
Functional Morphology of the Female Genital Organs in the Peruvian Red Uakari Monkey (Cacajao Calvus...
10
RESEARCH ARTICLE Functional Morphology of the Female Genital Organs in the Peruvian Red Uakari Monkey (Cacajao Calvus Ucayalii)* PEDRO MAYOR 1,2 *, MARK BOWLER 2,3,4 , AND CARLOS LÓPEZ‐PLANA 1 1 Department of Animal Health and Anatomy, Faculty of Veterinary, Universitat Autònoma de Barcelona, Bellaterra, Barcelona, Spain 2 YAVACUS, Yavarí Conservación y Uso Sostenible, Iquitos, Peru 3 San Diego Zoo Global Institute for Conservation Research, Behavioural Biology Unit, Escondido, California 4 Centre for Social Learning and Cognitive Evolution, School of Psychology, University of St. Andrews, Scotland, United Kingdom Functional morphology may provide important information that could improve methodologies for the diagnosis of the reproductive phase of females, and develop assisted breeding techniques for wildlife. This study examined features of genital organs in 19 Peruvian red uakari monkey (Cacajao calvus ucayalii) females in different reproductive stages, collected from wild animals hunted for food by rural communities in the North‐eastern Peruvian Amazon, in order to provide knowledge on the reproductive physiology of this species. The observed mean ovulation rate was 1.4 follicles, and the observed maximal follicle diameter was 0.8 cm. After ovulation, the matured follicle luteinizes resulting in functional CL. In case of oocyte fertilization, the pregnancy CL grows to a maximum of 1.2 cm in diameter, and luteal volume per female decreases related to the advance of pregnancy. Pregnant females also present follicular activity until late pregnancy, but non‐ovulated follicles do not undergo atretic processes and apparently transform to accessory CL, resulting in a contribution of 30% of the total luteal volume. All pregnant females delivered a single fetus at term, resulting in a rate of reproductive wastage of 20% of oocytes or embryos. The endometrium and the endometrial glands in non‐pregnant females in the follicular phase show a significant increase related to the follicular growth, reaching a high proliferation in non‐pregnant females in the luteal phase. The red uakari monkey showed different vaginal epithelium features in accordance with the reproductive state of the female, suggesting that vaginal cytology could be a successful methodology with which to characterize the estrous cycle of this species. The present reproductive evaluation of the Peruvian red uakari monkey provides important information that could improve the development of assisted reproductive techniques in non‐human primates. Am. J. Primatol. 75:545–554, 2013. © 2013 Wiley Periodicals, Inc. Key words: red uakari; reproduction; Cacajao calvus; ovary; tubular genital organs INTRODUCTION The Peruvian red uakari monkey (Cacajao calvus ucayalii) is a medium‐bodied platyrrhine considered Vulnerable by the IUCN [Veiga et al., 2008]. Uakari monkeys (genus Cacajao) generally occur at low densities and have patchy distributions [Bowler et al., 2009; Heyman & Aquino, 2010], and their populations may be apparently declining due to hunting and habitat loss [Puertas & Bodmer, 1993]. Functional morphology of the reproductive organs is a key component for the better understand- ing of reproductive patterns useful in maximizing reproductive efficiency in captivity and to develop assisted breeding techniques. However, assisted breeding techniques that are routine in domesticated species are not easily adapted to wildlife [Andrabi & Maxwell, 2007]. One of the major problems with the implementation of in situ and ex situ conservation programs is the lack of availability of the biological material which is required for a better understand- ing of reproductive patterns [Pukazhenthi & Wildt, 2004]. Since the red uakari is difficult to breed in captivity [Fontaine, 1981] and because there are few data available on reproductive rates in wild Contract grant sponsor: Universitat Autònoma de Barcelona; Contract grant sponsor: The Dirección General de Fauna y Flora Silvestre (DGFFS) of Perú. The authors have no conflict of interest to declare. * Correspondence to: Pedro Mayor, Department of Animal Health and Anatomy, Universitat Autònoma de Barcelona, E‐ 08193, Bellaterra, Spain. E‐mail: [email protected] Received 21 November 2012; revised 23 December 2012; revision accepted 7 January 2013 DOI 10.1002/ajp.22132 Published online 25 February 2013 in Wiley Online Library (wileyonlinelibrary.com). American Journal of Primatology 75:545–554 (2013) © 2013 Wiley Periodicals, Inc.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Functional Morphology of the Female Genital Organs in the Peruvian Red Uakari Monkey (Cacajao Calvus...

RESEARCH ARTICLE
Functional Morphology of the Female Genital Organs in the Peruvian RedUakari Monkey (Cacajao Calvus Ucayalii)*
PEDRO MAYOR1,2*, MARK BOWLER2,3,4, AND CARLOS LÓPEZ‐PLANA1
1Department of Animal Health and Anatomy, Faculty of Veterinary, Universitat Autònoma de Barcelona, Bellaterra,Barcelona, Spain2YAVACUS, Yavarí Conservación y Uso Sostenible, Iquitos, Peru3San Diego Zoo Global Institute for Conservation Research, Behavioural Biology Unit, Escondido, California4Centre for Social Learning and Cognitive Evolution, School of Psychology, University of St. Andrews, Scotland, United Kingdom
Functional morphology may provide important information that could improve methodologies for thediagnosis of the reproductive phase of females, and develop assisted breeding techniques for wildlife.This study examined features of genital organs in 19 Peruvian red uakari monkey (Cacajao calvusucayalii) females in different reproductive stages, collected from wild animals hunted for food by ruralcommunities in the North‐eastern Peruvian Amazon, in order to provide knowledge on the reproductivephysiology of this species. The observedmean ovulation rate was 1.4 follicles, and the observedmaximalfollicle diameter was 0.8 cm. After ovulation, the matured follicle luteinizes resulting in functional CL.In case of oocyte fertilization, the pregnancy CL grows to a maximum of 1.2 cm in diameter, and lutealvolume per female decreases related to the advance of pregnancy. Pregnant females also presentfollicular activity until late pregnancy, but non‐ovulated follicles do not undergo atretic processes andapparently transform to accessory CL, resulting in a contribution of 30% of the total luteal volume. Allpregnant females delivered a single fetus at term, resulting in a rate of reproductive wastage of 20% ofoocytes or embryos. The endometrium and the endometrial glands in non‐pregnant females in thefollicular phase show a significant increase related to the follicular growth, reaching a high proliferationin non‐pregnant females in the luteal phase. The red uakari monkey showed different vaginalepithelium features in accordance with the reproductive state of the female, suggesting that vaginalcytology could be a successful methodology with which to characterize the estrous cycle of this species.The present reproductive evaluation of the Peruvian red uakarimonkey provides important informationthat could improve the development of assisted reproductive techniques in non‐human primates. Am. J.Primatol. 75:545–554, 2013. © 2013 Wiley Periodicals, Inc.
Key words: red uakari; reproduction; Cacajao calvus; ovary; tubular genital organs
INTRODUCTION
ThePeruvian red uakarimonkey (Cacajao calvusucayalii) is a medium‐bodied platyrrhine consideredVulnerable by the IUCN [Veiga et al., 2008]. Uakarimonkeys (genus Cacajao) generally occur at lowdensities and have patchy distributions [Bowleret al., 2009; Heyman & Aquino, 2010], and theirpopulations may be apparently declining due tohunting and habitat loss [Puertas & Bodmer, 1993].
Functional morphology of the reproductiveorgans is a key component for the better understand-ing of reproductive patterns useful in maximizingreproductive efficiency in captivity and to developassisted breeding techniques. However, assistedbreeding techniques that are routine in domesticatedspecies are not easily adapted to wildlife [Andrabi &Maxwell, 2007]. One of the major problems with theimplementation of in situ and ex situ conservation
programs is the lack of availability of the biologicalmaterial which is required for a better understand-ing of reproductive patterns [Pukazhenthi &Wildt, 2004]. Since the red uakari is difficult to breedin captivity [Fontaine, 1981] and because there arefew data available on reproductive rates in wild
Contract grant sponsor: Universitat Autònoma de Barcelona;Contract grant sponsor: The Dirección General de Fauna y FloraSilvestre (DGFFS) of Perú.The authors have no conflict of interest to declare.
*Correspondence to: Pedro Mayor, Department of AnimalHealth and Anatomy, Universitat Autònoma de Barcelona, E‐08193, Bellaterra, Spain. E‐mail: [email protected] 21 November 2012; revised 23 December 2012;revision accepted 7 January 2013DOI 10.1002/ajp.22132Published online 25 February 2013 in Wiley Online Library(wileyonlinelibrary.com).
American Journal of Primatology 75:545–554 (2013)
© 2013 Wiley Periodicals, Inc.

populations [Bowler & Bodmer, 2009; 2011; Bowleret al., 2013], the reproductive biology of the reduakari is practically unknown. This study improvesthe reproductive knowledge of the Peruvian reduakari monkey, characterizing the functionality ofthe female genital organs.
METHODSStudy Area
This study was conducted on the Yavari‐MirínRiver, in the Northeastern Peruvian Amazon. Thestudy area spans 107,000 ha of continuous forest,predominantly non‐flooding terra firme forest. Theclimate in the region is typically equatorial with anannual temperature from 22 to 36°C, a relativehumidity from 80% to 100%, and an annual rainfallfrom 1,500 to 3,000 mm. Seasons are defined as dry(January–February and July–September) and wet(March–June and October–December).
Animal Collection and Sample AnalysisBetween2005and2012, hunters living in the area
where the study was conducted collected genitalorgans from 19 adult Peruvian red uakari monkeyfemales, as part of an ongoing participatory conserva-tion program that involves local hunters in imple-menting community‐based wildlife management.The research protocol was approved by the ResearchEthics Committee for Experimentation in Wildlife atthe Dirección General de Flora y Fauna Silvestrefrom Peru (0229‐2011‐DGFFS‐DGEFFS). No animalswere killed specifically for the research and hunterswere not paid for the sample collection. The hunterswere trained to remove all the abdominal andpelvic organs complete with the perineal region toavoid damage to the material. Researchers wereresponsible for the dissection of the reproductiveorgans.
The genital organs of adult females were exam-ined for evidence of embryos or fetuses. Females withone embryo or fetus were considered pregnant, andthe pregnancy stagewas defined as embryonic or fetal[International Committee on Veterinary Embryolog-ical Nomenclature, 1994]. Non‐pregnant adult ani-mals with ovaries containing active cyclic corporalutea (CL) were described as being in the luteal phaseof the estrous cycle, while females with ovariesbearing large antral follicles and lacking cyclic CLwere considered to be in the follicular phase of theestrous cycle. In the absence of either large antralfollicles or CL, the ovaries were considered inactive.
The macroscopic features of the fixed genitalorgans were analyzed in all the experimentalanimals. Ovarian sizes (r1, r2, and r3) were calculatedand the ovarian volume estimated by means of theformula for the volume of ovoid bodies: V ¼ (4/3)p(r1r2r3). Measurements of tubular organs includedmean outer diameter and length of the uterine tubes,
uterine body, cervix, vagina, and size of the externalurogenital opening. All measurements were deter-mined in three fields per tubular organ and female.
Samples of the reproductive organs were dehy-drated and embedded in paraffin wax. Ovaries wereserially sectioned at 3 mm sections along the longitu-dinal axis. Every sixth sectionwasmounted on a glassmicroscope slide. Three sections from the tubularorgans of each female were mounted onto a glassmicroscope slide. Periodic acid‐Schiff (PAS) stain wasused to stain connective tissues, mucus and basallaminae, and hematoxylin or Masson’s trichromestains were used to distinguish collagen, keratine,and muscle fibers. Sections were examined under alight microscope.
Ovarian CL were considered active after apregnancy was established or as indicated by lutealcell morphology. Depending on the diameter, viabili-ty of luteal cells, the presence or absence of an oocyte,and the reproductive state of females, CL wereclassified as cyclic CL, pregnancy CL, accessory CL,and atretic CL [Mayor et al., 2013].
Based on the number of pregnancy CL, wedetermined the ovulation rate, expressed as thenumber of CL per female with ovulations. Wecalculated the rate of ovum or embryo mortality asthe difference between the number of CL and theobserved embryos or fetuses.
Measurements of antral follicles and CL weretakenusing amicrometric ocular. Diameters (D) weremeasured as the mean length of the two maximalperpendicular axes. Luteal volumes were calculatedusing the formula: V ¼ (4/3)p(D/2)3. Recorded ovari-an variables were number and diameter of antralfollicles, and number, diameter, volume and type ofCL. Luteal volume per female was calculated as thesum of volumes from active luteal structures.
Recorded tubular organ variables were micro-scopic features and measurements of the epithelium,the tunica mucosa and the tunica muscularis of theuterine tubes and uterus, and epithelium thicknessand number of cell layers of the vaginal epithelium.The density of endometrial glands wasmeasured andranked from 1 to 5, according the average number ofglands countered in random 1‐mm2
fields at 100�.Significance scoring of density of endometrial glandswas 1 (0–5 glands/mm2), 2 (5–10 glands/mm2), 3 (10–15 glands/mm2), 4 (15–20 glands/mm2), and 5 (20–25 glands/mm2). Cornification of the vaginal epithe-lium was calculated as the thickness of the cornifiedlayers respect to the total epithelial thickness in eachfemale. These features were correlated to thereproductive state of the females. All variableswere determined in five randomly selected fields.
Diameters (D) of the materno‐fetal attachmentwere measured as the mean length of the twomaximal perpendicular axes, and the area of thematerno‐fetal attachment was calculated using theformula: V ¼ (4/3)p(D/2)3.
Am. J. Primatol.
546 / Mayor et al.

Statistical Analysis
Assessments of the relationships between mac-roscopic and microscopic features of the femalegenital organs and reproductive state were testedby ANOVA. Differences between means were testedusing the Tukey–Kramer multiple comparisons test.We used linear regression and Pearson’s correlationin order to determine the relationships between thelargest follicle and number of antral follicles, betweenthe largest follicle and density of glands of endome-trium, between fetal length and total luteal volume,and between fetal length and area of the materno‐fetal attachment. Statistical analyses were per-formed usingGraphPadPrism5 (GraphPad SoftwareInc., San Diego, CA: www.graphpad.com). Differ-ences with a probability value of 0.05 or less wereconsidered significant. All values are expressed asthe mean � standard deviation (SD).
All research conducted was in compliance withAmerican Society of Primatologists’ guidelines for theethical treatment of non‐human primates and ad-hered to the legal requirements of Perú.
RESULTS
Of the 19 adult Peruvian red uakari monkeyfemales included in this study, 9 were active non‐pregnant females (4 in the follicular and 5 in theluteal phase), and 6 were pregnant females at
different stages of pregnancy (2 at the embryonicand 4 at the fetal stage of pregnancy). Four inactivefemales lacked large antral follicles and CL.
OvariesThe creamy white ovaries of adult red uakari
monkey females were ovoid bodies with a smoothsurface. The ovarian size did not show significantdifferences related to the reproductive state and theside of the body (Table I). Only large antral follicleswere visible as transparent bodies but did notprotrude above the surface.
Ovarian follicles were observed in both pregnantand non‐pregnant females. Non‐pregnant females inthe follicular phase showed a higher number of antralfollicles >2 mm (F3,211 ¼ 21.612; P < 0.0001) and alarger antral‐follicle diameter (F3,211 ¼ 30.926;P < 0.0001) than other females (Table II). Thelargest antral follicle observed had a diameter of8.06 mm. Non‐pregnant females in the follicularphase showed a non‐statistical decrease in thenumber of antral follicles related to the largest antralfollicle diameter (r2 ¼ 0.446; P ¼ 0.332). Females inthe luteal phase had fewer antral follicles than otherfemales (F3,211 ¼ 28.892; P < 0.0001). Atretic fol-licles and thecal‐type interstitial tissue were widelypresent in the ovary.
The CL did not protrude above the surface,presented a very lobed appearance and was
TABLE I. Average Volume of Ovaries in the Red Uakari (N ¼ 19)
n Left ovary (cm3) Right ovary (cm3)
Inactive females 4 0.50 � 0.71 0.67 � 0.94Non‐pregnant females in the follicular phase 4 3.13 � 1.93 3.52 � 1.87Non‐pregnant females in the luteal phase 5 4.48 � 2.76 3.56 � 1.41Pregnant females 6 3.59 � 4.78 3.54 � 4.33
F3,15 ¼ 1.207; P ¼ 0.341 F3,15 ¼ 1.134; P ¼ 0.367
Differences in ovarian volume among females in different reproductive states were tested using 1‐way ANOVA and Tukey–Kramer multiple comparisonstest.
TABLE II. Follicular Characterization According to Reproductive Status of the Female Red Uakari (N ¼ 19)
Reproductive status n
Number ofantral folliclesper female
Number oflarge antral
follicles >2 mm
Average largestantral‐folliclediameter (mm)
Largestantral folliclediameter (mm)
Inactive females 85 21.2 � 9.4a 0.00 � 0.00a 1.19 � 0.05a 1.25Non‐pregnant females inthe follicular phase
49 12.2 � 9.5a 0.75 � 0.96b 3.70 � 3.10b 8.06
Non‐pregnant females inthe luteal phase
27 5.2 � 5.7b 0.20 � 0.45a 1.64 � 0.68a 2.51
Pregnant females 54 9.0 � 11.4a,b 0.17 � 0.41a 1.31 � 0.76a 2.11F3,211 ¼ 28.892;
P < 0.0001F3,211 ¼ 21.612;
P < 0.0001F3,211 ¼ 30.926;
P < 0.0001
a,bValues appearing in columns with different superscripts are significantly different (P < 0.05).Differences in number and diameter of antral follicles were tested using 1‐way ANOVA and Tukey–Kramer multiple comparisons test.
Am. J. Primatol.
Female Genital Organs of the Red Uakari Monkey / 547
externally bounded by a zone of condensed stromaltissue representing the theca externa of the ovulatoryfollicle. Largest CL occupied almost the wholeovarian stroma.
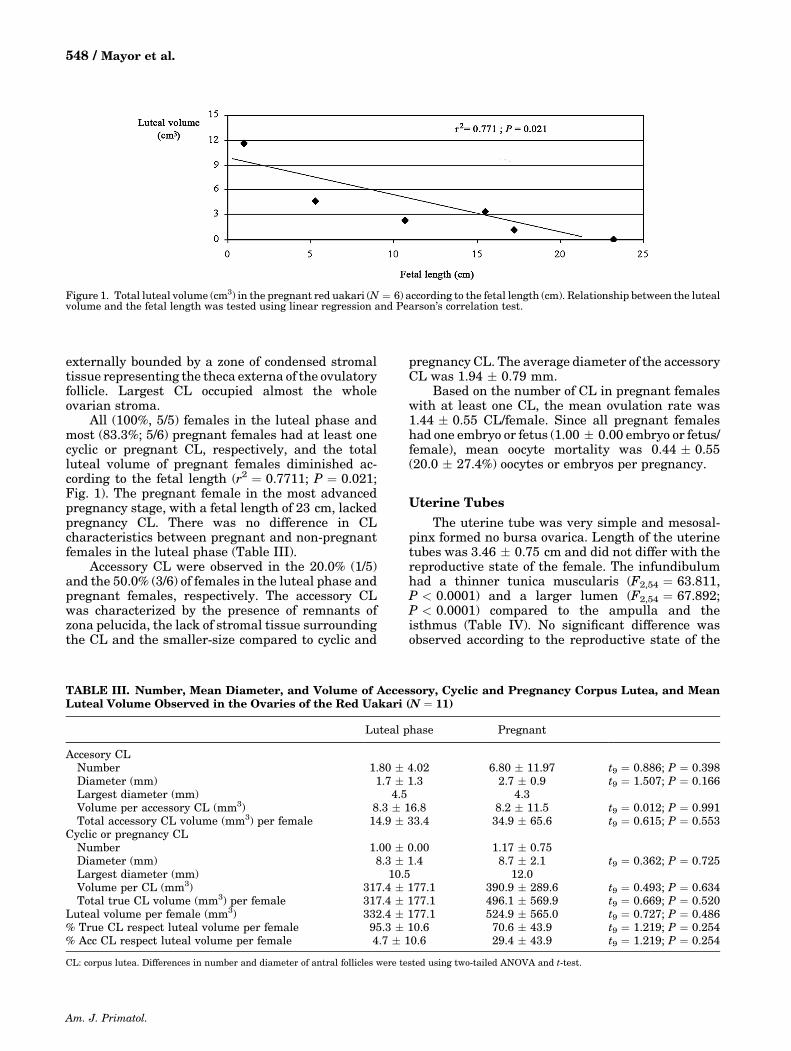
All (100%, 5/5) females in the luteal phase andmost (83.3%; 5/6) pregnant females had at least onecyclic or pregnant CL, respectively, and the totalluteal volume of pregnant females diminished ac-cording to the fetal length (r2 ¼ 0.7711; P ¼ 0.021;Fig. 1). The pregnant female in the most advancedpregnancy stage, with a fetal length of 23 cm, lackedpregnancy CL. There was no difference in CLcharacteristics between pregnant and non‐pregnantfemales in the luteal phase (Table III).
Accessory CL were observed in the 20.0% (1/5)and the 50.0% (3/6) of females in the luteal phase andpregnant females, respectively. The accessory CLwas characterized by the presence of remnants ofzona pelucida, the lack of stromal tissue surroundingthe CL and the smaller‐size compared to cyclic and
pregnancy CL. The average diameter of the accessoryCL was 1.94 � 0.79 mm.
Based on the number of CL in pregnant femaleswith at least one CL, the mean ovulation rate was1.44 � 0.55 CL/female. Since all pregnant femaleshad one embryo or fetus (1.00 � 0.00 embryo or fetus/female), mean oocyte mortality was 0.44 � 0.55(20.0 � 27.4%) oocytes or embryos per pregnancy.
Uterine Tubes
The uterine tube was very simple and mesosal-pinx formed no bursa ovarica. Length of the uterinetubes was 3.46 � 0.75 cm and did not differ with thereproductive state of the female. The infundibulumhad a thinner tunica muscularis (F2,54 ¼ 63.811,P < 0.0001) and a larger lumen (F2,54 ¼ 67.892;P < 0.0001) compared to the ampulla and theisthmus (Table IV). No significant difference wasobserved according to the reproductive state of the
Figure 1. Total luteal volume (cm3) in the pregnant red uakari (N ¼ 6) according to the fetal length (cm). Relationship between the lutealvolume and the fetal length was tested using linear regression and Pearson’s correlation test.
TABLE III. Number, Mean Diameter, and Volume of Accessory, Cyclic and Pregnancy Corpus Lutea, and MeanLuteal Volume Observed in the Ovaries of the Red Uakari (N ¼ 11)
Luteal phase Pregnant
Accesory CLNumber 1.80 � 4.02 6.80 � 11.97 t9 ¼ 0.886; P ¼ 0.398Diameter (mm) 1.7 � 1.3 2.7 � 0.9 t9 ¼ 1.507; P ¼ 0.166Largest diameter (mm) 4.5 4.3Volume per accessory CL (mm3) 8.3 � 16.8 8.2 � 11.5 t9 ¼ 0.012; P ¼ 0.991Total accessory CL volume (mm3) per female 14.9 � 33.4 34.9 � 65.6 t9 ¼ 0.615; P ¼ 0.553
Cyclic or pregnancy CLNumber 1.00 � 0.00 1.17 � 0.75Diameter (mm) 8.3 � 1.4 8.7 � 2.1 t9 ¼ 0.362; P ¼ 0.725Largest diameter (mm) 10.5 12.0Volume per CL (mm3) 317.4 � 177.1 390.9 � 289.6 t9 ¼ 0.493; P ¼ 0.634Total true CL volume (mm3) per female 317.4 � 177.1 496.1 � 569.9 t9 ¼ 0.669; P ¼ 0.520
Luteal volume per female (mm3) 332.4 � 177.1 524.9 � 565.0 t9 ¼ 0.727; P ¼ 0.486% True CL respect luteal volume per female 95.3 � 10.6 70.6 � 43.9 t9 ¼ 1.219; P ¼ 0.254% Acc CL respect luteal volume per female 4.7 � 10.6 29.4 � 43.9 t9 ¼ 1.219; P ¼ 0.254
CL: corpus lutea. Differences in number and diameter of antral follicles were tested using two‐tailed ANOVA and t‐test.
Am. J. Primatol.
548 / Mayor et al.
female (Table V). Whereas the mucosal lining of thefimbriated infundibulum presented a labyrinth ofbranching, longitudinal folds with primary, second-ary, and tertiary folds, only primary folds wereobserved in the isthmus.
UterusThe simplex uterus was an elongated organ
characterized by a globular fundic portion and a longcervix that protruded into the vaginal canal (Fig. 2).The uterus of non‐pregnant females was completely
located inside the pelvic cavity, but in pregnantfemales the uterine body extended to abdominalcavity. Table V shows macroscopic measurements ofthe uterine body and cervix.
Figure 3 shows the changes in the endometrialglands related to the reproductive phase of thefemale. In non‐pregnant females in the follicularphase, the endometrial glands showed a significantincrease related to the follicular growth (r2 ¼ 0.771;P ¼ 0.021). Non‐pregnant females in the luteal phaseshowed well‐developed endometrial glands andendometrium. Non‐pregnant females in the follicular
TABLE IV. Microscopic Measurements of the Uterine Tubes in the Female Red Uakari (N ¼ 19)
Uterine tubes Epithelium thickness (mm) Muscular thickness (mm) Lumen thickness (mm)
Infundibulum 14.7 � 2.4 53.1 � 18.2a 1.45 � 0.57a
Ampulla 14.2 � 2.3 86.1 � 34.6b 0.54 � 0.11b
Isthmus 15.7 � 2.2 191.3 � 55.9c 0.22 � 0.07c
F2,54 ¼ 2.093; P ¼ 0.133 F2,54 ¼ 63.811; P < 0.0001 F2,54 ¼ 67.892; P < 0.0001
a,b,cValues appearing in columns with different superscripts are significantly different (P < 0.05).Differences in microscopic measurements of the uterine tube were tested using 1‐way ANOVA and Tukey–Kramer multiple comparisons test.
TABLE V. Macroscopic and Microscopic Measurements of the Uterine Tubes, Uterus, Vagina, and ExternalUrogenital Opening in the Female Uakari (N ¼ 19)
Inactive
Non‐pregnantin the
follicular phase
Non‐pregnantin the
luteal phase Pregnant
n 4 4 5 6Uterine tubes
Infundibulum diameter (mm) 2.43 � 0.87 2.53 � 0.67 2.68 � 0.65 2.23 � 0.73 F3,15 ¼ 0.365; P ¼ 0.779Ampulla diameter (mm) 1.86 � 0.55 1.98 � 0.54 2.37 � 0.92 2.05 � 0.76 F3,15 ¼ 0.409; P ¼ 0.749Isthmus diameter (mm) 1.93 � 0.23 1.78 � 0.61 2.15 � 0.81 1.85 � 0.75 F3,15 ¼ 0.276; P ¼ 0.842Total length (cm) 3.60 � 1.39 3.03 � 0.84 3.60 � 0.42 3.12 � 0.56 F3,15 ¼ 0.629; P ¼ 0.608
Uterine bodyLength (cm) 1.72 � 0.86 1.47 � 0.53 1.86 � 0.85 6.83 � 5.65 F3,15 ¼ 2.906; P ¼ 0.069Diameter (cm) 1.48 � 0.97a 1.36 � 0.36a 1.37 � 0.48a 4.22 � 2.29b F3,15 ¼ 5.381; P ¼ 0.010Total thickness (mm) 1.23 � 0.18a,b 2.12 � 1.23a,b 2.58 � 1.45a 0.83 � 0.62b F3,15 ¼ 3.323; P ¼ 0.048Endometrium (mm) 0.42 � 0.25a,b 1.35 � 0.96a,b 1.78 � 1.22a 0.28 � 0.54b F3,15 ¼ 3.808; P ¼ 0.033Density of endometrial glands 0.7 � 0.3a 4.0 � 1.4b,c 4.5 � 0.6c 1.6 � 2.3a,b F3,15 ¼ 6.788; P ¼ 0.0041Myometrium (mm) 0.80 � 0.07 0.76 � 0.27 0.55 � 0.14 0.78 � 0.26 F3,15 ¼ 1.524; P ¼ 0.241
Uterine cervixLength (cm) 0.93 � 0.38 0.98 � 0.19 1.09 � 0.21 1.15 � 0.42 F3,15 ¼ 0.453; P ¼ 0.719Diameter (cm) 1.22 � 0.49 1.38 � 0.64 1.37 � 0.48 1.36 � 0.29 F3,15 ¼ 0.107; P ¼ 0.954Density of cervical glands 1.33 � 0.58a 3.00 � 0.87a,b 3.25 � 1.50a,b 3.60 � 0.89b F3,15 ¼ 4.141; P ¼ 0.025Myometrium (mm) 0.68 � 0.03 0.69 � 0.04 0.69 � 0.15 0.71 � 0.30 F3,15 ¼ 0.022; P ¼ 0.996
VaginaLength (cm) 3.18 � 0.86 3.31 � 0.30 3.32 � 0.38 3.43 � 0.81 F3,15 ¼ 0.124; P ¼ 0.947Diameter (cm) 1.41 � 0.88 1.05 � 0.31 1.25 � 0.24 1.15 � 0.49 F3,15 ¼ 0.361; P ¼ 0.782Thickness epithelium (mm) 55.0 � 16.7a 132.0 � 54.6b 120.7 � 21.7b 67.8 � 51.8a F3,15 ¼ 3.888; P ¼ 0.031Number of epithelial cell layers 6.7 � 0.8 14.5 � 10.8 11.0 � 2.7 5.5 � 2.4 F3,15 ¼ 2.87; P ¼ 0.071Cornification of the epithelium 0.0 � 0.0a 3.3 � 1.1b 0.9 � 0.7a 0.5 � 0.7a F3,15 ¼ 16.51; P < 0.0001
External urogenital opening.Dorso‐ventral diameter (cm)
2.44 � 0.76 2.13 � 0.64 2.51 � 0.42 2.45 � 0.38 F3,15 ¼ 0.425; P ¼ 0.738
a,b,cValues appearing in columns with different superscripts are significantly different (P < 0.05).Differences inmacroscopic andmicroscopicmeasurements of tubular organswere tested using 1‐wayANOVAandTukey–Kramermultiple comparisons test.
Am. J. Primatol.
Female Genital Organs of the Red Uakari Monkey / 549

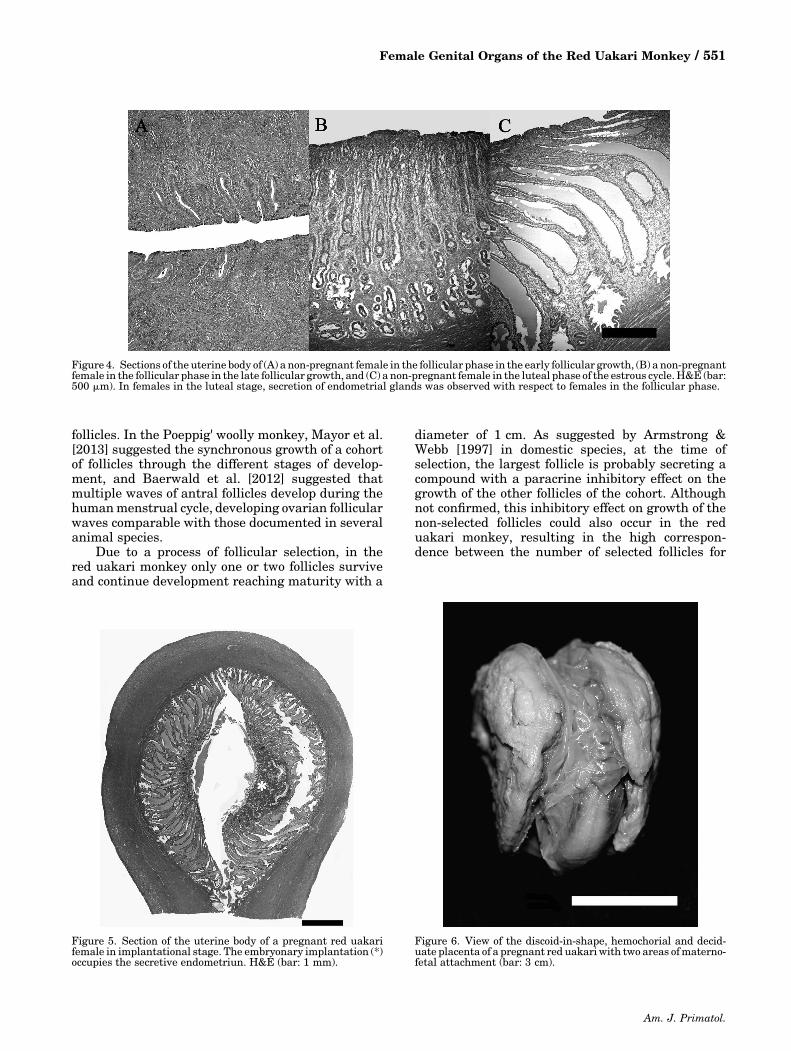
and the luteal phase had a greater proliferation ofendometrial glands (F3,15 ¼ 6.788; P ¼ 0.0041) and athicker endometrium (F3,15 ¼ 3.808; P ¼ 0.033),compared to pregnant females (Fig. 4). In pregnantfemales at the embryonic stage, the embryo implan-tation occupied the secretive endometriun (Fig. 5)and pregnant females at the fetal stage presented analmost non‐existent endometrium. Females in thefollicular phase did not present amounts of collagenbeneath the endometrial epithelium.
The mucosa of the endocervix had many primaryfolds, and the epithelium was monostratified withlarge columnar PAS‐stained mucous‐secreting cells.Although glandular development in pregnant fe-males was greater than that in inactive females(F3,15 ¼ 4.141; P ¼ 0.025), there was no significant
difference related to the reproductive state of activefemales.
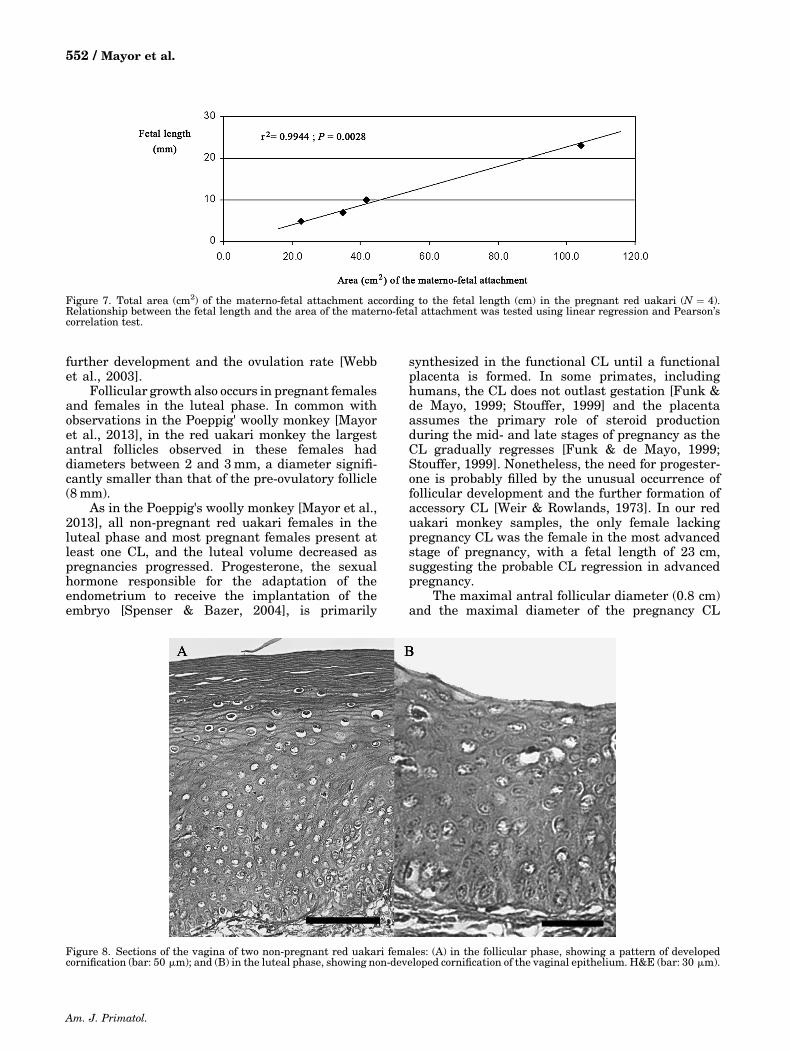
Pregnant red uakari monkey females had adiscoid‐in‐shape, hemochorial and deciduate placen-ta, with two discs as a region of maternal–fetalattachment (Fig. 6), showing a progressive surfaceincrease related to the fetal length (r2 ¼ 0.9944;P ¼ 0.0028; Fig. 7).
Vagina
The vaginal process of the cervix formed a0.38 � 0.28 cm enfolding protruding into the vagina.Macroscopic features did not show significant differ-ences according to the reproductive state of thefemale (Table V). The vaginal mucosa was character-ized by a stratified squamous epithelium withnumerous lamina propria papilla. Non‐pregnantfemales in the follicular phase and in the lutealphase had a thicker epithelium than pregnant andinactive females (F3,15 ¼ 3.888; P ¼ 0.031). Non‐pregnant females in the follicular phase had a greaterepithelial cornification compared to other females(F3,15 ¼ 16.51; P < 0.0001; Fig. 8).
DISCUSSIONOne of the major problems with the implementa-
tion of in situ and ex situ conservation programs isthe poor knowledge of reproductive performance inwild species [Andrabi & Maxwell, 2007]. Studyingreproduction in Vulnerable and endangered speciesis difficult due to the access to a large enough samplesize. In this study, genital organs were recoveredfrom 19 female uakari monkeys to provide newinformation on the reproductive physiology of thePeruvian red uakari monkey.
Non‐pregnant uakari females in the follicularphase showed an apparent decrease in the number ofantral follicles related to the largest antral folliclediameter. The determination of follicular wavesrequires frequent evaluations of follicular growthwithin individuals. Although these evaluations werenot conducted in the present study, the results of thisstudy suggest a synchronous growth of a cohort of
Figure 2. Dorsal view of the genital organs of a non‐pregnantfemale red uakari. Ovaries (O), Oviduct (Ov), uterine body (UB),cervix (CX), vagina (VG), perineal region (Pr), and urinarybladder (Ub) (bar: 2 cm).
Figure 3. Changes in the endometrial glands related to the reproductive phase of the red uakari female (N ¼ 15). Relationship betweenthe largest follicle and the density of endometrial glands was tested using linear regression and Pearson’s correlation test.
Am. J. Primatol.
550 / Mayor et al.
follicles. In the Poeppig' woolly monkey, Mayor et al.[2013] suggested the synchronous growth of a cohortof follicles through the different stages of develop-ment, and Baerwald et al. [2012] suggested thatmultiple waves of antral follicles develop during thehumanmenstrual cycle, developing ovarian follicularwaves comparable with those documented in severalanimal species.
Due to a process of follicular selection, in thered uakari monkey only one or two follicles surviveand continue development reaching maturity with a
diameter of 1 cm. As suggested by Armstrong &Webb [1997] in domestic species, at the time ofselection, the largest follicle is probably secreting acompound with a paracrine inhibitory effect on thegrowth of the other follicles of the cohort. Althoughnot confirmed, this inhibitory effect on growth of thenon‐selected follicles could also occur in the reduakari monkey, resulting in the high correspon-dence between the number of selected follicles for
Figure 4. Sections of the uterine body of (A) a non‐pregnant female in the follicular phase in the early follicular growth, (B) a non‐pregnantfemale in the follicular phase in the late follicular growth, and (C) a non‐pregnant female in the luteal phase of the estrous cycle. H&E (bar:500 mm). In females in the luteal stage, secretion of endometrial glands was observed with respect to females in the follicular phase.
Figure 5. Section of the uterine body of a pregnant red uakarifemale in implantational stage. The embryonary implantation (*)occupies the secretive endometriun. H&E (bar: 1 mm).
Figure 6. View of the discoid‐in‐shape, hemochorial and decid-uate placenta of a pregnant red uakariwith two areas ofmaterno‐fetal attachment (bar: 3 cm).
Am. J. Primatol.
Female Genital Organs of the Red Uakari Monkey / 551
further development and the ovulation rate [Webbet al., 2003].
Follicular growth also occurs in pregnant femalesand females in the luteal phase. In common withobservations in the Poeppig' woolly monkey [Mayoret al., 2013], in the red uakari monkey the largestantral follicles observed in these females haddiameters between 2 and 3mm, a diameter signifi-cantly smaller than that of the pre‐ovulatory follicle(8mm).
As in the Poeppig's woolly monkey [Mayor et al.,2013], all non‐pregnant red uakari females in theluteal phase and most pregnant females present atleast one CL, and the luteal volume decreased aspregnancies progressed. Progesterone, the sexualhormone responsible for the adaptation of theendometrium to receive the implantation of theembryo [Spenser & Bazer, 2004], is primarily
synthesized in the functional CL until a functionalplacenta is formed. In some primates, includinghumans, the CL does not outlast gestation [Funk &de Mayo, 1999; Stouffer, 1999] and the placentaassumes the primary role of steroid productionduring the mid‐ and late stages of pregnancy as theCL gradually regresses [Funk & de Mayo, 1999;Stouffer, 1999]. Nonetheless, the need for progester-one is probably filled by the unusual occurrence offollicular development and the further formation ofaccessory CL [Weir & Rowlands, 1973]. In our reduakari monkey samples, the only female lackingpregnancy CL was the female in the most advancedstage of pregnancy, with a fetal length of 23 cm,suggesting the probable CL regression in advancedpregnancy.
The maximal antral follicular diameter (0.8 cm)and the maximal diameter of the pregnancy CL
Figure 7. Total area (cm2) of the materno‐fetal attachment according to the fetal length (cm) in the pregnant red uakari (N ¼ 4).Relationship between the fetal length and the area of the materno‐fetal attachment was tested using linear regression and Pearson’scorrelation test.
Figure 8. Sections of the vagina of two non‐pregnant red uakari females: (A) in the follicular phase, showing a pattern of developedcornification (bar: 50 mm); and (B) in the luteal phase, showing non‐developed cornification of the vaginal epithelium. H&E (bar: 30 mm).
Am. J. Primatol.
552 / Mayor et al.

(1.2 cm) resulted in an approximately 1.5‐fold in-crease between both ovarian structures. Zuckerman& Weir [1977] concluded that the final size of the CLis related to the diameter of the mature follicle fromwhich it arose, but there is variation from nearequality to a twofold increase over that of the follicle.
Formation of accessory CL occurred both inpregnant and non‐pregnant females in the lutealphase. Some follicles are apparently so sensitive tothe luteinizing factors that they respond beforeovulation takes place, developing the luteinizationof non‐ovulated follicles [Stansfield & Allen, 2012]. Inthe pregnant red uakari monkey, the presence ofaccessory CL resulted in a contribution up to 30% ofthe total luteal volume but was not related to theprogress of pregnancy, suggesting a probable relationwith the particular activity of the functional CL ineach pregnant female [Rowlands et al., 1970].
The observed mean ovulation rate was 1.44oocytes and the litter size was 1.00 fetus per pregnantfemale, resulting in a rate of reproductive wastageaveraging 20.0% oocytes or embryos per pregnancy,similar to the 33.3% observed in the Poeppig’s woollymonkey [Mayor et al., 2013]. This apparently highpercentage of reproductive wastage means the loss ofa maximal one oocyte or embryo. The reducedovulation rate seems to be compatible with the largeuterine spatial requirements of the placenta and theembryo, resulting in a lower rate of oocyte mortality[Mayor et al., 2004].
As in other New World primates [Mayor et al.,2012; Veras et al., 2009], the uterus of the red uakarimonkey is composed of a globular fundic portion and along cervix that protrudes into the vaginal canal.During the estrous cycle, changes in the endometri-um proceed through two distinct phases: the prolif-erative or follicular phase, and the secretory or lutealphase [Burton, 1980; Dempsey, 1939; Kraemer et al.,1977; Mayor et al., 2012; Veras et al., 2009; Zucker-man & Parkes, 1932].
In the proliferative or follicular phase, theendometrium prepares the epithelium and stromafor implantation, showing a progressive developmentof endometrial glands related to the follicular growth.In the luteal phase, the endometrium transformsfrom a proliferative to a secretory type, showing athicker endometrium with proliferation and activesecretion of endometrial uterine glands. Under theinfluence of progesterone, the non‐pregnant uterustransforms into an enriched environment suited forthe developing embryo and for maintenance of thepregnant state [Lyngset, 1968a]. In the absence offertilization, the endometrium cannot bemaintained,collapses and probably most of it is shed during aperiod of bleeding, resulting in the onset ofmenstruation.
NewWorld primates apparently show importantdifferences related to the physiology of uterinechanges. Whereas Veras et al. [2009] suggested
that the howler monkey does not exhibit visiblemenstrual bleeding, Mayor et al., [2012] reported inthe Poeppig’s woolly monkey a dramatic loss of thefunctional zone of the endometrium and a menstrualbleeding that could stand as an external sign for thediagnosis of the reproductive phase. In the reduakari, the small sample size does not permit us todraw conclusive results, but the lack of large amountsof collagen, as endometrial remodeling [Tanaka et al.,2009], suggests a non‐visible menstrual bleeding.
In the case of fertilization, the discoid‐in‐shape,hemochorial and deciduate placenta progressivelycollapses the endometrial glands and the endometri-um disappears. As in most New World platyrrhinemonkeys, and contrary to observations in the howlermonkey [Benirschke & Miller, 1982; Miller &Benirschke, 1985] and the woolly monkey [Mayoret al., 2012], the red uakari monkey presents abidiscoid placenta. The region of maternal–fetalattachment shows a progressive increase in surfacearea as pregnancy progresses.
As in the brown howler monkey [Veras et al.,2009] and the woolly monkey [Mayor et al., 2012], thepregnant red uakari female presents an evidentmucous secretion from the endocervical glands,closing the endocervical canal so that foreignmaterial cannot enter the uterus during gestation[Senger, 1997]. Females in the follicular phase alsopresented a high density and secretion of endocer-vical glands. We could not quantify the endocervicalsecretion, but rising levels of estrogens during thefollicular phase probably promote a fluid secretion sothat spermatozoa may enter into the uterus aroundthe time of ovulation [Lyngset, 1968b].
In the red uakari, the non‐glandular, stratifiedand squamous vaginal epithelium shows changingfeatures in accordance with the reproductive state ofthe female. Whereas females in the follicular phaseshow a thick vaginal epitheliumand a clear pattern ofcornification, females in the luteal phase maintainthe epithelial thickness but lose the epithelialcornification. Finally, pregnant and inactive femaleshave a thin and non‐cornified epithelium. As ob-served in other NewWorldmonkeys [Dempsey, 1939;Gluckman et al., 2004; Goodman & Wislocki, 1935;Mayor et al., 2012] but not in the howler monkey[Veras et al., 2009], the vaginal features observedsuggest that vaginal cytology could be a successfulmethodology in order to characterize the estrous cycleof the red uakari monkey.
In practice, current reproductive biotechnologiesare inefficient for many endangered animals due toinsufficient knowledge of basic reproductive factors[Andrabi &Maxwell, 2007]. The present reproductiveevaluation of the Peruvian red uakari monkeyprovides important morphological and physiologicalinformation that could be directed towardsthe development of management strategies for thespecies. Furthermore, this study may improve the
Am. J. Primatol.
Female Genital Organs of the Red Uakari Monkey / 553

development of imaging diagnoses for monitoringfemales.
ACKNOWLEDGMENTS
We sincerely thank all the people from thecommunity of Nueva Esperanza in the Yavarí‐MirínRiver, who actively participated in data collection,which shows that communal participation is animportant step in the development of wildlifemanagement. We also extend our thanks toDr. Richard Bodmer and the Wildlife ConservationSociety for their kind assistance during the fieldwork.
REFERENCESAndrabi SMH, Maxwell WMC. 2007. A review on reproductive
biotechnologies for conservation of endangered mammalianspecies. Anim Reprod Sci 99:223–243.
Armstrong DG, Webb R. 1997. Ovarian follicular dominance:the role of intraovarian growth factors and novel proteins.Rev Reprod 2:139–146.
Baerwald R, Adams GP, Pierson RA. 2012. Ovarian antralfolliculogenesis during the humanmenstrual cycle: a review.Hum Reprod Update 18:73–91.
Benirschke K, Miller CJ. 1982. Anatomical and functionaldifferences in the placenta of primates. Biol Reprod 26:29–53.
Bowler M, Barton C, McCann‐Wood S, Puertas P, Bodmer R.2013. Annual variation in breeding success and changes inpopulation density of Cacajao calvus ucayalii in the LagoPreto conservation concession, Peru. In: Veiga LM, BarnettAA, Ferrari SF, Norconk MA, editors. Evolutionary biologyand conservation of Titis, Sakis and Uacaris. Cambridge,UK: Cambridge University Press. p 173–178.
Bowler M, Bodmer R. 2009. Social behavior in fission–fusiongroups of red uakari monkeys (Cacajao calvus ucayalii). AmJ Primatol 71:976–987.
Bowler M, Bodmer RE. 2011. Diet and food choice in PeruvianRed Uakaris (Cacajao calvus ucayalii): selective or opportu-nistic seed predation? Int J Primatol 32:1109–1112.
Bowler M, Noriega J, Recharte M, Puertas PE, Bodmer RE.2009. Peruvian red uakari monkeys (Cacajao calvusucayalii) in the Pacaya‐Samiria National Reserve—a rangeextension across a major river barrier. Neotrop Primates16:34–37.
Burton GJ. 1980. Light microscopy of the endometrium of thedusky leaf‐monkey (Presbytis obscura). J Reprod Fert59:107–111.
Dempsey EW. 1939. The reproductive cycle of the new worldmonkeys. Am J Anat 64:381–405.
Fontaine R. 1981. The Uakaris, genus Cacajao. In: Coimbra‐Filho AF, Mittermeier RA, editors. Ecology and behavior ofneotropical primates. Rio de Janeiro: AcademiaBrasileira deCiencias. p 443–493.
Funk CR, de Mayo FJ. 1999. Progesterone actions onreproductive tract. In: Knobil E, Neill JD, editors. Encyclo-pedia of reproduction. San Diego, CA: Academic Press.p 6–16.
Gluckman TL, Walz SE, Schultz‐Darken N, Bolton ID. 2004.Cytologic assessment of the vaginal epithelium in thecommon marmoset (Callithrix jacchus): a preliminary newapproach to reproductive screening. Contemp Top Lab AnimSci 43:28–31.
Goodman L, Wislocki GB. 1935. Cyclical uterine bleeding in aNew world monkey (Ateles geoffroyi). Anat Rec 61:379–387.
Heymann EW, Aquino R. 2010. Peruvian red uakaris (Cacajaocalvus ucayalii) are not flooded‐forest specialists. Int JPrimatol 31:751–758.
International Committee on Veterinary Embryological No-menclature. 1994. Nomina embryologica veterinaria. NewYork: World Association of Veterinary Anatomists.
Kraemer DC, Maqueo M, Hendrickx AG, Vera‐Cruz NC. 1977.Histology of the baboon endometrium during the menstrualcycle and pregnancy. Fert Steril 28:482–487.
Lyngset O. 1968a. Studies on reproduction in the goat. I. Thenormal genital organs of the pregnant goat. Acta Vet Scand9:242–256.
Lyngset O. 1968b. Studies on reproduction in the goat. I. Thenormal genital organs of the non‐pregnant goat. Acta VetScand 9:208–222.
Mayor P, Bowler M, López‐Plana C. 2013. Ovarian functionali-ty in Poeppig’s woolly monkey (Lagothrix poeppigii). AnimReprod Sci 136:310–316.
Mayor P, Bowler M, López‐Plana C. 2012. Anatomicohisto-logical characteristics of the tubular genital organs of thefemale woolly monkey (Lagothrix poeppigii). Am J Primatol74:1006–1016.
Mayor P, Jori F, López‐Béjar M. 2004. Anatomicohistologicalcharacteristics of the tubular genital organs of the femalecollared peccary (Tayassu tajacu) from north‐eastern Ama-zon. Anat Histol Embryol 33:65–74.
Miller PW, Benirschke K. 1985. A large allantoic sac in theplacenta of the spider monkey Ateles geoffroyi. Placenta6:423–426.
Puertas P, Bodmer RE. 1993. Conservation of a high diversityprimate assemblage. Biodivers Conserv 2:586–593.
Pukazhenthi BS, Wildt DE. 2004. Which reproductive technol-ogies are most relevant to studying, managing and conserv-ing wildlife? Reprod Fertil Dev 16:33–46.
Rowlands IW, Tam WH, Kleiman DG. 1970. Histological andbiochemical studies on the ovary and of progesterone levelsin the systemic blood of the green acouchi (Myoprocta pratti).J Fertil 22:533–545.
Senger PL. 1997. Pathways to pregnancy and parturition.Pullman; WA: Current Conceptions Inc.
Spenser TE, Bazer FW. 2004. Conceptus signals for theestablishment and maintenance of pregnancy. Reprod BiolEndocrinol 2:49.
Stansfield FJ, Allen WR. 2012. Luteal maintenance ofpregnancy in the African elephant (Loxodonta africana).Reproduction 143:845–854.
Stouffer RL. 1999. Corpus luteum of pregnancy. In: Knobil E,Neill JD, editors. Encyclopedia of reproduction. San Diego,CA: Academic Press. p 709–717.
Tanaka T, Wang C, Umesaki N. 2009. Remodelling of thehuman endometrial is regulated by laminin and type IVcollagen. Int J Mol Med 23:173–180.
Veiga LM, Bowler M, Silva JS, et al. 2008. Cacajao calvus. In:IUCN 2011. IUCN Red list of threatened species. Version2011.2. http://www.iucnredlist.org Accessed May 30, 2012.
Veras MM, Marqués KV, Miglino MA, García E. 2009.Observations on the female internal reproductive organs ofthe brown howler monkey (Alouatta guariba clamitans). AmJ Primatol 71:145–152.
Webb R, Nicholas B, Gong JG, et al. 2003. Mechanismsregulating follicular development and selection of thedominant follicle. Reprod Suppl 61:71–90.
Weir BJ, Rowlands IW. 1973. Reproductive strategies ofmammals. Annu Rev Ecol Syst 4:139–163.
Zuckerman L, Weir JJ. 1997. The ovary. New York: AcademicPress.
Zuckerman S, Parkes AS. 1932. The menstrual cycle of theprimates: the cycle of the baboon. Proc Zool Soc London102:139–192.
Am. J. Primatol.
554 / Mayor et al.




















