н ы х ко нку р ент ны х о т н о ш е н и я х м е ж д у видами, что способствует д и ф ф е р е н ц и а ц и и экониш.

З А К Л Ю Ч Е Н И Е 255
Эти факты заставляют нас предположить , что почти на всем п р о т я ж е н и и градиента влажности почвы нет местообитаний, которые принято называть «стрессовыми» . Экологические условия этих местообитаний , как правило , соответствуют физиологич е с к и м н о р м а м р е а к ц и й р а с т е н и й - д о м и н а н т о в и эдификаторов с о о б щ е с т в . В и д ы в каждом местообитании полностью соответствуют экологической и фитоценотической обстановке . И з м е н е н и я интенсивности или степени проявления п р о ц е с с о в функционирования , с и с т е м н о с т и (сложности организации , ф о р м и р о в а н и я и р а з в и т и я регуля-торных явлений , в заимосвязи функций) в направлении как ар идиз ации , т а к и гуми-дизации условий п роизрастания отражают скорее т е н д е н ц и и э в о л ю ц и о н н ы х изменений растений по мере освоения и м и новых экологических н и ш .
Эти изменения могут происходить под одновременным давлением экотопического отбора и конкурентных о т н о ш е н и й между р а с т е н и я м и . Однако сила их совместного действия неодинакова в р а з н ы х частях г р а д и е н т а в л а ж н о с т и п о ч в ы (рис . 12.1). В аридной части велик пресс экотопического отбора, н о н а п р я ж е н н о с т ь ко нку р ент ны х отношений находится на м и н и м а л ь н о м уровне . Н апр о т ив , в г у м и д н о й части градиента усиливается конкурентное взаимодействие растений, но напряженность экотопического- отбора снижается . В средней части градиента совместное действие этих процессов м и н и м а л ь н о .
Можно предположить, что именно в таких условиях, «не гумидных и не аридных» произрастали наименее специализированные травы, у которых бьши все потенции для структурно-функциональной специализации в аридных экотопах, и для завоевания разнообразных н и ш — в г у м и д н ы х , что также требовало у величения с п е ц и а л и з а ц и и в д и ф ф е р е н ц и р о в а н н о м использовании почвенной влаги. Таким образом , зоной нача-
Гумидные экотопы Аридные экотопы
Р и с . 12.1. Н а ч а л о э к с п а н с и и т р а в я н и с т ы х р а с т е н и й , в о з м о ж н о , б ы л о п р и у р о ч е н о к м е с т о о б и т а н и я м , где с о в м е с т н о е д е й с т в и е экотопического о т б о р а и к о н к у р е н ц и и
м и н и м а л ь н о .
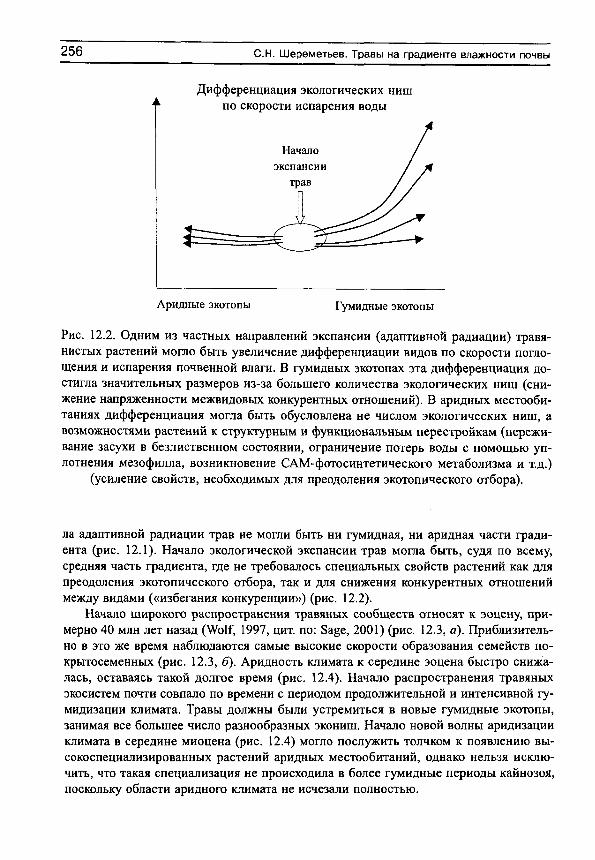
256 С . Н . Ш е р е м е т ь е в . Травы н а г р а д и е н т е в л а ж н о с т и почвы
Д и ф ф е р е н ц и а ц и я экологических н и ш п о скорости испарения воды
Аридные экотопы Гумидные экотопы
Рис. 12.2. О д н и м из ч а с т н ы х направлений экспансии ( адаптивной р а д и а ц и и ) травянистых растений могло б ы т ь увеличение д и ф ф е р е н ц и а ц и и в и д о в п о скорости погао-щения и испарения п о ч в е н н о й влаги. В г у м и д н ы х экотопах эта д и ф ф е р е н ц и а ц и я достигла зн ач и т ельн ых р а з м е р о в из-за большего количества экологических н и ш (снижение напряженности м е ж в и д о в ы х конкурентных отношений) . В а р и д н ы х местообитаниях д и ф ф е р е н ц и а ц и я могла быть обусловлена не число м э к о л о г и ч е с к и х н и ш , а возможностями р а с т е н и й к с т р у к т у р н ы м и ф у н к ц и о н а л ь н ы м п е р е с т р о й к а м (переживание засухи в б е з л и с т в е н н о м состоянии , о граничение потерь воды с п о м о щ ь ю у п лотнения м е з о ф и л л а , в о з н и к н о в е н и е С А М - ф о т о с и н т е т и ч е с к о г о м е т а б о л и з м а и т.д.)
(усиление свойств , необходимых для преодоления экотопического отбора) .
ла адаптивной р а д и а ц и и трав не могли быть ни гумидная , ни аридная части градиента (рис . 12.1). Н а ч а л о экологической экспансии т р а в могла б ы т ь , судя по всему, средняя часть г р а д и е н т а , где не т р е б о в а л о с ь с п е ц и а л ь н ы х свойств р а с т е н и й как д л я преодоления экотопического отбора, так и для с н и ж е н и я к о н к у р е н т н ы х о т н о ш е н и й между видами («избегания конкуренции») (рис. 12.2).
Начало ш и р о к о г о р а с п р о с т р а н е н и я травяных с о о б щ е с т в о т н о с я т к эоцену, п р и мерно 40 млн лет назад (ХУоЩ 1997, цит. по: 8а§е , 2001) (рис. 12.3, а). Приблизитель но в это же время н а б л ю д а ю т с я с а м ы е высокие скорости образования семейств п о крытосеменных (рис . 12.3, б) . А р и д н о с т ь климата к сер едине э о ц е н а быст р о снижалась , оставаясь такой д о л г о е в р е м я (рис . 12.4). Н а ч а л о р а с п р о с т р а н е н и я т р а в я н ы х экосистем почти совпало по времени с периодом продолжительной и интенсивной гу-мидизации кли мата . Травы д о л ж н ы б ы л и у с т р е м и т ь с я в н о в ы е г у м и д н ы е экотопы, занимая все б о л ь ш е е число разнообразных экониш. Н а ч а л о новой в о л н ы аридизации климата в середи не м и о ц е н а (рис . 12.4) могло п о с л у ж и т ь толчком к п о я в л е н и ю выс о к о с п е ц и а л и з и р о в а н н ы х р а с т е н и й а р и д н ы х м е с т о о б и т а н и й , однако нельзя и с к л ю чить , что такая с п е ц и а л и з а ц и я не происходила в более г у м и д н ы е п е р и о д ы кайнозоя , поскольку области аридного климата не исчезали п о л н о с т ь ю .

З А К Л Ю Ч Е Н И Е 257
160 -,
120 •
I 80 •
I 40 -
80 60 40 20
10 -,
Е
Е
60 40 20 Возраст, млн, лет
РИ Р1е
Ра/ Ео а
Рис. 123. И з м е н е н и е числа с е м е й с т в п о к р ы т о с е м е н н ы х р а с т е н и й (а ) и скорость их образования (б) в к а й н о з о е ( сплошная кривая по д а н н ы м МиИег, 1981, п р е р ы в и с т а я
кривая по д а н н ы м Веп1оп, 1993).
Стрелками показано (1) начало широкого распространения многолетних трав (\Уо1Г, 1997, цит. по: За^е, 2001) и (2) сообществ с С^-растениями (8а§е, 2004).
Если сопоставить средние д л я семейств скорости и с п а р е н и я воды ' с их возрас том (по МиПег, 1981), то становится видна очень яркая т е н д е н ц и я у в е л и ч е н и я скорости по мере умен ьшения возраста таксонов (рис. 12.5). М о ж н о б ы л о б ы провести разнообразные статистические сопоставления , однако, у ч и т ы в а я некоторую неоднознач-
' В расчет принимались только те семейства, для которых имелось не меньше 400 наблюдений, а их представители встречались не менее, чем в 4-5 сообществах.
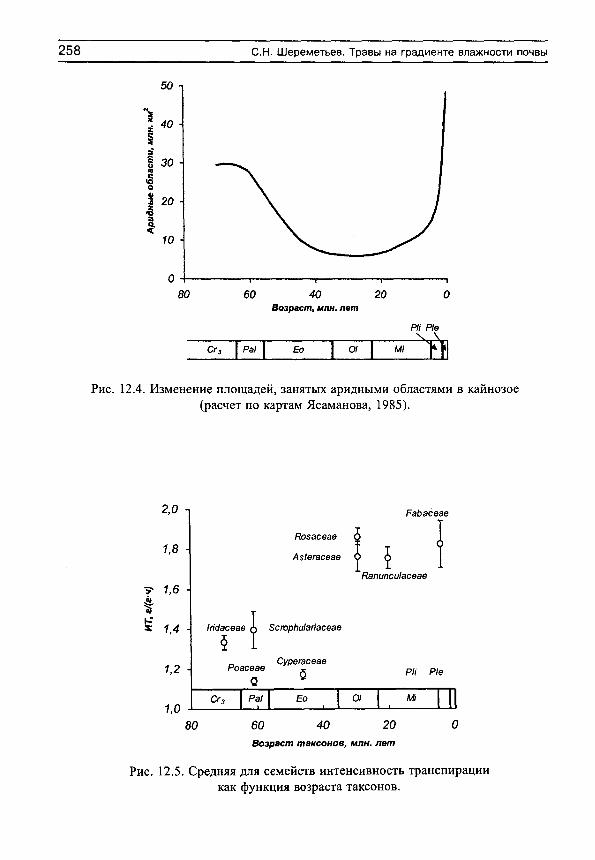
258 С . Н . Ш е р е м е т ь е в . Т р а в ы на г р а д и е н т е в л а ж н о с т и почвы
Рис . 12.4. И з м е н е н и е п л о щ а д е й , занятых а р и д н ы м и областями в кайнозое (расчет по картам Ясаманова , 1985).
2,0 -1
1,8 •
1,6 •
1,4 •
1,0
Шасвае 'асвае л
Новасеае
АаШасвэв
ЗсюрЫапасеав
РаЬасвае
НапипсШасвае
Роасеав Сурегасвае
РИ Р1в
Сг, Ра1 Ео 01
80 60 40 20
Возраст таксонов, млн. лет
Рис . 12.5. Средняя для семейств и н т е н с и в н о с т ь т р а н с п и р а ц и и как ф у н к ц и я возраста таксонов .

З А К Л Ю Ч Е Н И Е 259
Рис. 12.6. И з м е н е н и я содержания углекислоты в а т м о с ф е р е ( в ы р а ж е н н о й как отношение концентрации СО^ в п р о ш л о м к современной — КСО^) в кайнозое по д а н н ы м
(1) — В е т е г , 1998; (2) — ВеегИпё е{ а1., 1998; (3) — Сго\у1еу, Вегпег, 2001.
ность в определении времени появления семейств в палеонтологической летописи, это вряд ли целесообразно . О т м е т и м , например , что по интенсивности т р а н с п и р а ц и и семейства РаЬасеае и Капипси1асеае м о г у т б ы т ь з н а ч и т е л ь н о с т а р ш е , а 1гШасеае и 8сгорНи1апасеае— моложе , ч е м это указано в сводке I . МиИег (1981). Те м не менее , м ы можем констатировать, что у более молодых таксонов скорости транспирации значительно в ы ш е , чем у старых . Эти в ы с о к и е скорости с о п о с т а в и м ы п о возрасту с пе риодом гумидизации климата и, как м ы считаем, процессом экспансии трав в г у мидные экотопы (рис . 12.2, 12.4 и 12.5).
Но почему растения более молодых таксонов испаряли воду быстрее? П о д а н н ы м р а з н ы х а в т о р о в ( Б у д ы к о , Р о н о в , 1979; Б у д ы к о , 1980; В е е г 1 т § , \\ 'оо(1\уаг(1, 1997; В е е г 1 т § е1 а1., 1998; Вегпег, 1998; М с Е 1 \ у а т , 1998; Сго\у1еу, В е т е г , 2001; К о Л т а п , 2001, 2002; 2асЬо8 е! а1., 2001; К.е1а11аск, 2002 и др.) к о н ц е н т р а ц и я углекислого газа в атмосфере имеет х о р о ш о в ы р а ж е н н у ю т е н д е н ц и ю к с н и ж е н и ю (рис . 12.6). П о мнен и ю I . А. К.ауеп (2002) г е н е т и ч е с к и д е т е р м и н и р о в а н н ы й у с т ь и ч н ы й и н д е к с (и /или плотность) бьш подвержен изменениям со стороны в н е ш н и х факторов , особенно парциального давления СО^. П о э т о м у (по ископаемым л и с т ь я м ) наблюдается т е н д е н ц и я к у в е л и ч е н и ю п лот ности у с т ь и ц почти на всем п р о т я ж е н и и кайнозоя (рис . 12.7, а). О. I . ВеегИп§ и Р. I . \Уоод\уагс1 (1997), на основании ряда т е о р е т и ч е с к и х п р е д п о с ы л о к и э к с п е р и м е н т а л ь н ы х д а н н ы х (см. также В е е г 1 т § , Коуег, 2002) в о с с т а н о в и л и н е к и й о б о б щ е н н ы й ход и н т е н с и в н о с т и фотосинтеза и э ф ф е к т и в н о с т и испо ль з о вания воды

260 С . Н . Ш е р е м е т ь е в . Т р а в ы на г р а д и е н т е в л а ж н о с т и почвы
60 40 20 Возраст, илн. лет
60 40 20 Возрэст, млн, лет
Рис. 12.7. Предполагаемые изменения структурно-функциональных характеристик листьев растений в кайнозое : плотности расположения устьиц ( о г и б а ю щ а я кривая) (а), интенсивности фотосинтеза (б), эффективности использования воды ( Э И В ) (в), интен
с и в н о с т и т р а н с п и р а ц и и (г) (по д а н н ы м ВеегИп§, ХУоойлуагд, 1997).
Точками (г) показаны те же величины интенсивности транспирации, что и на рис. 12.5.
в фанерозое ( с о о т в е т с т в у ю щ и е участки кривых для кайнозоя п о к а з а н ы н а рис . 12.7, б и 12.7, в). О т с ю д а н е с л о ж н о рассчитать такой же о б о б щ е н н ы й ход и н т е н с и в н о с т и транспирации (рис . 12.7, г). Сопоставление этих расчетов с н а ш и м и д а н н ы м и показывает почти п о л н о е совпадение тенденций (рис. 12.7, г). Следовательно , увеличение скорости испарения воды (рис. 12.5, 12.7, г) могло быть обусловлено с н и ж е н и е м концентрации углекислоты в атмосфере и увеличением п л о т н о с т и р а с п о л о ж е н и я у с т ь и ц на листьях кайнозойских травянистых растений .
Развитие этой т е н д е н ц и и совпало с у в л а ж н е н и е м климата в эоцене и могло способствовать экспансии трав в г у м и д н ы е экотопы. Ш и р о к а я экспансия в а р и д н ы е местообитания происходила , возможно , в более позднее в р е м я и могла б ы т ь пр иу р о че на к периоду у с и л е н и я аридизации климата , начавшемуся в мио цене .

З А К Л Ю Ч Е Н И Е 261
Таким образом, м о ж н о предположить , что экспансия т р а в я н ы х с о о б щ е с т в началась из районов у м е р е н н о г о климата , средних по у с л о в и я м у в л а ж н е н и я (рис . 12.1, 12.2), во второй п о л о в и н е э о ц е н а (рис . 12.3) сначала в г у м и д н ы е области и затем, в середине миоцена, — в аридные. Адаптивная радиация во влажные местообитания сопровождалась д и ф ф е р е н ц и а ц и е й растений по степени использования почвенной влаги, направленной на снижение м е ж в и д о в о й конкуренции . П р о н и к н о в е н и е в а р и д н ы е экотопы стало в о з м о ж н ы м благодаря р а з в и т и ю свойств , о г р а н и ч и в а ю щ и х использо вание воды, необходимых для преодоления экотопического отбора .

ЛИТЕРАТУРА
Абачев К. Ю. 1990. О малом жизненном цикле Сагех сокМса (Сурегасеае) II Ботан. журн. Т. 75, № 11. С. 1547-1554.
Алексеев Ю. Е. 1996. Осоки (морфология, биология, онтогенез, эволюция). М. 251 с. Алексеенко Л. Н. 1976. Водный режим луговых растений в связи с условиями среды. Л. 198 с. Бабушкин Л. Н. 1976. Поглощение паров воды из межклеточного пространства листьев. Ки
шинев. 142 с. Багриновский К. А., Багинская Н. В., Баженова А. Ф. 1973. Математический анализ циркад-
ных систем организма на основании процедуры «Косинор» // Кибернетические подходы к биологии. Новосибирск. С. 196-209.
Байтулин И. О. 1979. Корневая система аридной зоны Казахстана. Алма-Ата. 184 с. Банникова И. А., Измайлова Н. Н., Максимович С. В. 1982. Водный режим и продуктивность
разнотравно-злаковой карагановой степи в Восточном Хангае (МНР) // Экология. № 3. С. 6-12.
Бейдеман И. Н. 1983. Справочник по расходу воды растениями в природных зонах СССР (геоботаническая и экологическая характеристики). Новосибирск. 256 с.
Бейдеман И. Н., Паутова В. Н. 1969. Водный режим растений на островах и берегах озера Байкал и методика его изучения // Тр. лимнол. ин-та. Т. 9 (29), вып. 2. 382 с.
Бигон М., Харпер Дж., Таунсенд К. 1989. Экология. Особи, популяции и сообщества. Т. 2. М. 477 с.
Бобровская Н. И. 1968. Краткая характеристика водного режима некоторых растений Центрально-Казахстанского мелкосопочника // Ботан. журн. Т. 53, № 10. С. 1400-1407.
Бобровская Н. И. 1985. Водный режим деревьев и кустарников пустынь. Л. 96 с. Бобровская Н. И. 1988. Водный режим. Тырсово-холоднополынно-житняково-змеевковое с
караганами сообщество // Сухие степи МНР. Ч. 2. Стационарные исследования (сомон Унджул). Л. С. 31-48.
Бобровская Н. И. 1991. Водный режим растений степей и пустынь Монголии. СПб. 154 с. Бобровская Н. И. 1994. О различных приспособлениях к засухе у доминантов степных и пу
стынных фитоценозов Монголии // Ботан. журн. Т. 79, № 5. С. 91-99. Бобровская Н. И., Попова Т. А. 1978. Сравнительная экофизиологическая и биологическая
характеристика АШит ро1угИпшп Тигсг. ех К.е§е1, А. топ^оНсит Ке§е1 // Пробл. освоения пустынь. № 1. С. 65-69.
Будыко М. И. 1980. Климат в прошлом и будущем. Л. 350 с. Будыко М. И., Ронов А. Б. 1979. Эволюция химического состава атмосферы в фанерозое//
Геохимия. № 5. С. 643-653. Буевич 3. Г. 1983. Жизненные формы растений // Горная лесостепь Восточного Хангая (МНР).
Природные условия (сомон Тэвшрулэх). М. С. 130-135. Вальтер Г. 1969. Растительность земного шара. Эколого-физиологическая характеристика. Т. 1.
Тропические и субтропические зоны. М. 551 с. Вальтер Г. 1974. Растительность земного шара. Эколого-физиологическая характеристика. Т. 2.
Леса умеренной зоны. М. 423 с. Вальтер Г. 1975. Растительность земного шара. Эколого-физиологическая характеристика. Т. 3.
Тундры, луга, степи, внетропические пустыни. М. 428 с.

Л И Т Е Р А Т У Р А 263
Василевич В.И. 1983. Очерки теоретической фитоценологии. Л. 248 с. Василевская В. К. 1979. Развитие экологической анатомии в СССР // Ботан. журн. Т. 64, № 5.
С. 654-664. Васильев Б. Р. 1988. Строение листа древесных растений различных климатических зон. Л.
208 с. Васильев С. В., Долгопольская Н. Л., Зархидзе В. А., Сергеев Г. Е. 1973. Квантильные диаг
раммы и их использование для характеристики особенностей динамики численности животных // Тр. ВИЗР В. 39. С. 107-118.
Васильев С. В., Левина С. И., Сергеев Г. Е. 1982. Система математико-статистических приемов для классификации сельскохозяйственных территорий по ряду динамических характеристик вредоносных объектов. М. 104 с.
Витко К. Р. 1963. Экологическая характеристика гырнецовой дубравы в южной Молдавии: Автореф. дис. ... канд. биол. наук. Кишинев. 18 с.
Вознесенский В. Л. 1977. Фотосинтез пустынных растений: Юго-Восточные Каракумы. Л. 256 с.
Гамалей Ю. В. 1983. Анатомическая характеристика ксерофитов Гоби// Комплексная характеристика пустынных экосистем Заалтайской Гоби. Пущине: Инст. эвол. морфол. и эколог животных. С. 40-45.
Гамалей Ю. В. 1984. Анатомия листа у растений пустыни Гоби/ / Ботан. журн. Т. 69, № 5. С. 569-584.
Гамалей Ю. В. 1988. Признаки ксероморфизма// Пустыни Заалтайской Гоби: Характеристика растений-доминантов. Л. С. 67-84.
Гамалей Ю. В. 1997. Надклеточная организация растений // Физиол. раст. Т. 44, № 6. С. 819-846.
Гамалей Ю. В. 2002а. Транспорт и распределение ассимилятов. Направления, подходы и методы исследований // Физиол. раст. Т. 49, № 1. С. 22-39.
Гамалей Ю. В. 20026. Экологическая дифференциация двудольных. I . Деревья и травы // Успехи совр. биол. Т. 122, № 1. С. 55-72.
Гамалей Ю. В. 2004. Транспортная система сосудистых растений (Происхождение, структура, функции, регуляция, разнообразие типов, эволюция и экологическая специализация, анализ специфики по таксономическим и эколого-географическим группам растений). СПб. (в печати).
Гамалей Ю. В., Шийрэвдамба I I . 1988. Структурные типы пустынных растений/ / Пустыни Заалтайской Гоби. Характеристика растений-доминантов. Л. С. 45-66.
Гамалей Ю. В., Глаголева Т. А., Кольчевский К. Г., Чулановская М. В. 1992. Экология и эволюция типов С^-синдрома в связи с филогенией семейств СкепоросИасеае и Роасеае II Ботан. журн. Т. 77, № 2. С. 1-12.
Генкель П. А. 1969. Научная деятельность Н. А. Максимова и его роль в создании экологической физиологии растений // Проблемы физиологии растений. М.: Наука. С. 306-331.
Гильманов Т. Г. 1978. Математическое моделирование биогеохимических циклов в травяных экосистемах. М. 168 с.
Гороновский И. Т., Назаренко Ю. П., Некряч Е. Ф. 1974. Краткий справочник по химии. Киев. 992 с.
Горшкова А. А. 1970. Эколого-морфологические особенности и водный режим степных растений Забайкалья: Автореф. дис. ... д-ра биол. наук. Новосибирск. 57 с.
Горшкова А. А. 1971. Экология водного режима степных растений Забайкалья/ / Экология флоры Забайкалья. Иркутск. С. 5-ИЗ.
Горшкова А. А. 1966. Биология степных пастбищных растений Забайкалья. М. 274 с. Горшкова А. А., Буркова В. М. 1955. Интенсивность транспирации у степных растений Бу
рят-Монгольской АССР // Ботан. журн. Т. 40, № 5. С. 709-715.

264 С . Н . Ш е р е м е т ь е в . Т р а в ы на г р а д и е н т е в л а ж н о с т и почвы
Горшкова А. А., Буркова В. М. 1962. Интенсивность транспирации травянистых растений Средней Сибири // Тр. Вост.-Сиб. фил-ла СО АН СССР. № 35. С. 66-85.
Горшкова А. А., Зверева Г. К. 1982. Экология степных сообществ Центральной Тувы // Степная растительность Сибири и некоторые черты ее экологии. Новосибирск. С. 1 9 ^ 1 .
Горшкова А. А., Яблокова Л. П., Назын-Оол О. А. 1982. Основные черты водного режима степных растений Тувы // Растительные сообщества Тувы. Новосибирск. С. 140-154.
Горышина Т. К. 1979. Экология растений. М. 368 с. Грубов В. И. 1982. Определитель сосудистых растений Монголии. Л. 443 с. Губанов И. А., Ганболд Э. 1983. Сосудистые растения // Флора Восточного Хангая (МНР). М.
С. 102-163. Губанов И. А., Киселева К. В., Новиков В. С , Тихомиров В. Н. 1990. Луговые травянистые
растения. Биология и охрана. М. 183 с. Джиллер П. 1988. Структура сообществ и экологическая ниша. М. 184 с. Добровольский Г. В., Шеремет Б. В., Афанасьева Т. В., Палечек Л. А. 1998. Почвы. Энцик
лопедия природы России. М.: АВР. 368 с. Еленевский А. Г. 1978. Систематика и география вероник СССР и прилежащих стран. М. 259 с. Емельянов И. П. 1976. Формы колебаний в биоритмологии. Новосибирск. 127 с. Емельянов И. П. 1986. Структура биологических ритмов человека в процессе адаптации. Но
восибирск. 180 с. Жатканбаев Ж. Ж. 1961. Транспирация и расход воды растениями-эдификаторами основных
сообществ в пустынных степях Центрального Казахстана: Автореф. дис. ... канд. биол. наук. Л. 17 с.
Зайцев Г. Н. 1984. Математическая статистика в экспериментальной ботанике. М. 424 с. Заугольнова Л. Б. 1974. Образование двух биоморф в ценопопуляции СаИит гиЛетсит \Уй1с1. //
Научн. докл. высш. школы. Биологические науки. № 8. С. 65-72. Зозулин Г. М. 1959. Подземные части основных видов травянистых растений и ассоциаций
плакоров Средне-Русской лесостепи в связи с вопросами формирования растительного покрова// Тр. Центрально-Черноземного гос. запов. Курск. Вып. 5. С. 1-314.
Зялалов А. А. 1984. Физиолого-термодинамический аспект транспорта воды по растению. М. 136 с.
Иванов Л. А., Силина А. А., Цельникер Ю. Л. 1953. О транспирации полезащитных полос в условиях Деркульской степи // Ботан. журн. Т. 38, № 2. С. 167-184.
Измайлова Н. Н. 1977. О водном режиме Апетша /п^Ша ^111(1. // Проблемы экологии, геоботаники, ботанической географии и флористики. Л. С. 207-214.
Измайлова Н. Н. 1978. Водный режим растений горной лесостепи Восточного Хангая // География и динамика растительного и животного мира МНР. М. С. 115-119.
Измайлова Н. Н. 1986. Водный режим // Степи Восточного Хангая. М. С. 99-126. Казанцева Т. И. 1986. Распределение и динамика продуктивности надземной фитомассы/ /
Пустыни Заалтайской Гоби. Ч. 1. М. С. 106-114. Кайбияйнен Л. К. 1984. Ритмологические и параметрические аспекты адаптации растений к
конкретным условиям среды // Адаптация древесных растений к экстремальным условиям среды. Петрозаводск: Кар. фил-ал АН СССР. С. 53-65.
Карамышева 3. В. 1969. Ареал и фитоценотическая характеристика АПетша/п§Ша \У11Ы. // Биокомплексная характеристика основных ценозообразователей Казахстана. Л. С. 116-118.
Касьянова Л. Н., Погодаева Н. Н. 1979. Транспирация и продуктивность растений Забайкалья. Новосибирск. 198 с.
Ковалев О. В. 2000. Эволюция С^-синдрома фотосинтеза у цветковых растений // Ботан. журн. Т 85. № 11. С. 7-20.
Копытова Л. Д. 1974. Запас воды в степных сообществах Забайкалья и ее расход на транспи-рацию: Автореф. дис. ... канд. биол. наук. Иркутск. 22 с.

Л И Т Е Р А Т У Р А 265
Копытова Л. Д., Горшкова А. А. 1977. Запас воды в сообществах и ее расход на транспира-цию растений // Экология и пастбищная дигрессия степных сообществ Забайкалья. Новосибирск. С. 53-94.
Куркин К. А. 1987. Опыт экологической классификации растительности пойменных лугов // Ботан. журн. Т. 72, № 12. С. 1605-1616.
Лавренко Е. М. 1978. О растительности степей и пустынь Монгольской Народной Республики // Пробл. освоения пустынь. № 1. С. 3-9.
Лакин Г Ф. 1980. Биометрия. М. 294 с. Лопатин В. Д. 1973. Определение относительной конкурентоспособности и ее изменение на
сеяных лугах и под воздействием удобрений // Ботан. журн. Т. 58, № 6. С. 841-847. Лутова Л. А., Проворов Н. А., Тиходеев О. Н., Тихонович И. А., Ходжайова Л. Т., Шишко
ва С. О. 2000. Генетика развития растений. СПб.: Наука. 538 с. Максимов Н. А. 1952. Избранные работы по засухоустойчивости и зимостойкости растений.
Т. I . Водный режим и засухоустойчивость растений. М. 576 с. Мелещенко С. Н. 1997. Система водного транспорта высшего растения и ее элементы. 2. Ме
тод определения упругих свойств стенок одиночных и находящихся в составе тканей клеток растений // Физиология растений. Т. 44, № 2. С. 256-263.
Мелещенко С. Н. 2001. О распространенных физических ошибках при описании водных потоков в системе почва-растение-атмосфера // Физиология растений. Т. 48, № 2. С. 298-305.
Нахуцришвили Г. Ш. 1972. Экология высокогорных травянистых растений и фитоценозов Центрального Кавказа: Автореф. дис. ... д-ра биол. наук. Тбилиси. 52 с.
Нобел П. 1973. Физиология растительной клетки (физико-химический подход). М. 288 с. Паутова В. Н. 1969. Транспирация растений лесных и степных сообществ на Большом Уш-
каньем острове и побережьях озера Байкал. М. С. 144-384. Паутова В. Н., Копытова Л. Д. 1970. Транспирация растений степных сообществ на Большом
Ушканьем острове (оз. Байкал) // Ботан. журн. Т. 55, № 5. С. 669-687. Полюшкин А. П., Горшкова А. А. 1979. Способность степных пастбищных растений к отра
станию // Биоморфология и продуктивность степных растений Забайкалья. Новосибирск. С. 50-116.
Попов Н. Н. 1971. Влияние освещенности на процессы водного обмена растений // Состояние воды и водный обмен у культурных растений. М. С. 154-157.
Попова Т. А. 1977. О биологии плотнодерновинных злаков {АШит ро1уггН12ит Тигсг., А. ЬШеп-1а1ит Р18сЬ.) Монголии // Проблемы экологии, геоботаники, ботанической географии и флористики. Л. С. 165-172.
Работнов Т. А. 1973. Факторы устойчивости наземных фитоценозов // Бюлл. МОИП. Отд. биол. Т. 78, вып. 4. С. 67-76.
Рачковская Е. И., Гунин П. Д. 1980. Комплексные стационарные исследования в Заалтайской Гоби // Пробл. освоения пустынь. № 2. С. 5-12.
Рачковская Е. И., Федорова И. Т. 1983. Растительные сообщества // Комплексная характеристика пустынных экосистем Заалтайской Гоби. Пущино: Инст. эвол. морфол. и экол. животных. С. 30-33.
Реуцкий В. Г. 1984. Адаптивная регуляция водообмена растений: Автореф. дис. ... д-ра биол. наук. Минск. 45 с.
Сабиров Г. 1982. Биолого-экологические особенности и водный режим основных кормовых растений Устюрта: Автореф. дис. ... канд. биол. наук. Ташкент. 19 с.
Савинкин А. П. 1973. Водный режим пустынных пастбищ и богарных посевов в подзоне бурых почв Казахстана: Автореф. дис. ... канд. биол. наук. Алма-Ата. 28 с.
Санжид Ж., Федорова И. Т. 1983. Динамика видового состава и особенности фенологичесюго развития растений пустынных сообществ // Комплексная характеристика пустынных экосистем Заалтайской Гоби. Пущино: Инст. эвол. морфол. и экологии животных. С. 33-37.

266 С . Н . Ш е р е м е т ь е в . Травы н а г р а д и е н т е в л а ж н о с т и почвы
Свешникова В. М. 1956а. Осмотическое давление у высокогорных растений/ / Труды АН ТаджССР. Т. 45. 54 с.
Свешникова В. М. 19566. О водном режиме почв и растений высокогорных лугов // Труды Бот. ин-та АН ТаджССР. Т. 18. С. 234-275.
Свешникова В. М. 1962. Водный режим растений и почв высокогорных пустынь Памира. Душанбе. 247 с.
Свешникова В. М. 1963. Материалы по водному режиму полыней Казахстана // Стационарные исследования растительности. Тр. БИН АНСССР. Сер. Ш. В. 15. С. 174-190.
Свешникова В. М. 1979. Доминанты казахстанских степей (эколого-физиологическая характеристика). Л. 192 с.
Свешникова В. М. 1988. Водный режим. Вострецово-бескильницевый чиевник // Сухие степи МНР. Ч. 2. Стационарные исследования (сомон Унджул). Л. С. 48-52.
Свешникова В. М. 1993. Водный режим дерновинных злаков Евразиатской степной области // Ботан. журн. Т. 78, № 8. С. 67-79.
Свешникова В. М., Бобровская Н. И. 1969. Водный режим // Биокомплексная характеристика основных ценозообразователей Центрального Казахстана. Ч. 2. Л. С. 126-127.
Свешникова В. М.., Бобровская Н. И., Чойжамц Б. 1971. Первые сведения о водном режиме растений южной части МНР // Ботан. журн. Т. 56, № 12. С. 1797-1805.
Серебрякова Т. И., Кагарлицкая Т. Н. 1972. Большой жизненный цикл и эволюционные отношения жизненных форм некоторых видов Уеготса Ь. секции Р^еиМу^таскшт КосЬ // Бюлл. МОИП, отд. биол. Т 77, № 6. С. 81-98.
Слейчер Р. 1970. Водный режим растений. М. 366 с. Слемнев Н.Н. 1988. Фотосинтез // Сухие степи МНР. Ч. 2. Стационарные исследования (со
мон Унджул). Л. С. 6-31. Слемнев Н. П., Болд Д., Казанцева Т. И., Федорова И. Т., Якунин Г. Н. 1983. Опыт повыше
ния продуктивности пастбищ остепненных пустынь в Заалтайской Гоби (МНР) // Ботан. журн. Т 68, № П. С. 1533-1538.
Слемнев Н. Н., Мокроносов А. Д. 1988. Объекты и некоторые методические особенности исследований//Пустыни Заалтайской Гоби: характеристика растений-доминантов. Л. С. 137-142.
Слемнев Н. Н., Цоож Ш. 1988. Дневные и вегетационные изменения фотосинтеза растений// Пустыни Заалтайской Гоби: характеристика растений-доминантов. Л. С. 142-159.
Титов Ю. В., Печенюк Е. В. 1990. Динамика травяной растительности поймы реки Хопер. Л. 140 с.
Тулашвили Н. И. 1976. Водный режим некоторых высокогорных растений Центрального Кавказа в разных экологических условиях: Автореф. дис. ... канд. биол. наук. Тбилиси. 25 с.
Убугунова В. И. 1999. Экологические условия формирования речных пойм Монголии и их свойств // Автореф. дис. ... д-ра биол. наук. Иркутск. 46 с.
Федорова И. Т. 1980. Растительность восточной части Заалтайской Гоби/ / Пробл. осв. пустынь. № 2. С. 46-57.
Федоскин Н. В. 1979. Морфологическая структура побегов степных растений // Биоморфология и продуктивность степных растений Забайкалья. Новосибирск. С. 13-39.
Цвелев Н. Н. 1988. Флора Хоперского государственного заповедника. Л. 191 с. Чойжамц Б. 1974. Краткая характеристика ковылей лесостепной зоны МНР // Структура и
динамика степных экосистем МНР. М. С. 156-160. Чойжамц Б. 1978. К экологии злаков Восточного Хангая (МНР): Автореф. дис. ... канд. биол.
наук. Л. 20 с. Шалыт М. С. 1950. Подземная часть некоторых луговых, степных и пустынных растений и
фитоценозов. Часть I . Травянистые и полукустарничковые растения и фитоценозы лесной (луга) и степной зон // Тр. БИН АН СССР Сер. 3. Геоботаника. Вып. 6. М.-Л. С. 205-442.

Л И Т Е Р А Т У Р А 267
Шереметьев С. Н. 1991. Экология водного режима пустынных, степных и луговых растений Монголии // Автореф. дис. ... д-ра биол. наук. СПб. 33 с.
Шереметьев С. Н. 2002. Водный режим травянистых растений на градиенте влажности почвы. I . Интенсивность транспирации // Ботан. журн. Т. 87, № 8. С. 46-68.
Шереметьев С.Н., Жаргалсайхан Л. 1990. Экология водного режима степных растений Монголии. Л. 110 с.
Шереметьев С. Н., Филатова И. И., Чеботарева К. Е., Нескрябина Е. С. 2001. Структурно-функциональная организация видов разнотравно-осокового сообщества поймы реки Хопер (Воронежская область) // Ботан. журн. Т. 86, № 7. С. 39-52.
Эдлефсен И. Е., Андерсон А. Б. С. 1966. Термодинамика почвенной влаги/ / Термодинамика почвенной влаги. Л. С. 5-273.
Юнатов А. А. 1950. Основные черты растительного покрова Монгольской Народной Республики // Тр. Монгольской комиссии АН СССР. В. 39. 223 с.
Юнатов А. А. 1954. Кормовые растения пастбищ и сенокосов МНР. М.-Л. 351 с. Якимова Е. П., Павлова Н. Е. 1989. О водном режиме степных растений Забайкалья в связи
с их происхождением, биоморфологическими и эколого-физиологическими особенностям и / / Ботан. журн. Т. 74, № 12. С. 1774-1779.
Ян Баочжень. 1960. Экология и водный режим некоторых сообществ и их эдификаторов сухих степей Центрального Казахстана: Автореф. дис. ... канд. биол. наук. Л. 22 с.
Ясаманов Н. А. 1985. Древние климаты Земли. Л. 296 с. АЬгат8 М. В., КиЬ15ке М. Е., Мо5(о11ег 8 .А. 1994. КеЫ^щ \уе1 апй сЗгу уеаг есорЬу51о1оёу Ю
81гис1иге 1п соп1та81т§ 1етрега1е {гее 8рес1е8 // Есо1о§у. Уо1. 75, № 1. Р. 123-133. Аскег1у В. В. , Яе1сЬ Р. В. 1999. Сопуег§епсе апё согге1айоп8 а т о п § каГ 512е апё йдпсИоп 1п зеей
р1ап18: а сотрагаНуе 1е51 и81п§ 1пс1ереп(1еп{ соп1га818 // А т . I . Во1. Уо1. 86, Ко. 9. Р. 1272-1281. ВавНйе В. , 81ре8 В . , Напп Т\щ I . Р. 1993. ЕСГес1 о{ веусге \уа1ег 51ге88 оп азресгв оГ
сга88и1асеап ас1с1 те1аЬо118т ш Хегохгсуох II Р1ап1 РЬу81о1. Уо1. 103, N0. 4. Р. 1089-1096. ВескеП К. Р. 1997. Рге88иге-Уо1ите апа1у818 о!" а гап§е оГ ро1к11оЬус1пс р1ап15 1тр11ез Ле ех181епсе
о^ пе§а11Уе Шг§ог т уе§е1;а{1уе сеИз // Апп. Во!. Уо1. 79, N0. 2. Р. 145-152. ВеегИп§ В . I . , \\^оо(1\уаг(1 Р. I . 1997. СЬап§е5 оГ 1ап(1 р1ап1 й т с Н о п оуег Л е РЬапего201с:
гесопз1гис1юпз Ьазес! оп Л е ГоззИ гесогй // Во1. I . Ь т . 8ос. Уо1. 124, N0. 2. Р. 137-153. Веег1т§ В. '\̂ оос1№агс1 Р I . , Ь о т а з М. Я., АУШв М. А., ди1ск XV. Р , УаШез Р I . 1998. ТЬе
1пПиепсе оГ СагЬопх^'егоиз ра1аеоа1то5рЬегез оп р1ап йтсйоп: ап ехрептеШа! апё тос1е1Ип§ аззеззтеШ // РЫ1. Тгапз. Я. 8ос. Ьопё. В. Уо1. 353, N0. 1365. Р 131-140.
ВеегНп§ В. 1., Яоуег В . Е. 2002. ЯеасЬпе а СО^ 81ёпа1 й о т ГоззИ з1ота{а // Кеш РЬу1о1. Уо1. 153. 188ие 3. Р 387-397.
В е т е г Я. Р. 1998. ТЬе сагЬоп сус1е апё СО^ оуег РЬапего201с Ите : 1Ье го1е оГ 1апс1 ркШз // РЫ1. Тгапв. Я. 8ос. Ьопй. В. Уо1. 353, N0. 1365. Р 75-82.
Вепеоп М. У (е±). 1993. ТЬе РоззИ Яесогс! 2. Ьопс1оп. 845 р. В1п§Ьат С , АтЬо^аз! В. , СогпеИззеп О. С , Еее I . К., На1Ьегё Р. 1982. 1п1'егепйа1 5Ш1811са1
т е Л о й з Гог езНтаПп^ апй сотраг:п§ со81пог рагате1ег8 // СЬгопоЫо1о§1а. Уо1. 9, N0. 4. Р 397^39.
Воуег 8. 1985. ^Vа^е^ {гапзроП// Апп. Яеу. Р1ап1 РЬу81о1. Уо1. 36. Р 473-516. Вгау Е. А. 1993. Мо1еси1аг гезропзез Ю \уа*ег йейск / / Р1ап1 РЬ1зуо1. Уо1. 103, N0. 4. Р 1035-
1040. Саппу М. I . 1995. Арор1а8Йс \уа1ег апё зо1и1е т о у е т е п ! : пе\ ш к з {'ог ап оМ зрасе // Апп. Яеу.
Р1ап1 РЬуз. Р1ап1 Мо1. В1о1. Уо1. 46. Р 215-236. СЬеипе V. N . 8., Тугее М. Т., Ват1у I . 1976. 8оте роззМе зоигсез оС еггог ш с1е1егт1пт§ Ьи1к
е1аз11с тойиЬ апё оЛег рагате1егз й о т ргеззиге-УоЫте сигуез оГ 8Ьоо18 апё 1еауе8 // Сап. I . Во1. Уо1. 54, N0. 8. Р 758-765.
Сго\у1еу Т :., В е т е г Я. А. 2001. СО^ ап(1 сИта^е сЬапёе // 8с1епсе. V. 292. N0. 5518. Р 870-872.

268 С . Н . Ш е р е м е т ь е в . Т р а в ы н а г р а д и е н т е в л а ж н о с т и почвы
Сгох(1а1е I . Ь. 1998. 81ота1а1 раПет^Пё 1п топосо1у1е(1опз: Тгас1е5сапИа аз а тос1е1 5уз1ет // }. Ехр. Во1. Уо1. 49. 8рес1а1 Тззие. Р. 279-292.
СгохйаЬ I . Ь. 2000. 8{ота1а1 ра«егп1п§ 1п Апехозрегтз // Апп. Во1. Уо1. 87, N0. 8. Р. 1069-1080.
Вмйо Н. 2002. Р о р и к и о п (1упат1С5 о̂ " с1оск-соп1го11её Ь1о1о§1са1 зресхез: т о й е Ь апй \уЬу С1гса(11ап гЬу1Ьт5 аге С1гсас11ап / / 1 . ТЬеог. В1о1. Уо1. 217, N0. 4. Р. 425-442.
Оа1е I . Е., 8и1сИ{'Ге I . К 1986. \Уа{ег ге1аиоп8 о? р1ап1 се11з // Р1ап{ рЬу81о1оёу Уо1. IX. \Уа1ег апй 8о1и1ез 1п р1ап18. Ес1.: 81е\уаг(1 Р. С. Ьопйоп. Р. 1^8.
Е(1\уагс15 О.. Е., РигЬапк Р. Т., На1сЬ М. В . , Озтопй С. В. 2001. Ша1 <1ое8 к 1аке 1о Ье С^? Ьеззопз й-от Ле е у о М о п оГ С, рЬо1о8уп1Ьез18 // Р1ап1 РЬуз1о1. Уо1. 125, N0. 1. Р. 46-49.
ЕЫеппеег I . Р . , СегИпё Т. Е. 1995. А1то8рЬепс СО^ ап<1 га410 оГ 1п1егсе11и1аг 1о атЫеШ СО^ сопсеШгаИопз 1П р1ап18 // Тгее РЬу51о1. Уо1. 15, N0. 2. Р. 105-111.
Ро\у1ег N . 1986. ТЬе го1е оС сотрей110п 1п р1ап1 соттип1Йе8 1п апй апА 5ет1апс1 гедхопз // Апп. Яеу. Есо1. 8уз1. Уо1. 17, Р 89-110.
Оат1ег Е., Ьаигеп! О. 1994. ЬеаГ апа1оту, 8рес1Г1с т а з з апс1 \уа1ег соп1еп1 1п соп§епепс аппиа1 ап<1 регепша1 ^газз 8рес1е8 // Ые\ РЬу1о1. Уо1. 128, N0. 4. Р. 725-736.
Оагп1ег Е., 8а1а8ег 1.-Е., ЕаигеШ С , 8оп1е Ь. 1999. Ке1а110пзЫрз ЬеЬуееп рЬо1о8уп1Ье818, пИгоёеп апй \еа{ 51гис1иге 1п 14 ^газз зресхез апс11Ье1г ёерепйепсе оп Л е Ьазхз о̂ " ехргез51оп // Не\РЬуЫ. Уо1. 143, N0. 1. Р 119-129.
ОгаЬат Е. А., NоЬе1 Р. 8. 1999. Роо1 \уа1ег ир4аке, каГ \уа1ег 81ога§е апс! ваз ехсЬап§е о!" а йезеЛ 5исси1еп1: 1трИса110пз 1ог гоо1 з у з к т гейипйапсу// Апп. Во1. Уо1. 84, N0. 2. Р. 213-223.
Ога1ап1 Ь., ВотЬеШ А. 2000. СоггеЬИоп ЬеПусеп \еа{ а^е апс! оЛег 1еаГ 1га115 1п (Ьгее Мейкег-гапеап таяи18 зЬшЬ 8рес1е8: ^ие^си8 Иех, РНШугеа 1аа/оИа апй а^Ш^ тсапш II Епу. Ехр. Во1. Уо1. 43, N0. 1. Р 141-153.
О п т е I . Р. 1979. Р1ат 81га1е§1е8 апй уе§е1а110п ргосезз. Ке\ Уогк. 222 р. НаШеге Р 1969. СЬгопоЫо1оёу// Апп. Кеу. РЬу81о1. Уо1. 31. Р 675-725. Нетапйег М. Ь., Раззаз Н. I . , 8 т и Ь Ь. О. 1999. С1опа1 апа1уз18 оГ ер1с1егта1 раПет1п8 с1ипп§
та12е 1еаГ ёеуе1ортеп1 // Веуе1ортеп1а1 В1о1о§у. Уо1. 216, N0. 2. Р. 646-658. Неггега А., Регпапйег М. О., Тахзта М. А. 2000. ЕИссХ оГ (1гои§Ь1 оп САМ аЫ \уа1ег ге1а110П8
т р1ап18 оГ Ререгот1а сагпеуаИи II Апп. Во1. Уо1. 86, N0. 3. Р. 511-517. 15Ыс1а N . . Капеко М., А11ас1а К. 1999. Вю1ое1са1 с1оск8 // РКА8. Уо1. 969. Ьзие 16. Р 8819-8820. I2та^1оуа N . N . 1985. В1е Яе8и1ип§ йез ХУаззегЬаизЬаИз йег РЯапгеп иШег йеп бко1оё15сЬеп
Ве(11п§ипееп йез 081-СЬап§а1 (МУР) // РесМез РереПопит. Вс1 96. Н. 5-6. 8. 467-477. 1оЬпзоп I . М., РгИсЬагс! I . , ОогЬат ^., Тотоз А. В. 1996. ОГО\У1Ь, \уа1ег ге1а11опз апс1 5о1и1е
асситикПоп 1п 05то{1са11у зйеззес! зееёИп^з оГ 1Ье 1гор1са1 1гее Со1оркозрегтит торапе II Тгее РЬу81о1оёу Уо1. 16, N0. 8. Р 713-718.
Ка15ег Н., Карреп Е. 1997. 1п 81Ш оЬзегуаИопз оС з1ота1а1 т о у е т е Ш з 1п йИГегеШ Н§Ы-с1агк ге§1те5: Л е 1ПЙиепсе оГ епйо^епоиз гЬу1Ьт1с11у апб 1оп§-1егт а(1]и81теп18 // I . Ехр. Во1. Уо1.48, N0. 313. Р 1583-1589.
Ке11о§ Е. А. 1998. РекНопзЫрз о^ сегеа1 сгорз апс! о1Ьег дгаззез // Ргос. Nа^1. Асас1. 8с1. Уо1. 95, N0. 5. Р 2005-2010.
Ке11о§ Е. А. 2001. Еуо1ийопагу ЫзЮгу оПЪе ^газзез // Р1ап1 РЬузю!. Уо1. 125, N0. 3. Р 1198-1205. КЬап М. А., ип§аг I . А., 8Ьо\уаиег А. М. 2000. Е1Тес18 заИпку оп §ГО\У1Ь, \уа1ег ге1айопз апс! 10п
асситикИоп о{ 1Ье зиЫгор1са! регепп1а1 Ьа1орЬу1е, А1пр1ех §п//11кИ уаг. зЮскзИ II Апп. Во1. Уо1. 85, N0. 2. Р 225-232.
КосЬ К. 1996. СагЬоЫс1га1е-тос1и1а1е(! §епе ехрге881оп 1п р1ап48 // Аппи. Кеу. Р1ап1 РЬу81о1. Р1ап1 Мо1. В1о1. Уо1. 47. Р 509-540.
Ко1с!е Р. Т., РоЫсЬаих Р. Н., Могзе 8. Р., 8т11Ь С. М. 1989. Р1ап1 \уа1ег зкШз, ЬуйгаиЬс ге8181апсе апё сарас11апсе// Р1ап{ рЬу81о1о§1са1 есо1о§у. КеМ те1Ьос1з апс11пз1гитеп1;а11оп. Ес18.: Реагсу Р. XV., ЕЫег1П8ег 1. Р., Моопу Н. А., Рипёе! Р XV. Ьопйоп, Nе\ Уогк. Р 161-184.

Л И Т Е Р А Т У Р А 269
К б т е г С , Меутауег М., Мепепйег-ИесН 8.Р., 8тее15-8сЬее1 А. 1989. РипсИопа! тофЬо1оёу о '̂ Моип1ат Р1ап{5 // Р1ога. Вй 182. Н. 5/6. 8. 353-383.
Ь е т а М. А., Оо1отЬек В. А., ЕсЬауе I . 2000. Ве1ау тос1е1 оНЫ с1гсас11ап р а с е т а к е г / / 1 . ТЬеог. Вю1. Уо1. 204, N0. 4. Р 565-573.
Пп С. 2000. РЬо1огесер1ог5 апё ге§и1а110п оГ По\уег1П8 Ите // Р1ап1 РЬуз. Уо1. 123, Ыо. 1. Р 39-50. Ып1оп М. I . , КоЬе1 Р. 8. 1999. Ьозз о? \уа1ег {гапзроп сарас14у йие 1о ху1ет сау11аиоп 1п гоо15
оГ 4*0 САМ зисси1еп18 // А т . I Во1. Уо1. 86, Ыо. 11. Р 1538-1543, Ьоот15 Р . 8., Ат1Ьог I . 8. 1999. У1е1с1 ро1еп11а1, р1ап1 а831т11а1огу сарасИу, апй те1аЬоИс
еШс1епсе // Сгор 8с1. Уо1. 39, N0. 6. Р 1584-1596. ЕйП^е и. 2000. ТЬе гопор1а81 {'ипс11оп1п2 аз 1Ье т а 5 1 е г 5\Ь !'ОГ с1гсаё1ап ге^икНоп о!"
сгаззиксеап асМ те1аЬо11зт// Р1ап1а. Уо1. 211. 1з5ие 6. Р. 761-769, МагИп С , 01оуег В. I . 1998. Се11и1аг сИГГегепНаНоп 1п 1Ье зЬоо! ер1с1ет11з // СиггеШ Ор1п1оп 1п
Р1ат Вю1оеу Уо1. 1, N0. 6. Р 511-519. МсАШз^ег С. А., Кпарр А. К., Мага§п1 С. А. 1998. к каГ-куе! рЬо1о8уп1Ье513 ге1а1её 1о р1ап1
зиссезз т а Ы§Ыу рго(1ис11Уе §га5з1апс1? // Оесо1о§1а. Уо1, 118, N0. 1-2. Р. 40-46. МсС1ип8 С. Р . 2001. С1гсаа1ап гЬуЛтз т р1ап15 // Апп. Реу. Р1ап1 РЬуз. Уо1. 52. Р 139-162. МсЕ1\уа1п I . С. 1998. По ГоззН р]ап15 51§па1 ра1аеоа1то8рЬег1с СО^ сопсеп4га110п 1п (Ье §ео1о§1са1
раз!? // РЫ1. Тгапз. Р . 8ос. Ьопё. В. Уо1. 353, N0. 1365. Р 83-96. МеЛпа Е., Ргапсхзсо М. 1994. РЬо1о8уп1Ьез15 апс! \уа!ег ге1а110П8 о!" зауаппа 1гее зрес1е8 (11Яег1п§
1п 1еаГ рЬепо1о§у// Тгее РЬу51о1оеу Уо1. 14, N0. 12. Р 1367-1381. М1§аЫё М. А., ЕШаак М. А. 2001. ЕсорЬу81о1оё1са1 зШсИез оп з о т е с1езег1 р1ап1 8рес1е8 пайуе
1о 1Ье Меёкеггапеап аг1а 1п Е§ур1 / / 1 . Апй ЕПУ. УО1. 48, N0. 2. Р. 131-203. М111аг А. ^. 1998. Мо1еси1аг 1п1п§ие ЬеПуееп рЬо1о1гап8(1ис(10п апс! 1Ье С1гсаЙ1ап с!оск // Апп. Во1.
Уо1. 81, N0. 5. Р 581-587. МШаг А. I 1999. В'юЬ^каХ сЪскз т АгаЬШорш 1Ы1апа//'Ыеч^ ?Ьу1.Ш. 141 .188ие2 .Р 175-197. МиПег ] . 1981. РоззИ роИеп гесогё оГех1ап1 ап§108регт8 // Во1. Реу. Уо1. 47, N0. 1. Р 1-142. МигШз О., МШаг А. 1. 2000. Но\ р1ап1з 1е111Ье И т е // Сиггеп1 Ор1п10п 1п Р1ап1 В1о1оёу Уо1. 3,
N0. 1. Р 4 3 ^ 6 . Ые150п XV., Топ§ V. Ъ., Еее I . К., НаШегз Р. 1979. Ме1Ьос!з {'ог соз1пог-гЬу1Ьтоте1гу // СЬгопо-
Ыо1о§1а. Уо1. 6, N0. 4. Р 305-323. ЫоЬе! Р. 8. 1983. ВхорЬузюа! р1ап1 рЬузю1о§у апс! есо1о§у. 8ап Ргапс18со. 608 р. ЫоЬе! Р. 8. 1991. РЬу51сосЬет1са1 апс! епу)гоптеп1а! р1ап1 рЬу81о1о§у. Асас!ет1с Ргезз. 635 р. Рое^Ыпд Р . 8. 2003. РЬазе сЬап§е апс! Ше ге§и1а11оп оГ с!еуе1ортеп1 11т1п§ 1п р1ап1з // 8с1епсе.
Уо1. 301, N0. 5631. Р 334-336. Руапкоу V. I . , Копс1га1сЬик А. V., 8Ыр1еу В. Ееа{' 81гис1иге апс! зрес1Г1с 1еаГ т а з з : {Ье а1р1пе с1езег1
р1ап{з оГШе Еаз^ет Р а т к з , Тас!]1к134ап // Ыеш РЬу{о1. Уо1. 143, N0. 1. Р 131-142. РазсЬег I I . , НйП М.-Т., 81еЬке К., Озтопс! В., Веек К, Еййёе 11. 2001. 8ра11о1етрога1 уапаНоп
оГ те1аЬоН8т 1п а р1ап1 с1гсаё1ап гЬуЛт : ТЬе Ыо1оё1са1 с1оск аз ап аззетЫу о? соир1ес! тс!1У1с1иа1 08С111аЮг8// РКА8. Уо1. 98, N0. 20. Р 11804-11805.
Рауеп I . А. 2002. 8е1ес{1опргеззигез оп 51ота1а1 еуо1ийоп //Не\ РЬу{о1. Уо1. 153. кзие 3. Р. 371-386. Ре1а11аск О. I . 2002. СагЬоп сЬох1(1е апс1 с11та{е оуег Ле раз! 300 Муг // РЫ1. Тгапз. К. 8ос. Ьопс!.
А. Уо1. 360, N0. 1793. Р 659-673. РоеппеЬег§ Н., Мегго\ М. 2003. ТЬе пе1\уогк оГ {1те: ипс1егз1апс1т§ (Ье то1еси1аг С1гсас!1ап
зуз1ет // СиггеШ В1о1оЕу Уо1. 13. N0. 5. Р Р198-Р207. Р о Л т а п В. Н. 2001. 01оЬа1 Ыос11уегз1{у апё (Ье апс1еп( сагЬоп сус1е // РКА8. Уо1. 98, N0. 8.
Р 4305-4310. Ро1Ьтап В . Н. 2002. А(тозрЬег1с сагЬоп сЬох1с!е 1еуе18 {от (Ье 1аз( 500 тНИоп уеагз // РКА8.
Уо1. 99, N0. 7. Р 4167^171. РиоГГР, Репз1п8 Е. 1996, ТЬе (етрегатге-сотреп5а(ес! Оооё\У1п тос!е1 51ти1а(е8 т а п у с1гсасЬап
с1оск ргорег(1ез // ТЬеог, Вю1, Уо1. 179, N0. 4. Р 275-285.

270 С . Н . Ш е р е м е т ь е в . Травы на г р а д и е н т е в л а ж н о с т и почвы
8а§е К. Р. 2001. ЕпухгоптегНа! ап(1 еУо1и{10пагу ргесогкИйопк Сог 1Ье отщт апЛ сНуегзЮсаНоп о^ 1Ье рЬо1о8уп1Ье11с зупёготе // Р1ап1 Ь1о1. Уо1. 3. ккие 3. Р. 202-213.
8аёе Р . Р 2004. ТЬе еуо1и1юп оГС4 рЬо1о8уп1Ье515 // Ые\ РЬу1о1. Уо1. 161. Ьзие 2. Р 341-370. 8сЬи112 Т. Р., Кау 8. А. 2003. СхгсасЬап сЬскз т ёаНу ап<1 зеакопа! соп1го1 оГ с1еуе1ортеп1//
8стсе . Уо1. 301, N0. 5631. Р 326-328. 8Ые1с15 Ь. М. 1950. ЬеаГ хеготофЬу аз ге1а1ес1 1о рЬу51о1о§1са1 апс1 81гиси1га1 1пЯиепсе8// Во1.
Реу. Уо1. 16, N0. 8. Р 399-447. 8ЫпЬго1 Т., 8сагЬгои§Ь К. 1999. Пзхпе уаг1аЫИ1у Ю геёи1а1е 1оп§ 4егт Ыо1о§1са1 г Ь у Л т з / / ] .
ТЬеог. В1о1. Уо1. 196, N0. 4. Р 455^71 . 8 а а г а т а т У , МайЬауагао С. N . 1997. ТЬе епег§е11с Ьа818 о{ 08то1о1егапсе т р1ап18: рЬу81са1
рппс1р1е5//1. Леог. В1о1. Уо1. 189, N0. 4. Р 333-352. 8т11Ь XV. К., МсОеап Т. М. 1989. АёарНуе ге1айопзЫр Ье1\уееп \Ы V/а^ст гереИепсу, 81ота1а1
ё181пЬтюп, апА §а8 ехсЬап§е // А т . Во{. Уо1. 76, N0. 3. Р. 465^69. 8 т к Ь XV. К., Ве11 В. Т., 8Ьерегс1 К. А. 1998. Аз80С1а110пз Ье1\уееп \еа{ зШдсШге, опеп1айоп, апс1
зипНёЬ! ехрозиге 1п йуе ХУезйт Аиз1гаИап соттипШез // А т . I . Во1. Уо1. 85, N0. 1. Р. 56-63. 8отег8 В . 1999. ТЬе рЬу81о1оёу апй то1еси1аг Ьазез оГ 1Ье р1ап1 С1гсас11ап с1оск// Р1ап1 РЬуз.
Уо1. 121, N0. 1. Р 9-19. 8(а18о1'1, Тпс. Электронный учебник по статистике. Москва, 2001. 81а18оГ1. XVЕВ: ЬПр://
\у\у\у.81а18ой.ги/Ьоте/1ех(Ьоок/с1е{'аи11.Ыт. 81еис11е Е. 1989. XVа̂ ег По\ 1п р1ап18 апё Из соирИп§ Ю о1Ьег ргосеззез: ап оуегУ1е\у// МеЛойз
т еп2уто1о§у Уо1. 174. Р 183-225. ТЬогп1еу I . Н. М. 1996. Мос1е1т§ \уа1:ег т сгорз апс1 р1ап1 есозузГешз // Апп. Во1. Уо1. 77, N0. 3.
Р 261-275. ТЬогп1еу I . Н. М., Саппе1 М. О. Р . 1997. Тетрега1е егаззкпё гезропзе 1о сНта1е сЬап§е: ап
апа1у513 из1П8 Ниг1еу разШге тос1е1 // Апп. Во1. Уо1. 80, N0. 2. Р 205-221. Т^ап^и^11^п^ XV. 1954. ОЬег йеп Е^пйизз уоп ЦЪег1етрега(игеп йег ВШМег Ье! Ваиеге1П5сЫизз 1п
КйуеПеп аиГ с1е1сЬе бко1о§18сЬе СО^-АззхтИаИаптеззипб // Вег. В(8сЬ. Во{. Оез. В<1 67. Н. 6/7. 8. 191-204.
Т и т е г N . С. 1981. ТесЬп1яиез апА е х р е п т е п Ы арргоасЬ Гог 1Ье т е а з и г е т е п ! о!" р1ат ууа1ег з1а1и8 // Р1ап1 апй 8оИ Уо1. 58, N0. 1-3. Р 339-366.
Тугее М. Т., Н а т т е 1 Н. Т. 1972. ТЬе теазигетеп! оГ 1Ье 1иг§ог ргеззиге апй 1Ье \уа1ег ге1а110пз оГ р1ап1з Ьу 1Ье ргеззиге-ЬотЬ 1есЬп1яие / / 1 . Ехр. Во{. Уо1. 23, N0. 74. Р. 267-282.
Тугее М. Т., 1агУ18 Р. О. 1982. XVа̂ е̂ 1п Иззиез апё се11з // Епсус1ореЛа оГ р1ап1 рЬуз1о1о§у. Nе\8ег1ез. Уо1. 12 В. РЬу81о1о§1са1 р1ап1 есо1о§у. П. XVа̂ е̂ геЬНопз апй сагЬоп азз1т11а410п. Еёз.: Еапёе О. Т., ЫоЬе! Р 8., Озтопс! С. В., 21е81ег Н. ВегИп, Не!с!е1Ьег§, Nе^V Уогк. Р 35-77.
Тугее М. Т., К1сЬ1ег Н. 1981. АИетаНуе теШос! оГ апа1у81п8 \уа1ег рокпИа! хзоШегтз: з о т е саийопз апй с к п й с а й о п з . I . ТЬе 1трас1 о!' поп-1с!еаЬ1у апй оГ з о т е ехрегхтепЫ еггогз / / 1 . Ехр. В о 1 . Уо1. 32, Но. 128. Р 643-653.
Тугее М. Т., Р1сЬ1ег Н. 1982. А11ета11Уе т е Л о с ! оС апа1у81П8 \уа1ег ро4еп11а1 13о1Ьегт8: з о т е саиИопз апс! скпйсаНопз. I I . СигуШпеагИу 1п \уа1ег ро1еп11а1 1зо1Ьегт8 // Сап. ] . Во1. Уо1. 60, N0. 6. Р 911-916.
ХУагс! I . К., 81га1п В. Р . 1999. Е1еуа1ес1 СО^ з1ис!1ез: раз!, ргезеШ апс! {иШге // Тгее РЬузхо!. Уо1. 19, N0. 4-5. Р 211-220.
ХУеЬЬ А. А. Р . 2003. ТЬе рЬузюкёу о!'сксасНап гЬу1Ьтз т р к Ш з / / Не\ РЬу!. Уо1. 160. 1ззие 2. Р 281-303.
2асЬо8 I . , Ра§ап1 М. 81оап Е., ТЬотаз Е. ВШирз К. 2001. Тгепс!8, гЬу1Ьт8, апс! аЬеггаИопз 1П ё1оЬа1 сита!е 65 Ма 1о ргезеш // 8с1епсе. Уо!. 292, N0. 5517. Р 686-693.
2о18 С , XV п̂̂ ег К. 1994. РЬо1о8уп1Ье818 оГ а 1гор1са1 сапору 1гее, СегЬа реп1ап(1га, 1п а 1оуу1ап(1 Гогез! ш Рапата // Тгее РЬузю1оёу Уо1. 14, N0. 11. Р 1291-1301.

ОГЛАВЛЕНИЕ
Введение •. 3 Материал и методика 5 Определения 18
Часть I . С Е З О Н Н А Я Д И Н А М И К А В О Д Н О Г О Р Е Ж И М А Д О М И Н А Н Т О В И Э Д И Ф И К А Т О Р О В С О О Б Щ Е С Т В В РАЗНЫХ Ч А С Т Я Х Г Р А Д И Е Н Т А В Л А Ж Н О С Т И П О Ч В Ы 32
Глава 1. Аридная часть градиента 32 1.1. Разнотравно-днепровскоковыльное сообщество 32 1.2. Ковыльково-луково-баглуровое сообщество 44 1.3. Твердоосочково-змеевковое сообщество 54
Глава 2. Средняя часть градиента 60 2.1. Ковыльно-разнотравно-вострецовое сообщество 60 2.2. Разнотравно-осоковое сообщество 70 2.3. Злаково-осоковое сообщество 81
Глава 3. Гумидная часть градиента 89 3.1. Осоково-разнотравно-злаковое сообщество 89 3.2. Крупноразнотравно-дернистоосоковое сообщество 95 3.3. Злаково-богаторазнотравное сообщество 105 3.4. Остроосоковое сообщество 111
Часть I I . Д Н Е В Н А Я Д И Н А М И К А В О Д Н О Г О Р Е Ж И М А Д О М И Н А Н Т О В И Э Д И Ф И К А Т О Р О В С О О Б Щ Е С Т В В РАЗНЫХ Ч А С Т Я Х Г Р А Д И Е Н Т А В Л А Ж Н О С Т И П О Ч В Ы 122
Глава 4. Транспирация 125 Глава 5. Дефицит водного н а с ы щ е н и я 138 Глава 6. Содержание воды в листьях 144
Часть I I I . З А К О Н О М Е Р Н О С Т И С Т Р У К Т У Р Н О - Ф У Н К Ц И О Н А Л Ь Н О Й О Р Г А Н И З А Ц И И Р А С Т Е Н И Й Н А Г Р А Д И Е Н Т Е В Л А Ж Н О С Т И П О Ч В Ы 151
Глава 7. Интенсивность транспирации 151 Глава 8. Д е ф и ц и т водного н а с ы щ е н и я 172 Глава 9. Содержание воды 188 Глава 10. Осмотическое давление клеточного сока 209 Глава 11. М е ж в и д о в ы е структурно-функциональные корреляции 230
Заключение 253
Литература 262

Научное издание
Сергей Николаевич ШЕРЕМЕТЬЕВ
ТРАВЫ НА ГРАДИЕНТЕ ВЛАЖНОСТИ ПОЧВЫ (водный обмен и структурно-функциональная организация)
М.: Товарищество научных изданий КМК. 271 с.
Редактор издательства К. Г. Михайлов
Оригинал-макет Т. А. Горлиной
Для заявок: 123100, Москва, а/я 16, Издательство КМК;
электронный адрес ктк2000@оп1те.ги Ьйр: //\уеЬсеп1ег ги/~ктк2000
Подписано в печать 12.12.2005. Заказ № 2353 Формат 70x100/16. Объём 17 п.л. Бумага офсетная. Тираж 500 экз.
Отпечатано в ГУП ППП «Типография «Наука», 121099, Москва, Шубинский пер., 6

V.
>м<)*




















