
GARCÍA LlORCA, J. ; P. CAHIZA y JP AGUILAR 2010 “Análisis zooarqueológico de los componentes...
7
Transcript of GARCÍA LlORCA, J. ; P. CAHIZA y JP AGUILAR 2010 “Análisis zooarqueológico de los componentes...


TOMO V - 1765
ANÁLISIS ZOOARQUEOLÓGICO DE LOS COMPONENTES FORMATIVOS DEL
ALERO LAS TUMANAS, VALLE FÉRTIL. INFORME PRELIMINAR
Jorge Garcia Llorca∗, Pablo A. Cahiza∗ y Juan Pablo Aguilar∗ ∗ INTRO DUCCIÓN En este trabajo presenta mos los resul tados parciales del anális is del conjunto óseo del Alero Las Tumanas (LT1), un abrigo rocoso de aproximadamente 16 m², localizado en la marge n derecha del río Las Tumanas, en la Sierra de Rivero -departa mento de Valle Fértil , en el sector oriental de la provincia de S an Juan -. LT1 forma parte de un conjunto de sitios local izados en la cuenca del río Las Tumanas integra do por áreas de sitios residenciales o multipropósi tos al aire libre, si tios de propósitos especí ficos al aire libre -molienda-, si tios con representaciones rupest res y si tios residencial es o multipropósitos en abrigos rocosos. Como result ado de nuestra interve nción en el s itio, identificamos cuatro componentes correspondientes a múlt ipl es ocupaciones realizadas por individuos pertenecientes a sociedades de baja int ensi dad de producción. Uno de ellos fue definido como componente moderno, mient ras que los tres res tantes, como componentes formativos1: 1; 2 y 3. Estos últimos fueron datados con fechados 14C de contextos de fogón de 340 ± 40 AP Cal DC 1480-1640 (p = .05) 1450 a 1650 Cal DC (p = .95) (Beta -222618, carbón, δ 13C= -24.1‰) – asociado a conjuntos cerá micos de pastas naranjas y grises con decoración pintada - ; 520 ± 40 AP Cal DC 1410-1430 (p = .05) 1320 a 1340 – 1390 a 1440 Cal DC (p = .95) (Beta -222619, carbón, δ 13C= -24.8‰ ) –en conte xtos de conjuntos cerámicos de pastas marrones y grises con decoración incisa - (Cahiza 2007) y 1720± 20 AP Cal DC 250 a 390 (p = .05) 230 a 410 Cal D C (p = .95) (Calibrado a 2 sigma con el programa INTCAL 04) (Beta -245227, carbón, δ 13C= -24.0‰) – con presencia de cerámica grabada-. El área bajo estudio se encuent ra en la ecoregión “Chaco Seco”, con la vegetación característica como es el bosque xerófi lo, compuesto por quebrachos (Schinops is y As pi dosperma ), mistol (Zi zyphus m istol), entre otros (Montero y Autino 2004:28-29). La si tuación actual de l a fauna muestra índices altos de ret racción, incluso con peligro de ext inción en muchos casos. El caso más expresivo es el guanaco (L ama guani coe) erradicado de los ambie ntes de llanura y refugia do en las zonas altas de los cordones montañosos. De la misma forma sucede con otros mamíferos y aves. En cuanto a los procesos sedimentarios que intervinieron en la deposit ación y erosión de LT1, debemos tener en cuenta que durante e ven tos hidrológicos extremos o ext raordinarios, las inundaciones y aluviones en ríos de montaña no solo se relacionan con la ele vada al ime ntación líquida, sino ta mbién con el excesivo ingreso de sedimentos provenie ntes de sus numerosos afluentes, generalmente en forma de deslizamientos y flujos de detritos. La dimensión del paquete sedimentario parece ser el producto de fuerte s eventos agregativos, como lo demuestra los 2-2 ,5 m de acumulación e n un periodo de aproximadamente 1.700 años. En este sentido el es t rato o capa E que separa los componentes format ivos 1 y 2, de alrededor de 30 cm es uno de los más claros ej emplos ya que se trata de un nivel arenoso de deposi tación “normal” (grueso arriba, fino abajo) que responde a fases de relleno de playa de fenómenos aluviales intensos, comunes en ríos con áreas de captación serrana como Las Tuma nas. Otro componente importante de la p ila sedimentaria de LT1 está constituido por bloques de diverso tamaño –con un ra ngo que abarca desde rocas de más de 1m³ hasta clas tos pequeños-. Esta si tuación no se limita al área dentro de la línea de goteo, sino que es extensible al sec tor externo del alero y también la observamos e n toda la pared del cerro . Creemos que el flujo de sedimentos por gravedad, especialmente caídas de r ocas y deslizamientos de rocas , constituye el proceso sedimentario activo primario (B lair y McPherson 1994). Las raíces vegetales también han transformado algunas de las condiciones de deposit ación del sitio, especialmente en
* INCIHUSA-CONICET / FFyL-UNCuyo g llorca@ mendoza-conicet.gob.ar pcahiza@ mendoza-conic et.gov.ar ∗ ∗INCIH USA-CONICET [email protected]

TOMO V - 1766
Hemos regi strado la presencia de ele mentos óseos de roedores y ede ntados, sin embargo no detectamos galerías, túnele s ni madrigueras e n ninguno de los niveles de LT1. Aunque he mos observado huell as de roído de elementos óseos . El espacio cultivado colindante con el s itio y con el río presentó gra n cantidad de cuevas de roe dores. LT1 tiene ópt imas condiciones de acceso y entre la fauna local abundan los zorros y felinos, a pesar de ello no hemos encontrado rasgos positivos de su actividad (madrigueras, deyecciones, etc), aunque si, especímenes óseos con y sin alteración térmica de esas especies autóctonas . Esto último incluye la presencia de serpientes, muy probablemente Boa const rictor occidentalis . En tiempos recientes el alero ha servido como lugar alternativo de pernocte (especialme nte ante inclemencias climáticas) para el ganado caprino. En la superficie actual del alero se encontraron restos de t res cubiertas de automóvi les quemadas. S egún uno de los propietarios es to correspondió a un evento puntual para ahuyentar serpientes del abrigo. Este accionar ha ennegrecido paredes y techo del alero. El emplazamie nto de cuatro entierros en el sector oeste del alero (componente formativo 2) impl icó la remoción de sedimentos para el cavado de las tumbas, constatado por el registro de fosas (enti erros 1 y 2). Aunque este proceso constituye la mayor perturbación de LT1, creemos que el registro manti ene buenas condiciones de integridad. El remontaje de fragmentos cerámicos (n=18) no sólo aportó información de índole tecnológica s i no también tafonómica, en ta nto pudimos comprobar que éstos se pudieron realizar sólo entre elementos localizados en una misma unidad est ratigráfica. MATERIA LES Y MÉTOD OS El análisis tuvo en cuenta la diferenciación en los componentes es tablec idos, así como su consideración global. El contexto de excavación reveló sectores con ma yor acumulación de restos óseos , en volúmenes sedimentarios diferenciados , s iendo el componente formativo 2 el de mayor profundidad estratigráfica. Las excavaciones poseen una extensión cercana a los 16m², dis tribuidos 10m² dentro de la línea de goteo y 6m² fuera de el la. N ues tra intervención incidió ha sta 250cm de profundidad –sólo en las cuadríc ulas G 3 y E1-, debido a la gran cantidad de bloques, que han impedido la continuación de l a excavación. Se tuvo en cuenta la bibl iografía disponible a ctualmente para el anális is zooarque ológico en lo concerniente a los aspectos teóricos y metodológicos (Mengoni Goñalons 1988, 1999, 2009; Lyman 1994, 2008; Reitz and W ing 1999, entre otros). En la determinación de las categorías anatómicas y taxonómicas, se usaron manuales y at las específicos (Barone 1990; Galota y Galota 1988), as í como las colecciones osteológicas que posee la Unidad de Antropología (INCIHUS A). En el presente artículo indicamos la abunda ncia taxonómica y cuantificamos los resultados sobre la base del NISP y MN E, además del peso de cada resto óseo. Derivamos algunos cocientes de estas unida des de análi sis, con el objeto de obtener tendencias t afonómicas en relación con los componentes es tablecidos y del sit io en su conjunto. También analizamos la R iqueza Taxonómica (N TAX A) y los Índice s de Divers idad y de Equi tatividad (Lyman 2008, Mengoni Goñalons 2009). En la ident ifi cación si stemática del conjunto óseo se util izó bibliogra fía específica, y en particula r Montero y Autino (2004). Debido al impacto de la fra gmentación en la identificación de los especímenes óseos, recurrimos a diferenciar dis tintas ca tegorías tafonómicas en relación con el nivel de ident ifi cación. En el caso de los mamíferos, fueron divididos en comparación con el peso de especies vivientes: los ma mí feros grandes (Mamalia grande) e n correspondencia con el tamaño del guanaco (L am a guanicoe), con un peso aproximado entre 60 y 100 kg.; para los mamíferos medianos (Mamalia media no), con los ovinos (Ovis arie s)-caprinos (Capra hircus) o venados (Cervidae), y peso entre 5 y 45 kg.; en el caso de los mamíferos pequeños (Mamal ia pequeño) con la liebre (Lepus sp.), el gato montés (Oncifeli s geoffroyi), incluyendo especies de menor tamaño, tal el ca so de los roe dores en general y pesos de 5 kg. y menores. U na cate goría menos resolutiva incluye mamíferos (Mammalia) que de ma nera general , cubrieron res tos óseos de las dos categorías de mayor porte. En un escalón más bajo de identi fi cación, incluimos aquellos casos de difí cil determinación que en general cubren un rango de tamaño pequeño, sean mamíferos o aves. En el caso particular de las Aves, fueron consideradas de manera global, sin tipificación del tamaño, aunque en su mayoría se rela cionaron con aquel las aves de tamaño más pequeño, por ejemplo las martinetas (Eudromia elegans). F inalmente la categoría con menor rango de identificación, incluyeron aquellos res tos sin posibil idad de identificación anatómica o taxonómica (NI)
TOMO V - 1767
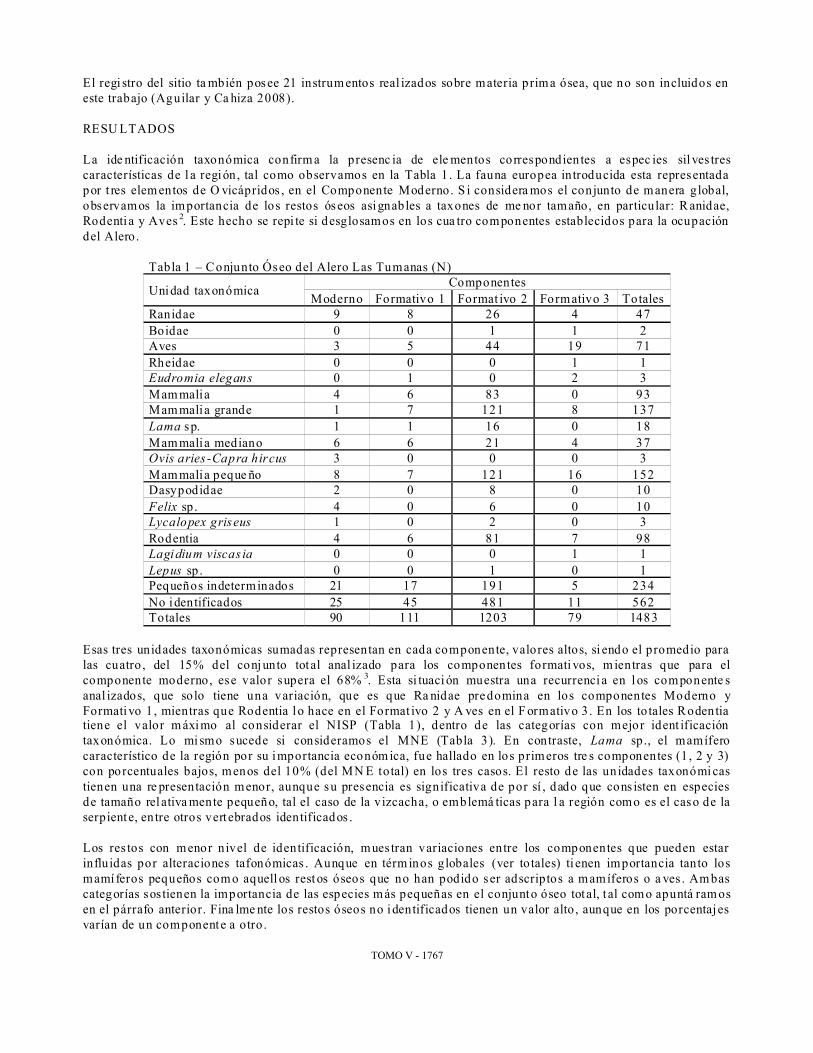
El regi stro del sitio ta mbién posee 21 instrum entos real izados sobre materia prima ósea, que no son incluidos en este trabajo (Aguilar y Ca hiza 2008). RESU LTADOS La ide ntificación taxonómica confirm a la presenc ia de ele mentos correspondien tes a espec ies sil ves tres características de l a región, tal como observamos en la Tabla 1 . La fauna europea introducida esta representada por t res elem entos de O vicápridos , en el Componente Moderno. S i considera mos el conjunto de manera global, observamos la importancia de los restos óseos asignables a taxones de me nor tamaño, en particular: R anidae, Rodenti a y Aves 2. Este hecho se repi te si desglosamos en los cua tro componentes establecidos para la ocupación del Alero.
Tabla 1 – C onjunto Óseo del Alero Las Tumanas (N)
Unidad taxonómica Componentes Moderno Formativo 1 Format ivo 2 Formativo 3 Totales
Ranidae 9 8 26 4 47 Boidae 0 0 1 1 2 Aves 3 5 44 19 71 Rheidae 0 0 0 1 1 Eudromia elegans 0 1 0 2 3 Mam malia 4 6 83 0 93 Mam malia grande 1 7 121 8 137 Lama sp. 1 1 16 0 18 Mam malia mediano 6 6 21 4 37 Ovis aries -Capra hircus 3 0 0 0 3 Mam malia peque ño 8 7 121 16 152 Dasypodidae 2 0 8 0 10 Felix sp . 4 0 6 0 10 Lycalopex griseus 1 0 2 0 3 Rodentia 4 6 81 7 98 Lagi dium viscas ia 0 0 0 1 1 Lepus sp . 0 0 1 0 1 Pequeños indeterminados 21 17 191 5 234 No identificados 25 45 481 11 562 Totales 90 111 1203 79 1483
Esas tres unidades taxonómicas sumadas representan en cada componente, valores altos, si endo el promedio para las cuatro , del 15% del conj unto tot al anal izado para los componentes formati vos, m ientras que para el componente moderno, ese valor supera el 68% 3. Esta si tuaci ón muestra una recurrenci a en l os componente s anal izados, que solo tiene una variación, que es que Ra nidae predomina en los componentes Moderno y Formativo 1 , mientras que Rodentia l o hace en el Format ivo 2 y A ves en el F ormativo 3. En los to tales R odentia tiene el valor m áximo al considerar el NISP (Tabla 1), dentro de las categorías con mejor ident ificación taxonómica. Lo mi smo sucede si consideramos el MNE (Tabla 3). En contraste, Lama sp ., el mamífero característico de la región por su importancia económ ica, fue hallado en los primeros tre s componentes (1 , 2 y 3) con porcentuales bajos, menos del 10% (del MN E total) en los tres casos. El resto de las un idades taxonómi cas tienen una re presentación menor, aunque su presencia es significativa de por sí , dado que cons isten en especies de tamaño rel ativa mente pequeño, tal el caso de la vizcacha, o emblemá ticas para l a región como es el caso de la serpiente, en tre otros vert ebrados identificados . Los res tos con menor nivel de identificación, mues tran variaciones entre los componentes que pueden estar influidas por alteraciones tafonómicas . Aunque en térm inos globales (ver totales) ti enen importancia tanto los mamí feros pequeños com o aquell os rest os óseos que no han podido ser adscriptos a mamíferos o a ves . Am bas categorías sostienen la importancia de las especies m ás pequeñas en el conjunto óseo tot al, t al como apuntá ram os en el párrafo anterior. Fina lme nte los restos óseos no identificados tienen un valor alto , aunque en los porcentaj es varían de un componente a otro .
TOMO V - 1768
Al discriminar el conjunto observamos que el Componente Moderno tiene com o dato interesante la presencia de especímenes identificados com o ovicápridos , Felix sp. y L am a sp ., entre un conjunto relativamente esca so, con predominio de taxones de tam año pequeño. De acuerdo a los índices de Abundancia R elativa (T abla 2), y en relación con los otros componentes, la muestra presenta una distribución más pareja (“even distribution”) en la abundancia de las d iferentes categorías taxonóm icas (Mengoni Goñalons 2009). El porcentaje de los restos óseos no identi ficados alcanza el 28% , mientras que el de alteración térmic a es bajo , alcanzando una media menor al 14% del total del conjunto . A su vez el Componente F ormativo 1, t ambién se caracteriza por ser una muestra pequeña, donde predominan las especies de menor port e y con presencia míni ma de camélidos y martinetas, y, a su vez, es el que tiene menor riqueza taxonómica (NTAXA) de lo s cuatro conjuntos (Tabla 2). Mientras que el Índice de Divers idad, es el que expresa una relación menos pareja de los cuatro com ponentes cons iderados. Los restos óseos con alteración térmica son poco más del 12% del to tal de la muestra.
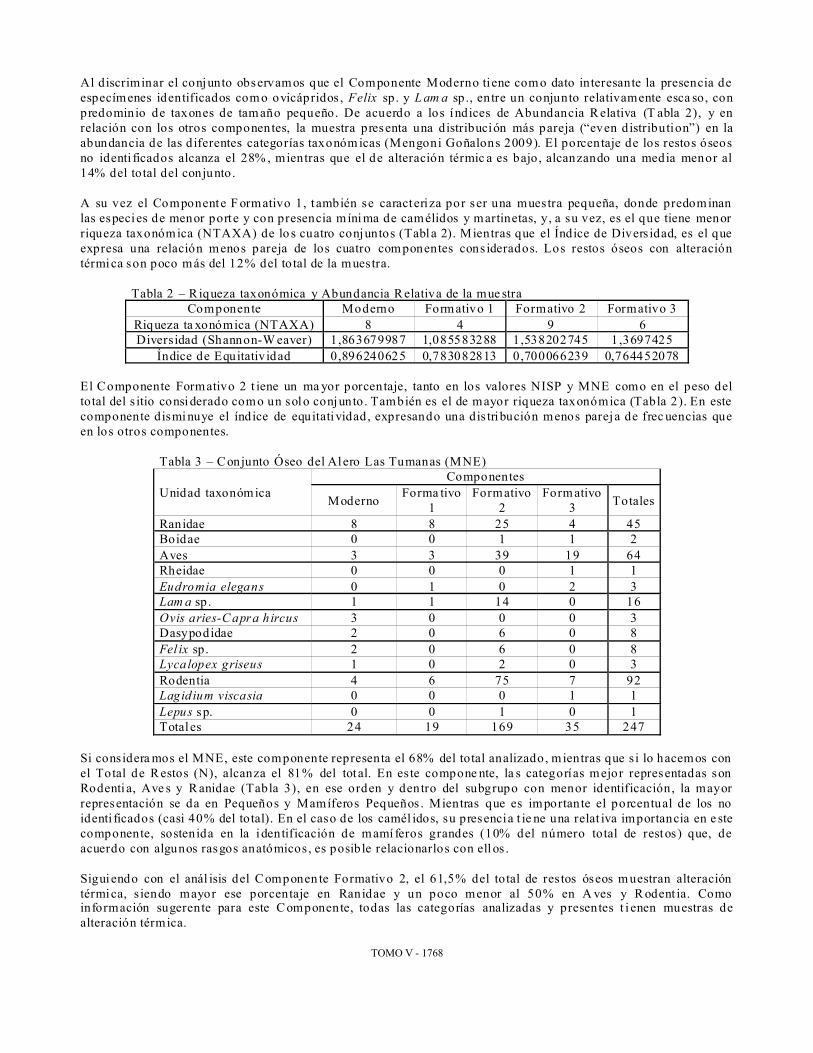
Tabla 2 – Riqueza taxonómica y Abundancia Relativa de la mue stra Componente Moderno Formativo 1 Formativo 2 Formativo 3
Riqueza ta xonómica (NTAXA) 8 4 9 6 Diversidad (Shannon-W eaver) 1,863679987 1,085583288 1,538202745 1,3697425
Índice de Equitativ idad 0,896240625 0,783082813 0,700066239 0,764452078 El Componente Formativo 2 t iene un ma yor porcentaje, tanto en los valores NISP y MNE como en el peso del total del s itio consi derado como un solo conjunto . También es el de mayor riqueza taxonómica (Tabla 2). En este componente dismi nuye el índice de equitatividad, expresando una dis tribución menos pareja de frec uencias que en los otros componentes.
Tabla 3 – Conjunto Óseo del Alero Las Tumanas (MNE)
Unidad taxonómica Componentes
Moderno Forma tivo 1
Formativo 2
Form ativo 3 Totales
Ranidae 8 8 25 4 45 Boidae 0 0 1 1 2 Aves 3 3 39 19 64 Rheidae 0 0 0 1 1 Eudromia elegans 0 1 0 2 3 Lam a sp . 1 1 14 0 16 Ovis aries-C apra h ircus 3 0 0 0 3 Dasypodidae 2 0 6 0 8 Fel ix sp. 2 0 6 0 8 Lyca lopex griseus 1 0 2 0 3 Rodentia 4 6 75 7 92 Lagid ium viscasia 0 0 0 1 1 Lepus sp. 0 0 1 0 1 Totales 24 19 169 35 247
Si considera mos el MNE, este componente representa el 68% del total analizado, mientras que s i lo hacemos con el Total de Restos (N), alcanza el 81% del tot al. En este compone nte, la s categorías mejor representadas son Rodenti a, Ave s y Ranidae (Tabla 3), en ese orden y dentro del subgrupo con menor identificación, la mayor representación se da en Pequeños y Mamíferos Pequeños . Mientras que es importante el porcentual de los no identi ficados (casi 40% del total). En el caso de los camél idos, su presencia t ie ne una relat iva importancia en e ste componente, sostenida en la identificación de mamí feros grandes (10% del número total de restos) que, de acuerdo con algunos rasgos anatómicos, es posible relacionarlos con ell os . Siguiendo con el anál isis del Componen te Formativo 2, el 61,5% del total de res tos óseos m uestran alteración térmica, s iendo mayor ese porcentaje en Ranidae y un poco menor al 50% en A ves y R odent ia. Como información sugerente para este Componente, todas las categorías analizadas y presentes t i enen muestras de alteración térmica.

TOMO V - 1769
El Componente Formativo 3, posee una riqueza taxonóm ica relativame nte baja (Tabla 2), s iendo las A ves las que sobresalen en la identificación en rel ación con el tot al analizado. En cuanto a la alte ración térmica, casi el 57% alcanzó esa categoría s i consideramos el conjunto total, mientras que en Aves, poco más del 15%. En particular e l anális is de los camélidos, que están representados en los Componentes Moderno y Formativo 1 y 2 -la mayoría de ellos se enc ontraron en el último (Tabla 1)-, permi te es tablecer que salvo dos especímenes atribuibles a vértebras torácicas y un espécimen relac ionado con la pelvis , el res to se correspondieron con manos y pies, es decir a las extremidades. Esta particularidad del regis tro del Alero Las Tumanas, está de acuerdo con los resultados observados en otros contextos arqueológicos regionales (Garcia Llorca y Cahiza 2007: 127). Por último el 68% de la muestra de caméli dos en el Componente Formativo 2, tiene al teración térmica y dos fal anges sin fus ionar, sobre un total de diez. El conjunto arqueofaunís tico analizado tiene una tendencia clara en la representación de especies pequeñas e n los cuatro componentes determinados. Por otra part e, los procesos tafonómicos actúan sobre estos conjuntos, de una ma nera diferente en relaci ón con las especies de mayor porte. De hecho, el índice de fragmentación estipulado por el cociente N ISP /MNE tiende a ser bajo, con un promedio cercano a uno. El Cociente entre el peso y e l número de restos , considerado e n promedio para cada componen te, visualiza valores muy bajos, ligeramente mayor a uno, siendo el componente moderno el de mayor valor (1,4), influido por el tamaño de la muestra y por la presencia de especímenes pertenecientes a ovi cápridos. Sin embargo el grado de fragmentación que presentaron los res tos óseos ha sido importante y difi cul ta la tarea de identificación anatómica y ta xonómica. CO NSIDERACIONES FINA LES En este informe preliminar el análi sis permite anticipar el significativo aporte a l a d ieta, de especies de tamaño pequeño (la mayoría corresponden a recursos r -Hayden 1990:33, ver tam bién Garcia Llorca y Cahiza 2007 -). Esta a severaci ón se muestra documentada no solo por el anál isis zooarqueológico específico, sino ade más por la información contextual, que se genera en la propia excavación. La contrastación con los fechados radiocarbónicos pe rmite efectuar una asociación directa con los componentes format ivos establecidos y la recurrencia de determinadas especies e n el consumo de los habi tantes del alero. En part icular los elementos anatómicos hallados en la s categorías Aves, Rodentia y Ranidae, así como la importancia de la al teraci ón térmica en esos restos óseos, indican una tendencia clara e n cuanto al consumo y el aprovechamiento de recursos de acceso relativame nte fác i l y de óptima disponibilidad. La muestra arqueofaunísti ca se complementa con abundante s fragmentos de cáscaras de huevo, principalmente de Rheidae, además de placas de Edentados, en ambos casos con alta incidencia de alteración térmica. En el caso de los camélidos, aunque su presencia es escasa, no deja de ser importante en el Componente Format ivo 2. El anális is de la muestra arqueofa unística permite es tablecer y contras tar los modos de vida sucedidos en el Alero La s Tumanas, con el res to de l a informació n arqueológica disponible. Los estudios actuales sobre estos conjuntos están dirigidos a analizar la influencia de la alteración térmica, así como profundizar el de marcas y huella s, además de discrim ina r algunos de los taxone s me nos inclusivos anal izados, en particular el de las Aves. AGRADECIMIENTOS Este trabajo se enmarca en los proyectos PIP CONICET y P ICT (ANPCyT) di rig idos por J . Roberto Bárcena. NOTAS 1 El término Formativo lo utiliza mos de una manera extensiva para posibil itar el diálogo disci plinar a nivel regional, y tiene como objeto caract erizar a las sociedades que habitaron durante este tiempo y espacio, a las cuales podríam os definir como de “baja intensidad de producción”. 2 En caso de los edentados , las placa s del caparazón no fueron incorporadas al conteo de los especímenes o elementos óseos. 3 Si consideramos el MNE, ese valor es superior al 80%.

TOMO V - 1770
REF ERENC IAS BIB LIOG RÁF IC AS
Aguilar, JP y P Ca hiza. 2008. Anál isis de la T ecnología ósea del Alero Las Tumanas”. V Taller Internacional de Zooarqueología de Camélidos Sudamericanos y Prim er Congreso Nacional de Zooarqueología Argentina. Malargüe. B arone, R.1990. Anatom ía comparada de l os mamíferos domésti cos . Tomo I. Osteología, Parte II: atlas, fascículo II. Buenos Aires . Cahiza, P. 2007. Las sociedades formativas tardías de Valle Fértil, San Juan. Comechingonia 10: 79-93. Galotta, D. R . y J . M. G alot ta. 1988. Os teología del llama (Llama glam a guanicoe f.d. glama Linnaeus, 1758). Revista de Ciencias Agrarias y Tecnología de Alimentos 9 (1-4):19-61. Garcia Llorca, J . y P . A. C ahiza. 2007. Aprovechamiento de Recursos Fa unístic os e n las Lagunas de Guanacache (Mendoza, Arge ntina). Anális is Zooarqueológico de La Empozada y Al tos de Melién II. Chungara 39 (1): 117-133. Hayden, B. 1990. Nimrods, piscators , pluckers , and planters: the emergence of food production . Journal of Anthropological Archaeology 9:31-69. Lyman, R. L. 1994. Vertebrate Taphonomy. Cambridge, Cambridge U niversity Press . -----. 2008. Quantitative Paleozoology. Cambridge, Cambridge Universi ty P ress. Mengoni Goñalons , G . L.1988. Anális is de materiale s fa uníst icos de si tios arqueológicos. Xam a I: 71-120. -----. 1999. Cazadores de guanacos de la estepa patagónic a. Sociedad Argentina de Antropología, B uenos Aires. -----. 2009. Zoología en la práctica: algunos temas metodológicos. Xama . En prensa. Montero, R. y A. Autino. 2004. Sist emát ica y Filogenia de los Vert ebrados. Universidad Nacional de Tucumán Rei tz, E. J. and E. S. W ing. 1999. Zooarchaeology. C ambridge, Cambridge Univers ity Press.




















