
Glutamate Uptake Impairment and Neuronal Damage in Young and Aged Rats In Vivo
10
Journal ofNeurochemistry Lippincott—Raven Publishers, Philadelphia © 1997 International Society for Neurochemistry Glutamate Uptake Impairment and Neuronal Damage in Young and Aged Rats In Vivo Lourdes Massieu and Ricardo Tapia Departamento de Neurociencias, Instituto de Fisiologla Celular, Universidad Nacional Autónoma de Mexico, Mexico D.F., Mexico Abstract: The extracellular concentration of glutamate increases during hypoxia/ischemia probably due to de- ficient uptake. Glutamate might contribute to neuronal damage associated with this disorder and to neurodegen- eration during aging. In the present study, we have tested the effect of two inhibitors of glutamate transport, L-trans- pyrrolidine-2,4-dicarboxylate and dihydrokainate, on the extracellular levels of glutamate and on neuronal damage, which was quantitatively studied by image analysis of histological brain sections. Drugs were administered by microdialysis and glutamate concentration was deter- mined by HPLC in the striatum and the hippocampus of 3-month-old and 22—24-month-old rats. In both regions studied, the basal concentration of extracellular gluta- mate was higher in aged than in young rats. Pyrrolidine dicarboxylate induced a substantial elevation of extracel- lular glutamate in both regions, and although this increase was almost twofold higher in old than in young animals, no neuronal damage was observed. In contrast, dihydro- kainate had a poor effect on glutamate levels, but induced clear neuronal damage in the striatum and the hippocam- pus in both groups of rats. The present results suggest that age appears not to be a significant factor in the sensi- tivity of neurons to the toxic effect of extracellular gluta- mate increase via blockade of its transport system. Key Words: Dihydrokainate—Glutamate uptake— Hippo- campus— Microdialysis— Neuronal damage— Pyrroli- dine dicarboxylate—Striatum. J. Neurochem. 69, 1151—1160 (1997). A large body of evidence suggests that excitatory amino acids, such as glutamate and aspartate, may play an important role in the process of neuronal death asso- ciated with several diseases. During cerebral ischemia (Benveniste et al., 1984; Globus et a!., 1988), hypo- glycemia (Sandberg et a!., 1986), and cerebral trauma (Faden et a!., 1989), the extracellular concentration of glutamate increases substantially, due probably to an exaggerated release (Drejer et a!., 1985) or an impair- ment of its high-affinity uptake system (Nicholls and Attwell, 1990). Moreover, neuronal death associated with these neurological disorders can be prevented ef- fectively by the administration of glutamate receptor antagonists (Simon et a!., 1984; Gill et a!., 1988; Swan and Meidrum, 1990). During aging, certain populations of neurons degen- erate, including the pyramidal cells in the hippocampus (Mann, 1994). In addition, some studies show that aged rats are more susceptible to neuronal damage induced by some toxic compounds. For example, ad- ministration of drugs impairing energy metabolism, such as 3-acetylpyridine, which inhibits glycolysis, or 3-nitropropionic acid, which inhibits oxidative metab- olism, induces an age-dependent neurodegeneration of the striatum (Bossi et a!., 1993; Brouillet et al., 1993; Schulz et a!., 1994). It is interesting that both types of lesions apparently lead to secondary excitotoxic mechanisms (Beal et a!., 1993; Schulz et a!., 1994). An increased vulnerability of aged individuals to the neurological or epileptogenic effects of domoic acid in humans and of kainic acid in rats has also been described (Teitelbaum et al., 1990; Wozniak et a!., 1991), although other reports show that aged rats are less susceptible to the excitotoxic action of kainic and quinolinic acids (Finn et al., 1991; Kesslak et a!., 1995). The mechanisms responsible for the age-related pro- pensity to neurodegeneration are not understood, but might involve alterations in glutamate metabolism, re- lease, and uptake resulting in increased extracellular levels of this amino acid. Some evidence suggests a putative role of glutamate in the pathogenesis of Alz- heimer’s disease (Greenamyre and Young, 1989; Matt- son, 1990), and pioneering studies showed that direct administration of the glutamate analogues kainic and quinolinic acids reproduces several of the neurochemi- cal changes observed in Huntington’s disease (Coyle Received November 5, 1996; revised manuscript received March 14, 1997; accepted April 16, 1997. Address correspondence and reprint requests to Dr. L. Massieu at Departamento de Neurociencias, Instituto de FisiologIa Celular, Universidad Nacional Autdnoma de Mexico, AP 70-253, 045 10- Mexico D.F., Mexico. Abbreviations used: DHK, dihydrokainate; PDC, L-trans-pyrrOli- dine-2,4-dicarboxylate. 1151
Transcript of Glutamate Uptake Impairment and Neuronal Damage in Young and Aged Rats In Vivo

Journal ofNeurochemistryLippincott—Raven Publishers, Philadelphia© 1997 International Society for Neurochemistry
Glutamate Uptake Impairment and Neuronal Damagein Young and Aged Rats In Vivo
Lourdes Massieu and Ricardo Tapia
Departamento de Neurociencias, Instituto de Fisiologla Celular, Universidad Nacional Autónoma de Mexico,Mexico D.F., Mexico
Abstract: The extracellular concentration of glutamateincreases during hypoxia/ischemia probably due to de-ficient uptake. Glutamate might contribute to neuronaldamage associated with this disorder and to neurodegen-eration during aging. In the present study, we have testedthe effect of two inhibitors of glutamate transport, L-trans-pyrrolidine-2,4-dicarboxylate and dihydrokainate, on theextracellular levels of glutamate and on neuronal damage,which was quantitatively studied by image analysis ofhistological brain sections. Drugs were administered bymicrodialysis and glutamate concentration was deter-mined by HPLC in the striatum and the hippocampus of3-month-old and 22—24-month-old rats. In both regionsstudied, the basal concentration of extracellular gluta-mate was higher in aged than in young rats. Pyrrolidinedicarboxylate induced a substantial elevation of extracel-lularglutamate in both regions, and although this increasewas almost twofold higher in old than in young animals,no neuronal damage was observed. In contrast, dihydro-kainate had a poor effect on glutamate levels, but inducedclear neuronal damage in the striatum and the hippocam-pus in both groups of rats. The present results suggestthat age appears not to be a significant factor in the sensi-tivity of neurons to the toxic effect of extracellular gluta-mate increase via blockade of its transport system. KeyWords: Dihydrokainate—Glutamate uptake— Hippo-campus— Microdialysis— Neuronal damage— Pyrroli-dine dicarboxylate—Striatum.J. Neurochem. 69, 1151—1160 (1997).
A large body of evidence suggests that excitatoryamino acids, such as glutamate and aspartate, may playan important role in the process of neuronal death asso-ciated with several diseases. During cerebral ischemia(Benveniste et al., 1984; Globus et a!., 1988), hypo-glycemia (Sandberg et a!., 1986), and cerebral trauma(Faden et a!., 1989), the extracellular concentration ofglutamate increases substantially, due probably to anexaggerated release (Drejer et a!., 1985) or an impair-ment of its high-affinity uptake system (Nicholls andAttwell, 1990). Moreover, neuronal death associatedwith these neurological disorders can be prevented ef-fectively by the administration of glutamate receptor
antagonists (Simon et a!., 1984; Gill et a!., 1988; Swanand Meidrum, 1990).
During aging, certain populations of neurons degen-erate, including the pyramidal cells in the hippocampus(Mann, 1994). In addition, some studies show thataged rats are more susceptible to neuronal damageinduced by some toxic compounds. For example, ad-ministration of drugs impairing energy metabolism,such as 3-acetylpyridine, which inhibits glycolysis, or3-nitropropionic acid, which inhibits oxidative metab-olism, induces an age-dependent neurodegeneration ofthe striatum (Bossi et a!., 1993; Brouillet et al., 1993;Schulz et a!., 1994). It is interesting that both typesof lesions apparently lead to secondary excitotoxicmechanisms (Beal et a!., 1993; Schulz et a!., 1994).An increased vulnerability of aged individuals to theneurological or epileptogenic effects of domoic acidin humans and of kainic acid in rats has also beendescribed (Teitelbaum et al., 1990; Wozniak et a!.,1991), although other reports show that aged rats areless susceptible to the excitotoxic action of kainic andquinolinic acids (Finn et al., 1991; Kesslak et a!.,1995).
The mechanisms responsible for the age-related pro-pensity to neurodegeneration are not understood, butmight involve alterations in glutamate metabolism, re-lease, and uptake resulting in increased extracellularlevels of this amino acid. Some evidence suggests aputative role of glutamate in the pathogenesis of Alz-heimer’s disease (Greenamyre and Young, 1989; Matt-son, 1990), and pioneering studies showed that directadministration of the glutamate analogues kainic andquinolinic acids reproduces several of the neurochemi-cal changes observed in Huntington’s disease (Coyle
Received November 5, 1996; revised manuscript received March14, 1997; accepted April 16, 1997.
Address correspondence and reprint requests to Dr. L. Massieuat Departamento de Neurociencias, Instituto de FisiologIa Celular,Universidad Nacional Autdnoma de Mexico, AP 70-253, 045 10-Mexico D.F., Mexico.
Abbreviations used: DHK, dihydrokainate; PDC, L-trans-pyrrOli-
dine-2,4-dicarboxylate.
1151

1152 L MASSIEU AND R. TAPIA
and Schwarcz, 1976; McGeer and McGeer, 1976;Schwarcz et al., 1983).
In a previous study, we found that in vivo intrastria-ta! microdialysis administration of two glutamate up-take inhibitors, dihydrokainate (DHK) and pyrroli-dine-2,4-dicarboxylate (PDC), to young rats led tosubstantial elevations of extrace!!u!ar glutamate con-centrations. However, striata! neurona! damage wasfound only when DHK was administered (Massieu eta!., 1995). To investigate whether the toxic effect ofelevated glutamate concentrations is age-dependent, inthe present work we have compared the vulnerabilityto glutamate uptake impairment in vivo in young (3-month-old) and aged (22—24-month-old) rats. Fur-thermore, we have extended the study to thehippocam-pus, a region that shows age-related neuropathologica!changes, such as those observed in Alzheimer’ s dis-ease.
MATERIALS AND METHODS
AnimalsThree month-old and 22—24-month-old Wistar male rats
(Camm Research Lab Animals, Wayne, NJ, U.S.A.) wereused throughout and handled in accordance with the Rulesfor Research in Health Matters (Mexico), with approval ofthe local animal care committee. They were anesthetizedwith 4—5% halothane in a 95% 02/5% CO2 mixture andplaced in a stereotaxic frame with the nose bar positionedat —3.3 mm. The skin was cut open to expose the skull,and a 1—2-mm hole was drilled. Animals were implantedunilaterally with microdialysis cannulas 2 mm long and 0.5mm in diameter (CMA12, Bioana!ytical Systems, West La-fayette, IN, U.S.A.) in either the striatum [coordinates: A+0.7 mm from bregma, L 2.8 mm from midline, and V 6.0mm from dura for young animals, and A +1.0 mm, L 2.9mm, and V 6.0 mm for aged rats (Paxinos and Watson,1986)1 or thehippocampus (coordinates: A —3.6 mm, L 2.0mm, and V 4.0 mm for young animals, and A —3.9 mm, L2.3 mm, and V 4.! mm for aged rats). Animals were main-tained under low anesthesia (0.5% halothane) throughoutthe experiment. Theprobes were perfused continuously witha Krebs—Ringersolution containing 118 mM NaCl, 4.5 mMKC!, 2.5 mM MgSO4, 4.0 mM NaH2PO4, 2.5 mM CaC!2,25 mM NaHCO3, and 10 mM glucose, pH 7.4, at aflux rateof 2 /i!/min, using a microinjection pump (Carnegie CMA/100). After a 60-mm equilibration period, 25-si! (12.5 mm)consecutive fractions of perfusate were collected continu-ously. After the first three fractions, which were analyzed todetermine the basal levels of amino acids, DHK (25 mM),PDC (25 and 100 mM), or Ringer solution (vehicle control)was perfused for seven consecutive fractions. Extrapolatingfrom the data of dialysis membrane efficiency for aminoacids, <10% of these inhibitor doses may reach the extracel-lular space, which is enough to increase extracellular gluta-mate substantially (Massieu et a!., 1995). At the end ofthe experiment, the skin was sutured and the animals weremaintained for 1 week under standard animal room condi-tions, with food and water ad libitum, and then used forhistological evaluation.
Stock solutions of DHK and PDC (obtained from TocrisCookson, St. Louis, MO, U.S.A.) were prepared by dissolv-
ing the compounds in 1 M NaOH and adjusting the pH to7—7.5. Aliquots of the stock solutions were added directlyto the Krebs—Ringer perfusion medium to give the concen-trations indicated above.
Amino acid analysisAmino acids were measured irs the dialysates by HPLC,
as previously described (Salazar et a!., 1994). In brief, the25-,ul collected fractions were derivatized with the samevolume of o-phthaldialdehyde and 3 mm later 20 ~tl wasinjected into aBeckman liquid chromatograph equipped withan ODS column (25 cm X 4 mm i.d.). The mobile phasewas methanol/potassium acetate (0.1 M, pH 5.5) and wasrun at a rate of 1.5 ml/min in a linear gradient (15-mmduration) from 25 to 75% methanol. Amino acid quantifica-tion was made by comparison with a standard mixture ofamino acids (Sigma, St. Louis, MO, U.S.A.) processed inthe same manner.
Histological evaluationSeven days after surgery, rats were anesthetized with intra-
peritonea! sodium pentobarbita! and transcardially perfusedwith 250 ml of 0.9% saline, followed by 250 ml of 4%paraformaldehyde in 0.! M phosphate buffer, pH 7.4. Brainswere removed, left in fixative for an additional 24 h, andtransferred successively to 10%, 20%, and 30% sucrose (24h each). Brains were paraffin-embedded, and 10-jim sectionswere obtained and stained with cresy! violet. Twelve to fif-teen sections from each brain were examined, and the le-sioned area was quantified with theaid ofan image analyzersystem (Macintosh Image 1.6). The average damaged areaper tissue section in each animal was calculated, and resultsare expressed as means of the damaged area per group oflesioned rats. It was not possible to analyze all the animalsin each series of experiments, because not all of them sur-viveduntil day 7 after the microdialysis. These animal losseswere due most probably to the surgical procedure rather thanto the drugs injected, because, as indicated in the figurelegends, no correlation was found within each experimentalgroup between the severity of the lesion and the number ofsurvivors.
Statistical analysis was carried out by aone-way ANOVAtest, followed by aFisher’s least significant difference multi-ple comparison test.
RESULTS
Extracellular basal levels of amino acidsBasal concentrations of amino acids collected in di-
alysate perfusates from young rats are in agreementwith previous data (Donzanti et a!., 1983; Cobo et a!.,1992; Massieu et a!., 1995). Glutamate values variedfrom 0.5 to 2 1uM in both the striatum and the hippo-campus and were significantly higher in aged animalsin both regions (Figs. 1 and 2).
As indicated in Table 2, in the striatum the basalconcentration of glutamine was significantly lower(7.9 versus 15.2 1tM) and that of taurine significantlyhigher (3.1 versus 2.3 jiM) in the aged rats comparedwith that in the young rats. In the hippocampus, basallevels of aspartate were lower (0.36 versus 0.75 jiM,Fig. 2) and those of glutamine higher (17.5 versus 9.0
J. Neurochem., Vol. 69, No. 3, 1997

GLUTAMATE UPTAKE INHIBITION AND AGING 1153
FIG. 1. Effect of PDC on the extracellularconcentration of glutamate and aspartate inthe striatum and the hippocampus of 3-month-old (young) and 22—24-month-old(aged) rats. Microdialysis was carried out asdescribed in Materials and Methods. The firstthree fractions correspondto the basal levelsof amino acids, and at the end of the thirdfraction, PDC (100 mM in the striatum and25 mM in the hippocampus) was per-fused for seven additional fractions. Insets:Graphs show the basal concentrations ofglutamate or aspartate, averaged from thefirst three fraction values. Data are means±SEM for the number of animals indicatedin parentheses. *p < 0.05.
jiM, Table 2) in the aged animals. The basal valuesof alanine and glycine were very similar in both experi-mental groups in the two regions studied. The basalconcentration of GABA was too low to be determinedreliably with the methodology used.
Changes in the extracellular levels of amino acidsinduced by administration of glutamate uptakeinhibitors
PDC effects. When 25 mM PDC was administeredin the striatum through the dialysis probe, a 10-fold
FIG. 2. Effect of DHK (25 mM) on theextracellular concentration of glutamateand aspartate in the striatum and thehippocampus of young and aged rats.Experimental details are as in Fig. 1. In-sets: Graphs show the basal concentra-tions of glutamate or aspartate, aver-aged from the first three fraction values.Data are means ±SEM for the numberof animals indicated in parentheses. *p< 0.05.
J. Neurochem., Vol. 69, No. 3, 1997

1154 L. MASSIEU AND R. TAPIA
TABLE 1. Bperfu
asal extracellular levels of amino acids and their changes induced by PDCsion in the striatum and the hippocampus of young and aged rats
Basal levels (1iM) Levels at peak stimulation (1iM)
Young Aged Young Aged
StriatumAlanineGlutamineGlycineTaurine
HippocampusAlanineGlutamineGlycineTaurine
1.60 ±0.12 1.36 ±0.22 4.12 ± 1.04 (157) 3.22 ±0.61 (136)10.1 ±1.13 7.89 ± 1.81 22.0 ±5.83 (117) 19.0 ±6.00 (140)1.07 ±0.12 1.61 ±0.20 2.81 ±0.52 (162) 3.78 ±0.69 (134)1.59 ±0.09 2.49 ±0.26” 12.0 ± 1.81 (655) 13.2 ±2.01 (432)
2.22 ±0.52 2.50 ±0.26 2.46 ±0.76 (11) 3.19 ±0.55 (27)11.7 ±2.37 17.4 ±2.32 21.6 ±4.05 (85) 24.2 ±5.55 (39)2.10 ±0.42 2.03 ±0.19 3.93 ±1.16 (87) 2.78 ±0.57 (37)4.36 ±0.46 5.7 ±0.53 10.2 ±1.31 (134) 12.8 ±4.62 (125)
Amino acid levels were determined by HPLC as described in Materials and Methods. Basal levelscorrespond to the pooled data of the first three fractions collected, and peak stimulation levels correspondto concentrations determined in fractions 5, 6, or 7. PDC concentration was 100 mM for the striatum and25 mM for the hippocampus. Values are means ±SEM of seven to nine rats for the striatum and five orsix animals for the hippocampus. The numbers in parentheses are the percent increase with respect to thecorresponding basal level.
“p < 0.05 as compared with young rats.
increase in the extrace!!u!ar concentration of glutamate(up to 10 jiM) was observed in young animals. Thisincrease was sustained as long as PDC was perfused(n = 8; data not shown). No age-dependent effect ofPDC was observed at this concentration, because avery similar increment in glutamate levels was ob-served in the presence of this inhibitor in the aged rats(up to 12.5 jiM, n = 5; data not shown). When theconcentration of PDC was raised to 100 mM, a re-markable effect of this inhibitor was observed in thestriatum in both experimental groups (Fig. 1). Inyoung rats, the extracellular levels of glutamate in-creased more than 20-fold its basal value, reachinga 17 jiM concentration, whereas in aged animals thestimulatory effect of PDC was also notable (15-foldits basal levels). The extracellular concentration ofglutamate reached in the presence of PDC in the agedgroup was as high as 31.2 jiM, almost twice the valueobserved in young animals (p < 0.05; Fig. 1). PDCat 100 mMalso induced a notable elevation of extracel-lular aspartate (>80-fold and >30-fold in young andaged rats, respectively), but the peak concentrationreached in both cases was similar (close to 10 jiM;Fig. 1).
PDC produced only relatively small changes in theextracellu!ar levels of other amino acids in the stria-tum: <200% increase was observed in alanine, g!uta-mine, and glycine concentrations in the two experi-mental groups; more important changes in taurine(>600% in young and >400% in aged rats) wereobserved (Table 1).
In contrast to the lack of an age-dependent effect of25 mM PDC in the striatum, in the hippocampus thisconcentration induced a significantly higher increaseof extrace!!ular glutamate in aged rats compared withyoung rats, reaching peak concentrations of 26.8 jiM
and 17.3 jiM, respectively (Fig. 1). PDC at 25 mMalso induced important changes in the extracellular lev-els of aspartate, which were elevated 11-fold relativeto its basal concentration in the young group and 26-fold in the aged group. However, the peak concentra-tion of aspartate was not statistically different betweenthe two experimental groups (Fig. 1). PDC inducedchanges smaller than or close to 100% in the extracel-lular levels of alanine, glutamine, glycine, and taurinein both the young and the aged group (Table I).
DHK effects. In contrast to PDC, DHK had a pooreffect on glutamate accumulation in the striatum. Peakconcentrations of 3.7 and 8.2 jiM were observed in the3-month-old and 22—24-month-old groups, respec-tively. Although in all collected fractions glutamateconcentration was higher in aged animals, this differ-ence was statistically significant only in fraction 6,where values were 6.63 ±1.2 jiM for aged rats and3.75 ±0.5 jiM for young rats (Fig. 2). Similarly, theeffect of DHK on aspartate levels was much weakerthan that of PDC. Increases of 1.4- and 2.9-fold wereobserved in young and aged rats, respectively.
In the hippocampus, DHK also had a weak effecton glutamate accumulation. This effect was higher inyoung (2.8-fold increase) than in aged animals (1.6-fold increase), but the maximum concentrationsreached were very similar, 6.2 and 5.9 jiM, respec-tively (Fig. 2). The changes induced by DHK in aspar-tate levels were also weaker than those induced byPDC. Although in all fractions the concentration ofaspartate was higher in the young group, these valueswere not statistically different from those in the agedanimals.
Changes in the other amino acids studied were<100%, except for taurine, whose levels were in-creased 700% in the striatum and 200% in the hippo-
J. Neurochem., Vol. 69, No. 3, 1997

GLUTAMATE UPTAKE INHIBITION AND AGING 1155
TABLE 2. Basal extracellular levels of amino acids and their changes induced by DHKperfusion in the striatum and the hippocampus ofyoung and aged rats
Basal levels (jiM) Levels at peak stimulati on (jiM)
Young Aged Young Aged
StriatumAlanineGlutamineGlycineTaurine
HippocampusAlanineGlutamineGlycineTaurine
2.6815.22.922.32
3.779.083.104.55
±0.55±1.03±0.41±0.18
±0.31±1.15±0.28±0.29
1.82 ±0.527.99 ± 1.21”2.05 ±0.343.17 ±0.25”
4.24 ± 1.0517.5 ±2.26”3.02 ±0.715.50 ±0.67
4.82 ±1.49 (80)27.6 ±7.13 (82)5.39 ± 1.18 (85)19.3 ±6.68 (731)
7.22 ±0.88 (91)12.5 ±3.72 (38)7.53 ±0.94 (143)14.2 ± 1.96 (213)
2.839.103.7825.9
5.2317.65.0915.2
±1.23 (55)±1.93” (14)±1.23 (84)±5.77 (717)
±1.45 (23)±2.40 (0.5)±1.54 (68)±5.61 (177)
Amino acid levels were determined by HPLC as described in Materials and Methods. Basal levelscorrespond to the pooled data of the first three fractions collected, and peak stimulation levels correspondto concentrations determined in fractions 5, 6, or 7. The concentration of DHK was 25 mM. Values aremeans ±SEM of five or six animals. The numbers in parentheses are the percent increase with respect tothe corresponding basal level.
“p < 0.05, as compared with young rats.
campus. No age-dependent significant difference wasfound in the peak concentrations of alanine, glycine,and taurine induced by DHK, in either the striatum orthe hippocampus, except for glutamine whose peaklevels were lower in aged animals in the striatum (Ta-ble 2).
Neuronal degenerationAs described in Materials and Methods, neuronal
damage was assessed by image analysis of cresy! vio-let-stained brain sections 7 days after the microdialysisprocedures.
As previously reported (Massieu et a!., 1995), noneurona! damage was observed when young rats wereperfused with 25 mM PDC in the striatum (data notshown). Similarly, none of the aged animals perfusedwith 25 mM PDC showed any damage (data notshown). When the concentration of PDC was raisedto 100 mM, again no neuronal damage occurred ineither young or aged animals. As shown in Fig. 3Cand F, in the two experimental groups normal largecells with clearly defined nuclei were present, even inclose vicinity to the perfusion site. The injected stria-turn looked identical to the contralateral side (compareFig. 3A and D with C and F). Quantification of thedamaged area showed no significant differences be-tween the PDC-treated tissue and the control tissueperfused with vehicle (Table 3), which indicates thatthe PDC-induced lesion is due to probe insertion. Infact, in both PDC and vehicle groups, the damage wasrestricted to the vicinity of the probe tract.
In the hippocampus, perfusion with 25 mM PDCalso failed to produce any lesion different from theRinger control group in both young and aged animals(Table 3). As indicated in Fig. 4C, D, G, and H, therewas no damage beyond the probe tract in both groupsof animals.
In contrast to PDC and in agreement with previousdata (Massieu et a!., 1995), perfusion with DHK re-sulted in neuronal damage in the striatum. Figure 3Bshows a representative striata! tissue from a younganimal with numerous cells with pyknotic nuclei, typi-cal of damaged neurons, and very few normal cells.Figure 3E shows a representative picture of the stria-tum of the aged group of rats. Although numerouspyknotic cells are present, in this group of animals thelesion was moderate, because many large cells withclear cytoplasm and well defined nuclei, similar tothose observed in the contralatera! noninjected stria-tum, can be seen as well (compare Fig. 3E and D).Quantitative image analysis confirmed the differencesbetween the extent of damage between young and agedrats (Table 3).
Perfusion with 25 mM DHK also induced neuronaldamage in the hippocampus of both groups of animals.A marked and widespread neurodegeneration was ob-served in the CAl region, and some damage was pres-ent in CA3 and CA4/hilus (Table 3), as well as in adiscrete zone of the dentate gyrus close to the tract ofthe dialysis probe. As shown in Fig. 4A, B, E, and F,numerous pyknotic cells are present all along the CAlregion and basically no normal cells can be found inthis area.
DISCUSSION
The most important findings of the present work arethe following: (a) basal extrace!!u!ar concentrationsof glutamate were significantly higher in aged than inyoung rats, in both the striatum and the hippocampus;(b) the extracellular concentrations of glutamate andaspartate were remarkably increased in the presenceof PDC in the striatum and the hippocampus of bothyoung and aged rats; (c) the percent stimulatory effect
J. Neurochem., Vol. 69, No. 3, 1997
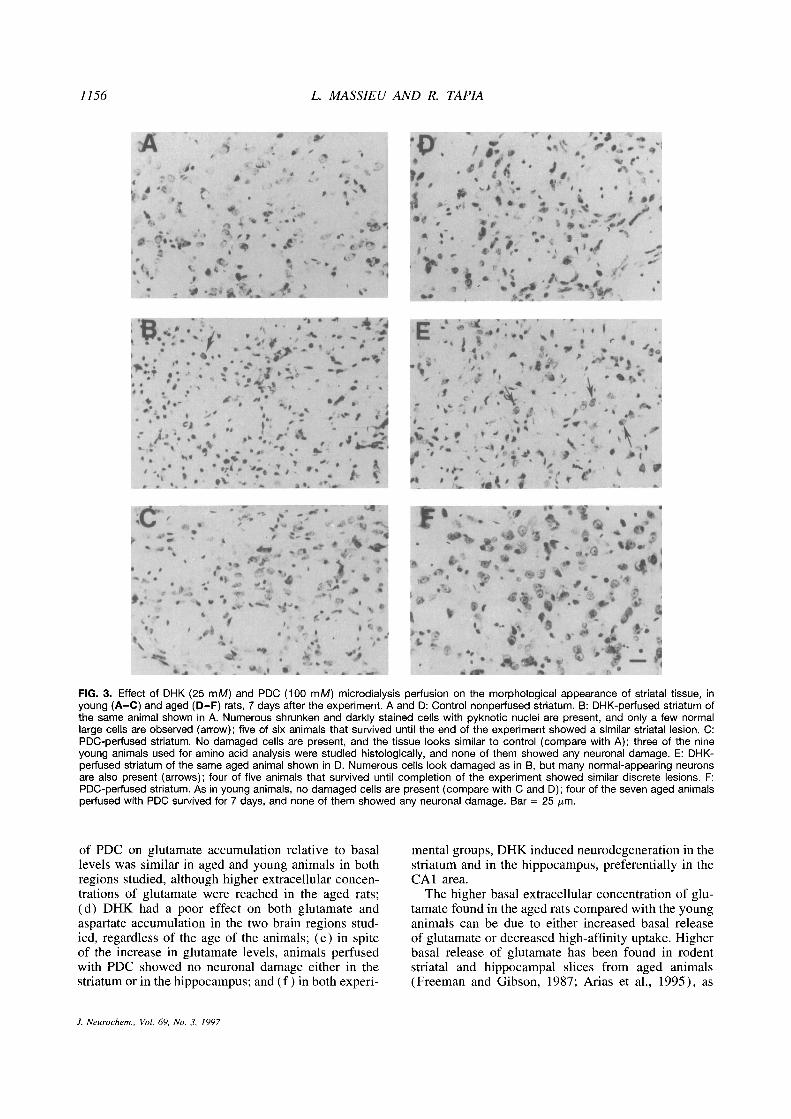
1156 L. MASSIEU AND R. TAPIA
FIG. 3. Effect of DHK (25 mM) and PDC (100 mM) microdialysis perfusion on the morphological appearance of striatal tissue, inyoung (A—C) and aged (D—F) rats, 7 days after the experiment. A and 0: Control nonperfused striatum. B: DHK-perfused striatum ofthe same animal shown in A. Numerous shrunken and darkly stained cells with pyknotic nuclei are present, and only a few normallarge cells are observed (arrow); five of six animals that survived until the end of the experiment showed a similar striatal lesion. C:PDC-perfused striatum. No damaged cells are present, and the tissue looks similar to control (compare with A); three of the nineyoung animals used for amino acid analysis were studied histologically, and none of them showed any neuronal damage. E: DHK-perfused striatum of the same aged animal shown in D. Numerous cells look damaged as in B, but many normal-appearing neuronsare also present (arrows); four of five animals that survived until completion of the experiment showed similar discrete lesions. F:PDC-perfused striatum. As in young animals, no damaged cells are present (compare with C and D); four of the seven aged animalsperfused with PDC survived for 7 days, and none of them showed any neuronal damage. Bar = 25 jim.
of PDC on glutamate accumulation relative to basallevels was similar in aged and young animals in bothregions studied, although higher extracellular concen-trations of glutamate were reached in the aged rats;(d) DHK had a poor effect on both glutamate andaspartate accumulation in the two brain regions stud-ied, regardless of the age of the animals; (e) in spiteof the increase in glutamate levels, animals perfusedwith PDC showed no neurona! damage either in thestriatumor in the hippocampus; and (f) in both experi-
mental groups, DHK induced neurodegeneration in thestriatum and in the hippocampus, preferentially in theCAl area.
The higher basal extracellular concentration of glu-tamate found in the aged rats compared with the younganimals can be due to either increased basal releaseof glutamate or decreased high-affinity uptake. Higherbasal release of glutamate has been found in rodentstriatal and hippocampal slices from aged animals(Freeman and Gibson, 1987; Arias et a!., 1995), as
J. Neurochem., Vol. 69, No. 3, 1997

GLUTAMATE UPTAKE INHIBITION AND AGING 1157
TABLE 3. Damaged area (,um2 X 1O~)per tissue
section ofyoung and aged rats 7 days after perfusion withDHK or PDC in the striatum and the hippocampus
Young Aged
StriatumDHK (25 mM) 2,672 ±667 (
5/5)~.b 931 ±131 (4/4)PDC (100 mM) 568 ±32 (3/3) 564 ± 56 (4/4)Ringer 407 ±57 (3/3) ND
HippocampusDHK (25 mM)
CAl 212 ±16.2 (5/5)b 206 ±14.5 (3/3)CA3 58.4 ±15.8 (4/5) 56.6 ±3.2 (3/3)CA4/hilus 54.6 ±10.6 (4/5) 50, 70 (2/3)
PDC (25 mM)CAl 30.7 ±3.4 (4/4) 26.0 ±9.1 (5/5)CA3 0 (0/4) 0 (0/5)CA4/hilus 0 (0/4) 0 (0/5)
RingerCAl 15.5 ±4.2 (3/3) NDCA3 0 (0/3) NDCA4/hilus 0 (0/3) ND
Values are means ±SEM from 12—15 sections for each damagedrat. The number of rats showing damage relative to the total numberof animals histologically analyzed is indicated in parentheses. ND,not determined.
“p < 0.05 vs. aged rats; b~< 0.05 vs. PDC or Ringer. PDC vs.Ringer, no significant differences in any case.
we!! as in the lateral striatum of aged rats in vivo(Donzanti et a!., 1983). However, glutamate uptakehas been found to be either decreased or increased insynaptosomes from aged rats compared with young(Price et al., 1981; Najierahim et a!., 1990), and nodifference has been found in hippocampa! slices(Palmer et a!., 1994).
Of the other amino acids measured, the basal con-centration of glutamine was significantly decreased inthe striatum and significantly increased in the hippo-campus in the aged group. An age-dependent decreasein the basal release of glutamine has been reported inhuman cortical prisms (Smith et a!., 1983), but in therat cortex in vivo, increased extracellular concentra-tions of g!utamine were found in 24—26-month-oldrats compared with young rats (Cobo et a!., 1992).These results suggest that basal extracellular levels ofglutamine might vary depending on the brain region,the age, and the species studied.
The finding that PDC induced a much more notableincrease in glutamate levels than DHK correlates we!!with the reported order of potency (PDC ~‘ DHK) ofthese compounds to inhibit glutamate uptake in vitro(Johnston et al., 1979; Bridges et a!., 1991; Robinsonet a!., 1991). The fact that the effect of PDC was farmore efficient for glutamate and aspartate than for theother amino acids suggests that, at the dose used, theaction of this compound is primarily on the excitatoryamino acid transporter. Part of this effect might bedue to a carrier-mediated PDC—glutamate exchange,because PDC, in contrast to DHK, is taken up by the
transporter (Bridges et al., 1991; Isaacson and Nico!!,1993; Sarantis et a!., 1993).
The most striking observation of the present studyis the lack of correlation between the increase in extra-cellular glutamate levels and neuronal damage. In aprevious study (Massieu et a!., 1995), we reported thatin the striatum of young rats glutamate levels rosesteeply when perfused with either PDC or DHK,whereas neuronal damage was observed only withDHK. In the present work, the same discrepancy wasfound in the hippocampus. The lack of toxic effect ofPDC could be explained by its limited diffusion, dueto removal by the carrier, as has been postulated forglutamate and treo-hydroxyaspartate in cerebellarslices (Garthwaite et a!., 1992). However, this possi-bility is unlikely, because a high concentration of extra-cellular glutamate was reached and no neurona! dam-age was observed, even in the tissue surrounding theprobe tract. The differential toxic effect of DHK andPDC might also be attributed to inhibition of differenttransporter molecules. At least four different glutamatetransporters have been cloned to date, with differentdistributions (neurona!versus glial) and sensitivities topharmacological inhibition (Kanai and Hediger, 1992;Pines et a!., 1992; Storck et a!., 1992; Fairman et a!.,1995). It is thus possible that these transporters maycontribute differently to the clearance of synaptic glu-tamate and therefore to glutamate toxicity. In supportof this hypothesis, it has been found recently that inhi-bition of the synthesis of the glutamate transportersin rats, by treatment with antisense oligonucleotides,results in differential glutamate uptake inhibition andneuronal damage (Rothstein et a!., 1996). Alterna-tively, the toxic effects of DHK might be due to addi-tional factors, such as a direct activation of glutamatepostsynaptic receptors, as suggested by the finding thatDHK can inhibit the sodium-independent [
3H]kainatebinding in vitro (Johnston et a!., 1979). However, theneurotoxic effect of the acute injection of DHK in vivois prevented by the previous administration of MK-801 in the striatum and in the hippocampus, whereasthat of kainate is not (Massieu et al., 1995). Thus, theneurotoxic effect of DHK appears to involve, eitherdirectly or indirectly, the activation of the NMDA glu-tamate receptor subtype.
It is interesting that, although basal levels of extra-cellular glutamate are higher in the aged rats and atwofold increase was reached in the presence of PDC(up to 31 jiM), which was sustained as long as thiscompound was perfused (>1 h), no neurona! damagewas observed. These results suggest that aged rats aremore tolerant to elevated extracellular glutamate invivo, which might be due to a decreased number orefficiency of glutamate receptors. In fact, it has beenshown that NMDA and a-amino-3-hydroxy-5-methy!i-soxazo!e-4-propionate (AMPA) /kainate receptors de-crease with age (Bonhaus et a!., 1990; Clark et a!.,1992; Magnusson and Cotman, 1993; Nagahara et a!.,1993), and aged rats are less susceptible to the toxic
.1. Neurochem., Vol. 69. No. 3, 1997
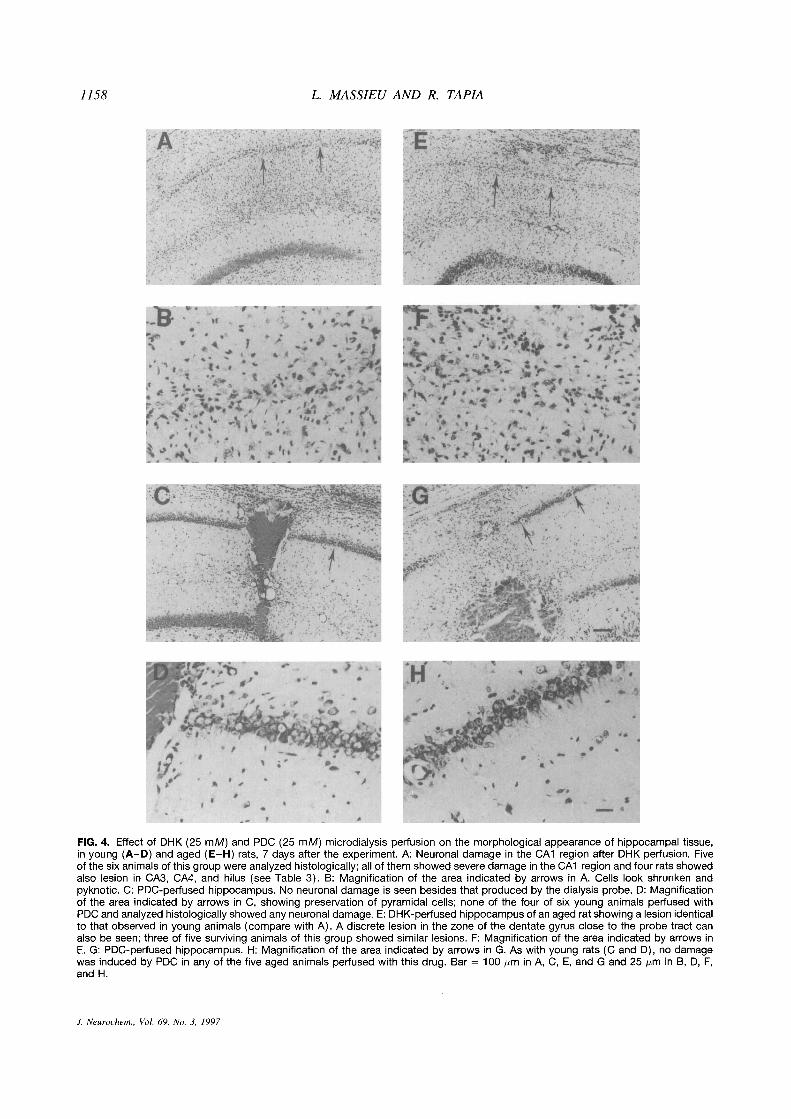
1158 L. MASSIEU AND R. TAPIA
FIG. 4. Effect of DHK (25 mM) and PDC (25 mM) microdialysis perfusion on the morphological appearance of hippocampal tissue,in young (A—D) and aged (E—H) rats, 7 days after the experiment. A: Neuronal damage in the CAl region after DHK perfusion. Fiveof the six animals ofthis group were analyzed histologically; all of them showed severe damage in the CAl region and four rats showedalso lesion in CA3, CA4, and hilus (see Table 3). B: Magnification of the area indicated by arrows in A. Cells look shrunken andpyknotic. C: PDC-perfused hippocampus. No neuronal damage is seen besides that produced by the dialysis probe. 0: Magnificationof the area indicated by arrows in C, showing preservation of pyramidal cells; none of the four of six young animals perfused withPDC and analyzed histologically showed any neuronal damage. E: DHK-perfused hippocampus of an aged rat showing a lesion identicalto that observed in young animals (compare with A). A discrete lesion in the zone of the dentate gyrus close to the probe tract canalso be seen; three of five surviving animals of this group showed similar lesions. F: Magnification of the area indicated by arrows inE. G: PDC-perfused hippocampus. H: Magnification of the area indicated by arrows in G. As with young rats (C and 0), no damagewas induced by PDC in any of the five aged animals perfused with this drug. Bar = 100 jim in A, C, E, and G and 25 jim in B, D, F,and H.
1. Neurochem., Vol. 69, No. 3, 1997

GLUTAMATE UPTAKE INHIBITION AND AGING 1159
effects of quinolinic (Finn et al., 1991) and kainic(Kesslak et al., 1995) acids. Our finding that the dam-age produced by DHK in the striatum was diminishedsignificantly in the aged group is in agreement withthis interpretation.
Acknowledgment: The authors thank Federico Jandetefor help in histological techniques and Arturo Franco forphotographic artwork. This work was supported by DGAPA(project 1N205095) of the Universidad Nacional Autónomade Mexico.
REFERENCES
Arias C., Arrieta I., and Tapia R. (1995) /3-Amyloid peptide frag-ment 25—35 potentiates the calcium-dependent release of excit-atory amino acids from depolarized hippocampal slices. J. Neu-rosci. Res. 41, 561—566.
Beal M. F., Brouillet E., Jenkins B. G., Ferrante R. J., Kowall N. W.,Miller J. M., Storey E., Srivastava R., Rosen B. R., and HymanB. T. (1993) Neurochemical and histologic characterization ofstriatal excitotoxic lesions produced by the mitochondrial toxin3-nitropropionic acid. J. Neurosci. 13, 4181—4192.
Benveniste H., Drejer J., Schousboe A., and Diemer N. H. (1984)Elevation of the extracellular concentrations of glutamate andaspartate in rat hippocampus during transient cerebral ischemiamonitored by intracerebral microdialysis. J. Neurochem. 43,1369—1374.
Bonhaus D. W., Perry W. B., and McNamara J. 0. (1990) Decreaseddensity, but not number, of N-methyl-u-aspartate. glycine andphencyclidine binding sites in hippocampus of senescent rats.Brain Res. 532, 82—86.
Bossi S. R., Simpson J. R., and Isacson 0. (1993) Age dependenceof striatal neuronal death caused by mitochondrial dysfunction.Neuroreport 4, 73—76.
Bridges R. J., Stanle M. S., Anderson M. W., Cotman C. W., andChamberli A. R. (1991) Conformationally defined neurotrans-mitter analogues. Selective inhibition of glutamate uptake byone pyrrolidine-2,4-dicarboxylate diastereomer. .1. Med. Chem.34, 717—725.
Brouillet E., Jenkins B. G., Hyman B. T., Ferrante R. J., KowallN. W., Srivastava R., Roy D. S., Rosen B. R., and Beal M. F.(1993) Age-dependent vulnerabilityof the striatum to the mito-chondrial toxin 3-nitropropionic acid. J. Neurochem. 60, 356—359.
Clark A. S., Magnusson K. R., and Cotman C. W. (1992) In vitroautoradiography of hippocampal excitatory amino acid bindingin aged Fisher 344 rats: relationship to performance on theMorris water maze. Behav. Neurosci. 106, 324—335.
Cobo M., Expdsito I., Porras A., and Mora F. (1992) Release ofamino acid neurotransmitters in different cortical areas of con-scious young and aged rats. Neurobiol. Aging 13, 705—709.
Coyle J. T. and Schwarcz R. (1976) Lesion of striatal neurons withkainic acid provides a model for Huntington’s chorea. Nature263, 244—246.
Donzanti B. A., Hite J. F., and Yamamoto B. K. (1983) Extracellularglutamate levels increase with age in the lateral striatum: poten-tial involvement of presynapticD-2 receptors. Synapse 13, 376—382.
Drejer J., Benveniste H., Diemer N. H., and Schousboe A. (1985)Cellular origin of ischemia-induced glutamate release frombrain tissue in vivo and in vitro. J. Neurochem. 45, 145— 151.
Faden A. I., Dernediuck P., Panter S. S., and Vink R. (1989) Therole ofexcitatory amino acids and NMDA receptors in traumaticbrain injury. Science 244, 798—800.
Fairman W. A., Vandenberg R. J., Arriza J. L., Kavanaugh M. P.,and Amara S. G. (1995) An excitatory amino-acid transporterwith properties of a ligand-gated chloride channel. Nature 375,599—603.
Finn S. F., Hyman B. T., Storey E., Miller J. M., and Beal M. F.(1991) Effects of aging on quinolinic acid lesions in rat stria-turn. Brain Res. 562, 278—280.
Freeman G. B. and Gibson G. E. (1987) Selective alteration ofmouse brain neurotransmitter release with age. Neurobiol.Aging 8, 147—152.
Garthwaite G., Williams G. D., and Garthwaite J. (1992) Glutamatetoxicity: an experimental and theoretical analysis. Eur. J. Neu-rosci. 4, 353—360.
Gill R., Foster A. C., and Woodruff G. N. (1988) MK8O1 is neuro-protective in gerbils when administered during the post-isch-ernie period. Neuroscience 25, 847—855.
Globus M. Y.-T., Busto R., Dietrich W. D., Martinez E., Valdes I., andGinsberg M. D. (1988) Effect of ischemia on the in vivo releaseof striatal dopamine, glutamate, and y-arninobutyric acid studiedby intracerebral microdialysis. J. Neurochem. 51, 1455—1464.
Greenamyre J. T. and Young A. B. (1989) Excitatory amino acidsand Alzheimer disease. Neurobiol. Aging 10, 593—602.
Isaacson J. S. and Nicoll R. A. (1993) The uptake inhibitor L-trans-PDC enhances responses to glutamate but fails to alter thekinetics of excitatory synaptic currents in the hippocampus. J.Neurophysiol. 70, 2187—2191.
Johnston G. A. R., Kennedy S. M. E., and Twitchin D. (1979) Ac-tion of the neurotoxin kainic acid on high affinity uptake of L-
glutamic acid in rat brain slices. J. Neurochem. 32, 121—127.Kanai Y. and Hediger M. (1992) Primary structure and functional
characterization of high-affinity glutamate transporter. Nature360, 467—471.
Kesslak J. P., Yuan D., and Cotman C. W. (1995) Vulnerability ofthe hippocampus to kainate excitotoxicity in the aged, matureand young adult rat. Neurosci. Lett. 188, 117—120.
Magnusson K. R. and Cotman C. W. (1993) Age-related changes inexcitatory amino acid receptors in two mouse strains. Neurobiol.Aging 14, 197—206.
Mann D. M. (1994) Vulnerability of specific neurons to aging, inNeurodegenerative Diseases (Calne D. B., ed), pp 15—31.W. B. Saunders, Philadelphia.
Massieu L., Morales-Villagrán A., and Tapia R. (1995) Accumula-tion of extracellular glutamate by inhibition of its uptake is notsufficient for inducing neuronal damage: an in vivo rnicrodialy-sis study. J. Neurochem. 64, 2262—2272.
Mattson M. P. (1990) Antigenic changes similar to those seen inneurofibrillary tangles are elicited by glutamate and calciuminflux in cultured hippocampal neurons. Neuron 4, 105—117.
McGeer E. G. and McGeer P. L. (1976) Duplication of biochemicalchanges of Huntington’s chorea by intrastriatal injections ofglutamic and kainic acids. Nature 263, 517—518.
Nagahara A. H., Nicolle M. M., and Gallagher M. (1993) Alterationsin [
3H]-kainate binding in the hippocampal formation of agedLong—Evans rats. Hippocampus 3, 269—277.
Najlerahim A., Francis P. T., and Bowen D. M. (1990) Age-relatedalteration in excitatory amino acid neurotransmission in ratbrain. Neurobiol. Aging 11, 155—158.
Nicholls D. and Attwell D. (1990) The release and uptake of excit-atory amino acids. Trends Pharmacol. Sci. 11, 462—468.
Palmer A. M., Robichaud P. J., and Reiter C. T. (1994) The releaseand uptake ofexcitatory amino acids in rat brain: effect of agingand oxidative stress. Neurobiol. Aging 15, 103—111.
Paxinos G. and Watson C. (1986) The Rat Brain in StereotaxicCoordinates. Academic Press, Sidney.
Pines G., Danbolt N. C., Bjørás M., Zhang Y., Bendahan A., EideL., Koepsell H., Storm-Mathisen J., Seeberg E., and KannerB. I. (1992) Cloning and expression of a rat brain L-glutamatetransporter. Nature 360, 464—467.
Price M. T., Olney J. W., and Haft R. (1981) Age-related changesin glutamate concentration and synaptosornal glutamate uptakein young rat striaturn. Life Sci. 28, 1365—1370.
Robinson M. B., Hunter-Ensor M., and Sinor J. (1991) Pharmaco-logically distinct sodium-dependent L- [3HI glutamate transportprocesses in rat brain. Brain Res. 44, 196—202.
Rothstein J. D., Dykes-Hoberg M., Pardo C. A., Bristol L. A., JinL., Kunc R. W., Kanai Y., Hediger M. A., Wang Y., Schielke
J. Neurochem., Vol. 69, No. 3, 1997

1160 L MASSIEU AND R. TAPIA
J. P., and Welty D. F. (1996) Knockout of glutamate transport-ers reveals a major role for astroglial transport in excitotoxicityand clearance of glutamate. Neuron 16, 675—686.
Salazar P., Montiel T., Brailowsky S., and Tapia R. (1994) Decreaseof glutamate decarboxylase activity after in vivo cortical infu-sion of y-aminobutyric acid. Neurochem. mt. 24, 363—368.
Sandberg M., Butcher S. P., and Hagberg M. (1986) Extracellularoverflow of neuroactive amino acids during severe insulin-in-duced hypoglycemia: in vivo dialysis of the rat hippocampus.J. Neurochem. 47, 178—184.
Sarantis M., Ballerini L., Miller B., Silver R. A., Edwards M., andAttwell D. (1993) Glutamate uptake from the synaptic cleftdoes not shape the decay of the non-NMDA component of thesynaptic current. Neuron 11, 541—549.
Schulz J. B., Henshaw D. R., Jenkins B. G., Ferrante R. J., KowallN. W., Rosen B. R., and Beal M. F. (1994) 3-Acetylpyridineproduces age-dependent excitotoxic lesions in rat striatum. J.Cereb. Blood Flow Metab. 14, 1024—1029.
Schwarcz R., Whetsell W. 0., and Mangano R. M. (1983) Quino-linic acid: an endogenous metabolite that produces axon-sparinglesions in rat brain. Science 219, 316—318.
Simon R. P., Swan J. H., Griffith T., and Meldrum B. 5. (1984)Blockade ofN-methyl-D-aspartate receptors may protect againstischernic damage in the brain. Science 226, 850—852.
Simpson J. R. and Isacson 0. (1993) Mitochondrial impairmentreduces the threshold for in vivo NMDA-mediated neuronaldeath in the striatum. Exp. Neurol. 121, 57—64.
Smith C. C. T., Bowen D. M., and Davison A. N. (1983) The evokedrelease of endogenous amino acids from tissue prisms ofhumanneocortex. Brain Res. 269, 103—109.
StorckT., Schulte S., Hofmann K., and Stoffel W. (1992) Structure,expression, and functional analysis of a Na + -dependent gluta-mate/aspartate transporter from rat brain. Proc. Nati. Acad. Sci.USA 89, 10955—10959.
Strong R., Sarnorajski T., and Gottesfeld Z. (1984) High-affinityuptake of neurotransmitters in rat neostriatum: effects of aging.J. Neurochem. 43, 1766—1768.
Swan J. and Meldrum B. S. (1990) Protection by NMDA antagonistsagainst selective cell loss following transient ischaemia. J.Cereb. Blood Flow Metab. 10, 343—351.
Teitelbaum J. S., Zatorre R. 1., Carpenter S., Gendron D., EvansA. C., Gjeddes A., and Cashman N. R. (1990) Neurologic se-quelae of domoic acid intoxication due to ingestion of contami-nated mussels. N. EngI. J. Med. 322, 1781—1787.
Wozniak D. F., Stewart G. R., Miller J. P., and Olney J. W. (1991)Age-related sensitivity to kainate neurotoxicity. Exp. Neural.114, 250—253.
I. Neurochem., Vol. 69, No. 3. /997




















