
Glutamate receptor metabotropic 7 is cis-regulated in the mouse brain and modulates alcohol drinking
13
Glutamate receptor metabotropic 7 is cis-regulated in the mouse brain and modulates alcohol drinking ☆ Csaba Vadasz a,b, ⁎ , Mariko Saito a,b , Beatrix M. Gyetvai b , Melinda Oros b,c , Istvan Szakall b,d , Krisztina M. Kovacs b , Vidudala V.T.S. Prasad b,e , Reka Toth b a Department of Psychiatry, New York University School of Medicine, New York, NY 10016, USA b Laboratory of Neurobehavioral Genetics, Nathan S. Kline Institute for Psychiatric Research, 140 Old Orangeburg Road, Orangeburg, NY 10962, USA c Department of Biochemistry, Debrecen University School of Medicine, Debrecen, Hungary d Department of Animal Health, Bayer Hungaria Kft., Alkotas u.50, Budapest, Hungary e Neurobiology Research Laboratory, VA Medical Center, 4801 E. Linwood Boulevard, Kansas City, MO 64128-2226, USA Received 2 May 2007; accepted 14 August 2007 Available online 23 October 2007 Abstract Alcoholism is a heritable disease that afflicts about 8% of the adult population. Its development and symptoms, such as craving, loss of control, physical dependence, and tolerance, have been linked to changes in mesolimbic, mesocortical neurotransmitter systems utilizing biogenic amines, GABA, and glutamate. Identification of genes predisposing to alcoholism, or to alcohol-related behaviors in animal models, has been elusive because of variable interactions of multiple genes with relatively small individual effect size and sensitivity of the predisposing genotype to lifestyle and environmental factors. Here, using near-isogenic advanced animal models with reduced genetic background interactions, we integrate gene mapping and gene mRNA expression data in segregating and congenic mice and identify glutamate receptor metabotropic 7 (Grm7) as a cis- regulated gene for alcohol consumption. Traditionally, the mesoaccumbal dopamine reward hypothesis of addiction and the role of the ionotropic glutamate receptors have been emphasized. Our results lend support to an emerging direction of research on the role of metabotropic glutamate receptors in alcoholism and drug addiction. These data suggest for the first time that Grm7 is a risk factor for alcohol drinking and a new target in addiction therapy. © 2007 Elsevier Inc. All rights reserved. Keywords: QTG; QTL; Recombinant QTL introgression; RQI; Genetics; Gene identification; Alcohol drinking; Alcoholism; Psychiatric disorders Genetic predisposition is a significant factor in the expression of most of the common diseases, such as high blood pressure, diabetes, cancer, depression, alcoholism, and Alzheimer disease. Although from a societal point of view common diseases are in the focus, in most common diseases progress in identifying genes has been limited to a small percentage of individuals with fully penetrant genes of large effect. Therefore, in general, most of the research efforts have focused on the biological effects of these major genes. The vast majority of a population with a common disease is character- ized by a complex phenotype with the involvement of a network of multiple interacting genes, each with individually small effect size against a background of variable environ- mental influences. Despite intensive research efforts, progress in identifying these minor genes has been slow. In alcoholism, even in animal models of alcohol-related behaviors [1–5], the contribution of defined genetic factors seems certain in only very few cases. Most of the preclinical gene mapping studies with animal models have been based on recombinant inbred (RI) or Mendelian segregating population designs. In RI or Mendelian designs in each strain or in each individual, respectively, the Available online at www.sciencedirect.com Genomics 90 (2007) 690 – 702 www.elsevier.com/locate/ygeno ☆ Microarray gene expression data have been deposited with the GEO repository under Series Entry GSE7155. ⁎ Corresponding author. Laboratory of Neurobehavioral Genetics, Nathan S. Kline Institute for Psychiatric Research, New York University School of Medicine, 140 Old Orangeburg Road, Orangeburg, NY 10962, USA. Fax: +1 845 398 5531. E-mail address: [email protected] (C. Vadasz). 0888-7543/$ - see front matter © 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.ygeno.2007.08.006
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Glutamate receptor metabotropic 7 is cis-regulated in the mouse brain and modulates alcohol drinking

Available online at www.sciencedirect.com
7) 690–702www.elsevier.com/locate/ygeno
Genomics 90 (200
Glutamate receptor metabotropic 7 is cis-regulated in the mouse brainand modulates alcohol drinking☆
Csaba Vadasz a,b,⁎, Mariko Saito a,b, Beatrix M. Gyetvai b, Melinda Oros b,c, Istvan Szakall b,d,Krisztina M. Kovacs b, Vidudala V.T.S. Prasad b,e, Reka Toth b
a Department of Psychiatry, New York University School of Medicine, New York, NY 10016, USAb Laboratory of Neurobehavioral Genetics, Nathan S. Kline Institute for Psychiatric Research, 140 Old Orangeburg Road, Orangeburg, NY 10962, USA
c Department of Biochemistry, Debrecen University School of Medicine, Debrecen, Hungaryd Department of Animal Health, Bayer Hungaria Kft., Alkotas u.50, Budapest, Hungary
e Neurobiology Research Laboratory, VA Medical Center, 4801 E. Linwood Boulevard, Kansas City, MO 64128-2226, USA
Received 2 May 2007; accepted 14 August 2007Available online 23 October 2007
Abstract
Alcoholism is a heritable disease that afflicts about 8% of the adult population. Its development and symptoms, such as craving, loss of control,physical dependence, and tolerance, have been linked to changes in mesolimbic, mesocortical neurotransmitter systems utilizing biogenic amines,GABA, and glutamate. Identification of genes predisposing to alcoholism, or to alcohol-related behaviors in animal models, has been elusivebecause of variable interactions of multiple genes with relatively small individual effect size and sensitivity of the predisposing genotype to lifestyleand environmental factors. Here, using near-isogenic advanced animal models with reduced genetic background interactions, we integrate genemapping and gene mRNA expression data in segregating and congenic mice and identify glutamate receptor metabotropic 7 (Grm7) as a cis-regulated gene for alcohol consumption. Traditionally, the mesoaccumbal dopamine reward hypothesis of addiction and the role of the ionotropicglutamate receptors have been emphasized. Our results lend support to an emerging direction of research on the role of metabotropic glutamatereceptors in alcoholism and drug addiction. These data suggest for the first time that Grm7 is a risk factor for alcohol drinking and a new target inaddiction therapy.© 2007 Elsevier Inc. All rights reserved.
Keywords: QTG; QTL; Recombinant QTL introgression; RQI; Genetics; Gene identification; Alcohol drinking; Alcoholism; Psychiatric disorders
Genetic predisposition is a significant factor in theexpression of most of the common diseases, such as highblood pressure, diabetes, cancer, depression, alcoholism, andAlzheimer disease. Although from a societal point of viewcommon diseases are in the focus, in most common diseasesprogress in identifying genes has been limited to a smallpercentage of individuals with fully penetrant genes of large
☆ Microarray gene expression data have been deposited with the GEOrepository under Series Entry GSE7155.⁎ Corresponding author. Laboratory of Neurobehavioral Genetics, Nathan S.
Kline Institute for Psychiatric Research, New York University School ofMedicine, 140 Old Orangeburg Road, Orangeburg, NY 10962, USA. Fax: +1845 398 5531.
E-mail address: [email protected] (C. Vadasz).
0888-7543/$ - see front matter © 2007 Elsevier Inc. All rights reserved.doi:10.1016/j.ygeno.2007.08.006
effect. Therefore, in general, most of the research efforts havefocused on the biological effects of these major genes. The vastmajority of a population with a common disease is character-ized by a complex phenotype with the involvement of anetwork of multiple interacting genes, each with individuallysmall effect size against a background of variable environ-mental influences. Despite intensive research efforts, progressin identifying these minor genes has been slow. In alcoholism,even in animal models of alcohol-related behaviors [1–5], thecontribution of defined genetic factors seems certain in onlyvery few cases.
Most of the preclinical gene mapping studies with animalmodels have been based on recombinant inbred (RI) orMendelian segregating population designs. In RI or Mendeliandesigns in each strain or in each individual, respectively, the

691C. Vadasz et al. / Genomics 90 (2007) 690–702
genetic background is unique and represents a specific source ofgenetic interaction. This interaction between the target gene andthe background may substantially contribute to the total geneticvariance and may limit detection of polygenes with small effectsize reliably. Recently, to reduce genetic background noise, wedeveloped a different design with 78 near-isogenic inbred strainsof N97% C57BL/6ByJ (B6By) genetic background identity andmapped six quantitative trait loci (QTLs) for alcohol consump-tion [6,7]. Given this and the availability of near-isogenic strainsas convenient precursors for rapid development of congenicstrains, we tested different genetic designs to confirm linkagebetween alcohol consumption and one of the QTLs, ethylalcohol consumption-2 locus (Eac2) mapped to distal Chr. 6, andused bioinformatic methods to examine the Eac2 region in asearch for the underlying quantitative trait gene (QTG).
Here, we report an integrated genome and transcriptomeanalysis of the Eac2 region on Chr. 6, which suggests that a genecoding for metabotropic glutamate receptor subtype-7 (Grm7) isa candidate QTG for Eac2.
Results
Alcohol consumption in a congenic strain
Using microsatellite marker-assisted selection and repeatedbackcrosses to the alcohol-preferring B6By background strain,we developed a new congenic strain, B6By.C.6.132.54 (B6By.C6). This congenic strain carries a BALB/cJ (C) donor chromo-
Fig. 1. Alcohol consumption in inbred congenic mice and segregating quasi-congeniccomposition in C57BL/6ByJ (background strain, black chromosomes), BALB/cJ (ddonor chromosome segments), and B6By.C6 (B6By.C.6.132.54, a congenic strain dwere the only chromosomes that harbored differential donor segments in C5A3. In a pwas significantly lower than that in the background C57BL/6ByJ (F1,13=6.21, p=0genotyped reciprocal F2 population derived from respectively alcohol-preferring andindependently derived quasi-congenic strains on the B6By background. C5B3 carriesof C5A3 and B6.C6 (see details under Methods in the supporting information). F2alcohol than mice with the T:T (BALB/cJ/BALB/cJ) genotype (independent samples36) is in intron7 of Grm7. G:G has been detected in alcohol-preferring C57BL/6J aBALB/cJ, BALB/cByJ, DBA/2J, and A/J (http://www.jax.org/phenome).
some segment on Chr. 6 with proximal and distal backgroundmarkers D6Mit275 (25.5 cM, 51.1 Mb, from UniSTS annota-tion of NCBI build 36) and D6Mit134 (57.5 cM, 125.3 Mb),respectively. The congenic strain in a two-bottle free-choicealcohol preference test demonstrated significantly lower con-sumption of 12% (v/v) alcohol in comparison to its backgroundpartner B6By, confirming a significant decreasing effect of thegenetic factor(s) residing on the donor segment (Fig. 1A). In anadditional independent test, using a two-bottle choice para-digm with intermittent access to alcohol, we found that con-sumption of 12% alcohol was significantly higher among B6Bymales (12.70±1.03, n=13) in comparison with B6By.C6 males[8.37±0.96 (g/kg/day, mean±SE), n=19, average of four 3-daytrials, t30=3.00, p=0.005, two-tailed].
To test the hypothesis that other undetected donor chromosomesegments are responsible for this effect, a genome scan with amouse single-nucleotide polymorphism (SNP) panel of 402markers was carried out. No additional donor segments weredetected (Fig. S1). Because the genome scan showed flankingbackground markers at 75.9 Mb (rs4226008; NCBI mouse build36/dbSNP build 126) and 122.3 Mb (rs3023093), and limitingdonor markers at 81.9 (rs4226024) and at 91.8 Mb (rs3712161),we concluded the segment size must be between 9.9 and 46.4Mb.
Exclusion of identical-by-descent regions
For narrowing the set of candidate genes, first, the donorsegment in the B6By.C6 congenic strain was subjected to
population. (A) Alcohol consumption (g/kg/day; mean±1 SE) and chromosomeonor strain, white chromosomes), C5A3 (a quasi-congenic RQI strain carryingerived from C5A3). Only chromosomes 1, 5, 6, and 7 are shown because thesereplanned comparison, alcohol consumption among the congenic B6By.C6 mice.028). (B) SNP allelic effects on oral alcohol self-administration in a selectivelyalcohol-nonpreferring inbred strains I5B25A and C5B3. I5B25A and C5B3 area BALB/cJ-derived chromosome segment that overlaps with the Chr. 6 segmentsindividuals with the G:G (B6By/B6By) genotype consumed significantly moret test, t27=2.1, p=0.048). SNP rs3723352 (Chr. 6: 111.318602 Mb; NCBI buildnd C57BL/6ByJ strains, while T:T has been detected in alcohol-nonpreferring
692 C. Vadasz et al. / Genomics 90 (2007) 690–702
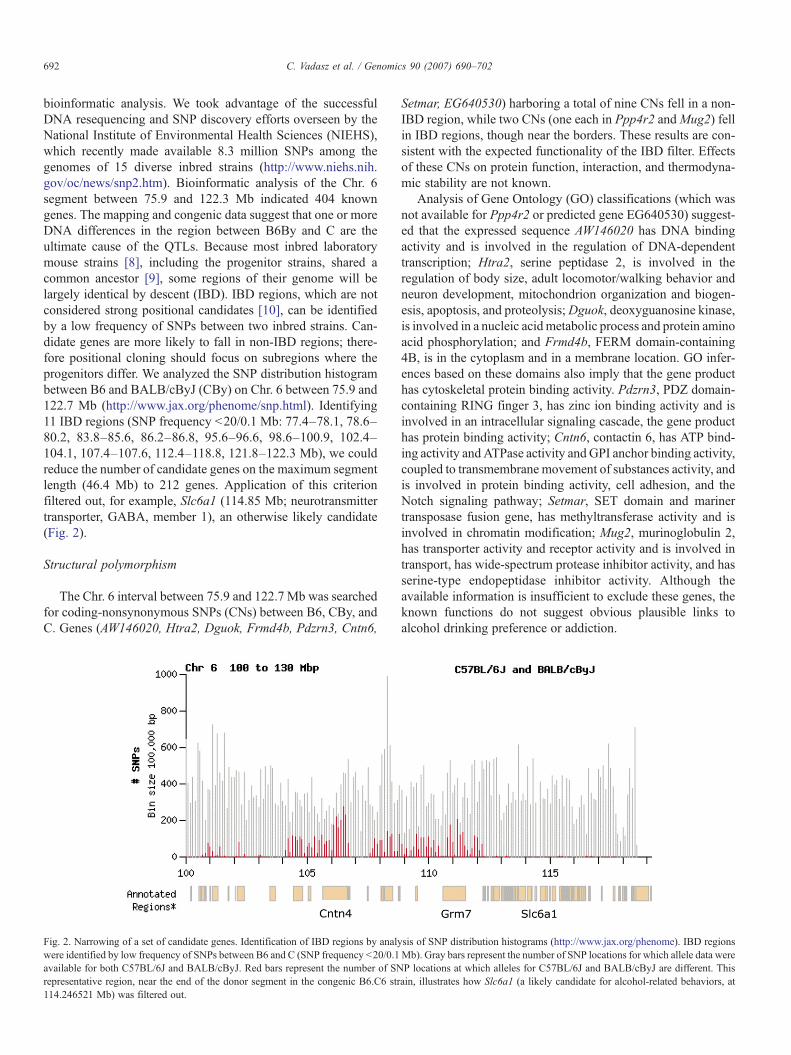
bioinformatic analysis. We took advantage of the successfulDNA resequencing and SNP discovery efforts overseen by theNational Institute of Environmental Health Sciences (NIEHS),which recently made available 8.3 million SNPs among thegenomes of 15 diverse inbred strains (http://www.niehs.nih.gov/oc/news/snp2.htm). Bioinformatic analysis of the Chr. 6segment between 75.9 and 122.3 Mb indicated 404 knowngenes. The mapping and congenic data suggest that one or moreDNA differences in the region between B6By and C are theultimate cause of the QTLs. Because most inbred laboratorymouse strains [8], including the progenitor strains, shared acommon ancestor [9], some regions of their genome will belargely identical by descent (IBD). IBD regions, which are notconsidered strong positional candidates [10], can be identifiedby a low frequency of SNPs between two inbred strains. Can-didate genes are more likely to fall in non-IBD regions; there-fore positional cloning should focus on subregions where theprogenitors differ. We analyzed the SNP distribution histogrambetween B6 and BALB/cByJ (CBy) on Chr. 6 between 75.9 and122.7 Mb (http://www.jax.org/phenome/snp.html). Identifying11 IBD regions (SNP frequency b20/0.1 Mb: 77.4–78.1, 78.6–80.2, 83.8–85.6, 86.2–86.8, 95.6–96.6, 98.6–100.9, 102.4–104.1, 107.4–107.6, 112.4–118.8, 121.8–122.3 Mb), we couldreduce the number of candidate genes on the maximum segmentlength (46.4 Mb) to 212 genes. Application of this criterionfiltered out, for example, Slc6a1 (114.85 Mb; neurotransmittertransporter, GABA, member 1), an otherwise likely candidate(Fig. 2).
Structural polymorphism
The Chr. 6 interval between 75.9 and 122.7 Mb was searchedfor coding-nonsynonymous SNPs (CNs) between B6, CBy, andC. Genes (AW146020, Htra2, Dguok, Frmd4b, Pdzrn3, Cntn6,
Fig. 2. Narrowing of a set of candidate genes. Identification of IBD regions by analywere identified by low frequency of SNPs between B6 and C (SNP frequency b20/0.1available for both C57BL/6J and BALB/cByJ. Red bars represent the number of SNrepresentative region, near the end of the donor segment in the congenic B6.C6 str114.246521 Mb) was filtered out.
Setmar, EG640530) harboring a total of nine CNs fell in a non-IBD region, while two CNs (one each in Ppp4r2 andMug2) fellin IBD regions, though near the borders. These results are con-sistent with the expected functionality of the IBD filter. Effectsof these CNs on protein function, interaction, and thermodyna-mic stability are not known.
Analysis of Gene Ontology (GO) classifications (which wasnot available for Ppp4r2 or predicted gene EG640530) suggest-ed that the expressed sequence AW146020 has DNA bindingactivity and is involved in the regulation of DNA-dependenttranscription; Htra2, serine peptidase 2, is involved in theregulation of body size, adult locomotor/walking behavior andneuron development, mitochondrion organization and biogen-esis, apoptosis, and proteolysis;Dguok, deoxyguanosine kinase,is involved in a nucleic acidmetabolic process and protein aminoacid phosphorylation; and Frmd4b, FERM domain-containing4B, is in the cytoplasm and in a membrane location. GO infer-ences based on these domains also imply that the gene producthas cytoskeletal protein binding activity. Pdzrn3, PDZ domain-containing RING finger 3, has zinc ion binding activity and isinvolved in an intracellular signaling cascade, the gene producthas protein binding activity; Cntn6, contactin 6, has ATP bind-ing activity andATPase activity andGPI anchor binding activity,coupled to transmembrane movement of substances activity, andis involved in protein binding activity, cell adhesion, and theNotch signaling pathway; Setmar, SET domain and marinertransposase fusion gene, has methyltransferase activity and isinvolved in chromatin modification; Mug2, murinoglobulin 2,has transporter activity and receptor activity and is involved intransport, has wide-spectrum protease inhibitor activity, and hasserine-type endopeptidase inhibitor activity. Although theavailable information is insufficient to exclude these genes, theknown functions do not suggest obvious plausible links toalcohol drinking preference or addiction.
sis of SNP distribution histograms (http://www.jax.org/phenome). IBD regionsMb). Gray bars represent the number of SNP locations for which allele data wereP locations at which alleles for C57BL/6J and BALB/cByJ are different. Thisain, illustrates how Slc6a1 (a likely candidate for alcohol-related behaviors, at

693C. Vadasz et al. / Genomics 90 (2007) 690–702
Cis-regulated genes
In several recent studies it has been recommended that a setof candidate genes can be narrowed by requiring the gene to beexpressed, to be correlated with the phenotype of interest, and tohave cis-regulated expression [11–17]. To filter the remaining212 genes for expression and genotype-dependent transcriptabundance we analyzed gene expression data obtained from theCXB RI strain set. Gene expression QTL mapping of the CXBRI strains is highly informative for our study because the pro-genitors of the RI and RQI strains are identical (C57BL/6By) orclosely related (BALB/cBy vs BALB/cJ). Interval QTL map-ping of mRNA abundance of the 212 genes was implementedusing the Hippocampus Consortium M430v2 CXB (Dec05)Robust Multiarray Average (RMA) database (http://www.genenetwork.org/public/WebQTL.py). The lod (logarithm ofthe odds to the base 10) scores for expression of the 212 geneswere tested for genome-wide significance (1000 permutations[18]). The criterion of cis-regulation was the identification of asingle peak of the transcript abundance QTL with genome-widesignificance (pb0.05) and a peak position identical or near tothe genomic location of the gene. Only 7 genes had genome-wide statistically significant lod scores: Lrrtm4, Sema4f, Aak1,Nfu1 (aka Hirip5), Chchd4, Grm7, and Cecr5 (Table 1). Peaksof the transcript abundance QTLs for these genes fell into binsthat included their genomic locations, indicating the possibilityof cis-regulation of these genes. In the CXB hippocampal dataset the B6 alleles increased gene expression for Lrrtm4, Sema4f,Aak1, Nfu1 (aka Hirip5), and Chchd4, while for Grm7 andCecr5 the C alleles increased expression (Table 1).
Polymorphism in cis-regulated genes
Focusing on the seven candidate genes, in a search forDNA polymorphisms in NCBI dbSNP build 126 (http://www.informatics.jax.org) between C57BL/6J (B6; reference strain)and BALB/cByJ, BALB/cJ, or BALB/c within coordinates ofthe candidates we found 67 matching SNPs for Lrrtm4, 0 forSema4f, 47 for Aak1 (6 mRNA-UTR, 1 coding-synonymous,40 intron), 11 for Nfu1 (6 mRNA-UTR, 5 intron), 14 for Chchd4
Table 1Cis-regulated genes in donor chromosome segment
Chr. cM Genome coordinates,NCBI build 36 (strand)
Symbol, name
6 Syntenic 79948557–80738698 (+) Lrrtm4, leucine-rich repea6 35 82877544–82905386 (−) Sema4f, sema domain, im
TM domain, and short cy6 35.5 86815474–86957498 (+) Aak1, AP2-associated kin6 86975523–86994094 (+) Nfu1, NFU1 iron–sulfur c
(Saccharomyces cerevisia6 Syntenic 91429780–91438927 (−) Chchd4, coiled-coil–helix
domain-containing 46 Syntenic 110611375–111533007 (+) Grm7, glutamate receptor6 54 120475114–120496924 (−) Cecr5, cat eye syndrome
candidate 5 homolog (hum
a Affymetrix oligonucleotide microarray, Mouse Genome 430 2.0, Hippocampus* pb0.05, genome-wide significance test (1000 permutations).
(2 mRNA-UTR and 12 intron), 218 for Grm7 (intron), and 0 forCecr5.
Gene expression differences in congenic and backgroundstrains
It has been suggested that by comparing gene expression instrains with well-defined small genomic differences cis-regulation of expression can be detected [7,19,20]. To test theexpected congenic vs background differences in expression, in amicroarray gene expression experiment we compared the sevencis-acting genes for differences in relative brain mRNA abun-dance between the background B6By, the congenic B6By.C6,and the donor C. As shown in Table 2, expression of Lrrtm4,Sema4f, Aak1, and Cecr5 was not significantly different amongthe strains, while significant genetic variation was detected forthree genes: Nfu1, Chchd4, and Grm7 (p=0.000–0.036; one-way ANOVA, SPSS version 13). For Nfu1, Chchd4, and Grm7significantly different expression was detected between thebackground B6By and the congenic B6By.C6 after correctionfor multiple comparisons (Table 2). The direction of the alleliceffects for whole brain transcript abundance between the pro-genitor and the congenic strains agreed with the results of theHippocampus Consortium M430v2 CXB (Dec05) RMA data-base analysis for most of the candidate genes.
Multistrain analysis
To see if any of these genes showed genotype–alcohol con-sumption correlation across commercially available commoninbred strains with publicly accessible data we estimated thecorrelation between genotypes of the four significant (Nfu1,Chchd4, andGrm7) or near-significant (Aak1) SNPs and alcoholconsumption in strains B6, AKR/J, C, A/J, DBA/2J, 129/SvJ,and C3H/J using previously published alcohol consumption data[21–23]. We found that alcohol consumption was highly cor-related with genotypes of Nfu1 (rs30605208, n=6, r=−0.96,p=0.001) and Aak1 (rs30912462, n=4, r=−0.95, p=0.02).Relatively high correlation was found for Grm7 (rs305571150,n=8, r=−0.57), which did not quite reach the p=0.05 level of
Probe set a Lod score B6 alleleeffect
t transmembrane neuronal 4 1455937_at 5.5* Increasemunoglobulin domain,toplasmic domain
1419328_at 4.3* Increase
ase 1 1435038_s_at 7.5* Increaseluster scaffold homologe), aka Hirip5
1418229_s_at 6.6* Increase
–coiled-coil–helix 1417233_at 11.5* Increase
, metabotropic 7 1459532_at 6* Decreasechromosome region,an)
1424816_at 4.3* Decrease
Consortium M430v2 CXB RMA data set.

Table 2Cerebral gene expression differences between congenic and progenitor strains
Gene on donorsegment
Probe set ID Strain Mean Standarddeviation
Standarderror
df F Significancea TukeyHSDb
Lrrtm4 1455937_at B6By 11.41 1.13 0.65 2 1.91 0.229 0.313BALB/cJ 9.76 0.63 0.36B6By.C6 9.9 1.5 0.87
Sema4f 1419328_at B6By 52.3 3.15 1.82 2 3.05 0.122 0.106BALB/cJ 48.08 1.98 1.14B6By.C6 44.12 5.97 3.44
Aak1 1435038_s_at B6By 357.46 55.46 32.02 2 3.25 0.111 0.24BALB/cJ 272 22.59 13.04B6By.C6 293.84 43.37 25.04
Nfu1 1418229_s_at B6By 138.17 5.47 3.16 2 10.09 0.012 0.036BALB/cJ 98.09 12.88 7.44B6By.C6 106.83 14.15 8.17
Chchd4 1417233_at B6By 171.16 21.76 12.56 2 39.62 0 0.001BALB/cJ 76.72 6.73 3.89B6By.C6 84.33 10.33 5.96
Grm7 1459532_at B6By 6.74 0.7 0.4 2 6.11 0.036 0.049BALB/cJ 9.34 1.52 0.88B6By.C6 9.46 0.83 0.48
Cecr5 1424816_at B6By 29.81 1.81 1.04 2 1.08 0.398 0.848BALB/cJ 31.29 1.15 0.66B6By.C6 28.91 2.74 1.58
Affymetrix oligonucleotide microarray, Mouse Genome 430 2.0. Vadasz–Saito Cerebral Gene Expression database, GEO repository series entry GSE7155.a Between-group difference, one-way ANOVA of RMA normalized data.b Post hoc test for B6By vs B6By.C6.
694 C. Vadasz et al. / Genomics 90 (2007) 690–702
significance (p=0.07). Chchd4 showed the lowest correlation(rs30014728, n=6, r=−0.27, p=0.3). Evaluating the results ofmultistrain analysis, we have to keep in mind its limitations insupport of QTG candidacy. In our view, it can be only tenta-tively useful in either supporting or refuting QTG candidacy ofa gene.
GO annotation summary of the candidate genes indicatedthat while the biological process in which Nfu1 is involved isnot known, it may have a housekeeping function; Aak1 may beinvolved in protein amino acid phosphorylation and endocy-tosis; Chchd4 appears to be involved in mitochondrial proteintransport; and Grm7 has been intensively characterized as a G-protein-coupled receptor protein that is involved in synaptictransmission, behavioral fear response, sensory perception, andstress (Table S3).
Promoter analysis
Analysis of the genotype–alcohol consumption correlationand the GO annotation summaries left Grm7 as the primarycandidate for testing the hypothesis that genetic variation inexpression is associated with a mutation in a transcription factorbinding site (TFBS). Analysis of Grm7 by the PromoterInspec-tor program predicted 13 promoters and two regulatory regions(http://www.genomatix.de). PromoterInspector is a programthat predicts eukaryotic Pol II promoter regions with about 85%specificity in mammalian genomic sequences. Accordingly, onemay expect PromoterInspector to find good matches but not allpromoters.
Comparing the positions of the 218 SNPs detected in Grm7we found 1 SNP (rs30557150) in a promoter region (fifth pro-
moter, 110,819,264–110,819,864 bp, Fig. 3). MatInspector, aprogram that detects TFBSs in nucleotide sequences using alarge library of weight matrices, identified a TFBS above a goodmatch threshold: V$SORY/SOX5.01 (matrix similarity=0.987;core similarity=1.0) (http://www.genomatix.de). The TFBSwas lost due to SNP rs30557150 (C/T) in the rs30557150_Callele of Grm7, which is consistent with the observation thatC57BL/6J carrying the rs30557150_C allele showed lowertranscript abundance of Grm7 than strains with thers30557150_T allele (Fig. 3). However, experimental evidenceis required to provide sufficient proof for the involvement ofthis TFBS in transcriptional function. In a test of differentmicroarray gene expression probe sets in WebQTL (http://www.genenetwork.org), results of eQTL mapping suggested cis-regulation for Sox5 in the mouse hippocampus in both BXD andCXB RI strain sets. Whole genome mapping with 1000permutation test yielded LOD=31.56 with peak locationbetween 144.01 and 144.82 Mb in the BXD population(database: Hippocampus Consortium M430v2 (Jun06) PDNN;Trait ID: 1446461_at); however, no significant peak was foundfor another probe set (1432189_a_at). No significant peak wasfound for 1446461_at in the Hippocampus ConsortiumM430v2CXB (Dec05) PDNN database. Whole genome mapping with1000 permutation test yielded LOD=3.0 for probe set1432189_a_at, but no significant peak was found for probeset 1446461_at, in the Hippocampus Consortium M430v2 CXB(Dec05) RMA database. We found 362 SNPs in Sox5 in acomparison of B6, DBA/2J, and BALB/cBy, including anmRNA-UTR insertion (rs6151967, http://www.informatics.jax.org), which is consistent with the hypothesis that probe setdifferences may reflect heterogeneity.

Fig. 3. Reference flanking sequence of SNP rs30557150. Cis-regulated Grm7 is polymorphic (C/T) in a regulatory region among mouse strains. rs30557150 (Chr. 6:110,819,852 inGrm7) is a putative promoter SNP. Strains with higher transcript abundance (BALB/cJ, BALB/cByJ, DBA/2J, A/J; Hippocampus ConsortiumM430v2CXB (Dec05) RMA data set at http://www.genenetwork.org; ProbeSetID=1459532) carry the “T” allele, while strains with lower transcript abundance (C57BL/6ByJ,C57BL/6J, AKR/J, NOD/LtJ; Hippocampus Consortium M430v2 BXD (Dec05) PDNN; ProbeSetID=1459532) carry the “C” allele.
695C. Vadasz et al. / Genomics 90 (2007) 690–702
No significant “genetic” (between-strain) correlation wasfound between hippocampal expression of Sox5 (1432189_a_at)and of Grm7 (1459532_at) in the CXB population (Pearson’scorrelation: r=0.498, n=15, p=0.059, two-tailed) or betweenhippocampal expression of Sox5 (1446461_at) and Grm7(1459532_at) in the BXD population (Pearson’s correlation:r=−0.178, n=69, p=0.147, two-tailed).
Confirmatory analysis of cis-regulation of Grm7
Although the hippocampus has been implicated inaddiction [24] and alcohol consumption [4], traditionally, ithas not been considered as a region critically involved inalcohol preference and dependence. Therefore, we performedadditional expression QTL mapping on two striatal data-bases created for BXD and B6D2F2 populations (seeMaterials and methods). To test the between-strain general-izability of the CXB hippocampal gene expression results,an additional BXD hippocampal data set was analyzed. Asshown in Fig. 4, the results of the confirmatory analysesprovided strong support for cis-regulation of Grm7 in bothbrain regions.
Confirmatory analysis of linkage in reciprocal F2 populations
We used an alternative genetic design for testing the asso-ciation between Grm7 and alcohol consumption. Reciprocal F2populations were created by crossing F1 hybrids of alcohol-
preferring (I5B25A) and alcohol-nonpreferring (C5B3) quasi-congenic RQI strains, which are on the same genetic back-ground (both strains carry N97% B6 genome [6]). F2 mice wereselectively (N1 SD above or below mean alcohol consumption)genotyped for a SNP marker (rs3723352, G/T) that resides inintron 7 of Grm7 at Chr. 6: 111,318,602 (NCBI build 36). Be-cause consumption of 12% (v/v) alcohol (g/kg/day, average offour 3-day trials) was not significantly different between theselectively genotyped reciprocal F2 groups (F1,56=0.73, p=0.79), data were combined. Comparison of the B6By/B6By (G:G) and BALB/cJ/BALB/cJ (T:T) homozygotes indicated thatindividuals with the B6By/B6By genotype consumed signifi-cantly more alcohol (independent samples t test, t27=2.1, p=0.048; Fig. 1B). These results were consistent with dataobtained from RQI QTL mapping [7], detection of T:Tgenotypes in other inbred strains (e.g., CByJ, 129/SvJ, DBA/2J) known to show low alcohol consumption, and results of thealcohol preference tests with B6By.C6 congenic animals (Fig.1A). Importantly, the quasi-congenic F2 experiments alsoprovided supportive evidence for the exclusion of competingcandidate genes Nfu1, Aak1, and Chchd4, because the alcohol-nonpreferring C5B3 quasi-congenic RQI strain did not carry theBALB/cJ-type alleles of Nfu1, Aak1, and Chchd4. To create theF2 on an ∼97% homogeneous background, RQI strain C5B3was chosen for the cross because it carries a BALB/cJ-derived Chr. 6 segment (with flanking background markersD6Mit230 at 98.97 Mb and D6Mit258 at 137.07 Mb), whichpartially overlaps the BALB/cJ-derived Chr. 6 segment in the

Fig. 4. Expression QTL mapping in four data sets. QTL mapping of mRNA abundance of candidate genes was carried out using WebQTL (http://www.genenetwork.org). Grm7 (ProbeSetID=1459532_at) had genome-wide statistically significant lod scores with peaks at (or near) its genomic location in the following data sets: CXB Hippocampus (LOD=6.0), BXD Striatum (LOD=7.3), B6D2F2 Striatum (LOD=21.1), BXDHippocampus (LOD=20.5). For data set details see Materials and methods.
696C.Vadasz
etal.
/Genom
ics90
(2007)690–702

697C. Vadasz et al. / Genomics 90 (2007) 690–702
congenic B6By.C6 (75.9–122.3 Mb, Fig. 1A). In thealcohol-preferring I5B25A quasi-congenic strain, BALB/cJ-derived alleles were detected only on Chrs. 8 and 13. InC5B3, the whole BALB/cJ-derived Chr. 6 segment waspositioned distal from the genomic positions of AW146020,Htra2, Dguok, and Frmd4b (genes identified with structuralpolymorphism) and Nfu1, Aak1, and Chchd4 (genesidentified as cis-regulated).
Discussion
In our recent studies composite interval mapping identifiedseveral significant linkages to ethyl alcohol consumption onmouse Chr. 6 and indicated that the RQI donor block region forEac2 spans 33.5–48.2 cM [7]. Eila2 (36.5 cM), Etlm3 (30 cM),Rear1 (36.5 cM), Bits2 (37 cM), and Taste5 (48 cM) arepreviously identified behavior and preference QTLs mappingto this interval. Interestingly, despite intensive efforts, mappingstudies on QTLs for murine alcohol preference drinkingbehavior using the most popular BXD recombinant inbredstrains and various genetic preparations derived from theprogenitor strains failed to identify QTLs in this chromosomeregion [1]. Reasons for the failure may include, but are notlimited to, the following: different mapping strategy (our quasi-congenic RQI strains have ∼97% “background homogeneity,”significantly reducing epistatic interaction variation, i.e.,genetic background noise, which can mask target allele signals[25–28]), different laboratories with differences in testingprocedures [29,30], and differences between DBA/2J andBALB/cJ in genomic architecture (different gene dispersionpatterns+different mode of gene action in each strain resultingin various increaser–decreaser linkages with different tightness,different interactions with the B6 genes and environmentalmodifiers, etc.). It is possible that the task of mapping andsequencing true polygenes has been underestimated [31]. Forexample, two polygenes with small effect size exhibitingantagonistic epistasis may be mapped to a single small QTLregion (b1 cM), such that their phenotypic effect depends onthe genetic background [32]. If similar complications aretypical for behavioral traits, it may explain the inability topredict results of new crosses based on RI or classicalintercross mapping results (cf. [33]).
In this study we integrated genetic, transcriptome, and bio-informatic approaches to identify quantitative trait genes for abehavioral phenotype, oral alcohol self-administration. Theresults suggest that Grm7, which encodes the group III meta-botropic glutamate receptor 7 (mGluR7), modulates alcoholdrinking: Mice carrying the genotype that predisposes for lowerexpression of Grm7 in the brain consume more alcohol in apreference drinking behavioral paradigm. Molecular, cellular,and biological functions of Grm7/mGluR7 strongly support thissuggestion.
Studies on human mGluR7 mRNA splicing variants have sofar revealed five isoforms [34,35] and suggest that structuraland functional diversity of this receptor is achieved byalternative exon usage affecting the immediate carboxy-terminus. This region interacts with defined proteins, such as
PICK [36], α-tubulin [37], and calmodulin [38], thus mediatingvarious receptor functions such as intracellular signal transduc-tion, axonal targeting, and synaptic clustering. Most variantsare expressed in brain and retina, while human isoformsGRM7_v3 and GRM7_v4 are expressed in nonneuronal tissues[35]. In rats, two splice variants (mGluR7a and mGluR7b)have been reported, which are defined by different C-termini[39]. Immunohistochemical studies using antibodies raisedagainst the C-termini of rat mGluR7a and human mGluR7bshowed similar, but not identical, distributions in the brains ofrats and mice [40]. Alternative splicing, in addition to probeambiguity (targeting multiple genes), heterogeneity (targetingdifferent transcript variants from one gene), and other factors,may explain our observation that only one of the fourAffymetrix Mouse 430 2.0 probe sets for Grm7 could detectcis-regulation [41].
Sox5 (SRY-box containing gene 5) is a novel SRY-relatedtranscription factor [42]. The genomic location of this TF (Chr.6, 69.5 cM, 143,790,051–144,166,639 bp) is just 33.178 Mbdistal from that of Grm7. One of its known functions is relatedto cell fate commitment and cartilage development [43]; how-ever, its functions in the brain or interactions with Grm7 are notknown, and its cis-regulated expression did not show a cor-relation with that of Grm7 in CXB or BXD strain sets (seeResults). Thus, the hypothesis that Sox5 may play a role as atranscription factor in the regulation of expression of Grm7 inthe brain needs further support.
mGluR7 is a presynaptic autoreceptor, or heteroreceptor, innonglutamatergic axon terminals, with an inhibitory action onthe release of various neurotransmitters such as glutamate,GABA, and monoamines [44]. There is a considerable overlapbetween the neuroanatomical distribution of mGluR7 and me-solimbic brain regions, suggesting that this receptor has a parti-cularly crucial role in processing information, which can modu-late neurotransmitter release in pathways involved in addictionand reinstatement. High representation of mGluR7 was ob-served in the amygdala [45], the periventricular zone of thehypothalamus, and the olfactory circuitry [46]. Simonyi et al.found the highest levels of mGluR7 expression in the dentategyrus, CA1, CA3, and piriform cortex [47]. Recently depositedmicroarray data indicated relatively high Grm7 expression inamygdala, hippocampus, and cerebellum in various mousestrains, including B6 and CByJ [48].
Genetic ablation of mGluR7 in mice provided furthersupportive data. Grm7 knockouts showed altered amygdala-dependent conditioned fear and aversion responses and reducedanxiety-related behaviors, suggesting a role for mGluR7 inthe modulation of stress-related behaviors [49,50]. Indeed, anew specific agonist of mGluR7 activated the hypothalamic–pituitary–adrenal (HPA) axis and dramatically increased stresshormone levels [51]. A possible glutamatergic mechanismunderlying the activation of the HPA axis, which may haverelevance to alcohol drinking, involves disinhibition of cortico-tropin-releasing factor (CRF)-containing paraventricular nuc-leus (PVN) neurons, which leads to increased ACTH and cor-ticosterone levels: Activation of the presynaptic inhibitorymGluR7 receptor can decrease L-glutamate release from the

698 C. Vadasz et al. / Genomics 90 (2007) 690–702
hippocampal–hypothalamic pathway, decreasing the excitatorytone on GABAergic neurons, or agonists may activate mGluR7heteroreceptors directly, decreasing the inhibitory tone on PVNneurons [50,52,53]. In stress-induced alcohol drinking and re-lapse behavior both the glutamate system and the corticotropin-releasing hormone system have been implicated [44,54,55]. Theproposed role for the amygdala in the relationship among stress,CRF, and alcohol abuse [56], and the role of Grm7 in stress, isconsistent with a link between the genetic variation in Grm7and predisposition to alcohol drinking. Our present observationof increased expression of Grm7 in BALB/cJ and earlier reportsproposing the BALB/cJ strain as an animal model for patholo-gical anxiety [57,58], characterized by increased “freezing” andemotionality in open-field test [59–61] and by elevated stress-induced corticosterone and emotionality [62], are also consis-tent with the anxiolytic effects of targeted disruption of Grm7[49].
Although the mesolimbic dopamine reward hypothesis ofaddiction has been traditionally important in psychostimulantuse and abuse [63–65], there is emerging evidence that gluta-mate systems play a critically important role in reinforcementand addiction [66]. It has been shown that glutamatergic inner-vation of the VTA and the nucleus accumbens regulates loco-motor activity [67]. mGluR7 may be implicated in regulation ofmotor activity, because administration of group III mGlureceptor agonist L-AP4 attenuates acute amphetamine [68]- andcocaine [69]-stimulated locomotor activity. In studies on thelink between alcohol effects and the glutamatergic neurotrans-mission the emphasis has been on the role of the ionotropicglutamate receptors; however, research on the role of metabo-tropic glutamate receptors in alcohol use is gaining impetus[70]. It has been shown that chronic exposure to alcohol reducedmGluR1 mRNA levels in cerebellar Purkinje cells in mice [71].In rats, chronic exposure to Lieber–DeCarli liquid diet withalcohol (5% w/v) decreased mRNA levels of mGluR3 andmGluR5 in dentate gyrus, while levels of mGluR1, mGluR5,and mGluR7 were decreased in the CA3 area of the hippo-campus [72]. Such adaptations in mGluR7 expression to long-term exposure to alcohol are also expected in excessive alcoholself-administration, and reduced mGluR7 expression maybecome a causative factor in excessive drinking, which is con-sistent with our results.
Use of pharmacological agents indicates that the mGluR5antagonist MPEP decreases relapse to alcohol self-administra-tion [70,73,74], blocks the discriminative stimulus effects ofalcohol [75], and attenuates latent inhibition in conditioned tasteaversion [76]. Based on the supporting psychopharmacologicaldata and its high expression in the nucleus accumbens, cortex,and hippocampus [77], it was proposed that mGluR5 receptorsmight specifically regulate ethanol reinforcement [70]. Recentexperiments indicated that the novel mGluR7-specific agonistAMN082 inhibited cocaine self-administration [78], andreverse dialysis studies demonstrated that group III agonist L-AP4, possibly acting on mGluR7, decreased in vivo extra-cellular glutamate in the nucleus accumbens by inhibitingnonvesicular glutamate release, and group III antagonist MSOPincreased extracellular accumbal glutamate [79]. Thus, it ap-
pears that the postsynaptic mGluR5 receptors need to be blocked,while the presynaptic mGluR7 receptors need to be activated, forinhibition of psychostimulant self-administration. Significantdownregulation of Grm7 has recently been reported in micelacking the subunit α1 of the GABA-A receptor (Gabra1) [80].Because GABRA1 is associated with alcohol dependence [81],there is the possibility of signaling and genetic interaction bet-ween the two receptors.
The source of tissue in our expression QTL mapping may bea source of concern, because the hippocampus is not generallyconsidered as tightly linked to addiction. Thus, the questionmay be raised “Are hippocampal expression data relevant toalcohol drinking?” Although debate continues over the biolo-gical mechanisms of reward and addiction, hippocampal ex-pression data are relevant to alcohol drinking because the hip-pocampus is an important part of the limbic circuitry, a neuralsubstrate of reinforcement and drug addiction. One of thefunctions of the limbic cortical–ventral striatopallidal thalamo-cortical loop circuitry is processing of contextual information bythe hippocampus [82]. For example, dorsal hippocampus inac-tivation abolishes contextual, but fails to alter explicit condi-tioned stimulus- or cocaine-induced, reinstatement of cocaine-seeking behavior [83]. It appears that the amygdala mediatesconditioning to discrete conditioned stimuli, and the hippo-campus mediates contextual or spatial stimuli and may underliethe motivational impact of contextual stimuli on drug seeking[82].
Using progenitor strain-derived mapping population andadditional expression data derived from amygdala, ventral anddorsal striatum, and prefrontal cortex would have been ideal;however, currently such databases are not available. The mostrelevant and available expression mapping database is derivedfrom hippocampal tissue of CXB RI mice. The progenitorstrains in this RI set are BALB/cByJ and C57BL/6ByJ; thus oneof the progenitors (BALB/cByJ) is a subline of BALB/cJ, thestrain that was used as progenitor in our experiments. As of July2007, the SNP-based genetic similarity between BALB/cByJand BALB/cJ cannot be assessed very reliably because thedensity of SNP coverage differs. BALB/cByJ has 7,675,370SNPs, BALB/cJ has 86,221 SNPs (http://phenome.jax.org).Selecting the entire genome, and requiring that SNP locationsmust have allele data present for both strains, 26,831 SNP lo-cation rows were retrieved. Selecting the entire genome, andrequiring that there must be polymorphism between BALB/cByJ and BALB/cJ for a SNP location to be displayed, 41 SNPlocation rows were retrieved, reflecting an average 0.15% dif-ference. A similar comparison for BALB/cByJ and C57BL/6ByJ yielded 33.28% difference. For Grm7 no SNPs could beretrieved in a BALB/cByJ vs BALB/cJ comparison. Thesesuggest that the genomic difference between BALB/cByJ andBALB/cJ and the risk of obtaining false information due to thestrain substitution are relatively small.
Taken together, the reviewed evidence establishes a sig-nificant functional relevance of mGluR7 in psychostimulant-related behaviors, including oral alcohol self-administration.
A potential source of error is introduced in the analysis bythe method used to filter candidate genes residing in the donor

699C. Vadasz et al. / Genomics 90 (2007) 690–702
chromosome segment of the congenic strain B6.C6. Althoughour use of the IBD concept does not assume that the genomes ofinbred strains follow a simple block-like structure, which couldbe used conveniently in QTL mapping [84], and we use thisconcept strictly in a bilineal comparison relying on a definedSNP frequency threshold (b20/100 kb), visual inspection ofeach 100-kb bin, and having access to high-quality genomesequence of the strains recently made available by the NIEHSDNA resequencing efforts, still, there is a chance that relevantgenes escaped detection. Another potential source of error is tofocus on cis-regulated genes exclusively. Gene expression is aphenotype subject to numerous developmental and environ-mental effects. Lack of attention to its spatiotemporal limita-tions may lead to erroneous conclusions.
To counteract these potential effects on the validity of geneidentification, we searched the donor segment for structuralpolymorphism and identified 10 genes with CNs. These genescannot be excluded as potential candidates but their knownfunctions do not suggest any obvious plausible links to alco-hol drinking preference or addiction. Moreover, allelic asso-ciation with alcohol consumption was shown in an additional,alternative genetic design using segregating near-isogenicanimals. Importantly, in this design the position of the target-ed donor segment excluded contribution from both cis-regu-lated and structurally polymorph candidate genes residing inproximal regions of chromosome 6. Finally, we presented alarge body of evidence in support of the functional rele-vance of Grm7 (mGluR7) to psychostimulant/alcohol-relatedbehaviors.
In summary, using near-isogenic advanced animal modelswith reduced genetic background interactions, gene mappingand gene mRNA expression data in segregating and congenicmice were integrated in a search for a gene underlying Eac2, aQTL for alcohol consumption. We identified glutamate receptormetabotropic 7 (Grm7) as a cis-regulated gene for alcohol con-sumption. Confirmation of its role requires further rigorouswork including fine mapping and mechanistic studies, whichare in progress in our laboratory. It is likely that the predisposingeffects of Grm7 polymorphism are not restricted to alcohol-related behaviors; it may play a modulatory role in other formsof addiction and comorbid psychiatric disorders.
Materials and methods
Animals
Progenitors of B6By, CXBI/ByJ (I), and C inbred mouse strains wereobtained from The Jackson Laboratory (Bar Harbor, ME, USA). The quasi-congenic RQI strains and the chromosome 6 congenic B6By.C6 mice weredeveloped in our research colony at The Nathan Kline Institute for PsychiatricResearch, Office of Mental Health, New York State (NKI) as described [6]. Allother mice were bred and maintained in our colony. Animals were housed ingroups (three to five per cage) under standard laboratory conditions (12 h light/12 h dark cycle) with food and water available ad libitum. At the time oftesting, animals were housed individually with lab chow and water conti-nuously available. All reported data refer to male mice. The care and use ofanimals met the standards and recommendations of the IACUC of NKI inaccordance with U.S. Department of Agriculture and U.S. Public HealthService guidelines.
Development of congenic strain B6By.C6
The alcohol-nonpreferring quasi-congenic RQI strain B6.Cb5i7-Alpha3(C5A3) was developed by five backcross–intercross cycles using the alcohol-avoiding C strain as donor and the alcohol-preferring B6By strain as background[6,28]. Using C5A3 as a progenitor, the congenic B6.C6 strain was developedby an additional five backcrosses to B6By with concomitant selection for Calleles of Chr. 6 microsatellite markers (e.g., D6Mit105). Other relevant markerswere selected from the genome-wide genotype database of RQI strains (http://rqigenetics.org/). Congenic status was tested by a genome scan with a mouseSNP panel of 402 markers (service provided by Genetics Division, Brigham andWomen’s Hospital, Harvard Medical School). Only the expected single Chr. 6donor segment was detected as of C origin (Fig. S1).
Oral alcohol self-administration
In all experiments a “two-bottle” choice paradigm with escalating alcoholconcentration was used. Male mice were individually housed in our drinkingstudy room for at least 1 week prior to beginning the study. Mice were tested foralcohol preference and consumption according to the procedure for oral self-administration as described [6,85]. Two versions of the “two-bottle” choiceparadigm were used, one with contiguous trials, and one with intermittent trials.(1) “Two-bottle” choice paradigm with contiguous trials. The test consisted ofsix 3-day trials, in which mice were allowed to choose between alcohol solutionand tap water. To acclimate the animals to the taste of alcohol, the alcoholsolution was offered in escalating concentrations: a 3% solution for trial 1 (days1–3) was increased to 6% in trial 2 (days 3–6), and further increased to 12% fortrials 3 (days 6–9), 4 (days 9–12), 5 (days 12–15), and 6 (days 15–18). Thisarrangement provided multiple measures of alcohol preference at 12%concentration (v/v). Because offering alcohol solutions of 3 and 6% servedthe primary purpose of acclimation of animals to 12% solution, we did not carryout experiments in duplicate for 3 and 6% alcohol consumption, and we did notuse these data for analysis. (2) “Two-bottle” choice paradigm with intermittenttrials. The test consisted of eight 3-day trials, in which mice were allowed tochoose between alcohol solution and tap water. As above, the alcohol solutionwas offered in escalating concentrations: a 3% solution for trial 1 and trial 2 wasincreased to 6% in trial 3 and trial 4 and further increased to 12% for trials 5–8.After every two trials, mice had access to water only for 24 h (from Mondaymorning through Tuesday morning). Thus, the total experiment with intermittentaccess lasted for 4 weeks. For analysis only trials 5–8 were used.
The liquids were offered in custom-made drinking tubes composed ofcentrifuge tubes fitted with single-hole rubber stoppers into which stainless steelsippers were inserted. We also used stainless-steel springs to fasten the tubesfirmly to the top of the cage covers. Two empty cages with alcohol and tap waterdrinking tubes were put on the racks to obtain control weights for leakage andevaporation. For each trial, the mean control values were subtracted from thealcohol and water consumption data. Food was located on the cover at the frontof the cage, with one drinking tube always nearer to the food than the other. Thepositions of the water and alcohol drinking tubes on the cage cover werealternated in each 3-day preference trial to avoid a position effect. The weightsof the drinking tubes were measured before and after a 3-day trial using an A&Delectronic analytical balance connected to a computer. Data were entered auto-matically using A&D software and spreadsheets. After correction for drippingand evaporation, data from the four 3-day trials were averaged for eachindividual.
Brain dissection, RNA labeling, and microarray processing
We used adult, male B6By, B6By.C6, and C mice (n=30; 90 mice in toto).Animals were killed by cervical dislocation, brains were quickly removed andhemisectioned. and alternate left or right hemispheres were collected in RNA-Later for expression studies. After each hemisphere was processed separately, 10hemispheres were pooled for one high-density oligonucleotide microarray(Mouse Genome 430 2.0; Affymetrix, Santa Clara, CA, USA). For each strainthree oligonucleotide microarrays were used (n=3). RNA was isolated usingTRIzol reagent (Invitrogen, Carlsbad, CA, USA) and RNeasy Mini Kits(Qiagen, Valencia, CA, USA). The yield of total RNA from each tissue sample

700 C. Vadasz et al. / Genomics 90 (2007) 690–702
was 45.3±7.6 μg. Fragmented cRNA probes were prepared from 7 μg of totalRNA according to the protocol of Affymetrix using Superscript Choice System(Invitrogen) and an RNA Transcript Labeling Kit (Enzo, Farmingdale, NY,USA). cRNA quality analysis on a 2100 Bioanalyzer, hybridization toGeneChips, and scanning of the arrays were carried out in accordance with themanufacturer’s protocol at the Genomics Facility of the New York UniversityCancer Institute using Gene Chip Operating Software (GCOS).
Microarray gene expression data acquisition and normalization
Raw data with MIAME-compliant annotations of experiments wereuploaded to Gene Traffic Database (http://genetraffic.med.nyu.edu/; IobionInformatics, LLC). We used .CEL files produced by GCOS for probe (cell)-levelRMA-based normalization of data [86]. All subsequent analyses were done onRMA-normalized data. Data from these studies are available at Gene ExpressionOmnibus (GEO series entry GSE7155). All public gene expression data werebased on Affymetrix Mouse Gene Expression Array 430 2.0. CXB RI hippo-campal, BXD hippocampal, BXD striatal, and B6D2F2 striatal mRNA data wereobtained from the Hippocampus Consortium M430v2 CXB (Dec05) RMAdatabase, Hippocampus Consortium M430v2 BXD (Dec05) PDNN database,HBP/Rosen Striatum M430V2 (Apr05) MAS5 Clean database, and OHSU/VAB6D2F2 Striatum M430v2 (Sep05) PDNN database, respectively, at http://www.genenetwork.org/public/WebQTL.
Bioinformatic analysis
Ongoing resequencing efforts made available large SNP genotype data setsfor numerous inbred strains, including B6 and CBy, closely related sublines ofour progenitor strains B6By and C (http://www.jax.org/phenome/snp.html).Because currently our progenitor strains B6By and C are not as wellcharacterized in bioinformatic databases as the closely related sublines B6 andCBy, in the present analysis we took advantage of the excellent bioinformaticcoverage and Mouse Phenome Database priority strain status of B6 and CBy. Ina comparison of C and CBy for all SNPs on Chr. 6 we found 3063 SNPs, whilethe number of different SNPs was 188. This suggests 188/3063=6.14% SNPdifferences on Chr. 6, the length of which is 149.4 Mb. Assuming evendistribution, a rough estimate of the average density of subline SNPs is 188/149.4=1.258, i.e., subline SNP density b2 SNP/Mb (http://www.informatics.jax.org/javawi2/servlet/WIFetch?page=snpQF). This is well below the thresholdapplied for detection of IBD regions (20 SNPs/0.1 Mb) by us and others. In SNPdatabase searches B6 and CBy were substituted for B6By and C if databaseinformation was limited for the progenitor strains. Promoter analysis of Grm7was carried out with the PromoterInspector program (http://www.genomatix.de;Eldorado 08-2006 database, NCBI build 36, extracted region: NC_000072between 110,571,933 and 111,571,932). Positions for the 218 SNPs in Grm7(detected between B6 and CBy or C) were obtained at http://www.informatics.jax.org/; NCBI build 36, 110,611,375–111,533,007. TFBSs were analyzed byMatInspector, the leading program for promoter analysis [87]. The promoteranalysis focused on a 41-bp region, which included the SNP rs30557150Y=C/T (Fig. 3).
Segregating F2 population studies
RQI quasi-congenic strains with significant differences in alcoholconsumption have been described [6,7]. Reciprocal F2 populations were createdby using alcohol-preferring (I5B25A) and alcohol-nonpreferring (C5B3) quasi-congenic progenitor strains. Both strains carry N97% B6 genome. The samplesizes of the (C5B3×I5B25A)F2 and (I5B25A×C5B3)F2 male mice were 159and 163, respectively. Phenotypic “extremes” were identified as 1 SD below orabove the mean alcohol consumption value and were subjected to selectivegenotyping. The selectively genotyped sample sizes were n=28 for (C5B3×I5B25A)F2 and n=30 for (I5B25A×C5B3)F2. To test the association of allelicvariants of Grm7 with alcohol consumption, the phenotypically selected F2mice were genotyped for SNP rs3723352, which resides in intron7 of Grm7 at111.513Mb (NCBI build 36). For allelic variation of rs3723352 in inbred strainswe searched the Mouse Phenome Database (http://phenome.jax.org/pubcgi/phenome/mpdcgi?rtn=snps%2Fretrieve&searchreq=rs3723352).
Multistrain correlation between alcohol consumption and SNPgenotype
Genotypes for Nfu1 (rs30605208), Aak1 (rs30912462), Grm7 (rs305571150), and Chchd4 (rs30014728) were obtained at http://www.informatics.jax.org/. For the correlation analysis SNP genotypes with the C57BL/6J-type allelewere assigned “0”, while the other allele was assigned “1” (genotypes for somestrains were not available). All published alcohol consumption data (g/kg/day)were used without adjustment [21–23].
Acknowledgments
Research described in this article was supported by NIHNIAAA Grant AA11031 and U.S. Army Medical Research andMateriel Command Grant DAMD 17-00-1-0578 to C.V. Wethank Ray Wang, Janos Piturca, Rui Mao, and Jessica Dai forexcellent assistancewith animal care, and genotyping, respectively.
Appendix A. Supplementary data
Supplementary data associated with this article can be found,in the online version, at doi:10.1016/j.ygeno.2007.08.006.
References
[1] J.K. Belknap, A.L. Atkins, The replicability of QTLs for murine alcoholpreference drinking behavior across eight independent studies, Mamm.Genome 12 (2001) 893–899.
[2] J.A. Treadwell, K.B. Pagniello, S.M. Singh, Genetic segregation of braingene expression identifies retinaldehyde binding protein 1 and syntaxin 12as potential contributors to ethanol preference in mice, Behav. Genet. 34(2004) 425–439.
[3] C. Fehr, et al., The syntaxin binding protein 1 gene (Stxbp1) is a candidatefor an ethanol preference drinking locus on mouse chromosome 2,Alcohol. Clin. Exp. Res. 29 (2005) 708–720.
[4] T. Liang, et al., alpha-Synuclein maps to a quantitative trait locusfor alcohol preference and is differentially expressed in alcohol-preferring and -nonpreferring rats, Proc. Natl. Acad. Sci. USA 100(2003) 4690–4695.
[5] R.L. Shirley, et al., Mpdz is a quantitative trait gene for drug withdrawalseizures, Nat. Neurosci. 7 (2004) 699–700.
[6] C. Vadasz, et al., Scanning of five chromosomes for alcohol consumptionloci, Alcohol 22 (2000) 25–34.
[7] C. Vadasz, et al., Mapping of QTLs for oral alcohol self-administration inB6.C and B6.I quasi-congenic RQI strains, Neurochem. Res. 32 (2007)1099–1112.
[8] J.A. Beck, et al., Genealogies of mouse inbred strains, Nat. Genet. 24(2000) 23–25.
[9] C.M. Wade, et al., The mosaic structure of variation in the laboratorymouse genome, Nature 420 (2002) 574–578.
[10] M. Mehrabian, et al., Integrating genotypic and expression data in asegregating mouse population to identify 5-lipoxygenase as asusceptibility gene for obesity and bone traits, Nat. Genet. 37 (2005)1224–1233.
[11] R.B. Brem, et al., Genetic dissection of transcriptional regulation in bud-ding yeast, Science 296 (2002) 752–755.
[12] J. Klose, et al., Genetic analysis of the mouse brain proteome, Nat. Genet.30 (2002) 385–393.
[13] S.A. Monks, et al., Genetic inheritance of gene expression in human celllines, Am. J. Hum. Genet. 75 (2004) 1094–1105.
[14] M. Morley, et al., Genetic analysis of genome-wide variation in humangene expression, Nature 430 (2004) 743–747.
[15] E.E. Schadt, et al., Genetics of gene expression surveyed in maize, mouseand man, Nature 422 (2003) 297–302.

701C. Vadasz et al. / Genomics 90 (2007) 690–702
[16] L. Bystrykh, et al., Uncovering regulatory pathways that affect hema-topoietic stem cell function using ‘genetical genomics’, Nat. Genet. 37(2005) 225–232.
[17] E.J. Chesler, et al., Complex trait analysis of gene expression uncoverspolygenic and pleiotropic networks that modulate nervous system func-tion, Nat. Genet. 37 (2005) 233–242.
[18] G.A. Churchill, R.W. Doerge, Empirical threshold values for quantitativetrait mapping, Genetics 138 (1994) 963–971.
[19] I.A. Eaves, et al., Combining mouse congenic strains and microarray geneexpression analyses to study a complex trait: the NOD model of type 1diabetes, Genome Res. 12 (2002) 232–243.
[20] M.K. Mulligan, et al., Toward understanding the genetics of alcoholdrinking through transcriptome meta-analysis, Proc. Natl. Acad. Sci. USA103 (2006) 6368–6373.
[21] J.K. Belknap, J.C. Crabbe, E.R. Young, Voluntary consumption of ethanolin 15 inbred mouse strains, Psychopharmacology 112 (1993) 503–510.
[22] L.A. Rodriguez, et al., Alcohol acceptance, preference, and sensitivity inmice. I. Quantitative genetic analysis using BXD recombinant inbredstrains, Alcohol. Clin. Exp. Res. 18 (1994) 1416–1422.
[23] K. Gill, et al., Alcohol preference in AXB/BXA recombinant inbredmice: gender differences and gender-specific quantitative trait loci, Mamm.Genome 9 (1998) 929–935.
[24] S.R. Vorel, et al., Relapse to cocaine-seeking after hippocampal theta burststimulation, Science 292 (2001) 1175–1178.
[25] C. Vadasz, et al., Analysis of the mesotelencephalic dopamine system byquantitative-trait locus introgression, Neurochem. Res. 23 (1998)1337–1354.
[26] C. Vadasz, et al., Transfer of brain dopamine system-specific quantitativetrait loci onto a C57BL/6ByJ background, Mamm. Genome 5 (1994)735–737.
[27] C. Vadasz, Development of congenic recombinant inbred neurologicalanimal model lines, Mouse Genome 88 (1990) 16–18.
[28] O. Abiola, et al., The nature and identification of quantitative trait loci: acommunity’s view, Nat. Rev., Genet. 4 (2003) 911–916.
[29] D. Wahlsten, et al., Different data from different labs: lessons fromstudies of gene–environment interaction, J. Neurobiol. 54 (2003)283–311.
[30] J.C. Crabbe, D. Wahlsten, B.C. Dudek, Genetics of mouse behavior:interactions with laboratory environment, Science 284 (1999) 1670–1672.
[31] R. Korstanje, B. Paigen, From QTL to gene: the harvest begins, Nat.Genet. 31 (2002) 235–236.
[32] J. Kroymann, T. Mitchell-Olds, Epistasis and balanced polymorphisminfluencing complex trait variation, Nature 435 (2005) 95–98.
[33] I.M. Stylianou, et al., Complex genetic architecture revealed by analysis ofhigh-density lipoprotein cholesterol in chromosome substitution strainsand F2 crosses, Genetics 174 (2006) 999–1007.
[34] P.J. Flor, et al., A novel splice variant of a metabotropic glutamate receptor,human mGluR7b, Neuropharmacology 36 (1997) 153–159.
[35] H.L. Schulz, H. Stohr, B.H. Weber, Characterization of three novelisoforms of the metabotropic glutamate receptor 7 (GRM7), Neurosci.Lett. 326 (2002) 37–40.
[36] K.K. Dev, S. Nakanishi, J.M. Henley, Regulation of mglu(7) receptors byproteins that interact with the intracellular C-terminus, Trends Pharmacol.Sci. 22 (2001) 355–361.
[37] J.A. Saugstad, et al., Cloning and expression of a new member of the L-2-amino-4-phosphonobutyric acid-sensitive class of metabotropic glutamatereceptors, Mol. Pharmacol. 45 (1994) 367–372.
[38] Y. Nakajima, et al., A relationship between protein kinase C phosphoryla-tion and calmodulin binding to the metabotropic glutamate receptorsubtype 7, J. Biol. Chem. 274 (1999) 27573–27577.
[39] C. Corti, et al., Cloning and characterization of alternative mRNA formsfor the rat metabotropic glutamate receptors mGluR7 and mGluR8, Eur. J.Neurosci. 10 (1998) 3629–3641.
[40] A. Kinoshita, et al., Immunohistochemical localization of metabotropicglutamate receptors, mGluR7a and mGluR7b, in the central nervoussystem of the adult rat and mouse: a light and electron microscopic study,J. Comp. Neurol. 393 (1998) 332–352.
[41] J. Lu, et al., Transcript-based redefinition of grouped oligonucleotide probe
sets using AceView: high-resolution annotation for microarrays, BMCBioinformatics 8 (2007) 108.
[42] V.M. Wunderle, et al., Cloning and characterization of SOX5, a newmember of the human SOX gene family, Genomics 36 (1996) 354–358.
[43] A.E. Brent, T. Braun, C.J. Tabin, Genetic analysis of interactions betweenthe somitic muscle, cartilage and tendon cell lineages during mousedevelopment, Development 132 (2005) 515–528.
[44] D.D. Schoepp, Unveiling the functions of presynaptic metabotropicglutamate receptors in the central nervous system, J. Pharmacol. Exp. Ther.299 (2001) 12–20.
[45] M. Masugi, et al., Metabotropic glutamate receptor subtype 7 ablationcauses deficit in fear response and conditioned taste aversion, J. Neurosci.19 (1999) 955–963.
[46] J.M. Kinzie, et al., Distribution of metabotropic glutamate receptor 7messenger RNA in the developing and adult rat brain, Neuroscience 69(1995) 167–176.
[47] A. Simonyi, L.A. Miller, G.Y. Sun, Region-specific decline in theexpression of metabotropic glutamate receptor 7 mRNA in rat brain duringaging, Brain Res. Mol. Brain Res. 82 (1–2) (2000) 101–106.
[48] J.J. Nadler, et al., Large-scale gene expression differences across brainregions and inbred strains correlate with a behavioral phenotype, Genetics174 (2006) 1229–1236.
[49] J.F. Cryan, et al., Antidepressant and anxiolytic-like effects in mice lackingthe group III metabotropic glutamate receptor mGluR7, Eur. J. Neurosci.17 (2003) 2409–2417.
[50] K. Mitsukawa, et al., Metabotropic glutamate receptor subtype 7 ablationcauses dysregulation of the HPA axis and increases hippocampal BDNFprotein levels: implications for stress-related psychiatric disorders,Neuropsychopharmacology 31 (2006) 1112–1122.
[51] K. Mitsukawa, et al., A selective metabotropic glutamate receptor 7agonist: activation of receptor signaling via an allosteric site modulatesstress parameters in vivo, Proc. Natl. Acad. Sci. USA 102 (2005)18712–18717.
[52] M.P. Johnson, G. Kelly, M. Chamberlain, Changes in rat serum corti-costerone after treatment with metabotropic glutamate receptor agonists orantagonists, J. Neuroendocrinol. 13 (2001) 670–677.
[53] J.G. Tasker, C. Boudaba, L.A. Schrader, Local glutamatergic andGABAergic synaptic circuits and metabotropic glutamate receptors inthe hypothalamic paraventricular and supraoptic nuclei, Adv. Exp. Med.Biol. 449 (1998) 117–121.
[54] Z. Sarnyai, Y. Shaham, S.C. Heinrichs, The role of corticotropin-releasingfactor in drug addiction, Pharmacol. Rev. 53 (2001) 209–243.
[55] I. Sillaber, et al., Enhanced and delayed stress-induced alcohol drinking inmice lacking functional CRH1 receptors, Science 296 (2002) 931–933.
[56] Z. Nie, et al., Ethanol augments GABAergic transmission in the centralamygdala via CRF1 receptors, Science 303 (2004) 1512–1514.
[57] C.F. Plappert, P.K. Pilz, Difference in anxiety and sensitization of theacoustic startle response between the two inbred mouse strains BALB/cANand DBA/2N, Genes Brain Behav. 1 (2002) 178–186.
[58] A.M. Depino, C. Gross, Simultaneous assessment of autonomic functionand anxiety-related behavior in BALB/c and C57BL/6 mice, Behav. BrainRes. 177 (2007) 254–260.
[59] J.C. DeFries, M.C. Gervais, E.A. Thomas, Response to 30 generations ofselection for open-field activity in laboratory mice, Behav. Genet. 8 (1978)3–13.
[60] C. Vadasz, G. Kobor, A. Lajtha, Motor activity and the mesotelencephalicdopamine function: II. Multivariate analysis of genetically segregatinggenerations, Behav. Brain Res. 48 (1992) 41–47.
[61] C. Vadasz, G. Kobor, A. Lajtha, Motor activity and the mesotelencephalicdopamine function: I. High-resolution temporal and genetic analysis ofopen-field behavior, Behav. Brain Res. 48 (1992) 29–39.
[62] K. Priebe, et al., Maternal influences on adult stress and anxiety-likebehavior in C57BL/6J and BALB/cJ mice: a cross-fostering study, Dev.Psychobiol. 47 (2005) 398–407.
[63] R.A. Wise, Addictive drugs and brain stimulation reward, Annu. Rev.Neurosci. 19 (1996) 319–340.
[64] R.A. Wise, M.A. Bozarth, A psychomotor stimulant theory of addiction,Psychol. Rev. 94 (1987) 469–492.

702 C. Vadasz et al. / Genomics 90 (2007) 690–702
[65] E.J. Nestler, Is there a common molecular pathway for addiction? Nat.Neurosci. 8 (2005) 1445–1449.
[66] P.W. Kalivas, Glutamate systems in cocaine addiction, Curr. Opin.Pharmacol. 4 (2004) 23–29.
[67] C.J. Swanson, P.W. Kalivas, Regulation of locomotor activity bymetabotropic glutamate receptors in the nucleus accumbens and ventraltegmental area, J. Pharmacol. Exp. Ther. 292 (2000) 406–414.
[68] L. Mao, Y.S. Lau, J.Q. Wang, Activation of group III metabotropicglutamate receptors inhibits basal and amphetamine-stimulated dopaminerelease in rat dorsal striatum: an in vivo microdialysis study, Eur. J.Pharmacol. 404 (2000) 289–297.
[69] L. Mao, J.Q. Wang, Distinct inhibition of acute cocaine-stimulated motoractivity following microinjection of a group III metabotropic glutamatereceptor agonist into the dorsal striatum of rats, Pharmacol. Biochem.Behav. 67 (2000) 93–101.
[70] C.W. Hodge, et al., The mGluR5 antagonist MPEP selectively inhibits theonset and maintenance of ethanol self-administration in C57BL/6J mice,Psychopharmacology (Berlin) 183 (2006) 429–438.
[71] A. Simonyi, et al., Chronic ethanol on mRNA levels of IP3R1, IP33-kinase and mGluR1 in mouse Purkinje neurons, NeuroReport 7 (1996)2115–2118.
[72] A. Simonyi, et al., Chronic ethanol-induced subtype- and subregion-specific decrease in the mRNA expression of metabotropic glutamate re-ceptors in rat hippocampus, Alcohol. Clin. Exp. Res. 28 (2004) 1419–1423.
[73] P. Backstrom, et al., mGluR5 antagonist MPEP reduces ethanol-seekingand relapse behavior, Neuropsychopharmacology 29 (2004) 921–928.
[74] J.P. Schroeder, D.H. Overstreet, C.W. Hodge, The mGluR5 antagonistMPEP decreases operant ethanol self-administration during maintenanceand after repeated alcohol deprivations in alcohol-preferring (P) rats, Psy-chopharmacology (Berlin) 179 (2005) 262–270.
[75] J. Besheer, C.W. Hodge, Pharmacological and anatomical evidence for aninteraction between mGluR5- and GABA(A) alpha1-containing receptorsin the discriminative stimulus effects of ethanol, Neuropsychopharmacol-ogy 30 (2005) 747–757.
[76] C. Bills, et al., Effects of metabotropic glutamate receptor 5 on latent
inhibition in conditioned taste aversion, Behav. Brain Res. 157 (2005)71–78.
[77] W.P. Spooren, et al., Novel allosteric antagonists shed light on mglu(5)receptors and CNS disorders, Trends Pharmacol. Sci. 22 (2001)331–337.
[78] X. Li, et al., Activation of metabotropic glutamate receptor 7 (mGluR7) byAMN082 attenuates the rewarding effects of cocaine by a DA-independentmechanism in rats, Society for Neuroscience Meeting, Atlanta, 2006,[Abstract 21.7].
[79] Z.X. Xi, et al., Inhibition of non-vesicular glutamate release by group IIImetabotropic glutamate receptors in the nucleus accumbens, J. Neuro-chem. 87 (2003) 1204–1212.
[80] I. Ponomarev, et al., Transcriptional signatures of cellular plasticity in micelacking the alpha1 subunit of GABAA receptors, J. Neurosci. 26 (2006)5673–5683.
[81] D.M. Dick, et al., Association between GABRA1 and drinking behaviorsin the collaborative study on the genetics of alcoholism sample, Alcohol.Clin. Exp. Res. 30 (2006) 1101–1110.
[82] B.J. Everitt, T.W. Robbins, Neural systems of reinforcement for drugaddiction: from actions to habits to compulsion, Nat. Neurosci. 8 (2005)1481–1489.
[83] R.A. Fuchs, et al., The role of the dorsomedial prefrontal cortex,basolateral amygdala, and dorsal hippocampus in contextual reinstate-ment of cocaine seeking in rats, Neuropsychopharmacology 30 (2005)296–309.
[84] B. Yalcin, et al., Unexpected complexity in the haplotypes of commonlyused inbred strains of laboratory mice, Proc. Natl. Acad. Sci. USA 101(2004) 9734–9739.
[85] C. Vadasz, et al., Ventral tegmental transcriptome response to intermittentnicotine treatment and withdrawal in BALB/cJ, C57BL/6ByJ, and quasi-congenic RQI mice, Neurochem. Res. 32 (2007) 457–480.
[86] R.A. Irizarry, et al., Summaries of Affymetrix GeneChip probe level data,Nucleic Acids Res. 31 (2003) e15.
[87] K. Cartharius, et al., MatInspector and beyond: promoter analysis based ontranscription factor binding sites, Bioinformatics 21 (2005) 2933–2942.


![In vivo positron emission tomography imaging with [ 11 C]ABP688: binding variability and specificity for the metabotropic glutamate receptor subtype 5 in baboons](https://static.fdokumen.com/doc/165x107/6316b26ad18b031ae106426d/in-vivo-positron-emission-tomography-imaging-with-11-cabp688-binding-variability.jpg)

















