
Effect of host and larval frass volatiles on behavioural response of the old house borer
Upload
independentCategory
view
1download
0
Function of defensive volatiles in pedunculate oak(Quercus robur) is tricked by the moth Tortrix viridanapce_2545 1..16
ANDREA GHIRARDO1,2, WERNER HELLER3, MATTHIAS FLADUNG2, JÖRG-PETER SCHNITZLER1 &HILKE SCHROEDER2
1Helmholtz Zentrum München, Institute of Biochemical Plant Pathology, Research Unit Environmental Simulation (EUS),85764 Neuherberg, Germany, 2Johann Heinrich von Thünen Institute (vTI), Institute of Forest Genetics, 22927Grosshansdorf, Germany and 3Helmholtz Zentrum München, Institute of Biochemical Plant Pathology (BIOP), 85764Neuherberg, Germany
ABSTRACT
The indirect defences of plants are comprised of herbivore-induced plant volatiles (HIPVs) that among other thingsattract the natural enemies of insects. However, the actualextent of the benefits of HIPV emissions in complexco-evolved plant-herbivore systems is only poorly under-stood. The observation that a few Quercus robur L. treesconstantly tolerated (T-oaks) infestation by a major pest ofoaks (Tortrix viridana L.), compared with heavily defoli-ated trees (susceptible: S-oaks), lead us to a combined bio-chemical and behavioural study. We used these evidentlydifferent phenotypes to analyse whether the resistance ofT-oaks to the herbivore was dependent on the amount andscent of HIPVs and/or differences in non-volatile polyphe-nolic leaf constituents (as quercetin-, kaempferol- andflavonol glycosides). In addition to non-volatile metabolicdifferences, typically defensive HIPV emissions differedbetween S-oaks and T-oaks. Female moths were attractedby the blend of HIPVs from S-oaks, showing significantlyhigher amounts of (E)-4,8-dimethyl-1,3,7-nonatriene(DMNT) and (E)-b-ocimene and avoid T-oaks with relativehigh fraction of the sesquiterpenes a-farnesene and germa-crene D. Hence, the strategy of T-oaks exhibiting directlyherbivore-repellent HIPV emissions instead of high emis-sions of predator-attracting HIPVs of the S-oaks appears tobe the better mechanism for avoiding defoliation.
Key-words: herbivore-induced plant volatiles; PTR-MS; ter-penes; volatile organic compounds; Y-tube-olfactometer.
INTRODUCTION
Interactions between trees and their pathogenic, parasiticand herbivorous enemies are among the driving factors ofco-evolutionary processes in forest ecosystems (Ehrlich &Raven 1964; Mauricio & Rausher 1997; Nosil, Crespl &Sandoval 2002; Becerra 2003). Plants are subjected toboth constant and seasonal pressures from pathogens (e.g.viruses, bacteria, fungi) as well as from vertebrate and
invertebrate animals.Therefore, plants exhibit well-adaptedand excellently regulated direct and indirect mechanismsof protection against abiotic and biotic factors.
One of the most prominent indirect defence strategies isoften referred to as ‘crying for help’ (Dicke & Baldwin2010). Here, when subjected to herbivore attack, sessileplants invoke aid from the surrounding insect communityby emitting volatile substances that attract predators toprey on the herbivores (Dicke & Sabelis 1988; Arimuraet al. 2000; Kessler & Baldwin 2001; Allison & Hare 2009;Degenhardt 2009). The signals communicated to the envi-ronment are herbivore-induced plant volatiles (HIPVs),which form an invisible cloud of odour around the plantthat is detectable for insects. These signals aim to elicitbehavioural changes in different ecosystem members withapparent beneficial consequences for the plant’s fitness(Dicke & Baldwin 2010). Thus, emissions of HIPV arebelieved to be beneficial for plants.
Monophagous insects can use volatile organic com-pounds (VOCs, syn. plant volatiles) to locate an appropriatehost (Feeny 1975) within a mixed forest stand composed ofdeciduous and coniferous trees (Schröder & Hilker 2008).During oviposition, adult female moths search for treeswhere they can lay their eggs to produce the next insectgeneration (Renwick & Chew 1994; Honda 1995).
VOCs are comprised of mono-, sesqui- and homo-terpenes, fatty-acid derivatives, alcohols, and other sub-stances, such as indole and methyl salicylate, that areproduced only in trace amounts concomitant with manyother constitutive plant VOCs, such as isoprene acetalde-hyde or methanol. VOCs generally exhibit multifunctionalpurposes, such as protection against abiotic stresses (Loreto& Schnitzler 2010), recruitment of pollinators and plant-to-plant signalling, and as noted previously, they appear toprotect plants against herbivore attacks (Pichersky & Ger-shenzon 2002; Dudareva, Pichersky & Gershenzon 2004;Reinhard, Srivivasan & Zhang 2004; Pichersky, Noel &Dudareva 2006; Gershenzon & Dudareva 2007). As plantsinvest many resources (carbon, energy) in the production ofVOCs, they must balance the benefits of VOC biosynthesiswith the cost of their production.Correspondence: H. Schroeder. E-mail: [email protected]
Plant, Cell and Environment (2012) doi: 10.1111/j.1365-3040.2012.02545.x
bs_bs_banner
© 2012 Blackwell Publishing Ltd 1

In the present work, we determined the HIPV emissionsof pedunculate oaks (Quercus robur L.) that were subjectedto different degrees of defoliation by the green oak leafroller (Tortrix viridana L., Lepidopera: Tortricidae) (Fig. 1).During severe outbreak episodes in Germany when T. viri-dana caused almost complete defoliation of oaks, we usedthe observation that a few individual oaks were remarkablyless defoliated than neighbouring trees. We defined theseless-defoliated phenotypes as resistant (T-oaks) and heavilydefoliated phenotypes from the same forest stand assusceptible (S-oaks).
Using grafted plants of these S-oak and T-oak pheno-types, we investigated in the laboratory the underlyingphysiological and biochemical mechanisms responsible forthe differences in susceptibility to the insect outbreaksobserved in their natural habitat. By employing a combina-tion of behavioural, biochemical, and molecular approachesat the larval and adult developmental stages, we tested thefollowing hypotheses:
1 Adult T. viridana females and larvae are able to differen-tiate between S-oaks and T-oaks.
2 The resistance of some phenotypes to T. viridana is dueto differences in the HIPV emission patterns of S-oaksand T-oaks or in other plant’s secondary metabolites.
3 Larvae developing on leaves from less defoliated T-oakscompared with larvae developing on heavily defoliatedS-oaks (1) require a longer period for development; (2)exhibit a higher mortality rate; and/or (3) gain less weightas larvae or pupae in either individual or in all develop-mental stages.
MATERIALS AND METHODS
Plant and insect material
During the last outbreak of the green oak leaf roller in2003–2005, we identified pedunculate oak trees in NorthRhine-Westphalia (western-central region of Germany)that were heavily (defined as susceptible: S-oaks) or slightly(defined as resistant:T-oaks) defoliated by estimation of thepercent defoliation using standard pictures for all kinds ofdefoliation levels (Evers et al. 1997).These trees belonged tothree genetically different populations (Table 1, details onthe oak stands reported in Schroeder & Degen 2008). Weselected late and early bud bursting individuals among bothresistant and susceptible oaks. Among all forest standsanalysed, the selected S-oaks and T-oaks from the popula-tion ‘Asbeck’ (Table 1) showed the most obvious differencesin defoliation rate. In July 2008,we took branches from thesetrees and grafted them onto Q. robur saplings (Schröder2010). Hybridization between Q. robur and Quercus petraeais quite common in natural oak populations and the hybridsare often difficult to distinguish based upon morphology(Curtu, Gailing & Finkeldey 2009).Therefore, the affiliationof the selected individuals with the species Q. robur (pedun-culate oak) was ascertained using eight microsatellitemarkers located in five different linkage groups for theirspecies pureness (Scotti-Saintagne et al. 2004). Five of the
eight grafted phenotypes resulted in being pure Q. robur.Thus, all experiments were carried out using the followingfive pure phenotypes of Q. robur-grafted plants: ASB2a,ASB14a, ASB17a, ASB13b and ASB47b; where a indicatesresistant and b indicates susceptible oaks (Table 1).
The moth T. viridana is an oligophagous, univoltine insectfeeding on different species of the genus Quercus with apreference for pedunculate oaks. Female adults lay eggs onbranches proximate to next year’s buds during June/July.The eggs overwinter on the branches, and hatching of thefirst instar larvae begins in mid-March to April, when newlyhatched caterpillars feed on the trees’ newly opened buds.Heavily infested trees become defoliated as early as May. Inlate May or beginning of June, the fully grown caterpillars(5th instar) go into the pupa stage on remaining leaves.Moths begin to emerge approximately 14 d later and startto search for appropriate host oaks to lay eggs on twigs forproduction of the next generation. The development of the1st instar larvae depends on the physiological state offreshly opened buds (Hunter 1990), while the later success-ful development of immature stages depends on the growthrate of host leaves (Ivashov, Boyko & Simchuk 2002).
In 2006, we sampled T. viridana larvae in 15 oak stands inNorth Rhine-Westphalia, Germany. After hatching of adultmoths, we crossed combinations: (1st) from the same treewithin a stand, (2nd) from different trees within a stand and(3rd) from different stands in an area of approximately30 ¥ 30 km. In 2006, we performed a total of 25 crosses.Since 2007, we have performed new crosses yearly (up to150 in the year 2011), and the eggs that emerged from thesecrosses were hibernated under natural field conditions.
All our experiments were performed using 3rd or 4thinstars because the gut conditions of younger instars (1stand 2nd) may still be influenced by host plant characteris-tics (Appel & Maines 1995), whereas later instars (5th) mayinfluence plant’s defence responses themselves.
Rearing conditions
Depending on the outside temperature in early to lateMarch, twigs with hibernated eggs were transferred fromnatural conditions to a cold-storage room of 2 to 4 °C and inApril and early May to an air-conditioned room to delaytheir hatching. In this culture room, the room temperaturewas 18–20 °C and air humidity was around 40%. Light wasprovided by artificial light from 0700 to 1700 h, with a lightintensity of approx. 50 mmol photons m-2 s-1 photosyntheti-cally active radiation (PAR).
The newly hatched larvae were set upon buds that justopen using a small brush. Up to two larvae were placed oneach bud. The small twigs with the buds were placed in1.5 mL water-filled plastic tubes to avoid desiccation. Up tofour buds were placed in 250 mL glass jars. Normally, the 1stinstars hide within the buds until they moulted to the 2ndinstars. At that stage, the larvae came out looking for freshbuds.When the 2nd instars left the buds after 4 d, they wereplaced on new ones. The glass jars were controlled daily forneed of fresh buds or for later instars fitting fresh young
2 A. Ghirardo et al.
© 2012 Blackwell Publishing Ltd, Plant, Cell and Environment
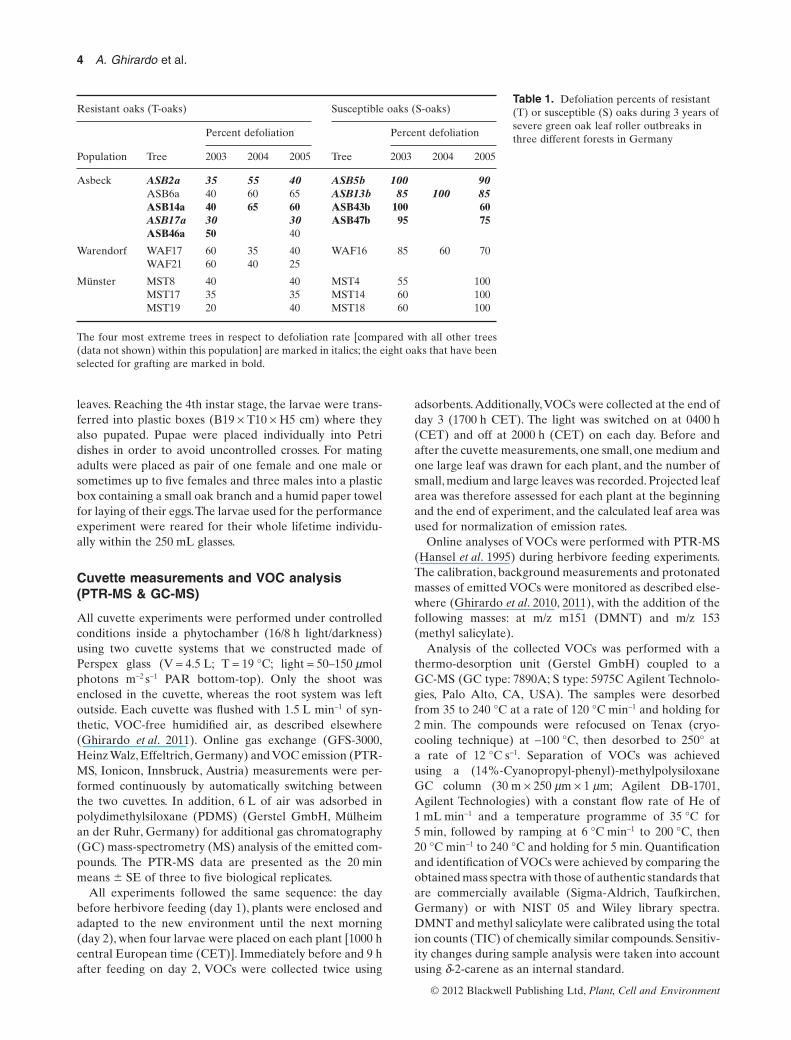
Figure 1. Behavioural choice of T. viridana adults and larvae between S-oaks and T-oaks. (a) Mated and naive female moths ofT. viridana were assayed in a Y-tube olfactometer for testing the preferences of S-oak ( ) and T-oak ( ), which were previously fed for24 h with larvae of T. viridana 2 d before (n = 62), and totally unfed (n = 35), and one fed versus an empty cuvette (n = 34). (b) Firstdecision taken from larvae previously grown up on leaves from S-oaks and T-oaks, and (c) amount of leaf-mass consumed. Amount of fedleaf area were classified in black ( ) when both leaves were untouched, in grey ( ) when both leaves showed feeding marks. Differencesin choice behaviour between S-oaks and T-oaks were tested by z-test; an asterisk means P < 0.01.
Detrimental effect of HIPV emissions in oaks 3
© 2012 Blackwell Publishing Ltd, Plant, Cell and Environment
leaves. Reaching the 4th instar stage, the larvae were trans-ferred into plastic boxes (B19 ¥ T10 ¥ H5 cm) where theyalso pupated. Pupae were placed individually into Petridishes in order to avoid uncontrolled crosses. For matingadults were placed as pair of one female and one male orsometimes up to five females and three males into a plasticbox containing a small oak branch and a humid paper towelfor laying of their eggs.The larvae used for the performanceexperiment were reared for their whole lifetime individu-ally within the 250 mL glasses.
Cuvette measurements and VOC analysis(PTR-MS & GC-MS)
All cuvette experiments were performed under controlledconditions inside a phytochamber (16/8 h light/darkness)using two cuvette systems that we constructed made ofPerspex glass (V = 4.5 L; T = 19 °C; light = 50–150 mmolphotons m-2 s-1 PAR bottom-top). Only the shoot wasenclosed in the cuvette, whereas the root system was leftoutside. Each cuvette was flushed with 1.5 L min-1 of syn-thetic, VOC-free humidified air, as described elsewhere(Ghirardo et al. 2011). Online gas exchange (GFS-3000,Heinz Walz, Effeltrich, Germany) and VOC emission (PTR-MS, Ionicon, Innsbruck, Austria) measurements were per-formed continuously by automatically switching betweenthe two cuvettes. In addition, 6 L of air was adsorbed inpolydimethylsiloxane (PDMS) (Gerstel GmbH, Mülheiman der Ruhr, Germany) for additional gas chromatography(GC) mass-spectrometry (MS) analysis of the emitted com-pounds. The PTR-MS data are presented as the 20 minmeans � SE of three to five biological replicates.
All experiments followed the same sequence: the daybefore herbivore feeding (day 1), plants were enclosed andadapted to the new environment until the next morning(day 2), when four larvae were placed on each plant [1000 hcentral European time (CET)]. Immediately before and 9 hafter feeding on day 2, VOCs were collected twice using
adsorbents.Additionally,VOCs were collected at the end ofday 3 (1700 h CET). The light was switched on at 0400 h(CET) and off at 2000 h (CET) on each day. Before andafter the cuvette measurements, one small, one medium andone large leaf was drawn for each plant, and the number ofsmall, medium and large leaves was recorded. Projected leafarea was therefore assessed for each plant at the beginningand the end of experiment, and the calculated leaf area wasused for normalization of emission rates.
Online analyses of VOCs were performed with PTR-MS(Hansel et al. 1995) during herbivore feeding experiments.The calibration, background measurements and protonatedmasses of emitted VOCs were monitored as described else-where (Ghirardo et al. 2010, 2011), with the addition of thefollowing masses: at m/z m151 (DMNT) and m/z 153(methyl salicylate).
Analysis of the collected VOCs was performed with athermo-desorption unit (Gerstel GmbH) coupled to aGC-MS (GC type: 7890A; S type: 5975C Agilent Technolo-gies, Palo Alto, CA, USA). The samples were desorbedfrom 35 to 240 °C at a rate of 120 °C min-1 and holding for2 min. The compounds were refocused on Tenax (cryo-cooling technique) at -100 °C, then desorbed to 250° ata rate of 12 °C s-1. Separation of VOCs was achievedusing a (14%-Cyanopropyl-phenyl)-methylpolysiloxaneGC column (30 m ¥ 250 mm ¥ 1 mm; Agilent DB-1701,Agilent Technologies) with a constant flow rate of He of1 mL min-1 and a temperature programme of 35 °C for5 min, followed by ramping at 6 °C min-1 to 200 °C, then20 °C min-1 to 240 °C and holding for 5 min. Quantificationand identification of VOCs were achieved by comparing theobtained mass spectra with those of authentic standards thatare commercially available (Sigma-Aldrich, Taufkirchen,Germany) or with NIST 05 and Wiley library spectra.DMNT and methyl salicylate were calibrated using the totalion counts (TIC) of chemically similar compounds. Sensitiv-ity changes during sample analysis were taken into accountusing d-2-carene as an internal standard.
Table 1. Defoliation percents of resistant(T) or susceptible (S) oaks during 3 years ofsevere green oak leaf roller outbreaks inthree different forests in Germany
Resistant oaks (T-oaks) Susceptible oaks (S-oaks)
Population Tree
Percent defoliation
Tree
Percent defoliation
2003 2004 2005 2003 2004 2005
Asbeck ASB2a 35 55 40 ASB5b 100 90ASB6a 40 60 65 ASB13b 85 100 85ASB14a 40 65 60 ASB43b 100 60ASB17a 30 30 ASB47b 95 75ASB46a 50 40
Warendorf WAF17 60 35 40 WAF16 85 60 70WAF21 60 40 25
Münster MST8 40 40 MST4 55 100MST17 35 35 MST14 60 100MST19 20 40 MST18 60 100
The four most extreme trees in respect to defoliation rate [compared with all other trees(data not shown) within this population] are marked in italics; the eight oaks that have beenselected for grafting are marked in bold.
4 A. Ghirardo et al.
© 2012 Blackwell Publishing Ltd, Plant, Cell and Environment

Y-tube olfactometer experiments withadult moths
The responses of T. viridana naive, but mated adult femalesto plant volatiles were tested in a Y-shaped glass tube olfac-tometer (a Y-tube) (2 cm i.d., main arm = 20 cm length, sidearms = 12.5 cm length, angle of 140°) made by a glassblower (Bruno Kummer GmbH, Freiburg, Germany). Theolfactometer generated two laminar airflows originatingfrom S-oak (one Y-tube arm) and T-oak (second Y-tubearm) plants (enclosed in cuvettes). Individual naive adultfemales of T. viridana were placed on the downwind side ofthe tube, and their choice for either odour source wasrecorded. A choice was recorded when females reached atleast the halfway point of either source arm and stayedthere for several minutes. The results from females that didnot satisfy these requisites within 30 min were classified asno choice. Each day, two new plants were enclosed in thecuvettes with opposite phenotypes (i.e. S-oaks took place ofT-oaks and vice versa). In 2010, the choice of 62 T. viridanafemales were tested in seven replicates with S-oak andT-oaks that had been fed upon 24 h before the Y-tubeexperiments started.
In 2011, additional combinations were tested: (1) 35females could choose between unfed plants of both pheno-types; (2) 30 females could select between fed S-oaksagainst an empty cuvette (blank); and (3) 4 females weretested with a fed T-oak against an empty cuvette (blank).
Estimation of egg-numbers from natural T-oaksand S-oaks
To clarify the question, whether the number of eggs differsbetween twigs of T-oaks and S-oaks, we harvested inNovember 2010 branches of three susceptible and fourresistant oaks from the ‘Asbeck’ population from four sidesof each tree (North, South, East and West). Each branch(length 2 m) was cut in pieces, deposited in bags and hiber-nated under natural conditions behind the institute atGroßhansdorf. In March 2011, branch sections (reflecting alength of 1.25 m) were placed in glass jars and stored atroom temperature to count the hatched larvae per branch.The other branch sections were stained for up to 60 minin a DAPI solution [5 mg mL-1 (w/v), DAPI stains DNAin different tissues, Kapuscinski 1995]. After staining,the branch sections were stored in an antifade solution(DABCO) and the number of DAPI-stained eggs was cal-culated under UV-light with a binocular microscope. Stain-ing of the eggs was necessary, as eggs of T. viridana have asticky surface and therefore dust, small algae and otherparticles became deposited during hibernation, makingdirect counting of eggs almost impossible (Gasow 1925).
Y-tube olfactory and optical experimentswith larvae
In the same Y-tube olfactometer used for the adults, thelarvae were tested using (1) olfactory and (2) optical stimuli
originating from leaves taken from S-oaks or T-oaks. Olfac-tory behaviour was assayed in the same way as for theadults. In total, 64 larvae were tested for olfactory stimuli.
The optical behaviour of T. viridana larvae was assayedusing the same Y-tube by placing artificial and naturalleaves of different colours upon its arms. The artificialleaves consisted of oak leaf-shaped coloured paper. Wetested a total of 72 larvae in three different treatments: (1)40 larvae were tested using one leaf from a S-oak versus oneleaf from a T-oak; (2) 16 larvae were used for decisionbetween one green paper leaf versus one red paper leaf; and(3) also, 16 larvae were presented one green paper leafversus one white paper leaf.As for the females, a choice wasrecorded when larvae reached at least the halfway point ofeither source arm and stayed there for several minutes. Theresults from larvae that did not satisfy these requisiteswithin 30 min were classified as no choice.
Performance of larvae and feedingchoice experiments
In 2010, 182 larvae were individually reared on S-leaves and145 on T-leaves. In 2011, 205 larvae each were reared onS-leaves or T-leaves. For each larva, four parameters weremeasured:
1 Stage of development (larval instar): The larvae wereobserved daily and the larval stage was noticed;
2 Larval weight: The larvae were weighed on an analyticalbalance every 4th day during their larval stages;
3 Timing of pupa formation: number of days from hatchingto the pupa stage. This was measured when the pupa wasfully coloured; and
4 Amount of fed leaf biomass (only in 2010). Additionally,the mortality rates of each developmental stage and thepupa were assessed.
For feeding choice experiments, only larvae fed duringtheir entire instar development either on S-leaves orT-leaves were used. Ninety-one larvae were tested using atypical cafeteria experiment: each larva (3rd or 4th instar)was carefully placed between one T-leaf and one S-leaf lyingon either side of a plastic box (B19 ¥ T10 ¥ H5 cm). Thelarvae remained in the boxes for 24 h and were continuouslyphotographed every 3 min by a webcam.We documented (1)the first choice and (2) occasional movement between thetwo leaves (for instance, whether larvae changed their pref-erence after biting either a T-leaf or S-leaf). Decision wasclassified as ‘no decision’ when larvae did not make a choicewithin 24 h and no leaf material was fed.Before and after theexperiment, the weight of each leaf was measured to esti-mate leaf area losses by feeding. Totally untouched leaveswere used to calculate dehydration.The mean of the weightloss of these control leaves was subtracted from the overallweight loss due to larvae feeding.
Analysis of phenolic compounds
Freshly harvested leaves were shock frozen in liquid nitro-gen and stored at -80 °C. For further analysis, the material
Detrimental effect of HIPV emissions in oaks 5
© 2012 Blackwell Publishing Ltd, Plant, Cell and Environment

was homogenized in a mortar under liquid nitrogen to afine powder. Aliquots of approximately 50 mg were thenextracted in an exactly 10-fold volume (w/v) methanol[high-performance liquid chromatography (HPLC) grade]at 4 °C overnight. For RP-HPLC analysis, 75 mL extract wasmixed with 25 mL pure H2O. Precipitated lipophilic materialwas removed by centrifugation for 10 min at 9000 g, and10 mL aliquots of supernatant were immediately analysedby HPLC (Beckman HPLC System Gold, Beckman,Munich, Germany; column 250 ¥ 4.5 mm Spherisorb ODScolumn (NC, 5 mm, Bischoff, Leonberg, Germany, at 20 °C,and a flow rate of 1 mL min-1) as described in Turunen et al.(1999). The following solvents were used for gradientelution: solvent A [980 mL H2O plus 20 mL 5% ammoniumformate in formic acid (98%, w/v)] and solvent B [882 mLmethanol plus 96 mL H2O plus 20 mL ammoniumformate in formic acid (98%, w/v)]. The gradient appliedwas 0–5 min, 100% A (isocratic); 5–45 min, 0–100% B,45–48 min, 100% B, 48–58 min 100–0% B and 58–60 min100% A. Detection was at 280 nm with a UV/visible diode-array detector (Beckman Model 168).
Analysis of terpene synthase (TPS) activities
Protein extracts were prepared using the buffers andprocedures described in Schnitzler et al. (2004). Analysisof monoterpenes and sesquiterpene synthase activitiesfrom geranyl diphosphate (GPP) and farnesyl diphosphate(FPP), respectively, was performed using a new protocoldescribed in Köksal et al. (2010). In this newly developedTPS assay, enzymatically produced mono- or sesquiterpe-nes were trapped from the aqueous matrix by stir barsorptive extraction (SBSE) with PDMS coated stir bars(Twister, PDMS film thickness 0.5 mm, Gerstel). Removingthe stir bars terminated the assays. After rinsing inde-ionized H2O, the twisters were analysed using TDU andGC-MS as described previously.
Statistical and multivariate data analysis
All gas exchange and VOC experiments were performedindependently with three to five biological replicates, andthe results are shown as the average � SE. In addition, eachTPS analysis was also performed with three technicalreplicates. Statistical significance was assessed using thesoftware package Sigma plot (v. 11.0, Systat Software Inc.,Chicago, IL, USA).
Principal component analysis (PCA) and partial leastsquare regression (PLSR) techniques and statisticalMartens’ test (Martens & Martens 2000) were performedusing the software package ‘The Unscrambler’ (v. 8.0,CAMO A/S, Trondheim, Norway).
All data were initially explored by PCA before process-ing the remaining data using the supervised MDA methodPLSR. Thus, we first ensured that the data could be clus-tered according to phenotype. In the PLSR,Y described thephenotype, with T-oaks = 1 and S-oaks = 0; and the X matrixcontained TPS activity, VOC emission rates or individual
relative phenolic compound abundance values. Datawere pre-processed by logarithmic transformation withX = |Log(X)| and weighted with 1 per SD to decrease thedifference between large and small values and provide thesame chance for each independent variable to influencethe calibration model. A random subset of the data (half ofthe overall number of samples) was used to create modelspredicting of oak phenotypes, whereas the second half wasused to validate these models.
RESULTS
Preferences of adults and larvae of T. viridana
To test hypothesis one, we observed the behaviour of naive,but mated adult females in a Y-tube olfactometer. Wheneach arm of the olfactometer was supplied with a headspaceof either S-oaks or T-oaks (experiment performed in 2010),the adult females significantly more often chose S-oaks[66% S versus 8% T (z-test, P < 0.01), 26% no decision]when plants were previously fed by larvae, but none of theadults made a choice if unfed plants were used (experimentperformed in 2011) (Fig. 1a).
The combinations either one fed S-oak or T-oak againstan empty cuvette of the year 2011 resulted in only twofemales choosing an empty cuvette, whereas 20 or 25%decided for the S-oak or T-oak, respectively (z-test:P < 0.01). The overall reaction of the females within thisexperiment was even low, resulting in up to nearly 80% ofthe females making no choice (Fig. 1a). In summary, theY-tube experiments with the adult females showed that theblend of HIPVs emitted by S-oaks was more attractive andpreferred by the females compared with the blend ofHIPVs released by T-oaks.
Olfactory and optical analyses using Y-tube assaysrevealed that the larvae employed optical stimuli ratherthan olfactory stimuli to locate leaves (Supporting Informa-tion Fig. S1). Out of 64 larvae, only 12 showed a reaction toan olfactory stimulus (data not shown). These results indi-cate that the olfactory capability of the larvae is weak. Tocheck this conjecture, we tested the optical capability oflarvae using the same set-up with empty cuvettes and bydepositing leaves outside of the Y-tube. Larvae moved inthe direction of the leaves within a time of 20 to 30 min, withno preference being observed between leaves from S-oaksor T-oaks [25% S versus 27% T (z-test, P = 0.83), 48% nodecision, Supporting Information Fig. S1a]. When differ-ently coloured leaves cut from paper (with an oak leafshape) were used, all 32 larvae chose the green artificialleaves, and none chose the red or white leaves [35% greenleaves, 0% white leaves, 0% red leaves (z-test, P < 0.001),65% no decision, Supporting Information Fig. S1b].
Estimation of egg numbers on branches ofT-oaks and S-oaks
In 2010, the susceptible oaks in the ‘Asbeck’ populationbecame heavily defoliated by T. viridana, thus we expected
6 A. Ghirardo et al.
© 2012 Blackwell Publishing Ltd, Plant, Cell and Environment

a high number of eggs, at least on branches of theS-phenotypes. On average, a branch of 50 cm in lengthcould carry up to 40 buds, thus offering a place for up to afew hundred first instar larvae (Schütte 1957). Counting ofDAPI-stained eggs resulted in high numbers of eggs: 0.35eggs cm-1 on S-oak twigs and 0.77 eggs cm-1 on T-oak twigs.Whereas counting of the hatched larvae resulted in vastlower numbers. On S-oaks, only 7 larvae that meant 0.008eggs per cm branch and on T-oaks 14 larvae (0.01 eggs cm-1)were counted.
Cafeteria experiment and performance of larvae
In cafeteria experiments, we investigated whether greenoak leaf roller larvae discriminate between S-oaks andT-oaks.When larvae developed on S-oaks, they significantly(z-test: P < 0.01) preferred S-leaves as their first choice(70% S versus 24% T; Fig. 1b) and consumed proportionallyless leaf material from T-oaks than from S-oaks (47% Sversus 17% T; z-test: P = 0.058; Fig. 1c). In contrast, whenlarvae developed on T-leaves, they chose S- and T-leavesalmost equally as their first choice (55% S versus 45% T;z-test: P = 0.766; Fig. 1b), and the amount of material con-sumed was also similar (36% S versus 34% T; z-test:P = 0.857; Fig. 1c).
Assimilation of secondary plant metabolites has a largeimpact on the performance of herbivores. Therefore, wetested the growth performance of larvae developing onS-oaks or T-oaks. The results showed no significant differ-ences in either 2010 or 2011 in any of the larval parametersanalysed. For the larvae developed on S-oaks, the averagetimespan of development from the 1st instar to the pupaestage was 17.1 � 1.9 d and for the larvae developed onT-oaks 16.8 � 1.6 d, respectively (t-test, P = 0.47). Theoverall development (until the adult stage) took 29 � 2.0 don S-oaks and 28.7 � 1.7 d on T-oaks, respectively (t-test,P = 0.53).
The mortality rate of larvae developing on either T-oaksor S-oaks was similar, with no significant difference beingdetected: in 2010, from 182 (on S-oaks) and 145 (on T-oaks)caterpillars, 45 each survived, meaning 75% mortality rateon S-oaks and 69% mortality rate on T-oaks. In 2011, from198 (on S-oaks) and 199 (on T-oaks) caterpillars, only 45and 40 survived on S-leaves and on T-leaves, respectively(mortality rate of 77% on S-oaks versus 80% on T-oaks).Pupal weight also did not differ between the two foodsources, being on average 33.1 � 6.1 mg for the pupaedeveloped on S-oaks and 33.9 � 7.8 mg for the pupaedeveloped on T-oaks (t-test: P = 0.68) (Fig. 2a). Instead, theamount of leaves ingested differed greatly: larvae thatdeveloped on T-oaks required significantly more leaf mate-rial (31.7 � 9.8 cm2 leaves) than larvae that developed onS-oaks (22.4 � 9.5 cm2 leaves) (t-test: P < 0.002) (Fig. 2b).Thus, the larvae needed to eat and digest more T-leaf mate-rial to reach the same pupal weight.This difference in eatenleaf biomass is not a result of better leaf quality of offspringproduction as 1st instars of all crosses were randomly dis-tributed among the experiments. A second evidence for
equal initial conditions is the fact that the weight as 3rdinstars did not differ at the beginning of the experiment(7.1 � 4.8 mg for larvae on S-oaks, 6.3 � 4.7 mg for larvaeon T-oaks, t-test: P = 0.67).
Phenolic compounds in S-oaks and T-oaks
Phenolic compounds play an important role in resistanceagainst pathogens and insects (Lattanzio, Lattanzio & Car-dinali 2006). We found that the phenolic compositions ofS-oak and T-oak phenotypes already differed prior tofeeding (Supporting Information Fig. S2). Three com-pounds with the spectra and retention time of a catechinderivate (proanthocyanidin), two quercetin glycosides withdifferent sugar moieties, and an unknown phenolic deriva-tive (Supporting Information Fig. S2b–d,f) were found tobe more abundant in T-leaves, while the levels of anotherquercetin 3-glycoside, a kaempferol 3-glycoside and amono-acylated flavonol 3-glycoside, were higher in S-leafextracts (Supporting Information Fig. S2e,g). No differ-ences were found in individual compounds between intact(systemic) and larvae-pitted (local) leaves 48 h after placingthe larvae on a plant (Supporting Information Fig. S2).
Herbivore-induced emissions in S-oaksand T-oaks
Plants subjected to herbivore attack produce specific vola-tile signals that efficiently call other insects to prey on cat-erpillars (e.g. Kessler & Baldwin 2001). When we analysedthe blend of VOCs released from S-oak and T-oak pheno-types, we observed typical herbivore-induced compounds,such as the homoterpene DMNT and the monoterpene(E)-b-ocimene. However, the emissions of these com-pounds were higher in S-oaks than in T-oaks (Figs 3 & 4).Conversely, the scent produced by T-oaks contained higheramounts of the sesquiterpenes (Fig. 3) a-farnesene, b-bourbonene and germacrene D (Fig. 4).
Using PTR-MS analysis, we followed online the rapidand transient changes in the emission profiles of oaksbefore, during and after larval feeding for 2.5 d. The VOCs
Figure 2. Performance of larvae. Bar plot data as the medianand 5th/95th percentiles of (a) pupal weight gained of allindividuals developed on leaves either from S-oaks ( ) or T-oaks( ) and (b) amount of leaf material from S-oaks and T-oaksconsumed by larvae (n = 53). Statistically significant differencesare marked with asterisk (t-test) (*P < 0.05).
Detrimental effect of HIPV emissions in oaks 7
© 2012 Blackwell Publishing Ltd, Plant, Cell and Environment

most relevant to larvae feeding were mono-, sesqui- andhomoterpenes (Fig. 3a–c), and the shikimate pathway-derived compound methyl salicylate (Fig. 3d). With a fewhours after addition of larvae to the plants, these com-pounds became detectable. Reduction of emission ratesduring the night reflected the scarcity of light-dependentdelivery of intermediates. Switching on the lights the nextmorning resulted in a very fast increase of emission rates,most remarkably observed for the very rapid and steepincrease in monoterpenes emission (Fig. 3a). Emissions ofgreen leaf volatiles (GLV), acetaldehyde and methanol alsoresponded rapidly to herbivore feeding, though showing nodifferences between the oak phenotypes (Supporting Infor-mation Fig. S3). In particular, GLV emissions fluctuatedstrongly, reflecting individual feeding events of the larvae.The strong increase of GLV emissions during the earlyhours of night indicated also that the larvae preferred to eatduring the darkness. Isoprene, the most common VOCemitted by deciduous oaks in a light-dependent manner,was not induced by herbivory but showed a steep increase
over time (Supporting Information Fig. S4) that is charac-teristic of developing leaves (Ghirardo et al. 2011). Beforethe addition of larvae, the leaves also emitted small,light-dependent amounts of monoterpenes. Duringlarval feeding, monoterpene emissions were induced morestrongly in the S-oaks compared with the T-oaks (Fig. 3a).The same behaviour, but with even stronger expression, wasobserved for DMNT, with significantly higher amountsemitted by the two S-phenotypes. In contrast, T-phenotypesexhibited higher emission rates for sesquiterpenes (Fig. 3b).This observation is in accordance with the GC-MS datasampled in parallel to the online analysis 48 h after onsetof caterpillar feeding (Fig. 4). Regarding the induction ofmethyl salicylate, no difference was detected between theS-oaks and T-oaks.
Because insects respond to the specific proportion ofan individual volatile in a blend of volatiles (Dicke 1994;
Figure 3. Temporal emission profile of HIPV before and duringT. viridana larval-feeding. (a) Monoterpene, (b) sesquiterpene,(c) DMNT and (d) methyl salicylate emissions of phenotypicallydifferent S-oaks (47b , 13b ) and T-oaks (17a , 2a ). Eachplant was fed with four larvae of T. viridana ( ). Dark phasesare symbolized in grey ( ). Cuvette conditions: T = 19 °C,light = 50–150 mmol photon m-2 s-1 PAR. Data represent means ofthree experiments � SE. CET, central European time; HIPV,herbivore-induced plant volatile; PAR, photosynthetically activeradiation.
Figure 4. Most relevant HIPV emissions. (a) The emissionsof S-oaks ( ) and T-oaks ( ) after 2 d of larvae feeding and(b) emission pattern normalized to DMNT. Nomenclature:1, (E)-b-ocimene; 2, (Z)-b-ocimene; 3, limonene; 4,(E)-b-caryophyllene; 5, a-cubebene; 6, a-farnesene; 7,b-bourbonene; 8, b-cubebene; 9, germacrene B; 10, germacreneD; 11, germacrene; 12, DMNT; 13, methyl salicylate; 14, indole.Data represent means of four experiments � SE. Statisticallysignificant differences between T-oaks and S-oaks are shown withasterisk (t-test) (*P < 0.05). HIPV, herbivore-induced plantvolatile.
8 A. Ghirardo et al.
© 2012 Blackwell Publishing Ltd, Plant, Cell and Environment

Takabayashi & Dicke 1996), we investigated the differencesbetween S-oaks and T-oaks with respect to DMNT (themost typical HIPV of oaks). The contribution of sesquiter-penes in the blend of volatiles was much higher in T-oakscompared with S-oaks (Fig. 4b).
In combination with the behavioural data on adult moths(Fig. 1a) and the performance of larvae (Fig. 2), theseanalyses indicated that the vulnerability of S-oaks was trig-gered by the ability of females to relocate S-oaks, whichbetter suit their diet, using their specific HIPV pattern.
Key compounds identified by MDA
We aimed to distinguish oak phenotypes based on TPSactivities (Supporting Information Fig. S5), herbivore-induced plant volatile emission patterns and phenolic com-position using multivariate data analysis (MDA). Themono- and sesquiterpene synthase (TPS) activity (Figs 5a &Supporting Information Fig. S5) and emission (Fig. 5d) datascores were successfully separated into two distinct clustersrepresenting resistant (marked red in Fig. 5) and suscep-tible (marked blue in Fig. 5) oaks belonging to the twophenotypes. The first and second principal components(PC1+PC2) together explained 82 and 37% of the variationin the enzymatic and emission data, respectively. Thisenabled the separation of S-samples from T-samples using79 and 87% of the phenotype information (Y-values),respectively. MDA revealed that the monoterpene (E)-b-ocimene and the homoterpene DMNT were more corre-lated with S-oaks, whereas T-oaks were more prominentlyassociated with the sesquiterpenes a-farnesene, germacreneD and b-caryophyllene (Fig. 5b,e). The compounds thatwere significant for describing the two clusters werea-farnesene, germacrene D, (E)-b-ocimene and DMNT(indicated with * on Fig. 5b,e).
Discriminative PLSR is often used to predict unknownresponse values (Esbensen 2002). Thus, we created aPLSR model (Fig. 5c,f) to test whether the phenotypes ofunknown oak samples could be predicted by measuringonly either VOC or TPS activity.The correlations and slopesobtained for both modelling (in blue) and validation (inred) were close to 1, indicating that the prediction ofunknown oak phenotypes based on their herbivore-inducedterpene biosynthesis and emission data might be highlyaccurate.
Application of PCA allowed us to discover intrinsic dif-ferences in the polyphenol patterns between S-oaks andT-oaks (Fig. 6a). The data also reflected the developmentalchanges in plants between the younger plants used for thelarval feeding experiments and the plants used 4 weekslater for the Y-tube experiments when T. viridana devel-oped into adult female moths. Most of the variation(PC1 = 33%) between samples could be ascribed to leafdevelopment; samples from trees at an early developmentalstage clustered to the left, while samples from trees thatwere 4 weeks older clustered to the right side of the plotshown in Fig. 6. The second greatest amount of variation(PC2 = 13%) described the differences between S-oaks and
T-oaks fairly well. Both of the phenotypes differing in sus-ceptibility to T. viridana could be separated by means ofdevelopmentally independent differences in the concentra-tions of phenolic compounds. PLSR analysis (Fig. 6b,c)revealed that the concentration differences of severalmethanol-soluble phenolic compounds were correlatedwith the T-oaks rather than the S-oaks. In particular, thequercetin 3-glycoside (peaks #20), kaempferol 3-glycoside(peak #21), two unknown compounds (peaks #17 and #18)and acylated flavonoid-3-glycoside (peak #29) coulddescribe the two oak types fairly well.
DISCUSSION
HIPVs used for host plant location byfemale insects
Plants are considered to emit HIPVs that carry informationthat can function as cues for organisms of all trophic levelsincluding plants, herbivores, and predators and parasitoidsfor the purpose of eliciting behavioural change in differentecosystem members with beneficial consequences forthe plant’s fitness (Dicke & Baldwin 2010; Kessler & Heil2011). Driven by these olfactory stimuli, adult femaleT. viridana used larval-induced defensive volatiles ofpedunculate oaks to locate trees that might feed their off-spring better in the following year. Their decision was trig-gered by a blend of HIPVs typically produced by plantsas defensive signals to attract species that prey on theirattackers (e.g. Arimura et al. 2000; Kessler & Baldwin2001; Kappers et al. 2005) but that is perceived by thepest to indicate suitable oaks for infestation (Fig. 7).
Chemical cues are widely used by mono- and oligopha-gous insects searching for host plants (Schoonhoven, vanLoon & Dicke 2005). In addition to host volatiles beingused to locate an appropriate host, recognition of non-hostsis essential in the process of host location by insects (Bruce& Pickett 2011). It is known that bark beetles are able todiscriminate between volatiles emitted by non-hosts andothers emitted by hosts to find an appropriate tree foroviposition (Schröder & Hilker 2008). Additionally, leafbeetles of the species Chrysomela populi L. (Coleoptera:Chrysomelidae) use VOCs to seek food for self-nutritionand oviposition. In this case, food seeking is driven by VOCsthat are typically emitted from young developing andhealthy poplar leaves (Brilli et al. 2009; Ghirardo et al.2010). In the case described herein, during oviposition, it isessential for female moths to relocate the plants where theypreviously fed as larvae or trees with similar benefits fortheir offspring. These potential host trees represent thecorrect site for the deposition of their eggs to produce thefollowing generation. Female moths, which are no longerfeeding, can choose the best host plant by searching forfamiliar odour profiles. Apparently, T. viridana femalesused the typical herbivore-induced molecules (E)-b-ocimene and DMNT (Arimura et al. 2000; Kappers et al.2005; Cha et al. 2008) to recognize the most appropriateQ. robur plants for their offspring (Fig. 7).
Detrimental effect of HIPV emissions in oaks 9
© 2012 Blackwell Publishing Ltd, Plant, Cell and Environment

Figure 5. PLSR of TPS activities and VOC emissions. (a, d) Scores, (b, e) X- and Y-loading, of PLSR computed from (a, b) in vitro TPSactivities and (d, e) VOCs emissions from S-oaks ( ) and T-oaks ( ) (for T, Y = 1; S, Y = 0) [* indicates the significant X-variable in themodel describing Y; significance tested with modified Jack-knife uncertainty test ‘Marten’s test’ (Martens & Martens 2000)]. (c, f) Linearrelationship between measured ( ) and predicted ( ) Y-values of model calibrated ( ) with random subset of data and validated ( )using Martens’s test. Nomenclature: (E)- b-O = (E)-b-ocimene; (Z)- b-O = (Z)-b-ocimene; L. = limonene; a-C = a-cubebene;b-B = b-bourbonene; a-F = a-farnesene; b-F = b-farnesene; (Z)-Fa = (Z)-farnesene; Ge = geraniol; G.D = germacrene D; G = germacrene;G.B = germacrene B; b-C = b-cubebene; b-Ca = (E)-b-caryophyllene; DMNT = (E)-4,8-dimethyl-1,3,7-nonatriene; I. = indole;M. = myrcene; M.S. = methyl salicylate. PLSR, partial least square regression; TPS, terpene synthase; VOC, volatile organic compound.
10 A. Ghirardo et al.
© 2012 Blackwell Publishing Ltd, Plant, Cell and Environment

The difference in the odour profiles between resistantand susceptible pedunculate oaks likely explains the clearpreference of female moths for S-oaks. The repellent char-acteristic of T-oaks could be ascribed to the higher ratio ofsesquiterpenes, such as a-farnesene and germacrene D ashas been shown for the forest tent caterpillar on poplars(Arimura, Huber & Bohlmann 2004), in relation to DMNT,which agrees well with the results of a recent study (Chaet al. 2011), demonstrating that increasing proportions ofsesquiterpenes in the odour profile directly decrease attrac-tiveness to Paralobesia viteana Clemens (Lepidoptera: Tor-tricidae) females. The importance of odour profiles forinsect decisions has also been reported in other studies(Dicke 1994; Bruce & Pickett 2011). Based on these studies,it is clear that insects react to specific ratios of odours, ratherthan to the absolute abundance of individual volatiles. Invitro analysis of TPS activities demonstrated that the differ-ences in the volatile bouquets of S-oaks and T-oaks coin-cided with different activation of TPS activities. Thus, thedifferences in induction capabilities with respect to TPSactivities seem to result in two blends of HIPVs that areperceived very differently by the moths and therefore rep-resent the reason for their preferences for S-oaks ratherthan T-oaks (Fig. 7).
Two oak phenotypes and two strategies: hidingversus attracting natural enemies of herbivores
The ecologically effective defence strategy against thegreen oak leaf roller appeared to have a lower capabilityof emitting attractant HIPVs combined with higher emis-sions of potentially repellent HIPVs. Our data leave us tosuggest that the ecologically effective defence mechanismof T-oaks is based on their specific, for female moths less-attractive, odour profile compared with the highly attrac-tive odour blend of S-oaks. Among the multifunction rolesof VOC, highly and rapidly responding trees will generallybenefit from the protective effects of VOC against abioticstress (Vickers et al. 2009; Loreto & Schnitzler 2010), theconsequences of pollinator attraction (Gershenzon &Dudareva 2007) and general defensive mutualism withanimals (Langenheim 1994). Conversely, they may be moresusceptible to insect infestation, with serious consequencesfor the plants’ survival. Thus, during insect outbreaks,plants that emit lower amounts of HIPVs for geneticreasons would paradoxically benefit from this general dis-advantage. This has been proven for the mountain pinebeetle: Pine populations with lower amounts of terpeneshave less beetle attacks than the ones with high amounts ofterpenes (Clark, Carroll & Huber 2010). Therefore, plantsmust find a balance between the beneficial and negativeeffects caused by VOC emissions (Allison & Hare 2009).This may somehow explain why plants, which are perma-nently subjected to selection, often exhibit large intra-species variability in constitutive (Staudt et al. 2001) andinducible (Huang et al. 2010; Snoeren et al. 2010) VOCemissions.
Figure 6. PCA and PLSR of phenolic compounds of leavesfrom S-oak and T-oaks. (a) Scores of PCA, (b) scores and (c) X-and Y-loading (for T, Y = 1; S, Y = 0) of PLSR computed fromphenolic composition of S-oaks ( ) and T-oaks ( ) harvestedfrom young (Y) and older (O) plants. Young (Y) leaves weretaken after the T. viridana larval-feeding experiments and old(O) leaves after Y-tube experiments [* indicates the significantX-variable in the model describing Y; significance tested withmodified Jack-knife uncertainty test (Martens & Martens 2000)].PCA, principal component analysis; PLSR, partial least squareregression.
Detrimental effect of HIPV emissions in oaks 11
© 2012 Blackwell Publishing Ltd, Plant, Cell and Environment

In this study, the blend of oak volatiles played the crucialrole as a defensive mechanism against T. viridana. This con-clusion is supported by the fact that larvae that fed on thedifferent oak phenotypes showed similar mortality ratesand performances.Thus, the lower defoliation rate of T-oakscompared with S-oaks could not be explained by the abilityof resistant trees to actively and directly reduce the numberof eggs or larvae with toxic, secondary metabolic com-pounds to directly benefit the plants through decreasingherbivore numbers (Summers & Felton 1993). The numberof hatched T. viridana larvae from branches of T-oaks andS-oaks did not differ, though unfortunately the numberswere too low for statistical analysis of significance.
Although the mortality of larvae developing on T-oaks orS-oaks was similar (Fig. 7), larvae that developed on sus-ceptible plants consumed less plant biomass for a similarpupal weight than larvae grown on resistant oaks. Thus, the
leaves of T-oaks provide a poorer quality for the moth’soffspring than the leaves of S-oaks.
Indeed, our analysis of soluble polyphenols revealedhigher constitutive concentrations of some quercetin3-glycosides and the proanthocyanidin precursor catechinin the leaves of the resistant phenotypes rather than inthe leaves of the susceptible phenotypes. Quercetin3-glycosides are flavonols with oxidative and antioxidativeproperties (Buer, Imin & Djordjevic 2010) that may haveconsequences for larval food digestion. Proanthocyanidins(condensed flavan-3-ols), which typically accumulate in oakleaves during development (Feeny & Bostock 1968), alsopossess antioxidant (Luximon-Ramma et al. 2002), antibi-otic (Karioti et al. 2011) and digestive-retardant (Hagerman& Butler 1981) properties. However, whether the statisti-cally observed links between leaf quality and constitutivepolyphenol content and between polyphenol and HIPV
Figure 7. Schematic overview of the life cycle of T. viridana and its interaction with oak phenotypes. Scheme shows in three differentphases the interaction between Q. robur and T. viridana. (1) Decision phase of naive adult female moths during oviposition. (2)Developmental phase of T. viridana; (right) mortality rate of larvae developed on leaves from either S-oaks or T-oaks; (left) larvalpreferences between leaves from S-oaks or T-oaks. (3) Herbivory-induced volatile defence responses phase and associated keycompounds. Insect’s and plant’s activities/responses are located on white and grey backgrounds, respectively. Susceptible (S) and resistant(T) oaks are abbreviated for convenience. HIPV, herbivore-induced plant volatile.
12 A. Ghirardo et al.
© 2012 Blackwell Publishing Ltd, Plant, Cell and Environment

patterns (and therefore the behaviour of adults) existmechanistically must be addressed in future experiments.Poorer leaf quality might also be due to a higher amount ofinhibitors of trypsin-like proteases in T-oaks, as it isassumed that protease inhibitors are important mediatorsof resistance of oaks towards the green oak leaf roller(Ivashov, Simchuk & Medvedkov 2001). An initial,however, not yet finished transcriptome analysis ofdamaged S-oaks and T-oaks revealed twofold higherexpression rates of trypsin-like inhibitors in T-oaks than inS-oaks (unpublished data).
Therefore, adult females do indeed select the best feedingplant for their progenies in the next year, and the blend ofvolatiles emitted from S-oaks seemed to be the apparentindicator of food quality. The present data indicate thatthese moths have evolved the ability to use chemical signalssent by the plants to another recipient (i.e. natural enemies)to precisely relocate their host. A similar system works inNicotiana attenuata plants where a transgenic approach hasbeen chosen. For this species, it could be shown thatsilenced GLV production reduces the plant’s apparency toherbivores (Halitschke et al. 2008). However, this group ofvolatiles does not differ in its emission pattern in T-oaks andin S-oaks.
Co-evolution of host plant and herbivores
Plant defensive mechanisms evolve continuously tocombat the constant insect pressure exerted against them.Insects constantly react to the defence mechanisms of thehost plants. Host trees and herbivores are species withextremely different generation times and mobility (Hamil-ton, Axelrod & Tanese 1990). Oaks can first reproduce atan age of approximately 30–50 years and any obtainedadaptations will only be manifested in its offspring at thattime. In contrast, the annual reproduction cycles of mothsallow them to adapt much more quickly. With respect tothe re-colonization event that took place after the lastice age, T. viridana has had many more generations toundergo adaptation than the pedunculate oaks. Therefore,T. viridana might have evolved how to recognize andselect the most appropriate plants through associationwith their volatile cues. It became obvious at the larvalstage that the behavioural decision depended (to a certainextent) on imprinting during development on a specificleaf type. It is known that in many herbivorous insects,dietary experience with specific plants induces food pref-erences (e.g. Bernays & Weiss 1996). Only recently, a‘tuning’ of the taste sensilla of the tobacco hornwormManduca sexta to the chemical composition of particularfoliage has been described (Glendinning et al. 2009). Thus,a preference of larvae reared on S-oaks for the S-oaks andreared on T-oaks for the T-oaks should be expected. ForS-oaks, our results confirm this assumption. However, theresults for the larvae reared on T-oaks may have revealedan intermediate reaction of both induced food preferencetogether with the repellent odour (or taste) of poorerfood quality.
Considering the lower defoliation rate of T-oaks, we mustask why the number of individuals of this phenotype in thenatural habitat is low compared with the high number ofS-oaks in the near surroundings. It is possible that this isbecause under non-outbreak conditions, HIPV emissionsrepresent a consistent advantage. One prediction of co-evolutionary theory is that plants are at a disadvantagewhen investing in defence traits in the absence of herbivo-rous insects (Whittaker & Feeny 1971). The resistant phe-notypes might express constitutive, cost-intensive defencemechanisms, which would potentially be indicated by dif-ferences in the pattern of bioactive phenolic compounds orsome other disadvantage, such as a lower germinationcapacity, hindering the more widespread distribution ofthese phenotypes. Additionally, hiding is much more effec-tive when only a few individuals are following this strategy.
In summary, we demonstrated that pedunculate oak’ssusceptibility to herbivory is triggered by the decision-making process of adult T. viridana females searching forappropriate host trees (Fig. 7). Pedunculate oak trees‘talking more loudly’ (Baldwin et al. 2006) were heardbetter and, consequently, were even more infested thansilent trees. Hence, oak fitness was not found to benefit fromtypical HIPVs in this study. The natural variation in thequantity and blend composition of VOCs emitted from agiven plant species (Dicke & Baldwin 2010) might repre-sent the innate ability to avoid pest infestation and rapidplant declines. Previously explained generic tritrophic inter-action mechanisms are therefore much more complex thanhas been thought. In natural ecosystems, beneficial effectsof plants’ volatile emissions need to be balanced againstoccasional negative costs.
ACKNOWLEDGMENTS
This project is financially supported by the ‘DeutscheForschungsgemeinschaft’ (DFG, no. FL 263/19-1), Bonn,Germany (http://www.dfg.de). We thank our studentsUlrike Hinz, Inken Michaelsen and Stefanie Thomsen (allvTI) for performing some of the behavioural experimentsand our technical assistants Susanne Jelkmann (vTI), InaZimmer (EUS) and Susanne Stich (BIOP) for their excel-lent laboratory work. We would like to give special thanksto Maaria Rosenkranz (EUS), André Kessler (CornellUniversity) and Marcel Dicke (Wageningen University)for their comments on the manuscript. The images inFig. 1 were provided by (from left to right) Saskia Lie-berei, Andrea Ghirardo, Stefanie Thomsen and HilkeSchroeder.
REFERENCES
Allison J.D. & Hare D.J. (2009) Learned and naïve natural enemyresponses and the interpretation of volatile organic compoundsas cues or signals. New Phytologist 184, 768–782.
Appel H.M. & Maines L.W. (1995) The influence of host plant ongut conditions of gypsy moth (Lymantria dispar) caterpillars.Journal of Insect Physiology 41, 241–246.
Detrimental effect of HIPV emissions in oaks 13
© 2012 Blackwell Publishing Ltd, Plant, Cell and Environment

Arimura G., Ozawa R., Shimoda T., Nishioka T., Boland W. &Takabayashi J. (2000) Herbivory-induced volatiles elicit defencegenes in lima bean leaves. Nature 406, 512–515.
Arimura G., Huber D.P.W. & Bohlmann J. (2004) Forest tent cat-erpillars (Malacosoma disstria) induce local and systemic diurnalemissions of terpenoid volatiles in hybrid poplar (Populus tri-chocarpa ¥ deltoides): cDNA cloning, functional characteriza-tion, and patterns of gene expression of (-)-germacrene Dsynthase, PtdTPS1. The Plant Journal 37, 603–616.
Baldwin I.T., Halitschke R., Paschold A., von Dahl C.C. & PrestonC.A. (2006) Volatile signaling in plant-plant interactions: ‘talkingtrees’ in the genomics era. Science 311, 812–815.
Becerra J.X. (2003) Synchronous coadaptation in an ancient case ofherbivory. Proceedings of the National Academy of Sciences ofthe United States of America 100, 12804–12807.
Bernays E.A. & Weiss M.R. (1996) Induced food preferences incaterpillars: the need to identify mechanisms. EntomologiaExperimentalis et Applicata 78, 1–8.
Brilli F., Ciccioli P., Frattoni M., Prestininzim M., Spanedda A.F. &Loreto F. (2009) Constitutive and herbivore-induced monoter-penes emitted by Populus x euroamericana leaves are keyvolatiles that orient Chrysomela populi beetles. Plant, Cell &Environment 32, 542–552.
Bruce T.J.A. & Pickett J.A. (2011) Perception of plant volatileblends by herbivorous insects – finding the right mix. Phytochem-istry 72, 1605–1611.
Buer C.S., Imin N. & Djordjevic M.A. (2010) Flavonoids: new rolesfor old molecules. Journal of Integrative Plant Biology 52,98–111.
Cha D.H., Nojima S., Hesler S.P., Zhang A., Linn C.E., Roelofs W.L.& Loeb G.M. (2008) Identification and field evaluation of grapeshoot volatiles attractive to female grape berry moth (Paralobe-sia viteana). Journal of Chemical Ecology 34, 1180–1189.
Cha D.H., Linn C.E. Jr, Teal P.E.A., Zhang A., Roelofs W.L. &Loeb G.M. (2011) Eavesdropping on plant volatiles by a special-ist moth: significance of ratio and concentration. PLoS ONE 6,e17033.
Clark E.L., Carroll A.L. & Huber D.P.W. (2010) Differences inlodgepole pine constitutive terpene profile across a geographicrange in British Columbia and the correlation to historical attackby mountain pine beetle, Dendroctonus ponderosae Hopkins(Coleoptera: Curculionidae). The Canadian Entomologist 142,557–573.
Curtu A.L., Gailing O. & Finkeldey R. (2009) Patterns of contem-porary hybridization inferred from paternity analysis in a four-oak-species forest. BMC Evolutionary Biology 9, 284.
Degenhardt J. (2009) Indirect defense responses to herbivory ingrasses. Plant Physiology 149, 96–102.
Dicke M. (1994) Local and systemic production of volatileherbivore-induced terpenoids: their role in plant-carnivoremutualism. Journal of Plant Physiology 143, 465–472.
Dicke M. & Baldwin I.T. (2010) The evolutionary context forherbivore-induced plant volatiles: beyond the ‘cry for help’.Trends in Plant Science 15, 167–175.
Dicke M. & Sabelis M.W. (1988) How plants obtain predatorymites as bodyguards. Netherland Journal of Zoology 38, 148–165.
Dudareva N., Pichersky E. & Gershenzon J. (2004) Biochemistry ofplant volatiles. Plant Physiology 135, 1893–1902.
Ehrlich P.R. & Raven P.H. (1964) Butterflies and plants: a study inco-evolution. Evolution 18, 586–608.
Esbensen K.H. (2002) Multivariate Data Analysis-in Practice. AnIntroduction to Multivariate Data Analysis and ExperimentalDesign 5th edn. CAMO Process AS, Oslo, Norway.
Evers J., Franz C., Körver F. & Ziegler C. (1997) Waldbäume –Bilderserien zur Einschätzung von Kronenverlichtungen beiWaldbäumen. M. Faste, Kassel, Germany.
Feeny P.P. (1975) Biochemical coevolution between plants andtheir insect herbivores. In Coevolution of Animals and Plants(eds L.E. Gilbert & P.H. Raven), pp. 1–19. University of TexasPress, Austin, TX, USA and London, UK.
Feeny P.P. & Bostock H. (1968) Seasonal changes in the tannincontent of oak leaves. Phytochemistry 7, 871–880.
Gasow H. (1925) Der grüne Eichenwickler (Tortrix viridana Linné)als Forstschädling. Arbeiten der Biologischen Reichsanstalt 12,355–508.
Gershenzon J. & Dudareva N. (2007) The function of terpenenatural products in the natural world. Nature Chemical Biology3, 408–414.
Ghirardo A., Koch K., Taipale R., Zimmer I., Schnitzler J.P. &Rinne J. (2010) Determination of de novo and pool emissions ofterpenes from four common boreal/alpine trees by 13CO2 label-ing and PTR-MS analysis. Plant, Cell & Environment 33, 781–792.
Ghirardo A., Gutknecht J., Zimmer I., Brüggemann N. & SchnitzlerJ.P. (2011) Biogenic volatile organic compound and respiratoryCO2 emissions after 13C-labeling: online tracing of C transloca-tion dynamics in poplar plants. PLoS ONE 6, e17393.
Glendinning J.I., Foley C., Loncar I. & Rai M. (2009) Inducedpreference for host plant chemicals in the tobacco hornworm:contribution of olfaction and taste. Journal of ComparativePhysiology A 195, 591–601.
Hagerman A.E. & Butler L.G. (1981) The specificity ofproanthcyanidin-protein interactions. Journal of BiologicalChemistry 256, 4494–4497.
Halitschke R., Stenberg J.A., Kessler D., Kessler A. & Baldwin I.T.(2008) Shared signals – ‘alarm calls’ from plants increase appar-ency to herbivores and their enemies in nature. Ecology Letters11, 24–34.
Hamilton W.D., Axelrod R. & Tanese R. (1990) Sexual reproduc-tion as an adaptation to resist parasites. Proceedings of theNational Academy of Sciences of the United States of America 87,3566–3573.
Hansel A., Jordan A., Holzinger R., Prazeller P., Vogel W. & Lind-inger W. (1995) Proton transfer reaction mass spectrometry:on-line trace gas analysis at ppb level. International Journal ofMass Spectrometry 149, 609–619.
Honda K. (1995) Chemical basis of differential oviposition by lepi-dopterous insects. Archives of Insect Biochemistry and Physiol-ogy 30, 1–23.
Huang M., Abel C., Sohrabi R., Petri J., Haupt I., Cosimano J.,Gershenzon J. & Tholl D. (2010) Variation of herbivore-induced volatile terpenes among Arabidopsis ecotypes dependson allelic differences and subcellular targeting of two terpenesynthases, TPS02 and TPS03. Plant Physiology 153, 1293–1310.
Hunter M.D. (1990) Differential susceptibility to variable plantphenology and its role in competition between two insect herbi-vores on oak. Ecological Entomology 15, 401–408.
Ivashov A.V., Simchuk A.P. & Medvedkov D.A. (2001) Possiblerole of inhibitors of trypsin-like proteases in the resistance ofoaks to damage by oak leaf roller Tortrix viridana L. andgypsy moth Lymantria dispar L. Ecological Entomology 26, 664–668.
Ivashov A.V., Boyko G.E. & Simchuk A.P. (2002) The role of hostplant phenology in the development of the oak leaf roller moth,Tortrix viridana L. (Lepidoptera: Tortricidae). Forest Ecologyand Management 157, 7–14.
Kappers I.F., Aharoni A., van Herpen T.W., Luckerhoff L.L.,Dicke M. & Bouwmeester H.J. (2005) Genetic engineering ofterpenoid metabolism attracts bodyguards to Arabidopsis.Science 309, 2070–2072.
Kapuscinski J. (1995) DAPI: A DNA-specific fluorescent probe.Biotechnic and Histochemistry 70, 220–232.
14 A. Ghirardo et al.
© 2012 Blackwell Publishing Ltd, Plant, Cell and Environment

Karioti A., Sokovic M., Koukoulitsa C., Bilia A.R. & Skaltsa H.(2011) Antimicrobial properties of Quercus ilex L. proanthocya-nidin dimers and simple phenolics: evaluation of their synergisticactivity with conventional antimicrobials and prediction of theirpharmacokinetic profile. Journal of Agricultural and FoodChemistry 59, 6412–6422.
Kessler A. & Baldwin I.T. (2001) Defensive function of herbivore-induced plant volatile emissions in nature. Science 1291, 2141–2144.
Kessler A. & Heil M. (2011) The multiple faces of indirect defencesand their agents of natural selection. Functional Ecology 25,348–357.
Köksal M., Zimmer I., Schnitzler J.P. & Christianson D.W. (2010)Structure of isoprene synthase illuminates the mechanism ofteragram atmospheric carbon emission. Journal of MolecularBiology 402, 363–373.
Langenheim J.H. (1994) Higher plant terpenoids: a phytocentricoverview of their ecological roles. Journal of Chemical Ecology20, 1223–1280.
Lattanzio V., Lattanzio V.M.T. & Cardinali A. (2006) Role of phe-nolics in the resistance mechanisms of plant against fungalpathogens and insects. In Phytochemistry: Advances in Research(ed. F. Imperato), pp. 23–67. Research Signpost, Kerala, India.
Loreto F. & Schnitzler J.P. (2010) Abiotic stresses and inducedBVOCs. Trends in Plant Science 15, 154–166.
Luximon-Ramma A., Bahorun T., Soobrattee M.A. & Aruoma O.I.(2002) Antioxidant activities of phenolic, proanthocyanidin, andflavonoid components in extracts of cassia fistula. Journal ofAgricultural and Food Chemistry 50, 5042–5047.
Martens H. & Martens M. (2000) Modified jack-knife estimationof parameter uncertainty in bilinear modelling (PLS-R). FoodQuality and Preference 11, 5–16.
Mauricio R. & Rausher M.D. (1997) Experimental manipulation ofputative selective agents provides evidence for the role ofnatural enemies in the evolution of plant defense. Evolution 51,1435–1444.
Nosil P., Crespl B.J. & Sandoval C.P. (2002) Host-plant adaptationdriver the parallel evolution of reproductive isolation. Nature417, 440–443.
Pichersky E. & Gershenzon J. (2002) The formation and functionof plant volatiles: perfumes for pollinator attraction and defense.Current Opinion in Plant Biology 5, 237–243.
Pichersky E., Noel J.P. & Dudareva N. (2006) Biosynthesis ofplant volatiles: nature’s diversity and ingenuity. Science 311, 808–811.
Reinhard J., Srivivasan M.V. & Zhang S. (2004) Scent-triggerednavigation in honeybees. Nature 427, 411.
Renwick J.A.A. & Chew F.S. (1994) Oviposition behavior inLepidoptera. Annual Review of Entomology 39, 377–400.
Schnitzler J.P., Steinbrecher R., Zimmer I., Steigner D. & FladungM. (2004) Hybridisation of European oaks (Quercus ilex ¥ Q.robur) results in a mixed isoprenoid emitter type. Plant, Cell &Environment 27, 585–593.
Schoonhoven L.M., van Loon J.J.A. & Dicke M. (2005) Insect-PlantBiology. Oxford University Press, Oxford, UK.
Schröder H. (2010) Sommerveredelung bei Eichen – eine Erfolgs-geschichte. AFZ-Der Wald 5, 16–17.
Schröder R. & Hilker M. (2008) The relevance of background odorin resource location by insects: a behavioral approach. Bio-science 58, 308–316.
Schroeder H. & Degen B. (2008) Genetic structure of the greenoak leaf roller (Tortrix viridana L.) and one of its hosts, Quercusrobur L. Forest Ecology and Management 256, 1270–1279.
Schütte F. (1957) Untersuchungen über die Populationsdynamikdes Eichenwicklers (Tortrix viridana L.). Zeitschrift fürAngewandte Entomologie 40, 285–331.
Scotti-Saintagne C., Mariette S., Porth I., Goicoechea P.G., Barre-neche T., Bodénès C., Burg K. & Kremer A. (2004) Genomescanning for interspecific differentiation between two closelyrelated oak species [Quercus robur L. and Q. petraea [(Matt.)Liebl.]. Genetics 168, 1615–1626.
Snoeren T.A., Kappers I.F., Broekgaarden C., Mumm R., Dicke M.& Bouwmeester H.J. (2010) Natural variation in herbivore-induced volatiles in Arabidopsis thaliana. Journal of Experimen-tal Botany 61, 3041–3056.
Staudt M., Mandl N., Joffre R. & Rambal S. (2001) Intra-specificvariability of monoterpene composition emitted by Quercus ilexleaves. Canadian Journal of Forest Research 31, 174–180.
Summers C.B. & Felton G.W. (1993) Prooxidant effects of phenolicacids on the generalist herbivore Helicoverpa zea (Lepidoptera:Noctuidae): potential mode of action for phenolic compounds inplant anti-herbivore chemistry. Insect Biochemistry and Molecu-lar Biology 24, 943–953.
Takabayashi J. & Dicke M. (1996) Plant-carnivore mutualismthrough herbivore-induced carnivore attractants. Trends in PlantScience 4, 109–113.
Turunen M., Heller W., Stich S., Sandermann H., Sutinen M.L. &Norokorpi Y. (1999) The effects of UV exclusion on the solublephenolics of young Scots pine seedlings in the subarctic. Envi-ronmental Pollution 106, 219–228.
Vickers C.E., Gershenzon J., Lerdau M.T. & Loreto F. (2009) Aunified mechanism of action for volatile isoprenoids in plantabiotic stress. Nature Chemical Biology 5, 283–291.
Whittaker R.H. & Feeny P.P. (1971) Allelochemicals – chemicalinteractions between species. Science 171, 157–770.
Received 9 March 2012; received in revised form 15 May 2012;accepted for publication 16 May 2012
SUPPORTING INFORMATION
Additional supporting information may be found in theonline version of this article.
Figure S1. Y-tube experiments with larvae to test theoptical stimulus from different coloured leaves. Larvaewere tested for preferences between leaves taken fromS-oaks ( ) or T-oaks ( ) or red and green or white andgreen paper-cut ( ) (with oak-leaf shape). Decision wasclassified as ‘no decision’ when larvae did not make a choicewithin 20 min. Differences in choice behaviour betweenS-oaks and T-oaks were tested by z-test; an asterisk meansP < 0.01.Figure S2. Analyses of phenolic compounds in leaves fromS-oak and T-oaks. (a) HPLC chromatogram of methanolicextract recorded at 280 nm (as described in Turunen et al.1999). Phenolic compounds were separated in four groups;gallates and hydroxycinnamic acid derivatives, flavonol gly-cosides, and acylated flavonol glycosides with respect topure standard references. Numbers indicate peaks wherediode array spectra (250–440 nm) were compared with ref-erence compounds and literature data. Peak intensitiesof phenolic compounds differed between S-oaks ( ) andT-oaks ( ). (b) peak #10 (catechin, as catechin equivalents);(c) peak #13 (unknown, as gallic acid equivalents); (d) peak#20 (quercetin glycoside, as quercetin 3-glucoside equiva-lents); (e) peak #21 (kaempferol glycoside, as kaempferol3-glucoside equivalents); (f) peak #23 (quercetin glycoside,
Detrimental effect of HIPV emissions in oaks 15
© 2012 Blackwell Publishing Ltd, Plant, Cell and Environment

as quercetin 3-glucoside equivalents); (g) peak #34 (mono-acylated flavonol glycoside, as tiliroside equivalents). Theassignment of peaks to substance groups is based on theabsorption spectra and retention time. ‘Young’ and ‘old’corresponds to leaves taken after the T. viridana larval-feeding and Y-tube experiments, respectively; intact anddamaged refers to healthy and larvae-penetrated leavesharvested from the same plant. (n = 4 (old)/8 (young) + SE;significant difference is given with different letter forP < 0.05; * = P < 0.01).Figure S3. Online emission profiles of constitutive andinducible VOCs before and during larval feeding. (a) Greenleaf volatiles (GLV) and (b) acetaldehyde of phenotypicallydifferent S-oaks (47b, 13b) and T-oaks (17a, 2a). Each plantwas fed with four larvae of T. viridana ( ). Dark phasesare symbolized in grey ( ). Cuvette condition: T = 19 °C,light = 50–150 PAR. Data represent means of threeexperiments � SE.Figure S4. Online emission profiles of constitutive VOCsbefore and during T. viridana larval feeding. (a) Ethanol, (b)methanol, and (c) isoprene of phenotypically differentS-oaks (47b, 13b) and T-oaks (17a, 2a). Each plant was fedwith four larvae of T. viridana ( ). Dark phases aresymbolized in grey ( ). Cuvette condition: T = 19 °C,
light = 50–150 PAR. Data represent means of threeexperiments � SE.Figure S5. Analysis of in vitro terpene synthase (TPS)activities. (a) Monoterpene, (b) sesquiterpene activities ofTPS extracted from herbivore-fed leaves of phenotypicallydifferent S-oaks (47b, 13b) and T-oaks (17a, 2a). Shadedbars represent TPS activities from leaves which weredirectly pitted by larvae. For the analysis of in vitro synthe-sis of terpenes from geranyl diphosphate (GPP) and farne-syl diphosphate (FPP), the assay described elsewhere(Köksal et al. 2010) was used. In this assay, enzymaticallyproduced terpenes were trapped from the aqueous matrixby stir bar sorptive extraction (SBSE) with polydimethylsi-loxane (PDMS) coated stir bars (Twister, PDMS film thick-ness 0.5 mm, Gerstel, Mülheim, Germany). Removing thestir bars terminated the enzymatic reactions. After rinsingin de-ionized H2O, the twisters were analysed using TDUand GC-MS as described in Materials and Methods.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materials sup-plied by the authors. Any queries (other than missing mate-rial) should be directed to the corresponding author for thearticle.
16 A. Ghirardo et al.
© 2012 Blackwell Publishing Ltd, Plant, Cell and Environment
Copyright © 2022 FDOKUMEN




















