Functional Characterization of Actinobacillus ... - CiteSeerX
26
1 Functional Characterization of Actinobacillus pleuropneumoniae Urea Transport Protein, ApUT Geeta Godara 1 , Craig Smith 3 , Janine Bosse 4 , Mark Zeidel 2 , John Mathai 2* 1# Department of Medicine, University of Pittsburgh, Pittsburgh, PA 15231. 2 Beth Israel Deaconess Medical Center and Harvard Medical School, 330 Brookline Avenue, Boston, MA 02215. 3 Faculty of Life Sciences, University of Manchester, Manchester M13 9NT, UK. 4 Molecular Infectious Disease Group, Department of Paediatrics, Imperial College London, St. Mary’s Campus, Norfolk Place, London W2 1PG, UK Running title: Bacterial Urea Transporter Address for correspondence to: John Mathai Beth Israel Deaconess Medical Center 840 Memorial Drive Cambridge, MA 02139 Ph: 617 754 1271 Email: [email protected] Articles in PresS. Am J Physiol Regul Integr Comp Physiol (January 14, 2009). doi:10.1152/ajpregu.90726.2008 Copyright © 2009 by the American Physiological Society.
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Functional Characterization of Actinobacillus ... - CiteSeerX

1
Functional Characterization of Actinobacillus pleuropneumoniae Urea Transport Protein, ApUT
Geeta Godara1, Craig Smith3, Janine Bosse4, Mark Zeidel2, John Mathai2*
1#Department of Medicine, University of Pittsburgh, Pittsburgh, PA 15231.
2Beth Israel Deaconess Medical Center and Harvard Medical School, 330 Brookline Avenue,
Boston, MA 02215. 3Faculty of Life Sciences, University of Manchester, Manchester M13 9NT, UK.
4 Molecular Infectious Disease Group, Department of Paediatrics, Imperial College London, St. Mary’s Campus, Norfolk Place, London W2 1PG, UK
Running title: Bacterial Urea Transporter
Address for correspondence to:
John Mathai
Beth Israel Deaconess Medical Center
840 Memorial Drive
Cambridge, MA 02139
Ph: 617 754 1271
Email: [email protected]
Articles in PresS. Am J Physiol Regul Integr Comp Physiol (January 14, 2009). doi:10.1152/ajpregu.90726.2008
Copyright © 2009 by the American Physiological Society.

2
ABSTRACT
Urea transporters (UTs) effect rapid flux of urea across biological membranes. In the mammalian
kidney, UT activity is essential for effective urine concentration. In bacteria, UT-mediated urea
uptake permits intracellular urease to degrade urea to ammonia and CO2, a process which either
buffers acid loads or provides nutrient nitrogen. We have characterized the urea transport channel
protein ApUT from Actinobacillus pleuropneumoniae. Kinetic analysis of bacterial inside-out
membranes enriched in ApUT showed ~28 fold increase in urea permeability (3.3 ± 0.4 x10-4
cm/s) compared to control vesicles (0.11 ± 0.02 x10-4 cm/s). In addition to urea ApUT also
conducts water. Urea and water transport across the channel was phloretin and mercury inhibitable
and the site of inhibition may be located on the cytoplasmic side of the protein. Glycerol and urea
analogues such as methylamine, dimethylurea, formamide, acetamide, methylurea, propanamide
and ethylamine did not permeate across ApUT.
Key Words:
Urea Permeability; Water permeability; bacterial vesicles; urea analogues

3
INTRODUCTION
Urea, an end product of nitrogen metabolism, is widespread in nature. Elasmobranchs and
mammals use urea as an osmoticant, permitting the elasmobranch to maintain its osmolality and
the mammalian kidney to effect maximal concentration of urine (9). Microbes metabolize urea as a
nitrogen source or as a mechanism to combat an acidic environment. Urea transporters mediate
bacterial acquisition of urea and are considered to be critical for survival.
Because urea is a small electroneutral molecule, it exhibits appreciable permeability across
lipid membranes. Although this passive urea permeability has been thought to be sufficient for its
biological roles, several specific urea transporter proteins have been identified in prokaryotes and
eukaryotes. In mammals UT-A (6 isoforms) and UT-B (2 isoforms) genes encode members of a
specialized family of urea transporters that play an important role in the renal urinary concentrating
mechanism and potentially other processes including spermatogenesis (9).
In bacteria two different types of transporters are known to exist for urea uptake. Under
conditions of nitrogen starvation, in Methylophilus methylotropus and Corynebacterium
glutamicaum, an ATP-dependent ABC type transporter is responsible for urea uptake into the cell
(2, 24). Passive or facilitated movement of urea is seen in H. pylori where urea uptake is mediated
by an acid activated urea channel, UreI, which was shown to be essential for survival in vivo (33).
UreI is a single component protein with 6 transmembrane spanning domains and does not share
sequence homology with mammalian urea transporters. Several ureI type genes have subsequently
been identified in other microbes such as Streptococcus salivarius, Brucella melitensis, and
Heliobacter hepaticus (3, 25). As an example, in H. pylori, the gastric pathogen responsible for
much of human peptic ulcer disease, activity of a urease system maintains a near neutral
periplasmic and cytoplasmic pH in the face of an ambient pH of 2 in the gastric lumen (26).

4
Following its uptake into the organism, urease breaks down urea to produce ammonia and carbon
dioxide; the buffering capacity of ammonia helps protect the organism from the gastric acid
environment. Recent studies also suggest that the conversion of CO2 to HCO3- by periplasmic α-
carbonic anhydrase is also essential for acid acclimation (20).
Therefore, urea metabolism and urease activity in particular have been implicated in the ability
of certain bacteria to infect and cause disease (4, 8, 38). Examination of the urease gene cluster of
the swine pathogen, Actinobacillus pleuropneumoniae (5) resulted in the identification and cloning
of a putative urea transporter cDNA encoding a 300 amino acid protein, ApUT (Gen bank ref
AAD49729). ApUT does not show any sequence homology to the acid activated bacterial urea
transporter UreI or the energy dependent urea uptake systems in some bacteria. However, it shows
26% identity to the mammalian urea transporter UT-A2 (Gen Bank ref Q62668, (34) and 21%
identity to mammalian members of the UT-B subfamily (Gen Bank ref Q13336, (27). Given the
lack of detailed functional data on mammalian urea transporters, and our ability to reconstitute
activity of ApUT in bacteria, we have examined ApUT function in detail as a model transporter for
the UT-A/UT-B family. ApUT was overexpressed in bacteria and using inside-out membrane
vesicles enriched in ApUT, a comprehensive kinetic and functional characterization was
performed.
MATERIALS AND METHODS
Bacterial Strains and Growth Conditions: Actinobacillus pleuropneumoniae strain Shope 4074
(serotype 1) was propagated at 37 °C with 5% CO2 on brain–heart infusion (BHI) agar (DIFCO)
supplemented with 0.01% β-nicotinamide adenine dinucleotide (NAD; Roche). Escherichia coli

5
strains Top10 and BL21-AI (Invitrogen) were cultured in Luria-Bertani (LB) broth or on LB agar,
supplemented with 50 µg/ml ampicillin when appropriate.
Plasmid construction: Genomic DNA was prepared from A. pleuropneumoniae Shope 4074 using
a QIAamp mini DNA kit (Qiagen). Polymerase chain reaction (PCR) was performed according to
standard procedures (29). Primers ApUT_F (5’-ATGAAGCTGCTTAATCCACCCTTA) and
ApUT_R (5- GACCGTTTTGACTAAGGATTCAGCA) were used to amplify a 901 bp fragment
containing the 816 bp ApUt gene. The resulting PCR product was cloned into pGEM-T (Promega)
to create pJ9ApUt which was transformed into E. coli Top10. Orientation of the insert was
determined by restriction digestion with PstI and PvuI (Roche). ApUT cDNA was subsequently
subcloned into an IPTG inducible overexpression plasmid, pRSET-A (Invitrogen) downstream of
the 6 His tag. The resulting plasmid, pRSETA-ApUT, was transformed into E. coli BL21-AI for
expression.
Bacterial expression studies: BL21-AI, E. coli strain (invitrogen) was cultured at 37°C in LB broth
at pH 7.4 supplemented with 50 µg/ml ampicillin when carrying the plasmid pRSET A containing
ApUT cDNA. Cells were grown up to OD600 of 0.8 and transcription from T7 promoter was
switched on by addition of 0.2 mM Isopropyl thio β-D-galactoside (IPTG) at 10° C. After 10 hour
of induction period, cells were harvested by centrifugation for 10 minutes at 3000 x g.
Inside-out vesicle Preparation: The bacterial pellet was resuspended in buffer containing 100 mM
NaCl, 100 mM Sucrose, 50 mM Tris-Hcl, pH 7.4 (buffer A). Cells were centrifugally washed once
in buffer A and resuspended in the buffer A containing 20 mM CF (Carboxyfluorescein), protease
inhibitor cocktail, 1 tablet/20ml and DNAase, 10 ug/ml . The cells were lysed by passing the
bacterial solution through a French press at 4°C and centrifuged at 10,000 g for 10 minutes. The
supernatant was collected and the pellet containing the nuclei, unbroken cells and debris was

6
discarded. The supernatant was centrifuged at 70,000 g for 45 minutes and the pellet was washed
twice by centrifugation and resuspended in 1 ml of buffer A. The vesicles were immediately used
for transport studies (13).
Spheroplast or right-side out vesicles: E. coli spheroplast were prepared by the method of Kaback
(11) with minor modifications. Briefly bacteria was harvested and washed as mentioned above for
preparation of inside-out vesicles. Bacteria was resuspended and washed by centrifugation in a
buffer made of 20% sucrose and 30 mM Tris-HCl, pH 8.0. The pellet was resuspended in the above
buffer containing lysozyme (500 ug/ml) and 10 mM EDTA. After 30 min of incubation with gentle
swirling at 25° C spheroplast were harvested by centrifugation at 13000 x g for 10 min. The pellet
was washed twice by centrifugation in the above buffer without lysozyne and EDTA and after final
wash the pellet was resuspended in the same buffer and used immediately for transport
measurements.
Permeability Measurements: Water permeability (Pf) was measured by exposure of vesicles to an
abrupt doubling of external osmotic pressure in a stopped-flow fluorimeter. The shrinkage of
vesicles in response to the osmotic gradient was measured as change in fluorescence over time due
to self quenching of entrapped CF. 8-12 curves were averaged and fit to single exponential. A
family of single exponential curves was generated by simulation of the water permeability equation
where only the Pf was varied using MathCad software.
d(Vt)/dt = (Pf)(SAV)(MVW)(Cin/Vt – Cout)
where, Vt , is the relative volume of the vesicle at time t, (i.e., volume at time t, divided by the
initial volume), Pf (cm/s) is the osmotic water permeability coefficient, SAV is the surface area to
volume ratio, MVW is the molar volume of water (18 cm3 /mol), Cin and Cout are initial solute

7
concentrations inside and outside the vesicle. Since the external solute volume is very large
compared to internal vesicle volume, the Cout is assumed to be constant.
Water permeability (Pf) was obtained by comparing the experimental single exponential with
that of single exponentials obtained from simulation (21). The permeability of urea, urea analogs
and glycerol was measured by pre-equilibrating the vesicles at a solute concentration of 200 mM
each in buffer A for 30 minutes and then rapidly diluting them two fold in a stopped flow device to
create an isoosmotic urea gradient. Solute efflux results in vesicle shrinkage and CF quenching
which was recorded over time. Vesicles sizes were determined by using quasi-elastic light scatter
using a DynaPro particle sizer. The average diameter of inside-out vesicels and spheroplast are in
the range of 0.24 to 0.32 microns and 1.2 to 3.0 microns respectively. Permeability coefficients
were calculated using Mathcad software as described earlier (21, 22). Urea transport inhibitor
studies were done by incubation of vesicles for 30 minutes in buffer A containing 1.0 mM
phloretin or a 10 min incubation in 0.5 mM mercuric chloride. Control vesicles were similarly
treated. Kinetic studies of ApUT were performed by incubating the vesicles in various urea
solutions ranging in concentrations from 100 mM to 1000 mM. In the stopped flow measurements,
appropriate amounts of NaCl were added to maintain isoosmotic conditions upon dilution. Proton
permeability was measured by exposure of vesicles to a gradient of 0.5 pH unit and and the internal
pH decrease due to entry of protons was monitored as a decrease in entrapped CF fluorescence
(22). Ammonia permeability was measured by exposure of vesicles incubated in buffer A at a pH
of 6.8 to a 10 mM ammonia gradient. Ammonia entry leads to increase in internal pH which is
monitored as a fluorescence increase of entrapped CF in a stopped-flow device as described earlier
(22). Groups were compared using unpaired t-test and P<0.05 was considered significant.

8
RESULTS
Expression of ApUT in bacterial membrane vesicles was detected by SDS-PAGE of membrane
vesicles probed using anti-HisG antibody, Fig.1. Inside-out vesicles isolated from bacteria
expressing ApUT show a major band of approximately 35 kDa. The faint higher molecular weight
bands seen on the blot may represent oligomers of ApUT. Control membrane vesicles obtained
from uninduced bacteria do not show any bands. Sequence analysis of ApUT including the 6-his
tag predicts a molecular weight of 34 kDa. The presence of the 6 His-Tag either at the N- or C-
terminus of the protein did not affect the expression or the function of the protein (data not shown).
Urea transport studies were performed by preloading the vesicles with urea and then abruptly
diluting them in the stopped-flow device into buffer of the same osmolality but half the
concentration of urea as the interior of the vesicles. Efflux of urea was followed by water efflux,
resulting in shrinking of the vesicles (18). This vesicle shrinkage was monitored by the self-
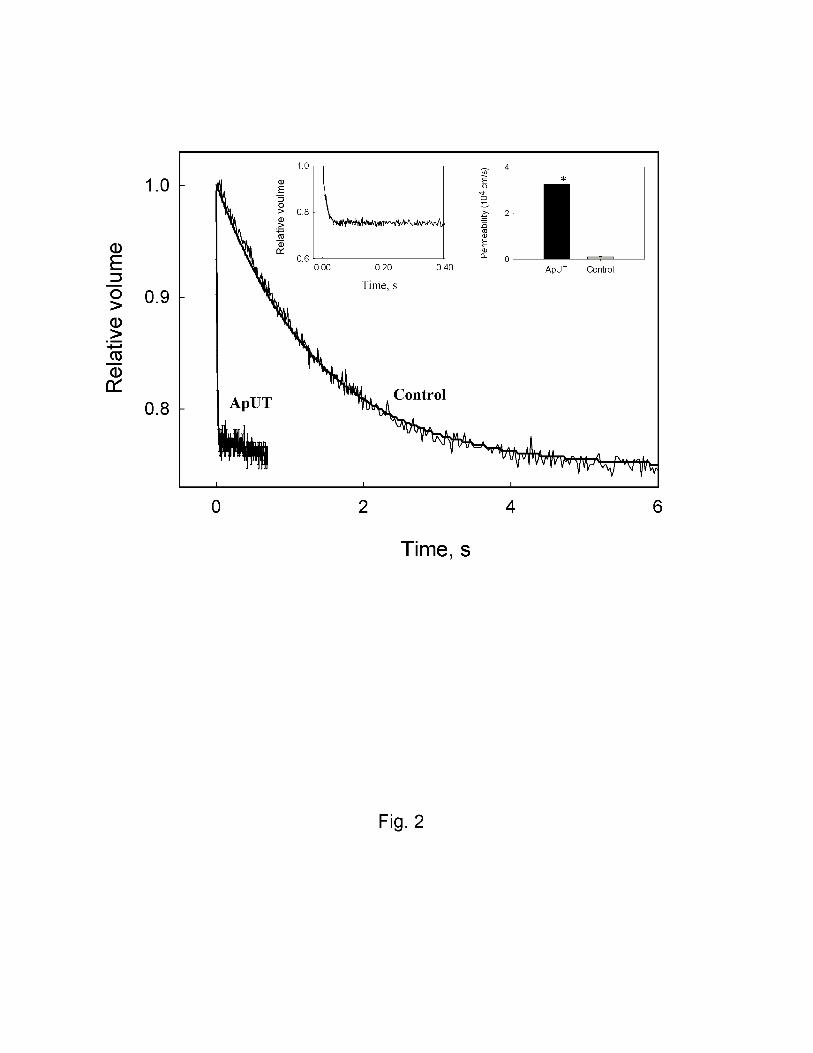
quenching of entrapped CF. Fig. 2 shows that ApUT containing vesicles equilibrate rapidly while
control vesicles without ApUT take a longer time to equilibrate. The urea permeability coefficient
of ApUT containing vesicles was ~ 28 fold higher (3.3 ± 0.4 x 10-4 cm/s) than that of the vesicles
lacking the urea transporter (0.11 ± 0.01 x 10-4 cm/s) (P<0.001). In order to check if the ApUT was
also permeable to water, ApUT containing vesicles were subjected to a doubling of external
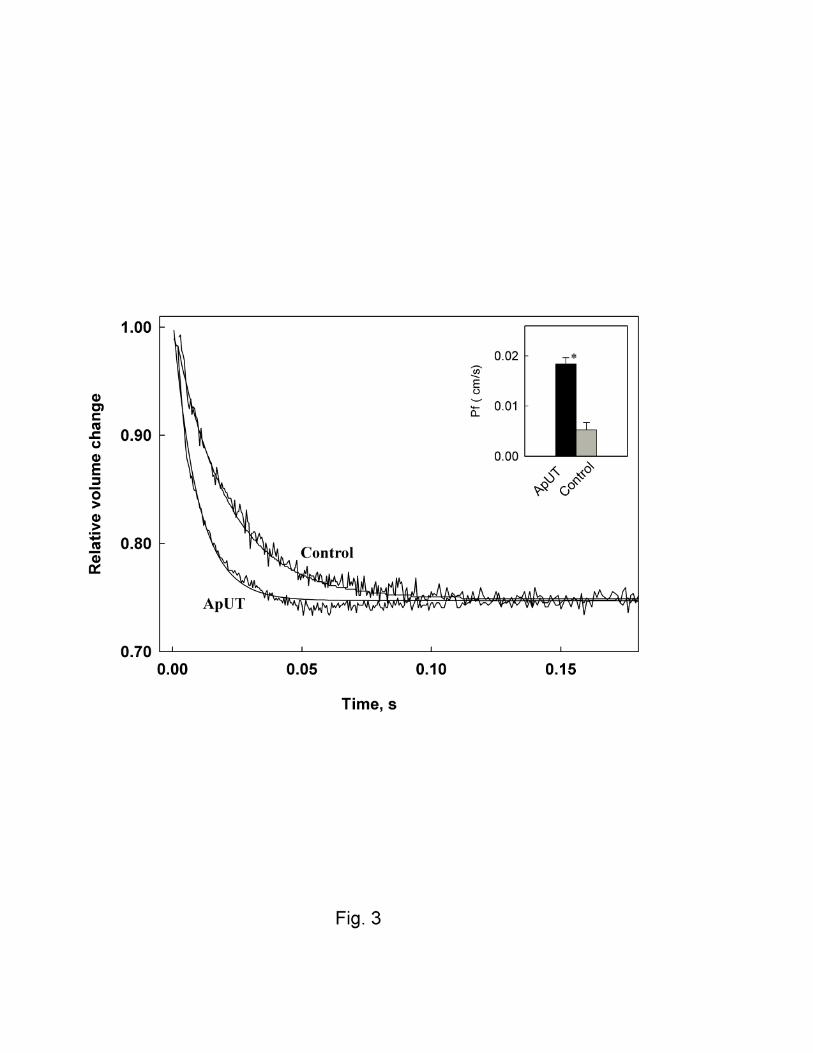
osmotic pressure and their volume change was monitored over time. Fig.3 shows that ApUT
containing vesicles shrank more rapidly than control vesicles on exposure to an osmotic gradient,
which suggested that ApUT was permeable to water. Osmotic water permeability coefficients of

9
ApUT vesicles were ~ 3.5 fold higher (Pf 1.83 ± 0.30 x 10-2 cm/s) compared to those of control
vesicles, 0.53 ± 0.14 x 10-2 cm/s (P<0.005).
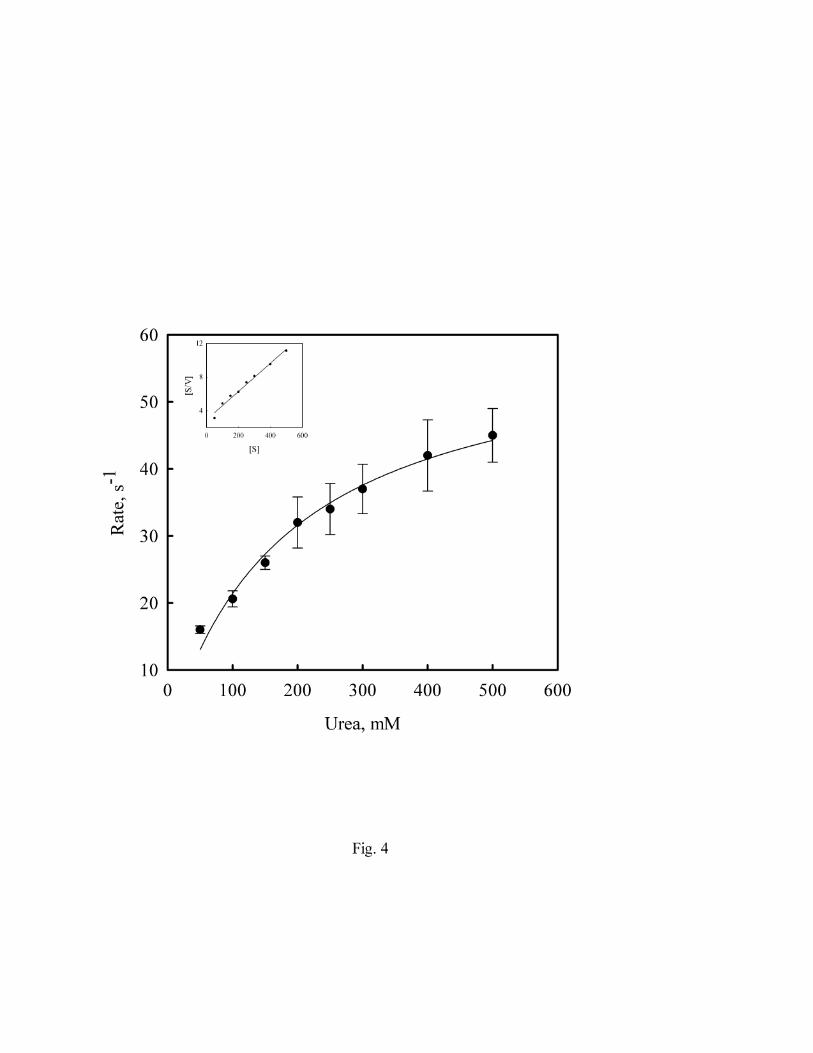
From prior work it was unclear whether urea transporters function more as urea pores or urea
carriers. These apparent mechanisms were distinguished by defining whether or not urea flux was
saturable. To determine whether ApUT exhibits saturability, and, if so, to define the Km and Vmax
of urea transport, vesicles containing ApUT were preloaded with various concentrations of urea
and the efflux of urea over time was monitored under isoosmotic conditions. Fig.4, shows the rate
of vesicle shrinkage due to urea efflux at various substrate concentrations. At higher substrate
concentrations the rate of urea efflux tended to saturate. The data was fitted by nonlinear regression
to the michalis-menton equation and the apparent Vmax and Km values are 60 and 182 mM. Inset
shows a Hanes plot and the Vmax and Km values calculated from this plot using linear regression
was 174 mM and 60 s-1. The Km value was in the same range as the Km of 218 mM measured for
mammalian erythrocyte urea transporter UT-B (23).
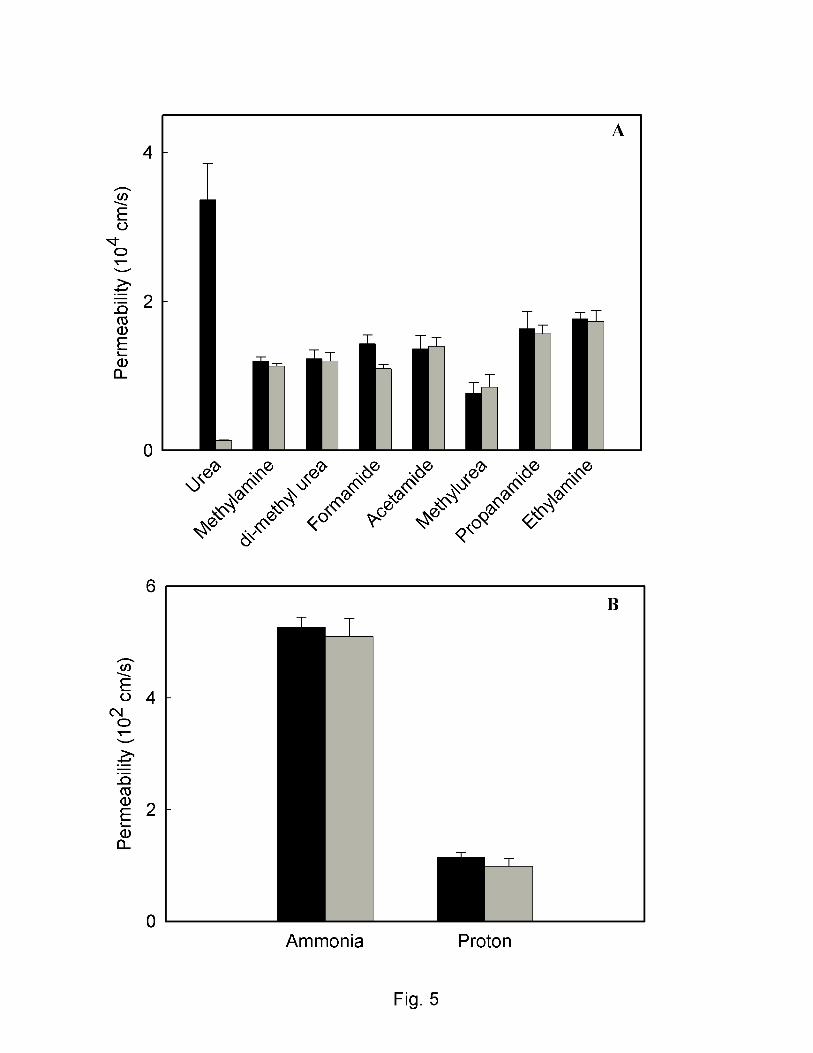
To determine the specificity of ApUT for urea, the permeabilities of several urea analogues
were measured by substituting the analogues for urea during preloading and measuring their efflux
under conditions identical to those used in the urea efflux assays. Fig.5A shows that the measured
permeability of various analogues (methylamine, dimethylurea, formamide, acetamide, methylurea,
propanamide and ethylamine) was similar for both the ApUT containing vesicles and control
vesicles lacking urea transporter. This suggests that none of the urea analogues tested permeate
through the ApUT pore. Since ammonia is a breakdown product of urease activity on urea, we also
checked if ApUT is permeable to ammonia. Fig.5B shows that ammonia does not permeate across
ApUT. Furthermore ApUT was also found to be impermeable to protons, Fig. 5B and glycerol

10
(data not shown). Together these results suggest that ApUT is specific for urea with a small water
conductance.
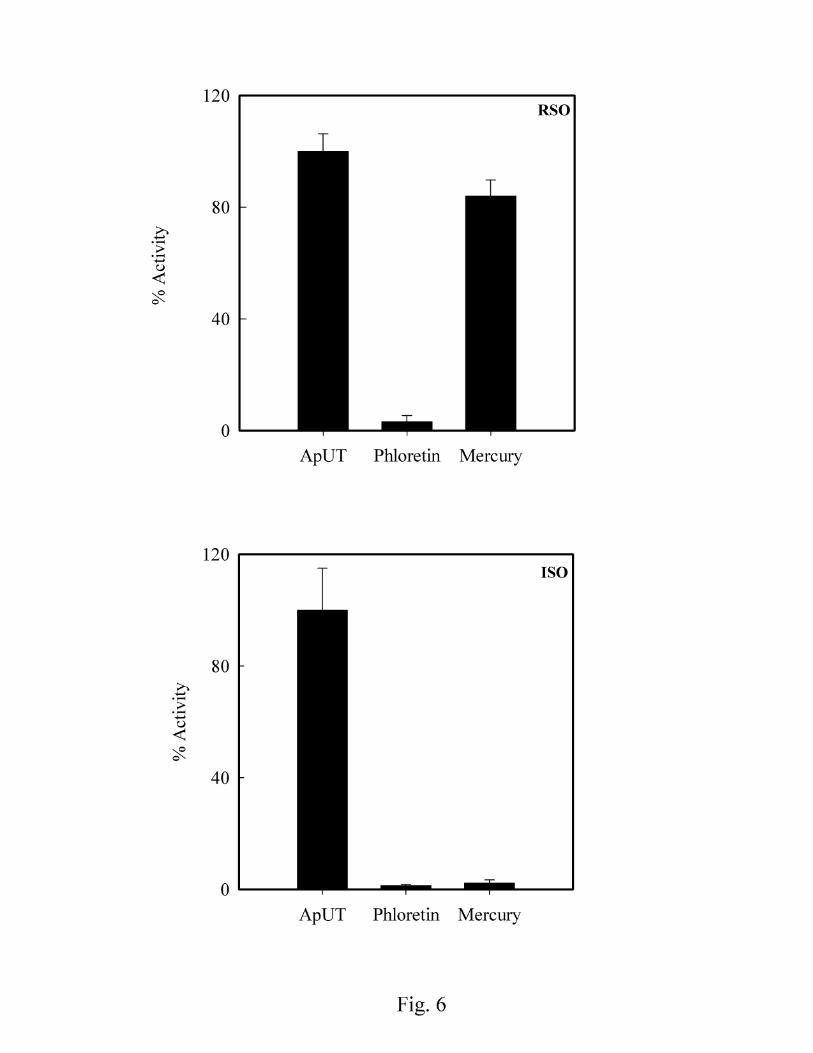
The classical urea transport inhibitor phloretin and mercury inhibited urea transport through
ApUT, Fig. 6. Interestingly mercury was found to inhibit urea transport only in inside-out vesicles.
The sidedness of urea transport inhibition by mercury suggests that the mercury binding site is
located on the intracellular side of the protein molecule. Water permeability of inside-out vesicles
was inhibited upto 75% (data not shown) upon incubation with mercury. However water
permeability in spheroplast showed very high background rate and therefore mercury inhibition
results for water permeability were not conclusive (unpublished observations). In addition, water
permeability of inside-out vesicles containing ApUT was also significantly inhibited by phloretin
(ApUT, 1.7 ± 0.2 x 10-2 cm/s and with phloretin 0.62 ± 0.22 x 10-2 cm/) (P<0.05), indicating that
both urea and water permeate through the same pathway.
DISCUSSION
Recent work has defined in some detail the genetics and molecular biology of mammalian and
bacterial urea transporters (25, 26, 30, 35). Efforts to develop a functional understanding of how
UT’s transport urea across the membrane will require a combination of detailed structural and
functional studies. Bacterial transporter proteins have in general proven to be better subjects than
their mammalian counterparts for reconstitution and subsequent detailed functional and structural
study. We have therefore performed a detailed functional characterization of a bacterial urea
transporter, ApUT from the swine pathogen, Actinobacillus pleuropneumoniae, because it bears
considerable sequence identity to mammalian UT’s. Although this microbe is a strict respiratory
tract pathogen, urease activity has been implicated in its virulence (1). Since the pathogenetic site

11
for this organism, the respiratory tract, does not have an acidic pH, it is not clear if urea uptake and
metabolism is needed as a source of nitrogen or if urea’s breakdown products, ammonia and carbon
dioxide, serve in some other capacity to establish or sustain the infection.
Several putative bacterial urea transporters have been identified by scanning the genome data
bank (25). Sequence analysis studies suggest that vertebrates acquired UT genes from bacteria and
gene duplication events gave rise to putatively 10 transmembrane segments for UTs A2 to A5 and
20 for UT-A1 (25). Sequence analysis of ApUT suggest that it is probably comprised of at least 7
transmembrane segments and shares ~26% sequence identity with mammalian urea transporters
where most of the homologues are proposed to comprise of 10 transmembrane spanning segments.
Furthermore, the tandem sequence repeats seen in eukaryotic urea transporters (LPXXTXPF) (28)
are also seen in ApUT. Like UT-A or UT-B proteins from higher organisms ApUT is not
homologous to the acid activated UreI family of urea transporters which are mostly comprised of 6
transmembrane segments. ApUT shares 33% sequence identity with a phylogenitically distant urea
transporter Yut from Yersinia pseudotuberculosis, that has been shown to transport urea (31).
Therefore, as we previously proposed based on sequence homology to characterized proteins,
ApUT functions as a urea transporter (5). Our further characterization of ApUT showed that
transport of urea was sensitive to phloretin, and mercury. Interestingly, mercury sensitivity was
observed to be polar, where urea transport of inside out vesicles was inhibited by mercury. In
contrast, in spheroplast or right-side out vesicles addition of mercury did not lead to inhibition of
urea flux suggesting that mercury inhibition site is accessible only from the cytoplasmic side. Frog
urea transporter, fUT, is inhibited by mercury and was rendered mercury insensitivity by mutating
residues C185 or H187 (37). ApUT exhibits 20% identity to fUT and 19% identity to mercury
sensitive mammalian urea transporter UT-B, where, site of mercury inhibition is not clear (15).

12
Urea analogues including acetamide, methylurea, and dimethyl urea have been shown to inhibit
UT-mediated urea transport (6, 40). Whether this is due to the capacity of the protein to transport
the analogue in question was not determined. Extending our functional analysis we observed that
ApUT did not transport a number of urea analogues, including methylamine, dimethylurea,
formamide, acetamide, methylurea, propanamide and ethylamine. Our data suggest that inhibition
is likely to be due to blockade of urea transport, without transport of the competing analogue.
However, this may not be the case for all urea transporters since Levin et. al reported transport of
acetamide, N-methylurea, and proprianamide by UT-B (14)
Whether urea transporters also transport water has been the subject of considerable interest (10,
14, 32, 39). Early studies by Macey suggested that the erythrocyte urea transporter, which we now
know to be UT-B, transports water (17). Our measurements of water flux indicate that ApUT can
increase water flow through membranes in which it is expressed, with water permeability rising to
1.83 x 10-2 cm/s at the density of expression obtained in our experiments. Similarly the mammalian
erythrocyte urea transporter, UT-B, was also shown to transport water efficiently (39). In contrast
mammalian urea transporters UT-A2 and UT-A3 do not transport water(18). In both ApUT and the
erythrocyte urea transporter, phloretin inhibition of both urea and water transport suggests that both
molecules permeate via the same pathway.
Having discovered that like UT-B, ApUT has the capacity to transport water we tested whether
ApUT mediated flux of ammonia or protons. This was important to evaluate so as to ascertain
whether ApUT might contribute to pH regulation. In particular, since urea is a polar molecule and
enters into hydrogen bonding with itself and water, there exists the potential for the formation of a
‘proton wire’ (7). We observed that the flux of ammonia or protons was not increased by
expression of ApUT, indicating that neither were transported via this protein. Interestingly, the

13
water transport protein Aquaporin 1 does not transport protons due to an electrostatic barrier within
the protein pore, therefore like water transport via AQP1, it is likely that a similar barrier exists in
UT’s and that transport of urea or water by ApUT involves hydrogen bond breakage (12). The
availability of an atomic structure for ApUT would permit this suggestion to be evaluated.
Previously, it has been difficult to assess kinetic parameters of urea transporters due to a
combination of the high concentration of urea required to approach saturation and the expression
systems used. Applying our method to this problem we were able to show that ApUT tends to
saturate, with an apparent Km of 182 mM and a Vmax of 60 s-1. In one set of experiments where
inside-out vesicles showed very high ApUT mediated urea flux, we observed Km and Vmax
values of 357 mM and 140 s-1. The Km and Vmax numbers should be treated with caution as
vesicles become unstable at higher urea concentrations. Studies performed on the mammalian
erythrocyte urea transporter UT-B have estimated the Km of UT-B to be 218 mM (16, 23).
Therefore, given the phylogenetic distance and relatively low sequence homology to UT-B our
estimate of ApUT Km seems reasonable. The measured turnover numbers for UT-B has been
reported to be 2-3 orders of magnitude faster than the fastest known carrier (erythrocyte anion
exchanger). Accordingly, a channel mediated mechanism was suggested for UT-B mediated urea
transport (19). The apparent saturability of ApUT indicates that rather than acting as a simple pore
urea is transported by this transporter either by a carrier-mediated mechanism or by a simple
channel process involving a binding site for urea. In order to distinguish between a channel or a
simple carrier mode of transport additional specific measurements of single channel fluxes or
turnover numbers are required (19, 36). The measured single channel fluxes of mammalian urea
transporters UT-A2 and UT-A3 are 100 fold less than UT-B, the erythrocyte urea transporter and

14
did not show apparent saturation kinetics, which suggest a channel mode of urea transport for
these transporters(18).
What can ApUT tell us about urea transporters from higher organisams? Given the lack of
detailed kinetic information for the UTs from higher organisms whether ApUT represents a true
paradigm of the UT family as a whole is at present difficult to determine. ApUT is a bacterial urea
transporter that functionally shares several characteristics with phylogenetically higher members of
the UT-A/UT-B family in terms of urea transport, phloretin-inhibition and transport of water.
Therefore, cautious extrapolation of the data in this paper should prove useful for future
experiments addressing detailed functional analysis of mammalian UTs. Future work will involve
reconstitution of ApUT into proteoliposomes to measure its single channel permeability and as
well as crystallization.
In summary, we have expressed and functionally reconstituted a bacterial urea transporter that
bears 26% homology to mammalian kidney urea transporters. ApUT transports urea highly
effectively, and can transport water. Urea flux exhibits saturable kinetics and is inhibited by
application of mercury to the intracellular face of the protein molecule. ApUT is highly specific
for urea and excludes urea-like solutes.

15
Text Foot Note:
# current address: Maine Medical Center, Portland, Maine 04102.
Acknowledgments:
This work was supported by a grant form NIH to MZ and JM (DK-43955, DK-48217). Funding for
JB and CPS was provided by the BBSRC. GS is funded by KR UK.

16
References:
1. Baltes N, Tonpitak W, Gerlach GF, Hennig-Pauka I, Hoffmann-Moujahid A, Ganter M, and Rothkotter HJ. Actinobacillus pleuropneumoniae iron transport and urease activity: effects on bacterial virulence and host immune response. Infect Immun 69: 472-478, 2001.
2. Beckers G, Bendt AK, Kramer R, and Burkovski A. Molecular identification of the urea uptake system and transcriptional analysis of urea transporter- and urease-encoding genes in Corynebacterium glutamicum. J Bacteriol 186: 7645-7652, 2004.
3. Beckwith CS, McGee DJ, Mobley HL, and Riley LK. Cloning, expression, and catalytic activity of Helicobacter hepaticus urease. Infect Immun 69: 5914-5920, 2001.
4. Belzer C, Stoof J, Beckwith CS, Kuipers EJ, Kusters JG, and van Vliet AH. Differential regulation of urease activity in Helicobacter hepaticus and Helicobacter pylori. Microbiology 151: 3989-3995, 2005.
5. Bosse JT, Gilmour HD, and MacInnes JI. Novel genes affecting urease acivity in Actinobacillus pleuropneumoniae. J Bacteriol 183: 1242-1247, 2001.
6. Chou CL and Knepper MA. Inhibition of urea transport in inner medullary collecting duct by phloretin and urea analogues. Am J Physiol 257: F359-365, 1989.
7. Deamer DW. Proton permeation of lipid bilayers. J Bioenerg Biomembr 19: 457-479, 1987. 8. Eaton KA, Brooks CL, Morgan DR, and Krakowka S. Essential role of urease in
pathogenesis of gastritis induced by Helicobacter pylori in gnotobiotic piglets. Infect Immun 59: 2470-2475, 1991.
9. Fenton RA, Chou CL, Stewart GS, Smith CP, and Knepper MA. Urinary concentrating defect in mice with selective deletion of phloretin-sensitive urea transporters in the renal collecting duct. Proc Natl Acad Sci U S A 101: 7469-7474, 2004.
10. Fenton RA, Howorth A, Cooper GJ, Meccariello R, Morris ID, and Smith CP. Molecular characterization of a novel UT-A urea transporter isoform (UT-A5) in testis. Am J Physiol Cell Physiol 279: C1425-1431, 2000.
11. Kaback HR. [13] Bacterial Membranes Methods in Enzymology. In: Enzyme purification and related techniques (Volume 22 ed.), edited by Jakoby WB: Academic Press, 1971, p. 99-120.
12. Kato M, Pisliakov AV, and Warshel A. The barrier for proton transport in aquaporins as a challenge for electrostatic models: the role of protein relaxation in mutational calculations. Proteins 64: 829-844, 2006.
13. Lancaster JR, Jr. and Hinkle PC. Studies of the beta-galactoside transporter in inverted membrane vesicles of Escherichia coli. II. Symmetrical binding of a dansylgalactoside induced by an electrochemical proton gradient and by lactose efflux. J Biol Chem 252: 7662-7666, 1977.
14. Levin MH, de la Fuente R, and Verkman AS. Urearetics: a small molecule screen yields nanomolar potency inhibitors of urea transporter UT-B. Faseb J 21: 551-563, 2007.
15. Lucien N, Sidoux-Walter F, Roudier N, Ripoche P, Huet M, Trinh-Trang-Tan MM, Cartron JP, and Bailly P. Antigenic and functional properties of the human red blood cell urea transporter hUT-B1. J Biol Chem 277: 34101-34108, 2002.
16. Macey RI. Transport of water and urea in red blood cells. Am J Physiol 246: C195-203, 1984.

17
17. Macey RI and Farmer RE. Inhibition of water and solute permeability in human red cells. Biochim Biophys Acta 211: 104-106, 1970.
18. Maciver B, Smith CP, Hill WG, and Zeidel ML. Functional characterization of mouse urea transporters UT-A2 and UT-A3 expressed in purified Xenopus laevis oocyte plasma membranes. Am J Physiol Renal Physiol 294: F956-964, 2008.
19. Mannuzzu LM, Moronne MM, and Macey RI. Estimate of the number of urea transport sites in erythrocyte ghosts using a hydrophobic mercurial. J Membr Biol 133: 85-97, 1993.
20. Marcus EA, Moshfegh AP, Sachs G, and Scott DR. The periplasmic alpha-carbonic anhydrase activity of Helicobacter pylori is essential for acid acclimation. J Bacteriol 187: 729-738, 2005.
21. Mathai JC, Mori S, Smith BL, Preston GM, Mohandas N, Collins M, van Zijl PC, Zeidel ML, and Agre P. Functional analysis of aquaporin-1 deficient red cells. The Colton-null phenotype. J Biol Chem 271: 1309-1313, 1996.
22. Mathai JC, Sprott GD, and Zeidel ML. Molecular mechanisms of water and solute transport across archaebacterial lipid membranes. J Biol Chem 276: 27266-27271, 2001.
23. Mayrand RR and Levitt DG. Urea and ethylene glycol-facilitated transport systems in the human red cell membrane. Saturation, competition, and asymmetry. J Gen Physiol 81: 221-237, 1983.
24. Mills J, Wyborn NR, Greenwood JA, Williams SG, and Jones CW. Characterisation of a binding-protein-dependent, active transport system for short-chain amides and urea in the methylotrophic bacterium Methylophilus methylotrophus. Eur J Biochem 251: 45-53, 1998.
25. Minocha R, Studley K, and Saier MH, Jr. The urea transporter (UT) family: bioinformatic analyses leading to structural, functional, and evolutionary predictions. Receptors Channels 9: 345-352, 2003.
26. Mobley HL, Island MD, and Hausinger RP. Molecular biology of microbial ureases. Microbiol Rev 59: 451-480, 1995.
27. Olives B, Neau P, Bailly P, Hediger MA, Rousselet G, Cartron JP, and Ripoche P. Cloning and functional expression of a urea transporter from human bone marrow cells. J Biol Chem 269: 31649-31652, 1994.
28. Rousselet G, Ripoche P, and Bailly P. Tandem sequence repeats in urea transporters: identification of an urea transporter signature sequence. Am J Physiol 270: F554-555, 1996.
29. Sambrook j, Fritsch, E.F., and Maniatis, T. Molecular cloning. A laboratory Manual. New York: Cold Spring Harbor Laboratory Press, 1989.
30. Sands JM. Mammalian urea transporters. Annu Rev Physiol 65: 543-566, 2003. 31. Sebbane F, Bury-Mone S, Cailliau K, Browaeys-Poly E, De Reuse H, and Simonet M. The
Yersinia pseudotuberculosis Yut protein, a new type of urea transporter homologous to eukaryotic channels and functionally interchangeable in vitro with the Helicobacter pylori UreI protein. Mol Microbiol 45: 1165-1174, 2002.
32. Sidoux-Walter F, Lucien N, Olives B, Gobin R, Rousselet G, Kamsteeg EJ, Ripoche P, Deen PM, Cartron JP, and Bailly P. At physiological expression levels the Kidd blood group/urea transporter protein is not a water channel. J Biol Chem 274: 30228-30235, 1999.
33. Skouloubris S, Thiberge JM, Labigne A, and De Reuse H. The Helicobacter pylori UreI protein is not involved in urease activity but is essential for bacterial survival in vivo. Infect Immun 66: 4517-4521, 1998.

18
34. Smith CP, Lee WS, Martial S, Knepper MA, You G, Sands JM, and Hediger MA. Cloning and regulation of expression of the rat kidney urea transporter (rUT2). J Clin Invest 96: 1556-1563, 1995.
35. Smith CP and Rousselet G. Facilitative urea transporters. J Membr Biol 183: 1-14, 2001. 36. Stein WD. Channels, Carriers, and Pumps. New York: Academic Press Inc., 1990. 37. Stewart GS, Smith CP, and Cooper GJ. Molecular characterization of the mercurial
sensitivity of a frog urea transporter (fUT). Am J Physiol Renal Physiol 290: F1437-1442, 2006.
38. Tsuda M, Karita M, Mizote T, Morshed MG, Okita K, and Nakazawa T. Essential role of Helicobacter pylori urease in gastric colonization: definite proof using a urease-negative mutant constructed by gene replacement. Eur J Gastroenterol Hepatol 6 Suppl 1: S49-52, 1994.
39. Yang B and Verkman AS. Analysis of double knockout mice lacking aquaporin-1 and urea transporter UT-B. Evidence for UT-B-facilitated water transport in erythrocytes. J Biol Chem 277: 36782-36786, 2002.
40. You G, Smith CP, Kanai Y, Lee WS, Stelzner M, and Hediger MA. Cloning and characterization of the vasopressin-regulated urea transporter. Nature 365: 844-847, 1993.

19
Figure legends:
Fig. 1. Immunoblot analysis of ApUT. Lysates from vesicles containing ApUT or control vesicles
lacking ApUT were analyzed by 8-16% SDS-PAGE immunoblot and protein was probed by using
anti-His antibody. The band at ~ 35 kDa represents the ApUT monomer and the higher molecular
weight band may represent ApUT multimers.
Fig.2 Urea permeability of ApUT expressing vesicles. Figure shows stopped-flow tracings
(average of 8 traces) of urea efflux from vesicles containing ApUT and control vesicles lacking
ApUT. Inset (top left) shows a magnified tracing of ApUT and inset to right shows the calculated
urea permeability. The average urea permeability of ApUT containing vesicles is ~28 fold higher
(Purea = 3.3 x10-4 cm/s) than that of the control vesicles (Purea 0.11 x 10-4). *, P<0.001.
Fig. 3. Osmotic water permeability of ApUT. Figure shows time trace of volume change of
vesicles with and without ApUT upon abrupt exposure of vesicles to a doubling of extravesicular
osmolarity. Water permeability of ApUT containing vesicles is ~ 3.5 fold higher (Pf 1.83 X 10-2
cm/s) than the control vesicles lacking ApUT (Pf 0.53 x 10-2 cm/s). * P <0.005
Fig. 4. Kinetics of urea transport. ApUT vesicles were preloaded with varying concentrations of
urea and its rate of efflux was measured. The apparent Km and Vmax values computed using
nonlinear regression method are 182 mM and 60 s-1. The Km and Vmax values computed from
Hanes plot (inset) are 174 mM and 60 s-1 respectively.
Fig. 5. Permeability of urea analogues, Ammonia and protons. Vesicles were preloaded with either
200 mM urea or various urea analogs as shown in the figure. Urea permeability is significantly
enhanced in ApUT containing vesicles (black bar) compared to control (grey bar). The
permeability of urea analogues is statistically similar for both ApUT containing vesicles and
control vesicles. B, Permeability of ApUT containing vesicles to ammonia or proton/hydroxide is

20
similar to that of control vesicles lacking ApUT. Permeability coefficients were calculated as
described in methods.
Fig. 6. Phloretin and Mercury sensitivity of urea permeability in ApUT expressing vesicles. Top,
urea permeability of right side out vesicles (RSO) is inhibited by 1 mM phloretin but not 0.5 mM
mercury. Bottom, urea permeability of inside out vesicles (ISO) is fully inhibited by mercury and
phloretin. Urea permeability (cm/s) in absence of any inhibitor is taken as 100% activity.
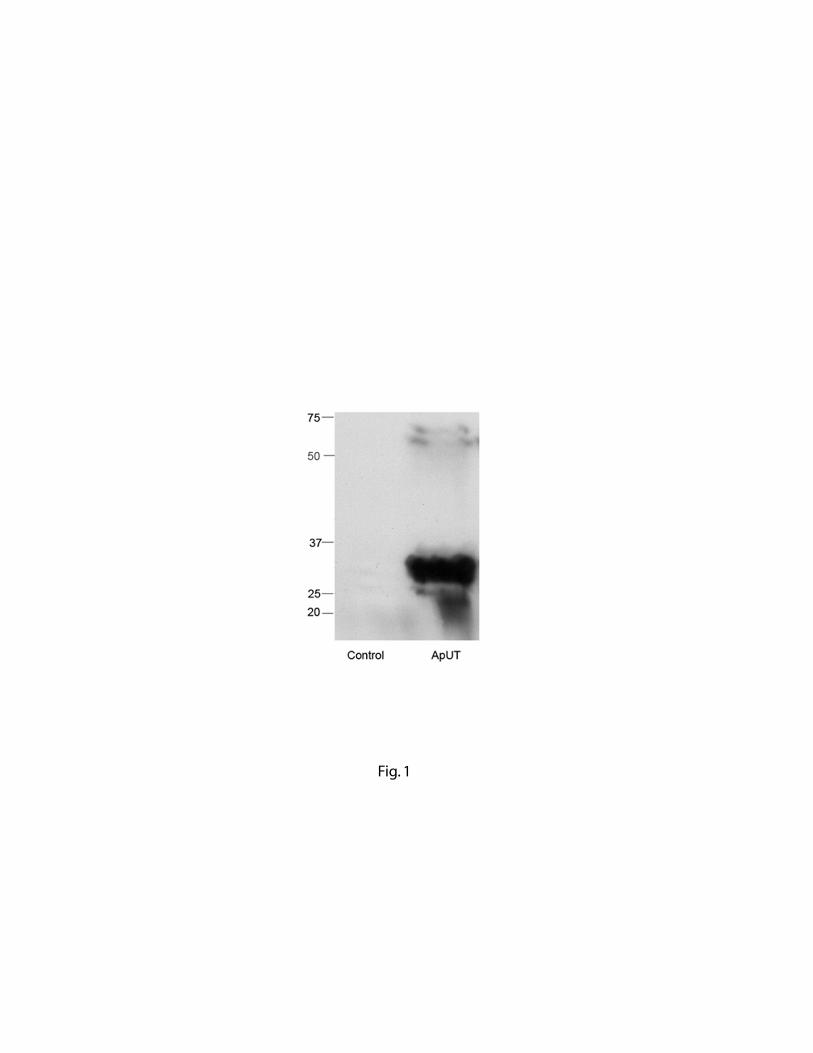
Fig. 1