
Genetic variability of Hong Kong (H3N2) influenza viruses: spontaneous mutations and their location...
13
Gene, 11 (1980) 319-331 319 © Elsevier/North-Holland BiomedicalPress Genetic variability of Hong Kong (H3N2) influenza viruses: spontaneous mutations and their location in the viral genome (T 1-oligonucleotide fingerprinting; genetic heterogeneity; gene isolation; virus evolution; mutation mapping) Juan Ort/n a, Rafael Nfijera b* , Cecilio L6pez b, Mercedes Dfivilaa and Esteban Domingo a* a Centro de Biologia Molecular, Facultad de Ciencias, UniversidadAut6noma, Canto Blanco, Madrid-34, and b Centro Nacional de Microbiologia, Virologia e Inmunologia San#arias, Ma]adahonda, Madrid (Spain) (Received May 6th, 1980) (Accepted July 20th, 1980) SUMMARY The genetic heterogeneity of five influenza A (H3N2) strains isolated between 1968 and 1977 has been estimated by Tl-oligonucleotide fingerprinting of a2p-labeled viral RNA. Assuming that the large Tl-resistant oligonucleotides represent a random sample of the viral RNA, the genetic differences observed would affect 0.3 to 10.7% of the RNA positions of the genes studied, depending on the pair of viruses considered. A smaller degree of genetic heterogeneity was observed when six coetaneous viral samples were compared. The distribution of spontaneous mutations among the viral genes was studied by fingerprinting individual RNA segments isolated either by gel electrophoresis or hybridization with plasmids containing influenza-specific DNA sequences. No statistically significant differences were detected in the distribution of mutations among the viral genes studied. The mutation frequency at the hemagglutinin RNA region coding for the HA1 subunit was found to be two times higher than that at the region encoding the HA2 subunit. Our results suggest that the antigenic variability of influenza viruses may be a consequence of a general genetic variability which affects many of the viral genes. INTRODUCTION Influenza A viruses are the causative agents of a major infectious disease of man. Their genome con- sists of eight segments of negative-strand RNA, each * Present addresses: (E.D.) Biogen S.A., 3, Route de Troinex, 1227 Carionge, Geneve (Switzerland); (R.N.) Virus Unit, WHO, Geneve (Switzerland). Abbreviations: HA, hemagglutinin; M, matrix protein; NA, neuraminidase; NP, nucleoprotein; NS, non-structuralprotein; PX' mixture of polymerases 1, 2, 3. one encoding for one or more of viral genes (Palese, 1977; Scholtissek, 1978; Webster and Bean, 1978; Inglis et al., 1979; Lamb et al., 1979). Influenza epidemiology relies on the ability of the virus to undergo variation in its surface antigens, some giving rise to new virus subtypes that cause pandemics of disease. Such variations have been termed a'ntigenic shift and may be the result of recom- bination of the circulating human strain with other influenza viruses from an undetermined reservoir (Webster and Laver, 1975; Desselberger et al., 1978; Young and Palese, 1979). However, the latest pande- mic of influenza originated in China in 1977 from an
Transcript of Genetic variability of Hong Kong (H3N2) influenza viruses: spontaneous mutations and their location...

Gene, 11 (1980) 319-331 319 © Elsevier/North-Holland Biomedical Press
Gene t i c variabil i ty o f Hong Kong (H3N2) inf luenza viruses: spon taneous mu ta t i ons and their
loca t ion in the viral genome
(T 1-oligonucleotide fingerprinting; genetic heterogeneity; gene isolation; virus evolution; mutation mapping)
Juan Ort/n a, Rafael Nfijera b* , Cecilio L6pez b, Mercedes Dfivila a and Esteban Domingo a*
a Centro de Biologia Molecular, Facultad de Ciencias, UniversidadAut6noma, Canto Blanco, Madrid-34, and b Centro Nacional de Microbiologia, Virologia e Inmunologia San#arias, Ma]adahonda, Madrid (Spain)
(Received May 6th, 1980) (Accepted July 20th, 1980)
SUMMARY
The genetic heterogeneity of five influenza A (H3N2) strains isolated between 1968 and 1977 has been estimated by Tl-oligonucleotide fingerprinting of a2p-labeled viral RNA. Assuming that the large Tl-resistant oligonucleotides represent a random sample of the viral RNA, the genetic differences observed would affect 0.3 to 10.7% of the RNA positions of the genes studied, depending on the pair of viruses considered. A smaller degree
of genetic heterogeneity was observed when six coetaneous viral samples were compared. The distribution of spontaneous mutations among the viral genes was studied by fingerprinting individual RNA segments isolated either by gel electrophoresis or hybridization with plasmids containing influenza-specific DNA sequences. No statistically significant differences were detected in the distribution of mutations among the viral genes studied. The mutation frequency at the hemagglutinin RNA region coding for the HA1 subunit was found to be two times higher than that at the region encoding the HA2 subunit. Our results suggest that the antigenic variability of influenza viruses may be a consequence of a general genetic variability which affects many of the viral genes.
INTRODUCTION
Influenza A viruses are the causative agents of a major infectious disease of man. Their genome con- sists of eight segments of negative-strand RNA, each
* Present addresses: (E.D.) Biogen S.A., 3, Route de Troinex, 1227 Carionge, Geneve (Switzerland); (R.N.) Virus Unit, WHO, Geneve (Switzerland).
Abbreviations: HA, hemagglutinin; M, matrix protein; NA, neuraminidase; NP, nucleoprotein; NS, non-structural protein; PX' mixture of polymerases 1, 2, 3.
one encoding for one or more of viral genes (Palese, 1977; Scholtissek, 1978; Webster and Bean, 1978; Inglis et al., 1979; Lamb et al., 1979).
Influenza epidemiology relies on the ability of the virus to undergo variation in its surface antigens,
some giving rise to new virus subtypes that cause pandemics of disease. Such variations have been termed a'ntigenic shift and may be the result of recom- bination of the circulating human strain with other influenza viruses from an undetermined reservoir (Webster and Laver, 1975; Desselberger et al., 1978; Young and Palese, 1979). However, the latest pande- mic of influenza originated in China in 1977 from an

320
H1N1 virus that was shown to be genetically almost identical to H1NI viruses circulating in the 1950's (Nakajima et al., 1978; Scholtissek et al., 1978). The possibility is still open that human influenza viruses are preserved in nature in some latent state and that they reappear by unknown mechanisms.
In addition to the antigenic shift, the influenza virus surface antigens change in a stepwise fastuon during the interpandemic periods, allowing for the emergence of epidemic strains. This antigenic drift seems to be caused by the accumulation of point mu- tations (Young et al., 1979; Min You et al., 1980) and the subsequent selection of antigenic variants by the immune host population (Webster and Laver, 1975). In fact, antigenic drift has been shown to occur in the laboratory (Fazekas de St. Groth and Hannoun, 1973; Gerhard and Webster, 1978).
Genetic polymorphism has been shown in a variety of RNA viruses. Sampling RNA sequences by T1 oligonucleotide fingerprinting revealed one to two changes in the RNA sequence between any two individual clones of a Q/3 bacteriophage population (Domingo et al., 1978), while foot-and-mouth disease viruses isolated during epizootic outbreaks show mu- tations at 0.7 to 2.2% of the positions of their RNAs (Domingo et al., 1980). Likewise, several laboratory strains of VSV show distinct oligonucleotide finger- prints (Clewley et al., 1977). By polyacrylamide gel electrophoresis, differences were reported in the mobility of the RNA segments of several rotaviruses (Rodger and Holmes, 1979) and reoviruses (Hrdy et al., 1979), suggesting heterogeneity in their nucleotide sequences.
In addition, a rapid accumulation of mutations was shown in the VSV genome during persistent infec- tions (Holland et al., 1979; Semler and Holland, 1979) and antigenic variants of Visna virus were selected after infection of single sheep with cloned virus (Scott et al., 1979), suggesting that in the course of these infections a seemingly fast evolution of the viral population can occur.
In the present study we have taken advantage of the increased sensitivity of the oligonucleotide finger- printing technique, when applied to RNAs or limited sequence complexity such as the individual influenza virus RNA segments, to address the following ques- tions concerning the genetic variability of this virus: What is the degree of genetic divergence between reference strain viruses of the H3N2 series? Is there
any detectable genetic heterogeneity between H3N2 viruses c0-circulating in a given area? Do the sequence differences accumulate in the surface antigen genes or, rather, are they distributed at random among the viral genes? Is there any preferential location of muta- tions in either the HA1 or HA2 moieties of the HA gene?
As previously reported for H1N1 viruses (Nakaji- ma et al., 1978; Young et al., 1979), we have detected considerable genetic differences between H3N2 reference strains. Depending on the pair of viruses considered, these differences are estimated to amount from 0.3 to 10.7% of the RNA positions of the genes studied and appear to be randomly distributed among them. The mutations observed in the HA gene are distributed in a 2 to 1 ratio between the regions coding for the HA1 and HA2 subunits. We also pro- vide evidence for some detectable genetic hetero- geneity in viruses co-circulating in the same area. Our data suggest that the spontaneous mutation rate may be the basis for antigenic variability of influenza viruses.
MATERIALS AND METHODS
(a) Viruses and cells
The canine kidney cell line MDCK was obtained from the American Type Culture Collection at passage 53 and subcultured twice weekly using the Dulbecco's modification of Eagle's medium (DMEM, Dulbecco and Vogt, 1964), supplemented with 5% fetal calf serum. Cells were used between passage 55 and 65.
Influenza reference strains A/Hong Kong/8/68, A/Victoria/3/35, A/Victoria/112/76 and A/Texas/1/77 were obtained from the Influenza Center, London and used after 3 -5 passages in egg. The viruses A/ Madrid/662/77, A/Madrid/672/77, A/Pamplona/1026/ 77, A/Pamplona/1028/77, A/Pamplona/1031/77 and A/Pamplona/1033/77 were isolated in these two cities (300 km apart) between March 10 and 25, 1977 and used for analysis after one egg passage. Viruses were titrated on MDCK monolayers essentially as described (Tobita et al., 1975).
Escherichia coli clones, containing the following A/Victoria/3/75 virus gene sequences: HA (pV4/14, pVHA101 and pVHA55) (Min Jou et al., 1980), NP

(pV5/39) and NA (pV6/21) were obtained from Dr.
W. Fiers. Bacterial cultures were incubated overnight
in the presence of 200/zg/ml of chloramphenicol to allow for plasmid amplification and plasmid DNA was purified according to published procedures (Cle- well and Helinski, 1969; Currier and Nester, 1976).
(b) Materials
Media and sera were from Grand Island Biochemi- cal Co., Scotland. [32P]Phosphate, carrier free, was purchased from the Radiochemical Center, Amers- ham, England. RNase T1 was bought from Calbio- chem, San Diego, CA, proteinase K from Merck, Darmstadt, and trypsin from Worthington, Freehold,
NJ. Acrylamide was obtained either from Serva, Hei- delberg, or from Ferosa, Barcelona, and chloram-
phenicol and tetracycline were purchased from Sigma Chemical Co., St. Louis, MO.
(c) Virus label ing and purif ication
For virus labeling, MDCK cells were infected at a
multiplicity of infection of 10 -a plaque-forming units per cell. After an adsorption period of 1 h, the inocu- lum was removed and the infected cells incubated in Hanks medium containing 200-400/aCi/ml of [32p].
phosphate ( 2 - 4 mCi//amol). When the cytopathic
effect was complete, labeled virus was purified from
the supernatant medium after removing the cellular debris by centrifugation for 15 min at 4000 × g. The virus was concentrated by centrifugation for 1 h at 256000 × g and 4°C through a 33% (w/v) sucrose cushion in 0.1 M NaC1-0.01 M Tris" HCI-O.001 M EDTA, pH 7.5. The virus-containing pellet was resuspended and centrifuged for 1 h at 256 000 × g in a 33 -50 (w/v) sucrose gradient in the same buffer.
(d) Isolation o f R N A
Purified virus preparations were incubated for 10 min at 37°C with 100/~g/ml of proteinase K and 0.5% sodium dodecylsulfate, and extracted three times with phenol-chloroform-isoamyl alcohol-8- hydroxyquinoline (Holmes and Bonner, 1973). Viral RNA was then precipitated twice with ethanol at -20°C. Separation of the individual segments of RNA was performed by one of the following proce-
dures:
321
(1)Viral RNA precipitates were resuspended in formamide, boiled for 1 min and applied to a 40 ×
20 × 0.2 cm slab gel containing 3% polyacrylamide and 6 M urea in 0.09 M Tris-borate-0.002 M EDTA, pH 8.3 (Peacock and Dingman, 1968). Electropho-
resis was carried out in the cold for 1 8 - 2 0 h at 40 mA. The RNA bands were visualized by autoradio- graphy, eluted from the gel in 1.0 M NaC1-0.05 M Tris "HC1-0.O01 M EDTA, pH 7.5 and the RNA
Px=:m
H A - - I
NP NA
Fig. 1. Separation of influenza virus RNA segments by poly- acrylamide gel electrophoresis. A/Victoria/3/75 virus RNA was labeled and purified as described in MATERIALS AND METHODS. Purified viral RNA (1 × l06 cpm) was heated for 1 rain at 100°C in 98% formamide and electrophoresis was carried out as described in MATERIALS AND METHODS. Assignment of RNA segments to viral genes was assumed to be as described by Desselberger and Palese (1978) and further confirmed for RNAs 4, 5 and 6 by comparison of fingerprints of these RNA segments isolated by either gel electrophoresis or hybridization with cloned viral DNA.

322
precipitated twice with ethanol. Fig. 1 shows a typical pattern of 32p-labeled RNA.
(2) Total viral RNA was hybridized to plasmid DNA containing viral sequences fixed on nitrocellulose filters. Incubation was for 3 6 4 8 h at 37°C in 1 ml of 5 × SSC-50% formamide-0.5% sodium dodecyl- sulfate. After hybridization, the f'dters were washed with 2 X SSC, the retained RNA was eluted by
incubation at 50°C in formamide and precipitated twice with ethanol after addition of 25/ag of carrier RNA.
For the selection of viral RNA sequences com- plementary to HA gene inserts contained in plasmids pVHA55 and pVHA101, hybrid containing f'llters were incubated for 30 min at 37°C with 5 units of T1 RNase in 0.5 ml of 0.5 M NaC1-0.01 M Tris • HC1, pH 7.5 containing 10/lg of carrier RNA. Resistant RNA was eluted and processed as described above.
(e) RNA fingerprinting
Total 32P-labeled influenza virus RNA or individual
RNA segments were digested for 30 min at 37°C with
one unit of T1 RNase/25 #g of RNA in 0.02 M Tris • HC1-0.002 M EDTA pH 7.5. Resistant oligonucleo-
tides were separated by two-dimensional electropho-
resis as described (de Wachter and Fiers, 1972; Do- mingo et al., 1980). Dried gels were exposed at -70°C with the aid of intensifying screens.
RESULTS
(a) Comparative analysis of reference strains and virus isolates by oligonucleotide fingerprinting of total genomic RNA
As a first step in the study of the genetic related- ness of H3N2 influenza viruses, we have compared the Tl-oligonucleotide fingerprints of total genomic
RNA from four reference strains and from six field isolates obtained in Spain, in 1977. The fingerprints
obtained from the total RNA of field isolates were indistinguishable from each other (data not shown), in spite of the differences detected among some of
them by hemagglutination-inhibition (Table I). Con-
siderable differences were detected, however, among the RNAs from reference strains or between them and the RNAs from field isolates. As an example, Fig. 2 shows the comparison between the fingerprints of A/Victoria/3/75 and A/Madrid/662/77 RNAs. Out of 40 well resolved T1 oligonucleotides in the latter, 7 were absent from A/Victoria/3/75 RNA, that showed 8 additional ones. Since the analyzed oligo-
nucleotides account for about 6.3% of the genome
and assuming that each change affects one nucleotide, it is estimated that the mutations affect about 1.5%
of the positions in the RNA. A similar number of dif- ferences were obtained when the RNAs from A/Vic- toria/ 112/76 and A/Texas/1/77 strains were included
TABLE I Hemogglutinatlon- inhibition reactions of human
influenza A viruses ( H 3 N 2 ) isolated in Spain in 1977
Antigen Ferret antisera
A/Victoria / 3 / 7 5 A/Victoria / 112 /76
Victor ia /3 /75 20 4800 5120
Victoria/112/76 1280 5120
Madrid/672177 1920 960
Madrid •662/77 10 240 - -
Pamplona/1028/77 1 920 1 280
Pomplona/1051/77 19 20 12 80
Pomplono/1026/77 480 3 20 Pomplono/1055/77 1280 12 8 0
a HI titers represent the reciprocal of the dilution inhibiting two of four hemaggluti- nating units of virus.
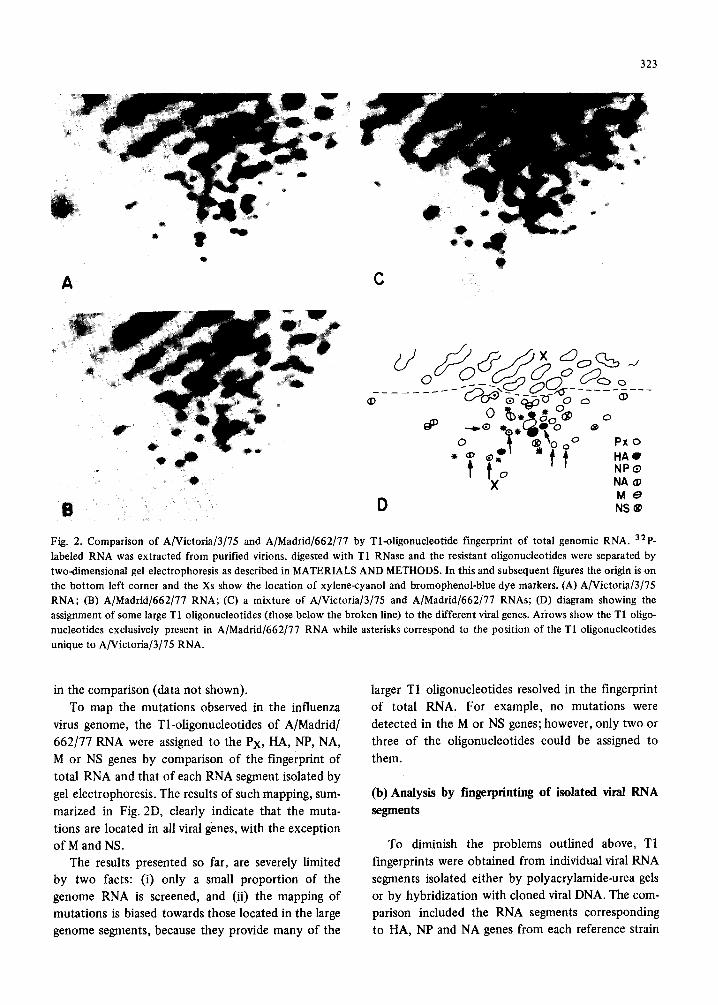
323
f e
A C
B
C)
e . _ . L i e o ,.
"=' ? e ~ f ~, HAe I ~ o " " NP(9
X NA (p M e
D NS®
Fig. 2. Comparison of A/Victoria/3/75 and A/Madrid/662/77 by Tl-oligonucleotide fingerprint of total genomic RNA. 32p. labeled RNA was extracted from purified virions, digested with T1 RNase and the resistant oligonucleotides were separated by two-dimensional gel electrophoresis as described in MATERIALS AND METHODS. In this and subsequent figures the origin is on the bottom left corner and the Xs show the location of xylene-cyanol and bromophenol-blue dye markers. (A) A/Victoria/3/75 RNA; (B) A/Madrid/662/77 RNA; (C) a mixture of A/Victoria/3[75 and A/Madrid/662/77 RNAs; (D) diagram showing the assignment of some large T1 oligonucleotides (those below the broken line) to the different viral genes. Arrows show the T1 oligo- nucleotides exclusively present in A/Madrid/662/77 RNA while asterisks correspond to the position of the T1 oligonucleotides unique to A/Victoria/3/75 RNA.
in the comparison (data not shown).
To map the mutations observed in the influenza
virus genome, the Tl-oligonucleotides of A/Madrid/
662/77 RNA were assigned to the PX, HA, NP, NA,
M or NS genes by comparison of the fingerprint of
total RNA and that of each RNA segment isolated by
gel electrophoresis. The results of such mapping, sum-
marized in Fig. 2D, clearly indicate that the muta-
tions are located in all viral genes, with the exception
of M and NS. The results presented so far, are severely limited
by two facts: (i) only a small proportion of the genome RNA is screened, and (ii) the mapping of mutations is biased towards those located in the large
genome segments, because they provide many of the
larger T1 oligonucleotides resolved in the fingerprint of total RNA. For example, no mutations were
detected in the M or NS genes; however, only two or
three of the oligonucleotides could be assigned to
them.
(b) Analysis by fingerprinting of isolated viral RNA
segments
To diminish the problems outlined above, T1 fingerprints were obtained from individual viral RNA
segments isolated either by polyacrylamide-urea gels or by hybridization with cloned viral DNA. The com- parison included the RNA segments corresponding to HA, NP and NA genes from each reference strain
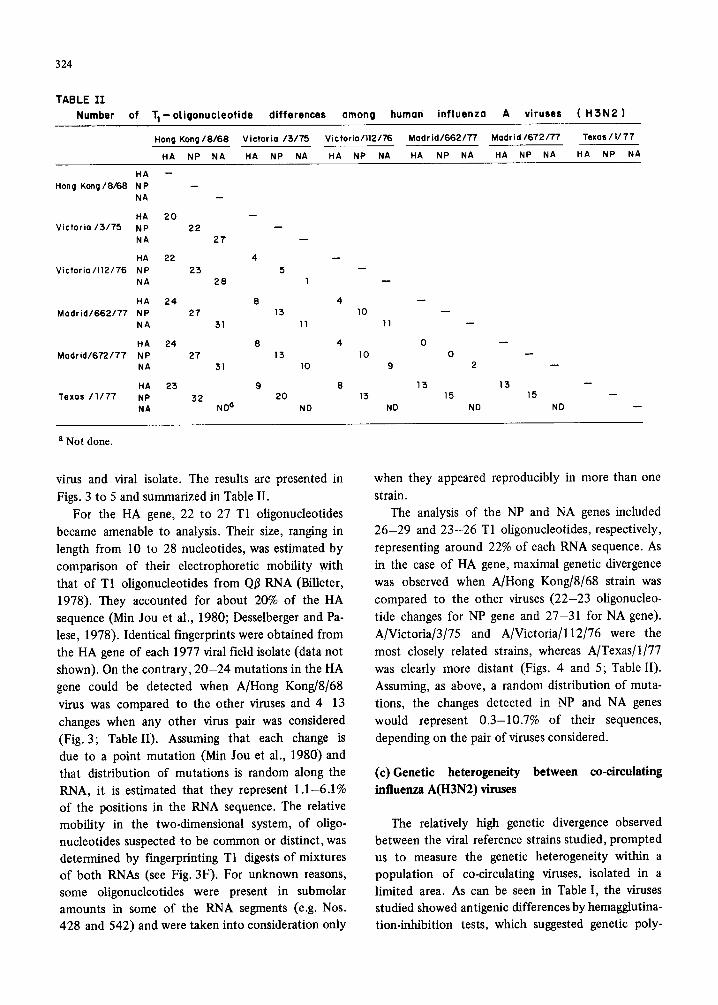
324
TABLE 11
Number of Tl-otigonucleotide differences among human influenza A viruses ( H 3 N 2 )
Hang Kong/8/68 Victoria /:5/75 Victoria/112/76 Modrid/662/T/ ' Madrid/672/77 Texas/ I /T7
HA NP NA HA NP NA HA NP NA HA NP NA HA NP NA HA NP NA
HA -- Hang Kong/8/68 NP
NA
HA 20 V ic to r ia /3 /75 NP 22 - -
NA 27 - -
HA 22 4 Victoria/112/76 NP 23 5
NA 28 l
HA 24 8 Madr id/662/77 NP 27 13
NA 31 11
HA 24 8 Madrid/672/77 NP 27 15
NA 31 10
HA 23 9 Texas /1177 NP 32 20
NA ND ° ND
10 11
4 0 10 0
9 2
8 13 13 13 15 15
ND ND ND
a N o t done .
virus and viral isolate. The results are presented in
Figs. 3 to 5 and summarized in Table II. For the HA gene, 22 to 27 T1 oligonucleotides
became amenable to analysis. Their size, ranging in length from 10 to 28 nucleotides, was estimated by comparison of their electrophoretic mobility with that of T1 oligonucleotides from Q/3 RNA (Billeter, 1978). They accounted for about 20% of the HA sequence (Min Jou et al., 1980; Desselberger and Pa- lese, 1978). Identical fingerprints were obtained from the HA gene of each 1977 viral field isolate (data not shown). On the contrary, 20-24 mutations in the HA gene could be detected when A/Hong Kong/8/68 virus was compared to the other viruses and 4 -13 changes when any other virus pair was considered (Fig. 3; Table II). Assuming that each change is due to a point mutation (Min Jou et al., 1980) and that distribution of mutations is random along the RNA, it is estimated that they represent 1.1-6.1% of the positions in the RNA sequence. The relative mobility in the two-dimensional system, of oligo- nucleotides suspected to be common or distinct, was determined by fingerprinting T1 digests of mixtures of both RNAs (see Fig. 3F). For unknown reasons, some oligonucleotides were present in submolar amounts in some of the RNA segments (e.g. Nos. 428 and 542) and were taken into consideration only
when they appeared reproducibly in more than one
strain. The analysis of the NP and NA genes included
26-29 and 23-26 T1 oligonucleotides, respectively, representing around 22% of each RNA sequence. As in the case of HA gene, maximal genetic divergence was observed when A/Hong Kong/8/68 strain was compared to the other viruses (22-23 oligonucleo- tide changes for NP gene and 27-31 for NA gene). A/Victoria/3/75 and A/Victoria/l12/76 were the most closely related strains, whereas A/Texas/I/77 was clearly more distant (Figs. 4 and 5; Table II). Assuming, as above, a random distribution of muta- tions, the changes detected in NP and NA genes would represent 0.3-10.7% of their sequences, depending on the pair of viruses considered.
(c)Genetic heterogeneity between co-circulating influenza A(H3N2) viruses
The relatively high genetic divergence observed between the viral reference strains studied, prompted us to measure the genetic heterogeneity within a population of co-circulating viruses, isolated in a limited area. As can be seen in Table I, the viruses studied showed antigenic differences by hemagglutina- tion-inhibition tests, which suggested genetic poly-

325
• b
I
0
Q qk °
4t3
A
B
g
Q
X
~u~ 1
~0
D
0
e
0
X
4~ 0
41
LD E x I
m '1t w 0
ip D •
0 C x q F
Fig. 3. Ti-oligonucleotide fingerprints of isolated hemagglutinin RNAs from several influenza viruses (H3N2). 32P-Labeled hemagglutinin RNAs were selected by hybridization with cloned viral DNA (pV4/14) and their T1 fingerprints obtained as described in MATERIALS AND METHODS. (A) A/Hong Kong/ 8•68 HA RNA; (B) A/Victoria/3[75 HA RNA; (C)A/Victoria/ 112/76 HA RNA; (D) A/Texas/I/77 HA RNA; (E) A/Madrid/662177 HA RNA; (F) mixture of A/Victoria/3/75 and A/Madrid/ 662/77 HA RNAs. In this and subsequent figures, large T1 oligonucleotides from A/Hong Kong/8/68 are given reference numbers (the 400 series for HA RNA, the 500 series for NP RNA and the 600 series for NA RNA). The numbers in the other fingerprints indicate the oligonucleotides that f'trst appear in any given virus, when considering them in chronological order. Arrows in (F) show oligonucleotides present in only one of the RNAs of the mixture.
morphism at the HA gene. However, the suspected
sequence changes were not detected by fingerprinting
either of total viral RNA or of HA gene RNA.
When the study was extended to the NP, NA and
M RNAs, two oligonucleotide changes were detected
in the fingerprints of NA RNA of the isolates A/
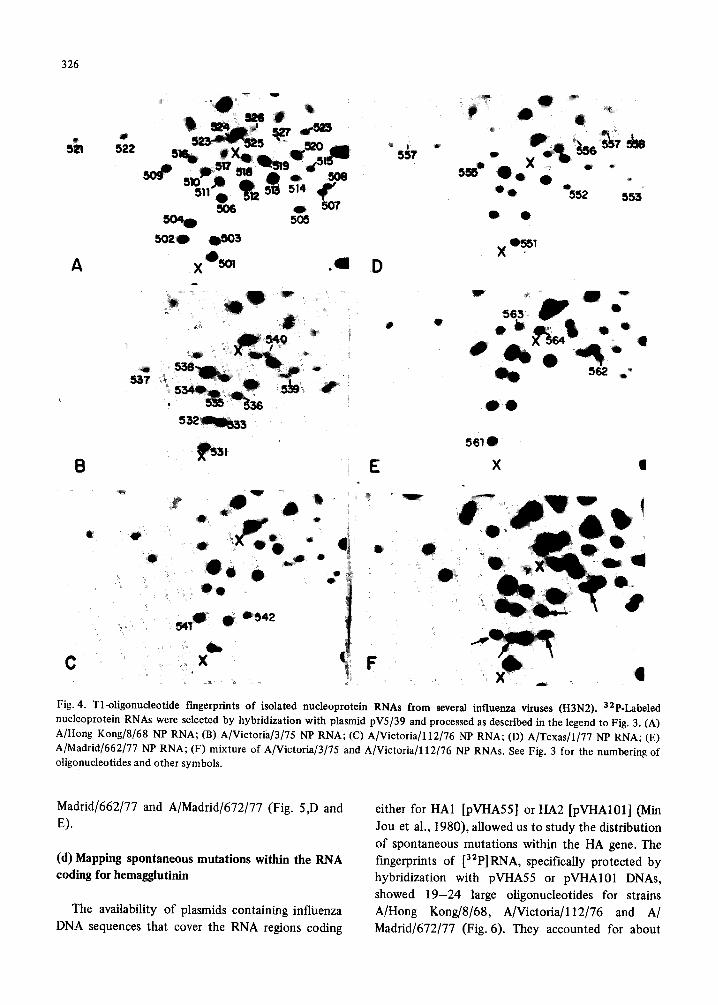
326
m ̧ 521 522
O ~ q
5o2,m, e,.~o~
A x °5°~ .41 D
l J w t 4
8
8
B E
I 55?
P II •
5~" O o o 0 qlk 0552
o •
0551 X
563 o b
e
0 O
O ~g~
Ik
L qlP Q
J• Q
562 ,,~
5614J
X
553
I 0
B
'qqmP" ! k -
Q
" iL a t
." 0
qlP
o
0 o v j
B e o e o
541 ° • D542
(Ib C x q F
q
O Q Q b 9
S
I
Fig. 4. Tl-oligonudeotide rmgerprints of isolated nucleoprotein RNAs from several influenza viruses (H3N2). 32P-Labeled nucleoprotein RNAs were selected by hybridization with plasmid pV5/39 and processed as described in the legend to Fig. 3. (A) A/Hong Kong/8/68 NP RNA; (B) A/Victoria/3/75 NP RNA; (C) A/Victoria/112/76 NP RNA; (D) A/Texas/l/77 NP RNA; (E) A/Madrid/662/77 NP RNA; (F) mixture of A/Victoria[3[75 and A/Victoria/112/76 NP RNAs. See Fig. 3 for the numbering of oligonucleotides and other symbols.
Madrid/662/77 and A/Madrid/672/77 (Fig. 5,D and E).
(d) Mapping spontaneous mutations within the RNA coding for hemagglutinin
The availability of plasmids containing influenza DNA sequences that cover the RNA regions coding
either for HA1 [pVHA55] or HA2 [pVHA101] (Min Jou et al., 1980), allowed us to study the distribution of spontaneous mutations within the HA gene. The fingerprints of [32p] RNA, specifically protected by hybridization with pVHA55 or pVHA101 DNAs, showed 19-24 large ohgonucleotides for strains A/Hong Kong/8/68, A/Victoria/112/76 and A/ Madrid/672/77 (Fig. 6). They accounted for about

A
m
X
-1~t2 O
O
I I
O
X
t
g
Q I
327
6~
e
w
C
0
X
t
O x
9 ~ e m ~ ~, i J
° L ° r ' -
Q
0
fill.N!
E X " ' F X
Fig. 5. Tl-oligonucleotide f'mgerprints of isolated neuraminidase RNAs from several influenza viruses (H3N2). 32P-Labeled neuraminidase RNAs were selected by hybridization with plasmid pV6/21 and processed as described in the legend to Fig. 3. (A) A/Hong Kong/8]68 NA RNA; (B) A/Victoria/112/76 NA RNA; (C) A[Victoria/3/75 NA RNA; (D)A/Madrid/672/77 NA RNA; (E) A/Madrid/662/77 NA RNA; (F) mixture of A/Madrid[672/77 and A[Victoria/3175 NA RNAs. Asterisks show the oligonu- cleotide differences observed when NA RNAs from A/Madrid[662~77 and A/Madrid/672/77 are compared. See Fiz. 3 for num- bering and other symbols.
3 0 - 4 0 % of the viral sequences contained in each
cloned DNA. The HA-specific oligonucleotides (Fig.
3) distributed themselves as follows: some were pro-
tected by DNA from pVHA55 clone, like Nos. 431,
436, 462 (Fig. 6,B and C) while some others were
protected by DNA from pVHA101 clone, like Nos.
4 0 2 , 4 0 4 , and 461 (Fig. 6 , D - F ) . In addit ion, a third
group of oligonucleotides were not protected either
by pVHA55 or by pVHA101 DNAs and can therefore
be assumed to derive from positions 750 to 1180 in

328
A i)
B
q ~ O
Q
tP'431
K
D
m P I O
x J
Fig. 6. Tl-oligonucleotide f'mgerprints of distinct sequences within hemagglutinin RNA. a2P-Labeled hemagglutinin RNA sequences complementary to pVHA55 and pVHA101 DNAs were selected and their fingerprint obtained as described in MATERIALS AND METHODS. The X indicates the position of the bromophenol blue-dye marker. (A), (B) and (C) show the fingerprints of pVHA55-specific RNA from A/Hong Hong/8/68, A/Victoria/112/76 and A/Madrid/672/77 strains. (D), (E) and (F) show the corresponding fingerprints of pVHA101 specific RNA. The arrows indicate the Tl-oligonucleotide changes detected when corresponding RNA sequences of the different viral strains are compared.
the HA sequence. In fact, oligonucleotides 401 and 403 could correspond to positions 1094-1123 and 806-830, respectively (Min Jou et al., 1980). By comparison of the fingerprints shown in Fig. 6, 14 oligonucleotide changes could be detected in the RNA sequence covered by pVHA55 plasmid and 8 in the corresponding pVHA101 sequence. Taking into account the insert sequence lengths and the frac- tion represented in the fingerprints, it is calculated that the mutations observed would correspond to 7.0 and 3.4% of the positions of each RNA region.
DISCUSSION
In the present study we have used the Tl-oligo- nucleotide fingerprinting technique to estimate the extent of RNA sequence heterogeneity among influenza virus strains and field isolates. This tech- nique was proved useful in the detection of genetic changes, even point mutations, in viral RNAs (Flavell et al., 1974; Domingo et al., 1976; 1978; 1980; Young et al., 1979). Since the specific radioactivity of viral RNA was relatively low, no sequence or com-

position data are as yet available for the T1 oligo- nucleotides analyzed, and therefore it has been assumed that the loss of any spot from a given posi-
tion in the fingerprint, as well as the appearance of a new one, represent independent mutations. The genetic distances among viral strains and isolates (Table II) show a fairly good correlation with the time intervals between their isolation dates. With the exception of the A/Texas/I/77 strain, the RNA oligo- nucleotide fingerprint data are compatible with the surmise that all other strains belong to the same evolutionary line; i.e. any oligonucleotide present in A/Hong Kong/8/68 RNA, but absent in A/Victoria/ 3/75 RNA, is no longer present in the corresponding fingerprints from any of the subsequent strains (e.g. spots 4 0 6 4 1 0 in Fig. 3). This is also true for muta- tions observed when any other two strains are com- pared in the evolutionary line (see spot 403). In addi- tion, oligonucleotides that first appear in any given virus strain, such as 431, are generally present in later strains, although this is not the case in a few instances, such as oligonucleotide 433. Strain A/Texas/I/77 has probably evolved from A/Hong Kong/8/68 along a different line, since it retained oligonucleotides 411 and 413 which are absent from any other RNA studied and had lost oligonucleotide 412, otherwise invariant.
The degree of genetic variation observed among viral strains is such that only 9-11 out of 22-29 oligo- nucleotides in the HA, NP or NA RNAs, remained constant in the 1968-77 period, suggesting that even a limited population of co-circulating influenza viruses are genetically heterogeneous. This prediction was tested both at the antigenic and genetic levels, and the results obtained (Table I and Fig. 5) indicate that heterogeneity can be detected among viruses isolated within a fortnight in a limited geographical area.
These observations are compatible with sponta- neous mutation of the influenza virus genes being responsible for the antigenic drift (Webster and Laver, 1975; Young et al., 1979;Min Jou et al., 1980; Laver et al., 1980), although it is not possible to ascertain the contribution of gene reassortment, on the fitness of any given antigenic variant (Desselberger et al., 1978;Young and Palese, 1979).
The relative variability of the different influenza virus genes has been previously estimated both at the protein and the RNA levels. Antigenic variation has been shown to occur, not only at the HA and NA proteins (Schild and Dowdle, 1975), but also at the
329
internal group-specific NP and M antigens (Dimmock
et al., 1977; Brand et al., 1977; Schild et al., 1979). Differences in RNA sequences have been estimated by determining the melting profiles of heterologous RNA hybrids (Scholtissek et al., 1978) or by T1- oligonucleotide fingerprinting of viral RNAs (Young et al., 1979). We have used the latter technique to measure the distribution of the observed mutations in the influenza virus genes. When Tl-oligonucleotide fingerprints of total genomic RNA were compared (Fig. 2), both the number of changes detected and the number of T1 oligonucleotides assigned to each viral gene were too low to enable us to quantitate the genetic variability of the different viral genes. How- ever, no preferential accumulation of mutations could be observed in any of them (Fig. 2). Statistically more significant results were obtained when the same technique was applied to isolated viral RNA segments.
22-29 gene-specific oligonucleotides, comprising 20 to 22% of each gene RNA sequence, were considered for comparisons and the results presented in Table II suggest that there is no significant difference in the genetic variability of HA, NP and NA RNAs. Similar conclusions can be derived from the comparison of t h e M RNA from some of the viral strains studied (data not shown). The differences observed among viruses isolated in Spain in 1977 were located in the HA and the NA genes, but the number of alterations is not large enough for a quantitation of the hetero- geneity of such population of field isolates.
The best known of the influenza virus genes is, by far, the one coding for hemagglutinin. Amino acid sequences for large portions of several hemagglutinin molecules are available (Ward and Dopheide, 1979; Waterfield et al., 1979) and its complete nucleotide sequence has been determined from cloned DNA both for Fowl Plaque and A/Victoria/3/75 viruses (Porter et al., 1979; Min Jou et al., 1980). The pro- rein sequence changes responsible for the selection of variants of A[Memphis/1/71 strain with mono- clonal antibodies, have been located on HA1 (Laver et al., 1979), suggesting that the antigenic properties of hemagglutinin lay on this region of the molecule. In fact, most of the amino acid sequence changes detected among H3N2 viruses during the period 1968-77 were located on the HA1 portion of the hemagglutinin (Laver et al., 1980). However, only a 2 to I ratio in the genetic variability of both HA subunits could be observed, when the Tl-oligonucleo-

330
tide fingerprints of RNA sequences complementary
to plasmids pVHA55 and pVHA101 were analyzed
(Fig. 6). Due to the low number of sequence changes
studied both at the protein and RNA levels, results
are difficult to compare. The possible discrepancy
could be accounted for, however, by assuming that
many of the observed mutat ions at the HA2 region
are silent.
The large genetic heterogeneity observed among
H3N2 influenza viruses suggests that the antigenic
variation of this virus may be a consequence of a
general genetic variability. In fact, the distribution of
spontaneous mutat ions among the viral genes indicates
that the surface antigen genes are not specially muta-
tion-prone regions, but rather that mutat ion frequency
is similar throughout the viral genome. The genetic
variability might be a proper ty of the viral replication
system and not inherent in any viral RNA sequence.
Population immunity has been suggested to account
for the accumulation of mutat ions at the antigenic
sites of hemagglutinin and neuraminidase (Webster
and Laver, 1975). The question remains about the
identi ty of the selection forces which determine a
similar accumulation for other RNA sequences with
no antigenic relevance. One possibility to explain
these experimental results would be that mutat ions
at antigenically irrelevant sequences are fixed in the
viral populations as "passenger" mutat ions due to the
immunological selection of hemagglutinin and/or
neuraminidase. In addition, the founder effect due
to the small size of infecting virus populations in
nature would allow a rapid random drift of muta-
tions.
Similar results for genetic heterogeneity among
viral isolates and distribution of spontaneous muta-
tions have been obtained for Q~ bacteriophage and
foot-and-mouth-disease virus (Domingo et al., 1978;
1980), supporting the notion that a collection of
molecules with slightly different sequences or "quasi-
species" is the true target for selection (Eigen and
Schuster, 1979).
ACKNOWLEDGEMENTS
We are indebted to Dr. E. Vifiuela for helpful sug-
gestions and support. We thank Dr. W. Fiers and
Dr. W. Min Jou for providing us unpublished informa-
tion and bacterial clones containing influenza
sequences. The technical assistance of E. Cano and
A. S~nchez is gratefully acknowledged. This research
was supported by grants from "Comisi6n Asesora
para el Desarrollo de la Investigaci6n Cientl'fica" and
"Comisi6n Administradora del Descuento Comple-
mentario".
REFERENCES
BiUeter, M.A.: Sequence and location of large RNase T1 oligo- nucleotides in bacteriophage Q/3 RNA. J. Biol. Chem. 253 (1978) 8381-8389.
Brand, C.M., Stealy, V.M. and Rowe, J.: Peptide mapping of 12 S i.labeUe d influenza virus proteins. Matrix protein as marker in recombination. J. Gen. Virol. 36 (1977) 385- 394.
CleweU, D.B. and Helinski, D.R.: Supercoiled circular DNA- protein complex in Escherichia coli. Purification and induced conversion to an open circular DNA form. Proc. Natl. Acad. Sci. USA 62 (1969) 1159-1166.
Clewley, J.P., Bishop, D.H.L., Kang, C.Y., Coffin, J., Schnitz- lein, W.M., Reichmann, M.E. and Shope, R.E.: Oligonu- cleotide fingerprints of RNA species obtained from rhabdoviruses belonging to the vesicular stomatitis virus subgroup. J. Virol. 23 (1977) 152-166.
Currier, T.C. and Nester, E.W. : Isolation of covalently closed circular DNA of high molecular weight from bacteria. Anal. Biochem. 76 (1976) 431-441.
Desselberger, U. and Palese, P.: Molecular weights of RNA segments of influenza A and B viruses. Virology 88 (1978) 394-399.
Desselberger, U., Nakajima, K., Alfino, P., Pedersen, F.S. Haseltine, W.A., Hannoun, C. and Palese, P.: Biochemical evidence that new influenza virus strains in nature may arise by recombination (reassortment). Proc. Natl. Acad. Sci. USA 75 (1978) 3341-3345.
de Wachter, R. and Fiers, W. : Preparative two-dimensional polyacrylamide gel electrophoresis of 32P-labelled RNA. Anal. Biochem. 49 (1972) 184-197.
Dimmock, N.J., Carver, A.S., Kennedy, S.I.T., Lee, M.R. and Luscombe, S. Internal proteins of influenza virus: [asS]- methionine peptide maps as genetic markers: J. Gen. Virol. 36 (1977) 503-513.
Domingo, E., FlaveU, R. and Weissmann, C.: In vitro site- directed mutagenesis: generation and properties of an infectious extracistronic mutant of bacteriophage Q#. Gene 1 (1976) 3-25.
Domingo, E., Sabo, D., Taniguchi, T. and Weissmann, C.: Nueleotide sequence heterogeneity of an RNA phage pop- ulation. Cell 13 (1978) 735-744.
Domingo, E., D~vila, M., Ortfn, J.: Nucleotide sequence heterogeneity of the RNA from a natural population of foot-and-mouth-disease virus. Gene 11 (1980) 347-357.

331
Dulbecco, R. and Vogt, M.: Plaque formation and isolation of pure lines with poliomyelitis viruses, J. Exp. Med. 99
(1954) 167-182. Eigen, M. and Schuster, P.: The hypercycle. A principle of
natural self-organization. Springer Verlag, Beling, Heidel- berg, New York, 1979.
Fazekas de St. Groth, S. and Hannoun, C.: S61ection par pression immunologique de mutants dominants du virus de la grippe A (Hong Kong). C.R. Acad. Sci. (Paris) 276D
(1973) 1917-1920. Flavell, R.A., Sabo, D.L., Bandle, E.F. and Weissmann, C.:
Site directed mutagenesis: generation of an extracistronic mutation in bacteriophage Q# RNA. J. Mol. Biol. 89 (1974) 255-272.
Gerhard, W. and Webster, R.G.: Antigenic drift in influenza A viruses. Selection and characterization of antigenic variants of A/PR/8/34 (HONI) influenza virus with monoclonal antibodies. J. Exp. Med. 148 (1978) 383-
392. Holland, J.J., Grabau, E.A., Jones, C.L. and Semier, B.L.:
Evolution of multiple genome mutations during longterm persistent infection by vesicular stomatiris virus. Cell 16
(1979) 495-504. Holmes, D.S. and Bonner, J.: Preparations, molecular weight,
base compositio n and secondary structure of giant nuclear ribonucleic acid. Biochemistry 12 (1973) 2330-2338.
Hrdy, D.B., Rosen, L. and Fields, B.N.: Polymorphism of the migration of double-stranded RNA genome segments of reovirus isolates from humans, cattle and mice. J. Virol. 31 (1979) 104-111.
Inglis, S.C., Barrett, T., Brown, C.M. and Almond, J.W.: The smallest genome RNA segment of influenza virus contains two genes that may overlap. Proc. Natl. Acad. Sci. USA
76 (1979) 3790-3794. Lamb, R.A. and Choppin, P.W. : Segment 8 of the influenza
virus genome is unique in coding for two polypeptides. Proc. Nail. Acad. Sci. USA 76 (1979)4908-4912.
Laver, W.G., Air, G.M., Webster, R.G., Gerhard, W., Ward, C.W. and Dopheide, T.A.: Antigenic drift in type A Influenza virus: sequence differences in the hemagglutinin of Hong Kong (H3N2) variants selected with mono- clonal hybridoma antibodies. Virology 98 (1979) 226-
237. Laver, W.G., Air, G.M., Dopheide, T.A. and Ward, C.W.:
Amino acid sequence changes in the hemagglutinin of A/Hong Kong (H3N2) influenza virus during the period 1968-77. Nature 283 (1980) 454-457.
Min Jou, W., Vethoeyen, M., Devos, R., Saman, E., Rongxi- ang, F., Huylebroeck, D., Fiefs, W., Threlfall, G., Barber, C., Carey, N. and Emtage, S.: Complete structure of the hemagglutinin gene from human influenza A/Victoria/3/ 75 (H3N2) strain as determined from cloned DNA. Cell
19 (1980) 683-696. Nakajima, K., Desselberger, U. and Palese, P.: Recent human
influenza A (H1N1) viruses are closely related generically to strains isolated in 1950. Nature 274 (1978) 334-339.
Palese, P.: The genes of influenza virus. Cell 10 (1977) 1-10. Peacock, A.C. and Dingman, C.W.: Molecular weight estima-
tion and separation of RNA by electrophoresis in poly- acrylamide-agarose gels. Biochemistry 7 (1968) 668-674.
Porter, A.G., Barber, C., Carey, N.H., Haliewell, R.A., Threl- fall, G. and Emtage, J.S.: Complete nucleotide sequence of an influenza virus hemagglurinin gene from cloned DNA. Nature 282 (1979) 471~t77.
Rodger, S.M. and Holmes, I.H.: Comparison of the genomes of simian, bovine and human rotavirus by gel electropho- resis and detection of genome variation among bovine isolates. J. Virol. 30 (1979) 839-846.
Schild, G.C. and Dowdle, W.R. : Influenza virus characteriza- tion and diagnostic serology. In E.D. Kilbourne (Ed.), The Influenza Virus and Influenza, Academic Press, New York, 1975, pp. 315-372.
Schild, G.C., Oxford, J.S. and Newman, R.W.: Evidence for antigenic variation in influenza A nudeoprotein. Virology 93 (1979) 569-573.
Scott, J.V., Stowring, L., Haase, A.T., Narayan, O. and Vigne, R.: Antigenic variation in Visna virus. Cell 18 (1979)
321-327. Scholrissek, C.: The genome of influenza virus. Curl Top.
Microbiol. Immunol. 80 (1978) 139-163. Scholtissck, C., yon Hoyningen, V. and Rott, R.: Genetic
relatedness between the new 1977 epidemic strains (HIN1) of influenza and human influenza strains isolated between 1947 and 1957 (H1N1). Virology 89 (1978) 613-617.
Semler, B.L. and Holland, J.J.: Persistent vesicular stomatitis virus infection mediates base substitutions in viral RNA termini. J. Virol. 32 (1979) 420-428.
Tobita, K., Sugiura, A., Enomoto, C. and Furuyama, M.: Plaque assay and primary isolation of influenza A viruses in an established line of canine kidney ceils (MDCK) in the presence of trypsin. Med. Microbiol. Immunol. 162 (1975) 9-14.
Ward, C.W. and Dopheide, T.A.: Primary structure of the Hong Kong (H3) hemagglurinin. Br. Med. Bull. 35 (1979)
51-56. Waterfleld, M.D., Espelie, K., Elder, K. and Skehel, J.J.:
Structure of the hemagglutinin of influenza virus. Br. Med. Bull. 35 (1979) 57-64.
~Vebster, R.G. and Bean Jr., W.J.: Generics of influenza virus. Annu. Rev. Genet. 12 (1978) 415-431.
Webster, R.G. and Laver, W.G.: Antigenic variation of influenza viruses, in E.D. Kilbourne (Ed.), The Influenza Viruses and Influenza, Academic Press, New York, 1975, pp. 269-314.
Young, J.F. and Palese, P.: Evolution of human influenza A viruses in nature: Recombination contributes to generic variation of H1N1 strains. Proc. Nail. Acad. Sci. USA 76 (1979) 6547-6551.
Young, J.F., Desselberger, U. and Palese, P.: Evolution of human influenza A viruses in nature: sequential mutations in the genomes of new HIN1 isolates. Cell 18 (1979) 7 3 -
83.
Communicated by W. Fiers.




















