
Functional and morphological evidence for a ventricular conduction system in zebrafish and Xenopus...
9
Functional and morphological evidence for a ventricular conduction system in zebrafish and Xenopus hearts David Sedmera, 1 Maria Reckova, 1,2,4 Angela deAlmeida, 1,2,4 Martina Sedmerova, 1 Martin Biermann, 2,3 Jiri Volejnik, 1 Alexandre Sarre, 4 Eric Raddatz, 4 Robert A. McCarthy, 1 Robert G. Gourdie, 1 and Robert P. Thompson 1 1 Department of Cell Biology and Anatomy, Medical University of South Carolina, Charleston, South Carolina 29425; 2 Klinik und Poliklinik fu ¨ r Nuklearmedizin, University of Mu ¨ nster, Mu ¨ nster, Germany 48161; 3 Krannert Institute of Cardiology, Indianapolis, Indiana 46202; and 4 Institute of Physiology, University of Lausanne, CH-1005 Lausanne, Switzerland Submitted 15 October 2002; accepted in final form 22 November 2002 Sedmera, David, Maria Reckova, Angela deAlmeida, Martina Sedmerova, Martin Biermann, Jiri Volejnik, Alexandre Sarre, Eric Raddatz, Robert A. McCarthy, Robert G. Gourdie, and Robert P. Thompson. Functional and morphological evidence for a ventricular conduction system in zebrafish and Xenopus hearts. Am J Physiol Heart Circ Physiol 284: H1152–H1160, 2003. First published November 27, 2002; 10.1152/ajpheart.00870.2002.—Zebrafish and Xeno- pus have become popular model organisms for studying verte- brate development of many organ systems, including the heart. However, it is not clear whether the single ventricular hearts of these species possess any equivalent of the specialized ventric- ular conduction system found in higher vertebrates. Isolated hearts of adult zebrafish (Danio rerio) and African toads (Xeno- pus laevis) were stained with voltage-sensitive dye and optically mapped in spontaneous and paced rhythms followed by histo- logical examination focusing on myocardial continuity between the atrium and the ventricle. Spread of the excitation wave through the atria was uniform with average activation times of 20 2 and 50 2 ms for zebrafish and Xenopus toads, respec- tively. After a delay of 47 8 and 414 16 ms, the ventricle became activated first in the apical region. Ectopic ventricular activation was propagated significantly more slowly (total ven- tricular activation times: 24 3 vs. 14 2 ms in zebrafish and 74 14 vs. 35 9 ms in Xenopus). Although we did not observe any histologically defined tracts of specialized conduction cells within the ventricle, there were trabecular bands with promi- nent polysialic acid-neural cell adhesion molecule staining forming direct myocardial continuity between the atrioventric- ular canal and the apex of the ventricle; i.e., the site of the epicardial breakthrough. We thus conclude that these hearts are able to achieve the apex-to-base ventricular activation pat- tern observed in higher vertebrates in the apparent absence of differentiated conduction fascicles, suggesting that the ventric- ular trabeculae serve as a functional equivalent of the His- Purkinje system. fish; frog; amphibian; electrophysiology; optical mapping; Rana; PSA-NCAM THE AFRICAN TOAD (Xenopus laevis) and zebrafish (Danio rerio) are becoming model organisms representing the amphibians and teleosteans, respectively. Recent ad- vances in molecular genetics, together with ease of experimental manipulations not feasible in higher ver- tebrates, make these species useful model systems for studying heart development and the role of different genes in heart (mal)formation and (mal)function. Al- though the structure of the heart in these species is well characterized (10, 11, 18, 32), and the frog heart has long been used as a model system for heart phys- iology (e.g., Refs. 9 and 25), it is not entirely clear how closely findings from such systems can be applied to understanding the human heart. One of the important questions about the lower ver- tebrate heart is whether it possesses pacemaking and conduction tissues comparable with those of higher vertebrates. The cardiac pacemaker of frogs and fish and its neurohumoral regulation have been found to be comparable functionally with the sinoatrial node of mammals (reviewed in Ref. 12). It is also agreed that the atrioventricular myocardium serves the same role as the atrioventricular node, i.e., to create a delay between the atrial and ventricular contraction and prevent atrial tachyarrhythmias from reaching the ventricle (1). However, controversy persists as to the existence of a fast-conducting component of the ven- tricular myocardium (21, 23, 27) or the functional equivalent of the His-Purkinje system. Most previous investigators (1, 23) searched conduc- tion system equivalents histologically, searching for pale cells arranged in bundles such as those found in the hearts of birds and mammals. This search was complicated by the fact that neither anurans nor fish possess a defined interventricular septum in which those fascicles are to be found (27). So far, we are unaware of any morphological study that would un- equivocally demonstrate the existence of specialized ventricular conduction tissue in lower vertebrates. However, the lack of a morphologically distinct structure does not necessarily imply that the function Address for reprint requests and other correspondence: D. Sed- mera, Dept. of Cell Biology and Anatomy, Medical Univ. of South Carolina, 173 Ashley Ave., BSB 601, Charleston, SC 29425 (E-mail: [email protected]). The costs of publication of this article were defrayed in part by the payment of page charges. The article must therefore be hereby marked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. Am J Physiol Heart Circ Physiol 284: H1152–H1160, 2003. First published November 27, 2002; 10.1152/ajpheart.00870.2002. 0363-6135/03 $5.00 Copyright © 2003 the American Physiological Society http://www.ajpheart.org H1152
Transcript of Functional and morphological evidence for a ventricular conduction system in zebrafish and Xenopus...

Functional and morphological evidence for a ventricularconduction system in zebrafish and Xenopus hearts
David Sedmera,1 Maria Reckova,1,2,4 Angela deAlmeida,1,2,4 Martina Sedmerova,1
Martin Biermann,2,3 Jiri Volejnik,1 Alexandre Sarre,4 Eric Raddatz,4
Robert A. McCarthy,1 Robert G. Gourdie,1 and Robert P. Thompson1
1Department of Cell Biology and Anatomy, Medical University of South Carolina,Charleston, South Carolina 29425; 2Klinik und Poliklinik fur Nuklearmedizin, University of Munster,Munster, Germany 48161; 3Krannert Institute of Cardiology, Indianapolis, Indiana 46202; and4Institute of Physiology, University of Lausanne, CH-1005 Lausanne, Switzerland
Submitted 15 October 2002; accepted in final form 22 November 2002
Sedmera, David, Maria Reckova, Angela deAlmeida,Martina Sedmerova, Martin Biermann, Jiri Volejnik,Alexandre Sarre, Eric Raddatz, Robert A. McCarthy,Robert G. Gourdie, and Robert P. Thompson. Functionaland morphological evidence for a ventricular conduction systemin zebrafish and Xenopus hearts. Am J Physiol Heart CircPhysiol 284: H1152–H1160, 2003. First published November27, 2002; 10.1152/ajpheart.00870.2002.—Zebrafish and Xeno-pus have become popular model organisms for studying verte-brate development of many organ systems, including the heart.However, it is not clear whether the single ventricular hearts ofthese species possess any equivalent of the specialized ventric-ular conduction system found in higher vertebrates. Isolatedhearts of adult zebrafish (Danio rerio) and African toads (Xeno-pus laevis) were stained with voltage-sensitive dye and opticallymapped in spontaneous and paced rhythms followed by histo-logical examination focusing on myocardial continuity betweenthe atrium and the ventricle. Spread of the excitation wavethrough the atria was uniform with average activation times of20 � 2 and 50 � 2 ms for zebrafish and Xenopus toads, respec-tively. After a delay of 47 � 8 and 414 � 16 ms, the ventriclebecame activated first in the apical region. Ectopic ventricularactivation was propagated significantly more slowly (total ven-tricular activation times: 24 � 3 vs. 14 � 2 ms in zebrafish and74 � 14 vs. 35 � 9 ms in Xenopus). Although we did not observeany histologically defined tracts of specialized conduction cellswithin the ventricle, there were trabecular bands with promi-nent polysialic acid-neural cell adhesion molecule stainingforming direct myocardial continuity between the atrioventric-ular canal and the apex of the ventricle; i.e., the site of theepicardial breakthrough. We thus conclude that these heartsare able to achieve the apex-to-base ventricular activation pat-tern observed in higher vertebrates in the apparent absence ofdifferentiated conduction fascicles, suggesting that the ventric-ular trabeculae serve as a functional equivalent of the His-Purkinje system.
fish; frog; amphibian; electrophysiology; optical mapping;Rana; PSA-NCAM
THE AFRICAN TOAD (Xenopus laevis) and zebrafish (Daniorerio) are becoming model organisms representing the
amphibians and teleosteans, respectively. Recent ad-vances in molecular genetics, together with ease ofexperimental manipulations not feasible in higher ver-tebrates, make these species useful model systems forstudying heart development and the role of differentgenes in heart (mal)formation and (mal)function. Al-though the structure of the heart in these species iswell characterized (10, 11, 18, 32), and the frog hearthas long been used as a model system for heart phys-iology (e.g., Refs. 9 and 25), it is not entirely clear howclosely findings from such systems can be applied tounderstanding the human heart.
One of the important questions about the lower ver-tebrate heart is whether it possesses pacemaking andconduction tissues comparable with those of highervertebrates. The cardiac pacemaker of frogs and fishand its neurohumoral regulation have been found to becomparable functionally with the sinoatrial node ofmammals (reviewed in Ref. 12). It is also agreed thatthe atrioventricular myocardium serves the same roleas the atrioventricular node, i.e., to create a delaybetween the atrial and ventricular contraction andprevent atrial tachyarrhythmias from reaching theventricle (1). However, controversy persists as to theexistence of a fast-conducting component of the ven-tricular myocardium (21, 23, 27) or the functionalequivalent of the His-Purkinje system.
Most previous investigators (1, 23) searched conduc-tion system equivalents histologically, searching forpale cells arranged in bundles such as those found inthe hearts of birds and mammals. This search wascomplicated by the fact that neither anurans nor fishpossess a defined interventricular septum in whichthose fascicles are to be found (27). So far, we areunaware of any morphological study that would un-equivocally demonstrate the existence of specializedventricular conduction tissue in lower vertebrates.
However, the lack of a morphologically distinctstructure does not necessarily imply that the function
Address for reprint requests and other correspondence: D. Sed-mera, Dept. of Cell Biology and Anatomy, Medical Univ. of SouthCarolina, 173 Ashley Ave., BSB 601, Charleston, SC 29425(E-mail: [email protected]).
The costs of publication of this article were defrayed in part by thepayment of page charges. The article must therefore be herebymarked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
Am J Physiol Heart Circ Physiol 284: H1152–H1160, 2003.First published November 27, 2002; 10.1152/ajpheart.00870.2002.
0363-6135/03 $5.00 Copyright © 2003 the American Physiological Society http://www.ajpheart.orgH1152

is absent, as demonstrated by developmental studiesperformed in higher vertebrates. In the chick, morpho-logically distinct bundles are not present before stage31 [7 days of incubation (33)], but specific staining canidentify the precursors of these structures much earlieras a diffuse network (6), and, indeed, the function of aHis-Purkinje network can be demonstrated by conver-sion of ventricular activation patterns at around stages29–31 [6–7 days (5)]. In the mouse, the situation issimilar, and apex-to-base activation pattern is estab-lished well before completion of ventricular septation(26), implicating ventricular trabeculae in the processof accelerated impulse propagation (34).
The question of presence or absence of functionallydefined specialized ventricular conduction tissue is ofimportance from both phylogenetic and practicalpoints of view. Is the fast-conducting component aneomorphic feature of homeothermic vertebrates ordoes it have more ancient roots? Similarly, its exis-tence would strengthen the case for using various ze-brafish mutants for the study of the genetic mecha-nisms of cardiac arrhythmias. We have therefore un-dertaken a functional study of the spread of normaland artificially induced activation in the adult heart ofzebrafish and Xenopus, coupled with histological andimmunohistochemical examination of atrioventricularconnections. We hypothesized that the existence ofspecialized conduction tissue would be revealed byapex-to-base activation pattern and by prolonged ven-tricular activation time of ectopic heartbeats. In bothspecies, we were able to document ventricular activa-tion rapidly spreading from apex to base, whereasectopic activation induced by electrical pacing tooksignificantly more time. Histological examination didnot reveal any morphologically distinct tracts of spe-cialized conduction cells. However, the ventricular tra-beculae formed straight myocardial continuity be-tween the myocardium of the atrioventricular canaland the ventricular apex, which was the earliest acti-vated region of the ventricle. We conclude that bothspecies possess the functional equivalent of a ventric-ular conduction system.
MATERIAL AND METHODS
Animals. This study conforms to the principles of Declara-tion of Helsinki and “Guiding Principles for Research Involv-ing Animals and Human Beings” of the American Physiolog-ical Society.
The zebrafish were maintained by standard methods (36).Three-year-old adult zebrafish (n � 10) were transferred to apetri dish and euthanized by rapid cutting between the headand vertebral column, followed by brain pithing. The pericar-dial cavity was opened, and the torso was transferred to0.0015% solution of 4-[�-[2-(di-n-butylamino)-6-naphthyl]vi-nyl]pyridinium (di-4-ANEPPS) with 40 �mol cytochalasin Din freshwater fish saline [composition (in g/l): 9.88 NaCl, 0.37KCl, 0.33 CaCl2, 0.14 MgCl2, 0.17 Na2HPO4, and 0.04NaH2PO4; pH 7.2] bubbled with 100% oxygen for 10 min.After a brief rinse in the same saline, the torso was pinned onthe silicone bottom of a custom-made dish, which was posi-tioned on the temperature-controlled fixed stage (Biostage
600, 20/20 Technology) of a Leica DMLB-FS upright micro-scope.
The Xenopus toads (females, 2–3 yr old, n � 5) weresupplied by Xenopus Express (Homosassa, FL) and main-tained under 12:12-h light-dark conditions. Instead of theusual euthansia by anesthetic overdose, they were rapidlydecapitated and despinalized, and their hearts were excisedand rinsed of excess blood in amphibian Ringer saline gassedwith 100% oxygen [composition (in g/l): 6.5 NaCl, 0.14 KCl,0.12 CaCl2, and 0.2 NaHCO3; pH 7.2] and stained for 10 minin 0.003% di-4-ANEPPS with 40 �mol cytochalasin D in thesame saline. The isolated heart was pinned through thedistal bulbus and apex to achieve the required orientation inthe same custom chamber positioned on a temperature-con-trolled stage of Leica MZFLIII epifluorescence microscope.For observation and additional illumination, we used aSchott KLD-1500 battery-powered light source with a greeninterference filter.
Pacing at the ventricular apex was performed after therecordings in spontaneous rhythm were obtained. A plati-num electrode (bipolar for Xenopus, pseudounipolar withanode in the tissue bath for zebrafish) was positioned at theapex of the ventricle by using a Narishige micromanipulator.The heart was stimulated in overdrive mode at 125% of theintrinsic rate with 2-ms pulses twice the diastolic threshold,which was between 1.5 and 2.5 mA for both species.
Experimental protocol. All experiments were performed at26°C (standard laboratory temperature to which the animalswere adapted) with continuous bubbling of the bath with100% oxygen, which was interrupted only during recording.Under these conditions, the hearts were electrophysiologi-cally stable (spontaneously beating in regular rhythm) for atleast 1 h; however, no more than 3 min of total illuminationwith the green light was possible because of the developmentof arrhythmias and photobleaching. For these reasons, onlythe recordings from the first 2 min were used for analysis;therefore, not all possible positions (dorsal, ventral, left, andright lateral views for Xenopus; ventral and both lateralviews for zebrafish) were obtained from all hearts. Addition-ally, five zebrafish hearts were cut open with fine scissors (2in frontal, 3 in sagittal plane) following initial imaging, andthe hearts were restained and imaged again to reveal endo-cardial activation patterns.
Optical mapping was performed by illuminating the spec-imen with green light (546 � 10 nm). Emitted light waspassed through a 590-nm LP barrier filter and detected witha 12-bit intensified Neurocam camera (EG&G-Wallac/Olym-pus). The decrease in intensity of emitted fluorescence corre-sponds to changes in membrane potential (17). The magnifi-cations used resulted in a spatial resolution of 78 �m/pixelfor Xenopus and 19 �m/pixel for zebrafish in the binned modeused for data analysis. The frame rates used were either 500(full resolution, 80 � 80 pixels) or 1,000 (with binning)frames per second depending on camera settings.
Data processing. The recordings were digitally processedby using UltraView and Universal Mapping software. Atypical cardiac cycle was selected (usually the first in eachrecording, because it usually had the best signal-to-noiseratio, but there were no beat-to-beat differences in any indi-vidual heart), and a time course of fluorescence intensityfrom individual pixels was digitally filtered by using a But-terworth low-pass (100 Hz) filter. The first derivative wascomputed (dV/dt), and its peak corresponding to maximaldV/dt was used for detection of the action potential upstroke.To account for virtual electrode effect, the pixels adjacent tothe stimulation site were excluded from analysis. Isochronalcontour maps of activation were constructed and superim-
H1153VENTRICULAR CONDUCTION SYSTEM IN LOWER VERTEBRATES
AJP-Heart Circ Physiol • VOL 284 • APRIL 2003 • www.ajpheart.org

posed on images of the hearts taken at maximal spatialresolution of the camera.
Electrophysiological recordings. To validate our data usingstandard electrophysiological techniques, five green frogs(Rana clamitans) were prepared by brain and spinal cordpithing for electrocardiographic recording. The active elec-trode was clamped on the ventricular apex, and a referenceelectrode was positioned in the mouth. The recordings wereperformed at room temperature (25°C), and the skin waskept moist with paper towels soaked with tap water. Ven-tricular extrasystole was delivered by using a silver pseudou-nipolar suction electrode positioned at the ventricular apexat the end of T wave during continuous ECG recordings usingcustom software running on a Macintosh LC II computer.ECG intervals were then measured on digital recordings ofspontaneous and paced beats.
Histological examination. For morphological evaluation,the hearts were fixed after the experiments in Dent’s fixative(80% methanol-20% DMSO) and processed into paraffin. Sec-tions at 5 �m thickness were cut in series in frontal, trans-verse, and sagittal planes and stained with hematoxylin-eosin. Selected sister sections were stained with antibodiesdirected against smooth muscle actin (1:1,000, Sigma) ormyosin (9:1, MF20, Developmental Studies HybridomaBank) diluted with 10 mM Tris buffer, 0.9% NaCl, 0.001%sodium azide, 2% bovine serum albumin, and 0.1% Triton-X.The antibody binding was detected using Cy2-coupled goat-anti-mouse secondary antibody (1:200, Jackson Immuno),and the nuclei were counterstained with propidium iodide(1:10,000). Polysaliac-acid neural adhesion molecule (PSA-NCAM) IgM monoclonal antibody (1:200, 5A5, Developmen-tal Studies Hybridoma Bank), a marker for developing con-duction system in the embryonic chick (6) was detected byCy5-coupled anti-mouse IgM secondary antibody (JacksonImmuno). Observations were made on a Bio-Rad MRC1024confocal microscope using appropriate lasers and filter sets.In addition, we refer to scanning electron micrographs fromour earlier studies (10, 11, 29). Because of its size, overviewpictures of the Xenopus heart were obtained by scanning theslides at 2,400 dpi in transparency mode using a UMAXPowerLook II scanner.
Statistical evaluation. All values are expressed asmeans � SD. For statistical comparisons of spontaneousversus paced activation time, we used a paired t-test withvalues of P � 0.05 considered as significant.
RESULTS
Zebrafish. The heart rate and other parameters ofisolated heart function are expressed in Table 1. Theactivation of the atria progressed from its junction withthe sinus venosus in an isotropic manner. The averagetime necessary for activation of the entire atrium (mea-sured as a temporal difference between activation ofthe first and the last pixel) was 20 � 2 ms (n � 10).After a delay caused by slow conduction through the
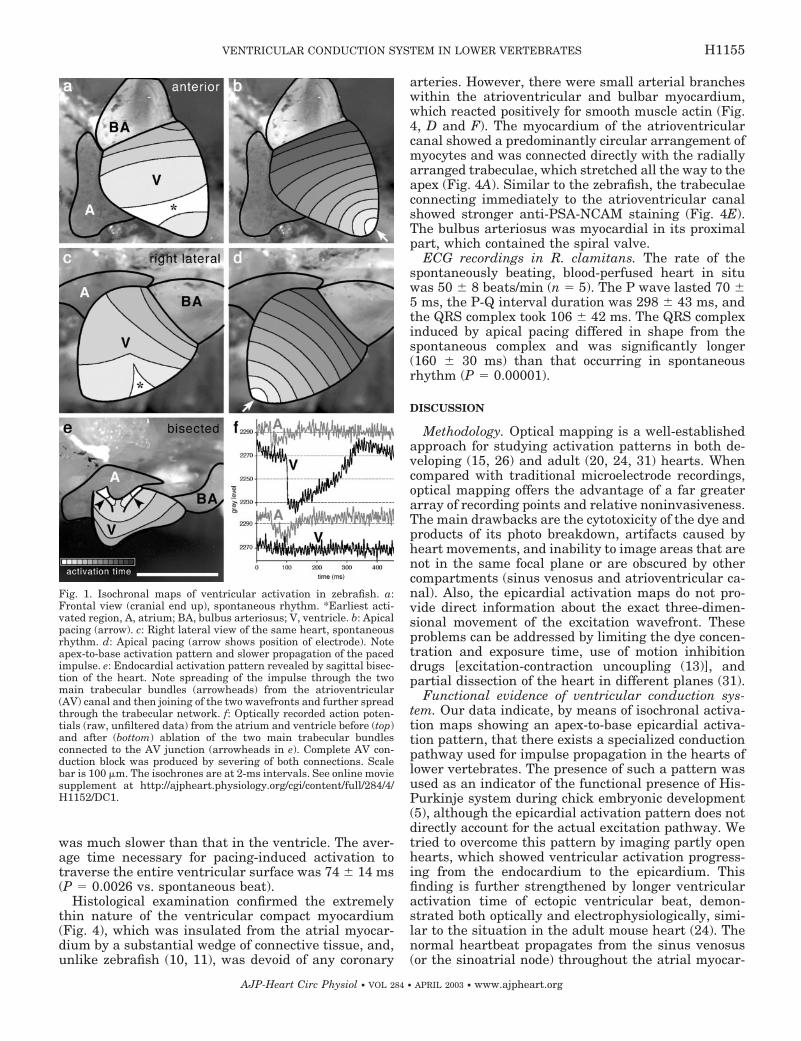
atrioventricular canal (Table 1), the ventricle first be-came activated in the apical region (Fig. 1). This direc-tionality could be discriminated in all views. On aver-age, 14 � 2 ms were necessary for the activation of theentire ventricular surface. The activation wave elicitedby ectopic ventricular pacing propagated across theventricular surface isotropically and took 24 � 3 ms(P � 0.0001 vs. spontaneous beat) to traverse theventricle. The propagation of the action potentialacross the ventricular surface can be seen in the onlinemovie supplement.1 Imaging of the exposed endocar-dial surfaces revealed the activation wave spreadingrapidly through the radial trabecular network towardthe outer contour of the heart (Fig. 1E). Attempts ofablation of the two connections between the atrioven-tricular canal myocardium and the main trabecularbundles (Figs. 1E and 2, E and F) showed rapid com-pensation by impulse propagating from the other con-nection, indicating a degree of redundancy. If bothconnections were severed, complete conduction blockensued (Fig. 1F).
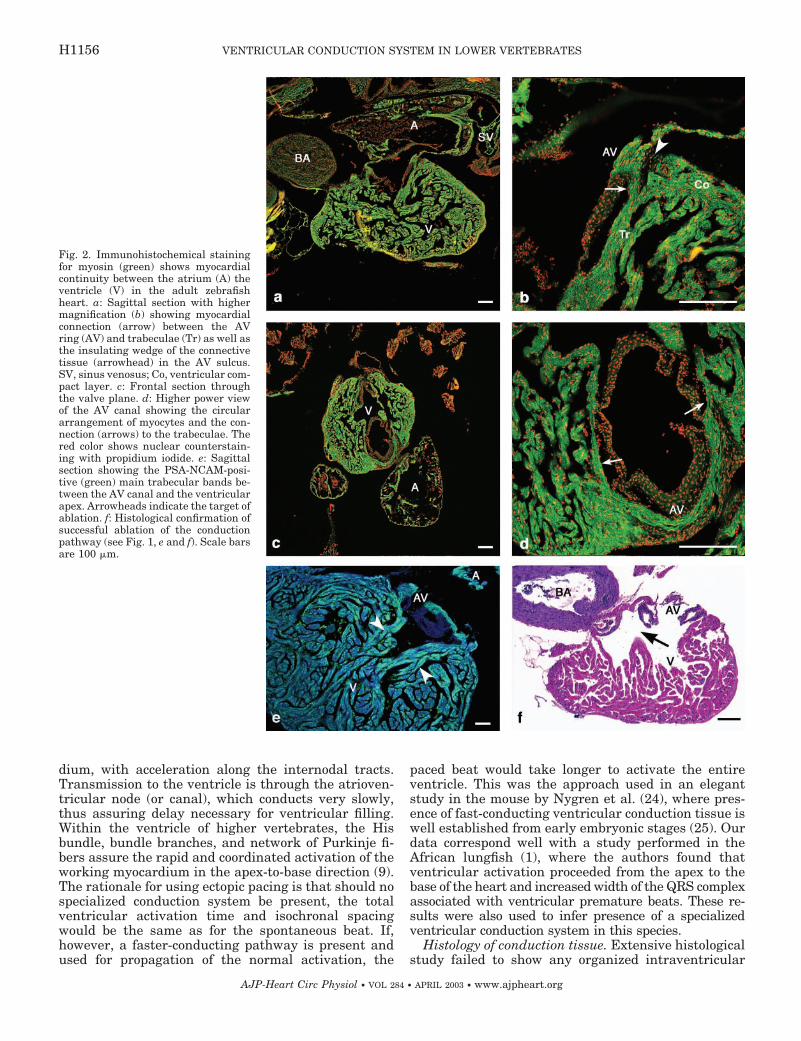
Histological examination showed myocardial conti-nuity among the sinus venosus, atrium, and the ven-tricle (Fig. 2). There were two distinct trabecular bandsrunning from the anterior and posterior side of theatrioventricular canal on each side, which then joinedtogether (Fig. 2E). These bands showed stronger PSA-NCAM staining than the remaining trabeculae. Thebulboventricular valve was surrounded by myocar-dium (Fig. 2C); however, the bulbus arteriosus wasentirely nonmyocardial (Fig. 2A). The arrangement ofthe myocytes within the atrioventricular canal waspredominantly circumferential and the canal con-nected directly to the ventricular trabeculae, althoughthere was a wedge of connective tissue separating itfrom the outer compact myocardium (Fig. 2B).
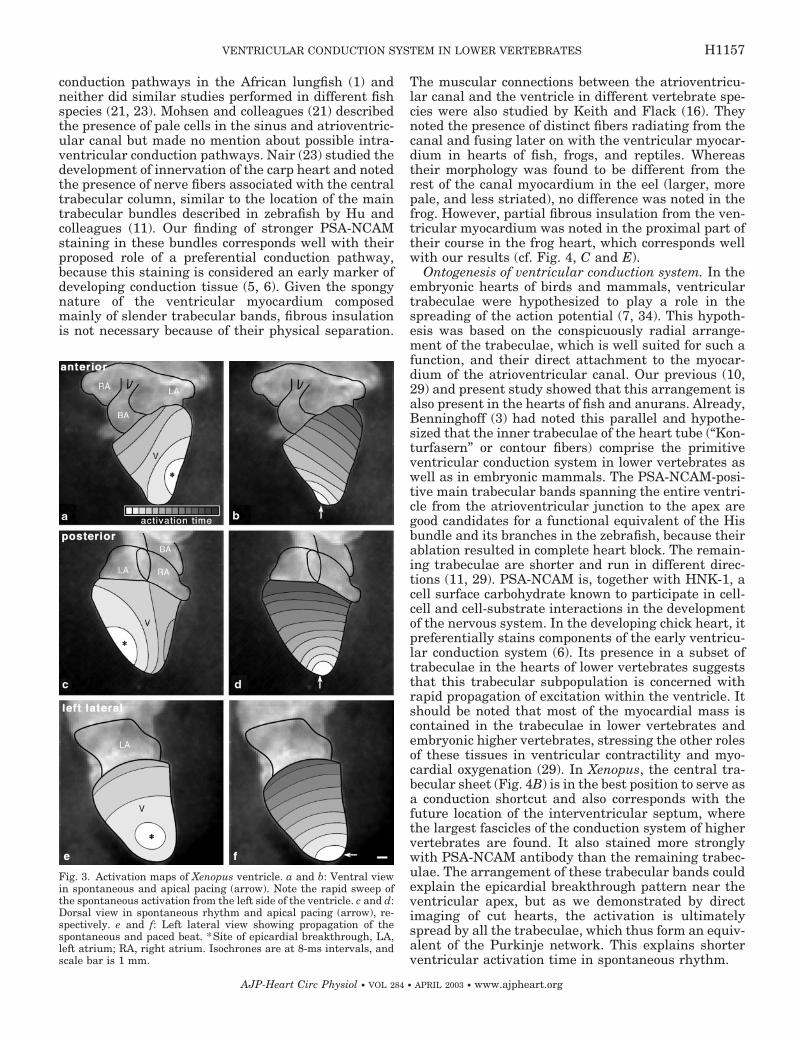
Xenopus. The functional parameters of isolatedhearts are summarized in Table 1. The activation ofthe atrium spread isotropically from the sinoatrialjunction toward the atrioventricular canal and took50 � 2 ms (n � 5). The first activated region of theventricle was located near the apex on the left side(Fig. 3). From there, the wave of excitation spread overand around the ventricle toward the bulbus, taking35 � 9 ms to activate the entire ventricular surface.The action potential obtained from the bulbus was ofsmall amplitude and difficult to interpret due to com-plex spatial arrangement, but the spread of activation
1See http://ajpheart.physiology.org/cgi/content/full/284/4/H1152/DC1.
Table 1. Electrophysiological parameters of adult isolated zebrafish and Xenopus hearts
SpeciesHeart Rate,beats/min
Atrial ActivationTime, ms
AV Delay,ms
Ventricular ActivationTime, ms
Spontaneous Paced
Zebrafish (Danio rerio) 123�12 20�2 47�8 14�2 24�3*African toad (Xenopus laevis) 67�10 50�2 414�16 35�9 74�14*
Values are means � SD; n � 10 zebrafish hearts 5 and Xenopus hearts. AV, atrioventricular. *P � 0.01 vs. spontaneous beat.
H1154 VENTRICULAR CONDUCTION SYSTEM IN LOWER VERTEBRATES
AJP-Heart Circ Physiol • VOL 284 • APRIL 2003 • www.ajpheart.org
was much slower than that in the ventricle. The aver-age time necessary for pacing-induced activation totraverse the entire ventricular surface was 74 � 14 ms(P � 0.0026 vs. spontaneous beat).
Histological examination confirmed the extremelythin nature of the ventricular compact myocardium(Fig. 4), which was insulated from the atrial myocar-dium by a substantial wedge of connective tissue, and,unlike zebrafish (10, 11), was devoid of any coronary
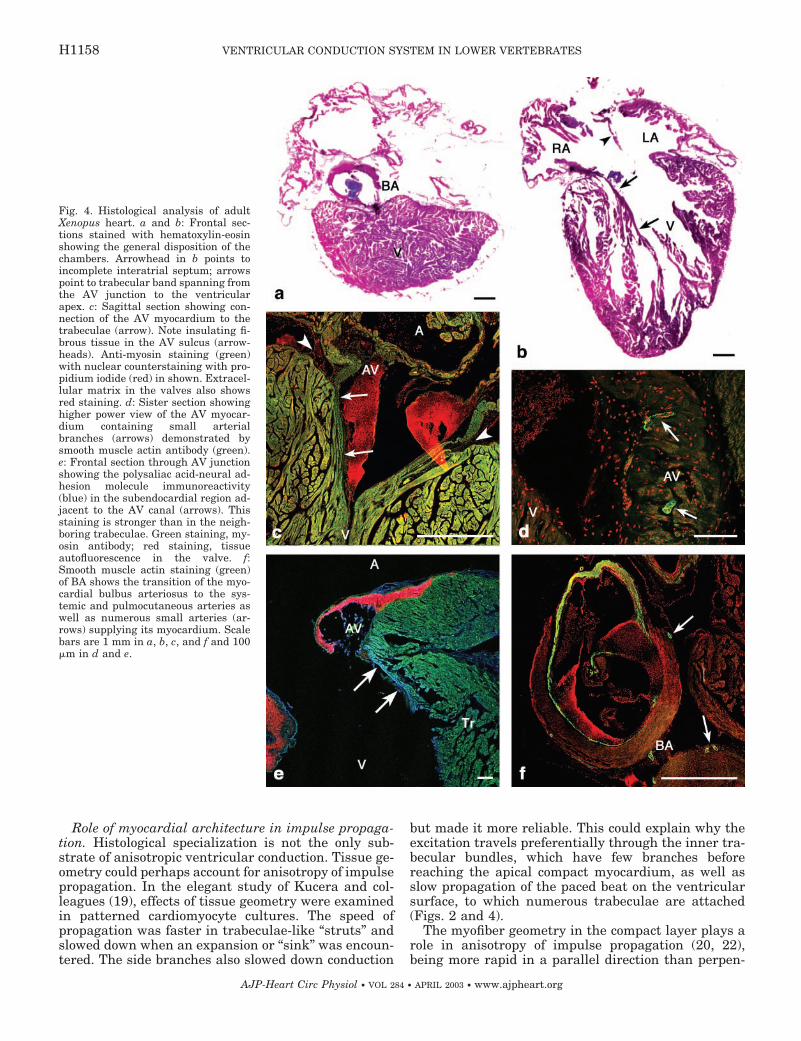
arteries. However, there were small arterial brancheswithin the atrioventricular and bulbar myocardium,which reacted positively for smooth muscle actin (Fig.4, D and F). The myocardium of the atrioventricularcanal showed a predominantly circular arrangement ofmyocytes and was connected directly with the radiallyarranged trabeculae, which stretched all the way to theapex (Fig. 4A). Similar to the zebrafish, the trabeculaeconnecting immediately to the atrioventricular canalshowed stronger anti-PSA-NCAM staining (Fig. 4E).The bulbus arteriosus was myocardial in its proximalpart, which contained the spiral valve.
ECG recordings in R. clamitans. The rate of thespontaneously beating, blood-perfused heart in situwas 50 � 8 beats/min (n � 5). The P wave lasted 70 �5 ms, the P-Q interval duration was 298 � 43 ms, andthe QRS complex took 106 � 42 ms. The QRS complexinduced by apical pacing differed in shape from thespontaneous complex and was significantly longer(160 � 30 ms) than that occurring in spontaneousrhythm (P � 0.00001).
DISCUSSION
Methodology. Optical mapping is a well-establishedapproach for studying activation patterns in both de-veloping (15, 26) and adult (20, 24, 31) hearts. Whencompared with traditional microelectrode recordings,optical mapping offers the advantage of a far greaterarray of recording points and relative noninvasiveness.The main drawbacks are the cytotoxicity of the dye andproducts of its photo breakdown, artifacts caused byheart movements, and inability to image areas that arenot in the same focal plane or are obscured by othercompartments (sinus venosus and atrioventricular ca-nal). Also, the epicardial activation maps do not pro-vide direct information about the exact three-dimen-sional movement of the excitation wavefront. Theseproblems can be addressed by limiting the dye concen-tration and exposure time, use of motion inhibitiondrugs [excitation-contraction uncoupling (13)], andpartial dissection of the heart in different planes (31).
Functional evidence of ventricular conduction sys-tem. Our data indicate, by means of isochronal activa-tion maps showing an apex-to-base epicardial activa-tion pattern, that there exists a specialized conductionpathway used for impulse propagation in the hearts oflower vertebrates. The presence of such a pattern wasused as an indicator of the functional presence of His-Purkinje system during chick embryonic development(5), although the epicardial activation pattern does notdirectly account for the actual excitation pathway. Wetried to overcome this pattern by imaging partly openhearts, which showed ventricular activation progress-ing from the endocardium to the epicardium. Thisfinding is further strengthened by longer ventricularactivation time of ectopic ventricular beat, demon-strated both optically and electrophysiologically, simi-lar to the situation in the adult mouse heart (24). Thenormal heartbeat propagates from the sinus venosus(or the sinoatrial node) throughout the atrial myocar-
Fig. 1. Isochronal maps of ventricular activation in zebrafish. a:Frontal view (cranial end up), spontaneous rhythm. *Earliest acti-vated region, A, atrium; BA, bulbus arteriosus; V, ventricle. b: Apicalpacing (arrow). c: Right lateral view of the same heart, spontaneousrhythm. d: Apical pacing (arrow shows position of electrode). Noteapex-to-base activation pattern and slower propagation of the pacedimpulse. e: Endocardial activation pattern revealed by sagittal bisec-tion of the heart. Note spreading of the impulse through the twomain trabecular bundles (arrowheads) from the atrioventricular(AV) canal and then joining of the two wavefronts and further spreadthrough the trabecular network. f: Optically recorded action poten-tials (raw, unfiltered data) from the atrium and ventricle before (top)and after (bottom) ablation of the two main trabecular bundlesconnected to the AV junction (arrowheads in e). Complete AV con-duction block was produced by severing of both connections. Scalebar is 100 �m. The isochrones are at 2-ms intervals. See online moviesupplement at http://ajpheart.physiology.org/cgi/content/full/284/4/H1152/DC1.
H1155VENTRICULAR CONDUCTION SYSTEM IN LOWER VERTEBRATES
AJP-Heart Circ Physiol • VOL 284 • APRIL 2003 • www.ajpheart.org
dium, with acceleration along the internodal tracts.Transmission to the ventricle is through the atrioven-tricular node (or canal), which conducts very slowly,thus assuring delay necessary for ventricular filling.Within the ventricle of higher vertebrates, the Hisbundle, bundle branches, and network of Purkinje fi-bers assure the rapid and coordinated activation of theworking myocardium in the apex-to-base direction (9).The rationale for using ectopic pacing is that should nospecialized conduction system be present, the totalventricular activation time and isochronal spacingwould be the same as for the spontaneous beat. If,however, a faster-conducting pathway is present andused for propagation of the normal activation, the
paced beat would take longer to activate the entireventricle. This was the approach used in an elegantstudy in the mouse by Nygren et al. (24), where pres-ence of fast-conducting ventricular conduction tissue iswell established from early embryonic stages (25). Ourdata correspond well with a study performed in theAfrican lungfish (1), where the authors found thatventricular activation proceeded from the apex to thebase of the heart and increased width of the QRS complexassociated with ventricular premature beats. These re-sults were also used to infer presence of a specializedventricular conduction system in this species.
Histology of conduction tissue. Extensive histologicalstudy failed to show any organized intraventricular
Fig. 2. Immunohistochemical stainingfor myosin (green) shows myocardialcontinuity between the atrium (A) theventricle (V) in the adult zebrafishheart. a: Sagittal section with highermagnification (b) showing myocardialconnection (arrow) between the AVring (AV) and trabeculae (Tr) as well asthe insulating wedge of the connectivetissue (arrowhead) in the AV sulcus.SV, sinus venosus; Co, ventricular com-pact layer. c: Frontal section throughthe valve plane. d: Higher power viewof the AV canal showing the circulararrangement of myocytes and the con-nection (arrows) to the trabeculae. Thered color shows nuclear counterstain-ing with propidium iodide. e: Sagittalsection showing the PSA-NCAM-posi-tive (green) main trabecular bands be-tween the AV canal and the ventricularapex. Arrowheads indicate the target ofablation. f: Histological confirmation ofsuccessful ablation of the conductionpathway (see Fig. 1, e and f). Scale barsare 100 �m.
H1156 VENTRICULAR CONDUCTION SYSTEM IN LOWER VERTEBRATES
AJP-Heart Circ Physiol • VOL 284 • APRIL 2003 • www.ajpheart.org
conduction pathways in the African lungfish (1) andneither did similar studies performed in different fishspecies (21, 23). Mohsen and colleagues (21) describedthe presence of pale cells in the sinus and atrioventric-ular canal but made no mention about possible intra-ventricular conduction pathways. Nair (23) studied thedevelopment of innervation of the carp heart and notedthe presence of nerve fibers associated with the centraltrabecular column, similar to the location of the maintrabecular bundles described in zebrafish by Hu andcolleagues (11). Our finding of stronger PSA-NCAMstaining in these bundles corresponds well with theirproposed role of a preferential conduction pathway,because this staining is considered an early marker ofdeveloping conduction tissue (5, 6). Given the spongynature of the ventricular myocardium composedmainly of slender trabecular bands, fibrous insulationis not necessary because of their physical separation.
The muscular connections between the atrioventricu-lar canal and the ventricle in different vertebrate spe-cies were also studied by Keith and Flack (16). Theynoted the presence of distinct fibers radiating from thecanal and fusing later on with the ventricular myocar-dium in hearts of fish, frogs, and reptiles. Whereastheir morphology was found to be different from therest of the canal myocardium in the eel (larger, morepale, and less striated), no difference was noted in thefrog. However, partial fibrous insulation from the ven-tricular myocardium was noted in the proximal part oftheir course in the frog heart, which corresponds wellwith our results (cf. Fig. 4, C and E).
Ontogenesis of ventricular conduction system. In theembryonic hearts of birds and mammals, ventriculartrabeculae were hypothesized to play a role in thespreading of the action potential (7, 34). This hypoth-esis was based on the conspicuously radial arrange-ment of the trabeculae, which is well suited for such afunction, and their direct attachment to the myocar-dium of the atrioventricular canal. Our previous (10,29) and present study showed that this arrangement isalso present in the hearts of fish and anurans. Already,Benninghoff (3) had noted this parallel and hypothe-sized that the inner trabeculae of the heart tube (“Kon-turfasern” or contour fibers) comprise the primitiveventricular conduction system in lower vertebrates aswell as in embryonic mammals. The PSA-NCAM-posi-tive main trabecular bands spanning the entire ventri-cle from the atrioventricular junction to the apex aregood candidates for a functional equivalent of the Hisbundle and its branches in the zebrafish, because theirablation resulted in complete heart block. The remain-ing trabeculae are shorter and run in different direc-tions (11, 29). PSA-NCAM is, together with HNK-1, acell surface carbohydrate known to participate in cell-cell and cell-substrate interactions in the developmentof the nervous system. In the developing chick heart, itpreferentially stains components of the early ventricu-lar conduction system (6). Its presence in a subset oftrabeculae in the hearts of lower vertebrates suggeststhat this trabecular subpopulation is concerned withrapid propagation of excitation within the ventricle. Itshould be noted that most of the myocardial mass iscontained in the trabeculae in lower vertebrates andembryonic higher vertebrates, stressing the other rolesof these tissues in ventricular contractility and myo-cardial oxygenation (29). In Xenopus, the central tra-becular sheet (Fig. 4B) is in the best position to serve asa conduction shortcut and also corresponds with thefuture location of the interventricular septum, wherethe largest fascicles of the conduction system of highervertebrates are found. It also stained more stronglywith PSA-NCAM antibody than the remaining trabec-ulae. The arrangement of these trabecular bands couldexplain the epicardial breakthrough pattern near theventricular apex, but as we demonstrated by directimaging of cut hearts, the activation is ultimatelyspread by all the trabeculae, which thus form an equiv-alent of the Purkinje network. This explains shorterventricular activation time in spontaneous rhythm.
Fig. 3. Activation maps of Xenopus ventricle. a and b: Ventral viewin spontaneous and apical pacing (arrow). Note the rapid sweep ofthe spontaneous activation from the left side of the ventricle. c and d:Dorsal view in spontaneous rhythm and apical pacing (arrow), re-spectively. e and f: Left lateral view showing propagation of thespontaneous and paced beat. *Site of epicardial breakthrough, LA,left atrium; RA, right atrium. Isochrones are at 8-ms intervals, andscale bar is 1 mm.
H1157VENTRICULAR CONDUCTION SYSTEM IN LOWER VERTEBRATES
AJP-Heart Circ Physiol • VOL 284 • APRIL 2003 • www.ajpheart.org
Role of myocardial architecture in impulse propaga-tion. Histological specialization is not the only sub-strate of anisotropic ventricular conduction. Tissue ge-ometry could perhaps account for anisotropy of impulsepropagation. In the elegant study of Kucera and col-leagues (19), effects of tissue geometry were examinedin patterned cardiomyocyte cultures. The speed ofpropagation was faster in trabeculae-like “struts” andslowed down when an expansion or “sink” was encoun-tered. The side branches also slowed down conduction
but made it more reliable. This could explain why theexcitation travels preferentially through the inner tra-becular bundles, which have few branches beforereaching the apical compact myocardium, as well asslow propagation of the paced beat on the ventricularsurface, to which numerous trabeculae are attached(Figs. 2 and 4).
The myofiber geometry in the compact layer plays arole in anisotropy of impulse propagation (20, 22),being more rapid in a parallel direction than perpen-
Fig. 4. Histological analysis of adultXenopus heart. a and b: Frontal sec-tions stained with hematoxylin-eosinshowing the general disposition of thechambers. Arrowhead in b points toincomplete interatrial septum; arrowspoint to trabecular band spanning fromthe AV junction to the ventricularapex. c: Sagittal section showing con-nection of the AV myocardium to thetrabeculae (arrow). Note insulating fi-brous tissue in the AV sulcus (arrow-heads). Anti-myosin staining (green)with nuclear counterstaining with pro-pidium iodide (red) in shown. Extracel-lular matrix in the valves also showsred staining. d: Sister section showinghigher power view of the AV myocar-dium containing small arterialbranches (arrows) demonstrated bysmooth muscle actin antibody (green).e: Frontal section through AV junctionshowing the polysaliac acid-neural ad-hesion molecule immunoreactivity(blue) in the subendocardial region ad-jacent to the AV canal (arrows). Thisstaining is stronger than in the neigh-boring trabeculae. Green staining, my-osin antibody; red staining, tissueautofluorescence in the valve. f:Smooth muscle actin staining (green)of BA shows the transition of the myo-cardial bulbus arteriosus to the sys-temic and pulmocutaneous arteries aswell as numerous small arteries (ar-rows) supplying its myocardium. Scalebars are 1 mm in a, b, c, and f and 100�m in d and e.
H1158 VENTRICULAR CONDUCTION SYSTEM IN LOWER VERTEBRATES
AJP-Heart Circ Physiol • VOL 284 • APRIL 2003 • www.ajpheart.org

dicular to the predominant bundle orientation. Theorientation of muscle fiber in large marine fish heartswas described by Sanchez-Quintana and Hurle (28),and the orderly alignment of the trabeculae in thedogfish shark supported their role in impulse propaga-tion, but the interspecies differences were consider-able. Unlike in mammals, neither species examinedshowed distinct spiraling in the compact layer, whichcorrelates with the observed isotropy of propagation ofthe paced beat. Furthermore, the compact myocardiumis considerably thinner in the species examined in ourstudy, making the existence of spiral systems thatusually arise with complex organization of the compactlayer (14) even less likely. Indeed, varying the positionof the stimulation electrode on the ventricular surfacedid not change the isochronal spacing or total ventric-ular activation time (unpublished data) in contrast tothe situation in the mouse (22).
Heart blood supply in lower vertebrates. The anuranventricle is considered to be “coronary less” (8); how-ever, two coronary trunks originating from the carotidarch were described to spread over the thick myocar-dial wall of bulbus. In this study, we report the exis-tence of additional branches, highlighted by anti-smooth muscle actin immunostaining, in the similarlythick myocardium of the atrioventricular canal. Thisfeature can explain the development of atrioventricu-lar block after 1 h in culture and larger atrioventricu-lar delay (over 400 ms) compared with the 300-ms P-Qinterval of the blood-perfused R. clamitans heart insitu at comparable heart rates. It seems that slowconduction through these two regions is more depen-dent on oxygen concentration. The alternating ar-rangement of fast (atria and ventricle) and slow-con-ducting segments (atrioventricular canal and bulbusarteriosus) is remarkably similar to the embryonicchick heart (7). Considerably longer atrioventriculardelay between atrial and ventricular contraction inanurans compared with zebrafish could be explainedby the necessity of maintaining strictly laminar bloodflow in these larger hearts to assure efficient compart-mentalization of oxygenated and deoxygenated bloodwithin the ventricle (4). On the other hand, the longerduration of the QRS complex in R. clamitans comparedwith Xenopus can be explained by methodological dif-ference; whereas ECG gives a more precise estimate oftotal activation time of the ventricular myocardium, itis impossible to image the entire ventricle (includinginside) from any given view using optical mapping.
Concluding comments. The existence of the func-tional equivalent of a specialized network of conductioncells (albeit diffuse) in the ventricle of lower verte-brates has interesting phylogenetic and mechanisticimplications. First, it suggests that the emergence ofpathways for preferential spread of electrical excita-tion in the ventricle is not a recent innovation in highervertebrates. In this respect, it is noteworthy that theexistence of myocytes specialized for conduction havebeen described in the primitive tube heart of the tuni-cate Ciona (2), a genus whose long extinct ancestorsare thought to sit at the base of the chordate family
tree. Second, the demonstration of an organized net-work of conduction cells in the lower vertebrate heartcould present new opportunities for deepening our un-derstanding of the developmental biology of such tis-sues. Indeed, because proper heart function is notrequired for initial survival in zebrafish, this modelcould be exploited to study morphological and electro-physiological heart phenotypes that would cause earlylethality in mammalian models (30, 35). Such innateadvantages, together with powerful tools already avail-able for probing genetics in zebrafish, could provide abasis for exciting new advances in this field in comingyears.
We thank Ambuja Bale and Dr. John Woodward for providing uswith the Xenopus for these experiments and Dr. Jose Maria Perez-Pomares for helpful comments on the paper. D. Sedmera is particu-larly thankful to Dr. Lukas Kappenberger for introduction to theidea of ectopic heart pacing.
This work was supported by National Institutes of Health (NIH)Grant RO1 HL-50582 (to R. P. Thompson), National Science Foun-dation Shared Instrumentation Grant and NIH Grant PO1 HD-39946 (to R. G. Gourdie), and a University Research Committeegrant (to D. Sedmera). D. Sedmera was also supported by SouthCarolina Center of Biomedical Research Excellence Grant RR 16434-01. M. Biermann was supported by NIH, Indiana University, Deut-sche Forschungsgemenischaft, and Westfalische Wilhems-Universi-tat, Munster.
REFERENCES
1. Arbel ER, Liberthson R, Langendorf R, Pick A, Lev M, andFishman AP. Electrophysiological and anatomical observationson the heart of the African lungfish. Am J Physiol Heart CircPhysiol 232: H24–H34, 1977.
2. Becker DL, Cook JE, Davies CS, Evans WH, and GourdieRG. Expression of major gap junction connexin types in theworking myocardium of eight chordates. Cell Biol Int 22: 527–543, 1998.
3. Benninghoff A. Uber die Beziehung des Reitzleitungssystemsund der Papillarmusklen zu den Konturfasern des Her-zschlauches. Anat Anz 57: 185–208, 1923.
4. Burggren WW. Cardiac design in lower vertebrates: what canphylogeny reveal about ontogeny? Experientia 44: 919–930,1988.
5. Chuck ET, Freeman DM, Watanabe M, and RosenbaumDS. Changing activation sequence in the embryonic chick heart.Implications for the development of the His-Purkinje system.Circ Res 81: 470–476, 1997.
6. Chuck ET and Watanabe M. Differential expression of PSA-NCAM and HNK-1 epitopes in the developing cardiac conductionsystem of the chick. Dev Dyn 209: 182–195, 1997.
7. de Jong F, Opthof T, Wilde AA, Janse MJ, Charles R,Lamers WH, and Moorman AF. Persisting zones of slow im-pulse conduction in developing chicken hearts. Circ Res 71:240–250, 1992.
8. de la Cruz MV, Moreno-Rodriguez R, and Angelini P.Phylogeny of the coronary arteries. In: Coronary Artery Anoma-lies: a Comprehensive Approach, edited by Angelini P. Philadel-phia, PA: Lippincott Williams & Wilkins, 1999, p. 1–9.
9. Gattuso A, Mazza R, Pellegrino D, and Tota B. Endocardialendothelium mediates luminal ACh-NO signaling in isolatedfrog heart. Am J Physiol Heart Circ Physiol 276: H633–H641,1999.
10. Hu N, Sedmera D, Yost HJ, and Clark EB. Structure andfunction of the developing zebrafish heart. Anat Rec 260: 148–157, 2000.
11. Hu N, Yost HJ, and Clark EB. Cardiac morphology and bloodpressure in the adult zebrafish. Anat Rec 264: 1–12, 2001.
12. Irisawa H. Comparative physiology of the cardiac pacemakermechanism. Physiol Rev 58: 461–498, 1978.
H1159VENTRICULAR CONDUCTION SYSTEM IN LOWER VERTEBRATES
AJP-Heart Circ Physiol • VOL 284 • APRIL 2003 • www.ajpheart.org

13. Jalife J, Morley GE, Tallini NY, and Vaidya D. A fungalmetabolite that eliminates motion artifacts. J Cardiovasc Elec-trophysiol 9: 1358–1362, 1998.
14. Jouk PS, Usson Y, Michalowicz G, and Grossi L. Three-dimensional cartography of the pattern of the myofibres in thesecond trimester fetal human heart. Anat Embryol (Berl) 202:103–118, 2000.
15. Kamino K. Optical approaches to ontogeny of electrical activityand related functional organization during early heart develop-ment. Physiol Rev 71: 53–91, 1991.
16. Keith A and Flack M. The form and nature of the musclarconnections between the primary divisions of the vertebrateheart. J Anat Physiol 41: 172–189, 1907.
17. Knisley SB, Hill BC, and Ideker RE. Virtual electrode effectsin myocardial fibers. Biophys J 66: 719–728, 1994.
18. Kolker SJ, Tajchman U, and Weeks DL. Confocal imaging ofearly heart development in Xenopus laevis. Dev Biol 218: 64–73,2000.
19. Kucera JP, Kleber AG, and Rohr S. Slow conduction incardiac tissue. II. Effects of branching tissue geometry. Circ Res83: 795–805, 1998.
20. Litchenberg WH, Norman LW, Holwell AK, Martin KL,Hewett KW, and Gourdie RG. The rate and anisotropy ofimpulse propagation in the postnatal terminal crest are corre-lated with remodeling of Cx43 gap junction pattern. CardiovascRes 45: 379–387, 2000.
21. Mohsen T, Anthonioz P, and Jadoun G. [Presence in theheart of Protopterus annectens (Fishes, Dipneuste) of nodal andconducting cells histologically distinct from common myocardialcells.] C R Seances Soc Biol Fil 170: 712–715, 1976.
22. Morley GE and Vaidya D. Understanding conduction of elec-trical impulses in the mouse heart using high-resolution videoimaging technology. Microsc Res Tech 52: 241–250, 2001.
23. Nair MG. The development of the nervous system in the heart ofthe Chinese carp, Cyprinus carpio (Linnaeus), with a specialreference to its conducting system. Mikroscopie 29: 1–7, 1973.
24. Nygren A, Clark RB, Belke DD, Kondo C, Giles WR, andWitkowski FX. Voltage-sensitive dye mapping of activation and
conduction in adult mouse hearts. Ann Biomed Eng 28: 958–967,2000.
25. Re A. Intervention of sulfurated waters in the onset and con-duction of excitation in the isolated frog heart. Boll Soc Ital BiolSper 40: 705–709, 1964.
26. Rentschler S, Vaidya DM, Tamaddon H, Degenhardt K,Sassoon D, Morley GE, Jalife J, and Fishman GI. Visual-ization and functional characterization of the developing murinecardiac conduction system. Development 128: 1785–1792, 2001.
27. Robb JS. Comparative Basic Cardiology. New York: Grune &Stratton, 1965.
28. Sanchez-Quintana D and Hurle JM. Ventricular myocardialarchitecture in marine fishes. Anat Rec 217: 263–273, 1987.
29. Sedmera D, Pexieder T, Vuillemin M, Thompson RP, andAnderson RH. Developmental patterning of the myocardium.Anat Rec 258: 319–337, 2000.
30. Stainier DY. Zebrafish genetics and vertebrate heart formation.Nat Rev Genet 2: 39–48, 2001.
31. Tamaddon HS, Vaidya D, Simon AM, Paul DL, Jalife J,and Morley GE. High-resolution optical mapping of the rightbundle branch in connexin40 knockout mice reveals slow con-duction in the specialized conduction system. Circ Res 87: 929–936, 2000.
32. Van Mierop LHS and Kutsche LM. comparative anatomy andembryology of the ventricles and arterial pole of the vertebrateheart. In: Congenital Heart Disease: Causes and Processes, ed-ited by Nora JJTA. New York: Futura, 1984, p. 459–479.
33. Vassall-Adams PR. The development of the atrioventricularbundle and its branches in the avian heart. J Anat 134: 169–183,1982.
34. Viragh S and Challice CE. The development of the conductionsystem in the mouse embryo heart. I. The first embryonic A-Vconduction pathway. Dev Biol 56: 382–396, 1977.
35. Warren KS and Fishman MC. “Physiological genomics”: mu-tant screens in zebrafish. Am J Physiol Heart Circ Physiol 275:H1–H7, 1998.
36. Westerfield M. The Zebrafish Book: a Guide for the LaboratoryUse of Zebrafish (Danio rerio). Eugene, OR: Univ. of OregonPress, 1995.
H1160 VENTRICULAR CONDUCTION SYSTEM IN LOWER VERTEBRATES
AJP-Heart Circ Physiol • VOL 284 • APRIL 2003 • www.ajpheart.org




















