
Genetic identification of intracellular trafficking regulators involved in Notch-dependent binary...
16
Journal of Cell Science Genetic identification of intracellular trafficking regulators involved in Notch-dependent binary cell fate acquisition following asymmetric cell division Ste ´ phanie Le Bras 1,2, *, Christine Rondanino 1,2,` , Ge ´ raldine Kriegel-Taki 1,2 , Aurore Dussert 1,2,§ and Roland Le Borgne 1,2, * 1 CNRS, UMR 6061, Institut Ge ´ne ´tique et De ´ veloppement de Rennes, 35043 Rennes, France 2 Universite ´ Rennes 1, UEB, IFR 140, Faculte ´ de Me ´ decine, 35043 Rennes, France ` Present addresses: GReD Laboratory, CNRS UMR 6293, INSERM U1103, Clermont Universite ´ , 63177 Aubie ` re, France; Universite ´ d’Auvergne, Faculte ´ de Me ´ decine, 63000 Clermont-Ferrand, France § Present address: Department of Developmental and Regenerative Biology, Mount Sinai School of Medicine, New York, USA *Authors for correspondence ([email protected]; [email protected]) Accepted 12 June 2012 Journal of Cell Science 125, 4886–4901 ß 2012. Published by The Company of Biologists Ltd doi: 10.1242/jcs.110171 Summary Notch signalling is involved in numerous cellular processes during development and throughout adult life. Although ligands and receptors are largely expressed in the whole organism, activation of Notch receptors only takes place in a subset of cells and/or tissues and is accurately regulated in time and space. Previous studies have demonstrated that endocytosis and recycling of both ligands and/or receptors are essential for this regulation. However, the precise endocytic routes, compartments and regulators involved in the spatiotemporal regulation are largely unknown. In order to identify intracellular trafficking regulators of Notch signalling, we have undertaken a tissue-specific dsRNA genetic screen of candidates potentially involved in endocytosis and recycling within the endolysosomal pathway. dsRNA against 418 genes was induced in the Drosophila melanogaster sensory organ lineage in which Notch signalling regulates binary cell fate acquisition. Gain or loss of Notch signalling phenotypes were observed in adult sensory organs for 113 of them. Furthermore, 26 genes were found to regulate the steady state localisation of Notch, Sanpodo, a Notch co-factor, and/or Delta in the pupal lineage. In particular, we identified 20 genes with previously unknown function in D. melanogaster intracellular trafficking. Among them, we identified CG2747 and we show that it regulates the localisation of clathrin adaptor AP-1 complex, a negative regulator of Notch signalling. Together, our results further demonstrate the essential function of intracellular trafficking in regulating Notch-signalling-dependent binary cell fate acquisition and constitute an additional step toward the elucidation of the routes followed by Notch receptor and ligands during signalling. Key words: Notch, Endocytosis, Recycling, Intracellular trafficking, AP-1 Introduction Notch cell-cell signalling is required in a vast majority of developmental processes and during the adult life of many organisms. It regulates cell fate specification as well as stem cell behaviour and defects can lead to numerous developmental pathologies and cancers underlying its crucial role (reviewed by Gridley, 2003; Miele et al., 2006). The challenging question is to understand the mechanisms allowing one cell to act as a signalling cell and the other one as the receiving cell, when both cells can potentially express both ligands and receptors. Although it can be performed through a spatial and temporal regulation of their expression, DSL (Delta, Serrate, Lag2) ligand and Notch receptor differential expression could not be sufficient to explain the subtle directionality of Notch signalling. In this context, regulation of the availability of both receptors and DSL ligands at the cell surface appears crucial to ensure a proper Notch signalling activation. Therefore ligand and receptor post- translational modifications and trafficking are emerging as crucial regulatory mechanisms. Several lines of evidence suggest that endocytic trafficking of DSL ligands enhances their signalling activity while receptor trafficking insures their steady state level at the cell surface thereby regulating their availability for ligand binding (reviewed by Bray, 2006; Fu ¨rthauer and Gonza ´lez-Gaita ´n, 2009; Kopan and Ilagan, 2009; Le Borgne, 2006; Weinmaster and Fischer, 2011; Yamamoto et al., 2010). Although recycling of DSL ligands is necessary to produce an active DSL ligand, the nature of this maturation is still poorly characterised and two models are actually favoured: endocytosis and pulling forces (Klueg and Muskavitch, 1999; Nichols et al., 2007; Windler and Bilder, 2010) versus endocytosis and recycling (Benhra et al., 2010; Emery et al., 2005; Jafar-Nejad et al., 2005; Le Borgne and Schweisguth, 2003; Rajan et al., 2009; Wang and Struhl, 2004). The cellular context dependence could account for these two non- mutually exclusive models and the Drosophila melanogaster sensory organ lineage, in which Notch unidirectional signalling is the only pathway involved (Heitzler and Simpson, 1991), represents an interesting study model in which the signal sending and receiving cells are easily distinguishable. Each sensory organ, present on the adult D. melanogaster notum, is derived from a single precursor cell (pI), which undergoes a stereotyped series of four asymmetric cell divisions 4886 Research Article
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Genetic identification of intracellular trafficking regulators involved in Notch-dependent binary...

Journ
alof
Cell
Scie
nce
Genetic identification of intracellular traffickingregulators involved in Notch-dependent binary cell fateacquisition following asymmetric cell division
Stephanie Le Bras1,2,*, Christine Rondanino1,2,`, Geraldine Kriegel-Taki1,2, Aurore Dussert1,2,§ andRoland Le Borgne1,2,*1CNRS, UMR 6061, Institut Genetique et Developpement de Rennes, 35043 Rennes, France2Universite Rennes 1, UEB, IFR 140, Faculte de Medecine, 35043 Rennes, France`Present addresses: GReD Laboratory, CNRS UMR 6293, INSERM U1103, Clermont Universite, 63177 Aubiere, France; Universite d’Auvergne, Faculte de Medecine, 63000Clermont-Ferrand, France§Present address: Department of Developmental and Regenerative Biology, Mount Sinai School of Medicine, New York, USA*Authors for correspondence ([email protected]; [email protected])
Accepted 12 June 2012Journal of Cell Science 125, 4886–4901� 2012. Published by The Company of Biologists Ltddoi: 10.1242/jcs.110171
SummaryNotch signalling is involved in numerous cellular processes during development and throughout adult life. Although ligands andreceptors are largely expressed in the whole organism, activation of Notch receptors only takes place in a subset of cells and/or tissuesand is accurately regulated in time and space. Previous studies have demonstrated that endocytosis and recycling of both ligands and/or
receptors are essential for this regulation. However, the precise endocytic routes, compartments and regulators involved in thespatiotemporal regulation are largely unknown. In order to identify intracellular trafficking regulators of Notch signalling, we haveundertaken a tissue-specific dsRNA genetic screen of candidates potentially involved in endocytosis and recycling within the
endolysosomal pathway. dsRNA against 418 genes was induced in the Drosophila melanogaster sensory organ lineage in which Notchsignalling regulates binary cell fate acquisition. Gain or loss of Notch signalling phenotypes were observed in adult sensory organs for113 of them. Furthermore, 26 genes were found to regulate the steady state localisation of Notch, Sanpodo, a Notch co-factor, and/orDelta in the pupal lineage. In particular, we identified 20 genes with previously unknown function in D. melanogaster intracellular
trafficking. Among them, we identified CG2747 and we show that it regulates the localisation of clathrin adaptor AP-1 complex, anegative regulator of Notch signalling. Together, our results further demonstrate the essential function of intracellular trafficking inregulating Notch-signalling-dependent binary cell fate acquisition and constitute an additional step toward the elucidation of the routes
followed by Notch receptor and ligands during signalling.
Key words: Notch, Endocytosis, Recycling, Intracellular trafficking, AP-1
IntroductionNotch cell-cell signalling is required in a vast majority ofdevelopmental processes and during the adult life of manyorganisms. It regulates cell fate specification as well as stem cell
behaviour and defects can lead to numerous developmentalpathologies and cancers underlying its crucial role (reviewed byGridley, 2003; Miele et al., 2006). The challenging question is to
understand the mechanisms allowing one cell to act as asignalling cell and the other one as the receiving cell, whenboth cells can potentially express both ligands and receptors.
Although it can be performed through a spatial and temporalregulation of their expression, DSL (Delta, Serrate, Lag2) ligandand Notch receptor differential expression could not be sufficient
to explain the subtle directionality of Notch signalling. In thiscontext, regulation of the availability of both receptors and DSLligands at the cell surface appears crucial to ensure a properNotch signalling activation. Therefore ligand and receptor post-
translational modifications and trafficking are emerging ascrucial regulatory mechanisms.
Several lines of evidence suggest that endocytic trafficking ofDSL ligands enhances their signalling activity while receptor
trafficking insures their steady state level at the cell surface
thereby regulating their availability for ligand binding (reviewedby Bray, 2006; Furthauer and Gonzalez-Gaitan, 2009; Kopan andIlagan, 2009; Le Borgne, 2006; Weinmaster and Fischer, 2011;Yamamoto et al., 2010). Although recycling of DSL ligands is
necessary to produce an active DSL ligand, the nature of thismaturation is still poorly characterised and two models areactually favoured: endocytosis and pulling forces (Klueg and
Muskavitch, 1999; Nichols et al., 2007; Windler and Bilder,2010) versus endocytosis and recycling (Benhra et al., 2010;Emery et al., 2005; Jafar-Nejad et al., 2005; Le Borgne and
Schweisguth, 2003; Rajan et al., 2009; Wang and Struhl, 2004).The cellular context dependence could account for these two non-mutually exclusive models and the Drosophila melanogaster
sensory organ lineage, in which Notch unidirectional signalling isthe only pathway involved (Heitzler and Simpson, 1991),represents an interesting study model in which the signalsending and receiving cells are easily distinguishable.
Each sensory organ, present on the adult D. melanogaster
notum, is derived from a single precursor cell (pI), whichundergoes a stereotyped series of four asymmetric cell divisions
4886 Research Article

Journ
alof
Cell
Scie
nce
to generate five different cells, four composing the
mechanosensory bristle and a glial cell (Fig. 1A,B). During each
division, Notch signalling is involved in cell fate acquisition. For
example, Notch is inhibited in the pI daughter cell, which adopts
the pIIb cell identity and eventually activates Notch signalling in
the adjacent daughter cell becoming the pIIa cell. Although data
from different laboratories have emphasised the role of
intracellular trafficking in the uni-directionality of Notch
signalling between these two daughter cells (Benhra et al., 2011;
Benhra et al., 2010; Berdnik et al., 2002; Coumailleau et al., 2009;
Couturier et al., 2012; Djiane et al., 2011; Emery et al., 2005;
Gallagher and Knoblich, 2006; Hutterer and Knoblich, 2005; Jafar-
Nejad et al., 2005; Langevin et al., 2005; Rajan et al., 2009;
Roegiers et al., 2005; Tong et al., 2010), little is known and
understood about the regulators and membrane compartments
involved in this process during the pI mitosis and/or in each of its
daughter cells. Nonetheless, some recent data have emphasised the
importance of a pI-daughter-cell-specific intracellular trafficking
of Delta, Notch and/or a Drosophila Notch co-factor, Sanpodo
(Spdo) (O’Connor-Giles and Skeath, 2003). In the signal sending
pIIb cell, both basal to apical transcytosis of Delta mediated by
Neuralized (see above and Benhra et al., 2010) and its trafficking
toward an apical Actin Rich Structure (ARS) driven by WASp and
the Arp2/3 complex (Rajan et al., 2009) are required for proper
Notch signalling activation. While in the receiving cell, the
clathrin adaptor complex AP-1 was genetically shown to be
required for the correct localisation of Notch and Spdo (Benhra
et al., 2011).
In order to identify novel regulators of the intracellular
trafficking of Notch signalling major components, we have
undertaken a tissue-specific double-strand RNA (dsRNA) genetic
screen of 418 genes potentially involved in endocytosis and/or
recycling within the endolysosomal pathway. To validate our
in vivo Notch-specific strategy, 50 previously known Notch
signalling regulators were screened, including 24 for which the
function has not yet been studied during sensory organ lineage
development. We took advantage of the fact that the genetic
impairment of Notch signalling directly affects the development
of external sensory organs and therefore allows for adult
phenotype screening (Hartenstein and Posakony, 1990). Among
the 113 Notch regulators identified based on adult phenotype, 61
were screened for, and 26 were found to cause a change in the
steady state localisation of Notch, Sanpodo and/or Delta, in the
pupal sensory organ lineage. In particular, we identified genes
with previously unknown function in intracellular trafficking in
Drosophila melanogaster such as CG27247 a regulator of AP-1
localisation, CG7787 putatively involved in the recycling
pathway and members of the Tetraspanin family.
Fig. 1. Sensory organ lineage and screen results. (A) Diagram
of the adult sensory organ composed of two external cells (shaft
and socket) and two internal cells (sheath and neuron). (B) Scheme
of the cell precursor pI pupal lineage leading to the specification of
the adult sensory organ cells and one apoptotic glial cell after four
asymmetric cell divisions. In A and B, blue nuclei indicate cells
responding to Notch signalling and red nuclei indicate cells
sending Notch signals. (C–F). Examples of Notch-like bristle
phenotype screened for, in the dsRNA genetic screen induced in the
Drosophila notum. (D9,F9) Scheme of putative pI pupal lineages in
case of a loss (D9) or gain (F9) of Notch signalling in all or some of
the asymmetric cell divisions. (G) Numbers of candidates with
dsRNA-induced adult phenotypes for each screen category (dark
grey box: candidates with phenotype; light grey box: known Notch
regulators with phenotype). Numbers into brackets indicate
candidate genes/known Notch regulators screened in each category.
Notch intracellular trafficking regulators 4887

Journ
alof
Cell
Scie
nce
ResultsPrinciple and validation of gene silencing-inducible screen
To screen specifically in the sensory organ lineage, we made use of
a well-characterised and previously described dsRNA in vivo
strategy (Mummery-Widmer et al., 2009). Taking advantage of the
GAL4–UAS binary expression system (Brand and Perrimon,
1993), we induced gene silencing of selected genes specifically in
the notum where the sensory organs develop. To do so, transgenic
females carrying GAL4 under the control of a sensory organ
promoter were crossed with males carrying an upstream activating
sequence (UAS)–dsRNA transgenic construct. In the F1 progeny,
GAL4 specifically activates the UAS and eventually induces gene
silencing in the fly notum during sensory organ development. For
each cross, two experimenters analysed at least 20 F1 progenies
blindly. In order to identify specific regulators of Notch signalling,
we scored for bristle phenotype on the notum (Fig. 1D–F). While a
loss of bristle and/or double shafts without a socket cell reflects a
loss of Notch signalling in the sensory organ lineage (Fig. 1D,D9),
an excess of socket cells and/or double shafts with socket cells is
correlated with a gain of Notch signalling in the sensory organ
lineage (Fig. 1F,F9). As Notch signalling is also involved in the
process of pI specification, we could, additionally, score for an
excess of sensory organs reflecting a loss of Notch signalling in
lateral inhibition (Fig. 1E). A genome-wide dsRNA screen was
previously performed to identify regulators of Notch signalling in
the sensory organs in which one sensory organ driver-GAL4 was
used: pannier (pnr)–GAL4 (Mummery-Widmer et al., 2009). In
this previous screen, we noticed that the phenotype observed for
360 (86%) of our genes could not be assessed as expression of the
dsRNA induced either lethality or a morphological defect of the
notum. This observation led us to modulate the strength of gene
silencing by placing the F1 progenies at 18, 25 or 29 C. As the
efficiency of the GAL4–UAS system is partially temperature
sensitive (Mondal et al., 2007), this allows inducing lower (at
18 C) or higher (at 29 C) dsRNA expression. Additionally, to
circumvent any technical bias of GAL4–UAS-induced phenotype
and further describe the Notch-like phenotype, we independently
used two GAL4 transgenic constructs which both drive expression
in the notum during development: apterous (ap)–GAL4 (Calleja
et al., 1996) and scabrous (sca)–GAL4 (Mlodzik et al., 1990).
In order to validate our Notch signalling-specific strategy
(Knoblich, 2010), we choose to screen 50 known Notch
signalling regulators and observe the same phenotype than
previously described for 24 of them (see supplementary material
Table S1). Although we could not reproduce the Notch loss-of-
function-like phenotype of only two known Notch regulators,
aristaless (Kojima et al., 2005) and Liquid facets (Wang and
Struhl, 2004), our data indicate that our strategy allows specific
screening for Notch regulators in the sensory organ lineage as
previously described (Mummery-Widmer et al., 2009).
Interestingly, we also observed a bristle phenotype for 14 of
the 24 known Notch regulators whose function in the sensory
organ lineage had not been previously described. Not all the
known Notch signalling regulators appear to be involved in the
Drosophila sensory organ lineage, which further highlights
the cellular context dependence of Notch signalling in vivo as
previously reported (Fuwa et al., 2006). For example, dsRNA
against Kurtz and Nedd4 did not induce an adult phenotype while
they are negative regulators of Notch signalling in the Drosophila
wing vein (Mukherjee et al., 2005; Sakata et al., 2004).
Identification of Notch signalling regulators
To identify intracellular trafficking regulators of Notch signallingin Drosophila melanogaster sensory organs, we specificallyscreened for 368 genes from the endolysosomal pathway (seesupplementary material Table S2). We selected these genes among
members of intracellular trafficking regulator families mostlyidentified from yeast genetics and involved in different traffickingaspects such as, coat components [clathrin mediated endocytosis
(Maldonado-Baez and Wendland, 2006)], lipid microdomainorganisation [non-clathrin mediated endocytosis (Simons andGerl, 2010)], cytoskeleton [actin, myosin and/or microtubules
(Hehnly and Stamnes, 2007)], small GTPases, ubiquitylation/deubiquitylation factors involved in vesicle targeting (Murphyet al., 2009; Wennerberg et al., 2005), Endosomal SortingComplex Required for Transport (ESCRT) complexes (Henne
et al., 2011), membrane recognition and/or fusion regulators [suchas SNAP receptors, SNAREs (Malsam et al., 2008), Exocyst (Hsuet al., 2004)] and ATPases (Forgac, 2007). We also based our
selection on gene ontology (GO) annotation from FlyBase (usingthe GO terms: endocytosis, endosomal sorting, secretion) andselected putative orthologue(s) of traffic regulators identified in a
Caenorhabditis elegans genetic screen (Balklava et al., 2007) ormammal proteomic screens (Baust et al., 2008; Baust et al., 2006).Noteworthy, the molecular function of 54 of these genes has not
yet been defined in D. melanogaster (‘novel unknown function’category in supplementary material Table S2).
To perform our screen we used 716 dsRNA lines, whichrepresent the 368 candidates and 50 known Notch regulators, as we
systematically screened with up to five different dsRNA lines,when available, in order to circumvent any effect due to thedsRNA construct insertion site. Expression of 264/716 (36.8%)
dsRNA lines induced a bristle phenotype or lethality with eitherboth or one of the GAL4 lines in our screen conditions. When twoor more dsRNA lines induced a bristle phenotype, it wasconsistently the same gain or loss of Notch signalling
phenotype(s), depending on the candidate, as we never observedopposite bristle phenotype between different dsRNA lines againstthe same candidate.
In order to confirm and validate the results, we reproduced thescreen, with all the positive and lethal dsRNA hits and somenegative ones as controls, using the same GAL4 lines and up totwo additional GAL4 also driving expression in the notum: Eq–
GAL4 (Pi et al., 2001) and/or pnr–GAL4 (Calleja et al., 1996).Among the dsRNA lines individually crossed with these severalGAL4 lines, we observed that 175/264 (66.3%) dsRNA lines,
representing 113 candidates, induced a reproducible bristlephenotype with one or more GAL4 (see supplementarymaterial Table S2). To further validate our results, we had
included 52 dsRNA lines, obtained from the National Institute ofGenetics Fly Stock Center (NIG-Fly) or the Transgenic RNAiProject (TRIP), which target different part of the candidate RNA
sequence than the dsRNA lines from the Vienna DrosophilaRNAi Center (VDRC). In doing so, we confirmed the specificNotch-like bristle or lack of phenotype observed with the VDRCdsRNA lines (see supplementary material Table S2).
In the end, we firmly identified 113 Notch regulators in thesensory organ lineage (Table 1), which belong to the differentscreening categories that we initially defined (Fig. 1G).
Specifically, we identified 77 previously unknown regulators ofthe Notch signalling pathway with a role in the sensory organlineage. These regulators belong to all our initial screen categories,
Journal of Cell Science 125 (20)4888

Journ
alof
Cell
Scie
nce
Table 1. Complete list of positive hits from the genetic dsRNA screen
Screen category/molecular function CG number Name
Notum adult phenotype
Excess of organs: lateralinhibition defect [Notch
signalling loss-of-function(LOF)-like]
Sensory organ defect: Notchsignalling gain-of-function(GOF)- or loss-of-function
(LOF)-like phenotypes
Coat components and accessory proteinsClathrin complex CG6948 Clathrin light chain LOF: excess of organs
(weak)n.o.
Clathrin binding CG2520 like-AP180 LOF: excess of organs(weak)
n.o.
Clathrin adaptor complex (AP-1) CG9113 AP-1c n.o. GOF: excess of socket cells,double shafts with socket cell
Clathrin adaptor complex (AP-1) CG5864 AP-1s n.o. GOF: excess of socket cells,double shafts with socket cell
Clathrin adaptor complex (AP-1) CG9388 AP-47 n.o. GOF: excess of socket cells,double shafts with socket cell
Clathrin adaptor complex (AP-2) CG7057 AP-50 n.o. GOF: excess of socket cells,double shafts with socket cell
Clathrin adaptor complex (AP-2) CG4260 a-adaptin n.o. GOF: excess of socket cells,double shafts with socket cell
Clathrin adaptor complex (AP-1 and AP-2) CG12532 b-adaptin n.o. GOF: excess of socket cellsUnknown (AP-1 accessory protein: putative
orthologue of S. cerevisiae Laa1p)CG2747 CG2747 n.o. GOF: excess of socket cells,
double shafts with socket cellLipid microdomain organization/interactors
Multispan membrane protein withunknown function
CG7740 prominin-like n.d. LOF: bristle loss
Tetraspanin CG10742 Tetraspanin 3A LOF: excess of organs(weak)
n.o.
Tetraspanin CG32136 Tetraspanin 68C LOF: excess of organs* LOF: bristle lossTetraspanin CG9496 Tetraspanin 29 Fb n.d. LOF: bristle lossTetraspanin CG12847 Tetraspanin 42Ec n.d. LOF: bristle lossTetraspanin CG12143 Tetraspanin 42Ej n.d. LOF: bristle lossTetraspanin CG12841 Tetraspanin 42Ek n.d. LOF: bristle lossTetraspanin CG9033 Tetraspanin 47F n.d.* LOF: bristle lossTetraspanin CG9494 Tetraspanin 29 Fa n.o. GOF: excess of socket cells
Cytoskeleton, regulators or interactorsActin binding CG5695 Jaguar n.d. GOF: excess of socket cellsActin binding CG1520 WASp n.d. LOF: bristle lossActin binding; microtubule-associated protein CG6450 lava lamp LOF: excess of organs n.o.WASp activation complex CG4931 specifically Rac1-associated
protein 1/CYFIPLOF: excess of organs* LOF: bristle loss
Arp2/3 actin binding complex CG4560 Arpc3A LOF: excess of organs n.o.Arp2/3 actin binding complex CG10954 Arc-p34 LOF: excess of organs LOF: bristle loss, double shaftsArp2/3 actin binding complex CG8978 Suppressor of profilin
2/Arpc1n.d. LOF: bristle loss
GTPase activity; actin binding,microtubule-associated protein
CG8705 peanut n.d. LOF: bristle loss
GTPase activity; microtubule-associated protein CG4173 Septin 2 n.d. LOF: bristle lossGTPase activity; microtubule-associated protein CG2916 Septin 5 n.d.* LOF: bristle lossDNA processing (replication) CG1584 Origin recognition
complex subunit 6n.d. LOF: bristle loss, double shafts
without socket cellRNA processing (Transcription factor) CG7238 Septin interacting protein 1 n.d. LOF: bristle lossDynein complex CG6998 cut up n.d. LOF: bristle lossEpsin-like CG42250 liquid facets-Related n.d.* LOF: bristle lossMicrotubule-associated protein CG3265 Eb1 n.d. LOF: bristle loss
Small GTPases, GEF/GAP or effectorsGTPase activity (Rab) CG17515 Rab21 LOF: excess of organs* LOF: bristle lossGTPase activity (Rab) CG9575 Rab35 LOF: excess of organs
(weak)LOF: bristle loss
GTPase activity (Rab) CG5771 Rab-protein 11 n.d. LOF: bristle lossGTPase activity (Ras) CG8416 Rho1 n.d. LOF: bristle lossGTPase activity (ARF) CG11027 ADP ribosylation factor 102F n.d.* LOF: bristle lossGTPase activity (ARF) CG7435 ADP ribosylation factor 84F n.o.* GOF: excess of socket cellsGTPase (Rap) activator activity (GAP) CG6975 gigas LOF: excess of organs
(weak)LOF: bristle loss
GTPase (Rho) activator activity (GAP) CG13345 tumbleweed n.d. LOF: bristle lossGuanyl-nucleotide exchange factor activity (GEF) CG7787 CG7787 LOF: excess of organs* LOF: bristle loss, double shaftsGuanyl-nucleotide exchange factor activity (GEF) CG15797 ric8a LOF: excess of organs n.o.
Table 1. Continued on next page
Notch intracellular trafficking regulators 4889

Journ
alof
Cell
Scie
nce
Screen category/molecular function CG number Name
Notum adult phenotype
Excess of organs: lateralinhibition defect [Notch
signalling loss-of-function(LOF)-like]
Sensory organ defect: Notchsignalling gain-of-function(GOF)- or loss-of-function
(LOF)-like phenotypes
Guanyl-nucleotide exchange factor activity (GEF) CG9491 Gef26 LOF: excess of organs(weak)
LOF: bristle loss
Guanyl-nucleotide exchange factor activity(GEF)
CG31048 CG31048 n.d. LOF: bristle loss
Guanyl-nucleotide exchange factor activity(GEF)
CG32239 Guanine nucleotide exchangefactor GEF64C
n.d. LOF: bristle loss
Guanyl-nucleotide exchange factor activity(GEF)
CG8114 Pebble n.d. LOF: bristle loss
GTPase (Rab5) effector protein CG8506 Rabenosyn-5 n.d. LOF: bristle lossGTPase (Ras) binding CG10255 Lap1 n.d. LOF: bristle loss
Ubiquitylation/deubiquitylationUbiquitin ligase complex (SCF); microtubule-
associated proteinCG16983 skpA n.d. LOF: bristle loss, double shafts
Ubiquitin ligase complex (SCF) CG8881 skpB n.d. LOF: bristle lossUbiquitin ligase complex (SCF) CG12700 skpD n.d. LOF: bristle lossUbiquitin ligase complex (SCF) CG11942 skpE n.d. LOF: bristle lossUbiquitin ligase (E3) CG5841 mind bomb 1 LOF: excess of organs n.o.Ubiquitin ligase (E3) CG11861 Cullin-3/Guftagu n.d.* LOF: bristle loss, double shaftsUbiquitin ligase (E3) CG11988 neuralized n.d. LOF: bristle loss, double shafts
ESCRT components and interactorsVacuolar protein sorting (Class E/ESCRT-0) CG2903 Hepatocyte growth factor
regulated tyrosine kinasesubstrate
n.d. LOF: bristle loss, double shafts
Vacuolar protein sorting (Class E/ESCRT-0) CG6521 Signal transducing adaptormolecule
LOF: excess of organs* GOF: excess of socket cells?
Vacuolar protein sorting (Class E/ESCRT-I) CG12770 Vacuolar protein sorting 28 n.o. GOF: excess of socket cellsVacuolar protein sorting (Class E/ESCRT-II) CG14750 Vacuolar protein sorting 25 LOF: excess of organs LOF: bristle lossVacuolar protein sorting (Class E/ESCRT-III) CG4071 Vacuolar protein sorting 20 LOF: excess of organs LOF: bristle lossVacuolar protein sorting (Class E/ESCRT-III) CG4108 Chmp1 LOF: excess of organs GOF: excess of socket cellsVacuolar protein sorting (Class E/ESCRT-III) CG14542 Vps2 LOF: excess of organs* GOF: excess of socket cells,
double shaftsATPase CG6842 Vacuolar protein sorting 4 n.o. GOF: excess of socket cells?
Membrane recognition and/or fusionExocyst complex CG6095 Exo84 LOF: excess of organs* LOF: bristle loss and GOF:
excess of socket cells, doubleshafts
Exocyst complex CG8843 Sec5 n.d.* GOF: double shafts with socketcell, loss of shaft cells
Exocyst complex CG5341 Sec6 n.d.* GOF: loss of shaft cellsExocyst complex CG7034 Sec15 LOF: excess of organs* LOF: bristle lossSNAP receptor activity (SNARE) CG1515 lethal (1) G0155 n.d.* LOF: bristle lossSNAP receptor activity (SNARE) CG4109 Syntaxin 8 n.d. LOF: bristle lossSNAP receptor activity (v-SNARE) CG12210 synaptobrevin n.d. LOF: bristle loss, double shaftsSNAP receptor activity (t-SNARE) CG5081 Syntaxin 7/Avalanche LOF: excess of organs GOF: excess of socket cells
ATPasesATPase CG9931 NEM-sensitive fusion protein 2 n.d.* LOF: bristle lossVacuolar ATPase (subunit c) CG3161 Vacuolar H+ ATPase subunit
16-1n.d. LOF: bristle loss
Vacuolar ATPase (subunit c) CG32089 Vacuolar H+ ATPase subunit16-2
n.d. LOF: bristle loss
Vacuolar ATPase (subunit c9) CG7007 Vacuolar H+ ATPase subunitPPA1-1
LOF: excess of organs LOF: bristle loss
Vacuolar ATPase (subunit A) CG3762 Vacuolar H+ ATPase subunit68-2
n.d. LOF: bristle loss
Vacuolar ATPase (subunit E) CG1088 Vacuolar H+ ATPase 26kD Esubunit
n.d. LOF: bristle loss
Vacuolar ATPase (subunit G) CG6213 Vacuolar H+ ATPase G-subunit n.d. LOF: bristle lossVarious
Calcium channel activity CG34146 bruchpilot n.d. LOF: bristle lossCalcium ion binding CG9126 Stromal interaction molecule n.d.* LOF: bristle loss, double shaftsCatalytic activity: Notch fucosyltransferase CG12366 O-fucosyltransferase 1 n.d.* LOF: bristle loss, double shaftsCatalytic activity: Notch glucosyltransferase CG31152 rumi n.d. LOF: bristle loss
Table 1. Continued
Table 1. Continued on next page
Journal of Cell Science 125 (20)4890

Journ
alof
Cell
Scie
nce
which cover various aspects of intracellular trafficking. The vast
majority of the observed phenotypes resemble those of a loss of
Notch signalling. Nevertheless, phenotypes similar to gain-of-
Notch signalling were observed for 20 genes from various screen
categories including members of coat components (AP-1 and AP-
2) or ESCRT complexes (0, I and III). Both AP-1 and AP-2
complexes had previously been identified as regulators of Spdo
trafficking and eventually as negative regulators of Notch
signalling pathway during binary cell fate decision (Benhra et al.,
2011; Berdnik et al., 2002; Tong et al., 2010). Therefore, our
genetic screen clearly led to the identification of potential
intracellular trafficking regulators directly involved in the
regulation of Notch signalling via its major components.
Steady-state localisation of Notch, Sanpodo, Delta and cell
fate identity
A Notch-like adult sensory organ phenotype could be due to a
defect in Notch signalling component traffic and/or induced by
unrelated defects such as in cell fate determinant segregation, cell
polarity, cell cycle control and/or general intracellular trafficking.
Out of the 113 candidates that we genetically identified as Notch
signalling regulators in the sensory organ lineage, we wanted to
identify those involved in the intracellular trafficking of the major
components of Notch signalling: Delta, Notch and its co-factor
Spdo. This study was made feasible as they present a specific
steady-state pattern of subcellular localisation in the sensory organ
pupal lineage during pI mitosis and at the pI daughter cell stage
Screen category/molecular function CG number Name
Notum adult phenotype
Excess of organs: lateralinhibition defect [Notch
signalling loss-of-function(LOF)-like]
Sensory organ defect: Notchsignalling gain-of-function(GOF)- or loss-of-function
(LOF)-like phenotypes
Catalytic activity: Notch c-secretase CG18803 presenilin n.d. LOF: bristle lossCatalytic activity: histone methyltransferase CG10955 Rtf1 n.d. LOF: bristle lossGuanylate protein kinase CG31349 polychaetoid LOF: excess of organs n.o.Serine/threonine protein kinase; chromosomal
passenger complex (CPC)CG6620 IplI-aurora-like kinase/Aurora B n.d. LOF: bristle loss
Protein binding: TGFb CG14026 thickveins LOF: excess of organs n.o.Protein binding CG9695 Disabled LOF: excess of organs n.o.Protein binding: Notch ligand activity CG3619 Delta LOF: excess of organs LOF: bristle lossProtein binding: Notch receptor activity CG3936 Notch n.d. LOF: bristle lossProtein binding: Notch CG31020 Sanpodo n.d. LOF: bristle loss, double shaftsProtein binding CG9834 endophilin B n.d. LOF: bristle lossProtein binding CG5820 Gp150 n.d. LOF: bristle loss
Cell polarityMyosin complex CG15792 zipper n.d. LOF: bristle lossMyosin binding CG2671 lethal (2) giant larvae n.o. GOF: excess of socket cellsProtein binding CG5055 bazooka n.d. LOF: bristle lossProtein binding CG6383 crumbs n.d. LOF: bristle lossProtein binding CG5884 par-6 n.o. GOF: excess of socket cellsSerine/threonine protein kinase CG3068 Aurora/aurora A LOF: excess of organs GOF: double shafts with socket
cellNovel unknown function
DNA processing (DNA helicase) CG18013 Psf2 n.d. LOF: bristle loss, double shaftswithout socket cell
Microtubule-associated protein CG8014 Receptor mediated endocytosis8
n.d. LOF: bristle loss
Protein binding CG6834 CG6834 n.d. LOF: bristle lossProtein binding: Notch CG2863 Notchless n.d. LOF: bristle lossRNA processing (snoRNA binding) CG10341 CG10341 n.o. GOF: double shafts with socket
cell, loss of shaft cellsRNA processing (transcription factor binding) CG7583 C-terminal binding protein n.o. GOF: excess of socket cellsRNA processing (transcription factor) CG5461 bunched n.d. LOF: bristle loss (weak)RNA processing (transcription factor) CG4882 CG4882 n.d. LOF: bristle lossRNA processing (transcription factor) CG4029 jumeau n.d. LOF: bristle loss, double shaftsRNA processing (translation elongation
factor); microtubule-associated proteinCG11901 Ef1c n.d. LOF: bristle loss
Unknown CG11295 lethal-(2)-denticleless n.d. LOF: bristle lossUnknown CG8435 CG8435 n.d.* LOF: bristle lossUnknown CG8639 Cirl n.d. LOF: bristle loss
Genes are listed in order of the screen category that they were initially selected in. The notum adult phenotypes are sub-divided into two major processescontrolled by Notch signalling: lateral inhibition regulating the number of sensory organ precursors specified, and sensory organ lineage, which controls themorphology of the adult organs. The adult phenotypes are indicated as LOF pI (excess of organs resulting from a lateral inhibition defect), LOF (loss of Notchsignalling-like phenotypes) and/or GOF (gain of Notch signalling-like phenotypes).
n.o., no phenotype observed; n.d., phenotype could not be determined because of bald cuticle (phenotype of bristle loss); *lateral inhibition defect detected onpupal notum by immunostaining.
Table 1. Continued
Notch intracellular trafficking regulators 4891

Journ
alof
Cell
Scie
nce
(Fig. 2A–F90) (also see Benhra et al., 2010). In a wild-type lineage,
while both apical Delta and Notch are mostly localised at the
cortex (Fig. 2A0,A90, D0,D90), basolateral Delta is found in vesicles
in mitotic pI and pIIb/pIIa cells (Fig. 2B0,C0,E0,F0). Spdo has a
more dynamic pattern of localisation: cytoplasmic in the mitotic pI
(Fig. 2A9–C9), its localisation becomes asymmetric in the pI
daughter cells. While Spdo is enriched along the apicobasal
interface of pI daughter cells (Fig. 2D9–F9), Spdo is mostly
localised in vesicles in the anterior pIIb cell but at the basolateral
plasma membrane in the posterior pIIa cell (Fig. 2E9). Changes in
Notch, Spdo and/or Delta localisation could either originate from
an aberrant cell-fate identity acquisition in the lineage (two pIIb or
pIIa-like cells) or reflect trafficking defect(s) causing a defective
Notch signalling pathway. As a proof of principle, we recently
demonstrated that the clathrin adaptor complex AP-1, identified in
this screen, controls Spdo and Notch trafficking in the sensory
organ lineage. In particular, a lack of AP-1 function induces Spdo
and Notch subcellular localisation changes and an adult gain of
Notch signalling phenotype (Benhra et al., 2011). Similarly, loss of
Neur, Sec15 or Arp2/3 functions induce changes in Spdo and/or
Delta subcellular localisation correlated with adult loss of Notch
signalling phenotypes (Benhra et al., 2010; Jafar-Nejad et al.,
2005; Le Borgne and Schweisguth, 2003; Rajan et al., 2009;
Roegiers et al., 2005).
Regulators of Notch, Sanpodo and/or Delta subcellular
localisation identified in the screen
Among the 113 Notch regulators we identified, we decided to
analyse those that were not previously known to cause
subcellular localisation changes and/or that do not have
described function in cell polarity or in asymmetric cell
division (see supplementary material Table S3). Among the 61
genes that we screened for a dsRNA-induced change in Delta,
Notch and/or Spdo steady-state localisation (using one dsRNA
line for each), 32 did not cause any visible defect while three
genes (gigas, CG31048 and CG8435) caused a lack of pI
specification (revealed by an absence of Spdo staining, our
sensory organ identity marker), which explains the observed
adult bristle loss phenotype (see supplementary material Table
S3). Twenty-six genes caused a phenotype of Notch, Spdo and/or
Delta mis-localisation at the pI and/or pI daughter cell stage (we
used the threshold of at least three, out of 20 analysed, sensory
organs presenting the same phenotype on two different nota).
Although a wide range of phenotypes was observed, they can be
subdivided into three major categories (Table 2; Figs 3–5).
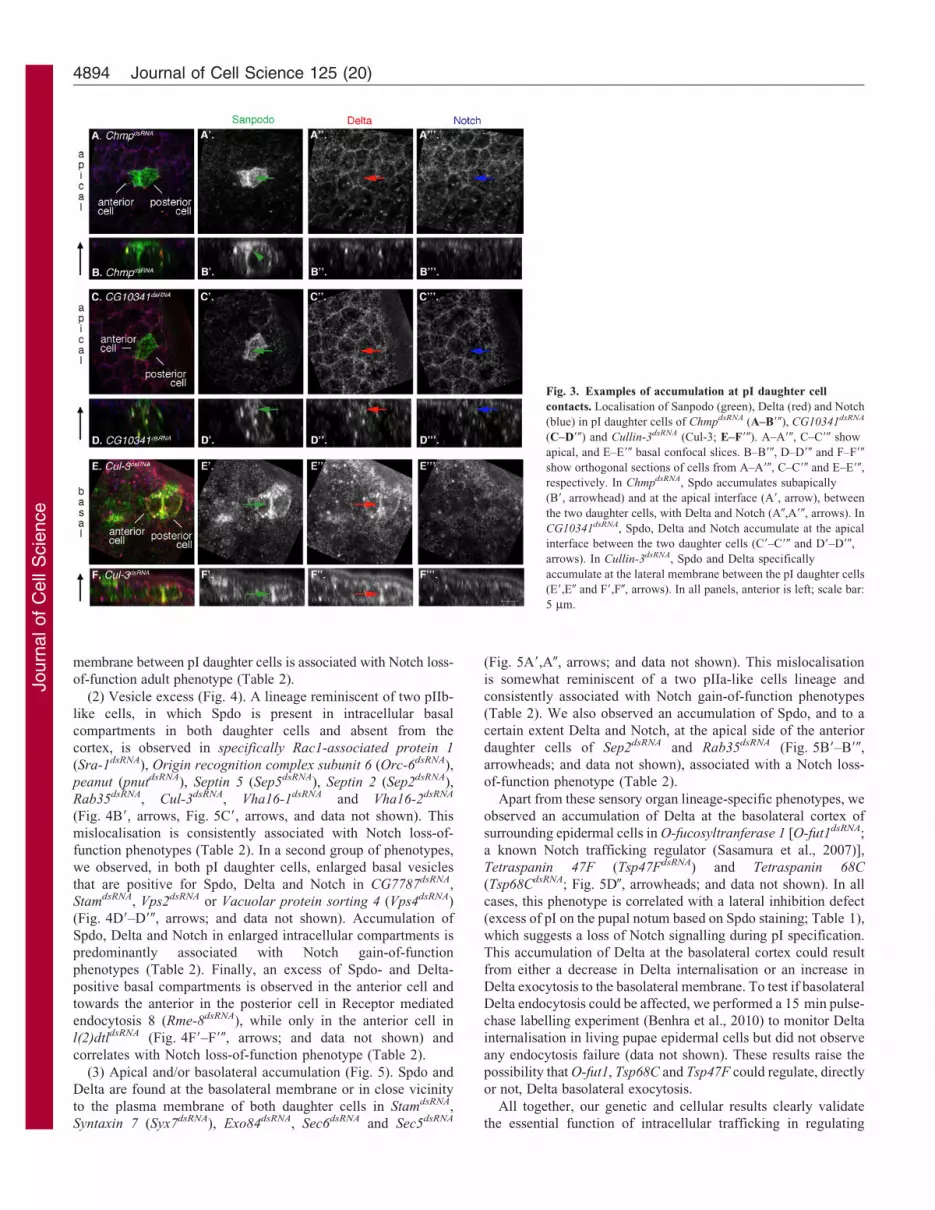
(1) Accumulation at pI daughter cell contact (Fig. 3). An excess
of Spdo is seen subapically between pI daughter cells of
CG2747dsRNA, Vacuolar protein sorting 28 (Vps28dsRNA) and
Chmp1dsRNA (Fig. 3B9, arrowhead; Fig. 6B9, arrowhead; and data
Fig. 2. The steady-state pattern of localisation of Sanpodo,
Delta and Notch. Localisation of Sanpodo (green), Delta (red)
and Notch (blue) in wild-type pI dividing cell (A–C90) and at
the pI daughter-cell stage (D–F90). A–A90, D–D90 show apical
and B–B90, E–E90 basal confocal slices. C–C90, F–F90 show
orthogonal sections of cells from A–B90 and D–E90,
respectively. The asymmetric localisation of Spdo in
endosomes in the anterior cell, and in endosomes and at the
basolateral cortex in the posterior cell reflects the differential
cell identity of the pI daughter cells (E9). In all panels, anterior
is left; scale bar: 5 mm.
Journal of Cell Science 125 (20)4892

Journ
alof
Cell
Scie
nce
not shown). We also observed an accumulation of Spdo, Notch and
Delta at the apical interface between the pI daughter cells in
CG2747dsRNA, Signal transducing adaptor molecule (StamdsRNA),
Vps28dsRNA, Chmp1dsRNA, Vps2dsRNA and CG10341dsRNA (Fig. 3A9–
A90,C9–C90,D9–D90, arrows; Fig. 6A9–A90, arrows; and data not
shown). Finally, we detected an accumulation of Spdo and Delta at
the lateral membrane between pI daughter cells of CG7787dsRNA
and Cullin-3 (Guftagu/Cul-3dsRNA) (Fig. 3E9,E0,F9,F0, arrows; and
data not shown). Strikingly, accumulation of Spdo subapically and/
or, with Notch and Delta, at the apical interface between pI
daughter cells correlates with a Notch gain-of-function adult
phenotype, while accumulation of Spdo and Delta at the lateral
Table 2. Complete list of genes that affect Notch, Sanpodo and/or Delta subcellular localisation at the pI daughter cell stage after
dsRNA induction
Screen category CandidateMolecularfunction
Adultsensoryorgan
phenotype2-cell stage localisation defects of Notch, Sanpodo and/or
Delta
Coat componentsand accessory proteins
CG2747 (putative orthologue ofSaccharomyces cerevisiae Laa1p)
AP-1 accessoryprotein
GOF Subapical accumulation of Spdo. Accumulationof Spdo, Notch and Delta at apical interface.
Lipid microdomainorganization
Tetraspanin 47F Tetraspanin LOF Cortex accumulation: Delta at basolateral cortex.
Tetraspanin 68C Tetraspanin LOF Cortex accumulation: Delta at basolateral cortex.Cytoskeleton,
regulators or interactorsspecifically Rac1-associated
protein 1 (Sra-1)/CYFIPWASp activation
complexLOF Lineage reminiscent to two pIIb-like cells.
Origin recognition complexsubunit 6 (Orc-6)
septin regulator LOF Lineage reminiscent to two pIIb-like cells.
peanut (pnut) septin LOF Lineage reminiscent to two pIIb-like cells.Septin 5 (Sep5) septin LOF Lineage reminiscent to two pIIb-like cells.Septin 2 (Sep2) septin LOF Lineage reminiscent to two pIIb-like cells. Cortex
accumulation: Spdo, Notch and Delta at apical cortex ofanterior cell.
Small GTPases,GEFs/GAPs
Rab35 Rab LOF Lineage reminiscent to two pIIb-like cells. Cortexaccumulation: Spdo, Notch and Delta at apical cortex ofanterior cell.
CG7787 RabGEF LOF Lateral accumulation of Spdo and Delta. Vesicle excess: largedonuts Spdo, Notch and Delta positive.
Ubiquitylation Cullin-3 (Cul-3)/Guftagu E3 ubiquitin ligase LOF Lateral accumulation of Spdo and Delta. Lineage reminiscentto two pIIb-like cells.
ESCRT componentsand interactors
Signal transducing adaptormolecule (Stam)
ESCRT O GOF ? Accumulation of Spdo, Notch and Delta at apical interface.Vesicle excess: Spdo, Notch and Delta positive. Lineagereminiscent to two pIIa-like cells.
Vacuolar proteinsorting 28 (Vps28)
ESCRT I GOF Subapical accumulation of Spdo. Accumulation of Spdo,Notch and Delta at apical interface. Weak vesicleexcess: Spdo, Notch and Delta positive.
Chmp1 ESCRT III GOF Subapical accumulation of Spdo. Accumulation of Spdo,Notch and Delta at apical interface. Weak vesicleexcess: Spdo, Notch and Delta positive.
Vps2 ESCRT III GOF Accumulation of Spdo, Notch and Delta at apicalinterface. Vesicle excess: large donuts Spdo, Notch andDelta positive.
Vacuolar protein sorting 4 (Vps4) ATPase GOF Vesicle excess: large donuts Spdo, Notch and Deltapositive.
Membrane recognitionand/or fusion
Syntaxin 7 (Syx7)/Avalanche t-SNARE GOF Lineage reminiscent to two pIIa-like cells.
Exo84 Exocyst complex LOF andGOF
Lineage reminiscent to two pIIa-like cells.
Sec6 Exocyst complex GOF Lineage reminiscent to two pIIa-like cells.Sec5 Exocyst complex GOF Lineage reminiscent to two pIIa-like cells.
Vacuolar ATPase Vacuolar H+ ATPasesubunit 16 -1 (Vha16-1)
vacuolar ATPase(Vo domain,subunit c)
LOF Lineage reminiscent to two pIIb-like cells.
Vacuolar H+ ATPasesubunit 16-2 (Vha16-2)
vacuolar ATPase(Vo domain,subunit c)
LOF Lineage reminiscent to two pIIb-like cells.
Various O-fucosyltransferase 1 Fucosyltransferase LOF Cortex accumulation: Delta at basolateral cortex.Novel unknown function CG10341 RNA biogenesis GOF Accumulation of Spdo, Notch and Delta at apical
interface.lethal-(2)-denticleless (l(2)dtl) unknown function LOF Vesicle excess in anterior cell (Spdo, Notch and Delta
positive).Recycling pathway,
unknown functionReceptor mediated
endocytosis 8 (Rme-8)Microtubule
associatedprotein
LOF Vesicle excess in anterior cell and toward the anterior inposterior cell (Spdo, Notch and Delta positive).
Genes are listed in order of the screen category they belong to.
Notch intracellular trafficking regulators 4893
Journ
alof
Cell
Scie
nce
membrane between pI daughter cells is associated with Notch loss-
of-function adult phenotype (Table 2).
(2) Vesicle excess (Fig. 4). A lineage reminiscent of two pIIb-
like cells, in which Spdo is present in intracellular basal
compartments in both daughter cells and absent from the
cortex, is observed in specifically Rac1-associated protein 1
(Sra-1dsRNA), Origin recognition complex subunit 6 (Orc-6dsRNA),
peanut (pnutdsRNA), Septin 5 (Sep5dsRNA), Septin 2 (Sep2dsRNA),
Rab35dsRNA, Cul-3dsRNA, Vha16-1dsRNA and Vha16-2dsRNA
(Fig. 4B9, arrows, Fig. 5C9, arrows, and data not shown). This
mislocalisation is consistently associated with Notch loss-of-
function phenotypes (Table 2). In a second group of phenotypes,
we observed, in both pI daughter cells, enlarged basal vesicles
that are positive for Spdo, Delta and Notch in CG7787dsRNA,
StamdsRNA, Vps2dsRNA or Vacuolar protein sorting 4 (Vps4dsRNA)
(Fig. 4D9–D90, arrows; and data not shown). Accumulation of
Spdo, Delta and Notch in enlarged intracellular compartments is
predominantly associated with Notch gain-of-function
phenotypes (Table 2). Finally, an excess of Spdo- and Delta-
positive basal compartments is observed in the anterior cell and
towards the anterior in the posterior cell in Receptor mediated
endocytosis 8 (Rme-8dsRNA), while only in the anterior cell in
l(2)dtldsRNA (Fig. 4F9–F90, arrows; and data not shown) and
correlates with Notch loss-of-function phenotype (Table 2).
(3) Apical and/or basolateral accumulation (Fig. 5). Spdo and
Delta are found at the basolateral membrane or in close vicinity
to the plasma membrane of both daughter cells in StamdsRNA,
Syntaxin 7 (Syx7dsRNA), Exo84dsRNA, Sec6dsRNA and Sec5dsRNA
(Fig. 5A9,A0, arrows; and data not shown). This mislocalisation
is somewhat reminiscent of a two pIIa-like cells lineage and
consistently associated with Notch gain-of-function phenotypes
(Table 2). We also observed an accumulation of Spdo, and to a
certain extent Delta and Notch, at the apical side of the anterior
daughter cells of Sep2dsRNA and Rab35dsRNA (Fig. 5B9–B90,
arrowheads; and data not shown), associated with a Notch loss-
of-function phenotype (Table 2).
Apart from these sensory organ lineage-specific phenotypes, we
observed an accumulation of Delta at the basolateral cortex of
surrounding epidermal cells in O-fucosyltranferase 1 [O-fut1dsRNA;
a known Notch trafficking regulator (Sasamura et al., 2007)],
Tetraspanin 47F (Tsp47FdsRNA) and Tetraspanin 68C
(Tsp68CdsRNA; Fig. 5D0, arrowheads; and data not shown). In all
cases, this phenotype is correlated with a lateral inhibition defect
(excess of pI on the pupal notum based on Spdo staining; Table 1),
which suggests a loss of Notch signalling during pI specification.
This accumulation of Delta at the basolateral cortex could result
from either a decrease in Delta internalisation or an increase in
Delta exocytosis to the basolateral membrane. To test if basolateral
Delta endocytosis could be affected, we performed a 15 min pulse-
chase labelling experiment (Benhra et al., 2010) to monitor Delta
internalisation in living pupae epidermal cells but did not observe
any endocytosis failure (data not shown). These results raise the
possibility that O-fut1, Tsp68C and Tsp47F could regulate, directly
or not, Delta basolateral exocytosis.
All together, our genetic and cellular results clearly validate
the essential function of intracellular trafficking in regulating
Fig. 3. Examples of accumulation at pI daughter cell
contacts. Localisation of Sanpodo (green), Delta (red) and Notch
(blue) in pI daughter cells of ChmpdsRNA (A–B90), CG10341dsRNA
(C–D90) and Cullin-3dsRNA (Cul-3; E–F90). A–A90, C–C90 show
apical, and E–E90 basal confocal slices. B–B90, D–D90 and F–F90
show orthogonal sections of cells from A–A90, C–C90 and E–E90,
respectively. In ChmpdsRNA, Spdo accumulates subapically
(B9, arrowhead) and at the apical interface (A9, arrow), between
the two daughter cells, with Delta and Notch (A0,A90, arrows). In
CG10341dsRNA, Spdo, Delta and Notch accumulate at the apical
interface between the two daughter cells (C9–C90 and D9–D90,
arrows). In Cullin-3dsRNA, Spdo and Delta specifically
accumulate at the lateral membrane between the pI daughter cells
(E9,E0 and F9,F0, arrows). In all panels, anterior is left; scale bar:
5 mm.
Journal of Cell Science 125 (20)4894

Journ
alof
Cell
Scie
nce
Notch-signalling-dependent binary cell fate acquisition. Indeed,
we identified 26 genes for which a Notch signalling adult
phenotype is associated with a change in intracellular localisation
of major Notch signalling components after the first asymmetric
cell division.
CG2747 regulates clathrin adaptor AP-1 intracellular
localisation
Among the genes isolated in the genetic and cellular screen,
CG2747dsRNA phenocopies the loss of AP-1 function (Benhra
et al., 2011). Indeed, we observed a subapical accumulation of
Spdo (Fig. 6B9, arrowhead) and an accumulation of Spdo, Delta
and Notch at the apical interface between pI daughter cells
(Fig. 6A–B90, arrows). To further validate the specificity of the
phenotype induced by dsRNA, we used two independent dsRNA
lines obtained from the NIG-Fly and TRIP Centers (lines 2747R-
3 and BL29322, respectively), which target different regions of
the transcript (regions 1081–1612 and 407–906 of the transcript
CG2747-RD, respectively), and observed the same phenotype
(data not shown). We also observed an excess of socket cells and/
or double shafts with socket cells with both dsRNA lines (Fig. 6C,
arrows; and data not shown), which is reminiscent of Notch
signalling gain-of-function phenotype observed in AP-1 loss of
function (Benhra et al., 2011).
Although CG2747 function has not yet been studied in D.
melanogaster, it belongs to the highly conserved HEAT repeat-
containing protein 5 (HEATR5) family (Fernandez and Payne,
2006), previously shown to physically interact with the ear-
domain of murine c-adaptin subunit of the AP-1 complex (Lui
et al., 2003). For instance CG2747 shares 49% of sequence
identities with the Homo sapiens HEATR5B and when we used
the p200 antibody against the mammal HEATR5B (Hirst et al.,
2005), we observed a staining in intracellular vesicles and at the
junction of wild-type notum epithelial cells (Fig. 6F,F0). In the
area of a notum where CG2747dsRNA is induced, p200 staining is
greatly reduced (Fig. 6E,E0 compared to Fig. 6F,F0), suggesting
Fig. 4. Examples of excess vesicles in pI daughter cells.
Localisation of Sanpodo (green), Delta (red) and Notch (blue) in pI
daughter cells of Vha16-2dsRNA (A–B90), Vps4dsRNA (C–D90) and
l(2)dtldsRNA (E–F90). A–A90, C–C90, E–E90 show apical and B–B90,
D–D90 and F–F90 basal confocal slices. In Vha16-2dsRNA, Spdo is
found in basal vesicles in both daughter cells (B9, arrows), a lineage
reminiscent of two pIIb-like cells. In Vps4dsRNA, Spdo, Delta and
Notch colocalise in enlarged basal vesicles in both daughter cells
(D9–D90, arrows). In l(2)dtldsRNA, an excess of Spdo-, Delta- and
Notch-positive basal compartments is observed in the anterior cell
(F9–F90, arrows). In all panels, anterior is left; scale bar: 5 mm.
Notch intracellular trafficking regulators 4895

Journ
alof
Cell
Scie
nce
that the mammal antibody recognises the Drosophila protein.
Furthermore, it was demonstrated in Saccharomyces cerevisiae,
that Laap1, sole member of the HEATR5 family, is necessary for
proper AP-1 intracellular localisation (Fernandez and Payne,
2006). To investigate whether CG2747 shares the same function
in D. melanogaster, we analysed the localisation of AP-1 on the
pupal notum. AP-1c staining is greatly reduced in the region of
the notum where UAS–CG2747dsRNA expression is induced in
comparison with the anterior head in which the ap–GAL4 driver
is not active (Fig. 6G). In the converse experiment, we observed
that p200 staining is not affected in AP-47 (gene encoding the mu
subunit of AP-1 complex) homozygote mutant cells (Fig. 6H–
H0). Thus CG2747 localisation is independent of AP-1 activity.
We concluded that CG2747dsRNA reduces the level of CG2747 at
a sufficient degree to prevent AP-1c membrane localisation and
phenocopies the AP-1 cellular and adult phenotypes. All these
results strongly support the model in which CG2747 is necessary
for AP-1 proper intracellular localisation and function in
Drosophila.
DiscussionOur dsRNA genetic screen, performed in D. melanogaster notum
using the GAL4–UAS system, allowed us to specifically identify
113 Notch signalling regulators among 418 candidates chosen for
previously described or suggested function in intracellular
trafficking (Table 1). Importantly, up to 76% of the regulators
we identified were not found in a similar genetic screen
performed at a genome-wide level (Mummery-Widmer et al.,
2009). Our study clearly shows that using multiple GAL4 drivers
and three different temperatures increases the efficiency of such
dsRNA genetic screen as it allows identifying optimal dsRNA
technical conditions for each construct. In particular, using
different GAL4 drivers limit the false positive or negative results
observed when the driver itself induces morphological defects
and/or when the expression of the dsRNA induces lethality or
notum morphological defects.
Recovering almost 30% of positive hits in a screen is unusual.
We interpret this high number first as a reflection of the tight
interconnection between membrane traffic and both Delta and
Notch signalling to ultimately ensure the proper spatiotemporal
control of the pathway. Second, this high number is also
explained by the fact that many regulators are acting as protein
complexes (APs, ESCRTs, Exocysts, ATPases, septins etc.). This
property could be used to confirm the specificity of the dsRNA
effect, as inactivation of gene products belonging to the same
protein complex is expected to give similar phenotype. If this
prediction is fully fulfilled for AP-1, AP-2, septins and the
ubiquitin ligase complex SCF, it is only partially fulfilled for the
ESCRT and Exocyst complexes (see below).
Although these results are novel and further validate the
concept of a regulation of Notch signalling by intracellular
trafficking (for review, see Fortini and Bilder, 2009; Furthauer,
Gonzalez-Gaitan, 2009; Kopan and Ilagan, 2009; Le Bras et al.,
2011; Musse et al., 2012; Yamamoto et al., 2010), we were
aiming at identifying novel regulators of the subcellular
localisation of three major Notch signalling components:
Notch, Spdo and Delta. Indeed, the observed adult phenotypes
could result from various and multiple defects in Notch signalling
during pI asymmetric cell division and/or at the pI daughter cell
stage. Furthermore, our identified genes could regulate various
molecular events such as cell fate determinants segregation,
Delta, Spdo and/or Notch proper subcellular localisation via
Fig. 5. Examples of apical and/or basal accumulation.
Localisation of Sanpodo (green), Delta (red) and Notch (blue) in
pI daughter cells of Exo84dsRNA (A–A90), Rab35dsRNA (B–C90) and
Tsp68CdsRNA (D–D90). B–B90 show apical and A–A90, C–C90, D–
D90 basal confocal slices. In Exo84dsRNA, Spdo and Delta are
found at the basolateral membrane or in close proximity to the
plasma membrane of both daughter cells (A9–A0, arrows), a
lineage somewhat reminiscent of two pIIa-like cells. In
Rab35dsRNA, Spdo, Delta and Notch accumulate at the apical side
of the anterior daughter cell (B9–B90, arrowheads). Spdo is also
found in basal vesicles in both daughter cells (C9, arrows), a
lineage reminiscent of two pIIb-like cells. In Tsp68CdsRNA, Delta
can be found at the basolateral cortex of epidermal cells (D0,
arrowheads). In all panels, anterior is left; scale bar: 5 mm.
Journal of Cell Science 125 (20)4896

Journ
alof
Cell
Scie
nce
endocytosis and/or recycling. Therefore, we decided to take
advantage of the fact that Spdo, Delta and Notch localisation are
dynamic during pI division and at the pI daughter cell stage in the
pupal notum. We were able to observe localisation changes for 26
out of the 61 genes that we studied to further understand the adult
phenotypes induced by dsRNA (Table 2). Three major categories
of localisation changes were identified at the pI daughter cell
stage: accumulation at pI daughter cell contact, vesicle excess
(and in some cases, enlarged vesicles), and apical and/or
basolateral accumulation. And, in each of these categories, we
observed pupal lineages which correlate either with an adult
Notch gain- or loss-of-function-like phenotype (as illustrated
with Spdo localisation in supplementary material Fig. S1).
For 10 genes, changes in Notch component localisation can be
a direct consequence of their inactivation or reflect a change in pI
daughter cell fate acquisition. Indeed, we observed a pupal
lineage somewhat reminiscent of two pIIa-like (Syx7, Exo84,
Sec6, Sec5) or two pIIb-like (Sra-1, Orc6, pnut, Sep5, Vha16-1,
Vha16-2) daughter cells (supplementary material Fig. S1B,F
respectively). And these lineages correlate with the observed
adult phenotypes i.e. Notch gain of function or loss of function,
respectively. Although these results do not elucidate the function
of these genes on Notch signalling regulation, they indicate that
Notch signalling can be similarly regulated in various cellular
contexts. For example, it was previously shown that the Vacuolar
ATPase functions to control the acidification of endosomes
required for Notch activation after binding by its ligand in the eye
imaginal disc and ovaries (Vaccari et al., 2010; Yan et al., 2009).
Surprisingly, our results suggest that part of the same complex
might regulate different aspects of Notch signalling in the
sensory organ lineage. Indeed, we observed a gain-of-Notch-
signalling-like adult phenotype for three Exocyst components
(Exo84, Sec6 and Sec5) but also a loss-of-Notch-signalling-like
adult one for Exo84. These results can either reflect a bias in the
RNAi silencing which might not be as effective for each Exocyst
subunits and/or suggest that the Exocyst might have several
functions during pI asymmetric cell division with opposite role in
the regulation of Notch signalling. Further studies will be
necessary to validate these results and to define if Exo84 could
function with Sec15, another Exocyst component, which
positively regulates Spdo and Delta post-endocytic trafficking
in pI daughter cells (Jafar-Nejad et al., 2005).
For the first time, we identified members of the septin family
(pnut and Sep5) and one of their regulator [Orc-6 (Chesnokov
Fig. 6. CG2747 regulates clathrin adaptor AP-1 intracellular
localisation. (A–B90). Localisation of Sanpodo (green), Delta
(red) and Notch (blue) in pI daughter cells of CG2747dsRNA.
A–A90 show apical confocal slice and B–B90 show orthogonal
section of cells from A–A90. Spdo accumulates subapically
(B9, arrowhead) and at the interface between the two daughter
cells with Delta and Notch (A9–A90, arrows). (C) Excess of
double shafts with sockets cells (arrows) observed on a
CG2747dsRNA notum. This adult phenotype is reminiscent of gain
of Notch signalling. (D–F0). Partial loss of HEATR5B staining
(p200, green) in the median part of the notum where UAS–
CG2747dsRNA is induced by ap–GAL4 (D,D9 left side, E–E0)
compared with the posterior part of the notum where ap–GAL4
does not drive UAS expression (D,D9 right side, asterisk, F–F0).
E–F9 are higher magnifications of D–D9, taken at the level of
adherens junctions (DE-CAD, red in E,E9 and F,F9). (G) Loss of
AP-1c staining in the median part of the notum where UAS–
CG2747dsRNA is induced by ap–GAL4 (right side) compared with
the anterior head in which ap–GAL4 does not drive UAS
expression (left side, star). (H–H0) HEATR5B staining is not
affected in AP-47SHE11 mutant cells (red, H and H0, inside dotted
lines). Mutant cells are identified by the absence of nls–GFP (H,
blue, inside dotted line). H–H0 are confocal slices taken at the
level of adherens junctions (DE-CAD, green in H and H9). In
panels A–B90, D–H0, anterior is left. Scale bar: 5 mm (A–B90,G),
200 mm (D,D9) and 15 mm (E–F0,H–H0).
Notch intracellular trafficking regulators 4897

Journ
alof
Cell
Scie
nce
et al., 2003)] as putative regulators of Notch signalling. Septincomplexes can act as scaffolds and/or diffusion barriers in
various cellular events such as cytoskeleton organisation,cytokinesis, membrane organisation and vesicle targeting whichcould potentially regulate the pI asymmetric cell division (for areview, see Cao et al., 2009). It is necessary to further decipher
the pupal phenotype to define if these septins directly regulateNotch signalling traffic and/or pI cytokinesis (N.B. Founounouand R.L.B. unpublished). While the pupal lineage is somewhat
reminiscent of two pIIb-like daughter cells, we also observed anaccumulation of Spdo, Notch and Delta at the apical side of theanterior pI daughter cell of Sep2dsRNA as well as of Rab35dsRNA
(supplementary material Fig. S1G). Their common phenotype isnot surprising knowing that human Rab35 was proposed to playan essential control on the terminal step of cytokinesis in part bycontrolling SEPT2 subcellular distribution during cell division
(Kouranti et al., 2006). Although it is not yet possible to decipherif this apical accumulation is a cause or a consequence of theNotch pupal and adult loss-of-function phenotype, this data
identify two putative regulators of apical localisation and confirmthat Notch signalling major components are differentiallytrafficked between pI daughter cells.
Inactivation of 14 genes led to localisation changes indicatingmultiple subcellular sites (plasma membrane, vesicularcompartments, interface between the two daughter cells), which
appear to be essential to the fine regulation of Notch signalling inthe Drosophila sensory organ lineage. Indeed, subapicalaccumulation of Spdo, localisation of Notch, Spdo and Delta atthe apical interface between pI daughter cells, and/or an excess of
enlarged endosomes in both daughter cells are associated withadult Notch gain-of-function-like phenotypes (supplementarymaterial Fig. S1C,D,E, respectively). However, accumulation of
Spdo and Delta at the lateral membrane between pI daughter cellscorrelates with Notch signalling loss-of-function phenotype(supplementary material Fig. S1H,I).
In a control situation, Notch accumulates transiently at theapical interface between pIIb and pIIa (Benhra et al., 2011;Couturier et al., 2012). Accumulation of Spdo together withNotch at this apical pIIb/pIIa interface has previously been
observed in AP-1 loss-of-function mutants and correlated with again of Notch signalling (Benhra et al., 2011). Because,impairment of Delta trafficking towards the pIIb/pIIa interface
in Arp2/3 mutants leads to a loss of Notch signalling (Rajan et al.,2009), it was proposed that Delta–Notch interaction resulting inNotch activation is taking place at the apical pIIb/pIIa interface
that could function as a signalling platform (Benhra et al., 2011).Nonetheless, this proposal awaits experimental demonstration.This proposal was recently challenged by F. Schweisguth and
colleagues, who generated a functional Notch construct taggedwith GFP and expressed at physiological level (NiGFP)(Couturier et al., 2012). Notch activation is reported to occur15–45 min after cytokinesis and productive signalling is
proposed to take place at the pIIa/pIIb interface of thecytokinetic furrow, where NiGFP accumulates in numb mutantbackground or when Dynamin-dependent endocytosis is blocked.
Adult and subcellular AP-1 loss-of-function-type phenotypesare observed when two genes with previously unknown functionin Drosophila, CG10341 and CG2747, were inactivated. Both of
them had been identified as putative membrane traffickingregulators in a C. elegans screen (Balklava et al., 2007).CG10341 belongs to the Nuclear Assembly Factor 1 (NAF1)
family involved in ribosome biogenesis, which suggests anindirect role, if so, in intracellular trafficking. On the contrary,
CG2747 belongs to the HEATR5 family and we were able toshow that CG2747 is required for the clathrin adaptor AP-1complex subcellular localisation, similarly to its putativeorthologue of the S. cerevisiae Laa1p (Fernandez and Payne,
2006). We observed that the human HEATR5B/p200 antibodystaining is affected in CG2747dsRNA, which supports anevolutionary conservation of the HEATR5 function among
metazoans. Nonetheless, the function of human HEATR5B/p200 remains unknown as no phenotype could be observed inp200-depleted cells maybe due to a poor silencing efficiency and/
or a redundancy with the other human HEATR5 member,HEATR5A (Lui et al., 2003). Therefore, we identified a newregulator of Notch signalling that acts as a major regulator of theclathrin adaptor complex, AP-1.
We also observed an accumulation of Notch, Spdo and Delta atthe interface and/or in endosomes in both daughter cells,correlated with adult Notch gain-of-function phenotypes, when
we inactivated members of the ESCRT complex (Stam, Vps28,Chmp1, Vps2 and Vps4). Accumulation at the interface and/orendosomes can result from a blockade in endosome maturation
when ESCRT function is impaired and it has already beendescribed that accumulation of Notch in endocytic compartmentscan result in an ectopic ligand-independent activation of Notch(Herz et al., 2009; Moberg et al., 2005; Thompson et al., 2005;
Vaccari and Bilder, 2005; Vaccari et al., 2009). Additionally,ESCRT complexes are involved in various cellular mechanisms:cargo engagement and/or deubiquitylation, maturation of multi
vesicular bodies, vesicle budding and/or cytokinesis (for areview, see Henne et al., 2011). In our screen, impairment ofdifferent ESCRT pathway components led to opposite adult
phenotypes i.e. loss- or gain-of-Notch-signalling-like onesdepending on the complex and/or its subunit(s) depleted bydsRNA (see Table 1). However, we did not observe any Spdo,
Delta or Notch localisation changes associated with these adultloss-of-function phenotypes. Further studies are, therefore,necessary to elucidate our genetics results by identifying whichsubcellular mechanisms and which cargo(es) are regulated by the
different identified ESCRT subunits to control Notch signalling.
Accumulation of Spdo and Delta at the lateral membranebetween pI daughter cells is correlated with adult Notch loss-of-
function phenotypes in Cul-3dsRNA and CG7787dsRNA
(supplementary material Fig. S1H,I respectively). This lateralmembrane was previously named apical actin-rich structure
‘stalk’ and identified as the lateral part of branched actin networkpresent at the interface and through which endocytosed Deltatraffic back (Rajan et al., 2009). Therefore, CG7787 and Cul-3might regulate Spdo and/or Delta basolateral endocytosis and/or
recycling required for Notch signalling as this accumulationphenotype is correlated with a loss of Notch signalling. WhileCul-3 is a subunit of E3 ubiquitin ligase, CG7787 putatively
encodes a guanyl-nucleotide exchange factor. CG7787 belongs tothe MSS4/DSS4 family proposed to function as chaperone formisfolded Rab proteins (Nuoffer et al., 1997) and in particular,
Rabs associated with the Exocyst pathway (Itzen et al., 2006).Therefore, CG7787 might be involved in the same recyclingpathway as Sec15 (see above) and positively regulates Notch
signalling. Other data support the general idea that a recyclingpathway positively regulates Notch signalling in the sensoryorgan lineage. Indeed, we observed an excess of Spdo-, Notch-
Journal of Cell Science 125 (20)4898

Journ
alof
Cell
Scie
nce
and Delta-positive vesicles either in the anterior or towards the
anterior cell in Rme-8dsRNA and l(2)dtldsRNA, which is associated
with an adult Notch loss-of-function phenotype. Although l(2)dtl
function in intracellular trafficking is still unknown, Rme-8 was
shown to regulate a recycling pathway (Shi et al., 2009). All
those results confirm that Spdo, Notch and Delta transiently
traffic through the lateral membrane and/or endosomes to ensure
a proper Notch signalling.
Finally, we observed a basolateral accumulation of Delta in
epithelial cells of O-fut1dsRNA, Tsp47FdsRNA and Tsp68CdsRNA
nota and that Delta endocytosis is not affected which suggest the
existence of basolateral Delta exocytosis. In support of this
hypothesis, it was already demonstrated that Delta can be
fucosylated by mammalian O-fut1 (Panin et al., 2002) but this
data remained to be demonstrated in vivo and in Drosophila.
When confirmed by further studies in classical genetic mutants,
these results will eventually highlight a function for Delta
basolateral exocytosis and also a new role of the tetraspanins
family on Notch signalling.
In conclusion, our screen led us to identify intracellular
trafficking regulators of major Notch signalling actors. Although
it is still debatable whether the subcellular localisation changes
observed are a cause or a consequence of the Notch signalling
phenotype, the screen we performed led to the identification of
11 previously unknown regulators of Notch signalling (CG2747,
Tsp47F, Orc-6, pnut, Sep2, Sep5, Rab35, CG7787, CG10341,
l(2)dtl and Rme-8). Without any doubt, further analyses of our
identified genes will bring a better understanding of their
trafficking function in regulating Notch-signalling-dependent
binary cell fate acquisition, as well as of their putative
molecular interactions.
Materials and MethodsDrosophila stocks and genetics
Unless otherwise stated, fly stocks were obtained from the Bloomington DrosophilaStock Center. Driver–GAL4 stocks used in this study were: ap–GAL4 (Calleja et al.,1996), sca–GAL4 (Mlodzik et al., 1990), Eq–GAL4 (Pi et al., 2001) and pnr–GAL4(Calleja et al., 1996). All dsRNA transgenic lines were supplied by the ViennaDrosophila RNAi Center [VDRC, (Dietzl et al., 2007)]; except lines (as indicated insupplementary material Tables S1–S3), which were obtained from the National Instituteof Genetics Fly Stock Center (NIG-FLY) or the Transgenic RNAi Project (TRIP) via theBloomington Drosophila Stock Center. For RNAi-induced phenotype study, crossesbetween UAS-hairpin RNAi males and driver-GAL4 females were raised at 18 C, 25 Cor shifted at 29 C during L2–L3 larval stages. For each cross in which the genotypeswere blinded for objectivity purpose, two experimenters examined at least 20 fliessensory organ distribution and/or morphological phenotypes. w1118 males were crossedwith driver-GAL4 females for control experiments. To obtain AP-47SHE11 mitoticclones, we used the FLP-FRT technique and the stocks (1) y w hs-FLP; FRT82B, Ubi-
GFP(S65T)nls and (2) FRT82B, AP-47SHE11/TM6 Tb Sb, as previously described(Benhra et al., 2011). Heat shocks were performed at L2 and L3 during 30 min.
Immunocytochemistry
Pupae were aged for 17 h to 20 h after puparium formation, dissected in 16PBS,fixed in 4% paraformaldehyde and stained as previously described (Le Borgne andSchweisguth, 2003). Primary antibodies used were mouse anti-Notch ExtraCellular Domain (NECD; DSHB, 1:100), rabbit anti-Spdo [a kind gift from J.Skeath; 1:1000 (O’Connor-Giles and Skeath, 2003)], guinea pig anti-Delta ExtraCellular Domain (GP582, a kind gift from M. Muskavitch; 1:2000), rabbit anti-HEATR5B [p200; a kind gift from M. Robinson; 1:20 (Hirst et al., 2005)], rat anti-DE-CAD (DCAD2; DSHB 1:250) and mouse anti-AP-1c [1:100 (Benhra et al.,2011)]. Cy2-, Cy3- and Cy5-coupled secondary antibodies (1:500) were fromJackson Laboratories. Delta 15 minutes internalisation assays were performed withmouse anti-Delta DSHB (1:100), as previously described (Benhra et al., 2011).
Images were acquired with a Leica SPE confocal microscope, which was noise-suppressed using the smooth function of ImageJ. In all figures, Notch (Cy5-)images were colour balanced using ImageJ. Defect in lateral inhibition wasacknowledged when more than four to five sensory organs were systematicallydetected with a 636 1.4 NA lens, zoom 3 on a notum.
AcknowledgementsWe thank M. Muskavitch, M. Robinson, J. Skeath, the BloomingtonStock Center, the Vienna Drosophila RNAi Center, the TRiP atHarvard Medical School (NIH/NIGMS R01-GM084947) and theNational Institute of Genetics Fly Stock Center for providingantibodies or fly stocks, as well as the Microscopy RennesImaging Center. The monoclonal antibody generated by S.Artavanis Tsakonas (NECD) was obtained from the DevelopmentalStudies Hybridoma Bank, generated under the auspices of theNational Institute of Child Health and Human Development, andmaintained by the University of Iowa Department of BiologicalSciences. We thank members of the Le Borgne laboratory for helpfuldiscussions. We thank A. Pacquelet and G. Michaux for criticalreading of the manuscript. Special thanks to Amy Winehouse for hermusic that accompanied us, while we screened around 100,000 flies.
FundingThis work was supported by the Action Thematique IncitativePrioritaire programme CNRS to R.L.B.; Region Bretagne(Programme Accueil de COMpetences en Bretagne ‘Notasid’[grant number 2168 to R.L.B.]; Fondation ARC pour la Recherchesur le Cancer [grant number 4905 to R.L.B.]; Fondation pour laRecherche Medicale to R.L.B., Rennes Metropole [grant ‘Aided’Installation Scientifique’ to S.L.B.]; and La Ligue contre le Cancer35 to R.L.B.
Supplementary material available online at
http://jcs.biologists.org/lookup/suppl/doi:10.1242/jcs.110171/-/DC1
ReferencesAbdelilah-Seyfried, S., Chan, Y. M., Zeng, C., Justice, N. J., Younger-Shepherd,
S., Sharp, L. E., Barbel, S., Meadows, S. A., Jan, L. Y. and Jan, Y. N. (2000).
A gain-of-function screen for genes that affect the development of the Drosophila
adult external sensory organ. Genetics 155, 733-752.
Acar, M., Jafar-Nejad, H., Takeuchi, H., Rajan, A., Ibrani, D., Rana, N. A., Pan,
H., Haltiwanger, R. S. and Bellen, H. J. (2008). Rumi is a CAP10 domain
glycosyltransferase that modifies Notch and is required for Notch signaling. Cell 132,
247-258.
Balklava, Z., Pant, S., Fares, H. and Grant, B. D. (2007). Genome-wide analysis
identifies a general requirement for polarity proteins in endocytic traffic. Nat. Cell
Biol. 9, 1066-1073.
Baust, T., Czupalla, C., Krause, E., Bourel-Bonnet, L. and Hoflack, B. (2006).
Proteomic analysis of adaptor protein 1A coats selectively assembled on liposomes.
Proc. Natl. Acad. Sci. USA 103, 3159-3164.
Baust, T., Anitei, M., Czupalla, C., Parshyna, I., Bourel, L., Thiele, C., Krause,
E. and Hoflack, B. (2008). Protein networks supporting AP-3 function in targeting
lysosomal membrane proteins. Mol. Biol. Cell 19, 1942-1951.
Ben-Yaacov, S., Le Borgne, R., Abramson, I., Schweisguth, F. and Schejter, E. D.
(2001). Wasp, the Drosophila Wiskott-Aldrich syndrome gene homologue, is required
for cell fate decisions mediated by Notch signaling. J. Cell Biol. 152, 1-13.
Benhra, N., Vignaux, F., Dussert, A., Schweisguth, F. and Le Borgne, R. (2010).
Neuralized promotes basal to apical transcytosis of delta in epithelial cells. Mol. Biol.
Cell 21, 2078-2086.
Benhra, N., Lallet, S., Cotton, M., Le Bras, S., Dussert, A. and Le Borgne, R. (2011).
AP-1 controls the trafficking of Notch and Sanpodo toward E-cadherin junctions in
sensory organ precursors. Curr. Biol. 21, 87-95.
Berdnik, D., Torok, T., Gonzalez-Gaitan, M. and Knoblich, J. A. (2002). The
endocytic protein alpha-Adaptin is required for numb-mediated asymmetric cell
division in Drosophila. Dev. Cell 3, 221-231.
Bogdan, S., Grewe, O., Strunk, M., Mertens, A. and Klambt, C. (2004). Sra-1
interacts with Kette and Wasp and is required for neuronal and bristle development in
Drosophila. Development 131, 3981-3989.
Brand, A. H. and Perrimon, N. (1993). Targeted gene expression as a means of altering
cell fates and generating dominant phenotypes. Development 118, 401-415.
Bray, S. J. (2006). Notch signalling: a simple pathway becomes complex. Nat. Rev. Mol.
Cell Biol. 7, 678-689.
Calleja, M., Moreno, E., Pelaz, S. and Morata, G. (1996). Visualization of gene
expression in living adult Drosophila. Science 274, 252-255.
Campbell, G. (2005). Regulation of gene expression in the distal region of the
Drosophila leg by the Hox11 homolog, C15. Dev. Biol. 278, 607-618.
Cao, L., Yu, W., Wu, Y. and Yu, L. (2009). The evolution, complex structures and
function of septin proteins. Cell. Mol. Life Sci. 66, 3309-3323.
Chen, C. M., Freedman, J. A., Bettler, D. R., Jr, Manning, S. D., Giep, S. N.,
Steiner, J. and Ellis, H. M. (1996). Polychaetoid is required to restrict segregation of
Notch intracellular trafficking regulators 4899

Journ
alof
Cell
Scie
nce
sensory organ precursors from proneural clusters in Drosophila. Mech. Dev. 57, 215-227.
Chesnokov, I. N., Chesnokova, O. N. and Botchan, M. (2003). A cytokinetic functionof Drosophila ORC6 protein resides in a domain distinct from its replication activity.Proc. Natl. Acad. Sci. USA 100, 9150-9155.
Chung, H. M. and Struhl, G. (2001). Nicastrin is required for Presenilin-mediatedtransmembrane cleavage in Drosophila. Nat. Cell Biol. 3, 1129-1132.
Coumailleau, F., Furthauer, M., Knoblich, J. A. and Gonzalez-Gaitan, M. (2009).Directional Delta and Notch trafficking in Sara endosomes during asymmetric celldivision. Nature 458, 1051-1055.
Couturier, L., Vodovar, N. and Schweisguth, F. (2012). Endocytosis by Numb breaksNotch symmetry at cytokinesis. Nat. Cell Biol. 14, 131-139.
David, N. B., Martin, C. A., Segalen, M., Rosenfeld, F., Schweisguth, F. and
Bellaıche, Y. (2005). Drosophila Ric-8 regulates Galphai cortical localization topromote Galphai-dependent planar orientation of the mitotic spindle duringasymmetric cell division. Nat. Cell Biol. 7, 1083-1090.
Dietzl, G., Chen, D., Schnorrer, F., Su, K. C., Barinova, Y., Fellner, M., Gasser,
B., Kinsey, K., Oppel, S., Scheiblauer, S. et al. (2007). A genome-wide transgenicRNAi library for conditional gene inactivation in Drosophila. Nature 448, 151-156.
Djiane, A., Shimizu, H., Wilkin, M., Mazleyrat, S., Jennings, M. D., Avis, J., Bray,
S. and Baron, M. (2011). Su(dx) E3 ubiquitin ligase-dependent and -independentfunctions of polychaetoid, the Drosophila ZO-1 homologue. J. Cell Biol. 192, 189-200.
Dobens, L., Jaeger, A., Peterson, J. S. and Raftery, L. A. (2005). Bunched sets aboundary for Notch signaling to pattern anterior eggshell structures during Drosophilaoogenesis. Dev. Biol. 287, 425-437.
Eid, J. P., Arias, A. M., Robertson, H., Hime, G. R. and Dziadek, M. (2008). TheDrosophila STIM1 orthologue, dSTIM, has roles in cell fate specification and tissuepatterning. BMC Dev. Biol. 8, 104.
Emery, G., Hutterer, A., Berdnik, D., Mayer, B., Wirtz-Peitz, F., Gaitan, M. G. and
Knoblich, J. A. (2005). Asymmetric Rab 11 endosomes regulate delta recycling andspecify cell fate in the Drosophila nervous system. Cell 122, 763-773.
Fernandez, G. E. and Payne, G. S. (2006). Laa1p, a conserved AP-1 accessory proteinimportant for AP-1 localization in yeast. Mol. Biol. Cell 17, 3304-3317.
Fetchko, M., Huang, W., Li, Y. and Lai, Z. C. (2002). Drosophila Gp150 is requiredfor early ommatidial development through modulation of Notch signaling. EMBO J.
21, 1074-1083.
Forgac, M. (2007). Vacuolar ATPases: rotary proton pumps in physiology andpathophysiology. Nat. Rev. Mol. Cell Biol. 8, 917-929.
Fortini, M. E. and Bilder, D. (2009). Endocytic regulation of Notch signaling. Curr.
Opin. Genet. Dev. 19, 323-328.
Fostier, M., Evans, D. A., Artavanis-Tsakonas, S. and Baron, M. (1998). Geneticcharacterization of the Drosophila melanogaster Suppressor of deltex gene: Aregulator of notch signaling. Genetics 150, 1477-1485.
Furthauer, M. and Gonzalez-Gaitan, M. (2009). Endocytic regulation of notchsignalling during development. Traffic 10, 792-802.
Fuwa, T. J., Hori, K., Sasamura, T., Higgs, J., Baron, M. and Matsuno, K. (2006).The first deltex null mutant indicates tissue-specific deltex-dependent Notch signalingin Drosophila. Mol. Genet. Genomics 275, 251-263.
Gallagher, C. M. and Knoblich, J. A. (2006). The conserved c2 domain protein lethal(2) giant discs regulates protein trafficking in Drosophila. Dev. Cell 11, 641-653.
Gridley, T. (2003). Notch signaling and inherited disease syndromes. Hum. Mol. Genet.
12, 9R-13R.
Hartenstein, V. and Posakony, J. W. (1990). A dual function of the Notch gene inDrosophila sensillum development. Dev. Biol. 142, 13-30.
Hehnly, H. and Stamnes, M. (2007). Regulating cytoskeleton-based vesicle motility.FEBS Lett. 581, 2112-2118.
Heitzler, P. and Simpson, P. (1991). The choice of cell fate in the epidermis ofDrosophila. Cell 64, 1083-1092.
Henne, W. M., Buchkovich, N. J. and Emr, S. D. (2011). The ESCRT pathway. Dev.
Cell 21, 77-91.
Herranz, H., Stamataki, E., Feiguin, F. and Milan, M. (2006). Self-refinement ofNotch activity through the transmembrane protein Crumbs: modulation of gamma-secretase activity. EMBO Rep. 7, 297-302.
Herz, H. M., Woodfield, S. E., Chen, Z., Bolduc, C. and Bergmann, A. (2009).Common and distinct genetic properties of ESCRT-II components in Drosophila.PLoS ONE 4, e4165.
Hirst, J., Borner, G. H., Harbour, M. and Robinson, M. S. (2005). The aftiphilin/p200/gamma-synergin complex. Mol. Biol. Cell 16, 2554-2565.
Hsu, S. C., TerBush, D., Abraham, M. and Guo, W. (2004). The exocyst complex inpolarized exocytosis. Int. Rev. Cytol. 233, 243-265.
Hutterer, A. and Knoblich, J. A. (2005). Numb and alpha-Adaptin regulate Sanpodoendocytosis to specify cell fate in Drosophila external sensory organs. EMBO Rep. 6,836-842.
Itzen, A., Bleimling, N., Ignatev, A., Pylypenko, O. and Rak, A. (2006). Purification,crystallization and preliminary X-ray crystallographic analysis of mammalian MSS4-Rab8 GTPase protein complex. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun.
62, 113-116.
Jafar-Nejad, H., Andrews, H. K., Acar, M., Bayat, V., Wirtz-Peitz, F., Mehta, S. Q.,
Knoblich, J. A. and Bellen, H. J. (2005). Sec15, a component of the exocyst,promotes notch signaling during the asymmetric division of Drosophila sensory organprecursors. Dev. Cell 9, 351-363.
Jankovics, F., Sinka, R. and Erdelyi, M. (2001). An interaction type of genetic screen
reveals a role of the Rab11 gene in oskar mRNA localization in the developing
Drosophila melanogaster oocyte. Genetics 158, 1177-1188.
Jekely, G. and Rørth, P. (2003). Hrs mediates downregulation of multiple signalling
receptors in Drosophila. EMBO Rep. 4, 1163-1168.
Justice, N., Roegiers, F., Jan, L. Y. and Jan, Y. N. (2003). Lethal giant larvae acts
together with numb in notch inhibition and cell fate specification in the Drosophila
adult sensory organ precursor lineage. Curr. Biol. 13, 778-783.
Klueg, K. M. and Muskavitch, M. A. (1999). Ligand-receptor interactions and trans-
endocytosis of Delta, Serrate and Notch: members of the Notch signalling pathway in
Drosophila. J. Cell Sci. 112, 3289-3297.
Knoblich, J. A. (2010). Asymmetric cell division: recent developments and their
implications for tumour biology. Nat. Rev. Mol. Cell Biol. 11, 849-860.
Kojima, T., Tsuji, T. and Saigo, K. (2005). A concerted action of a paired-type
homeobox gene, aristaless, and a homolog of Hox11/tlx homeobox gene, clawless, is
essential for the distal tip development of the Drosophila leg. Dev. Biol. 279, 434-445.
Kopan, R. and Ilagan, M. X. (2009). The canonical Notch signaling pathway:
unfolding the activation mechanism. Cell 137, 216-233.
Kouranti, I., Sachse, M., Arouche, N., Goud, B. and Echard, A. (2006). Rab35
regulates an endocytic recycling pathway essential for the terminal steps of
cytokinesis. Curr. Biol. 16, 1719-1725.
Langevin, J., Le Borgne, R., Rosenfeld, F., Gho, M., Schweisguth, F. and Bellaıche,
Y. (2005). Lethal giant larvae controls the localization of notch-signaling regulators
numb, neuralized, and Sanpodo in Drosophila sensory-organ precursor cells. Curr.
Biol. 15, 955-962.
Le Borgne, R. (2006). Regulation of Notch signalling by endocytosis and endosomal
sorting. Curr. Opin. Cell Biol. 18, 213-222.
Le Borgne, R. and Schweisguth, F. (2003). Unequal segregation of Neuralized biases
Notch activation during asymmetric cell division. Dev. Cell 5, 139-148.
Le Borgne, R., Remaud, S., Hamel, S. and Schweisguth, F. (2005). Two distinct E3
ubiquitin ligases have complementary functions in the regulation of delta and serrate
signaling in Drosophila. PLoS Biol. 3, e96.
Le Bras, S., Loyer, N. and Le Borgne, R. (2011). The multiple facets of ubiquitination
in the regulation of notch signaling pathway. Traffic 12, 149-161.
Lu, H. and Bilder, D. (2005). Endocytic control of epithelial polarity and proliferation
in Drosophila. Nat. Cell Biol. 7, 1139-1132.
Lui, W. W., Collins, B. M., Hirst, J., Motley, A., Millar, C., Schu, P., Owen, D. J.
and Robinson, M. S. (2003). Binding partners for the COOH-terminal appendage
domains of the GGAs and gamma-adaptin. Mol. Biol. Cell 14, 2385-2398.
Maldonado-Baez, L. and Wendland, B. (2006). Endocytic adaptors: recruiters,
coordinators and regulators. Trends Cell Biol. 16, 505-513.
Malsam, J., Kreye, S. and Sollner, T. H. (2008). Membrane fusion: SNAREs and
regulation. Cell. Mol. Life Sci. 65, 2814-2832.
Miele, L., Golde, T. and Osborne, B. (2006). Notch signaling in cancer. Curr. Mol.
Med. 6, 905-918.
Mistry, H., Wilson, B. A., Roberts, I. J., O’Kane, C. J. and Skeath, J. B. (2004).
Cullin-3 regulates pattern formation, external sensory organ development and cell
survival during Drosophila development. Mech. Dev. 121, 1495-1507.
Mlodzik, M., Baker, N. E. and Rubin, G. M. (1990). Isolation and expression of
scabrous, a gene regulating neurogenesis in Drosophila. Genes Dev. 4, 1848-1861.
Moberg, K. H., Schelble, S., Burdick, S. K. and Hariharan, I. K. (2005). Mutations in
erupted, the Drosophila ortholog of mammalian tumor susceptibility gene 101, elicit
non-cell-autonomous overgrowth. Dev. Cell 9, 699-710.
Mondal, K., Dastidar, A. G., Singh, G., Madhusudhanan, S., Gande, S. L.,
VijayRaghavan, K. and Varadarajan, R. (2007). Design and isolation of
temperature-sensitive mutants of Gal4 in yeast and Drosophila. J. Mol. Biol. 370,
939-950.
Mukherjee, A., Veraksa, A., Bauer, A., Rosse, C., Camonis, J. and Artavanis-
Tsakonas, S. (2005). Regulation of Notch signalling by non-visual beta-arrestin. Nat.
Cell Biol. 7, 1091-1101.
Mummery-Widmer, J. L., Yamazaki, M., Stoeger, T., Novatchkova, M., Bhalerao,
S., Chen, D., Dietzl, G., Dickson, B. J. and Knoblich, J. A. (2009). Genome-wide
analysis of Notch signalling in Drosophila by transgenic RNAi. Nature 458, 987-992.
Murphy, J. E., Padilla, B. E., Hasdemir, B., Cottrell, G. S. and Bunnett, N. W.
(2009). Endosomes: a legitimate platform for the signaling train. Proc. Natl. Acad.
Sci. USA 106, 17615-17622.
Musse, A. A., Meloty-Kapella, L. and Weinmaster, G. (2012). Notch ligand
endocytosis: mechanistic basis of signaling activity. Semin. Cell Dev. Biol. 23, 429-
436.
Nichols, J. T., Miyamoto, A., Olsen, S. L., D’Souza, B., Yao, C. and Weinmaster, G.
(2007). DSL ligand endocytosis physically dissociates Notch1 heterodimers before
activating proteolysis can occur. J. Cell Biol. 176, 445-458.
Nuoffer, C., Wu, S. K., Dascher, C. and Balch, W. E. (1997). Mss4 does not function
as an exchange factor for Rab in endoplasmic reticulum to Golgi transport. Mol. Biol.
Cell 8, 1305-1316.
O’Connor-Giles, K. M. and Skeath, J. B. (2003). Numb inhibits membrane
localization of Sanpodo, a four-pass transmembrane protein, to promote asymmetric
divisions in Drosophila. Dev. Cell 5, 231-243.
Ohshiro, T., Yagami, T., Zhang, C. and Matsuzaki, F. (2000). Role of cortical
tumour-suppressor proteins in asymmetric division of Drosophila neuroblast. Nature
408, 593-596.
Journal of Cell Science 125 (20)4900

Journ
alof
Cell
Scie
nce
Overstreet, E., Chen, X., Wendland, B. and Fischer, J. A. (2003). Either part of aDrosophila epsin protein, divided after the ENTH domain, functions in endocytosis ofdelta in the developing eye. Curr. Biol. 13, 854-860.
Overstreet, E., Fitch, E. and Fischer, J. A. (2004). Fat facets and Liquid facetspromote Delta endocytosis and Delta signaling in the signaling cells. Development
131, 5355-5366.Panin, V. M., Shao, L., Lei, L., Moloney, D. J., Irvine, K. D. and Haltiwanger, R. S.
(2002). Notch ligands are substrates for protein O-fucosyltransferase-1 and Fringe.J. Biol. Chem. 277, 29945-29952.
Pi, H., Wu, H. J. and Chien, C. T. (2001). A dual function of phyllopod in Drosophilaexternal sensory organ development: cell fate specification of sensory organ precursorand its progeny. Development 128, 2699-2710.
Rajan, A., Tien, A. C., Haueter, C. M., Schulze, K. L. and Bellen, H. J. (2009). TheArp2/3 complex and WASp are required for apical trafficking of Delta into microvilliduring cell fate specification of sensory organ precursors. Nat. Cell Biol. 11, 815-824.
Resino, J. and Garcıa-Bellido, A. (2004). Drosophila genetic variants that change cellsize and rate of proliferation affect cell communication and hence patterning. Mech.
Dev. 121, 351-364.Roegiers, F., Jan, L. Y. and Jan, Y. N. (2005). Regulation of membrane localization of
Sanpodo by lethal giant larvae and neuralized in asymmetrically dividing cells ofDrosophila sensory organs. Mol. Biol. Cell 16, 3480-3487.
Royet, J., Bouwmeester, T. and Cohen, S. M. (1998). Notchless encodes a novelWD40-repeat-containing protein that modulates Notch signaling activity. EMBO J.
17, 7351-7360.Sakata, T., Sakaguchi, H., Tsuda, L., Higashitani, A., Aigaki, T., Matsuno, K. and
Hayashi, S. (2004). Drosophila Nedd4 regulates endocytosis of notch and suppressesits ligand-independent activation. Curr. Biol. 14, 2228-2236.
Sasamura, T., Ishikawa, H. O., Sasaki, N., Higashi, S., Kanai, M., Nakao, S.,
Ayukawa, T., Aigaki, T., Noda, K., Miyoshi, E. et al. (2007). TheO-fucosyltransferase O-fut1 is an extracellular component that is essential for theconstitutive endocytic trafficking of Notch in Drosophila. Development 134, 1347-1356.
Seidner, G. A., Ye, Y., Faraday, M. M., Alvord, W. G. and Fortini, M. E. (2006).Modeling clinically heterogeneous presenilin mutations with transgenic Drosophila.Curr. Biol. 16, 1026-1033.
Shi, A., Sun, L., Banerjee, R., Tobin, M., Zhang, Y. and Grant, B. D. (2009).Regulation of endosomal clathrin and retromer-mediated endosome to Golgiretrograde transport by the J-domain protein RME-8. EMBO J. 28, 3290-3302.
Simons, K. and Gerl, M. J. (2010). Revitalizing membrane rafts: new tools andinsights. Nat. Rev. Mol. Cell Biol. 11, 688-699.
Tang, H., Rompani, S. B., Atkins, J. B., Zhou, Y., Osterwalder, T. and Zhong,
W. (2005). Numb proteins specify asymmetric cell fates via an endocytosis- andproteasome-independent pathway. Mol. Cell. Biol. 25, 2899-2909.
Tenney, K., Gerber, M., Ilvarsonn, A., Schneider, J., Gause, M., Dorsett, D.,Eissenberg, J. C. and Shilatifard, A. (2006). Drosophila Rtf1 functions in histone
methylation, gene expression, and Notch signaling. Proc. Natl. Acad. Sci. USA 103,11970-11974.
Thompson, B. J., Mathieu, J., Sung, H. H., Loeser, E., Rørth, P. and Cohen, S. M.(2005). Tumor suppressor properties of the ESCRT-II complex component Vps25 inDrosophila. Dev. Cell 9, 711-720.
Tong, X., Zitserman, D., Serebriiskii, I., Andrake, M., Dunbrack, R. and Roegiers,F. (2010). Numb independently antagonizes Sanpodo membrane targeting and Notchsignaling in Drosophila sensory organ precursor cells. Mol. Biol. Cell 21, 802-810.
Tuxworth, R. I., Vivancos, V., O’Hare, M. B. and Tear, G. (2009). Interactionsbetween the juvenile Batten disease gene, CLN3, and the Notch and JNK signallingpathways. Hum. Mol. Genet. 18, 667-678.
Vaccari, T. and Bilder, D. (2005). The Drosophila tumor suppressor vps25 preventsnonautonomous overproliferation by regulating notch trafficking. Dev. Cell 9, 687-698.
Vaccari, T., Lu, H., Kanwar, R., Fortini, M. E. and Bilder, D. (2008). Endosomalentry regulates Notch receptor activation in Drosophila melanogaster. J. Cell Biol.
180, 755-762.Vaccari, T., Rusten, T. E., Menut, L., Nezis, I. P., Brech, A., Stenmark, H. and
Bilder, D. (2009). Comparative analysis of ESCRT-I, ESCRT-II and ESCRT-IIIfunction in Drosophila by efficient isolation of ESCRT mutants. J. Cell Sci. 122,2413-2423.
Vaccari, T., Duchi, S., Cortese, K., Tacchetti, C. and Bilder, D. (2010). The vacuolarATPase is required for physiological as well as pathological activation of the Notchreceptor. Development 137, 1825-1832.
Wang, W. and Struhl, G. (2004). Drosophila Epsin mediates a select endocyticpathway that DSL ligands must enter to activate Notch. Development 131, 5367-5380.
Wang, W. and Struhl, G. (2005). Distinct roles for Mind bomb, Neuralized and Epsinin mediating DSL endocytosis and signaling in Drosophila. Development 132, 2883-2894.
Weinmaster, G. and Fischer, J. A. (2011). Notch ligand ubiquitylation: what is it goodfor? Dev. Cell 21, 134-144.
Wennerberg, K., Rossman, K. L. and Der, C. J. (2005). The Ras superfamily at aglance. J. Cell Sci. 118, 843-846.
Wilkin, M. B., Carbery, A. M., Fostier, M., Aslam, H., Mazaleyrat, S. L., Higgs, J.,
Myat, A., Evans, D. A., Cornell, M. and Baron, M. (2004). Regulation of notchendosomal sorting and signaling by Drosophila Nedd4 family proteins. Curr. Biol. 14,2237-2244.
Windler, S. L. and Bilder, D. (2010). Endocytic internalization routes required fordelta/notch signaling. Curr. Biol. 20, 538-543.
Yamamoto, S., Charng, W. L. and Bellen, H. J. (2010). Endocytosis and intracellulartrafficking of Notch and its ligands. Curr. Top. Dev. Biol. 92, 165-200.
Yan, Y., Denef, N. and Schupbach, T. (2009). The vacuolar proton pump, V-ATPase, isrequired for notch signaling and endosomal trafficking in Drosophila. Dev. Cell 17, 387-402.
Yeh, E., Zhou, L., Rudzik, N. and Boulianne, G. L. (2000). Neuralized functions cellautonomously to regulate Drosophila sense organ development. EMBO J. 19, 4827-4837.
Notch intracellular trafficking regulators 4901




















