
given in figure 1. The respiration without substrat - NCBI
11
THE OXIDATION OF GLYCEROL BY STREPTOCOCCUS FAECALIS I. C. GUNSALUS AND W. W. UMBREIT Laboratory of Bacteriology, College of Agriculture, Cornell University, Ithaca, New York' Received for publication October 30, 1944 It has been reported that certain of the lactic acid bacteria are capable of utilizing glycerol only under aerobic conditions (Gunsalus and Sherman, 1943). Inasmuch as these organisms are devoid of cytochrome (Frei et al., 1934; Farrell, 1935) and are reported to produce hydrogen peroxide during respiration, the presence of a functional respiration has been questioned. It therefore became of some importance to determine the nature of the reactions involved in the utilization of glycerol by these forms. METHODS The culture used, strain 24 from the departmental culture collection, had pre- viously been identified as Streptococcus faecalis, Lancefield serological group D. It was shown to utilize glycerol only under aerobic conditions (Gunsalus and Sherman, 1943) and to derive energy from this process. (Certain other strains are capable of using glycerol under both aerobic and anaerobic conditions.) Active cell suspensions can be prepared under the conditions suggested for the group B streptococci (Wood and Gunsalus, 1942). Suspensions so prepared (usually from a 1 per cent tryptone, 1 per cent yeast extract, 0.5 per cent K2H1P04, and 0.1 per cent glucose medium, the cells being washed with phosphate or saline) could be employed immediately in the respiration experiments or could be stored for 2 or 3 days at refrigerator temperatures. In storage the oxidative powers were gradually lost, but the loss in activity was not comparable for all substrates tested. The glucose concentration in the growth medium could not be increased (to 0.3 per cent, for example) without marked decrease in the oxidative power of the cells, especially on such substrates as glycerol and other alcohols. There was no evidence of adaptation to glycerol. Active cell preparations could be obtained by drying washed cells in vacuo over anhydrous calcium sulfate (dri- erite). The conventional Warburg techniques were used throughout. EXPERIMENTAL Data on the oxygen uptake of a cell suspension refrigerated for several hours are given in figure 1. The respiration without substrate was negligible (Qo,(N) = 10); lactate and pyruvate were not oxidized at an appreciable rate, whereas glucose was oxidized slowly (Qo2(N) -74). With glycerol, however, oxygen was consumed at a rapid rate (Qo,(N) 420) until 1 mole was used per mole of glycerol. Thus these streptococci possess a powerful respiratory system acting upon glycerol. A Qo,(N) of 600 to 700 is not unusual. 1 The authors are indebted to the members of the Department of Agricultural Bacteriol- ogy, University of Wisconsin, where preliminary work on this problem was done. 347
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of given in figure 1. The respiration without substrat - NCBI

THE OXIDATION OF GLYCEROL BY STREPTOCOCCUS FAECALIS
I. C. GUNSALUS AND W. W. UMBREIT
Laboratory of Bacteriology, College of Agriculture, Cornell University, Ithaca, New York'
Received for publication October 30, 1944
It has been reported that certain of the lactic acid bacteria are capable ofutilizing glycerol only under aerobic conditions (Gunsalus and Sherman, 1943).Inasmuch as these organisms are devoid of cytochrome (Frei et al., 1934; Farrell,1935) and are reported to produce hydrogen peroxide during respiration, thepresence of a functional respiration has been questioned. It therefore becameof some importance to determine the nature of the reactions involved in theutilization of glycerol by these forms.
METHODS
The culture used, strain 24 from the departmental culture collection, had pre-viously been identified as Streptococcus faecalis, Lancefield serological group D.It was shown to utilize glycerol only under aerobic conditions (Gunsalus andSherman, 1943) and to derive energy from this process. (Certain other strainsare capable of using glycerol under both aerobic and anaerobic conditions.)
Active cell suspensions can be prepared under the conditions suggested forthe group B streptococci (Wood and Gunsalus, 1942). Suspensions so prepared(usually from a 1 per cent tryptone, 1 per cent yeast extract, 0.5 per cent K2H1P04,and 0.1 per cent glucose medium, the cells being washed with phosphate or saline)could be employed immediately in the respiration experiments or could be storedfor 2 or 3 days at refrigerator temperatures. In storage the oxidative powerswere gradually lost, but the loss in activity was not comparable for all substratestested. The glucose concentration in the growth medium could not be increased(to 0.3 per cent, for example) without marked decrease in the oxidative powerof the cells, especially on such substrates as glycerol and other alcohols. Therewas no evidence of adaptation to glycerol. Active cell preparations could beobtained by drying washed cells in vacuo over anhydrous calcium sulfate (dri-erite). The conventional Warburg techniques were used throughout.
EXPERIMENTAL
Data on the oxygen uptake of a cell suspension refrigerated for several hours aregiven in figure 1. The respiration without substrate was negligible (Qo,(N) =
10); lactate and pyruvate were not oxidized at an appreciable rate, whereasglucose was oxidized slowly (Qo2(N) -74). With glycerol, however, oxygenwas consumed at a rapid rate (Qo,(N) 420) until 1 mole was used per mole ofglycerol. Thus these streptococci possess a powerful respiratory system actingupon glycerol. A Qo,(N) of 600 to 700 is not unusual.
1 The authors are indebted to the members of the Department of Agricultural Bacteriol-ogy, University of Wisconsin, where preliminary work on this problem was done.
347

348 I. C. GUNSALUS AND 'X. WI. UMBRFIT
Flydrogen Peroxide Formation
The data of figure 1 show that 1 oxygen is utilized per glycerol. The productsof the reaction were therefore studied. When higher levels of glycerol were added(figure 2), only a partial oxidation occurred. For example, when 2.5, 5, and 10micromoles of glycerol were supplied, the oxygen used per mole of glycerol was,respectively, 0.97, 1.01, and 0.67 moles. The low value at 10 micromoles couldbe due to the accumulation of toxic amounts of hydrogen peroxide which thenwould form one of the products of the reaction.
LSw
zw0
x0
U,0:w
-J0C)cr
1 0 2/GYE _ - - '5ALM GLYCEROL
GLCROL _-
60
40
211-10 5A^M GLUCOSESUBSTR ATE A M PYRUVATEADDED 5 M LACTATE
0 __NDOG_.EN9U I0 10 20 30 40 50
MINIUTESFIG. 1. OXIDATIONS BY STREPTOCOCCUS FAECALIS, STRAIN 24
0.650 mg bacterial nitrogen per cup.M/30 phosphate buffer pH 7.0.Total volume 3.0 ml + 0.2 ml 20% KOH in center well.Temperature 37 C.
The occurrence of hydrogen peroxide was tested for in three ways.First, by addition of catalase which would decompose the peroxide formed and
thus should permit the complete oxidation of 10 or more micromoles of glycerolwith an oxygen uptake of 2 mole of oxygen per mole of glycerol. In the presenceof crystalline catalase, 1 part per 100,000 (Sumner and Dounce, 1939),2 10micromoles of glycerol were oxidized with an oxygen uptake of 138 JAI (0.6102/glycerol), whereas 20 micromoles took up 223 IAI (0.50 02/glycerol) (curves Aand C, figure 3). The addition of catalase alone had no effect upon the endoge-nous respiration.
2 We are indebted to Professor J. B. Sumner for the crystalline catalase employed.

OXIDATION OF GLYCEROL BY STREPTOCOCCUS FAECALIS
Second, by increasing the volume of fluid in the Warburg flask to 4 ml insteadof the usual 3. Thus one-third more oxygen should be taken up before theconcentration of hydrogen peroxide reached an inhibitory level. On additionof 10 micromoles of glycerol to a flask with 3 ml of fluid, 138 ,ud of 02 were used(46 per ml), whereas in a flask with 4 ml of fluid, 183 Al of 02 were used (46per ml). This would indicate that the concentration of hydrogen peroxide whichinhibits the oxidation of glycerol is close to 2 micromoles per ml. The concen-tration of peroxide necessary to inhibit glycerol oxidation varies slightly fromone suspension to the next.
2401
a
.C
x
:(A~it
200J~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~1OoM GLYCEROL____ t(.67 0 /MOLE)L160
> t20 / 5A1M GLYCEROL-1.01 02/MOLE)
Ij,/tv />/2.5,4M GLYCEROL
40 211-A ENDOGENOUS0 15 30 45 60 75
MINUTESFIG. 2. GLYCEROL OXIDATION BY STREPTOCOCCUS FAECALIS, STRAIN 24
0.45 mg bacterial nitrogen per cup.Other conditions as in figure 1.
Third, by the addition of pyruvate which is chemically oxidized by hydrogenperoxide to acetic acid and carbon dioxide (Sevag, 1932). This removal ofperoxide would allow the oxidation of higher levels of glycerol. In the presenceof 5 micromoles of pyruvate, 10 micromoles of glycerol were oxidized with theutilization of 1.02 moles of oxygen per glycerol (curve B, figure 3). This quan-tity of pyruvate was not sufficient to remove all of the peroxide formed, but didreduce its concentration below the toxic level. That pyruvate did not enter thereaction except to remove the peroxide is illustrated by the data in curves Dand E, figure 3, which show that virtually identical amounts of oxygen weretaken up by 5 micromoles of glycerol in the presence or absence of pyruvate.
Thus, one of the products of glycerol oxidation is hydrogen peroxide, whichaccumulates and inhibits the oxidation when its concentration reaches approxi-mately 0.002 molar.
349
i

I. C. GUNSALUS AND W. W. UMBREIT
Acid Production
The other product or products of oxidation were somewhat more difficult todetermine. When glycerol was oxidized in Ringer's bicarbonate buffer withan atmosphere of 5 per cent CO2 in air, one mole of acid (or acid plus carbondioxide) was obtained per mole of glycerol. Only traces of carbon dioxide assuch resulted from the oxidation. The lactic acid found, upon analysis of theflask contents by the method of Barker and Summerson (1941), accounted foronly part of the acid formed. Later it became apparent that lactic acid wasindeed the other product of the reaction, although in some suspensions the lactate
-240 | A 220,ULM GLYCERI ~~~+GATALASE
10,^4.M GLYCEROL4//DMPYR1JVKF
awCl)D
z
w
X0
Cf)(rw
:3i0
0
1601 -
20
80
40 _0 1 1 1 1~~~~~~~~~~~~~~~
0 15 30 45 60 75MINUTES
FIG. 3. GLYCEROL OXIDATION WITH REMOVAL OF HYDROGEN PEROXIDE BYPYRUVATE OR CATALASE
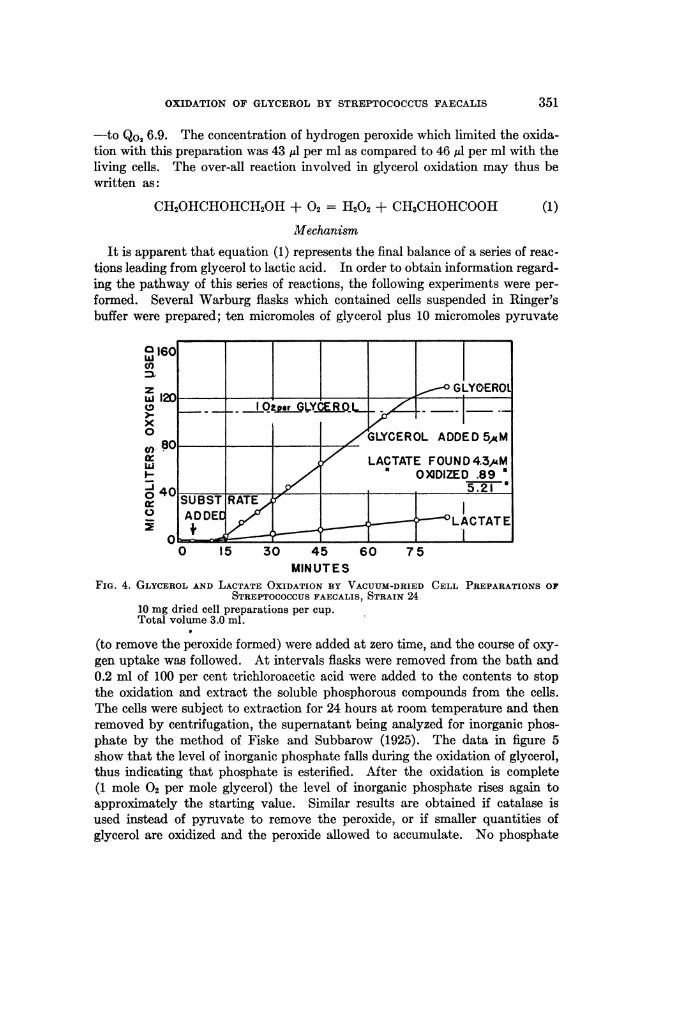
was further utilized (or its formation prevented) by reactions not concerned di-rectly with glycerol oxidation. Recourse was then had to cell preparations inwhich some of these side reactions could be eliminated. Data upon the oxida-tion of glycerol with a dried cell preparation are given in figure 4. This prepara-
tion oxidized glycerol to slightly beyond the theoretical uptake of one 02 per
glycerol, but was also capable of oxidizing lactate slightly. The amount oflactic acid found upon analysis, plus the amount of lactate oxidized in an equiva-lent time, entirely accounts for the glycerol added. With this preparation therate of glycerol oxidation was greatly decreased in comparison with the livingcell (i.e., Qo2 10.3 for the preparation as compared to values of 50 to 70 for livingcells). The rate of oxidation was reduced still further by the addition of catalase
// - IOAM GLYCEROLC 5 CATALASE
D oM GLYCEROL___ >t4-5,^M PYRUVrEEY^ 5,M GLYCEROL
zoo I x T.
350)
v --"J'Am VI nv VfAi u-
OXIDATION OF GLYCEROL BY STREPTOCOCCUS FAECALIS
-to Qo2 6.9. The concentration of hydrogen peroxide which limited the oxida-tion with this preparation was 43 ,ul per ml as compared to 46 ,l per ml with theliving cells. The over-all reaction involved in glycerol oxidation may thus bewritten as:
CH20HCHOHCH20H + 02 = H202 + CH3CHOHCOOH (1)
MechanismIt is apparent that equation (1) represents the final balance of a series of reac-
tions leading from glycerol to lactic acid. In order to obtain information regard-ing the pathway of this series of reactions, the following experiments were per-
formed. Several Warburg flasks which contained cells suspended in Ringer'sbuffer were prepared; ten micromoles of glycerol plus 10 micromoles pyruvate
Co.160 1
z _ GLY_EROLo - j.Q~1zoerGL.YSQEPQ - ___
0 GLYCEROL ADDED 5,AM)00
LACTATE FOUND 4.3AMI- OX~~~~~~~~0QIDZED .89
_J 40 5_______ 5.210 SUBST RATEo ADDE LACTATE
0 15 30 45 60 75MINUTES
FIG. 4. GLYCEROL AND LACTATE OXIDATION BY VACUUM-DRIED CELL PREPARATIONS OFSTREPTOCOCCUS FAECALIS, STRAIN 24
10 mg dried cell preparations per cup.Total volume 3.0 ml.
(to remove the peroxide formed) were added at zero time, and the course of oxy-gen uptake was followed. At intervals flasks were removed from the bath and0.2 ml of 100 per cent trichloroacetic acid were added to the contents to stopthe oxidation and extract the soluble phosphorous compounds from the cells.The cells were subject to extraction for 24 hours at room temperature and thenremoved by centrifugation, the supernatant being analyzed for inorganic phos-phate by the method of Fiske and Subbarow (1925). The data in figure 5show that the level of inorganic phosphate falls during the oxidation of glycerol,thus indicating that phosphate is esterified. After the oxidation is complete(1 mole 02 per mole glycerol) the level of inorganic phosphate rises again toapproximately the starting value. Similar results are obtained if catalase isused instead of pyruvate to remove the peroxide, or if smaller quantities ofglycerol are oxidized and the peroxide allowed to accumulate. No phosphate
351

I. C. GUNSALUS AND W. W. UMBREIT
is taken up by cells respiring endogenously. The cell preparations used infigure 4 also show a phosphate uptake. In this case 0.13 ,u mole of phosphate wasesterified during glycerol oxidation, whereas 0.22 jrmole was released duringthe same interval in the presence of lactate. Therefore, the intermediate reac-tions involve phosphorylation.
In the figures so far provided it will be noted that a lag of approximately sevenminutes occurs from the time glycerol is added until the oxidation begins at ameasurable, and maximal, rate. This suggests that the glycerol is convertedinto some other substance before oxidation. From the phosphate uptake one
240
202 0 X 7 =0 0
Zot~ ~~~~~~~~~~1oOI1 2 0 0 50 6 1300
z w
o ~*-PHOSPHATEci)zH~~~~~~~~~~~
0
40 -P
0 IC 20 30 40 50 60 100MINUTES
FIG. 5. PHOSPHOROUS UPTAKE DURING GLYCEROI, OXIDATION0.9 mg bacterial nitrogen.10 4 moles glycerol (224 ,ul).10 moles pyruvate, to remove H202.Ringer's solution pH 7.5.
would suspect the conversion of glycerol into glycer ol phosphates before oxida-tion, especially since glycerol-phosphate-oxidizing enzymes are known (Green,1936; Euler et al., 1937). We therefore tested purified commercial glycerolphosphate, consisting of 48 per cent A-, and 52 per cent a-glycerol-phosphate.This mixture, however, was not oxidized at an appreciable rate by the living cell.As phosphate esters generally do not enter living cells, the ability of the driedcell preparation to oxidize the glycerol phosphates was also tested. The resultswere again negative. This fact seemed to exclude the glycerol phosphates asintermediates in the reaction. However, upon further study, two remainingpossibilities became apparent: first, that the drying process had not destroyed
352

OXIDATION OF GLYCEROL BY STREPTOCOCCUS FAECALIS
the permeability properties of the cell membrane; and, second, that the presenceof the glycerol phosphates, other than the form oxidized (presumably the1-(-)-a; see Baer and Fisher, 1939), inhibited the oxidation of the active form.
Therefore, to avoid permeability problems, studies were made with the cellpreparations. A number of treatments applied to dried cells w\Nere not suitableinasmuch as they either had no effect upon the preparation, or inactivated itentirely. For example, treatment with acetone, or washing with M/15 phosphateat pH 5, 7, or 8 did not alter the activity. Autolysis of the preparation at 37 Cfor 24 hours (after which time very few cells remained) permitted the oxidationof the glycerol phosphates at a measurable rate (Qo2 = 1.28), but the prepara-tion also oxidized glycerol (Qo2 = 1.43). Finally, preparations capable ofoxidizing glycerol phosphate were obtained by grinding dried cells in vacuo in amicro ball mill.
TABLE 1Effect of varied treatments on the ability of vacuum-dried cells of Streptococcus faecalis (no. 24)
to oxidize glycerol and glycerol phosphates
TREATMENTG OL GLYCEROL REMARKSGLCRLPHOSPHATES
Living cells........................... 42.0 0.1Same cells dried in vacuo............. 11.7 1.14 All intact cells, all gram-
positive
Ground* {in air...................... 0 0 A few cells left, but mostlyin vacuo........ 0.55 0.36 gram-negative debris
Groundt with yeast extract {fin air.... 6.9 1.18 A few cells left, but mostlyin vacuo.. 4.6 5.5 gram-negative debris
* Ground 18 hours in micro ball mill.t Ground with an equal weight of yeast extract.
Typical data on the cell preparations obtained by this procedure are givenin table 1. The original dried cell preparation oxidizes glycerol at a rapid rate,but glycerol phosphate very slowly, if at all. By grinding with yeast extractin vacuo, the rate of glycerol oxidation may be decreased to 40 per cent of theoriginal value for dried cells, whereas the oxidation of glycerol phosphate isincreased almost 5 times.With most of the preparations employed it was necessary to have the yeast
extract present during the grinding process. The addition of an equivalentquantity of yeast extract to the preparation after grinding did not restore theactivity. Presumably, the yeast extract plays some part in the grinding processand is not merely a source of diffusible cofactors. An occasional preparationground in vacuo without yeast extract could be activated by the addition of ATP(adenosine-5-triphosphate), as illustrated in figure 6. With such preparationsglycerol addition does not result in oxygen uptake, but the further addition of
353

I. C. GUNSALUS AND W. W. UMBREIT
ATP causes oxidation (stopping at the toxic concentration of H202). WhenATP is added without glycerol, no oxidation occurs. The commercial glycerolphosphates are oxidized slowly by this preparation; the addition of muscleadenylic acid does not improve the respiration. This may be due to the inhibi-tory action of the inactive forms of glycerol phosphate on the oxidation of theactive forms, since the addition of the mixed glycerol phosphates to cell prepara-
tions oxidizing glycerol results in an inhibition. The activation of glyceroloxidation by ATP (figure 6) indicates that the glycerol is converted into thenatural glycerol phosphate before oxidation.
50
20 /~~~~
o40 IH202 CONc0 2 43408LMLD~~~~~~~~~~~
z
0
wI-
01
0
I 5~ TIP 5MG ATP
GLYCEROL0 20 40 60 80 100 120
MINUTE SFIG. 6. ATP ACTIVATION OF GLYCEROL OXIDATION BY GROUND CELL PREPARATION
100 mg vacuum-dried cells (prep. 5) ground 18 hours in micro ball mill undervacuum and suspended in 3.3 ml M/15 phosphate buffer pH 7.1.
0.5 ml (15 mg) suspension added per flask.0.1 ml M/20 glycerol (5,u moles) and0.5 ml Na ATP (10 mg/ml) added to side arm.Side arms tipped as indicated, 37 C.
From these data the pathway of glycerol oxidation may be formulated withrelative clarity. Glycerol reacts with ATP to form a glycerol phosphate,(presumably 1-(-)-a; Baer and Fisher, 1939). This is the compound which israpidly oxidized, probably to triose phosphate. The latter is converted intolactic acid through the same system followed by triose phosphates resulting fromglucose fermentation. The glycerol oxidation system is thus part of a consider-ably more complex system of reactions.
It therefore becomes difficult to attribute the effect of an inhibitor to a reac-tion at a single point in this series. In the living cell, inhibition of any one ofthe reactions seems to result in the inactivation of the glycerol-oxidizing system.Therefore, it has not been possible to trap intermediates. Oxygen uptake on
354

OXIDATION OF GLYCEROL BY STREPTOCOCCUS FAECALIS
glycerol is not inhibited by cyanide in the usual concentrations (i.e., M/100 toM/1,000) but is inhibited by cyanide at levels of 0.03 M (80% inhibition) and at0.1 M (100% inhibition). Iodoacetate (which was used in an attempt to blockthe system at the triose phosphate stage) completely inhibited the oxidation ofglycerol at 0.001 M. Sodium fluoride (in attempts to set up a similar block atthe phosphoglyceric acid stage) showed 20 per cent inhibition of glycerol oxida-tion at 0.01 M, 80 per cent at 0.03 M, and 100 per cent at 0.075 M. However, theaction of fluoride appears to be on the glycerol system as such since pyruvatedoes not relieve the inhibition, as should be the case if transformations of thephosphoglyceric acids were inhibited.
Since the material reacting with oxygen appears to be a glycerol phosphate,it is of interest to compare this system with that of the animal. Two a-glycerol-phosphate dehydrogenases have been reported: that of Euler et al. (1937) whichis a coenzyme I system, and that of Green (1936) which is linked to cytochrome.If the enzyme system with which we are dealing is coenzyme I linked, it shouldbe possible to obtain a conversion of glycerol to lactic acid under anaerobicconditions, providing pyruvate is supplied. However, when this was tried withresting cells, no acid was produced from glycerol nor from glycerol plus pyruvateunder anaerobic conditions. Furthermore, with this strain (no. 24) other hydro-gen acceptors (for example, fumarate) do not permit growth or acid productionunder anaerobic conditions. It appears that the oxidizing system is tied di-rectly to oxygen, and we have been unable to find other substrates with whichit will react. In a sense, therefore, this system resembles the enzyme of Green,but differs in its direct reaction with oxygen without the participation of thecytochrome system.
It is rather remarkable that a noncytochrome system is capable of the rapidreaction with oxygen exhibited during glycerol oxidation. When one considersthat the preparations with which we are dealing are far from pure, and muchinactive matter is contributing to the dry weight and the nitrogen, the capacityof the enzyme to utilize oxygen must be very great indeed. Although one wouldpresume that the pure enzyme is a flavoprotein, it differs from known flavopro-teins in the rapidity with which it reacts with oxygen. The rate of oxidation ofglycerol is not increased by the use of pure oxygen instead of air nor by theaddition of methylene blue.The dried cells may be treated with ammonium sulfate at pH 3 to 4 (as is used
in the separation of the flavin from the protein with Warburg and Christian'sold yellow enzyme), or subjected to the procedures used by Lipmann (1939)and by Straub (1939) for separating the flavin component from their prepara-tions, without removing activity from the cell particles. The separation used onthe amino acid oxidase (pH 5, 1/2 saturated (NH4)2SO4) completely inactivatesthe glycerol system (although it is less drastic than the ones previously men-tioned), and we have been unable to restore the activity. Also, the relation be-tween the activity of the cells on glycerol and the flavin content of the mediumis not a direct one. Hence, detailed knowledge of the nature of the enzymes in-volved in glycerol oxidation must await their isolation.
355

I. C. GUNSALUS AND W. W. UMBREIT
The evidence presented to show the course of glycerol oxidation also bearsupon the functional nature of this respiration. Certainly the glycerol oxidationprovides energy to the cell since this streptococcus will grow well when glycerolis supplied aerobically, whereas very little growth occurs in the base medium(Gunsalus and Sherman, 1943). However, the respiration need not yield energydirectly to the cell, for, after glycerol is converted into triose phosphate, the lattercan be fermented to lactate and provide energy. The powerful respiratory systemoperating on glycerol may, therefore, be merely a means which the cell employsto remove two extra hydrogens, and the cell may derive its energy, not from therespiration as such, but by fermenting the&product of that respiration. There isno critical evidence at present that the cell derives energy from the respirationas such.The uptake of exactly one mole of oxygen per mole of glycerol suggests that
there is no very direct pathway to oxygen from the coenzyme I stage, and thelow oxidation of glucose and lactate as shown in figure 1 strengthens this conclu-sion. However, in working with this organism over the past three years, wehave noted a considerable variation in the ability of different suspensions tooxidize glucose and lactate. A similar variability was noted by Barron and Jacobs(1938). Suspensions which oxidize glucose and lactate at the more rapid rates(or preparations therefrom, for example as in figure 4) take up slightly morethan one 02 per glycerol-the extra oxygen uptake being correlated with lactateor glucose oxidation which presumably proceeds through reduced coenzyme I.The data in figure 4 illustrate the type of result obtained. While this variabilityhas not yet been traced to its cause, its occurrence does not influence the con-clusions obtained with the glycerol system.
SUMMARY
Glycerol is oxidized by Streptococcus faecalis, strain 24, at a rapid rate (Qo,(N)= 400-700), the over-all reaction being:
Glycerol + 02- lactate + H202
The pathway of this oxidation involves phosphorylation. The glycerol isphosphorylated (with ATP) to glycerol phosphate, which is oxidized to triosephosphate. The latter is converted into lactic acid.The oxidation is inhibited by the accumulation of H202 to a concentration of
0.002 molar.Various properties of the enzyme system are described.
REFERENCES
BAER, E., AND FISHER, H. 0. L. 1939 VI. Synthesis of biological 1-(-)--a-glycerophos-phorie acid. J. Biol. Chem., 128, 491-500.
BARKER, S. B., AND SUAIMERSON. W. H. 1941 The colorimetric determination of lactic acidin biological material. J. Biol. Chem., 138, 535-554.
BARRON, E. S. G., AND JACOBS, H. R. 1938 Oxidations produced by hemolytic strepto-cocei. J. Bact., 36, 433-449.
EULER, H., ADLER, E., AND GUNTHER, G. 1937 Zur Kenntnis der Dehydrierung vona-Glycerin-phosphorsiiure Tierk6rper. Z. physiol. Chem., 249, 1-15.
356

OXIDATION OF GLYCEROL BY STREPTOCOCCUS FAECALIS 357
FARRELL, M. A. 1935 Studies on the respiratory mechanisms of streptococci. J. Bact.,29, 411-435.
FISKE, C. H., AND SUBBAROW, Y. 1925 The colorimetric determination of phosphorus.J. Biol. Chem., 66, 375-400.
FREI, W., RIEDMULLER, L., AND ALMASY, F. 1934 Uber Cytochrom und das Atmungs-system der Bakterien. Biochem. Z., 274, 253-267.
GREEN, D. E. 1936 a-Glycero-phosphate dehydrogenase. Biochem. J., 30, 629-641.GUNSALUS, I. C., AND SHER-MAN, J. M. 1943 The fermentation of glycerol by streptococci.
J. Bact., 45, 155-162.LIPMANN, F. 1939 An analysis of the pyruvic acid oxidation system. Cold Spring Harbor
Symposia Quant. Biol., 7, 248-257.SEVAG, M. G. 1932 tYber den Atmungsmechanismus der Pneumococcen. Biochem. Z.,
267, 211-237.STRAUB, F. B. 1939 Isolation and properties of a flavoprotein from heart muscle tissue.
Biochem. J., 33, 787-792.SUMNER, J. B., AND DOUNCE, A. L. 1939 Catalase. II. J. Biol. Chem., 127, 439-447.WARBURG, O., AND CHRISTIAN, W. 1932 ttber ein neues Oxidationsferment und sein
Absorptionsspektrum. Biochem. Z., 245, 438-458.WOOD, A. J., AND GUNSALUS, I. C. 1942 The production of active resting cells of strepto-
cocci. J. Bact., 44, 333-341.




















