
Generation of HLA-B*1516/B*1567/B*1595 and B*1517 alleles (B15 specific group) by transpecies...
8
Generation of HLA-B*1516/B*1567/B*1595 and B*1517 alleles (B15 specific group) by transpecies evolution Angela Román a,b , Isabel Cervera a , Jacqueline Head b , Miriam Rodríguez a , Jorge Martínez-Laso a, * a Unidad de Inmunoterapia Celular, Instituto de Salud Carlos III, Centro Nacional de Microbiología, Majadahonda, Madrid, Spain b Microbiología, Hospital Clínico de San Carlos, Madrid, Spain Received 16 August 2007; received in revised form 10 October 2007; accepted 17 October 2007 Summary The generation of the human leukocyte antigen (HLA)–B*1516, B*1517, B*1567, and B*1595 alleles has been analyzed using exon 1, intron 1, exon 2, intron 2, and exon 3 sequences from human and non-human primates. Results showed that at the first place three evolutionary steps would have been necessary for the generation of HLA-B*1516 and B*1517 alleles: (1) a non-human primate step with the generation of a major histocompatibility complex (MHC)– B*1516/1517–like allele; (2) a human or non-human primate step with two different ways of evolution generating a MHC-B*1516 and a MHC-B*1517 ancestors; and (3) a human step consisting of the generation of HLA-B*1516 and HLA-B*15170101 alleles. After that, HLA-B*1567, B*1595 B*151701012, and B*151702 alleles would be generated by point mutation events. In conclusion these alleles are generated by two different evolutionary pathways. The generation of these alleles points out the importance of the exons/introns in the generation of the evolution of HLA alleles. © 2007 American Society for Histocompatibility and Immunogenetics. Published by Elsevier Inc. All rights reserved. KEYWORDS Gene conversion; Point mutation; HLA-B; MHC; Primates Introduction A total of 125 different B15 subtypes including B*15010101- B*1599 and B*9501-B*9521 alleles have been admitted by the World Health Organization (WHO) Nomenclature Committee for the HLA system [1-3], establishing the largest HLA-B number of subtypes described in the HLA system. Taking into account the great complexity of the generation of the dif- ferent subtypes of B15 alleles, a characteristic sequence (from position 394 to the end of exon 2) exists only in the The contributions of Angela Roman and Isabel Cervera are equal, and the order of authorship is arbitrary. The name listed for the HLA-B*15010101, B*15170101, B*270502, B*4501, and B*5601 sequences has been officially assigned as a con- firmatory by the WHO Nomenclature Committee. This follows the agreed policy that, subject to the conditions stated in the most recent Nomenclature report [see Refs. 1–3 in text], names will be assigned to new sequences as they are identified. List of such new names will be published in the following WHO Nomenclature Report. The nucleotide sequence data reported in this paper have been sub- mitted to the GenBank nucleotide sequence database and have been assigned the following accession numbers: EF203076 (HLA- B*15010101), EF203077 (HLA-B*15170101), EF203078 (HLA-B*270502), EF203079 (HLA-B*4501), and EF203080 (HLA-B*5601). * Corresponding author. Fax: 34-918223423. E-mail address: [email protected] (J. Martínez-Laso). Human Immunology (2007) 68, 1001–1008 0198-8859/$ -see front matter © 2007 American Society for Histocompatibility and Immunogenetics. Published by Elsevier Inc. All rights reserved. doi:10.1016/j.humimm.2007.10.010
Transcript of Generation of HLA-B*1516/B*1567/B*1595 and B*1517 alleles (B15 specific group) by transpecies...

Ga
AJ
a
Sb
R
I
ABW
a
Bfiara
Human Immunology (2007) 68, 1001–1008
0d
eneration of HLA-B*1516/B*1567/B*1595 and B*1517lleles (B15 specific group) by transpecies evolution
ngela Romána,b, Isabel Cerveraa, Jacqueline Headb, Miriam Rodrígueza,orge Martínez-Lasoa,*
Unidad de Inmunoterapia Celular, Instituto de Salud Carlos III, Centro Nacional de Microbiología, Majadahonda, Madrid,painMicrobiología, Hospital Clínico de San Carlos, Madrid, Spain
eceived 16 August 2007; received in revised form 10 October 2007; accepted 17 October 2007
Summary The generation of the human leukocyte antigen (HLA)–B*1516, B*1517, B*1567, andB*1595 alleles has been analyzed using exon 1, intron 1, exon 2, intron 2, and exon 3 sequencesfrom human and non-human primates. Results showed that at the first place three evolutionarysteps would have been necessary for the generation of HLA-B*1516 and B*1517 alleles: (1) anon-human primate step with the generation of a major histocompatibility complex (MHC)–B*1516/1517–like allele; (2) a human or non-human primate step with two different ways ofevolution generating a MHC-B*1516 and a MHC-B*1517 ancestors; and (3) a human step consisting ofthe generation of HLA-B*1516 and HLA-B*15170101 alleles. After that, HLA-B*1567, B*1595B*151701012, and B*151702 alleles would be generated by point mutation events. In conclusionthese alleles are generated by two different evolutionary pathways. The generation of these allelespoints out the importance of the exons/introns in the generation of the evolution of HLA alleles.© 2007 American Society for Histocompatibility and Immunogenetics. Published by Elsevier Inc. Allrights reserved.
KEYWORDSGene conversion;
Point mutation;
HLA-B;
MHC;
Primates
fnaf(
nTmaBE
ntroduction
total of 125 different B15 subtypes including B*15010101-*1599 and B*9501-B*9521 alleles have been admitted by theorld Health Organization (WHO) Nomenclature Committee
The contributions of Angela Roman and Isabel Cervera are equal,nd the order of authorship is arbitrary.
The name listed for the HLA-B*15010101, B*15170101, B*270502,*4501, and B*5601 sequences has been officially assigned as a con-rmatory by the WHO Nomenclature Committee. This follows thegreed policy that, subject to the conditions stated in the mostecent Nomenclature report [see Refs. 1–3 in text], names will be
ssigned to new sequences as they are identified. List of such new198-8859/$ -see front matter © 2007 American Society for Histocompatibioi:10.1016/j.humimm.2007.10.010
or the HLA system [1-3], establishing the largest HLA-Bumber of subtypes described in the HLA system. Taking intoccount the great complexity of the generation of the dif-erent subtypes of B15 alleles, a characteristic sequencefrom position 394 to the end of exon 2) exists only in the
ames will be published in the following WHO Nomenclature Report.he nucleotide sequence data reported in this paper have been sub-itted to the GenBank nucleotide sequence database and have been
ssigned the following accession numbers: EF203076 (HLA-*15010101), EF203077 (HLA-B*15170101), EF203078 (HLA-B*270502),F203079 (HLA-B*4501), and EF203080 (HLA-B*5601).
* Corresponding author. Fax: 34-918223423.
E-mail address: [email protected] (J. Martínez-Laso).lity and Immunogenetics. Published by Elsevier Inc. All rights reserved.

BdoBsthpc
7SBBaa
S
S32bta
PPBsG4Ip
PpapEiqM
R
Ftt
N
TcB
baln3f3fIaiTpi
t3ttMm
H
TgaBcpoaBBwwpMalucm
H
Ast7astBttraa3
1002 A. Román et al.
*1517, B*1516, B*1567, and B*1595 alleles, suggesting aifferent and specific pathway of evolution respect to thether B15 alleles. This characteristic motif is shared with*57 and B*58 alleles and their generation has been de-cribed coming from HLA-B*1516 or B*1517 [4]. The origin ofhis motif still remains unclear taking into account only theuman sequences but a possible explanation could be hy-othesized considering the non-human primate major histo-ompatibility complex (MHC) sequences.
Alleles in this family encode molecules carrying B62, 63,5, 76, and 77 (subgroups of B15) serologic specificities.erologically distinct molecules, B71 and B72 (subtypes of70), are also encoded by alleles in the B15 family. Although15/70 is found at high frequencies in all ethnic groups,lleles encoding these specificities and their frequencies arept to be different in distinct populations [5-7].
ubjects and methods
equences comprising partial exon 1, intron 1, exon 2, intron 2, exon, and partial intron 3 of B*15010101, B*15170101, B*4501, B*5601,70502 were sequenced in five unrelated healthy umbilical cordlood units of Caucasian origin (Madrid, Spain) obtained from full-erm normal deliveries, after ethical committee approval. Intron 1nd intron 2 of B*5601 were described as new sequences.
Non-human primate sequences Gogo-B*0503, Popy-B*0602,opy-B*0901, Papa-B*05, Papa-B*06, Papa-B*07 and Patr-B*0203atr-B*0101, Patr-B*0401, Patr-B*0501, Patr-B*2001 and Popy-*0401 were taken from Martínez-Laso et al. [8]. The correspondingequences from HLAB*1504, 1515, 1539, 4901 were obtained fromomez Casado et al. [5,6] and HLA-B*15170102, 151702, 1566, 4504,7010101, 5001, 5401, and 5512 allele sequences were taken fromMGT-HLA database (www.ebi.ac.uk/imgt/hla) [1]. The intron com-ilation by Cereb et al. [9] was also used.
Genomic DNA was isolated from peripheral blood using REALURE SPIN KIT (REALTM, Durviz, Spain) following the manufacturer’srotocol. HLA-B sequences were obtained by polymerase chain re-ction (PCR) using specific primers (5=BE1c, 5=BE1d, and 3=BCI3) asreviously described [5,10]. PCR products were purified usingXOSAP-IT® (USB Corp., Cleveland, OH) methodology and automat-cally sequenced in a 3700 ABI sequencer. Alignment of the se-uences and neighbor-joining dendrograms were conducted usingEGA 3.1 software [11].
esults
rom the results obtained, three different steps related tohe trans-species evolution could be defined in the genera-ion of the alleles studied, as described below.
on-human primate evolution step
his step took place before the speciation and consisted in twoonsecutive events. First, a Non-Human Primate (NHP)–MHC-
ABBREVIATIONS
HLA human leukocyte antigenMHC major histocompatibility complexPCR polymerase chain reaction
*15 ancestor allele was generated by a gene conversion event M
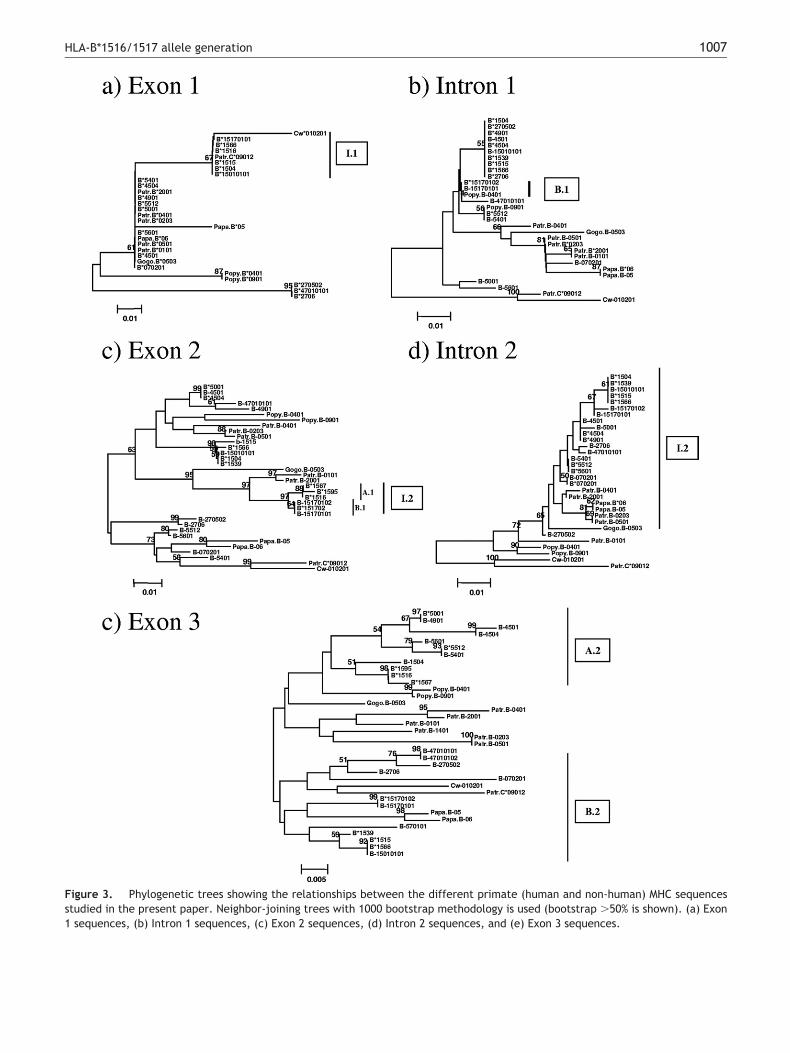
etween a NHP-MHC-C allele as a donor and a NHP-MHC-Bllele as a receptor of the characteristic GA sequence motifocated at exon 1 (denoted as I.1 in Figures 1 and 2). Phyloge-etic tree of exon 1 group alleles involving in this event (Figurea). Second, this NHP-MHC-B*15 ancestor allele received aragment comprising at least the end of exon 2 (nucleotides94–471) and the beginning of intron 2 (nucleotides 472–487)rom a sequence similar to Patr-MHC-B*0101/2001 (denoted as.2; Figures 1 and 2) generating a NHP-MHC-B*1516/1517–likellele. Phylogenetic tree of exon 2 shows the group of allelesnvolved in the corresponding donated fragment (Figure 3c).he intron 1 dendrogram is not able to show the same effect,robably because of the size of the corresponding fragmentnvolved (Figure 3d).
In addition the sequence motif GA is present in every one ofhe HLA-B*15 alleles, and the fragment comprising nucleotides94-487 appears only in some Patr-MHC-B alleles. Thereforehe donation of GA must have occurred in a step of the evolu-ion previous to the donation of the fragment mentioned in aHC-B*15 ancestor allele, discarding every possibility of a hu-an donation of the GA present in the HLA-Cw alleles.
uman or non-human primate evolution step
wo different evolutionary pathways would be necessary toenerate the two groups of alleles (HLA-B*1516/1567/1595nd B*1517; A and B in Figures 1 and 2) from the NHP-MHC-*1516/1517–like allele. The first pathway would be a geneonversion event consisting in a G nucleotide donation atosition 293 (exon 2) from a NHP-MHC-B allele, (this nucle-tide is the most common at this position) or some HLA-B*15llele as B*1503 as a possible donor and the NHP-MHC-*1516/1517–like allele as a receptor, generating the MHC-*1516 ancestor (Figures 1 and 2; A.1). The second pathwayould be another, different gene conversion event thatould take place, whereby a donation of the fragment fromositions 85–89 of intron 1 would be donated by a NHP-HC-B allele or some HLA-B allele belonging to the B*54, 55,nd 56 group over the same NHP-MHC-B*1516/1517–like al-ele as a receptor, yielding the MHC-B*1517 ancestor (Fig-res 1 and 2; B.1). Phylogenetic analysis of these eventsannot be observed because of the short size of the frag-ents involved (Figures 3b and 3c).
uman evolution step
fter the establishment of the two groups, the followingtep is theorized. (A) The MHC-B*1516 ancestor would behe receptor of a characteristic fragment from position27–752 of exon 3 of the HLA-B*45/49/50 alleles, gener-ting an HLA-B*1516 –like allele (Figures 1 and 2; A.2). Thisequence had a point mutation event changing C at posi-ion 211 (exon 2) by T (which only appears in B*1516,*1567, and B*1595; Figures 1 and 2; A.3). With this eventhe HLA-B*1516 would be created. However a point muta-ion event at positions 879 (G ¡ A) and 385 (G ¡ A),espectively, would be necessary to generate the B*1567nd B*1595 alleles (Figures 1 and 2; A.4). (B) In this case,donation of a fragment from position 782 to 792 of exonof alleles as HLA-B*27/47 would take place over the
HC-B*1517 ancestor, creating the HLA-B*15170101 (Fig-
FAbB
H
igure 1. Complete exon 1, intron 1, exon 2, intron 2, exon 3, and partial intron 3 sequences of the HLA-B alleles analyzed.sterisks (*) reflect unknown sequences. Dots (.) indicate deleted nucleotides. Nucleotide agreement with the consensus is indicatedy a hyphen (-). Black region indicates the shared sequences between different alleles involved in the genetic events to generate the
1003LA-B*1516/1517 allele generation
*1517 group of alleles and the B*1516, B*1567, and B*1595 alleles. (For additional information see text and Figure 2.)

1004 A. Román et al.
Figure 1. (Continued)

1005HLA-B*1516/1517 allele generation
Figure 1. (Continued)

uw53a(
D
TBpltspimoeau
aricsaimp
A
TMSgdt
F(N
1006 A. Román et al.
res 1 and 2; B.2). Alleles B*151701012 and B*151702ould be generated by point mutation events at positions28 (C ¡ G) of intron 2 and 874 (C ¡ T) of exon, respectively (Figures 1 and 2; B.3). These two groupsre confirmed by the corresponding phylogenetic treeFigure 3e).
iscussion
his study offers a hypothesis on how the HLA-B*1516 and*1517 subgroups were formed by different evolutionaryathways belonging even to the same group. In general si-ent mutations or mutations in introns can be maintained inhe generation of new alleles because they do not affect theequence of the amino acids or protein function. In theresent study exon fragments belonging to other HLA spec-ficities also are exchanged, conferring new antigenic deter-inants. This fact is important in the immunologic behaviorf the HLA alleles, in which the compatibility among differ-nt alleles in a common group (i.e., the HLA-B15 group oflleles) is not enough to avoid immunologic responses (i.e.,
igure 2. A possible scheme of the HLA-B*1516, 1567, 1595, 1GC) and point mutation (PM) events. (1) Primate MHC-B includeHP-MHC-B alleles or HLA-B*54/B*55/B*56 alleles.
nrelated transplants). I
In summary some studies of the generation of HLA-And -B alleles have shown that both gene conversion andecombination events are mechanisms commonly involvedn the polymorphism generation; these events would in-lude noncoding intron 1 and intron 2 regions [4,5,12,13],tressing the importance of introns for new alleles gener-tion. The study of HLA-B intron sequences is increasinglymportant to understand their role in evolution and poly-orphism generation (Martínez-Laso et al., manuscript inreparation) and to design strategies for HLA genotyping.
cknowledgments
his work was supported in part by grants from the Spanishinistry of health (PI05/0338, CP04/00026), and Instituto dealud Carlos III (MPY1180/06). Support was also provided byrants from the Fundación para la Investigación Biomédicael Hospital Clínico San Carlos (to A.R. and J.H.). The au-hors also thank the Fundación para la Cooperación y Salud
101, and 151702 allele generation, comprising gene conversion-MHC-B alleles or HLA-B*15 alleles; (2) primate MHC-B includes
5170s NHP
nternacional Carlos III for its excellent technical support.
Fs1
1007HLA-B*1516/1517 allele generation
igure 3. Phylogenetic trees showing the relationships between the different primate (human and non-human) MHC sequencestudied in the present paper. Neighbor-joining trees with 1000 bootstrap methodology is used (bootstrap �50% is shown). (a) Exon
sequences, (b) Intron 1 sequences, (c) Exon 2 sequences, (d) Intron 2 sequences, and (e) Exon 3 sequences.
R
[
[
[
[
1008 A. Román et al.
eferences
[1] Marsh SGE. Nomenclature for factors of the HLA system updateSeptember 2006. Tissue Antigens 2006;68:540.
[2] Marsh SGE, Albert ED, Bodmer WF, Bontrop RE, Dupont B, ErlichHA, et al. Nomenclature for factors of the HLA system, 2004.Tissue Antigens 2005;65:301.
[3] Robinson J, Waller MJ, Parham P, de Groot N, Bontrop R,Kennedy LJ, et al. IMGT/HLA and IMGT/MHC: Sequence data-bases for the study of the major histocompatibility complex.Nucl Acids Res 2003;31:311.
[4] Martinez-Laso J, Moscoso J, Zamora J, Martin-Villa M, Lowy E,Vargas-Alarcon G, et al. Different evolutionary pathway ofB*570101 and B*5801 (B17 group) alleles based in intron se-quences. Immunogenetics 2004;55:866.
[5] Gomez-Casado E, Vargas-Alarcón G, Martínez-Laso J, Perez-Blas M, Granados J, Layrisse Z, et al. Generation of the HLA-B35, -B5, -B16, and -B15 groups of alleles studied by intron 1and 2 sequences analysis. Immunogenetics 1997;46:469.
[6] Gomez-Casado E, Vargas-Alarcón G, Martínez-Laso J, GranadosJ, Varela P, Alegre R, et al. Evolutionary relationship betweenHLA-B alleles as indicated by an analysis of intron sequences.
Tissue Antigens 1999;53:153.[7] Lee KW, Jeon H, Park JY. HLA-B*15 diversity in the Koreanpopulation. Tissue Antigens 2000;56:428.
[8] Martínez-Laso J, Gomez-Casado E, Arnaiz-Villena A. Descrip-tion of seven new non-human primate MHC-B alleles. TissueAntigens 2006;67:85.
[9] Cereb N, Kong Y, Lee S, Maye P, Yang SY. Nucleotide sequencesof MHC class I introns 1, 2 and 3 in humans and intron 2 innonhuman primates. Tissue Antigens 1996;49:498.
10] Dunn PP, Day S, Williams S, Bendukidze N. DNA sequencing as atissue-typing tool. Methods Mol Med 2004;91:233.
11] Kumar S, Tamura K, Nei M. MEGA3: Integrated software formolecular evolutionary genetics analysis and sequence align-ment. Brief Bioinform 2004;5:150.
12] Vargas-Alarcón G, Gómez-Casado E, Martínez-Laso J, Granados J,Layrisse Z, Alegre R, et al. Differences in intron 2 sequences be-tween B*39061 and B*39062 in Amerindians: Comparison withthose of B*3901, B*5101, and B*52012 alleles. Immunogenetics1997;45:436.
13] Vargas-Alarcón G, Martínez-Laso J, Gomez-Casado E, Perez-Blas M, Granados J, Alegre R, et al. Description of HLA-A*6803and A*68N in Mazatecan Indians from Mexico. Immunogenetics
1997;46:446.



















