
Heading-vector navigation based on head-direction cells and path integration
24
Heading-Vector Navigation Based on Head-Direction Cells and Path Integration John L. Kubie 1 * and Andre ´ A. Fenton 2 ABSTRACT: Insect navigation is guided by heading vectors that are computed by path integration. Mammalian navigation models, on the other hand, are typically based on map-like place representations pro- vided by hippocampal place cells. Such models compute optimal routes as a continuous series of locations that connect the current location to a goal. We propose a ‘‘heading-vector’’ model in which head-direction cells or their derivatives serve both as key elements in constructing the optimal route and as the straight-line guidance during route execution. The model is based on a memory structure termed the ‘‘shortcut ma- trix,’’ which is constructed during the initial exploration of an environ- ment when a set of shortcut vectors between sequential pairs of visited waypoint locations is stored. A mechanism is proposed for calculating and storing these vectors that relies on a hypothesized cell type termed an ‘‘accumulating head-direction cell.’’ Following exploration, shortcut vectors connecting all pairs of waypoint locations are computed by vec- tor arithmetic and stored in the shortcut matrix. On re-entry, when local view or place representations query the shortcut matrix with a current waypoint and goal, a shortcut trajectory is retrieved. Since the trajectory direction is in head-direction compass coordinates, navigation is accomplished by tracking the firing of head-direction cells that are tuned to the heading angle. Section 1 of the manuscript describes the properties of accumulating head-direction cells. It then shows how accumulating head-direction cells can store local vectors and perform vector arithmetic to perform path-integration-based homing. Section 2 describes the construction and use of the shortcut matrix for computing direct paths between any pair of locations that have been registered in the shortcut matrix. In the discussion, we analyze the advantages of heading-based navigation over map-based navigation. Finally, we survey behavioral evidence that nonhippocampal, heading-based navigation is used in small mammals and humans. V V C 2008 Wiley-Liss, Inc. KEY WORDS: hippocampus; navigation; head direction cell; place cell INTRODUCTION How does a navigator construct an efficient path that extends from its current location to a goal? There are two general solutions. One map- based approach suggests that the navigator constructs a path that represents a continuous series of locations along the optimal route. A second heading-vector approach suggests that the navigator constructs a vec- tor of a particular length and direction that extends from the current location to the goal. The discovery of place cells led to enthusiasm for map-based neural models for mammalian navigation based on place cells. We argue that the head-direction cells should reorient our thinking. Specifically, we suggest that head-direction cells are keys to thinking of heading- vector models for navigation. This article has two foci. The first is to present an overall approach to navigation organized around head- direction cells and heading vectors rather than place cells and cognitive maps. The second purpose is to present a particular instantiation of a heading-vector navigation model. The rationale for creating a com- puter-based model is not so much to argue this partic- ular instantiation is correct, but to discover strengths and weaknesses, which may guide the search both for better models and for neural candidates of model components. The Heading-Vector Approach to Navigation Head-direction cells have the remarkable property of firing at maximal rates when the animal’s head is in a particular azimuthal heading angle (Ranck, 1985; Taube et al., 1990). This demonstrates that the rat brain contains a compass ideal for several forms of navigation. This article proposes a model for how this compass signal is used in combination with place in- formation and path integration (PI) to implement a heading-vector mechanism of navigation. In the model, place information is only used to select the or- igin and goal of the heading vector. PI is used to con- struct the heading vector. PI, the use of self-motion cues to compute a vector between a navigator’s current location and an origin (Mittelstaedt, 2000), is a key component of many navigational systems. We find it useful to divide PI into two categories referred to as PI homing and PI localization (Fuhs and Touretzky (2006) refer to sepa- rate functions of path integration for locale navigation and for praxis navigation. These appear to be equiva- lent to PI localization and PI homing, respectively). 1 Department of Anatomy and Cell Biology, The Robert F. Furchgott Center for Neural and Behavioral Science, S.U.N.Y. Downstate Medical Center, Brooklyn, New York; 2 Department of Physiology and Pharma- cology, The Robert F. Furchgott Center for Neural and Behavioral Sci- ence, S.U.N.Y. Downstate Medical Center, Brooklyn, New York Grant sponsor: National Science Foundation; Grant number: IOS- 0725001. *Correspondence to: John Kubie, Department of Anatomy and Cell Biol- ogy, S.U.N.Y. Downstate Medical Center, 450 Clarkson Ave., Brooklyn, NY 11203, United States. E-mail: [email protected] Accepted for publication 9 October 2008 DOI 10.1002/hipo.20532 Published online 12 December 2008 in Wiley InterScience (www. interscience.wiley.com). HIPPOCAMPUS 19:456–479 (2009) V V C 2008 WILEY-LISS, INC.
Transcript of Heading-vector navigation based on head-direction cells and path integration

Heading-Vector Navigation Based on Head-Direction Cellsand Path Integration
John L. Kubie1* and Andre A. Fenton2
ABSTRACT: Insect navigation is guided by heading vectors that arecomputed by path integration. Mammalian navigation models, on theother hand, are typically based on map-like place representations pro-vided by hippocampal place cells. Such models compute optimal routesas a continuous series of locations that connect the current location toa goal. We propose a ‘‘heading-vector’’ model in which head-directioncells or their derivatives serve both as key elements in constructing theoptimal route and as the straight-line guidance during route execution.The model is based on a memory structure termed the ‘‘shortcut ma-trix,’’ which is constructed during the initial exploration of an environ-ment when a set of shortcut vectors between sequential pairs of visitedwaypoint locations is stored. A mechanism is proposed for calculatingand storing these vectors that relies on a hypothesized cell type termedan ‘‘accumulating head-direction cell.’’ Following exploration, shortcutvectors connecting all pairs of waypoint locations are computed by vec-tor arithmetic and stored in the shortcut matrix. On re-entry, whenlocal view or place representations query the shortcut matrix with acurrent waypoint and goal, a shortcut trajectory is retrieved. Since thetrajectory direction is in head-direction compass coordinates, navigationis accomplished by tracking the firing of head-direction cells that aretuned to the heading angle. Section 1 of the manuscript describes theproperties of accumulating head-direction cells. It then shows howaccumulating head-direction cells can store local vectors and performvector arithmetic to perform path-integration-based homing. Section 2describes the construction and use of the shortcut matrix for computingdirect paths between any pair of locations that have been registered inthe shortcut matrix. In the discussion, we analyze the advantages ofheading-based navigation over map-based navigation. Finally, we surveybehavioral evidence that nonhippocampal, heading-based navigation isused in small mammals and humans. VVC 2008 Wiley-Liss, Inc.
KEY WORDS: hippocampus; navigation; head direction cell; place cell
INTRODUCTION
How does a navigator construct an efficient path that extends from itscurrent location to a goal? There are two general solutions. One map-
based approach suggests that the navigator constructsa path that represents a continuous series of locationsalong the optimal route. A second heading-vectorapproach suggests that the navigator constructs a vec-tor of a particular length and direction that extendsfrom the current location to the goal. The discoveryof place cells led to enthusiasm for map-based neuralmodels for mammalian navigation based on placecells. We argue that the head-direction cells shouldreorient our thinking. Specifically, we suggest thathead-direction cells are keys to thinking of heading-vector models for navigation.
This article has two foci. The first is to present anoverall approach to navigation organized around head-direction cells and heading vectors rather than placecells and cognitive maps. The second purpose is topresent a particular instantiation of a heading-vectornavigation model. The rationale for creating a com-puter-based model is not so much to argue this partic-ular instantiation is correct, but to discover strengthsand weaknesses, which may guide the search both forbetter models and for neural candidates of modelcomponents.
The Heading-Vector Approachto Navigation
Head-direction cells have the remarkable propertyof firing at maximal rates when the animal’s head is ina particular azimuthal heading angle (Ranck, 1985;Taube et al., 1990). This demonstrates that the ratbrain contains a compass ideal for several forms ofnavigation. This article proposes a model for how thiscompass signal is used in combination with place in-formation and path integration (PI) to implement aheading-vector mechanism of navigation. In themodel, place information is only used to select the or-igin and goal of the heading vector. PI is used to con-struct the heading vector.
PI, the use of self-motion cues to compute a vectorbetween a navigator’s current location and an origin(Mittelstaedt, 2000), is a key component of manynavigational systems. We find it useful to divide PIinto two categories referred to as PI homing and PIlocalization (Fuhs and Touretzky (2006) refer to sepa-rate functions of path integration for locale navigationand for praxis navigation. These appear to be equiva-lent to PI localization and PI homing, respectively).
1Department of Anatomy and Cell Biology, The Robert F. FurchgottCenter for Neural and Behavioral Science, S.U.N.Y. Downstate MedicalCenter, Brooklyn, New York; 2Department of Physiology and Pharma-cology, The Robert F. Furchgott Center for Neural and Behavioral Sci-ence, S.U.N.Y. Downstate Medical Center, Brooklyn, New YorkGrant sponsor: National Science Foundation; Grant number: IOS-0725001.*Correspondence to: John Kubie, Department of Anatomy and Cell Biol-ogy, S.U.N.Y. Downstate Medical Center, 450 Clarkson Ave., Brooklyn,NY 11203, United States. E-mail: [email protected] for publication 9 October 2008DOI 10.1002/hipo.20532Published online 12 December 2008 in Wiley InterScience (www.interscience.wiley.com).
HIPPOCAMPUS 19:456–479 (2009)
VVC 2008 WILEY-LISS, INC.

The function of PI homing is to calculate routes an animalmight take. In PI homing, the vector is computed by vectorsummation of the navigator’s outbound route from the originto the current location (Fig. 1A). One obvious use of PI hom-ing is in solving the navigational problem of getting back tothe origin. This is done by inverting the PI vector to computea heading vector that is a direct path from the animal’s currentlocation to the origin, which is commonly referred to as‘‘home.’’ Animals as small-brained as ants perform efficient PI-homing over large distances using this method (Wehner,2003). PI homing is also used by sailors to compute a returnroute to a home location. The heading-vector model for navi-gation described in this article uses memory mechanisms toextend the simple homing vector system to serve as a mecha-nism to create numerous ‘‘local’’ heading vectors that connectsalient locations.
In comparison, the principal function of PI localization is toupdate the animal’s estimate of its location on a map. Thecomputational mechanism for PI localization is for the naviga-tor to start with an estimate of its location on a map, move,and use the short PI-movement vector to update the estimateof current location on the map. The process is continuous,with each new estimate of current location starting a cycle. PIvectors are short, and PI resets with each updated estimate ofthe animal’s location (Hasselmo, 2007). On occasion, perhapsat pause points, the animal updates its location estimate by tak-ing a sighting from external cues. Sightings reset the integratorand eliminate drift. Although a navigator using PI localizationcan perform efficient navigation from the map, PI-vectors playno direct role. Examples of PI localization are a rat using PI toupdate its current location on a cognitive map or a sailor, usingrecent history of movement through the sea, to update the esti-mate of location on a navigational chart. The heading-vectormodel described in this article uses PI homing, not PIlocalization.
As shown in Figure 1B, the PI vector (global vector) is sim-ply the algebraic sum of x and y components of the path seg-ments (local vectors). As shown, vector summation in Cartesiancoordinates is a simple calculation. The question of how ananimal can do PI without a Cartesian frame of reference isproblematic and is addressed in various models (Wan et al.,1993; O’Keefe and Burgess, 2005; Hasselmo et al., 2007)including a process related to sinusoidal addition that will bedescribed in Section 2, below. Figure 1B also illustrates a con-cept termed ‘‘Path-segment arithmetic,’’ which refers to the factthat the sequence order of adding local vectors has no effect onthe resultant global vector. Path-segment arithmetic of this sortcan lead to flexible computation of several routes from a singleoutbound journey, but requires that local vectors be stored sep-arately from the global vector (Cheng, 1989; Etienne et al.,1998; Etienne and Jeffery, 2004).
PI appears to rely on three fundamental components: (1) Anintegrator that sums movements in distance units; (2) a reset-ting mechanism, that permits the integrator to start at zero atthe origin; and (3) a directional representation that keeps trackof the direction of the subject’s movement with reference to aninertial frame (such as north) (Gallistel, 1980; Gallistel andCramer, 1996; Etienne et al., 2004). Elegant behavioral workby Wehner and colleagues has shown each of these componentsin ants, although the brain mechanisms are only partiallyunderstood (Wehner, 2003).
PI homing, as described in insects, is part of a broader cate-gory of navigation termed ‘‘vector’’ or ‘‘heading-vector’’ naviga-tion. In heading-vector navigation, locomotion is guided bydirectional heading and linear distance. Virtually all insect navi-gation is analyzed in a heading-vector conceptual framework.With the notable exception of the work of Collett et al., whoused this approach to analyze gerbil navigation, the approachhas had modest impact on models of mammalian navigation(Collett et al., 1986).
FIGURE 1. (A) Insects such as ants can take an indirect out-bound journey (ABC) and return via a direct path (homing path).(B) The return path is performed by computing a global vector (G)which is the vector sum of the path segments A, B, and C. Path seg-
ments are termed ‘‘local vectors.’’ Due to the commutative propertiesof vector addition, G is the same for any sequence of adding localvectors. In the illustration, G(ABC) 5 G(CBA). The homing vectorof 1A is computed by inverting the global vector (G).
VECTORS AND NAVIGATION 457
Hippocampus

A different conceptual scheme is the cognitive map. Fueledby O’Keefe and Dostrovsky’s discovery of place cells in the hip-pocampus, O’Keefe and Nadel developed the cognitive map-ping model (O’Keefe and Dostrovsky, 1971; O’Keefe andNadel, 1978). According to the theory, the hippocampal maphas at least two functions: helping an animal recognize its cur-rent location and navigation. The essential feature of map-based navigation is that it flexibly computes direct, efficientroutes from any location to any other location in the mapped-space. Map-based navigation permits an animal to pass severalwell-known tests of complex, goal-directed navigation. Specifi-cally, when an animal is using map-based navigation it can tra-verse unvisited regions of space, take advantage of shortcuts,and efficiently circumvent detours.
The cognitive map theory of O’Keefe and Nadel spawnedthe idea that map-based navigation depends on hippocampalplace cells. Numerous attempts have been made to show howmap-like functions may emerge from hippocampal processing.Many of these are reviewed in the Discussion. A brief summaryis that early models focused solely on hippocampal place cells(O’Keefe, 1991; Brown and Sharp, 1995; Muller et al., 1996).These suffered from an inability to compute paths that traverseunvisited regions of space. More recent theories have incorpo-rated PI and, to a lesser degree, head-direction signals. Thesetheories (Touretzky and Redish, 1996; Samsonovich andMcNaughton, 1997), as well as more recent versions thatmoved the map to the network of grid cells in the medial ento-rhinal cortex all focus on self-localization to identify currentposition rather than navigation between distant locations(O’Keefe and Burgess, 2005; Fuhs and Touretzky, 2006;McNaughton et al., 2006; Hasselmo, 2007; Hasselmo andBrandon, 2008). Navigation requires more than knowing cur-rent position, and when navigation between two places isexplained, it is restricted to highly familiar environments (Bur-gess and O’Keefe, 1996). All of these theories are wed to self-localization; each requires that at all times an animal tracks itslocation on the cognitive map. To the degree that these theoriesexplain navigation, paths are restricted to small, familiar envi-ronments, such as elevated mazes and small enclosures.
The theory outlined in this article suggests a different con-ceptual framework. We propose that the principal guidance fornavigation is a heading vector rather than a map, and that thebest candidate for such a vector is the head-direction cell. Inthe model, place cells are used to select a heading vector, butare not used during movement along the vector. In manyrespects, this theory parallels ideas that are well developed ininsect research and nautical navigation. We imagine a sequencewhere during a single, brief environmental exposure a sparse setof efficient straight-line paths connecting salient locations iscomputed and stored. The salient locations may have denseplace cell representations, resembling a local patch of the largerspace. Importantly, for navigation through the space, a patch isused as a navigational waypoint, rather than part of a Euclideanmap. The representation of the environment for navigation is aset of waypoint locations connected by shortcut paths. Thepaths would be the complete set of direct routes connecting all
pairs of waypoints. This amounts to a representation ofstraight-line paths between key places in the environment, butnot a map for continuously charting locations through thespace.
The heading-vector framework being proposed offers solu-tions to several shortcomings of the cognitive mapping mod-els. There are four attractive features to the heading-vectorframework. First, it works. We will show how heading vectorscomputed from head-direction cells, place cells, and PI can beused to efficiently navigate between pairs of locations in amodestly familiar environment. Second, the theory is consist-ent with the ways rats explore environments. We find that,when a rat is placed in a novel enclosure it moves along walledges and between prominent landmarks. Such a pattern, alsoobserved in humans (Bohbot et al., 2002), is optimal for cre-ating waypoints and heading vectors, but not optimal for cre-ating a representation of continuous space. Third, heading-vector navigation is consistent with recent studies of humannavigation. For example, Spiers and Maguire (2006) find thatdrivers engage their hippocampus when selecting a routeacross a city, but the hippocampus is not activated duringroute execution. In another example, ancient Polynesianislanders had no maps, but rather used heading vectors guidedby stars to navigate among dispersed islands (Lewis, 1994).Finally, this theory unifies various forms of navigation fromrodent exploration in small, laboratory environments to long-distance travel. For example, bird migration appears to beguided by heading vectors calculated with the aid of a suncompass (Bingman and Jones, 1994). We propose that a rep-resentation of waypoints and heading vectors is the initialnavigation system in all environments; with increased familiar-ity, however, a continuous, map-like representation of theenvironment may be constructed permitting map-basednavigation.
The Heading-Vector Model
High-level navigation permits an animal to take efficientroutes to unmarked or invisible goals. Mammalian models forhigh-level navigation published over the past 30 years fall intotwo categories: those that are maps and those that rely on vec-tors. Most map-based theories have two features in common:(1) place cells signal to an animal where it is in space and (2)to navigate from one location to another, an animal constructsand follows a sequence of place-cell firing fields and the direc-tions that connect them. Here, we propose a vector modelmore closely based on the work of Collett et al. and discoveriesfrom insect navigation (Collett et al., 1986, 1998; Wehner,2003). In the vector model, when an animal navigates fromone location to another it follows a path-direction templaterather than a list of places. The path-direction template we pro-pose is either a set of head-direction cells or cells with proper-ties similar to a head-direction cell. Figure 2 depicts the essen-tial difference between cognitive map-based models and theheading-vector model presented in this article. In the heading-
458 KUBIE AND FENTON
Hippocampus

vector model, although place cell firing has an essential role inidentifying an animal’s current location and the goal, it doesnot serve a critical function during path execution. This role isserved by an element we call a ‘‘path vector.’’
Optimal routes are straight-line paths. Although computingstraight lines while traveling in a moving, egocentric frame isnot trivial, it is clear that insects can do this in simple nervoussystems. Theoretical work in insects postulates the existence of‘‘compass neurons’’ (Wehner, 2003). Although the behavioralwork in mammals is far less complete, the physiology isadvanced. The head-direction cells discovered by Ranck andcolleagues are compass neurons (Ranck, 1985; Taube et al.,1990; Wehner, 2003). An individual head-direction cell firesrapidly when the animal’s head is at a particular angle withrespect to the earth’s inertial frame (e.g., an azimuth such as anorth–south line). We propose that a mammal computes anoptimal route in much the same way an ant homes: it followsa homing vector. We further propose that the homing vector orpath vector has the compass properties of a head-direction cell.Figure 3 illustrates that, if an animal maximized the firing of aparticular head-direction cell (or a set of similar cells) whilemoving, it will move in a straight path. One thing is missing,however: a distance metric. A complete path vector should havea directional signal and a distance metric. To create the explicitmodel we postulate a ‘‘head-direction accumulator’’ (HDA) cellwhich, in addition to acting as a compass, provides a distancemetric. HDA cells are head-direction cells whose firing rate isan integral function of distance traveled in a particular direc-tion (Fig. 4). We note that, although HDA cells have not beendiscovered, velocity integrators have been found in other sys-tems (Aksay et al., 2000), and PI requires this type of mecha-nism, and intracellular integration has been found in slicerecordings of EC (Egorov et al., 2002; Fransen et al., 2006).Another important point is that we do not believe that singlecells drive behavior. When we use terminology that suggeststhat single cells are critical, such as references to ‘‘a place cell’’or a ‘‘head-direction cell,’’ we are referring to a set of cells withsimilar firing characteristics and connectivity.
The manuscript is divided into three sections. Section 1presents a model by which HDA cells compute the global vectorfor PI homing. Although we consider the model described inSection 1 to be a strong model for mammalian homing, we arealso presenting it for its heuristic value. It was designed, in part,to demonstrate concepts, such as path-segment arithmetic, whichare exploited in Section 2. Section 2 extends Section 1 by illus-trating how a vector-navigation model can be used to computepath vectors (shortcuts) between any pair of visited locations(waypoints). Although the shortcut matrix of Section 2 can, inprincipal, be built with one of the several mechanisms for PI, inthe computer model we use the path integrator built fromHDAs. First, we illustrate how a simple initial exploratory jour-ney through an environment can be broken down into path seg-ments that are stored with reference to their start and end loca-tions. Second, we show how the connecting path (shortcut) canbe recalled using a lookup table based on start and stop locationsas the keys. We call the storage and retrieval entity a ‘‘shortcutmatrix.’’ Finally, in Section 3, we analyze two special cases thatmay use the shortcut matrix: extended space and bounded space.In extended space, we suggest that the heading vectors producedby a shortcut matrix may serve for long-distance navigation andto connect widely separate environments. In much smallerregions of bounded space, a shortcut matrix may serve to de-velop a special set of routes that connect waypoints adjacent toboundaries and register the shape of the environment.
GENERAL METHOD
Simulations were performed with a graphical computer pro-gram called ‘‘ziggy.’’ One portion of the program implementsthe head-direction-accumulator PI model of Section 1, and asecond portion implements the shortcut matrix of Section 2.Macintosh and Windows PC versions of the software can bedownloaded from the site http://www.downstate.edu/neuro-science_materials/downloads/Kubie-FentonSoftware.html.
FIGURE 2. (A) Navigation with place cell guidance. A set ofhypothetical place cell firing fields is shown laid on the surface ofa square enclosure. The rat is currently in the firing field marked‘‘rat’s location’’ and desires to get to the goal, which is in the firingfield marked ‘‘Goal.’’ In a ‘‘place cell guidance’’ model, the hippo-campus computes a path as a series of firing fields that will lead
the rat from its current location to the goal. Path-vector guidanceis illustrated in (B). The rat is in the firing field of one cell anddesires to get to the goal, which is in the firing field of anothercell. In this model, the animal follows a path vector to travel fromits current location to the goal.
VECTORS AND NAVIGATION 459
Hippocampus

SECTION 1. PI HOMING WITH HEAD-DIRECTION ACCUMULATORS
In the Introduction, we described a mechanism where hom-ing vectors (global vectors) could be computed by adding thevectors of path segments (local vectors). Although this is easyto compute with a Cartesian representation, it is considerablymore difficult using the head-oriented representation providedby sensory systems. In this section, we show how a homingvector can be computed with head-oriented computation. Spe-cifically, we describe HDAs and show how they can reliablycompute global vectors. In addition, we will lay the ground-work for more flexible route computation by showing how thestorage of path-segment vectors can be achieved.
PI homing uses PI to compute a homing vector on an out-bound journey and follow the inverse home. At least five compo-nents are assumed to exist: (i) an ability to measure the distancetraveled; (ii) an ability to detect the travel direction as an anglefrom the inertial reference frame (e.g., true north); (iii) a vectorsum that stores the distance and direction of the outbound jour-
FIGURE 3. Head-direction cells as heading guides. If an ani-mal moves while maximizing the firing of a particular head-direc-tion cell, it will move on a straight path in the cell’s preferreddirection: it will be following the path guided by the head-direc-tion cell. Top row. The tuning curves of three hypothetical head-direction cells, HD1 has a preferred firing angle of 1058, HD2 of
408, and HD3 of 3308. The bottom illustrates that if, from a startlocation, an animal proceeds in a path that maintains one of thesecells at maximal firing, it will be moving on a straight path head-ing in the preferred direction of the head-direction cell. The figureis a simplification in that we believe that in each instance behaviorwould be guided by a set of cells with similar firing characteristics.
ney; (iv) the ability to follow the inverse of the vector sum toreturn home; and (v) a reset mechanism. In searching for neuro-nal evidence for these functions, the most important clue hasbeen the discovery of head-direction cells (Ranck, 1985; Taubeet al., 1990). Each head-direction cell is tuned to a preferreddirection that is the angle of the rat’s head with respect to the in-ertial reference frame of the earth. Thus, head-direction cells arestrong candidates for the most complicated operator in PI hom-ing. Although most head-direction cells are narrowly tuned, someare broadly tuned sinusoids, especially those found in the lateralmammillary nucleus (Stackman and Taube, 1998). We postulatethe existence of a cell type called a ‘‘HDA’’ (Fig. 4). In the sim-plest scheme, each HDA is downstream to a broadly tuned head-direction cell and integrates its input until reset. HDAs come insets with preferred directions that sample all head angles. The cellsof a HDA set have the property that they are reset together.HDAs are similar to the ‘‘graded persistent firing cells’’ describedin page 4, equation 3 of Hasselmo and Brandon (2008).
If the inputs to a set of HDAs have cosine input functions,then the set of accumulators is itself a sinusoid. This meansthat if we plot each element as a vector on a radial plot (r,u),
460 KUBIE AND FENTON
Hippocampus

(r is the accumulator value and theta is the cell’s preferred fir-ing angle) we will obtain a circle whose angle of connectionto the origin is phase and the diameter is amplitude. In theset of radial accumulators, the element with the largest valuewill have a length reflecting amplitude for the set and a pre-ferred angle that reflects phase for the set. In other words,within a set, the radial accumulator with the largest value willreflect both phase and amplitude of the set. We call the mem-ber of a HDA set that has the largest value the ‘‘largest HDAelement.’’
We created a computer model to test and illustrate thisproperty. The input to the program is a path that is entered asa series of x and y coordinates. The path is a series of linearsegments that lead from start to end. For each segment, a setof radial accumulators is computed. Initially, a set consists of36 radial accumulators with 108 angles between neighbors. Thevalue stored in each accumulator is d ! cos u, where d is thedistance of the segment and theta is the deviation of the angleof the segment from the preferred angle of the accumulator. Asanticipated, when the set of radial accumulators for a segment
is graphed on a radial plot, a circle is produced. The HDA ele-ment with highest activation corresponds to the angle of thepath segment and the magnitude of this value is the length ofthe line. Small errors occur when the angle of the path segmentlies between two radial accumulators. In those cases, the closeraccumulator has the larger value.
At this point, we argue that each HDA set is an accuraterepresentation of the length and angle of each linear path seg-ment. The process of combining sets of HDAs to produce asummed vector is simple: the value of all corresponding com-ponents is summed. This is identical to the analytical methodfor adding sinusoids. The resultant is a sinusoid that has a dif-ferent phase and amplitude of any of its components. Figure 5shows the result when two HDA sets have corresponding com-ponents added to form a new HDA set. The circularity of theresultant plot shows that it is a sinusoid. The largest HDAmember in this set has a preferred direction that correspondsto the angle of the vector sum and a magnitude that corre-sponds to the length of the vector sum. In other words, sum-ming the HDA sets of the path segments produces an HDA
FIGURE 4. Radial Accumulators. (A) The firing of a radial ac-cumulator whose preferred direction is 08, while an animal moveson a 3-segment path. The slope of the firing rate function is high-est when the rat moves in the preferred direction, is lower at 458from the preferred direction, and is zero when the rat moves at908 to the preferred direction. (1) Each accumulator, like a head-direction cell, is most sensitive at a particular head angle. (2)When an animal moves in a cell’s preferred direction, the cell’s ac-tivity increases and accumulates. (3) As movement direction devi-ates from a cell’s preferred direction, the amount of the incrementfalls off as cos h. (B) The slope of activation plotted as a function
of angular deviation from the cell’s preferred firing angle. This is asinusoid function plotted in radial and Cartesian coordinates. (C)A set of radial accumulators, each with a preferred direction 108from its neighbors. Each dark line represents the activity of onecell plotted in its preferred direction. With movement in the 408direction (arrow), activity is greatest in the 408 accumulator andfalls off as a cell’s preferred direction deviates from 408. Negativecosine values are not plotted. The radial accumulator with thehighest value is the one tuned at 408. Its value corresponds to thevalue of the input vector.
VECTORS AND NAVIGATION 461
Hippocampus

set that is the vector sum. This demonstrates that we can useHDA-set arithmetic to perform vector addition.
In insect models of PI homing, a global vector, the vectorsum of local vectors (path segments) is continuously updated.If only a homing vector is calculated, only one vector sumneeds to be maintained. Although it seems likely that compactstorage is a necessity in small-brained animals such as the ant,even in ants there is evidence that both global and local vec-tors are retained during PI (Collett et al., 1998). We proposethat in larger-brained animals, such as mammals several vectorsare computed and stored during initial exploration. Specifi-cally, that each locomotor pause marks the end of a path seg-ment, that an HDA set is computed and stored for each pathsegment (local vector) and that each of these HDA sets isstored at the end of exploration. When PI homing is calledupon, a homing vector is calculated by adding all of the HDAsets representing local vectors. Figures 5 and 8B show that for
complex journeys, adding HDA sets computes accurate com-posite homing vectors.
Several of the properties of HDAs and HDA sets have beenexplored. The most sensitive property is the cosine input func-tion. Recall that in the HDA model, accumulator sensitivity isbroadly tuned; it drops off with the cosine of deviation fromthe preferred head angle. Figure 6 shows the effects of usingother input functions. Only a cosine input function does a per-fect job. Other broadly tuned functions do adequate jobs, whilenarrowly tuned functions lead to serious errors. There arenumerous ways that the nervous system could cope with thisproblem, but the most obvious is to provide cosine-tunedinputs. Although most head-direction cells are narrowly tuned(<908), the cells in the lateral mammillary nucleus are broadlytuned, commonly resembling complete sinusoids, suggestingthat these may be the HDA inputs. It is important to note thatin a model similar to ours, Hasselmo and Brandon (2008) also
FIGURE 5. Radial accumulators on a multisegment path. (A)A 3-segment path, (123) is illustrated. The endpoint is 398 fromthe start and 280 pixels away. This is the global vector. (B) Threesets of radial accumulators are illustrated, one for each path seg-ment. The illustrated accumulator values are 50% the scale of thepath segment. In each set, the radial accumulator tuned to 408 isblack. (C) The radial accumulator that corresponds to the globalvector. This represents the global vector and was obtained at theend of the path by adding the corresponding segments of the threeaccumulators that represent local vectors. To the right of the radial
plot is a line illustrating that adding the 408 lengths of the localvectors produces the 408 length of the global vector. The 408 vec-tor is the longest of the set and is closest in angle and length tothe true displacement. D, E, and F illustrate that a head-directionaccumulator set that operates as the running sum produces thesame result as one that adds the path segments. D is a repeat ofthe local vectors for the three segments. E shows the global vectorvalues, as represented by a set of HD accumulators at the end ofeach path segment, and F shows the global vector values at theend of the journey.
462 KUBIE AND FENTON
Hippocampus
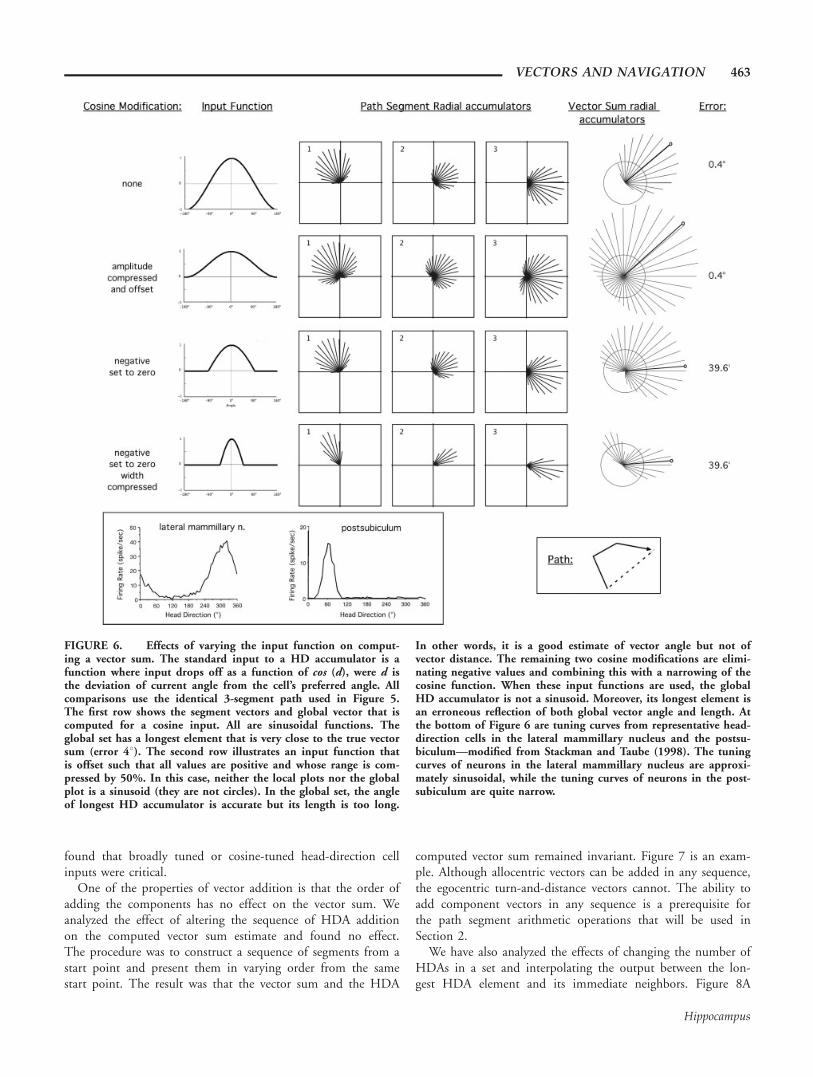
found that broadly tuned or cosine-tuned head-direction cellinputs were critical.
One of the properties of vector addition is that the order ofadding the components has no effect on the vector sum. Weanalyzed the effect of altering the sequence of HDA additionon the computed vector sum estimate and found no effect.The procedure was to construct a sequence of segments from astart point and present them in varying order from the samestart point. The result was that the vector sum and the HDA
computed vector sum remained invariant. Figure 7 is an exam-ple. Although allocentric vectors can be added in any sequence,the egocentric turn-and-distance vectors cannot. The ability toadd component vectors in any sequence is a prerequisite forthe path segment arithmetic operations that will be used inSection 2.
We have also analyzed the effects of changing the number ofHDAs in a set and interpolating the output between the lon-gest HDA element and its immediate neighbors. Figure 8A
FIGURE 6. Effects of varying the input function on comput-ing a vector sum. The standard input to a HD accumulator is afunction where input drops off as a function of cos (d), were d isthe deviation of current angle from the cell’s preferred angle. Allcomparisons use the identical 3-segment path used in Figure 5.The first row shows the segment vectors and global vector that iscomputed for a cosine input. All are sinusoidal functions. Theglobal set has a longest element that is very close to the true vectorsum (error 48). The second row illustrates an input function thatis offset such that all values are positive and whose range is com-pressed by 50%. In this case, neither the local plots nor the globalplot is a sinusoid (they are not circles). In the global set, the angleof longest HD accumulator is accurate but its length is too long.
In other words, it is a good estimate of vector angle but not ofvector distance. The remaining two cosine modifications are elimi-nating negative values and combining this with a narrowing of thecosine function. When these input functions are used, the globalHD accumulator is not a sinusoid. Moreover, its longest element isan erroneous reflection of both global vector angle and length. Atthe bottom of Figure 6 are tuning curves from representative head-direction cells in the lateral mammillary nucleus and the postsu-biculum—modified from Stackman and Taube (1998). The tuningcurves of neurons in the lateral mammillary nucleus are approxi-mately sinusoidal, while the tuning curves of neurons in the post-subiculum are quite narrow.
VECTORS AND NAVIGATION 463
Hippocampus
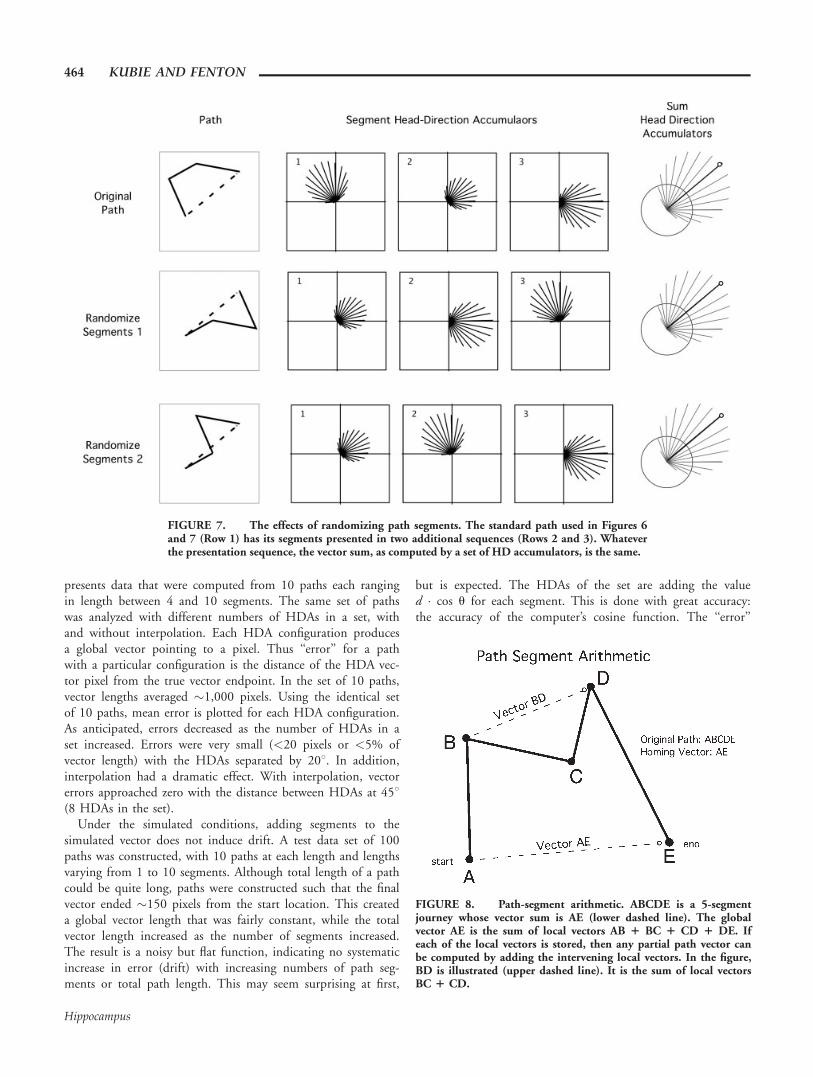
presents data that were computed from 10 paths each rangingin length between 4 and 10 segments. The same set of pathswas analyzed with different numbers of HDAs in a set, withand without interpolation. Each HDA configuration producesa global vector pointing to a pixel. Thus ‘‘error’’ for a pathwith a particular configuration is the distance of the HDA vec-tor pixel from the true vector endpoint. In the set of 10 paths,vector lengths averaged "1,000 pixels. Using the identical setof 10 paths, mean error is plotted for each HDA configuration.As anticipated, errors decreased as the number of HDAs in aset increased. Errors were very small (<20 pixels or <5% ofvector length) with the HDAs separated by 208. In addition,interpolation had a dramatic effect. With interpolation, vectorerrors approached zero with the distance between HDAs at 458(8 HDAs in the set).
Under the simulated conditions, adding segments to thesimulated vector does not induce drift. A test data set of 100paths was constructed, with 10 paths at each length and lengthsvarying from 1 to 10 segments. Although total length of a pathcould be quite long, paths were constructed such that the finalvector ended "150 pixels from the start location. This createda global vector length that was fairly constant, while the totalvector length increased as the number of segments increased.The result is a noisy but flat function, indicating no systematicincrease in error (drift) with increasing numbers of path seg-ments or total path length. This may seem surprising at first,
but is expected. The HDAs of the set are adding the valued ! cos u for each segment. This is done with great accuracy:the accuracy of the computer’s cosine function. The ‘‘error’’
FIGURE 8. Path-segment arithmetic. ABCDE is a 5-segmentjourney whose vector sum is AE (lower dashed line). The globalvector AE is the sum of local vectors AB 1 BC 1 CD 1 DE. Ifeach of the local vectors is stored, then any partial path vector canbe computed by adding the intervening local vectors. In the figure,BD is illustrated (upper dashed line). It is the sum of local vectorsBC 1 CD.
FIGURE 7. The effects of randomizing path segments. The standard path used in Figures 6and 7 (Row 1) has its segments presented in two additional sequences (Rows 2 and 3). Whateverthe presentation sequence, the vector sum, as computed by a set of HD accumulators, is the same.
464 KUBIE AND FENTON
Hippocampus

that occurs is due to a lack of correspondence between the realvector angle and the longest HDA angle. This error occursonly once, on the final segment, when we estimate the HDAvector. With interpolation this error essentially disappears, andis independent of the number of path segments that contributeto the final vector.
After a homing vector is computed, how does an animal fol-low it home? At this point the HDA set that represents thevector sum is complete, and its longest member is the homingvector. We will describe three candidate mechanisms:
i. The animal finds the head-direction cells with preferreddirections in the direction opposite to the homing vector. Theanimal tracks this vector home by (a) maintaining the head-direction cell set firing at a maximal rate; and (b) moving for-ward until the HDA that is the homing vector reduces its valueto zero. All elements in the HDA set ought to return to zero aswell; this composite could be the signal a subject monitors toreach the target.
ii. The animal moves in a direction that causes the HDAcells that represent the homing vector to decrease at a maximalrate until it reaches zero. This mechanism is inefficient in thatit appears to require vicarious body and head movements awayfrom the most direct path.
iii. HDA cells are more complex than we have proposed.They are head-direction cells whose overall gain is modulatedby displacement in the direction of the preferred head angle.Additionally, these cells would be organized in opposed pairs.Homing would involve tracking the complement HDA, maxi-mizing firing rate until a ‘‘home’’ rate occurred, and indicatingreturn to the origin of the path segment.
The first mechanism seems most efficient and parsimonious.Other, relatively simple mechanisms are possible. We imaginethat, in order for homing to follow a straight line, theanimal should either be tracking a head-direction cell set’sactivity (or a set of similarly tuned HD cells) or the decrementin activity of the HDA cell or population that represents thehoming vector.
Other assumptions in the model are as follows:
A. Each path segment is linear. Although the model startswith linear path segments, the process of creating a linear seg-ment from a meandering path can clearly be accomplished byan HDA set.
B. The preferred angles of a HDA set are evenly separated.Although not carefully tested, preliminary tests suggest thateven spacing is not essential.
C. The vector sum is taken as the longest HDA memberrather than an interpolated value between the longest memberand its two nearest neighbors. As indicated above, interpolationgreatly increases accuracy, and interpolation can be achieved byrepresenting the accumulating head-direction value in the activ-ity of a population of HDA cells.
D. The HDA set for each leg of the outbound journey isstored until needed. This is not essential for computing thehoming vector, but, as we will show in Section 2, storing out-
bound components is of great value for flexible use of the PIhoming mechanism.
E. HDA values can be decremented and can be negative.Since firing rate cannot go negative, it is difficult to create acos u function in firing rate. Negative inputs could be repre-sented in three ways: (1) as inhibition; (2) as causing the accu-mulator to leak (a form of inhibition); or (3) by only permit-ting positive inputs, but pairing each HDA with one pointingin the opposite direction. On readout, the difference in thesetwo HDA would be computed for each downstream element.This mechanism is suggested by several authors (Wan et al.,1993; Vickerstaff and Di Paolo, 2005).
F. The dynamic range of HDA values is limited. For exam-ple, what happens when an HDA reaches the maximal value(firing rate)? Can it accumulate any more? This is a problemfor neurons that have a narrow dynamic range. We suggest twosolutions: (1) implementing a nonlinear activation function, (2)having sets of HDA accumulators with varying granularity(sensitivity) to accommodate PI homing over a range of distances.
Discussion: PI-Homing With Head-DirectionAccumulators
The HDA method is an efficient mechanism for PI homing,but is it implemented in mammalian brains? The best argu-ment for HDAs is the widespread existence of head-directioncells in mammalian brains. We argue that the creation ofHDAs is a minor developmental adjustment from the creationof a head-direction cell. Although the discovery of cells withthe firing properties of this cell type would dramaticallystrengthen the model, such cells would be hard to identifyusing the spatial recording methods frequently employed.Clearly, we feel that a search for this cell type is warranted.Additionally, it would be useful to find evidence that head-direction cell activity is gated by locomotor distance or speed.Indeed, head-direction cells have been found to increase theirfiring rates when an animal is moving compared with beinghand-held (Zugaro et al., 2001). The findings of unusual com-binations of head direction, velocity, location, and other corre-lates in retrosplenial cortex (Cho and Sharp, 2001), entorhinalcortex (Sargolini et al., 2006), and other locations (Sharp andGreen, 1994) are encouraging. An additional requirement isintegration. True neuronal integrators are found in the oculo-motor system (Major et al., 2004). The discovery of cells inthe entorhinal cortex with membrane properties that wouldsupport nonleaky integration (nonleaky Ca21 channels) is anencouraging finding (Egorov et al., 2002; Fransen et al., 2006).
The HDA model of PI homing relies on undiscovered ele-ments. This is not unusual—every navigational model that weare aware of posits one or several undiscovered elements. Forexample, the two radial-array models described above eachposit the existence of a radial array that acts as an accumulator,an operator that has not been found. Most other mammalianmodels are less explicit, but all make assumptions about neuralmechanisms that are undocumented.
VECTORS AND NAVIGATION 465
Hippocampus

One of the interesting features of the HDA model is that itdoes not make use of attractor dynamics. Although HDAs areorganized in sets, there is no requirement for synaptic interac-tion among set members. Nor is there any requirement that theHDA members in a set ‘‘learn’’ their roles. The output of the setis simple polling. Most other models make use of attractor dy-namics and Hebbian learning rules. One likely alternative to theHDA model presented here would be a modification of the ra-dial attractors described by Touretzky and colleagues (Touretzkyet al., 1993; Wan et al., 1993). In this scenario, a single radialattractor would replace a set of HDAs. To serve these needs, theTouretzky model would have to be modified to become a sum-ming attractor, such that each input would move the attractor inthe direction of the input. Such an attractor would also need (1)a reset mechanism and (2) a gain mechanism to signal the dis-tance of the current vector sum. Exploring modifications of theTouretzky sinusoidal attractor or similar models is beyond thescope of this manuscript, but would be a worthwhile endeavor.It should be noted that the sinulsoidal-attractor model has manysimilarities to the HDA model presented here. In particular, thediscussions of these articles describe how circular attractors couldcompute path-integration functions.
A final point is that the HDA homing mechanism makes nouse of place cells or any known cell type in the hippocampus.We imagine that HDAs will be found directly downstream tothe head-direction cells that serve as input. This suggests thatthey will be found at the projection sites of head-direction cells,especially those that are broadly tuned like the cells in the lateralmammillary nucleus. The projections of the HDA system shouldalso include populations of head-direction cells to permit an ani-mal to follow a head-direction cell home. Favored sites to lookfor HDA cells are the entorhinal cortex, subiculum, lateral mam-millary nucleus, anterior thalamic nucleus, and other retro-hip-pocampal areas, but not the hippocampus or dentate gyrus.
SECTION 2. CREATING AND USINGA SHORTCUT MATRIX
‘‘Piloting’’ refers to an animal’s ability to compute a naviga-tional route (Whishaw and Tomie, 1997; Mittelstaedt, 2000).PI homing is a simple example of piloting, but animals can, inprincipal use remembered vectors to compute optimal routesbetween visited locations. Having described path-segment arith-metic earlier in this article, it is straightforward to apply path-segment arithmetic to a multisegment path. The goal is tocompute a direct path between any pair of waypoint locations.
In the computer model and the figures for this article, pathsegments between waypoint locations are linear. A linear globalvector is equivalent to the sum of an indirect series of path seg-ments. This follows from the basic features of vector addition,and was demonstrated in Section 1. It should be clear there-fore, that although we have used linear path segments for sim-plicity, real-world path segments need not be linear. In Section2, we will define a path segment as a continuous locomotorpath between two pauses. We propose that, at the pause loca-
tion, the animal registers its position. Although the path con-necting pauses may not be straight, any path integrator, includ-ing the set of radial accumulators described above, will produceoutput that is identical to the straight path between the pair ofpause locations.
Figure 8 illustrates how path segment arithmetic can be usedto compute a shortcut vector between any pair of waypoints. Inthis section of the article, we describe a series of operations where
1. Shortcuts connecting every waypoint on a path to everyother waypoint are computed.2. The set of shortcuts is stored in a ‘‘shortcut matrix,’’ indexedby the start location and the stop location of each path. Eachshortcut is a heading vector.3. Subsequently, when an animal is in a start location, it can‘‘lookup’’ the stop locations available (every other waypoint onthe original journey) and select a direct path to the waypoint.The ‘‘path’’ is a vector, representing the head-direction and dis-tance the animal should travel from its current waypoint loca-tion to reach the goal waypoint. Although any number of neu-ral mechanisms for computing PI-homing could be used tobuild the shortcut matrix, our implementation will use theHDA method of Section 1.
Computational procedure—A path is entered as described inSection 1. The path segments are individually analyzed andstored as HDA sets, which are equivalent to local vectors forthe path. Each node (waypoint) can be considered a location inspace and is given an arbitrary name (e.g., ‘‘rotten log’’ or ‘‘elmtree’’). The direct vector is computed with vector arithmetic byadding the local vectors that lie between the current locationand goal. For example, if we have a path ABCDE and we wantto know the shortcut vector connecting location B to locationD, we add the local vectors BC 1 CD. After a path iscompleted, each node is considered a potential start or stoplocation. Shortcuts between all permutations of the nodes arecomputed and the path vectors are stored in a 2-keyed lookuptable. The two keys are the arbitrary names of the start andend locations, and the value returned is the shortcut pathconnecting them.
The following pseudocode gives a sense of the retrieval oper-ation. A dictionary is a lookup table, and by using dictionariesembedded within dictionaries, the two-key lookup isimplemented.
dim LocalDictionaryForStart as dictionarydim shortcut as vectorstart 5 <string name of start location>stop 5 <string name of destination>LocalDictionaryForStart 5 MasterDict.lookup(start)shortcut 5 LocalDictionaryForStart.lookup(stop)Figure 9 illustrates the rich set of paths and shortcuts that
are available to an animal after even a single exploratory excur-sion through an apparatus. Several features are apparent andrequire comment. First, while the shortcuts are generallyadequate, there are a number of shortcut vectors with errors.These are due to the 108 separation between neighbors in anHDA set. Perhaps the most important point is that, although
466 KUBIE AND FENTON
Hippocampus

FIGURE 9. Contents of the shortcut matrix. The first fourpanels illustrate the contents of the shortcut matrix for four shortexploratory journeys. Each shortcut matrix contains an estimate ofthe direct path connecting each pair of nodes. Shortcuts werecomputed using the path segment arithmetic method illustrated inFigure 9. Local vectors were estimated using the Head DirectionAccumulator method. In panels (A) through (D) the dark line isthe path and gray lines are the computed shortcuts. Path-segment
arithmetic was done using 36-element Head Direction Accumula-tor sets (108 separation of neighbors). There is a small, but notice-able error for many shortcuts. C2 and D2 repeat the calculationusing 180-element HDA sets, resulting in elimination of percepti-ble error. It should be noted that the local vectors were estimatedusing the longest element method. If we had used interpolation,the errors would have been dramatically decreased.
VECTORS AND NAVIGATION 467
Hippocampus

the paths appear to cover much of the floor of the apparatus,they come from a restricted set of start and stop waypoint loca-tions. These waypoints represent the only locations that canuse the shortcut matrix. For example, in Figure 10A, there arefive paths that cross part of the center of the floor. Althoughthese paths cross the center of the apparatus, an animal canonly access them from the endpoints. If the animal is in theapparatus center it has no access to any of the paths.
The computer program was designed to mimic how a navi-gating subject would use the shortcut matrix. Figure 10 illus-trates its operation. The left column lists the set of start loca-tions. Positioning the navigator at one start location is done byclicking one of the entries in the left column. This reveals theset of destinations that are available from the current location.Selecting a goal by clicking an entry in the ‘‘destination’’ col-umn elicits the suggested shortcut. In the first example in Fig-ure 10, the navigator is at the elm tree and chooses as a desti-nation the wine cellar. The shortcut matrix goes into thelookup table for elm tree, retrieves the shortcut vector for winecellar, and displays it to the navigator. The path vector that ispresented suggests to the navigator that if it faces 2858 (inhead-direction coordinates) and proceeds on a straight path fora distance that corresponds to the length of the arrow, it willbe at the wine cellar. If the navigator chooses to take that path,
it can find a set of head-direction cells that are tuned to thatdirection and move while maintaining peak firing of the head-direction cell set. A similar scenario is shown on the secondline, where the navigator is standing at the rotten log and sendsin inquiry to the shortcut matrix for the route to the entry towoods. The matrix suggests a path vector with a heading of408 and a distance of 200 pixels. If the navigator so chooses, itcan follow that path vector to the woods.
An important point about the shortcut matrix is that it doesnot contain a map. The maps in the right column of the figureare not available to the navigator. They serve to check that thepaths suggested by the shortcut matrix are valid.
In the computer implementation, arbitrary place names wereused as location identifiers. This is meant to indicate that theidentifier carries no spatial information, such as its x and ycoordinates. Which location identifiers could serve this functionin an animal’s brain? We will suggest two answers, which mayoverlap. The first is location-specific sensory data. The smell ofa particular tree, the appearance of a fork in the road, the viewand sound of the origin of a stream from a pond, the retinalimage of a set of landmarks. A second and more interestinglocation identifier may be the firing of a set of place cells whosefields are in a particular location. Figure 11 is a repeat of thebottom row of Figure 10, with place-cell identifiers replacing
FIGURE 10. Illustration of shortcut selection. The navigatorhas completed an exploration with five path segments created froma start location and five other waypoints. The path is in the shapeof an irregular pentagon. Using the method illustrated in Figure10, a shortcut matrix is computed and the values are stored. Thematrix is stored such that a pair of waypoints refers to the shortcutthat connects them. The waypoint coordinates are not stored andthe map (right) is not stored. (In the map depiction, dark lines arethe path and light lines are shortcuts.) The result is a lookup Tableindexed by the identities of two waypoints. The traveler uses thematrix by entering current location (left column) and then clicking
on destination. The matrix delivers the vector (shortcut) that willtake the traveler to the destination. The vector is in head-directionand distance coordinates. In the first example, the traveler is stand-ing at the elm tree and inquires for the shortcut to the wine cellar.A 2808 200 pixel vector is retrieved as the path vector. In the sec-ond example, the traveler is at the rotten log and inquires for theshortcut to the entry to woods. A 438 180 pixel heading-vector isretrieved. The right column recalls the shape of the original path,showing that the proposed paths are accurate. [Colorfigure can beviewed in the online issue which is available at www.interscience.wiley.com.]
468 KUBIE AND FENTON
Hippocampus

name identifiers (‘‘field 73’’ replaces ‘‘rotten log’’ and ‘‘field 92’’replaces ‘‘entry to woods’’). The point is that the firing of aunique set of location-specific neurons is an adequate spatialidentifier. Two facts make place cells excellent candidates. First,we know that place cells exist. Second, damage to the set ofhippocampal place cells eliminates flexible and efficientnavigation.
For the shortcut matrix to be useful, it must be recalled andused whenever an animal re-enters the environment. In otherwords, on each environmental entry, the brain state shouldreturn to the brain state that existed when the matrix wasformed or last updated. Minimally, three items must be part ofthis state. First, the shortcut matrix must be recalled. Second,the set of place cells (or location identifiers) that represent way-points should be recalled. Finally, the system of head-directioncells (HD cells and HD accumulators) should be in registerwith their original orientation. There is solid evidence thatplace cells and HD cells return to their original configurationson each entry into a familiar environment (Taube et al., 1990;Thompson and Best, 1990). It is interesting to note that Wangand Spelke, in studying human spatial processing (2002), focuson the necessity of matching the rotational orientation of thecurrent environment to a stored orientation, a process thatprobably relies of the head-direction system.
Discussion: Shortcut Matrix
The shortcut matrix may be one of a number of flexible andefficient navigational mechanisms available to mammals. Wehave attempted to show the strengths and weaknesses of thistype of implementation. The strengths are clear. With little ex-posure to an environment, an animal could learn a large num-ber of direct and efficient shortcuts. These invariably involvestraight paths. The paths are frequently novel and can traverseunexplored regions. The weaknesses are also clear. The paths
can only connect previously visited regions and they cannotanticipate what lies along the path.
The weakest element in the shortcut matrix we have presentedis its apparent reliance on a hypothetical neuron, the HDA.This weakness is not profound for two reasons. First, we thinkit likely that HDAs, or their equivalents, will be found. Theyare closely related to head-direction cells and, on a theoreticalbasis, it is highly likely that some sort of accumulator or integra-tor accounts for PI. The more salient reason is that the shortcutmatrix can be formed by any PI homing-type PI mechanism.For example, the circular attractor—PI scheme developed byTouretzky and students in the early 1990s should work in anequivalent fashion (Touretzky et al., 1993; Wan et al., 1993).Recently it has been argued that grid cells are path integrators(Fuhs and Touretzky, 2006; McNaughton et al., 2006).Although grid cells are modeled to perform PI-localization,minor modifications ought to demonstrate that they can com-pute homing vectors. In short, any mechanism capable of takingan outward, multilegged journey and computing a compositevector is capable of using the computed vectors as the basis for ashortcut matrix. The HDA was used because it provides a con-crete implementation, and because it shares some propertieswith the shortcut matrix, but other methods of adding local vec-tors should work equally well (Wan et al., 1993; Fuhs and Tour-etzky, 2006; Hasselmo and Brandon, 2008).
Since the shortcut matrix has a profile of strengths andweaknesses, use of this type of navigation ought to have detect-able effects on navigational performance. Below, we list a set ofattributes of navigational behavior that are suggested by theproperties of the shortcut matrix. Some predictions can betested with available data. Other predictions could be tested infuture studies.
1. The shortcut matrix should support one trial learning. Onetrial exploratory exposure ought to be sufficient for moderatelyflexible paths.
FIGURE 11. Shortcut selection with place-cell input. This fig-ure is a replot of the bottom row of Figure 11, with place celllabels replacing place name labels. This illustrates the notion thatthe lookup table can respond to queries from two place cells in afashion similar to queries of other place identifiers. In this case,we imagine that the traveler is at the elm tree causing place cell 73
to fire and present one input to the matrix. The traveler wants togo to the wine cellar, causing place cell 92 to fire and present asecond input to the shortcut matrix. With two place cell inputs,the matrix returns an accurate heading vector. [Color figure can beviewed in the online issue which is available at www.interscience.wiley.com.]
VECTORS AND NAVIGATION 469
Hippocampus

2. When the shortcut matrix is employed it ought to be diffi-cult for the animal to navigate to or from unvisited locations.3. Use of the shortcut matrix depends on reliable orientationto the inertial frame. Aspects of an animal’s initial behavior onentry into a familiar environment ought to reflectreorientation.4. The shortcut matrix depends heavily on head-direction cells.Any manipulation that alters the function of head-directioncells ought to affect shortcut navigation.5. The shortcut matrix depends on PI. If PI is manipulated ordisrupted, one should see parallel changes in navigationalbehavior.6. Use of the shortcut matrix ought to result in straight-pathnavigation, even during errors and uncertainty.7. Use of the shortcut matrix ought to induce behavior patternssuch that, on entry into a familiar environment, a rat first movesto a familiar location—a waypoint—before navigating to the goal.
Although the beginnings of a navigational model have beenpresented, many issues remain unsettled. What happens whenan animal explores more than once? Do multiple exploratorypaths merge or are they treated separately? Are all pause pointsin all entries into an environment recorded as waypoints?What happens when an animal pauses in the same locationtwice? Can a shortcut matrix complement other types ofspatial and navigational learning? Can a shortcut matrix aidin avoiding barriers or obstacles? These should be addressedin future work.
Where is the shortcut matrix? Candidates cover the hippo-campal formation and retro-hippocampal areas. The require-ments are place cell inputs and projection to a head-directioncell region or a region containing the hypothesized HDAs.
Notions similar to the shortcut matrix can be found in theinsect literature, especially the work on bees (Collett et al., 1998;Wehner, 2003), and in Hasselmo and Brandon’s discussion of theproperties of cells that exhibit graded persistent firing (2008).
SECTION 3. USE OF THE SHORTCUT MATRIXIN BOUNDED SPACE AND EXTENDED SPACE
Bounded space refers to relatively small spatial regions sepa-rated from larger spatial frames by a clear boundary. An ani-mal’s representation of a bounded space is extremely sensitiveto the shape of the enclosure, making the rectangular and cy-lindrical enclosures frequently used in place-cell studies goodmodels of different spaces (Cheng, 1986; Gallistel and Cramer,1996). There is abundant evidence that after an animal hasmoderate exposure to this type of environment, place cell firingfields will cover all available locations (Muller and Kubie,1987). We argue, however, that shortcut paths will fill the spaceeven more rapidly, and an animal will use shortcut navigationfor early navigational challenges within the enclosure.
In Section 2, we considered the problem of successive explo-rations creating indefinite expansion of the shortcut matrix.
Partially to limit this problem we will postulate three things asfollows:
1. Initial exploration is concentrated on spatial boundaries, onsalient features that mark the boundaries, and on landmarkswithin the space.2. The shortcut matrix is largely frozen after the exploratoryperiod.3. Addition or manipulation of landmarks induces a briefadditional period of plasticity.
Exploration
If we follow these rules then the shortcut matrix will beextremely sensitive to the shape and size of a bounded space.We ran a simple test to examine the relationship between ex-ploratory behavior and the hypothetical shortcut matrix. It iscommonly stated that, on investigating a novel environment,rats will explore the perimeter and salient objects. To test theseideas, we obtained a set of exploratory paths in novel environ-ments to see the types of shortcut matrices that could becreated.
Method
Five rats were tested in each of the two novel enclosures.One enclosure was triangular (80 cm on a side), the other rec-tangular (90 cm 3 60 cm), and each had a novel toy objectaffixed to the floor toward the center of the enclosure. The sub-jects had been trained on the ‘‘place accuracy task’’ (Kubieet al., 2007) and given months of handling and exposure to acylindrical enclosure. Each rat was given a 3-min exposure toeach novel enclosure. A rat’s behavior was recorded with digitalvideo, and paths and pauses were automatically traced. Eachpause was matched with a segment of the video and scored forrearing. For each trial, the set of pauses was used as a series ofwaypoints in constructing a shortcut matrix.
Results
Exploratory patterns were consistent and as predicted. Pathsfocused on the perimeter of accessible space, outlining theshape of the apparatus. Invariably, there was at least one jour-ney to the object. Pauses occurred at the corners, at one or twopositions along the wall and at the object. Most importantly,the shortcut matrices constructed from each pause-waypoint se-ries effectively traversed accessible space and constructed severalpaths to the central object.
Figure 12A illustrates the paths for a rat in the two enclo-sures, and 12B shows the hypothetical shortcut matrices thatthese paths would produce. If the purpose of exploration wasto set up the firing of the complete set of place cells, one mightexpect initial exploration to be widespread exploration resem-bling either a random walk or an organized effort to cover eacharea of space. This is not what is seen. Rather, initial explora-tion is characterized by exploration of the boundaries, especially
470 KUBIE AND FENTON
Hippocampus

boundary corners, and exploration of salient landmarks. Openspace is minimally traversed. Such an exploratory pattern isexcellent for setting up a set of shortcuts and for sensitive rep-resentation of the shape of the apparatus. Similar patterns ofboundary and landmark exploration can be seen in mammalsranging from rodents to humans (Bohbot et al., 2002; Leveret al., 2006).
Extended space
Extended space refers to regions of space much larger thanlaboratory space. The most dramatic examples are the migra-tory environments of birds and various sea animals. Avianmigration is particularly instructive. Birds appear to make use
of a ‘‘sun compass’’ for general heading information and land-mark recognition for path correction (Bingman and Cheng,2005). Hippocampal damage appears to spare the sun compass,which leaves heading signals intact, while waypoint (landmark)recognition is affected, especially waypoints that define spacenear the roost (Bingman and Jones, 1994). Navigation by thesun compass and the shortcut matrix share a critical feature incommon: both are accomplished by a heading vector. We feelit highly likely that both rely heavily on head-direction cells forheading, and suggest that they share other common mecha-nisms. Waypoint recognition appears to be a hippocampalfunction. If this is correct then navigation based on vectorheadings would be a global navigational mechanism, playing asignificant role both in extended space and bounded space.
FIGURE 12. Exploratory behavior in bounded space. Exam-ples of rat exploration of a rectangular enclosure (left) and a trian-gular enclosure (right) (method details are in the following para-graph). These examples are representative of the five exploratorysessions in each apparatus. (A) Paths during the first 30 s of explo-ration with pauses and rears marked with circles. Note that pathsparallel the walls, with occasional excursions toward the object.Pauses are always exhibited at the object and each of the visited
corners. In addition, in each exploratory session there were atmost two additional pauses, which were always at select pointsalong a wall. (B) Hypothetical shortcut matrix created from thepaths and pause locations of part A. (C) Path plots are extendedto 180 s. Note that the preferred pauses and paths of the first 30 sare regularly revisited, with the center of the apparatus rarelyvisited.
VECTORS AND NAVIGATION 471
Hippocampus

Speculating further, we can imagine the situation depicted inFigure 13, where heading vectors have two functions. Theyconnect regions within a bounded space by efficient shortcutvectors; and they form connections between regions ofbounded space. According to this notion in an extended-space,heading-vector systems will tie together scattered spatial regionsand, in a similar manner, connect heterogeneous representationsystems into a single global entity.
GENERAL DISCUSSION
The inspiration for the model presented here comes fromvarious navigational models for insects, particularly ants andbees (Collett and Collett, 2002; Wehner, 2003). Analysis ofinsect navigation spans a century, and has culminated in elegantstudies of foraging and homing. Ant homing behavior is partic-ularly well studied and understood. Noteworthy in the insectliterature is that insects have no function resembling hippocam-pal place recognition. Rather, insect navigation is computedfrom and guided by heading vectors.
Two features differentiate the heading-vector model pre-sented here and other models of mammalian navigation. First,the model is based on heading vectors provided by head-direc-tion cells. Second, the heading signal originates outside of thehippocampus.
Since the publication of ‘‘The Hippocampus as a CognitiveMap,’’ (O’Keefe and Nadel, 1978) studies of the neurobiologyof place recognition and navigation have been dominated bythe hippocampus. At the time of the book’s publication, the re-markable and dominant actor was the hippocampal place cell.In the interim, two additional remarkable cell types have beendiscovered: head-direction cells (Ranck, 1985; Taube et al.,1990) and grid cells (Hafting et al., 2005). Not surprisingly, invirtually every model place cells convey the self-localization sig-nal (‘‘you are here’’). This seems sensible. More surprising, inthe models for navigation, place cells continue to play thedominant role and navigational computations are performedlargely within the hippocampus. This is likely a mistake. Theextrahippocampal head-direction cells are ideal candidates toserve as a long-distance heading signal.
The closest reference to a heading-based model is in theoriginal O’Keefe and Nadel proposition, which has only a one-paragraph description of navigational models. The end of theparagraph reads as follows:
These two will sum, bringing representation of the placecontaining food in that environment close to activation. Sincethe place that the animal occupies will also be receiving afferentdrive, there will be two sets of place representations that arepotentiated. The programming system can now search for theappropriate motor program that activates both of these repre-sentations simultaneously; this program will take the animalfrom where it is to where the food is.
FIGURE 13. Extended space example. Four regions ofbounded space are shown embedded in extended space. Eachregion of bounded space has both place cell firing fields and head-ing vectors (small arrows). The larger arrows connecting thebounded regions illustrate heading vectors in extended space. A
navigator with this type of composite map would know the head-ings and distances connecting the four bounded regions (travelnorth west to go from the court house to forest park), but neednot have the intervening space mapped with place cells.
472 KUBIE AND FENTON
Hippocampus
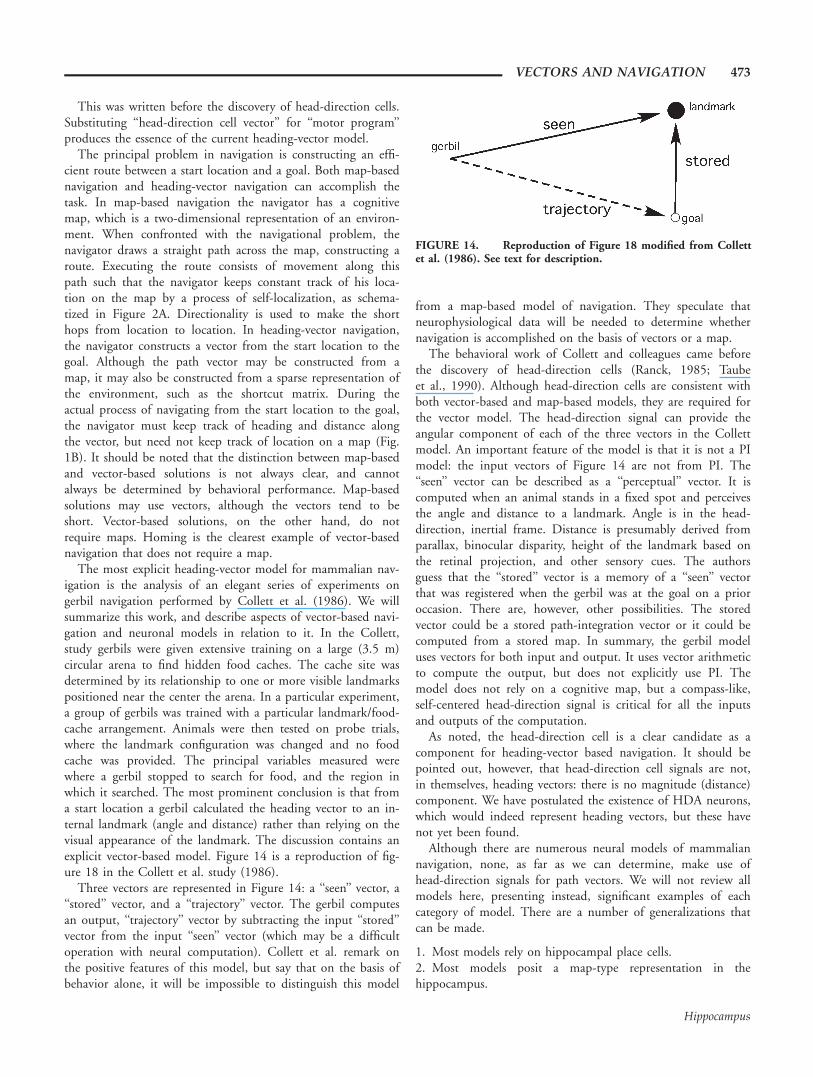
This was written before the discovery of head-direction cells.Substituting ‘‘head-direction cell vector’’ for ‘‘motor program’’produces the essence of the current heading-vector model.
The principal problem in navigation is constructing an effi-cient route between a start location and a goal. Both map-basednavigation and heading-vector navigation can accomplish thetask. In map-based navigation the navigator has a cognitivemap, which is a two-dimensional representation of an environ-ment. When confronted with the navigational problem, thenavigator draws a straight path across the map, constructing aroute. Executing the route consists of movement along thispath such that the navigator keeps constant track of his loca-tion on the map by a process of self-localization, as schema-tized in Figure 2A. Directionality is used to make the shorthops from location to location. In heading-vector navigation,the navigator constructs a vector from the start location to thegoal. Although the path vector may be constructed from amap, it may also be constructed from a sparse representation ofthe environment, such as the shortcut matrix. During theactual process of navigating from the start location to the goal,the navigator must keep track of heading and distance alongthe vector, but need not keep track of location on a map (Fig.1B). It should be noted that the distinction between map-basedand vector-based solutions is not always clear, and cannotalways be determined by behavioral performance. Map-basedsolutions may use vectors, although the vectors tend to beshort. Vector-based solutions, on the other hand, do notrequire maps. Homing is the clearest example of vector-basednavigation that does not require a map.
The most explicit heading-vector model for mammalian nav-igation is the analysis of an elegant series of experiments ongerbil navigation performed by Collett et al. (1986). We willsummarize this work, and describe aspects of vector-based navi-gation and neuronal models in relation to it. In the Collett,study gerbils were given extensive training on a large (3.5 m)circular arena to find hidden food caches. The cache site wasdetermined by its relationship to one or more visible landmarkspositioned near the center the arena. In a particular experiment,a group of gerbils was trained with a particular landmark/food-cache arrangement. Animals were then tested on probe trials,where the landmark configuration was changed and no foodcache was provided. The principal variables measured werewhere a gerbil stopped to search for food, and the region inwhich it searched. The most prominent conclusion is that froma start location a gerbil calculated the heading vector to an in-ternal landmark (angle and distance) rather than relying on thevisual appearance of the landmark. The discussion contains anexplicit vector-based model. Figure 14 is a reproduction of fig-ure 18 in the Collett et al. study (1986).
Three vectors are represented in Figure 14: a ‘‘seen’’ vector, a‘‘stored’’ vector, and a ‘‘trajectory’’ vector. The gerbil computesan output, ‘‘trajectory’’ vector by subtracting the input ‘‘stored’’vector from the input ‘‘seen’’ vector (which may be a difficultoperation with neural computation). Collett et al. remark onthe positive features of this model, but say that on the basis ofbehavior alone, it will be impossible to distinguish this model
from a map-based model of navigation. They speculate thatneurophysiological data will be needed to determine whethernavigation is accomplished on the basis of vectors or a map.
The behavioral work of Collett and colleagues came beforethe discovery of head-direction cells (Ranck, 1985; Taubeet al., 1990). Although head-direction cells are consistent withboth vector-based and map-based models, they are required forthe vector model. The head-direction signal can provide theangular component of each of the three vectors in the Collettmodel. An important feature of the model is that it is not a PImodel: the input vectors of Figure 14 are not from PI. The‘‘seen’’ vector can be described as a ‘‘perceptual’’ vector. It iscomputed when an animal stands in a fixed spot and perceivesthe angle and distance to a landmark. Angle is in the head-direction, inertial frame. Distance is presumably derived fromparallax, binocular disparity, height of the landmark based onthe retinal projection, and other sensory cues. The authorsguess that the ‘‘stored’’ vector is a memory of a ‘‘seen’’ vectorthat was registered when the gerbil was at the goal on a prioroccasion. There are, however, other possibilities. The storedvector could be a stored path-integration vector or it could becomputed from a stored map. In summary, the gerbil modeluses vectors for both input and output. It uses vector arithmeticto compute the output, but does not explicitly use PI. Themodel does not rely on a cognitive map, but a compass-like,self-centered head-direction signal is critical for all the inputsand outputs of the computation.
As noted, the head-direction cell is a clear candidate as acomponent for heading-vector based navigation. It should bepointed out, however, that head-direction cell signals are not,in themselves, heading vectors: there is no magnitude (distance)component. We have postulated the existence of HDA neurons,which would indeed represent heading vectors, but these havenot yet been found.
Although there are numerous neural models of mammaliannavigation, none, as far as we can determine, make use ofhead-direction signals for path vectors. We will not review allmodels here, presenting instead, significant examples of eachcategory of model. There are a number of generalizations thatcan be made.
1. Most models rely on hippocampal place cells.2. Most models posit a map-type representation in thehippocampus.
FIGURE 14. Reproduction of Figure 18 modified from Collettet al. (1986). See text for description.
VECTORS AND NAVIGATION 473
Hippocampus

3. Models fall into two nonoverlapping categories: those thatcompute routes and those that use vectors.
A. Models that compute routes create lists of place cellsbetween pairs of locations. They do not use vectors.B. Models that rely on vectors do PI for self-localization;they do not do route calculations. Thus, the vectors in theseprior models are principally used for self-localization.
4. Route calculations are not closely tied to head-directioncells. The notion of tracking the firing of a head-direction cellor a head-direction cell ensemble to follow a straight path to-ward a goal is not incorporated in any model.
We will use the framework of these four features to compareand contrast the present model with prior work. Note that themodel we present is distinct in all four of these categories. It doesnot rely on place cells; it neither posits nor depends on a map-like representation; it relies on heading vectors for navigationbetween places but not for self-localization; it is based on head-direction cells for computing and following navigation vectors.
Relevance of Place Cells
Almost all current models of rodent navigation exploit the dis-coveries that a population of hippocampal place cells provides aplace code (O’Keefe and Dostrovsky, 1971), and that lesions ofthe hippocampus impair flexible navigation (Morris et al., 1982).The basic problem of navigation is to compute the way from thecurrent location, ‘‘here’’ to a target location, ‘‘there’’ (Fig. 15).Although place cell firing can signal ‘‘here’’ (Wilson andMcNaughton, 1993), no clear signal for ‘‘there’’ has beendescribed. Naturally, models of navigation derive the ‘‘here’’ sig-nal from place cell activity. It was proposed that location-specificplace cell firing arises when perception of an environmental fea-ture matches the stored value of that feature (Zipser, 1985; Sharp,1991). In contrast, a series of models from the McNaughtongroup postulate that synaptically connected place cells with adja-cent firing fields have stronger excitatory synapses than cells withdistant firing fields. Such connectivity with global inhibitionforms a self-perpetuating continuous attractor network (Samso-novich and McNaughton, 1997). Networks of this type will onlyallow cells with firing fields at a single location to be coactive.Such a network can perform PI without any environmental stim-ulation, because the coactive set of cells will change smoothly asthe rat moves from one location to the next (McNaughton et al.,1996; Samsonovich and McNaughton, 1997; Tsodyks, 1999;Stringer et al., 2002). It is important to recall the distinctionbetween PI-self-localization and PI-homing, because in thesemodels, PI is explicitly for self-localization and it is not specifiedhow navigating from one place to another is accomplished. Mul-ler et al. (1996) pointed out that if the real-world distancebetween a pair of firing fields is stored as the inverse of the synap-tic strength (‘‘synaptic resistance’’) between the cells, then the in-formation needed for navigation is available in the matrix of syn-aptic strengths. In such models, navigation would be accom-plished by a hypothetical mechanism that could search for thelowest synaptic resistance pathway between the cells with firing
fields at the current and target locations. Hasselmo et al. (2002)implement a mechanism similar to Muller et al., but involvingseveral hippocampal regions and exploiting theta rhythmicity.
Other models of navigation have used place cell firing torepresent information in addition to the current location.O’Keefe (1991) proposed that collective place-cell firing definesthe environment’s centroid, providing the origin of an allocen-tric coordinate system for navigational computations. Touretzkyand Redish (1996) used place cells to associate sensory and idi-othetic cues. This would serve the important function of keep-ing self-localization by PI in register with the environment.Models by Sharp and colleagues (Brown and Sharp, 1995;Sharp et al., 1996) as well as those of Touretzky and Redish(1996) proposed that the place signal is an input to the motorsystem, localized in the nucleus accumbens. When rewards areencountered during exploration, the reinforcement strengthensassociations between the recent useful movements and the loca-tions in which they occurred. This could mediate a form ofinstrumental learning for navigation driven by the stored asso-ciation of place cell firing with useful local movements. Burgessand O’Keefe used a related idea for navigation. They hypothe-sized that there are goal cells downstream of the hippocampusand suggested that during exploration, the presence of rewardstrengthens the association between recently active place cellsand the active goal cell (1996). Subsequent navigation to thegoal is accomplished by moving to increase the firing of the
FIGURE 15. Schematic of the problem of finding an optimalpath when the direct path crosses an unvisited region of space. Inthis example, mapped space is U-shaped. Several models that findthe shortest path through a series of place-cell firing fields will notbe able to cross the unvisited region.
474 KUBIE AND FENTON
Hippocampus

goal cell by enlarging the subset of active place cells that arestrongly associated with the particular goal cell.
A common theme of all these navigation models is that oneway or another, before initiating movement or during travel, asequence of place cells must be activated to navigate betweentwo locations. With the exception of the McNaughton models,the models require extensive exploration of the environmentprior to navigation, because a navigation route driven by placecells cannot cross a previously unexplored expanse. This isbecause the cells with firing fields in the void had not been pre-viously active. Navigating through uncharted territory is possi-ble in the McNaughton model, because it hypothesizes that thesynaptic structure of the place cell network is innately precon-figured prior to any experience in the environment. All thesefeatures contrast with the model we have presented, because inour model, place cell firing indicates current location but is notrequired for any aspect of the computation of the way fromhere to there.
Relevance of a Map-Like Representation
Many navigation models utilize a map-like representation ofspace, conforming to the cognitive mapping theory, incorporat-ing the properties of place cells and consequences of hippocam-pal lesions. The models designed to form continuous attractornetworks are a particularly robust embodiment of a cognitivemap, because they always signal one location and faithfullytrack position through the encoded space, even in the absenceof external inputs (Samsonovich and McNaughton, 1997; Tso-dyks, 1999; Kali and Dayan, 2000; Stringer et al., 2002; Con-klin and Eliasmith, 2005; Fuhs and Touretzky, 2006). It is veryimportant to consider that continuous attractor models of navi-gation were designed to build a map-like representation of theenvironment, mimic and explain place cell firing properties,and perform robust PI-localization. As pointed out by Mittel-staedt (2000), these models, however, do not perform PI-hom-ing or other forms of efficient navigation. Although theyexplain how spatial information can be stored and organized(Muller et al., 1996), these models do not provide many cluesabout how the information for piloting can be extracted tomediate getting from ‘‘here’’to ‘‘there’’ by straight-line paths.
Another set of models assumes the existence of a map-likerepresentation of the environment and focuses instead on howinformation in the map can be used to navigate between places(Mittelstaedt, 2000). The most elaborate models of this typethat are constrained by known physiological signals, are thoseof Touretzky and Redish (Wan et al., 1993; Redish and Tour-etzky, 1998). The map-like place cell representation serves as amechanism to maintain registration between PI-localization,and environmental stimuli that are processed in a local viewmodule. Head-direction cells (discussed below) also serve aregistration function by providing a directional bearing for thesystem of navigation networks. Recce and Harris (1996) takeadvantage of map-like representations to perform coordinatetransformations between an egocentric neocortical map derivedfrom PI and an allocentric map derived from place cell firing.
Note, that PI is used to derive information for self-localizationbut not for planning or finding efficient ways to a target. Sincethe information for navigation is contained in the map, theauthors, like many, used an arbitrary, physiologically unrealisticalgorithm to search the map for the shortest distance betweenthe current location and a location of a stored goal (Hether-ington and Shapiro, 1993; Muller et al., 1996).
A handful of authors have proposed physiologically plausiblesolutions that can extract the information from a place-cell net-work. One group proposed a gradient ascent mechanism onthe basis of hypothetical goal cells (Burgess et al., 1994; Bur-gess and O’Keefe, 1996, 2003). These extra-hippocampal goalcells are activated when the rat encounters reward, which trig-gers the goal cell to store the direction the rat is facing and thelocation that is being signaled by a set of cells that are broadlytuned to position like cells in the subiculum or ventral hippo-campus. The result is that the activity in the population of goalcells signals the direction and distance to a target location, andthis activity increases as the subject approaches the goal. Theuse of goal cells allows the system to take efficient detours, anddirect shortcuts across uncharted spaces. Although a map-likerepresentation was used to tune the firing of goal cells, the goalcells themselves did not form a map-like representation. Bilkeyand Clearwater (2005) used an essentially similar gradientascent mechanism by ignoring the spatial signal in place cellfiring and taking advantage of the finding that there is anincreased density of place cell firing fields at a goal in the watermaze (Hollup et al., 2001). Similarly, other models that pro-posed physiologically plausible methods for goal-directed navi-gation used location-specific and direction-specific signals totune motor output elements that themselves were not organizedin a map-like manner (Brown and Sharp, 1995; Sharp et al.,1996; Touretzky and Redish, 1996; Redish and Touretzky,1998). An interesting generalization from this work is thatwhile a map-like representation is useful for storing, organizing,and manipulating spatial information, models that can use thisinformation to navigate in a physiologically plausible mannerall have relied on directed representations of goals that werethemselves decidedly not map-like. In general, these modelspermit efficient navigation to a single goal in the environmentand use firing-rate gradients rather than vectors to guide move-ment. The navigation model we propose does not depend on amap-like representation of the environment. The shortcut ma-trix representation we propose stores only the straight-linepaths between the subset of visited regions that were designatedas waypoints, and based on monitoring heading from a head-direction signal, the model provides an explicit, simple, maxi-mum-rate mechanism for selecting the direct path to an arbi-trarily distant target waypoint location.
We propose that a shortcut matrix is setup to store directpaths between all pairs of registered locations in an environ-ment. Although the shortcut matrix provides a form of repre-sentation of the environment, it does not have important prop-erties of a map. The distances between arbitrary locations arenot represented, because the matrix only stores the heading vec-tors between the subset of locations that have been registered
VECTORS AND NAVIGATION 475
Hippocampus

during exploration. The shortcut matrix permits piloting acrossuncharted territory from a charted start location to a chartedtarget location. However, the shortcut matrix has no informa-tion for a navigator that starts out from an uncharted location.
Relevance of Route and Vector Computations
Most models of navigating from one place to another haverelied on computing routes through a definite sequence of pla-ces. Distance is coded in synaptic strength in these models.The network either comes preconfigured (Samsonovich andMcNaughton, 1997) or relies on an extensive exploratory pe-riod to visit all parts of the environment, so that the synapticweights can be appropriately set between place cells with neigh-boring firing fields (Hetherington and Shapiro, 1993; Mulleret al., 1996). Models with this arrangement of synaptic weightsare able to do PI, which is inherently vector-based. However,PI is used for self-localization in these models, rather than forroute calculation, or determining a heading vector. These mod-els capture important features of the place cell representation,but they are poor mechanisms for navigation. Learning requiresextensive environmental exploration, there is no guarantee thatthe path of least synaptic resistance will be a straight line andthese systems cannot find paths that cross regions of unvisitedspace, without some additional mechanism (Burgess andO’Keefe, 1996; Muller et al., 1996).
A minority of navigation models have used explicit vectorcomputations. A vector in polar coordinates (r,u) can be repre-sented as a phasor, which is a sinusoidal function of fixed fre-quency with the form: f (x) 5 r ! cos(xx 1 f), x is a variablelike location or time, where r is the amplitude, e is the (con-stant) frequency, and f is the phase. O’Keefe (1991) suggestedthat rats may use phasors to encode perceptual vectors to envi-ronmental features. The phase was set by the theta rhythm inthe hippocampal electroencephalogram. A major advantage ofthe phasor formulation is that vector addition is accomplishedsimply by summing across the relevant population of phase-tuned neurons. Vectors in this early O’Keefe model are percep-tual and are used to aid self-localization rather than navigation.Touretzky et al. (1993) demonstrate that an array of sinusoidsof differing phase can be used to do phasor arithmetic. In addi-tion, they describe how phasors can compute trajectories as for-mulated by Collett et al. (1986). This Touretzky et al. modelhas some of the features of our heading vector model, althoughthey are not well fleshed out. Our navigation model’s vectorarithmetic is similar to that described by Touretzky et al.(1993), but is implemented by summing the fixed-frequencyelements of an array of sinusoids of differing phase. Phase isdetermined by the preferred head direction of each sinusoid inthe heading-vector model we present.
Relevance of Head-Direction Cells
Many navigation models have incorporated head-directioncells. One role for these cells is to maintain registrationbetween internal representations of place and the physical land-
marks in the environment (McNaughton et al., 1991). In themodels of Redish and Touretzky (1998), head-direction cellsprovide inputs to the local view and path integrator modules,which maintain registration with external cues through theirinteractions with the place cell system. A second role for head-direction cells is in PI itself. Directional cells provide bearinginformation to a path integrator, which along with movementinformation is used for self-localization by PI.
The concept of a continuous attractor and its importance fornavigation was initially developed on the basis of head-direc-tion cells. Skaggs et al. (1995) and Zhang (1996) describedthat a one-dimensional attractor system could be realized by aset of recurrently connected head-direction cells, if strong mu-tual excitation connects the cells with similar preferred direc-tions and mutual inhibition connects the cells with differentpreferred directions. Although the recurrent connectivity wasneither necessary, nor realistic for these brain regions (Song andWang, 2005), the concept was important. Indeed, they are theconceptual basis of the two-dimensional continuous attractornetworks that are used to account for location-specific placecell properties and self-localization by PI (Samsonovich andMcNaughton, 1997).
Head-direction cells are central to the heading-vectormodel we present. Head-direction cells provide the onlyinput to the path integrator. Unlike prior models, thehypothesized path integrator is itself a type of head-directioncell, the HDA. We hypothesize the existence of a set of accu-mulating head-direction cells for PI to compute a headingbetween two locations. A complete set of these cells performsvector arithmetic that is formally described by phasor calcula-tions. The head-direction cell is also the output of our navi-gation model, providing the heading vector from a locationto a goal. We hypothesize that the set of heading vectorsbetween all pairs of registered locations (pauses or waypoints)forms a shortcut matrix. The start and target locations arethe two keys used to retrieve a heading from the shortcut ma-trix. These keys are the only nondirectional elements that areused in the heading-vector model. Places, as provided by theplace-cell code, would be natural keys for querying the short-cut matrix, but other items of knowledge, like a local view,will work just as well. Although we have described the inputs,outputs, and computations of the navigation system in termsof single head-direction cell-like elements, we note that themodel works just as well, and in fact more robustly, if head-direction information is provided by a network of cells withattractor network properties. In particular, the model benefitsfrom the attractor properties of stability and stationarity.These features are characteristic of real head-direction cellnetworks, which is why head-direction cells are the archetypeof a continuous attractor neural network.
Location of the Path Integrator
Our model places PI outside of the hippocampus, in regionsincluding or related to head-direction cells. Two laboratorygroups have addressed the issue of whether the path integrator
476 KUBIE AND FENTON
Hippocampus

is in the hippocampus and have come to different conclusions.Alyan and McNaughton (1999) trained rats to follow acrooked tunnel, emerge from the tunnel and choose a route to-ward the start location. They found that both normal rats andrats with hippocampal lesions took direct return paths.Whishaw and colleagues trained rats to follow a string-guidedindirect path from the home nest to a food stash. Rats invaria-bly carry the food stash back to the nest. In a well-lit room,rats with hippocampal lesions and control rats took a straightreturn path to the home nest. However, in darkness, only thecontrol rats could return directly to the nest (Whishaw andGorny, 1999; Clark et al., 2005). Although we would like tosee a definitive answer on this issue, we take the strong resultsof Alyan and McNaughton as supportive. In related studies,Whishaw group’s finding that damage to retrosplenial corteximpairs PI homing supports a critical role for extrahippocampalhead-direction cells (Whishaw et al., 2001). Finally, Worsleyet al. (2001) found that patients with right temporal lobe dam-age performed poorly on PI homing tasks. Since the affectedgroup had damage to both the hippocampus and parahippo-campal gyrus, the results do not localize the PI system. A highpriority for future studies should be localization of PI circuitry.Current work with grid cells, suggests the medial entorhinalcortex is a good candidate (Sargolini et al., 2006).
Human Navigation
Behavioral studies in humans support the proposition thatnavigation is based on heading vectors rather than step-by-stepmovement along a path of identifiable locations (Wang andSpelke, 2002; Waller and Haun, 2003; Muehl and Sholl, 2004;Spiers and Maguire, 2007). For example, Bailenson et al.(2000) found that, given a choice between equal-distance routesbetween two points, subjects reliably chose the route whose ini-tial segment direction was closer to the global vector to thegoal. Spiers and Maguire have studied patterns of activity inthe human brain during navigation, and correlated these withthe navigators’ perception of the process (2006). The authors’summary is that navigation alternates between planning a route(or a route segment) and executing the plan. Functional imag-ing suggests that the hippocampus is involved in the planningphase, while nonhippocampal structures such as subiculum,entorhinal cortex, parietal lobe, and prefrontal cortex are activeduring execution. These results and speculations fit with ourmodel.
Limitations, Speculations, and Future Directions
We acknowledge that the vector-heading model we have pre-sented is a simple, initial step. In particular, the shortcut-matrixmodel only applies to navigation after a single limited exposureto a new environment. Among the issues not addressed are:what happens with multiple exposures? Does the shortcut ma-trix contribute to formation of place cell representations? Canthe shortcut matrix, or modified versions, solve detour prob-
lems? Our review of the human and animal behavioral litera-ture suggests that most navigation is guided by heading vectorsrather than intrahippocampal route-based mechanisms. A parsi-monious suggestion is that head-direction cells guide manyforms of navigation.
Acknowledgments
The authors thank Eduard Kelemen, James Ranck, and twoanonymous reviewers for helpful comments. They also thankKhaula Tauqeer who gathered the path data in Section 3.
REFERENCES
Aksay E, Baker R, Seung HS, Tank DW. 2000. Anatomy and dis-charge properties of pre-motor neurons in the goldfish medullathat have eye-position signals during fixations. J Neurophysiol84:1035–1049.
Alyan S, McNaughton BL. 1999. Hippocampectomized rats are capa-ble of homing by path integration. Behav Neurosci 113:19–31.
Bailenson JN, Shum MS, Uttal DH. 2000. The initial segment strat-egy: A heuristic for route selection. Mem Cognit 28:306–318.
Bilkey DK, Clearwater JM. 2005. The dynamic nature of spatialencoding in the hippocampus. Behav Neurosci 119:1533–1545.
Bingman V, Cheng K. 2005. Mechanisms of animal global navigation:Comparative perspectives and enduring challenges. Ethol Ecol Evol17:295–318.
Bingman VP, Jones TJ. 1994. Sun compass-based spatial learningimpaired in homing pigeons with hippocampal lesions. J Neurosci14 (11 Part 1):6687–6694.
Bohbot VD, Jech R, Ruzicka E, Nadel L, Kalina M, Stepankova K,Bures J. 2002. Rat spatial memory tasks adapted for humans:Characterization in subjects with intact brain and subjects withselective medial temporal lobe thermal lesions. Physiol Res 51(Suppl 1):S49–S65.
Brown MA, Sharp PE. 1995. Simulation of spatial learning in theMorris water maze by a neural network model of the hippocampalformation and nucleus accumbens. Hippocampus 5:171–188.
Burgess N, O’Keefe J. 1996. Neuronal computations underlying thefiring of place cells and their role in navigation. Hippocampus6:749–762.
Burgess N, O’Keefe J. 2003. Spatial models of the hippocampus. In:Arbib MA, editor. The Handbook of Brain Theory and NeuralNetworks, 2nd ed. Cambridge: MIT press. pp 539–543.
Burgess N, Recce M, O’Keefe J. 1994. A model of hippocampal func-tion. Neural Netw 7:1065–1081.
Cheng K. 1986. A purely geometric module in the rat’s spatial repre-sentation. Cognition 23:149–178.
Cheng K. 1989. The vector sum model of pigeon landmark use.J Exp Psychol Anim Behav Process 15:366–375.
Cho J, Sharp PE. 2001. Head direction, place, and movementcorrelates for cells in the rat retrosplenial cortex. Behav Neurosci115:3–25.
Clark BJ, Hines DJ, Hamilton DA, Whishaw IQ. 2005. Movementsof exploration intact in rats with hippocampal lesions. Behav BrainRes 163:91–99.
Collett M, Collett TS, Bisch S, Wehner R. 1998. Local and globalvectors in desert ant navigation. Nature 394:269–272.
VECTORS AND NAVIGATION 477
Hippocampus

Collett TS, Collett M. 2002. Memory use in insect visual navigation.Nat Rev Neurosci 3:542–552.
Collett TS, Cartwright BA, Smith BA. 1986. Landmark learning andvisuo-spatial memories in gerbils. J Comp Physiol [A] 158:835–851.
Conklin J, Eliasmith C. 2005. A controlled attractor network modelof path integration in the rat. J Comput Neurosci 18:183–203.
Egorov AV, Hamam BN, Fransen E, Hasselmo ME, Alonso AA. 2002.Graded persistent activity in entorhinal cortex neurons. Nature420:173–178.
Etienne AS, Jeffery KJ. 2004. Path integration in mammals. Hippo-campus 14:180–192.
Etienne AS, Maurer R, Berlie J, Reverdin B, Rowe T, GeorgakopoulosJ, Seguinot V. 1998. Navigation through vector addition. Nature396:161–164.
Etienne AS, Maurer R, Boulens V, Levy A, Rowe T. 2004. Resettingthe path integrator: A basic condition for route-based navigation.J Exp Biol 207 (Part 9):1491–1508.
Fransen E, Tahvildari B, Egorov AV, Hasselmo ME, Alonso AA. 2006.Mechanism of graded persistent cellular activity of entorhinal cor-tex layer v neurons. Neuron 49:735–746.
Fuhs MC, Touretzky DS. 2006. A spin glass model of path integrationin rat medial entorhinal cortex. J Neurosci 26:4266–4276.
Gallistel CR. 1980. The Organization of Action: A New Synthesis,Vol. 13. Hillsdale N.J., New York: L. Erlbaum Associates; distrib-uted by Halsted Press. 432 p.
Gallistel CR, Cramer AE. 1996. Computations on metric maps inmammals: Getting oriented and choosing a multi-destination route.J Exp Biol 199 (Part 1):211–217.
Hafting T, Fyhn M, Molden S, Moser MB, Moser EI. 2005. Micro-structure of a spatial map in the entorhinal cortex. Nature436:801–806.
Hasselmo ME. 2007. Arc length coding by interference of thetafrequency oscillations may underlie context-dependent hippocam-pal unit data and episodic memory function. Learn Mem 14:782–794.
Hasselmo ME, Brandon MP. 2008. Linking cellular mechanisms tobehavior: Entorhinal persistent spiking and membrane potentialoscillations may underlie path integration, grid cell firing, and epi-sodic memory. Neural Plast 2008:658323.
Hasselmo ME, Hay J, Ilyn M, Gorchetchnikov A. 2002. Neuromodu-lation, theta rhythm and rat spatial navigation. Neural Netw 15:689–707.
Hasselmo ME, Giocomo LM, Zilli EA. 2007. Grid cell firing mayarise from interference of theta frequency membrane potentialoscillations in single neurons. Hippocampus 17:1252–1271.
Hetherington PA, Shapiro ML. 1993. A simple network model simu-lates hippocampal place fields. II. Computing goal-directed trajec-tories and memory fields. Behav Neurosci 107:434–443.
Hollup SA, Molden S, Donnett JG, Moser MB, Moser EI. 2001.Accumulation of hippocampal place fields at the goal location inan annular watermaze task. J Neurosci 21:1635–1644.
Kali S, Dayan P. 2000. The involvement of recurrent connections inarea CA3 in establishing the properties of place fields: A model.J Neurosci 20:7463–7477.
Kubie JL, Fenton A, Novikov N, Touretzky D, Muller RU. 2007.Changes in goal selection induced by cue conflicts are in registerwith predictions from changes in place cell field locations. BehavNeurosci 121:751–763.
Lever C, Burton S, O’Keefe J. 2006. Rearing on hind legs, environ-mental novelty, and the hippocampal formation. Rev Neurosci17:111–133.
Lewis D. 1994. We, the Navigators: The Ancient Art of Landfind-ing in the Pacific, Vol. 21. Honolulu: University of Hawaii Press.442 p.
Major G, Baker R, Aksay E, Mensh B, Seung HS, Tank DW. 2004.Plasticity and tuning by visual feedback of the stability of a neuralintegrator. Proc Natl Acad Sci USA 101:7739–7744.
McNaughton BL, Chen LI, Markus EJ. 1991. ‘‘Dead Reckoning,’’landmark learning, and the sense of direction: A neurophysiologicaland computational hypothesis. J Cogn Neurosci 3:190–202.
McNaughton BL, Barnes CA, Gerrard JL, Gothard K, Jung MW,Knierim JJ, Kudrimoti H, Qin Y, Skaggs WE, Suster M, WeaverKL. 1996. Deciphering the hippocampal polyglot: The hippocam-pus as a path integration system. J Exp Biol 199 (Part 1):173–185.
McNaughton BL, Battaglia FP, Jensen O, Moser EI, Moser MB. 2006.Path integration and the neural basis of the ‘cognitive map’. NatRev Neurosci 7:663–678.
Mittelstaedt H. 2000. Triple-loop model of path control by headdirection and place cells. Biol Cybern 83:261–270.
Morris RG, Garrud P, Rawlins JN, O’Keefe J. 1982. Place navigationimpaired in rats with hippocampal lesions. Nature 297:681–683.
Muehl KA, Sholl MJ. 2004. The acquisition of vector knowledge andits relation to self-rated direction sense. J Exp Psychol Learn MemCogn 30:129–141.
Muller RU, Kubie JL. 1987. The effects of changes in the environ-ment on the spatial firing of hippocampal complex-spike cells.J Neurosci 7:1951–1968.
Muller RU, Stead M, Pach J. 1996. The hippocampus as a cognitivegraph. J Gen Physiol 107:663–694.
O’Keefe J. 1991. The hippocampal cognitive map and navigationalstrategies. In: Paillard J, editor. Brain and Space. Oxford, UK:Oxford University Press. pp 273–295.
O’Keefe J, Burgess N. 2005. Dual phase and rate coding in hippocam-pal place cells: Theoretical significance and relationship to entorhi-nal grid cells. Hippocampus 15:853–866.
O’Keefe J, Dostrovsky J. 1971. The hippocampus as a spatial map.Preliminary evidence from unit activity in the freely-moving rat.Brain Res 34:171–175.
O’Keefe J, Nadel L. 1978. The Hippocampus As a Cognitive Map,Vol. 14. Oxford, New York: Clarendon Press; Oxford UniversityPress.570 p.
Ranck JB Jr. 1985. Head direction cells in the deep cell layer of dorsalpresubiculum in freely moving rats. In: Buzsaki GaV CH, editor.Electrical Activity of the Archicortex. Budapest: Akademiai Kiadeo.pp 217–220.
Recce M, Harris KD. 1996. Memory for places: A navigational modelin support of Marr’s theory of hippocampal function. Hippocam-pus 6:735–748.
Redish AD, Touretzky DS. 1998. The role of the hippocampus insolving the Morris water maze. Neural Comput 10:73–111.
Samsonovich A, McNaughton BL. 1997. Path integration and cogni-tive mapping in a continuous attractor neural network model.J Neurosci 17:5900–5920.
Sargolini F, Fyhn M, Hafting T, McNaughton BL, Witter MP, MoserMB, Moser EI. 2006. Conjunctive representation of position,direction, and velocity in entorhinal cortex. Science 312:758–762.
Sharp PE. 1991. Computer simulation of hippocampal place cells.Psychobiology 19:103–115.
Sharp PE, Green C. 1994. Spatial correlates of firing patterns of singlecells in the subiculum of the freely moving rat. J Neurosci14:2339–2356.
Sharp PE, Blair HT, Brown M. 1996. Neural network modeling ofthe hippocampal formation spatial signals and their possible role innavigation: A modular approach. Hippocampus 6:720–734.
Skaggs WE, Knierim JJ, Kudrimoti HS, McNaughton BL. 1995. A modelof the neural basis of the rat’s sense of direction. In: Tesauro G, Tour-etzky D, Leen T, editors. Advances in Neural Information ProcessingSystems 7. Cambridge: MIT Press, Massachusetts. p 173–180.
Song P, Wang XJ. 2005. Angular path integration by moving ‘‘hill ofactivity’’: A spiking neuron model without recurrent excitation ofthe head-direction system. J Neurosci 25:1002–1014.
Spiers HJ, Maguire EA. 2006. Thoughts, behaviour, and brain dynam-ics during navigation in the real world. Neuroimage 31:1826–1840.
478 KUBIE AND FENTON
Hippocampus

Spiers HJ, Maguire EA. 2007. A navigational guidance system in thehuman brain. Hippocampus 17:618–626.
Stackman RW, Taube JS. 1998. Firing properties of rat lateral mam-millary single units: Head direction, head pitch, and angular headvelocity. J Neurosci 18:9020–9037.
Stringer SM, Rolls ET, Trappenberg TP, de Araujo IE. 2002. Self-organizing continuous attractor networks and path integration:Two-dimensional models of place cells. Network 13:429–446.
Taube JS, Muller RU, Ranck JB Jr. 1990. Head-direction cellsrecorded from the postsubiculum in freely moving rats. I. Descrip-tion and quantitative analysis. J Neurosci 10:420–435.
Thompson LT, Best PJ. 1990. Long-term stability of the place-field ac-tivity of single units recorded from the dorsal hippocampus offreely behaving rats. Brain Res 509:299–308.
Touretzky DS, Redish AD. 1996. Theory of rodent navigation basedon interacting representations of space. Hippocampus 6:247–270.
Touretzky DS, Redish AD, Wan HS. 1993. Neural representations ofspace using sinusoidal arrays. Neural Comput 5:869–884.
Tsodyks M. 1999. Attractor neural network models of spatial maps inhippocampus. Hippocampus 9:481–489.
Vickerstaff RJ, Di Paolo EA. 2005. Evolving neural models of pathintegration. J Exp Biol 208 (Part 17):3349–3366.
Waller D, Haun DB. 2003. Scaling techniques for modeling direc-tional knowledge. Behav Res Methods Instrum Comput 35:285–293.
Wan HS, Touretzky DS, Redish AD. 1993. Towards a ComputationalTheory of Rat Navigation. Proceedings of the 1993 ConnectionistModels Summer School pp 11–19. Erlbaum, Hillsdale, NJ.
Wang R, Spelke E. 2002. Human spatial representation: Insights fromanimals. Trends Cogn Sci 6:376.
Wehner R. 2003. Desert ant navigation: How miniature brains solvecomplex tasks. J Comp Physiol A Neuroethol Sens Neural BehavPhysiol 189:579–588.
Whishaw IQ, Gorny B. 1999. Path integration absent in scent-track-ing fimbria-fornix rats: Evidence for hippocampal involvement in‘‘sense of direction’’ and ‘‘sense of distance’’ using self-movementcues. J Neurosci 19:4662–4673.
Whishaw IQ, Tomie JA. 1997. Piloting and dead reckoning dissoci-ated by fimbria-fornix lesions in a rat food carrying task. BehavBrain Res 89:87–97.
Whishaw IQ, Maaswinkel H, Gonzalez CL, Kolb B. 2001. Deficits inallothetic and idiothetic spatial behavior in rats with posterior cin-gulate cortex lesions. Behav Brain Res 118:67–76.
Wilson MA, McNaughton BL. 1993. Dynamics of the hippocampalensemble code for space. Science 261:1055–1058.
Worsley CL, Recce M, Spiers HJ, Marley J, Polkey CE, Morris RG.2001. Path integration following temporal lobectomy in humans.Neuropsychologia 39:452–464.
Zhang K. 1996. Representation of spatial orientation by the intrinsicdynamics of the head-direction cell ensemble: A theory. J Neurosci16:2112–2126.
Zipser D. 1985. A computational model of hippocampal place fields.Behav Neurosci 99:1006–1018.
Zugaro MB, Tabuchi E, Fouquier C, Berthoz A, Wiener SI. 2001.Active locomotion increases peak firing rates of anterodorsal tha-lamic head direction cells. J Neurophysiol 86:692–702.
VECTORS AND NAVIGATION 479
Hippocampus




















