
Maladaptive anticipatory saccades in schizophrenia
16
BiOL PSYCHIATRY "/79 1991 ;30:"ncjLT~ Maiadaptivc Anticipatory Saccades in Schizophrenia Daniel W. Hommer, Thomas Clem, Robert Litman, and David Pickar We compared the saccades made by 8 neuroleptic-treated and 7 drug-free schizophrenic inpvtients with those made by ! ! normal controls during two eye movement tasks. The first task was designed to elicit visually guided but not internally guided saccades. The second task was designed so that optimal performance required saccades be guided on the basis of an internal representation of target behavior. During the first task, schizo- phrenics made visually guided saccades that were as accurate as those made by control, but both drug-free and neuroleptic-treated schizophrenics made intrusive saccades at a significantly higher rate than control subjects. Most of these maladaptive saccades ap- peared to be premature attempts to anticipate target jump. During the second eye move- ment task, which for optimal performance required use of an internal representation to guide eye movements, most patients learned to anticipate target jump as well as controls. However, neuroleptic-treated patients made significantly smaller adaptive anticipatory saccades than either drug-free schizophrenic patients or normal subjects. These finding are discussed as they relate to the prefrontal cortex-basal ganglia circuits involved in the regulation of behavior by representational knowledge and the idea that the abnormal anticipatory saccades we observed represent a failure in the sensorimotor gating of information derived from internal representations. Introduction Since the rediscovery of the phenomena 18 years ago by Holzman et al (1973), the study of smooth pursuit eye movement dysfunction has become an important focus for research in o..k:......~...~..:.~ u ....... o,. e~,.,.~a;,- mnu~,m~ntc of schDnphrenic'g have nnt ken I ll.J¢ ~ t., ¥ ¢~1 9 11 l~h.7 v ~ i i ,KI~ i I $~.P a ~ u v a u v a a ~ v w ~ . ~ . . . . . . studied as extensively as their smooth pursuit. Despite this, there seems to be general agreement on scvcrai aspects of saccadic function in schizophrenia. Most investigators have found thatthc latencyand velocityof visuallyguided saccadcs arc normal (Dicfcndorf and Dodge 1908; Lcvin ct al 1982; Mathcr and Putchat 1983; van dcn Bosch 1984). In contrast, aspects of saccadic function related to the ability to inhibit nonfunctional, inappropriatesaccadcs appear to bc disordcrcd. Schizophrenics makc many more intrusivc saccadcs whcn thcy should bc fi):ing tlieir gaze on a nonmoving targetthan normal subjects From the Geriatrics Research Education and Clinical Center, Seattle VA Medical Center and the Depa.,Iment of Psychiatry and Behavioral Sciences, University of Washington, Seattle. WA (DWH); the Biomedical Engineering Branch, National Institutes of Health, Bethesda, MD ('rc); and the Clinic~ Neuroscience Branch, National Institutes of Mental Health, Betbesda, MD (RT,DP). Received October 29, 1990, revised April 5, 1991. Address reprint requests to Daniel W. Hommer, M.D., ',he Geriatrics Research Education and Clinical Center, Seattle VA Medical Center, 1660 S. Columbian Way, Seattle, WA 98108. © 1991 Society of Biological Psychiatry 0006-3223/91/$03.50
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Maladaptive anticipatory saccades in schizophrenia

BiOL PSYCHIATRY "/79 1991 ;30:"ncjLT~
Maiadaptivc Anticipatory Saccades in Schizophrenia
Daniel W. Hommer, Thomas Clem, Robert Litman, and
David Pickar
We compared the saccades made by 8 neuroleptic-treated and 7 drug-free schizophrenic inpvtients with those made by ! ! normal controls during two eye movement tasks. The first task was designed to elicit visually guided but not internally guided saccades. The second task was designed so that optimal performance required saccades be guided on the basis of an internal representation of target behavior. During the first task, schizo- phrenics made visually guided saccades that were as accurate as those made by control, but both drug-free and neuroleptic-treated schizophrenics made intrusive saccades at a significantly higher rate than control subjects. Most of these maladaptive saccades ap- peared to be premature attempts to anticipate target jump. During the second eye move- ment task, which for optimal performance required use of an internal representation to guide eye movements, most patients learned to anticipate target jump as well as controls. However, neuroleptic-treated patients made significantly smaller adaptive anticipatory saccades than either drug-free schizophrenic patients or normal subjects. These finding are discussed as they relate to the prefrontal cortex-basal ganglia circuits involved in the regulation of behavior by representational knowledge and the idea that the abnormal anticipatory saccades we observed represent a failure in the sensorimotor gating of information derived from internal representations.
Introduction
Since the rediscovery of the phenomena 18 years ago by Holzman et al (1973), the study of smooth pursuit eye movement dysfunction has become an important focus for research in o..k:......~...~..:.~ u ....... o,. e~,.,.~a;,- mnu~,m~ntc of schDnphrenic'g have nnt k e n I l l . J ¢ ~ t . , ¥ ¢ ~ 1 9 1 1 l~h.7 v ~ i i ,KI~ i I $~.P a ~ u v a u v a a ~ v w ~ . ~ . . . . . .
studied as extensively as their smooth pursuit. Despite this, there seems to be general agreement on scvcrai aspects of saccadic function in schizophrenia. Most investigators have found that thc latency and velocity of visually guided saccadcs arc normal (Dicfcndorf and Dodge 1908; Lcvin ct al 1982; Mathcr and Putchat 1983; van dcn Bosch 1984). In contrast, aspects of saccadic function related to the ability to inhibit nonfunctional, inappropriate saccadcs appear to bc disordcrcd. Schizophrenics makc many more intrusivc saccadcs whcn thcy should bc fi):ing tlieir gaze on a nonmoving target than normal subjects
From the Geriatrics Research Education and Clinical Center, Seattle VA Medical Center and the Depa.,Iment of Psychiatry and Behavioral Sciences, University of Washington, Seattle. WA (DWH); the Biomedical Engineering Branch, National Institutes of Health, Bethesda, MD ('rc); and the Clinic~ Neuroscience Branch, National Institutes of Mental Health, Betbesda, MD (RT,DP).
Received October 29, 1990, revised April 5, 1991. Address reprint requests to Daniel W. Hommer, M.D., ',he Geriatrics Research Education and Clinical Center, Seattle VA
Medical Center, 1660 S. Columbian Way, Seattle, WA 98108.
© 1991 Society of Biological Psychiatry 0006-3223/91/$03.50

780 BIOL PSYCHIATRY 1991 ;30:779-794
D.W. Hommer et al
do (Cegalis et al 1983; lacono et al 1981; Mailet and Pichot 1981; Matsue et al 1986; Schmid-Burgk et~ al 1982).
Levin (1984) has suggested that this increase in intrusive saccades results from a failure of the frontal cortex to inhibit subcortical structures involved in oculomotor control, particularly the superior colliculus (SC). From this poi,~' of view, intrusive saccades could be considered a manifestation of a more general defect in schizophrenic brain function, possibly inadequate sensorimotor gating (Braff and Geyer 1990). Thaker et al (1989) have proposed that reduced activity hi hie basal ganglia gamma-amino-butyric acid (GABA)- containing output pathway from substantia nigra to the SC is responsible for the increased saccadic distractibility they observed among schizophrenics during an antisaccade task. On the basis of single unit recording studies in behavior nonhuman primates, Hikosaka (1989) has argued that this nigral-collicular pathway controls a "gate" in the SC through which multiple cortical inputs converge to trigger saccades.
Although the precise nature of the intrusive saccades that interrupt fixation in schizo- phrenia has not been determined, it has been generally assumed that they are random phenomena, perhaps a kind "motor noise." However, two groups of investigators have noted that some intrusive saccades made by schizophrenics appear to be premature an- ticipations of future target behavior (Levin et al 1982; Mather and Putchat 1983). If many of the intrusive saccades made by schizophrenics are anticipatory in nature, this suggests that in addition to supporting ideas about frontal lobe dysfunction and impaired senso- rimotor gating, saccadic abnormalities may also provide a way of exploring Goldman- Rakic's (1987) recent proposal that schizophrenia can be characterized as a dysfunction affecting the prefrontal cortex and resulting in an impairment in the use of representational knowledge to guide behavior. Anticipatory saccades are not visually guided; they must be triggered on the basis of an internal representation of future target behavior. For this reason, analysis of the nature of intrusive saccades made by schizophrenics may provide information relevant to Goldman-Rakic's proposal. In addition, examination of the specific types of saccadic errors made by schizophrenics may provide a way of understanding the oculomotor dysfunction of schizophrenia within the general framework that Fuster (1989) has employed to analyze failure of the prefrontal cortex.
In this report, we compare the saccades made by neuroleptic-treated and drug-free schizophrenics with those made by normal controls during two eye movement tasks, each designed to provide a different context related to anticipation. In one of these tasks, anticipatory -- ~--' - sac~:aues were adaptive and indicated effective utilization of internal repre- sentations. In the other task, anticipatory saccades were maladaptive and indicated a failure to properly use internal representations to guide behavior. Both tasks were per- formed in complete darkness in order to reduce the likelihood that saccades would be guided by visual stimuli other than the target.
The first task was designed to encourage visually guided saccades while discouraging internally guided, anticipatory saccades. During the task, the next location was always predictable but the timing of the next target step was unpredictable. During this type of task, if a subject attempts to guide saccades on the basis of an internal representation of the target's partially regular behavior, the result will be premature and maladaptive anticipatory saccades that actually decrease the amount of time the target is centered on the fovea. We refer to this task as the visually guided saccade-maladaptive anticipatory saccade (VGS-MAS) task.
The second task was designed to encourage nonvisually guided (internally generated) saccades that interrupt fixation to accurately anticipate the next change in target position.

Saccades in Schizophrenia BIOL PSYCHIATRY 78 i 1991 ;30:'J79-794
We refer to this task as the nonvisually guided saccade-adaptive anticipatory saccade (NVGS-AAS) task. During the NVGS-AAS task, the target's behavior is completely predictable and cues are provided to make learning to predict easy. The most adaptive way to engage in this task (i.e., the way to maximize the amount of time the target is on the fovea) is for the subject to base his or her eye movements on an internalized representation of the target's completely regular behavior. Consistent with the proposed functional neuroanatomy of internal representations (Goldman-Rakic 1987), lesions of the frontal eye field (FEF) impair the ability of monkeys to make adaptive anticipatory saccades during a task identical to our NVGS-AAS task but have no effect on purely visually guided saccades (Bruce and Borden 1986).
If schizophrenia is a disorder affecting utilization of representatioaal knowledge, as Goldman-Rakic suggests, then the visually guided saccades made by schizophrenics in either task should be csse,hally normal, as this type of saccade is not guided on the basis of an internal representation. In contrast, saccades generated on the basis of an internal representation should be dysfunctional. A dysfunction of internal representation could take three possible forms which correspond to the three core aspects of prefrontal cortex failure, as described by Fuster (1989): (l) an impairment in the ability to make adaptive anticipatory saccades during the NVGS-AAS task. This deficit corresponds to what Fuster has called a failure of preparatory set; (2) an impaired accuracy of adaptive anticipatory saccades which corresponds to a deficiency in what Fuster has called provisional short- term memory (other investigators have called this working memory (Goldman-Rakic 1987); (3) a failure to inhibit inappropriate attempts to make anticipatory saccades during the VGS-MAS task which corresponds to what Fuster has called a failure to suppress internal sources of interference in file formation of behavioral structures. This also could be cor, sidered a failure in the sensorimotor gating of information derived from an internal repr,.'~entation.
If schizophrenics have no impairment in either the generation or appropriate utilization of internal representations, then they should make accurate anticipatory saccades during the NVGS-AAS task and refrain from making anticipatory saccades during the VGS- MAS task.
Subjects and Methods
~ J U I I L I U I " " ! ~ 1 [ ; ¥ [ ; ! 1 i i ~ f i l U l ~ 3 u u J ~ . , t 3 n i l u I J i i l l J t Z l , l ~ l i t ~ ) W I U | O~.,itlIZ...~-WI~I~IUI-VaRAf~ L J m L ' ~ ' I ' ~ . .rl,~j,l'~'b
study. Informed consent was obtained prior to eye movement recording. The schizophrenic patients were all inpatients at the Clinical Center of the National Institutes of Health in Bethesda, Maryland. All schizophrenic patients met DSM-III-R (APA 1987) criteria for chronic schizophrenia and had been hospitalized for at least I month. Eight of the schizophrenics had been receiving flupbenazine hydrochloride in stable doses of 10-40 rag/day (mean = 26.25 rag/day, SD = 14.07) and benzotropine (1-4 rag/day) for at least 3 weeks. No patient had clinically apparent akathisia, rigidity, tremor, or any evidence of tardive dyskinesia. The drug-free schizophrenic patients had not received any neuroleptic or other psychotropic medicatk, for at least 3 weeks. The healthy control subjects had been free of any medication for at least 3 weeks. All subjects, both patients and normal volunteers, were right-handed.
All the schizophrenic patients had been ill for at least 4 years and had histories of multiple hospitalizations for the treatment of psychosis, and at the time of recording, were suffering from moderately severe psychosis. The mean age of the subject groups

782 BIOL PSYCHIATRY 1991 ;30:779-794
D.W. Hommer et al
were, for fluphenazine-treated patients 25.7 years (SD 3.1); drug-free patients 30.6 years (SD 3.2); normal controls 32.5 years (SD 5.9). Six of the subjects in each group were
men.
Eye Movement Recording Apparatus Eye movements were recorded using an infrared limbus detection eye tracking device (Eye Trac model 210, Applied Sciences Laboratories, Waltham, MA) which has a re- sponse time constant of 4 msec, an accuracy of better than 0.25*, an~l a range of 15" to the left and fight of center. The subjects were seated a distance of 43 cm in front of a video monitor on which the visual target was displayed. The target was a small, bright square that subtended a visual angle of less than 0.25*. It was displayed against a dark background in a darkened room. The subject's head was secured by a bite bar. Only horizontal eye movements were recorded. The analog output of the tracking device was sampled at 1000 Hz with 8 bit resolution and stored for subsequent display and analysis using an IBM AT type microcomputer. Before each run of data collection, the system was electronically calibrated, Data were collected only from the eye for which the most rapid and accurate calibration could be obtained.
Eye Movement Tasks Subjects were instructed to keep their eyes on the target as it moved on the screen and follow it as rapidly and accurately as possible. They were not told anything about the predictability of the target's motion, nor were they told to attempt to anticipate. The first (VGS-NAS) task was designed to elicit visually guided saccades and discourage attempts at anticipation (anticipatory saccades were detrimental to performance). In this task, the target stepped back and forth between l0 ° to the right of center and 10" to the left at a random interval of 1.5-2.5 sec. The mean duration of the interval between target steps was 1.97 _+ 0.08 sec and there were no significant differences between the intervals for the three groups (normals 1.96 sec, neuroleptic-treated schizophrenia 1.98 sec, and drug- free schizophrenics 1.96 sec). A subject could accurately predict the location of the next target appearance, but not its timing. Twenty target steps were presented.
After a brief rest, the subjects engaged in a second (NVGS-AAS) task designed to elicit adaptive anticipatory saccades. This task differed from the VGS-MAS task only in respect to the regular timing of target steps and the cue provided to encourage anticipation. In this task, the target appeared at 10 ° to the right of the center and remained on for 0.75 sec. It then disappeared aod the screen remained completely dark for 0.25 sec. After this gap, the target reappeared at 10 ° to the lea of center and remained on for 0.75 sec, followed again by a gap of 0.25 sec. Finally, the target stepped back to its original location. This cycle was repeated 10 times. In this task, the subject could accurately predict both the next target location and the timing of its jump. The gap served to cue the subject that the target would change location in 250 msec.
Eye Movement Data Analysis
Eye movement data were analyzed using an interactive graphics program developed by one of the authors (TC). The eye and target position were displayed on a computer screen as a plot of position versus time. Each data point represented the mean eye position value

Saccades in Schizophrenia toOL PSYCm~TRV 783 1991 ;~:7";-~-794
22
in 20
0 " 0 18
a u . I " = : a (.,3 ~) 16 0 1 - -
°') I~L. 14
12
oSro O~ 0 O0
0 0 0
mE I 4 ' I I
m • ~
10 NORMALS SCHIZOPHRENICS DRUG-FREE
'O
O O
!
O
O
SCHIZOPHRENICS FLUPHENAZINE
Figure !. During the r i s k y guided eye movement task groups of schizophrenics made as accurate visually guided sac- cades as normal controls.
over an interval of 5 msec. Instantaneous eye velocity and position, as well as time intervals and amplitude related to any eye movement, could be directly measured from the screen display by appropriate placement of the cursors. Eye blink art;eacts could be distinguished from saccades on the basis of their characteristic pattern (a high velocity interval followed immediately by another high velocity interval in the opposite direction (see Figure 5).
Primarily, visually guided saccades were defined as the initial saccade directed towards a target, beginning at least 100 msec but not more than 500 msec after a change in target position. Small saccades occurring less than 560 msec after a change in target position and correcting for inaccuracy (over or undershot) of the initial visually guided saccade were considered corrective saccades. Any saccade larger than 1 ° that occurred 500 msec or more following a change in target position and before the next change in target position was considered an intrusive saccade. Saccades that increased the mismatch (position error) between target and eye position by directing the gaze away from the target, as well as saccades that corrected the mismatch by returning gaze to the target, were counted as intrusions. Intrusive saccades that increased position error and were in the direction of the next target appearance were considered anticipatory.
All data are expressed as the mean ± the standard deviation. Statistical analyses were by univariate analysis of variance (ANOVA) followed by Duncan's multiple range test to evaluate differences among groups.
Results
Visually Guided Saccades during the VGS-MAS Task
The amplitude of primary visually guided saccades made during the VGS-MAS task did not differ significantly among the groups. However, neuroleptic-treated schizophrenics made slightly smaller primary visually guided saccades than either drug-free group. The mean amplitude of the initial visually guided saccade was 18.4 ___ 0.9 ° for controls, 18.7 ___ 1.0 ° for drug-free schizophrenics, and 17.5 ± 2.8 ° for fluphenazine-~e.ated schizo- phrenics (Figure 1).
Both drug-free and fluphenazine-treated schizophrenic patients had a significar.=Sy shorter latency for primary visually guided saccades than control subjects (ANOVA, F(2.23) = 3.75, p = 0.04; Duncan's Multiple Range test, p < 0.05.) (Figure 2). The mean saccadic

784 BIOL PSYCHIATRY 1991 ;30:779-794
D.W. Hommer et al
240
220
Z .200
8 ,°o <
140 O
120
<)
¢1¢
NORMALS SCHIZOPHRENICS SCHIZOPHRENICS 100 DRUG-FREE FLUPHENAZINE
Figure 2. Both groups of schizophrenics showed a modest but significant reduction in the latency of visually guided saccades compared with normal controls.
latency among drug-free schizophrenics was 163 ___ 11 msec; among fluphenazine-treated patients it was 159 _.+ 27 msec; controls had a saccadic latency of 185 ± 25 msec. Both groups of schizophrenic patients made many more primary visually guided saccades with a latency of less than 150 msec than did control subjects (drug-free schizophrenics 40%; fluphenazine-treated schizophrenics 35%; controls 20%).
Intrusive Saccades during the VGS-MAS Task (Maladaptive Anticipation)
Both drug-free and fluphenazine-treated schizophrenics made significantly more frequent intrusive saccades than control subjects during the VGS-MAS task while they should have been maintaining fixation and waiting for the target to move (ANOVA, F(2,23) = 11.3;p < 0.001, Duncan's Multiple Range test p < 0.05). The rates of saccadic intrusions during fixation were as follows: controls 0.26 ___ 0.25 saccades/target step; drug-free schizophrenics 1.04 _ 0.49 saccades/target step; fluphenazine-treated patients 0.83 ± 0.38 saccades/target step (Figure 3). In each of the three groups, more than 95% of the intrusive saccades that took the gaze away from the target were anticipatory, that is, they were in the same direction as the next target appearance.
Drug-free schizophrenic patients made significantly larger saccadic intrusions than
1.0 U3 z 0 ~o0 .8 ~ Z r r 0 !-- O0.6 ___,,, o c o
m,=,,0, O n O <~ J.2 O9
0.0
¢
:t ° ,+ ¢¢.I.¢,0 NORMALS
Ill
Ill B
B m Im
SCHIZOPHRENICS DRUG-FREE
O
¢ ¢
¢ ¢
O
SCHIZOPHRENICS FLUPHENAZlNE
Figure 3. Both drug-free and neu- roleptic-treated schizophrenics made significantly more frequent intrusive saccades during the VGS- MAS task while they should have been maintaining fixation and waiting for the target to move. Sixty percent of the schizophrenics made this type of saccade at a rate 2 SD above the mean rate of the normal controls.
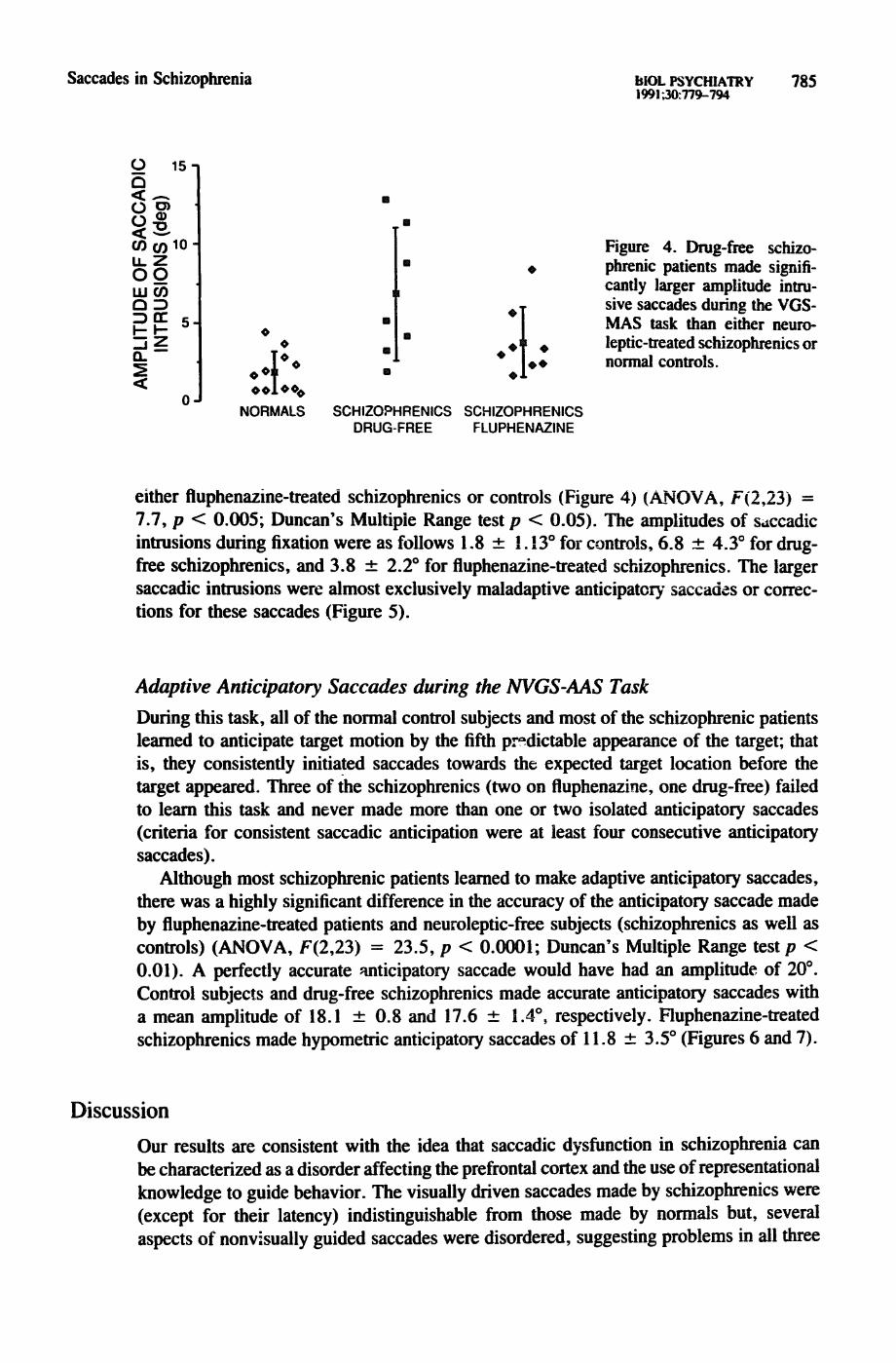
Saccades in Schizophrenia hlOL PSYCHIATRY 7 8 5 1991;30:779-794
f~ 15
< P .
"O
O) if) 10 U . Z o g w m a ~
F - P - -n__ Q.
< .
B
II
O
" "I 0 0 II
~0 0 m OQ' • OO m O14'O
OO OO O
SCHIZOPHRENICS DRUG-FREE
NORMALS SCHIZOPHRENICS FLUPHENAZINE
Figure 4. Drug-free schizo- phrenic patients made signifi- cantly larger amplitude intru- sive saccades during the VGS- MAS task than either neuro- leptic-treated schizophrenics or normal controls.
either fluphenazine-treated schizophrenics or controls (Figure 4) (ANOVA, F(2,23) = 7.7, p < 0.005; Duncan's Multiple Range test p < 0.05). The amplitudes of s,tccadic intrusions during fixation were as follows 1.8 _+ I. 13 ° for controls, 6.8 -4- 4.3 ° for drug- free schizophrenics, and 3.8 _+ 2.2 ° for fluphenazine-treated schizophrenics. The larger saccadic intrusions were almost exclusively maladaptive anticipator)" saccades or correc- tions for these saccades (Figure 5).
Adaptive Anticipatory Saccades during the NVGS-AAS Task
During this task, all of the normal control subjects and most of the schizophrenic patients learned to anticipate target motion by the fifth p~dictable appearance of the target; that is, they consistently initiated saccades towards the expected target location before the target appeared. Three of the schizophrenics (two on fluphenazine, one drug-free) failed to learn this task and never made more than one or two isolated anticipatory saccades (criteria for consistent saccadic anticipation were at least four consecutive anticipatory saccades).
Although most schizophrenic patients learned to make adaptive anticipatory saccades, there was a highly significant difference in the accuracy of the anticipatory saccade made by fluphenazine-treated patients and neuroleptic-free subjects (schizophrenics as well as controls) (ANOVA, F(2,23) = 23.5, p < 0.0001; Duncan's Multiple Range test p < 0.01). A perfectly accurate anticipatory saccade would have had an amplitude of 20 °. Conu'ol subjects and drug-free schizophrenics made accurate anticipatory saccades with a mean amplitude of 18.1 +_- 0.8 and 17.6 ___ 1.4 °, respectively. Fluphenazine-treated schizophrenics made hypometric anticipatory saccades of 11.8 +__ 3.5 ° (Figures 6 and 7).
Discussion
Our results are consistent with the idea that saccadic dysfunction in schizophrenia can be characterized as a disorder affecting the prefrontal cortex and the use of representational knowledge to guide behavior. The visually driven saccades made by schizophrenics were (except for their latency) indistinguishable from those made by normals but, several aspects of nonvisually guided saccades were disordered, suggesting problems in all three

786 BIOL PSYCHIATRY 1991:30:779-794
D.W. Hommer et al
A
W
'r
4 SECONDS
B
F---
4 SECONOS
C D
-~ . - - - - - - - - / -
4 SECONDS 0.5 SECONDS
Figure 5. This figure shows the target (upper) and eye (lower) position recordings of one normal subject (A) and three schizophrenics (B,C,D) during the VGS-MAS task. The schizophrenics made frequent intrusive saccades that appear to be maladaptive, premature anticipations of the target's next step. The star in record B indicates a small eye-blink artifact.
20
t~ U~
O
jmj.
NORMALS
II .]-. 4)
O
4) O
SCHIZOPHRENICS SCHIZOPHRENICS DRUG-FREE FLUPHENAZINE
Figure 6. Neuroleptic-treated schizophrenic patients made significantly smaller and less accurate adaptive anticipatory saccades daring the NVGS-AAS task th~-n either drug-free schizophrenics or normal con- trol su~ects.

Saccades in Schizophrenia BIOL PSYCHIATRY 787 1991;30:779-7~.
NEUROLEPTIC TREATED 2 SECONDS
DRUG FREE 2SECONDS
c~ w
w O o
l u}
Figure 7. This figure shows an eye po- sition recording of a subject, from each of the three groups during the NVGS- AAS task. Target position is indicated by the bars above and below the eye position record. The arrows indicate the beginning and end of nonvisually guided, adaptive anticipatory saccades. The neuroleptic-treated patient makes con- sistenfly smaller anticipatory saccades than subjects from either of the neuro- leptic-free groups.
CONTROL
2SECONDS
e~ o
of Fuster's core areas of prefrontal cortex function. The most dramatic eye movement difference between schizophrenics and normals was in the area of suppression of inter- ference or sensorimotor gating; schizophrenics failed to inhibit maladaptive anticipatory saccades during the VGS-MAS task. In addition, 20% of the schizophrenics, but none of the normals, showed an inability to use a preparatory set to guided oculomotor behavior since during the NVGS-AAS task they failed to make anticipatory saccades. Finally, it appears that neuroleptics and/or anticholinergics produce an impairment in working mem- ory; neurpleptic/antich°linergic'treatecl' but not drug-free, schizophrenic patients made hypometric anticipatory saccades during the NVGS-AAS task.
Although our results show that saccadic dysfunction in schizophrenia can be understood using concepts developed to describe the function of the prefrontal cortex, they should not be taken to constitute compelling evidence for localization of saccadic dysfunction to his brain region. As will be discussed below, the abnormalities we have observed

788 BIOL PSYCHIATRY 1991 ;30:779-794
D.W. Hommer et al
could stem from any level of a neural system involving the basal ganglia and thalamus, as well as the prefrontal cortex. However, we do believe that our findings and the conceptual framework in which we place them can help to further oculomotor research in schizophrenia by providing a new way of seeing eye-movement abnormalities.
For example, during the VGS-MAS task, schizophrenics make many more intrusive saccades than normals do; this is not a new finding. What we have done that we believe is new is, instead of simply considering the excess of intrusive saccades an indication of some type of nonspecific failure of inhibition, we have asked what is the likely function of these clearly maladaptive saccades. Over 95% of the intrusive saccades that take the gaze prematurely off the target are directed toward its next location; the intrusions seemed to be attempts to anticipate target behavior. Any anticipation requires an internal repre- sentation of that which is anticipated. An internal representation of target behavior during the VGS-MAS task can be used to predict only the direction, but not the timing, of the next target step. Our results suggest that although schizophrenics were able to generate an internal representation of the target's future behavior, they were unable to suppress maladaptive anticipatory saccades triggered on the basis of this necessarily incomplete and contextually inappropriate representation. We consider this a failure in the use of an internal representation to guide behavior.
Although schizophrenics appeared unable to effectively inhibit inappropriate antici- patory saccades during the VGS-MAS task, the majority of patients learned to make anticipatory saccades when they were appropriate to the behavioral context. However, despite adequate performance by the majority of patients, fully 20% of the schizophrenics completely failed to anticipate during the NVGS-AAS task. Their failure to learn this simpl~ task represents a severe level of h~pairy ent. Normal monkeys can readily perform the NVGS-AAS task (although animals with lesions of the FEF fail to learn it) (Bruce and Borden 1986). In addition, our NVGS-AAS task is considerably simpler than the oculomotor delayed-response tasks used to evaluate prefrontal cortex function in non- human primates (Funahashi et al 1990) which requires that the correct response be constantly updated by new information. The NVGS-AAS task simply required that the correct response alternate from side to side. As representational processing is most dra- matically impaired when testing with a delayed-response task (Friedman 1989), perfor- mance on the NVGS-AAS task may underestimate the magnitude of representational processing dysfunction among schizophrenics. Whether patients who failed to a_--:t~cipate represent a distinct subgroup of schizophrenics remains to be determined.
Effect of Neuroleptics Neither the frequency of maladaptive anticipatory saccades during the VG~/-MAS task nor the ability to make adaptive anticipatory saccades during the NVGS-AAS task appear to be affected by neuroleptic medication, but the amplitude of these types of saccades is lower among patients receiving neuroleptics and anticholinergics than among medication- free patients. We have interpreted this reduction in amplitude as reflecting an inadequacy of provisional/short-term or working memory. In other words, the representation of the target location (or saccade to be made) held in working memory decays to such an extent over the brief interval that it must be retained, that only hypometric saccades can be generated from it. Sawaguchi and Goldman-Rakic (1991) found hypometric memory- guided saccades in monkeys following an infusion of a dopamine D-1 receptor antagonist directly into the dorsolateral prefrontal cortex and concluded that blockade of cortical

Saccades in Schizophrenia BIOL PSYCHIATRY 789 1991;30:779-794
D-1 dopamine receptors leads to the type of impairment of working memory described above.
Patients with idiopathic parkinsonism sometim s make hypometric saccades even with visual guidance, but show a much greater impai;ment when they are required to make saccades that anticipate target jumps (Bronstein and Kennard 1985; Teriiv~iinen and Calne 1980). Behaviors that rely on inter~,al rather than external guidance seem to be especially vulnerable to the disruptive effects of a reduction in dopaminergic neurotransmission. Cools et al (1984) found that parkinsonian patients are deficient in their ability to sequence motor behavior on the basis of internal, anticipatory cues. Brown and Marsden (1988) demonstrated that parkinsonian patients have a defect in internally cued visual attention as well as in internally cued motor behavior. These findings among parkinsonian patients suggest that some of their saccadic and other deficits could be considered f~ilures of working memory. However, reduction in dopaminergic neurotransmission may not be the entire explanation for the hypometric saccades we observed.
All of our neuroleptic-treated patients were also receiving anticholinergics for the treatment of drug-induced parkinsonism; anticholinergics are known to affect a variety of cognitive functions (Caine et al 1981). Furthermore, verbal episodic memory impair- ment has been found to be directly related to serum anticholinergic levels among schizo- phrenic patients (Perlick et al 1986; Tune et al 1982). However, comparison of schizo- phrenic patients receiving anticholinergics in addition to neuroleptics with patients receiving neuroleptics alone have yielded mixed results. Cleghorn et al (1990) found anticholinergics had no effect on any aspect of memory whereas Calev (1984) found impaired recall. Despite this uncertainty, possible effects of anticholinergics on any psychobiological measure must be carefully considered (Spohn and Strauss 1989). A recent study comparing parkinsonian patients receiving anticholinergics with matched patients receiving only L- Dopa suggests that anticholinergics specifically impair frontal cortex function (Dubois et al 1990). Impairment of frontal cortex should adversely affect working memory and the accuracy of anticipatory saccades. At present, it is impossible to say whether anticholin- ergie~, neuroleptics, or both lead to hypometric saccades among drug-treated patients.
Nature of Saccadic Responses in Schizophrenia The preceding interpretation of saccadic dysfunction in terms of behavior dependent on the prefrontal cortex can be correct only if the intrusive saccades that distinguish schizo- phrenics from controls were generated on the basis of an internal representation and ff visually guided saccades made during the same task were normal. We can be relatively certain that the intrusive saccades were internally generated because the eye movement tasks were performed in the dark and the only object available to visually guide saccades was the target itself. However, simply because a saccade is internally rather than visually guided does not guarantee that it was guided on the basis of a representational model. Internally generated intrusive saccades could simply be manifestations of a fixation in- ctability secondary to random activity in the oculomotor pathways.
Pairs of saccades known as square wave jerks (SWJ) are a commonly observed example of apparently random oculomotor events that interrupt fixation (Elidan et al 1984; Her- ishanu and Sharpe 1981; Ohtsuka et al 1986; Shallo-Hoffman et al 1989). SWJs resemble what we have argued are maladapti'~e anticipatory saccades followed by a corrective saccade (Figure 5). However, there are two differences between SWJs and the saccades we found interrupting schizophrenics' fixation that lead us to believe they represent distinct

790 BIOL PSYCHIATRY 1991:30:779-794
D.W. Hommer et al
phenomena. First, SWJs ~c randomly directed whereas nearly all of the intrusive saccades we observed were in the direction of the next target step. Second, SWJs are generally small saccades with an amplitude usually no greater than 2 ° or 3 ° whereas most of the intrusive saccades made by schizophrenics were considerably larger than this. For these reasons we believe that the intrusive saccades during the VGS-MAS task are manifestations of a failure to properly gate signals from brain circuits involved in the generation of internal representations of target behavior a~.d not simply random fluctuations in fixation stability.
The other requirement for being able to claim that Goldman-Rakic's and Fuster's ideas can be extended to describe saccadic dysfunction in schizophrenia is that saccades not requiring internal representations for their generation be normal. We found that primary visually guided saccades made by schizophrenics (particularly neuroleptic-free patients) were as accurate as those made by controls. However, saccadic latency was significantly shorter amon~ both groups of schizophrenics. This is puzzling because short reaction time usually :,mpiies superior performance and because, during manual reaction time tasks, schizophrenics generally demonstrate prolonged iatencies (Nuechtedein 1977). Furthermore, most investigators who have examined saccadic latency in schizophrenia have found that it does not differ from that of normals (lacono 1981; Levin 1982; Mather and Putchat 1983; van den Bosch 1984).
Our finding of a decreased latency is probably related to the task we used to elicit visually guided saccades. Our task allowed subjects to anticipate future target position but not time of target movement. These conditions may have led to more rapid oculomotor reactions than usually occur when both timing and position are random. Furthermore, a reduction in saccadic latency is consistent with the excessive anticipation that we believe characterizes schizophrenic performance during the VGS-MAS task.
Fischer and Breitmeyer (1987) found that when visual attention is withdrawn from the current fixation target before the next target appears, the visually guided saccade made to the next target has a shorter ia~e,cy than when attention remains focused, if during the VGS-MAS task schizophrenic patients are using an internal (but incomplete) repre- sentation of the behavior of the target to trigger anticipatory saccades, as our patients appear to be doing, then it is also likely that they would use the same representation to trigger a premature withdrawal of attention from the current fixation target, thus allowing them to make short latency visually guided saccades.
Sensorimotor Gating and Basal Ganglia-Thalamocortical Circuits
In her recent reviews of prefrontal cortex function, Goldman-Rakic t 1987, 1988) presented evidence that although specific regions of the prefrontal cortex may be necessary for the full expression of behavior based on inter~d representations, the representations them- selves as the product of neural activity distributed over several interconnected cortical regions. In the case of control of saccadic eye movements, these regions include two areas of the prefrontal cortex the frontal eye fields and the dorsolateral prefrontal cor- tex as well as a region outside the prefrontal cortex-the posterior parietal cortex (Cavada and Goldman-Rakic 1989; Selemon and Goldman-Rakic 1988). These regions share reciprocal connections with each other and they each project to the caudate nucleus (Kunzle and Akert 1977; Parent et al 1984; Selemon and Goldman-Rakic 1985; Yeterian and van Hoesen 1978) forming the cortical origin of what Alexander et al (1986) have called the

Saccades in Schizophrenia BIOL PSYCHIATRY 791 1991;30:'/79-794
oculomotor basal ganglia thalamocortical loop. In addition to their striatal and intr'~ortical connections, all three regions also project monosynaptically to the superior colliculus (SC) (Fries 1984). The SC is a midbrain structure involved in the orientation of attention to external stimuli (Wurtz and Albano 1977) and which, under usual circumstances, serves as the site where multiple inputs converge to form a final common pathway for the generation of saccades (van Opstal and van Gisbergen 1990).
Hikosaka (1989) has suggested that basal ganglia output irom the substantial nigra acts on the SC to regulate a "gate" that controls passage of motor comm~ds from regions of the cortex, including those that generate internal representations used in the control of eye movements, to the brainstem neurons that code directly for saccade metrics. Evidence that the SC performs this gating function comes in part from a study in which Hikosaka and Wurtz (1985) disrupted basal ganglia output by injections of muscimol, a synthetic agonist of the inhibitory neurotransmitter GABA, into the substantia nigra of monkeys engaged in an eye movement task similar to the VGS-MAS task. After the muscimol injections, the monkeys made frequent, premature anticipatory saccades and had a decreased saccadic latency. This pattern is virtually identical to what we have observed among schizophrenics. Hikosaka (1989) interprets the abnormal saccades he and Wurtz found as reflecting a disinhibition of the SC, which leaves the neural gate wide open to multiple cortical inputs. The excessive and maladaptive anticipatory saccades we observed among schizophrenics could result from a similar failure of basal ganglia output to properly gate the multiple inputs to the superior colliculus from the cortical regions involved in generation of an internal representation of target motion. Because the basal ganglia thalamocortical circuit is a loop, it is possible that dysfunction in any of its levels from the cortex to the substantia nigra could lead to gating failure. A gating failure could result in the defective control of interference considered by Fuster (1989) to be characteristics of prefrontal cortex dysfunction.
It may be possible to develop animal models for the saccadic abnormalities we have observed in schizophrenia. However, it is uncertain if such models could be general- ized to account for the linguistic, cognitive, and emotional symptoms of schizophrenia. On a behavioral level, the generalizability of such models would depend on how sa¢- cadic abnormalities are related to the symptoms of schizophrenia, at present, this is not known.
On a neurological level, the utility of an animal model of saceadic dysfunction de- pends on whether basal ganglia thalamocortical circuits similar to the one implicated in saccadic dysfunction are involved in the control of language, cognition, and emo- tion. This may turn out to be the case. Alexander et al (1986) have summarized the evidence for a family of parallel and structurally similar based ganglia thaiamocortical circuits. Some of these circuits play a motor or oculomotor role but other circuits link- ing temporal and cingulate cortices with the limbic striatum most likely play a role in the control of affective, linguistic, and cognitive function. In light of this neuroana- tomy, it is reasonable to suggest that an abnormality of structure or neurotransmission affecting one particular level of these circuits (such as the medial thalamus, globus pallidus/substantia nigra, or striatum) might lead to formally similar abnormalities that could be described in terms of prefrontal cortex dysfunction, and would involve a wide range of behaviors from the oculomotor to the linguistic. These ideas are speculative but they lead to a variety of hypotheses that could be investigated in schizophrenic and normal subjects as well as nonhuman primates.

792 BIOL PSYCHIATRY 1991 ;30:779-794
D.W. Hommer et al
The authors would like to thank Dr. P. Goldman-Rakic for her valuable comments on an earlier version of this manuscript.
References Alexander GE, DeLong MR, Strick PL t!986): Parallel organization of functionally segregated
circuits linking basal ganglia and cortex. Annu Rev Neurosci 9:357-381.
American Psychiatric Association (1987): Diagnostic and Statistical Manual of Mental Disorders 3rd ed, rev. Washington, DC: APA Press.
Bronstein AM, Kennard C (1985): Predictive ocular motor control in Parkinson's disease. Brain 108:925-940.
Braff DL, Geyer MA (1990): Sensorimotor gating in schizophrenia: Human and animal model studies. Arch Gen Psychiatry 47:181-188.
Brown RG, Marsden CD (1988): Internal versus external cues and control of attention in Parkinson's disease. Brain 111:323-346.
Bruce CJ, Borden JA (1986J: The primate frontal eye fields are necessary for predictive saccadic tracking. Soc Neurosci (Abstract) 12:1086.
Caine ED, Weingartner H, Ludlow DL, Cudahy EA, Wehry S (1981): Qualitative anelysis of scopolamine-induced amnesia. Psychopharmacology 74:74-80.
Calev A (1984): Recall and recognition in mildly disturbed schizophrenics: Use of matched tests. Psychol Med 14:425--429.
Cavada C, Goldman-Rakic PS (1989): Posterior parietal cortex in rhesus monkeys: I. Parcellation of areas based on distinctive limbic and sensory, corticocortical connections. J Comp Neurol 287:393-421.
Cegalis JA, Hefez H, Wong PS (1983): What is deviant about deviant smooth pursuit eye movemeLts in schizophrenia. Psychiatry Res 10:47-58.
Cleghorn JM, Kaplan RD, Szechtman B, Szechtman H, Brown GM (1990): Neuroleptic chug effects on cognitive function in schizophrenia. Schizophrenia Res 3:211-219.
Cools, AR, Den Bercken JH, Horstink MW, Van Spaendonck KP, Berger HJ (1984): Cognitive and motor shifting aptitude in Parkinson's disease. J Neuro! Neurosurg Psychiatry 47:443-453.
Diefendorf AR, Dodge R (1908): An experimental study of the ocular reactions of the insane from photographic records. Brain 31:451-489.
Dubois B, Pillon B, Lhermitte F, Agid Y (1990): Cholinergi: deficiency and frontal dysfunction i n P o r t - i . e n . ' e ~ne,~e,~ A . , . M.,,..-.-,I OR- I f " / 191
Elidan J, Gay I, l_~v S (1984): Square wave jerks: Incidence, characteristics and significance. J Otolaryngol ] 3:278-28 I.
Fischer B, Breitmeyer B (1987): Mechanisms of visual attention revealed by saccadic eye move- ments. Neuropsychologia 25:73-83.
Friedman HR, Janas JD, Goldman-Rakic PS (1989): Enhancement of metabolic activity in the diencephalon of monkeys performing working memory tasks: A 2-deoxyglucose study in be- having Rhesus monkeys. J Cognitive Neurosci 2:18-31.
Fries W (1984): Cortical projections to the superior collicu!us in the macaque monkey: A retrograde study using horseradish per,,xidase. J Comp Neurol 230:55-76.
Funahashi S, Bruce CJ, Goldman-Rakic PS (1990): Visuospatial coding in primate prefrontal neurons revealed by oculomotor paradigms. J Neurophysiol 63:814-831.
Fuster JM (1989): The Prefrontal Cortex. New York: Raven Press.
Goldman-Rakic PS (1987): Ch'cuitry of primate prefrontal cortex and the regulation of behavior by representational memoq,,. In Plum F, Mountcastle V (eds), Handbook of Physiology, The Nervous System, Vol. V. Bethesda, MD: American Physiological Society, pp 373-417.

Saccades in Schizophrenia BIOL PSYCHIATRY 793 1991;30:779-794
Goldman-Rakic PS (1988): Changing concepts of cortical connectivity: Parallel distributed cortical networks. In Rakic P, Singer W (eds), Chichester Dahlem Konferenzen, Neurobiology of Neocortex. New York: John Wiley and Sons, p 177.
Herishanu YO, Sharpe JA (1981): Normal square wave jerks. Invest Ophthalmol Vis Sci 20:268- 272.
Hikosaka O (1989): Role of basal ganglia in saccades. Rev Neurol (Paris) 145:580-586.
Hikosaka O, Wurtz RH (1985): Modification of saccadic eye movements by GABA-related sub- stances. I1. Effects of muscimol in monkey substantia nigra pars reticulata. J Neurophysiol 53:292-308.
Holzman PS, Proctor LR, Hughes DW (1973): Eye tracking patterns in schizophrenia. Science 181:179-181.
lacono WG, Tuason VB, Johnson RA (1981): Dissociation of smooth-pursuit and saccadic eye tracking in remitted schizophrenics. Arch Gen Psychiatry 38:991-996.
Kunzle H, Akert K (1977): Efferent connections of cortical area (frontal eye field) in Macaca Fascicularis. A reinvestigation using autoradiographic technique. J Comp Neurol 173:147-164.
Levin S (1984): Frontal lobe dysfunction in schizophi'enia !. Eye movement impairments. J Psychiatr Res 18:27-55.
Levin S, Jones A, Stark L, Merrin EL, Holzman PS (1982): Identification of abnormal patterns in eye movements of schizophrenic patients. Arch Gen Psychiatry 39:1125-1130.
Mallet JP, Pichot P (1981): Eye-tracking patterns in schizopbaenia, an analysis based on the incidence of saccades. Arch Gen Psychiatry 38:183-186.
Mather JA, Putchat C (1983): Motor control of schizophrenics-I. Oculomotor control of schizo- phrenics: A deficit in sensory processing, not strictly in motor control. J Psychiatr Res 17:343- 360.
Matsue Y, Okuma T, Saito H, et al (1986): Saccadic eye movements in tracking, fixation, and rest in schizophrenic and normal subjects. Biol Psychiatry 21:382-389.
Nuechterlein KH (1977): Reaction time and attention in schizophrenia: ~. critical evaluation of the data and theories. Schizophr Bull 3:373-428.
Ohtsuka K, Mukono K, Ukai K, Ishikawa S (1986): The origin of square wave jerks: Conditions of fixation and microsaccades. Jpn J Ophthalmol 30:209-215.
Parent A, Bouchard C, Smith Y (1984): the striatopallidal and striatonigral projections: Two distinct fiber systems in the primate. Brain Res 303:385.
Perlick D, Stastny P, Katz I, Mayer M, Mattis S (1986): Memory deficits and anticholinergic levels in chronic schizophrenia. Am J Psychiatry 143:230-232.
Sawaguchi T, Goldman-Rakic PS (1991): D- 1 dopamine receptors in prefrontal cortex: Involvement in working memory. Science z~ 1 :~,,~ t-~3u.
Schmid-Burgk W, Becker W, Diekmann V, Jurgens R, Kornhuber HH (1982): Disturbed smooth pursuit and saccadic eye movements in schizophrenia. Arch Psychiatr Nervenkr 232:381-389.
Selemon LD, Goldman-Rakic PS (1985): Longitudinal topography and interdigilation of cortico- striatal projection in the rhesus monkey. J Neurosci 5:776-794.
Selemon LD, Goldman-Ramie PS (1988): Common cortical and subcortical targets of the dorsolateral prefrontal and posterior parietal cortices in the rhesus monkey: Evidence for a distributed neural network subserving spatially guided behavior. J Neurosci 8:4049-4068.
Shallo-Hoffman J, Petersen J, Muhlendyck H (1989): How "normal" are square wave jerks? Invest Ophthalmol Vis Sci 30:1009-1011.
Spohn HE, Strauss ME (1989): Relation of neuroleptic and anticholinergic medication to cognitive function in schizophrenia. J Abnorm Psychol 98:367-380.
Ter~iv~iinen H, Calne DB (1980): Studies of parkinsonian movement: 1. Programming and execution of eye movement. Acta Neurol Scand 62:137-148.
Thaker GK, Nguyen JA, Tamminga CA (1989): Increased saccadic distractibility in tardive dys- kinesia: Functional evidence for subcortical GABA dysfunction. Biol Psychiatry 25:49-59.

794 BIOL PSYCHIATRY 1991:30:779-794
D.W. Hommer et al
Tune LE, Strauss MS, Lew MF (1982): Serum levels of anticholinergic drugs and impaired recent memory in clh-onic, gchizophrenic patients. Am 3 Psychiatry 139:1460-1462.
van den Bosh ILl (1984): Eye tracking impairment: Attentional and psychometric correlates in psychiatric patients. J Psychiatr Res 18:227-286.
van Opstal A J, van Gisbergen JAM (1990): Role of monkey superior colliculus in saccade averaging. Exp Brain Res 79:143-149.
Wurtz RH, Albano JE (1980): Visual-motor functions of the primate superior colliculus. Annu Rev Neurosci 3:189-226.
Yeterian EH, van Hoesen GW (1978): Cortico-striate projections in the rhesus monkey: The organization of certain cortico-caudate connections. Brain Res 139:43-63.




















