
Gene expression profile for schizophrenia: discrete neuron transcription patterns in the entorhinal...
10
Gene Expression Profile for Schizophrenia Discrete Neuron Transcription Patterns in the Entorhinal Cortex Scott E. Hemby, PhD; Stephen D. Ginsberg, PhD; Brian Brunk, PhD; Steven E. Arnold, MD; John Q. Trojanowski, MD, PhD; James H. Eberwine, PhD Background: Several lines of evidence indicate the al- tered function of the temporal lobe, including the hip- pocampus and entorhinal cortex (EC), is associated with schizophrenia. We used single-cell gene expression tech- nologies to assess coordinate changes in the expression of multiple genes, including neuronal signaling and syn- aptic-related markers in EC layer II stellate neurons. Methods: We used a single-neuron microdissection tech- nique coupled with linear antisense RNA amplification and high density/candidate gene arrays to assess coordi- nate changes in gene expression. The expression and rela- tive abundance of more than 18 000 messenger RNAs were assessed from EC layer II stellate neurons from postmor- tem samples of schizophrenic and age-matched control brains. Results of this initial screen were used to per- form a more specific secondary messenger RNA screen for each subject. Results: Data disclosed marked differences in expres- sion of various G-protein–coupled receptor-signaling transcripts, glutamate receptor subunits, synaptic pro- teins, and other transcripts. Results of secondary screen- ing showed significant decreases in levels of G-protein subunit i1, glutamate receptor 3, N-methyl-D-aspartate receptor 1, synaptophysin, and sensory nerve action potentials 23 and 25 in the stellate neurons of schizo- phrenic patients. We observed down-regulation of phos- pholemman (a phosphoprotein associated with anion channel formation) messenger RNA and protein levels in layer II/III stellate neurons in the population with schizophrenia. Conclusions: These results provide a preliminary ex- pression profile of schizophrenia in defined neuronal populations. Understanding the coordinated involve- ment of multiple genes in human disease provides in- sight into the molecular basis of the disease and offers new targets for pharmacotherapeutic intervention. Arch Gen Psychiatry. 2002;59:631-640 S CHIZOPHRENIA IS a chronic, de- bilitating psychiatric illness affecting approximately 1% of the general population. Clini- cal manifestations appear dur- ing late adolescence to early adulthood. Characteristic features of schizophrenia in- clude a mixture of positive (distortions of inferential thought, perception, language/ communication, and behavioral monitor- ing) and negative (blunted affect, alogia, and avolition) symptoms. 1 The temporal lobe, including the hippocampus, subiculum, and entorhinal cortex (EC), is a primary brain region associated with schizophrenia. The EC is integral to the function of the hip- pocampus, regulating the interaction of the hippocampus with other brain regions. Dis- ruption of neuronal functioning in this re- gion could affect information processing be- tween the hippocampus and various cortical areas. Dysregulation of temporal lobe func- tion is associated with symptoms that are similar to those found in individuals diag- nosed as having schizophrenia. For ex- ample, results of functional neuroimaging studies and neuropsychological assess- ment of patients with schizophrenia re- port significant deficits in temporal lobe function. 2 Results of most structural imag- ing studies in schizophrenia indicate a slight but significant reduction in hippocampal volume, 3-11 although other studies failed to observe these differences. 12-14 A relative pau- city of neurodegeneration, cell death, or gliosis is observed in temporal lobe struc- tures in schizophrenic brains. 15,16 Although several abnormalities have been identified in the brains of schizo- phrenic patients, alterations in neuronal or- ganization and connectivity in the tempo- ral lobe represent a subtle neuropathologic feature of the disease. 17 Several studies have reported decreased abundance of synaptic protein messenger RNAs (mRNAs) and pro- tein levels in schizophrenic patients, 18-24 ORIGINAL ARTICLE Author affiliations are listed at the end of this article. (REPRINTED) ARCH GEN PSYCHIATRY/ VOL 59, JULY 2002 WWW.ARCHGENPSYCHIATRY.COM 631 ©2002 American Medical Association. All rights reserved.
Transcript of Gene expression profile for schizophrenia: discrete neuron transcription patterns in the entorhinal...

Gene Expression Profile for Schizophrenia
Discrete Neuron Transcription Patterns in the Entorhinal Cortex
Scott E. Hemby, PhD; Stephen D. Ginsberg, PhD; Brian Brunk, PhD; Steven E. Arnold, MD;John Q. Trojanowski, MD, PhD; James H. Eberwine, PhD
Background: Several lines of evidence indicate the al-tered function of the temporal lobe, including the hip-pocampus and entorhinal cortex (EC), is associated withschizophrenia. We used single-cell gene expression tech-nologies to assess coordinate changes in the expressionof multiple genes, including neuronal signaling and syn-aptic-related markers in EC layer II stellate neurons.
Methods: We used a single-neuron microdissection tech-nique coupled with linear antisense RNA amplificationand high density/candidate gene arrays to assess coordi-nate changes in gene expression. The expression and rela-tive abundance of more than 18000 messenger RNAs wereassessed from EC layer II stellate neurons from postmor-tem samples of schizophrenic and age-matched controlbrains. Results of this initial screen were used to per-form a more specific secondary messenger RNA screenfor each subject.
Results: Data disclosed marked differences in expres-sion of various G-protein–coupled receptor-signaling
transcripts, glutamate receptor subunits, synaptic pro-teins, and other transcripts. Results of secondary screen-ing showed significant decreases in levels of G-proteinsubunit i�1, glutamate receptor 3, N-methyl-D-aspartatereceptor 1, synaptophysin, and sensory nerve actionpotentials 23 and 25 in the stellate neurons of schizo-phrenic patients. We observed down-regulation of phos-pholemman (a phosphoprotein associated with anionchannel formation) messenger RNA and protein levelsin layer II/III stellate neurons in the population withschizophrenia.
Conclusions: These results provide a preliminary ex-pression profile of schizophrenia in defined neuronalpopulations. Understanding the coordinated involve-ment of multiple genes in human disease provides in-sight into the molecular basis of the disease and offersnew targets for pharmacotherapeutic intervention.
Arch Gen Psychiatry. 2002;59:631-640
S CHIZOPHRENIA IS achronic,de-bilitating psychiatric illnessaffecting approximately 1% ofthe general population. Clini-cal manifestations appear dur-
ing late adolescence to early adulthood.Characteristic features of schizophrenia in-clude a mixture of positive (distortions ofinferential thought, perception, language/communication, and behavioral monitor-ing)andnegative(bluntedaffect, alogia, andavolition) symptoms.1 The temporal lobe,includingthehippocampus,subiculum,andentorhinal cortex (EC), is a primary brainregion associated with schizophrenia. TheEC is integral to the function of the hip-pocampus, regulating the interaction of thehippocampuswith otherbrain regions. Dis-ruption of neuronal functioning in this re-gioncouldaffect informationprocessingbe-tweenthehippocampusandvariouscorticalareas. Dysregulation of temporal lobe func-tion is associated with symptoms that are
similar to those found in individuals diag-nosed as having schizophrenia. For ex-ample, results of functional neuroimagingstudies and neuropsychological assess-ment of patients with schizophrenia re-port significant deficits in temporal lobefunction.2 Results of most structural imag-ingstudies inschizophrenia indicatea slightbut significant reduction in hippocampalvolume,3-11 although other studies failed toobserve thesedifferences.12-14 Arelativepau-city of neurodegeneration, cell death, orgliosis is observed in temporal lobe struc-tures in schizophrenic brains.15,16
Although several abnormalities havebeen identified in the brains of schizo-phrenic patients, alterations in neuronal or-ganization and connectivity in the tempo-ral lobe represent a subtle neuropathologicfeature of the disease.17 Several studies havereported decreased abundance of synapticprotein messenger RNAs (mRNAs) and pro-tein levels in schizophrenic patients,18-24
ORIGINAL ARTICLE
Author affiliations are listedat the end of this article.
(REPRINTED) ARCH GEN PSYCHIATRY/ VOL 59, JULY 2002 WWW.ARCHGENPSYCHIATRY.COM631
©2002 American Medical Association. All rights reserved.

indicating decreased synaptic density in this region andother possible alterations in synaptic circuitry. In addi-tion, catecholaminergic and glutamatergic signaling ab-normalities have been reported in the temporal lobe ofschizophrenic patients, suggesting alterations in the struc-ture and connectivity of this region.25-27
Within the temporal lobe, EC layer II stellate neu-rons exhibit alterations, including aberrant cytoarchitec-tural arrangement,28-30 smaller neuron size with normal neu-ron density,31 and decreased expression of the microtubule-associated protein 2.32 The EC layer II stellate neuronsconstitute an integral component of the conduit throughwhich information flows to the hippocampus, which helpsto regulate cortical-hippocampal-subcortical interac-
tions.33 Disruption of the functional integrity of these neu-rons may contribute to the aberrant behaviors associatedwith schizophrenia. The strategic location of EC layer IIstellate neurons and the previously identified biologicalcorrelates in these neurons make them an excellent can-didate for probing disease-related differences in gene ex-pression associated with schizophrenia.
Although several studies have provided insight intothe roles of particular genes, assessments have been lim-ited to 1 or a few transcripts; however, the multigenic na-ture of schizophrenia is probably due to the coordinatedysregulation of several genes.34 Recently, Mirnics et al35
used complementary DNA (cDNA) microarray technol-ogy to assess alterations in the expression of multiple genes
SUBJECTS AND METHODS
SUBJECTS
Brains from 8 patients who underwent long-term hospi-talization for schizophrenia and 9 age-matched neurologi-cally normal controls were used. Postmortem brain tissuefrom schizophrenic patients was obtained from the estab-lished brain collection of the Mental Health Clinical Re-search Center on Schizophrenia at the University of Penn-sylvania, Philadelphia (Table 1). Control tissue wasobtained via the Center for Neurodegenerative Disease Re-search at the University of Pennsylvania. Controls had nohistory of neurological or major psychiatric illness. We per-formed gross and microscopic diagnostic neuropatho-logic examinations, which included examination of mul-tiple cortical and subcortical regions, in all cases, and noneuropathologic abnormalities relevant to mental statuswere found. Schizophrenic subjects were elderly, “poor-outcome” patients who were participants in clinicopatho-logical studies at the University of Pennsylvania School ofMedicine in collaboration with 8 state hospitals in easternand central Pennsylvania. All patients were prospectivelyaccrued, underwent clinical interviews and assessments, andwere diagnosed according to DSM-IV criteria1 by researchpsychiatrists of the Mental Health Clinical Research Cen-ter.36 In general, clinical features included prominent nega-tive symptoms, relatively mild positive symptoms, mod-erate to severe cognitive dysfunction, and impairments inbasic self-care activities that warranted the long-term hos-pitalization of these patients. Antipsychotic treatment wascalculated as mean daily chlorpromazine equivalents fromdose intervals ranging from no greater than 72 hours, 1month, and 1 year before death.
IMMUNOCYTOCHEMISTRY
Tissue blocks, which included the middle portion of theEC, were dissected from the temporal lobe at autopsy,fixed in a solution of 70% ethanol/150mM sodium chlo-ride, embedded in paraffin, and cut in 6-mm sections asdescribed previously.37 A section from each individualwas stained with acridine orange to verify the presence ofnucleic acids in the tissue.38 To identify individual neu-rons for subsequent single-cell analysis, we performedimmunocytochemistry for the sections using a monoclo-nal antibody to nonphosphorylated neurofilament
(RmdO20).39 The antibody was labeled by means of theavidin-biotin method (ABC Vectastain; Vector Laborato-ries, Burlingame, Calif) and visualized by means of 3,3�-diamino benzidine.
SINGLE-CELL GENE EXPRESSION
After immunolabeling, an oligo(dT)-T7- primer/promoter(AAACGACGGCCAGTGAATTGTAATACGACTCACTAT-AGGCGC[T]24) was hybridized to poly A+ mRNA over-night in a solution consisting of 50% formamide/5� silversulfadiazine and chlorhexidine (SSC) at 25°C. Complimen-tary DNA was synthesized directly on the tissue sections(in situ transcription) using avian myeloblastosis virus re-verse transcriptase (0.5 U/µL) (Seikagaku America, Fal-mouth, Mass) in Tris buffer containing 6mM magnesiumchloride, 120mM potassium chloride, 7mM dithio-threitol, 250µM each of deoxyadenosine triphosphate, de-oxycytidine triphosphate, deoxyguanosine triphosphate, andthymidine triphosphate, and 0.12 U/µL of RNAsin.40 Sec-tions were incubated at 37°C for 90 minutes. Next, sec-tions were washed twice in 2� SSC, 25°C for 5 minutes,and stored at 4°C in 0.5� SSC up to 72 hours. After in situtranscription, layer II stellate neurons were dissected us-ing a micropipette attached to a micromanipulator undera high-power objective field (original magnification �40).Contents were collected in the pipette and emptied into1.5-mL microcentrifuge tubes for second-strand cDNA syn-thesis and subsequent antisense RNA (aRNA) amplifica-tion. The amplification and reamplification procedures aredescribed in detail elsewhere.41,42 Samples were pooled im-mediately before second-round amplification. We incor-porated phosphorus 33-labeled cytidine triphosphate in thepooled sample probes destined for hybridization of hu-man gene discovery arrays (GDA; Genome Systems, Inc,St Louis, Mo). The RNA samples were pooled for each sub-ject and labeled with phosphorus 32 cytidine triphos-phate for candidate array hybridization. Under optimal con-ditions, the first round of aRNA amplification results in anapproximately 1000-fold yield and an approximately 106-fold yield after 2 rounds. The aRNA procedure is a linearamplification process with minimal change in the relativeabundance of the mRNA population in the native state ofthe neuron. Messenger RNA can be reliably amplified fromsmall amounts of fixed tissue, including individual neu-rons and neuronal processes.37,41-43
For initial screening of the GDAs (�18000 genes),aRNA from 6 neurons from each of 4 schizophrenic
(REPRINTED) ARCH GEN PSYCHIATRY/ VOL 59, JULY 2002 WWW.ARCHGENPSYCHIATRY.COM632
©2002 American Medical Association. All rights reserved.

in the prefrontal cortex. Tissue samples were obtained frompostmortem brains of schizophrenic patients and age-matched control subjects. Regional assessments of geneexpression create an informative mosaic of expression levelchanges. Identifying specific molecular correlates of schizo-phrenia has been complicated by several factors, includ-ing clinical heterogeneity, cellular heterogeneity of corti-cal and subcortical regions, and the difficulty in assessingmultiple genes in discrete neuronal populations. Meth-ods of single-cell gene expression combined with cDNAmicroarray technology can overcome some of the ana-tomical and molecular limitations by assessing multipletranscripts in target neuronal populations. In the presentstudy, we report coordinate changes in the relative ex-
pression levels of more than 18000 genes in EC layer IIstellate neurons from schizophrenic patients and age-matched, nonpsychiatric control subjects using high-density cDNA microarrays.
RESULTS
DEMOGRAPHIC DATA
No significant difference was seen between the schizo-phrenic and control groups in age (t15=−1.14; P=.27), post-mortem interval (t14=0.68; P= .51), or brain weight(t14=−0.45; P=.66), indicating these factors do not con-tribute to the observed changes in differentially ex-
patients and 4 controls were pooled (eg, 24 neurons percondition for each array) before the second round of am-plification. Tissue from the schizophrenic patients was se-lected on the basis that none had been treated with anti-psychotic medication for at least 1 year before death.
CONSTRUCTION OF CANDIDATE ARRAYS
Candidate arrays were prepared on nylon membranes con-taining, but not limited to, dopamine receptors (eg, D1, D2,D4, D5, and DAT), G-protein subunits (i�1, i�2, �s, �z, �q,�o, �, �1, and �2), transcription factors (CREB, CRE�2,CREM, junB, and juD c-fos), glutamate receptor mRNAs(AMPA [GluR1-4], kainite [GluR5-7], and N-methyl-D-aspartate receptor 1 [NMDA R1]), and synaptic proteins(�-synuclein, synaptophysin 1 and 2, synaptobrevin, syn-aptobrevin 2, synaptogyrin 1a and 3, synaptic vesicle–associated protein [SNAP] 23 and 25, postsynaptic den-sity 95, and synaptotagmin VII). Inserts were amplified in96-well plates using polymerase chain reaction analysis withM13 forward and reverse primers under the following con-ditions: 95°C for 5 minutes (1 cycle); 95°C for 30 sec-onds, 52°C for 45 seconds, and 72°C for 2 minutes (40 cyclesof this combination); and 72°C for 10 minutes (1 cycle).After polymerase chain reaction analysis, aliquots under-went electrophoresis on a 1% agarose gel (1� Tris-borate–EDTA pH 8.0 and 0.05% ethidium bromide) at 5 V/cm, andthe polymerase chain reaction band size was verified. Gelimages were captured by means of a digital camera and ar-chived on a computer. We spotted 250 ng of each ampli-fied insert on a net nylon transfer membrane (HyBond XL;Amersham Pharmacia Biotech, Minneapolis, Minn) usinga 96-well dot-blot apparatus (Minifold I; Schleicher & Sch-uell, Inc, Baltimore, Md). The DNA was crosslinked to themembrane by means of UV radiation.
GDA AND CANDIDATE ARRAY HYBRIDIZATION
Arrays were hybridized for 24 hours at 44°C in a rotisseriehybridization oven (Hybaid, Boston, Mass) with the fol-lowing solution: 50% formamide, 5� SSC, 5� Denhardtsolution, 0.1% sodium dodecyl sulfate (SDS), 200 ng ofsheared salmon sperm, and 1.0mM sodium pyrophos-phate. After hybridization, membranes were washedsequentially with solutions consisting of 2� SSC/0.1%SDS, 0.5� SSC/0.1% SDS, and 0.1� SSC/0.1% SDS for 20minutes each at 44°C. We detected labeled hybridizedproducts using phosphoimager cassettes, and we analyzed
hybridization signal intensities using ImageQuant soft-ware (Amersham Pharmacia/Molecular Dynamics, MenloPark, Calif).
DATA ANALYSIS
The specific signal (minus background) of probe bound toeach clone is expressed as a ratio of the total hybridizationintensity of the array, thereby minimizing variations dueto differences in the specific activity of the probe and theabsolute quantity of probe present.43 Differential expres-sion of greater than 2-fold is accepted as above back-ground and relevant for further examination. Two-foldchanges are considered a conservative limit. Data from thecandidate gene arrays were analyzed by t test, and the nullhypothesis was rejected when P�.05.
RELATIONAL DATABASE
Data were imported into the RNA Abundance Database, anOracle relational database developed at the University of Penn-sylvania. The RNA Abundance Database is designed to cap-ture information on RNA abundance assays for any type ofhigh-throughput gene expression experiment, including mi-croarrays, macroarrays, and serial analysis of gene expres-sion tags. For each experiment, hybridization signal inten-sity for each data point was expressed as a percentage of thetotal intensity on the array. This enabled comparison of datagenerated under different conditions and across experimen-tal platforms. To identify genes by functional role or chro-mosomal location, queries were performed against the da-tabase of transcribed sequences (DoTS),44 a component ofthe Genomics Unified Schema relational database also de-veloped at the University of Pennsylvania and imple-mented in Oracle. The DoTS contains known and putativetranscripts from human and mouse tissues. Each transcripthas a consensus sequence assembled by computational analy-sis of the expressed sequence tag (EST) and known mRNAsequences available in the public databases. These DoTS tran-scripts were then annotated to assign such things as pre-dicted cellular roles, GO functions, and chromosomal loca-tions.44 The spots in the array experiments can be linked toDoTS transcripts through their respective EST sequences,allowing the assignment of cellular roles of 13510 and chro-mosomal location to 11591 clones. Data sets were selectedby means of SQL queries joining the DoTS and RNA Abun-dance Database, and scattergrams were generated usingSigmaPlot software (SPSS Science, Chicago, Ill).
(REPRINTED) ARCH GEN PSYCHIATRY/ VOL 59, JULY 2002 WWW.ARCHGENPSYCHIATRY.COM633
©2002 American Medical Association. All rights reserved.

pressed genes. The schizophrenic group included 5 womenand 3 men with an average±SEM age of 83±9.3 years,whereas the age of disease onset was 23.4±3.4 years. Theage-matched controls (average±SEM age, 77.7±12.2 years)consisted of 5 women and 4 men.
IMMUNOCYTOCHEMISTRY AND RNA RECOVERY
Examination of tissue sections after immunolabeling withRmdO20 disclosed a distinct laminar pattern of immuno-reactivity that was confined to the somatodendritic regionof neurons in layers II/III and V of the EC (Figure 1A-B).No distinct differences in the intensity or pattern were ap-parent between the groups. Immunostaining was used todelineate layer II stellate neurons for microdissection (Fig-ure 1C-D). As in previous studies, no apparent differencewas seen in mRNA recovery between the groups.45
GENE EXPRESSION
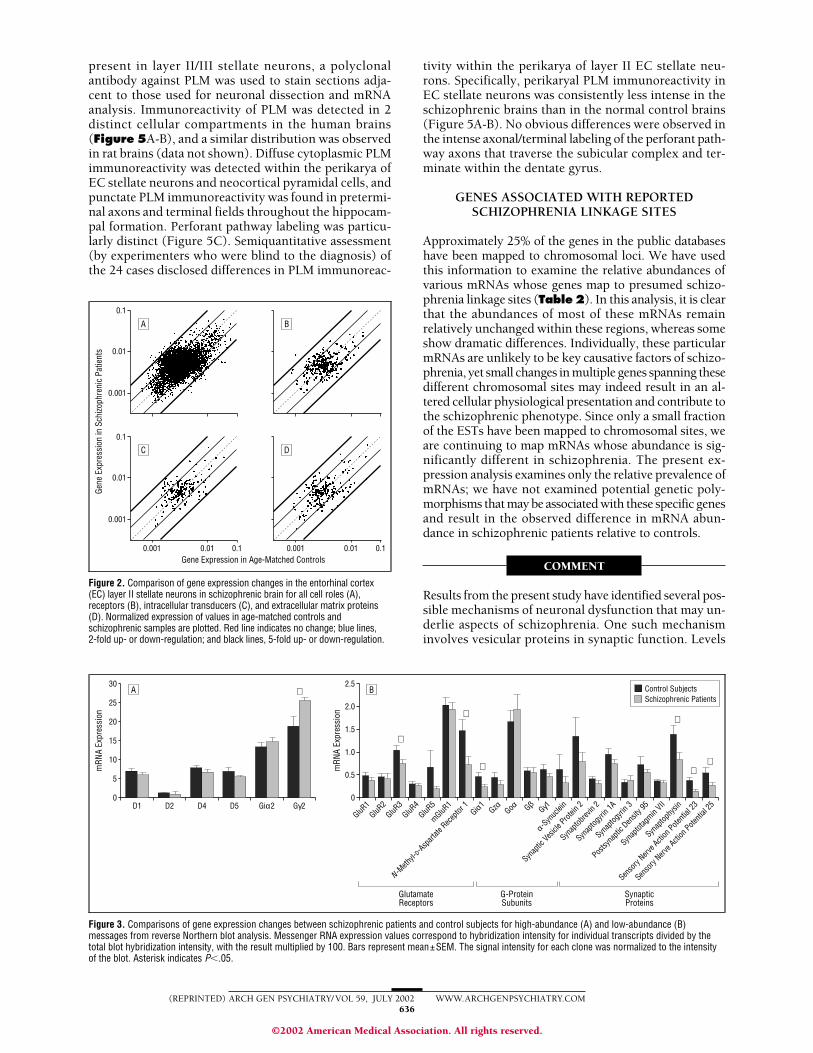
The GDA format contained 18240 genes, of which 2574(14%) were up-regulated more than 2-fold in the schizo-phrenic group and 1565 (9%) were down-regulated. Inaddition, we examined a subgroup of transcripts that en-code proteins (13510 mRNAs) of known function andare designated as all cell roles in the Institute for Ge-nomic Research database. Changes in gene expressionwere assessed by the degree of differential expression inspecific functional families encoding all cell roles, recep-tors (292 clones), intracellular transducers (169 clones,including G proteins and second-messenger systems), andextracellular matrix proteins (199 clones, including syn-aptic proteins). The subsets were selected because of thekey role members of each of these families play in cel-
lular functioning, not necessarily because of their sig-nificance in schizophrenia. For convenience, differ-ences in mRNA levels for these categories are shown inFigure 2. The complete expression profiles generatedin this study are available in Excel format via e-mail (avail-able at: [email protected]).
RECEPTORS
The absence of dopamine receptor subtypes on the GDAarrays necessitated inclusion of these clones of the custom-designed candidate arrays. No significant difference wasseen in mRNA abundance for D1, D2, D4, or D5 receptorsubunits between the schizophrenic and control groups(Figure 3A). Analysis showed an up-regulation in se-rotonin receptor mRNA (+3.0-fold). The �2-adrenergicreceptor mRNA was down-regulated 2.1-fold, a findingconsistent with the reduced �2-adrenergic receptor bind-ing in the limbic system of the schizophrenic brain.46 Sec-ondary screening of G-protein subunits that couple tomonoamine receptors disclosed a significant decrease inGi�1 subunit mRNA (t15=2.37; P=.03) and a significantincrease in G�2 subunit mRNA (t15=−2431; P=.03) lev-els in schizophrenic patients (Figure 3).
Several groups have reported alterations in �-ami-nobutyric acid and glutamate receptor protein and mRNAsubunits in the schizophrenic brain.26,27,47-55 Consistentwith these findings, we found a 4.2-fold increase in �-ami-nobutyric acid A�1 subunit mRNA in schizophrenic pa-tients. No significant differences were detected in NMDAR2A, GluR1, GluR2, or GluR6 on the GDA arrays, andnone were detected for GluR1, GluR4, and GluR5 on thecustom-designed arrays. However, GluR3 was found tobe significantly down-regulated on the GDA arrays (−2.2-
Table 1. Case Information*
Age, y Sex PMI, h Age of Onset, y Antipsychotic Dose†
Schizophrenic PatientsS1 81 F 13 26 NAS2 88 F 17.5 20 75S3 67 F 8.5 16 800S4 86 F 12 19 75S5 89 M 15 26 0S6 74 F 6.5 23 0S7 99 M 10.5 23 0S8 99 M 6.5 27 0Mean (SD) 83.9 (3.5) . . .‡ 11.2 (1.4) 23.4 (3.4) . . .
Nonpsychiatric controlsC1 81 F 8 . . . . . .C2 98 M 15 . . . . . .C3 85 F 11 . . . . . .C4 91 F 11.5 . . . . . .C5 72 F 4 . . . . . .C6 71 M 13 . . . . . .C7 76 F 13 . . . . . .C8 63 M 15 . . . . . .C9 62 F 24 . . . . . .Mean (SD) 77.7 (4.1) . . .§ 12.9 (2.1) . . . . . .
*PMI indicates postmortem interval; NA, not available; and ellipses, not applicable.†Indicates daily antipsychotic medication dose 1 month before death, expressed in chlorpromazine equivalents.‡Includes 3 male and 5 female patients.§Includes 3 male and 6 female controls.
(REPRINTED) ARCH GEN PSYCHIATRY/ VOL 59, JULY 2002 WWW.ARCHGENPSYCHIATRY.COM634
©2002 American Medical Association. All rights reserved.

fold) and confirmed using the custom-designed arrays(t15=2.18; P=.045). In addition, NMDA R1 was signifi-cantly down-regulated in the schizophrenic group(t15=2.55; P=.02; Figure 4B) using the custom arrays.
Cholinergic dysfunction has also been implicated inschizophrenia, including decreased nicotinic receptorbinding in the hippocampus56 and demonstration of a di-nucleotide polymorphism at chromosome 15q13-14, thesite of the �7 subunit of the nicotinic receptor.57 Extend-ing these reports, we found a 2.7-fold increase in expres-sion of the �7 subunit mRNA in EC stellate neurons inour schizophrenic population.
GENES ASSOCIATED WITH SYNAPTIC PROTEINS
Several synaptic protein mRNAs were differentially regu-lated between the schizophrenic and control groups, in-cluding down-regulation in schizophrenia of �-adaptin(−5.5-fold), synaptic vesicle amine transporter (−3.5-fold), synaptotagmin I (−3.1-fold), synaptotagmin IV(−2.5-fold), and SNAP 25 (−4.4-fold). An example of thedifferential hybridization intensity for SNAP 25 on a GDA
filter is provided in Figure 4. In addition, syntaxin mRNAwas up-regulated (+4.4-fold) in schizophrenic patients.Assessment of several synaptic protein mRNAs using thecandidate arrays showed significant decreases in synap-tophysin (t15=2.22; P=.04), SNAP 23 (t15=2.94; P=.01),and SNAP 25 (t15=2.09; P=.055) mRNA levels in schizo-phrenic patients (Figure 3B).
PHOSPHOLEMMAN EXPRESSION
Differential hybridization to the cDNAs encoding sev-eral ESTs was noted, in addition to genes of known func-tion. One of the most highly regulated ESTs corre-sponded to phospholemman (PLM), a phosphoproteininvolved in the formation and/or regulation of a chlo-ride anion channel. Expression levels of PLM mRNA insingle EC stellate neurons were lower in schizophrenicbrains than in those of matched controls (−4.5-fold). Wewere unable to perform secondary screening on PLMmRNA abundance because of the lack of clone in thehuman clone set (Emory Functional Genomics Facility,Atlanta, Ga). To determine whether PLM protein was
I
II
III
C D
A B
Figure 1. Immunoreactivity of a monoclonal antibody to nonphosphorylated neurofilament (RmdO20) in the entorhinal cortex (EC). Distribution and staining ofmedium-weight neurofilament immunoreactive stellate neurons from control brain tissue in layer II of the EC under low- (A; scale bar, 50 µm) and high-power(B; scale bar, 10 µm) magnification are seen. The arrow in panel A indicates the area of high-power magnification shown in panel B. Section immunolabeled withRmdO20 with a representative stellate neuron (C; scale bar, 25 µm) and after microdissection of the indicated neuron (D; scale bar, 25 µm).
(REPRINTED) ARCH GEN PSYCHIATRY/ VOL 59, JULY 2002 WWW.ARCHGENPSYCHIATRY.COM635
©2002 American Medical Association. All rights reserved.
present in layer II/III stellate neurons, a polyclonalantibody against PLM was used to stain sections adja-cent to those used for neuronal dissection and mRNAanalysis. Immunoreactivity of PLM was detected in 2distinct cellular compartments in the human brains(Figure 5A-B), and a similar distribution was observedin rat brains (data not shown). Diffuse cytoplasmic PLMimmunoreactivity was detected within the perikarya ofEC stellate neurons and neocortical pyramidal cells, andpunctate PLM immunoreactivity was found in pretermi-nal axons and terminal fields throughout the hippocam-pal formation. Perforant pathway labeling was particu-larly distinct (Figure 5C). Semiquantitative assessment(by experimenters who were blind to the diagnosis) ofthe 24 cases disclosed differences in PLM immunoreac-
tivity within the perikarya of layer II EC stellate neu-rons. Specifically, perikaryal PLM immunoreactivity inEC stellate neurons was consistently less intense in theschizophrenic brains than in the normal control brains(Figure 5A-B). No obvious differences were observed inthe intense axonal/terminal labeling of the perforant path-way axons that traverse the subicular complex and ter-minate within the dentate gyrus.
GENES ASSOCIATED WITH REPORTEDSCHIZOPHRENIA LINKAGE SITES
Approximately 25% of the genes in the public databaseshave been mapped to chromosomal loci. We have usedthis information to examine the relative abundances ofvarious mRNAs whose genes map to presumed schizo-phrenia linkage sites (Table 2). In this analysis, it is clearthat the abundances of most of these mRNAs remainrelatively unchanged within these regions, whereas someshow dramatic differences. Individually, these particularmRNAs are unlikely to be key causative factors of schizo-phrenia, yet small changes in multiple genes spanning thesedifferent chromosomal sites may indeed result in an al-tered cellular physiological presentation and contribute tothe schizophrenic phenotype. Since only a small fractionof the ESTs have been mapped to chromosomal sites, weare continuing to map mRNAs whose abundance is sig-nificantly different in schizophrenia. The present ex-pression analysis examines only the relative prevalence ofmRNAs; we have not examined potential genetic poly-morphisms that may be associated with these specific genesand result in the observed difference in mRNA abun-dance in schizophrenic patients relative to controls.
COMMENT
Results from the present study have identified several pos-sible mechanisms of neuronal dysfunction that may un-derlie aspects of schizophrenia. One such mechanisminvolves vesicular proteins in synaptic function. Levels
0.1
0.001
0.01
0.1
0.001
0.01
0.001 0.01 0.1
Gene
Exp
ress
ion
in S
chizo
phre
nic
Patie
nts
0.001 0.01 0.1Gene Expression in Age-Matched Controls
A B
C D
Figure 2. Comparison of gene expression changes in the entorhinal cortex(EC) layer II stellate neurons in schizophrenic brain for all cell roles (A),receptors (B), intracellular transducers (C), and extracellular matrix proteins(D). Normalized expression of values in age-matched controls andschizophrenic samples are plotted. Red line indicates no change; blue lines,2-fold up- or down-regulation; and black lines, 5-fold up- or down-regulation.
30
10
∗
∗
20
25
15
5
0D1 D2 D4 Giα2D5 Gγ2
mRN
A Ex
pres
sion
2.5
0.5
1.5
2.0
1.0
0
GluR1
GluR2
GluR3
GluR4
GluR5
mGluR1
N -Meth
yl-D-A
spart
ate Rec
eptor
1Giα1
Gzα Goα GβGγ1
α-Syn
uclei
n
Synap
tic Ves
icle P
rotein
2
Synap
tobrev
in 2
Synap
togyri
n 1A
Synap
togyri
n 3
Postsy
napti
c Den
sity 9
5
Synap
totag
min VII
Synap
tophy
sin
Senso
ry Nerv
e Acti
on Pote
ntial
23
Senso
ry Nerv
e Acti
on Pote
ntial
25
GlutamateReceptors
G-ProteinSubunits
SynapticProteins
mRN
A Ex
pres
sion
A B Control SubjectsSchizophrenic Patients
∗
∗
∗
∗∗
Figure 3. Comparisons of gene expression changes between schizophrenic patients and control subjects for high-abundance (A) and low-abundance (B)messages from reverse Northern blot analysis. Messenger RNA expression values correspond to hybridization intensity for individual transcripts divided by thetotal blot hybridization intensity, with the result multiplied by 100. Bars represent mean±SEM. The signal intensity for each clone was normalized to the intensityof the blot. Asterisk indicates P�.05.
(REPRINTED) ARCH GEN PSYCHIATRY/ VOL 59, JULY 2002 WWW.ARCHGENPSYCHIATRY.COM636
©2002 American Medical Association. All rights reserved.

of mRNAs encoding synaptic vesicle proteins (synpato-physin and synaptotagmin I and IV) and synaptic plasmamembrane proteins (SNAP 23 and SNAP 25) were foundto be significantly decreased in EC layer II stellate neu-rons of schizophrenic patients, whereas another plasmamembrane protein syntaxin was up-regulated greater than4-fold. The proteins encoded by these mRNAs serve dif-ferent functions at different functional steps in the syn-aptic vesicle cycle, and it is reasonable to conclude thatalterations in the levels of the proteins encoded by thesemRNAs may lead to decreased neurotransmitter releasefrom the layer II stellate neurons. For exocytosis to oc-cur, a trimeric core complex must be formed consistingof 2 synaptic plasma membrane proteins and 1 synapticvesicle protein.58 Decreased levels of SNAP 25 may pre-vent the establishment of the anchor complex for vesicu-lar docking to the plasma membrane. Furthermore, de-creased levels of synaptotagmin I and IV, which bind thecalcium2+ ion and possibly serve as a sensor for exocy-tosis,59 indicate another potential means of decreased neu-rotransmitter release. These findings are paralleled bystudies demonstrating decreased synaptic vesicle pro-tein mRNA and protein levels in the temporal cor-tex18,20-24 and other brain regions.22,35,60-64 The altered ex-pression of SNAP 25 and syntaxin are not likely due tolong-term antipsychotic treatment, since long-term halo-peridol decanoate administration in rodents does not affectSNAP 25 mRNA expression and decreases syntaxin andsynaptophysin mRNA expression.65,66 However, the ob-served decreases in synaptotagmin I and IV mRNA lev-els in schizophrenic patients may be attributable in partto the treatment history, since long-term haloperidol ad-ministration also decreases synaptotagmin mRNA lev-els,65 although extrapolations of these data to humansshould be made with caution.
Results of high-density array analysis indicate down-regulation of �2-adrenergic receptor mRNAs46 and up-regulation of the �-aminobutyric acid A�152-55 subunitand serotonin receptor mRNA, findings that are consis-tent with those of previous studies. No significant dif-ferences were observed for the dopamine receptor sub-type mRNAs in the present study. However, Gi�1 andG�2 subunit mRNA levels were significantly reduced andelevated, respectively, in the schizophrenic population,a finding consistent with Gi� immunoreactivity in the
temporal cortex of schizophrenic patients.67 Glutama-tergic dysfunction is yet another possible mechanism un-derlying the neuropathophysiology of schizophrenia, spe-
A B
Figure 4. Magnified portions of human gene discovery microarray imagesafter hybridization with control (A) and schizophrenic (B) samples. Redarrows indicate doublets representing sensory nerve action potential (SNAP)25. In the control samples, labeled antisense RNA hybridized to bothcomplementary DNAs corresponding to SNAP 25 with moderate intensity.In contrast, hybridization intensity was less intense in the schizophrenicsamples.
*
*
*
A
B
C
Figure 5. Phospholemman (PLM) immunoreactivity in the hippocampalformation. A polyclonal antibody against PLM protein was used to stainsections adjacent to those used for neuronal dissection and messenger RNAanalysis. Distribution and staining intensity of PLM-immunoreactive stellatecells in layer II of entorhinal cortex (EC) in a cognitively normal control brain(A; scale bar, 50 µm). The high-power inset shows diffuse cytoplasmic PLMimmunoreactivity throughout the somatic domain of stellate cells (scale bar,25 µm). The PLM immunoreactivity is less intense within the entorhinalcortex of a patient diagnosed as having schizophrenia (B; scale bar, 50 µm).The inset depicts, at higher power, less intense PLM immunoreactivity in thesomatic compartment compared with the control brain (scale bar, 25 µm).The PLM-immunoreactive preterminal axons and terminals are observedthroughout the hippocampal formation, including the perforant path (C; scalebar, 50 µm). Intense, punctate labeling of the perforant path is observedtraversing the subiculum, with some varicosities terminating in pericellularbasketlike arrangements. The inset depicts the area labeled by the asterisk onthe low-power image (scale bar, 10 µm).
(REPRINTED) ARCH GEN PSYCHIATRY/ VOL 59, JULY 2002 WWW.ARCHGENPSYCHIATRY.COM637
©2002 American Medical Association. All rights reserved.

cifically, the gene and protein expression of the ionotropicsubtypes in human postmortem tissue.47-51 For ex-ample, previous studies have demonstrated decreased ex-pression of GluR1 and GluR2 mRNAs in hippocampalsubfields27 and NMDA R1 mRNA in the temporal cor-tex.68 Extending these findings, NMDA R1 and GluR3were down-regulated in EC layer II stellate neurons inthe present study. Dysregulation in ionotropic gluta-mate receptors may have profound downstream effects,including alterations in excitatory neurotransmission andsubsequent cognitive and behavioral sequelae believedto be driven by glutamatergic circuitry.
In addition to genes known to be involved in syn-aptic function, array analysis led to the identification ofPLM mRNA in the EC layer II stellate neurons. Phos-pholemman is a phosphoprotein involved in the forma-tion and/or regulation of a chloride anion channel69 en-riched in cardiac and skeletal muscle, although resultsof Northern blot analysis have demonstrated moderatemRNA expression in total brain homogenates.70,71 Peri-karyal PLM immunoreactivity in EC stellate neurons wasconsistently less intense in the schizophrenic brains thanin the normal control brains. The observed EC stainingpattern is not selective to our brain collection popula-tion; it was replicated in EC tissue sections from 2 schizo-phrenic patients obtained from the Stanley FoundationBrain Bank, Bethesda, Md. Further studies are war-ranted to characterize the neuroanatomical distributionof PLM, to delineate the functional role of this proteinin the brain, and to further assess the contribution of PLMdown-regulation in schizophrenia.
Chromosomal mapping of genes that are altered inschizophrenia may provide insight into how the chro-mosomal abnormality is manifested in the symptomatol-ogy of schizophrenia. These genes may map directly achromosomal breakage, but more likely are adjacent geneswhose regulation is affected in schizophrenia. Such regu-latory differences may be associated with polymor-phisms in the promoter regions of these genes that, inturn, alter transcription rates leading to changes in mRNAabundance. Individual mRNAs are unlikely to be singu-lar causal factors for schizophrenia. However, small
changes in multiple genes spanning these different chro-mosomal loci may result in an altered cellular physiol-ogy, thus contributing to the schizophrenic phenotype.
A common confound in using human tissue for neu-ropathophysiological examinations lies in the clinical di-agnosis of the individual patient. In the present study, theuse of a prospective collection of brains from subjects whounderwent clinical assessment during life obviates this prob-lem. Since the pharmacological course of treatment forschizophrenia may influence gene expression, initial screen-ing of arrays used brain tissue from patients who had notreceived antipsychotic medication for at least 1 year be-fore death, followed by secondary screening of all subjectsin the sample population regardless of medication his-tory. The observed consistency in these hybridization pat-terns is likely due to the long-term treatment histories ofall subjects in the study. Nevertheless, the influence of medi-cation exposure on gene expression cannot be dis-counted. The postmortem interval was similar to or lessthan that of other studies and is unlikely to grossly influ-ence the molecular analysis presented herein.18-22,27,35,45,66
Nonetheless, the utility of an expression profile specificfor schizophrenia can be envisioned. For example, differ-entially expressed transcripts could serve as an addi-tional postmortem diagnostic tool. Application of similartechnologies to generate peripheral markers may enablemore rapid diagnosis and pharmacological intervention tai-lored to the patient’s specific symptoms. In addition, con-firmation of corresponding changes at the protein level mayprovide novel targets for drug discovery and/or a refine-ment of existent pharmacotherapies. In the future, disease-related transcripts might also be targets for gene therapyinterventions.
Submitted for publication January 21, 2000; final revisionreceived September 26, 2001; accepted October 18, 2001.
From the Departments of Pharmacology and Psychia-try and Behavioral Sciences, Yerkes Regional Primate Re-search Center, Neuroscience Division, Emory UniversitySchool of Medicine, Atlanta, Ga (Dr Hemby); the Demen-tia Research Program, Department of Psychiatry, NathanKline Institute, New York University School of Medicine,
Table 2. Number and Abundances of mRNAs Whose Abundance Is Altered in Schizophreniaand Whose Genes Map to Presumed Schizophrenia Linkage Sites*
Linkage Site
Schizophrenic�Control†
No Change
Control�Schizophrenic‡
�4 � �2-4 � 1.5-2 � 1.5-2 � �2-4 � �4 �
5q11 0 6 7 28 3 1 05p14 1 0 1 9 1 1 06p22-23 2 8 5 35 3 1 08p21-22 0 3 2 26 1 3 010q21-22 2 17 15 58 8 9 013q14 0 4 4 22 4 3 013q32 2 6 6 28 2 4 115q13-14 0 2 0 14 1 1 022q12 2 18 12 69 15 8 1Xq24 2 10 13 41 8 8 0
*Data are given as number of messenger RNAs (mRNAs) at greater than 4-fold, greater than 2- to 4-fold, and 1.5- to 2-fold differences.†Indicates abundance was greater in the schizophrenic patients.‡Indicates abundance was greater in normal controls.
(REPRINTED) ARCH GEN PSYCHIATRY/ VOL 59, JULY 2002 WWW.ARCHGENPSYCHIATRY.COM638
©2002 American Medical Association. All rights reserved.

Orangeburg (Dr Ginsberg); and the Center for Bioinfor-matics (Dr Brunk) and the Departments of Psychiatry (DrsArnold and Eberwine), Neurology (Drs Arnold and Eber-wine), Pharmacology (Dr Eberwine), and Pathology andLaboratory Medicine (Dr Trojanowski), University of Penn-sylvania School of Medicine, Philadelphia. Dr Hemby is aconsultant for Solvay Pharmaceuticals, Utrecht, the Neth-erlands. Dr Eberwine is on the Scientific Advisory Board ofIncyte Pharmaceuticals, Sunnyvale, Calif, which owns Ge-nome Systems. Drs Eberwine and Trojanowski are Found-ing Scientists, consultants and stockholders for Layton Bio-Science, Inc, Sunnyvale, which has licensed the aRNAamplification and in situ transcription methods.
This study was supported by the Walter Sonneborn KatzNational Alliance for Research on Schizophrenia and De-pression (NARSAD) Young Investigator Award (Great Neck,NY) and a National Alliance for Autism Research Award(Princeton, NJ) (Dr Hemby), a NARSAD Distinguished In-vestigator Award (Dr Eberwine), grants AG10124 andAG09215 from the National Institute on Aging (Bethesda,Md) (Dr Trojanowski), and grants MH55199 (Dr Arnold)and MH43880 (Drs Trojanowski and Arnold) from theNational Institute of Mental Health (Bethesda).
The Functional Genomics Facility of the Emory Uni-versity School of Medicine, Atlanta, Ga, provided the cDNAclones for secondary screening, and the Stanley Foundation,Bethesda, kindly provided schizophrenic and normal tissuesections from brains in their brain bank. Larry Jones, PhD,kindly provided the phospholemman antibody. The authorsthank the staff of the Mental Health Clinical Research Cen-ter on Schizophrenia and the Department of Pathology andLaboratory Medicine of the University of Pennsylvania fortheir assistance in case accrual and evaluation.
Corresponding author and reprints: James H. Eber-wine, PhD, Department of Pharmacology, University of Penn-sylvania, 3620 Hamilton Walk, Philadelphia, PA 19104.
REFERENCES
1. American Psychiatric Association. Diagnostic and Statistical Manual of Mental Dis-orders, Fourth Edition. Washington, DC: American Psychiatric Association; 1994.
2. Gur RE. Functional brain-imaging studies in schizophrenia. In: Psychopharma-cology: The Fourth Generation of Progress. 4th ed. New York, NY: Raven Press;1995:1185-1192.
3. Bogerts B, Ashtari M, Degreef G, Alvir JM, Bilder RM, Lieberman JA. Reducedtemporal limbic structure volumes on magnetic resonance images in first epi-sode schizophrenia. Psychiatry Res. 1990;35:1-13.
4. Breier A, Buchanan RW, Elkashef A, Munson RC, Kirkpatrick B, Gellad F. Brain mor-phology and schizophrenia: a magnetic resonance imaging study of limbic, pre-frontal cortex, and caudate structures. Arch Gen Psychiatry. 1992;49:921-926.
5. Shenton ME, Kikinis R, Jolesz FA, Pollak S, Lemay M, Wible C, Hokama H, Mar-tin J, Metcalf D, Coloman M. Abnormalities of the left temporal lobe and thoughtdisorder in schizophrenia: a quantitative magnetic resonance imaging study.N Engl J Med. 1992;327:604-612.
6. Altshuler LL, Bartzokis G, Grieder T, Curran J, Jimenez T, Leight K, Wilkins J,Gerner R, Mintz J. An MRI study of temporal lobe structures in men with bipolardisorder or schizophrenia. Biol Psychiatry. 2000;48:147-162.
7. Suddath RL, Christison GW, Torrey EF, Casanova MF, Weinberger DR. Anatomi-cal abnormalities in the brains of monozygotic twins discordant for schizophre-nia [published correction appears in N Engl J Med. 1990;322:1616]. N Engl JMed. 1990;322:789-794.
8. Becker T, Elmer K, Mechela B, Schneider F, Taubert S, Schroth G, Grodd W, Bar-tels M, Beckmann H. MRI findings in medial temporal lobe structures in schizo-phrenia. Eur Neuropsychopharmacol. 1990;1:83-86.
9. Rossi A, Stratta P, Mancini F, Gallucci M, Mattei P, Core L, Di Michele V, Casac-chia M. Magnetic resonance imaging findings of amygdala-anterior hippocampusshrinkage in male patients with schizophrenia. Psychiatry Res. 1994;52:43-53.
10. Gur RE, Turetsky BI, Cowell PE, Finkelman C, Maany V, Grossman RI, ArnoldSE, Bilker WB, Gur RC. Temporolimbic volume reductions in schizophrenia. ArchGen Psychiatry. 2000;57:769-775.
11. Flaum M, Swayze VW II, O’Leary DS, Yuh W, Ehrhardt J, Arndt S, Andreasen N.Effects of diagnosis, laterality, and gender on brain morphology in schizophre-nia. Am J Psychiatry. 1995;152:704-714.
12. Buchanan RW, Breier A, Kirkpatrick B, Elkashef A, Munson RC, Gellad F, Car-penter WT Jr. Structural abnormalities in deficit and nondeficit schizophrenia.Am J Psychiatry. 1993;150:59-65.
13. Turetsky BT, Cowell PE, Gur RC, Grossman RI, Shtasel DL, Gur RE. Frontal andtemporal lobe brain volumes in schizophrenia: relationship to symptomatologyand clinical subtype. Arch Gen Psychiatry. 1995;52:1061-1070.
14. Hoff AL, Wieneke M, Faustman WO, Horon R, Sakuma M, Blankfeld H, EspinozaS, DeLisi LE. Sex differences in neuropsychological functioning of first-episodeand chronically ill schizophrenic patients. Am J Psychiatry. 1998;155:1437-1439.
15. Falkai P, Honer WG, David S, Bogerts B, Majtenyi C, Bayer TA. No evidence forastrogliosis in brains of schizophrenic patients: a post-mortem study. Neuro-pathol Appl Neurobiol. 1999;25:48-53.
16. Arnold SE, Trojanowski JQ, Gur RE, Blackwell P, Han L, Choi C. Absence of neu-rodegeneration and neural injury in the cerebral cortex in a sample of elderly pa-tients with schizophrenia. Arch Gen Psychiatry. 1998;55:225-232.
17. Trojanowski JQ, Arnold SE. In pursuit of the molecular neuropathology of schizo-phrenia. Arch Gen Psychiatry. 1995;2:274-276.
18. Eastwood SL, Harrison PJ. Detection and quantification of hippocampal synap-tophysin messenger RNA in schizophrenia using autoclaved, formalin-fixed, par-affin wax–embedded sections. Neuroscience. 1999;93:99-106.
19. Eastwood SL, Harrison PJ. Hippocampal and cortical growth-associated pro-tein-43 messenger RNA in schizophrenia. Neuroscience. 1998;86:437-448.
20. Eastwood SL, Harrison PJ. Decreased synaptophysin in the medial temporal lobein schizophrenia demonstrated using immunoautoradiography. Neuroscience.1995;69:339-343.
21. Eastwood SL, Burnet PW, Harrison PJ. Altered synaptophysin expression as a markerof synaptic pathology in schizophrenia. Neuroscience. 1995;66:309-319.
22. Thompson PM, Sower AC, Perrone-Bizzozero NI. Altered levels of the synapto-somal associated protein SNAP-25 in schizophrenia. Biol Psychiatry. 1998;43:239-243.
23. Sokolov BP, Tcherepanov AA, Haroutunian V, Davis K. Levels of mRNAs encod-ing synaptic vesicle and synaptic plasma membrane proteins in the temporal cor-tex of elderly schizophrenic patients. Biol Psychiatry. 2000;48:184-196.
24. Young CE, Arima K, Xie J, Hu L, Beach T, Falkai P, Honer W. SNAP-25 deficitand hippocampal connectivity in schizophrenia. Cereb Cortex. 1998;8:261-268.
25. Akil M, Lewis DA. The catecholaminergic innervation of the human entorhinal cor-tex: alterations in schizophrenia [abstract]. Soc Neurosci Abstr. 1995;21:238.
26. Longson D, Deakin JF, Benes FM. Increased density of entorhinal glutamate-immunoreactive vertical fibers in schizophrenia. J Neural Transm. 1996;103:503-507.
27. Eastwood SL, McDonald B, Burnet PW, Beckwith J, Kerwin R, Harrison P. De-creased expression of mRNAs encoding non-NMDA glutamate receptors GluR1and GluR2 in medial temporal lobe neurons in schizophrenia. Mol Brain Res. 1995;29:211-223.
28. Arnold SE, Han L-Y, Ruscheinsky DD. Further evidence of cytoarchitectural ab-normalities of the entorhinal cortex in schizophrenia using spatial point patternanalyses. Biol Psychiatry. 1997;42:639-647.
29. Arnold SE, Hyman BT, Hoesen GWV, Damasio AR. Some cytoarchitectural ab-normalities of the entorhinal cortex in schizophrenia. Arch Gen Psychiatry. 1991;48:625-632.
30. Jakob H, Beckmann H. Prenatal developmental disturbances in the limbic allo-cortex in schizophrenics. J Neural Transm. 1986;65:303-326.
31. Arnold SE, Franz BR, Gur RC, Gur RE, Shapiro R, Moberg P, Trojanowski J. Smallerneuron size in schizophrenia in hippocampal subfields that mediate cortical-hippocampal interactions. Am J Psychiatry. 1995;152:738-748.
32. Arnold SE, Lee VMY, Gur RE, Trojanowski J. Abnormal expression of two mi-crotubule-associated proteins (MAP2 and MAP5) in specific subfields of the hip-pocampal formation in schizophrenia. Proc Natl Acad Sci U S A. 1991;88:10850-10854.
33. Jones RSG. Entorhinal-hippocampal connections: a speculative view of their func-tion. Trends Neurosci. 1993;16:58-64.
34. Eberwine J, Crino P, Arnold S, et al. Molecular analysis of the single cell: impor-tance in the study of psychiatric disorders. In: Psychopharmacology: Fifth Gen-eration of Progress [book on CD-ROM]. Philadelphia, Pa: Lippincott-Raven Pub-lishers; 1998.
35. Mirnics K, Middelton FA, Marquez A, et al. Molecular characterization of schizo-phrenia viewed by microarray analysis of gene expression in prefrontal cortex.Neuron. 2000;28:53-67.
(REPRINTED) ARCH GEN PSYCHIATRY/ VOL 59, JULY 2002 WWW.ARCHGENPSYCHIATRY.COM639
©2002 American Medical Association. All rights reserved.

36. Arnold SE, Gur RE, Shapiro RM, Fisher K, Moberg P, Gibney M, Gur RC, Black-well P, Trojanowski J. Prospective clinicopathologic studies of schizophrenia:accrual and assessment of patients. Am J Psychiatry. 1995;152:731-737.
37. Ginsberg SD, Crino PB, Hemby SE, Weingarten J, Lee U, Eberwine J, Tro-janowski J. Predominance of neuronal mRNAs in individual Alzheimer’s diseasesenile plaques. Ann Neurol. 1999;45:174-181.
38. Mikel UV, Becker RL Jr. A comparative study of quantitative stains for DNA inimage cytometry. Anal Quant Cytol Histol. 1991;13:253-260.
39. Lee VMY, Carden MJ, Schlaepfer WW, Trojanowski J. Monoclonal antibodies dis-tinguish several differentially phosphorylated states of the two largest rat neu-rofilament subunits (NF-H and NF-M) and demonstrate their existence in the nor-mal nervous system of adult rats. J Neurosci. 1987;7:3474-3488.
40. Tecott L, Barchas J, Eberwine J. In situ transcription: specific synthesis of cDNAin fixed tissue sections. Science. 1988;240:1661-1664.
41. Van Gelder RN, von Zastrow ME, Yool A, Dement W, Barchas J, Eberwine J. Am-plified RNA synthesized from limited quantities of heterogeneous cDNA. ProcNatl Acad Sci U S A. 1990;87:1663-1667.
42. Eberwine J, Yeh H, Miyashiro K, Cao Y, Nair S, Finnell R, Zettel M, Coleman P.Analysis of gene expression in single live neurons. Proc Natl Acad Sci U S A.1992;89:3010-3014.
43. Ginsberg SD, Hemby SE, Weintgarten JE, Lee V, Eberwine J, Trojanowski J. Ex-pression profile of transcripts in Alzheimer’s disease tangle bearing CA1 neu-rons. Ann Neurol. 2000;48:77-87.
44. DoTS program of Genomics Unified Schema. Allgenes.org Web site. Availableat: http://www.allgenes.org. Accessed 2000.
45. Harrison PJ, Burnet PW, Falkai P, Bogerts B, Eastwood S. Gene expression andneuronal activity in schizophrenia: a study of polyadenylated mRNA in the hip-pocampal formation and cerebral cortex. Schizophr Res. 1997;6:93-102.
46. Joyce JN, Lexow N, Kim SJ, Artymyshyn R, Senzon S. Lawrence D, CassanovaM, Winokur A. Distribution of beta-adrenergic receptor subtypes in human post-mortem brain: alterations in limbic regions of schizophrenics. Synapse. 1992;10:228-246.
47. Gao XM, Sakai K, Roberts RC, Conley R, Dean B, Tamminga C. Ionotropic glu-tamate receptors and expression of N-methyl-D-aspartate receptor subunits insubregions of human hippocampus: effects of schizophrenia. Am J Psychiatry.2000;157:1141-1149.
48. Healy DJ, Haroutunian V, Powchik P, Davidson M, Davis K, Watson S, Meador-Woodruff J. AMPA receptor binding and subunit mRNA expression in prefrontalcortex and striatum of elderly schizophrenics. Neuropsychopharmacology. 1998;19:278-286.
49. Noga JT, Hyde TM, Herman MM, Spurney C, Bigelow L, Weinberger D, Klein-man J. Glutamate receptors in the postmortem striatum of schizophrenic, sui-cide, and control brains. Synapse. 1997;27:168-176.
50. Sokolov BP. Expression of NMDAR1, GluR1, GluR7, and KA1 glutamate recep-tor mRNAs is decreased in frontal cortex of “neuroleptic-free” schizophrenics:evidence on reversible up-regulation by typical neuroleptics. J Neurochem. 1998;71:2454-2464.
51. Meador-Woodruff JH, Healy DJ. Glutamate receptor expression in schizo-phrenic brain. Brain Res Brain Res Rev. 2000;31:288-294.
52. Benes FM, Vincent SL, Marie A, Khan Y. Up-regulation of GABAA receptor bind-ing on neurons of the prefrontal cortex in schizophrenic subjects. Neurosci-ence. 1996;75:1021-1031.
53. Benes FM, Khan Y, Vincent SL, Wickramasinghe R. Differences in the subre-gional and cellular distribution of GABAA receptor binding in the hippocampalformation of schizophrenic brain. Synapse. 1996;22:338-349.
54. Dean B, Hussain T, Hayes W, Scarr E, Kitsoulis S, Hill C, Opeskin K, Copolow D.
Changes in serotonin2A and GABA(A) receptors in schizophrenia: studies on thehuman dorsolateral prefrontal cortex. J Neurochem. 1999;72:1593-1599.
55. Ohnuma T, Augood SJ, Arai H, McKenna P, Emson P. Measurement of GABAergicparameters in the prefrontal cortex in schizophrenia: focus on GABA content, GABA(A)receptor alpha-1 subunit messenger RNA and human GABA transporter-1 (HGAT-1)messenger RNA expression. Neuroscience. 1999;93:441-448.
56. Freedman R, Hall M, Adler LE, Leonard S. Evidence in postmortem brain tissuefor decreased numbers of hippocampal nicotinic receptors in schizophrenia. BiolPsychiatry. 1995;38:22-33.
57. Freedman R, Coon H, Myles-Worsley M, Orr-Urtreger A, Olincy A, Davis A, Poly-meropoulos M, Holik J, Hopkins J, Hoff M, Rosenthal J, Waldo MC, Reimherr F,Wender P, Yaw J, Young DA, Breese CR, Adams C, Patterson D, Adler LE, Krug-lyak L, Leonard S, Byerley W. Linkage of a neurophysiological deficit in schizo-phrenia to a chromosome 15 locus. Proc Natl Acad Sci U S A. 1997;94:587-592.
58. Sudhof TC. The synaptic vesicle cycle: a cascade of protein-protein interactions.Nature. 1995;375:645-653.
59. Shao X, Li C, Fernandez I, Zhang X, Sudhof TC, Rizo J. Synaptotagmin-syntaxininteraction: the C2 domain as a Ca2+-dependent electrostatic switch. Neuron. 1997;18:133-142.
60. Gabriel SM, Haroutunian V, Powchik P, Honer WG, Davidson M, Davies P, DavisKL. Increased concentrations of presynaptic proteins in the cingulate cortex ofsubjects with schizophrenia. Arch Gen Psychiatry. 1997;54:559-566.
61. Karson CN, Mrak RE, Schluterman KO, Sturner WQ, Sheng JG, Griffin WS. Al-terations in synaptic proteins and their encoding mRNAs in prefrontal cortex inschizophrenia: a possible neurochemical basis for “hypofrontality.” Mol Psy-chiatry. 1999;4:39-45.
62. Perrone-Bizzozero NI, Sower AC, Bird ED, Benowitz LI, Ivins KJ, Neve RL. Levelsof the growth-associated protein GAP-43 are selectively increased in associationcortices in schizophrenia. Proc Natl Acad Sci U S A. 1996;93:14182-14187.
63. Glantz LA, Lewis DA. Reduction of synaptophysin immunoreactivity in the pre-frontal cortex of subjects with schizophrenia: regional and diagnostic specific-ity. Arch Gen Psychiatry. 1997;54:943-952.
64. Honer WG, Falkai P, Young C, Hashimoto K, Hondo H, Hisatomi S, Motomura K,Uchimura H. Cingulate cortex synaptic terminal proteins and neural cell adhe-sion molecule in schizophrenia. Neuroscience. 1997;78:99-110.
65. Nakahara T, Nakamura K, Tsutsumi T, et al. Effect of chronic haloperidol treat-ment on synaptic protein mRNAs in the rat brain. Mol Brain Res. 1998;61:238-242.
66. Eastwood SL, Heffernan J, Harrison PJ. Chronic haloperidol treatment differen-tially affects the expression of synaptic and neuronal plasticity-associated genes.Mol Psychiatry. 1997;2:322-329.
67. Yang CQ, Kitamura N, Nishino N, Shirakawa O, Nakai H. Isotype-specific G pro-tein abnormalities in the left superior temporal cortex and limbic structures ofpatients with chronic schizophrenia. Biol Psychiatry. 1998;43:12-19.
68. Humphries C, Mortimer A, Hirsch S, de Belleroche N. NMDA receptor mRNA cor-relation with antemortem cognitive impairment in schizophrenia. Neuroreport.1996;7:2051-2055.
69. Chen LS, Lo CF, Numann R, Cuddy M. Characterization of the human and ratphospholemman (PLM) cDNAs and localization of the human PLM gene to chro-mosome 19q13.1. Genomics. 1997;41:435-443.
70. Moorman JR, Ackerman SJ, Kowdley GC, Griffin MP, Mounsey JP, Chen Z, CalaSE, O’Brian JJ, Szabo G, Jones LR. Unitary anion currents through phospho-lemman channel molecules. Nature. 1995;377:737-740.
71. Chen Z, Jones LR, O’Brian JJ, Moorman JR, Cala SE. Structural domains in phos-pholemman: a possible role for the carboxyl terminus in channel inactivation.Circ Res. 1998;82:367-374.
(REPRINTED) ARCH GEN PSYCHIATRY/ VOL 59, JULY 2002 WWW.ARCHGENPSYCHIATRY.COM640
©2002 American Medical Association. All rights reserved.



















