
three -dimensional ultrasound monitoring the utero-ovarian ...
Upload
independentCategory
view
4download
0
doi:10.1006/mthe.2002.0599, available online at http://www.idealibrary.com on IDEAL
Gene Transfer to Ovarian Cancer Versus Normal Tissueswith Fiber-Modified Adenoviruses
Anna Kanerva,1 Minghui Wang,1 Gerd J. Bauerschmitz,1 John T. Lam,1 Renee A. Desmond,2
Snehal M. Bhoola,3 Mack N. Barnes,3 Ronald D. Alvarez,3 Gene P. Siegal,1,4 David T. Curiel,1
and Akseli Hemminki1,5,*
1Division of Human Gene Therapy, Departments of Medicine, Pathology and Surgery, 2Department of Medicine, 3Obstetrics and Gynecology, 4Pathology, CellBiology and Surgery, 5Biomedical Engineering, University of Alabama at Birmingham, Birmingham, Alabama 35294, USA
*To whom correspondence and reprint requests should be addressed. Fax: (205) 975-8565. E-mail: [email protected].
Adenovirus serotype 5 (Ad5) displays unparalleled gene transfer efficacy to cells with high cox-sackie-adenovirus receptor (CAR) expression. Unfortunately, cells isolated from clinical humancancers, both ovarian and other types, express highly variable and often low levels of CAR.Fortunately, native Ad5 tropism can be modified to circumvent CAR deficiency and to enhanceinfectivity. Ad5/3luc1 incorporates the serotype 3 fiber knob and binds to a receptor distinctfrom CAR, while the fiber of Ad5lucRGD is modified with an RGD-4C motif, allowing CAR-inde-pendent binding to integrins. We studied the liver tropism and blood clearance of these virusesafter intravenous (i.v.) injection, and biodistribution after intraperitoneal (i.p.) injection totumor-bearing mice. To estimate efficacy, we assessed gene transfer to purified human primaryovarian cancer cells, and in a mouse model of ovarian cancer. Ad5/3luc1 achieved improvedgene transfer over Ad5lucRGD, and both infectivity-enhanced viruses were superior to the iso-genic control with an unmodified Ad5 capsid. In the presence of malignant ascites, gene trans-fer was improved with both Ad5/3luc1 and Ad5lucRGD. Thus, retargeting to the Ad3 receptorenhances gene transfer to clinically relevant ovarian cancer substrates, while the mouse toxi-city and biodistribution profile of both fiber-modified Ad vectors is comparable to Ad5.
Key Words: cancer, gene therapy, ovarian cancer, adenovirus, pseudotyping, toxicity, biodistribution
ARTICLE
INTRODUCTION
The adenovirus (Ad) is a useful cancer gene therapy vec-tor, because it infects both proliferating and quiescent cellsand its immunogenicity may help to evoke an immuneresponse toward potentially remaining tumor cells [1].Most importantly, > 700 cancer patients have received Adintratumorally, intraarterially, intraperitoneally, andintravenously in clinical trials, with few severe side effectsand generally mild toxicity [2,3]. Entry of Ad into cellsinitiates with binding of the fiber knob to the primaryreceptor, which is the coxsackie-adenovirus receptor (CAR)for most serotypes, including the ubiquitously usedserotype 5 (Ad5) [1]. Binding is followed by interaction ofthe Ad penton base with cellular �v� group integrins,which leads to internalization. Recent data suggest thatbinding is the rate-limiting step [4–9]. Thus, although Adhas a number of characteristics that make it an appealingagent for the treatment of cancer, lack of the primaryreceptor could make target tissues refractory.
MOLECULAR THERAPY Vol. 5, No. 6, June 2002Copyright © The American Society of Gene Therapy1525-0016/02 $35.00
Most ovarian cancer patients have peritoneally dis-seminated disease, which is associated with a dismal prog-nosis. However, the compartmentalized nature of the dis-ease creates a rationale for local treatment.Intraperitoneally administered gene therapy with Ad hasbeen attempted in several trials [10–13]. Importantly, thesafety data have been excellent, with doses as high as 7.5� 1013 viral particles (VP)/day administered for 5 consec-utive days, without dose-limiting toxicity [12]. In contrast,gene transfer data have been minimal, and there is littleevidence supporting a potential clinical benefit.
The reason for the low gene transfer efficacy to clini-cal tumors is becoming increasingly clear, because there ismounting evidence that tumor cells extracted from humanovarian cancer patient samples express highly variable andoften low amounts of CAR [14–18]. This phenomenon isnot restricted to ovarian cancer and could be a generalphenomenon associated with malignant progression[4,5,7,19,20]. In contrast, CAR is expressed ubiquitously on
695
ARTICLE doi:10.1006/mthe.2002.0599, available online at http://www.idealibrary.com on IDEAL
most normal epithelial tissues [21].Thus, without retargeting, low CARexpression on tumor cells maycause low gene transfer efficiencyto cancer cells, but high transduc-tion and consequent toxicity tononmalignant cells. The functionof CAR is not well understood, butit may be associated with adhesionand tight junctions [22–24]. It hasbeen suggested that CAR levels cor-relate inversely with the aggressive-ness of the tumor and that the pres-ence of CAR causes atumor-suppressing effect [23].
Fortunately, Ad can be directedto receptors not downregulated in c
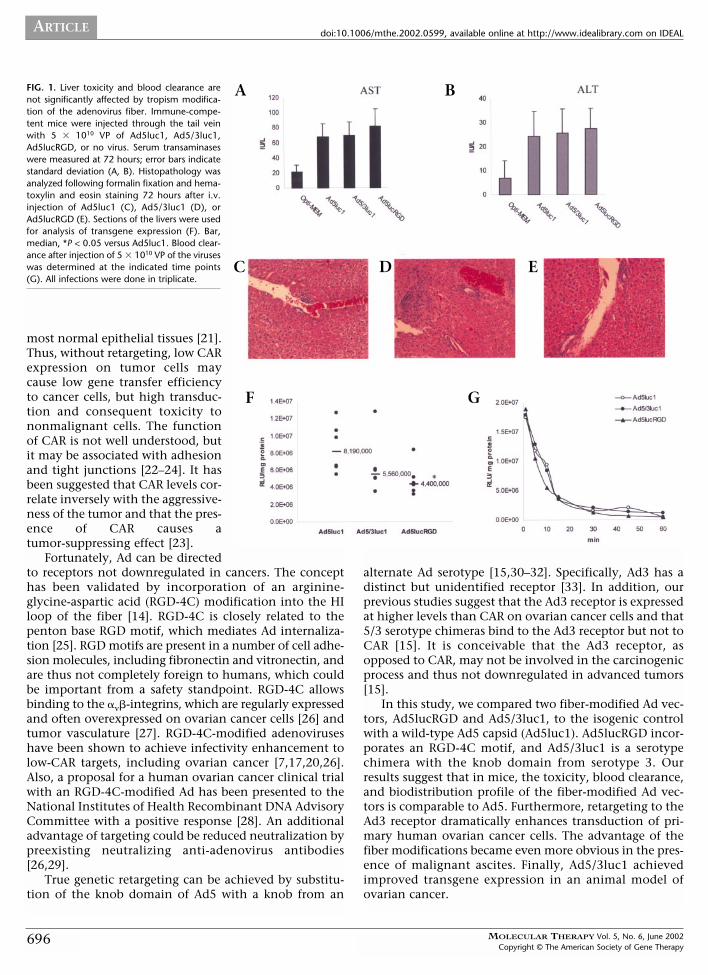
FIG. 1. Liver toxicity and blood clearance arenot significantly affected by tropism modifica-tion of the adenovirus fiber. Immune-compe-tent mice were injected through the tail veinwith 5 � 1010 VP of Ad5luc1, Ad5/3luc1,Ad5lucRGD, or no virus. Serum transaminaseswere measured at 72 hours; error bars indicatestandard deviation (A, B). Histopathology wasanalyzed following formalin fixation and hema-toxylin and eosin staining 72 hours after i.v.injection of Ad5luc1 (C), Ad5/3luc1 (D), orAd5lucRGD (E). Sections of the livers were usedfor analysis of transgene expression (F). Bar,median, *P < 0.05 versus Ad5luc1. Blood clear-ance after injection of 5 � 1010 VP of the viruseswas determined at the indicated time points(G). All infections were done in triplicate.
696
A B
C D E
F G
ancers. The concepthas been validated by incorporation of an arginine-glycine-aspartic acid (RGD-4C) modification into the HIloop of the fiber [14]. RGD-4C is closely related to thepenton base RGD motif, which mediates Ad internaliza-tion [25]. RGD motifs are present in a number of cell adhe-sion molecules, including fibronectin and vitronectin, andare thus not completely foreign to humans, which couldbe important from a safety standpoint. RGD-4C allowsbinding to the �v�-integrins, which are regularly expressedand often overexpressed on ovarian cancer cells [26] andtumor vasculature [27]. RGD-4C-modified adenoviruseshave been shown to achieve infectivity enhancement tolow-CAR targets, including ovarian cancer [7,17,20,26].Also, a proposal for a human ovarian cancer clinical trialwith an RGD-4C-modified Ad has been presented to theNational Institutes of Health Recombinant DNA AdvisoryCommittee with a positive response [28]. An additionaladvantage of targeting could be reduced neutralization bypreexisting neutralizing anti-adenovirus antibodies[26,29].
True genetic retargeting can be achieved by substitu-tion of the knob domain of Ad5 with a knob from an
alternate Ad serotype [15,30–32]. Specifically, Ad3 has adistinct but unidentified receptor [33]. In addition, ourprevious studies suggest that the Ad3 receptor is expressedat higher levels than CAR on ovarian cancer cells and that5/3 serotype chimeras bind to the Ad3 receptor but not toCAR [15]. It is conceivable that the Ad3 receptor, asopposed to CAR, may not be involved in the carcinogenicprocess and thus not downregulated in advanced tumors[15].
In this study, we compared two fiber-modified Ad vec-tors, Ad5lucRGD and Ad5/3luc1, to the isogenic controlwith a wild-type Ad5 capsid (Ad5luc1). Ad5lucRGD incor-porates an RGD-4C motif, and Ad5/3luc1 is a serotypechimera with the knob domain from serotype 3. Ourresults suggest that in mice, the toxicity, blood clearance,and biodistribution profile of the fiber-modified Ad vec-tors is comparable to Ad5. Furthermore, retargeting to theAd3 receptor dramatically enhances transduction of pri-mary human ovarian cancer cells. The advantage of thefiber modifications became even more obvious in the pres-ence of malignant ascites. Finally, Ad5/3luc1 achievedimproved transgene expression in an animal model ofovarian cancer.
MOLECULAR THERAPY Vol. 5, No. 6, June 2002Copyright © The American Society of Gene Therapy
ARTICLEdoi:10.1006/mthe.2002.0599, available online at http://www.idealibrary.com on IDEAL
RESULTS
Liver Toxicity and Blood Clearance Rates of FiberKnob–Modified AdenovirusesTo assess liver toxicity, 5 � 1010 VP were injected intra-venously (i.v.) into immune-competent mice. After 72hours, serum asparate aminotransferase (AST) (Fig. 1A) andalanine aminotransferase (ALT) (Fig. 1B) levels were meas-ured. All viruses caused moderate release of transaminases,with readings threefold higher than without virus. Therewere no significant differences between the groups.
To evaluate toxicity further, livers were harvested andanalyzed histopathologically. Generally, the changes were
TABLE 1: Histopathologic analysis of liver toxicity with tropism-modified adenovirusesa
n Normal Nonspecific hepatitis (%)
Opti-MEM onlyb 5 4 1 (20%)
Ad5luc1 6 3 3 (50%)
Ad5/3luc1 6 5 1 (17%)
Ad5lucRGD 6 3 3 (50%)
aC57BL/6 mice were injected i.v. with 5 � 1010 VP. At 72 hours, livers were harvested,fixed, and stained with hematoxylin and eosin. Histopathology was scored by twopathologists independently and in a blinded manner.bViruses were diluted into 0.3 ml of Opti-MEM.
MOLEC
Copyrigh
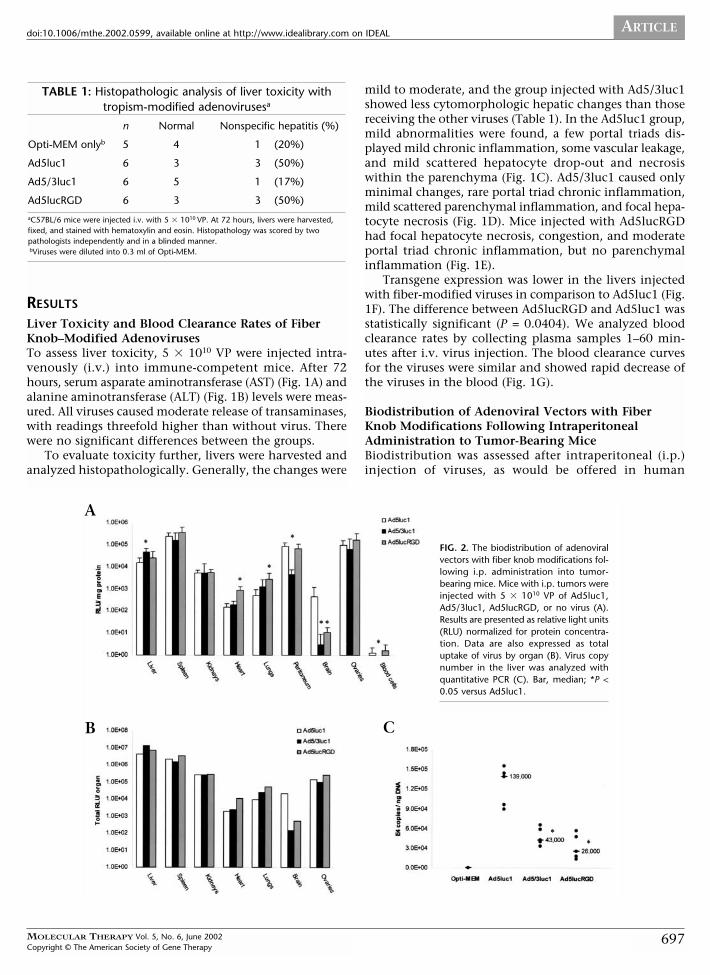
FIG. 2. The biodistribution of adenoviralvectors with fiber knob modifications fol-lowing i.p. administration into tumor-bearing mice. Mice with i.p. tumors wereinjected with 5 � 1010 VP of Ad5luc1,Ad5/3luc1, Ad5lucRGD, or no virus (A).Results are presented as relative light units(RLU) normalized for protein concentra-tion. Data are also expressed as totaluptake of virus by organ (B). Virus copynumber in the liver was analyzed withquantitative PCR (C). Bar, median; *P <0.05 versus Ad5luc1.
A
B C
ULAR THERAPY Vol. 5, No. 6, June 2002t © The American Society of Gene Therapy
mild to moderate, and the group injected with Ad5/3luc1showed less cytomorphologic hepatic changes than thosereceiving the other viruses (Table 1). In the Ad5luc1 group,mild abnormalities were found, a few portal triads dis-played mild chronic inflammation, some vascular leakage,and mild scattered hepatocyte drop-out and necrosiswithin the parenchyma (Fig. 1C). Ad5/3luc1 caused onlyminimal changes, rare portal triad chronic inflammation,mild scattered parenchymal inflammation, and focal hepa-tocyte necrosis (Fig. 1D). Mice injected with Ad5lucRGDhad focal hepatocyte necrosis, congestion, and moderateportal triad chronic inflammation, but no parenchymalinflammation (Fig. 1E).
Transgene expression was lower in the livers injectedwith fiber-modified viruses in comparison to Ad5luc1 (Fig.1F). The difference between Ad5lucRGD and Ad5luc1 wasstatistically significant (P = 0.0404). We analyzed bloodclearance rates by collecting plasma samples 1–60 min-utes after i.v. virus injection. The blood clearance curvesfor the viruses were similar and showed rapid decrease ofthe viruses in the blood (Fig. 1G).
Biodistribution of Adenoviral Vectors with FiberKnob Modifications Following IntraperitonealAdministration to Tumor-Bearing MiceBiodistribution was assessed after intraperitoneal (i.p.)injection of viruses, as would be offered in human
697
ARTICLE doi:10.1006/mthe.2002.0599, available online at http://www.idealibrary.com on IDEAL
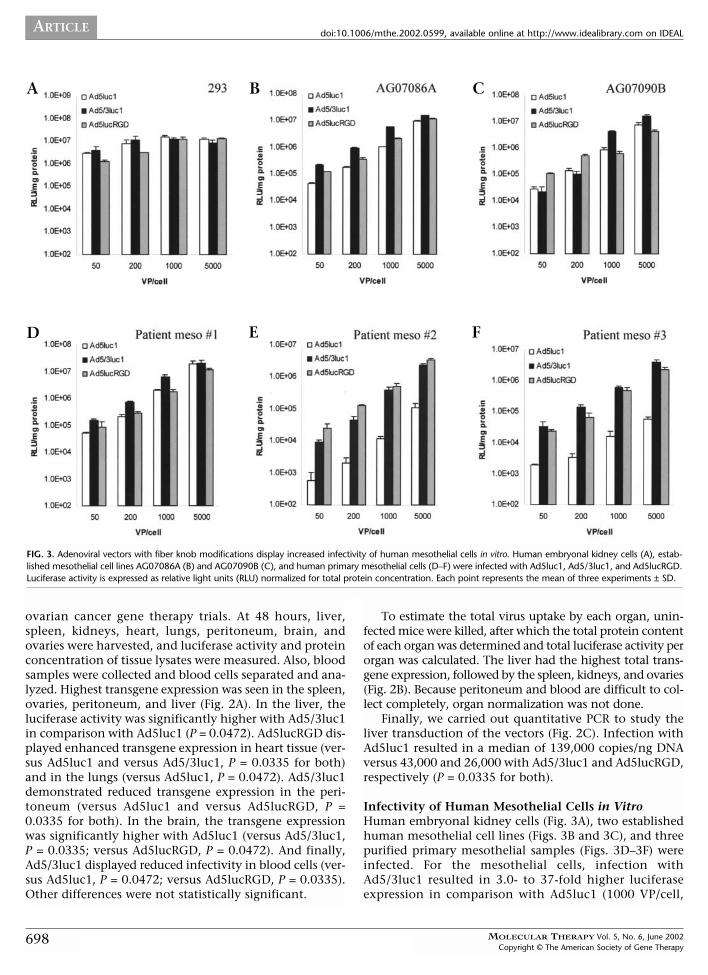
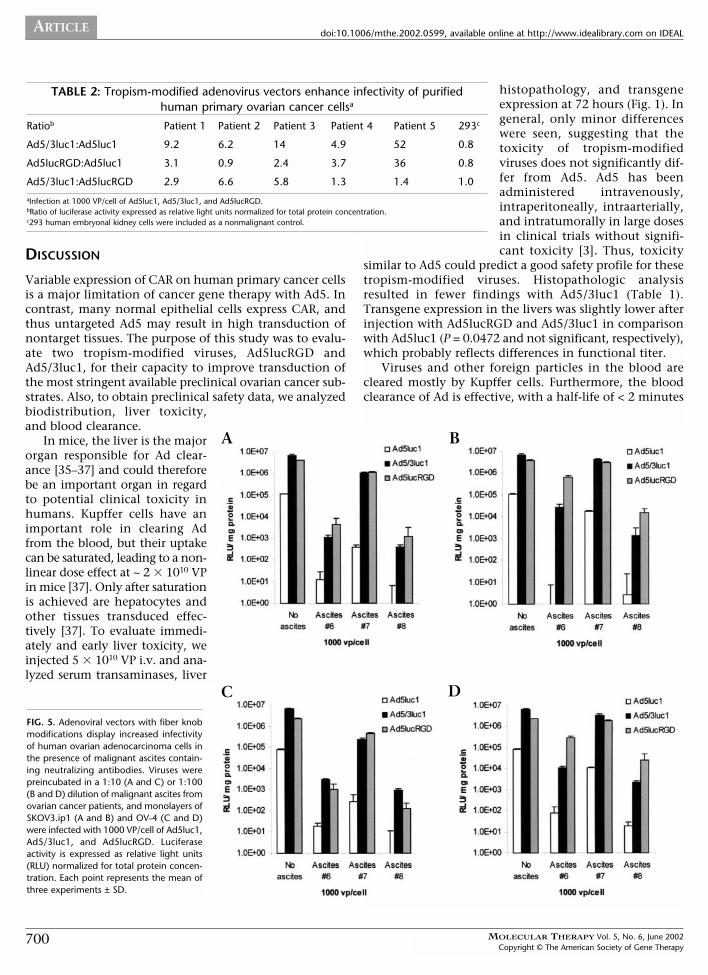
FIG. 3. Adenoviral vectors with fiber knob modifications display increased infectivity of human mesothelial cells in vitro. Human embryonal kidney cells (A), estab-lished mesothelial cell lines AG07086A (B) and AG07090B (C), and human primary mesothelial cells (D–F) were infected with Ad5luc1, Ad5/3luc1, and Ad5lucRGD.Luciferase activity is expressed as relative light units (RLU) normalized for total protein concentration. Each point represents the mean of three experiments ± SD.
A B C
D E F
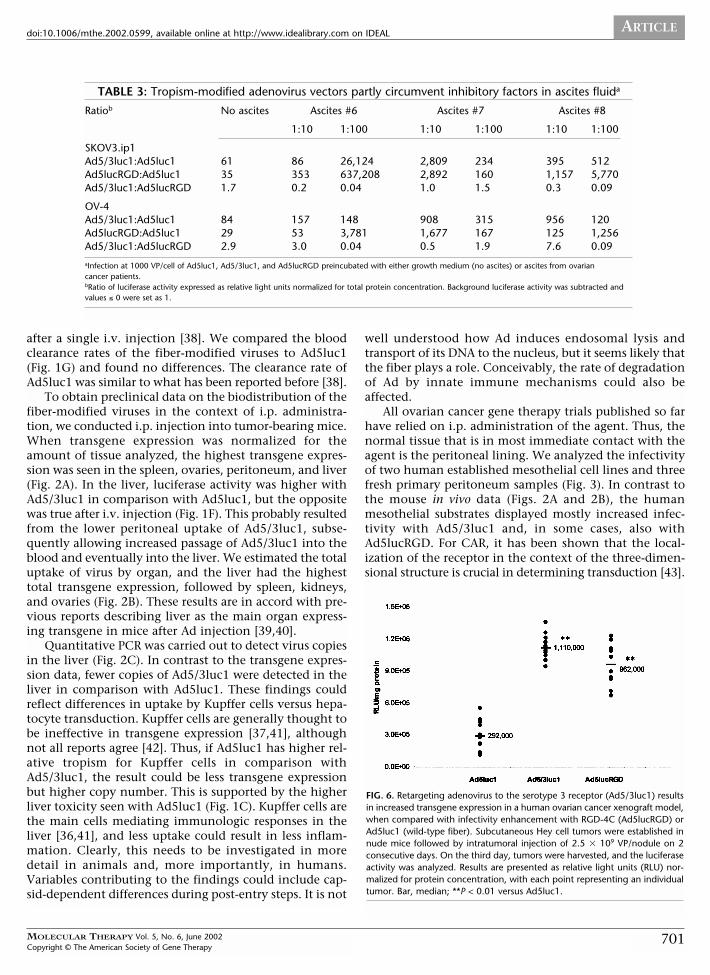
ovarian cancer gene therapy trials. At 48 hours, liver,spleen, kidneys, heart, lungs, peritoneum, brain, andovaries were harvested, and luciferase activity and proteinconcentration of tissue lysates were measured. Also, bloodsamples were collected and blood cells separated and ana-lyzed. Highest transgene expression was seen in the spleen,ovaries, peritoneum, and liver (Fig. 2A). In the liver, theluciferase activity was significantly higher with Ad5/3luc1in comparison with Ad5luc1 (P = 0.0472). Ad5lucRGD dis-played enhanced transgene expression in heart tissue (ver-sus Ad5luc1 and versus Ad5/3luc1, P = 0.0335 for both)and in the lungs (versus Ad5luc1, P = 0.0472). Ad5/3luc1demonstrated reduced transgene expression in the peri-toneum (versus Ad5luc1 and versus Ad5lucRGD, P =0.0335 for both). In the brain, the transgene expressionwas significantly higher with Ad5luc1 (versus Ad5/3luc1,P = 0.0335; versus Ad5lucRGD, P = 0.0472). And finally,Ad5/3luc1 displayed reduced infectivity in blood cells (ver-sus Ad5luc1, P = 0.0472; versus Ad5lucRGD, P = 0.0335).Other differences were not statistically significant.
698
To estimate the total virus uptake by each organ, unin-fected mice were killed, after which the total protein contentof each organ was determined and total luciferase activity perorgan was calculated. The liver had the highest total trans-gene expression, followed by the spleen, kidneys, and ovaries(Fig. 2B). Because peritoneum and blood are difficult to col-lect completely, organ normalization was not done.
Finally, we carried out quantitative PCR to study theliver transduction of the vectors (Fig. 2C). Infection withAd5luc1 resulted in a median of 139,000 copies/ng DNAversus 43,000 and 26,000 with Ad5/3luc1 and Ad5lucRGD,respectively (P = 0.0335 for both).
Infectivity of Human Mesothelial Cells in VitroHuman embryonal kidney cells (Fig. 3A), two establishedhuman mesothelial cell lines (Figs. 3B and 3C), and threepurified primary mesothelial samples (Figs. 3D–3F) wereinfected. For the mesothelial cells, infection withAd5/3luc1 resulted in 3.0- to 37-fold higher luciferaseexpression in comparison with Ad5luc1 (1000 VP/cell,
MOLECULAR THERAPY Vol. 5, No. 6, June 2002Copyright © The American Society of Gene Therapy
ARTICLEdoi:10.1006/mthe.2002.0599, available online at http://www.idealibrary.com on IDEAL
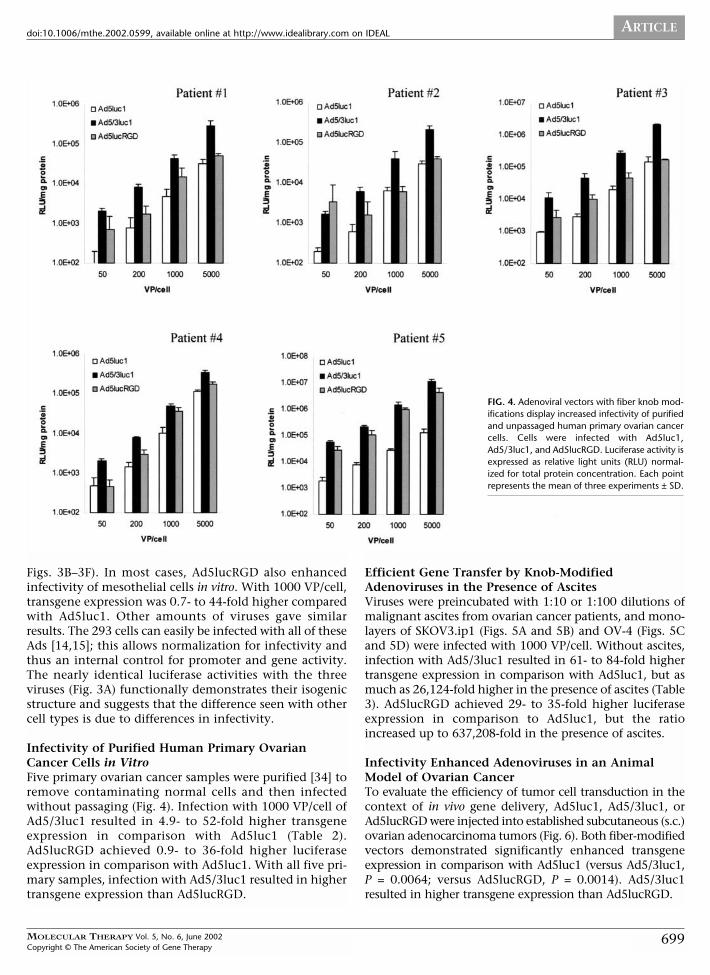
FIG. 4. Adenoviral vectors with fiber knob mod-ifications display increased infectivity of purifiedand unpassaged human primary ovarian cancercells. Cells were infected with Ad5luc1,Ad5/3luc1, and Ad5lucRGD. Luciferase activity isexpressed as relative light units (RLU) normal-ized for total protein concentration. Each pointrepresents the mean of three experiments ± SD.
Figs. 3B–3F). In most cases, Ad5lucRGD also enhancedinfectivity of mesothelial cells in vitro. With 1000 VP/cell,transgene expression was 0.7- to 44-fold higher comparedwith Ad5luc1. Other amounts of viruses gave similarresults. The 293 cells can easily be infected with all of theseAds [14,15]; this allows normalization for infectivity andthus an internal control for promoter and gene activity.The nearly identical luciferase activities with the threeviruses (Fig. 3A) functionally demonstrates their isogenicstructure and suggests that the difference seen with othercell types is due to differences in infectivity.
Infectivity of Purified Human Primary OvarianCancer Cells in VitroFive primary ovarian cancer samples were purified [34] toremove contaminating normal cells and then infectedwithout passaging (Fig. 4). Infection with 1000 VP/cell ofAd5/3luc1 resulted in 4.9- to 52-fold higher transgeneexpression in comparison with Ad5luc1 (Table 2).Ad5lucRGD achieved 0.9- to 36-fold higher luciferaseexpression in comparison with Ad5luc1. With all five pri-mary samples, infection with Ad5/3luc1 resulted in highertransgene expression than Ad5lucRGD.
MOLECULAR THERAPY Vol. 5, No. 6, June 2002Copyright © The American Society of Gene Therapy
Efficient Gene Transfer by Knob-ModifiedAdenoviruses in the Presence of AscitesViruses were preincubated with 1:10 or 1:100 dilutions ofmalignant ascites from ovarian cancer patients, and mono-layers of SKOV3.ip1 (Figs. 5A and 5B) and OV-4 (Figs. 5Cand 5D) were infected with 1000 VP/cell. Without ascites,infection with Ad5/3luc1 resulted in 61- to 84-fold highertransgene expression in comparison with Ad5luc1, but asmuch as 26,124-fold higher in the presence of ascites (Table3). Ad5lucRGD achieved 29- to 35-fold higher luciferaseexpression in comparison to Ad5luc1, but the ratioincreased up to 637,208-fold in the presence of ascites.
Infectivity Enhanced Adenoviruses in an AnimalModel of Ovarian CancerTo evaluate the efficiency of tumor cell transduction in thecontext of in vivo gene delivery, Ad5luc1, Ad5/3luc1, orAd5lucRGD were injected into established subcutaneous (s.c.)ovarian adenocarcinoma tumors (Fig. 6). Both fiber-modifiedvectors demonstrated significantly enhanced transgeneexpression in comparison with Ad5luc1 (versus Ad5/3luc1, P = 0.0064; versus Ad5lucRGD, P = 0.0014). Ad5/3luc1resulted in higher transgene expression than Ad5lucRGD.
699
ARTICLE doi:10.1006/mthe.2002.0599, available online at http://www.idealibrary.com on IDEAL
TABLE 2: Tropism-modified adenovirus vectors enhance infectivity of purified human primary ovarian cancer cellsa
Ratiob Patient 1 Patient 2 Patient 3 Patient 4 Patient 5 293c
Ad5/3luc1:Ad5luc1 9.2 6.2 14 4.9 52 0.8
Ad5lucRGD:Ad5luc1 3.1 0.9 2.4 3.7 36 0.8
Ad5/3luc1:Ad5lucRGD 2.9 6.6 5.8 1.3 1.4 1.0
aInfection at 1000 VP/cell of Ad5luc1, Ad5/3luc1, and Ad5lucRGD.bRatio of luciferase activity expressed as relative light units normalized for total protein concentration.c293 human embryonal kidney cells were included as a nonmalignant control.
DISCUSSION
Variable expression of CAR on human primary cancer cellsis a major limitation of cancer gene therapy with Ad5. Incontrast, many normal epithelial cells express CAR, andthus untargeted Ad5 may result in high transduction ofnontarget tissues. The purpose of this study was to evalu-ate two tropism-modified viruses, Ad5lucRGD andAd5/3luc1, for their capacity to improve transduction ofthe most stringent available preclinical ovarian cancer sub-strates. Also, to obtain preclinical safety data, we analyzedbiodistribution, liver toxicity,
and blood clearance.In mice, the liver is the majororgan responsible for Ad clear-ance [35–37] and could thereforebe an important organ in regardto potential clinical toxicity inhumans. Kupffer cells have animportant role in clearing Adfrom the blood, but their uptakecan be saturated, leading to a non-linear dose effect at ~ 2 � 1010 VPin mice [37]. Only after saturationis achieved are hepatocytes andother tissues transduced effec-tively [37]. To evaluate immedi-ately and early liver toxicity, weinjected 5 � 1010 VP i.v. and ana-lyzed serum transaminases, liver
FIG. 5. Adenoviral vectors with fiber knobmodifications display increased infectivityof human ovarian adenocarcinoma cells inthe presence of malignant ascites contain-ing neutralizing antibodies. Viruses werepreincubated in a 1:10 (A and C) or 1:100(B and D) dilution of malignant ascites fromovarian cancer patients, and monolayers ofSKOV3.ip1 (A and B) and OV-4 (C and D)were infected with 1000 VP/cell of Ad5luc1,Ad5/3luc1, and Ad5lucRGD. Luciferaseactivity is expressed as relative light units(RLU) normalized for total protein concen-tration. Each point represents the mean ofthree experiments ± SD.
700
A B
C D
M
histopathology, and transgeneexpression at 72 hours (Fig. 1). Ingeneral, only minor differenceswere seen, suggesting that thetoxicity of tropism-modifiedviruses does not significantly dif-fer from Ad5. Ad5 has beenadministered intravenously,intraperitoneally, intraarterially,and intratumorally in large dosesin clinical trials without signifi-cant toxicity [3]. Thus, toxicitydict a good safety profile for these
similar to Ad5 could pretropism-modified viruses. Histopathologic analysisresulted in fewer findings with Ad5/3luc1 (Table 1).Transgene expression in the livers was slightly lower afterinjection with Ad5lucRGD and Ad5/3luc1 in comparisonwith Ad5luc1 (P = 0.0472 and not significant, respectively),which probably reflects differences in functional titer.
Viruses and other foreign particles in the blood arecleared mostly by Kupffer cells. Furthermore, the bloodclearance of Ad is effective, with a half-life of < 2 minutes
OLECULAR THERAPY Vol. 5, No. 6, June 2002Copyright © The American Society of Gene Therapy
MOLECUL
Copyright ©
ARTICLEdoi:10.1006/mthe.2002.0599, available online at http://www.idealibrary.com on IDEAL
TABLE 3: Tropism-modified adenovirus vectors partly circumvent inhibitory factors in ascites fluida
Ratiob No ascites Ascites #6 Ascites #7 Ascites #8
1:10 1:100 1:10 1:100 1:10 1:100
SKOV3.ip1Ad5/3luc1:Ad5luc1 61 86 26,124 2,809 234 395 512Ad5lucRGD:Ad5luc1 35 353 637,208 2,892 160 1,157 5,770Ad5/3luc1:Ad5lucRGD 1.7 0.2 0.04 1.0 1.5 0.3 0.09
OV-4Ad5/3luc1:Ad5luc1 84 157 148 908 315 956 120Ad5lucRGD:Ad5luc1 29 53 3,781 1,677 167 125 1,256Ad5/3luc1:Ad5lucRGD 2.9 3.0 0.04 0.5 1.9 7.6 0.09
aInfection at 1000 VP/cell of Ad5luc1, Ad5/3luc1, and Ad5lucRGD preincubated with either growth medium (no ascites) or ascites from ovarian cancer patients.bRatio of luciferase activity expressed as relative light units normalized for total protein concentration. Background luciferase activity was subtracted andvalues ≤ 0 were set as 1.
after a single i.v. injection [38]. We compared the bloodclearance rates of the fiber-modified viruses to Ad5luc1(Fig. 1G) and found no differences. The clearance rate ofAd5luc1 was similar to what has been reported before [38].
To obtain preclinical data on the biodistribution of thefiber-modified viruses in the context of i.p. administra-tion, we conducted i.p. injection into tumor-bearing mice.When transgene expression was normalized for theamount of tissue analyzed, the highest transgene expres-sion was seen in the spleen, ovaries, peritoneum, and liver(Fig. 2A). In the liver, luciferase activity was higher withAd5/3luc1 in comparison with Ad5luc1, but the oppositewas true after i.v. injection (Fig. 1F). This probably resultedfrom the lower peritoneal uptake of Ad5/3luc1, subse-quently allowing increased passage of Ad5/3luc1 into theblood and eventually into the liver. We estimated the totaluptake of virus by organ, and the liver had the highesttotal transgene expression, followed by spleen, kidneys,and ovaries (Fig. 2B). These results are in accord with pre-vious reports describing liver as the main organ express-ing transgene in mice after Ad injection [39,40].
Quantitative PCR was carried out to detect virus copiesin the liver (Fig. 2C). In contrast to the transgene expres-sion data, fewer copies of Ad5/3luc1 were detected in theliver in comparison with Ad5luc1. These findings couldreflect differences in uptake by Kupffer cells versus hepa-tocyte transduction. Kupffer cells are generally thought tobe ineffective in transgene expression [37,41], althoughnot all reports agree [42]. Thus, if Ad5luc1 has higher rel-ative tropism for Kupffer cells in comparison withAd5/3luc1, the result could be less transgene expressionbut higher copy number. This is supported by the higherliver toxicity seen with Ad5luc1 (Fig. 1C). Kupffer cells arethe main cells mediating immunologic responses in theliver [36,41], and less uptake could result in less inflam-mation. Clearly, this needs to be investigated in moredetail in animals and, more importantly, in humans.Variables contributing to the findings could include cap-sid-dependent differences during post-entry steps. It is not
AR THERAPY Vol. 5, No. 6, June 2002 The American Society of Gene Therapy
well understood how Ad induces endosomal lysis andtransport of its DNA to the nucleus, but it seems likely thatthe fiber plays a role. Conceivably, the rate of degradationof Ad by innate immune mechanisms could also beaffected.
All ovarian cancer gene therapy trials published so farhave relied on i.p. administration of the agent. Thus, thenormal tissue that is in most immediate contact with theagent is the peritoneal lining. We analyzed the infectivityof two human established mesothelial cell lines and threefresh primary peritoneum samples (Fig. 3). In contrast tothe mouse in vivo data (Figs. 2A and 2B), the humanmesothelial substrates displayed mostly increased infec-tivity with Ad5/3luc1 and, in some cases, also withAd5lucRGD. For CAR, it has been shown that the local-ization of the receptor in the context of the three-dimen-sional structure is crucial in determining transduction [43].
FIG. 6. Retargeting adenovirus to the serotype 3 receptor (Ad5/3luc1) resultsin increased transgene expression in a human ovarian cancer xenograft model,when compared with infectivity enhancement with RGD-4C (Ad5lucRGD) orAd5luc1 (wild-type fiber). Subcutaneous Hey cell tumors were established innude mice followed by intratumoral injection of 2.5 � 109 VP/nodule on 2consecutive days. On the third day, tumors were harvested, and the luciferaseactivity was analyzed. Results are presented as relative light units (RLU) nor-malized for protein concentration, with each point representing an individualtumor. Bar, median; **P < 0.01 versus Ad5luc1.
701

ARTICLE doi:10.1006/mthe.2002.0599, available online at http://www.idealibrary.com on IDEAL
The situation could be similar for the receptor of serotype3. In the intact peritoneum, the receptor may not be acces-sible and thus infectivity is low (Figs. 2A and 2B), whereaswith the isolated cells, the receptor could be exposed.Alternatively, the process of enzymatic disaggregation ofthe mesothelial cells and initiation of culture could reducethe expression of CAR, and therefore increase the relativetransgene expression mediated by the fiber-modifiedviruses. Finally, it is possible that human and mousemesothelial cells display different expression patterns ofthe Ad3 receptor and/or CAR. In any case, it is unclear iftransduction of the peritoneum is likely to cause severeside effects to patients. In the ovarian cancer trials pub-lished thus far, doses of up to 7.5 � 1013 VP of untargetedvirus have been administered, and although abdominalpain was reported regularly, it was not dose-limiting [12].
An important recent revelation has been that primarytumor cells express highly variable and often low amountsof CAR [4,5,7,14–20]. Thus, it is crucial to analyze the moststringent available clinical substrates for reliable preclinicalestimation of efficacy. For this purpose, we have developeda purification method that typically yields a > 95% purepopulation of cancer cells [34]. We analyzed the purified pri-mary cells without passaging, which could help avoid con-founding due to genotypic and phenotypic changesinvolved in the clonal selection process. In this case, fivepatient samples were infected (Fig. 4). In comparison withAd5luc1, transgene expression increased 0.9- to 36-foldwhen samples were infected with Ad5lucRGD. This is inaccord with previous reports that have demonstrated thatthe RGD-4C modification increases the infectivity of pri-mary ovarian cancer cells [14,17,26]. However, it should benoted that the actual level of infectivity enhancement con-ferred by the RGD-4C motif can only be demonstrated whenthe comparison is between isogenic viruses, that is, virusescontaining identical expression cassettes, as were used inthis case and in one other study [26]. Ad5/3luc1 achieved5- to 50-fold higher transgene expression in comparison toAd5luc1 (Fig. 4), and in all cases, Ad5/3luc was superior toAd5lucRGD (Table 2). In an animal model of ovarian can-cer, infection with Ad5/3luc1 and Ad5lucRGD resulted inhigher transgene expression than with Ad5luc1. Xenograftscontain variable amounts of host stromal cells, whose recep-tor expression levels influence the infectivity of thexenograft as a whole. This and the high local concentrationof virus may have contributed to the relatively good infec-tivity seen with Ad5luc1. These results indicate that modi-fication of the Ad5 fiber with an RGD-4C motif in the HIloop or swapping the knob for a serotype 3 knob can leadto significant enhancements in transduction of purified pri-mary ovarian cancer cells and in vivo. Also, because the 5/3chimera was superior to the RGD-4C, it is the best availablegenetic retargeting moiety for ovarian cancer. Our previousdata suggest that transgene expression correlates closelywith binding and transduction as opposed to possible down-stream effects [15].
702
At diagnosis, most ovarian cancer patients have ascitesfluid. Previous studies have suggested partial escape ofRGD-4C-modified viruses from neutralizing anti-aden-ovirus antibodies present in malignant ascites [26,29].Here, we have shown that Ad5/3luc1 is also able to cir-cumvent neutralization (Fig. 5 and Table 3). In fact, in thepresence of ascites, gene transfer was improved > 3 ordersof magnitude in comparison with Ad5luc1. The level ofneutralizing anti-Ad3 antibodies in ascites is unknown,but the prevalence of total serum anti-Ad3 antibodiesseems to be comparable to that of anti-Ad5 [44]. However,in parallel to what has been suggested for RGD-4C-modi-fied Ads [26,29], the chimeric nature of the Ad5/3luc1 fibermay allow escape from many of the anti-Ad3-fiber neu-tralizing antibodies, as such antibodies are often confor-mation-sensitive [45].
In conclusion, we have shown that the mouse biodis-tribution, liver toxicity, and blood clearance of Ad5/3luc1and of Ad5lucRGD are not significantly different fromthose of Ad5luc1. Both Ad5/3luc1 and Ad5lucRGDincreased transgene expression in the most stringentavailable preclinical substrates, purified primary cancercells, in the presence of neutralizing antibodies and in ananimal model. These approaches could be useful forincreasing transduction in the context of mutation com-pensation, delivery of genes coding for prodrug convert-ing enzymes (molecular chemotherapy), or conditionallyreplicating viruses. Conditionally replicating viruses haverecently shown promise in clinical trials [3], albeit themost impressive result is in combination with chemother-apy [46]. Importantly, it has been clearly demonstratedthat the oncolytic potency of replicating agents is directlydetermined by the capability of infecting target cells [47].Thus, infectivity enhancement, such as described here,could help achieve higher clinical efficacy. Progress inclinical gene therapy of other diseases, such as cardio-vascular disease, hemophilia, and severe combinedimmunodeficiency syndrome has been dependent onimprovements in the vectors [48–50]. Clinical trials willultimately show if preclinical advances such as reportedhere can help achieve similar progress in the field of ovar-ian cancer gene therapy.
MATERIALS AND METHODS
Cells and tissues. Human mesothelial cell lines AG07086A and AG07090Bwere obtained from the Coriell Cell Repositories (Camden, NJ). The 293cells were purchased from Microbix (Toronto, ON, Canada). SKOV3.ip1,Hey, and OV-4 cells were obtained from Janet Price, Judy Wolf (both ofM.D. Anderson Cancer Center, Houston, TX), and Timothy J. Eberlein(Harvard Medical School, Boston, MA), respectively. All cell lines were cul-tured in recommended conditions.
Fresh malignant ascites fluid samples from patients with pathologicallyconfirmed ovarian adenocarcinoma were obtained from the University ofAlabama at Birmingham (UAB) Hospital. Cancer cells were purified by apreviously described immunomagnetic-based method [34]. Briefly, ovariancancer cells were initially bound with a mouse anti-TAG-72-antibody, andthen collected with magnetic beads coated with anti-mouse-IgG.
MOLECULAR THERAPY Vol. 5, No. 6, June 2002Copyright © The American Society of Gene Therapy

ARTICLEdoi:10.1006/mthe.2002.0599, available online at http://www.idealibrary.com on IDEAL
Primary mesothelial cells were obtained by the enzymatic disaggrega-tion of fresh peritoneal strips collected from female patients undergoingintraabdominal surgery with a strategy modified from previous reports[51]. The peritoneum strips were washed several times with PBS, and excessadipose tissue was removed. The sample was incubated with 15 ml of a solu-tion containing 0.125% (wt/vol) trypsin (Mediatech, Herndon, VA), 0.01%(wt/vol) EDTA (Fisher Scientific, Fair Lawn, NJ), and 0.1% glucose (Sigma,St Louis, MO) for 20 minutes at 37°C on a rocker. Then, the suspensionwas centrifuged at 50g for 5 minutes at 4�C. The supernatant was discarded,and the cell pellet was washed once with 5 ml of Ham’s F-12 supplementedwith 10% FBS, 2 mM L-glutamine, penicillin (100 IU/ml), streptomycin(100 �l/ml), and 10 ng/ml of recombinant human epidermal growth fac-tor (EGF) (Life Technologies, Grand Island, NY). The pellet was resuspendedin the same medium, and seeded in a 25-cm2 tissue culture flask. Half ofthe medium was changed 24 hours later, and thereafter fully replaced onceevery 3 days. Cells were cultured without EGF at least 48 hours beforeinfection to induce quiescence. For each sample, the cell type and puritywas confirmed by immunohistochemistry (monoclonal antibodies recog-nizing calretinin, cytokeratin 5/6, and vimentin). To exclude tumor-con-taminated mesothelial samples, H&E-stained sections of each piece of peri-toneum were reviewed by a pathologist.
Recombinant adenoviruses. The replication-incompetent Ad vectors havebeen described previously. Ad5luc1 [15], Ad5/3luc1 [15], and Ad5lucRGD[14] were propagated on 293 cells and purified on cesium chloride gradi-ents. The viral particle (VP) concentration was determined at 260 nm, andstandard plaque assay on 293 cells was done to determine infectious par-ticles. The ratios of VP:infectious particles were 5.24, 45.7, and 51.1 forAd5luc1, Ad5/3luc1, and Ad5lucRGD, respectively.
Ad-mediated gene transfer assays. Cells were infected for 30 minutes atroom temperature at 50, 200, 1000, and 5000 VP/cell by adding Ad5luc1,Ad5/3luc1, or Ad5RGDluc1 diluted in 200 �l of growth medium with 2%FBS. Cells were washed once, and complete medium was added. After 24hours of incubation at 37�C, luciferase assay was done (Luciferase AssaySystem, Promega, Madison, WI). The protein concentration was deter-mined using a Bio-Rad DC protein assay kit (Bio-Rad, Hercules, CA) toallow normalization of the gene expression data for the number of cells.Background luciferase activities were subtracted from the readings.
For the results shown in Figure 5, viruses were preincubated for 10minutes at room temperature with malignant ascites fluid diluted 1:10 or1:100 in growth medium with 2% FBS. Then, cells were infected at 1000VP/cell for 1 hour at 37�C, followed by incubation and luciferase assay asbefore.
Animals. Mice were obtained at 4–6 weeks of age and quarantined for atleast 1 week before the study. Mice were kept under pathogen-free condi-tions according to the American Association for Accreditation of LaboratoryAnimal Care guidelines. Animal protocols were reviewed and approved bythe Institutional Animal Care and Use Committee of UAB.
In vivo liver toxicity. Female C57BL/6 mice (Charles River Laboratories,Wilmington, MA; n = 6/virus, n = 5/Opti-MEM), aged 8–10 weeks, wereinjected i.v. through the lateral tail vein with 5 � 1010 VP of Ad5luc1,Ad5/3luc1, or Ad5lucRGD in a volume of 300 �l of Opti-MEM, or Opti-MEM only. After 3 days, small aliquots of blood (50 �l) were collected fromthe tail vein and allowed to clot. Serum samples were separated by cen-trifugation, and transaminases were analyzed using GO- & GP-Transaminase Kit 505 (Sigma). Then the mice were killed and pieces of thelivers fixed in 10% neutral buffered formalin. Serial paraffin-embedded sec-tions were taken and stained with H&E. Histopathology was scored blindedby two independent pathologists. A second liver sample was snap-frozen,and ground to a fine powder using a mortar and pestle cooled in a dryice–ethanol bath. Liver powders were lysed using Cell Culture Lysis Buffer(Promega) at room temperature for 20 minutes. Lysates were frozen once,then centrifuged at 10,000g for 15 minutes. Luciferase activity and proteincontent in the supernatant was measured as before. Mean backgroundluciferase activity was subtracted from the data.
Blood clearance. Female C57BL/6 mice (Charles River Laboratories; n =3/virus), 8–10 weeks old, were injected i.v. through the proximal lateral tailvein with 5 � 1010 VP of Ad5luc1, Ad5/3luc1, or Ad5lucRGD in a volume
MOLECULAR THERAPY Vol. 5, No. 6, June 2002Copyright © The American Society of Gene Therapy
of 300 �l of Opti-MEM. Blood samples (20 �l) were drawn from the tailvein into heparinized tubes at 1, 5, 10, 15, 30, 45, and 60 minutes postinfection. Clotting was prevented with heparin, and bleeding betweenintervals was suppressed with pressure. Samples were briefly centrifuged toseparate the plasma. Aliquots of 1 �l were used for triplicate infections of293 cells, seeded in 96-well plates on the day before at 15,000 cells/well in100 �l of growth medium with 5% FBS. After 36 hours of incubation,luciferase activity was measured as before.
Biodistribution. To study biodistribution, we injected 6-week-old CB17SCID mice (UAB Center for AIDS Research, CFAR SCID Mouse Core Facility;n = 5/group) i.p. with 1 � 107 SKOV3.ip1 cells. After 10 days, 5 � 1010 VPof Ad5luc1, Ad5/3luc1, Ad5lucRGD, or no virus were injected i.p. in 1 mlof Opti-MEM. After 48 hours, blood samples were collected by cardiacpuncture, blood cells were separated immediately by centrifugation, andthe pellet was frozen. Organs were harvested and representative sectionswere snap-frozen. Transgene expression was analyzed as before. Mean back-ground luciferase activity was subtracted from data. To be able to expressresults also as total RLU/organ, the total amount of protein in the organsof three uninjected mice was determined.
To quantitate the E4 copy number, liver DNA was purified using aDNeasy Tissue Kit (Qiagen, Valencia, CA). The primers used for amplify-ing the E4 gene were: forward (5�-GGAGTGCGCCGAGACAAC-3�), andreverse (5�-ACTACGTCCGGCGTTCCAT-3�), and the probe (5�-TGGCAT-GACACTACGACCAACACGATCT-3�). The mouse �-actin (an endogenouscontrol) gene was amplified with primers: forward (5�-CGAGCGGTTCC-GATGC-3�), and reverse (5�-TGGATGCCACAGGATTCCAT-3�), and detectedwith a probe (5�-AGGCTCTTTTCCAGCCTTCCTTCTTGG-3�). The PCR con-ditions were as described earlier [26]. For deduction of the actual copynumber, the sample fluorescence readings were compared with the stan-dard curve created by amplifying known amounts of DNA (108, 106, 104,and 102 copies). Normalization for the amount of total DNA was carriedout by comparing the PCR results with mouse �-actin primers. Mouse �-actin was amplified to generate a standard curve in a known amount oftotal DNA (200, 20, 2, and 0.2 ng).
Subcutaneous (s.c.) tumor xenograft model in nude mice. Subcutaneoustumors were established by injecting 107 Hey cells into both flanks of 6-week-old female athymic CD-1 nude mice (Charles River Laboratories; n =5/group). When the nodules reached 5 mm diameter, 2.5 � 109 VP/nod-ule of Ad5luc1, Ad5/3luc1, or Ad5RGDluc1, or no virus, were injected intra-tumorally in a 30-�l volume of Opti-MEM on 2 consecutive days (days 1and 2), the total dose being 5 � 109 VP/nodule. On the third day, the micewere killed, and the tumors harvested and snap-frozen. The tumors wereground, and luciferase activity analyzed as before. Mean backgroundluciferase activity was subtracted from data.
Statistics. Statistical differences among groups were assessed with theKruskal–Wallis test. When significant differences were detected, pairwisecomparisons between groups were done with the Wilcoxon two-sampletest.
ACKNOWLEDGMENTS
We thank Ramon Alemany (Institut Catala d’Oncologia, Barcelona, Spain) andPaul N. Reynolds (Division of Human Gene Therapy, University of Alabama atBirmingham) for helpful suggestions and discussions, and Bin Liu (Division ofHuman Gene Therapy, University of Alabama at Birmingham) for help with ani-mal experiments. This work was supported by NIH R01 grants (CA68245,CA83821, CA74242, CA86881, HL67962, CA90547-01), NCI CO97110, NIHSPORE grants (P50 CA89019, P50 CA83591), Deutsche ForschungsgemeinschaftGrants (BA2076/1-1, BA2076/1-2), the Damon Runyon–Walter Winchell CancerResearch Fund, the Sigrid Juselius Foundation, the Emil Aaltonen Foundation,the Maud Kuistila Foundation, and the Finnish Medical Foundation.
RECEIVED FOR PUBLICATION DECEMBER 12, 2001; ACCEPTED MARCH 13, 2002.
REFERENCES1. Russell, W. C. (2000). Update on adenovirus and its vectors. J. Gen. Virol. 81: 2573–2604.2. Hemminki, A., and Alvarez, R. D. (2000). Adenovirus in oncology: a viable option?
703

ARTICLE doi:10.1006/mthe.2002.0599, available online at http://www.idealibrary.com on IDEAL
Biodrugs 16: 77–87.3. Kirn, D. (2001). Clinical research results with dl1520 (Onyx-015), a replication-selective
adenovirus for the treatment of cancer: what have we learned? Gene Ther. 8: 89–98.4. Li, Y., et al. (1999). Loss of adenoviral receptor expression in human bladder cancer cells:
a potential impact on the efficacy of gene therapy. Cancer Res. 59: 325–330.5. Miller, C. R., et al. (1998). Differential susceptibility of primary and established human
glioma cells to adenovirus infection: targeting via the epidermal growth factor receptorachieves fiber receptor-independent gene transfer. Cancer Res. 58: 5738–5748.
6. Fechner, H., et al. (2000). Trans-complementation of vector replication versus Coxsackie-adenovirus-receptor overexpression to improve transgene expression in poorly permis-sive cancer cells. Gene Ther. 7: 1954–1968.
7. Cripe, T. P., et al. (2001). Fiber knob modifications overcome low, heterogeneous expres-sion of the coxsackievirus-adenovirus receptor that limits adenovirus gene transfer andoncolysis for human rhabdomyosarcoma cells. Cancer Res. 61: 2951–2960.
8. Hutchin, M. E., Pickles, R. J., and Yarbrough, W. G. (2000). Efficiency of adenovirus-medi-ated gene transfer to oropharyngeal epithelial cells correlates with cellular differentia-tion and human coxsackie and adenovirus receptor expression. Hum. Gene Ther. 11:2365–2375.
9. Nalbantoglu, J., et al. (2001). Muscle-specific overexpression of the adenovirus primaryreceptor car overcomes low efficiency of gene transfer to mature skeletal muscle. J. Virol.75: 4276–4282.
10. Alvarez, R. D., et al. (2000). A cancer gene therapy approach utilizing an anti-erbB-2single-chain antibody-encoding adenovirus (AD21): a phase I trial. Clin. Cancer Res. 6:3081–3087.
11. Alvarez, R. D., et al. (2000). Adenoviral-mediated suicide gene therapy for ovarian can-cer. Mol. Ther. 2: 524–530, doi: 10.1006/mte.2000.0194.
12. Buller, R. E., et al. (2001). Multi-dose intraperitoneal (IP) rad/p53 (SCH58500) genereplacement combined with chemotherapy in heavily pretreated recurrent ovarian can-cer is associated with prolonged survival. Proc. Am. Soc. Clin. Oncol. 20: 262a.
13. Hasenburg, A., et al. (2000). Thymidine kinase gene therapy with concomitant topote-can chemotherapy for recurrent ovarian cancer. Cancer Gene Ther. 7: 839–844.
14. Dmitriev, I., et al. (1998). An adenovirus vector with genetically modified fibers demon-strates expanded tropism via utilization of a coxsackievirus and adenovirus receptor-inde-pendent cell entry mechanism. J. Virol. 72: 9706–9713.
15. Kanerva, A., et al. (2002). Targeting adenovirus to the serotype 3 receptor increases genetransfer efficiency to ovarian cancer cells. Clin. Cancer Res. 8: 275–280.
16. Kelly, F. J., et al. (2000). Selectivity of TAG-72-targeted adenovirus gene transfer to pri-mary ovarian carcinoma cells versus autologous mesothelial cells in vitro. Clin. Cancer Res.6: 4323–4333.
17. Vanderkwaak, T. J., et al. (1999). An advanced generation of adenoviral vectors selec-tively enhances gene transfer for ovarian cancer gene therapy approaches. Gynecol.Oncol. 74: 227–234.
18. Khuu, H. (1999). Detection of coxsackie-adenovirus receptor (CAR) immunoreactivity inovarian tumors of epithelial derivation. Appl. Immunohistochem. Mol. Morphol. 7:266–270.
19. Hemmi, S., Geertsen, R., Mezzacasa, A., Peter, I., and Dummer, R. (1998). The presenceof human coxsackievirus and adenovirus receptor is associated with efficient adenovirus-mediated transgene expression in human melanoma cell cultures. Hum. Gene Ther. 9:2363–2373.
20. Kasono, K., et al. (1999). Selective gene delivery to head and neck cancer cells via anintegrin targeted adenoviral vector. Clin. Cancer Res. 5: 2571– 2579.
21. Tomko, R. P., Xu, R., and Philipson, L. (1997). HCAR and MCAR: the human and mousecellular receptors for subgroup C adenoviruses and group B coxsackieviruses. Proc. Natl.Acad. Sci. USA 94: 3352– 3356.
22. Honda, T., et al. (2000). The coxsackievirus-adenovirus receptor protein as a cell adhe-sion molecule in the developing mouse brain. Mol. Brain Res. 77: 19– 28.
23. Okegawa, T., et al. (2001). The mechanism of the growth-inhibitory effect of coxsackieand adenovirus receptor (CAR) on human bladder cancer: a functional analysis of CARprotein structure. Cancer Res. 61: 6592–6600.
24. Cohen, C. J., et al. (2001). The coxsackievirus and adenovirus receptor is a transmem-brane component of the tight junction. Proc. Natl. Acad. Sci. USA 98: 15191–15196.
25. Wickham, T. J., Mathias, P., Cheresh, D. A., and Nemerow, G. R. (1993). Integrins �v�3and �v�5 promote adenovirus internalization but not virus attachment. Cell 73: 309–319.
26. Hemminki, A., et al. (2001). An adenovirus with enhanced infectivity mediates molecu-lar chemotherapy of ovarian cancer cells and allows imaging of gene expression. Mol.
704
Ther. 4: 223–231, doi: 10.1006/mthe.2001.0446.27. Arap, W., Pasqualini, R., and Ruoslahti, E. (1998). Cancer treatment by targeted drug
delivery to tumor vasculature in a mouse model. Science 279: 377–380.28. US Department of Health and Human Services, NIH, Recombinant DNA Advisory
Committee (2001). Minutes of meeting, December 13 & 15, 2000. Hum. Gene Ther.12: 1573–1575.
29. Blackwell, J. L., et al. (2000). Using a tropism-modified adenoviral vector to circumventinhibitory factors in ascites fluid. Hum. Gene Ther. 11: 1657–1669.
30. Havenga, M. J., et al. (2001). Improved adenovirus vectors for infection of cardiovascu-lar tissues. J. Virol. 75: 3335–3342.
31. Gall, J., Kass-Eisler, A., Leinwand, L., and Falck-Pedersen, E. (1996). Adenovirus type 5and 7 capsid chimera: fiber replacement alters receptor tropism without affecting pri-mary immune neutralization epitopes. J. Virol. 70: 2116–2123.
32. Knaan-Shanzer, S., et al. (2001). Highly efficient targeted transduction of undifferenti-ated human hematopoietic cells by adenoviral vectors displaying fiber knobs of subgroupB. Hum. Gene Ther. 12: 1989–2005.
33. Stevenson, S. C., Rollence, M., White, B., Weaver, L., and McClelland, A. (1995). Humanadenovirus serotypes 3 and 5 bind to two different cellular receptors via the fiber headdomain. J. Virol. 69: 2850–2857.
34. Barker, S. D., et al. (2001). An immunomagnetic-based method for the purification ofovarian cancer cells from patient-derived ascites. Gynecol. Oncol. 82: 57–63.
35. Lieber, A., et al. (1997). The role of Kupffer cell activation and viral gene expression inearly liver toxicity after infusion of recombinant adenovirus vectors. J. Virol. 71:8798–8807.
36. Zhang, Y., et al. (2001). Acute cytokine response to systemic adenoviral vectors in miceis mediated by dendritic cells and macrophages. Mol. Ther. 3: 697–707, doi:10.1006/mthe.2001.0329.
37. Tao, N., et al. (2001). Sequestration of adenoviral vector by Kupffer cells leads to a non-linear dose response of transduction in liver. Mol. Ther. 3: 28–35, doi:10.1006/mthe.2000.0227.
38. Alemany, R., Suzuki, K., and Curiel, D. T. (2000). Blood clearance rates of adenovirustype 5 in mice. J. Gen. Virol. 81 Pt 11: 2605–2609.
39. Wood, M., et al. (1999). Biodistribution of an adenoviral vector carrying the luciferasereporter gene following intravesicle or intravenous administration to a mouse. CancerGene Ther. 6: 367–372.
40. Reynolds, P., Dmitriev, I., and Curiel, D. (1999). Insertion of an RGD motif into the HIloop of adenovirus fiber protein alters the distribution of transgene expression of thesystemically administered vector. Gene Ther. 6: 1336–1339.
41. Wolff, G., et al. (1997). Enhancement of in vivo adenovirus-mediated gene transfer andexpression by prior depletion of tissue macrophages in the target organ. J. Virol. 71:624–629.
42. Wheeler, M. D., Yamashina, S., Froh, M., Rusyn, I., and Thurman, R. G. (2001). Adenoviralgene delivery can inactivate Kupffer cells: role of oxidants in NF-κB activation andcytokine production. J. Leukoc. Biol. 69: 622–630.
43. Walters, R. W., et al. (1999). Basolateral localization of fiber receptors limits adenovirusinfection from the apical surface of airway epithelia. J. Biol. Chem. 274: 10219–10226.
44. Vogels, R., et al. (2000). Improved adenoviral vectors for gene therapy and vaccination.J. Gene Med. 2: S51.
45. Gahery-Segard, H., et al. (1998). Immune response to recombinant capsid proteins ofadenovirus in humans: antifiber and anti-penton base antibodies have a synergistic effecton neutralizing activity. J. Virol. 72: 2388–2397.
46. Khuri, F. R., et al. (2000). A controlled trial of intratumoral ONYX-015, a selectively repli-cating adenovirus, in combination with cisplatin and 5-fluorouracil in patients with recur-rent head and neck cancer. Nat. Med. 6: 879–885.
47. Douglas, J. T., Kim, M., Sumerel, L. A., Carey, D. E., and Curiel, D. T. (2001). Efficientoncolysis by a replicating adenovirus (ad) in vivo is critically dependent on tumor expres-sion of primary ad receptors. Cancer Res. 61: 813–817.
48. Isner, J. M., and Asahara, T. (1999). Angiogenesis and vasculogenesis as therapeuticstrategies for postnatal neovascularization. J. Clin. Invest. 103: 1231–1236.
49. Cavazzana-Calvo, M., et al. (2000). Gene therapy of human severe combined immun-odeficiency (SCID)-X1 disease. Science 288: 669–672.
50. Kay, M. A., et al. (2000). Evidence for gene transfer and expression of Factor IX inhaemophilia B patients treated with an AAV vector. Nat. Genet. 24: 257–261.
51. Casado, E., et al. (2001). Strategies to accomplish targeted expression of transgenes inovarian cancer for molecular therapeutic applications. Clin. Cancer Res. 7: 2496–2504.
MOLECULAR THERAPY Vol. 5, No. 6, June 2002Copyright © The American Society of Gene Therapy
Copyright © 2022 FDOKUMEN




















