
Glacio-eustatic Control of Continental–Shallow Marine Cyclicity from Late Quaternary Deposits of...
13
Glacio-eustatic Control of Continental–Shallow Marine Cyclicity from Late Quaternary Deposits of the Southeastern Po Plain, Northern Italy Alessandro Amorosi, Maria Luisa Colalongo, and Fabio Fusco Dipartimento di Scienze della Terra e Geologico-Ambientali, Universita ` di Bologna, Via Zamboni 67, 40127 Bologna, Italy Giancarlo Pasini Istituto di Geologia Marina, C.N.R., Via Gobetti 101, 40129 Bologna, Italy and Flavia Fiorini Dipartimento di Scienze della Terra e Geologico-Ambientali, Universita ` di Bologna, Via Zamboni 67, 40127 Bologna, Italy Received September 10, 1998 Integrated sedimentological and micropaleontological (forami- nifera, ostracods, pollens) investigations of a 173-m-long continu- ous core in the subsurface of the SE Po Plain reveal a cyclic facies distribution of continental to shallow-marine deposits. Radiomet- ric and pollen data enable an age attribution of the study units to the late Middle Pleistocene–Holocene. Glacioeustasy appears to have exerted a major control on facies development, as docu- mented by the close relationships between cyclic lithofacies pat- terns and the curves of eustatic fluctuation. Particularly, relative sea-level falls and lowstand phases were characterized by the development of alluvial sedimentation, whereas major transgres- sive pulses and subsequent highstands are reflected by abrupt shifts to paralic and shallow-marine deposits. This facies architec- ture is paralleled by a distinctive pollen distribution, reflecting the alternation of a glacial type of vegetation, dominated by Pinus and nonarboreal pollen types, with interglacial mixed deciduous broad-leaved forests. © 1999 University of Washington. Key Words: cyclostratigraphy; continental deposits; marine deposits; benthic foraminifera; ostracods; palynology; Po Plain, Italy. INTRODUCTION The Po Plain is the surface expression of a perisutural basin bounded by two mountain chains (the Apennines to the S and the Alps to the N), with its prolongation in the Adriatic Sea (Fig. 1). The repeated alternation, a few hundreds of meters thick, of shallow-marine and alluvial sediments is the basic motif of the subsurface of the SE Po Plain (Regione Emilia- Romagna and ENI-AGIP, 1998). The detailed facies architecture of the uppermost 30 m of the SE Po Plain has been recently shown by Amorosi et al. (1999), who documented that facies distribution strongly reflects glacio-eustatic fluctuations. Particularly, at this location late Pleistocene alluvial deposits are unconformably overlain by a transgressive–regressive cycle (Ravenna Allomember) of Ho- locene age (Fig. 2). Facies distribution within the Ravenna Allomember reflects the landward migration of a transgressive barrier–lagoon system (back-barrier and lower beach ridge deposits of Fig. 2), followed by the progradation of a delta system (marsh and upper beach ridge deposits of Fig. 2). As part of the geological mapping project of the Quaternary deposits of the southern Po Plain to scale 1:50,000, promoted by the Geological Survey of the Regione Emilia-Romagna and the Geological Survey of Italy, a continuously cored borehole (S17), 173 meters deep, near the village of Mandriole (15 km N of Ravenna), was studied in detail through integrated sedi- mentological and micropaleontological (foraminifera, ostra- cods, pollens) investigations (Fig. 1). The stratigraphic frame- work of the uppermost 30 m of this core has been discussed by Amorosi et al. (1999) (Fig. 2). In this paper we present the first example of detailed facies characterization in the Po Plain of the deposits that accumu- lated during the three past glacial/interglacial cycles and dis- cuss the link between the sedimentary evolution of the SE Po Plain and the Late Quaternary sea-level fluctuations. MATERIAL AND METHODS Sediment color and texture, vertical lithofacies relationships, and the type and concentration of accessory materials, includ- ing paleosols, roots, plant fragments, and organic matter, were Quaternary Research 52, 1–13 (1999) Article ID qres.1999.2049, available online at http://www.idealibrary.com on 0033-5894/99 $30.00 Copyright © 1999 by the University of Washington. All rights of reproduction in any form reserved. 1
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Glacio-eustatic Control of Continental–Shallow Marine Cyclicity from Late Quaternary Deposits of...

nodrthmtsdsstanb
dI
bt(tmR
Quaternary Research52, 1–13 (1999)Article ID qres.1999.2049, available online at http://www.idealibrary.com on
Glacio-eustatic Control of Continental–Shallow Marine Cyclicityfrom Late Quaternary Deposits of the Southeastern
Po Plain, Northern Italy
Alessandro Amorosi, Maria Luisa Colalongo, and Fabio Fusco
Dipartimento di Scienze della Terra e Geologico-Ambientali, Universita` di Bologna, Via Zamboni 67, 40127 Bologna, Italy
Giancarlo Pasini
Istituto di Geologia Marina, C.N.R., Via Gobetti 101, 40129 Bologna, Italy
and
Flavia Fiorini
Dipartimento di Scienze della Terra e Geologico-Ambientali, Universita` di Bologna, Via Zamboni 67, 40127 Bologna, Italy
Received September 10, 1998
basa
eaeteasiilia
Sw ectsg lateP by at f Ho-l nnaA siveb ridged eltas .
aryd otedb andt hole( kmN sedi-m tra-c me-w ed byA
ciesc umu-l dis-c E PoP
ips,a clud-
Integrated sedimentological and micropaleontological (forami-ifera, ostracods, pollens) investigations of a 173-m-long continu-us core in the subsurface of the SE Po Plain reveal a cyclic faciesistribution of continental to shallow-marine deposits. Radiomet-ic and pollen data enable an age attribution of the study units tohe late Middle Pleistocene–Holocene. Glacioeustasy appears toave exerted a major control on facies development, as docu-ented by the close relationships between cyclic lithofacies pat-
erns and the curves of eustatic fluctuation. Particularly, relativeea-level falls and lowstand phases were characterized by theevelopment of alluvial sedimentation, whereas major transgres-ive pulses and subsequent highstands are reflected by abrupthifts to paralic and shallow-marine deposits. This facies architec-ure is paralleled by a distinctive pollen distribution, reflecting thelternation of a glacial type of vegetation, dominated by Pinus andonarboreal pollen types, with interglacial mixed deciduousroad-leaved forests. © 1999 University of Washington.
Key Words: cyclostratigraphy; continental deposits; marineeposits; benthic foraminifera; ostracods; palynology; Po Plain,taly.
INTRODUCTION
The Po Plain is the surface expression of a perisuturalounded by two mountain chains (the Apennines to the S
he Alps to the N), with its prolongation in the Adriatic SFig. 1). The repeated alternation, a few hundreds of mhick, of shallow-marine and alluvial sediments is the botif of the subsurface of the SE Po Plain (Regione Emomagna and ENI-AGIP, 1998).The detailed facies architecture of the uppermost 30 m o
f thi1
innd
rsc-
e
E Po Plain has been recently shown by Amorosiet al. (1999),ho documented that facies distribution strongly refllacio-eustatic fluctuations. Particularly, at this locationleistocene alluvial deposits are unconformably overlain
ransgressive–regressive cycle (Ravenna Allomember) oocene age (Fig. 2). Facies distribution within the Ravellomember reflects the landward migration of a transgresarrier–lagoon system (back-barrier and lower beacheposits of Fig. 2), followed by the progradation of a dystem (marsh and upper beach ridge deposits of Fig. 2)As part of the geological mapping project of the Quatern
eposits of the southern Po Plain to scale 1:50,000, promy the Geological Survey of the Regione Emilia-Romagna
he Geological Survey of Italy, a continuously cored boreS17), 173 meters deep, near the village of Mandriole (15
of Ravenna), was studied in detail through integratedentological and micropaleontological (foraminifera, os
ods, pollens) investigations (Fig. 1). The stratigraphic fraork of the uppermost 30 m of this core has been discussmorosi et al. (1999) (Fig. 2).In this paper we present the first example of detailed fa
haracterization in the Po Plain of the deposits that accated during the three past glacial/interglacial cycles anduss the link between the sedimentary evolution of the Slain and the Late Quaternary sea-level fluctuations.
MATERIAL AND METHODS
Sediment color and texture, vertical lithofacies relationshnd the type and concentration of accessory materials, in
ng paleosols, roots, plant fragments, and organic matter,
were0033-5894/99 $30.00Copyright © 1999 by the University of Washington.
All rights of reproduction in any form reserved.

u ssc ding rvC asu ano wei ofet a,t erc wec amn (191 91B n-c no ev( n( ega
tef ata 99Mc sa one rago cet ths umi tesw
A P1,A ent P2,a
ntf . 3).T
0
andfi aneseo ilabled pro-p -r entra-t
outh-e han-n n aw f thef PoR ege-t
2
dary
AMOROSI ET AL.2
sed as basic tools for identification of the major facies aiations. Primary sedimentary structures, especially bedeometries in coarse-grained deposits, are poorly preseharacterization of depositional environments was also bpon detailed micropaleontologic (benthic foraminiferastracods) investigation of 434 samples (Fig. 3). Samples
nitially dried for 8 h at60°C. Two hundred and fifty gramsach sample was washed with either H2O or 12% H2O2,
hrough sieves of 63-mm mesh. For counting the microfaunhe .63-mm size fractions of the whole wash residues wonsidered. Three hundred specimens for sampleounted. The paleoenvironmental significance of the foriferal assemblages is based upon papers by Jorissen988), Van der Zwaan and Jorissen (1991), Verhallen (19armawidjajaet al.(1992), Langer (1993), Sgarrella and Moharmont Zei (1993), and Bellottiet al. (1994). Interpretatiof ostracod assemblages is based upon papers by D1965), Socac (1978), Athersuchet al. (1989), Henderso1990), Van Frausum and Wouters (1990), and Montennd Pugliese (1995).Sixty-five samples from clays and silty clays were collec
or pollen analysis. Ten grams of dried sediments was treccording to standard methods (Faegri and Iversen, 1oore et al., 1992), using ZnCl2 (density, 2.05 g/cm3) to
oncentrate pollen. A tablet ofLycopodiumsp. spores wadded to each sample at the start of sample preparatistimate pollen concentration (Stockmarr, 1971). An avef 400 pollen grains per sample was counted. Pollen con
rations are based upon the total amount of grains. Onlyynthetic pollen diagram is shown, in which the pollen sncludes all pollens, with the exception of hygro-hydrophyhich are represented apart (Fig. 3), and indeterminate g
FIG. 1. Location of well S17. The dash
raiw
o-g
ed.eddre
erei-87,),
oto
ro
ded2;
toen-e
,ns.
rboreal pollen types (AP) are split into four classes (AP2, Alnus group, andPinus), whereas nonarboreal poll
ypes (NAP) are subdivided into three classes (NAP1, NAndPoaceae).
SEDIMENTOLOGY AND MICROPALEONTOLOGYOF CORE S17
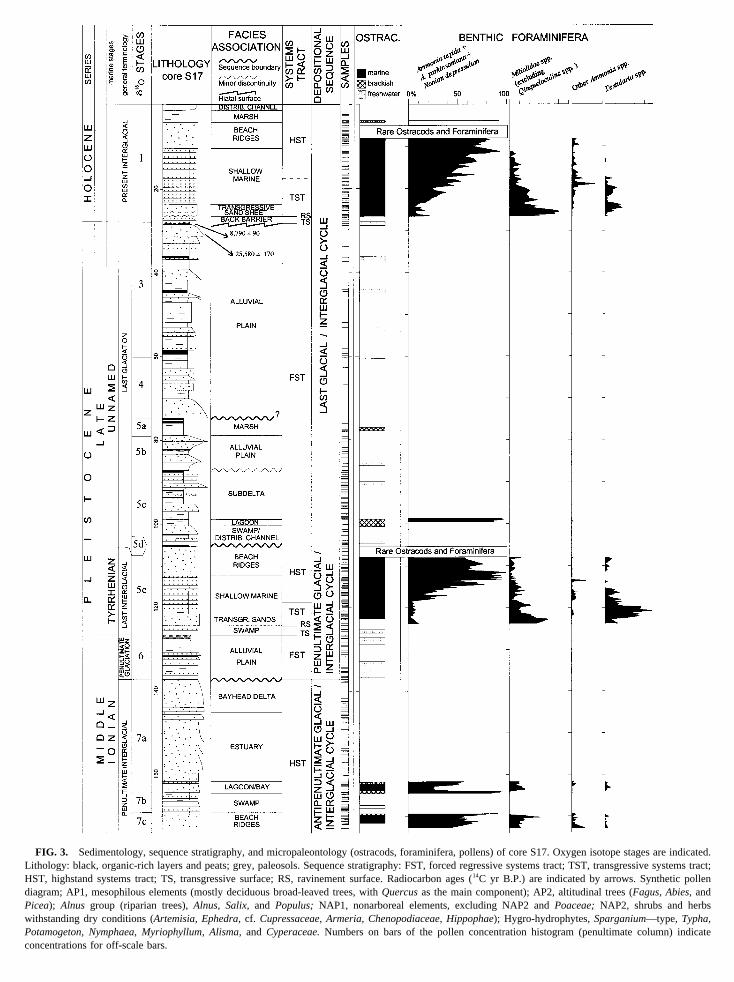
Twenty-two stratigraphic intervals, including 12 differeacies associations, were identified within core S17 (Fighey are described in descending order.
.00–2.30 m: Distributary Channel Facies Association
This facies association includes brownish silty sandsne sands, with root traces and abundant iron and mangxides. No fossils are present. The pollen spectrum avaisplays a varied association with AP and NAP in equalortions. AP are dominated byQuercus,while NAP are repesented by many taxa in equal percentages. Pollen concion is very low.
This association is interpreted as levee deposits of the srnmost distributary channel of the paleo-Po delta. This cel was active until 800 yr B.P. (Veggiani, 1974), wheell-known avulsion event caused the abandonment o
ormer Po delta lobe and the northward migration of theiver toward its present position. Pollen data indicate a v
ation possibly related to human activity.
.30–4.60 m: Marsh Facies Association
This facies association, showing a gradational bounith the overlying deposits, consists dominantly of grey c
lines refer to the fence diagram of Figure 2.
edlays

a t. Tb -t sfrs ntsS (i.eP hea sisw .
ssil int
4
fins vel esa
dsp s.R them datad riant
grad-i eltas sia shore( lying,f its.
1
icala seds fos-
lain, withs Ra
GLACIO-EUSTATIC CONTROL OF SEDIMENTATION, NORTHERN ITALY 3
nd peats. Wood fragments and plant debris are abundanrackish ostracodCyprideis torosaand rareLoxoconcha ellip
ica characterize this interval. Remarkable concentrationoraminifera Ammonia parkinsonianaand A. tepidaare alsoecorded. The dominance (65%) ofPoaceaein the pollenpectrum could indicate proximity to marsh environmecattered pollen grains possibly related to human activitylantago, Poaceae. 50 mm) are present in this interval. Tge of these deposits is constrained by radiocarbon analyood to between 22306 6014C yr B.P. (Fig. 2) and 800 yr B.PThis association corresponds to brackish marsh and po
agoonal deposits that accumulated in the lower delta plahe paleo-Po delta (Amorosiet al., 1999).
.60–10.30 m: Beach Ridge Facies Association
This facies association includes well-sorted very fine toands, with abundant shelly material, that are gradually oain by silty sands. The lower part of this interval includbundantAmmonia tepidaandA. parkinsoniana,with subor-
FIG. 2. Fence diagram (after Amorosiet al., 1999) showing stratal arctratigraphic correlations between core S17 and the neighboring wells.
s
he
of
.
.,
of
blyof
er-
inateElphidiumspp. (consisting almost entirely ofE. grano-um and E. complanatum), and several species ofCribroel-hidium,among whichC. decipiensis the dominant specieare reworked foraminifera and ostracods characterizeiddle and upper parts of this facies association. Pollenisplay a rich association, with high percentages of ripa
reesAlnusandSalix.This association has been interpreted to represent a pro
ng shoreline of a beach ridge in a wave-dominated dystem (delta front deposits of the paleo-Po delta) (Amoroetl., 1999). The fossiliferous sands are interpreted as nearshoreface and foreshore) deposits, whereas the overossil-free silty sands are interpreted as backshore depos
0.30–23.60 m: Shallow-Marine Facies Association
This facies association is characterized by the rhythmlternation of grey clay–silty clay and very fine, sharp-baand layers (generally less than 2 cm thick). Marine macroils are abundant and commonly concentrated at distinct s
cture of the uppermost Pleistocene–Holocene deposits of the SE Po Pdiocarbon dates (14C yr B.P.) are indicated.
hite
trati-
re indicateL t;H llendP swP dicatec
FIG. 3. Sedimentology, sequence stratigraphy, and micropaleontology (ostracods, foraminifera, pollens) of core S17. Oxygen isotope stages ad.ithology: black, organic-rich layers and peats; grey, paleosols. Sequence stratigraphy: FST, forced regressive systems tract; TST, transgressive systems tracST, highstand systems tract; TS, transgressive surface; RS, ravinement surface. Radiocarbon ages (14C yr B.P.) are indicated by arrows. Synthetic poiagram; AP1, mesophilous elements (mostly deciduous broad-leaved trees, withQuercusas the main component); AP2, altitudinal trees (Fagus, Abies,andicea); Alnus group (riparian trees),Alnus, Salix,and Populus; NAP1, nonarboreal elements, excluding NAP2 andPoaceae;NAP2, shrubs and herbithstanding dry conditions (Artemisia, Ephedra,cf. Cupressaceae, Armeria, Chenopodiaceae, Hippophae); Hygro-hydrophytes,Sparganium—type, Typha,otamogeton, Nymphaea, Myriophyllum, Alisma,and Cyperaceae.Numbers on bars of the pollen concentration histogram (penultimate column) inoncentrations for off-scale bars.
GLACIO-EUSTATIC CONTROL OF SEDIMENTATION, NORTHERN ITALY 5
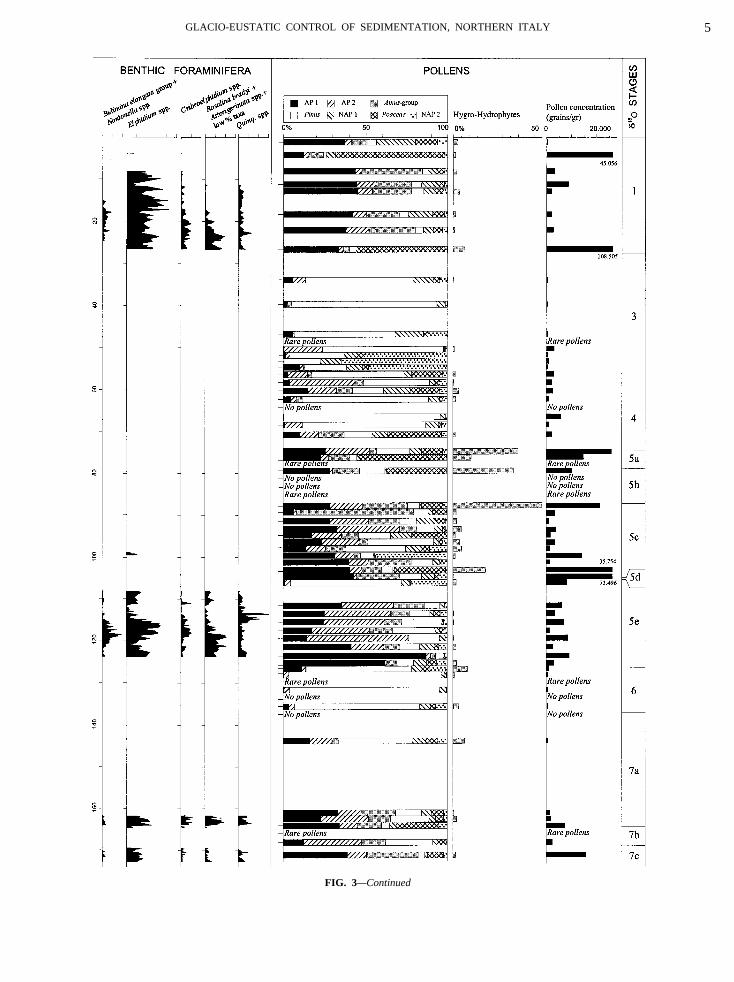
FIG. 3—Continued

g uw ) iAtQa re-c enb ri-a L.t -t ot( ciat thelt vee sith
nvr lows disc floe
2
ions witr riaCs inifes thsso ata hisi sith1
wes stad ssc wif1 asw etfl loc
2
n bs frot
f ts ofaoe val,w fA uousb -richs rangeb
baya of ab -c
2
sistso me-d 1.50,3 ero-s ten-d eredb Lowm istincts .50–5 thisfsy g. 2)h -f theo -g ssa,C yo-c ris,C Inc erizet taxa,w eousv .At o-n m.R ntra-tn ron-m
vialp andc sedi-m tincts ithine s and
AMOROSI ET AL.6
raphic levels. The foraminifera are characterized by theard increase (up to 93% at the top of this interval; Fig. 3. parkinsoniana, A. tepida,and Nonion depressulum.This
rend is paralleled by the abrupt decrease inMiliolidae spp. anduinqueloculinaspp. High concentrations ofElphidiumspp.,mong whichE. granosumis the dominant species, areorded throughout this interval. The ostracods are represy rare valves ofPontocythere turbida, Callistocythere adtica, Leptocythere levis, L. cf. multipunctata, L. rara,
enera, Loxoconcha rubritincta,andL. stellifera.Pollen specra from this interval indicate a relatively high abundancehermophilous deciduous forests. High values ofJuglans15%) at 11.20 m allow age attribution of this facies assoion to historical time. The pollen spectrum available inower part of this interval shows very low values ofPinusandhe relative abundance of mixed deciduous broad-leavedtation. Radiometric dating of wood from correlative depoas provided an age of 33056 60 14C yr B.P. (Fig. 2).This association formed in a shallow-marine, prodelta e
onment during an interglacial period, under increasingalinity stressed conditions, probably influenced by fluvialharge. The sand layers are interpreted as the result ofvents.
3.60–26.60 m: Transgressive Sand Sheet
The base of this facies association is defined by an erosurface, which is overlain by a few meters of silty sandsare intercalations of silty clays and abundant shelly mateoncentration of marine bivalves and gastropods (Turritellap.) is common at distinct stratigraphic levels. The foramral assemblage is dominated byMiliolidae spp. andElphidiumpp. (among whichE. crispumis the dominant species), wiubordinateRosalina bradyi, Asterigerinataspp., Textulariapp., Cribroelphidium spp., andQuinqueloculinaspp. Thestracod assemblage is dominated byP. turbida, Loxoconch
umida,and L. turbida, with subordinateC. adriatica and C.ff. flavidofusca.No pollen analyses were carried out in t
nterval. Radiometric dating of wood from correlative depoas provided an age of 76056 130 14C yr B.P. (Amorosiet al.,999).This association formed in a high-energy (possibly lo
horeface) littoral environment, under interglacial or interial conditions. The basal erosion surface of this facies aiation is interpreted as a ravinement surface (RS) (S968; Nummedal and Swift, 1987). The facies associationhole has been interpreted as a transgressive sand sheecting the landward migration of a barrier during the Hoene transgression (Amorosiet al., 1999).
6.60–27.90 m: Back-Barrier Facies Association
This facies association consists of silty sands overlaiilts, with peat intercalations. No samples were collectedhis interval for microfaunal analysis. Correlative depo
sitpp-n
ted
f
-
g-s
i---od
alhl.
-
s
r-o-t,a
, re--
yms
rom adjacent wells have shown that the microfauna consisforaminiferal assemblage dominated byA. tepida and an
stracod assemblage dominated byCyprideis torosa(Amorosit al., 1999). The pollen spectrum available for this interith very low values ofPinus and relatively high values oP1, indicates the dominance of an arboreal, mixed decidroad-leaved vegetation. Radiometric dating of organicediment from correlative deposits has provided an ageetween 87906 90 and 81706 50 14C yr. B.P. (Fig. 2).This association formed in either a marsh or a protected
s back-barrier deposits, during the Holocene migrationarrier–lagoon system (Amorosiet al., 1999), under interglaial or interstadial climate conditions.
7.90–75.00 m: Alluvial Plain Facies Association
This facies association, approximately 47 m thick, conf bioturbated grey silts and clays, with common fine toium sand intercalations. The major sand bodies (28.15–39.10–42.00, and 70.10–75.00 m) commonly displayional bases and normal grading. A coarsening-upwardency from silts to silt–sand alternations is locally encountelow these upward-fining successions (31.50–34.70 m).ature paleosols and organic-rich layers are present at d
tratigraphic levels (36.55–37.05, 44.00–44.60, and 589.00 m). Stiff black silts constitute the uppermost part of
acies association (27.90–28.15 m), wherea . 15,000-yrtratigraphic hiatus (between 25,5806 170 and 87906 90 14Cr B.P., on the basis of dates on organic-rich sediment (Fias been documented by Amorosiet al. (1999). The micro
auna of this association is restricted almost entirely torganic-rich layers, where freshwater ostracods (Candona anulata, C. neglecta, C. albicans, C. candida, C. compreypria ophtalmica, Heterocypris salinus, Cypris pubera, Ilypris gibba, I. bradyi, I. aff. monstrifera, Cytherissa lacustyclocypris ovum, C. levis) are commonly encountered.ontrast, a reworked microfauna or lack of fossils characthe sand bodies. The pollen flora is composed of a fewith Pinus as the major arboreal element and herbacegetation dominated byArtemisia, Asteraceae,andPoaceaeP1 and AP2 are generally either scarce or absent, andPicea,
ogether withAlnusandBetula, is the major arboreal compent within the 56.40- to 60.15-m interval and at 70.70eworked grains are often encountered and pollen conce
ions are low to very low. High percentages (.60%) of Che-opodiaceaeindicate the probable presence of marsh envients in adjacent areas.This association is interpreted to have formed on an allu
lain under glacial or stadial climate conditions. The siltslays are considered to represent floodplain and leveeents. The accumulation of organic-rich layers at dis
tratigraphic levels reflects low sedimentation rates wphemeral standing bodies of water (freshwater swamponds). Major sand bodies including reworked benthic for
ami-
n tc vas fci hisf f thC t ths dac
7
mop f thb blep onA ofw
asp
7
ius ces nag entad eb
woc eflc wb tion
8
mop rsc grs rdT ves arem andA eryh n-d tor
atee iete ned inc rdt inom
9
up-p siltyc dt erat .p hE anti thisi -a tom l thep on ina
therl l ori ra int rinei thea sis-t esh-w
1
k siltl la-t sandb g-u esh-w efi faunai y theh llens bun-d Ar ecid-u g de-p 0 m,w
anu . Thes nel-r k siltl ersm ando l ori
1
nds.
GLACIO-EUSTATIC CONTROL OF SEDIMENTATION, NORTHERN ITALY 7
ifera are interpreted as channel-fill deposits, whereasoarsening-upward successions are interpreted as creplay deposits. Samples displaying relative abundance oPi-ea, Alnus,andBetula,with parallel scarcity ofPinus,indicate
mproving climate conditions. The lower boundary of tacies association is tentatively correlated with the base oastenaso Unit (Amorosi and Farina, 1995), identified aouthern margin of the Po Plain on the basis of boreholeorrelations.
5.00–79.25 m: Marsh Facies Association
This facies association includes grey clays with comeat intercalations. The microfauna consists dominantly orackish ostracodC. torosa.Pollen spectra show remarkaollen concentrations, with AP1 as the dominant group amP, and an abundance ofPoaceae,the high percentageshich could reflect proximity to paludal areas.This association is interpreted to have formed on a co
lain (marsh) during an interglacial or interstadial period.
9.25–87.75 m: Alluvial Plain Facies Association
This facies association consists of silts and fine to medands, forming two distinct coarsening–fining upward sucions, separated by a low mature paleosol. The microfauenerally absent or represented by scattered reworked bnd planktic foraminifera. Rare freshwater ostracods (C. can-ida, Pseudocandona sucki,and P. sarsi) are present at thase of this association. Pollens are scarce or absent.This association formed on an alluvial plain. The t
oarsening–fining upward successions are interpreted to rhannel entrenchment upon levee/floodplain deposits, folloy channel abandonment and return to floodplain deposi
7.75–97.90 m: Subdelta Facies Association
This facies association includes grey clays with comeat intercalations. Sharp-based fine to coarse sand layeommonly encountered and a general upward increase inize and in the abundance of sand intercalations is recohe microfauna consists of a freshwater ostracod fauna,imilar to that of the overlying interval. Pollen spectraainly composed of AP, with equal proportions of AP1P2. Pinus displays very low values. Samples showing vigh percentages ofAlnus (at 89.30 m) and remarkable abuance ofSparganiumtype (at 87.85 m) indicate proximityivers and paludal areas.
This association probably formed in enclosed shallow-wnvironments of an upper delta plain, characterized by quven stagnant waters, in proximity to a distributary chanuring an interglacial or interstadial period. The upwardrease in sand abundance suggests partial filling of inteributary areas by flood waters, leading to formation of mouth bars (subdelta).
Thesse
eeta
ne
g
tal
ms-ishic
ected.
nare
ained.ry
rorl,-is-r
7.90–100.10 m: Lagoonal Facies Association
This facies association, showing gradational lower ander boundaries with the adjacent deposits, consists of greylays with abundant mollusc shells. The brackish ostracoC.orosa is present throughout the interval. Benthic foraminifhat are tolerant of low salinity conditions, such asA. tepida, Aarkinsoniana,and subordinateElphidiumspp. (among whic. granosum granosumis the dominant species), are abund
n the middle part of the interval. The pollen spectrum ofnterval is characterized by high percentages ofChenopodiceae (40%), which are interpreted to reflect proximityarsh environments. Remarkable values of AP1 revearesence of a mixed deciduous broad-leaved vegetatidjacent areas.This association is interpreted to have formed in ei
agoons or bays of a lower delta plain during an interglacianterstadial period. Abundance peaks of benthic foraminifehe middle part of the unit suggest a slightly higher manfluence. Lack of foraminifera at the base and top ofssociation implies typical brackish conditions and is con
ent with vertical transitions to deposits characteristic of frater environments.
00.10–105.35 m: Swamp/Distributary Channel FaciesAssociation
This facies association is characterized by a basal blacayer, which is overlain by grey silts with local peat intercaions and abundant wood fragments. An erosional-basedody, approximately 2.5 m thick, with an internal fininpward tendency, is present at the top of this interval. Frater ostracodsC. neglectaandC. compressacharacterize thner-grained deposits, whereas a scarce reworked micros present in the sands. This interval is characterized bighest values of pollen concentration. Particularly, the popectrum of the black silt layer (105.30 m) shows an aance ofPinus, Artemisia,and NAP and a scarcity of AP.emarkable change in the pollen association, to mixed dous broad-leaved vegetation, characterizes the overlyinosits. The transition is recorded by the sample at 104.5hereBetula,pioneer tree, species attain 20%.This association is interpreted to have formed within
pper delta plain, dominated by paludal areas (swamps)harp-based sand body is likely to reflect distributary chanelated deposits. The pollen spectrum of the basal blacayer is typical for stadial or glacial conditions and diff
arkedly from the underlying (see following sections)verlying deposits, both showing evidence of interglacia
nterstadial climate conditions.
05.35–112.20 m: Beach Ridge Facies Association
This facies association includes very fine to medium sahe microfauna is very similar to that described for the 4
.60-
t pla y Aw
ssib teg
1
at . Tm to2oN -r adr is.P y A( lowvlo get ne
ta?e ndt
1
una dam thal otGst ofd
wis -2 ifera elom ntirc
1
ndw stco a itc
t thb
1
om-m nt int withint thosel ofP ev nlyf nta -a
vel-o ationo bankfi
1
und-a yero end.W mi-c n ofs isp s ofA highv ur-r
er ap er-l tionale mpler ial ors
1
m toc rangedi arep fera,wa u-lCt op
ralice nneld istics.
1
nds,
AMOROSI ET AL.8
o 10.30-m interval. The pollen spectrum from the samvailable indicates a dense forest vegetation dominated bith low values ofPinusand NAP.This association represents beach ridge deposits, po
elonging to a delta front, that accumulated during an inlacial or interstadial period.
12.20–121.60 m: Shallow-Marine Facies Association
This facies association consists of silty sand–silty clayernations, with abundant marine bivalves and gastropods
icrofauna is similar to that described for the 10.30-3.60-m interval, differing for the higher percentages ofMili-lidae spp., Textularia spp., Bulimina elongatagroup, andonionellaspp. and the presence of the ostracodsCarinocythe
eis carinata, Loxoconcha turbida, L. tumida, Cytherettaiatica, Cytheridea neapolitana,and Xestoleberis communollen data indicate a dense forest vegetation dominated b
AP1 and AP2 are in equal proportions), with extremelyalues ofPinusand NAP.QuercusandCarpinus(from low-
ands to lower altitudes) andFagusandAbies(from montain-us zones) are the major components of the arboreal ve
ion. A marked increase ofAbies at 119.45 m indicates apisode of changed climate conditions toward cooling.This association formed in a shallow-marine (prodel
nvironment under interglacial or interstadial climate coions.
21.60–124.20 m: Transgressive Sand Sheet
This facies association, showing an erosional lower bory, is characterized by very fine to fine sands, with abunarine macrofossils. The microfauna is very similar to
isted for the 23.60- to 26.60-m interval, with the exceptionhe higher abundance ofTextulariaspp. andBulimina elongataroup and the lower abundance of the foraminiferaReussella
pinulosaandTrifarina angulosaand the ostracodCytherop-eron monoceros.This interval displays the highest valueseciduousQuercusandQ. ilex-cocciferatype.This association formed on a transgressive shoreline,
lightly deeper waters compared to those of the 23.606.60-m interval, based on the distinctive benthic foraminssemblages. Pollen spectra indicate the maximum devent of thermophilous broad-leaved forests within the e
ore.
24.20–126.70 m: Swamp Facies Association
This facies association consists of clays, silts, and saith abundant woods and plant fragments. Freshwater oods (C. neglecta, P. sucki,andIlyocypris gibba) constitute thenly microfauna present in the interval. Two pollen spectr
his interval show different patterns, with an increase ofQuer-us that is paralleled by a decrease ofPinus.This association formed in swamps of a coastal plain a
eginning of an interglacial or interstadial period.
seP,
blyr-
l-he
-
P
ta-
)i-
d-nttf
thtoap-e
s,ra-
n
e
26.70–137.20 m: Alluvial Plain Facies Association
This facies association includes silts and clays, with con very fine sand intercalations. No microfauna is prese
he sands, whereas freshwater ostracods are abundanthe fine-grained deposits. The species correspond toisted for the 27.90- to 75.00-m interval, with the additionaralimnocytere teneraandScottia tumida.Pollen spectra arery similar to those of the 27.90- to 75.00-m interval. Oew taxa are present, withPinusas the major arboreal elemend herbaceous vegetation dominated byArtemisia, Asterceae,andPoaceae.A few samples yielded no pollen.This association corresponds to an alluvial plain that de
ped under glacial or stadial climate conditions. Sedimentccurred predominantly as levee accumulations, overnes, and swamp deposits.
37.20–145.00 m: Bayhead Delta Facies Association
This facies association, showing a transitional lower bory to the underlying deposits, is composed of a silt laverlain by fine to coarse sands, with a fining-upward trood and plant fragments are common. No macro- and
rofossils are present in this association, with the exceptiocattered reworked foraminifera. A varied pollen floraresent uniquely at 143.80 m, where similar proportionP1 and AP2 are encountered, together with relativelyalues ofPinus.In coincidence of this sample, the last occence ofPterocaryawas noted.
This association formed in a paralic environment undredominant fluvial influence. Vertical transition to the und
ying deposits suggests a bayhead delta as a likely deposinvironment. The pollen spectrum of the analyzed saeflects the transition between warm-temperate and glactadial types of vegetation.
45.00–161.70 m: Estuary Facies Association
This facies association consists predominantly of mediuoarse sands, with subordinate fine sands. These are arn crudely upward-fining successions. No microfossilsresent, with the exception of scattered reworked foraminihereas macrofossils diagnostic of both marine (Mactra cor-llina, Bittium reticulatum, Spisula subtruncata, Hinia retic
ata) and brackish environments (species ofHydrobiidae,erastoderma glaucum) are abundant. The presence ofLen-
idium mediterraneumsuggests proximity to river mouths. Nollen analyses were carried out in this interval.This association formed in a shallow, high-energy pa
nvironment. An estuary, possibly including active chaeposits, could account for the observed facies character
61.70–164.50 m: Lagoon/Bay Facies Association
This facies association is characterized by very fine sahowing traces of parallel- and cross-lamination, with su
bor-
d fercmp nsor eci uof
ctei app unw
1
wic aceT shwt ues
pa
1
gres ves ithh ttE -t e-b ra,Cm ishw hed tror
rine resw orb tho waf tho
nds cos aredg
d and1 ciesc vials and1s
rn off thats ial/i -j d byA cialcd cialp ixedd( rkedb P2).B nc tionc ons,w P.
datap S17( d byA theu o theH orre-s topec
tionw 1.55a rt oft theh rlainb ncet es ofs g top ajorc tra,11 eille,1 stage5 nds withS levelo1 hicf eent
est ac theo .
GLACIO-EUSTATIC CONTROL OF SEDIMENTATION, NORTHERN ITALY 9
inate silts. In the middle part of the association foraminionsist predominantly ofA. parkinsonianaandA. tepida,withinor concentrations ofMiliolidae spp.,Textularia spp.,El-hidium spp., andQuinqueloculinaspp. Scattered specimef Nonion depressulumare also present. AbundantC. torosaisecorded at the base and top of the interval. Pollen spndicate a greater development of thermophilous decidorests relative to mountain forests.
This association reflects a paralic environment, charazed by alternating brackish and low salinity waters, andears to have been deposited in either a lagoon or a bayarm-temperate climate conditions.
64.50–169.00 m: Swamp Facies Association
This facies association includes silt–sand alternations,ommon peat intercalations, wood fragments, and root trhe microfauna of this interval consists uniquely of rare freater ostracods, such asC. neglecta, C. compressa,andScottia
umida.Pollen spectra are characterized by high AP2 valThis association is interpreted to have formed in a swamdelta plain during a cool-temperate period.
69.00–173.40 m: Beach Ridge Facies Association
This facies association consists of very fine to fine darkands with abundant shell fragments. The microfauna isimilar to those of the 161.70- to 164.50-m interval, wigher values ofA. parkinsonianaandA. tepida(up to 91% a
he top of the interval) and lower abundance ofMiliolidae spp.,lphidiumspp.,Rosalina bradyi,andAsterigerinataspp. Os
racods include rare instars ofPontocytere turbida, Xestoleris dispar, Loxoconcha turbida, L. tumida, L. stellifeytheridea neapolitana,andLeptocythere bacescoi.The top-ost part of the interval consists exclusively of the brackater ostracodC. torosa. The pollen spectrum reflects tevelopment of thermophilous deciduous forests and the seduction of mountain arboreal vegetation.
This association formed in a high-energy, shallow-manvironment, probably under considerable influence by faters, during an interglacial or interstadial period. Nearsheach ridge deposits of a delta front could account forbserved characteristics. The occurrence of a brackish
auna at the top of the interval marks the transition toverlying facies association.
AGE OF CORE S17 AND CORRELATION BETWEENFACIES EVOLUTION AND GLOBAL
SEA-LEVEL CHANGES
The distinctive cyclic arrangement of alluvial, paralic, ahallow-marine deposits within core S17 enables the retruction of the patterns of shoreline migration in the studyuring the Late Quaternary. Particularly, three majortrans-ressivepulsations, marked by the onset of marginal ma
rinPa
traus
r--
der
ths.-
.of
yry
-
ng
ehe,etere
n-a
e
eposits, are readily identified in the core (at 4.60–26.6005.35–124.20 m and below 169.00 m, in Fig. 3). These fahanges contrast markedly with the abrupt shifts to alluedimentation (intervals between 27.90 and 75.00 m26.70 and 137.20 m, in Fig. 3), recording two majorregres-ivephases.The close relationship between this cyclic stacking patte
acies and pollen distribution within core S17 suggestsedimentary evolution was mostly controlled by glac
nterglacial fluctuations. Thetransgressiveintervals record maor peaks in pollen concentration and are characterizeP-dominated pollen types, which are typical for interglaonditions (Fig. 3). Within these deposits,Pinus and NAPisplay very low values. The early stages of the interglaeriods were characterized by the abrupt increase in meciduous broad-leaved vegetation, dominated byQuercusAP1 in Fig. 3), whereas the late stages were generally may the progressive increase in mountain belt vegetation (Ay contrast, theregressiveintervals record very low polleoncentrations (indicating a strong reduction in vegetaover) and pollen spectra that are typical for glacial conditiith dominance ofPinusand NAP and scarcity or lack of ARadiometric dates from correlative deposits and pollen
rovide important constraints to determine the age of coreFig. 4). On the basis of the radiocarbon dates reportemorosiet al. (1999), the onset of marine sedimentation inppermost part of the core (4.60–26.60 m) is assigned tolocene (see Fig. 2), i.e., to the transgressive phase cponding to the Stage 2/1 transition in the oxygen-isourve of Martinsonet al. (1987).The AP profile of core S17 shows a distinctive culminaithin coastal and shallow-marine deposits between 11nd 125.88 m, with maximum percentages in the lower pa
his interval (Fig. 4). This culmination, which recordsighest AP values below the Holocene culmination, is ovey two minor oscillations, forming a characteristic seque
hat has been documented from all long-cored pollen seriouthern Europe (Italy, France, and Greece). Accordinrevious work (Fig. 4), the pollen zones including this mulmination should be attributed to the Eemian (Wijms969; Wijmstra and Smit, 1976; Woillard, 1978; Follieriet al.,988; Reille and de Beaulieu, 1990; de Beaulieu and R992; Tzedakis, 1993, 1994) and correlated to isotope Sube (see Tzedakiset al., 1997). The correlation of coastal ahallow-marine deposits between 105.35 and 124.20 mubstage 5e, corresponding to the highest position of seaf the last 260 ka (Aharon and Chappell, 1986; Bardet al.,990; Pirazzoli, 1993), is fully consistent with the stratigrap
ramework, showing no intervening marine deposits betwhis stratigraphic interval and the Holocene deposits.
Using the above data as tie points, it is possible to suggorrelation between facies distribution within core S17 andxygen-isotope curve of Martinsonet al. (1987) (Fig. 4)articularly, the minor transgressive phases marked in the
core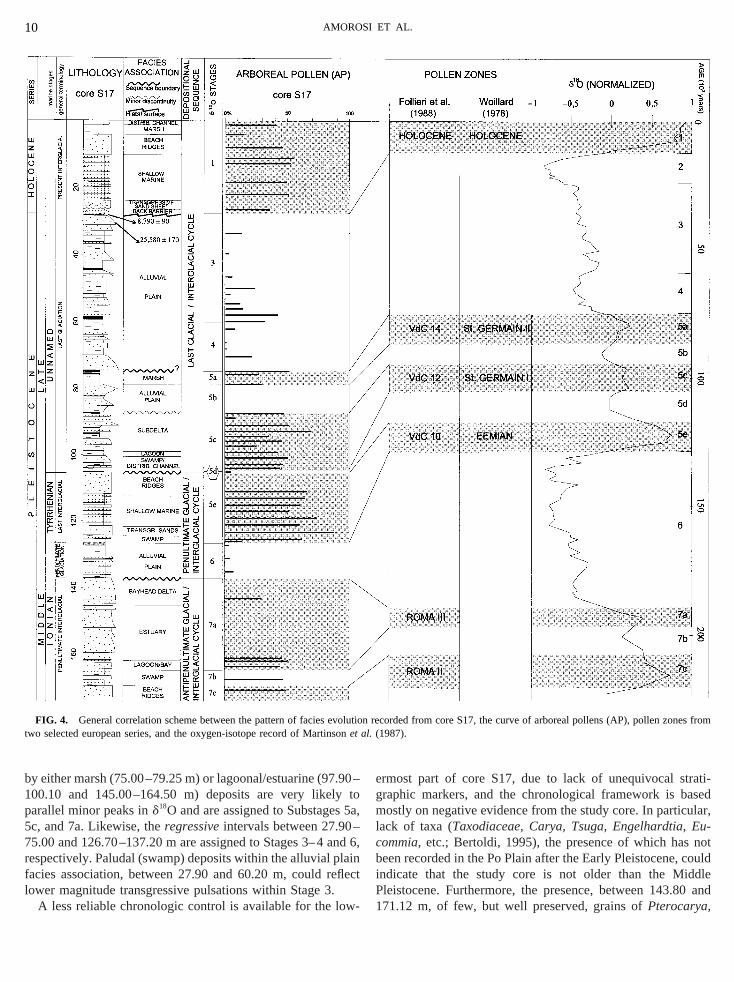
b .901 y tp 5a5 –7 anr laif eflel
w-
e rati-g asedm ular,l Eu-c notb couldi dleP 0 and1 ,
zones fromt
AMOROSI ET AL.10
y either marsh (75.00–79.25 m) or lagoonal/estuarine (9700.10 and 145.00–164.50 m) deposits are very likelarallel minor peaks ind18O and are assigned to Substagesc, and 7a. Likewise, theregressiveintervals between 27.905.00 and 126.70–137.20 m are assigned to Stages 3–4espectively. Paludal (swamp) deposits within the alluvial pacies association, between 27.90 and 60.20 m, could rower magnitude transgressive pulsations within Stage 3.
A less reliable chronologic control is available for the lo
FIG. 4. General correlation scheme between the pattern of facies evwo selected european series, and the oxygen-isotope record of Martinet
–o,
d 6,nct
rmost part of core S17, due to lack of unequivocal straphic markers, and the chronological framework is bostly on negative evidence from the study core. In partic
ack of taxa (Taxodiaceae, Carya, Tsuga, Engelhardtia,ommia,etc.; Bertoldi, 1995), the presence of which haseen recorded in the Po Plain after the Early Pleistocene,
ndicate that the study core is not older than the Midleistocene. Furthermore, the presence, between 143.871.12 m, of few, but well preserved, grains ofPterocarya
tion recorded from core S17, the curve of arboreal pollens (AP), pollen1987).
olusonal. (

w n rcF( thes comb ssie lda e7 rapi
ost ciai epos thes
es nct ndc edm thf ck( hes de( ro-g stet
l cyc cet da( br uoo s dt lds inc ba( ndG ntap tho wit ciag Suse ita
5dm ciew atv ssS revo1
h yclei ansi-t tives ccu-m tages5 essivep Stage3 king( het bed atl ge1
ateg clec e 7/6t vialp low-m pretedt
topeS ly, ar near-s ther ver-l eltaf ingh
m isi mateg uldb trends llow-m tandp
nnaa SEP Thep topeS
(fo-r yclicp andp vals( ecordo als( hift toc
llena nated
GLACIO-EUSTATIC CONTROL OF SEDIMENTATION, NORTHERN ITALY 11
hose presence in Latium in similar amounts has beeorded up to the Roma II forest phase (Follieriet al.,1988, inig. 4), with an estimated age of 187–1953 103 yr B.P.Follieri et al., 1989), could restrict the maximum age oftudy units to the late Middle Pleistocene. These data,ined with stratigraphic position, suggest that the transgrevent recorded by marine deposits below 169.00 m coussigned to the next maximum ind18O (Fig. 4), i.e., Substagc. This implies that core S17 presumedly extends stratig
cally down to approximately Substage 7c.
SEQUENCE STRATIGRAPHY
In terms of sequence stratigraphy, two fourth-order depional sequences (corresponding to the past two glanterglacial cycles) and the upper part of an underlying ditional sequence are present in the study core. Each ofequences spans an interval of approximately 105 years.The last (post-Substage 5e) glacial/interglacial cycl
trongly asymmetric, with a long period of sea-level fall (puuated by higher frequency relative sea-level cycles) aomparatively short interval of sea-level rise (Fig. 4). Sentation during relative sea-level falls corresponds to
orced regressive systems tracts (FST) of Hunt and Tu1992) and Posamentieret al.(1992). These are overlain by tuiteof systems tracts defined in the original sequence moPosamentieret al.,1988), corresponding to the lowstand prading wedge systems tract (LST), the transgressive sy
ract (TST), and the highstand systems tract (HST).The boundary between the past two glacial/interglacia
les corresponds to the abrupt sea-level fall that took plahe end of the Eemian (Fig. 4). Locating the sequence bounSB) within a strongly asymmetric cycle, characterizedepeated phases of falling sea level, is not an unambigperation, since more than one unconformable surface i
ected in the core (Fig. 3) and each of these surfaces coutrictly regarded as a potential SB. In theoretical modelsluding the FST, the SB has been placed either at thePosamentieret al. (1992) or at the top (Helland-Hansen ajelberg, 1994) of the FST. Although this is not a fundameoint of our discussion, in the case of core S17 we followriginal sequence model and place the SB in coincidence
he discontinuity surface developed at the last interglalacial transition, i.e., at the boundary between isotopetages 5e and 5d of the curve of Martinsonet al. (1987). Anqually logical solution is represented by the unconformssigned to the Stage 5/4 transition.The transition between isotope Substages 5e andarked in the study core by a sharp basinward shift of fahich is paralleled by the abrupt onset of a cold climegetation (sample at 105.30 m). The very reduced thickneubstage 5d deposits within core S17 is consistent with pus work from several parts of Europe (see Tzedakiset al.,997), suggesting the presence of a generalized stratigr
apbe-
-vebe
h-
i-l/-se
is-a
i-eer
ls
ms
-atry
yuse-be-se
lethl/b-
y
iss,eofi-
hic
iatus at the SB. The FST of the last glacial/interglacial cncludes the deposits that formed from Substage 5e/5d trion to Stage 3/2 transition (Fig. 3). Higher frequency relaea-level cycles within the FST include the deposits that aulated during the various steps of the falling stage (Subsd and 5b and Stage 4) and the subsequent minor transgrhases (Substages 5c and 5a and peak transgressions in). Deposits from the last glacial interval (Stage 2) are lacAmorosiet al.,1999); this implies that the LST is absent. Transgressive–regressive wedge of Holocene age, descriength by Amorosiet al.(1999) and corresponding to the Sta, includes the TST and the HST (Fig. 3).A similar facies architecture characterizes the penultim
lacial/interglacial cycle. The lower boundary of this cyoincides with the major sea-level fall recorded at the Stagransition that caused the abrupt shift from paralic to allulain sedimentation (FST and, possibly, LST). The shalarine deposits assigned to Substage 5e (Fig. 3) are inter
o correspond to the TST and the HST.Facies architecture of marine deposits related to iso
tages 5e and 1 displays very similar trends. Particularetrogradational stacking pattern of transgressive paralic,hore, and shallow-marine deposits, possibly reflectingapid landward migration of barrier–lagoon systems, is oain by the progradational stacking pattern of prodelta, dront, and delta plain sediments, formed during the followighstands.The stratigraphic interval between 137.20 and 173.40
nterpreted to represent the upper part of the antipenultilacial/interglacial cycle, the lower boundary of which shoe placed at the Stage 9/8 transition. The progradationaluggested by superposition of paralic deposits above shaarine sands is consistent with deposition during a highshase (Fig. 3).
CONCLUSIONS
The detailed study of a 173-m-thick core from the Raverea reveals for the first time the depositional history of theo Plain over the past three glacial/interglacial cycles.ollen records for the study core date back to oxygen isotage 7.
(1) Detailed sedimentological and micropaleontologicalaminifera and ostracods) investigations show a vertical cattern of facies, including the alternation of continentalaralic/shallow-marine deposits. Particularly, glacial interStages 6, 4, and 3) are characterized by a discontinuous rf alluvial plain deposits. In contrast, interglacial intervStage 7, Substages 5e, and Stage 1) reflect the abrupt soastal, shallow-marine, and deltaic sedimentation.(2) Facies architecture is paralleled by distinctive po
ssociations, showing the alternation of a vegetation domiy Pinusand NAP, characteristic of glacial and stadial in
ter-
v oped
nr orrs thu poi cleR mag tas
iliaR fun( plec d SP
A eser th
A latthe
A tartal p
A an.rk/
B edpa
B G.Ad
B thiclta(R
,
B nino
d ran
D her
F y &
F rom
F
H vari--
H hook
H forced-
J riatic
J inci-
L
M andice
tigra-
M Orbe-o-
M 2nd
N hy atocenen”260.ralo-
P ment.
P ls onndall,2, pp.s and
P orcedles andm
R erra-A.,
R pperance).
S f thea
S sin in
S
S
AMOROSI ET AL.12
als, with mixed deciduous broad-leaved forests, develuring interglacial and interstadial periods.(3) In terms of sequence stratigraphy, the study units ca
elated to two fourth-order depositional sequences (cponding to the past two glacial/interglacial cycles) and topper part of an underlying sequence. Falling sea-level de
ts constitute the bulk of the last glacial–interglacial cyetrogradational to progradational stacking patterns ofinal marine deposits form the transgressive and highsystems tracts, respectively.
ACKNOWLEDGMENTS
This work was supported by the Geological Survey of the Regione Emomagna (thanks are due to R. Pignone) and COFIN. MURST 1997
coordinator: D. Rio). We are grateful to S. C. Vaiani for help in samollection. The paper benefited from careful reviews by G. Orombelli anorter.
REFERENCES
haron, P., and Chappell, J. (1986). Oxygen isotopes, sea level changthe temperature history of a coral reef environment in New Guinea ovlast 105 years. Palaeogeography, Palaeoclimatology, Palaeoecology56,337–379.
morosi, A., and Farina, M. (1995). Large-scale architecture of a thrust-realluvial complex from subsurface data: The Quaternary succession ofBasin in the Bologna area (northern Italy).Giornale di Geologia57, 3–16.
morosi, A., Colalongo, M. L., Pasini, G., and Preti, D. (1999). Sedimenresponse to Late Quaternary sea-level changes in the Romagna coas(northern Italy).Sedimentology46, 99–121.
thersuch, J., Horne, D. J., and Whittaker, J. E. (1989). MarineBrackish Water Ostracods.In “Synopses of the British Fauna” (D. MKermack and R. S. K. Barnes, Eds.), Vol. 43. Brill, Leiden/New YoKopenaghen/Ko¨ ln.
ard, E., Hamelin, B., and Fairbanks, R. G. (1990). U-Th ages obtainmass spectrometry in corals from Barbados: Sea level during the130000 years.Nature346,456–458.
armawidjaja, D. M., Jorissen, F. J., Puskaric, S., and Van der Zwaan,(1992). Microhabitat selection by benthic foraminifera in the northernriatic Sea.Journal of Foraminiferal Research22, 297–317.
ellotti, P., Carboni, M. G., Di Bella, L., and Palagi, I. (1994). Benforaminiferal assemblages in the depositional sequence of the Tiber DeIn“Studies on Ecology and Palaeoecology of Benthic Communities”Matteucci et al., Eds.), Bollettino della Societa` Paleontologica ItalianaSpecial Volume2, pp. 29–40.
ertoldi, R. (1995). Palinostratigrafia dei bacini intermontani dell’Appennordorientale.Il Quaternario 8, 61–70.
e Beaulieu, J. J., and Reille, M. (1992). The last climatic cycle at La GPile (Vosges, France): A new pollen profile.Quaternary Science Review11,431–438.
evoto, G. (1965). Lacustrine Pleistocene in the Lower Liri Valley (SoutLatium). Geologica Romana4, 291–368.
aegri, K., and Iversen, J. (1992). “Text Book in Pollen Analysis.” WileSons, Chichester.
ollieri, M., Magri, D., and Sadori, L. (1988). 250,000-year pollen record fValle di Castiglione (Roma).Pollen et Spores30, 329–356.
ollieri, M., Magri, D., and Sadori, L. (1989). Pollen stratigraphical s
yn-d
bee-es-
.r-nd
-ds
.
ande
edPo
ylain
d
byst
J.-
..
d
n
thesis from Valle di Castiglione (Roma).Quaternary International3/4,81– 84.
elland-Hansen, W., and Gjelberg, J. G. (1994). Conceptual basis andability in sequence stratigraphy: A different perspective.Sedimentary Geology 92, 31–52.
enderson, P. A. (1990). Freshwater ostracods.In “Synopses of the BritisFauna” (D. M. Kermack and R. S. K. Barnes, Eds.), Vol. 42. Universal BServices, Dr. W. Backhuys, Oegstgeest, The Netherlands.
unt, D., and Tucker, M. E. (1992). Stranded parasequences and theregressive wedge systems tract: Deposition during base-level fall.Sedimentary Geology81, 1–9.
orissen, F. J. (1987). The distribution of benthic foraminifera in the AdSea.Marine Micropalaeontology12(1), 21–48.
orissen, F. J. (1988). Benthic foraminifera from the Adriatic Sea: Prples of phenotypic variation.Utrecht Micropaleontological Bulletins37,1–176.
anger, M. R. (1993). Epiphytic foraminifera.Marine Micropalaeontology20,235–265.
artinson, D. G., Pisias, N. G., Hayes, J. D., Imbrie, J., Moore, T. C.,Shackleton, N. J. (1987). Age dating and the orbital theory of theages—Development of a high-resolution 0 to 300,000 year chronostraphy. Quaternary Research27, 1–29.
ontenegro, M. E., and Pugliese, N. (1995). Ostracodi della laguna ditello: Tolleranza ed opportunismo.Atti del Museo di Geologia e Paleontlogia di Monfalcone, Quaderno speciale3, 71–80.
oore, P. D., Webb, J. A., and Collinson, M. E. (1992). “Pollen Analysis,”ed. Blackwell Sci., Oxford.
ummedal, D., and Swift, D. J. P. (1987). Transgressive stratigrapsequence-bounding unconformities: Some principles derived from Holand Cretaceous examples.In “Sea-Level Fluctuation and Coastal Evolutio(D. Nummedal, O. H. Pilkey, and J. D. Howard, Eds.), Vol. 41, pp. 241–Special Publication of Society of Economic Paleontologists and Minegists.
irazzoli, P. A. (1993). Global sea-level changes and their measureGlobal and Planetary Change8, 135–148.
osamentier, H. W., Jervey, M. T., and Vail, P. R. (1988). Eustatic controclastic deposition. I. Conceptual framework.In “Sea Level Changes: AIntegrated Approach” (C. K. Wilgus, B. S. Hastings, C. G. St. C. KenH. W. Posamentier, C. A. Ross, and J. C. Van Wagoner, Eds.), Vol. 4109–124. Special Publication of Society of Economic PaleontologistMineralogists.
osamentier, H. W., Allen, G. P., James, D. P., and Tesson, M. (1992). Fregressions in a sequence stratigraphic framework: Concepts, exampsequence stratigraphic significance.American Association of PetroleuGeologists Bulletin76, 1687–1709.
egione Emilia-Romagna and ENI-AGIP (1998). Riserve idriche sottnee della Regione Emilia-Romagna. A cura di G. Di Dio. S.EL.CFirenze.
eille, M., and de Beaulieu, J.-L. (1990). Pollen analysis of a long uPleistocene continental sequence in a Velay maar (Massif Central, FrPalaeogeography, Palaeoclimatology, Palaeoecology80, 35–48.
garrella, F., and Moncharmont Zei, M. (1993). Benthic Foraminifera oGulf of Naples (Italy): Systematics and autoecology.Bollettino della Societ̀Paleontologica Italiana32, 145–264.
ocac, A. (1978). Pleistocene ostracode fauna of the Pannonian baCroatia.Palaeontologica Jugoslavica20, 1–51.
tockmarr, J. (1971). Tablets with spores in absolute pollen analysis.Pollen etSpores13(4), 615–621.
wift, D. J. P. (1968). Coastal erosion and transgressive stratigraphy.Journal
of Geology76, 444–456.
T rou
T yclee
T , M.ans oe
V rivea”gica
V alca
, pp.
V e Po
V aneanom-
W ction
W –78.
W for
GLACIO-EUSTATIC CONTROL OF SEDIMENTATION, NORTHERN ITALY 13
zedakis, P. C. (1993). Long-term tree population in northwest Greece thmultiple Quaternary climatic cycles.Nature364,437–440.
zedakis, P. C. (1994). Vegetation change through glacial-interglacial cA long pollen sequence perspective.Philosophical Transactions of thRoyal Society of London, B345,403–432.
zedakis, P. C., Andrieu, V., de Beaulieu, J.-L., Crowhurst, S., FollieriHooghiemstra, H., Magri, D., Reille, M., Sadori, L., Shackleton, N. J.,Wijmstra, T. A. (1997). Comparison of terrestrial and marine recordchanging climate of the last 500,000 years.Earth and Planetary SciencLetters150,171–176.
an der Zwaan, G. J., and Jorissen, F. J. (1991). Biofacial patterns ininduced shelf anoxia.In “Modern and Ancient Continental Shelf Anoxi(R. V. Tyson and T. H. Pearson, Eds.), Vol. 58, pp. 65–82. GeoloSociety Special Publication.
an Frausum, A., and Wouters, K. (1990). Ostracoda from Holocene c
eous tufa deposits in southern Belgium: A palaeoenvironmental analysis.Ingh
s:
,df
r-
l
r-
“Ostracoda and Global Events” (R. Whatley and C. Maybury, Eds.)505–514. Chapman & Hall, London/New York.
eggiani, A. (1974). Le variazioni idrografiche del basso corso del fiumnegli ultimi 3000 anni. Padusa, 1–2, Rovigo, 39–60.
erhallen, P. J. J. M. (1991). Late Pliocene to early Pleistocene Mediterrmud-dwelling foraminifera: Influence of a changing environment on cmunity structure and evolution.Utrecht Micropaleontological Bulletins40,1–220.
ijmstra, T. A. (1969). Palynology of the first 30 m of a 120 m deep sein Northern Greece.Acta Botanica Neerlandica18, 511–527.
ijmstra, T. A., and Smit, A. (1976). Palynology of the middle part (30metres) of the 120 m deep section in Northern Greece (Macedonia)ActaBotanica Neerlandica25, 297–312.
oillard, G. M. (1978). Grand Pile peat bog: A continuous pollen recordthe last 140.000 years.Quaternary Research9, 1–21.




















