
Characterization and Functional Validation of Tobacco PLC Delta for Abiotic Stress Tolerance
Upload
independentCategory
view
0download
0
Functional Analysis of Arabidopsis Ethylene-ResponsiveElement Binding Protein Conferring Resistance to Baxand Abiotic Stress-Induced Plant Cell Death1
Taro Ogawa, Ling Pan, Maki Kawai-Yamada, Li-Hua Yu, Saburo Yamamura, Tomotsugu Koyama,Sakihito Kitajima2, Masaru Ohme-Takagi, Fumihiko Sato, and Hirofumi Uchimiya*
Institute of Molecular and Cellular Biosciences, The University of Tokyo, Tokyo 113–0032, Japan (T.O., L.P.,M.K.-Y., L.-H.Y., H.U.); Iwate Biotechnology Research Center, Kitakami, Iwate 024–0003, Japan (S.Y., H.U.);Gene Function Research Laboratory, National Institute of Advanced Industrial Science and Technology,Tsukuba 305–8562, Japan (T.K., M.O.-T.); and Department of Applied Life Sciences, Kyoto University,Kitashirakawa, Sakyo, Kyoto 606–8502, Japan (S.K., F.S.)
Arabidopsis (Arabidopsis thaliana) ethylene-responsive element binding protein (AtEBP) gene was isolated as a suppressor ofBax-induced cell death by functional screening in yeast (Saccharomyces cerevisiae). To further examine the cell death suppressiveaction of AtEBP in plant cells, we established transgenic tobacco (Nicotiana tabacum) plants overexpressing AtEBP as well astransgenic tobacco plants ectopically expressing mouse Bax protein under a dexamethasone-inducible promoter. We preparedthe crosses of the selective lines of each transgenic plant, which were evaluated in terms of cell death suppression activity.Results indicate that AtEBP suppressed Bax-induced cell death in tobacco plants, an action also associated with a lowered levelof ion leakage. Furthermore, tobacco Bright Yellow-2 cells overexpressing AtEBP conferred resistance to hydrogen peroxide(H2O2) and heat treatments. AtEBP protein localized in the nucleus and functioned as an in vivo transcription activator asconfirmed in transient assays and experiments using stable transgenic system. Up-regulation of defense genes was observed intransgenic Arabidopsis plants overexpressing AtEBP. Based on the analysis of mRNA accumulation in ethylene-relatedmutants, the position of AtEBP in signaling pathway is presented.
The proapoptotic mammalian protein Bax can in-duce cell death in the budding yeast, Saccharomycescerevisiae (Sato et al., 1994; Greenhalf et al., 1996; Zhaet al., 1996) as well as in mammalian cells (Oltvai et al.,1993). Bax forms channels in the outer membrane ofthe mitochondrion and triggers the release of cyto-chrome c, which activates a series of caspases, initiat-ing a cascade of protease activation. Commonaltiesin the mechanism of Bax-induced cell death werefound in yeast and mammalian cells (for review, seeReed, 1997). Furthermore, Bax-induced cell death inyeast can be suppressed by expression of mammalian(Xu and Reed, 1998), and plant (Kawai et al., 1999;Kampranis et al., 2000; Pan et al., 2001) genes, respec-tively. In our previous work, we attempted to screengenes from an Arabidopsis (Arabidopsis thaliana)cDNA library, which suppressed the lethal effects ofmouse Bax in yeast (Pan et al., 2001). The mostabundant gene (82% of isolated genes) screened wasfound to be AtEBP, known as Arabidopsis ethylene-
responsive element binding protein gene. AtEBP hasone APETALA2/ethylene responsive element bindingprotein (AP2/EREBP) domain, which is known to bindGCC-box (AGCCGCC), an ethylene responsive cis-element (Allen et al., 1998). Buttner and Singh (1997)also isolated AtEBP as an in vitro interacting partnerof OBF4/TGA4, a basic Leu zipper transcription factor.However, the biological significance of AtEBP in plantais not fully understood.
The AP2/EREBP family is a unique group of planttranscriptional factors. The AP2/EREBP domain con-sists of about 60 conserved amino acids (Allen et al.,1998). The APETALA2 family, containing two AP2/EREBP domains, regulates plant development (Jofukuet al., 1994; Elliot et al., 1996; Klucher et al., 1996;Boutilier et al., 2002). On the other hand, the ethylene-responsive factor (ERF) family, containing one AP2/EREBP domain, is one of the largest groups of tran-scriptional factors (Riechmann et al., 2000). Thesegenes can work in various conditions: downstreamof ethylene, jasmonate, and abscisic acid signalingpathways; and environmental-stress responses such ascold, dry, and salt (Ohme-Takagi and Shinshi, 1995;Stockinger et al., 1997; Finkelstein et al., 1998; Liu et al.,1998; Solano et al., 1998; Menke et al., 1999; van der Fitsand Memelink, 2000, 2001; Gu et al., 2002). It is knownthat two types of transcriptional factors, namely a tran-scriptional activator and a repressor, are included inERF family proteins (Fujimoto et al., 2000; Ohta et al.,
1 This work was supported by Research for the Future from theJapan Society for the Promotion of Science.
2 Present address: Department of Applied Biology, Kyoto Instituteof Technology, Matsugasaki Sakyo-ku, Kyoto 606–8585, Japan.
* Corresponding author; e-mail [email protected]; fax81–3–5841–8466.
Article, publication date, and citation information can be found atwww.plantphysiol.org/cgi/doi/10.1104/pp.105.063586.
1436 Plant Physiology, July 2005, Vol. 138, pp. 1436–1445, www.plantphysiol.org � 2005 American Society of Plant Biologists www.plant.org on June 4, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2005 American Society of Plant Biologists. All rights reserved.

2000, 2001). Furthermore, several ERF proteins arereported to interact with other proteins such as tran-scriptional factor, nitrilase-like protein, and ubiquitin-conjugated enzyme (Buttner and Singh, 1997; Xu et al.,1998; Koyama et al., 2003). These facts suggest that thephysiological role of each ERF may be divergent.
In this study, we demonstrate that plant cellsoverexpressing AtEBP are resistant to Bax-induced celldeath and abiotic stresses such as hydrogen peroxide(H2O2) and heat. Furthermore, AtEBP functions asa transcriptional activator as demonstrated in tran-sient assays and experiments using stable transgenicsystems. Based on the analysis of gene expressionlevels in ethylene-related mutants, we also present theposition of AtEBP in the ethylene signaling pathway.
RESULTS
AtEBP Suppresses Bax-Induced Cell Death inTobacco Plants
AtEBP gene was previously isolated as a suppressorof Bax-induced cell death by functional screening inyeast (Pan et al., 2001). To confirm that AtEBP sup-presses cell death in plant cells, we established trans-genic tobacco (Nicotiana tabacum) plants expressingAtEBP and ectopically expressing Bax using a dexa-methasone (DEX)-inducible system (Aoyama and Chua,1997; Fig. 1A). We generated five independent trans-genic tobacco plants expressing AtEBP under cauli-flower mosaic virus (CaMV) 35S promoter, followedby northern-blot analysis using AtEBP cDNA as aprobe (data not shown). We selected two lines (EBP8and EBP20) for the following experiments. We alsogenerated about 20 independent transgenic tobaccoplants ectopically expressing Bax. When transgenicBax plants (2-week-old) were transferred to a mediumcontaining 10 mM DEX, extensive etiolation at thewhole-plant level was observed at 4 d after DEXtreatment (Fig. 1B). One of these lines, namely Bax21,showed Bax expression immunologically at 6 h afterDEX treatment (Fig. 1C). We also selected this line forthe following cross experiment.
To evaluate whether AtEBP suppresses Bax-inducedcell death in plants, we prepared a hybrid line bycrossing these transgenic tobacco plants expressingAtEBP (EBP20) with plants ectopically expressingBax (Bax21). Only F1 hybrid plants were used in thefollowing experiments. Constitutive expression ofAtEBP and DEX-induced expression of Bax in the hy-brid line were confirmed by reverse transcription (RT)-PCR (Fig. 1D, a). Furthermore, the expression of Baxprotein in the hybrid line was also detected immuno-logically (Fig. 1D, b). Although an apparent chlorosis ofleaves in the parent line was seen at 4 d after DEXtreatment, leaves of the hybrid line retained green color(Fig. 1E). To quantitatively evaluate cell death, ionleakage from leaf discs was measured. Ion leakage hasbeen used as an indicator of plant cell death (Mitsuhara
et al., 1999; Rizhsky et al., 2002; Kawai-Yamada et al.,2004). The level of electrolyte leakage of each transgenicline was increased after DEX treatment, but the hybridline showed lower levels of ion leakage compared withthe parent line (Fig. 1F).
We already reported that production of reactiveoxygen species (ROS) production was involved in celldeath triggered by overexpression of Bax in Arabi-dopsis plants (Kawai-Yamada et al., 2004). To testwhether or not AtEBP expression affects ROS pro-duction by Bax, these transgenic tobacco lines (Bax21and Bax21 3 EBP20) and nontransgenic tobacco (SR1)were treated with 1 mM DEX for 7 h and the leaveswere stained with nitroblue tetrazolium (NBT) and3, 3#-diaminobenzidine (DAB) for the detection ofsuperoxide (O2
2) and H2O2, respectively. As shownin Figure 1G, subcellular localizations of O2
2 and H2O2were observed in tobacco plant cells expressing Baxdespite AtEBP expression. NBT and DAB failed tostain nontransgenic tobacco plant cells.
Tobacco Bright Yellow-2 Cells Overexpressing AtEBPConfer Resistance to H2O2 and Heat Stresses
We established tobacco Bright Yellow-2 (BY-2)cells overexpressing AtEBP by the Agrobacterium-mediated method (Shaul et al., 1996). AtEBP expressionwas detected only in the cell lines transformed witha vector possessing AtEBP (Fig. 2A).
Using such cell lines, we evaluated H2O2-inducedcell death. Ten-day-old cells were transferred to a freshmedium and then treated with H2O2 (0, 2, and 5 mM)for 4 h. The extent of cell death was determined byselective staining of dead cells with Evans blue bymeasurement of A600 (Fig. 2B). Compared with thecontrol lines (C1 and C2), repression of cell death inthe transgenic lines (T1 and T2) was apparent at 2 and5 mM H2O2.
A brief heat treatment of suspension-cultured cellshas also previously been demonstrated to induce celldeath evident by apoptotic phenotype such as cellshrinkage and DNA laddering (McCabe et al., 1997;Swidzinski et al., 2002). We also investigated theresistance to heat stress using the same transgeniclines. Namely, 10-d-old cells were incubated at 55�Cfor 2 h and kept for 12 h at 27�C and shrunken cellswere counted under a microscope (Fig. 2C). Thecontrol lines showed 50% to 80% of shrunken cells,whereas the transgenic lines showed only 30% to 40%of shrunken cells.
Localization of Green Fluorescence Protein-AtEBPFusion Protein Localizes in the Nucleus, andAtEBP Acts as a Transcriptional Activator
A putative nuclear localization signal was found infront of the AP2/EREBP domain of AtEBP protein. Wepreviously reported that the nuclear localization ofAtEBP was essential for the cell death suppressionactivity in yeast cells (Pan et al., 2001). To clarify the
Arabidopsis Ethylene-Responsive Element Binding Protein
Plant Physiol. Vol. 138, 2005 1437 www.plant.org on June 4, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2005 American Society of Plant Biologists. All rights reserved.

Figure 1. Suppression of Bax-induced cell death in tobacco plants overexpressing AtEBP. A, Schematic diagrams of the DEX-inducible vector construct carrying Bax (pTA-Bax) and the constitutive expression vector construct carrying AtEBP (pBIG-AtEBP).63UAS, Glucocoriticoid-regulated transcription factor-regulated promoter; Bax, mouse Bax-coding sequence; 3A, pea rbc-3Apoly A addition sequence; E9, pea rbc-E9 poly A addition sequence; GVG, chimeric glucocoriticoid-regulated transcriptionalfactor; 35S, CaMV 35S promoter; V, tobacco mosaic virus transcriptional enhancer sequence; AtEBP, AtEBP coding sequence;NOS polyA, nopaline synthetase polyadenylation sequence. B, Bax-induced chlorosis in tobacco plant. Two-week-old tobaccoplant was treated in a medium containing 10 mM DEX for 4 d. Bar5 1 cm. C, Western-blot analysis of Bax protein in transgenictobacco plants. Two-week-old tobacco plants (Bax21 line) were treated in a medium containing 10 mM DEX for 1 d. Totalproteins (20 mg) obtained from shoots of the plants at each time period (0, 6, 12, and 24 h) were used. D, a, RT-PCR analysis ofAtEBP and Bax in the hybrid line. Two-week-old plants (Bax21 and Bax213EBP20) were treated in a medium containing 1 mM
DEX for 24 h. Total RNAs from shoots of plants and a pair of specific oligonucleotide primers of AtEBP and Bax were used inRT-PCR analysis. b, Western-blot analysis of Bax proteins in the hybrid line. Two-week-old tobacco plants (Bax21 and Bax213EBP20) were treated in a medium containing 10 mM DEX for 2 d. Total proteins (20 mg) obtained from shoots of the plants at eachtime period (0, 24, and 48 h) were used. E, Repressed Bax-induced chlorosis in the hybrid line expressing AtEBP. Two-week-oldtobacco plants (Bax21 and Bax21 3 EBP20) were treated in a medium containing 10 mM DEX for 4 d. Bar 5 1 cm. F, Bax-induced ion leakage in the hybrid line. Leaf sections obtained from 2-week-old tobacco plants (Bax21 and Bax21 3 EBP20)were incubated with 0, 0.1, and 1.0 mM DEX in distilled water at 27�C. The resulting values are indicated as relatives. Datarepresent mean6 SE obtained from four independent measurements. G, Cytological observation of O2
2 and H2O2 production intobacco lines. Two-week-old tobacco plants (Bax21, Bax21 3 EBP20, and nontransgenic plant, SR1) were treated with 1 mM
DEX for 7 h to induce Bax, and the detached leaves were stained by NBT (top) and DAB (bottom). Bar5 50 mm. The experimentswere repeated at least three times, and representative photographs are shown.
1438 Plant Physiol. Vol. 138, 2005 www.plant.org on June 4, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2005 American Society of Plant Biologists. All rights reserved.
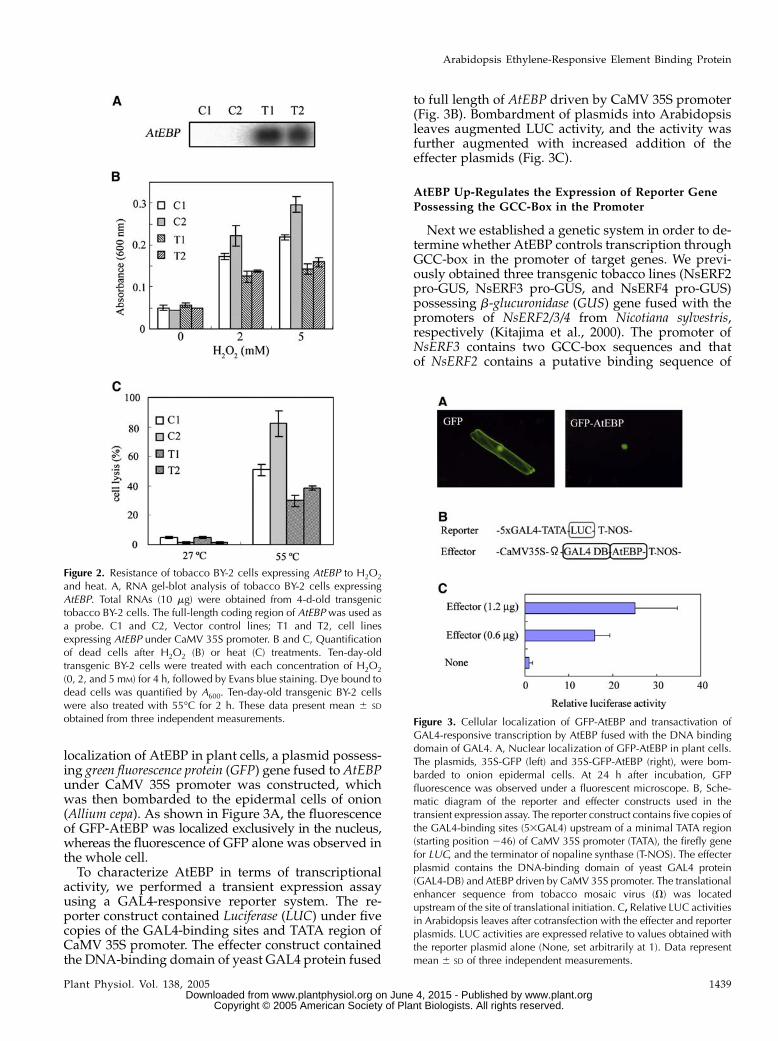
localization of AtEBP in plant cells, a plasmid possess-ing green fluorescence protein (GFP) gene fused to AtEBPunder CaMV 35S promoter was constructed, whichwas then bombarded to the epidermal cells of onion(Allium cepa). As shown in Figure 3A, the fluorescenceof GFP-AtEBP was localized exclusively in the nucleus,whereas the fluorescence of GFP alone was observed inthe whole cell.
To characterize AtEBP in terms of transcriptionalactivity, we performed a transient expression assayusing a GAL4-responsive reporter system. The re-porter construct contained Luciferase (LUC) under fivecopies of the GAL4-binding sites and TATA region ofCaMV 35S promoter. The effecter construct containedthe DNA-binding domain of yeast GAL4 protein fused
to full length of AtEBP driven by CaMV 35S promoter(Fig. 3B). Bombardment of plasmids into Arabidopsisleaves augmented LUC activity, and the activity wasfurther augmented with increased addition of theeffecter plasmids (Fig. 3C).
AtEBP Up-Regulates the Expression of Reporter Gene
Possessing the GCC-Box in the Promoter
Next we established a genetic system in order to de-termine whether AtEBP controls transcription throughGCC-box in the promoter of target genes. We previ-ously obtained three transgenic tobacco lines (NsERF2pro-GUS, NsERF3 pro-GUS, and NsERF4 pro-GUS)possessing b-glucuronidase (GUS) gene fused with thepromoters of NsERF2/3/4 from Nicotiana sylvestris,respectively (Kitajima et al., 2000). The promoter ofNsERF3 contains two GCC-box sequences and thatof NsERF2 contains a putative binding sequence of
Figure 2. Resistance of tobacco BY-2 cells expressing AtEBP to H2O2
and heat. A, RNA gel-blot analysis of tobacco BY-2 cells expressingAtEBP. Total RNAs (10 mg) were obtained from 4-d-old transgenictobacco BY-2 cells. The full-length coding region of AtEBP was used asa probe. C1 and C2, Vector control lines; T1 and T2, cell linesexpressing AtEBP under CaMV 35S promoter. B and C, Quantificationof dead cells after H2O2 (B) or heat (C) treatments. Ten-day-oldtransgenic BY-2 cells were treated with each concentration of H2O2
(0, 2, and 5 mM) for 4 h, followed by Evans blue staining. Dye bound todead cells was quantified by A600. Ten-day-old transgenic BY-2 cellswere also treated with 55�C for 2 h. These data present mean 6 SD
obtained from three independent measurements. Figure 3. Cellular localization of GFP-AtEBP and transactivation ofGAL4-responsive transcription by AtEBP fused with the DNA bindingdomain of GAL4. A, Nuclear localization of GFP-AtEBP in plant cells.The plasmids, 35S-GFP (left) and 35S-GFP-AtEBP (right), were bom-barded to onion epidermal cells. At 24 h after incubation, GFPfluorescence was observed under a fluorescent microscope. B, Sche-matic diagram of the reporter and effecter constructs used in thetransient expression assay. The reporter construct contains five copies ofthe GAL4-binding sites (53GAL4) upstream of a minimal TATA region(starting position 246) of CaMV 35S promoter (TATA), the firefly genefor LUC, and the terminator of nopaline synthase (T-NOS). The effecterplasmid contains the DNA-binding domain of yeast GAL4 protein(GAL4-DB) and AtEBP driven by CaMV 35S promoter. The translationalenhancer sequence from tobacco mosaic virus (V) was locatedupstream of the site of translational initiation. C, Relative LUC activitiesin Arabidopsis leaves after cotransfection with the effecter and reporterplasmids. LUC activities are expressed relative to values obtained withthe reporter plasmid alone (None, set arbitrarily at 1). Data representmean 6 SD of three independent measurements.
Arabidopsis Ethylene-Responsive Element Binding Protein
Plant Physiol. Vol. 138, 2005 1439 www.plant.org on June 4, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2005 American Society of Plant Biologists. All rights reserved.

EIN3/TEIL (Kosugi and Ohashi, 2000), but that ofNsERF4 does not contain any known binding site re-lated to the ethylene signaling (Fig. 4A). We also gen-erated transgenic tobacco plants expressing AtEBPunder CaMV 35S promoter.
To obtain hybrid plants, transgenic tobacco lineexpressing AtEBP (EBP8) and NsERF2/3/4 pro-GUS
lines were crossed to each other. AtEBP expression inthe hybrid plants (F1) was confirmed by RT-PCR (Fig.4B). Accordingly, we measured GUS activities in thesehybrid lines. The results indicated that the hybrid line(NsERF3 pro-GUS 3 EBP8) containing the GCC-boxin the promoter and expressing AtEBP, showed en-hanced GUS activity compared with the control line
Figure 4. Up-regulation of the reporter genepossessing GCC-box in transgenic tobacco plantsexpressing AtEBP. A, Schematic diagram of thestructures of NsERF2/3/4 pro-GUS. The promoterregions of NsERF2 (NsERF2 pro), NsERF3(NsERF3 pro), and NsERF4 (NsERF4 pro) werefused with GUS gene and the terminator ofnopaline synthase (T-NOS). Translational initia-tion is indicated as 11. B, RT-PCR analysis ofAtEBP in the hybrid lines. AtEBP expression inshoots of 2-week-old hybrid lines was confirmedby RT-PCR using a pair of specific oligonucleo-tide primers of AtEBP. SR1, Nontransgenic to-bacco plants; EBP8, a transgenic tobacco lineexpressingAtEBP; NsERF2/4 pro-GUS, transgenictobacco lines expressing GUS under NsERF2/4promoter lacking GCC-box; NsERF3 pro-GUS,a transgenic tobacco line expressing GUS underNsERF3 promoter containing GCC-box (Kitajimaet al., 2000). C, Analysis of GUS activity in eachhybrid line. GUS activities were measured usingshoots of 2-week-old tobacco plants. Data rep-resent mean 6 SD obtained from at least fourindependent measurements. Numerals corre-spond to Figure 2B. Plus sign (1) and minussign (2), presence or absence of AtEBP expres-sion or GCC-box, respectively.
Ogawa et al.
1440 Plant Physiol. Vol. 138, 2005 www.plant.org on June 4, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2005 American Society of Plant Biologists. All rights reserved.

(NsERF3 pro-GUS3 SR1; Fig. 4C). In contrast, despiteAtEBP expression, the other hybrid lines (NsERF2/4pro-GUS 3 EBP8) lacking GCC-box in the promoterdid not show any change of GUS activity comparedwith the control lines (NsERF2/4 pro-GUS3SR1). Theseresults suggest that AtEBP functions as a transcrip-tional activator through the GCC-box in plant cells.
AtEBP Overexpression Causes the Up-Regulation ofDefense Genes
In order to identify the genes that are regulated byAtEBP in plant cells, several transgenic Arabidopsislines overexpressingAtEBPunder CaMV 35S promoterwere established. Using such lines, we investigated theexpression of defense genes including pathogenesis-related (PR) genes. As shown in Figure 5, expressions ofPDF1.2, a plant defensin gene (Penninckx et al., 1996),and GST6 (Chen et al., 1996), which is related to ROSdetoxification, were increased in Arabidopsis linesoverexpressing AtEBP. On the other hand, the expres-sions of PR-1/2/3 in Arabidopsis lines overexpressingAtEBPwere not different from the control line. The datawere obtained as replicates with the same results.
The Position of AtEBP in the EthyleneSignaling Pathway
Many isolated ethylene-related mutants have beenanalyzed for their epistasis relationships, and theframework of the ethylene signaling pathway is al-most established (for review, see Ecker, 1995; Wanget al., 2002). Nevertheless, the precise positions of over100 ERF genes, including AtEBP in the ethylene sig-naling pathway, have not been identified, except for
ERF1 (Solano et al., 1998). To clarify the position ofAtEBP in the ethylene signaling pathway, we analyzedmRNA accumulation in ethylene-related mutants suchas ethylene-insensitive mutants (ethylene resistant 1-1[etr1-1], ethylene insensitive 2-1 [ein2-1], and ethyleneinsensitive 3-1 [ein3-1]), a constitutive active mutant inthe ethylene signal transduction (constitutive tripleresponse 1-1 [ctr1-1]), and an ethylene overproducingmutant (ethylene overproducer 3-1 [eto3-1]). As shownin Figure 6, the level of AtEBP mRNA was lower inein2-1 but higher in ctr1-1 and equal in etr1-1, ein3-1,and eto3-1 than in wild type. ERF1 mRNA wasdetected only in ctr1-1. Same results of the data wereobtained in the separate experiments. Thus, AtEBPmay be located downstream of EIN2 and CTR1, butnot under EIN3.
DISCUSSION
AtEBP Confers Resistance to Cell Death
We demonstrated previously that AtEBP sup-pressed Bax-induced cell death in yeast, where nucleartransport of AtEBP was essential (Pan et al., 2001).However, we were unable to identify the precisemechanism associated with this phenomenon. Thus,this study was designed to determine whether or notAtEBP confers the suppressive ability of Bax-inducedplant cell death. The research showed that overexpres-sion of AtEBP confers resistance to not only Bax-induced cell death at the whole-plant level but alsoH2O2- and heat-induced cell death in plant cells.
Both ROS generation and the release of cytochromec from the mitochondria cause the activation of celldeath in mammalian cells (Green and Reed, 1998).ROS production by ecotopic expression of Bax occursin plant cells (Kawai-Yamada et al., 2004, and thisstudy). Following Bax-induced ROS production, celldeath at the whole-plant level with cell shrinkage,membrane destruction, and other apoptotic pheno-type has been observed (Kawai-Yamada et al., 2001,2004). Such phenotypes of plant cells were also ob-served by exogenous following treatments of H2O2and heat (Levine et al., 1996; McCabe and Leaver,2000; Houot et al., 2001; Swidzinski et al., 2002). Thus,both Bax expression and exogenous treatments (H2O2and heat) induce a similar cell death phenomenon.
The scheme of Bax-induced cell death is consistentwith our previous reports (Kawai-Yamada et al., 2001,2004). In tobacco plants, Bax-induced ROS genera-tion was observed at 7 h after DEX treatment, loss ofmembrane permeability shown as ion leakage wasobserved up to 24 h after DEX treatment, and anapparent chlorosis of leaves was observed at 4 d afterDEX treatment. In Arabidopsis, ROS generation, ionleakage, and chlorosis by Bax were also observed insimilar time scale (Kawai-Yamada et al., 2004). Thesedata indicate that the lethal effect of Bax protein iscommon in different plant species.
Figure 5. Up-regulation of defense genes in Arabidopsis lines over-expressing AtEBP. Total RNAs (10 mg) obtained from shoots of 35-d-oldplants were loaded. The coding region of AtEBP excepting AP2/EREBPdomain, and the 3#-UTR of PR-1, PR-2, PR-3, PDF1.2, and GST6 wereused as probes. A gel stained with ethidium bromide (which indicatesrRNA) is shown as a control.
Arabidopsis Ethylene-Responsive Element Binding Protein
Plant Physiol. Vol. 138, 2005 1441 www.plant.org on June 4, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2005 American Society of Plant Biologists. All rights reserved.

AtEBP expression did not influence the process ofO2
2 and H2O2 production caused by Bax, but thephenomena of ion leakage and apparent chlorosis ofleaves were late or repressed compared to the controlline. We assume that the suppressive activity of AtEBPon ROS-induced cell death is due to the regulation ofROS-related genes (Fig. 7). Consistent with this conclu-sion, overexpression of AtEBP resulted in up-regulationof GST6, which is related to ROS metabolism. It maybe interesting to point out that other plant GSThomologs were also isolated as functional suppressorsof Bax-induced cell death in yeast (Kampranis et al.,2000; Pan et al., 2001).
AtEBP Is an in Vivo Transcriptional Activator
It was demonstrated that AtEBP can bind GCC-boxin vitro (Buttner and Singh, 1997). To clear the functionof AtEBP as a transcriptional factor, we performedseries of the following experiments. Microscopic ob-servation of GFP-AtEBP fusion protein confirmedthat AtEBP localizes in the nucleus. The transient ex-pression assay by bombardment also suggested thatAtEBP is a transcriptional activator. Furthermore, ge-netic experiments using transgenic tobacco plantsexpressing AtEBP and constructs possessing respec-tive reporter genes suggested that AtEBP up-regulatesthe reporter gene possessing the GCC-box in thepromoter.
Overexpression of AtEBP coordinately accumulatedPDF1.2 and GST6 mRNAs. PDF1.2 is a typical down-stream gene of the ethylene/jasmonic acid signaling(Penninckx et al., 1996, 1998). The GCC-box in thepromoter of PDF1.2 is known to be important fortranscriptional regulation by methyl jasmonic acid(Brown et al., 2003). Indeed, overexpression of ERFgenes such as Arabidopsis ERF1 and AtERF2, andtomato (Lycopersicon esculentum) Pti4 up-regulatedPDF1.2 expression (Gu et al., 2002; Brown et al.,2003; Lorenzo et al., 2003). Thus, it is likely that AtEBPpromotes transcription of PDF1.2 through in vivobinding to the GCC-box. However, PDF1.2 expressionwas not always increased according to a level of AtEBPmRNA. An appropriate dosage of AtEBP expressionmay stimulate PDF1.2 expression, whereas excessexpression may inhibit this expression by unknown
mechanism. On the other hand, GST6 expression levelwas dependent on the level of AtEBP expression. GST6is one of H2O2- and salicylic acid-inducible genes(Chen et al., 1996; Chen and Singh, 1999). The GST6promoter 2.0 kb upstream from ATG site did notpossess GCC-box. However, the cis-element of TGA4,a basic Leu zipper transcriptional factor known tointeract with AtEBP (Buttner and Singh, 1997), wasfound in the promoter. AtEBP and TGA4 may regulatethe expression of several genes cooperatively. Someconcerns regarding equivocal gene expression lead usto conclude that we cannot exclude the possibility ofphysiological and environmental status of plant ma-terials, which may affect to RNA-blot analysis. Furtherexploitation of such problems not only by transcriptanalysis but also by protein analysis needs to bepursued.
Expression Pattern of AtEBP
Various expression patterns were reported in ERFgenes, although these genes are known to be regulatedby the same hormone, ethylene. For instance, Solanoet al. (1998) found rapid induction of ERF1 in response
Figure 6. Comparison of AtEBP mRNA accumulation in ethylene-relatedmutants. Total RNAs (10 mg) from shoots of 30-d-old plants wereloaded. The coding region of AtEBP excepting AP2/EREBP domain andthe 3#-UTR of ERF1 were used as probes. A gel stained with ethidiumbromide (which indicates rRNA) is shown as a control.
Figure 7. Proposed model for the position and role of AtEBP in theethylene signaling pathway. This study suggests that AtEBP does notseem to interact with EIN3 and that AtEBP regulates the expressionlevels of several plant defense genes such as PDF1.2 and GST6 asindicated by white arrow.
Ogawa et al.
1442 Plant Physiol. Vol. 138, 2005 www.plant.org on June 4, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2005 American Society of Plant Biologists. All rights reserved.

to ethylene gas within 15 min. ERF1 was constitutivelyexpressed in ctr1, and ethylene induction of ERF1 wastotally dependent on a functional EIN3 protein (Fig. 7).In the case of AtERF genes, Fujimoto et al. (2000) alsodemonstrated late induction of AtERF1, AtERF2, andAtERF5 at 12 h after ethylene treatment.
Analysis of AtEBP expression in ethylene-relatedmutants indicated that AtEBP is located downstreamof CTR1 and EIN2, but not ETR1 and EIN3 (Fig. 7).Since four ETR1 homologs and five EIN3 homologs arefound in Arabidopsis genome (Riechmann et al., 2000;Wang et al., 2002), AtEBP expression may be directedby these genes. The accumulation of AtEBP mRNA ineto3-1 was almost equaled to that in wild type, becauseeto3-1 mutant was overproducing only at the seedlingstage (Woeste et al., 1999). One-month-old matureplants were used in that experiment, and the amountof ethylene may not be overproduced in eto3-1.
ERF1, a key factor of the ethylene signaling pathway,is known to be located the downstream of EIN3(Fig. 7). Solano et al. (1998) showed that transgenicArabidopsis overexpressing EIN3 expressed ERF1mRNA highly, supporting the position of ERF1 beinga direct target of EIN3. Transgenic Arabidopsis over-expressing ERF1 showed constitutive ethylene re-sponses such as inhibition of hypocotyl elongation,while transgenic plants overexpressing AtEBP didnot show such phenotype (data not shown). Thus,AtEBP and ERF1 may be distinctly separate from eachother.
In conclusion, we demonstrated in this study thatAtEBP conferred the resistance to Bax-induced celldeath in tobacco plants and the resistance to exoge-nous treatments such as H2O2 and heat in plant cells.Further studies are planned to dissect the molecularregulation controlled by AtEBP and to clear the linkbetween the physiological and developmental pheno-type.
MATERIALS AND METHODS
Plant Materials
The Columbia ecotype of Arabidopsis (Arabidopsis thaliana), and tobacco
(Nicotiana tabacum) cv Petit Havana SR1 and cv Samsun NN were used. All
plants were cultivated in growth chambers at 23�C (Arabidopsis) and 27�C(tobacco) under continuous light. The ethylene-related mutants, etr1-1, ein2-1,
ein3-1, and eto3-1, were obtained from the Arabidopsis Biological Resource
Center (Columbus, OH).
Suspension-cultured cells of tobacco BY-2 were cultured weekly in liquid
Murashige and Skoog (1962) medium supplemented with Suc (30 g/L),
KH2PO4 (200 mg/L), thiamine hydrochloride (1 mg/L), and 2,4-dichlorophe-
noxyacetic acid (0.2 mg/L) as described previously (Nagata et al., 1992). To
induce cell death by H2O2, 10-d-old tobacco BY-2 cells were treated with H2O2
(0, 2, and 5 mM) and cultured under continuous shaking at 27�C for 4 h. The
viability of BY-2 cells was measured by the addition of 0.05% Evans blue
(Nakalai, Kyoto) for 15 min, followed by extensive wash with distilled water
to remove unbound dye. Dye bound to dead cells was solubilized with 50%
methanol and 1% SDS for 30 min at 50�C, and quantified by A600. To induce cell
death by heat treatment, 10-d-old tobacco BY-2 cells were treated with 55�C for
2 h and returned to 27�C for 12 h. The percentage of shrunken cells was
calculated by observations of the cells under a microscope with cytoplasmic
shrinkage.
Transformation of Arabidopsis, Tobacco Plants, and
Tobacco BY-2 Cells
The open reading frame of AtEBP was inserted into plasmids, pBIN
(CaMV 35S) and pBIG (CaMV 35S-V), and the open reading frame of mouse
Bax was inserted into the DEX-inducible vector pTA7002 as described pre-
viously (Kawai-Yamada et al., 2001). The resulting plasmid, pBIN-AtEBP, was
transformed into Arabidopsis plants (Bechthold et al., 1993) and tobacco BY-2
cells (Shaul et al., 1996) by Agrobacterium-mediated transformation. The
plasmids, pBIG-AtEBP and pTA-Bax, were transformed into tobacco cv Petite
Havana SR1 as described by Gallois and Marinho (1995). Transgenic tobacco
(cv Samsun NN) plants, NsERF2 pro-GUS, NsERF3 pro-GUS, and NsERF4
pro-GUS (Kitajima et al., 2000) were also used.
Northern-Blot Analysis
Plant tissues were homogenized with liquid nitrogen in the extrac-
tion buffer (200 mM Tris-HCl, pH 8.0, 10 mM EDTA, 100 mM NaCl, 0.1%
SDS, and 0.1% mercapthoethanol). Total RNAs (10 mg) were fractionated on
1.2% agarose gel containing 5% formaldehyde, and transferred to a nylon
membrane (Biodyne B, Pall, Washington, NY). The 32P-labeled probes, the
3#-untranslated region (UTR) were used for PR-1 (Metzler et al., 1991), PR-2
(Uknes et al., 1992), PR-3 (Samac et al., 1990), PDF1.2 (Penninckx et al., 1996),
and GST6 (Chen et al., 1996). The full-length coding region was used for
AtEBP in the experiments using BY-2 cells, shown in Figure 2A, and tobacco
plants, and the coding region of AtEBP (Buttner and Singh, 1997), except for
AP2/EREBP domain, was used for the other experiments using Arabidopsis
plants.
Hybridization was performed in 10% dextran sulfate solution containing
1 M NaCl, 1% SDS, and 10 mg/mL heat-denatured salmon sperm DNA at
65�C for overnight. Washing was performed with 23 SSC for 10 min, with 13
SSC with 0.1% SDS at 65�C for 30 min, and with 0.13 SSC with 0.1% SDS at
65�C for 30 min. The membranes were analyzed by a BAS1500 imaging plate
scanner (Fuji Photo Film, Tokyo).
Western-Blot Analysis
Plant tissues were homogenized in the extraction buffer (50 mM HEPES,
pH 8.0, 2 mM EDTA, 330 mM sorbitol, and 0.8% mercaptoethanol). Total
proteins (20 mg per lane) separated by SDS-15% PAGE were transferred onto
a polyvinylidene fluoride membrane (Milipore, Belford, MA). After blocking
with phosphate-buffered saline buffer (137 mM NaCl, 8.10 mM Na2HPO4,
2.68 mM KCl, and 1.47 mM KH2PO4) containing 5% skim milk for overnight
at 4�C. The membranes were treated with a polyclonal antibody for Bax
(09–499, Upstate Biotechnology, Lake Placid, NY), followed by treatment
with horseradish peroxidase-conjugated anti-rabbit IgG (1:2,000 dilution;
Amersham Pharmacia Biotech, Piscataway, NJ). Detection was accomplished
by an enhanced chemiluminescence kit (Amersham Pharmacia) with expo-
sure to x-ray film (Fuji Photo Film).
Analysis of Ion Leakage
Three leaf discs from 2-week-old tobacco plants were floated on 2 mL
water with different concentrations (0, 0.1, and 1.0 mM) of DEX. Following
incubation at 27�C, the conductivity of the bathing solution was measured
with an electrical conductivity meter (B-173, Hitachi, Tokyo).
Histochemical Detection of O22 and H2O2
Histochemical detection of O22 and H2O2 was performed by treating
leaves with NBT, as described by Rao and Davis (1999), and with DAB, as
described by Thordal-Christensen et al. (1997). At 7 h after treatment with 1 mM
DEX, leaves detached from seedlings were used. For NBT staining, samples
were vacuum infiltrated with 10 mM NaN3 in 10 mM potassium phosphate
buffer, pH 7.8, for 2 min, and incubated in 0.1% NBT in 10 mM potassium
phosphate buffer, pH 7.8, for 30 min at room temperature. For DAB staining,
samples were incubated with 0.1% DAB in 10 mM potassium phosphate
buffer, pH 7.8 for 6 h. These stained samples were cleared by boiling in acetic
acid:glycerol:ethanol (1:3:3, [v/v/v]) solution before photographs were taken.
Arabidopsis Ethylene-Responsive Element Binding Protein
Plant Physiol. Vol. 138, 2005 1443 www.plant.org on June 4, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2005 American Society of Plant Biologists. All rights reserved.

Transient Expression Assay
To express GFP-AtEBP fusion protein in plant cells, AtEBP was inserted
into the GFP cassette vector, pUC18-35S-GFP (Niwa et al., 1999) at the
downstream of GFP in frame. The resulting plasmid (35S-GFP-AtEBP, 0.8 mg)
was transfected into the onion epidermis with the bombardment method
by described by Ohta et al. (2000). After 24 h of incubation, GFP fluorescence
was observed using a fluorescence microscopy with a filter set providing 455 to
490 nm excitation and emission above 515 nm.
In cotransfection assays, the reporter plasmid (1.6 mg) and the effecter
plasmid (1.2 mg) were bombarded into Arabidopsis leaves in each experiment.
The pUC18 was used to adjust the total amount of bombarded DNA. LUC
assay was performed with the Dual-Luciferase Reporter Assay System and
a luminescence reader (TD-20/20; Promega, Madison, WI). To normalize
values after each transfection, 0.4 mg of the plasmid pPTRL, which included
LUC from Renilla under the control of the CaMV 35S promoter, was used as an
internal control (Fujimoto et al., 2000; Ohta et al., 2001). Normalized LUC
activity recorded after transfection with the reporter plasmid alone was set
arbitrarily at 1.
GUS Assay
GUS assay was performed by using the substrate, 4-methylumbelliferyl-b-
D-glucuronide (Jefferson et al., 1987). Briefly, leaves were homogenized in the
extraction buffer (50 mM phosphate buffer, pH 7.0, 10 mM EDTA, 0.1% Triton
X-100, 0.1% sarcosyl, and 10 mM mercapthoethanol). After the crude solution
was incubated with 4-methylumbelliferyl-b-D-glucuronide at 37�C for 30 min,
the reaction was stopped by the addition of Na2CO3 solution. The amount of
4-methylumbelliferone, the resulting product, was measured with a fluores-
cence spectrophotometer (F4500, Hitachi) with excitation at 365 nm and
emission at 455 nm.
Distribution of Materials
Upon request, all novel materials described in this publication will be
made available in a timely manner for noncommercial research purposes,
subject to the requisite permission from any third-party owners of all or parts
of the material. Obtaining any permission will be the responsibility of the
requestor.
ACKNOWLEDGEMENT
We are grateful to Dr. H. Shinshi for providing us with the plasmid
containing the tobacco mosaic virus enhancer.
Received March 30, 2005; revised March 30, 2005; accepted April 13, 2005;
published June 24, 2005.
LITERATURE CITED
Allen MD, Yamasaki K, Ohme-Takagi M, Tateno M, Suzuki M (1998)
A novel mode of DNA recognition by a b-sheet revealed by the solu-
tion structure of the GCC-box binding domain in complex with DNA.
EMBO J 17: 5484–5496
Aoyama T, Chua NH (1997) A glucocorticoid-mediated transcriptional
induction system in transgenic plants. Plant J 11: 605–612
Bechthold N, Ellis J, Pelletier G (1993) In planta Agrobacterium-mediated
gene transfer by infiltration of adult Arabidopsis thaliana plants. C R
Acad Sci Paris Life Sci 316: 1194–1199
Boutilier K, Offringa R, Sharma VK, Kieft H, Ouellet T, Zhang L, Hattori
J, Liu CM, van Lammeren AA, Miki BL, et al (2002) Ectopic expression
of BABY BOOM triggers a conversion from vegetative to embryonic
growth. Plant Cell 14: 1737–1749
Brown RL, Kazan K, McGrath KC, Maclean DJ, Manners JM (2003) A role
for the GCC-box in jasmonate-mediated activation of the PDF1.2 gene of
Arabidopsis. Plant Physiol 132: 1020–1032
Buttner M, Singh KB (1997) Arabidopsis thaliana ethylene-responsive
element binding protein (AtEBP), an ethylene-inducible, GCC box
DNA-binding protein interacts with an ocs element binding protein.
Proc Natl Acad Sci USA 94: 5961–5966
Chen W, Chao G, Singh KB (1996) The promoter of a H2O2-inducible,
Arabidopsis glutathione S-transferase gene contains closely linked OBF-
and OBP1-binding sites. Plant J 10: 955–966
Chen W, Singh KB (1999) The auxin, hydrogen peroxide and salicylic acid
induced expression of the Arabidopsis GST6 promoter is mediated in part
by an ocs element. Plant J 19: 667–677
Ecker JR (1995) The ethylene signal transduction pathway in plants.
Science 268: 667–675
Elliot RC, Betzner AS, Huttner E, Oakes MP, Tucker WQ, Gerentes D,
Perez P, Smyth DR (1996) AINTEGUMENTA, an APETALA2-like gene of
Arabidopsis with pleiotropic roles in ovule development and floral
organ growth. Plant Cell 8: 155–168
Finkelstein RR, Wang ML, Lynch TJ, Rao S, Goodman HM (1998) The
Arabidopsis abscisic acid response locus ABI4 encodes an APETALA2
domain protein. Plant Cell 10: 1043–1054
Fujimoto SY, Ohta M, Usui A, Shinshi H, Ohme-Takagi M (2000)
Arabidopsis ethylene-responsive element binding factors act as tran-
scriptional activators or repressors of GCC box-mediated gene expres-
sion. Plant Cell 12: 393–404
Gallois P, Marinho P (1995) Leaf disk transformation using Agrobacterium
tumefaciens-expression of heterologous genes in tobacco. Methods Mol
Biol 49: 39–48
Green DR, Reed JC (1998) Mitochondria and apoptosis. Science 281:
1309–1312
Greenhalf W, Stephan C, Chaudhuri B (1996) Role of mitochondria and
C-terminal membrane anchor of Bcl-2 in Bax induced growth arrest
and mortality in Saccharomyces cerevisiae. FEBS Lett 380: 169–175
Gu YQ, Wildermuth MC, Chakravarthy S, Loh YT, Yang C, He X, Han Y,
Martin GB (2002) Tomato transcription factors pti4, pti5, and pti6
activate defense responses when expressed in Arabidopsis. Plant Cell
14: 817–831
Houot V, Etienne P, Petitot AS, Barbier S, Blein JP, Suty L (2001)
Hydrogen peroxide induces programmed cell death features in cultured
tobacco BY-2 cells, in a dose-dependent manner. J Exp Bot 52: 1721–1730
Jefferson RA, Kavanagh TA, Bevan MW (1987) GUS fusions: beta-
glucuronidase as a sensitive and versatile gene fusion marker in
higher plants. EMBO J 6: 3901–3907
Jofuku KD, den Boer BG, Van Montagu M, Okamuro JK (1994) Control of
Arabidopsis flower and seed development by the homeotic gene
APETALA2. Plant Cell 6: 1211–1225
Kampranis SC, Damianova R, Atallah M, Toby G, Kondi G, Tsichlis PN,
Makris AM (2000) A novel plant glutathione S-transferase/peroxidase
suppresses Bax lethality in yeast. J Biol Chem 275: 29207–29216
Kawai M, Pan L, Reed JC, Uchimiya H (1999) Evolutionally conserved
plant homologue of the Bax inhibitor-1 (BI-1) gene capable of suppress-
ing Bax-induced cell death in yeast. FEBS Lett 464: 143–147
Kawai-Yamada M, Jin L, Yoshinaga K, Hirata A, Uchimiya H (2001)
Mammalian Bax-induced plant cell death can be down-regulated by
overexpression of Arabidopsis Bax Inhibitor-1 (AtBI-1). Proc Natl Acad Sci
USA 98: 12295–12300
Kawai-Yamada M, Ohori Y, Uchimiya H (2004) Dissection of Arabidopsis
Bax inhibitor-1 suppressing Bax-, hydrogen peroxide-, and salicylic acid-
induced cell death. Plant Cell 16: 21–32
Kitajima S, Koyama T, Ohme-Takagi M, Shinshi H, Sato F (2000)
Characterization of gene expression of NsERFs, transcription factors
of basic PR genes from Nicotiana sylvestris. Plant Cell Physiol 41: 817–824
Klucher KM, Chow H, Reiser L, Fischer RL (1996) The AINTEGUMENTA
gene of Arabidopsis required for ovule and female gametophyte
development is related to the floral homeotic gene APETALA2. Plant
Cell 8: 137–153
Kosugi S, Ohashi Y (2000) Cloning and DNA-binding properties of
a tobacco Ethylene-Insensitive3 (EIN3) homolog. Nucleic Acids Res 28:
960–967
Koyama T, Okada T, Kitajima S, Ohme-Takagi M, Shinshi H, Sato F
(2003) Isolation of tobacco ubiquitin-conjugating enzyme cDNA in
a yeast two-hybrid system with tobacco ERF3 as bait and its character-
ization of specific interaction. J Exp Bot 54: 1175–1181
Levine A, Pennell RI, Alvarez ME, Palmer R, Lamb C (1996) Calcium-
mediated apoptosis in a plant hypersensitive disease resistance re-
sponse. Curr Biol 6: 427–437
Liu Q, Kasuga M, Sakuma Y, Abe H, Miura S, Yamaguchi-Shinozaki K,
Shinozaki K (1998) Two transcription factors, DREB1 and DREB2, with
an EREBP/AP2 DNA binding domain separate two cellular signal
Ogawa et al.
1444 Plant Physiol. Vol. 138, 2005 www.plant.org on June 4, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2005 American Society of Plant Biologists. All rights reserved.

transduction pathways in drought- and low-temperature-responsive
gene expression, respectively, in Arabidopsis. Plant Cell 10: 1391–1406
Lorenzo O, Piqueras R, Sanchez-Serrano JJ, Solano R (2003) ETHYLENE
RESPONSE FACTOR1 integrates signals from ethylene and jasmonate
pathways in plant defense. Plant Cell 15: 165–178
McCabe PF, Leaver CJ (2000) Programmed cell death in cell cultures. Plant
Mol Biol 44: 359–368
McCabe PF, Levine A, Meijer P-J, Tapon NA, Pennell RI (1997) A
programmed cell death pathway activated in carrot cells cultured at
low cell density. Plant J 12: 267–280
Menke FL, Champion A, Kijne JW, Memelink J (1999) A novel jasmonate-
and elicitor-responsive element in the periwinkle secondary metabolite
biosynthetic gene Str interacts with a jasmonate- and elicitor-inducible
AP2-domain transcription factor, ORCA2. EMBO J 18: 4455–4463
Metzler MC, Cutt JR, Klessig DF (1991) Isolation and characterization of
a gene encoding a PR-1-like protein from Arabidopsis thaliana. Plant
Physiol 96: 346–348
Mitsuhara I, Malic KA, Miura M, Ohashi Y (1999) Animal cell-death
suppressors Bcl-x(L) and Ced-9 inhibit cell death in tobacco plants. Curr
Biol 9: 775–778
Murashige T, Skoog F (1962) A revised medium for rapid growth and
bioassays with tobacco tissue culture. Physiol Plant 15: 473–497
Nagata T, Nemoto T, Hasezawa S (1992) Tobacco BY2 cell line as the ‘‘Hela’’
cell line in the cell biology of higher plants. Int Rev Cytol 132: 1–30
Niwa Y, Hirano T, Yoshimoto K, Shimizu M, Kobayashi H (1999) Non-
invasive quantitative detection and applications of non-toxic, S65T-type
green fluorescent protein in living plants. Plant J 18: 455–463
Ohme-Takagi M, Shinshi H (1995) Ethylene-inducible DNA binding
proteins that interact with an ethylene-responsive element. Plant Cell
7: 173–182
Ohta M, Matsui K, Hiratsu K, Shinshi H, Ohme-Takagi M (2001) Re-
pression domains of class II ERF transcriptional repressors share an
essential motif for active repression. Plant Cell 13: 1959–1968
Ohta M, Ohme-Takagi M, Shinshi H (2000) Three ethylene-responsive
transcription factors in tobacco with distinct transactivation functions.
Plant J 22: 29–38
Oltvai ZN, Milliman CL, Korsmeyer SJ (1993) Bcl-2 heterodimerizes in
vivo with a conserved homolog, Bax, that accelerates programmed cell
death. Cell 74: 609–619
Pan L, Kawai M, Yu LH, Kim KM, Hirata A, Umeda M, Uchimiya H (2001)
The Arabidopsis thaliana ethylene-responsive element binding protein
(AtEBP) can function as a dominant suppressor of Bax-induced cell
death of yeast. FEBS Lett 508: 375–378
Penninckx IA, Eggermont K, Terras FR, Thomma BP, De Samblanx GW,
Buchala A, Metraux JP, Manners JM, Broekaert WF (1996) Pathogen-
induced systemic activation of a plant defensin gene in Arabi-
dopsis follows a salicylic acid-independent pathway. Plant Cell 8:
2309–2323
Penninckx IA, Thomma BP, Buchala A, Metraux JP, Broekaert WF (1998)
Concomitant activation of jasmonate and ethylene response pathways is
required for induction of a plant defensin gene in Arabidopsis. Plant
Cell 10: 2103–2113
Rao MY, Davis KR (1999) Ozone-induced cell death occurs via two
distinct mechanisms in Arabidopsis: the role of salicylic acid. Plant J 17:
603–614
Reed JC (1997) Cytochrome c: can’t live with it, can’t live without it. Cell 91:
559–562
Riechmann JL, Heard J, Martin G, Reuber L, Jiang C, Keddie J, Adam L,
Pineda O, Ratcliffe OJ, Samaha RR, et al (2000) Arabidopsis transcrip-
tion factors: genome-wide comparative analysis among eukaryotes.
Science 290: 2105–2110
Rizhsky L, Hallak-Herr E, Van Breusegem F, Rachmilevitch S, Barr JE,
Rodermel S, Inze D, Mittler R (2002) Double antisense plants lacking
ascorbate peroxidase and catalase are less sensitive to oxidative stress than
single antisense plants lacking ascorbate peroxidase or catalase. Plant J 32:
329–342
Samac DA, Hironaka CM, Yallaly PE, Shah DM (1990) Isolation and
characterization of the genes encoding basic and acidic chitinase in
Arabidopsis thaliana. Plant Physiol 93: 907–914
Sato T, Hanada M, Bodrug S, Irie S, Iwama N, Boise LH, Thompson CB,
Golemis E, Fong L, Wang HG, et al (1994) Interactions among members
of the Bcl-2 protein family analyzed with a yeast two-hybrid system.
Proc Natl Acad Sci USA 91: 9238–9242
Shaul O, Mironov V, Burssens S, Van Montagu M, Inze D (1996) Two
Arabidopsis cyclin promoters mediate distinctive transcriptional oscilla-
tion in synchronized tobacco BY-2 cells. Proc Natl Acad Sci USA 93:
4868–4872
Solano R, Stepanova A, Chao Q, Ecker JR (1998) Nuclear events in
ethylene signaling; a transcriptional cascade mediated by ETHYLENE-
INSENSITIVE3 and ETHYLENE-RESPONSE-FACTOR1. Genes Dev 12:
3703–3714
Stockinger EJ, Gilmour SJ, Thomashow MF (1997) Arabidopsis thaliana
CBF1 encodes an AP2 domain-containing transcriptional activator that
binds to the C-repeat/DRE, a cis-acting DNA regulatory element that
stimulates transcription in response to low temperature and water
deficit. Proc Natl Acad Sci USA 94: 1035–1040
Swidzinski JA, Sweetlove LJ, Leaver CJ (2002) A custom microarray
analysis of gene expression during programmed cell death in Arabi-
dopsis thaliana. Plant J 30: 431–446
Thordal-Christensen H, Zhang Z, Wei Y, Collinge DB (1997) Subcellular
localization of H2O2 in plants: H2O2 accumulation in papillae and
hypersensitive response during the barley-powdery mildew interac-
tion. Plant J 11: 1187–1194
Uknes S, Mauch-Mani B, Moyer M, Potter S, Williams S, Dincher S,
Chandler D, Slusarenko A, Ward E, Ryals J (1992) Acquired resistance
in Arabidopsis. Plant Cell 4: 645–656
van der Fits L, Memelink J (2000) ORCA3, a jasmonate-responsive
transcriptional regulator of plant primary and secondary metabolism.
Science 289: 295–297
van der Fits L, Memelink J (2001) The jasmonate-inducible AP2/ERF-
domain transcription factor ORCA3 activates gene expression via in-
teraction with a jasmonate-responsive promoter element. Plant J 25: 43–53
Wang KL, Li H, Ecker JR (2002) Ethylene biosynthesis and signaling
networks. Plant Cell (Suppl) 14: S131–S151
Woeste KE, Ye C, Kieber JJ (1999) Two Arabidopsis mutants that over-
produce ethylene are affected in the posttranscriptional regulation of
1-aminocyclopropane-1-carboxylic acid synthase. Plant Physiol 119:
521–530
Xu P, Narasimhan ML, Samson T, Coca MA, Huh GH, Zhou J, Martin GB,
Hasegawa PM, Bressan RA (1998) A nitrilase-like protein interacts with
GCC box DNA-binding proteins involved in ethylene and defense
responses. Plant Physiol 118: 867–874
Xu Q, Reed JC (1998) Bax inhibitor-1, a mammalian apoptosis suppressor
identified by functional screening in yeast. Mol Cell 1: 337–346
Zha H, Fisk HA, Yaffe MP, Mahajan N, Herman B, Reed JC (1996)
Structure-function comparisons of the proapoptotic protein Bax in yeast
and mammalian cells. Mol Cell Biol 16: 6494–6508
Arabidopsis Ethylene-Responsive Element Binding Protein
Plant Physiol. Vol. 138, 2005 1445 www.plant.org on June 4, 2015 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2005 American Society of Plant Biologists. All rights reserved.
Copyright © 2022 FDOKUMEN




















