
HABITAT LOSS AND FRAGMENTATION IN DYNAMIC LANDSCAPES: AVIAN PERSPECTIVES FROM THE BOREAL FOREST *
15
April 2002 375 HABITAT FRAGMENTATION: A REASSESSMENT 375 Ecological Applications, 12(2), 2002, pp. 375–389 q 2002 by the Ecological Society of America HABITAT LOSS AND FRAGMENTATION IN DYNAMIC LANDSCAPES: AVIAN PERSPECTIVES FROM THE BOREAL FOREST FIONA K. A. SCHMIEGELOW 1,3 AND MIKKO MO ¨ NKKO ¨ NEN 2 1 Department of Renewable Resources, University of Alberta, Edmonton, Alberta, Canada T6G 2H1 2 Department of Biology, University of Oulu, P.O. Box 3000, FIN-90014 Oulu, Finland Abstract. Although habitat loss and fragmentation are widely regarded as major factors contributing to the decline of many populations, the relative importance of each phenomenon is seldom evaluated. Some researchers have questioned the generality of responses to habitat fragmentation, given variation in life history characteristics, the natural dynamics of sys- tems, and land use patterns. Furthermore, a fundamental mismatch may exist between ecological theory, with its emphasis on the spatial configuration of habitats, and empirical observations of population response. Nevertheless, the paucity of quantitative land man- agement guidelines often leads to inappropriate generalizations of conservation paradigms to regional issues. We reviewed the empirical evidence for true fragmentation effects in boreal bird communities in Fennoscandia and Canada, and concluded that most responses may be attributed to pure habitat loss in landscapes where forest harvesting is the dominant land use practice. In these dynamic landscapes, total forest cover may not change, and predicting patterns of species decline requires identification of the habitats and species of concern. We constructed simple empirical models of benchmark communities in boreal forests of Finland and Canada based on species composition, species abundance distribution, and habitat requirements, in order to identify features of bird species sensitive to the loss of older forests. These models require a solid understanding of the underlying structure of the community of interest, and predict species loss based on a random-sample hypothesis. Our results were consistent with observed patterns of bird population decline and species loss in these regions. This approach provides null models for comparison with habitat remnants in order to test for fragmentation effects, and a basis for more detailed exploration of population dynamics and persistence in these systems. The results of our review and analyses indicated that system- and species-specific considerations are important when assessing the potential outcome of habitat loss and fragmentation on regional biota. Indis- criminate application of conservation paradigms may lead to misguided research efforts and poor management guidelines. Key words: boreal birds; boreal forest; Canada; dynamic landscapes; Finland; forest harvesting; habitat fragmentation; habitat loss; random sample hypothesis. INTRODUCTION Habitat loss and fragmentation are widely regarded as major factors contributing to the decline of forest bird populations (e.g., Rappole 1996). Although re- search has focused for more than three decades on the ecological effects of fragmentation, recently there has been considerable debate as to the relative influence of habitat loss and changes in spatial configuration on population trajectories in disturbed landscapes (e.g., With 1999, Fahrig 2002). As well, the context in which habitat loss and fragmentation occur is being consid- ered more carefully (e.g., Haila 1999). A recent review by Harrison and Bruna (1999) indicated a lack of con- gruence between ecological theory and results from Manuscript received 31 May 2000; revised 18 April 2001; accepted 6 May 2001; final version received 1 June 2001. For reprints of this Invited Feature, see footnote 1, p. 319. 3 E-mail: [email protected] empirical studies of fragmentation. Theory emphasizes the role of dispersal, landscape connectivity, and spatial configuration of landscape elements for population per- sistence, whereas existing empirical evidence high- lights the importance of habitat degradation (e.g., through physical and biological edge effects). Destruction of natural habitats involves three factors, often acting in concert: (1) habitat loss, (2) area and edge effects, and (3) habitat isolation. The last two factors are termed habitat fragmentation (Andre ´n 1994, Fahrig 1997). It is important to distinguish between pure habitat loss and true fragmentation effects when assessing population responses, because resultant land management recommendations may differ. When hab- itat is lost from the landscape, individuals are also lost, resulting in population declines. Habitat loss also im- plies a corresponding increase in other landscape ele- ments (e.g., Rolstad 1991), which may affect the dy- namics in the remaining, original habitat. Fragmenta-
Transcript of HABITAT LOSS AND FRAGMENTATION IN DYNAMIC LANDSCAPES: AVIAN PERSPECTIVES FROM THE BOREAL FOREST *

April 2002 375HABITAT FRAGMENTATION: A REASSESSMENT
375
Ecological Applications, 12(2), 2002, pp. 375–389q 2002 by the Ecological Society of America
HABITAT LOSS AND FRAGMENTATION IN DYNAMIC LANDSCAPES:AVIAN PERSPECTIVES FROM THE BOREAL FOREST
FIONA K. A. SCHMIEGELOW1,3 AND MIKKO MONKKONEN2
1Department of Renewable Resources, University of Alberta, Edmonton, Alberta, Canada T6G 2H12Department of Biology, University of Oulu, P.O. Box 3000, FIN-90014 Oulu, Finland
Abstract. Although habitat loss and fragmentation are widely regarded as major factorscontributing to the decline of many populations, the relative importance of each phenomenonis seldom evaluated. Some researchers have questioned the generality of responses to habitatfragmentation, given variation in life history characteristics, the natural dynamics of sys-tems, and land use patterns. Furthermore, a fundamental mismatch may exist betweenecological theory, with its emphasis on the spatial configuration of habitats, and empiricalobservations of population response. Nevertheless, the paucity of quantitative land man-agement guidelines often leads to inappropriate generalizations of conservation paradigmsto regional issues. We reviewed the empirical evidence for true fragmentation effects inboreal bird communities in Fennoscandia and Canada, and concluded that most responsesmay be attributed to pure habitat loss in landscapes where forest harvesting is the dominantland use practice. In these dynamic landscapes, total forest cover may not change, andpredicting patterns of species decline requires identification of the habitats and species ofconcern. We constructed simple empirical models of benchmark communities in borealforests of Finland and Canada based on species composition, species abundance distribution,and habitat requirements, in order to identify features of bird species sensitive to the lossof older forests. These models require a solid understanding of the underlying structure ofthe community of interest, and predict species loss based on a random-sample hypothesis.Our results were consistent with observed patterns of bird population decline and speciesloss in these regions. This approach provides null models for comparison with habitatremnants in order to test for fragmentation effects, and a basis for more detailed explorationof population dynamics and persistence in these systems. The results of our review andanalyses indicated that system- and species-specific considerations are important whenassessing the potential outcome of habitat loss and fragmentation on regional biota. Indis-criminate application of conservation paradigms may lead to misguided research effortsand poor management guidelines.
Key words: boreal birds; boreal forest; Canada; dynamic landscapes; Finland; forest harvesting;habitat fragmentation; habitat loss; random sample hypothesis.
INTRODUCTION
Habitat loss and fragmentation are widely regardedas major factors contributing to the decline of forestbird populations (e.g., Rappole 1996). Although re-search has focused for more than three decades on theecological effects of fragmentation, recently there hasbeen considerable debate as to the relative influence ofhabitat loss and changes in spatial configuration onpopulation trajectories in disturbed landscapes (e.g.,With 1999, Fahrig 2002). As well, the context in whichhabitat loss and fragmentation occur is being consid-ered more carefully (e.g., Haila 1999). A recent reviewby Harrison and Bruna (1999) indicated a lack of con-gruence between ecological theory and results from
Manuscript received 31 May 2000; revised 18 April 2001;accepted 6 May 2001; final version received 1 June 2001. Forreprints of this Invited Feature, see footnote 1, p. 319.
3 E-mail: [email protected]
empirical studies of fragmentation. Theory emphasizesthe role of dispersal, landscape connectivity, and spatialconfiguration of landscape elements for population per-sistence, whereas existing empirical evidence high-lights the importance of habitat degradation (e.g.,through physical and biological edge effects).
Destruction of natural habitats involves three factors,often acting in concert: (1) habitat loss, (2) area andedge effects, and (3) habitat isolation. The last twofactors are termed habitat fragmentation (Andren 1994,Fahrig 1997). It is important to distinguish betweenpure habitat loss and true fragmentation effects whenassessing population responses, because resultant landmanagement recommendations may differ. When hab-itat is lost from the landscape, individuals are also lost,resulting in population declines. Habitat loss also im-plies a corresponding increase in other landscape ele-ments (e.g., Rolstad 1991), which may affect the dy-namics in the remaining, original habitat. Fragmenta-

376 INVITED FEATURE Ecological ApplicationsVol. 12, No. 2
tion effects (decreases in patch size and increases inthe amount of edge and in patch isolation) can com-pound the effects of pure habitat loss, producing aneven greater population decline (Bender et al. 1998),which may result in population extinction even if suit-able habitat still exists.
In highly modified forest systems, where forest landshave been converted for agricultural production or ur-ban expansion, the identification of remnant naturalhabitats and associated edges is often much clearer thanin similar forest systems managed for timber produc-tion. Forest conversion results in near static landscapeconfigurations; in contrast, forestry practices result indynamic landscapes that remain primarily forested,while undergoing spatial and temporal changes in com-position and age structure. In this context, the sharpnessof edges varies with forest regeneration and succession.Therefore, it seems reasonable to expect that biotic andabiotic responses to these human disturbances mightdiffer among forest systems with varying landscapecontexts. There is also considerable variation in thetypes and rates of natural disturbances within forestsystems, which might influence the response of biotato human-mediated habitat change. Despite increasingrecognition that naive application of conservation par-adigms to regional land management issues is mis-guided, a paucity of quantitative guidelines can resultin inappropriate generalizations. We use the applicationof fragmentation concepts in boreal forest research andmanagement as a case in point.
The circumpolar boreal forest is one of the earth’smost extensive terrestrial systems. Historically, large-scale natural disturbances, such as fire and insect out-breaks, resulted in forests that were both spatially andtemporally dynamic. Fire still acts as a major agent ofdisturbance in Canada and Russia, whereas intensivesuppression has largely removed it from boreal forestsin the Nordic countries. Although some conversion ofboreal forest for agricultural production has occurredalong the southern periphery of its distribution, muchof the boreal forest is actively managed for timber pro-duction. Because they have a relatively short historyof exploitation, the boreal forests of Russia and Canadacontain almost half of the world’s intact pristine forest(Bryant et al. 1997).
Our objective is to evaluate the application of frag-mentation paradigms, based largely on studies of tem-perate forests in agricultural mosaics, to managed bo-real forests. We begin with a brief overview of broad-scale population trends and bird community structurein Fennoscandia and Canada. We then review the em-pirical evidence for fragmentation effects on borealbird communities in these regions. Secondly, we pre-sent a null model approach for rapid assessment of theoutcome of habitat loss and the existence of fragmen-tation effects in these dynamic landscapes. We use datafrom Finland and Canada to generate predictive models
for bird population decline and species loss in borealforests of these regions, based on the identification ofcommon features of sensitive species. We concludewith some general guidelines for research and man-agement in dynamic forested systems.
BROAD-SCALE TRENDS IN BOREAL BIRD POPULATIONS
Birds are the richest vertebrate taxon in boreal for-ests, comprising .75% of all terrestrial vertebrate spe-cies (Smith 1993, Monkkonen and Viro 1997). How-ever, this diversity fluctuates seasonally, as many birdspecies that breed in the north migrate to winter farthersouth. In Canada, long-distance migrants (species thatwinter in the tropics) comprise 50% of the breedingspecies, followed by short-distance migrants (specieswintering in temperate areas) at 30%, and permanentresidents at 20% (Erskine 1977). In Fennoscandia,short-distance migrant species dominate (45%), fol-lowed by long-distance migrants (30%) and permanentresidents (Niemi et al. 1998). In both regions, residentspecies are disproportionately rare; migrants comprise;90% of all breeding individuals.
There is a long history of monitoring bird popula-tions in Fennoscandia, particularly in Finland. The old-est population estimates for birds date back to the early20th century, and broad-scale population data existsince the 1940s. Finnish data indicate that more specieshave increased in abundance than have decreased withthe conversion of relatively natural forest landscapesto intensively managed landscapes (Jarvinen et al.1977). The general trend between the 1940s and 1990shas been for many of the common species to becomemore numerous, while many of the rarer species havedeclined. In northern Finland, where the changes inlandscape structure have been the most dramatic duringthe 20th century, data suggest 16 significant populationdecreases and 16 increases among forest birds sincethe 1940s (Haila and Jarvinen 1990).
Helle and Jarvinen (1986) concluded that a substan-tial proportion of the population changes of bird speciesin northern Finland could be traced to a change in theavailability of preferred habitats. Although species pre-ferring edges or early successional habitats have in-creased, species requiring old forest and avoiding edgeshave decreased with the extensive harvesting of north-ern Finnish boreal forests. Among declining species,certain life history traits are particularly pronounced(Imbeau et al. 2001). These include habitat-relatedtraits, such as large area requirements, association withold-growth forests, and dependence on decaying woodfor foraging, as well as nesting-related traits—cavitynesting, in particular. These traits were especially com-mon among resident species, which are prominentamong species that have declined. However, patternsobserved at the scale of individual forest remnants arenot always consistent with regional population trendsin northern Finland (Vaisanen et al. 1986). For ex-

April 2002 377HABITAT FRAGMENTATION: A REASSESSMENT
ample, Virkkala (1991) found that population declinesof birds in managed forests of northern Finland werenot observed in virgin forest remnants. Although pop-ulations of Capercaillie (Tetrao urogallus), Three-toedWoodpecker (Picoides tridactylus), Siberian Tit (Paruscinctus), Siberian Jay (Perisoreus infaustus), and PineGrosbeak (Pinicola enucleator) have declined overallin northern Finland, they have remained relatively sta-ble in large blocks of virgin forest.
Comparable data for bird populations in boreal Can-ada are lacking. Breeding bird atlases have been con-ducted in some regions (e.g., Cadman et al. 1987, Se-menchuk 1992), but these are one-time snapshots ofgeneral distributions. Some North American BreedingBird Surveys (BBS) have been conducted in borealforests (USGS Patuxent Wildlife Research Center2000), but visitation has been erratic and, because theyare roadside counts, they do not sample forest specieswell (F. K. A. Schmiegelow, unpublished data). In gen-eral, existing broad-scale coverage is sparse and is con-centrated at the southern edge of the boreal region. Aswell, most ecological studies are relatively recent, lo-calized, and/or short in duration.
A summary of the 70 BBS routes that fall within theCanadian boreal forest found that over the past 30 yr(1966–1996), five bird species had experienced signif-icant population declines, while two had increased(Niemi et al. 1998). None of the declining species ex-hibit strong affinities for older forests (perhaps due tosampling bias), but all are long-distance (neotropical)migrants. Trends from the Ontario Forest Bird Moni-toring Program (Welsh 1995) for the 1987–1997 timeperiod show five species increasing significantly andthree declining (in Niemi et al. 1998). Two of the de-clining species are short-distance migrants associatedwith older forest. In a recent review, Imbeau et al.(2001) suggest that in conifer forests of Quebec, theboreal species most sensitive to forest change are res-idents, most of which are cavity nesters reliant on olderforests.
Despite a recent increase in avian research in Ca-nadian boreal forests, documentation of broad-scalepatterns in bird population trends for this region isseriously hampered by a paucity of data and lack ofstandardized sampling techniques. Generalizations arefurther confounded by geographic variation in forestcover types, natural disturbance histories, and associ-ated bird communities. This is of significant concern,given the recent acceleration of harvesting throughoutthese forests.
EVIDENCE FOR FRAGMENTATION EFFECTS
IN BOREAL FORESTS
Our goal is to assess the empirical evidence for frag-mentation effects, but most studies we reviewed didnot clearly distinguish between the effects of fragmen-tation (changes in habitat configuration) and those due
to habitat loss, nor were they designed to. McGarigaland Cushman (2002) provide guidelines on experi-mental approaches to address this problem. We attemptto separate these effects by classifying studies intothose examining potential edge effects (biological andphysical), area effects, and isolation effects. Withineach category, we critically evaluate the interpretationof results with respect to the strength of the evidencefor fragmentation per se, vs. patterns that might alsobe explained by habitat loss. We briefly review thepotential effects of additional changes in species in-teractions that could result from modified abundancerelations, due to either habitat loss or fragmentation.
Biological edge effects
Nest predation.—Recent compilations of data fromtemperate forests show a general increase in nest pre-dation rates with decreases in forest cover (Robinsonet al. 1995, Hartley and Hunter 1998), which could beattributed to either habitat loss or fragmentation. How-ever, many temperate-zone studies have demonstratedincreased predation on bird nests at forest edges (e.g.,Gates and Gysel 1978, Wilcove 1985, Small and Hunter1988). In the boreal forest, this edge effect appears tobe more variable. A general pattern in boreal forestsseems to be that increased nest predation occurs atforest–agricultural land edges (Andren et al. 1985, Kui-tunen and Helle 1988, Andren 1995, Bayne and Hobson1997, Hannon and Cotterill 1998). At forest–clearcutedges, however, an increase in nest predation is notevident in most cases (Andren 1995, Hanski et al. 1996,Huhta et al. 1996, 1998, Bayne and Hobson 1997, Songand Hannon 1999; but see Monkkonen et al. 2000).
Disturbance by forest harvesting and by forest clear-ing for agriculture differ in their consequences for nest-ing birds in boreal landscapes. This has been attributedto larger changes that take place in nest predator com-munities with habitat loss due to agriculture rather thanto forest harvesting, due to the introduction of novelhabitat. Andren (1992) showed that corvid densitiesare higher in agricultural landscapes, resulting in in-creased predation rates close to forest–farmland edgesand in small forest fragments surrounded by agricul-tural land. Likewise, in boreal Canada, Hannon andCotterill (1998) showed increased predation rates bycorvids, but not by small mammals, at forest–farmlandedges. Bayne and Hobson (1997) attributed higher ratesof predation in agricultural vs. managed forest land-scapes to a greater diversity of predators at edges andto higher densities of red squirrels (Tamiasciurus hud-sonicus) in remnant forest patches of the agriculturallandscape.
It is widely appreciated that the effects of predationon the breeding success of birds are dependent, amongother things, on the landscape context and the predatorcommunity (Martin 1993, Andren 1995, Bayne et al.1997, Tewksbury et al. 1998). Angelstam (1986) and

378 INVITED FEATURE Ecological ApplicationsVol. 12, No. 2
Andren (1992) suggested that the steepness in the pro-ductivity gradient between forests and the adjacent ma-trix is the main determinant of predation rates at edges.The sharper the gradient, the more pronounced theedge-related predation. Accordingly, nests in forestpatches in agricultural landscapes suffer from higherpredation rates because the matrix supports a high den-sity and diversity of nest predators.
Although increased nest predation is often implicat-ed in the decline of temperate forest bird populations(Whitcomb et al. 1981, Terborgh 1989), direct evidenceis scanty. In boreal birds, the most convincing supportfor the role of increased nest predation in relation topopulation declines concerns forest grouse in Finland.Habitat change in Fennoscandian forest landscapes hasincreased the abundance of generalist predators (Kurkiet al. 1998) and reduced the breeding success ofground-nesting forest grouse (Kurki et al. 1997). Si-multaneous decline of all forest grouse species in Fin-land since the 1960s (Linden and Rajala 1981, Tornberget al. 1999), despite different habitat associations(Swenson and Angelstam 1993), suggests a commonreason. Increased predation seems the most plausiblefactor contributing to declines in these ground-nestingbirds; however, these effects are exacerbated by habitatfragmentation only where forests have been convertedfor agriculture (Kurki et al. 2000). Nevertheless, whileKurki et al. (2000) conclude that increased forest frag-mentation results in reduced breeding success of forestgrouse, and is a probable cause of population declines,most of the variation in breeding success could be at-tributed to increasing percentages of agricultural landin southern areas, and a decreasing percentage of olderforests due to forest harvesting in northern areas.
The general pattern in boreal forests is that forestharvesting does not usually lead to elevated nest pre-dation rates at edges (Andren 1995), but may result inan overall increase in nest predation rates in harvestedlandscapes (Kurki et al. 1997). Most nest predators inboreal landscapes are habitat generalists, which onlysecondarily feed on bird eggs and nestlings. Their spa-tial distribution and abundance varies according to thedistribution and abundance of their main food. Nestpredation appears to be linked to vole cycles or seedcrops and to overall increases in small-mammal den-sities in harvested landscapes. For example, Cotterilland Hannon (1999) found significant temporal varia-tion in predation rates in boreal forests of western Can-ada that had been recently harvested. Similarly, a three-year study in northern Finland revealed high between-year variation, as well as significant spatial variationin predation rates (E. Huhta, J. Jokimaki, M. Monk-konen, and M. Korhonen, unpublished data). In thisstudy, predation pressure varied as predicted by thealternative prey hypothesis (Angelstam et al. 1984,1985), with highest rates in clear-cut areas during avole peak year, and in forest habitats in vole crash years
(see also Huhta 1996). Although increased predationmay contribute to long-term bird population declinesin northern boreal forests, as suggested for forestgrouse in Finland, this may be explained by changesin available habitat due to forest harvesting, rather thanto fragmentation effects per se.
Brood parasitism.—The Brown-headed Cowbird(Molothrus ater) is a brood parasite of many speciesof North American passerines, and has been implicatedin the decline of migratory forest songbirds in tem-perate forests of eastern and central North America(e.g., Bohning-Gaese et al. 1993). Although there issome evidence that parasitism rates increase with de-creasing forest cover and increasing fragmentation(Robinson et al. 1995), Hahn and Hatfield (1995) sug-gest that the proximity of agricultural infrastructures(fields, cattle, farms) can result in high parasitism ratesin large forested areas, even when forest cover is high.
In a recent review of edge effects in western NorthAmerican forests, Kremsater and Bunnell (1999) con-clude that, even in agricultural settings, patterns ofcowbird parasitism are not consistent, and that there isno evidence for elevated parasitism rates in managedforests of northwestern North America. Cowbirds are,in fact, very rare in these systems (Schieck et al.1995a). Similarly, Schmiegelow and Hannon (1999)report that cowbirds are rare and parasitism rates verylow in recently harvested North American boreal for-est. Cowbirds are recent colonists in the boreal forest,having been associated with shortgrass prairies histor-ically, and potential host species may exhibit few be-havioral defense mechanisms to avoid parasitism (e.g.,Hobson and Villard 1998). However, unless clearing offorests is associated with conversion for agriculture,cowbirds seem unlikely to reach high abundances inNorth American boreal forests.
The Cuckoo (Cuculus canorus), also a brood para-site, is a relatively common species in Fennoscandianforests (Vaisanen et al. 1998), but there is no indicationthat parasitism rates vary with habitat loss or frag-mentation. Cuckoos have been present in these forestsfor a long time. Thus it is likely that host species haveevolved avoidance behaviors to reduce parasitism rates(Davies 2000). In fact, Brooke and Davies (1987) sug-gest that the Cuckoo’s reproductive success is limitedby its ability to find and deceive suitable hosts. Forexample, von Haartman (1981) suggested that FinnishCuckoo populations are tracking the annual fluctuationsin populations of the Redstart (Phoenicurus phoeni-curus), its main host, rather than causing them.
Physical edge effects
Forest harvesting has resulted in an increasedamount of edges in boreal forest landscapes (Angelstam1992), causing an increase in physical edge effects inremnant forests (e.g., Esseen 1994). Physical edge ef-fects include such factors as the increased amount of

April 2002 379HABITAT FRAGMENTATION: A REASSESSMENT
sunlight, wind, temperature variation, and other chang-es to microclimatic conditions (Matlack 1993, 1994,Chen et al. 1995). Esseen (1994) showed that changesin microclimatic conditions could result in an increasedmortality of trees, ground herbs, and epiphytic lichensat edges. Indirectly, physical edge effects may haveimportant consequences for forest birds. Increasedamounts of forest edges in harvested landscapes mayaffect food availability. For example, in central Swed-ish boreal forests, the density of arboreal insects (themain food of many forest birds) was reduced in man-aged, selectively logged forests in comparison with un-managed stands (Pettersson 1996, Pettersson et al.1995). The reduction in arthropod densities was relatedto reduced arboreal lichen abundance in the managedstands. These studies were conducted in late winter,when invertebrate abundance is lowest and small dif-ferences may be critical to foraging birds. Althoughresults from selectively harvested stands cannot be di-rectly compared to forest edges, Esseen (1994) notesthat arboreal lichens are particularly sensitive to phys-ical edge effects; therefore, food availability may alsobe reduced for insectivorous birds in winter in edge-dominated landscapes.
In contrast, some studies have reported increasedarthropod densities at forest edges in summer. For ex-ample, Jokimaki et al. (1998) collected arthropods innorthern Finnish boreal forests and concluded that ar-thropod abundance decreased from forest edge to forestinterior; larger forest patches (.5 ha) had lower abun-dance than smaller patches. The total number of ar-thropods was positively correlated with the density ofthe shrub layer, which in turn was denser at forest edgesthan in forest interiors. Likewise, Helle and Muona(1985) and Halme and Niemela (1993) have shown thatColeoptera species were more abundant at forest edgesthan in forest interior habitats. In boreal forests of west-ern Canada, Song (1998) found no difference in thebiomass of flying insects between clearcut edges andinterior forest, but increases in the biomass of ground-dwelling arthropods at edges. The contrasting result forflying insects may be explained by differences in foresttype. Although most Fennoscandian studies have beenconducted in conifer-dominated forests, Song’s (1998)study focused on deciduous-dominated forest. In thislatter forest system, creation of edges by clear-cuttingresulted in decreases in shrub cover at edges, due tothe prolific suckering of trembling aspen, Populus tre-muloides, and balsam poplar, P. balsamifera, (Harper1999). In summary, food availability (arthropod den-sities) at edges may be reduced in winter, but possiblyenhanced in summer, as a result of forest harvesting.If winter conditions are regulating populations of birds(for a review, see Newton 1994), net effects of forestlandscape changes are likely to be negative, at least forboreal forest residents. Such negative effects could be
attributed to habitat loss, fragmentation, or an inter-action of the two.
Bird densities at edges
A logical question arising from the patterns in bio-logical and physical edge effects is whether or not theyaffect habitat selection by birds. Villard (1998) con-cluded that area-sensitive bird species do not avoidedges in most forested areas of North America. Onecould, in fact, hypothesize that in some boreal forestlandscapes, bird densities at forest–clearcut edgesshould be elevated because of increased food avail-ability, without concomitant increases in nest predationor parasitism. This pattern should be particularly clearamong migrant insectivores in coniferous forests. Birddensities were ;40% higher at forest edges than inforest interior habitats in a northern Finnish study(Helle 1983). All species clearly preferring edges were,as expected, insectivorous. Very few species clearlyavoided edges; among them were both insectivores(e.g., Song Thrush, Turdus philomelos, and Pied Fly-catcher, Ficedula hypoleuca) and seed eaters (cross-bills, Loxia spp., and Siskin, Carduelis spinus). How-ever, Helle and Jarvinen (1986) suggest that sedentaryspecies of old forests (titmice, Parus spp., Tree Creeper,Certhia familiaris, and Siberian Jay, Perisoreus in-faustus), as a group, clearly avoid edges. Nevertheless,Sklepkovych (1997) showed that nesting success in theSiberian Jay was negatively correlated with distance toedge, and indicated that jays may, in fact, gain foragingbenefits from forest edges. In western Canada, Song(1998) found no evidence for increased densities ofboreal birds at deciduous forest–clearcut edges.
Species interactions
If habitat loss and degradation result in modifiedcommunity structure, ecological interactions may alsobe altered. As in the earlier example of the relationshipbetween overall landscape productivity, increased pri-mary prey of generalist predators, and their effects onbreeding productivity of alternative prey, altered abun-dance relations could also affect the outcome of com-petitive or mutualistic interactions. Evidence of suchindirect effects is limited, but several studies suggestthat colonization of new habitat patches can be depen-dent on the presence of other individuals (Reed andDobson 1993). Among forest birds, migrants selectingbreeding sites often prefer settling close to conspecific(Svardson 1949) or heterospecific individuals; resi-dents in particular (Monkkonen et al. 1999). Time con-straints associated with a short breeding season andlarge year-to-year variation in environmental condi-tions, both characteristics of northern environments,presumably render these habitat selection tactics prof-itable (Monkkonen et al. 1997). Consequently, absenceof resident birds from remnant boreal forests could re-duce the rate of colonization of these patches by other

380 INVITED FEATURE Ecological ApplicationsVol. 12, No. 2
species, such as migrants, resulting in an increased ex-tinction probability in degraded landscapes.
Area effects
Although the magnitude of biological and physicaledge effects on bird populations is related to the areaof remnant habitats, additional area-related phenome-na, such as the effects of decreased patch size on theoccurrence of species with large home ranges, or den-sity-related patch size effects, may also be important.In general, migrant species have a smaller average bodysize than resident species (Monkkonen 1992). Bodysize is also correlated with other life history attributes,such as home range size (Southwood 1976, Peters1983). Because larger species will generally requirelarger home ranges, resident species should exhibit thegreatest area effects. Furthermore, because residentspecies may adjust their winter territory sizes to reflectfood availability (e.g., Enoksson and Nilsson 1983),and some residents form single- or mixed-speciesflocks in winter, resident boreal birds may also requirelarger forested areas during nonbreeding periods (seeSchmiegelow 1997). Both habitat loss and fragmen-tation could contribute to population declines as a resultof area effects.
Area effects could be exacerbated by fragmentationat an individual level if patches were smaller than thearea requirement of an individual, or at the populationscale if persistence of local populations is related topatch size and isolation (Andren 1994). However, thisassumes a binary landscape of suitable and unsuitablehabitat, and that individuals use only one habitat patch.In boreal forests of Fennoscandia, Capercaillie (Tetraourogallus), Northern Goshawk (Accipiter gentilis), andBlack Woodpecker (Dryocopus martius) can compen-sate for reductions in preferred habitat by includingmultiple patches of older forest within larger territories(Wegge and Rolstad 1986, Rolstad 1989, Widen 1989,Tjernberg et al. 1993). Similarly, Bonar (2001) dem-onstrated that the Pileated Woodpecker (Dryocopus pi-leatus), an ecological equivalent to the Black Wood-pecker, also includes multiple patches of preferred hab-itat within territories in managed boreal forests in west-ern Canada.
In Andren’s (1994) compilation of studies, no patchsize (area-related changes in density) or isolation ef-fects were found in studies from boreal forest land-scapes (see also Monkkonen and Reunanen 1999). Ina more recent study from northern Sweden, Edeniusand Sjoberg (1997) report that small patches of forest(,5 ha) supported fewer species and large patches sup-ported more than expected by sample size. However,at the species level, no patch size effects were evident.Bender et al. (1998) analyzed data for a variety ofspecies and systems and found no evidence that patchsize effects were related to landscape characteristics,including amount of habitat, median patch size, or scale
of investigation. However, they report greater meanpatch size effects for resident than for migrant species.The only boreal sites included in their analyses werefrom Finland and Sweden, and no such patterns wereapparent among migrant and resident birds from theseareas (data extracted from Bender et al. 1998: AppendixC). Moreover, although there was considerable vari-ability within each group, the mean patch size effectwas negative for both, i.e., density was higher in small-er patches. Similarly, Connor et al. (2000) concludedfrom their meta-analysis of patch size effects for a va-riety of taxa and systems that birds generally exhibitedmoderately large to large, positive effects, and that re-lationships were stronger in temperate and boreal en-vironments than in tropical environments. However, themean relationship for boreal birds was, again, negative(Connor et al. 2000: Table 2).
Nevertheless, the patch size effects reported here forboreal sites should be interpreted cautiously, as we didnot undertake a formal reanalysis of the data reportedin either Bender et al. (1998) or Connor et al. (2000),and mean effect sizes within taxa mask individual spe-cies variability. Further, the sample size from borealsystems was low, included only Fennoscandian sites,did not distinguish between naturally isolated (islandand wetland) habitat patches and those isolated due tohuman activities, and failed to consider landscape con-text.
Isolation effects
Although most fragmentation-related theory empha-sizes the role of habitat isolation and dispersal capa-bilities on population persistence in remnant habitats(e.g., MacArthur and Wilson 1967, Levins 1969), em-pirical tests of the importance of these attributes in reallandscapes are few. Debate continues to rage over theefficacy of corridors as a conservation tool in allevi-ating the effects of habitat fragmentation (for a review,see Simberloff et al. 1992). However, corridor presenceis often correlated with other habitat or landscape var-iables (e.g., patch size and landscape context), therebyconfounding assessments (Beier and Noss 1998).
In an experimental study of recently harvested borealforest in western Canada, Machtans et al. (1996) re-ported elevated use of riparian forest corridors by ju-venile birds immediately following harvest of the ad-jacent forest stands. However, a longer term study ofone of these corridors showed that use diminished rap-idly over time, as adjacent harvested areas regenerated(Robichaud et al., in press). Several studies in borealforests of eastern Canada have demonstrated that birdmovements across forest gaps are significantly reducedrelative to movement rates through forest, both in sum-mer (Desrochers and Hannon 1997) and winter (St.Clair et al. 1998). Nevertheless, in a recent analysis ofthe persistence of bird species in remnant patches ofolder forest, Hannon and Schmiegelow (in press) found

April 2002 381HABITAT FRAGMENTATION: A REASSESSMENT
that habitat affinity was a better predictor of abundanceresponse to fragmentation by forest harvesting thangap-crossing behavior (a surrogate of dispersal ability).Old-forest specialists were most sensitive to fragmen-tation resulting from habitat loss, and corridors didlittle to alleviate these effects. Similarly, Andren(1994) found no evidence of isolation effects in studiesfrom boreal forest landscapes.
Monkkonen et al. (2000) found that species numberand abundance of resident old-forest species in north-ern Finnish forest reserves declined with distance fromcontinuous forests in Russian Karelia. This may indi-cate that isolation from the extensive source areas inthe east has a negative impact on resident forest birds.However, the contribution of the landscape structurearound the reserves was not properly accounted for:the observed patterns may also result from habitat loss,as there is a gradient from east to west in northernFinland along which the proportion of old forest in thelandscape decreases.
HABITAT LOSS IN DYNAMIC LANDSCAPES
In boreal landscapes where forest harvesting is thedominant land use, population trajectories of birds ap-pear to be more strongly related to habitat loss anddegradation, and their affects on species abundancesand interactions, than to fragmentation sensu strictu.Here, we aim to derive null expectations about the rel-ative effects of habitat loss and fragmentation, basedlargely on the structure of the underlying community.In the absence of fragmentation effects, our hypothesisis that species loss, due to overall habitat loss, shouldbe predicted by the regional species abundance distri-bution (after Preston 1948). Reduction of particularhabitat types, in conjunction with habitat specializa-tion, results in the loss of rarer species by chance alone.If additional factors compound habitat loss, then spe-cies loss should be more rapid than predicted. Thismodel is consistent with the random-sample hypothesis(Connor and McCoy 1979, Haila 1983). We use thisapproach to establish benchmark communities andidentify common features of boreal bird species sen-sitive to loss of a specific habitat type.
In managed forest landscapes, quantification of hab-itat loss and degradation requires more explicit con-sideration of habitats of concern. In many cases, totalforest cover may remain constant, while the age struc-ture and relative proportions of different habitat typeschange dramatically, resulting in habitat degradationfor some species. In boreal forests, two habitat typesare becoming increasingly scarce: old forest, particu-larly in large tracts (e.g., Niemela 1999), and early-successional, post-burn stands (e.g., Murphy and Lehn-hausen 1998, Kouki et al. 2001). Here, we focus onthe potential implications of loss of older, pristine for-est (.80 yr old) for boreal birds. We use 80 yr as ourthreshold, as most commercial forest types are mer-
chantable at this age, and are harvested, unless pro-visions for protection exist.
METHODS
We generated bird species abundance models for old-er forests using long-term census data from referenceareas in Oulanka National Park, Finland (1977–1997)and Alberta, Canada (1993–1999). Oulanka NationalPark (267 km2) is located in northern Finland at theFinnish–Russian border (668 N, 298 E). Oulanka is aforested peninsula in a harvested landscape, stretchingwest from the continuous forests in Russia (Russiangreen belt). It is one of the few remaining referenceareas of pristine forest bird assemblages in Finland,due to extensive harvesting elsewhere during the past50 yr. The dominant tree species are Scotch pine (Pinussylvestris) and Norway spruce (Picea abies), with birch(Betula spp.), aspen (Populus tremula), and goat wil-low (Salix caprea) present in mixed stands. The Ca-nadian site is located in northeastern Alberta, in borealmixed-wood forest near Calling Lake (558 N, 1138 W).Boreal forests in western Canada have only recentlybeen allocated for timber harvest, and thus exist in arelatively pristine state. The Calling Lake referencearea remains contiguous with adjacent tracts of olderforest; a portion acts as a control for an experimentalfragmentation study (Schmiegelow and Hannon 1993,Schmiegelow et al. 1997). This forest is dominated bytrembling aspen (Populus tremuloides), white spruce(Picea glauca), and balsam poplar (Populus balsami-fera), with lesser amounts of white birch (Betula pa-pyrifera) and balsam fir (Abies balsamea).
Finnish bird data were derived from line-transect sur-veys, whereas point counts were used in the Canadiansurveys. We included all forest passerine, woodpecker,and grouse species known to breed in these areas inour community descriptions. We used aggregated datafrom all years to represent the regional species pooland to construct cumulative species abundance distri-butions. We used detection frequencies of species indifferent habitat types (from additional, unpublisheddata sets) to identify species clearly favoring forests.80 yr old (after Raivio and Haila 1990). Where pos-sible, we validated our classifications against publishedaccounts (Helle 1985, Raivio and Haila 1990, Schiecket al. 1995b, Jokimaki and Huhta 1996, Kirk et al.1996). Species identified as mature-forest specialistswere found primarily in older forest, and were detectedat very low frequencies in silvicultural mosaics, where-as old-forest specialists were never observed in themosaics. These are consistent with two of the fivegroups identified by Raivio and Haila (1990), but arecombined in all analyses that we report.
We created rank abundance plots (e.g., May 1975)from repeated, random subsamples (n 5 10 and n 5100) to assess the distribution of mature- and old-forestspecialists in the resultant communities. Species-spe-

382 INVITED FEATURE Ecological ApplicationsVol. 12, No. 2
PLATE 1. Resident species, such as the Boreal Chickadee and the Three-toed Woodpecker, are threatened by the loss of olderboreal forests, due to habitat specialization and rarity. Photographs by Chris Fisher, Curtis Stambaugh, and Fiona Schmiegelow.
TABLE 1. Bird community attributes of older forests (.80 yr old) in reference areas in Finland(1977–1997, Oulanka National Park) and Canada (1993–1999, near Calling Lake, Alberta).
Attribute Finland Canada
Number of speciesMean annual breeding density (no. pairs/km2)Number (and %) of old-forest specialist speciesDensity of old-forest specialists (no. pairs/km2)Number (and %) of resident speciesNumber (and %) resident of old-forest specialist species
48100–120
27 (56)30–4017 (35)14 (52)
61400–550
19 (31)120–170
14 (23)8 (42)
cific probabilities of absence were then compared be-tween old-forest specialist and generalist species, andbetween resident and migrant species, using Mann-Whitney U tests. Finally, we used rarefaction (e.g.,Hurlbert 1971, James and Rathbun 1981) to estimatethe expected number of species (all and mature–oldforest specialists only) in samples of varying size (seePlate 1). The resultant species accumulation curves rep-resent null models to which species lists from remnanthabitats can be compared.
RESULTS
Bird communities of older boreal forest
The bird species abundance models for old forestsin Finland and Canada fit a lognormal distribution (P5 0.200 for both areas). There was no evidence oftruncation of the curve for either system, indicatingthat our data provide good representations of the truedistributions. Bird species richness and mean annualabundance were higher in the Canadian reference area
than in Finland (Table 1), and the percentage of speciesidentified as old-forest specialists differed markedlybetween the systems
Rank abundance plots (Fig. 1) illustrate distributionsof the old-forest specialists within the larger bird com-munities. A notable pattern in the Finnish data is theextreme dominance of two species: the Brambling,Fringilla montifringilla, and Willow Warbler, Phyllos-copus trochilus (for a discussion, see Greenberg et al.1999). Most of the Finnish old-forest specialists aremoderately rare to rare, whereas the Canadian spe-cialists are relatively evenly distributed among abun-dance ranks. However, despite the very different rich-ness and distribution of this group in the two systems,their density relative to the larger community was re-markably consistent (Finland 31%, Canada 30%). Res-ident species in Finland, although a richer componentof the community than in Canada, tend to be rarer.Nevertheless, in both systems, resident species areoverrepresented among old-forest specialists, relativeto their richness in the larger community (Table 1).
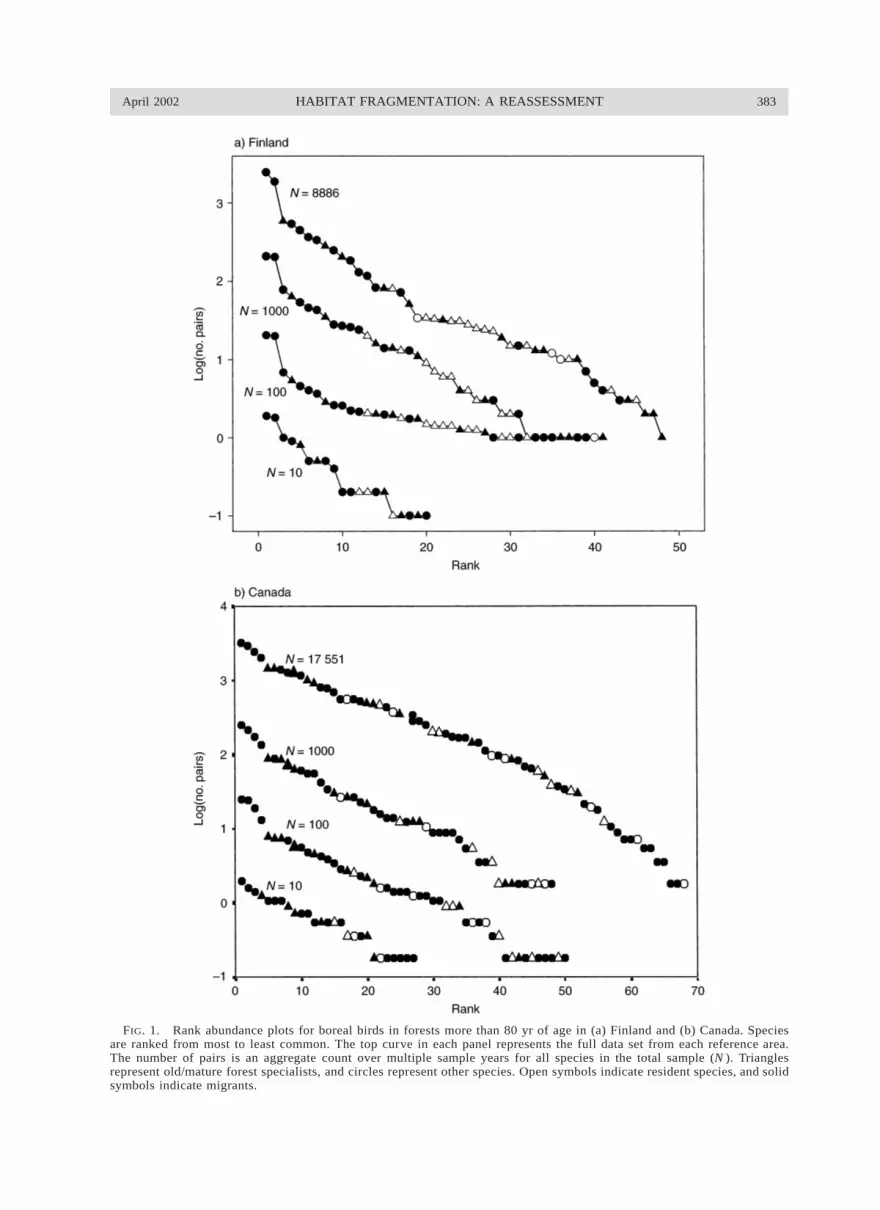
April 2002 383HABITAT FRAGMENTATION: A REASSESSMENT
FIG. 1. Rank abundance plots for boreal birds in forests more than 80 yr of age in (a) Finland and (b) Canada. Speciesare ranked from most to least common. The top curve in each panel represents the full data set from each reference area.The number of pairs is an aggregate count over multiple sample years for all species in the total sample (N ). Trianglesrepresent old/mature forest specialists, and circles represent other species. Open symbols indicate resident species, and solidsymbols indicate migrants.

384 INVITED FEATURE Ecological ApplicationsVol. 12, No. 2
FIG. 2. Rarefaction estimates of the expected number ofold-forest specialists in samples of forest birds from Finlandand Canada. Numbers on the x-axis correspond to the totalsample of individuals (specialist and nonspecialist species).The two horizontal lines indicate the 90th percentile for spe-cialist species in Finland and Canada.
Species loss from random samples
We expected subsamples from the larger distributionto exhibit veil lines (e.g., Preston 1948), inferred fromthe shape and truncation of the lower curves in eachrank plot (Fig. 1). Moderately rare to rare species arebetter represented with increasing sample size. Old-forest specialists were missing more often than non-specialists in sample sizes of 10 and 100 from Finland(P 5 0.019 and 0.009, respectively), but not fromequivalent-sized samples from Canada (P 5 0.54 and0.71, respectively). Resident species in both boreal ar-eas generally had a higher probability than migrants ofbeing absent in random samples of 10 and 100 indi-viduals drawn from a pool representing the larger com-munity (P 5 0.017 and 0.021, Finland; P 5 0.045 and0.014, Canada).
Asymptotic values for inclusion of old-forest spe-cialists are reached at different sample sizes in Finlandand Canada (Fig. 2) because of differing abundancedistributions and ranks of old-forest species. The 90thpercentiles for old-forest specialists occur at total sam-ple sizes (i.e., samples including all species in the re-gional pool) of ;2000 pairs in Finland and 1100 pairsin Canada. These represent null models for species loss,in the absence of significant fragmentation effects. Be-low these asymptotic values, species loss in conjunc-tion with habitat loss is predicted to occur due to sam-pling effects alone.
DISCUSSION
In forested landscapes where timber harvesting isthe dominant land use practice, forest cover cannot beconsidered equivalent to habitat amount for specialistspecies (i.e., a binary classification of landscapes is notappropriate). Predicting the outcome of habitat deg-
radation requires identification of habitat types and spe-cies of concern. Using long-term bird census data fromboreal forests in Finland and Canada, we demonstratedhow simple species abundance models can be used toidentify vulnerable species and test hypotheses abouthabitat loss and fragmentation.
Predicting species loss from older boreal forests
Old-forest specialists account for almost one-thirdof all birds breeding in older boreal forests in bothFinland and Canada. Although the richness and rankedabundances of members of this group varied betweenregions, we were able to identify some common fea-tures of sensitive species. Among the boreal bird spe-cies that specialize on older forests, residents shouldexhibit the greatest sensitivity to habitat loss, due totheir lower abundance. This rarity falls in one of the12 categories in the three-way classification scheme ofRabinowitz et al. (1986): boreal residents generallyhave a broad geographic distribution (several are cir-cumpolar), demonstrate habitat specificity, and are sup-ported as relatively small populations wherever theyare found. Populations of resident species in the north-ern boreal forest are regulated by winter conditions(e.g., Lahti et al. 1998). Although this may also be truefor residents in other systems (e.g., Newton 1994), lowfood availability and severe winter weather result inlower population sizes in the northern boreal zone thanelsewhere (Forsman 2000). For example, in the UnitedStates and Britain, resident species are typically morethan twice as abundant as migrants (O’Connor 1981,1992). Residents in other systems also have broadertaxonomic representation and evolutionary historiesthan do boreal residents, many of which have behav-ioral, morphological, and physiological adaptations forwinter conditions (e.g., food-caching, specializedplumage, and regulated hypothermia). Additionally,many boreal residents are reliant on older forests be-cause of nesting (availability of suitable trees for cav-ities) and foraging (dead or decaying wood) require-ments (Imbeau et al. 2001).
The migrant old-forest specialists in both Finland andCanada are not so easily compartmentalized. Some spe-cies are relatively abundant, whereas others exhibit char-acteristics of several of the rarity categories of Rabi-nowitz et al. (1986). As a group, they are not consistentin either their nesting or foraging requirements on thebreeding grounds (e.g., Schieck et al. 1995b, Kirk et al.1996, Vaisanen et al. 1998). However, many of the mi-grant species specializing on older, boreal forests havesimilar breeding and wintering habitats (i.e., they dependon forested habitats year-round), which may make themparticularly vulnerable to habitat loss and fragmentation,due to a lack of behavioral plasticity.
A null model for assessing species loss
In both Finnish and Canadian boreal forests, ouranalyses indicate that species gain over small areas will

April 2002 385HABITAT FRAGMENTATION: A REASSESSMENT
be rapid because of the relatively even species distri-butions represented in small samples. This is a generalcharacteristic of communities characterized by log-se-ries or lognormal distributions: the slope of the spe-cies–area relationship will be steepest when calculatedover small areas. Very steep slopes for species–areacurves have been reported from boreal forests in Swe-den (Bostrom and Nilsson 1983) and Canada (Schmie-gelow et al. 1997). However, apparent increases in di-versity with area were spurious, resulting simply fromlarger sample sizes (Rosenzweig 1995, Schmiegelowet al. 1997). Such results highlight the danger of draw-ing erroneous conclusions about fragmentation. Al-though steep species–area slopes have been associatedwith insular systems (e.g., MacArthur and Wilson1967), the shape of the species–area curve is a functionof the underlying species abundance distribution, andthus is sensitive to sample size. This suggests a scaledependency that may also apply to density-relatedpatch-size effects. In either case, patterns from habitatremnants must be compared to equivalent-sized areasfrom contiguous reference areas before any conclusionsabout fragmentation can be drawn.
We generated bird species accumulation curves forolder boreal forests using rarefaction, in order to con-struct null models to which species lists from remnanthabitats can be compared. Similar to approaches aimedat identifying thresholds in the population persistenceof individual species (e.g., With and Crist 1995, Fahrig1998), identifying the inflection point in species ac-cumulation curves for communities of concern couldprovide useful conservation guidelines. For example,given known densities of breeding birds in referenceareas, one could extrapolate from sample sizes requiredfor representation of old-forest species to estimate thehabitat threshold below which species loss is predictedto occur due to sampling effects alone. Such thresholdestimates do not represent the minimum area requiredfor regional population persistence; reserve sizes nec-essary to support self-sustaining populations would un-doubtedly be several magnitudes larger (e.g., Preston1962). However, they do provide a benchmark againstwhich to assess the effects of habitat fragmentation onspecies occurrence. This approach provides for moreefficient allocation of research efforts and managementactions through identification of the conditions underwhich the spatial configuration of habitats really mat-ters (sensu Fahrig 1998). Where potential fragmenta-tion effects are identified, mechanistic studies may berequired to assess the cause of population declines.Otherwise, conservation and management interestswould be better served by greater intellectual invest-ment in determining the area requirements of com-munities of concern.
CONCLUSIONS
In the research–management interface, ecologists areoften challenged to provide guidelines in the absence of
complete understanding. We demonstrated that, for bo-real birds, application of conservation paradigms fromother systems do not provide a very useful frameworkfor projecting the outcome of habitat loss and fragmen-tation in forested boreal landscapes managed primarilyfor timber production. Natural dynamics, land use pat-terns, and management issues differ between systems,and so too will responses (see also Haila 2002). Further,there is likely to be both inter- and intrataxal variationin response within systems, due to differing life historycharacteristics (e.g., Schmiegelow and Nudds 1987,Hansen and Urban 1992, Monkkonen et al. 1992, Monk-konen 1994, Monkkonen and Welsh 1994).
In dynamic landscapes, such as those managed forforestry, habitat loss is specific to habitat types andfragmentation effects may not be readily apparent be-cause of the transient nature of the matrix. Forest landsare not permanently converted for other land uses; rath-er, they undergo continuous state changes from harvestthrough succession. The vulnerable components of theforest are those with high initial conversion value, suchas pristine older forests, or those with diminishing com-mercial return, or lost opportunity costs; again, theseare older forests and habitats created by large, naturaldisturbances, such as postfire, early-successional for-ests. In these cases, a random-sample hypothesis (e.g.,Connor and McCoy 1979, Haila 1983) may adequatelypredict the outcome of selective habitat loss, but iden-tification of the elements of concern is crucial. Sampleeffects alone may be sufficient to cause regional pop-ulation declines and a loss of sensitive species, high-lighting the importance of quantifying the effects ofhabitat loss (e.g., Fahrig 1997).
We focused on boreal bird communities in Finlandand Canada and constructed simple empirical modelsfor projecting patterns of species loss with reductionsin the area of older forest. Our predictions were gen-erally consistent with observed patterns of populationdecline and species loss from these areas. These nullmodels provide a basis for comparison with habitatremnants, and for more detailed exploration of popu-lation dynamics and species persistence in these sys-tems. Nevertheless, we do not advocate widespread ap-plication of the particular null models presented here:that would contradict the very premise of our argu-ments. Whereas Harrison and Bruna (1999) highlightthe mismatch between ecological theory and empiricalstudies of fragmentation, we lament the indiscriminategeneralization of empirical observations. However, wedo encourage a revival of one of the fundamental con-cepts of insular ecology and foundations of samplingdesign, namely, a firm understanding of the underlyingstructure of the community of interest. In some cases,exploration of species abundance distributions mayprovide the most parsimonious explanation for the ef-fects of habitat loss and a useful conduit for commu-nicating potential ecological responses to land man-

386 INVITED FEATURE Ecological ApplicationsVol. 12, No. 2
agers. Detailing the composition and structure ofbenchmark communities in reference sites is thereforecritical to facilitate the rapid assessment of assem-blages from comparative sites (e.g., Colwell and Cod-dington 1994). In this manner, the relative influence ofhabitat loss and fragmentation on species occurrencemay be discriminated and research and managementpriorities may be established.
ACKNOWLEDGMENTS
We thank the many researchers whose work made this re-view possible, and the field assistants who collected the Finn-ish and Canadian bird census data presented here. S. Cum-ming provided analytical advice. This manuscript was sub-stantially improved by the thoughtful comments of M.-A.Villard and two anonymous reviewers. This research was sup-ported by the Academy of Finland (project #45626) and theCanadian Sustainable Forest Management Network.
LITERATURE CITED
Andren, H. 1992. Corvid density and nest predation in re-lation to forest fragmentation: a landscape perspective.Ecology 73:794–804.
Andren, H. 1994. Effects of habitat fragmentation on birdsand mammals in landscapes with different proportions ofsuitable habitat: a review. Oikos 71:355–366.
Andren, H. 1995. Effects of landscape composition on pre-dation rates at habitat edges. Pages 225–255 in L. Hansson,L. Fahrig, and G. Merriam, editors. Mosaic landscapes andecological processes. Chapman and Hall, London, UK.
Andren, H., P. Angelstam, and P. Widen. 1985. Differencesin predation pressure in relation to habitat fragmentation:an experiment. Oikos 45:273–277.
Angelstam, P. 1986. Predation on ground-nesting birds’ nestsin relation to predator densities and habitat edge. Oikos 47:365–373.
Angelstam, P. 1992. Conservation of communities—the im-portance of edges, surroundings, and landscape mosaicstructure. Pages 9–70 in L. Hansson, editor. Nature con-servation by ecological principles. Elsevier Applied Sci-ence, London, UK.
Angelstam, P., E. Lindstrom, and P. Widen. 1984. Role ofpredation in short-term population fluctuations some birdsand mammals in Fennoscandia. Oecologia 62:199–208.
Angelstam, P., E. Lindstrom, and P. Widen. 1985. Synchro-nous short-term population fluctuations of some birds andmammals in Fennoscandia—occurrence and distribution.Holarctic Ecology 8:285–298.
Bayne, E. M., and K. A. Hobson. 1997. Comparing the effectsof landscape fragmentation by forestry and agriculture onpredation of artificial nests. Conservation Biology 11:1418–1429.
Bayne, E. M., K. A. Hobson, and P. Fargey. 1997. Predationon artificial nests in relation to forest type: contrasting theuse of quail and plasticine eggs. Ecography 20:233–239.
Beier, P., and R. Noss. 1998. Do habitat corridors provideconnectivity? Conservation Biology 12:1241–1252.
Bender, D. J., T. A. Contreras, and L. Fahrig. 1998. Habitatloss and population decline: a meta-analysis of the patchsize effect. Ecology 79:517–533.
Bohning-Gaese, K., M. L. Taper, and J. H. Brown. 1993. Aredeclines in North American insectivorous songbirds due tocauses on the breeding range? Conservation Biology 7:76–86.
Bonar, R. 2001. Pileated Woodpecker, Dryocopus pileatus,habitat ecology in the Alberta foothills. Dissertation. Uni-versity of Alberta, Edmonton, Alberta, Canada.
Bostrom, U., and S. G. Nilsson. 1983. Latitudinal gradients
and local variations in species richness and structure ofbird communities on raised peat-bogs in Sweden. OrnisScandinavica 14:213–226.
Brooke, M. L. de, and N. B. Davies. 1987. Recent changesin host usage by cuckoos, Cuculus canorus, in Britain. Jour-nal of Animal Ecology 56:873–883.
Bryant, D., D. Nielsen, and L. Tangley. 1997. The last fron-tier forests: ecosystems and economies on the edge. WorldResources Institute, Washington, D.C., USA.
Cadman, M. D., P. D. F. Eagles, and F. M. Helleiner. 1987.Atlas of the breeding birds of Ontario. Federation of On-tario Naturalists and University of Waterloo Press, Water-loo, Ontario, Canada.
Chen, J., J. F. Franklin, and T. A. Spies. 1995. Vegetationresponses to edge environments in old-growth Douglas-firforests. Ecological Applications 2:387–396.
Colwell, R. K., and J. A. Coddington. 1994. Estimating ter-restrial biodiversity through extrapolation. PhilosophicalTransactions of the Royal Society of London, Series B,Biological Sciences 345:101–118.
Connor, E. F., A. C. Courtney, and J. M. Yoder. 2000. In-dividuals–area relationships: the relationship between an-imal population density and area. Ecology 81:734–748.
Connor, E. F., and E. D. McCoy. 1979. The statistics andbiology of the species–area relationship. American Natu-ralist 113:791–833.
Cotterill, S. E., and S. J. Hannon. 1999. No evidence of short-term effects of clear-cutting on artificial nest predation inboreal mixedwood forests. Canadian Journal of Forest Re-search 29:1900–1910.
Davies, N. B. 2000. Cuckoos, cowbirds and other cheats. T.and A. D. Poyser, London, UK.
Desrochers, A., and S. J. Hannon. 1997. Gap crossing de-cisions by forest songbirds during the post-fledging period.Conservation Biology 11:1204–1210.
Edenius, L., and K. Sjoberg. 1997. Distribution of birds innatural landscape mosaics of old-growth forests in northernSweden: relations to habitat area and landscape context.Ecography 20:425–431.
Enoksson, B., and S. G. Nilsson. 1983. Territory size andpopulation density in relation to food supply in the nuthatchSitta europaea (Aves). Journal of Animal Ecology 52:929–935.
Erskine, A. J. 1977. Birds in boreal Canada: communities,densities, and adaptations. Minister of Supply and Services,Ottawa, Canada. Canadian Wildlife Service Report Number41.
Esseen, P.-A. 1994. Tree mortality patterns after experimentalfragmentation of an old-growth conifer forest. BiologicalConservation 68:19–28.
Fahrig, L. 1997. Relative effects of habitat and fragmentationon population extinction. Journal of Wildlife Management61:603–610.
Fahrig, L. 1998. When does fragmentation of breeding hab-itat affect population survival? Ecological Modelling 105:273–292.
Fahrig, L. 2002. Effect of habitat fragmentation on the ex-tinction threshold: a synthesis. Ecological Applications 12:346–353.
Forsman, J. T. 2000. Heterospecific attraction in breedingbird communities. Implications to habitat selection and spe-cies interactions in a landscape perspective. Acta Univer-sitatis Ouluensis A 345, Oulu, Finland.
Gates, J. E., and L. W. Gysel. 1978. Avian nest dispersionand fledging success in field–forest ecotones. Ecology 59:871–883.
Greenberg, R., V. Pravosudov, J. Sterling, A. Kozlenko, andV. Kontorshchikov. 1999. Tits, warblers and finches: fo-liage-gleaning birds of Nearctic and Palearctic boreal for-ests. Condor 101:299–310.

April 2002 387HABITAT FRAGMENTATION: A REASSESSMENT
Haartman, L. von. 1981. Co-evolution of the Cuckoo Cuculuscanorus and a regular Cuckoo host. Ornis Fennica 58:1–10.
Hahn, D. C., and J. S. Hatfield. 1995. Parasitism at the land-scape scale: cowbirds prefer forests. Conservation Biology9:1415–1424.
Haila, Y. 1983. Land birds on northern islands: a samplingmetaphor for insular colonisation. Oikos 41:334–351.
Haila, Y. 1999. Islands and fragments. Pages 234–264 in M.L. Hunter, editor. Maintaining biodiversity in forest eco-systems. Cambridge University Press, Cambridge, UK.
Haila, Y. 2002. A conceptual genealogy of fragmentationresearch: from island biogeography to landscape ecology.Ecological Applications 12:321–334.
Haila, Y., and O. Jarvinen. 1990. Northern conifer forestsand their bird species assemblages. Pages 61–85 in A.Keast, editor. Biogeography and ecology of forest bird com-munities. SPB Academic Publishing, The Hague, The Neth-erlands.
Halme, E., and J. Niemela. 1993. Carabid beetles in frag-ments of coniferous forest. Annales Zoologici Fennici 30:17–30.
Hannon, S. J., and S. E. Cotterill. 1998. Nest predation inaspen woodlots in an agricultural area in Alberta: the enemyfrom within. Auk 115:16–25.
Hannon, S. J., and F. K. A. Schmiegelow. In press. Corridorsmay not improve the conservation value of small reservesfor most boreal birds. Ecological Applications.
Hansen, A. J., and D. L. Urban. 1992. Avian response tolandscape patterns: the role of species’ life histories. Land-scape Ecology 7:163–180.
Hanski, I. K., T. J. Fenske, and G. J. Niemi. 1996. Lack ofedge effect in nesting success of breeding birds in managedforest landscapes. Auk 113:578–585.
Harper, K. A. 1999. Variability in forest structure and com-position along edge-to-interior gradients from lakeshoreand clearcut edges and in riparian buffers in the aspen-dominated mixedwood boreal forest. Dissertation. Univer-sity of Alberta, Edmonton, Alberta, Canada.
Harrison, S., and E. Bruna. 1999. Habitat fragmentation andlarge-scale conservation: what do we know for sure? Ecog-raphy 22:225–232.
Hartley, M. J., and M. L. Hunter. 1998. A meta-analysis offorest cover, edge effects, and artificial nest predation rates.Conservation Biology 12:465–469.
Helle, P. 1983. Bird communities in open ground–climaxforest edges in northeastern Finland. Oulanka Reports 3:39–46.
Helle, P. 1985. Habitat selection of breeding birds in relationto forest succession in Northeastern Finland. Ornis Fennica62:113–123.
Helle, P., and O. Jarvinen. 1986. Population trends of NorthFinnish land birds in relation to their habitat selection andchanges in forest structure. Oikos 46:107–115.
Helle, P., and J. Muona. 1985. Invertebrate numbers in edgesbetween clear-fellings and mature forests in northern Fin-land. Silva Fennica 19:281–294.
Hobson, K. A., and M.-A. Villard. 1998. Forest fragmenta-tion affects the behavioral response of American Redstartsto the threat of cowbird parasitism. Condor 100:389–394.
Huhta, E. 1996. Effects of spatial scale and vegetation coveron predation of artificial ground nests. Wildlife Biology 1:73–80.
Huhta, E., J. Jokimaki, and P. Helle. 1998. Predation on ar-tificial nests in a forest dominated landscape—the effectsof nest type, patch size and edge structure. Ecography 21:464–471.
Huhta, E., T. Mappes, and J. Jokimaki. 1996. Predation onartificial ground nests in relation to forest fragmentation,
agricultural land and habitat structure. Ecography 19:85–91.
Hurlbert, S. H. 1971. The nonconcept of species diversity:a critique and alternative parameters. Ecology 52:577–586.
Imbeau, L., M. Monkkonen, and A. Desrochers. 2001. Long-term impacts of forestry on birds of the eastern Canadianboreal spruce forests: what can we learn from the Fennos-candian experience? Conservation Biology 15:1151–1162.
James, F. C., and S. Rathbun. 1981. Rarefaction, relativeabundance, and diversity of avian communities. Auk 98:785–800.
Jarvinen, O., K. Kuusela, and R. A. Vaisanen. 1977. Effectsof modern forestry on the number of breeding birds inFinland in 1945–1975. Silva Fennica 11:284–294.
Jokimaki, J., and E. Huhta. 1996. Effects of landscape matrixand habitat structure on a bird community in northern Fin-land: a multi-scale approach. Ornis Fennica 73:97–113.
Jokimaki, J., E. Huhta, J. Itamies, and P. Rahko. 1998. Dis-tribution of athropods in relation to forest patch size, edge,and stand characteristics. Canadian Journal of Forest Re-search 28:1068–1072.
Kirk, D. A., A. W. Diamond, K. A. Hobson, and A. R. Smith.1996. Breeding bird communities of the western and north-ern Canadian boreal forest: relationship to forest type. Ca-nadian Journal of Zoology 74:1749–1770.
Kouki, J., S. Lofman, P. Martikainen, S. Rouvinen, and A.Uotila. 2001. Forest fragmentation in Fennoscandia: link-ing habitat requirements of wood-associated threatenedspecies to landscape and habitat changes. ScandinavianJournal of Forest Research, Supplement 3:27–37.
Kremsater, L., and F. L. Bunnell. 1999. Edge effects: theory,evidence and implications to management of western NorthAmerican forests. Pages 117–153 in J. A. Rochelle, L. A.Lehmann, and J. Wisniewski, editors. Forest fragmentation:wildlife and management implications. Brill, Leiden, TheNetherlands.
Kuitunen, M., and P. Helle. 1988. Relationship of the com-mon treecreeper Certhia familiaris to edge and forest frag-mentation. Ornis Fennica 65:150–155.
Kurki, S., P. Helle, H. Linden, and A. Nikula. 1997. Breedingsuccess of black grouse and capercaillie in relation to mam-malian predator density on two spatial scales. Oikos 79:301–310.
Kurki, S., A. Nikula, P. Helle, and H. Linden. 1998. Abun-dances of red fox and pine marten in relation to compositionof boreal forest landscapes. Journal of Animal Ecology 67:874–886.
Kurki, S., A. Nikula, P. Helle, and H. Linden. 2000. Land-scape fragmentation and forest composition effects ongrouse breeding success in boreal forests. Ecology 81:1985–1997.
Lahti, K., M. Orell, S. Rytkonen, and K. Koivula. 1998.Time- and food-dependence in Willow Tit winter survival.Ecology 79:2904–2916.
Levins, R. 1969. Some demographic and genetic consequenc-es of environmental heterogeneity for biological control.Bulletin of the Entomological Society of America 15:237–240.
Linden, H., and P. Rajala. 1981. Fluctuations and long-termtrends in the relative densities of tetraonid populations inFinland 1964–1977. Finnish Game Research 39:13–34.
MacArthur, R. H., and E. O. Wilson. 1967. The theory ofisland biogeography. Princeton University Press, Princeton,New Jersey, USA.
Machtans, C. S., M.-A. Villard, and S. J. Hannon. 1996. Useof riparian buffer strips as movement corridors by forestbirds. Conservation Biology 10:1366–1379.
Martin, T. E. 1993. Nest predation among vegetation layersand habitat types: revising the dogmas. American Naturalist141:897–913.

388 INVITED FEATURE Ecological ApplicationsVol. 12, No. 2
Matlack, G. R. 1993. Microenvironment variation within andamong forest edge sites in the eastern United States. Bi-ological Conservation 66:185–194.
Matlack, G. R. 1994. Vegetation dynamics of the forestedge—trends in space and successional time. Journal ofEcology 82:113–123.
May, R. M. 1975. Patterns of species abundance and diver-sity. Pages 81–120 in M. L. Cody and J. M. Diamond,editors. Ecology and evolution of communities. HarvardUniversity Press, Cambridge, Massachusetts, USA.
McGarigal, K., and S. Cushman. 2002. Comparative evalu-ation of experimental approaches to the study of habitatfragmentation effects. Ecological Applications 12:335–345.
Monkkonen, M. 1992. Life history traits of Palearctic andNearctic migrant passerines. Ornis Fennica 69:161–172.
Monkkonen, M. 1994. Diversity patterns in Palaearctic andNearctic forest bird assemblages. Journal of Biogeography21:183–195.
Monkkonen, M., R. Hardling, J. T. Forsman, and J. Tuomi.1999. Evolution of heterospecific attraction: using otherspecies as cues in habitat selection. Evolutionary Ecology13:91–104.
Monkkonen, M., P. Helle, G. J. Niemi, and K. Montgomery.1997. Heterospecific attraction affects community struc-ture and migrant abundances in northern breeding bird com-munities. Canadian Journal of Zoology 75:2077–2083.
Monkkonen, M., P. Helle, and D. A. Welsh. 1992. Perspec-tives on Palaearctic and Nearctic bird migration: compar-isons and overview of life-history and ecology of migrantpasserines. Ibis 134 Supplement 1:7–13.
Monkkonen, M., and P. Reunanen. 1999. On critical thresh-olds in landscape connectivity: a management perspective.Oikos 84:302–305.
Monkkonen, M., R. Tornberg, and P. Vaisanen. 2000. Prox-imity to Goshawk nest may reduce predation rates on birds’nests. Wildlife in Finland 46:27–36. [In Finnish with En-glish summary.]
Monkkonen, M., and P. Viro. 1997. Taxonomic diversity ofthe terrestrial bird and mammal faunas in temperate andboreal biomes of the Northern Hemisphere. Journal of Bio-geography 24:603–612.
Monkkonen, M., and D. A. Welsh. 1994. A biogeographicalhypothesis of the effects of human caused landscape chang-es on the forest bird communities of Europe and NorthAmerica. Annales Zoologici Fennici 31:61–70.
Murphy, E. C., and W. A. Lehnhausen. 1998. Density andforaging ecology of woodpeckers following a stand-re-placement fire. Journal of Wildlife Management 62:1359–1372.
Newton, I. 1994. Experiments on the limitation of bird breed-ing densities: a review. Ibis 136:397–411.
Niemela, J. 1999. Management in relation to disturbance inthe boreal forest. Forest Ecology and Management 115:127–134.
Niemi, G., J. Hanowski, P. Helle, R. Howe, M. Monkkonen,L. Venier, and D. Welsh. 1998. Ecological sustainabilityof birds in boreal forests. Conservation Ecology 2(2):17.[online]. URL: http://www.consecol.org/vol2/iss2/art17
O’Connor, R. J. 1981. Comparisons between migrant andnon-migrant birds in Britain. Pages 167–195 in D. J. Aid-ley, editor. Animal migration. Cambridge University Press,Cambridge, UK.
O’Connor, R. J. 1992. Population variation in relation tomigrancy status in some North American birds. Pages 64–74 in J. M. Hagan and D. W. Johnston, editors. Ecologyand conservation of neotropical migrant landbirds. Smith-sonian Institution Press, Washington, D.C., USA.
Peters, R. H. 1983. The ecological implications of body size.Cambridge University Press, Cambridge, UK.
Pettersson, R. B. 1996. Effect of forestry on the abundanceand diversity of arboreal spiders in the boreal spruce forest.Ecography 19:221–228.
Pettersson, R. B., J. P. Ball, K. E. Renhorn, P. A. Esseen, andK. Sjoberg. 1995. Invertebrate communities in boreal for-est canopies as influenced by forestry and lichens withimplications for passerine birds. Biological Conservation74:57–63.
Preston, F. W. 1948. The commonness, and rarity, of species.Ecology 29:254–283.
Preston, F. W. 1962. The canonical distribution of commonessand rarity. Part II. Ecology 43:410–432.
Rabinowitz, D., S. Cairns, and T. Dillon. 1986. Seven formsof rarity and their frequency in the flora of the British Isles.Pages 182–204 in M. J. Soule, editor. Conservation biol-ogy: the science of scarcity and diversity. Sinauer Asso-ciates, Sunderland, Massachusetts, USA.
Raivio, S., and Y. Haila. 1990. Bird assemblages in silvi-cultural habitat mosaics in southern Finland during thebreeding season. Ornis Fennica 67:73–83.
Rappole, J. H. 1996. The importance of forest for the world’smigratory bird species. Pages 389–406 in R. M. Degraafand R. I. Miller, editors. Conservation of faunal diversityin forested landscapes. Chapman and Hall, New York, NewYork, USA.
Reed, J. M., and A. P. Dobson. 1993. Behavioural constraintsand conservation biology: conspecific attraction and re-cruitment. Trends in Ecology and Evolution 8:253–256.
Robichaud, I., M.-A. Villard, and C. S. Machtans. In press.Effects of forest regeneration on songbird movements inan industrial forest landscape. Landscape Ecology.
Robinson, S. K., F. R. Thompson, T. M. Donovan, D. R.Whitehead, and J. Faaborg. 1995. Regional forest frag-mentation and the nesting success of migratory birds. Sci-ence 267:1987–1990.
Rolstad, J. 1989. Effects of logging on capercaillie (Tetraourogallus) leks: I. Cutting experiments in southcentral Nor-way. Scandinavian Journal of Forest Research 4:99–110.
Rolstad, J. 1991. Consequences of forest fragmentation forthe dynamics of bird populations: conceptual issues andthe evidence. Biological Journal of the Linnean Society 42:149–163.
Rosenzweig, M. L. 1995. Species diversity in space and time.Cambridge University Press, Cambridge, UK.
Schieck, J., K. Lertzman, B. Nyberg, and R. Page. 1995a.Effects of patch size on birds in old-growth montane for-ests. Conservation Biology 9:1072–1084.
Schieck, J., M. Nietfeld, and J. B. Stelfox. 1995b. Differencesin bird species richness and abundance among three suc-cessional stages of aspen-dominated boreal forests. Cana-dian Journal of Zoology 73:1417–1431.
Schmiegelow, F. K. A. 1997. The effect of experimental frag-mentation on bird community dynamics in the boreal mix-edwood forest. Dissertation. University of British Colum-bia, Vancouver, British Columbia, Canada.
Schmiegelow, F. K. A., and S. J. Hannon. 1993. Adaptivemanagement, adaptive science and the effects of forest frag-mentation on boreal birds in northern Alberta. Transactionsof the North American Wildlife and Natural ResourcesConference 58:584–598.
Schmiegelow, F. K. A., and S. J. Hannon. 1999. Forest-leveleffects of management on boreal songbirds: the CallingLake fragmentation studies. Pages 201–221 in J. A. Ro-chelle, L. A. Lehmann, and J. Wisniewski, editors. Forestfragmentation: wildlife and management implications.Brill, Leiden, The Netherlands.
Schmiegelow, F. K. A., C. S. Machtans, and S. J. Hannon.1997. Are boreal birds resilient to fragmentation? An ex-perimental study of short-term community responses. Ecol-ogy 78:1914–1932.

April 2002 389HABITAT FRAGMENTATION: A REASSESSMENT
Schmiegelow, F. K. A., and T. D. Nudds. 1987. Island bio-geography of vertebrates in Georgian Bay Islands Nationalpark. Canadian Journal of Zoology 65:3041–3043.
Semenchuk, G. P. 1992. The Atlas of breeding birds of Al-berta. Federation of Alberta Naturalists. Edmonton, Al-berta, Canada.
Simberloff, D., J. A. Farr, J. Cox, and D. W. Mehlman. 1992.Movement corridors: conservation bargains or poor in-vestments? Conservation Biology 6:493–504.
Sklepkovych, B. 1997. Kinship and conflict—resource com-petition in a proto-cooperative species: the Siberian Jay.Dissertation. University of Stockholm, Akademitryck, Eds-bruk, Sweden.
Small, M. F., and M. L. Hunter. 1988. Forest fragmentationand avian nest predation in forested landscapes. Oecologia76:62–64.
Smith, A. 1993. Ecological profiles of birds in the borealforest of western Canada. Pages 14–26 in D. H. Kuhnke,editor. Birds in the boreal forest. Proceedings of a Work-shop held 10–12 March 1992 in Prince Albert, Saskatch-ewan. Northern Forestry Centre, Forestry Canada, Edmon-ton, Alberta, Canada.
Song, S. J. 1998. Effect of forest edge on songbirds breedingin the boreal forest of northern Alberta. Dissertation. Uni-versity of Alberta, Edmonton, Alberta, Candada.
Song, S. J., and S. J. Hannon. 1999. Predation in heteroge-nous forests: a comparison at natural and anthropogenicedges. Ecoscience 6:521–530.
Southwood, T. R. E. 1976. Bionomic strategies and popu-lation parameters. Pages 30–52 in R. M. May, editor. The-oretical ecology. Blackwell Scientific, Oxford, UK.
St. Clair, C. C., M. Belisle, A. Desrochers, and S. J. Hannon.1998. Winter responses of forest birds to habitat corridorsand gaps. Conservation Ecology 2(2):13. [Online, URL:^http://www.consecol.org/Journal/vol2/iss2/art13&.]
Svardson, G. 1949. Competition and habitat selection inbirds. Oikos 1:157–174.
Swenson, J., and P. Angelstam. 1993. Habitat separation bysympatric forest grouse in Fennoscandia in relation to forestsuccession. Canadian Journal of Zoology 71:1303–1310.
Terborgh, J. 1989. Where have all the birds gone? PrincetonUniversity Press, Princeton, New Jersey, USA.
Tewksbury, J. J., S. J. Hejl, and T. E. Martin. 1998. Breedingproductivity does not decline with increasing fragmentationin a western landscape. Ecology 79:2890–2903.
Tjernberg, M., K. Johnsson, and S. G. Nilsson. 1993. Densityvariation and breeding success of the black woodpecker
Dryocopus martius in relation to forest fragmentation. Or-nis Fennica 70:155–162.
Tornberg, R., M. Monkkonen, and M. Pahkala. 1999. Chang-es in diet and morphology of Finnish goshawks from 1960sto 1990s. Oecologia 121:369–376.
USGS Patuxent Wildlife Research Center. 2000. NorthAmerican Breeding Bird Survey Internet data set. [Online,URL: ^http://www.mp2-pwrc.usgs.gov/bbs/retrieval/ &.]
Vaisanen, R. A., O. Jarvinen, and P. Rauhala. 1986. How areextensive, human-caused habitat alterations expressed onthe scale of local bird populations in boreal forest? OrnisScandinavica 17:282–292.
Vaisanen, R. A., E. Lammi, and P. Koskimies. 1998. Distri-bution, numbers and population changes of Finnish breed-ing birds. Otava, Helsinki, Finland. [In Finnish with En-glish summary.]
Villard, M.-A. 1998. On forest-interior species, edge avoid-ance, area sensitivity, and dogmas in avian conservation.Auk 115:801–805.
Virkkala, R. 1991. Population trends of forest birds in a Finn-ish Lapland landscape of large habitat blocks: consequenc-es of stochastic environmental variations or regional habitatalteration? Biological Conservation 56:223–240.
Wegge, P., and J. Rolstad. 1986. Size and spacing of cap-ercaillie leks in relation to social behavior and habitat. Be-havioral Ecology and Sociobiology 19:401–408.
Welsh, D. A. 1995. An overview of the Forest Bird Moni-toring Program in Ontario, Canada. Pacific Southwest Re-search Station, Albany, California, USA. U.S. Forest Ser-vice PSW-GTR-149.
Whitcomb, R. F., C. S. Robbins, J. F. Lynch, B. L. Whitcomb,M. K. Klimkiewicz, and D. Bystrak. 1981. Effects of forestfragmentation on avifauna of the eastern deciduous forest.Pages 125–205 in R. L. Burgess and D. M. Sharpe, editors.Forest island dynamics in man-dominated landscapes.Springer-Verlag, New York, New York, USA.
Widen, P. 1989. The hunting habits of Goshawk Accipitergentiles in boreal forests of central Sweden. Ibis 131:205–213.
Wilcove, D. S. 1985. Nest predation in forest tracts and thedecline of migratory songbirds. Ecology 66:1211–1214.
With, K. A. 1999. Is landscape connectivity necessary andsufficient for wildlife management? Pages 97–115 in J. A.Rochelle, L. A. Lehmann, and J. Wisniewski, editors. For-est fragmentation: wildlife and management implications.Brill, Leiden, The Netherlands.
With, K. A., and T. O. Crist. 1995. Critical thresholds inspecies’ response to landscape structure. Ecology 76:2446–2459.




















